ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится большей частью к способам и структурам для получения и использования слитых белков, содержащих ингибитор ангиогенеза (развития кровеносных сосудов). Более конкретно, изобретение относится к способам и структурам для получения и использования слитых белков, называемых “иммунофузинами”, которые содержат область Fc иммуноглобулина и ингибитор ангиогенеза.
УРОВЕНЬ ТЕХНИКИ
Были открыты два активных ингибитора ангиогенеза: ангиостатин (O’Reilly и др. // Cell. 1994. Т.79. С.315) и эндостатин (O’Reilly и др. // Cell. 1997. Т.88. С. 277). Было обнаружено, что они естественным образом продуцируются первичными опухолями. Оба белка специфически подавляют пролиферацию клеток эндотелия и ингибируют рост опухоли путем блокирования ангиогенеза - образования новых кровеносных сосудов, которые питают опухоль. Исследования показали, что эти ингибиторы ангиогенеза нетоксичны даже в очень высоких дозах и могут подавлять рост метастазов, а первичные опухоли могут регрессировать до размера, неразличимого под микроскопом. Оба ингибитора были идентифицированы как полученные протеолизом фрагменты намного больших интактных молекул. Было обнаружено, что ангиостатин является фрагментом плазминогена, а эндостатин - фрагментом коллагена XVIII.
Эти два белка привлекли пристальное внимание онкологов, так как было показано, что они подавляют у мышей рост многих различных типов опухолей, без видимых побочных эффектов или появления лекарственной устойчивости. Традиционная химиотерапия обычно приводит к приобретенной лекарственной устойчивости, которая главным образом вызвана генетической нестабильностью раковых клеток. Терапия с использованием ингибиторов ангиогенеза действует не столько на раковые клетки, сколько на нормальные клетки эндотелия, которые поддерживают рост опухоли. Поскольку клетки эндотелия генетически стабильны, методы терапии с помощью ингибиторов ангиогенеза могут приводить к меньшей лекарственной устойчивости. Исследования показывают, что лекарственная устойчивость не развивалась у мышей, подвергавшихся длительной антиангиогенной терапии с помощью эндостатина. Более того, повторные циклы лечения мышей эндостатином приводили к длительной “спячке” опухолей, и после прекращения терапии рецидивы опухолей не наблюдались (Boehm и др. // Nature. 1997. Т.390. С.404).
Несмотря на многообещающие результаты исследований на мышах, растворимые активные ангиостатин и эндостатин с квалификацией, пригодной для клинического использования, не удалось получить с помощью систем экспрессии на основе Е.соli, бакуловирусов, клеток дрожжей и млекопитающих. Результатом экспрессии в Е.соli были нерастворимые белковые агрегаты неопределенного состава, непригодные для инъекции людям. Для других способов продукции, включая системы экспрессии на основе бакуловирусов и клеток млекопитающих, был характерен очень низкий уровень продукции рекомбинантных белков (O’Reilly и др. // Cell. 1997. Т.88. С.277).
Низкий выход в использованных до настоящего времени системах экспрессии можно объяснить тем, что и ангиостатин, и эндостатин являются внутренними фрагментами белков намного большего размера. Усеченные белки не могут сворачиваться нужным образом в отсутствие аминокислотных остатков, отсекаемых от молекул-предшественников. Например, ангиостатин содержит 26 остатков цистеина, образующих множественные дисульфидные связи. Экспрессия собственно ангиостатина может не обеспечивать оптимальных условий для того, чтобы эти множественные дисульфидные связи сформировались нужным образом на стадии секреции. Что касается рекомбинантного белка эндостатина, при продуцировании клетками Е.соli он осаждается в ходе диализа - по-видимому, вследствие гидрофобности эндостатина (O’Reilly и др. // Cell. 1997. Т.88. С.277).
Основным препятствием в использовании ангиостатина и эндостатина в их нынешнем виде является то, что для достижения нужного клинического эффекта необходимо ежедневно вводить большие количества белков в течение длительного срока, от нескольких недель до нескольких месяцев. Например, в общепринятых схемах опытов на мышах для получения оптимальной эффективности необходимы дозировки эндостатина 20 мг/кг ежедневно (Boehm и др. // Nature. 1997. Т.390. С.404). Поскольку требуется срочная проверка клинической эффективности эндостатина и ангиостатина, важно разработать способ продукции, дающий большие количества материала клинической квалификации.
Одна из систем экспрессии, используемых для обеспечения высокого уровня экспрессии слитых белков в клетках млекопитающих, представляет собой конструкцию ДНК, которая кодирует сигнальную последовательность, участок Fc (константную часть) иммуноглобулина и целевой белок. Слитый продукт такой конструкции имеет общепринятое название “иммунофузин”. В виде иммунофузинов были успешно экспрессированы несколько целевых белков, включая IL2 (интерлейкин 2), CD26, Tat, Rev, OSF-2, (β-IG-H3, рецептор IgE, PSMA и gp120. Эти экспрессируемые конструкции раскрыты в патентах США №5541087 и №5726044, описания которых включены сюда ссылкой.
Основной целью экспрессии рекомбинантных слитых белков в клетках млекопитающих было попытаться придать гибридным молекулам новые или полезные свойства, например нужное сворачивание, повышенную растворимость, специфическое взаимодействие in vivo с цитокином или токсином, связывание с рецептором Fc, фиксацию комплемента, связывание с белком А, увеличенное время полужизни в кровотоке и повышенную способность проникновения через гематоэнцефалический барьер. Примеры рекомбинантных слитых белков, продуцируемых в клетках млекопитающих, включают иммуноконъюгаты цитокинов (Gillies и др. // Proc. Natl. Acad. Sci. USA. 1992. Т.89. С.1428; Gillies и др. // Bioconjugate Chemistry. 1993. Т.4. С.230), иммуноадгезины (Capon и др. // Nature. 1989. Т.337. С.525), иммунотоксины (Chaudhary и др. // Nature. 1989. Т.339. С.394) и конъюгат фактора роста нерва (Friden и др. // Science. 1993. Т.259. С.373).
Цель настоящего изобретения состоит в том, чтобы получить новые ДНК, обеспечивающие эффективную продукцию и секрецию ингибиторов ангиогенеза в разнообразных хозяевах - клетках млекопитающих. Другой целью настоящего изобретения является создание способов лечения животных нуклеиновыми кислотами, которые кодируют, или аминокислотными последовательностями, которые определяют белки-ингибиторы ангиогенеза, включая ненативные, биосинтетические или такие иным образом полученные искусственные белки, как белки, созданные путем рационального конструирования.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение предлагает способы и структуры, пригодные для получения и использования слитых белков, содержащих белок-ингибитор ангиогенеза. Слитые белки способны обеспечить высокий уровень экспрессии биологически активных белков-ингибиторов ангиогенеза. Белки-ингибиторы ангиогенеза могут быть затем отщеплены от слитого белка и объединены с фармацевтически пригодным носителем перед введением млекопитающему, например человеку. В качестве альтернативы нуклеотидные последовательности, кодирующие, или аминокислотные последовательности, определяющие содержащие ингибитор ангиогенеза слитые белки, могут быть объединены с фармацевтически пригодным носителем и введены млекопитающему.
В одном из аспектов изобретение предоставляет молекулы нуклеиновой кислоты, например молекулы ДНК или РНК, кодирующие слитый белок по данному изобретению. Молекула нуклеиновой кислоты кодирует сигнальную последовательность, область Fc (константную часть) иммуноглобулина и по меньшей мере один целевой белок, обозначаемый здесь также как белок-ингибитор ангиогенеза, выбранный из группы, состоящей из ангиостатина, эндостатина, фрагмента плазминогена с активностью ангиостатина, фрагмента коллагена XVIII с активностью эндостатина и их комбинаций. В предпочтительном осуществлении молекула нуклеиновой кислоты кодирует последовательно, в направлении от 5’-конца к 3’-концу, сигнальную последовательность, область Fc иммуноглобулина и последовательность целевого белка. В другом предпочтительном осуществлении молекула нуклеиновой кислоты кодирует последовательно, в направлении от 5’-конца к 3’-концу, сигнальную последовательность, последовательность целевого белка и область Fc иммуноглобулина.
Еще в одном предпочтительном осуществлении область Fc иммуноглобулина содержит шарнирную область иммуноглобулина и предпочтительно содержит по меньшей мере одну константную часть тяжелой цепи иммуноглобулина, например домен 2 константной части тяжелой цепи (CH2), домен 3 константной части тяжелой цепи (СН3) и, в зависимости от типа иммуноглобулина, используемого для формирования области Fc, но не обязательно, - домен 4 константной части тяжелой цепи (СН4). В более предпочтительном осуществлении область Fc иммуноглобулина содержит шарнирную область, домен CH2 и домен СН3. При определенных обстоятельствах область Fc иммуноглобулина предпочтительно лишена домена CH1. Хотя области Fc иммуноглобулина могут иметь основой любой класс иммуноглобулинов, например IgA, IgD, IgE, IgG и IgM, предпочтительными являются области Fc иммуноглобулина на основе IgG (гамма-иммуноглобулина).
В ином осуществлении нуклеиновая кислота по настоящему изобретению может быть включена в функциональной связи в способный к репликации экспрессирующий вектор, который затем можно трансфецировать в клетку-хозяин млекопитающего. В другом предпочтительном осуществлении настоящее изобретение предоставляет клетки-хозяева, воспринимающие такие последовательности нуклеиновой кислоты, соответствующие данному изобретению.
В другом аспекте настоящее изобретение относится к слитому белку, содержащему область Fc иммуноглобулина, соединенную (либо непосредственно пептидной связью, либо через полипептидный мостик) с целевым белком, выбранным из группы, состоящей из ангиостатина, эндостатина, имеющего активность ангиостатина фрагмента плазминогена, имеющего активность эндостатина фрагмента коллагена XVIII, и их комбинаций. Целевой белок может быть сцеплен своим С-концом с N-концом области Fc иммуноглобулина. Однако в более предпочтительном варианте осуществления целевой белок сцеплен своим N-концом с С-концом области Fc иммуноглобулина.
В другом варианте осуществления слитый белок может содержать второй целевой белок, выбранный из группы, состоящей из ангиостатина, эндостатина, имеющего активность ангиостатина фрагмента плазминогена и имеющего активность эндостатина фрагмента коллагена XVIII. В конструкции этого типа первый и второй целевые белки могут быть одним и тем же белком или же разными белками. Например, в предпочтительном осуществлении слитый белок содержит первый целевой белок - ангиостатин, область Fc иммуноглобулина и второй целевой белок - эндостатин. Первый и второй целевые белки могут быть соединены вместе либо непосредственно, либо посредством полипептидного мостика. В качестве альтернативы оба целевых белка могут быть соединены либо непосредственно, либо с помощью полипептидного мостика с областью Fc иммуноглобулина. В последнем случае первый целевой белок присоединен к N-концу области Fc иммуноглобулина, а второй целевой белок присоединен к С-концу области Fc иммуноглобулина.
В другом варианте осуществления два слитых белка могут быть соединены или ковалентно, например, дисульфидной или пептидной связью или нековалентно, с получением мультимерного белка. В предпочтительном варианте осуществления два слитых белка соединены ковалентно с помощью одной или нескольких дисульфидных связей между остатками цистеина, предпочтительно расположенными внутри шарнирных областей иммуноглобулина, находящихся внутри областей Fc иммуноглобулина обоих цепей.
