Предпосылки изобретения
Существующие подходы к лечению заболевания путем введения терапевтических белков включают в себя получение in vitro терапевтических белков для общепринятой фармацевтической доставки (например, посредством внутривенной, подкожной или внутримышечной инъекции) и, более недавно, генной терапии.
Белки терапевтического значения в общем получают путем введения экзогенной ДНК, кодирующей белок, представляющий интерес для терапии, в подходящие клетки. Например, экзогенную ДНК, кодирующую желаемый терапевтический белок, вводят в клетки, такие как иммортализованные ("бессмертные") клетки, в векторе, таком как плазмида, из которого экспрессируется кодируемый белок. Далее было сделано предположение, что эндогенные клеточные гены и их экспрессия могут быть модифицированы генным нацеливанием. См., например, US Patent №5 272071, WO 91/06666, WO 91/06667 и WO 90/11354.
Доступные в настоящее время подходы к генной терапии используют инфекционные векторы, такие как ретровирусные векторы, содержащие генетический материал, который должен быть экспрессирован. Такие подходы имеют ограничения, такие как возможность генерирования компетентного в отношении репликации вируса во время получения вектора; рекомбинация между геномом терапевтического вируса и эндогенным ретровирусным геномом, потенциально генерирующая инфекционные агенты с новой клеточной специфичностью, диапазонами хозяев или увеличенными вирулентностью и цитотоксичностью; независимая интеграция в большие количества клеток, увеличивающая риск онкогенного инсерционного события; лимитированная клонирующая емкость в ретровирусе (которая ограничивает терапевтическую применимость) и непродолжительная экспрессия in vivo целевого продукта. Ценным был бы лучший подход для обеспечения генных продуктов, в частности подход, который не имел бы ограничений и риска, связанных с доступными в настоящее время способами.
Краткое изложение существа изобретения
Данное изобретение касается усовершенствованных способов как получения in vitro терапевтических белков, так и получения и доставки терапевтических белков посредством генной терапии. В данном способе экспрессию желаемого гена-мишени в клетке (т.е. желаемого эндогенного клеточного гена) изменяют введением посредством гомологичной рекомбинации в заранее выбранный сайт клеточного генома ДНК, которая включает в себя по меньшей мере регуляторную последовательность, экзон и донорный сайт сплайсинга. Эти компоненты вводят в хромосомную (геномную) ДНК таким образом, что это приводит в сущности к получению новой транскрипционной единицы (в которой регуляторная последовательность, экзон и донорный сайт сплайсинга, присутствующие в конструкции ДНК, оперативно соединены с эндогенным геном). В результате введения этих компонентов в хромосомную ДНК изменяется экспрессия желаемого эндогенного гена.
Измененная экспрессия гена, в применении этого выражения здесь, включает в себя активацию (или индукцию экспрессии) гена, который в норме является молчащим (неэкспрессируемым) в полученной клетке, увеличенную экспрессию гена, который не экспрессируется при физиологически значимых уровнях, в полученной клетке, изменение характера регуляции или индукции, который отличается от имеющего место в этой клетке, и снижение (в том числе исключение) экспрессии гена, экспрессируемого в этой клетке.
Далее данное изобретение касается конструкций ДНК, применимых в способе изменения экспрессии целевого гена. Эти конструкции ДНК содержат:
(а) направляющую последовательность,
(b) регуляторную последовательность;
(с) экзон и
(d) неспаренный донорный сайт сплайсинга.
Направляющая последовательность в конструкции ДНК направляет интеграцию элементов (а)-(d) в ген-мишень в клетке, так что элементы (b) и (d) оперативно связываются с последовательностями эндогенного гена-мишени.
В другом варианте конструкции ДНК включают в себя:
(а) направляющую последовательность,
(b) регуляторную последовательность,
(с) экзон,
(d) донорный сайт сплайсинта,
(е) интрон и
(f) акцепторный сайт сплайсинга,
причем направляющая последовательность направляет интеграцию элементов (а)-(f) таким образом, что элементы (b)-(f) оперативно связываются с эндогенным геном. Направляющая последовательность гомологична заранее выбранному сайту в клеточной хромосомной ДНК, с которым должна иметь место гомологичная рекомбинация. В этой конструкции экзон обычно находится в 3'-положении от регуляторной последовательности, а донорный сайт сплайсинга находится 3'- от экзогена.
Дальнейшее описание служит для иллюстрации двух вариантов данного изобретения, в которых последовательности, расположенные против хода транскрипции от гена эритропоэтина человека h(ЕРО), изменяют для возможности экспрессии hEPO в первичных, вторичных или иммортализованных клетках, которые не экспрессируют ЕРО в детектируемых количествах в их нетрансфицированном состоянии. В варианте 1 направляющая конструкция содержит две направляющие последовательности. Первая направляющая последовательность гомологична последовательностям 5' второй направляющей последовательности и обе последовательности находятся против хода транскрипции от кодирующего hEPO района. Направляющая конструкция также содержит регуляторный район (промотор mMT-1), экзон (экзон 1 гормона роста человека (hGH) и неспаренный донорный сайт сплайсинга. Продукт гомологичной рекомбинации с этой направляющей конструкцией показан на Фиг.1.
В варианте 2 направляющая конструкция также содержит две направляющие последовательности. Первая направляющая последовательность гомологична последовательностям внутри регуляторного района эндогенного hEPO, а вторая направляющая последовательность гомологична интрону 1 hEPO. Эта направляющая конструкция содержит также регуляторный район (промотор mMT-1), экзон (экзон 1 hGH) и неспаренный донорный сайт сплайсинга. Продукт гомологичной рекомбинации с этой направляющей конструкцией показан на Фиг.2.
В этих двух вариантах продукты событий нацеливания представляют собой химерные транскрипционные единицы, которые генерируют зрелую мРНК, в которой первый экзон гена гормона роста человека (hGH) расположен против хода транскрипции от экзонов 2-5 hEPO. Продукт транскрипции, сплайсинга и трансляции является белком, в котором аминокислотые остатки 1-4 сигнального пептида hEPO заменены аминокислотными остатками 1-3 hGH. Эти два варианта отличаются как по относительным положениям регуляторных последовательностей направляющей конструкции, которые вводятся, так и по специфическому характеру сплайсинга, который должен иметь место для получения конечного процессированного транскрипта.
Далее это изобретение касается способа получения белка in vitro или in vivo посредством введения описанной выше конструкции в хромосомную ДНК клетки хозяина гомологичной рекомбинацией для получения гомологично рекомбинантной клетки. Гомологично рекомбинантная клетка поддерживается затем при условиях, позволяющих транскрипцию, трансляцию и секрецию, что приводит к продуцированию целевого белка.
Данное изобретение касается трансфицированных клеток, таких как трансфицированные первичные или вторичные клетки (т.е. неиммортализованные клетки) и трансфицированные иммортализованные клетки, применимые для получения белков, в частности терапевтических белков, способов получения таких клеток, способов использования этих клеток для получения белков in vitro и способов генной терапии. Клетки данного изобретения являются клетками, происходящими из позвоночных, в частности из млекопитающих, и, более конкретно, из человека. Клетки, полученные по способу данного изобретения, содержат ДНК, кодирующую терапевтический продукт, ДНК, которая сама является терапевтическим продуктом, и/или ДНК, которая заставляет трансфицированные клетки экспрессировать ген на более высоком уровне или с характером регуляции или индукции, который отличается от имеющего место в соответствующей нетрансфицированной клетке.
Данное изобретение касается также способов, при помощи которых клетки, такие как первичные, вторичные и иммортализованные клетки, трансфицируют для включения экзогенного генетического материала, способов получения клональных клеточных штаммов или гетерогенных клеточных штаммов и способов иммунизации животных или получения антител в иммунизированных животных с использованием трансфицированных первичных, вторичных или иммортализованных клеток.
Данное изобретение касается, в частности, способа генного нацеливания или гомологичной рекомбинации в эукариотических клетках, таких как клетки, происходящие из грибов, растения или животного, например позвоночного животного, в частности млекопитающего, и, даже более конкретно, из человека. Т.е. оно касается способа введения ДНК в первичные, вторичные или иммортализованные клетки, происходящие из позвоночных, посредством гомологичной рекомбинации, так что эту ДНК вводят в геномную ДНК первичных, вторичных или иммортализованных клеток в заранее выбранном сайте. Используемые направляющие последовательности выбирают относительно сайта, в который должна быть вставлена ДНК в направляющей конструкций ДНК. Далее данное изобретение касается гомологично рекомбинантных первичных, вторичных или иммортализованных клеток, называемых гомологично рекомбинантными (HR) первичными, вторичными или иммортализованными клетками, полученных данным способом, и применений этих HR, первичных, вторичных или иммортализованных клеток.
В одном варианте данного изобретения, в котором изменяют экспрессию гена, этот ген активируется. То есть ген, присутствующий в первичных, вторичных или иммортализованных клетках, происходящих из позвоночных, который обычно (в норме) не экспрессируется в этих клетках, активируется и в результате экспрессируется кодируемый им белок. В этом варианте гомологичную рекомбинацию используют для вытеснения, выведения из строя или разрушения регуляторного района, обычно ассоциированного с этим геном в клетках, путем введения регуляторной последовательности, которая заставляет ген экспрессироваться при более высоких уровнях, чем уровни в соответствующей нетрансфицированной клетке.
В одном варианте активированный ген может быть амплифицирован включением амплифицируемого селективного маркерного гена, который обладает таким свойством, что клетки, содержащие амплифицированные копии селектируемого маркерного гена, могут отбираться путем культивирования их в присутствии подходящего селектирующего агента. Активированный эндогенный ген амплифицируется в тандеме с амплифицируемым селективным маркерным геном. Клетки, содержащие много копий активированного эндогенного гена, применимы для получения белка in vitro и генной терапии.
Генное нацеливание и амплификация, описанные в данном изобретении, применимы, в частности, для активации экспрессии генов, которые образуют транскрипционные единицы, которые достаточно велики, так что их трудно выделять и экспрессировать, или для активации генов, для которых недоступен или не был клонирован полный кодирующий район белка.
В дальнейшем варианте экспрессия гена, который экспрессируется в клетке как таковой, усиливается или обнаруживает характер регуляции или индукции, который отличается от наблюдаемого в соответствующей нетрансфицированной клетке. В другом варианте экспрессию гена, который экспрессируется в клетке как таковой, уменьшают (т.е. ослабляют или исключают). Данное изобретение описывает также способ, при помощи которого гомологичную рекомбинацию используют для превращения гена в кДНК-копию, не содержащую интронов, для переноса в дрожжевые или бактериальные клетки для получения белка in vitro.
Трансфицированные клетки данного изобретения применимы в ряде приложений в человеке и животных. В одном варианте эти клетки можно имплантировать в человека или в животное для доставки белка человеку или животному. Например, hGH, hEPO, инсулинотропин человека и другие белки могут доставляться системно или локально в больных для терапевтических целей. Кроме того, могут быть получены трансфицированные клетки, не происходящие из человека, продуцирующие гормон роста, эритропоэтин, инсулинотропин и другие белки, имеющие иное происхождение (не из человека).
Для удерживания клеток в фиксированном положении in vivo или для защиты и изолирования этих клеток от иммунной системы хозяина могут быть использованы барьерные устройства. Барьерные устройства, в частности, применимы для имплантации трансфицированных иммортализованных клеток, трансфицированных ксеногенных клеток или трансфицированных аллогенных клеток для лечения патологических состояний человека или животного или для сельскохозяйственнных применений (например, применения бычьего гормона роста для производства молочных продуктов). Барьерные устройства позволяют также проводить кратковременную (т.е. временную) терапию путем обеспечения легкого доступа к клеткам для удаления в случае прекращения режима лечения по той или иной причине. Кроме того, трансфицированные ксеногенные и аллогенные клетки можно использовать в отсутствие барьерных устройств для кратковременной генной терапии, так чтобы продукт гена, продуцируемый клетками, доставлялся in vivo до отторжения этих клеток иммунной системой хозяина.
Трансфицированные клетки данного изобретения применимы также для индуцирования продуцирования антител или для иммунизации людей и животных против патогенных агентов. Имплантированные трансфицированные клетки можно использовать для доставки иммунизирующих антигенов, что приводит к стимуляции клеточного и гуморального иммунного ответа хозяина. Эти иммунные ответные реакции могут предназначаться для защиты хозяина от инфекционных агентов в будущем (т.е. для вакцинации) для стимуляции и повышения способности противостояния инфекции в будущем или для продуцирования антител, направленных против антигена, продуцируемого in vivo трансфицированными клетками, что может быть полезным для терапевтических или диагностических целей. Удаляемые барьерные приспособления, содержащие эти клетки, можно использовать для обеспечения простого средства прекращения экспонирования с антигеном. Альтернативно, использование клеток, которые будут в конце концов отторгнуты (ксеногенных или аллогенных трансфицированных клеток), можно применять для ограничения контакта с антигеном, поскольку продуцирование антигена будет прекращаться при отторжении этих клеток.
Способы данного изобретения можно использоваться для получения первичных, вторичных или иммортализованных клеток, продуцирующих широкое разнообразие терапевтически применимых продуктов, в том числе (но не только): гормонов, цитокинов, антигенов, антител, ферментов, свертывающих кровь факторов, транспортных белков, рецепторов, регуляторных белков, структурных белков, факторов транскрипции, рибозимов или антисмысловых РНК. Дополнительно, способы данного изобретения можно использовать для получения клеток, которые продуцируют не встречающиеся в природе рибозимы, белки или нуклеиновые кислоты, которые применимы для получения in vitro терапевтического продукта или для генной терапии.
Краткое описание чертежей
Фиг.1 является схематической диаграммой стратегии транскрипционной активации гена hEPO; жирные линии, промотор мышиного металлотионеина-1; заштрихованный блок, 5' нетранслируемый район hGH; сплошной блок, экзон 1 hGH; полосатый блок, донорная последовательность 10 п.н. сплайсинга из интрона 1 hEPO; перекрестно заштрихованный блок, 5'-нетранслируемый район hEPO; многочисленные незаштрихованные (открытые) блоки, кодирующие hEPO последовательности; блок с диагональными полосами, 3'-нетранслируемые последовательности hEPO; HIII, сайт Hind III.
Фиг.2 является схемой стратегии транскрипционной активации гена hEPO; жирные линии, промотор мышиного металлотионеина-1; заштрихованный блок, 5'-нетранслируемый район hGH; сплошной блок, экзон 1 hGH; многочисленные незаштрихованные блоки, кодирующие hEPO последовательности; блок с диагональными полосами, 3'-нетраслируемые последовательности hEPO; HIII, Hind III сайт.
Фиг.3 является схематическим представлением плазмиды pXGH5, которая включает в себя ген hGH под контролем мышиного промотора металлотионеина.
Фиг.4 является схематическим представлением плазмиды рЕ3 neoERO. Указаны положения гена эритропоэтина человека и гена неомицинфосфотрансферазы (neo) и генов устойчивости к ампициллину (amp). Стрелки указывают направления транскрипции различных генов. pm МТ1 обозначает промотор мышиного маталлотионеина (регулирующий экспрессию hEPO) и рТК обозначает промотор тимидинкиназы вируса простого герпеса (регулирующий экспрессию neo). Покрытые точками районы карты отмечают положения последовательностей, происходящих из локуса гипоксантин-гуанин-фосфорибозилтрансферазы (HPRT) человека. Указаны относительные положения сайтов узнавания рестриктаз.
Фиг.5 является схематическим изображением плазмиды рс DNEO, которая включает в себя кодирующий neo район (фрагмент Bam HI-Bgl II) из плазмиды рSV2neo, вставленный в сайт BamHI плазмиды pcD; последовательности Amp-R и pBR3220 ri из pBR322; и полиА, границы сплайсинга 16S и районы раннего промотора из SV 40.
Фиг.6 является схематическим изображением плазмиды рRЕРO4.
Фиг.7 является графическим изображением экспрессии эритропоэтина в клеточной линии-мишени человека, подвергнутой ступенчатому отбору в 0,02, 0,05, 0,1, 0,2 и 0,4 мкМ метотрексате.
Фиг.8 является схематическим изображением плазмиды pREPO15. Фрагменты, полученные из геномных последовательностей hEPO, показаны заштрихованными блоками. Район между Bam HI (3537) и Bgl II /Hind III соответствует последовательностям при положениях 1-4008 во вводе HUMERPALU Genbank. Район между Bql II/Hind III (11463) соответствует последовательностям ДНК в положениях 4009-5169 во вводе HUMERPALU Genbank. Район между Hind III (11463) и XhoI (624) содержит последовательность, соответствующую положениям 7-624 Genbank ввода HUMERPA. Последовательности промотора CMV показаны в виде открытого блока и содержат последовательность из нуклеотидов 546-2105 последовательности HS5М1ЕР Genbank. Ген neo показан в виде открытого блока стрелкой. Промотор тимидинкиназы (tk), регулирующий ген neo, показан в виде заштрихованного блока. Последовательности рВ S II S K+, включающие в себя ген amp, указаны тонкой линией.
Фиг.9А представляет рестрикционные карты и схематические изображения продуктов, наблюдаемых при расщеплении эндогенного гена hEPO (сверху) и активированного гена hEPO после гомологичной рекомбинации нацеливающим фрагментом из pREPO15 (внизу).
Фиг.9В представляет результаты рестрикционного анализа и гибридизационного анализа по Саузерну необработанного (HF) и активированного (Т1) клона фибробластов человека НF342-15 (см. Пример 7).
Фиг.10 является схематическим изображением плазмиды pREPO18. Заштрихованными блоками показаны фрагменты, произведенные из геномных последовательностей hEPO. Район между BamHI (3537) и ClaI (7554) соответствует последовательностям в положениях 1-4008 в GEnbank вводе HUMERPALU. Район между ATG (12246) и Hind III (13426) соответствует последовательности ДНК в положениях 4009-5169 в Genbank вводе HUMERPALU. Район между Hind III (13426) и XhoI (624) содержит последовательность, соответствующую положениям 7-624 Genbank ввода HUMERPA. Последовательности промотора CMV показаны в виде незаштрихованного (открытого) блока и содержат последовательность из нуклеотидов 546-2015 Genbank последовательности HS 5М1ЕР. Единица транскрипции дигидрофолатредуктазы (dhfr) показана в виде заштрихованного блока стрелкой. Ген neo показан в виде открытого блока стрелкой. Промотор tk, регулирующий ген neo, показан в виде заштрихованного блока. Последовательности pBS II S К+, включающие в себя ген amp, указаны тонкой линией.
Фиг.11 является схематической иллюстрацией конструкции изобретения для активации и амплификации не содержащего интронов гена, гена α-интерферона, содержащей первую направляющую последовательность (1), амплифицируемый маркерный ген (AM), селектируемый маркерный ген (SM), регуляторную последовательность, сайт CAP, донорный сайт сплайсинга (SD), интрон (тонкие линии), акцепторный сайт сплайсинга (SA) и вторую направляющую последовательность (2). Черный блок обозначает кодирующую ДНК, а заштрихованные блоки обозначают нетранслируемые последовательности.
Фиг.12 является схематической иллюстрацией конструкции изобретения для активации и амплификации эндогенного гена, в котором первый экзон содействует образованию сигнального пептида, гена колониестимулирующего фактора гранулоцитов-макрофагов (GM-CSF), причем эта конструкция содержит первую направляющую последовательность (1), амплифицируемый маркерный ген (AM), селектируемый маркерный ген (SM), регуляторную последовательность, сайт CAP, донорный сайт сплайсинга (SD) и вторую направляющую последовательность (2). Черные блоки обозначают кодирующую ДНК и заштрихованные блоки обозначают нетранслируемые последовательности.
Фиг.13 является схематической иллюстрацией конструкции изобретения для активации и амплификации эндогенного гена, в котором первый экзон дает сигнальный пептид, гена колониестимулирующего фактора гранулоцитов человека, причем эта конструкция содержит первую направляющую последовательность (1), амплифицируемый маркерный ген (AM), селектируемый маркерный ген (SM), регуляторную последовательность, сайт CAP, донорный сайт сплайсинга (SD) и вторую направляющую последовательность (2). Черные блоки обозначают кодирующую ДНК, а заштрихованные блоки обозначают нетранслируемые последовательности.
Фиг.14 является схематической иллюстрацией конструкции изобретения для активации и амплификации эндогенного гена, в котором первый экзон является некодирующим, гена FSHβ, причем эта конструкция содержит первую направляющую последовательность (1), амплифицируемый маркерный ген (AM), селектируемый маркерный ген (SM), регуляторную последовательность, сайт CAP, донорный сайт сплайсинга (SD) и вторую направляющую последовательность (2). Черные блоки обозначают кодирующую ДНК и заштрихованные блоки обозначают нетранслируемые последовательности.
Подробное описание изобретения
Данное изобретение основано на открытии, что регуляция или активность эндогенных представляющих интерес генов в клетке может быть изменена встраиванием в клеточный геном в заранее заданном сайте посредством гомологичной рекомбинации конструкций ДНК, содержащих:
(а) направляющую последовательность;
(b) регуляторную последовательность;
(с) экзон и
(d) неспаренный донорный сайт сплайсинга,
в которых направляющая последовательность направляет интеграцию элементов (а)-(d) таким образом, что элементы (b)-(d) оперативно соединяются с эндогенным геном.
В другом варианте эти конструкции ДНК содержат:
(а) направляющую последовательность,
(b) регуляторную последовательность,
(c) экзон,
(d) донорный сайт сплайсинга,
(e) интрон и
(f) акцепторный сайт сплайсинга,
причем направляющая последовательность направляет интеграцию элементов (а)-(f) таким образом, что элементы (b) и (f) оперативно соединяются с первым экзоном эндогенного гена. Используемые направляющие последовательности выбирают относительно сайта, в который должна быть вставлена эта ДНК. В обоих вариантах используют событие нацеливания для создания новой транскрипционной единицы, которая представляет собой слитый продукт последовательностей, введенных направляющими конструкциями дНК, и эндогенного клеточного гена. Как обсуждалось здесь, например, образование новой транскрипционной единицы позволяет активировать транскрипционно молчащие гены (гены, не экспрессируемые в клетке перед трансфекцией) в клетках хозяина путем введения в геномную ДНК клетки хозяина конструкций ДНК данного изобретения. Как также обсуждалось здесь, экспрессия эндогенного гена, который экспрессируется в клетки как таковой, может быть изменена таким образом, что она увеличивается, уменьшается, в том числе устраняется, или характер регуляции или индукции может быть изменен через использование этого способа и конструкций ДНК данного изобретения.
Данное изобретение, как описано выше, касается способа генного или дНК-нацеливания в клетках эукариотического происхождения, таких как клетки, происходящие из грибов, растения или животного, такого как позвоночного животного, в частности, млекопитающего, и, даже более конкретно, человека. То есть оно касается способа введения ДНК в клетку, такую как первичные, вторичные или иммортализованные клетки, происходящие из позвоночных, посредством гомологичной рекомбинации или нацеливания этой ДНК, которую вводят в геномную ДНК клеток в заранее выбранном сайте. Оно касается, в частности, гомологичной рекомбинации, в которой продукты транскрипции и/или трансляции эндогенных генов модифицируют путем использования конструкций ДНК, содержащих направляющую последовательность, регуляторную последовательность, экзон и донорный сайт сплайсинга. Далее данное изобретение касается гомологично рекомбинантных клеток, получаемых данным способом, и применений гомологично рекомбинантных клеток.
Данное изобретение касается также способа активации гена, присутствующего в первичных клетках, вторичных клетках или иммортализованных клетках позвоночных животных, но обычно не экспрессируемого в этих клетках. Гомологичную рекомбинацию или нацеливание используют для введения в геном клетки последовательностей, которые заставляют ген экспрессироваться в реципиентных клетках. В следующем варианте усиливается экспрессия гена в клетке или изменяется характер регуляции или индукции гена посредством введения такой конструкции ДНК. В результате кодируемый продукт экспрессируется на более высоких уровнях, чем в соответствующей нетрансфицированной клетке. Данный способ и конструкции ДНК применимы также для получения клеток, в которых экспрессия желаемого продукта ниже в трансфицированной клетке, чем в соответствующей нетрансфицированной клетке. То есть в трансфицированной клетке продуцируется меньше белка (или вообще не продуцируется этот белок), чем в нетрансфицированных клетках.
В другом варианте обычно молчащий ген, кодирующий желаемый продукт, активируется в трансфицированных первичных, вторичных или иммортализованных клетках и амплифицируется. Этот вариант представляет собой способ введения гомологичной рекомбинацией с геномной ДНК последовательностей ДНК, которые обычно не соединены функционально с эндогенным геном и которые
(1) при введении в геном хозяина при эндогенном гене или вблизи от него изменяют (например, активируют) экспрессию эндогенного гена и, кроме того,
(2) делают возможным отбор клеток, в которых амплифицируется активированный эндогенный ген. Альтернативно, экспрессия гена, обычно экспрессируемого в клетке как таковой, усиливается и ген амплифицируется.
Далее следует описание конструкций ДНК данного изобретения, способов, в которых они применяются для получения трансфицированных клеток, трансфицированные клетки и применения этих клеток.
Конструкция ДНК
Конструкция ДНК данного изобретения включает в себя по меньшей мере следующие компоненты: направляющую последовательность; регуляторную последовательность; экзон и неспаренный донорный сайт сплайсинга. В этой конструкции экзон находится в 3'-положении от регуляторной последовательности и неспаренный донорный сайт сплайсинга находится 3' от экзона. Кроме того, могут присутствовать множественные экзоны и/или интроны перед этим экзоном (5' от него), фланкированным неспаренным донорным сайтом сплайсинга. Как описано здесь, часто присутствуют дополнительные компоненты конструкции, такие как селектируемые маркеры или амплифицируемые маркеры.
Находящаяся в конструкции ДНК может быть названа экзогенной. Термин "экзогенная" определяется здесь как ДНК, которую вводят в клетку по способу данного изобретения, например, с конструкциями ДНК, описанными здесь. Экзогенная ДНК может иметь последовательности, идентичные эндогенным последовательностям ДНК, присутствующим в клетке перед трансфекцией, или отличающиеся от них.
Направляющая последовательность или направляющие последовательности
Направляющими последовательностями или последовательностями являются последовательности ДНК, которые делают возможной приемлемую гомологичную рекомбинацию в геном выбранной клетки, содержащей представляющий интерес ген. Направляющими последовательностями являются, в общем, последовательности ДНК, которые гомологичны (т.е. идентичны или достаточно близки клеточной ДНК, так что направляющая последовательность и клеточная ДНК могут подвергаться гомологичной рекомбинации) последовательностям ДНК, обычно присутствующим в геноме самих клеток (например, кодирующей или некодирующей ДНК, лежащим против хода транскрипции от сайта инициации транскрипции, внутри или по ходу транскрипции от сайта терминации транскрипции представляющего интерес гена, или последовательностям, присутствующим в геноме вследствие предшествующей модификации). Используемые направляющие последовательность или последовательности выбирают с учетом сайта, в который должна быть вставлена ДНК в конструкции ДНК.
