Область техники, к которой относится изобретение
Настоящее изобретение касается геномной ДНК.
Предпосылки для изобретения
Современные подходы к лечению заболеваний, основанные на применении белков, обладающих лечебными свойствами, включают как прямое введение таких белков, выработанных in vitro, так и генотерапевтические методы. В принципе, выработка белка in vitro включает внесение экзогенной ДНК, кодирующей представляющий интерес белок, в подходящие культивируемые клетки-хозяева. С другой стороны, методы генотерапии включают введение пациенту генетически модифицированных клеток, плазмид или вирусов, которые включают последовательность, кодирующую конкретный интересующий терапевтический белок.
Некоторые терапевтические белки также могут быть выработаны в результате изменения экспрессии кодирующих их эндогенных генов желательным образом с помощью методов “направления генов”: см., например, патенты США №№5641670, 5733761 и 5272071 и международные патентные заявки WO 91/06666, WO 91/06667 и WO 90/11354; их полное содержание включено здесь для сведения в виде библиографических ссылок.
Резюме изобретения
Настоящее изобретение основывается на идентификации и секвенировании участка геномной ДНК, расположенного с 5’-стороны от кодирующей последовательности гена фолликулостимулирующего гормона-β (“FSHβ”) человека. Эта ДНК может быть использована, например, в составе ДНК-конструкции, которая изменяет (например, интенсифицирует) экспрессию эндогенного гена FSHβ в клетке млекопитающего за счет встраивания в состав генома этой клетки по механизму гомологичной рекомбинации. Термин “эндогенный ген FSHβ” обозначает геномную (т.е. хромосомную) копию гена, который кодирует белок FSHβ. Такая конструкция включает “направляющую” (“метящую”) последовательность, составленную заявляемой 5’-некодирующей последовательностью или производной от нее, а также последовательность, регулирующую транскрипцию. Последовательность, регулирующая транскрипцию, предпочтительно отличается по своей нуклеотидной последовательности от транскрипционной регуляторной последовательности эндогенного гена FSHβ. Направляющая последовательность обеспечивает встраивание регуляторной последовательности в участок в пределах кодирующей последовательности гена-мишени FSHβ или выше ее таким образом, что эта регуляторная последовательность оказывается функциональной присоединенной к эндогенной кодирующей последовательности. Термин “функционально присоединенный” обозначает то, что регуляторная последовательность способна обеспечивать экспрессию кодирующей последовательности эндогенного гена, кодирующего FSHβ. Дополнительно данная конструкция может включать ген селективного маркера, что облегчает отбор клеток, которые несут стабильно встроенную конструкцию, и (или) иную кодирующую последовательность, функциональным образом соединенную с промотором.
В одном варианте данная ДНК-конструкция включает: (а) направляющую последовательность; (b) регуляторную последовательность; (с) экзон; (d) донорный сайт сплайсинга. Направляющая последовательность обеспечивает встраивание ее самой и элементов (b)-(d) в участок, находящийся в пределах кодирующей последовательности гена-мишени FSHβ или выше ее. После такого встраивания элемент (b) может обеспечивать транскрипцию элементов (с) и (d) и всех остальных нижерасположенных кодирующих последовательностей эндогенного гена. В данной конструкции экзон обычно находится с 3’-стороны от регуляторной последовательности, а донорный сайт сплайсинга находится с 3’-стороны данного экзона.
В другом варианте ДНК-конструкция включает: (а) направляющую последовательность; (b) регуляторную последовательность; (с) экзон; (d) донорный сайт сплайсинга; (е) интрон; (f) акцепторный сайт сплайсинга, при том, что направляющая последовательность обеспечивает встраивание ее самой и элементов (b)-(f) таким образом, чтобы элементы (b)-(f) располагались в пределах данного эндогенного гена или выше его. После этого регуляторная последовательность обеспечивает образование транскрипта, который включает не только элементы (c)-(f), но также и кодирующую последовательность эндогенного гена FSHβ. Предпочтительно интрон и акцепторный сайт сплайсинга в составе данной конструкции расположены ниже донорного сайта сплайсинга.
Направляющая последовательность гомологична заранее выбранному сайту-мишени в геноме, по которому происходит гомологичная рекомбинация. Она включает по крайней мере 20 (например, по крайней мере 30, 50, 100 или 1000) подряд расположенных нуклеотидов из состава SEQ ID NO 4, что соответствует нуклеотидам [-7454]-[-1417] геномной последовательности FSHβ человека (нумерация по отношению к старт-кодону), или из состава SEQ ID NO 5, что соответствует нуклеотидам [-696]-[-155] геномной последовательности FSHβ человека. Термин “гомологичная” указывает на то, что направляющая последовательность идентична или в существенной степени сходна со своим геномным сайтом-мишенью на таком уровне, что направляющая последовательность и сайт-мишень могут претерпевать гомологичную рекомбинацию в клетке человека. Допустим небольшой процент несоответствий нуклеотидов, лишь бы гомологичная рекомбинация протекала с необходимой частотой. Для облегчения гомологичной рекомбинации длина направляющей последовательности предпочтительно составляет примерно по крайней мере 20 (например, по крайней мере 50, 100, 250, 400 или 1000) пар нуклеотидов (“п.н.”). Также направляющая последовательность может включать геномные последовательности из участков, находящихся за пределами последовательностей SEQ ID NO 4 или 5, лишь бы они включали по крайней мере 20 нуклеотидов из пределов одного из этих двух участков. Например, дополнительная направляющая последовательность может быть производной от последовательности, находящейся между SEQ ID NO 4 и сайтом инициации транскрипции гена FSHβ.
Из-за возможного полиморфизма в геномном локусе гена FSHβ небольшие изменения в нуклеотидном составе по данному геномному сайту-мишени могут иметь место у любого данного вида млекопитающего. Направляющие последовательности, которые соответствуют таким полиморфным вариантам последовательностей SEQ ID NO 4 или 5 (в частности, полиморфным вариантам, встречающимся у человека), попадают в объем настоящего изобретения.
В результате гомологичной рекомбинации регуляторная последовательность данной конструкции встраивается в заданный участок выше кодирующей последовательности гена FSHβ в хромосоме клетки. Полученный в результате новый транскриптон, включающий производную от конструкции регуляторную последовательность, изменяет экспрессию гена-мишени FSHβ. Вырабатываемый в результате белок FSHβ может быть идентичен по своей последовательности белку FSHβ, кодируемому неизмененным эндогенным геном, или же он может включать добавочные, замененные или сокращенные аминокислотные остатки по сравнению с белком FSHβ дикого типа, что обусловливается изменениями, внесенными в процессе гомологичной рекомбинации.
Изменение экспрессии гена включает в себя и активацию (т.е. побуждение к экспрессии) гена, который в норме является “молчащим” (т.е. по существу неэкспрессирован) в данной исходно полученной клетке, повышение или снижение уровня экспрессии гена и изменение параметров регуляции гена так, что в результате эти параметры отличаются от тех, которые имеются в исходно полученной клетке. Термин “исходно полученная клетка” обозначает клетку до прохождения гомологичной рекомбинации в ней.
Также в объем настоящего изобретения входит способ использования представляемой ДНК-конструкции, нацеленный на изменение экспрессии эндогенного гена FSHβ в клетке млекопитающего. Этот способ включает следующие этапы: (1) внесение ДНК-конструкции в клетку млекопитающего; (2) содержание клетки в условиях, которые способствуют гомологичной рекомбинации между данной конструкцией и геномным сайтом-мишенью, гомологичным данной направляющей последовательности, с образованием гомологично рекомбинантной клетки; (3) поддержание гомологично рекомбинантной клетки в условиях, которые способствуют экспрессии кодирующей последовательности FSHβ под контролем регуляторной последовательности, происходящей от данной конструкции. По крайней мере отчасти геномный сайт-мишень расположен с 5’-стороны от кодирующей последовательности эндогенного гена FSHβ. Следовательно, такой геномный сайт-мишень может включать кодирующую последовательность, равно как и 5’-некодирующую последовательность.
Также настоящее изобретение представляет трансфицированные или инфицированные клетки, в которых конструкция претерпела гомологичную рекомбинацию с геномной ДНК, находящейся выше эндогенного старт-кодона ATG одного или обоих аллелей эндогенного гена FSНβ. Такие трансфицированные или инфицированные клетки, также определяемые как претерпевшие гомологичную рекомбинацию клетки, характеризуются измененными параметрами экспрессии FSHβ. Эти клетки, особенно, применимы для выработки FSHβ in vitro и для доставки FSHβ методами генотерапии. Способы получения и применения таких клеток также охватываются настоящим изобретением. Такие клетки могут происходить от позвоночного животного, такого как млекопитающее (например, человек, не являющийся человеком примат, корова, свинья, лошадь, коза, овца, домашняя кошка, домашняя собака, кролик, мышь, морская свинка, хомячок или крыса).
Далее настоящее изобретение представляет способ получения белка FSHβ млекопитающего in vitro или in vivo путем внесения описанной выше конструкции в геном клетки-хозяина по механизму гомологичной рекомбинации. Затем претерпевшую гомологичную рекомбинацию клетку выдерживают в условиях, которые способствуют транскрипции, трансляции и, что необязательно, секреции белка FSHβ.
Также настоящее изобретение представляет выделенные нуклеиновые кислоты, включающие последовательность длиной по крайней мере 20 (например, по крайней мере 30, 50, 100, 200 или 1000) расположенных подряд нуклеотидов из состава SEQ ID NO 4 или по крайней мере 20 (например, по крайней мере 30, 50, 100 или 200) расположенных подряд нуклеотидов из состава SEQ ID NO 5, или сходного размера последовательность, идентичную участку SEQ ID NO 4 или 5, за исключением полиморфных вариаций или других небольших изменений (например, менее чем по 5% этой последовательности), которые не предотвращают гомологичной рекомбинации с последовательностью-мишенью.