В предпочтительном варианте осуществления целевой белок содержит фрагмент плазминогена, имеющий молекулярный вес около 40 кДа и может содержать аминокислотную последовательность, как указано далее в последовательности SEQ ID NO: 3. В другом предпочтительном варианте осуществления целевой белок содержит фрагмент коллагена XVIII, имеющий аминокислотную последовательность, представленную далее в последовательности SEQ ID NO: 1. Кроме того, целевой белок может быть ангиостатином или эндостатином полной длины или же их биологически активными фрагментами. Источник целевого белка для создания определенных слитых белков зависит от намечаемого использования целевого белка. Например, если целевой белок должен вводиться людям, целевой белок предпочтительно должен быть человеческого происхождения.
В ином аспекте настоящее изобретение предусматривает способы получения слитого белка, содержащего область Fc иммуноглобулина и целевой белок, выбранный из группы, состоящей из ангиостатина, эндостатина, имеющего активность ангиостатина фрагмента плазминогена и имеющего активность эндостатина фрагмента коллагена XVIII. Способ включает этапы: (а) получения клетки млекопитающего, содержащей молекулу ДНК, кодирующую такой слитый белок, с сигнальной последовательностью или без нее, и (б) культивирования клетки млекопитающего для получения слитого белка. Полученный целевой белок может быть затем собран, упорядочен, если необходимо, иным образом и очищен с использованием обычных методов очистки, хорошо известных и используемых в данной области. Если допустить, что слитый белок содержит сайт протеолитического расщепления, расположенный между областью Fc иммуноглобулина и целевым белком, тогда целевой белок может быть выщеплен из слитого белка с помощью обычных протеолитических ферментов и при необходимости очищен перед использованием.
В ином аспекте настоящее изобретение предусматривает способы лечения млекопитающих, например людей, при необходимости применения терапии на основе ингибиторов ангиогенеза. Например, предполагается, что ингибиторы ангиогенеза в соответствии с настоящим изобретением можно вводить людям, имеющим опухоль. Лечение ингибитором ангиогенеза может замедлять или останавливать рост опухоли и, при определенных условиях, может приводить к регрессии опухоли. Лечение может включать введение млекопитающему ингибитора ангиогенеза в количестве, достаточном для замедления или остановки роста опухоли. Ингибитор ангиогенеза может быть в форме слитого белка или в виде нуклеиновой кислоты, предпочтительно функционально соединенной с экспрессирующим вектором, в комбинации с фармацевтически пригодным носителем.
Изложенное выше, а также другие цели, особенности и преимущества настоящего изобретения будут более понятными из нижеследующих подробного описания, графических материалов и формулы.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1A-1F представляют собой схематическое изображение типичных слитых белков с ингибитором ангиогенеза, сконструированных в соответствии с настоящим изобретением (см. примеры 10-15). На них изображены следующие конструкции.
Фиг.1А: Fc-1-й домен Крингла (Kringle) ангиостатина;
Фиг.1В: Fc-внутренняя часть 1-го домена Крингла ангиостатина;
Фиг.1С: Fc-эндостатин-мостик Gly-Ser (глицин-серин)-внутренняя часть 1-го домена Крингла ангиостатина;
Фиг.1D: Fc-эндостатин-мостик Gly-Ser-1-й домен Крингла ангиостатина;
Фиг.1Е: Fc-эндостатин-мостик Gly-Ser-ангиостатин;
Фиг.1F: ангиостатин-Рс-эндостатин.
Вертикальными линиями обозначены возможные дисульфидные связи, соединяющие остатки цистеина (С), находящиеся внутри шарнирной области молекулы Fc.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение предусматривает слитые белки, называемые здесь иммунофузинами, которое пригодны для производства в коммерческих количествах ингибиторов ангиогенеза с квалификацией, пригодной для клинического применения. Ингибиторы ангиогенеза перед использованием могут быть отщеплены от белковой конструкции иммунофузина. Однако предполагается, что иммунофузины или нуклеиновые кислоты, кодирующие иммунофузины, можно прямо вводить млекопитающим при необходимости лечения ингибитором ангиогенеза.
Поэтому настоящее изобретение предусматривает слитые белки, содержащие область Fc иммуноглобулина и по меньшей мере один целевой белок, называемый здесь ингибитором ангиогенеза. Ингибитор ангиогенеза предпочтительно выбран из группы, состоящей из ангиостатина, эндостатина, имеющего активность ангиостатина фрагмента плазминогена, имеющего активность эндостатина фрагмента коллагена XVIII. Предполагается однако, что и другие полипептиды, имеющие активность ингибитора ангиогенеза, которые известны в настоящее время или будут открыты позднее, могут быть экспрессированы как слитые белки описанного здесь типа.
Фиг.1A-1F иллюстрируют шесть типичных вариантов осуществления белковых конструкций, воплощающих настоящее изобретение. Поскольку предпочтительными являются димерные конструкции, все они изображены как димеры, сшитые парой дисульфидных связей между цистеинами, расположенными на соседствующих субъединицах. На чертежах показаны дисульфидные мостики, которые связывают участки двух областей Fc иммуноглобулина в шарнирной области иммуноглобулина и, таким образом, являются характерными для нативных форм этих молекул. Хотя конструкции, содержащие шарнирную область Fc, предпочтительны и многообещающи как терапевтические агенты, изобретение предполагает, что при необходимости может быть выбрана сшивка в других местах. Кроме того, при некоторых обстоятельствах димеры или мультимеры, полезные в практике применения изобретения, могут быть получены путем нековалентного связывания, например с помощью гидрофобных взаимодействий.
Так как гомодимерные конструкции являются важными вариантами осуществления изобретения, Фиг.1 иллюстрирует такие конструкции. Следует подчеркнуть, что гетеродимерные конструкции также пригодны, но они, как это известно специалистам в данной области, часто могут плохо поддаваться очистке. Однако могут быть сконструированы пригодные для ингибирования ангиогенеза у различных видов млекопитающих жизнеспособные конструкции, представляющие собой смесь гомодимеров и гетеродимеров. Например, одна цепь гетеродимерной структуры может содержать эндостатин, а другая может содержать ангиостатин.
На Фиг.1А представлена димерная конструкция, полученная в соответствии с процедурой, описанной далее в примере 10. Каждый мономер димера содержит область Fc иммуноглобулина (1), включающую шарнирную область, домен СН2 и домен СН3. Непосредственно к С-концу области Fc (1) прикреплен первый участок Крингла ангиостатина (2), включающий и внутреннее, и внешнее кольца. На Фиг.1В представлен второй вариант осуществления изобретения (см. пример 11). В нем имеется такая же область Fc, как на Фиг.1А, но на этот раз к С-концу области Fc (1) присоединено только внутреннее кольцо первой области ангиостатина (3). Фиг. с 1С по 1Е показывают различные варианты осуществления белковых конструкций в соответствии с настоящим изобретением, которые включают в качестве целевого белка несколько ингибиторов ангиогенеза, расположенных тандемом и соединенных мостиком. На Фиг.1С целевой белок содержит эндостатин полной длины (4), полипептидный мостик (5) и внутреннее кольцо первого участка Крингла ангиостатина (3). Фиг.1D показывает белок, содержащий такую же область Fc, как на Фиг.1А, и целевой белок, содержащий эндостатин полной длины (4), полипептидный мостик (5) и полную область 1-го участка Крингла ангиостатина (как внутреннее, так и внешнее кольца) (2). Фиг.1Е отличается от конструкции, изображенной на Фиг.1D, тем, что большая часть домена С-концевой части белка представляет собой копию ангиостатина полной длины (7).
Хотя Фиг.1А-1Е представляют конструкции типа Fc-X, где Х - целевой белок, предполагается, что конструкции типа X-Fc могут также быть полезными в практике осуществления изобретения. Кроме того, предполагается, что полезные белки в соответствии с настоящим изобретением могут быть обозначены также формулой X-Fc-X, где знаки Х могут обозначать одни и те же или различные целевые белки. Фиг.1F показывает такую конструкцию, которая содержит в направлении от N-конца к С-концу ангиостатин человека полной длины (7), область Fc иммуноглобулина человека (6), включая шарнирную область, и домен эндостатина человека полной длины (4).
Термин “ингибитор ангиогенеза”, как он использован здесь, обозначает любую полипептидную цепь, которая уменьшает или подавляет у млекопитающего образование новых кровеносных сосудов. При противораковой терапии ингибитор ангиогенеза уменьшает или подавляет образование новых кровеносных сосудов в или на опухоли, предпочтительно в или на солидной опухоли. Предполагается, что с помощью различных хорошо известных методов анализа можно идентифицировать и использовать в данной области полезные ингибиторы ангиогенеза. Такие методы анализа включают, например, тест на пролиферацию клеток эндотелия капилляров крупного рогатого скота, анализ с помощью хориоаллантоисной мембраны кур (ХАМ) или анализ на роговой оболочке глаз мышей. Однако предпочтителен анализ на ХАМ (см., например, работы O’Reilly и др. // Cell. 1994. Т.79. С.315-328 и O’Reilly и др. // Cell. 1997. Т.88. С.277-285, изложение которых включено сюда ссылкой). Вкратце, зародыши с интактными желтками извлекали из трехдневных оплодотворенных белых яиц и помещали в чашки Петри. После инкубации в течение 3 дней при 37°С в атмосфере 3% СО2 к хориоаллантоисной мембране индивидуального зародыша прикладывали метилцеллюлозный диск с нанесенным на него предполагаемым ингибитором ангиогенеза. После инкубации в течение приблизительно 48 час хориоаллантоисные мембраны исследовали под микроскопом на наличие зон ингибирования.
Предпочтительными ингибиторами ангиогенеза, пригодными в практике использования настоящего изобретения, являются, например, ангиостатин (O’Reilly и др. // Cell. 1994. Т.79. С.315-328 и патенты США №5733876, 5837682 и 5885795) и эндостатин (O’Reilly и др. // Cell. 1997. Т.88. С.277-285 и патент США №5854205). Как установлено ранее, ангиостатин и эндостатин являются специфическими ингибиторами пролиферации клеток эндотелия и способны подавлять рост опухолей путем блокирования ангиогенеза, то есть образования новых кровеносных сосудов, питающих опухоли.
Ангиостатин был идентифицирован как протеолитический фрагмент плазминогена (O’Reilly и др. // Cell. 1994. Т.79. С.315-328 и патенты США №5733876, 5837682 и 5885795, изложение которых включено сюда ссылкой). Конкретно, ангиостатин - это внутренний фрагмент плазминогена с молекулярной массой 38 кДа, содержащий по меньшей мере три из областей Крингла плазминогена. Эндостатин был идентифицирован как протеолитический фрагмент коллагена XVIII (O’Reilly и др. // Cell. 1997. Т.88. С.277-285 и патент США №5854205, изложение которых включено сюда ссылкой). Конкретно, эндостатин является С-концевым фрагментом коллагена XVIII с молекулярной массой 20 кДа. Термины “ангиостатин” и “эндостатин”, как они использованы здесь, означают не только белки полной длины, но также и их варианты и биологически активные фрагменты, а также биологически активные фрагменты соответственно плазминогена и коллагена XVIII. Термин “биологически активный фрагмент” в применении к ангиостатину обозначает любой белковый фрагмент плазминогена или ангиостатина, который имеет по меньшей мере 30%, более предпочтительно по меньшей мере 70% и наиболее предпочтительно по меньшей мере 90% активности ангиостатина полной длины, определенной тестом в ХАМ. Термин “биологически активный фрагмент” в применении к эндостатину обозначает любой белковый фрагмент коллагена XVIII или эндостатина, который имеет по меньшей мере 30%, более предпочтительно по меньшей мере 70% и наиболее предпочтительно по меньшей мере 90% активности эндостатина полной длины, определенной тестом в ХАМ.