Могут быть использованы одна или более направляющих последовательностей. Например, в кольцевой плазмиде или во фрагменте ДНК предпочтительно используют одну направляющую последовательность. В линейной плазмиде или ее фрагменте ДНК предпочтительно используют две направляющие последовательности. Направляющие последовательности или последовательности могут, независимо одна от другой, находиться внутри представляющего интерес гена (например, последовательности экзона и/или интрона), непосредственно рядом с представляющим интерес геном (т.е. без дополнительных нуклеотидов между направляющей последовательностью и кодирующим районом представляющего интерес гена), против хода транскрипции от представляющего интерес гена (например, последовательности находящегося против хода транскрипции некодирующего района или эндогенные промоторные последовательности) или против хода транскрипции от представляющего интерес гена и на расстоянии от него (например, последовательности, находящиеся против хода транскрипции от эндогенного промотора). Направляющие последовательность или последовательности могут включать в себя районы направляемого гена, известные или секвенированные в настоящее время, и/или районы, расположенные далее против хода транскрипции, которые структурно не охарактеризованы, но могут быть картированы при помощи рестриктаз и определены специалистами в этой области.
Как описано здесь, генное нацеливание можно использовать для введения регуляторной последовательности, выделенной от отличающегося гена, составленной из компонентов, выделенных из различных клеточных и/или вирусных источников или синтезированной в виде новой регуляторной последовательности методами генной инженерии, внутрь, непосредственно рядом с эндогенным клеточным геном, против хода транскрипции от него или на значительном расстоянии от эндогенного клеточного гена. Альтернативно или дополнительно, последовательности, влияющие на структуру или стабильность продуцируемых РНК или белка, могут быть заменены, удалены, добавлены или иным образом модифицированы посредством нацеливания. Например, элементы стабильности РНК, сайты сплайсинга и/или лидерные последовательности молекул РНК могут быть модифицированы для улучшения или изменения функции, стабильности и/или транслируемости молекулы РНК. Могут быть также изменены белковые последовательности, такие как сигнальные последовательности, пропептидные последовательности, активные сайты и/или структурные последовательности для усиления или модификации транспорта, секреции или функциональных свойств белка. В соответствии с этим способом введение экзогенной ДНК приводит к изменению свойств обычной экспрессии гена и/или структурных свойств белка или РНК.
Регуляторная последовательность
Регуляторная последовательность конструкции ДНК может быть составлена из одного или более промоторов (таких как конститутивный или индуцируемый промотор), энхансеров, районов присоединения "лесов" или сайтов присоединения матрикса, отрицательных регуляторных элементов, сайтов связывания факторов транскрипции или комбинаций этих последовательностей.
Регуляторная последовательность может содержать индуцируемый промотор, что приводит к результату, что полученные клетки или введенные в индивидуума клетки не экспрессируют продукт, но могут быть индуцированы для его экспрессии (т.е. экспрессия индуцируется после получения трансфицированных клеток, но перед имплантацией или после имплантации). ДНК, кодирующую желаемый продукт, можно конечно вводить в клетки таким образом, что она экспрессируется при введении (например, при конститутивном промоторе). Регуляторная последовательность может быть выделена из клеточного или вирусного геномов (такие регуляторные последовательности включают в себя последовательности, которые регулируют экспрессию ранних или поздних генов SV 40, основных поздних генов аденовируса, мышиного гена металлотионеина-1-, гена фактора-1α элонгации, генов цитомегаловируса, генов коллагена, генов актина, генов иммуноглобулина или гена HMG-СоА-редуктазы). Регуляторная последовательность предпочтительно содержит сайты связывания факторов транскрипции, таких как ТАТА-бокс, ССААТ-бокс, сайты связывания AP1, Sp1 или HF-кВ.
Дополнительные элементы конструкции ДНК
Конструкция ДНК дополнительно содержит один или более экзонов. Экзон определяется здесь как последовательность ДНК, которая копируется в РНК и присутствует в зрелой молекуле мРНК. Экзоны могут, необязательно, содержать ДНК, которая кодирует одну или более аминокислот и/или частично кодирует аминокислоту (т.е. имеет одно или два основания кодона). Альтернативно, экзон содержит ДНК, которая соответствует 5'-некодирующей области. В случае, когда экзогенный экзон или экзоны кодируют одну или более аминокислот и/или часть аминокислоты, конструкция ДНК конструируется таким образом, чтобы при транскрипции и сплайсинге рамки считывания были внутри рамки считывания со вторым экзоном или кодирующим районом нацеленного гена. В применении здесь выражение "внутри рамки считывания" означает, что кодирующие последовательности первого экзона и второго экзона при слиянии соединяют вместе нуклеотиды таким образом, чтобы не изменять рамку считывания части мРНК, произведенной из второго зкзона.
В случае, когда первый экзон гена-мишени содержит последовательность ATG для инициации трансляции, экзогенный экзон конструкции предпочтительно содержит ATG и, если необходимо, один или более нуклеотидов, так что полученный кодирующий район мРНК, включающий в себя второй и последующие экзоны гена-мишени, находится внутри рамки считывания. Примеры таких генов-мишеней, в которых первый экзон содержит ATG, включают в себя гены, кодирующие hEPO, hGH, колониестимулирующий фактор гранулоцитов/макрофагов человека (hGM-CSF) и гранулоцитарный колониестимулирующий фактор человека (hG-CSF).
Донорный сайт сплайсинга является последовательностью, которая регулирует сплайсинг одного экзона с другим экзоном. В типичном случае первый экзон лежит 5' от второго экзона и донорный (левый) сайт сплайсинга, перекрывающий и фланкирующий первый экзон на его 3'-стороне, узнает акцепторный (правый) сайт сплайсинга, фланкирующий второй экзон на 5'-стороне второго экзона. Донорные сайты сплайсинга имеют характерную консенсусную последовательность, представленную как: (A/C) AGGU RAGU (где R обозначает пуриновый нуклеотид) с GU в четвертом и пятом положениях, что является необходимым (Jackson, I.J., Nucleic Acids Research, 19: 3715-3798 (1991)). Первые три основания донорного консенсусного сайта сплайсинга являются последними тремя основаниями экзона. Донорные сайты сплайсинга функционально определяются по их способности осуществлять соответствующую реакцию в пути сплайсинга мРНК.
Неспаренный донорный сайт сплайсинга определяется здесь как донорный сайт сплайсинга, который присутствует в направляющей конструкции и не сопровождается в этой конструкции акцепторным сайтом сплайсинга, расположенным 3' по отношению к неспаренному донорному сайту сплайсинга. Этот неспаренный донорный сайт сплайсинга приводит к сплайсингу до эндогенного акцепторного сайта сплайсинга.
Акцепторный сайт сплайсинга является последовательностью, которая, подобно донорному сайту сплайсинга, регулирует сплайсинг одного экзона с другим экзоном. Действуя вместе с донорным сайтом сплайсинга, механизм сплайсинга использует акцепторный сайт сплайсинга для осуществления удаления интрона. Акцепторные сайты сплайсинга имеют характерную последовательность, представленную как: YYYYYYYYYYNyAG, где Y обозначает любой пиримидин и N обозначает любой нуклеотид (Jackson, I.J., Nucleic Acids Research 19:3715-3798 (1991)).
Интрон определяется как последовательность из одного или более нуклеотидов, лежащая между двумя экзонами и удаляемая сплайсингом из молекулы-предшественника РНК при образовании молекулы мРНК.
Регуляторная последовательность, например, оперативно соединена с инициирующим кодоном ATG, который инициирует трансляцию. Необязательно сайт CAP (специфический инициирующий сайт мРНК, который связан с регуляторным районом и используется им) оперативно соединен с регуляторной последовательностью и инициирующим кодоном ATG. Альтернативно, сайт CAP, соединенный с регуляторной последовательностью и используемый ею, не включается в направляющую конструкцию и аппарат транскрипции будет определять новый сайт CAP. Для большинства генов сайт CAP обычно обнаруживается приблизительно как 25 нуклеотидов 3' от ТАТА-бокса. В одном варианте донорный сайт сплайсинга расположен непосредственно рядом с ATG, например, когда присутствие одного или более нуклеотидов не требуется, для того, чтобы экзогенный экзон находился внутри рамки со вторым экзоном нацеленного гена. Предпочтительно ДНК, кодирующая одну или более аминокислот или части аминокислоты внутри рамки считывания с кодирующей последовательностью нацеленного гена, расположена непосредственно рядом с ATG на его 3'-стороне. В таком варианте донорный сайт сплайсинга расположен непосредственно рядом с кодирующей ДНК на ее 3'-стороне.
Выражения "оперативно соединенные" или "функционально расположенные" определяются как конфигурация, в которой экзогенная регуляторная последовательность, экзон, донорный сайт сплайсинга и, необязательно, последовательность и акцепторный сайт сплайсинга подходящим образом ориентированы в положении по отношению к эндогенному гену таким образом, что регуляторный элемент регулирует образование первичного РНК-транскрипта, который инициируется при сайте CAP (необязательно включенном в направляющую конструкцию) и включает в себя последовательности, соответствующие экзону и донорному сайту сплайсинга направляющей конструкции, ДНК, лежащую против хода транскрипции от регуляторного района эндогенного гена (если он присутствует ), регуляторный район эндогенного гена (если он присутствует), нетранскрибируемый 5'-район эндогенного гена (если он присутствует) и экзоны и интроны (если они присутствуют) эндогенного гена. В оперативно соединенной конфигурации донорный сайт сплайсинга направляющей конструкции регулирует событие сплайсинга до акцепторного сайта сплайсинга, фланкирующего один из экзонов эндогенного гена, так что желаемый белок может быть образован из полностью сплайсированного зрелого транскрипта. В одном варианте акцепторный сайт сплайсинга является эндогенным, так что сплайсинг регулируется до эндогенного экзона, например эндогенного гена. В другом варианте, когда акцепторный сайт сплайсинга включен в направляющую конструкцию, событие сплайсинга удаляет интрон, введенный направляющей конструкцией.
Используемая кодирующая ДНК (например, в экзоне 1 направляющей конструкции) может необязательно кодировать одну или несколько аминокислот и/или часть аминокислоты, которые одинаковы с аминокислотами эндогенного белка. Используемая в этом изобретении кодирующая последовательность ДНК может, например, соответствовать первому экзону представляющего интерес гена. Альтернативно, кодирующая ДНК может кодировать одну или более аминокислот или часть аминокислоты, отличающихся от первого экзона представляющего интерес белка. Такой вариант представляет особый интерес, когда аминокислоты первого экзона представляющего интерес белка не являются решающими для активности или активностей этого белка. Например, при конструировании слитых конструкций с эндогенным геном hEPO можно использовать последовательности, кодирующие первый экзон hGH. В этом примере слияния экзона 1 hGH с экзоном 2 hEPO приводит к образованию гибридного сигнального пептида, который является функциональным. В родственных конструкциях эта техника может быть применена также для коррекции мутации, обнаруженной в гене-мишени.
В случае, когда желаемым продуктом является слитый белок эндогенного белка и продукта кодирующих последовательностей в направляющей конструкции, экзогенная кодирующая ДНК, включенная в клетки согласно данному способу, включает в себя ДНК, которая кодирует один или более экзонов или последовательность кДНК, соответствующую продукту трансляции или транскрипции, который должен быть слит с продуктом эндогенного гена мишени. В этом варианте нацеливание используют для получения химерных или многофункциональных белков, которые объединяют структурные, ферментативные или лиганд- или рецепторсвязывающие свойства двух или более белков в один полипептид. Например, экзогенная ДНК может кодировать "якорь" для мембраны для белка-мишени или сигнальный пептид для обеспечения или улучшения клеточной секреции, лидерные последовательности, ферментативные районы, районы трансмембранного домена, связывающие кофактор районы или другие функциональные районы. Примеры белков, которые обычно не секретируются, но которые могли бы быть слиты с сигнальным белком для обеспечения секреции, включают в себя допадекарбоксилазу, транскрипционные регуляторные белки, β-галактозидазу и тирозингидроксилазу.
В случае, когда первый экзон гена-мишени соответствует некодирующему району (например, первый экзон гена фолликулостимулирующего гормона бета (FSHβ)), экзогенный ATG не является необходимым и, предпочтительно, отсутствует.
ДНК этой конструкции может быть получена из источников, в которых она присутствует в природе, или она может быть получена способами генной инженерии или синтетическими способами.
Ген-мишень и образующийся продукт
Конструкция ДНК при трансфекции в клетки, такие как первичные, вторичные или иммортализованные клетки, может регулировать экспрессию желаемого продукта, например активной или функциональной части белка или РНК. Этот продукт может представлять собой, например, гормон, цитокин, антиген, антитело, фермент, фактор свертывающей системы крови, транспортный белок, рецептор, реугляторный белок, структурный белок, фактор транскрипции, антисмысловую РНК или рибозим. Дополнительно этот продукт может быть белком или нуклеиновой кислотой, не встречающейся в природе (т.е. слитым белком или слитой нуклеиновой кислотой).
Описанный в этом изобретении способ может производить один или более терапевтических продуктов, таких как эритропоэтин, кальцитонин, гормон роста, инсулин, инсулинотропин, инсулин-подобные факторы роста, паратиреоидный гормон, интерферон β и интерферон γ, факторы роста нерва, фолликулостимулирующий гормон (FSHβ), TGF-β, фактор некроза опухоли, глюкагон, фактор роста костей 2, фактор роста костей 7, тиреотропный гормон бета (TSH-β), интерлейкин 1, интерлейкин 2, интерлейкин 3, интерлейкин 6, интерлейкин 11, интерлейкин 12, гранулоцитарный колониестимулирующий фактор, макрофагальный колониестимулирующий фактор, гранулоцитарный/макрофагальный колониестимулирующий фактор, иммуноглобулины, каталитические антитела, протеинкиназа С, глюкоцереброзидаза, супероксиддисмутаза, тканевый активатор плазминогена, урокиназа, антитромбин III, ДНКаза, α-галактозидаза, тирозингидроксилаза, факторы свертывания крови V, фактор свертывания крови VII, фактор свертывания крови VIII, фактор свертывания крови IX, фактор свертывания крови X, фактор свертывания крови XIII, аполипопротеин Е или аполипопротеин А-1, глобины, рецептор липопротеина низкой плотности, рецептор IL-2, антагонисты IL-2, альфа-1-антитрипсин, модификаторы иммунного ответа и растворимый CD4.
Селектируемые маркеры и амплификация
Обнаружение нацеливающего события может быть облегчено путем использования одного или более селектируемых маркерных генов. Эти маркеры могут быть включены в направляющую конструкцию или присутствовать на разных конструкциях. Селектируемые маркеры могут быть разделены на две категории: позитивно селектируемые и негативно селектируемые (другими словами, маркеры либо для позитивного, либо для негативного отбора). В позитивном отборе клетки, экспрессирующие позитивно селектируемый маркер, способны выживать при обработке селективным (отбирающим) агентом (при таких маркерах, как neo, ксантин-гуанин-фосфорибозилтрансфераза (gpt), аденозиндезаминаза (ada), пуромицин (рас), гигромицин (hyg), CAD, кодирующий карбамилфосфатсинтазу, аспартаттранскарбамилазу и дигидрооротазу, глутаминсинтеза (GS), устойчивость к множественным лекарственным средствам 1 (mdrf) и гистидин D (his D), что позволяет отбирать клетки, в которых направляющая конструкция интегрирована в геном клетки-хозяина. В негативном отборе клетки, экспрессирующие негативно селектируемый маркер, разрушаются в присутствии отбирающего агента. Идентификация события нацеливания (попадания в мишень) может облегчаться путем использования одного или более маркерных генов, проявляющих свойство негативного отбора, так что негативно селектируемый маркер соединен с экзогенной ДНК, но имеет такую конфигурацию, что негативно селектируемый маркер фланкирует направляющую последовательность и точная гомологичная рекомбинация с последовательностями в геноме клетки-хозяине не приводит к стабильной интеграции негативно селектируемого маркера (Mansour, S.L. et al., Nature 336:348-352 (1988)). Маркеры, применимые для этой цели, включают в себя ген тимидинкиназы вируса простого герпеса (ТК) или бактериальный gpt ген.
Множество селектируемых маркеров можно включать в первичные, вторичные или иммортализованные клетки. Например, можно использовать селектируемый маркер, который придает селектируемый фенотип, такой как устойчивость к лекарствам, пищевую ауксотрофию, устойчивость к цитотоксическому агенту или экспрессия поверхностного белка. Селектируемыми маркерными генами, которые можно использовать, являются neo, gpt, ada, dhfr, pac, hyd, CAD, GS, mdrf и hisD, Приобретенный селектируемый фенотип позволяет идентифицировать и выделить реципиентные клетки.
Амплифицируемые гены, кодирующие селектируемые маркеры (например, ada, GS, dhfr и многофункциональный ген CAD) имеют дополнительное свойство, заключающееся в том, что они позволяют отбор клеток, содержащих амплифицированные копии селектируемого маркера, введенного в геном. Эта особенность обеспечивает механизм значительного увеличения копийности смежного или связанного гена, для которого желательна амплификация. Можно также использовать мутированные версии этих последовательностей, обнаруживающие улучшенные селектируемые свойства, и другие амплифицируемые последовательности.
Порядок компонентов в конструкции ДНК может вариировать. Если конструкция является кольцевой плазмидой, порядок элементов в полученной структуре может быть следующим: направляющая последовательность - плазмидная ДНК (состоящая из последовательностей для отбора и/или репликации направляющей плазмиды в микробном или ином подходящем хозяине) - селектируемый маркер (маркеры) - регуляторная последовательность - экзон - донорный сайт сплайсинга. Предпочтительно плазмида, содержащая направляющую последовательность и экзогенные элементы ДНК, расщепляется рестриктазой, которая разрезает один или несколько раз внутри направляющей последовательности для образования линейной или содержащей пробелы (бреши) молекулы перед введением в реципиентную клетку, так что свободные концы ДНК увеличивают частоту событий гомологичной рекомбинации, как описано в этом изобретении. Кроме того, свободные концы ДНК могут быть обработаны экзонуклеазой для создания выступающих 5'- или 3'-одноцепочечных концов ДНК для увеличения частоты желаемого события гомологичной рекомбинации. В этом варианте гомологичная рекомбинация между направляющей последовательностью и клеточной мишенью будет приводить к двум копиям направляющих последовательностей, фланкирующих элементы, содержащиеся внутри введенной плазмиды.
Если конструкция является линейной, порядок может быть следующим: первая направляющая последовательность - селектируемый маркер - регуляторная последовательность - экзон - донорный сайт сплайсинга - вторая направляющая последовательность или, альтернативно, первая направляющая последовательность - регуляторная последовательность - экзон - донорный сайт сплайсинга - ДНК, кодирующая селектируемый маркер - вторая направляющая последовательность. Клетки, которые стабильно интегрируют эту конструкцию, будут выдерживать обработку отбирающим агентом; субпопуляция стабильно трансфицированных клеток будет представлять собой гомологично рекомбинированные клетки. Гомологично рекомбинированные клетки могут быть идентифицированы разнообразными способами, в том числе ПЦР (PCR), гибридизацией по Саузерну и фенотипическим скринингом.
В другом варианте порядок элементов конструкции может быть следующим: первая направляющая последовательность - селектируемый маркер - регуляторная последовательность - экзондонорный сайт сплайсинга - интрон - акцепторный сайт сплайсинга - вторая направляющая последовательность.
Аальтернативно, порядок компонентов в конструкции ДНК может быть следующим: первая направляющая последовательность - селектируемый маркер 1 - регуляторная последовательность - экзон - донорный сайт сплайсинга - вторая направляющая последовательность - селектируемый маркер 2 или, альтернативно, первая направляющая последовательность - регуляторная последовательность - экзон - донорный сайт сплайсинга - селектируемый маркер 1 - вторая направляющая последовательность - селектируемый маркер 2. В этом варианте селектируемый маркер 2 обнаруживает свойство негативного отбора. Т.е. продукт гена селектируемого маркера 2 может быть отобран при росте в подходящей среде, содержащей агент (обычно лекарственное средство или аналог метаболита), который убивает клетки, экспрессирующие селектируемый маркер 2. Рекомбинация между направляющими последовательностями, фланкирующими селектируемый маркер 1, с гомологичными последовательностями в геноме клетки-хозяина приводит к нацеленной интеграции селектируемого маркера 1, тогда как селектируемый маркер 2 не интегрируется. Такая рекомбинация генерирует клетки, которые стабильно трансфицированы селектируемым маркером 1, но нестабильно трансфицированы селектируемым маркером 2, и такие клетки могут отбираться по росту в средах, содержащих селективный (отбирающий) агент, который отбирает по селектируемому маркеру 1, и селективный агент, который отбирает против селектируемого маркера 2.
Конструкция ДНК может также включать в себя позитивно селектируемый маркер, который позволяет отбирать клетки, содержащие амплифицированные копии этого маркера. Амплификация такого маркера приводит к совместной амплификации фланкирующих последовательностей ДНК. В этом варианте порядок компонентов конструкции является, например, следующим: первая направляющая последовательность - амплифицируемый позитивно селектируемый маркер - второй селектируемый (необязательный) маркер - регуляторная последовательность - экзон - донорный сайт сплайсинга - вторая направляющая последовательность ДНК.
В этом варианте активированный ген может дополнительно амплифицироваться включением селектируемого маркерного гена, который обладает таким свойством, что клетки, содержащие амплифицированные копии селектируемого маркерного гена, могут отбираться путем культивирования этих клеток в присутствии подходящего селектирующего агента. Активированный эндогенный ген будет амплифицироваться в тандеме с амплифицируемым селектируемым маркерным геном. Клетки, содержащие много копий активированного эндогенного гена, могут продуцировать очень высокие уровни желаемого белка и применимы для получения белка in vitro и в генной терапии.
В любом варианте селектируемые и амплифицируемые маркерные гены не должны лежать непосредственно рядом друг с другом.
Необязательно, конструкция ДНК может включать в себя бактериальное начало репликации и бактериальные маркеры устойчивости к антибиотику или другие селектируемые маркеры, которые делают возможным широкомасшабное размножение плазмид в бактериях или любых других подходящих для этого системах клонирования/хозяина. Конструкция ДНК, включающая в себя ДНК, кодирующую селектируемый маркер, вместе с дополнительными последовательностями, такими как промотор и границы сплайсинга, может быть использована для придания селектируемого фенотипа трансфицированным клеткам (например, плазмида рс DN ЕО, схематически изображенная на фиг.4). Такая конструкция ДНК может котрансфицироваться в первичные или вторичные клетки вместе с направляющей последовательностью ДНК с применением описанных здесь способов.
Трансфекция и гомологичная рекомбинация
В соответствии со способом данного изобретения конструкцию ДНК вводят в клетку, такую как первичную, вторичную или иммортализованную клетку, в виде единой конструкции ДНК или в виде отдельных последовательностей ДНК, которые становятся включенными в хромосомную или ядерную ДНК трансфицированной клетки.
Направляющая конструкция ДНК, включающая в себя направляющие последовательности, регуляторную последовательность, экзон, донорный сайт сплайсинга и селектируемые маркерные гены (ген), может быть введена в клетки на одной конструкции ДНК или на отдельных конструкциях. Общая длина конструкции ДНК будет вариировать в зависимости от числа компонентов (направляющих последовательностей, регуляторных последовательностей, экзонов, селектируемых маркерных генов и других элементов, например) и длины каждого из элементов. Полная длина конструкции в общем будет, по меньшей мере, приблизительно 200 нуклеотидов. Далее эта ДНК может быть введена в виде линейной, двухцепочечной (с одноцепочечными районами на одном или обоих концах или без них), одноцепочечной или кольцевой ДНК.
Конструкцию любого типа данного изобретения вводят затем в клетку для получения трансфицированной клетки. Трансфицированную клетку поддерживают при условиях, позволяющих гомологичную рекомбинацию, как это известно в этой области знаний (Capecchi M.R., Science 244: 1288-1292 (1989)). При поддержании гомологично рекомбинантной клетки в условиях, достаточных для транскрипции ДНК, регуляторный район, введенный нацеливающей конструкцией, как в случае промотора, будет активировать транскрипцию.
Конструкции ДНК могут быть введены в клетки разнообразными физическими или химическими способами, в том числе электропорацией, микроинъекцией, бомбардировкой микроснарядами, осаждением фосфатом кальция и мидиируемой липосомами, полибреном или ДЭАЭ-декстраном трансфекцией. Альтернативно, для введения этой ДНК можно использовать инфекционные векторы, такие как ретровирусы, вирус герпеса, аденовирус, аденовирусассоциированные, полиовирусные векторы и вирус эпидемического паротита.
Иногда (необязательно) направляющая ДНК может вводиться в клетки в виде двух или более отдельных фрагментов ДНК. В случае использования двух фрагментов эти два фрагмента имеют общую гомологию последовательности ДНК (перекрывание) при 3'-конце одного фрагмента и 5'-конце другого фрагмента, в то время как один фрагмент несет первую направляющую последовательность, а другой несет вторую направляющую последовательность. При введении в клетку эти два фрагмента могут подвергаться гомологичной рекомбинации с образованием единого фрагмента с первой и второй направляющими последовательностями, фланкирующими район перекрывания между двумя исходными фрагментами. Затем образовавшийся фрагмент находится в форме, пригодной для гомологичной рекомбинации с клеточными последовательностями-мишенями. Можно использовать более двух фрагментов, сконструированных таким образом, что они подвергаются гомологичной рекомбинации друг с другом с образованием в конце концов продукта, пригодного для гомологичной рекомбинации с клеточными последовательностями-мишенями, как описано выше.
Гомологично рекомбинантные клетки
Событие нацеливания приводит к вставке (инсерции) регуляторной последовательности направляющей (нацеливающей) конструкции, помещению эндогенного гена под их контроль (например, путем инсерции промотора или энхансера или обоих против хода транскрипции эндогенного гена или регуляторного района). Необязательно, событие нацеливания может одновременно приводить к делеции эндогенного регуляторного элемента, такого как тканеспецифический негативный регуляторный элемент. Нацеливание может вытеснять существующий элемент; например, тканеспецифический энхансер может быть вытеснен энхансером, имеющим более широкую или отличающуюся специфичность в отношении типа клеток, чем природно встречающиеся элементы, или обнаруживать характер регуляции или индукции, который отличается от встречающегося в соответствующей нетрансфицированной клетке. В этом варианте природно существующие последовательности делетируются, а новые последовательности добавляются. Альтернативно, эндогенные регуляторные элементы не удаляются или не заменяются, а разрушаются или выводятся из строя событием нацеливания, таким как направление экзогенных последовательностей внутрь эндогенных регуляторных элементов.
После введения этой ДНК в клетку, клетку поддерживают при условиях, пригодных для гомологичной рекомбинации между геномной ДНК и частью введенной ДНК, как это известно в данной области (Capecchi M.R., Science 244:1288-1292 (1989)).
Гомологичная рекомбинация между геномной ДНК и введенной ДНК приводит к образованию гомологично рекомбинантной клетки, такой как клетка гриба, растения или. животного и, в частности, первичная, вторичная или иммортализованная клетка человека или другого млекопитающего, в которой последовательности, которые изменяют экспрессию эндогенного гена, оперативно соединены с эндогенным геном, кодирующим продукт, с образованием новой транскрипционной единицы с экспрессией и/или кодирующим потенциалом, который отличается от кодирующего потенциала эндогенного гена. В частности, данное изобретение включает в себя гомологично рекомбинантную клетку, содержащую регуляторные последовательности и экзон, фланкированные донорным сайтом сплайсинга, которые вводят при определенном сайте при помощи нацеливающей конструкции ДНК и которые оперативно соединены со вторым экзоном эндогенного гена. Необязательно могут присутствовать множественные экзогенные экзоны (кодирующие или некодирующие) и интроны, оперативно соединенные с каким-либо экзоном эндогенного гена. Полученные гомологично рекомбинантные клетки культивируют при условиях, которые проводят отбор на амплификацию, если нужно, ДНК, кодирующей амплифицируемый маркер, и новую транскрипционную единицу. С амплификацией или без нее клетки, получаемые этим способом, можно культивировать при условиях, известных в этой области, пригодных для экспрессии целевого белка, получая тем самым этот белок in vitro, или эти клетки можно использовать для доставки in vivo терапевтического белка (т.е. для генной терапии).