В одном из вариантов выделенная нуклеиновая кислота по настоящему изобретению включает непрерывный 100-нуклеотидный блок из последовательности SEQ ID NO 4 или 5. Например, выделенная ДНК может включать нуклеотиды 1-100, 101-200, 201-300, 301-400, 401-500, 501-600, 601-700, 701-800, 801-900, 901-1000, 1001-1100, 1101-1200, 1201-1300, 1301-1400, 1401-1500, 1501-1600, 1601-1700, 1701-1800, 1801-1900, 1901-2000, 2001-2100, 2101-2200, 2201-2300, 2301-2400, 2401-2500, 2501-2600, 2601-2700, 2701-2800, 2801-2900, 2901-3000, 3001-3100, 3101-3200, 3201-3300, 3301-3400, 3401-3500, 3501-3600, 3601-3700, 3701-3800, 3801-3900, 3901-4000, 4001-4100, 4101-4200, 4201-4300, 4301-4400, 4401-4500, 4501-4600, 4601-4700, 4701-4800, 4801-4900, 4901-5000, 5001-5100, 5101-5200, 5201-5300, 5300-5400, 5401-5500, 5501-5600, 5601-5700, 5701-5800, 5801-5900, 5901-6000 или 5939-6038 из последовательности SEQ ID NO 4 или ее комплемента. С другой стороны, выделенная нуклеиновая кислота по настоящему изобретению может включать нуклеотиды 1-100, 101-200, 201-300, 301-400, 401-500 или 443-542 из последовательности SEQ ID NO 5. Эти блоки SEQ ID NO 4 или 5 или их комплементы применимы в качестве направляющих последовательностей в составе конструкций по настоящему изобретению.
В выделенной ДНК непрерывная нуклеотидная последовательность не связана с последовательностью, кодирующей полноразмерный FSHβ, или по крайней мере не связана в той же конфигурации (т.е. отделена такой же последовательностью), как в любом из нативных геномов. Таким образом, по использованию в данном тексте термин “выделенная ДНК” не обозначает хромосому или крупный фрагмент геномной ДНК (которая должна включаться в состав космиды или искусственной хромосомы дрожжей), который бы включал не только часть или полную последовательность SEQ ID NO 4 или 5, но также и интактную кодирующую последовательность FSHβ и все последовательности, которые находятся между кодирующей последовательностью FSHβ и последовательностью, соответствующей SEQ ID NO 4 или 5 в соответствии с расположением их в геноме клетки. Она включает, тем самым не ограничиваясь, ДНК, (i) которая встроена в плазмиду или вирус; или (ii) которая существует в виде отдельной молекулы, независимой от других последовательностей, например в виде фрагмента, полученного в методе полимеразной цепной реакции (“ПЦР”) или в результате расщепления рестриктазами. Предпочтительно выделенная ДНК не включает последовательность, которая кодирует интактный предшественник FSHβ (т.е. FSHβ с включением эндогенного сигнального сегмента, обеспечивающего секрецию).
Также настоящее изобретение представляет выделенную ДНК, включающую цепь, в составе которой имеется последовательность, которая включает по крайней мере 100 (например, по крайней мере 200, 400 или 1000) нуклеотидов и которая гибридизует либо в жестких условиях, либо в условиях средней степени жесткости с последовательностью SEQ ID NO 4 или 5 или с комплементом SEQ ID NO 4 или 5. Эта последовательность не связана с кодирующей последовательностью FSHβ или по крайней мере не связана в той конфигурации, которая имеет место в естественном геноме. Под условиями средней степени жесткости понимается проведение гибридизации при 50°С в буфере Черча (7% SDS, 0,5% NaHPO4, 1 M EDTA, 1% бычьего сывороточного альбумина) и промывки при 50°С в буфере 2×SSC. Высокая степень жесткости соответствует гибридизации при 42°С в присутствии 50% формамида, первой промывке при 65°С в 2×SSC, содержащем 1% SDS, и последующей второй промывке при 65°С в 0,1×SSC.
Также настоящим изобретением представляется выделенная ДНК, включающая цепь, в составе которой находится последовательность, которая (i) состоит по крайней мере из 100 (например, по крайней мере 200, 400 или 1000) нуклеотидов и (ii) проявляет по крайней мере 80%-ный уровень идентичности последовательности (например, 85%, 90%, 95% или 98%) с сегментом равной длины из последовательности SEQ ID NO 4 или 5 или комплемента SEQ ID NO 4 или 5. Эта последовательность не связана с кодирующей последовательностью гена FSHβ или по крайней мере не связана с ней в той же конфигурации, которая имеет место в любом нативном геноме.
Когда о конкретном полипептиде или молекуле нуклеиновой кислоты говорят, что она характеризуется определенным процентом идентичности или консерватизма по отношению к сравниваемому полипептиду или молекуле нуклеиновой кислоты, то такой процент идентичности или консерватизма определяют на основании алгоритма, предложенного Майерсом-Миллером (Myers & Miller, 1989, CABIOS), который встроен в программу ALIGN (версия 2.0) или в ее аналог, при том что величины штрафных баллов за точечный разрыв и разрыв последовательности по необходимости их использования установлены на уровне 4 и 12 соответственно. Все остальные параметры программы использованы по умолчанию. Программа ALIGN доступна из Интернета по адресу, например: http://www2.igh.cnrs.fr/bin/align-guess.cgi.
Также настоящее изобретение представляет способ введения животному FSHβ (например, млекопитающему, такому как человек, не являющийся человеком примат, корова, свинья, лошадь, коза, овца, домашняя кошка, домашняя собака, кролик, мышь, морская свинка, хомячок или крыса) путем получения клетки, у которой эндогенный ген FSHβ активирован в соответствии с описанным в данном тексте, с последующим имплантированием этой клетки данному животному, в организме которого данная клетка секретирует FSHβ. Также настоящим изобретением представляется способ выработки FSHβ путем получения клетки, у которой эндогенный ген FSHβ был активирован в соответствии с описанным в данном тексте, с последующим культивированием этой клетки in vitro в условиях, способствующих экспрессии и секреции FSHβ этой клеткой.
Выделенная ДНК по настоящему изобретению может быть использована, например, в качестве источника для прямой ПЦР-затравки с целью получения (путем сочетания с подходящей обратной затравкой) регуляторных и (или) кодирующих сегментов эндогенного гена FSHβ или в качестве гибридизационного зонда для указания на присутствие хромосомы 11 на препарате хромосом человека. Она также может быть использована в соответствии с описанным здесь далее в способе изменения экспрессии эндогенного гена FSHβ в клетке позвоночного животного.
За исключением специально оговариваемых случаев, все используемые в данном тексте методические и научные термины имеют те же значения, которые известны специалистам в той области техники, к которой относится настоящее изобретение. Ниже описаны методы и материалы, которые были взяты в качестве примеров, хотя методы и материалы, сходные или эквивалентные тем, что описаны в данной заявке, также могут быть использованы на практике осуществления или проверки настоящего изобретения. Все научные публикации, патентные заявки, патенты и другие ссылки, упоминающиеся здесь, включены во всей своей полноте только для сведения в виде библиографии. При возникновении каких-либо конфликтов настоящая заявка, включая терминологию, подлежит урегулированию. Упомянутые материалы, методы и примеры являются исключительно иллюстративными и не призваны ввести какие бы то ни было ограничения.
Другие параметры и преимущества настоящего изобретения будут ясны из нижеследующего подробного описания и прилагаемой формулы изобретения.
Краткое описание чертежей
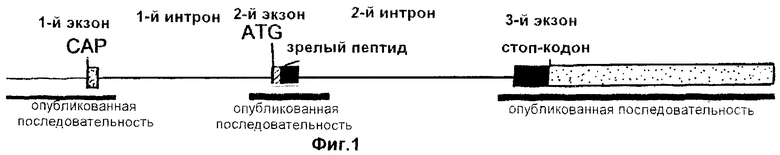
На фиг.1 схематически изображена геномная структура гена FSHβ человека.
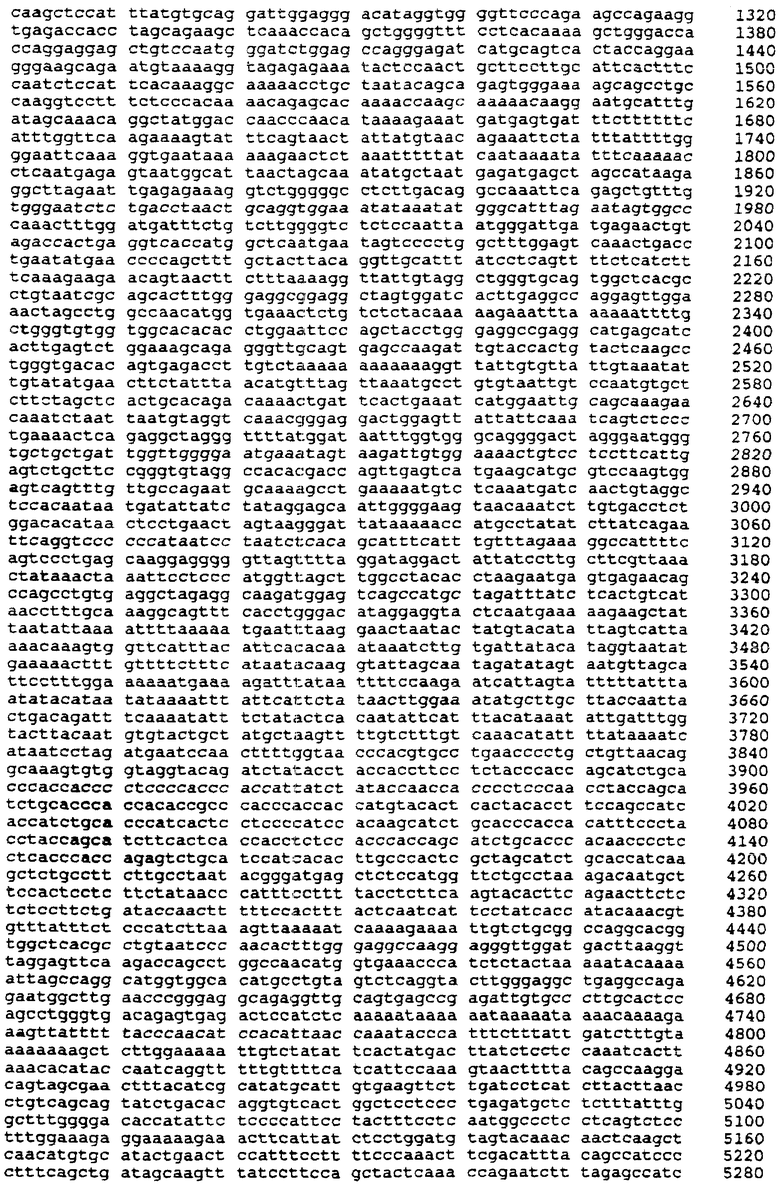
На фиг.2 схематически показан участок генома человека, в котором находится ген FSHβ (вверху), охватываемый вставкой (внизу) в составе плазмиды pHFB2. Три черные полосы в центре соответствуют геномным участкам данного гена, нуклеотидные последовательности которых опубликованы.