Термин “варианты” включает особые разновидности и аллельные варианты, а также другие существующие в природе или возникающие отличным от природного способа варианты, например полученные обычными генетико-инженерными методами, которые по меньшей мере на 70% подобны или на 60% идентичны, более предпочтительно по меньшей мере на 75% подобны или на 65% идентичны, а наиболее предпочтительно по меньшей мере на 80% подобны или на 70% идентичны любой из существующих природных последовательностей эндостатина или ангиостатина, раскрытых здесь.
Чтобы определить, имеет ли рассматриваемый полипептид требуемую степень подобия или идентичности с полипептидом сравнения (референсным полипептидом), прежде всего проводят параллельное сопоставление рассматриваемой аминокислотной последовательности и аминокислотной последовательности сравнения, используя алгоритм динамического программирования, описанный в работе Smith и Waterman // J. Mol. Biol. 1981. Т.147. С.195-197, в комбинации с матрицей замещения BLOSUM62, описанной на Фиг.2 работы Henikoff и Henikoff “Amino acid substitution matrices from protein blocks” // Proc. Natl. Acad. Sci. USA. 1992. Т.89. С.10915-10919. Для настоящего изобретения подходящее значение критерия недостоверности заполнения пробела равно - 12, а подходящее значение критерия недостоверности удлинения просвета равно - 4. Компьютерные программы, выполняющие параллельные сопоставления последовательностей с использованием алгоритма Смита-Уотермана (Smith-Waterman) и матрицы BLOSUM62, например программное обеспечение GCG (Oxford Molecular Group, Oxford, Англия), являются коммерческой продукцией и широко используются специалистами в данной области.
После того как проведено параллельное сопоставление рассматриваемой и референсной последовательностей, можно рассчитать величину процента подобия. Последовательно сопоставляются индивидуальные аминокислоты в каждой последовательности с определением их подобия друг другу. Если значение матрицы BLOSUM62, соответствующее двум сопоставляемым аминокислотам, равно нулю или является отрицательной величиной, то подобие в пределах пары обозначается как ноль. В ином случае подобие в пределах пары обозначается как 1,0. Общее значение подобия является суммой подобий в пределах пар для сопоставляемых параллельно аминокислот. Затем общее значение подобия нормализуют путем его деления на число аминокислот в меньшей рассматриваемой или в референсной последовательности. Нормализованное общее значение и представляет собой процент подобия. В качестве альтернативы для вычисления процента идентичности снова поочередно сопоставляют выстроенные в ряд параллельно аминокислоты каждой последовательности. Если аминокислоты не идентичны, значение идентичности в пределах пары равно нулю; в ином случае значение идентичности в пределах пары равно 1,0. Общее значение идентичности является суммой идентичных выстроенных в ряд параллельно аминокислот. Затем общее значение идентичности нормализуют путем его деления на число аминокислот в меньшей рассматриваемой или в референсной последовательности. Нормализованное общее значение и представляет собой процент идентичности. Для вычисления процента подобия и идентичности вставками и делециями пренебрегают. Соответственно, в этих вычислениях не используют величины критерия недостоверности заполнения пробела, хотя они используются при первичном выстраивании последовательностей.
Заявленные здесь целевые белки экспрессируются как слитые белки, имеющие область Fc иммуноглобулина. Как известно, константная часть каждой тяжелой цепи иммуноглобулина состоит из 4 или 5 доменов. Домены обозначаются последовательно следующим образом: СН1-шарнир-СН2-СН3(-СН4). Последовательности ДНК доменов тяжелой цепи имеют перекрестную гомологию среди классов иммуноглобулинов, например домен CH2IgG гомологичен домену CH2IgA и IgD, а также домену СН3IgM и IgE.
Понятно, что в использованном здесь смысле термин “область Fc иммуноглобулина” означает часть карбоксильного конца константной области цепи иммуноглобулина, предпочтительно константной области тяжелой цепи иммуноглобулина, или ее части. Например, область Fc иммуноглобулина может содержать (1) домен CH1, домен CH2 и домен СН3; (2) домен CH1 и домен СН2; (3) домен CH1 и домен СН3; (4) домен CH2 и домен СН3 или (5) комбинацию двух или более доменов и шарнирную область иммуноглобулина. В предпочтительном осуществлении область Fc, отраженная в конструкции ДНК, включает по меньшей мере шарнирную область иммуноглобулина, домен CH2 и домен СН3 и предпочтительно лишена по меньшей мере домена CH1.
Общепринято, что предпочтительным классом иммуноглобулина, из которого происходит константная область тяжелой цепи, является IgG (Igγ) (γ подклассы 1, 2, 3 или 4). Могут быть использованы другие классы иммуноглобулинов: IgA (Igα), IgD (Igδ), IgE (Igε) и IgM (Igμ). Выбор подходящих константных областей тяжелой цепи иммуноглобулина подробно описан в патентах США №5541087 и 5726044. Считается, что квалификация специалистов в данной области достаточна при выборе конкретных последовательностей константной области тяжелой цепи иммуноглобулина среди определенных классов и подклассов иммуноглобулинов для достижения конкретного результата. Часть конструкции ДНК, кодирующая область Fc иммуноглобулина, предпочтительно содержит последовательности по меньшей мере для части шарнирного домена и по меньшей мере части домена СН3 области Fсγ или гомологичных доменов любого иммуноглобулина из IgA, IgD, IgE или IgM.
В зависимости от применения могут быть использованы гены константной области не только человека, но и других видов, например мыши или крысы. Обычно область Fc, используемая как партнер для соединения в ДНК-конструкции для иммунофузина, может происходить из любого вида млекопитающих. В том случае, когда нежелательно получение в клетке-хозяине или в животном иммунного ответа на область Fc, область Fc может происходить из того же вида, что и клетка-хозяин или животное. Например, если животное-хозяин или клетка-хозяин - человек, можно использовать Fc человека; подобным же образом в том случае, когда животное-хозяин или клетка-хозяин - мышь, можно использовать мышиный Fc. Кроме того, можно использовать также замещение или делецию конструкций этих константных областей, где один или более аминокислотных остатков доменов константной области замещены или делетированы. Одним из примеров может служить введение аминокислотных замен в верхнюю область CH2 для получения варианта Fc с пониженным сродством к рецепторам Fc (Cole и др. // J. Immunol. 1997. Т.159. С.3613). Имеющие обычный опыт в данной области могут приготовить такие конструкции, применяя хорошо известные методы молекулярной биологии.
Использование Fcγ1 человека в качестве последовательности области Fc имеет несколько преимуществ. Например, если слитый белок, содержащий ингибитор ангиогенеза вместе с Fc, должен быть использован как биологически активное лекарство, домен Fcγ1 может придавать слитому белку активность в смысле эффекторных функций. Такие эффекторные функции включают такие типы биологической активности, как связывание комплемента, реализуемую через антитела клеточную цитотоксичность, транспорт в плаценте и увеличенное время полужизни в сыворотке. Домен Fc делает возможным также обнаружение методом твердофазного иммуноферментного анализа ELISA и очистку связыванием с белком A Staphylococcus aureus (Protein А). В некоторых применениях, однако, может оказаться желательным снять с области Fc некоторые эффекторные функции, такие, как связывание с рецепторами Fc или связывание комплемента.
В случае иммунофузинов - ингибиторов ангиогенеза одной из функций Fc иммуноглобулина в качестве партнера по связыванию является стимуляция должного сворачивания белка - ингибитора ангиогенеза для получения активного белка - ингибитора ангиогенеза и для придания активным компонентам растворимости, хотя бы во внеклеточной среде. Поскольку Fc как партнер по слиянию гидрофилен, иммунофузин с ингибитором ангиогенеза хорошо растворим, в отличие, например, от рекомбинантного эндостатина, продуцируемого в Е.coli (O’Reilly // Cell. 1997. Т.88. С.277). Во всех приведенных здесь примерах были получены высокие уровни продукции иммунофузинов. Иммунофузины - ингибиторы ангиогенеза выделялись (секретировались) в среду с концентрацией обычно в пределах от 30 до 100 мкг/мл и могли быть с успехом очищены до гомогенности с помощью хроматографии с белком А. Кроме того, иммунофузины - ингибиторы ангиогенеза можно было расщепить и дополнительно очистить с помощью стандартных методов очистки, включающих, например, очистку на гепарин-сефарозе, лизин-сефарозе или аффинную очистку.
В дополнение к высоким уровням экспрессии слитые белки в соответствии с настоящим изобретением обнаруживают также более длительное время полужизни в сыворотке, по-видимому, благодаря большему размеру их молекул. Например, ангиостатин человека, связанный с Fc человека, имеет время полужизни в сыворотке мыши 33 ч, а ангиостатин человека 4-6 ч (O’Reilly и др. // Nature Medicine. 1996. Т.2. С.689). Считается, что ангиостатин с молекулярным весом 40 кДа и эндостатин с молекулярным весом 20 кДа, благодаря их малому размеру, быстро выводятся из организма путем фильтрации через почки. Напротив, димерные формы Fc-ангиостатина и димерный Fc-эндостатин имеют молекулярный вес соответственно 145 и 100 кДа, поскольку имеются две области Fc иммуноглобулина, связанные либо с двумя молекулами ангиостатина, либо с двумя молекулами эндостатина. Такая бивалентная конструкция может проявлять более высокую активность в связывании с рецепторами ангиостатина или эндостатина. Если активность в подавлении ангиогенеза осуществляется через рецепторы, слитые белки, содержащие Fc, потенциально более эффективны в подавлении опухолей, чем моновалентный ангиостатин или моновалентный эндостатин сами по себе. Кроме того, если ангиостатин и/или эндостатин принадлежат к классу димерных белковых лигандов, физическое воздействие, оказываемое Fc на ангиостатин или эндостатин, будет превращать димеризацию во внутримолекулярный процесс, сдвигая таким образом равновесие в сторону образования димера и усиливая его связывание с рецептором. Для стабилизации димера путем образования ковалентных дисульфидных связей в мономер с помощью стандартной технологии рекомбинантной ДНК могут быть введены в подходящих местах остатки цистеина.
В том смысле, как он использован здесь, термин “мультивалентный” обозначает рекомбинантную молекулу, которая включает два или более биологически активных сегмента. Белковые фрагменты, образующие мультивалентную молекулу, могут быть соединены через полипептидный мостик, который соединяет составляющие части, не приводя к сдвигу рамки считывания, и позволяет каждой части функционировать независимо.