В применении здесь, термин "первичная клетка" включает в себя клетки, присутствующие в суспензии клеток, выделенных из источника ткани позвоночного животного (перед их посевом, т.е. присоединенные к субстрату для культуры ткани в чашке или колбе), клетки, присутствующие в эксплантате, полученном из ткани, клетки обоих типов, высеянные в первый раз, и клеточные суспензии, полученные из этих посеянных клеток. Термин "вторичная клетка или клеточный штамм" относится к клеткам на всех последующих стадиях в культивировании. Т.е. посеянную в первый раз первичную клетку удаляют с клеточного субстрата и пересевают (пассируют), ее называют здесь вторичной клеткой, так же как и все клетки в последующих пассажах. Вторичные клетки являются клеточными штаммами, которые состоят из вторичных клеток, которые были пассированы один или более раз. Клеточный штамм состоит из вторичных клеток, которые:
1) были пассированы один или более раз;
2) обнаруживают конечное число средних удвоений популяции в культуре;
3) обнаруживают свойства ингибируемого контактом, зависимого от прикрепления роста (зависимость от прикрепления не приложима к клеткам, которые размножаются в суспензионной культуре); и
4) не являются иммортализованными.
Иммортализованные клетки представляют собой клеточные линии (в противоположность клеточным штаммам, с обозначением "штамм", резервируемым для первичных и вторичных клеток), решающим признаком которых является то, что они обнаруживают явно неограниченную продолжительность жизни в культуре.
Клетки, выбранные для описываемого способа, могут подразделяться на 4 типа или категории:
1) клетки, которые сами по себе не производят или не содержат данного белка или продукта (такого, как белок, который обычно не экспрессируется этой клеткой, или такого, как слитый белок, обычно не обнаруживаемый в природе),
2) клетки, которые продуцируют или содержат этот белок или продукт, но в количествах иных, чем желательные количества (например, в количествах, меньших, чем физиологически нормальный низший уровень для полученной клетки perse),
3) клетки, которые продуцируют этот белок или продукт при физиологически нормальных уровнях для самой клетки, но должны иметь увеличенное или усиленное содержание или продуцирование этого белка, и
4) клетки, в которых желательно изменить характер регуляции или индукции гена, кодирующего этот белок.
Первичные, вторичные и иммортализованные клетки для трансфекции данным способом могут быть получены из разнообразных тканей и включают в себя все клеточные типы, которые могут поддерживаться в культуре. Например, первичные и вторичные клетки, которые можно трансфицировать данным способом, включают в себя фибробласты, кератиноциты, эпителиальные клетки (например, эпителиальные клетки молочной железы, кишечные эпителиальные клетки), эндотелиальные клетки, глиальные клетки, нервные клетки, форменные элементы крови (например, лимфоциты, клетки костного мозга), мышечные клетки и предшественники этих типов соматических клеток. При использовании гомологично рекомбинантных клеток в генной терапии первичные клетки предпочтительно получают из индивидуума, которому вводят трансфицированные первичные или вторичные клетки. Однако первичные клетки могут быть получены из донора (а не из реципиента) того же вида.
Гомологично рекомбинантные иммортализованные клетки можно также получать данным способом и использовать либо для получения белка, либо для генной терапии. Примеры иммортализованных человеческих клеточных линий для получения белка или генной терапии по способу данного изобретения включают в себя (но не ограничены ими) клетки НТ1080 (АТСС CCL 121), клетки HeLa и производные клеток HeLa (АТСС CCL 2, 2.1 и 2.2), клетки рака молочной железы MCF-7 (ATCC BTH 22), клетки лейкоза К-562 (АТСС CCL 243), клетки карциномы KB (ATCC CCL 17), клетки рака яичника 2780АD (Van der Blick, A.M. et al., Cancer Res. 48:5927-5932 (1988)), клетки Raji (ATCC CCL 86), клетки Jurkat (ATCC T1B 152), клетки Namalwa (ATCC CRL 1432), клетки HL-60 (ATCC CCL 240), клетки Daudi (ATCC CCL 213), клетки RPM1 8226 (ATCC CCL 155), клетки U-937 (ATCC CRL 1593), клетки меланомы Боуеса (ATCC CRL 9607), клетки подлинии 2R4 W1-38V A13 (ATCC CLL 75.1) и клетки MOLT-4 (ATCC CRL 1582), а также клетки гетерогибридомы, полученной слиянием клеток человека и клеток других видов. Могут быть использованы вторичные штаммы человеческих фибробластов, такие как W1-38 (ATCC CCL 75) и MRC-5 (ATCC CCL 171). Кроме того, первичные, вторичные или иммортализованные клетки человека, а также первичные, вторичные или иммортализованные клетки из других видов, проявляющие свойства амплификации генов in vitro можно использовать для получения белка in vitro или для генной терапии.
Способ превращения гена в кДНК-копию
Данное изобретение касается также способа, при помощи которого гомологичную рекомбинацию используют для превращения гена в кДНК-копию (копию гена, не содержащую интронов). кДНК-копия может быть перенесена в дрожжи или бактерии для получения белка in vitro, или кДНК-копия может быть введена в клетку млекопитающего для получения белка in vitro или in vivo. Если эта кДНК должна быть перенесена в микробные клетки, две ДНК-конструкции, содержащие направляющие последовательности, вводят гомологичной рекомбинацией, причем одна конструкция против хода транскрипции и одна конструкция по ходу транскрипции гена человека кодирует терапевтический белок. Например, последовательности, введенные против хода транскрипции, включают в себя последовательности ДНК, гомологичные геномным последовательностям ДНК при или против хода транскрипции ДНК, кодирующей первую аминокислоту зрелого процессированного терапевтического белка; ретровирусный длинный концевой повтор (LTR); последовательности, кодирующие маркер для отбора в микробных клетках; регуляторный элемент, функционирующий в микробных клетках; и ДНК, кодирующую лидерный пептид, ускоряющий секрецию из микробных клеток, с донорным сайтом сплайсинга. Последовательности, вводимые против хода транскрипции, вводят вблизи или против хода транскрипции геномной ДНК, кодирующей первую аминокислоту зрелого, процессированного терапевтического белка. Последовательности, вводимые по ходу транскрипции, включают в себя последовательности ДНК, гомологичные последовательностям геномной ДНК при или по ходу транскрипции ДНК, кодирующей последнюю аминокислоту зрелого, процессированного белка; микробную последовательность терминации транскрипции; последовательности, способные управлять репликацией ДНК в микробных клетках; и ретровирусный длинный концевой повтор (LTR). Последовательности, вводимые по ходу транскрипции, вводят смежно или по ходу транскрипции ДНК, кодирующей термирующий кодон зрелого, процессированного терапевтического белка. После введения этих двух конструкций ДНК в клетки полученные клетки поддерживают при условиях, пригодных для гомологичной рекомбинации между введенной ДНК и геномной ДНК, с образованием в результате рекомбинантных клеток. Иногда (необязательно) одна или обе конструкции ДНК могут кодировать один или более маркеров для позитивного или негативного отбора клеток, содержащих конструкцию ДНК, и стадию отбора можно добавить к способу, после того как одна или обе конструкции ДНК были введены в клетки. Альтернативно, последовательности, кодирующие маркер для отбора в микробных клетках, и последовательности, способные управлять репликацией ДНК в микробных клетках, могут присутствовать против хода транскрипции или по ходу транскрипции направляющей конструкции или маркер для отбора в микробных клетках может присутствовать по ходу транскрипции в направляющей конструкции, а последовательности, ответственные за репликацию ДНК, могут присутствовать против хода транскрипции направляющей конструкции. Затем гомологично рекомбинантные клетки культивируют при условиях, пригодных для регулируемой LTR транскрипции, процессинга и обратной транскрипции РНК-продукта гена, кодирующего терапевтический белок. Продукт обратной транскрипции представляет собой конструкцию ДНК, содержащую не имеющую интронов ДНК-копию, кодирующую терапевтический белок, оперативно связанную с последовательностями ДНК, содержащими две описанные выше экзогенные конструкции ДНК. Не содержащую интронов конструкцию ДНК, полученную данным способом, вводят затем в микробную клетку. Затем микробную клетку культивируют при условиях, пригодных для экспрессии и секреции терапевтического белка.
Получение белка in vivo
Гомологично рекомбинантные клетки данного изобретения применимы в виде популяций гомологично рекомбинантных клеточных линий, в виде популяций гомологично рекомбинантных первичных или вторичных клеток, гомологично рекомбинантных клональных клеточных штаммов или линий, гомологично рекомбинантных гетерогенных клеточных штаммов или линий и в виде клеточных смесей, в которых по меньшей мере присутствует одна репрезентативная клетка одной из четырех указанных выше категорий гомологично рекомбинантных клеток. Такие клетки можно использовать в системе доставки для лечения индивидуума с патологическим или нежелательным состоянием, отвечающим на доставку терапевтического продукта, представляющего собой:
1) терапевтический белок (например, белок, который отсутствует, продуцируется на низком уровне по отношению к физиологическим потребностям индивидуума, является дефектным или утилизируется неэффективно или неправильным образом в индивидууме; белок с новым функциями, такими как ферментативные или транспортные функции) или
2) терапевтическую нуклеиновую кислоту (например, РНК, ингибирующую экспрессию гена или имеющую собственную ферментативную активность). В способе данного изобретения обеспечения терапевтического белка или нуклеиновой кислоты гомологически рекомбинантные первичные клетки, клональные клеточные штаммы или гетерогенные клеточные штаммы вводят индивидууму, которому необходимо лечение или предотвращение патологического или нежелательного состояния, в достаточном количестве и подходящим способом для экспрессии или создания доступности этого белка или экзогенной ДНК при соответствующих физиологических уровнях. Физиологически приемлемым уровнем является уровень, который либо приближается к уровню, при котором продукт обычно продуцируется в теле, либо ведет к улучшению патологического или нежелательного состояния. Согласно варианту изобретения, описанному здесь, вводимые гомологично рекомбинантные иммортализованные клеточные линии могут быть заключены в одно или более полупроницаемых барьерных (защитных) приспособлений. Свойства проницаемости этого приспособления таковы, что клетки не могут выходить из приспособления при имплантации в животное, но терапевтический продукт легко проникает через него и может покидать барьерное устройство и входить в локальное пространство, окружающее имплантат, или вступать в системную циркуляцию. Например, hGH, hEPO, инсулинотропин человека, hGM-CSF, hG-CSF, α-интерферон человека или FSHβ человека могут доставляться системно больным для получения терапевтических преимуществ.
Барьерные устройства, в частности, применимы и позволяют имплантировать гомологично рекомбинантные иммортализованные клетки, гомологично рекомбинантные клетки из других видов (гомологично рекомбинантные ксеногенные клетки) или клетки из не имеющего гистосовместимости донора (гомологично рекомбинантные аллогенные клетки) для лечения состояний человека или животных или для сельскохозяйственных целей (т.е. для производства мяса и молока). Барьерные устройства позволяют также осуществлять краткосрочную (т.е. временную) терапию путем обеспечения легкого доступа к клеткам для их удаления в случае, когда режим лечения должен быть по тем или иным причинам прекращен.
Множество синтетических, полусинтетических или природных фильтрующих мембран можно использовать для этой цели, в том числе (но не только) целлюлозу, ацетат целлюлозы, нитроцеллюлозу, полисульфон, поливинилдендифторид, поливинилхлоридные полимеры и полимеры производных поливинилхлорида. Барьерные устройства можно использовать для возможности применения первичных, вторичных или иммортализованных клеток из других видов в генной терапии человека.
Получение белка in vitro.
Гомологично рекомбинантные клетки из человека или других видов согласно этому изобретению можно использовать также для получения белка in vitro. Эти клетки поддерживают при условиях, хорошо известных в данной области знаний, которые приводят к экспрессии белка. Белки, экспрессируемые при помощи описанных способов, могут быть очищены из клеточных лизатов или клеточных супернатантов для получения очищенного желаемого белка. Белки, полученные согласно этому способу, включают в себя терапевтические белки, которые могут доставляться в человека или в животное общепринятыми фармацевтическими способами, известными в этой области (например, перорально, внутривенно, внутримышечно, интраназально или кодкожно). Такие белки включают в себя hGH, hEPO, инсулинотропин человека, hGM-CSF, hG-CSF, FSHβ или α-интерферон. Эти клетки могут быть иммортализованными, первичными или вторичными клетками. Использование клеток из других видов может быть желательным в случаях, когда предпочтительны клетки не из человека для целей получения белка, если белок не из человека является терапевтически или коммерчески применимым, например использование клеток, полученных из лосося, для получения кальцитонина лосося, использование клеток, полученных из свиней, для получения свиного инсулина и использование бычьих клеток для получения бычьего гормона роста.
Преимущества
Методологии, конструкции ДНК, клетки и образующиеся белки этого изобретения обладают разносторонностью и многими другими преимуществами по сравнению со способами, применяемыми в настоящее время в этой области в генном нацеливании. Способность активации эндогенного гена помещением экзогенной регуляторной последовательности при различных положениях, вариирующих от непосредственно смежного положения по отношению к представляющему интерес гену (непосредственно слитой с транскрибируемым районом обычного гена) до положения 30 т.п.н. или далее против хода транскрипции от транскрибируемого района эндогенного гена или внутри интрона индогенного гена, является выгодной для экспрессии гена в клетках. Например, способ можно использовать для помещения регуляторного элемента против хода транскрипции или по ходу транскрипции районов, которые обычно молчат или негативно регулируют ген. Помещение регуляторного элемента против хода транскрипции или по ходу транскрипции такого района может преодолеть такие доминантные негативные эффекты, которые обычно ингибируют транскрипцию. Кроме того, районы ДНК, которые обычно ингибируют транскрипцию или имеют пагубное действие на экспрессию гена, могут быть делегированы при помощи направляющих конструкций, описанных здесь.
Дополнительно, т.к. известно, что промоторная функция сильно зависит от локального окружения, можно испытывать широкие диапазоны положений для нахождения локальных окружений, оптимальных для функционирования промотора. Однако, поскольку обнаружено, что стартовые кодоны ATG находятся часто внутри ДНК млекопитающего (приблизительно один на 48 п.н.), транскрипция не может просто инициироваться при любом положении против хода транскрипции гена и продуцировать транскрипт, содержащий длинную лидерную последовательность, предшествующую правильному стартовому кодону ATG, т.к. частое присутствие кодонов ATG в такой лидерной последовательности будет предотвращать трансляцию корректного продукта гена и делать матрицу бесполезной. Таким образом, введение экзогенного экзона, донорного сайта сплайсинга и, необязательно, интрона и акцепторного сайта сплайсинга в нацеливающие конструкции, содержащие регуляторный район, позволяет оптимизировать экспрессию гена посредством идентификации оптимального сайта для функции регуляторного района, без ограничения, налагаемого необходимостью избежать неподходящих стартовых кодонов ATG в продуцируемой мРНК. Это обеспечивает значительно увеличенную гибкость в помещании конструкции и делает возможной активацию широкого диапазона генов. Конструкции ДНК данного изобретения применимы также, например, в способах получения слитых белков, кодируемых рекомбинантными, или экзогенными, последовательностями и эндогенными последовательностями.
Генное нацеливание и амплификация генов, описанные выше, применимы, в частности, для изменения экспрессии генов, которые образуют транскрипционные единицы, которые настолько большие, что их трудно выделять и экспрессировать, или для включения генов, для которых вся кодирующая область белка не доступна или не была клонирована. Таким образом, эти конструкции ДНК, описанные выше, применимы для оперативного соединения экзогенных регуляторных элементов с эндогенными генами способом, который точно определяет транскрипционную единицу, обеспечивает гибкость в относительном размещении экзогенных регуляторных элементов и эндогенных генов в конечном счете и делает возможной высококонтролируемую систему для получения и регуляции экспрессии генов, представляющих терапевтический интерес.
Объяснение примеров
Как описано здесь, заявители показали, что ДНК может быть введена в клетки, такие как первичные, вторичные или иммортализованные клетки позвоночных, и интегрироваться в геном трансфицированных клеток посредством гомологичной рекомбинации. Далее они продемонстрировали, что экзогенная ДНК имеет желаемую функцию в гомологично рекомбинированных (HR) клетках и что точно "пораженные" клетки-мишени могут быть идентифицированы на основе детектируемого фенотипа, сообщаемого правильно нацеленной ДНК.
Заявители описывают конструирование плазмиды, применимой для нацеливания (направления) в определенный локус (локус HPRT) в геноме человека и отбора на основе устойчивого к лекарственному средству фенотипа (Пример 1a). Эта плазмида названа рЕ3 Neo и ее интеграция в клеточный геном при локусе HPRT дает клетки, которые имеют hprt, 6-TG-устойчивый фенотип и являются также устойчивыми к G418. Как описано, заявители показали, что плазмида pE3Neo функционирует правильно в нацеливании гена в установленной клеточной линии фибробластов человека (Пример 1b) путем демонстрации локализации ДНК, введенной в клетки этой линии, внутри экзона 3 гена HPRT.
Кроме того, заявители демонстрируют генное нацеливание в первичных и вторичных фибробластах кожи человека с использованием pE3Neo (Пример 1с). Данная заявка демонстрирует далее, что модификация концов ДНК усиливает направление ДНК в геномную ДНК (Примеры 1с и 1e).
Заявители описывают также способы, при помощи которых ген может быть вставлен в заранее выбранном сайте в геноме клетки, такой как первичная, вторичная или иммортализованная клетка, посредством генного нацеливания (Пример 1d).
Кроме того, данное изобретение касается способа получения белка с применением трансфицированных клеток. Способ использует трансфицированные клетки, такие как первичные клетки, вторичные клетки или иммортализованные клетки, с экзогенной ДНК, которая кодирует терапевтический продукт, или с ДНК, которая может быть нацелена на эндогенный ген, кодирующий терапевтический продукт. Например, Примеры 1g, 1h, 1j, 1k, 2, 3, 4 и 6-9 описывают получение белка нацеливанием (направлением) на выбранный экзогенный ген элементов последовательности ДНК, которые будут изменять экспрессию этого эндогенного гена.
Заявители также описывают конструкции ДНК и способы амплификации эндогенного клеточного гена, который был активирован генным нацеливанием (Примеры 3, 6, 8 и 9).
Примеры 1f-1h, 2, 4 и 6 иллюстрируют варианты, в которых обычные регуляторные последовательности против хода транскрипции от гена ЕРО человека изменяют для обеспечения экспрессии hEPO в первичных или вторичных фибробластных штаммах, которые не экспрессируют ЕРО в детектируемых количествах в их нетрансфицированном состоянии. В одном варианте продукт нацеливания оставляет обычный белок ЕРО интактным, но под контролем мышиного промотора металлотионеина. Примеры 1i и 1j демонстрируют использование сходных направляющих конструкций для активации эндогенного гена гормона роста в первичных и вторичных фибробластах человека. В других вариантах для активации экспрессии ЕРО в фибробластах человека продукты событий нацеливания являются химерными транскрипционными единицами, в которых первый экзон гена гормона роста человека расположен против хода транскрипции от экзонов 2-5 ЕРО. Продуктом транскрипции (регулируемой промотором мышиного металлотионеина), сплайсинга и трансляции является белок, в котором аминокислоты 1-4 сигнального пептида ЕРО заменены аминокислотными остатками 1-3 hGH. Химерная часть этого белка, сигнальный пептид удаляются перед секрецией из клеток. Пример 5 описывает направляющие конструкции и способы получения клеток, которые будут превращать ген (с интронами) в экспрессируемую кДНК-копию этого гена (без интронов), и извлечение таких экспрессируемых кДНК-молекул в микробных (например, дрожжевых или бактериальных) клетках. Пример 6 описывает конструирование нацеливающего вектора, названного рRЕРO4, для двойного отбора и отбора клеток, в которых амплифицировался ген dhfr. Плазмиду PREPO4 использовали для амплификации локуса ЕРО человека в клетках НТ1080 (иммортализованной линии клеток человека) после активации эндогенного гена hEPO гомологичной рекомбинацией. Как указано, ступенчатый отбор в содержащих метотрексат средах привел к 70-кратному увеличению в образовании hEPO в клетках, устойчивых к 0,4 мкМ метотрексату.
Примеры 7 и 8 описывают способы введения регуляторной последовательности против хода транскрипции обычного промотора ЕРО и способы получения ЕРО с использованием такой конструкции. Кроме того, Пример 8 описывает амплификацию гена-мишени ЕРО, полученного по способу Примера 7. Пример 9 описывает способы нацеливания генов α-интерферона, GM-CSF, G-CSF и FSHβ человека для создания клеток, применимых для получения белков.
Эти примеры обеспечивают способы активации или активации и амплификации эндогенных генов генным нацеливанием, которое не требует манипуляции или других использований кодирующих белки районов генов-мишеней. При помощи описанных здесь способов и конструкций ДНК или плазмид или их модификаций, которые очевидны для специалистов с обычной квалификацией в этой области, экспрессия генов может быть изменена в клетках, которые обладают свойствами, желательными для получения белков in vitro (например, в фармации), или для способов доставки белков in vivo (например, в генной терапии). Примеры 5 и 6 иллюстрируют две стратегии транскрипционной активации гена hEPO.
При помощи способов и конструкций ДНК или плазмид, описанных здесь, или их модификаций, которые очевидны для среднего специалиста в этой области, экзогенная ДНК, кодирующая терапевтический продукт (например, белок, рибозим, антисмысловую РНК), может быть вставлена при заранее выбранных сайтах в геноме первичных или вторичных клеток позвоночных (например, млекопитающих, таких как человек, так и других видов).
Данное изобретение будет теперь иллюстрироваться следующими далее примерами, которые не предназначены для какого-либо ограничения изобретения.
ПРИМЕРЫ
Пример 1. Получение трансфицированных клеточных штаммов генным нацеливанием
Генное нацеливание имеет место, когда трансфицированная ДНК либо интегрируется в хромосомные последовательности ДНК, либо частично заменяет хромосомные последовательности ДНК в результате гомологичной рекомбинации. Хотя такие события могут иметь место в ходе любого конкретного эксперимента по трансфекции, они обычно маскируются огромным избытком событий, в которых плазмидная ДНК интегрируется посредством негомологичной или неправильной рекомбинацией.
а. Генерирование конструкции, применимой для отбора событий генного нацеливания в клетках человека
Одним подходом для отбора нацеленных событий является генетический отбор на потерю функции гена, вызываемую интеграцией трансфицирующей ДНК. Локус HPRT человека кодирует фермент гипоксантин-фосфорибозилтрансферазу. Клетки hprt- могут быть отобраны по росту в среде, содержащей нуклеозидный аналог 6-тиогуанин (6-TG): клетки с аллелем дикого типа (HPRT+) убиваются 6-TG, тогда как клетки с мутантными аллелями (hprt-) могут выживать. Таким образом, клетки подвергнутые нацеленным событиям, которые разрушают функцию гена HPRT, можно отбирать в среде 6-TG.
Для конструирования плазмиды для нацеливания на локус HPRT фрагмент 6,9 т.п.н. Hind III, простирающийся от положений 11960-18869 в последовательности HPRT (Genebank name HUMHPRTB: Edwards, A., et al., Genomics 6:593-608 (1990)) и включающий в себя экзоны 2 и 3 гена HPRT, субклонировали в сайт Hind III рUС12. Полученный клон расщепляют при уникальном сайте Xhol в экзоне 3 фрагмента гена HPRT и фрагмент 1,1 т.п.н. SalI-XhoI, содержащий ген neo из pMC1Neo (Stratagene), вставляют, разрушая кодирующую последовательность экзона 3. Была выбрана и обозначена рЕ3Neo одна ориентация с направлением транскрипции neo, противоположным направлению транскрипции HPRT. Замена обычного экзона 3 HPRT neo-разрушенной версией будет приводить к фенотипу hprt-, 6-TG устойчивому. Такие клетки будут также устойчивы к G418.
b. Генное нацеливание в установленной клеточной линии фибробластов человека
В качестве демонстрации нацеливания в иммортализованных клеточных линиях и для установления, что pE3Neo функционирует правильно в генном нацеливании линию клеток фибросаркомы человека НТ1080 (АТСС CCL 121) трансфицировали рЕ3Neo посредством электропорации.
Клетки НТ1080 поддерживали в HAT (гипоксантин/аминоптерин/ксантин) с добавлением DMEM с 15% телячьей сывороткой (Hyclone) перед электропорацией. За два дня перед электропорацией клетки переносили на ту же самую среду без аминоптерина. Экспоненциально растущие клетки трипсинизировали, ресуспендировали и разбавляли в DMEM/15 % телячья сыворотка, центрифугировали и ресуспендировали в PBS (фосфатно-солевом буфере) при конечном клеточном объеме 13,3 млн клеток на мл. рЕ3Neo расщепляли Hind III, отделяя фрагмент 8 т.п.н. HPRT-neo от структуры рUС12, очищали фенольной экстракцией и осаждением этанолом и ресуспендировали при концентрации 600 мкг/мл. 50 мкл (30 мкг) добавляли в кювету для электропорации (расстояние между электродами 0,4 см; Bio-Rad Laboratories) вместе с 750 мкл клеточной суспензии (10 млн клеток). Электропорацию выполняли при 450 вольт, 250 мкФ (Bio-Rad Gene Pulser; Bio-Rad Laboratories). Содержимое кюветы сразу же добавляли к DMEM с 15% телячьей сывороткой, получая клеточную суспензию 1 млн. клеток на 25 мл среды. 25 мл обработанной клеточной суспензии высевали на чашки для культуры ткани с диаметром 150 мм и инкулировали при 37°С и 5% СО2. Через 24 часа добавляли раствор G418 непосредственно в чашки, получая конечную концентрацию 800 мкг/мл G418. Через 5 дней среду заменяли смесью ДМЕМ/15% телячья сыворотка/800 мкг/мл 6418. Через 9 дней после электропорации среду заменяли DMEM/15% телячья сыворотка/800 мкг/мл G418 и 10 мкМ 6-тиогуанином. Колонии, устойчивые к G418 и 6-TG, выскребали при помощи клонирующих цилиндров через 14-16 дней после инициации двойного отбора. Результаты 5 репрезентативных экспериментов по нацеливанию в клетках НТ1080 показаны в Таблице 1.
6-TG-клонов
Для трансфекции 5 контрольные чашки, предназначенные для определения общего выхода G418r-колонии, показали, что 33700 G418r-колонии могли генерироваться из первоначальных 1×107 обработанных клеток. Таким образом, отношение нацеленных событий к ненацеленным равно 66/33700 или 1 к 510. В 5 объединенных экспериментах нацеленные события происходят при частоте 3,6×106 или 0,00036% обработанных клеток.
Эксперименты с рестриктазами и с гибридизацией по Саузерну с применением зондов, произведенных из генов neo и HPRT, показали, что ген neo локализован на локусе HPRT при предсказанном сайте внутри экзона 3 HPRT.