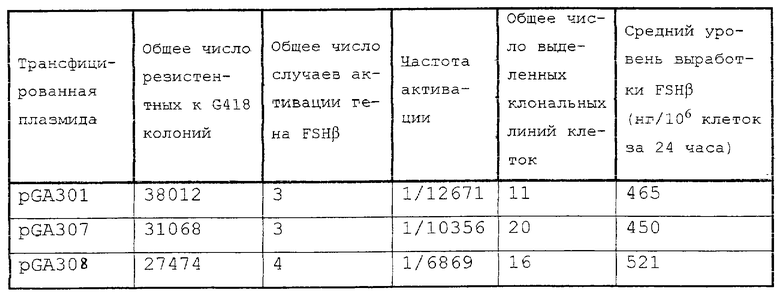
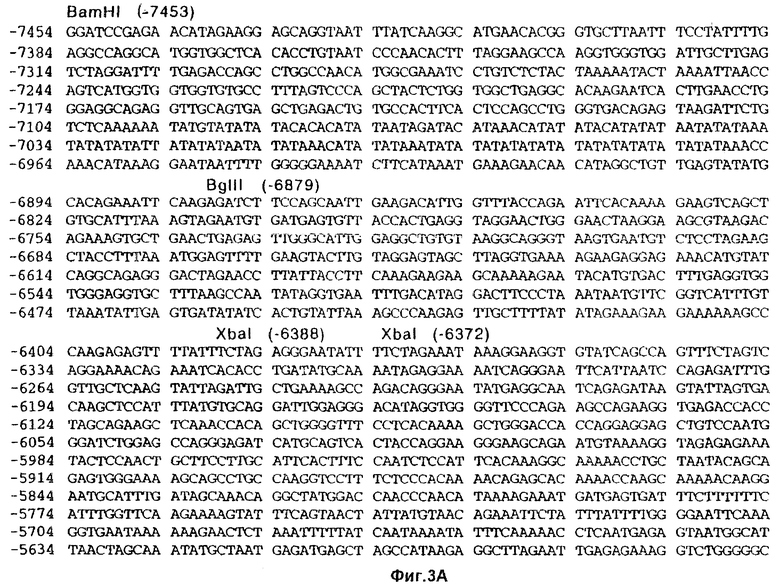
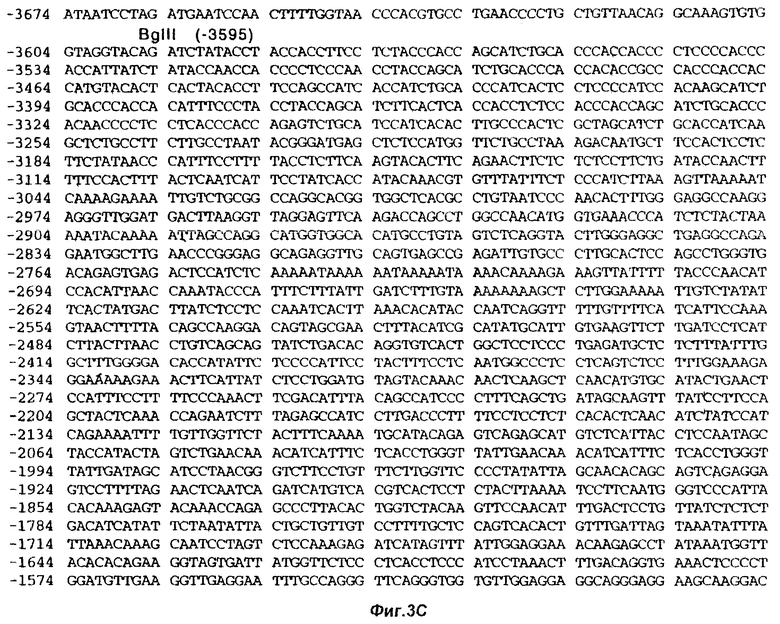
На фиг.3 представлена частичная нуклеотидная последовательность (SEQ ID NO 1) гена FSHβ человека, включая 7454 нуклеотида последовательности, расположенной с 5’-стороны от старт-кодона ATG. Также показана частичная полипептидная последовательность (SEQ ID NO 2), кодируемая данной кодирующей последовательностью. Опубликованные последовательности подчеркнуты. Метками “SD” и “SА” обозначены, соответственно, донорный и акцепторный сайты сплайсинга. Знак “зрелый” указывает на начало зрелого (процессированного) белка FSHβ.
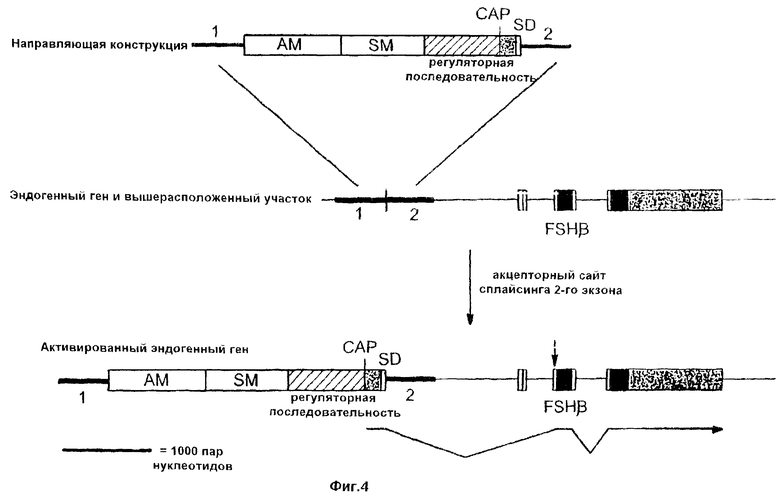
На фиг.4 схематически показана конструкция по настоящему изобретению. Эта конструкция включает первую направляющую последовательность (1); амплифицируемый маркерный ген (AM); селективный маркерный ген (SM); регуляторную последовательность; САР-сайт; последовательность, идентичную первому некодирующему экзону гена FSHβ человека; неспаренный донорный сайт сплайсинга (SD); вторую направляющую последовательность (2). Зачерненными участками показана кодирующая ДНК, а заштрихованными участками - нетранслируемые последовательности.
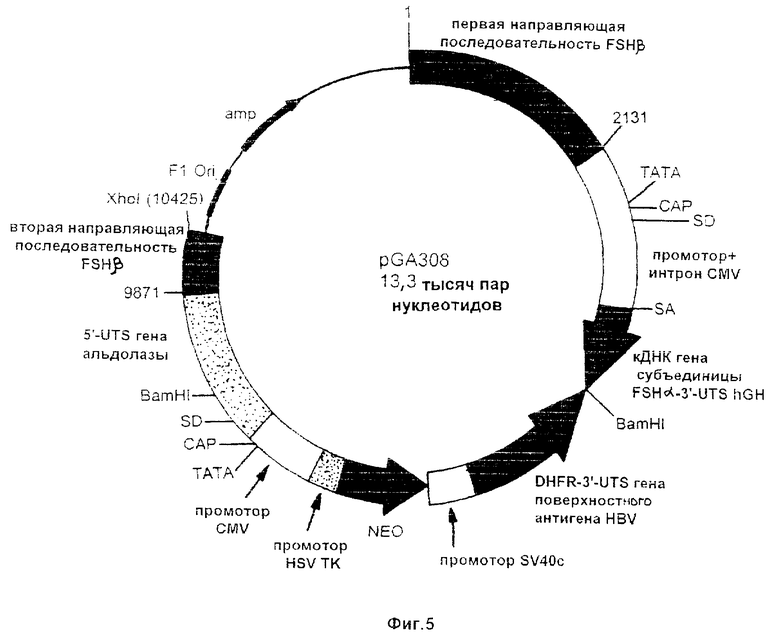
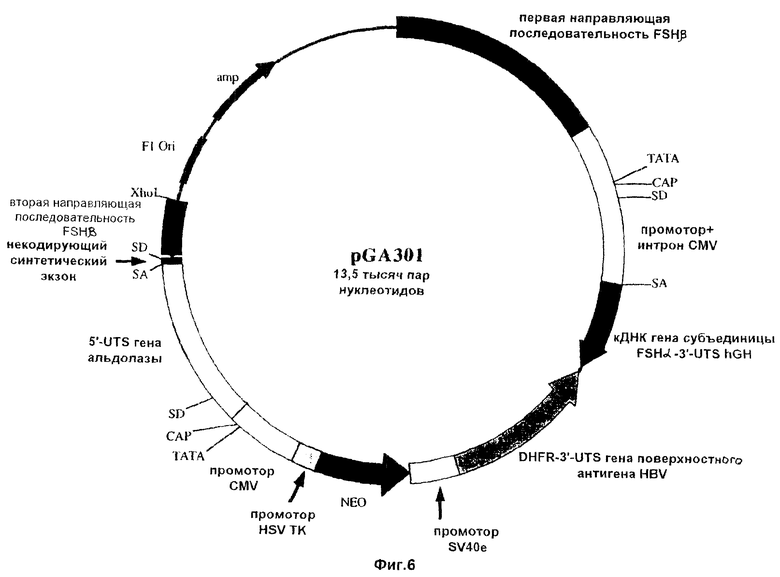
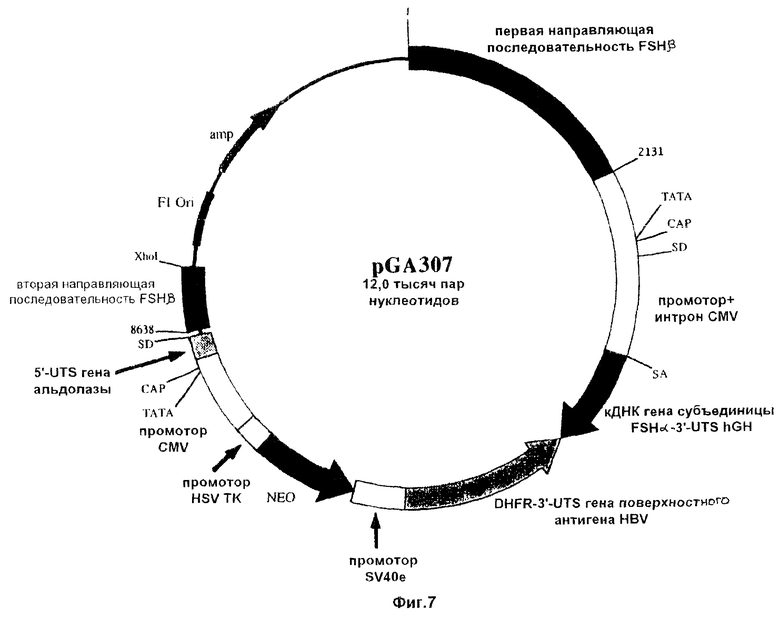
На фиг.5-7 схематически проиллюстрированы три конструкции по настоящему изобретению. Эти конструкции отличаются друг от друга по размеру 5’-нетранслируемого (5’-UTS) сегмента альдолазного гена, включаемого в состав плазмиды. Эти конструкции включают последовательность, кодирующую гликопротеиновую α-субъединицу ФСГ (т.е. FSHα), соединенную с промотором цитомегаловируса (CMV). На схеме использованы такие сокращенные обозначения: UTS - нетранслируемая последовательность; аmр - ампициллин; Ori - начало репликации; SD - донорный сайт сплайсинга; HSV ТК - ген тимидинкиназы простого герпесвируса; DHFR - дигидрофолатредуктаза; HBV - вирус гепатита-В; hGH - ген гормона роста человека.