В том смысле, как он использован здесь, термин “бивалентный” обозначает рекомбинантную молекулу, содержащую в слитой конструкции по настоящему изобретению два целевых белка, например молекулу Fc-X, где Х выбран независимо из ангиостатина, эндостатина или их варианта. Поскольку имеются два участка X, соединенные с областью Fc иммуноглобулина (которая сама обычно представляет собой димер фрагментов тяжелой цепи, включающих по меньшей мере часть шарнирной области и домен СН3, а также возможно, но необязательно домен СН2), молекула бивалентна (см., например, Фиг.1А). Если слитая конструкция в соответствии с настоящим изобретением имеет форму Fc-X-X, полученная молекула с димером Fc тетравалентна. Два белка, образующих молекулу Fc-X-X, могут быть соединены через пептидный мостик. Бивалентная молекула может повышать кажущееся сродство связывания между молекулой и ее рецептором. Например, если один участок эндостатина в конструкции Fc-эндостатин может связываться с рецептором на клетке с определенной величиной сродства, второй участок эндостатина той же самой конструкции Fc-эндостатин может намного активнее связаться со вторым рецептором на той же клетке (кажущееся сродство). Это является следствием физической близости второго участка эндостатина к рецептору после того, как первый участок эндостатина уже связался. В случае связывания антитела с антигеном кажущееся сродство увеличивается приблизительно в 104 раз.
В том смысле, как они использованы здесь, термины “мультимер” и “мультимерный” относятся к стабильной ассоциации двух или более полипептидных цепей: либо ковалентным способом, например, путем ковалентного взаимодействия, например, образования дисульфидной связи, или нековалентным способом, например, с помощью гидрофобного взаимодействия. Имеется в виду, что термин “мультимер” включает как гомомультимеры, где полипептиды одинаковы, так и гетеромультимеры, где полипептиды различны.
В том смысле, как он использован здесь, термин “димерный” обозначает специфическую мультимерную молекулу, где две белковых полипептидных цепи стабильно ассоциированы с помощью ковалентных или нековалентных взаимодействий. Следует понимать, что сами область Fc иммуноглобулина и фрагмент Fc обычно представляют собой димер фрагментов тяжелой цепи, включающий по меньшей мере часть шарнирной области и домен СН3, и возможно, но необязательно, домен СН2. Известно, что многие белковые лиганды связываются со своими рецепторами в виде димеров. Если белковый лиганд Х естественным образом димеризуется, то участки Х в молекуле Fc-X будут димеризоваться гораздо сильнее, поскольку процесс димеризации зависит от концентрации. Физическая сближенность двух участков X, соединенных прикрепленной областью Fc иммуноглобулина, превращает димеризацию во внутримолекулярный процесс, сильно сдвигая равновесие в сторону образования димеров и повышая их связывание с рецепторами.
Понятно, что настоящее изобретение использует обычные методы рекомбинантной ДНК для получения содержащих Fc слитых белков, пригодных для практического применения изобретения. Слитые конструкции с Fc предпочтительно создаются на уровне ДНК, а полученные ДНК интегрируют в экспрессирующие векторы и экспрессируют для получения иммунофузинов. Следует понимать, что термин “вектор” в том смысле, как он использован здесь, обозначает любую нуклеиновую кислоту, содержащую нуклеотидную последовательность, способную быть внедренной в клетку-хозяина и рекомбинировать с геномом и интегрироваться в геном клетки-хозяина или автономно реплицироваться в виде эписомы. Такие векторы включают линейные нуклеиновые кислоты, плазмиды, фагемиды, космиды, РНК-векторы, вирусные векторы и т.п. Примеры вирусных векторов, не ограничивающие все возможности, включают ретровирус, аденовирус и аденоассоциированный вирус. Понятно, что в том смысле, как он здесь использован, термин “генетическая экспрессия” или “экспрессия” целевого белка обозначает транскрипцию последовательности ДНК, трансляцию мРНК-транскрипта и секрецию продукта - слитого белка, содержащего Fc.
Подходящим экспрессирующим вектором является pdCs (Lo и др. // Protein Engineering. 1988. Т.11. С.495, изложение включено сюда ссылкой), в котором для транскрипции гена Fc-X используются энхансер/промотор цитомегаловируса человека и сигнал полиаденилирования SV40. Используемая последовательность энхансера и промотора цитомегаловируса человека была получена из нуклеотидов от -601 до +7 последовательности, предложенной в работе Boshart и др. // Cell. 1985. Т.41. С.521, изложение которой включено сюда ссылкой. Вектор содержит также мутантный ген дигидрофолат-редуктазы в качестве селектирующего маркера (Simonsen и Levinson // Proc. Natl. Acad. Sci. USA. 1983. Т.80. С.2495, изложение включено сюда ссылкой).
Подходящая клетка-хозяин может быть трансформирована или трансфецирована последовательностью ДНК настоящего изобретения и использована для экспрессии и секреции целевого белка. Общепринятые предпочтительные клетки-хозяева для использования в настоящем изобретении включают бессмертные клетки гибридом, клетки миеломы NS/0, клетки 293, клетки яичника китайского хомячка, клетки HeLa и клетки COS.
Слитые белки в соответствии с настоящим изобретением предпочтительно создаются обычными методами рекомбинантной ДНК. Слитые белки предпочтительно продуцируются при экспрессиии в клетке-хозяине молекулы ДНК, кодирующей сигнальную последовательность, область иммуноглобулина Fc и целевой белок (называемый здесь также ингибитором ангиогенеза). Предпочтительные конструкции могут кодировать в направлении 5’→3’ сигнальную последовательность, область Fc иммуноглобулина и целевой белок. В качестве альтернативы конструкции могут кодировать в направлении 5’→3’ сигнальную последовательность, целевой белок и область Fc иммуноглобулина.
В том смысле, как он использован здесь, термин “сигнальная последовательность” означает пептидный сегмент, который обеспечивает секрецию белка иммунофузина - ингибитора ангиогенеза и затем отщепляемый после трансляции в клетке-хозяине. Сигнальная последовательность в соответствии с настоящим изобретением представляет собой полинуклеотид, который кодирует аминокислотную последовательность, инициирующую транспорт белка через мембрану эндоплазматического ретикулума. Сигнальные последовательности, которые пригодны в настоящем изобретении, включают сигнальные последовательности легкой цепи антител, например антител 14.18 (Gillies и др. // J. Immunil. Meth. 1989. Т.125. С.191-202), сигнальные последовательности тяжелой цепи антител, например сигнальные последовательности тяжелой цепи антител МОРС141 (Sakano и др. // Nature. 1980. Т.286. С.5774) и любые другие известные в данной области сигнальные последовательности (например, см. Watson // Nucleic Acids Research. 1984. Т.12. С.5145). Все цитированные работы включены сюда ссылкой.
Сигнальные последовательности хорошо исследованы в данной области. Известно, что они обычно содержат от 16 до 30 аминокислотных остатков и могут содержать большее или меньшее число аминокислотных остатков. Типичный сигнальный пептид состоит из трех участков: основного N-концевого участка, центрального гидрофобного участка и более полярного С-концевого участка. Центральный гидрофобный участок содержит от 4 до 12 гидрофобных остатков, которые закрепляют сигнальный пептид в липидном бислое мембраны при транспортировке синтезирующегося полипептида. После инициирования сигнальный пептид обычно расщепляется в просвете эндоплазматического ретикулума клеточными ферментами, известными как сигнальные пептидазы. Потенциальные сайты расщепления сигнального пептида, как правило, подчиняются “правилу (-3, -1)”. Это означает, что типичный сигнальный пептид содержит небольшие нейтральные аминокислотные остатки в положениях -1 и -3 и не содержит в этой области остатков пролина. Сигнальная пептидаза расщепляет такой сигнальный пептид между аминокислотами -1 и +1. Итак, сигнальная последовательность как часть последовательностей, кодируемых ДНК, может быть отщеплена в процессе секреции от амино-конца белка иммунофузина. Это приводит к секреции белка иммунофузина, состоящего из области Fc и целевого белка. Детальное обсуждение последовательностей сигнального пептида дано von Heijne // Nucleic Acids Res. 1986. Т.14. С.4683. Содержание этой работы включено сюда ссылкой.
Как очевидно специалистам в данной области, пригодность конкретной сигнальной последовательности для использования в настоящем изобретении может потребовать некоторой обычной экспериментальной проверки. Такая экспериментальная проверка включает определение способности сигнальной последовательности управлять секрецией иммунофузина и также определение оптимальной конфигурации кодирования - геномной ДНК или кДНК - той последовательности, которую можно использовать для достижения эффективной секреции иммунофузинов. Дополнительно к этому специалисты в данной области могут создать синтетический сигнальный пептид, следуя правилам, установленным von Heijne (см. ссылку выше), и проверить эффективность такой сигнальной последовательности в обычных экспериментах. Сигнальная последовательность может также иметь название “сигнальный пептид”, “лидирующая последовательность” или “лидирующий пептид”.
Соединение сигнальной последовательности и области Fc иммуноглобулина иногда здесь обозначается как секреционная кассета. Примерная секреционная кассета, пригодная в практике применения настоящего изобретения, представляет собой полинуклеотид, кодирующий в направлении 5’→3’ сигнальную последовательность гена легкой цепи иммуноглобулина и область Fcγ1 гена γ1 иммуноглобулина человека. Область Fcγ1 гена иммуноглобулина γ1 предпочтительно содержит по меньшей мере часть шарнирного домена и по меньшей мере часть домена СН3, или в качестве альтернативы - по меньшей мере части шарнирного домена, домена СН2 и домена СН3. ДНК, кодирующая секреционную кассету, может быть в конфигурации геномной ДНК или в конфигурации кДНК.
В другом осуществлении последовательность ДНК кодирует сайт протеолитического расщепления, помещенный между секреционной кассетой и белком - ингибитором ангиогенеза. Сайт расщепления, обеспечивающий протеолитическое расщепление кодируемого слитого белка, обеспечивает таким образом отделение домена Fc от белка - ингибитора ангиогенеза. Понятно, что термин “сайт протеолитического расщепления”, как он использован здесь, означает аминокислотные последовательности, которые преимущественно расщепляются протеолитическим ферментом или другими агентами протеолитического действия. Подходящие сайты протеолитического расщепления включают аминокислотные последовательности, распознаваемые протеолитическими ферментами, такими, как трипсин, плазмин или энтерокиназа К. Известны многие пары сайт расщепления/расщепляющий агент. Например, см. патент США №5726044, раскрытие которого включено сюда ссылкой. Если последовательность целевого белка представляет собой молекулу-предшественник ангиостатина, эндостатина или их активных вариантов, необходимый белковый продукт может быть получен расщеплением с помощью протеолитического фермента эндогенного действия, такого, как эластин или плазмин, или урокиназа.
Настоящее изобретение включает также слитые белки, содержащие различные комбинации рекомбинантных ангиостатина и эндостатина или их фрагментов, которые могут быть произведены в больших количествах. Несмотря на продемонстрированную эффективность в подавлении роста опухолей, механизм блокирования ангиогенеза ангиостатином и эндостатином не вполне ясен. Ангиостатин имеет несколько структур Крингла, а эндостатин имеет несколько структурных мотивов, каждый из которых может быть единственно ответственным за связывание или способствующим связыванию белков с клетками эндотелия и проявлению антиангиогенного эффекта. Поэтому это изобретение включает целевые белки, которые являются биологически активными фрагментами ангиостатина, такими, как 1-й домен Крингла, 2-й домен Крингла, 3-й домен Крингла и их комбинации, и эндостатина, которые физиологически действуют подобно существующим в природе ангиостатину и эндостатину полной длины.