с. Генное нацеливание в первичных и вторичных фибробластах кожи человека
рН3Neo расщепляли Hind III, отделяя фрагмент 8 т.п.н. HPRT- neo от структуры рUС12, и очищали фенольной экстракцией и осаждением этанолом. ДНК суспендировали при 2 мг/мл. 3 млн вторичных клеток фибробластов крайней плоти человека в объеме 0,5 мл подвергали электропорации при 250 вольт и 960 мкФ с 100 мкг Hind III рЕ3Neo (50 мкл). Выполняли три отдельные трансфекции для 9 млн обработанных клеток в целом. Клетки процессировали и проводили отбор на устойчивость к G418. 500000 клеток на 150 мм-чашку для культуры высевали для отбора на устойчивость к G418. После 10 дней в присутствии G418 культуральную среду заменяли питательной средой для фибробластов человека, содержащей 400 мкг/мл G418 и 10 мкМ 6-TG. Отбор с комбинацией двух лекарственных средств продолжали еще в течение 10 дней. Чашки сканировали микроскопически для локализации колоний фибробластов человека, устойчивых к обоим лекарственным средствам. Фракция G418r 6-TGr-колоний составляла 4 на 9 млн обработанных клеток. Эти колонии составляют 0,0001% (или 1 в млн) всех клеток, способных к образованию колоний. Контрольные чашки, предназначенные для определения общего выхода G418r-колоний, показали, что 2850 G418r-колоний могли образоваться из первоначальных 9×106 обработанных клеток. Таким образом, отношение нацеленных событий к ненацеленным равно 4/2850 или 1/712. Эксперименты с рестриктазой и гибридизацией по Саузерну с применением зондов, полученных из генов neo и HPRT, использовали для локализации гена neo в локусе HPRT при предсказанном сайте внутри экзона 3 HPRT и демонстрации того, что нацеливание имело место в этих 4 клональных клеточных штаммах. Колонии, устойчивые к обоим лекарственным средствам, также выделяли посредством трансфекции первичных клеток (1/3,0×107).
Результаты нескольких экспериментов с нацеливанием при помощи рЕ3Neo суммированы в Таблице 2. Расщепленную Hind III плазмиду рЕ3Neo либо трансфицировали непосредственно, либо обрабатывали экзонуклеазой III для генерирования 5'-одноцепочечных выступов перед трансфекцией (см. Пример 1с). Испытывали препараты ДНК с одноцепочечными районами от 175 до 930 п.н. длиной. При использовании только рЕ3Neo, расщепленной Hind III, идентифицировали 1/799 G418r-устойчивых колоний при помощи рестрикционного анализа и гибридизации по Саузерну, имеющих нацеленную инсерцию гена neo при локусе HPRT (всего выделили 24 нацеленных клона). Нацеливание максимально стимулировалось (приблизительно в 10 раз) при использовании выступов длиной 175 п.н. с 1/80 G418r-колониями, обнаруживающими фрагменты рестрикции, которые были диагностическими для нацеливания при HPRT (всего выделили 9 нацеленных клонов). Таким образом, при помощи условий и рекомбинантных конструкций ДНК, описанных здесь, нацеливание действительно имело место в обычных фибробластах человека и общая частота нацеливания (число обнаруживающих нацеливание клонов, разделенное на общее число клонов, стабильно трансфицированных до устойчивости к G418) могла стимулироваться трансфекцией нацеливающими конструкциями, содержащими одноцепочечные выступающие концы, способом, описанным в Примере 1е.
350 п.н.
п.н.
d. Образование конструкции для направленнной инсерции представляющего терапевтический интерес гена в геном человека и ее использование в генном нацеливании
Вариант pE3Neo, в котором представляющий терапевтический интерес ген вставлен в кодирующую HPRT область, смежную с геном neo или вблизи гена neo, можно использовать для направления представляющего интерес в терапии гена в специфическое положение в реципиентном геноме первичной или вторичной клетки. Такой вариант рЕ3Neo можно сконструировать для нацеливания гена hGH на локус HPRT.
рХGН5 (схематически представленную на фиг.3) расщепляют EcoRI и выделяют фрагмент 4,1 т.п.н., содержащий ген hGH и соединенный с ним мышиный промотор металлотионеина (mMT). Выступающие концы EcoRI заполняют при помощи фрагмента Кленова из ДНК-полимеразы E.Coli. Отдельно рЕ3Neo расщепляют XhoI, которая разрезает при границе фрагмента neo и экзона 3 HPRT (3'-соединение вставки в экзон 3). XhoI выступающие концы линеаризованной плазмиды заполняют фрагментом Кленова из ДНК-полимеразы E.coli и полученный фрагмент лигируют с фрагментом hGH-mMT 4,1 т.п.н. с затупленными концами. Бактериальные колонии, полученные из смеси для лигирования, скринируют рестрикционным анализом на инсерцию одной копии фрагмента hGH-mMT, и одну ориентацию, при которой ген hGH транскрибирован в том же направлении, что и ген neo, выбирают и обозначают как рЕ3Neo/hGH. Эту плазмиду расщепляют Hind III, получая фрагмент 12,1 т.п.н., содержащий последовательности HPRT, neo и mMT-hGH. Расщепленную ДНК обрабатывают и трансфицируют в первичные или вторичные фибробласты человека, как описано в Примере 1с. Отбирают G418r TGr-колонии и анализируют их на инсерцию последовательностей mMT-GSH и neo в гене HPRT, как описано в Примере 1с. Отдельные колонии анализируют на экспрессию hGH с использованием коммерчески доступного иммутотеста (Nichols Institute).
Вторичные фибробласты человека трансфицировали рЕ3Neo/hGH и устойчивые к тиогуанину колонии анализировали на стабильную экспрессию hGH и с применением рестрикционного анализа и гибридизации по Саузерну. Из 13 анализированных TGr-колоний 8 колоний имели инсерцию гена hGH в локусе эндогенного HPRT. Все 8 штаммов стабильно экспрессировали значительные количества hGH со средним уровнем экспрессии 22,7 мкг/106 клеток/24 часа. Альтернативно, плазмиду рЕ3NeoERO (см. фиг.4) можно использовать для направления ЕРО в локус HPRT человека.
Использование гомологичной рекомбинации для направления представляющего терапевтический интерес гена в специфическое положение в геномной ДНК клетки может быть расширено и сделано более применимым для получения продуктов для терапевтических целей (например, для фармацеи, генной терапии) посредством инсерции (вставки) гена, через который клетки, содержащие амплифицированные копии этого гена, могут быть отобраны путем выдерживания их в подходящем режиме отбора при помощи лекарственных средств. Например, плазмида рЕ3Neo/hGH (Пример 1d) может быть модифицирована встраиванием гена dhfr, ada или CAD в положение непосредственно рядом с генами hGH или neo в рЕ3Neo/hGH. Первичные, вторичные или иммортализованные клетки трансфицируют такой плазмидой и идентифицируют точно нацеленные события. Затем эти клетки обрабатывают увеличивающимися концентрациями лекарственных средств, пригодных для отбора клеток, содержащих амплифицированные гены (для dhfr селектирующим агентом является метотрексат, для CAD селектирующим агентом является N-(фосфонацетил)-L-аспартат (PALA) и для ada селектирующим агентом является адениновый нуклеозид (например, аденозин). В этом случае интеграция терапевтического гена будет коамплифицироваться вместе с геном, амплифицированные копии которого отбираются. Таким образом, генетическое конструирование клеток для получения генов для терапевтический целей может легко контролироваться посредством предварительного выбора сайта, при котором происходит интеграция направляющей конструкции и при котором амплифицированные копии находятся в амплифицированных клетках.
е. Модификация концов ДНК для усиления нацеливания
Несколько линий доказательства позволяют предположить, что выступающие 3'-концы участвуют в некоторых путях гомологичной рекомбинации E.coli, бактериофага, S.cerevisiae и Xenopus laevis. В ооцитах Xenopus laevis молекулы с выступающими 3'-концами длиной в несколько сотен пар оснований подвергались рекомбинации с подобным образом обработанными молекулами гораздо более быстро после микроинъекции, чем молекулы с очень короткими выступающими концами (4 п.н.), образованными при расщеплении рестриктазами. В дрожжах образование выступающих 3'-концов длиной в несколько сотен п.н. является, по-видимому, ограничивающей скорость стадией в мейотической рекомбинации. Не было сообщений об участии выступающих 3'-концов в рекомбинации в клетках человека и ни в одном случае не было показано, что модифицированные ДНК-субстраты какого-либо сорта усиливают нацеливание (одну форму гомологичной рекомбинации) в каком-либо виде. Эксперимент, описанный в следующем примере и в Примере 1с, позволяет предположить, что выступающие 5'-концы эффективны для стимуляции нацеливания в первичных, вторичных и иммортализованных фибробластах человека.
Не было сообщений об усилении нацеливания путем модификации концов трансфицирующих молекул ДНК. Этот пример служит для иллюстрации того, что модификация концов линейных молекул ДНК путем превращения концов молекул из двухцепочечной формы в одноцепочечную форму может стимулировать нацеливание и включение в геном первичных и вторичных фибробластов человека.
1100 мкг плазмиды рЕ3Neo (Пример 1а) расщепляли Hind III. Эту ДНК можно использовать сразу после фенольной экстракции и осаждения этанолом или фрагмент Hind III 8 т.п.н., содержащий только HPRT и ген neo, может быть отделен от последовательностей вектора рUС12 гель-электрофорезом. Расщепление при помощи ExoIII расщепленной Hind III ДНК приводит к интенсивному экзонуклеолитическому расщеплению на каждом конце, начинающемуся на каждом свободном 3'-конце и оставляющему выступающие 5'-концы. Степень экзонуклеолитического действия и, следовательно, длина образующихся 5'-выступов могут регулироваться вариированием времени расщепления при помощи Exo III. Расщепление при помощи Exo III 100 мкг расщепленной Hind III рЕ3Neo проводят в соответствии с рекомендуемыми поставщиком условиями в течение времени 30 с, 1 мин, 1,5 мин, 2 мин, 2,5 мин, 3 мин, 3,5 мин, 4 мин, 4,5 мин, 5 мин. Для наблюдения степени расщепления аликвоту из каждой временной точки, содержащую 1 мкг обработанной Exo III ДНК, обрабатывают нуклеазой маша (Promega) при рекомендуемых поставщиком условиях и пробу фракционируют гель-электрофорезом. Измеряют различие в размере между необработанной, расщепленной Hind III рЕ3Neo и теми же самыми молекулами, обработанными Exo III и нуклеазой маша. Это различие в размере, разделенное на 2, дает среднюю длину 5'-выступа на каждом конце молекулы. При использовании указанных выше временных точек и расщепления при 30°С полученный 5'-выступы должны находиться в пределах от 100 до 1000 оснований.
60 мкг обработанной ExoIII ДНК (общего продукта расщепления рЕ3Neo при помощи HindIII) из каждой временной точки очищали и вводили электропорацией в первичные, вторичные или иммортализованные фибробласты человека при условиях, описанных в Примере 1с. Степень, до которой усиливается нацеливание каждым обработанным ExoIII препаратом, определяют количественно подсчетом числа G418r 6-TGr-колоний и сравнением этих чисел с нацеливанием расщепленной HindIII рЕ3Neo, не обработанной ExoIII.
Действие выступающих 3'-концов можно также оценить количественно при помощи аналогичной системы. В этом случае расщепленную Hind III рЕ3Neo обрабатывают экзонуклеазой гена 6 бактериофага Т7 (United States Biochemicals) в течение разных временных интервалов при рекомендуемых поставщиком условиях. Определение степени расщепления (средней длины 3'-выступа, образуемого на один конец) и условий электропорации соответствовали описанным для обработанной ExoIII ДНК. Степень, до которой усиливается нацеливание каждый обработанным экзонуклеазой гена 6 Т7 препаратом, определяют количественно путем подсчета числа 6418r 6-TGr-колоний и сравнения этих чисел с нацеливанием расщепленной HindIII рЕ3Neo, не обработанной экзонуклеазой гена 6 Т7.
Возможны другие способы образования выступающих 5'- и 3'-концов, например денатурация и отжиг двух линейных молекул, которые частично перекрываются друг с другом, будут создавать смесь молекул, имеющих 3'-выступ на обоих концах или 5'-выступ на обоих концах, а также повторно гибридизованных фрагментов, неотличимых от исходных линейных молекул. Длину этих выступающих концов определяют по длине ДНК, которая не является общей между двумя фрагментами ДНК.
f. Конструирование направляющих плазмид для помещения гена эритропоэтина человека под контроль мышиного промотора металлотионеина в первичных, вторичных и иммортализованных фибробластах человека
Дальнейшее описание служит для иллюстрации одного варианта данного изобретения, в котором обычные позитивные и негативные регуляторные последовательности, расположенные против хода транскрипции от гена эритропоэтина человека (hEPO), изменяют для обеспечения экспрессии эритропоэтина человека в первичных, вторичных или иммортализованных фибробластах человека, которые обычно не экспрессируют hEPO в значительных количествах.
Район, лежащий исключительно против хода транскрипции от кодирующего района ЕРО человека, может быть амплифицирован при помощи ПЦР. Три набора праймеров, применимых для этой цели, были сконструированы после анализа опубликованной последовательности ЕРО человека [Genebank designation HUMERPA; Lin. F-k., et al., Proc. Natl. Acad. Sci. USA 82:75807584 (1985)]. Эти пары праймеров могут амплифицировать фрагменты 609, 603 или 590 п.н. (см. табл. 3).
 20
20
(SEQ ID №1)595
(SEQ ID №2)24
(SEQ ID №3)59540
(SEQ ID №4)595
Эти три фрагмента существенно перекрываются и взаимозаменяемы для поставленных целей. Фрагмент 609 п.н., простирающийся от -623 до -14 по отношению к сайту инициации трансляции (положения нуклеотидов HUMERPA 2-610), лигируют на обоих концах с линкерами ClaI. Полученный фрагмент с линкерами ClaI расщепляют ClaI и встраивают в сайт ClaI pBluescript II S k/+ (Stratagene) с такой ориентацией, что положение нуклеотида HUMERPA 610 находится рядом с сайтом SalI в плазмидном полилинкере). Эта плазмида, р5'ЕРО, может быть расщеплена отдельно при уникальных сайтах FSpI или SfiI в расположенном против хода транскрипции фрагменте ЕРО человека (положения нуклеотидов HUMERPA 150 и 405, соответственно) и лигирована с промотором мышиного металлотионеина. В типичном случае этот фрагмент EcoRI-BglII 1,8 т.п.н. из гена mMT-1 (не содержащий кодирующих последовательностей mMT; Hamer, D.H. u Walling M., J. MOL. Appl. Gen. 1:273-288 (1982); этот фрагмент можно также выделить известными способами из мышиной геномной ДНК с применением праймеров ПЦР, сконструированных на основании анализа последовательностей mMT, доступных из Genbank; т.е. MUS-MTI, MUSMTIP, MUSMTIPRM] делают фрагментом с затупленными концами при помощи известных способов и лигируют с расщепленной SfiI (также с затупленными концами) или расщепленной FSpI плазмидой р5'ЕРО. Анализируют ориентацию полученных клонов и те клоны, в которых первый сайт Bgl II mMT является проксимальным к сайту SalI в плазмидном полилинкере, используют для нацеливания первичных и вторичных фибробластов человека. Эта ориентация направляет транскрипцию mMT в направлении положения нуклеотида HUMERPA 610 в конечной конструкции. Полученные плазмиды обозначены р5'ЕРО-mMTF и p5'EPO-mMTS для встраивания mMT в сайты FSpI и Sfi I, соответственно.
Дополнительные последовательности против хода транскрипции применимы в случаях, когда желательно модифицировать, делегировать и/или заменить негативные регуляторные элементы или энхансеры, которые лежат против хода транскрипции от исходной последовательности-мишени. В случае ЕРО может быть делегирован негативный регуляторный элемент, который ингибирует экспрессию ЕРО во внепеченочных и внепочечных тканях [Semenza, J.Z., et al., mol. all Biol. 10:930938 (1990)]. Получен ряд делеций во фрагменте 6 т.п.н. Делетированные районы могут быть заменены энхансером с широкой активностью (спектром) клеток-хозяев [например, энхансером из Cytomegalovirus (cmv)].
Ориентация фрагменга 5'ЕРО 609 т.п.н. в векторе pBluescript II Sk/+ была выбрана, т.к. перед последовательностями HUMERPA на их 5'-конце находится сайт BamHI (дистальный) и сайт Hind III (проксимальный). Таким образом, фрагмент 6 т.п.н. BamHI-Hind III, обычно лежащий против хода транскрипции от фрагмента 609 п.н. [Semenza, J.Z. et al., Mol. Cell Biol. 10:930-938 (1990)], может быть выделен из геномной ДНК известными способами. Например, бактериофаговую, космидную или дрожжевую искусственную хромосомную библиотеку можно скринировать амплифицированным при помощи ПЦР фрагментом 609 п.н. в качестве зонда. Желаемый клон будет иметь фрагмент 6 т.п.н. BamHI-HindIII и его идентичность может быть подтверждена сравнением его рестрикционной карты с рестрикционной картой вокруг гена ЕРО человека, определенной известными способами. Альтернативно, конструирование рестрикционной карты генома человека против хода транскрипции от гена ЕРО с использованием фрагмента 609 п.н. в качестве зонда может идентифицировать ферменты, которые образуют фрагмент, начинающийся между координатами HUMERPA 2 и 609 и простирающийся за сайт BamHI против хода транскрипции; этот фрагмент можно выделить при помощи гель-электрофореза из подходящего продукта расщепления геномной ДНК человека и лигировать в бактериальный или дрожжевой клонирующий вектор. Точный клон будет гибридизоваться с 609 п.н. 5'ЕРО-зондом и содержать фрагмент 6 т.п.н. BamHI-Hind III. Выделенный фрагмент 6 т.п.н. встраивают в правильной ориентации в р5'ЕРО, р5'EPOmMTF или р5'ЕРО-mMTS (так, чтобы сайт Hind III находился рядом с нуклеотидным положением HUMERPA 2). Дополнительные последовательности против хода транскрипции могут быть выделены известными способами, использующими способы прогулки по хромосоме, или выделением дрожжевых искусственных хромосом, гибридизующихся с зондом 609 п.н. 5'ЕРО.
Стратегии клонирования, описанные выше, позволяют модифицировать in vitro последовательности против хода транскрипции ЕРО для последующей направленной трансфекции первичных, вторичных или иммортализованных фибробластов человека. Эти стратегии описывают простые встраивания промотора mMT, а также делецию негативного регуляторного района и делецию негативного регуляторного района и замену его энхансером с широкой активностью клеток-хозяев.
g. Активация гена ЕРО человека и выделение получивших нацеленные конструкции ДНК первичных, вторичных и иммортализованных фибробластов человека при помощи скрининга
Для нацеливания плазмиды разрезают рестриктазами для освобождения вставки (инсерции) от плазмидного каркаса. В случае плазмиды р5'EPO-mMTS расщепление Hind III и Sal I высвобождает направляющий фрагмент 2,4 т.п.н., состоящий из промотора mMT 1,8 т.п.н., фланкированного на 5'- и 3'-сторонах последовательностями из 405 п.н. и 204 п.н., соответственно, ДНК для направления этой конструкции к регуляторному району гена ЕРО человека. Эту ДНК или направляющий фрагмент 2,4 т.п.н. очищают фенольной экстракцией и осаждением этанолом и трансфицируют в первичные или вторичные фибробласты человека при условиях, описанных в Примере 1с. Трансфицированные клетки высевают на чашки с диаметром 150 мм в питательную среду для фибробластов человека. Через 48 часов эти клетки высевают в 24-луночные чашки при плотности 10000 клеток/см2 [приблизительно 20000 клеток на лунку; если нацеливание происходит при скорости 1 событие нацеливания на 106 клонируемых клеток (Пример 1с), то приблизительно 50 лунок нужно будет проанализировать для выделения одной экспрессирующей колонии]. Клетки, в которых трансфицирующая ДНК попала в гомологичный район против хода транскрипции гена ЕРО человека, будут экспрессировать hEPO под контролем промотора mMT. После 10 дней луночные супернатанты тестируют на экспрессию ЕРО с использованием коммерчески доступного набора для иммунотеста (Amgen). Клоны из лунок, обнаруживающих синтез hEPO, выделяют известными способами, обычно тестированием фракций гетерогенных популяций клеток, распределенных по отдельным лункам, тестированием фракций позитивных лунок с повторением, если это необходимо, и выделением в конце концов колонии со встроенной ДНК путем скрининга 96-луночных микротитрационных планшетов, засеянных при концентрации одна клетка на лунку. ДНК из лизатов всего планшета может быть также подвергнута анализу с применением ПЦР на амплификацию фрагмента и применением mMT-специфического праймера вместе с праймером, лежащим против хода транскрипции от положения нуклеотида HUMERPA 1. Эта пара праймеров должна амплифицировать фрагмент ДНК с размером, точно предсказанным на основе последовательности ДНК. Позитивные чашки трипсинизируют и пересевают при последовательно более низких разведениях и получение ДНК и стадии ПЦР повторяют для выделения получивших нацеленные конструкции ДНК клеток.
Схемы нацеливания, описанные здесь, можно также использовать для активации экспрессии hGH в иммортализованных клетках человека (например, клетках НТ1080 (АТСС CCL 121), клетках HeLa и производных клеток HeLa (АТСС CCL 2,2.1 и 2.2), клетках рака молочной железы MCF-7 (АТСС НВТ 22), клетках лейкоза К-562 (АТСС CCL 232), клетках карциномы (АТСС CCL 17), клетках рака яичника 2780AD (Van Der Blick, A.M., et al., Cancer Res., 48:5927-5932 (1988), клетках Raji (АТСС CCL 86), клетках Jurkat (АТСС T1B 152), клетках Namalwa (АТСС CRL 1432), клетках HL-60 (АТСС CCL 240), клетках Daudi (АТСС CCL 213), клетках RPM1 8226 (АТСС CCL 155), клетках U-937 (АТСС CRL 1593), клетках меланомы Боуеса (АТСС CRL 9607), подлинии 2R4 клеток W1-38VА13 (АТСС CLL 75.1), клетках MOLT-4 (АТСС CRL 1582) и разнообразных гетерогибридомных клетках) для целей получения hGH для общепринятой фармацевтической доставки.
h. Активация гена ЕРО человека и выделение получивших нацеленные конструкции ДНК первичных, вторичных и иммортализованных фибробластов человека при помощи системы позитивного или комбинированного позитивного/негативного отбора
Стратегия конструирования плазмид р5'ЕРО-mMTF, р5'ЕРО-mMTS и их производных с дополнительным фрагментом BamH I-Hind III 6 т.п.н. против хода транскрипции может иметь дополнительную стадию встраивания гена neo рядом с промотором mMT. Кроме того, негативный селектируемый маркер, например gpt [из pMSG (Pharmacia) или другого подходящего источника] может быть встроен рядом с HUMERPA последовательностями в pBluescript IISk/+ полилинкере. В первом случае G418r-колонии выделяют и скринируют с применением ПЦР-амплификации или рестрикционного и гибридизационного анализа по Саузерну ДНК, полученной из пулов колоний, для идентификации получивших нацеленные конструкции колоний. В последнем случае G418r-колонии помещают в среду, содержащую 6-тиоксантин, для отбора против интеграции гена gpt (негативный отбор) [Besnard, C. et al., Mol. Cell Biol. 7:4139-4141 (1989)]. Кроме того, ген HSV-ТК может быть помещен на противоположной стороне этой инсерции в виде gpt, что позволяет проводить отбор на neo и против как gpt, так и ТК растущими клетками в среде для фибробластов человека, содержащей 400 мкг/мл G418, 100 мкМ 6-тиоксантин и 25 мкг/мл ганцикловира. Двойной негативный отбор обеспечивает почти абсолютный отбор на истинные события нацеливания, а блоттинг по Саузерну обеспечивает окончательное подтверждение.
Описанные здесь схемы нацеливания для гомологичной рекомбинации могут быть также использованы для активации экспрессии hEPO в иммортализованных клетках человека (например, клетках НТ1080 (АТСС CCL 121), клетках HeLa и производных клеток HeLa (АТСС CCL 2, 2,1 и 2,2), клетках рака молочной железы MCF-7 (АТСС НВТ 22), клетках лейкоза К-562 (АТСС CCL 232), клетках карциномы KB (АТСС CCL 17), клетках рака яичника 2780-AD (Van der Blick, A.M., et al., Cancer Res., 48:5927-5932 (1988), клетках Raji (АТСС CCL 86), клетках Jurkat (атсс Т1В 152), клетках Namalwa (АТСС CRL 1432), клетках HL-60 (АТСС CCL 240), клетках Daudi (АТСС CCL 213), клетках RРМ1 8226 (АТСС CCL 155), клетках U-937 (АТСС CRL 1593), клетках меланомы Bowes (АТСС CRL 9607), подлинии 2R4 W1-38VA13 (АТСС CLL 75.1), клетках MOLT-4 (АТСС CLL 1582) и разнообразных гетерогибридомных клетках) для целей получения hEPO для общепринятой фармацевтической доставки.
i. Конструирование направляющих (нацеливающих) плазмид для помещения гена гормона роста человека под контроль мышиного промотора металлотионеина в первичных, вторичных или иммортализованных фибробластах человека
Следующий пример служит для иллюстрации одного варианта данного изобретения, в котором обычные регуляторные последовательности против хода транскрипции от гена гормона роста человека изменяют для обеспечения экспрессии гормона роста человека в первичных, вторичных или иммортализованных фибробластах человека.
Направляющие молекулы, похожие на описанные в Примере 1f, для нацеливания на регуляторный район гена ЕРО получают с применением клонированных фрагментов ДНК, полученных из 5'-конца гена N гормона роста человека. Фрагмент с длиной приблизительно 1,8 т.п.н., охватывающий положения нуклеотидов HUMGHCSA ( Genbank Entry) 3787-5432 (положения двух сайтов EcoRI, которые образуют фрагмент удобного размера для клонирования или диагностического расщепления включающих в себя этот фрагмент субклонов), амплифицируют при помощи праймеров ПЦР, сконструированных согласно анализу последовательности HUMGHCSA в этом районе. Этот район простирается от середины интрона 1 гена NhGH до положения против хода транскрипции приблизительно 1,4 т.п.н. 5' по отношению к сайту старта трансляции. pUC12 расщепляют EcoRI и BamHI, обрабатывают с фрагментом Кленова для образования тупых концов и повторно замыкают в кольцо при разбавленных условиях, получая плазмиды, которые потеряли сайты EcoRI и BamHI. Такая плазмида названа рUС12ХЕВ. Hind III-линкеры лигируют на амплифицированный фрагмент hGH и полученный фрагмент расщепляют Hind III и лигируют с расщепленной Hind III плазмидой pUC12XEB. Полученную плазмиду, рUС12ХЕВ-5'hGH, расщепляют EcoFRI и BamHI для удаления фрагмента 0,5 т.п.н., лежащего непосредственно против хода транскрипции от инициирующего сайта транскрипции hGH. Расщепленную ДНК лигируют с фрагментом EcorI-Bgl II 1,8 т.п.н. из гена mMT1 [не содержащего кодирующих mMT последовательностей; Hamer, D.H., and Walling, M., J. Mol. Appl. Gen. 1:273-288 (1982); этот фрагмент можно также выделить известными способами из мышиной геномной ДНК с применением ПЦР-праймеров, сконструированных на основании анализа последовательностей mMT, доступных из Genbank, т.е. MUSMTI, MUSMTIP, MUSMTIPRM]. Эта плазмида р5' hGH-mMT имеет промотор mMT, фланкированный на обеих сторонах расположенными против хода транскрипции последовательностями hGH.