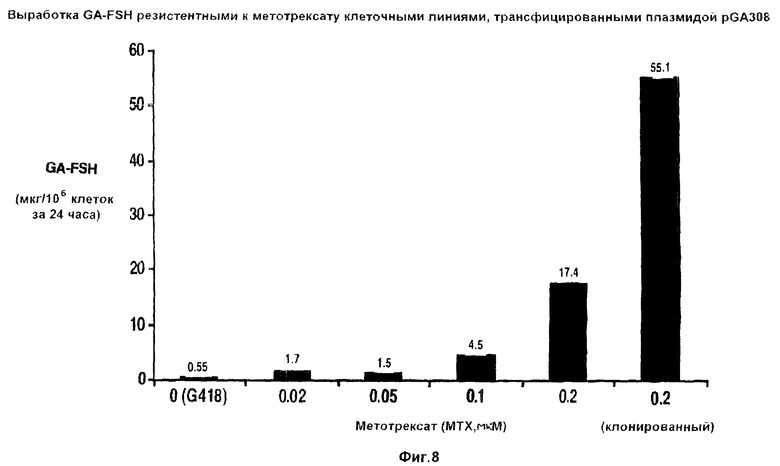
На фиг.8 показана диаграмма, отражающая выработку FSH клетками НТ-1080, трансфицированными плазмидой pGA308 (фиг.5) и отобранными по способности расти в присутствии различных концентраций метотрексата.
На фиг.9 показана последовательность SEQ ID NO 4 -последовательность, находящаяся выше сайта инициации транскрипции гена FSHβ человека.
На фиг.10 показана последовательность SEQ ID NO 5 -последовательность, находящаяся выше сайта инициации транскрипции гена FSHβ человека.
На фиг.11 показана первая направляющая последовательность (SEQ ID NO 6), используемая в конструкции по настоящему изобретению.
На фиг.12 показана вторая направляющая последовательность (SEQ ID NO 5), используемая в конструкции по настоящему изобретению.
Подробное описание
Настоящее изобретение основано на открытии нуклеотидного состава последовательности, расположенной выше кодирующей последовательности гена FSHβ человека.
FSH (ФСГ) является гонадотропином, который играет ключевую роль в росте и развитии ооцитов и сперматоцитов при нормальных физиологических условиях воспроизводства. Гормон ФСГ состоит из двух субъединиц - α и β, причем вторая из них обеспечивает биологическую специфичность ФСГ.
Ген FSHβ человека кодирует белок-предшественник, состоящий из 129 аминокислот, из которых 16 аминокислот приходятся на сигнальный пептид. Данный ген составлен тремя экзонами и двумя интронами, причем первый экзон является некодирующим экзоном. Геномная карта гена FSHβ человека показана на фиг.1. Эта карта выстроена на основе опубликованных последовательностей (HUMFSHBQ1, депозитарные №№ в базе данных GenBank - М54912, М38644, М21219 и М18536), которые соответствуют трем отдельным сегментам генома (фиг.1). Первый сегмент состоит из 720 пар нуклеотидов - 503 п.н. нетранслируемой последовательности и 1-й экзон (63 п.н.; некодирующий) и 127 п.н. 1-го интрона. Второй сегмент начинается с нуклеотида [-152] и заканчивается нуклеотидом [+367] (все номера положений, используемые здесь, даны, за исключением специально оговариваемых случаев, по отношению к сайту инициации трансляции). Этот сегмент включает 146 п.н. 1-го интрона, весь 2-й экзон (165 п.н.) и 208 п.н. 2-го интрона. Третий сегмент включает 102 п.н. 2-го интрона и весь 3-й экзон, а также 1480 пар нуклеотидов после стоп-кодона.
Конкретные последовательности с 5’-стороны от кодирующей последовательности FSHβ и их использование для целей изменения экспрессии эндогенного гена FSHβ.
Для получения геномной ДНК, включающей последовательность вверх от гена FSHβ, скринингу с использованием 40-нуклеотидного зонда ВЕТА2 подвергали геномную библиотеку лейкоцитов человека, встроенную в λЕMBL2 (Clontech, № по каталогу HL1006d). Этот зонд является производным 23 п.н. 1-го экзона и 17 п.н. 1-го интрона и имеет следующую последовательность:
5’-TTGGCATCTACCGTTTTCAAGTGGTGACAGCTACTTTTGA-3’ (SEQ ID NO 3).
С использованием радиоактивно помеченного зонда ВЕТА2 были проскринированы примерно миллион рекомбинантных фагов. Была выделена одна фаговая бляшка, обозначенная 8-1-1-1. HindIII/KpnI-фрагмент длиной 7600 пар нуклеотидов из состава фага 8-1-1-1 был субклонирован в состав pBluescript-II-SK+ (Stratagene, La Jolla, CA) с получением плазмиды, включающей примерно 6600 пар нуклеотидов верхней последовательности, 1-й экзон, 1-й интрон, 2-й экзон и 9 п.н. 2-го интрона (фиг.2). Эту плазмиду обозначили как pHFB2.
Плазмиду pHFB2 секвенировали по методу Сэйнджера. Путем сопоставления полученных данных секвенирования получили полную последовательность фаговой вставки 8-1-1-1. Эта нуклеотидная последовательность (SEQ ID NO 1) показана на фиг.3.
Как было показано, эта вставка охватывает участок гена FSHβ длиной 7622 пары нуклеотидов, начиная с нуклеотида [-7454] (фиг.3). Последовательности, охватывающие нуклеотиды [-7454]-[-1417] (6038 п.н. верхней последовательности: SEQ ID NO 4), и нуклеотиды [-696]-[-155] (542 п.н. 1-го интрона: SEQ ID NO 5) ранее опубликованы не были.
Для изменения экспрессии эндогенного гена FSHβ был применен базовый подход, проиллюстрированный на фиг.4. Роль первой направляющей последовательности (5’) выполняли нуклеотиды 3860-5784 последовательности SEQ ID NO 4, в то время как второй направляющей последовательностью (3’) являлась последовательность SEQ ID NO 5. ДНК-фрагменты, включающие такие последовательности, затем субклонировали в состав плазмид с получением направляющих конструкций pGA308, pGA301 и pGA307, которые изображены на фиг.5-7 соответственно. Каждая из этих плазмид включает примерно по 3200 п.н. 5’-направляющей последовательности и примерно по 500 п.н. 3’-направляющей последовательности.
Клетки НТ-1080 были раздельно трансфицированы каждой из указанных плазмид и помещены в селективную среду с антибиотиком G418. Примерно через 2 недели подсчитывали резистентные к G418 колонии в 6-луночных планшетах. Кроме того, кондиционированную среду в каждой лунке тестировали методом ТИФА на экспрессию GA-FSH. Клетки, характеризующиеся выработкой GA-FSH, обрабатывали трипсином и подсчитывали. Затем эти клетки разбавляли и вносили на 96-луночные планшеты для формирования клонов. Примерно через 2 недели культивирования клональные клеточные популяции тестировали методом ТИФА на экспрессию GA-FSH. Колонии, для которых устанавливалась выработка GA-FSH, воспроизводили в культуре и запасали для дальнейшего анализа. В таблице обобщены данные по частотам активации эндогенного гена и другим наблюдениям по описанной выше процедуре клонирования.
Более подробно были исследованы клетки, трансфицированные плазмидой pGA308. На фиг.8 показан диапазон уровней выработки ФСГ, проявляемых клетками НТ1080, трансфицированными pGA308 и культивировавшимися в среде, содержащей метотрексат в различных концентрациях. Линия, помеченная как “0,2 (клонированная)”, соответствует выработке ФСГ клеточной линией, клонированной с помощью ограничивающего разведения клеток, резистентных к 0,2 мкМ метотрексата. Полученные результаты, отображенные на фиг.8, отчетливо указывают на то, что при более высоких концентрациях метотрексата можно выделить такие клеточные линии, которые вырабатывают по крайней мере 50 мкг ФСГ на миллион клеток в день.