Другое осуществление настоящего изобретения предусматривает бифункциональные гибридные конструкции ингибиторов ангиогенеза. В использованном здесь смысле бифункциональная гибридная молекула или конструкция означает белок, получаемый объединением двух белковых субъединиц, причем две субъединицы могут происходить из различных белков. Каждая белковая субъединица имеет свою независимую функцию, так что в гибридной молекуле функции двух субъединиц могут быть аддитивными или синергичными. Такие белки с гибридными функциями позволят исследовать синергический эффект ангиостатина и эндостатина на животных моделях. Предпочтительный бифункциональный гибрид может содержать по меньшей мере два различных ингибитора ангиогенеза, соединенных в тандем либо непосредственно, либо с помощью полипептидного мостика. Например, в предпочтительном осуществлении соответствующая целевому белку последовательность кодирует по меньшей мере часть ангиостатина, соединенную в одной рамке считывания с по меньшей мере частью эндостатина, и оба домена - ангиостатинный и эндостатинный - проявляют антиангиогенную активность или ингибирование ангиогенеза. Две субъединицы могут быть соединены полипептидным мостиком.
Следует понимать, что в использованном здесь смысле термин “полипептидный мостик” означает пептидную последовательность, которая может связывать вместе два белка или белок с областью Fc. Полипептидный мостик предпочтительно содержит большей частью такие аминокислоты, как глицин и/или серин. Предпочтительно полипептидный мостик содержит набор пептидов глицина и серина длиной приблизительно 10-15 звеньев. Например, см. патент США №5258698, раскрытие которого включено сюда ссылкой. Однако предполагается, что оптимальные длина мостика и аминокислотный состав могут быть определены обычными экспериментальными исследованиями.
Было обнаружено, что если различные части ангиостатина экспрессируются как слитые молекулы с Fc, достигаются высокие уровни экспрессии предположительно благодаря тому, что часть Fc действует как носитель, помогающий правильному сворачиванию полипептида на С-конце. Кроме того, область Fc может быть гликозилирована и при физиологических значениях рН иметь высокий заряд. Поэтому область Fc может способствовать растворению гидрофобных белков.
Настоящее изобретение также предусматривает способы продукции ангиостатина и эндостатина других, кроме человека, видов, в виде слитых белков с Fc. Такие слитые белки - ингибиторы ангиогенеза иных, кроме человека, видов полезны для доклинического изучения ингибиторов ангиогенеза, так как исследования активности и токсичности белкового лекарства перед испытаниями на людях должны быть проведены в системах с животными моделями. Белок человека может не иметь активности у мышей в модельных экспериментах, так как белок может стимулировать иммунный ответ и/или проявлять другую фармакокинетику, искажающую результаты испытаний. Поэтому эквивалентный белок мыши является наилучшим заменителем человеческого белка для модельных испытаний на мышах.
Для сравнения растворимых слитых белков Fс(hu)-ангиостатин(hu), Fc(hu)-эндостатин(hu), Fс(mu)-ангиостатин(mu), Fс(mu)-эндостатин(mu) (hu означает человеческий, a mu - мышиный) с нерастворимыми белками, продуцируемыми в системе экспрессии E.coli, была использована стандартная модель карциномы легких Льюиса у мышей (O’Reilly и др. // Cell. 1997. Т.88. С.277). Растворимые слитые белки с Fc были более активны в подавлении роста опухоли в модели легочной карциномы Льюиса, чем соответствующие белки, полученные в Е.соli. Кроме того, лабораторные мыши являются линейными и их опухоли индуцированнные, а не спонтанные. Поэтому эффективность в мышиной модели может не соответствовать возможной эффективности против опухолей человека. Доклинические исследования на собаках дают более точную информацию об эффективности действия этих ингибиторов ангиогенеза на спонтанные опухоли, так как у собак существует большое количество естественно возникающих, спонтанных опухолей. Способы получения мышиных (mu) слитых белков Fc(mu)-ангиостатин(mu) и Fс(mu)-эндостатин(mu) и собачьих (са) слитых белков Fc(ca)-ангиостатин(са) и Fс(са)-эндостатин(са) по настоящему изобретению будет содействовать доклиническому изучению ингибиторов ангиогенеза как в мышиных, так и в собачьих системах.
Настоящее изобретение предусматривает способы лечения болезненных состояний, опосредованных ангиогенезом, путем введения ДНК, РНК или белков по настоящему изобретению. Опосредованные ангиогенезом болезненные состояния включают, например, плотные опухоли, порожденные кровеносной системой опухоли, метастазы опухолей, доброкачественные опухоли, включая гемангиомы, акустические невромы, нейрофибромы, трахомы и пирогенные гранулемы; ревматоидный артрит; псориаз; заболевания глазных кровеносных сосудов (диабетическая патология сетчатки - ретинопатия, ретинопатия преждевременного созревания, перерождение роговицы с появлением пятен, отторжение пересаженной роговицы, связанная с новообразованием сосудов глаукома); возвратная замедленная фиброплазия, краснуха, синдром Ослера-Веббера; ангиогенез миокарда; неоваскуляризация бляшек; телангиэктазия; связанные с гемофилией ангиофиброма и грануляция ран; избыточная или ненормальная стимуляция клеток эндотелия, спайки кишечника, артериосклероз, склеродермальные и гипертрофические шрамы, т.е. келоиды.
Конструкции ДНК, раскрытые здесь, могут быть полезны для процедур генотерапии, в которых ген ангиостатина или эндостатина вводится в клетку одним из многих способов, например, в виде нативной ДНК, соединенной с промотором, или в виде ДНК в вирусном векторе. Попав в клетку, ген ангиостатина и/или эндостатина экспрессируется, и in vivo продуцируется белок, имеющий нормальную биологическую функцию. ДНК-конструкция по настоящему изобретению дает высокие уровни экспрессии слитого белка. Слитые белки по настоящему изобретению могут также быть полезными для лечения болезненных состояний, опосредованных ангиогенезом, и могут иметь более высокую клиническую эффективность, чем природные ингибиторы ангиогенеза и другие рекомбинантные ингибиторы ангиогенеза, так как иммунофузины - ингибиторы ангиогенеза по настоящему изобретению - имеют более продолжительное время полужизни в сыворотке, чем сами по себе другие рекомбинантные ингибиторы ангиогенеза или природные ингибиторы ангиогенеза. Бивалентные и димерные формы, предложенные в настоящем изобретении, должны иметь более высокое сродство связывания благодаря бивалентной и димерной структуре. Бифункциональные гибридные молекулы по настоящему изобретению могут иметь более высокую клиническую эффективность вследствие возможных синергических эффектов двух различных ингибиторов ангиогенеза, связанных сцепленной областью Fc или гибким полипептидным мостиком.
Составы по настоящему изобретению могут быть введены животному любым из подходящих способов: непосредственно (например, путем инъекции, имплантации или аппликацией на поверхность участка ткани) или системно (например, парентерально или орально). Если состав должен быть введен парентерально, например внутривенно, подкожно, офтальмологически, внутрибрюшинно, внутримышечно, в ротовую полость, ректально, вагинально, в глазную впадину, внутрицеребрально, внутричерепным способом, в спинной мозг, внутрь желудочка, внутрь оболочки, внутрь полости, внутрикапсульно, интраназально или аэрозольно, состав предпочтительно содержит часть суспензии (водной суспензии или суспензии в физиологически совместимой жидкости) или раствора. Таким образом, носитель или переносчик физиологически приемлем, так что при его добавлении в ходе введения необходимого состава пациенту он не влияет неблагоприятным образом на баланс электролитов или объемов у пациента. Поэтому жидкая среда для агента может представлять собой нормальный физиологический солевой раствор (например, водный раствор 9,85% NaCI, 0,15 М, рН 7-7,4).
Предпочтительные дозы иммунофузинов для введения находятся в интервале от 50 нг/м2 до 1 г/м2, более предпочтительно от 5 мкг/м2 до 200 мг/м2 и наиболее предпочтительно от 0,1 мг/м2 до 50 мг/м2. Предпочтительные дозы на одно введение нуклеиновых кислот, кодирующих иммунофузины, находятся в пределах от 1 мкг/м2 до 100 мг/м2, более предпочтительно от 20 мкг/м2 до 10 мг/м2 и наиболее предпочтительно от 400 мкг/м2 до 4 мг/м2. Подчеркивается, однако, что оптимальные пути введения и дозировки могут быть определены обычным экспериментальным исследованием, что соответствует уровню квалификации в данной области.
Далее изобретение иллюстрируется следующими не ограничивающими его примерами.
Пример 1. Экспрессия слитого белка Fс(hu)-эндостатин(hu).
Эндостатин человека был экспрессирован как слитый белок, состоящий из Fc человека и эндостатина человека (huFc-huEndo), в соответствии с указаниями Lo и др. // Protein Engineering. 1998. Т.11. С.495. Fc обозначает фрагмент Fc гамма-иммуноглобулина человека (последовательность ДНК указана далее как последовательность SEQ ID NO: 1; аминокислотная последовательность приведена далее как последовательность SEQ ID NO: 2). Для приспособления кДНК (комплементарной ДНК) эндостатина для экспрессии с получением слитого белка Fc-Endo использовали полимеразную цепную реакцию (ПЦР) (последовательность кДНК приведена как последовательность SEQ ID NO: 3; соответствующая ей аминокислотная последовательность раскрыта в последовательности SEQ ID NO: 4). Прямым праймером служил либо 5’-СС CCG GGT ААА САС AGC САС CGC GAC ТТС С (последовательность SEQ ID NO: 5; кодируемые аминокислоты раскрыты в последовательности SEQ ID NO: 6) или 5’-С AAG СТТ САС AGC САС CGC GAC ТТС С (последовательность SEQ ID NO: 7; кодируемые аминокислоты раскрыты в последовательности SEQ ID NO: 8), где за сайтом XmaI или сайтом HindIII следует последовательность, кодирующая N-конец эндостатина. Праймер с сайтом XmaI делает возможным лигирование (сшивку лигазой) кДНК эндостатина к сайту XmaI на конце последовательности, кодирующей домен СН3 области Fc иммуноглобулина G (IgG). Праймер с сайтом HindIII делает возможным лигирование кДНК эндостатина к сайту HindIII вектора pdCs-Fc(D4K), который содержит сайт распознавания энтерокиназой Asp4-Lys (La Vallie и др. // J. Biol. Chem. 1993. Т.268. С.23311-23317) в точке соединения слитого белка. Обратным праймером служил 5’-С СТС GAG СТА СТТ GGA GGC AGT CAT G (последовательность SEQ ID NO: 9), который был сконструирован так, чтобы ввести стоп-кодон трансляции (антикодон, СТА) сразу после С-конца эндостатина, а за этим следовал сайт XhoI. Продукты ПЦР были клонированы и секвенированы, и фрагмент XmaI-XhoI был лигирован с полученным вектором pdCs-Fc, расщепленным рестриктазами XmaI и XhoI. Подобным же образом фрагмент HindIII-XhoI, кодирующий эндостатин, был лигирован в соответственно расщепленный вектор pdCs-huFc(D4K). Стабильные клоны, экспрессирующие Fc-Endo или Fс(D4К)-эндостатин, были получены электропорацией клеток NS/0 с последующей селекцией в ростовой среде, содержащей 100 нМ метотрексат. Уровень экспрессии белка определяли методом ELISA для Fc человека (пример 3) и подтверждали электрофорезом в полиакриламидном геле с додецилсульфатом натрия (SDS), который показывал наличие белкового продукта с молекулярным весом около 52 кДа. Клоны с наилучшей продукцией были субклонированы методом исчерпывающего разбавления.