Описанные здесь стратегии клонирования позволяют модифицировать in vitro последовательности против хода транскрипции от hGH для последующей направленной трансфекции первичных, вторичных или иммортализованных фибробластов человека. Эта стратегия описала простое встраивание промотора mMT. Могут быть рассмотрены другие стратегии, например, в которых энхансер с широкой специфичностью клеток-хозяев встраивают против хода транскрипции от встроенной последовательности mMT.
j. Активация гена hGH человека и выделение получивших нацеленные конструкции ДНК первичных, вторичных и иммортализованных фибробластов человека при помощи скрининга
Для нацеливания, плазмиды разрезают рестриктазами, которые освобождают инсерцию от плазмидного каркаса. В случае плазмиды р5' hGH-mMT расщепление Hind III высвобождает нацеливающий фрагмент 2,9 т.п.н., состоящий из промотора mMT 1,8 т.п.н., фланкированного на 5'- и 3'-сторонах ДНК, предназначенной для направления этой конструкции к регуляторному району гена hGH. Эту ДНК или только нацеливающий фрагмент 2,9 т.п.н. очищают фенольной экстракцией и осаждением этанолом и трансфицируют в первичные или вторичные фибробласты человека при условиях, описанных в Примере 1. Трансфицированные клетки высевают на чашки с диаметром 150 мм в питательную среду для фибробластов человека. Через 48 часов клетки высевают в 24-луночные чашки при плотности 10000 клеток/см2 [приблизительно 20000 клеток на лунку; если нацеливание происходит при скорости 1 событие на 106 клонируемых клеток (Пример 1с), то нужно анализировать приблизительно 50 лунок для выделения одной экспрессирующей колонии]. Клетки, в которых трансфицирующая ДНК направлялась к гомологичному району против хода транскрипции от hGH, будут экспрессировать hGH под контролем промотора mMT. После 20 дней все луночные супернатанты анализировали на экспрессию hGH с использованием коммерчески доступного набора для иммунотеста (Nichols). Клоны из лунок, обнаруживающих синтез hGH, выделяют известными способами, обычно тестированием фракций гетерогенных популяций клеток, распределенных в отдельные лунки или планшеты, анализом этих позитивных лунок и повторением этого, если необходимо, и в конце концов выделяют колонии, получившие нацеленные конструкции ДНК, посредством скрининга 96-луночных микротитрационных планшетов, засеянных по одной клетке на лунку. ДНК из полных лизатов планшетов можно также анализировать при помощи ПЦР для амплификации фрагмента с использованием mMT-специфического праймера вместе с праймером, лежащим по ходу транскрипции от положения 5432 HUMGHCSA. Эта пара праймеров должна амплифицировать фрагмент ДНК с размером, точно предсказанным на основе последовательности ДНК. Позитивные чашки трипсинизировали и повторно засевали при последовательно снижающихся разведениях, и получение ДНК и стадии ПЦР повторяли, если необходимо, для выделения получивших нацеленную конструкцию ДНК клеток.
Схемы нацеливания, описанные здесь, можно также использовать для активации экспрессии hGH в иммортализованных клетках человека (например, клетках НТ1080 (АТСС CCL 121), клетках HeLa и производных клеток HeLa (АТСС CCL 2, 2.1 и 2.2), клетках рака молочной железы MCF-7 (АТСС НВТ 22), клетках лейкоза К-562 (АТСС CCL 232), клетках карциномы KB (АТСС CCL 17), клетках рака яичника 2780AD (Van der Blick, A.M., et al., Cancer Res. 48:5927-5932 (1988), клетках Raji (АТСС CCL 86), клетках Jurkat (АТСС Т1В 152), клетках Namalwa (АТСС CRL 1432), клетках HL-60 (АТСС CCL 240), клетках Daudi (АТСС CCL 213), клетках RPM1 8226 (АТСС CCL 155), клетках U-937 (АТСС CRL 1593), клетках меланомы Bowes (АТСС CLL 9607), подлинии 2R4 клеток W1-38VA13 (АТСС CRL 75.2), клетках MOLT-4 (АТСС CRL1582) и разнообразных гетерогибридомных клетках) для целей получения hGH для общепринятой фармацевтической доставки.
к. Активация гена hGH человека и выделение получивших нацеленные конструкции ДНК первичных, вторичных и иммортализованных фибробластов человека при помощи системы позитивного или комбинированного позитивного/негативного отбора
Стратегия конструирования плазмиды р5'hGH-mMT может иметь дополнительную стадию встраивания гена neo рядом с промотором mMT. Кроме того, маркер негативного отбора, например gpt [из pMSG (Pharmacia) или другого подходящего источника], может быть встроен рядом с HUMGHCSA последовательностями в рUС12 полилинкер. В первом случае G418r-колонии выделяют и скринируют при помощи ПЦР-амплификации или рестрикционного и гибридизационного анализа по Саузерну ДНК, полученной из пулов колоний для идентификации получивших нацеленные конструкции колоний.
В последнем случае G418r-колонии помещают в среду, содержащую тиоксантин, для отбора против интеграции гена gpt (негативный отбор) (Besnard, C. et al., Mol. Cell Biol. 7:4139-4141 (1987)]. Кроме того, ген HSV-ТК может быть помещен на противоположной стороне инсерции в виде gpt, что позволяет проводить отбор на neo и против как gpt, так и ТК растущими клеткам в питательной среде для фибробластов человека, содержащей 400 мкг/мл G418, 100 мкМ 6-тиоксантин и 25 мкг/мл ганцикловира. Двойной негативный отбор должен обеспечивать почти абсолютный отбор на правильные события нацеливания. Гибридизационный анализ по Саузерну является подтверждающим.
Описанные здесь схемы нацеливания могут быть использованы также для активации экспрессии hGH в иммортализованных клетках человека (например, клетках НТ1080 (ATCC CCL 121), клетках HeLa и производных клеток HeLa (АТСС CCL 2, 2.1 и 2.2), клетках рака молочной железы MCF-7 (АТСС НВТ 22), клетках лейкоза К-562 (АТСС CCL 232), клетках карциномы KB (ATCC CCL 17), клетках рака яичника 2780АD (Van der Blick, A.M. et al., Cancer Res. 48:5927-5932 (1988), клетках Raji (АТСС CCL 86), клетках Jurkat (АТСС Т1В 152), клетках Namalwa (ATCC CRL 1432), клетках HL-60 (АТСС CCL 240), клетках Daudi (ATCC CCL 213), клетках RPM1 8226 (АТСС CCL 155), клетках U-937 (АТСС CRL 1593), клетках меланомы Bowes (АТСС CRL 9607), клетках сублинии 2R4 W1-38 V А13 (АТСС CLL 75.1), клетках MOLT-4 (ATCC CRL 1582) и разнообразных гетерогибридомных клетках) для целей получения hSH для общепринятой фармацевтической доставки.
Направляющие конструкции, описанные в Примерах 1f и 1i и использованные в Примерах 1g, 1h, 1i и 1j, могут быть модифицированы для включения в них амплифицируемого селектируемого маркера (например, ada, dhfr или CAD), который применим для отбора клеток, в которых амплифицируются активированный эндогенный ген и амплифицируемый селектируемый маркер. Такие клетки, экспрессирующие или способные экспрессировать эндогенный ген, кодирующий терапевтический продукт можно использовать для получения белков (например, hGH и hEPO) для общепринятой фармацевтической доставки или для генной терапии.
l. Трансфекция первичных и вторичных фибробластов экзогенной ДНК и селектируемым маркерным геном посредством электропорации
Экспоненциально растущие фибробласты или фибробласты ранней стационарной фазы трипсинизируют и споласкивают (смывают) с пластиковой поверхности питательной средой. Аликвоту этой клеточной суспензии берут для счета, а остальные клетки подвергают центрифугированию. Супернатант отсасывают и осадок ресуспендируют в 5 мл буфера для электропорации (20 мМ HEPES рН 7,3, 137 мМ NaCl, 5 мМ KCl, 0,7 мМ Na2HPO4, 6 мМ декстроза). Эти клетки повторно центрифугируют, супернатант отсасывают и клетки ресуспендируют в буфере для электропорации, содержащем 1 мг/мл ацетилированного бычьего сывороточного альбумина. Конечная клеточная суспензия содержит приблизительно 3×106 клеток/мл. Электропорация должна проводиться сразу после ресуспендирования.
Суперспирализованную плазмидную ДНК добавляют в стерильную кювету с интервалом между электродами 0,4 см (Bio-Rad). Конечная концентрация ДНК обычно по меньшей мере 120 мкг/мл. Затем в кювету добавляют 0,5 мл клеточной суспензии (содержащей приблизительно 1,5×106 клеток) и клеточную суспензию и растворы ДНК осторожно перемешивают. Электропорацию проводят в устройстве Gene-Pulser (Bio-Rad). Емкость и напряжение устанавливают на 960 мкФ и 250-300 В, соответственно. По мере увеличения напряжения выживание клеток уменьшается, но процент выживших клеток, которые стабильно включают введенную ДНК в их геном, сильно увеличивается. С учетом этих параметров должен наблюдаться период импульсов 14-20 мс.
Подвергнутые электропорации клетки поддерживают при комнатной температуре в течение приблизительно 5 минут и затем содержимое кюветы осторожно удаляют стерильной пипеткой. Клетки добавляют непосредственно к 10 мл предварительно нагретой питательной среды (с 15% телячьей сывороткой, как указано выше) в чашку с диаметром 10 см и инкубируют, как описано выше. На следующий день среду отсасывают и заменяют 10 мл свежей среды и инкубируют еще 16-24 часа. Субкультивирование клеток для определения эффективности клонирования и для отбора устойчивых с G418 колоний выполняют на следующий день. Клетки трипсинизируют, считывают и высевают; обычно фибробласты высевают при 103 клеток/10 см-чашку для определения эффективности клонирования и при 1-2×104 клеток на 10 см-чашку для G418-отбора.
Фибробласты человека отбирают на устойчивость к G418 в среде, состоящей из 300-400 мкг/мл G418 (Geneticin, соль дисульфат, приблизительно 50% раствор; Gibco) в питательной среде для фибробластов (с 15% телячьей сывороткой). Эффективность клонирования определяют в отсутствие G418. Посеянные клетки инкубируют в течение 12-14 дней, затем колонии фиксируют формалином, окрашивают кристаллическим фиолетовым красителем и считывают (для посеянной эффективности клонирования) или выделяют при помощи клонирующих цилиндров (для чашек с G418). Электропорацию и отбор фибробластов кролика выполняют в основном, как описано для фибробластов человека, за исключением того, что используют другие условия отбора. Кроличьи фибробласты отбирают на устойчивость к G418 в среде, содержащей 1 г/мл G418.
Фибробласты выделяют из свежесрезанной крайней плоти человека. Культуры засевают при 50000 клеток/см в среде DMEM + 10% телячья сыворотка. Когда культуры становятся конфлюэнтными, фибробласты собирают трипсинизацией и трансфицируют посредством электропорации. Условия электропорации оценивали с использованием трансфекции плазмидой pcDNEO (фиг.5). Характерный эксперимент по электропорации с использованием приблизительно оптимальных условий (60 мкг плазмиды рс DNEO при напряжении электропорации 250 В и емкости, установленной на 960 мкФ) приводил к одной G418r-колонии на 588 обработанных клеток (0,17% от всех обработанных клеток) или 1 G418r-колоний на 71 клонируемую клетку (1,4%).
При проведении 9 отдельных электропораций при близких к оптимальным условиям (60 мкг плазмиды pcDNEO при напряжении при электропорации 300 В и установке емкости при 960 мкФ) наблюдали в среднем одну G418r-колонию на 1899 обработанных клеток (0,05%) с диапазоном 1/882-1/7500 обработанных клеток. Это соответствует в среднем одной G418r-колонии на 38 клонируемых клеток (2,6%).
Первичные фибробласты человека с низким числом пассажей превращали в экспрессирующие hGH клетки котрансфекцией с плазмидами pcDNEO и pXGH5. В типичном случае 60 мкг эквимолярной смеси двух плазмид трансфицировали при близких к оптимальным условиям (электропорация при напряжении 300 В и емкости 960 мкФ). Результаты такого эксперимента дали одну G418-колонию на 14705 обработанных клеток.
Данные по экспрессии hSH для этих и других клеток, выделенных при идентичных условиях трансфекции, суммированы ниже. В конечном счете, 98% всех G418-колоний можно было размножать для образования массовой культуры.
Стабильные трансфекты можно было получить также электропорацией первичных или вторичных фибробластов человека с pXGH301, конструкцией ДНК, в которой гены neo и hGH присутствуют на одной и той же плазмидной молекуле. Плазмиду pXGH301 конструировали двухступенчатой процедурой. Фрагмент SalI-ClaI из pBR322 (положения 23-651 в pBR322) выделяли и встраивали в расщепленную SalI-ClaI плазмиду pcDNEO, вводя сайт Bam HI против хода транскрипции от района раннего промотора SV40 pcDNEO. Эту плазмиду pBNEO расщепляли Bam HI и фрагмент 2,1 т.п.н., содержащий ген neo под контролем раннего промотора SV40, выделяли и встраивали в расщепленную BamHI pXGH5. Выделяли плазмиду с одной вставкой фрагмента 2,1 т.п.н. BamHI, в которой neo и hGH транскрибируются в одном и том же направлении по отношению друг к другу. Эта плазмида была названа рХGН301. Например, 1,5×106 клеток подвергали электропорации с 60 мкг рХGН301 при 300 В и 960 мкФ. G418r-колонии выделяли из трансфицированных вторичных фибробластов при частоте 652 G418r-колоний на 1,5×106 обработанных клеток (1 на 2299 обработанных клеток). Приблизительно 59% этих колоний экспрессируют hGH.
ПРИМЕР 2. Конструирование нацеливающих плазмид, приводящих к образованию химерных транскрипционных единиц, в которых слиты последовательности ДНК гормона роста человека и эритропоэтина
Дальнейшее описание служит для иллюстрации двух следующих вариантов данного изобретения, в которых обычные регуляторные последовательности против хода транскрипции от гена ЕРО человека изменяют для обеспечения экспрессии hEPO в первичных или вторичных штаммах фибробластов, которые обычно не экспрессируют hEPO в детектируемых количествах в их нетрансфицированном состоянии. В этих вариантах продукты событий нацеливания представляют собой химерные транскрипционные единицы, в которых первый экзон гена гормона роста человека расположен против хода транскрипции экзонов 2-5 hEPO. Продукт транскрипции, сплайсинга и трансляции является белком, в котором аминокислоты 1-4 сигнального пептида hEPO заменены аминокислотными остатками 1-3 hGH. Эти два варианта отличаются в отношении обоих относительных положений чужеродных регуляторных последовательностей, которые встроены, и в отношении специфического характера сплайсинга, который должен иметь место для получения конечного процессированного транскрипта.
Плазмида рХЕРО-10 сконструирована для замены экзона 1 hEPO экзоном 1 hGH посредством генного нацеливания на эндогенный ген hEPO на хромосоме 7 человека. Плазмиду рХЕРО-10 конструируют следующим образом. Сначала конструируют промежуточную плазмиду рТ163 встраиванием фрагмента 6 т.п.н. Hind III - BamHI (см. Пример 1f), лежащего против хода транскрипции от кодирующего района hEPO, в расщепленную Hind III - Bam HI плазмиду pBluescript IISk+ (Stratagene, La Jolla, CA). Продукт этого лигирования расщепляют XhoI и HindIII и лигируют с фрагментом 1,1 т.п.н. HindIII-XhoI из pMC1 neo Poly A [Thomas, K.R. and Capecchi, M.R. Cell 51:503-512 (1987), доступной из Stratagene, La Jolla, CA], для создания рТ163. Олигонуклеотиды 13.1-13.4 используют в полимеразных цепных реакциях для образования слитого фрагмента, в котором последовательности мышиного промотора металлотионеина 1 (mMT-1) - экзона 1 hGH дополнительно слиты с последовательностями интрона 1 hEPO. Сначала олигонуклеотиды 13.1 и 13.3 используют для амплификации фрагмента промотор приблизительно 0,73 т.п.н. тМТ - экзон 1 hGH из рХGН5 (фиг.5). Затем олигонуклеотиды 13.2 и 13.4 используют для амплификации фрагмента приблизительно 0,57 т.п.н., содержащего преимущественно интрон 1 hEPO из геномной ДНК человека. Наконец, эти два амплифицированных фрагмента смешивают и амплифицируют далее с олигонуклеотидами 13.1 и 13.4 для образования конечного слитого фрагмента (слитого фрагмента 3), фланкированного сайтом SalI на 5'-стороне mMT-1-части и сайта-XhoI на 3'-стороне последовательности интрона 1 hEPO. Слитый фрагмент 3 расщепляют XhoI и SalI лигируют с расщепленной XhoI плазмидой рТ163. Смесь для лигирования трансформируют в E.coli и клон, содержащий одну инсерцию слитого фрагмента 3, в котором сайт XhoI регенерирован на 3'-стороне последовательностей интрона 1 hEPO, идентифицируют и обозначают как рХЕРО-10.

He выделенный жирными буквами район олигонуклеотида 13.1 идентичен промотору mMT-1, с природным сайтом Kpn I в виде его 5'-границы. Жирные буквы обозначают конец сайта SalI для превращения 5'-границы в сайт SalI. Жирный район олигонуклеотидов 13.2 и 13.3 обозначает последовательности hSH, тогда как нежирные районы представляют собой последовательности интрона 1 из гена hEPO. Нежирный район олигонуклеотида 13.4 идентичен последним 25 основаниям интрона 1 hEPO. Жирный район включает в себя конец сайта XhoI для превращения 3'-границы амплифицированного фрагмента в сайт XhoI.
Плазмида рХЕРО-11 сконструирована для помещения, генным нацеливанием, промотора mMT-1 и экзона 1 hGH против хода транскрипции от структурного гена hEPO и промоторного района при эндогенном локусе hEPO на хромосоме 7 человека. Плазмиду рХЕРО-11 конструируют следующим образом. Олигонуклеотиды 13.1 и 13.5 - 13.7 используют в полимеразных цепных реакциях для образования слитого фрагмента, в котором последовательности промотора мышиного металлотионеина 1 (mMT-1) - экзона 1 hGH дополнительно слиты с последовательностями hEPO от -1 до -630 по отношению к кодирующему району hEPO. Сначала олигунуклеотиды 13.1 и 13.6 используют для амплификации фрагмента промотор mMT-1 приблизительно 0,75 т.п.н. - экзон 1 hGH из рХGH5 (фиг.5). Затем олигонуклеотиды 13.5 и 13.7 используют для амплификации, из геномной ДНК человека, фрагмента приблизительно 0,65 т.п.н., состоящего преимущественно из последовательностей hEPO от -1 до -620 по отношению к кодирующему району hEPO. Оба олигонуклеотида 13.5 и 13.6 содержат линкерную последовательность 10 п.н., расположенную при районе интрон 1 hGH - промотор hEPO, который соответствует донорному сайту сплайсинга. Наконец, эти два амплифицированных фрагмента смешивают и амплифицируют далее с олигонуклеотидами 13.1 и 13.7 для образования конечного слитого фрагмента (слитого фрагмента 6), фланкированного сайтом SalI при 5'-стороне mMT-1-части и сайтом XhoI при 3'-стороне района промотора hEPO. Слитый фрагмент 6 расщепляют XhoI и SalI и лигируют с расщепленной XhoI плазмидой рТ163. Смесь для лигирования трансформируют в E.coli и клон, содержащий одну инсерцию слитого фрагмента 6, в которой сайт XhoI регенерирован при 3'-стороне последовательностей промотора hEPO, идентифицируют и обозначают как рХЕРО-11.

Районы с жирными буквами олигонуклеотидов 13.5, 13.6 обозначают последовательности hGH. Районы с буквами курсивом соответствуют первым 10 п.н. интрона 1 hEPO. Остальная часть этих олигонуклеотидов соответствует последовательностям hEPO от -620 до -597 по отношению к кодирующему району hEPO. Нежирный район олигонуклеотида 13.7 идентичен основаниям (-1) - (-24) по отношению к кодирующему району hEPO. Район с жирными буквами включает в себя конец сайта XhoI для превращения 2'-границы амплифицированного фрагмента в сайт XhoI.
Плазмиду рХЕРО-10 можно использовать для генного нацеливания расщеплением BamI и XhoI для высвобождения фрагмента 7,3 т.п.н., содержащего слитый фрагмент mMT-1/hGH, фланкированный на обеих сторонах последовательностями hEPO. Этот фрагмент (нацеливающий фрагмент 1) не содержит кодирующих последовательностей hEPO, имея только последовательности, лежащие между -620 и приблизительно -6620 против хода транскрипции от кодирующего района hEPO, и последовательности интрона 1 hEPO для регуляции нацеливания на локусе ЕРО человека. Нацеливающий фрагмент 1 трансфицируют в первичные или вторичные фибробласты кожи человека при условиях, сходных с описанными в Примере 1с. Устойчивые к G418 колонии выскребают в отдельные лунки 96-луночных микротитрационных планшетов и скринируют на экспрессию ЕРО при помощи твердофазного иммуноферментного анализа (ELISA) (R&D Systems, Minneapolis MN). Клетки, в которых трансфицированная ДНК интегрируется произвольно в геном человека, не могут продуцировать ЕРО. Клетки, в которых трансфицированная ДНК подверглась гомологичной рекомбинации с интроном 1 эндогенного hEPO и последовательностями hEPO против хода транскрипции, содержат химерный ген, в котором последовательности промотора mMT-1 и нетранскрибируемые последовательности и 5'-нетранслируемые последовательности hGH и экзон 1 hGH заменяют обычный промотор hEPO и экзон 1 hEPO (см. Фиг.1). Последовательности, не относящиеся к hEPO, в нацеливающем фрагменте 1 соединены с последовательностями hEPO по ходу транскрипции интрона 1 hEPO. Замена нормального регуляторного района hEPO промотором mMT-1 будет активировать ген ЕРО в фибробластах, которые обычно не экспрессируют hEPO. Замена экзона 1 hEPO экзоном 1 hGH приводит к образованию белка, в котором первые 4 аминокислоты сигнального пептида hEPO заменены аминокислотами 1-3 hGH, что создает функциональный химерный сигнальный пептид, который удаляется посттрансляционным процессингом из зрелого белка и секретируется из экспрессирующих клеток.
Плазмиду рХЕРО-11 можно использовать для генного нацеливания расщеплением BamHI и XhoI для высвобождения фрагмента 7,4 т.п.н., содержащего слияние mMT-1/hGH, фланкированное на обеих сторонах, последовательностями hEPO. Этот фрагмент (нацеливающий фрагмент 2) не содержит кодирующих последовательностей hEPO, имея только последовательности, лежащие между -1 и приблизительно -6620 против хода транскрипции от кодирующего района hEPO, для управления нацеливанием на локус ЕРО человека. Нацеливающий фрагмент 2 трансфицируют в первичные или вторичные фибробласты кожи человека при условиях, сходных с описанными в Примере 1g. Устойчивые к G418 колонии помещают в отдельные лунки 96-луночных планшетов и скринируют на экспрессию ЕРО при помощи твердофазного иммуноферментного анализа (ELISA) (R&D Systems, Minneapolis, MN). Клетки, в которых трансфицирующая ДНК интегрируется в геном человека произвольно, не могут продуцировать ЕРО. Клетки, в которых трансфицирующая ДНК подверглась гомологичной рекомбинации с эндогенным промотором hEPO и последовательностями против хода транскрипции, содержат химерный ген, в котором промотор mMT-1 и нетранскрибируемые последовательности, 5'-нетранслируемые последовательности hGH и экзон 1 hGH и линкер из 10 п.н., состоящий из первых 10 оснований интрона 1 hEPO, встроены при сайте HindIII, лежащем при положении -620 по отношению к кодирующему району hEPO (см. фиг.2). Локализация промотора mMT-1 против хода транскрипции от обычно молчащего промотора hEPO будет управлять синтезом, в первичных или вторичных фибробластах кожи человека, мРНК, считывая  нетранслируемые последовательности металлотионеина и hGH, экзон 1 hGH, 10 оснований ДНК, идентичных первым 10 основаниям интрона 1 hEPO, и обычный промотор hEPO и экзон 1 hEPO (-620 - +13 по отношению к кодирующей последовательности hEPO). Линкерная последовательность из 10 п.н. из интрона 1 hEPO действует как донорный сайт сплайснга для слияния экзона 1 hGH со следующим по ходу транскрипции акцепторным сайтом сплайсинга, который лежит непосредственно против хода транскрипции от экзона 2 hEPO. Процессинг образующегося транскрипта будет, следовательно, сплайсировать промотор hEPO, экзон 1 и последовательности интрона 1. Замена экзона 1 hEPO экзоном 1 hGH приводит к образованию белка, в котором первые 4 аминокислоты сигнального пептида hEPO заменены аминокислотами 1-3 hGH, что создает функциональный, химерный сигнальный пептид, который удаляется пост-трансляционным процессингом из зрелого белка и секретируется из экспрессирующих клеток.
нетранслируемые последовательности металлотионеина и hGH, экзон 1 hGH, 10 оснований ДНК, идентичных первым 10 основаниям интрона 1 hEPO, и обычный промотор hEPO и экзон 1 hEPO (-620 - +13 по отношению к кодирующей последовательности hEPO). Линкерная последовательность из 10 п.н. из интрона 1 hEPO действует как донорный сайт сплайснга для слияния экзона 1 hGH со следующим по ходу транскрипции акцепторным сайтом сплайсинга, который лежит непосредственно против хода транскрипции от экзона 2 hEPO. Процессинг образующегося транскрипта будет, следовательно, сплайсировать промотор hEPO, экзон 1 и последовательности интрона 1. Замена экзона 1 hEPO экзоном 1 hGH приводит к образованию белка, в котором первые 4 аминокислоты сигнального пептида hEPO заменены аминокислотами 1-3 hGH, что создает функциональный, химерный сигнальный пептид, который удаляется пост-трансляционным процессингом из зрелого белка и секретируется из экспрессирующих клеток.
Ряд конструкций, относящихся к рХЕРО-10 и рХЕРО-11, можно сконструировать при помощи известных способов. В этих конструкциях относительные положения промотора mMT-1 и последовательностей hGH, а также положение, при котором последовательности mMT-1/hGH встраивают в последовательности против хода транскрипции hEPO, вариируют для создания других химерных транскрипционных единиц, которые облегчают генное нацеливание, приводят к более эффективной экспрессии слитых транскриптов или имеют другие желаемые свойства. Такие конструкции будут давать сходные результаты, так что слитый ген hGH-EPO помещают под контроль экзогенного промотора генным нацеливанием на нормальный локус hEPO. Например, фрагмент HindIII-BamHII 6 т.п.н. (см. Пример 1f) против хода транскрипции гена hEPO имеет многочисленные последовательности узнавания рестриктаз, которые могут использоваться как сайты встраивания гена neo и слитого фрагмента промотор mMT1/ hGH. Один такой сайт, Bql II, лежащий приблизительно 1,3 т.п.н. по ходу транскрипции от сайта HindIII, является единственным в этом районе и может быть использован для встраивания одного или более селектируемых маркеров и регуляторного района, происходящего из другого гена, который будет служить для активации экспрессии hEPO в первичных, вторичных или иммортализованных клетках человека.