Базовые методы
Изменение уровня экспрессии эндогенного гена FSHβ
С использованием описанных выше верхних последовательностей гена FSHβ можно изменять экспрессию эндогенного гена FSHβ человека с помощью способа, принцип которого был описан в патенте США №5641670. Одна из таких стратегий изображена на фиг.4. В этом случае направляющая конструкция создается таким образом, чтобы она включала первую направляющую последовательность, гомологичную первому сайту-мишени на участке выше данного гена, амплифицируемый маркерный ген, ген селективного маркера, регуляторный сегмент, САР-сайт, экзон, неспаренный донорный сайт сплайсинга и вторую направляющую последовательность, соответствующую второму сайту-мишени, находящемуся ниже первого сайта-мишени и заканчивающемуся в пределах кодирующей последовательности FSHβ или выше ее. В этом подходе первый и второй сайты-мишени до гомологичной рекомбинации непосредственно примыкают друг к другу на хромосоме, хотя такая конфигурация и не является обязательной (см. ниже). Претерпевшие гомологичную рекомбинацию клетки будут вырабатывать мРНК-предшественник, который соответствует экзогенным экзону и донорному сайту сплайсинга, а также всем последовательностям, находящимся между этим донорным сайтом сплайсинга и последовательностью терминации транскрипции гена FSHβ, включая интроны, экзоны и 3’-нетранслируемый сегмент гена FSHβ (фиг.4). В результате сплайсинга такой про-мРНК образуется мРНК, в которой экзогенный экзон слит со 2-м экзоном эндогенного гена FSHβ. В результате трансляции этой мРНК образуется предшественник FSHβ.
Также могут быть использованы и другие подходы. Например, первый и (или) второй сайты-мишени могут находиться в пределах первого интрона гена FSHβ. С другой стороны, ДНК-конструкция может быть сформирована таким образом, чтобы включать (в направлении от 5’ к 3’) первую направляющую последовательность, амплифицируемый маркерный ген, ген селективного маркера, регуляторный сегмент, САР-сайт, экзон, донорный сайт сплайсинга, интрон, акцепторный сайт сплайсинга и вторую направляющую последовательность. При таком подходе 5’-конец второго направляющего сайта предпочтительно расположен менее чем в 40 п.н. выше нативного сайта инициации транскрипции гена FSHβ, что имеет целью исключить нежелательные старт-кодоны ATG. После гомологичной рекомбинации в этом локусе образующаяся про-мРНК будет включать экзогенный экзон, экзогенный донорный сайт сплайсинга, экзогенный интрон, экзогенный акцепторный сайт сплайсинга и все последовательности, находящиеся между экзогенным акцепторным сайтом сплайсинга и сайтом терминации транскрипции эндогенного гена FSHβ. В результате сплайсинга такого транскрипта будет образовываться мРНК, которая может транслироваться с получением предшественника FSHβ человека, включающий либо нормальную сигнальную последовательность секреции FSHβ, либо генетически сконструированную сигнальную последовательность секреции. Размер экзогенного интрона, а следовательно, и положение экзогенного регуляторного сегмента по отношению к кодирующей последовательности эндогенного гена могут варьироваться с целью оптимизации функций этого регуляторного сегмента.
При любой стратегии активации нет необходимости в непосредственном примыкании или даже близком соседстве первого и второго сайтов-мишеней. Когда они не находятся в непосредственной близости друг от друга, участок нормального верхнего сегмента гена FSHβ и (или) участок кодирующего сегмента могут быть делегированы в процессе гомологичной рекомбинации.
Если желательно, продукт активированного гена FSHβ может быть получен в клетке такого типа, который характеризуется экспрессией гена, кодирующего гликопротеиновую α-субъединицу человека ФСГ (FSHα), продукт которой образует гетеродимер с продуктом гена FSHβ. Это может быть нативный клеточный штамм или клеточная линия. С другой стороны, ген гликопротеиновой α-субъединицы человека (GenBank: последовательность № HUM-GLYCAl) может быть коэкспрессирована с продуктом гена FSHβ, при том что такая коэкспрессия достигается в результате экспрессии гена гликопротеиновой α-субъединицы человека или кДНК под контролем подходящего промотора или путем активации гена гликопротеиновой α-субъединицы человека с применением описанных в данной заявке способов.
В качестве примера последовательность, кодирующая гликопротеиновую α-субъединицу, может быть включена в состав ДНК-конструкции. Эта кодирующая последовательность помещается под транскрипционный контроль, осуществляемый регуляторной последовательностью, нуклеотидный состав которой может быть идентичен регуляторной последовательности, которая обеспечивает экспрессию эндогенного гена FSHβ, или может отличаться от нее. На фиг.5-7 показаны примеры таких конструкций.
ДНК-конструкция
ДНК-конструкция по настоящему изобретению по крайней мере включает направляющую последовательность и регуляторную последовательность. Дополнительно она может включать экзон; или экзон и неспаренный донорный сайт сплайсинга; или экзон, донорный сайт сплайсинга, интрон и акцепторный сайт сплайсинга. Экзон, если он присутствует, расположен с 3’-стороны регуляторной последовательности, а неспаренный донорный сайт сплайсинга находится с 3’-стороны экзона. Интрон и акцепторный сайт сплайсинга, если они присутствуют, находятся с 3’-стороны донорного свита, сплайсинга. Кроме того, могут присутствовать множественные экзоны и интроны (с подходящими донорными и акцепторными сайтами), предвосхищающими (т.е. находящимися с 5’-конца от) экзон, фланкированный неспаренным донорным сайтом сплайсинга. ДНК в составе данной конструкции обозначают как “экзогенную”, т.е. эта ДНК не является исходной частью генома клетки-хозяина. Экзогенная ДНК может включать последовательности, идентичные частям эндогенной геномной ДНК, присутствующей в клетке перед трансфекцией, или инфекции вирусным вектором, или отличающиеся от них. По использованию в данном тексте термин “трансфекция” обозначает внесение плазмиды в клетку с помощью химических и физических способов, таких как совместная преципитация с фосфатом кальция или хлоридом кальция, трансфекция, опосредованная DEAE-декстраном, липофекция, электропорация, микроинъекция, стрельба микрочастицами или бомбардировка. По использованию в данном тексте термин “инфекция” обозначает внесение вирусной нуклеиновой кислоты в клетку путем заражения вирусом. Различные элементы, входящие в состав ДНК-конструкции по настоящему изобретению, подробно описаны далее.
ДНК-конструкция также может включать цис-активные или транс-активные вирусные последовательности (например, сигналы упаковки), что тем самым обеспечивает доставку конструкции в ядро клетки в процессе вирусной инфекции. При необходимости ДНК-конструкция может быть “освобождена” от различных стадий жизненного цикла вируса, таких как ретровирусное встраивание в геном, обусловливаемое активностью интегразы, или формирование эписомы. Такое “освобождение” может быть осуществлено путем подходящих делений или мутаций вирусных последовательностей, таких как делеция участка, кодирующего интегразу, в ретровирусном векторе. Дополнительные подробности, касающиеся конструирования и использования вирусных векторов, можно найти у Robbins et al., 1998, Phamacol. Ther., 80, 35-47; Gunzburg et al., 1995, Mol. Med. Today, 1, 410-417: включены здесь для сведения в виде библиографических ссылок.
Направляющие последовательности
Направляющие последовательности обеспечивают гомологичную рекомбинацию желательной последовательности по выбранному сайту генома организма-хозяина. Направляющие последовательности гомологичны (т.е. способны к гомологичной рекомбинации с) соответствующим сайтам-мишеням генома организма-хозяина.
В состав кольцевой ДНК-конструкции может входить единственная направляющая последовательность или же две или большее число направляющих последовательностей. В линейной ДНК-конструкции может иметься две или большее число направляющих последовательностей. Сайт-мишень, которому гомологична данная направляющая последовательность, может находиться в пределах экзона и (или) интрона гена FSHβ, выше кодирующего участка гена FSHβ и в непосредственном соседстве с ним или выше кодирующего участка гена FSHβ и на некотором удалении от него.
Первая из двух направляющих последовательностей данной конструкции (или целая направляющая последовательность, если она является единственной направляющей последовательностью в данной конструкции) по крайней мере отчасти происходит от новых представленных геномных участков, расположенных выше кодирующих последовательностей гена FSHβ. Эта направляющая последовательность включает участок SEQ ID NO 1, например, по крайней мере 20 подряд расположенных нуклеотидов последовательности, которая соответствует положениям [-7454]-[-1417] (SEQ ID NO 4) или положениям [-696]-[-155] (SEQ ID NO 5). Вторая из двух направляющих последовательностей в данной конструкции может направлять участок генома выше кодирующей последовательности (например, также включать участок SEQ ID NO 4 или 5) или направлять экзон или интрон данного гена.
Направляющая последовательность (последовательности) может дополнительно включать последовательность, являющуюся производной от известного сегмента гена FSHβ, включая те, которые описаны в настоящей заявке, равно как и от участков, находящихся еще выше, структура которых пока не установлена, но которые могут быть картированы специалистами в данной области техники.
Геномные фрагменты, которые могут быть использованы в качестве направляющих последовательностей, могут быть идентифицированы по их способности гибридизовать с зондом, включающим полностью или частично SEQ ID NO 4 или 5. Такой зонд может быть сформирован с помощью метода ПЦР с использованием затравок, производных от SEQ ID NO 1.
Регуляторная последовательность
Регуляторная последовательность в составе ДНК-конструкции может включать один или несколько промоторов (например, конститутивный, тканеспецифичный или индуцибельный промотор), энхансеров, сайтов прикрепления к ядерному скелету или матриксу, негативные регуляторные элементы, сайты связывания транскрипционных факторов или сочетания этих элементов.
Регуляторная последовательность может являться производной от эукариотического (например, млекопитающего) или вирусного генома. Применимыми регуляторными последовательностями являются, тем самым не ограничиваясь, те, которые регулируют экспрессию ранних или поздних генов вируса SV40, гены цитомегаловируса и главные поздние гены аденовируса. Они также включают регуляторные сегменты, производные от генов, кодирующих металлотионеин-I мыши, фактор-1α элонгации, коллаген (например, субъединицы коллагенов Iα1 и Iα2 и коллаген-IV), актин (например, γ-актин), иммуноглобулин, HMG-KoA-редуктазу, глицеральдегидфосфатдегидрогеназу, 3-фосфоглицераткиназу, коллагеназу, стромелизин, фибронектин, виментин, ингибитор-I активатора плазминогена, тимозин-β4, тканевые ингибиторы металлопротеиназ, рибосомные белки, полипептиды главного комплекса гистосовместимости и лейкоцитарные антигены человека.
Предпочтительно регуляторная последовательность включает сайт связывания транскрипционного фактора, такой как сайт связывания бокса ТАТА, бокса ССААТ, AP1, Sp1 или NF-κВ.