Пример 2. Культура клеток и трансфекция.
Для временной трансфекции плазмиду вводили в клетки 293 почки человека методом соосаждения плазмидной ДНК с фосфатом кальция (Sambrook и др. // Molecular Cloning - A Laboratory Manual, Cold Spring Harbor, NY) или методом липофекции с использованием LipofectAMINE Plus (Life Technologies, Gaithersburg, MD) в соответствии с указаниями поставщика.
Для получения стабильно трансфецированных клонов плазмидную ДНК вводили электропорацией в клетки NS/0 миеломы мыши. Клетки NS/0 выращивали в среде Игла в модификации Дальбеко с добавкой 10% сыворотки теленка. Приблизительно 5×106 клеток промывали один раз фосфатно-буферным солевым раствором (PBS) и ресуспендировали в 0,5 мл PBS. Затем 10 мкг линеаризованной плазмидной ДНК инкубировали с клетками в кювете прибора Gene Pulser (просвет между электродами 0,4 см, фирма BioRad, Hercules, CA, США) с охлаждением льдом в течение 10 мин. В установке Gene Pulser (BioRad, Hercules, CA) проводили электропорацию с параметрами 0,25 В и 500 мкФ. Клетки выдерживали 10 мин во льду, после чего их ресуспендировали в ростовой среде и высевали на планшеты с 96 ячейками. Селекцию стабильно трансфецированных клонов проводили по росту в присутствии 100 нМ метотрексата (МТК), который добавляли через 2 дня после трансфекции. Реагент добавляли еще 3 раза каждые 3 дня, устойчивые к МТК клоны появлялись через 2-3 недели. Для идентификации высокоактивных продуцентов надосадочные жидкости клонов анализировали методом ELISA для Fc. Высокопродуктивные клоны выделяли и поддерживали в ростовой среде, содержащей 100 нМ МТК.
Пример 3. Методические приемы выполнения анализа ELISA.
Для определения концентрации белковых продуктов в надосадочных жидкостях устойчивых к МТК клонов использовали 3 различные модификации метода ELISA. Для измерения количества белков, содержащих Fc человека (huFc), применяли ELISA для huFc. Для измерения количества белков, содержащих Fc мыши (muFc) и Fc собаки (caFc) использовали модификации метода ELISA с антителами соответственно к muFc и к caFc. Методика проведения ELISA для huFc подробно описана ниже.
А. Подготовка (покрытие) планшетов для проведении реакции.
На планшеты ELISA с 96 ячейками наносили козьи антитела аффинной степени чистоты к IgG человека - AffiniPure Goat anti-Human IgG (H+L) (Jackson ImmunoResearch Laboratories, West Grove, PA, USA) при концентрации 5 мкг/мл в PBS в объеме 100 мкл в каждую из 96 ячеек планшета (Nunc-Immuno plate MaxiSorp™, Nalge Nunc International, Rochester, NY, USA). Покрытые антителами планшеты промывали 4 раза 0,05% твином-20 в PBS и блокировали раствором 1% бычьего сывороточного альбумина (BSA) и 1% козьей сыворотки в PBS (200 мкл на ячейку). После инкубации с блокирующим буфером в течение 2 ч при 37°С планшеты промывали 4 раза 0,05% твином в PBS и осушали бумажными полотенцами.
В. Инкубация с испытуемыми пробами и вторичными антителами.
Исследуемые пробы разбавляли до нужной концентрации в тест-буфере, содержащем в PBS 1% BSA, 1% козью сыворотку и 0,05% твин. С химерными антителами известной концентрации (имеющими анти-huFc активность) получали стандартную калибровочную кривую. Для получения калибровочной кривой в интервале концентраций от 125 до 3,9 нг/мл готовили последовательные разбавления в тест-буфере. Разбавленные пробы и стандарты вносили в ячейки планшета, 100 мкл в каждую ячейку, и затем планшет инкубировали в течение 2 ч при 37°С. После окончания инкубации планшет промывали 8 раз 0,05% раствором твина в PBS. В каждую ячейку затем добавляли 100 мкл вторичных антител - конъюгированных с пероксидазой хрена (ПХ) козьих антител к IgG человека (horse radish peroxidase (HPR)-conjugated goat anti-human IgG, Jackson ImmunoResearch Laboratories, West Grove, PA, USA), разбавленных приблизительно 1:120000 в тест-буфере. Для каждой партии конъюгированных с ПХ антител к IgG человека необходимо было определять точное разбавление вторичных антител. После инкубации в течение 2 ч при 37°С планшет промывали 8 раз 0,05% раствором твина в PBS.
С. Проявление.
Раствор субстрата готовили растворением 30 мг (1 таблетка) о-фенилендиамина дигидрохлорида (ОФД) в 15 мл раствора: 0,025 М лимонная кислота/0,05 М Na2HPO4 буфер, рН 5, содержащем 0,03% непосредственно перед употреблением добавленной Н2О2. Раствор субстрата добавляли в планшет в объеме 100 мкл на ячейку. Для проявления окраски выдерживали 30 мин в темноте при комнатной температуре. Время проявления можно менять в зависимости от различий в партиях красителя, вторичных антител и т.д. Реакцию останавливали добавлением 4 н. H2SO4, 100 мкл на ячейку. Окраску в ячейках регистрировали автоматическим считывающим устройством (“plate reader”) с установкой длин волн как 490, так и 650 нм, запрограммированным на вычитание фоновой оптической плотности при 650 нм из измеренной оптической плотности при 490 нм.
Процедура при ELISA для muFc была такой же, за исключением того, что планшет ELISA покрывали козьими антителами к мышиному иммуноглобулину G (AffiniPure Goat anti-murine IgG (H+L), Jackson ImmunoResearch, West Grove, PA, USA) при концентрации 5 мкг/мл в PBS, 100 мкл на ячейку, а вторичными антителами были конъюгированные с пероксидазой хрена козьи антитела к фрагменту Fcγ мышиного IgG (horse radish peroxidase-conjugated goat anti-muIgG, Fсγ, Jackson ImmunoResearch, West Grove, PA, USA), использованные в разведении 1:5000. Подобным же образом в ELISA для caFc планшеты ELISA покрывали кроличьими антителами, специфичными к фрагменту Fc IgG собаки (AffiniPure Rabbit anti-dot IgG, Fc Fragment specific, Jackson ImmunoResearch, West Grove, PA, USA), при концентрации 5 мкг/мл в PBS, 100 мкл на ячейку, а вторичными антителами были конъюгированные с пероксидазой хрена кроличьи антитела к фрагменту Fc собачьего IgG (horse radish peroxidase-conjugated AffiniPure rabbit anti-dog IgG, Fc fragment specific, Jackson ImmunoResearch, West Grove, PA, USA), использованные в разведении 1:5000.
Пример 4. Экспрессия слитого белка Fc человека - ангиостатин человека.
Ангиостатин человека (последовательность ДНК приведена ниже как последовательность SEQ ID NO: 10; аминокислотная последовательность приведена далее как последовательность SEQ ID NO: 11) экспрессировали как слитый белок Fc человека - ангиостатин человека (huFc-huAngio), no существу как описано в примере 1. Чтобы приспособить кДНК ангиостатина (последовательность SEQ ID NO: 3) для экспрессии в векторах pdCs-huFc или pdCs-huFc(D4K), применяли ПЦР. Соответствующими прямыми праймерами были 5’-СС CCG GGT AAG AAA GTG TAT CTC TCA GAG (последовательность SEQ ID NO: 12; кодируемые аминокислоты приведены в последовательности SEQ ID NO: 13) и 5’-С ССС AAG CTT AAA GTG TAT CTC TCA GAG (последовательность SEQ ID NO: 14; кодируемые аминокислоты раскрыты в последовательности SEQ ID NO: 15), где за сайтом Xmal или сайтом HindIII следовала последовательность, кодирующая N-конец ангиостатина. Обратным праймером был 5’-ССС CTC GAG СТА CGC TTC TGT ТСС TGA GCA (последовательность SEQ ID NO: 16), который был сконструирован таким образом, чтобы поместить стоп-кодон трансляции (антикодон, СТА) немедленно после С-конца ангиостатина, а затем следовал сайт Xhol. Продукты ПЦР клонировали и секвенировали, и полученные фрагмент XmaI-XhoI и фрагмент HindIII-XhoI, кодирующие ангиостатин, были лигированы с векторами соответственно pdCs-huFc и pdCs-huFc(D4K). Стабильные клоны NS/0, экспрессирующие huFc-huAngio и huFc(D4K)-huAngio, селектировали и анализировали, как описано в примерах 2 и 3.
Пример 5. Экспрессия слитого белка Fc мыши - эндостатин мыши.
Эндостатин мыши (последовательность ДНК представлена ниже как последовательность SEQ ID NO: 17; аминокислотная последовательность SEQ ID NO: 18) и Fc мыши (последовательность ДНК представлена ниже как последовательность SEQ ID NO: 19, кодируемые аминокислоты представлены в последовательности SEQ ID NO: 20) были экспрессированы как слитый белок Fc мыши - эндостатин мыши (muFc-muEndo), как в существенных деталях описано в примере 1. Чтобы приспособить кДНК эндостатина (последовательность SEQ ID NO: 4) для экспрессии в векторе pdCs-muFc(D4K), использовали ПЦР. Прямым праймером был 5’-С ССС AAG CTT CAT ACT CAT CAG GAC TTT С (последовательность SEQ ID NO: 21; кодируемые аминокислоты раскрыты в последовательности SEQ ID NO: 22), где после сайта HindIII расположена последовательность, кодирующая N-конец эндостатина. Обратным праймером был 5’-ССС СТС GAG СТА ТТТ GGA GAA AGA GGT С (последовательность SEQ ID NO: 23), который был сконструирован таким образом, чтобы поместить стоп-кодон трансляции (антикодон, СТА) немедленно после С-конца эндостатина, после чего следовал сайт XhoI. Продукт ПЦР клонировали и секвенировали, и полученный фрагмент HindIII-XhoI, кодирующий эндостатин, лигировали в вектор pdCs-muFc(D4K). Стабильные клоны NS/0, экспрессирующие muFc(D4K)-muEndo, селектировали и анализировали методом ELISA для muFc, как описано в примерах 2 и 3.
Пример 6. Экспрессия слитого белка Fc мыши - ангиостатин мыши.