Сначала конструируют промежуточную плазмиду рТ164 встраиванием фрагмента HindIII-BamHI 6 т.п.н. (Пример 1f), лежащего против хода транскрипции от кодирующего района hEPO, в расщепленную HindIII-BamHI плазмиду pBluescript II SK+ (Stratagene, La Jolla, CA).
Плазмиду pMC1neoPolyA (Thomas, K.R. and Capecchi, M.R. Cell 51:503-512 (1987); доступную из Stratagene, La Jolla, СА) расщепляют Bam HI и XhoI, затупляют концы при помощи фрагмента Кленова ДНК-полимеразы E.coli и полученный фрагмент 1,1 т.п.н. очищают рТ164 расщепляют BglII и затупляют концы с фрагментом Кленова ДНК-полимеразы E.coli. Эти два фрагмента с тупыми концами лигируют вместе и трансформируют в компетентную E.coli. Клоны с единственной инсерцией фрагмента neo 1,1 т.п.н. выделяют и анализируют рестрикционным анализом для идентификации тех клонов, в которых сайт BglII, воссозданный слиянием тупых сайтов XhoI и BglII, локализован на расстояние 1,3 т.п.н. от единственного сайта HindIII, присутствующего в плазмиде pТ164. Полученную плазмиду, рТ165, можно теперь расщеплять при единственном сайте BglII, фланкирующем 5'-сторону транскрипционной единицы neo.
Олигонуклеотиды 13.8 и 13.9 используют в полимеразных цепных реакциях для образования фрагмента, в котором последовательности мышиный промотор металлотионеина 1 (mMT-1) - экзон 1 hGH дополнительно слиты с фрагментом 10 п.н., содержащим донорный сайт сплайсинга. Выбранный донорный сайт сплайсинга соответствует донорному сайту сплайсинга природного интрона 1 hEPO, хотя можно использовать большее число донорных сайтов сплайсинга или консенсусные донорные сайты сплайсинга. Олигонуклеотиды (13.8 и 13.9) используют для амплификации фрагмента из pXGH5 (фиг. 5) промотор mMT-1 приблизительно 0,73 т.п.н. - экзон 1 hGH. Амплифицированный фрагмент (фрагмент 7) расщепляют Bgl II и лигируют с расщепленной BglII плазмидой рТ165. Смесь для лигирования трансформируют в E.coli и клон, содержащий одну инсерцию фрагмента 7, в котором сайт Kpn I в промоторе mMT-1 находится рядом с 5'-концом гена neo и промотор mMT-1 ориентирован таким образом, что транскрипция направляется по направлению к уникальному сайту HindIII, идентифицируют и обозначают как рХЕРО-12.

Район с нежирными буквами олигонуклеотида 13.8 идентичен промотору mMT-1, с природным сайтом KpnI в качестве его 5'-границы. Жирные буквы обозначают конец сайта BglII для превращения 5'-границы в сайт BglII.

Жирный район олигонуклеотида 13.9 обозначает последовательности hGH. Курсив соответствует первым 10 п.н. интрона 1 hEPO. Подчеркнутый сайт BglII добавлен для целей конструирования плазмиды.
Плазмиду рХЕРО-12 можно использовать для генного нацеливания расщеплением BamHI и HindIII для высвобождения фрагмента 7,9 т.п.н., содержащего ген neo и слияние mMT-1/hGH, фланкированное на обеих сторонах последовательностями hEPO. Этот фрагмент (нацеливающий фрагмент 3) не содержит кодирующих hEPO последовательностей, имея только последовательности, лежащие между приблизительно -620 и приблизительно -6620 против хода транскрипции от кодирующего района hEPO, для регуляции нацеливания против хода транскрипции локуса ЕРО человека. Нацеливающий фрагмент 3 трансфицируют в первичные, вторичные или иммортализованные фибробласты кожи человека при условиях, сходных с описанными в Примерах 1b и 1с. Устойчивые к G418 колонии выскребают и помещают в отдельные лунки 96-луночных планшетов и скринируют на экспрессию ЕРО твердофазным иммуноферментым анализом ELISA (R&D Systems, Minneapolis MN). Клетки, в которых трансфицирующая ДНК интегрируется произвольно в геном человека, не могут продуцировать hEPO. Клетки, в которых трансфицирующая ДНК подвергалась гомологичной рекомбинации с эндогенным промотором hEPO и последовательностями против хода транскрипции, содержат химерный ген, в котором промотор mMT-1 и нетранскрибируемые последовательности, 5'-нетранслируемые последовательности hSH и экзон 1 hSH и линкер 10 п.н., состоящий из первых 10 п.н. интрона 1 hEPO, встроены при сайте BglII, лежащем в положении приблизительно -1920 по отношению к кодирующего району hEPO. Локализация промотора mMT-1 против хода транскрипции обычно молчащего промотора hEPO будет направлять в первичных, вторичных или иммортализованных фибробластах человека синтез транскрипта, считывая последовательности нетранслируемого металлотионеина и hGH, экзон 1 hSH, 10 п.н., идентичных первым 10 п.н. интрона 1 hEPO, и район против хода транскрипции hEPO и экзон 1 hEPO (от приблизительно -1920 до +13 по отношению к кодирующей ЕРО последовательности). Линкерная последовательность 10 п.н. из интрона 1 hEPO действует как донорныи сайт сплайсинга для слияния экзона 1 hGH с расположенным по ходу транскрипции акцепторным сайтом сплайсинга, который лежит непосредственно слева (против хода транскрипции) от зкзона 2 hEPO. Процессинг полученного транскрипта, таким образом, будет сплайсировать последовательности против хода транскрипции hEPO, промоторный район, экзон 1 и последовательности интрона 1. При использовании рХЕРО-10, -11 и -12 пост-транскрипционный процессинг транскрипта может быть улучшен посредством мутагенеза in vitro для исключения акцепторных сайтов сплайсинга, лежащих в последовательностях против хода транскрипции hEPO между промотором mMT-1 и экзоном 1 hEPO, которые снижают уровень продуктивных событий сплайсинга, необходимых для образования желаемого транскрипта. Замена экзона 1 hEPO экзоном 1 hGH приводит к образованию белка, в котором первые 4 аминокислоты сигнального пептида hEPO заменены аминокислотами 1-3 hGH, что приводит к образованию функционального, химерного сигнального пептида, который удаляется пост-трансляционным процессингом из зрелого белка и секретируется из экспрессирующих клеток.
ПРИМЕР 3. Направленная модификация последовательностей против хода транскрипции и амплификация получившего нацеленную конструкцию ДНК гена
Клетки человека, в которых ген hEPO был активирован описанными выше способами, может быть индуцирован для амплификации транскрипционной единицы neo/mMT-1/EPO, если нацеливающая плазмида содержит маркерный ген, который придает устойчивость к высокому уровню цитотоксичного агента, посредством феномена амплификации генов. Было документировано, что среди других генов селектируемые маркерные гены, такие как ген дигидрофолатредуктазы (dhfr, селектирующий агент - метотрексат), мультифункциональный ген CAD [кодирующий карбамилфосфатсинтазу, аспартаттранскарбамилазу, дигидрооротазу; селектирующий агент - N-(фосфоноацетил)-L-аспартат (PALA)], ген глутаминсинтазы (селектирующий агент - метионинсульфоксимин (MSX)) и ген аденозиндезаминазы (ada; селектирующий агент - адениннуклеозид), являются амплифицируемыми в иммортализованных клеточных линиях человека (Wright, J.A., et al. Proc. Natl. Acad. Sci. USA 87:1791-1795 (1990); Cockett, M.I. et al., Biol. Technology 8:662-667 (1990)). В этих исследованиях было документировано, что амплификация генов происходит в большом числе иммортализованных клеточных линиях человека (НТ1080, HeLa, клетки рака молочной железы MCF-7, клетки лейкоза К-562, клетки карциномы KB или клетки рака яичника 2780AD), среди других клеток, обнаруживают амплификацию при подходящих условиях отбора.
Плазмиды рХЕРО-10 и рХЕРО-11 могут быть модифицированы встраиванием нормального или мутантного гена dhfr в уникальные сайты HindIII этих плазмид. После трансфекции клеток НТ1080 подходящей ДНК, отбора на устойчивость к G418 (сообщаемую геном neo) и идентификации клеток, в которых ген hEPO был активирован генным нацеливанием последовательностей neo, dhfr и mMT-1, эти клетки могут быть подвергнуты ступенчатому отбору в метотрексате (МТХ) для отбора на амплификацию dhfr и коамплификацию связанных последовательностей neo, mMT-1 и последовательностей hEPO (Kaufman, JR.J., Technique 2: 221-236 (1990)). Используют схему ступенчатого отбора, в котором клетки сначала выдерживают с низкими уровнями МТХ (0,01-0,08 мкМ) с последующим последовательным выдерживанием при возрастающих концентрациях МТХ до 250 мкМ или выше. Линейные возрастающие стадии 0,04-0,08 мкМ МТХ и последующие 2-кратные увеличения в концентрации МТХ будут эффективны в отборе на амплифицированные трансфицированные клеточные линии, хотя различные относительно неглубокие приращения будут также эффективными. Амплификацию наблюдают по увеличению в числе копий гена и подтверждают измерением экспрессии hEPO in vitro. Этой стратегией может быть достигнута значительная сверхэкспрессия hEPO посредством направленной модификации последовательностей, лежащих полностью вне кодирующего района hEPO.
Конструкции, похожие на описанные конструкции (Примеры 1f, 1h, 1i, 1k, 2 и 7), для активации экспрессии hGH в клетках человека также могут быть модифицированы далее для включения гена dhfr с целью получения клеток, которые сверхэкспрессируют ген hGH, посредством генного нацеливания на некодирующие последовательности и последующей амплификации.
ПРИМЕР 4. Нацеливание и активация локуса ЕРО человека в иммортализованной линии фибробластов человека
Была изготовлена нацеливающая (направляющая) конструкция рХЕРО-13 для проверки гипотезы, что эндогенный ген hEPO мог бы быть активирован в фибробластной клетке человека. Сначала конструировали плазмиду рТ22.1, содержащую геномную последовательность hEPO 63 п.н. против хода транскрипции первого кодона гена hEPO, слитого с мышиным промотором металлотионеина (mMT-1). Олигонуклеотиды 22.1-22.4 использовали в ПЦР для слияния последовательностей mMT-1 и hEPO. Свойства этих праймеров следующие: 22.1 представляет собой олигонуклеотид из 21 оснований, гомологичный сегменту промотора mMT-1, начинающемуся 28 п.н. против хода транскрипции от сайта Kpn I mMT-1; 22.2 и 22.3 являются комплементарными праймерами из 58 нуклеотидов, которые определяют слияние последовательностей hEPO и mMT-1 таким образом, что это слияние содержит 28 п.н. последовательностей hEPO, начинающихся 35 оснований против хода транскрипции от первого кодона гена hEPO, и последовательности mMT-1, начинающиеся при основании 29 олигонуклеотида 22.2, содержащие природный сайт Bgl II mMT-1 и простирающиеся на 30 оснований в последовательность mMT-1; 22.4 представляет собой 21 нуклеотид в длину и является гомологичным последовательностям hEPO, начинающимся при 725 п.н. по ходу транскрипции от первого кодона гена hEPO. Эти праймеры используют для амплификации фрагмента ДНК 1,4 т.п.н., содержащего слияние последовательностей mMT-1 и hEPO, описанное выше. Полученный фрагмент расцепляют Kpn I (фрагмент ПЦР, содержащий два сайта Kpn I: один природный сайт Kpn I в районе промотора mMT-1 и один природный сайт Kpn I в последовательности hEPO) и очищают. Плазмиду рХЕРО1 также расщепляли Kpn I, высвобождая фрагмент 1,4 т.п.н. и фрагмент 6,4 т.п.н. Фрагмент 6,4 т.п.н. очищали и лигировали со слитым фрагментом 1,4 т.п.н. Kph 1 ПЦР. Полученная конструкция была названа рТ22.1. Вторую промежуточную плазмиду, рТ22.2, конструировали лигированием фрагмента HindIII-BamHI 6 т.п.н., лежащего против хода транскрипции структурного гена hEPO (см. Пример 1f) с расщепленной BamHI и HindIII плазмидой рВ SIISК+ (Stratagene, La Jolla, CA). Третью промежуточную плазмиду, рТ22.3, конструировали вырезанием сначала фрагмента XhoII/BamHI 1,1 т.п.н. из рМС1NЕРроly A (Stratagene, La Jolla, CA), содержащего ген неомицинфосфотрансферазы. Затем концы этого фрагмента затупляли при помощи фрагмента Кленова ДНК-полимеразы 1 (New England Biolabs).
После этого этот фрагмент лигировали с сайтом HindIII pBS II S К+ (также затупленный ДНК-полимеразой 1) с образованием рТ22.3. Четвертую промежуточную плазмиду, рТ22.4, получали очисткой фрагмента XhoI/HindIII 1,1 т.п.н., содержащего ген neo из рТ22.3, и лигированием этого фрагмента с расщепленной XhoI и HindIII pT22.2. Таким образом, рТ22.4 содержит ген neo рядом со стороной HindIII фрагмента BamHI-HindIII фрагмента против хода транскрипции hEPO. Наконец, рХЕРО-13 получали вырезанием сначала фрагмента EocRI-AccI 2,0 т.п.н. из плазмиды рТ22.1, Сайт EcoRI этого фрагмента определяет 5'-границу промотора mMT-1, тогда как сайт AccI этого фрагмента лежит внутри экзона 5 hEPO. Таким образом, фрагмент AccI/EcoRI содержит почти полную экспрессионную единицу hEPO, не имея только части экзона 5 и природного сайта полиаденилирования. Этот фрагмент EcoRI/AccI 2,0 т.п.н. очищали, концы затупляли обработкой фрагментом Кленова ДНК-полимеразы 1 и лигировали с расщепленной XhoI, имеющей затупленные концы рТ22.4.
Клетки НТ1080 трансфицировали расщепленной PVU1-BamHI плазмидой рХЕРО-13. рХЕРО-13, расщепленная таким образом, образует три фрагмента; векторный фрагмент 1 т.п.н., включающий в себя ген amp; векторный фрагмент 1,7 т.п.н. из оставшихся векторных последовательностей и фрагмент приблизительно 9 т.п.н., содержащий последовательности hEPO, neo и mMT-1. Этот фрагмент Bam HI/PVUI из 9 т.п.н. содержал следующие последовательности в указанном порядке от сайта Bam HI: геномную последовательность против хода транскрипции hEPO приблизительно 5,2 т.п.н., транскрипционную единицу neo 1,1 т.п.н., промотор mMT-1 0.7 т.п.н. и фрагмент 2,0 т.п.н., содержащий кодирующую последовательность hEPO, укороченную в зкзоне 5. 45 мкг рХЕРО-13, расщепленную таким образом, использовали в электропорации 12 млн клеток (условия электропорации описаны в Примере 1b). Эту электропорацию повторяли в целом 8 раз, получая электропоряцию в целом 96 млн клеток. Клетки смешивали со средой, получая плотность клеток 1 млн клеток на мл, и аликвоты по 1 мл распределяли в целом на 96 чашек для культивирования ткани с диаметром 150 мм (Falcon), причем каждая чашка содержала как минимум 35 мл среды DMEM/15% телячья сыворотка. На следующий день эту среду отсасывали и заменяли свежей средой, содержащей 0,8 мг/мл G418 (Gibco). После 10 дней инкубирования из сред из каждой чашки брали пробы на hEPO для твердофазного иммуноферментного анализа ELISA (R&D Systems). Шесть из 96 чашек содержали по меньшей мере 10 мЕ/мл hEPO. Одну из этих чашек, №18, выбрали для очистки экспрессирующих hEPO колоний. Каждая из 96 чашек с диаметром 150 мм содержала приблизительно 600 устойчивых к G418 колоний (всего 57600 устойчивых к G418 колоний определено на всех 96 чашках). Приблизительно 600 колоний на чашке №18 трипсинизировали и пересевали при 50 клеток на мл в 364-луночные планшеты (Sterilin). После одной недели инкубирования отдельные колонии были видимыми при приблизительно 10 колониях на большую лунку 364-луночных планшетов (эти планшеты состоят из 16 малых лунок внутри каждой из 24 больших лунок). Каждую лунку подвергали скринингу на экспрессию hEPO в этой точке. Две из больших лунок содержали среды, по меньшей мере, с 20 мЕ/мл ЕРО. Лунка № А2 содержала 15 колоний, распределенных среди 16 малых лунок. Содержимое каждой из этих малых лунок трипсинизировали и переносили в 16 отдельных лунок 96-луночного планшета, после 7 дней инкубирования из сред из каждой из этих лунок брали пробы для анализа ELISA на hEPO. Только одна лунка, лунка №10, содержала hEPO. Этот клеточный штамм был назван НТ165-18А2-10 и его размножали в культуре для количественного анализа hEPO, выделения PHK и выделения ДНК. Количественное измерение продуцирования hEPO дало величину 2500 миллиединиц на миллион клеток за 24 часа.
ДНК-зонд 0,2 т.п.н., простирающийся от сайта AccI в экзоне 5 hEPO до сайта Bql II в 3'-нетраслируемом районе, использовали для зондирования РНК, выделенной из клеток НТ165-18А2-10. Направляющая конструкция, рХЕРО-13, укороченная при сайте AccI в экзоне 5, не содержит эти последовательности AccI/BglII и, следовательно, является диагностической для нацеливания при локусе hEPO. Только клеточные штаммы, которые рекомбинировались гомологично с природными последовательностями hEPO, продуцировали бы мРНК hEPO, содержащую последовательность, гомологичную последовательностям AccI/BglII. Было обнаружено, что НТ165-1842-10 экспрессирует мРНК предсказанного размера, гибридизующуюся с 32Р-меченым зондом AccI/BqIII hEPO на нозерн-блотах. Рестрикционный анализ и блоттинг по Саузерну подтвердили, что ген neo и промотор mMT-1 попали в одну из двух аллелей hEPO в клетках НТ165-18А2-10.
Эти результаты демонстрируют, что гомологичную рекомбинацию можно использовать для нацеливания и включения регуляторного района в ген, который в норме является молчащим в фибробластах человека, что приводит к функциональной активации этого гена.

ПРИМЕР 5. Получение не содержащих интронов генов
Генное нацеливание можно также использовать для получения проциссированного гена, не содержащего интронов, для переноса в дрожжи или бактерии для экспрессии гена и получения белка in vitro. Например, hGH можно продуцировать в дрожжах при помощи описанного ниже подхода.
Получают две отдельные направляющие конструкции. Направляющая конструкция 1 (ТС1) включает в себя последовательность LTP, например LTR из вируса мышиной лейкемии Moloney (MoMLV) маркер для отбора в клетках человека (например, ген neo из Тn5), маркер для отбора в дрожжах (например, дрожжевой ген URA3), регуляторный район, способный регулировать генную экспрессию в дрожжах (например, промотор GAL4) и иногда (необязательно) последовательность, которая при слиянии с геном hGH будет обеспечивать секрецию hGH из дрожжевых клеток (лидерную последовательность). Этот вектор может также включать в себя последовательность ДНК, которая делает возможной ретровирусную упаковку в клетках человека. Эта конструкция организована таким образом, что указанные выше последовательности фланкированы на обеих сторонах геномными последовательностями hGH, которые при гомологичной рекомбинации с N-последовательностями геномного гена hGH будут интегрировать эти экзогенные последовательности в ТС1 непосредственно слева от N-кодона 1 гена hGH (соответствующего положению аминокислоты 1 в зрелом, процессированном белке). Порядок последовательностей ДНК при интеграции является следующим: последовательности против хода транскрипции hGH и регуляторные последовательности, ген neo, LTR, ген URА3, промотор GAL4, дрожжевая лидерная последовательность, последовательности hGH, включающие в себя аминокислоту 1 зрелого белка и по ходу транскрипции hGH. Нацеливающая конструкция 2 (ТС2) включает в себя последовательности, достаточные для репликации плазмиды в дрожжах (например, кольцо в 2 микрона или последовательности ARS), терминирующую последовательность транскрипции држжжей, вирусный LTR и маркерный ген для отбора в клетках человека (например, бактериальный ген gpt). Эта конструкция организована таким образом, что указанные выше последовательности фланкированы на обеих сторонах геномными последовательностями hGH, которые при гомологичной рекомбинации с N-последовательностями геномного гена hGH будут интегрировать экзогенные последовательности в ТС2 непосредственно по ходу транскрипции от N-стоп-кодона гена hGH. Порядок последовательностей ДНК при интеграции является следующим: последовательности экзона 5 hGH, последовательности терминации транскрипции дрожжей, последовательности репликации плазмиды в дрожжах, LTR, ген gpt, 3'-нетранслируемые последовательности hGH.
Линейные фрагменты, полученные из TC1 и ТС2, последовательно направляются для включения в их соответствующие положения, фланкирующие ген hSH. После суперинфекции этих клеток с хелперным ретровирусом регулируемая LTR транскрипция через этот район будет приводить к образованию РНК с последовательностями LTR на обоих концах. Сплайсинг этой РНК будет приводить к образованию молекулы, в которой удалены обычные интроны hGH. Обратная транскрипция процессированного транскрипта приведет к накоплению двухцепочечных ДНК-копий процессированного слитого гена hGH. ДНК выделяют из двукратно получивших нацеленные конструкции, инфицированных ретровирусом клеток и расщепляют ферментом, который расщепляет эту транскрипционную единицу один раз в LTR. Расщепленный материал лигируют при условиях, усиливающих циркуляризацию, вводят в дрожжевые клетки и эти клетки затем подвергают отбору на ген РА3. Расти могут только клетки, получившие ген URA3 (соединенный с последовательностями, вводимыми ТС1 и ТС2 и процессированным геном hGH). Эти клетки содержат плазмиду, которая будет экспрессировать белок hGH при галактозной индукции и секретировать белок hGH из клеток благодаря слитой последовательности дрожжевого лидерного пептида, которая отщепляется при секреции, давая зрелую, биологически активную молекулу hGH.
Экспрессия в бактериальных клетках достигается простой заменой, в ТС1 и ТС2, геном устойчивости к ампициллину из pBR322 дрожжевого гена URA3, промотором tac (de Boer et al., Proc. Natl. Acad. Sci. 80:21-25 (1983)) дрожжевого промотора GAL4, бактериальной лидерной последовательностью дрожжевой лидерной последовательности, началом репликации pBR322 2-микронного кольца или последовательности ARS и бактериальной терминацией транскрипции (например, терминатором транскрипции trpA; Christle, G.E. et al., Proc. Natl. Acad. Sci. 78:4180-4184 (1981)) последовательности для дрожжевой терминации транскрипции. Подобным образом hEPO может экспрессироваться в дрожжах и бактериях простой заменой нацеливающих последовательностей hGH нацеливающими последовательностями hEPO, так чтобы дрожжевая или бактериальная лидерная последовательность была расположена непосредственно против хода транскрипции кодона 1 hEPO (соответствующего положению аминокислоты 1 в зрелом процессированном белке).
ПРИМЕР 6. Активация и амплификация гена ЕРО в иммортализованной клеточной линии человека
Включение экспрессионной единицы dhfr в уникальный сайт HindIII pXEPO-13 (см. Пример 4) приводит к образованию нового нацеливающего вектора, способного к двойному отбору и к отбору клеток, в которых амплифицируется ген dhfr. Единственный сайт HindIII в pXEPO-13 определяет место соединения гена neo и геномной последовательности, природно наводящейся против хода транскрипции гена ЕРО человека. Помещение гена в этом сайте обеспечивает конструкцию с генами neo и dhfr, окруженными последовательностью ДНК, происходящей из природного локуса ЕРО. Подобно pXEPO-13, производные с встроенным геном dhfr применимы для нацеливания на локус hEPO посредством гомологичной рекомбинации. Такая конструкция, названная pREPO4, представлена на Фиг.6. Эта плазмида включает в себя экзоны 1-4 и часть экзона 5 гена ЕРО человека, а также фрагмент HindIII-BamHI, лежащий против хода транскрипции кодирующего района hEPO. pSVe, pTK и pmMT-1 соответствуют промоторам из раннего района SV 40, гена тимидинкиназы (ТК) вируса простого герпеса (HSV) и гена мышиного металлотионеина-1. Ее получали следующим образом: расщепленную HindIII плазмиду рХЕРО-13 очищали, концы ее затупляли при помощи фрагмента Кленова ДНК-полимеразы 1. Для получения экспрессионной единицы dhfr плазмидную конструкцию pF8C1S9080 (Eaton et al., Biochemistry 25:8343-6347 (1986)) расщепляли EcoRI и SalI. Фрагмент 2 т.п.н., содержащий экспрессионную единицу, очищали из продукта расщепления и концы его затупляли фрагментом Кленова ДНК-полимеразы 1. Этот содержащий dhfr фрагмент затем лигировали с затупленным сайтом HindIII рХЕРО-13. Аликвоту смеси для лигирования трансформировали в E.coli и высевали на чашки для отбора на устойчивость к ампициллину. После инкубации в течение ночи при 37°С отдельные колонии наблюдали, выскребали и выращивали. Препараты миниплазмид готовили из этих культур и полученную ДНК подвергали рестрикционному расщеплению ферментами BqlII-HindIII и Sfi I для определения ориентации встроенных фрагментов dhfr. Было обнаружено, что плазмидная ДНК из одного из этих препаратов содержит такую инсерцию 2 т.п.н. фрагмента dhfr. Было найдено, что ориентация транскрипции экспрессионной единицы dhfr в этой плазмиде была противоположна ориентации смежного гена neo.
Плазмиду pREPO4 использовали для амплификации локуса hEPO в клетках после активации эндогенного гена hEPO гомологичной рекомбинацией. Активация гена такой конструкцией делает возможным отбор на увеличенную экспрессию DHFR с использованием лекарственного средства метотрексата (МТХ). В типичном случае усиленная экспрессия DHFR могла бы иметь место в результате увеличения числа копий вследствие амплификации ДНК. Чистым результатам была бы коамплификация активированного гена hEPO вместе с последовательностями dhfr. Коамплификация активированного локуса ЕРО должна приводить к увеличенной экспрессии ЕРО.
Эсперименты по нацеливанию проводили в клетках НТ1080 с плазмидой pREPO4. Выделяли линию HTREPO-52, экспрессирующую hEPO. Эту линию анализировали количественно на продуцирование ЕРО и при помощи блоттинга по Саузерну и нозерн-блоттинга. Было обнаружено, что этот штамм получал нацеленную одну копию последовательностей dhfr /neo/ mMT-1. Полученные уровни экспрессии при отборе с 0,8 мг/мл G418 были приблизительно 1300 мЕ/млн клеток/день. Поскольку нацеленный локус ЕРО содержал экспрессионную единицу dhfr, был возможен отбор на увеличенную экспрессию DHFR с антифолатом, МТХ. Таким образом, этот штамм подвергали ступенчатому отбору в 0,02, 0,05, 0,1, 0,2 и 0,4 мкМ МТХ. Результаты исходного отбора этого штамма представлены в Таблице 4 и на фиг.7.