Маркерные гены
Если желательно, данная конструкция может включать последовательность, кодирующую желательный полипептид, функционально присоединенную к его собственному промотору. Примером этому может являться ген селективного маркера, который может быть использован для облегчения идентификации этапа направления. Амплифицируемый маркерный ген также может быть использован для облегчения отбора клеток, несущих коамплифицированные фланкирующие последовательности ДНК. Клетки, несущие амплифицированные копии амплифицируемого маркерного гена, могут быть идентифицированы по их росту в присутствии агента, являющегося селективным в отношении экспрессии амплифицируемого гена. Активированный эндогенный ген обычно амплифицируется вместе амплифицируемым селективным маркерным геном. Клетки, несущие множественные копии активированного эндогенного гена, могут вырабатывать очень высокие титры белка FSHβ и, следовательно, применимы для выработки белка in vitro и генотерапии.
Гены селективного и амплифицируемого маркеров не должны быть расположены в непосредственной близости друг от друга. Ген амплифицируемого маркера и ген селективного маркера могут являться одним и тем же геном. Один или оба маркерных гена могут быть расположены в последовательности интрона ДНК-конструкции. Подходящие гены амплифицируемых маркеров и гены селективных маркеров описаны в патенте США №5641670.
Экзогенный экзон
Далее ДНК-конструкция может включать экзон, т.е. последовательность ДНК, которая транскрибируется в РНК и присутствует в зрелой (процессированной) молекуле мРНК. По использованию в данном тексте понятие “экзон” в составе конструкции относится к экзогенному или производному от конструкции экзону. Экзогенный экзон может быть некодирующим, как первый экзон гена FSHβ человека, причем на самом деле, но не обязательно, может быть идентичен по последовательности указанного экзона. С другой стороны, экзогенный экзон кодирует один или большее число аминокислотных остатков или хотя бы отчасти кодирует аминокислоту (т.е. включает 1-2 нуклеотида одного кодона). Когда экзон включает кодирующую последовательность, ДНК-конструкция должна выстраиваться таким образом, чтобы в процессе транскрипции и сплайсинга его кодирующая рамка попадала в общую открытую рамку гена-мишени FSHβ. Следовательно, экзогенный экзон после сплайсинга соединяется с эндогенным экзоном так, что это не приводит к изменению соответствующей рамке считывания части мРНК, происходящей от эндогенного экзона.
Включение кодирующего экзона в состав ДНК-конструкции обеспечивает выработку химерного белка, который включает и эндогенную последовательность белка FSНβ, и экзогенную белковую последовательность. Такой химерный белок может сочетать в составе одного полипептида структурные, каталитические свойства или свойства по связыванию с лигандом или рецептором, производные от двух или большего числа белков. Например, экзогенный экзон может кодировать поверхностно-клеточный “якорь”, сигнальный пептид, улучшающий клеточную секрецию, лидерную последовательность, каталитический домен, сайт связывания кофактора или эпитопную метку, необходимую для облегчения очистки химерного белка FSHβ, выработанного данным рекомбинантным генным локусом.
Донорный сайт сплайсинга
Экзогенный экзон фланкирован по своему 3’-концу донорным сайтом сплайсинга. Донорным сайтом сплайсинга является последовательность, которая обеспечивает сплайсинг одного экзона в РНК-транскрипте с акцепторным сайтом сплайсинга другого экзона РНК-транскрипта. Обычно первый экзон находится с 5’-стороны от второго экзона, а донорный сайт сплайсинга, расположенный с 3’-конца первого экзона, спарен с акцепторным сайтом 5’-конца второго экзона. Донорные сайты сплайсинга характеризуются консервативной консенсусной последовательностью - (A/C)AGGURAGU (где R обозначает пуриновый нуклеотид), при том что GU в 4-м и 5-м положениях являются обязательными (Jackson, 1991, Nucl. Acids Res., 19, 3715-3798). Первые три нуклеотида в составе консенсусного донорного сайта сплайсинга являются тремя последними нуклеотидами экзона: т.е. они непосредственно не сплайсируются. Донорные сайты сплайсинга функционально определяются по их способности обеспечивать соответствующую реакцию в механизме сплайсинга мРНК.
В качестве примера донорный сайт сплайсинга может быть помещен с 3’-конца и непосредственно за старт-кодоном ATG, когда присутствие одного или нескольких промежуточных нуклеотидов не является обязательным для того, чтобы экзогенный экзон попадал в фазу считывания рамки второго экзона гена-мишени. Когда экзогенный экзон кодирует одну или большее число аминокислот в фазе кодирующей последовательности гена-мишени, донорный сайт сплайсинга предпочтительно может быть помещен непосредственно встык к экзогенной кодирующей последовательности с ее 3’-конца.
Донорный сайт сплайсинга, фланкирующий экзогенный экзон, в составе конструкции не спарен, т.е. в самой конструкции снизу от донорного сайта сплайсинга отсутствует акцепторный сайт сплайсинга, с которым впоследствии должно происходить сплайсинговое присоединение. После гомологичной рекомбинации по сайту-мишени, находящемуся выше кодирующей последовательности гена FSHβ, имеющийся в конструкции неспаренный донорный сайт сплайсинга функционально присоединяется (“спаривается”) с эндогенным акцепторным сайтом сплайсинга из последовательности эндогенного экзона FSHβ. В результате процессинга транскрипта, считанного с претерпевшего гомологичную рекомбинацию гена FSHβ, происходит сплайсинг экзогенного экзона с акцепторным сайтом сплайсинга эндогенного экзона.
Также конструкция по настоящему изобретению может включать акцепторный сайт сплайсинга. Этот сайт в сочетании с донорным сайтом сплайсинга обеспечивает сплайсинг (“сшивание”) одного экзона с другим. Акцепторные сайты сплайсинга характеризуются последовательностью (Y)10NYAG (SEQ ID NO 7), где Y обозначает любой пиримидин и N обозначает любой нуклеотид (Jackson, 1991, Nucl. Acids Res., 19, 3715-3798).
Интроны
Необязательно ДНК-конструкция может включать интрон. Интрон - это последовательность, состоящая из одного или большего числа нуклеотидов, находящаяся между донорным сайтом сплайсинга и акцепторным сайтом сплайсинга и удаляемая при сплайсинге из молекулы про-РНК в ходе образования зрелой молекулы мРНК.
САР-сайт
Необязательно ДНК-конструкция включает САР-сайт. САР-сайт - это специфичный сайт инициации транскрипции, который связан с регуляторным сегментом и используется им. Такой САР-сайт находится в таком положении конструкции по отношению к регуляторной последовательности, чтобы после прохождения гомологичной рекомбинации регуляторная последовательность могла обеспечивать синтез транскрипта, который начинается от САР-сайта. С другой стороны, при отсутствии САР-сайта в составе конструкции транскрипционный аппарат должен располагаться по умолчанию так, чтобы подходящий сайт гена-мишени мог быть использован в качестве САР-сайта.
Дополнительные ДНК-элементы
Дополнительно конструкция может включать последовательности, которые влияют на структуру или стабильность РНК или белка, образующегося после гомологичной рекомбинации. Необязательно ДНК-конструкция может включать бактериальный сайт начала репликации и бактериальные маркерные гены резистентности к антибиотикам или иные селективные маркеры, которые обеспечивают крупномасштабное воспроизводство плазмид в бактериальных клетках или любых других подходящих системах клонирования и организмов-хозяев.
Все описывавшиеся выше элементы ДНК-конструкции функциональным образом соединяют друг с другом или размещают функционально по отношению друг к другу. В результате после гомологичной рекомбинации между конструкцией и геномной ДНК-мишенью регуляторная последовательность может обеспечивать синтез первичного РНК-транскрипта, который инициируется по САР-сайту (необязательно входящему в состав данной конструкции) и включает: (1) последовательность, соответствующую экзону и донорному сайту сплайсинга данной конструкции, если они присутствуют, и (2) последовательность, находящуюся между донорным сайтом сплайсинга и сайтом терминации транскрипции эндогенного гена. Последняя из указанных последовательностей может включать регуляторный сегмент эндогенного гена FSHβ, равно как и последовательности, являющиеся для этого сегмента соседними, но в норме не транскрибируемые. В конфигурации функционального соединения донорный сайт сплайсинга направляющей конструкции обеспечивает процесс сплайсинга с акцепторным сайтом сплайсинга, фланкирующим один из экзонов эндогенного гена FSHβ так, чтобы желательный белок мог быть выработан на основе полностью сплайсированного зрелого транскрипта. Акцепторный сайт сплайсинга может быть эндогенным таким образом, что процесс сплайсинга регулирует эндогенный экзон. В другом варианте, в котором в состав направляющей конструкции включается акцепторный сайт сплайсинга, в результате процесса сплайсинга удаляется экзогенный интрон, вносимый направляющей конструкцией.
Порядок расположения элементов в составе ДНК-конструкции может варьироваться. Когда данная конструкция является кольцевой плазмидой или вирусным вектором, относительное расположение элементов в образуемой структуре может быть, например, таким: направляющая последовательность, плазмидная ДНК (состоящая из последовательностей, используемых для отбора и/или обеспечения репликации направляющей плазмиды в бактериальном или ином подходящем организме-хозяине), селективный(ые) маркер(ы), регуляторная последовательность, экзон и неспаренный донорный сайт сплайсинга.
Когда конструкция является линейной, порядок может быть, например, таким: первая направляющая последовательность, ген селективного маркера, регуляторная последовательность, экзон, донорный сайт сплайсинга и вторая направляющая последовательность; или, как вариант, первая направляющая последовательность, регуляторная последовательность, экзон, донорный сайт сплайсинга, ген селективного маркера и вторая направляющая последовательность. Порядок расположения этих элементов также может быть таким: первая направляющая последовательность, селективный маркер, регуляторная последовательность, экзон, донорный сайт сплайсинга, интрон, акцепторный сайт сплайсинга, необязательно внутренний сайт прикрепления к рибосоме и вторая направляющая последовательность.