Ангиостатин мыши (последовательность ДНК представлена ниже как последовательность SEQ ID NO: 24; аминокислотная последовательность SEQ ID NO: 25) был экспрессирован как слитый белок Fc мыши - ангиостатин мыши (muFc-muAngio), как в существенных деталях описано в примере 1. Чтобы приспособить кДНК ангиостатина (последовательность SEQ ID NO: 6) для экспрессии в векторе pdCs-Fc(D4K), использовали ПЦР. Прямым праймером был 5’-С ССС AAG СТТ GTG TAT CTG ТСА GAA TGT AAG ССС ТСС TGT СТС TGA GCA (последовательность SEQ ID NO: 26; кодируемые аминокислоты раскрыты в последовательности SEQ ID NO: 27), где после сайта HindIII расположена последовательность, кодирующая N-конец ангиостатина. Обратным праймером был 5’-ССС СТС GAG СТА ССС ТСС TGT СТС TGA GCA (последовательность SEQ ID NO: 28), который был сконструирован таким образом, чтобы поместить стоп-кодон трансляции (антикодон, СТА) немедленно после С-конца ангиостатина, после чего следовал сайт XhoI (CTCGAG). Продукт ПЦР клонировали и секвенировали, и полученный фрагмент HindIII-XhoI, кодирующий ангиостатин, лигировали в вектор pdCs-muFc(D4K). Стабильные клоны NS/0, экспрессирующие muFc(D4K)-muAngio, селектировали и анализировали методом ELISA для muFc, как описано в примерах 2 и 3.
Пример 7. Экспрессия Fc собаки (caFc).
Для приготовления мРНК использовали моноцитные клетки периферической крови собак (РВМС), выделенные из крови собак. После синтеза первой цепи кДНК с обратной транскриптазой и олиго(дТ) проводили ПЦР для амплификации Fc собаки (Kazuhiko и др. заявка Японии №1992040894-А1) с использованием прямого праймера 5’-СС ТТА AGC GAA ААТ GGA AGA GTT CCT CGC (последовательность SEQ ID NO: 29; кодируемые аминокислоты раскрыты в последовательности SEQ ID NO: 30), в котором сразу вверх от последовательности, кодирующей шарнирную область Fc собаки, введен сайт AflII, а обратный праймер -5’-С СТС GAG ТСА ТТТ АСС CGG GGA ATG GGA GAG GGA TTT CTG (последовательность SEQ ID NO: 31), в котором после стоп-кодона трансляции Fc собаки (антикодон, ТСА) введен сайт XhoI. Обратный праймер вводит также молчащую мутацию для создания сайта рестрикции XmaI, облегчающего конструирование вектора pdCs-caFc(D4K) с помощью линкера-адаптора и пришивки к ДНК-конструкциям, кодирующим эндостатин или ангиостатин собаки. Подобно конструированию pdCs-huFc, которое было подробно описано в работе Lo и сотрудников (Lo и др. // Protein Engineering. 1998. Т.11. С.495), экспрессирующий вектор pdCs-caFc был сконструирован следующим образом. Фрагмент AflII-XhoI, кодирующий Fc собаки, был сшит лигазой с фрагментом XbaI-AflII, кодирующим сигнальный пептид легкой цепи, и с вектором pdCs, расщепленным XbaI-XhoI. Полученный экспрессирующий вектор pdCs-caFc использовали затем для трансфекции клеток 293. Примерно через 3 дня после трансфекции культуральную жидкость очищали хроматографией с белком А. Экспрессия Fc собаки (последовательность ДНК представлена дальше как последовательность SEQ ID NO: 32; аминокислотная последовательность приведена как последовательность SEQ ID NO: 33) была подтверждена электрофорезом в полиакриламидном геле с SDS и последующим анализом вестерн-блотингом с использованием конъюгированных с пероксидазой кроличьих антител к IgG собаки, специфичных к фрагменту Fc (peroixidase-cohjugated rabbit anti-dog IgG, Fc fragment specific, Jackson ImmunoResearch, West Grove, PA, USA).
Пример 8. Экспрессия слитого белка Fc собаки - эндостатин собаки.
Кодирующую последовательность для эндостатина собаки (последовательность ДНК представлена далее как последовательность SEQ ID NO: 34; аминокислотная последовательность представлена далее как последовательность SEQ ID NO: 35) превращали во фрагмент HindIII-XhoI для экспрессии в виде слитого белка с Fc, как в существенных чертах описано в примере 5. На 3’-конце, сразу после кодона, кодирующего С-концевой остаток лизина, был введен стоп-кодон, например, с помощью ПЦР, а за ним следовал сайт рестрикции NotI. На 5’-конце, однако, имелся сайт рестрикции DraIII, удобный для реконструкции. Был химически синтезирован олигонуклеотидный дуплекс, содержащий липкие концы HindIII и DraIII; он был использован для пришивки к рестрикционному фрагменту DraIII-XhoI, который кодировал остальную часть кДНК эндостатина собаки. Использованный дуплекс показан ниже:
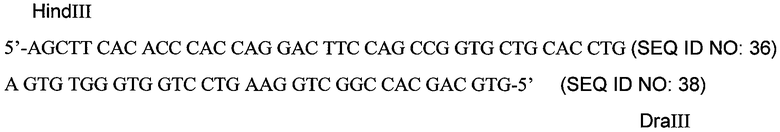
Первый триплет САС в дуплексе кодирует N-концевой остаток гистидина эндостатина собаки. Фрагмент HindIII-XhoI, кодирующий эндостатин собаки полной длины, был поэтому для экспрессии пришит лигазой к вектору pdCs-caFc, расщепленному HindIII-XhoI (см. пример 7). Стабильные клоны NS/0, экспрессирующие caFc - caEndo, селектировали и анализировали методом ELISA для caFc, как описано в примерах 2 и 3. Наличие белкового продукта определяли электрофорезом в полиакриламидном геле с SDS и подтверждали анализом с использованием вестерн-блотинга.
Пример 9. Экспрессия слитого белка Fc собаки - ангиостатин собаки.
Кодирующую ангиостатин собаки полной длины кДНК (последовательность ДНК представлена далее как последовательность SEQ ID NO: 39; аминокислотная последовательность представлена далее как последовательность SEQ ID NO: 40) приспосабливали для экспрессии в виде слитого белка с caFc, как в существенных чертах описано в приведенных выше примерах. Вкратце, на 3’-конце, непосредственно за кодоном, кодирующим С-концевой остаток лизина, вводили стоп-кодон (например, с помощью ПЦР), за которым вместо сайта XhoI следовал сайт рестрикции NotI, потому что в кДНК ангиостатина собаки имелся внутренний сайт рестрикции XhoI. На 5’-конце непосредственно вверх от N-конца ангиостатина был введен сайт HindIII с сохранением рамки считывания. Затем фрагмент HindIII-NotI, кодирующий ангиостатин собаки полной длины, для экспрессиии вшивали в вектор pdCs-caFc, расщепленный HindIII-NotI (сайт NotI был введен в него возле сайта Xhol путем лигирования через мостик). Стабильные клоны NS/0, экспрессирующие caFc-caAngio, селектировали и анализировали методом ELISA для caFc, как описано в примерах 2 и 3. Наличие белкового продукта определяли электрофорезом в полиакриламидном геле с SDS и подтверждали анализом с использованием вестерн-блотинга.
Пример 10. Экспрессия слитого белка muFc-домен К1 muAngio.
Ангиостатин содержит первые четыре из пяти доменов Крингла плазминогена. Чтобы определить, являются ли какой-либо один или несколько доменов Крингла ответственными за наблюдаемую антиангиогенную активность ангиостатина, можно для тестирования продуцировать либо единичные домены Крингла, либо их комбинации. Для демонстрации пригодности Fc как партнера в слитом белке экспрессию первого домена Крингла ангиостатина мыши (К1) осуществляли следующим образом. Первый домен Крингла в мышином ангиостатине оканчивается в 87-м положении остатком глутаминовой кислоты Glu-87 (последовательность SEQ ID NO: 25). В кДНК у этого положения имеется удобный сайт рестрикции NsiI, так что после расщепления рестриктазой Nsil 3’-концевую избыточность одной нити длиной в 4 нуклеотида удаляли Т4 полимеразой, чтобы создать конец с одинаково обрезанными нитями ДНК. Непосредственно вслед за триплетом GAA, кодирующим Glu-87, был введен в направлении вниз стоп-кодон трансляции пришивкой лигазой к палиндромному мостику TGA CTC GAG ТСА (последовательность SEQ ID NO: 41), где за стоп-кодоном TGA следует сайт XhoI. Фрагмент HindIII-XhoI, кодирующий этот усеченный ангиостатин, т.е. только первый домен Крингла, затем был вшит для экспрессии в вектор pdCs-muFc(D4K). Анализ методом ELISA для muFc и электрофорезом в полиакриламидном геле с SDS показал, что были получены высокие уровни экспрессии как при временной, так и при стабильной экспрессии.
Пример 11. Экспрессия слитого белка muFc -внутренняя часть К1 muAngio.
Домен Крингла состоит из множественных петель, включая внешнюю петлю и внутреннюю петлю. В первом домене Крингла ангиостатина мыши внутренняя петля ограничивается остатками цистеина Cys-55 и Cys-79, которые образуют друг с другом дисульфидную связь в основании петли. Cys-67 внутренней петли образует другую дисульфидную связь с остатком Cys внешней петли, формируя структуру Крингла. Чтобы проверить, имеет ли внутренняя петля какую-либо антиангиогенную активность, она была экспрессирована в виде слитого белка, содержащего muFc и внутреннюю петлю первого домена Крингла (К1), как описано ниже. Для мутирования (мутационного превращения) TGC (Cys-67) в AGC (Ser) с помощью ПЦР был использован в качестве матрицы фрагмент ДНК, кодирующий первый домен Крингла, вместе с мутагенным праймером, имеющим последовательность 5’ GGG CCT TGG AGC TAC ACT АСА (последовательность SEQ ID NO: 42; кодируемые аминокислоты раскрыты в последовательности SEQ ID NO: 43). Это давало гарантию, что Cys-67 не образует дисульфидную связь, если внутренняя петля первого домена Крингла экспрессирована без внешней петли. Для введения сайта HindIII в рамку считывания непосредственно с 5’-стороны кодона для Ser-43 (AGT) был использован восходящий праймер, имеющий последовательность 5’GCGGATCCAAGCTT AGT АСА CAT ССС ААТ GAG GG (последовательность SEQ ID NO: 44; кодируемые аминокислоты раскрыты в последовательности SEQ ID NO: 45). Сразу за сайтом HindIII в направлении вверх был также введен сайт BamHI. Сайт BamHI полезен для пришивки лигазой к сайту BamHI на конце последовательности для гибкого мостика Gly-Ser, показанного ниже в примере 12. Таким путем фрагмент ДНК HindIII-XhoI, кодирующий участок ангиостатина мыши от Ser-43 до Glu-87, был вшит для экспрессии в вектор pdCs-muFc(D4K). Анализ методом ELISA для muFc и электрофорезом в полиакриламидном геле с SDS показал, что были получены высокие уровни экспрессии слитого белка muFc с внутренней петлей К1 как при временной, так и при стабильной экспрессии.
Пример 12. Экспрессия слитого белка muFc-muEndo-мостик GlySer-внутренняя петля домена К1 muAnqio.
Гибридная молекула muFc-muEndo-внутренняя петля домена К1 состоит из слитого белка muFc-muEndo, присоединенного полипептидным мостиком, содержащим остатки глицина и серина, к внутренней петле первого домена Крингла мышиного эндостатина. Конструкцию ДНК составляли следующим образом.
На 3’-конце кДНК эндостатина мыши имеется сайт BspHI. Чтобы ввести на С-конце мышиного эндостатина гибкий мостик из остатков глицина и серина, фрагмент HindIII-BspHI длиной 540 пар оснований, кодирующий эндостатин, пришивали лигазой к олигонуклеотидному дуплексу с перекрыванием, образованному олигонуклеотидами, указанными в последовательностях SEQ ID NO: 46 и SEQ ID NO: 48. Аминокислотный мостик, кодируемый последовательностью SEQ ID NO: 46, показан в последовательности SEQ ID NO: 47.