Отбор с повышающимися уровнями МТХ показал увеличение экспрессии в линии HTREPO-52, с 70-кратным увеличением в продуцировании ЕРО, наблюдаемым в клеточной линии, устойчивой к 0,4 мкМ МТХ. Подтверждение амплификации локуса hEPO получали блоттингом по Саузерну в устойчивых к МТХ клеточных линиях, которые обнаружили приблизительно 10-кратное увеличение в числе копий активированного локуса hEPO по сравнению с исходным (не получившим нацеленной конструкции) аллелем hEPO.
ПРИМЕР 7. Получение слитого гена hEPO встраиванием промотора CMV на 1,8 т.п.н. против хода транскрипции кодирующего района геномного hEPO
Конструирование нацеливающей плазмиды рRЕРO15:
рRЕРO15 конструировали слиянием сначала промотора CMV с экзоном 1 hGH при помощи ПЦР-амплификации. Фрагмент 1,6 т.п.н. амплифицировали из экспрессионной конструкции hGH pXGH308, которая имеет район промотора CMV, начинающийся при нуклеотиде 546 и заканчивающийся при нуклеотиде 2105 последовательности Genbank HS5M1EP, слитый, с последовательностями hGH, начинающимися при нуклеотиде 5225 и заканчивающимися при нуклеотиде 7322 Genbank последовательности HUMGHCSA, с использованием олигонуклеотидов 20 и 35. Олигонуклеотид 20 (35 п.н., SEQ ID №18) гибридизовался с промотором CMV при -614 по отношению к кэп-сайту (в последовательности Genbank HEHCMV Р1) и включал в себя, сайт SalI при его 5'-конце. Олигонуклеотид 35 (42 п.н., SEQ ID №19), гибридизовался с промотором CMV при +966 и смежном экзоне 1 hGH и включал в себя первые 10 п.н. интрона 1 hEPO (содержащие часть донорного сайта сплайсинга) и сайт HindIII при его 5'-конце. Полученный фрагмент ПЦР расщепляли HindIII и SalI и очищали на геле. Плазмиду рТ163 (Пример 2) расщепляли XhoI и HindIII и фрагмент приблизительно 1,1 т.п.н., содержащий экспрессионную единицу neo, очищали на геле. Фрагмент CMV промотор 1,6 т.п.н./экзон 1 hGH/донорный сайт сплайсинга и фрагмент neo 1,2 т.п.н. лигировали вместе и встраивали в сайт HindIII PBSIISK+ (Stratagene, Inc.). Полученная промежуточная плазмида (названная pBNCHS ) содержала экспрессионную единицу neo в транскрипционной ориентации, противоположной ориентации фрагмента CMV промотор/ экзон 1 hGH/донорный сайт сплайсинга. Вторую промежуточную плазмиду, рRЕРO5Δ HindIII, констуировали расшеплением сначала pREPO5 при помощи HindIII. Это высвобождало два фрагмента 1,9 т.п.н. и 8,7 т.п.н., и фрагмент 8,7 т.п.н., содержащий нацеливающие последовательности ЕРО, очищали на геле и циркуляризовали рециркуляцией. Полученная плазмида, рRЕРO5-HindIII, содержала только некодирующие последовательности геномной ДНК, обычно находящиеся против хода транскрипции гена hEPO. Она включала последовательность от -6786 до -1 по отношению к экзону 1 ЕРО. Фрагмент 2,8 т.п.н., содержащий neo, промотор CMV, экзон 1 hGH и донорный сайт сплайсинга, вырезали из pBNCHS при помощи Hind III и очищали на геле. Этот фрагмент затупляли на концах при помощи фрагмента Кленова ДНК-полимеразы 1 (New England Biolabs, Inc.) и лигировали с расщепленной BglII и имеющей затупленный концы плазмидной рRЕРO5Δ HindIII. BqlII разрезают при положении -1779 против хода транскрипции экзона 1 hEPO в рRЕРO5Δ Hind III. Полученная конструкция, pREPO15 (Фиг. 8) содержала последовательности ЕРО против хода транскрипции от -5786 до -1779 по отношению к кодирующей области hEPO, экспрессионную единицу neo, промотор CMV, экзон 1 hGH, донорный сайт сплайсинга и последовательности от -1778 до -1 п.н. против хода транскрипции от кодирующего района hEPO, с различными собранными элементами, в перечисленном порядке, по отношению к нуклеотидной последовательности района против хода транскрипции hEPO. Для трансфекции клеток человека pREPO15 расщепляли Not 1 и Pvu 1 для высвобождения нацеливающего фрагмента 8,6 т.п.н. Нацеливающий фрагмент содержал первую и вторую направляющие последовательности 4,0 т.п.н. и 1,8 т.п.н., соответственно, с гомологией к ДК против хода транскрипции гена hEPO.
Культура клеток, трансфекция и идентификация экспрессирующих ЕРО получивших нацеленные конструкции клонов
Все клетки поддерживали при 37°С, 5% СО2 и 98% влажности в среде DMEM, содержащей 10% телячью сыворотку (DMEM/10, Hyclone Laboratories). Трансфекцию вторичных фибробластов крайней плоти человека выполняли электропорацией 12×10 клеток в PBS (G1BCO) с 100 мкг ДНК при 250 В и 960 мкФ. Обработанные клетки высевали при 1×106 клеток на чашки с диаметром 150 мм. На следующий день среды заменяли на DMEM/10, содержащую 0,8 мг/мл G418 (G1BCO). Отбор протекал в течение 14 дней, после чего из сред брали пробы для определения продуцирования ЕРО. Все колонии на чашках, проявляющие значимые уровни hEPO (более 5 мЕ/мл) при определении ЕРО при помощи ELISA (Genzyme), выделяли стерильными стеклянными клонирующими цилиндрами (Bellco) и переносили в отдельные лунки 96-луночного планшета. После инкубации в течение 1-2 дней из этих лунок брали пробы на продуцирование hEPO, определяемое при помощи ELISA. Полученные продуцирующие hEPO клеточные штаммы размножали в культуре для замораживания, выделения нуклеиновых кислот и определений количества продуцируемого ЕРО.
Трансфекцию клеток НТ1080 (АТСС CCL 121) проводили обработкой 12×106 клеток в PBS (G1BCO) с 45 мкг ДНК при 450 В и 250 мкФ. Рост и идентификация клонов вторичных фибробластов крайней плоти человека описана выше. Выделение продуцирующих hEPO клональных клеточных линий проводили лимитирующим разведением. Его выполняли первым посевом колоний, собранных из исходных чашек для отбора, в виде пулов из 10-15 колоний на лунку 24-луночного планшета. Затем продуцирующие hEPO пулы высевали при плотности клеток, приводящей к числу колоний менее 1 на лунку 96-луночного планшета. Отдельные клоны размножали для дальнейшего анализа, как описано для фибробластов крайней плоти человека выше.
Характеристика экспрессирующих ЕРО клонов:
Плазмида pREPO15 не содержит каких-либо кодирующих последовательностей hEPO. При получении нацеленной конструкции с фрагментом neo/CMV - промотор/экзон 1 hGH/донорный сайт сплайсинга против хода транскрипции экзона 1 hEPO экспрессия hEPO происходит инициацией транскрипции от промотора CMV, с образованием первичного транскрипта, который включает в себя последовательности CMV, экзон 1 hEPO и донорный сайт сплайсинга, последовательности 1,8 т.п.н. hEPO против хода транскрипции и обычные экзоны hEPO, интроны и 3' - нетранслируемые последовательности. Сплайсинг этого транскрипта имеет место от донорного сайта сплайсинга, находящегося рядом с экзоном 1 hGH, до следующего по ходу транскрипции акцепторного сайта сплайсинга, который расположен рядом с экзоном 2 hEPO. Это приводит к образованию нового интрона, состоящего из геномной последовательности против хода транскрипции гена hEPO, обычного промотора hEPO, экзона 1 hEPO и интрона 1 hEPO. В зрелом транскрипте экзон 1 hGH будет заменять экзон hEPO. Экзон 1 hEPO кодирует только первые 4 и 1/3 аминокислот из 26 аминокислот сигнального пептида, который отщепляется от белка-предшественника перед секрецией из клеток. Экзон 1 hGH кодирует первые 3 и 1/3 аминокислот сигнального пептида hGH, который также отщепляется от белка-предшественника перед секрецией из клетки. Трансляция этого транскрипта, в котором экзон 1 hGH заменяет экзон 1 hEPO, привело бы к белку, в котором сигнальный пептид является химерой последовательности hGH и hEPO. Удаление сигнального пептида посредством нормального события пост-трансляционного расщепления будет продуцировать зрелую молекулу hEPO, первичная последовательность которой неотличима от нормального продукта.
Трансфекция pREPO15 в фибробласты человека приводила к экспрессии ЕРО этими клетками. Таблица 5 показывает результаты экспериментов по нацеливанию с pREPO15 в фибробластах человека и клетках НТ1080. Частота получения нацеленных конструкций в нормальных фибробластах человека была 1/264 G418r-колоний и частота получения нацеленных конструкций в случае клеток НТ1080 была 1/450 G418-колоний. Уровни продуцирования hEPO из каждого из этих клеточных штаммов оценивали количественно. Было обнаружено, что продуцент hEPO, полученный в результате трансфекции фибробластов человека, секретировал 7679 мЕ/106 клеток/день (Таблица 5). Активированная по hEPO клеточная линия из клеток НТ1080 продуцировала 12582 мЕ/106 клеток/день (Таблица 5). Эти результаты показали, что активация локуса hEPO была эффективной и вызвала конститутивное продуцирование hEPO при относительно высоких уровнях. Рестрикционный анализ и гибридизационный анализ по Саузерну использовали для подтверждения, что событие нацеливания имело место при локусе ЕРО.
Блоттинг по Саузерну клонов фибробластов человека и клеток НТ1080, которые получили нацеленную pREPO15, был проведен. Фиг.9А показывает рестрикционную карту исходного и получившего нацеленную конструкцию локуса hEPO, а фиг. 9В показывает результаты рестрикционного анализа и гибридизационного анализа по Саузерну нацеленного (получившего нацеленную конструкцию) клона фибробластов человека. Продукты расщепления BglII/EcoRI и ВаmH1 обнаружили фрагменты 5,9 и 6,6 т.п.н., соответственно, как результат события нацеливания при локусе hEPO (дорожки Т1). Оба эти фрагмента происходили из встраивания 2,7 т.п.н. ДНК, содержащей ген neo и последовательности промотора CMV. Поскольку только один из двух аллелей hEPO получал нацеленные конструкции, фрагменты 4,3 т.п.н. (BqlII /EcoRI) или 10,6 т.п.н. (BamHI), отражающие неизменный локус hEPO, были видны в этих штаммах и в исходной ДНК (дорожки HF). Эти результаты подтверждают, что событие гомологичной рекомбинации имело место при локусе hEPO, приводя к образованию новой транскрипционной единицы, которая управляла образованием эритропоэтина человека.
(SEQ 10 №18)
GGTGGATCCCGT
(SEQ ID №19)
b из среды в чашках с G418r-колониями брали пробы для анализа ЕРО (ELISA) и пробы, обнаруживающие уровни hEPO больше 5 мЕ/мл, считали как событие активации ЕРО
с продуцирование ЕРО определяли количественно из штамма фибробластов человека, клеточной линии НF342-15 или клеточной линии НТ1080, HTREPO15-1-6-6
ПРИМЕР 8. Получение и амплификация слитого гена hEPO встраиванием промотора CMV 1,8 т.п.н. против хода транскрипции кодирующего района геномного hEPO
Конструирование нацеливающей плазмиды рRЕРO18:
Плазмиду рRЕРO18 (Фиг.10) конструировали встраиванием экспрессионной единицы dhfr при сайте ClaI, локализованном при 5'-конце гена neo pREPO15. Для получения экспрессионной единицы dhfr плазмидную конструкцию рF8С1S9080 [Eaton, et al., Biochemistry, 25: 8343-8347 (1986)] расщепляли EcoRI и SalI. Фрагмент 2 т.п.н., содержащий экспрессионную единицу dhfr, очищали из продукта расщепления и концы его затупляли при помощи фрагмента Кленова ДНК-полимеразы 1. Затем линкер ClaI (New England Biolabs) лигировали с затупленным фрагментом dhfr. Продукты этого лигирования расщепляли затем ClaI, лигированной с расщепленной ClaI плазмидой pREPO15. Аликвоту этой смеси для лигирования трансформировали в E.coli. и высевали на чашки для отбора на устойчивость к амплициллину. Бактериальные колонии анализировали расщеплением рестриктазами для определения ориентации встроенного фрагмента dhfr. Одну плазмиду с dhfr в транскрипционной ориентации, противоположной ориентации гена neo, назвали pREPO18(-). Вторую плазмиду с dhfr в той же самой транскрипционной ориентацией, что и ориентация гена neo, назвали pREPO18(+).
Культура клеток, трансфекция и идентификация экспрессирующих ЕРО получивших нацеленные конструкции клонов:
Все клетки поддерживали при 37°С, 5% СО2 и 98% влажности в ДМЕМ, содержащей 10% телячью сыворотку (DMEM/10, HyClone Laboratories). Трансфекция клеток НТ1080 (АТСС CCL 121) происходила обработкой 12×106 клеток в PBS (G1BCO) с 45 мкг ДНК при 450 В и 250 мкФ. Обработанные клетки высевали при 1×106 клеток на 150 мм-чашку. На следующий день среду заменяли на ДМЕМ/10, содержащую 0,8 мг/мл G418 (G1ВСО). Отбор продолжали в течение 14 дней, после чего брали пробы среды для определения образования hEPO. Чашки, обнаруживающие значимые уровни образования hEPO (более 5 мЕ/мл) при определении hEPO твердофазным иммуноферментным анализом ELISA (Genzyme Inc.), трипсинизировали и клетки пересевали для выделения клонов. Выделение продуцирующих hEPO клональных клеточных линий происходило с применением лимитирующего разведения путем посева сначала клонов в пулах из 10 - 15 колоний на лунку 24-луночного планшета с последующим высевом клеток из продуцирующих ЕРО пулов при плотностях клеток, приводящих к менее чем 1 колонии, на лунку 96-луночного планшета. Отдельные клоны размножали для замораживания, выделения нуклеиновых кислот и количественного определения продуцирования hEPO.
Выделение клеток, содержащих амплифицированные последовательности dhfr отбором с метотрексатом:
Получившие нацеленные конструкции устойчивые к G418 клеточные линии, продуцирующие hEPO после трансфекции рRЕРO18, высевали при разных плотностях клеток для отбора в метотрексате (МТХ). По мере определения новых клонов после отбора при одной концентрации МТХ эти клоны анализировали на продуцирование hEPO и пересевали при разных плотностях клеток в более высокую концентрацию МТХ (обычно с удвоением предыдущей концентрации). Этот процесс повторяли до тех пор, пока не достигался желаемый уровень продуцирования hEPO. При каждой стадии МТХ устойчивости выделяли ДНК и РНК для соответствующих анализов с использованием блоттинга по Саузерну и нозерн-блоттинга.
Характеристика экспрессирующих ЕРО клонов:
pREPO18 с двумя различными ориентациями dhfr трансфицировали в клетки НТ1080. Перед трансфекцией pREPO18(+) и рRЕРO18(-) расщепляли Хba1, высвобождая нацеливающий фрагмент 7,9 т.п.н., содержащий, в следующем порядке, район 2,1 т.п.н. геномной ДНК против хода транскрипции от экзона 1 hEPO (от -3891 до -1779 по отношению к стартовому кодону ATG hEPO), район 2 т.п.н., содержащий ген dhfr район 1,1 т.п.н., содержащий ген neo, район 1,5 т.п.н., содержащий промотор CMV, слитый с экзоном 1 hGH, 10 п.н. интрона 1 hEPO (содержащего донорный сайт сплайсинга) и затем район 1,1 т.п.н. геномной ДНК против хода транскрипции экзона 1 hEPO (от -1778 до -678 по отношению к стартовому кодону ATG ЕРО). Трансфекция и частота нацеливания (получения нацеленных конструкций) из двух экспериментов показаны в Таблице 6. 5 первичных устойчивых к G418 клонов выделили из этих экспериментов. Их размножали в культуре для количественного анализа экспрессии hEPO (Таблица 7). Поскольку pREPO18 содержала ген dhfr, можно было проводить отбор на клетки, содержащие амплифицированные копии направляющей конструкции с использованием МТХ, как описано в Примере 6. Устойчивые к G418 клоны использовали для подтверждения присутствия в них нацеленных конструкций ДНК в локусе hEPO при помощи рестрикционного анализа и гибридизационного анализа по Саузерну, после чего их подвергали ступенчатому отбору в МТХ, как описано выше.
ПРИМЕР 9. Активация и амплификация эндогенных генов α-интерферона, GM-CSF, G-CSF и FSHβ в иммортализованных клетках человека
Широкое разнообразие эндогенных клеточных генов можно активировать и амплифицировать с использованием способов и конструкций ДНК этого изобретения. Следующее далее описание раскрывает общую стратегию для активации и амплификации генов α-интерферона человека (лейкоцитарного интерферона), GM-CSF (колониестимулирующего фактора гранулоцитов/макрофагов), G-CSF (колониестимулирующего фактора гранулоцитов) и FSHβ (β-субъединицы фолликулостимулирующего гормона).
α-интерферон
Ген α-интерферона человека (Genbank Seguence HUMIFNAA) кодирует белок-предшественник из 188 аминокислот, содержащий сигнальный пептид из 23 аминокислот. Этот ген не содержит интронов. Фиг.11 схематически иллюстрирует одну стратегию для активации гена α-интерферона. Сконструирована нацеливающая конструкция для включения первой направляющей последовательности, гомологичной последовательностям против хода транскрипции этого гена, амплифицируемого маркерного гена, селектируемого маркерного гена, регуляторного района, сайта CAP, донорного сайта сплайсинга, интрона, акцепторного сайта сплайсинга и второй направляющей последовательности, соответствующей последовательностям по ходу транскрипции первой направляющей последовательности. Вторая направляющая последовательность не должна простираться дальше против хода транскрипции, чем до положения - 107, по отношению к нормальному стартовому кодону, во избежание образования нежелательных стартовых кодонов ATG.
В этой стратегии первая и вторая нацеливающая последовательности находятся непосредственно рядом друг с другом в обычном гене-мишени, но это не является необходимым (см. ниже). Здесь описаны амплифицируемые маркерные гены и селектируемые маркерные гены. Амплифицируемый маркерный ген и селектируемый маркерный ген может быть одним и тем же геном, их положения могут быть обращенным и один или оба этих гена могут находиться в интроне нацеливающей конструкции. Селектируемый маркерный ген является необязательным, а амплифицируемый маркерный ген необходим только в том случае, когда желательна амплификация. Включение в констуркцию специфического сайта CAP является необязательным. Необязательно, последовательности экзона из второго гена могут быть включены 3' до акцепторного сайта сплайсинга и 5' до второй направляющей последовательности в нацеливающей конструкции. Регуляторный район, сайт CAP, донорный сайт сплайсинга, интрон и акцепторный сайт сплайсинга могут быть выделены в виде полной единицы из гена фактора элонгации 1α человека (EF-1α; Genbank Seguence HUMEFIA) или немедленно-раннего (предраннего) района цитомегаловируса (CMV; Genbank seguence HEHCMVPI), или эти компоненты могут быть собраны из соответствующих компонентов, выделенных из различных генов.
Геномная ДНК, соответствующая району против хода транскрипции гена α-интеферона для использования в качестве направляющих последовательностей, и сборка нацеливающей конструкции могут быть выполнены с применением способов рекомбинантных ДНК, известных специалистам в этой области знаний. Как описано здесь, множество селектируемых и амплифицируемых маркеров можно использовать в нацеливающих конструкциях, и активацию и амплификацию можно выполнять в большом числе клеточных типов. Трансфекцию первичных, вторичных или иммортализованных клеток человека и выделение гомологично рекомбинантных клеток, экспрессирующих α-интерферон, можно выполнять с использованием способов, описанных в Примере 4, с применением твердофазного иммуноферментного анализа ELISA для α-интерферона человека (Biosource International, Camarillo, CA). В другом варианте гомологично рекомбинантные клетки можно идентифицировать скринингом с использованием полимеразной цепной реакции (ПЦР), как описано в Примере 1g и 1j. Выделение клеток, содержащих амплифицированные копии амплифицируемого маркерного гена и активированного локуса α-интерферона, выполняют, как описано в Примере 6.
В гомологично рекомбинантных клетках продуцируется предшественник мРНК, который включает в себя экзогенный экзон, донорный сайт сплайсинга, интрон, акцепторный сайт сплайсинга, вторую направляющую последовательность и кодирующий район α-интерферона человека и 3' нетранслируемые последовательности (Фиг.11). Сплайсинг этого транскрипта будет генерировать функциональную мРНК, которая может транслироваться с образованием α-интерферона человека.
Размер интрона и, следовательно, положение регуляторного района по отношению к кодирующему району этого гена могут вариировать для оптимизации функции регуляторного района. В нацеливающей конструкции могут присутствовать многочисленные экзоны. Кроме того, вторая направляющая последовательность не обязательно должна лежать смежно с первой направляющей последовательностью или вблизи нее в нормальном гене, так что части нормального района против хода транскрипции гена делетируются при гомологичной рекомбинации.
GM-CSF
Ген GM-CSF человека (Genbank seguence HUMGMCSFG) кодирует белок-предшественник из 144 аминокислот, содержащий сигнальный пептид из 17 аминокислот. Этот ген содержит 4 экзона и 3 интрона, и N-концевые 50 аминокислот предшественника кодируются в первом экзоне. Фиг.12 схематически иллюстрирует стратегию для активации гена GM-CSF. В этой стратегии конструируют направляющую (нацеливающую) конструкцию для включения в нее первой направляющей последовательности, гомологичной последовательностям против хода транскрипции этого гена, амплифицируемого маркерного гена, селектируемого маркерного гена, регуляторного района, гена CAP, экзона, который кодирует аминокислотную последовательность, которая идентична или функционально эквивалентна последовательности первых 50 аминокислот GM-CSF, донорного сайта сплайсинга и второй направляющей последовательности, соответствующей последовательностям по ходу транскрипции первой направляющей последовательности. По этой стратегии гомологично рекомбинантные клетки продуцируют предшественник мРНК, который соответствует экзогенному экзону и донорному сайту сплайсинга, второй направляющей последовательности, любым последовательностям между второй направляющей последовательностью и стартовым кодоном гена GM-CSF, и экзоны, интроны и 3'-нетранслируемый район гена GM-CSF (Фиг.11). Сплайсинг этого транскрипта приводит к слиянию экзогенного экзона с экзоном 2 эндогенного гена GM-CSF, которое при трансляции будет продуцировать GM-CSF.
В этой стратегии первая и вторая направляющая последовательности находятся непосредственно рядом в нормальном гене-мишени, но это не является необходимым (см. ниже). Здесь описаны амплифицируемые маркерные гены и селектируемые маркерные гены, пригодные для отбора. Амплифицируемый маркерный ген и селектируемый маркерный ген могут быть одним и тем же геном или их положения могут быть обращенными. Селектируемый маркерный ген является необязательным, а амплифицируемый маркерный ген необходим только тогда, когда требуется амплификация. Селектируемый маркерный ген и/или амплифицируемый маркерный ген могут быть расположены между донорным сайтом сплайсинга и второй направляющей последовательностью в нацеливающей конструкции. Введение специфического сайта CAP является необязательным. Регуляторный район, сайт CAP и донорный сайт сплайсинга могут быть выделены в виде полной единицы из гена фактора элонгации 1α человека (EF-1α; Genbank seguence HUMEFIA) или немедленно-раннего (предраннего) района цитомегаловируса (CMV; Genbank seguence HEHCMVP1) или эти компоненты могут быть собраны из подходящих компонентов, выделенных из разных генов (таких как промотор mMT-1 и сайт CAP, и экзон 1 и донорный сайт сплайсинга из генов hGH или hEPO).
Можно использовать другие подходы, например, первая и вторая направляющие последовательности могут соответствовать последовательностям в первом интроне гена GM-CSF. В другом варианте, можно использовать направляющую конструкцию, подобную описанной для α-интерферона, которая сконструирована таким образом, что она включает в себя первую направляющую последовательность, гомологичную последовательностям против хода транскрипции гена GM-CSF, амплифицируемый маркерный ген, селектируемый маркерный ген, регуляторный район, сайт CAP, донорный сайт сплайсинга, интрон, акцепторный сайт сплайсинга и вторую направляющую последовательность, соответствующую последовательностям по ходу транскрипции первой направляющей последовательности.
В любом случае вторая направляющая последовательность не должна обязательно лежать непосредственно рядом с первой направляющей последовательностью в нормальном гене или вблизи нее, так что части нормального района против хода транскрипции этого гена делетируются при гомологичной рекомбинации. Кроме того, в нацеливающей конструкции могут присутствовать многочисленные некодирующие или кодирующие экзоны. Получение геномной ДНК, соответствующей районам против хода транскрипции или интронам гена GM-CSF человека, для использования в качестве направляющих последовательностей и сборку нацеливающей конструкции можно выполнять при помощи способов рекомбинантных ДНК, известных специалистам в этой области знаний. Как описано здесь, множество селектируемых и амплифицируемых маркеров можно использовать в нацеливающих конструкциях и активация можно быть достигнута в большом количестве клеточных типов. Трансфекцию первичных, вторичных или иммортализовнных клеток, экспрессирующих GM-CSF, можно выполнять с использованием способов, описанных в Примере 4, с применением твердофазного иммуноферментного анализа ELISA для GM-CSF человека (R&D Systems, minneapolis, MN). Альтернативно, гомологично рекомбинантные клетки можно идентифицировать ПЦР-скринингом, как описано выше. Выделение клеток, содержащих амплифицированные копии амплифицируемого маркерного гена и активированного локуса GM-CSF, выполняют, как описано выше.
G-CSF
Ген G-CSF человека (Genbank seguence HUMGCSFG) кодирует белок-предшественник из 204-207 аминокислот, содержащий сигнальный пептид из 30 аминокислот. Этот ген содержит 5 экзонов и 4 интрона, первый экзон кодирует 13 аминокислот сигнального пептида. Фиг. 13 схематически иллюстрирует стратегию активации гена G-CSF. Конструировали нацеливающую конструкцию, которая включает в себя первую направляющую последовательность, гомологичную последовательностям против хода транскрипции этого гена, амплифицируемый маркерный ген, селектируемый маркерный ген, регуляторный район, сайт CAP, экзон, кодирующий аминокислотную последовательность, которая идентичная или функционально эквивалентна последовательности первых 13 аминокислот сигнального пептида G-CSF, донорный сайт сплайсинга и вторую направляющую последовательность, соответствующую последовательности по ходу транскрипции первой направляющей последовательности. При помощи этой стратегии гомологично рекомбинантные клетки продуцируют предшественник мРНК, который соответствует экзогенному экзону и донорному сайту сплайсинга, второй направляющей последовательности, любым последовательностям, между второй направляющей последовательностью и стартовым кодоном гена G-CSF и экзоном, интронам и 3'-нетранслируемому району гена G-CSF (Фиг.13). Сплайсинг этой матрицы приводит к слиянию экзогенного экзона с экзоном 2 эндогенного гена G-CSF, которое при трансляции будет продуцировать G-CSF. Возможность функциональной замены первых 13 аминокислот сигнального пептида нормального G-CSF аминокислотами, присутствующими в экзогенном экзоне, позволяет модифицировать сигнальный пептид и, следовательно, секреторные свойства продуцируемого белка.