С другой стороны, порядок расположения может быть таким: первая направляющая последовательность, первый ген селективного маркера, регуляторная последовательность, экзон, донорный сайт сплайсинга, вторая направляющая последовательность и второй ген селективного маркера; или первая направляющая последовательность, регуляторная последовательность, экзон, донорный сайт сплайсинга, первый ген селективного маркера, вторая направляющая последовательность и второй ген селективного маркера. Рекомбинация между направляющими последовательностями, фланкирующими первый селективный маркер, с гомологичными последовательностями генома организма-хозяина обусловливает направленное встраивание данного селективного маркера, в то время как второй селективный маркер не встраивается. Желательными трансфицированными или инфицированными клетками являются клетки, которые были стабильно трансфицированы или инфицированы первым селективным маркером, но не вторым селективным маркером. Такие клетки могут быть отобраны по их росту в культуральной среде, содержащей агент, являющийся селективным по экспрессии первого маркера, и другой агент, являющийся селективным по отношению к другому маркеру. Трансфицированные или инфицированные клетки, которые характеризуются неправильным встраиванием направляющей конструкции, обусловленным иным, нежели гомологичная рекомбинация, механизмом, как можно ожидать, будут экспрессировать второй маркерный ген и поэтому будут уничтожаться в культуральной среде.
Позитивный селективный маркер иногда вносят в состав конструкции для того, чтобы обеспечить отбор клеток, несущих амплифицированные копии такого маркера. В таком варианте порядок расположения элементов конструкции, например, таков: первая направляющая последовательность, позитивный амплифицируемый селективный маркер, второй селективный маркер (необязательный), регуляторная последовательность, экзон, донорный сайт сплайсинга и вторая направляющая последовательность ДНК.
Различные элементы данной конструкции могут быть получены из естественных источников (например, из геномной ДНК) или могут быть получены с применением генно-инженерных методов или синтетическим путем. Регуляторный сегмент, САР-сайт, экзон, донорный сайт сплайсинга и необязательно интрон и акцепторный сайт сплайсинга в данной конструкции могут быть выделены в виде цельной единицы, например, из последовательностей гена фактора-1α элонгации человека (GenBank, последовательность HUMEF1A) или сегмента немедленно раннего гена цитомегаловируса (GenBank, последовательность HEHCMVP1). Эти компоненты также могут быть выделены из разных генов.
Трансфекция или инфекция и гомологичная рекомбинация
ДНК-конструкция по настоящему изобретению может быть внесена в клетку, такую как первичная, вторичная или иммортализованная клетка, в виде единичной ДНК-конструкции или в виде отдельных последовательностей ДНК, которые встраиваются в хромосомную иди ядерную ДНК трансфицированной или инфицированной клетки. ДНК может быть внесена в виде линейной, двухцепочечной (включая одноцепочечные участки по одному или обоим концам или без них), одноцепочечной или кольцевой молекулы. ДНК-конструкция или ее РНК-эквивалент также могут быть внесены в виде вирусной нуклеиновой кислоты.
Когда конструкцию вносят в клетки-хозяева в виде двух раздельных ДНК-фрагментов, эти два фрагмента имеют участки гомологии последовательностей ДНК (т.е. “перекрываются”) по 3’-концу одного фрагмента и 5’-концу другого фрагмента, при том что один из них включает первую направляющую последовательность, другой включает вторую направляющую последовательность. После внесения в клетку эти два фрагмента претерпевают гомологичную рекомбинацию с образованием единой молекулы, в которой первая и вторая направляющие последовательности фланкируют сегмент перекрывания двух исходных фрагментов. После этого образовавшаяся молекула становится пригодной для гомологичной рекомбинации с клеточными сайтами-мишенями. Может быть использовано более двух фрагментов, при том что каждый из них формируют таким образом, чтобы они могли претерпевать гомологичную рекомбинацию друг с другом с образованием однозначного продукта, подходящего для гомологичной рекомбинации с клеточными сайтами-мишенями в соответствии с описанным выше.
ДНК-конструкция по настоящему изобретению, если сама она не включает селективный маркер, может быть трансфицирована или инфицирована одновременно с другой конструкцией, включающей такой маркер. Направляющая плазмида может быть расщеплена рестриктазой по одному или нескольким сайтам с целью получения линейной или разорванной молекулы перед трансфекцией или инфекцией. Полученные в результате свободные концы ДНК обеспечивают более высокую частоту желательного процесса гомологичной рекомбинации. Кроме того, свободные концы ДНК могут быть обработаны экзонуклеазой с целью формирования “выступающих” 5’- или 3’-одноцепочечных концов ДНК (например, по крайней мере 30 нуклеотидов в длину, а предпочтительно 100-1000 нуклеотидов в длину) с целью повышения частоты желательного процесса гомологичной рекомбинации. В этом варианте гомологичная рекомбинация между направляющей последовательностью и геномной мишенью должна обусловливать появление двух копий направляющих последовательностей, фланкирующих элементы, имеющиеся в составе внесенной плазмиды.
ДНК-конструкции могут быть трансфицированы в клетки (предпочтительно in vitro) с применением различных физических или химических приемов, включая электропорацию, микроинъекции, бомбардировку микрочастицами, преципитацию с фосфатом кальция, липофекцию или трансфекцию с использованием DEAE-декстрана или полибрена.
Трансфицированные или инфицированные клетки поддерживают в условиях, которые способствуют гомологичной рекомбинации в соответствии с известным в данной области техники (см., например, Capecchi, 1989, Science, 24, 1288-1292). Под “трансфицированной клеткой” понимается клетка, в которую (или в предшественник которой) молекула ДНК была внесена иным, нежели использование вирусного вектора, способом. Под “инфицированной клеткой” понимается клетка, в которую (или в предшественник которой) молекула ДНК или РНК была внесена с использованием вирусного вектора. Вирусами, для которых известно использование в качестве векторов, являются аденовирусы, адено-ассоциированные вирусы, герпесвирусы, вирус свинки, полиовирусы, лентивирус, ретровирусы, вирус Sindbis и вирусы коровьей оспы, такие как птичий поксивирус. Когда претерпевшую гомологичную рекомбинацию клетку выдерживают в условиях, достаточных для обеспечения транскрипции ДНК, регуляторный сегмент, включенный в состав ДНК-конструкции, должен обусловливать изменение транскрипции гена FSHβ.
Претерпевшие гомологичную рекомбинацию клетки (т.е. клетки, в которых произошла желательная гомологичная рекомбинация) могут быть идентифицированы по скринированию их фенотипа или путем тестирования надосадочной фракции культуральной среды методами твердофазного иммуноферментного анализа (ТИФА) на присутствие FSHβ. Промышленные наборы реактивов для детекции FSНβ доступны от фирмы Accurate Chemical & Scientific (Westbury, NY). Претерпевшие гомологичную рекомбинацию клетки также могут быть идентифицированы методами Саузерн- и Нозерн-блоттинга или с применением полимеразной цепной реакции (ПЦР).
По использованию в данном тексте термин “первичные клетки” включает: (1) клетки, присутствующие в суспензии клеток, выделенных из ткани позвоночного животного (перед тем как они будут высеяны, т.е. прикреплены к субстрату для культивирования, такому как чашка или колба); (2) клетки, присутствующие в эксплантате, взятом из ткани; (3) клетки, высеянные в первый раз; и (4) клеточные суспензии, производные от таких высеянных клеток. Первичные клетки также могут являться клетками, которые естественным образом находятся в теле человека или животного.
Вторичными клетками являются клетки на всех последующих стадиях их культивирования. Следовательно, в первый раз, когда высеянные первичные клетки изымают с культурального субстрата и пересевают (пассивируют), по обозначению в данном тексте они становятся вторичными клетками, равно как ими являются и клетки всех последующих пересевов. Штаммы вторичных клеток состоят из вторичных клеток, которые были пересеяны один или большее число раз. Обычно вторичные клетки характеризуются определенным конечным уровнем увеличения размера популяции в культуре и свойством зависимости роста от прикрепления при подавлении роста при контактах (зависимость от прикрепления не относится к тем клеткам, которые содержат в суспензионной культуре). Первичные и вторичные клетки не являются иммортализованными.
Иммортализованными клетками являются линии клеток (в противопоставлении клеточным “штаммам”, когда термин “штамм” относится только к первичным и вторичным клеткам), которые проявляют очевидно не ограниченную продолжительность жизни в культуре.
Клетки, отбираемые для трансфекции или инфекции, могут быть подразделены на 4 группы или категории: (1) клетки, в которых при их анализе белок FSHβ не обнаруживается или не продуцируется вовсе или он имеется или продуцируется не более чем в следовых количествах; (2) клетки, в которых этот белок имеется или продуцируется, но в таком количестве, которое не соответствует желаемому уровню (например, в количестве, которое меньше того уровня, который является физиологически нормальным для анализируемого типа клеток); (3) клетки, в которых этот белок характеризуется уровнем, физиологически нормальным для данного типа клеток, однако имеется необходимость в повышении уровня его содержания или интенсивности его выработки; и (4) клетки, в которых желательным является изменить параметры регуляции или индукции гена, кодирующего данный белок.
Первичные, вторичные и иммортализованные клетки, предназначенные к трансфекции или инфекции с помощью способа по настоящему изобретению, могут быть получены из различных тканей и включают все пригодные клеточные типы, которые можно культивировать. Например, подходящими первичными и вторичными клетками являются фибробласты, кератиноциты, эпителиальные клетки (например, эпителиальные клетки молочных желез, эпителиальные клетки кишечника), эндотелиальные клетки, глиальные клетки, нервные клетки, форменные элементы крови (например, лимфоциты, клетки костного мозга), мышечные клетки и предшественники этих типов соматических клеток. В случае, когда претерпевшие гомологичную рекомбинацию клетки предполагается использовать в генотерапевтических целях, первичные клетки предпочтительно получают от пациента, которому трансфицированные или инфицированные первичные или вторичные клетки должны быть введены. Однако первичные клетки могут быть получены и от донора (т.е. от другого, нежели данный пациент, индивидуума), принадлежащего тому же биологическому виду.