Фрагмент HindIII-BamHI, кодирующий мышиный эндостатин и мостик Gly-Ser, субклонировали в стандартный клонирующий вектор. Затем для введения фрагмента BamHI-XhoI, кодирующего внутреннюю петлю домена К1 в примере 11, использовали сайт BamHI. Полученный фрагмент HindIII-XhoI, кодирующий слитый белок muEndo-мостик Gly-Ser-внутренняя петля К1, был вшит для экспрессии в вектор pdCs-muFc(D4K). Анализ методом ELISA для muFc и электрофорезом в полиакриламидном геле с SDS показал, что были получены высокие уровни экспрессии слитого белка muFc-muEndo-мостик Gly-Ser-внутренняя петля К1 как при временной, так и при стабильной экспрессии.
Пример 13. Экспрессия слитого белка muFc-muEndo-мостик GlvSer-домен К1 muAngio.
Гибридная молекула muFc-muEndo-KI состоит из слитого белка rnuFc-muEndo, присоединенного полипептидным мостиком, содержащим остатки глицина и серина, к первому домену Крингла мышиного эндостатина. Конструкцию ДНК составляли следующим образом.
Конец BamHI фрагмента HindIII-BamHI, кодирующего muEndo-мостик Gly-Ser (пример 12), был пришит к фрагменту HindIII-XhoI, кодирующему первый домен Крингла мышиного ангиостатина (пример 10), через следующий адаптор:

Адаптор имеет липкий конец HindIII’, который при лигировании не формирует сайт HindIII. Поэтому полученный фрагмент HindIII-XhoI, кодирующий muEndo-мостик GlySer-K1, был вшит для экспрессии в вектор pdCs-muFc(D4K). Анализ методом ELISA для muFc и электрофорезом в полиакриламидном геле с SDS показал, что были получены высокие уровни экспрессии слитого белка muFc-muEndo-мостик Gly-Ser-K1 как при временной, так и при стабильной экспрессии.
Пример 14. Экспрессия слитого белка muFc-muEndo-мостик GIySer-muAnqio.
Гибридная молекула muFc-muEndo-мостик GIySer-muAngio представляет собой muFc-muEndo, присоединенный содержащим остатки глицина и серина полипептидным мостиком к мышиному ангиостатину. Конструкция ДНК была в существенных чертах собрана следующим образом. Конец BamHI фрагмента HindIII-BamHI, кодирующего muEndo-мостик Gly-Ser (пример 12) был пришит к фрагменту HindIII-XhoI, кодирующему мышиный ангиостатин, через адаптор, описанный в примере 13. Полученный фрагмент HindIII-XhoI, кодирующий muEndo-мостик GlySer-muAngio, был вшит для экспрессии в вектор pdCs-muFc(D4K). Анализ методом ELISA для muFc и электрофорезом в полиакриламидном геле с SDS показал, что были получены высокие уровни экспрессии слитого белка muFc-muEndo-мостик Gly-Ser-muAngio как при временной, так и при стабильной экспрессии.
Пример 15. Экспрессия слитого белка huAngio-huFc-huEndo.
Гибридная молекула huAngio-huFc-huEndo представляет собой ангиостатин человека, присоединенный пептидной связью к huFc-huEndo. Конструкция ДНК была собрана следующим образом. Фрагмент HindIII-XhoI, кодирующий ангиостатин человека без стоп-кодона, был сначала получен с помощью ПЦР, так что за кодоном для первого аминокислотного остатка ангиостатина непосредственно следовала последовательность CTCGAG сайта Xhol. HindIII на 5’-конце пришивали к фрагменту XbaI-AflII, кодирующему сигнальный пептид легкой цепи (Lo и др. // Protein Engineering. 1998. Т. 11. С. 495), через адаптор AflII-HindIII’:
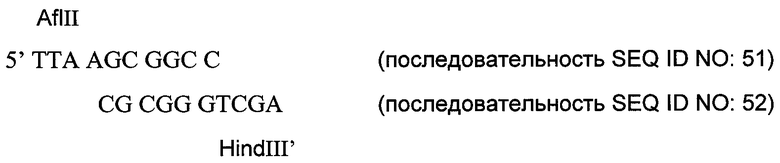
Липкий конец HindIII’ адаптора при лигировании не воссоздает сайт HindIII. На 3’-конце сайт XhoI пришивали к сайту AflII фрагмента AflII-XhoI, кодирующего huFc-huEndo, через адаптор XhoI’-AflII:
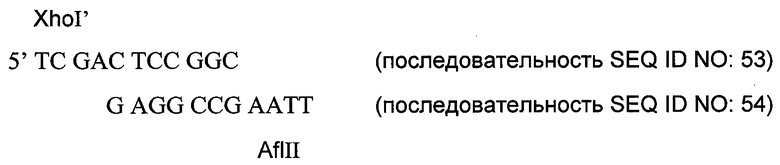
Липкий конец XhoI’ адаптора не воссоздает при лигировании сайт XhoI. Полученный фрагмент XbaI-XhoI, кодирующий слитый белок: сигнальный пептид-ангиостатин человека-huFс-эндостатин человека, клонировали для экспрессирования в вектор pdCs. Анализ с помощью ELISA для huFc и электрофореза в полиакриламидном геле с SDS показал, что были получены высокие уровни экспрессии слитого белка huAngio-huFc-huEndo как при временной, так и при стабильной экспрессии.
Пример 16. Фармакокинетика.
В одном из циклов фармакокинетического изучения линейным мышам с имплантированными опухолями легкого Льюиса размером 100-200 мм3 вводили инъекцией в хвостовую вену слитый белок huFc-huAngio в дозе 720 мкг на мышь. Размер опухолей и дозировку использованного в этих исследованиях huFc-huAngio выбирали для моделирования реальной методики лечения, описанной O’Reilly (O’Reilly // Nature Mediicine. 1996. Т.2. С.689). Кровь собирали кровопусканием из обратной петли через 1/2, 1, 2, 4, 8, 24 и 48 ч после инъекции. Образцы крови анализировали методом ELISA для huFc с последующим анализом вестерн-блоттингом. Было установлено, что время полужизни huFc-huAngio в кровеносном русле мышей составляет около 32 ч, а вестерн-анализ показал, что свыше 90% молекул huFc-huAngio остаются при циркуляции в кровяном русле интактными.
Фармакокинетические исследования были повторены также на швейцарских (Swiss) мышах без опухолей при дозировке 200 мкг на мышь. В этом случае было установлено, что huFc-huAngio имеет время полужизни в кровяном русле около 33ч.
Эквиваленты
Изобретение может быть осуществлено в других специфических формах без отхода от его идеи или существенных характеристик. Поэтому приведенные выше примеры во всех аспектах рассматриваются как иллюстративные, но не ограничительные для описанного здесь изобретения. Круг охвата изобретения поэтому определяется приложенной формулой, а не предшествующим описанием, и имеется в виду, таким образом, что сюда включены все могущие появиться изменения, которые находятся в рамках смысла и области эквивалентности формулы.

































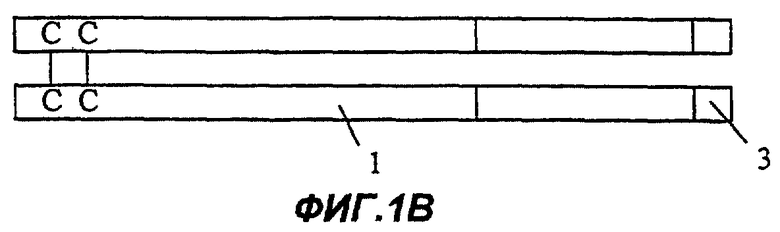
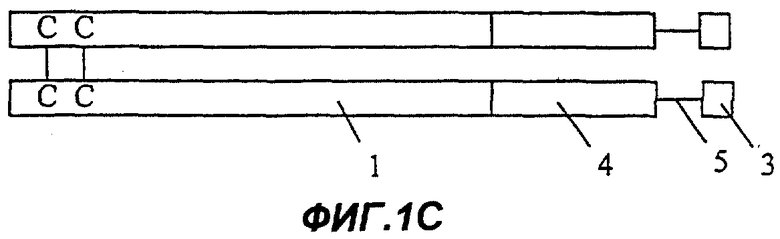
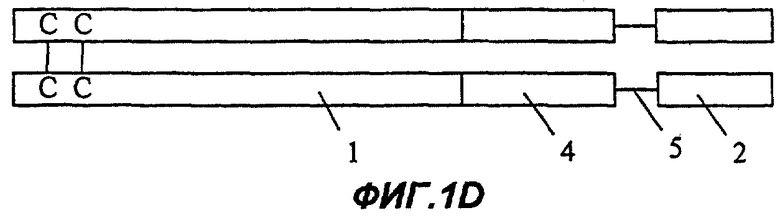
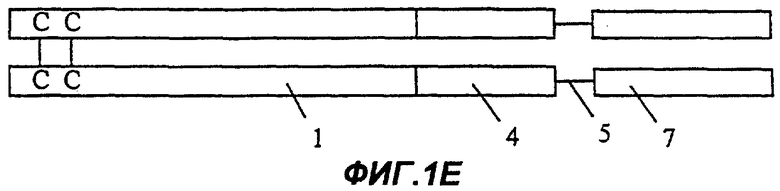
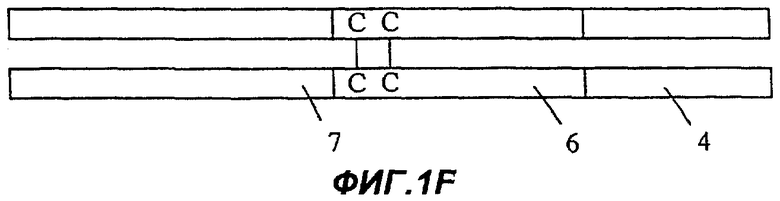
Изобретение относится к области биотехнологии и может быть использовано для получения слитых белков - ингибиторов ангиогенеза. Слитый белок представляет собой гомодимер, в котором область Fc иммуноглобулина гамма слита с целевым белком, включающим одну или более молекул с активностью ингибитора ангиогенеза, которая происходит от ангиостатина или эндостатина. Вариант гомодимерного слитого белка включает также второй целевой белок, содержащий одну или более молекул, имеющих активность ингибитора ангиогенеза, выбранных из ангиостатина, эндостатина, фрагмента плазминогена, обладающего активностью ангиостатина, и фрагмента коллагена XVIII, обладающего активностью эндостатина. Молекулы в составе гомодимерного слитого белка связаны с областью Fc иммуноглобулина гамма и между собой непосредственно или с помощью полипептидного мостика. Гомодимерный слитый белок получают путем трансфекции клетки млекопитающего вектором, содержащим молекулу ДНК, кодирующую целевой белок, культивирования клетки млекопитающего для продуцирования слитого белка и выделения слитого белка. Изобретение обеспечивает эффективную продукцию и секрецию ингибиторов ангиогенеза в разнообразных хозяевах – клетках млекопитающих, 6 н. и 7 з.п. ф-лы, 6 ил.