Примерами линий иммортализованных клеток человека, применимых для выработки белка или для генотерапии, являются, тем самым не исчерпываясь, клетки овариальной карциномы 2780AD (Van der Blick et al., 1988, Cancer Res., 48, 5927-5932), A549 (Американская коллекция типовых культур - АТСС; CCL 185), BeWo (ATCC CCL 98), клетки меланомы Боуса (АТТС CRL 9607), CCRF-CEM (АТСС CCL 119), CCRF-HSB-2 (АТСС CCL 120.1), COLO201 (АТСС CCL 224), COLO205 (АТСС CCL 222), COLO320DM (АТСС CCL 220), COLO 320HSR (АТСС CCL 220.1), клетки линии Daudi (АТСС CCL 213), Detroit-562 (ATCC CCL 138), клетки HeLa и производные от клеток HeLa линии (АТСС CCL 2, 2.1 и 2.2), НСТ116 (АТСС CCL 247), клетки HL-60 (АТСС CCL 240), клетки НТ1080 (АТСС CCL 121), IMR-32 (АТСС CCL 127), клетки линии Jurkat (ATCC TIB 152), лейкозные клетки К-562 (АТСС CCL 243), карциномные клетки KB (ATCC CCL 17), KG-1 (АТСС CCL 246), KG-1a (ATCC CCL 246.1), LS123 (АТСС CCL 255), LS17-4T (АТСС CCL CL-188), LS180 (АТСС CCL CL-187), клетки карциномы молочной железы MCF-7 (АТСС ВТН 22), клетки MOLT-4 (АТСС CRL 1582), клетки Namalwa (ATCC CRL 1432), NCI-Н498 (АТСС CCL 254), NCI-H508 (АТСС CCL 253), NCI-H548 (АТСС CCL 249), NCI-H716 (АТСС CCL 251), NCI-H747 (АТСС CCL 252), NCI-H1688 (АТСС CCL 257), NCI-H2126 (АТСС CCL 256), клетки линии Raji (АТСС CCL 86), RD (АТСС CCL 136), RPMI-2650 (АТСС CCL 30), клетки RPMI-8226 (АТСС CCL 155), SNU-C2A (АТСС CCL 250.1), SNU-C2B (АТСС CCL 250), SW-13 (АТСС CCL 105), SW48 (АТСС CCL 231), SW403 (АТСС CCL 230), SW480 (АТСС CCL 227), SW620 (АТСС CCL 227), SW837 (АТСС CCL 235), SW948 (АТСС CCL 237), SW1116 (АТСС CCL 233), SW1417 (АТСС CCL 238), SW1463 (ATCC CCL 234), T84 (ATCC CCL 248), клетки U-937 (ATCC CRL 1593), WiDr (ATCC CCL 218) и сублиния WI38VA13 клеток 2R4 (ATCC CLL 75.1), равно как и гетерогибридомные клетки, образующиеся путем слияния клеток человека и клеток других видов. Могут быть использованы линии вторичных фибробластов человека, такие как WI-38 (ATCC CCL 75) и MRC-5 (ATCC CCL 171). Кроме того, для выработки данного белка in vitro и в генотерапевтических целях могут быть использованы первичные, вторичные или иммортализованные клетки человека, равно как и первичные, вторичные или иммортализованные клетки других видов организмов.
Клетки, экспрессирующие FSHβ
Претерпевшие гомологичную рекомбинацию клетки по настоящему изобретению экспрессируют FSHβ на желаемом уровне и применимы для выработки FSHβ in vitro и в генотерапевтических целях.
Выработка белка
Претерпевшие гомологичную рекомбинацию клетки по настоящему изобретению могут быть использованы для выработки белка FSHβ in vitro. Эти клетки, с учетом известного в данной области техники, поддерживают в таких условиях, которые обеспечивают экспрессию белков. Белок FSHβ может быть очищен из клеточных лизатов или культуральных супернатантов. Фармацевтическая композиция, содержащая белок FSHβ, может быть введена человеку или животному с помощью стандартных фармацевтических путей, известных в данной области техники (например, пероральным, внутривенным, внутримышечным, интраназальным, внутрилегочным, через слизистые, внутрикожным, трансдермальным, ректальным, внутриоболочечным, подкожным, внутрибрюшинным путем или в повреждения). Пероральное введение может потребовать защиты белка от расщепления в пищеварительном тракте, например, с помощью помещения его в полимерные микрокапсулы.
Генотерапия
Претерпевшие гомологичную рекомбинацию клетки по настоящему изобретению применимы в качестве популяций рекомбинантных клеточных линий, в качестве рекомбинантных первичных или вторичных клеток, в качестве рекомбинантных штаммов или линий клональных клеток, в качестве рекомбинантных гетерогенных штаммов или линий клеток и в качестве клеточных смесей, в которых присутствует по крайней мере одна клетка, представляющая четыре указанные ранее категории претерпевших гомологичную рекомбинацию клеток. Такие клетки могут быть использованы для введения при лечении бесплодия, для повышения фертильности у человека или животного или в лечении любых других состояний, восприимчивых к FSHβ.
Претерпевшие гомологичную рекомбинацию первичные клетки, клональные клеточные штаммы или гетерогенные клеточные штаммы вводят пациенту с целью профилактики или лечения у него нежелательного или патологического состояния, используя достаточное количество и подходящий путь введения, с целью экспрессии и прямого предоставления белка или экзогенной ДНК на физиологически приемлемых уровнях. Физиологически приемлемым уровнем является такой уровень, который либо близок к уровню, при котором данный продукт в норме вырабатывается в организме, либо обусловливает улучшение патологического или нежелательного состояния. Если данные клетки являются сингенными данному иммунокомпетентному реципиенту, то они могут быть введены или имплантированы внутривенным, внутриартериальным, подкожным, внутрибрюшинным, внутрисальниковым, внутриоболочечным, внутричерепным или внутримышечным путем или в виде почечных капсул.
Если же клетки не являются сингенными, а реципиент иммунокомпетентен, то претерпевшие гомологичную рекомбинацию клетки, которые предстоит ввести, могут быть включены в барьерные приспособления, имеющие полупроницаемое покрытие. Свойства проницаемости у таких приспособлений таковы, что клетки защищены от выхода из них до момента имплантации в тело пациента, но лечебный белок легко проходит через покрытие и выходит из приспособления, попадая в ограниченное пространство вокруг имплантата или же попадая в циркуляционную систему всего организма: см., например, патенты США №№5641670, 5470731, 5620883 и 5487737 и одновременно поданную патентную заявку на патент США “Доставка терапевтических белков” (изобретатели J.C.Lamsa, D.A.Treco) от 16 апреля 1999 г.: все они приведены здесь для сведения в виде библиографических ссылок. Барьерное приспособление может быть имплантировано в любое подходящее место, например внутрибрюшинно, внутриоболочечно, подкожно, внутримышечно, в виде почечных капсул или в сальник.
В частности, барьерные приспособления применимы для обеспечения имплантации претерпевших гомологичную рекомбинацию иммортализованных клеток, претерпевших гомологичную рекомбинацию ксеногенных клеток (т.е. клеток, происходящих от другого вида организма) или претерпевших гомологичную рекомбинацию аллогенных клеток (т.е. клеток, происходящих от донора, не совместимого по признакам тканевой совместимости) с целью лечения пациента. Такие приспособления сохраняют данные клетки в фиксированном положении in vivo, защищая тем самым эти клетки от активности иммунной системы хозяина. Также барьерные приспособления применимы для целей простого краткосрочного (т.е. непостоянного) лечения за счет возможности легкого изъятия данных клеток тогда, когда по каким-либо показаниям лечебный цикл должен быть остановлен. Также трансфицированные или инфицированные ксеногенные и аллогенные клетки могут быть использованы в отсутствие барьерных приспособлений для краткосрочной генотерапии. В этом случае выработанный этими клетками белок FSHβ будет доставляться in vivo до тех пор, пока эти клетки не окажутся отторгнутыми иммунной системой реципиента.
Различные синтетические, полусинтетические или натуральные фильтрующие мембраны могут быть использованы для данной цели, включая, но тем самым не ограничиваясь, целлюлозу, ацетат целлюлозы, нитроцеллюлозу, полисульфон, поливинилидендифторид, полимеры на основе поливинилхлорида и полимеры на основе производных поливинилхлорида. Барьерные приспособления могут быть использованы в отношении первичных, вторичных или иммортализованных клеток отличающегося вида организма для целей генотерапии в применении к человеку.
Другим типом приспособления, применимого в генотерапии в соответствии с настоящим изобретением, является имплантируемый коллагеновый матрикс, в который погружены данные клетки. Такое приспособление, которое может включать шарики, к которым прикрепляются данные клетки, описано в международной патентной заявке WO 97/15195, которая включена здесь для сведения в виде библиографической ссылки.
Количество клеток, необходимое для конкретной дозы или для имплантата, зависит от различных факторов, включая уровень экспрессии данного белка, размер тела и состояние животного-хозяина и ограничения, накладываемые самой процедурой имплантации. Обычно количество клеток, имплантируемых взрослому человеку или животному со сходным размером тела, находится в диапазоне от 10 тыс. до 50 млрд., а предпочтительно от 100 млн. до 1 млрд. Если желательно, они могут быть имплантированы в нескольких участках тела пациента, причем как одновременно, так и в течение месяцев или лет. При необходимости дозировки могут быть повторены.
Другие варианты
Должно быть понятно, что, хотя настоящее изобретение было представлено в виде его подробного описания, тем не менее предшествующее его описание призвано лишь проиллюстрировать настоящее изобретение, но не в чем-либо ограничить его масштаб, который определяется нижеследующей формулой изобретения.
Другие аспекты, преимущества и модификации находятся в масштабе, определяемом нижеследующей формулой изобретения.













Изобретение относится к области молекулярной генетики. Сущность изобретения состоит в том, что представлена выделенная молекула нуклеиновой кислоты, которая гибридизует в жестких условиях или проявляет 80%-ный уровень идентичности с определенным геномным участком, расположенным выше кодирующего сегмента гена FSHβ, а также ДНК-конструкция, включающая эту молекулу нуклеиновой кислоты в качестве последовательности, являющейся направляющей последовательностью для обеспечения гомологичной рекомбинации. Технический результат - расширение арсенала генетических терапевтических средств. 8 с. и 10 з.п. ф-лы, 12 ил., 1 табл.
| HIRAI el al | |||
| The gene for the beta subunit of porsine PSH..., J | |||
| of Molec | |||
| Endocrinol., 1990, v.5 | |||
| Раздвижной паровозный золотник со скользящими по его скалке поршнями и упорными для них шайбами | 1922 |
|
SU147A1 |
| US 5639640 A, 17.06.1997 | |||
| "Биохимия гормонов и гормональная регуляция” под ред | |||
| Н.Л.Юдаева | |||
| Москва | |||
| "Наука" | |||
| Планшайба для точной расточки лекал и выработок | 1922 |
|
SU1976A1 |