В этой стратегии первая и вторая направляющие последовательности находятся непосредственно рядом в нормальном гене-мишени, но это не является обязательным. Вторая направляющая последовательность не должна обязательно лежать смежно или вблизи от первой направляющей последовательности в нормальном гене, так что части района против хода транскрипции этого гена делетируются при гомологичной рекомбинации. Амплифицируемый маркерный ген и селектируемый маркерный ген могут быть одним и тем же геном или их положения могут быть обращенными. Селектируемый маркерный ген является необязательным, а амплифицированный маркерный ген необходим только в том случае, когда желательна амплификация. Селектируемый маркер и/или амплифицируемый маркер могут быть расположены между донорным сайтом сплайсинга и второй направляющей последовательностью в нацеливающей конструкции. Введение специфического сайта CAP является необязательным. Регуляторный район, САР-сайт и донорный сайт сплайсинга могут быть выделены в виде полной единицы из гена фактора элонгации 1α человека (EF-1α; Genbank seguence HUMEFIA) или немедленно-раннего (предраннего) района цитомегаловируса (CMV; Genbank seguence HEHCMVP1), или эти компоненты могут быть собраны из соответствующих компонентов из различных генов, таких как промотор mMT-1 и сайт CAP, и экзон 1 и донорный сайт сплайсинга из генов hGH или ЕРО. Множественные экзогенные экзоны, кодирующие или некодирующие, могут быть использованы в нацеливающей конструкции при условии, что стартовый кодон ATG, который при сплайсинге должен быть внутри рамки со зрелым белком, был включен в один из этих экзонов.
Можно использовать другие подходы, например, первая и вторая направляющие последовательности могут соответствовать последовательностям в первом интроне гена G-CSF. Альтернативно, можно использовать нацеливающую конструкцию, подобную описанной для α-интерферона, которая сконструирована таким образом, что она включает в себя первую направляющую последовательность, гомологичную последовательностям против хода транскрипции гена G-CSF, амплифицируемый маркерный ген, селектируемый маркерный ген, регуляторный район, сайт CAP, донорный сайт сплайсинга, интрон, акцепторный сайт сплайсинга и вторую направляющую последовательность, соответствующую последовательностям по ходу транскрипции первой направляющей последовательности.
Получение геномной ДНК, соответствующей районам против хода транскрипции или интронам гена G-CSF человека, для использования в качестве направляющих последовательностей и сборку нацеливающей конструкции можно выполнять при помощи способов рекомбинантных ДНК, известных специалистам в этой области знаний. Как описано здесь, большое количество селектируемых и амплифицируемых маркеров можно использовать в нацеливающих конструкциях и активация может быть достигнута в большом числе клеточных типов. Трансфекцию первичных, вторичных или иммортализованных клеток человека и выделение гомологично рекомбинантных клеток, экспрессирующих G-CSF, можно выполнять при помощи способов, описанных в Примере 4, с использованием анализа ELISA для G-CSF человека (R&D Systems, MN). В другом варианте, гомологично рекомбинантные клетки могут быть идентифицированы ПЦР-скринингом, как описано выше. Выделение клеток, содержащих амплифицированные копии амплифицируемого маркерного гена и активированного локуса G-CSF, выполняют, как описано выше.
FSHβ
Ген FSHβ человека (Genbank seguence HUMFSHI) кодирует белок-предшественник из 129 аминокислот, содержащий сигнальный пептид из 16 аминокислот. Этот ген содержит три экзона и два интрона, причем первый экзон является некодирующим. Активацию гена FSHβ можно выполнять при помощи ряда стратегий. Одна стратегия представлена на Фиг.14. В этой стратегии, конструируют нацеливающую конструкцию, которая включает в себя первую направляющую последовательность, гомологичную последовательностям против хода транскрипции этого гена, амплифицируемый маркерный ген, селектируемый маркерный ген, регуляторный район, сайт CAP, экзон, донорный сайт сплайсинга и вторую направляющую последовательность, соответствующую последовательностям по ходу транскрипции первой направляющей последовательности. По этой стратегии, гомологично рекомбинантные клетки продуцируют предшественник мРНК, который соответствует экзогенному экзону и донорному сайту сплайсинга, второй направляющей последовательности, любым последовательностям между второй направляющей последовательностью и стартовым кодоном гена FSHβ, и экзоны, интроны и 3'-нетранслируемые районы гена FSHβ (Фиг.14). Сплайсинг этого транскрипта приводит к слиянию экзогенного экзона с экзоном 2 эндогенного гена FSHβ, которое при трансляции может продуцировать FSHβ. В этой стратегии первая и вторая направляющие последовательности находятся непосредственно рядом в нормальном гене-мишени, но это не является обязательным (см. ниже).
Могут быть использованы другие подходы, например, первая и вторая направляющие последовательности могут соответствовать последовательностям первого интрона гена FSHβ. В другом варианте можно использовать нацеливающую конструкцию, подобную описанной для α-интерферона. В этой стратегии нацеливающая конструкция сконструирована таким образом, что она включает в себя первую направляющую последовательность, гомологичную последовательностям против хода транскрипции гена FSHβ, амплифицируемый маркерный ген, селектируемый маркерный ген, регуляторный район, сайт CAP, донорный сайт сплайсинга, интрон, акцепторный сайт сплайсинга и вторую направляющую последовательность, соответствующую последовательностям по ходу транскрипции от первой направляющей последовательности. Вторая направляющая последовательность не должна простираться дальше против хода транскрипции, чем до положения -40 по отношению к сайту инициации транскрипции нормального FSHβ, для того чтобы избежать появления нежелательных стартовых кодонов ATG. В этих гомологично рекомбинантных клетках продуцируется предшественник мРНК, который включает в себя экзогенный экзон, донорный сайт сплайсинга, интрон, акцепторный сайт сплайсинга, вторую направляющую последовательность и кодирующие экзоны FSHβ человека, интрон и 3'-нетранслируемые последовательности. Сплайсинг этой матрицы будет генерировать функциональную мРНК, которая может транслироваться с образованием FSHβ человека. Размер интрона и, следовательно, положение регуляторного района относительно кодирующего района этого гена могут изменяться для оптимизации функции регуляторного района.
В любой стратегии активации вторая направляющая последовательность не должна обязательно лежать непосредственно смежно или близко к первой направляющей последовательности в нормальном гене, так что части района против хода транскрипции нормального гена делетируются при гомологичной рекомбинации. Кроме того, одна направляющая последовательность может находиться против хода транскрипции гена, а одна может находиться в экзоне или интроне гена FSHβ.
Амплифицируемый маркерный ген и селектируемый маркерный ген могут быть одним и тем же геном, их положения могут быть обращенными и один из них или оба могут быть расположены в интроне нацеливающей конструкции. Здесь описаны амплифицируемые маркерные гены и селектируемые маркерные гены, пригодные для отбора. Селектируемый маркерный ген является необязательным, а амплифицируемый маркерный ген необходим только в том случае, когда желательна амплификация. Введение специфического сайта CAP является необязательным. Иногда (необязательно) последовательности экзонов из другого гена могут быть включены 3' относительно акцепторного сайта сплайсинга и 5' относительно второй направляющей последовательности в нацеливающей конструкции. Регуляторный район, сайт CAP, экзон, донорный сайт сплайсинга могут быть выделены в виде полной единицы из гена фактора элонгации 1α человека (ЕР-1α; Genbank seguence HUMEFIA) или немедленно-раннего (предраннего) района цитомегаловируса (CMV; Genbank seguence HEHCMVPI) или эти компоненты могут быть собраны из соответствующих компонентов, выделенных из разных генов. В любом случае экзогенный экзон может быть таким же или отличающимся от первого экзона нормального гена FSBβ, и в нацеливающей конструкции могут присутствовать многочисленные экзоны.
Получение геномной ДНК, соответствующей району против хода транскрипции гена FSHβ, для использования в качестве направляющих последовательностей и сборку нацеливающей конструкции можно выполнять при помощи способов рекомбинантных ДНК, известных специалистам в этой области. Как описано здесь, большое количество селектируемых и амплифицируемых маркеров можно использовать в нацеливающих конструкциях, и активация может быть достигнута в большом количестве клеточных типов. Если желательно, продукт активированного гена FSHβ может быть получен в клеточном типе, который экспрессирует α-субъединицу гликопротеина человека, продукт которой образует гетеродимер с продуктом гена FSHβ. Может быть использован встречающийся в природе клеточный штамм или линия. Альтернативно, ген α-субъединицы гликопротеина человека (Genbank seguence HUMG1УCA1 ) может КО-ЭКСПРЕССИРОВАТЬСЯ С ПРОДУКТОМ ГЕНА FSHβ, причем такая совместная экспрессия выполняется путем экспрессии гена α-субъединицы гликопротеина человека или кДНК под контролем подходящего промотора, или посредством активации гена α-субъединицы гликопротеина человека описанными здесь способами. Трансфекцию первичных, вторичных или иммортализованных клеток человека и выделение гомологично рекомбинированных клеток, экспрессирующих FSHβ, можно выполнять при помощи описанных выше способов с применением анализа ELISA для FSHβ человека (accurate Chemical and Scientific, Westbury, NY). В другом варианте, гомологично рекомбинантные клетки можно идентифицировать при помощи ПЦР-скрининга, описанного выше. Выделение клеток, содержащих амплифицированные копии амплифицируемого маркерного гена и активированного локуса FSHβ, выполняют, как описано выше.
Эквиваленты
Специалисты в этой области смогут найти или будут способны установить не более чем рутинным экспериментированием многие эквиваленты специфических вариантов этого изобретения, описанных здесь. Такие эквиваленты должны охватываться следующей формулой изобретения.
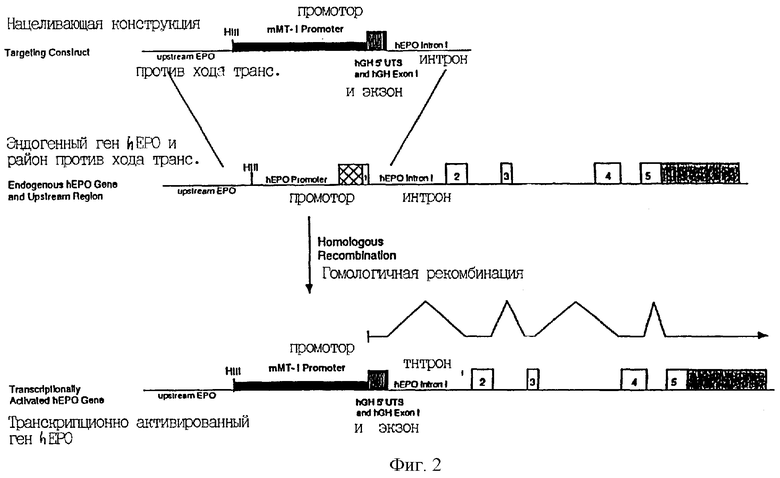
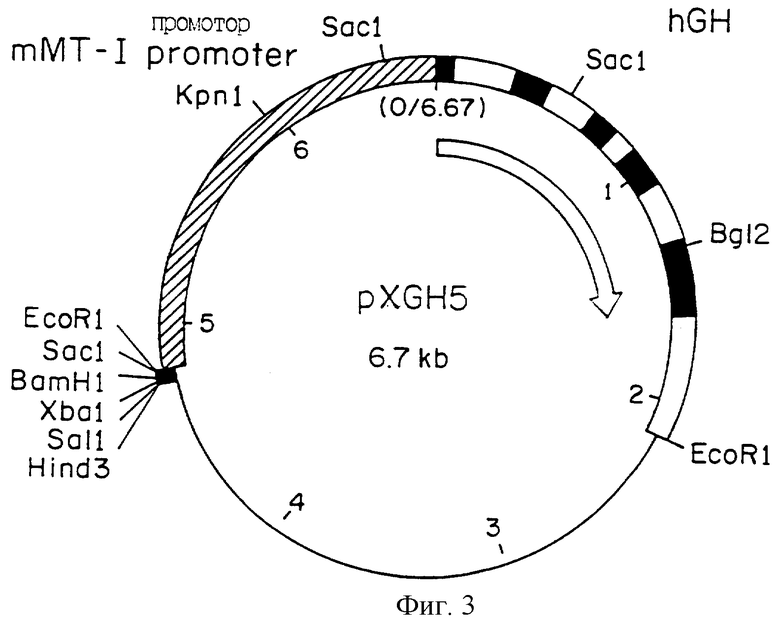
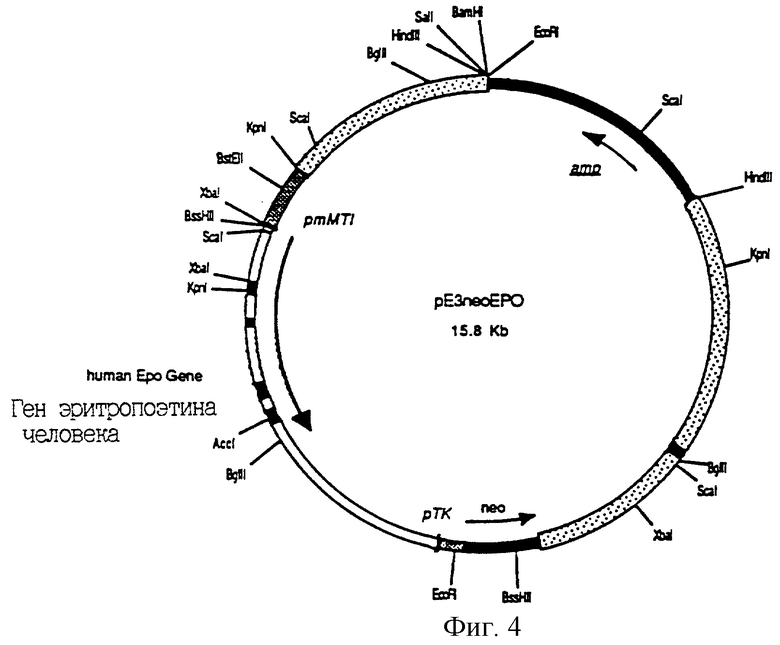
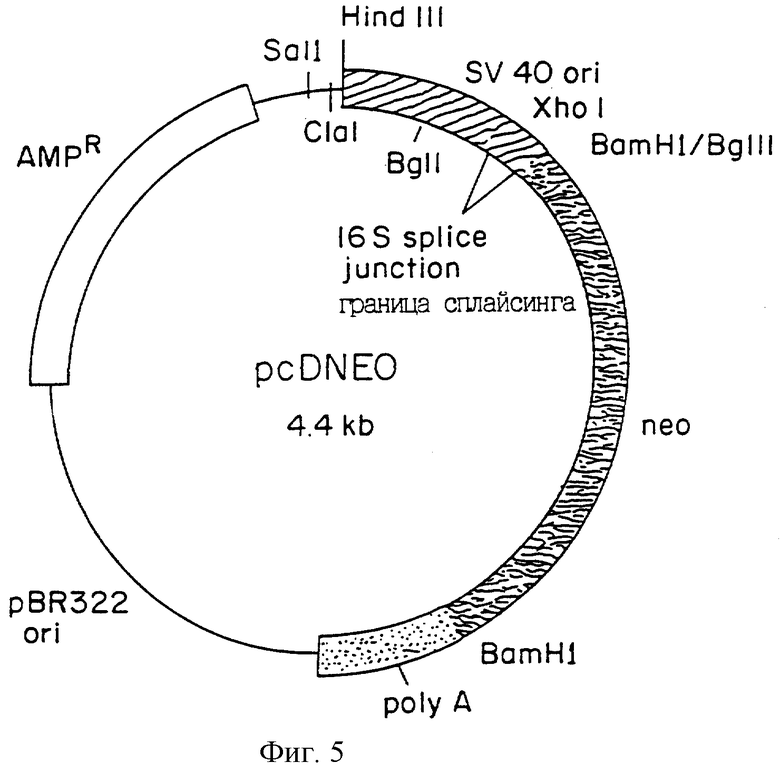

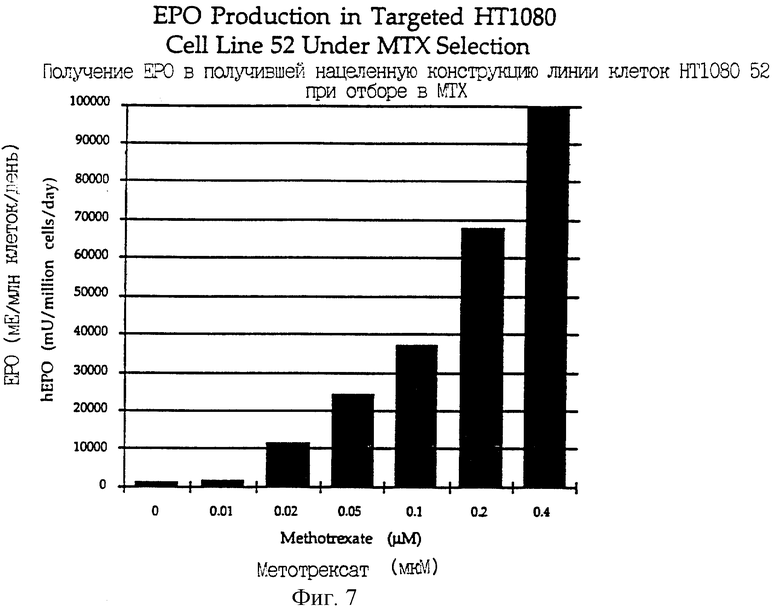
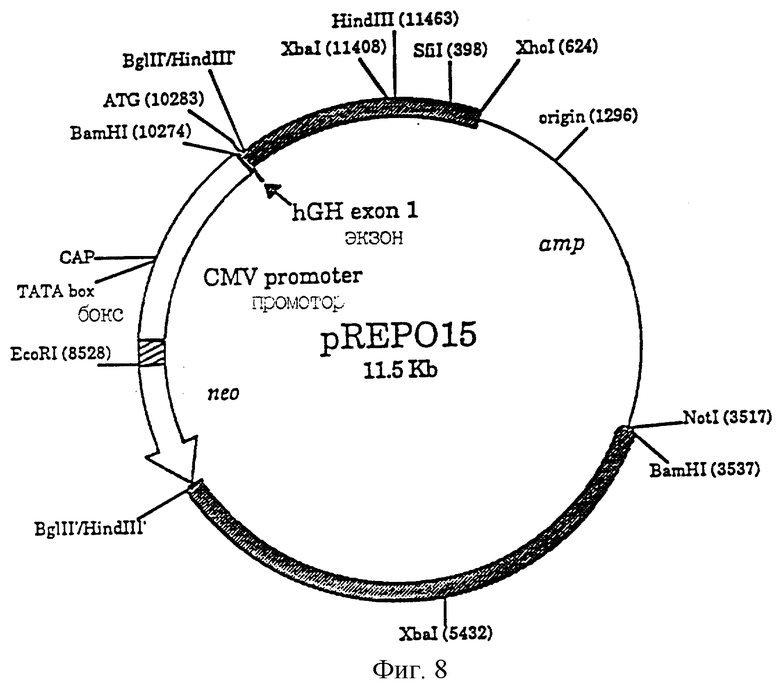
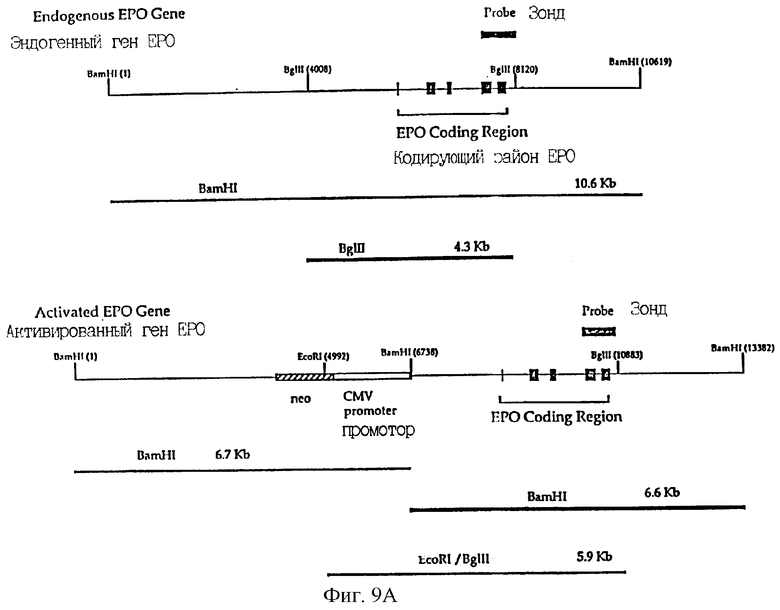
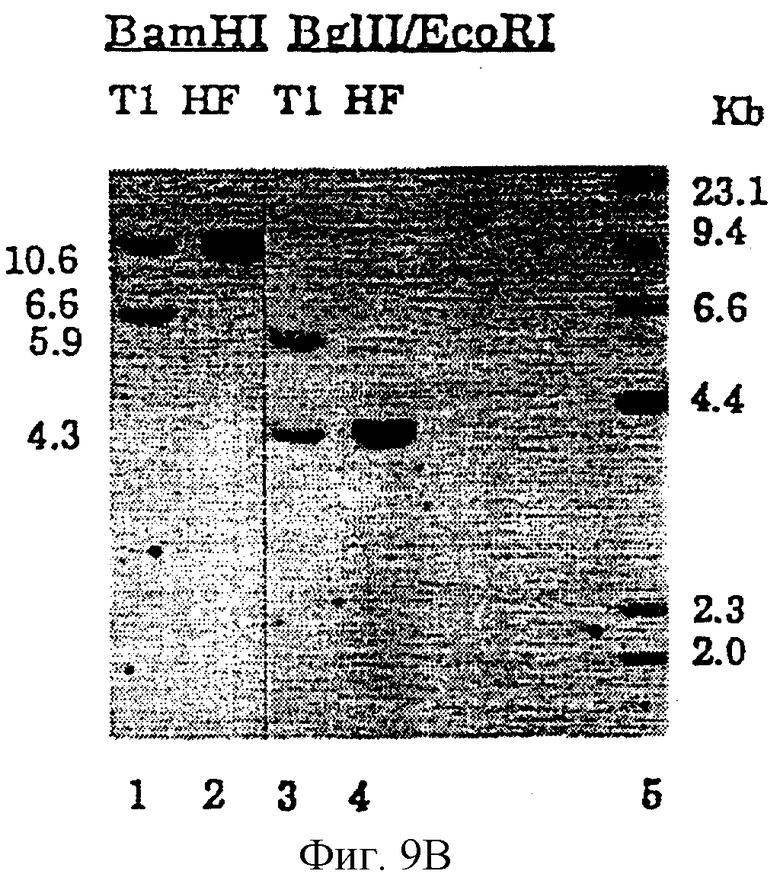
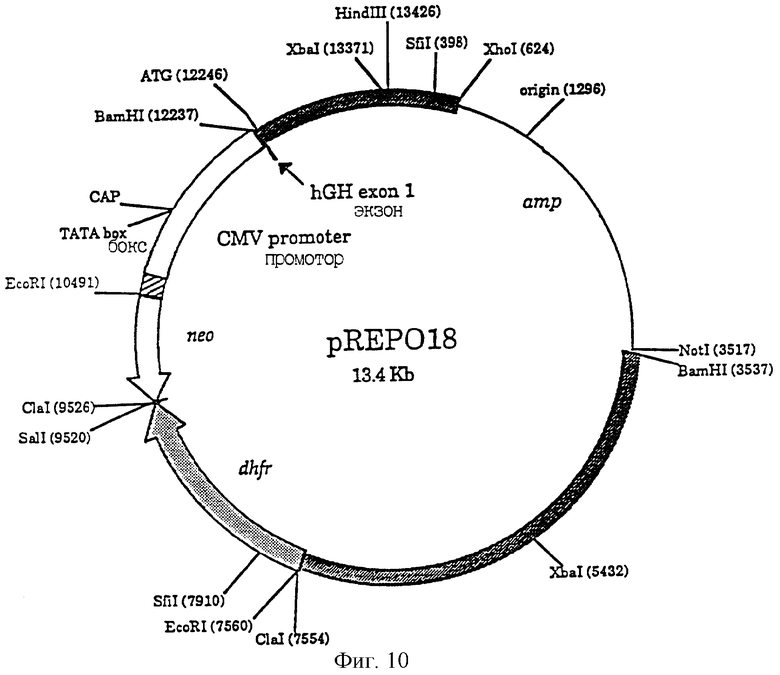
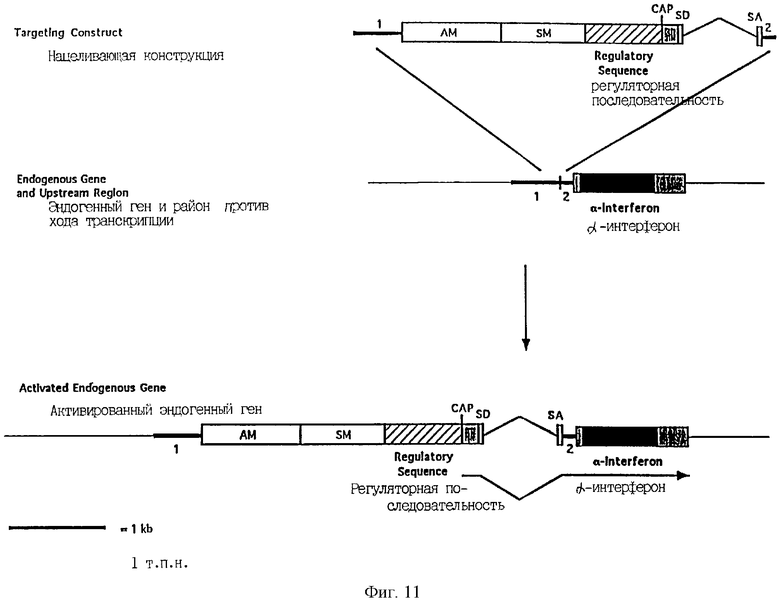
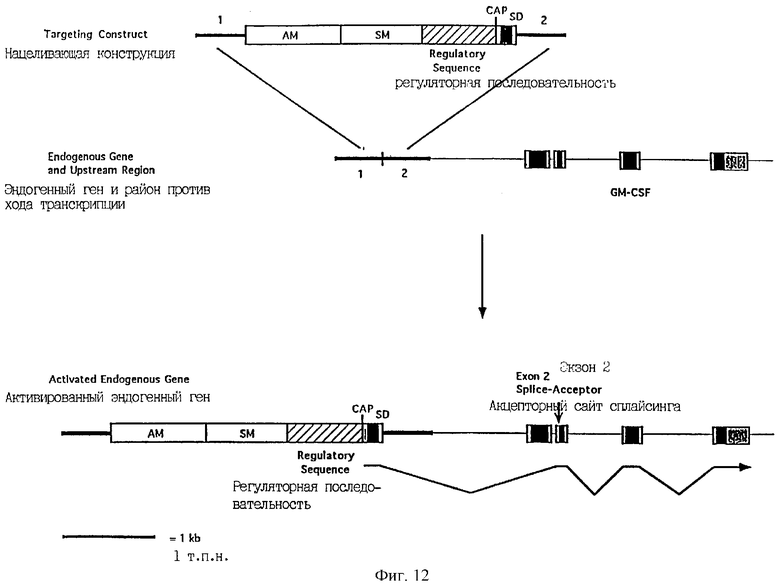
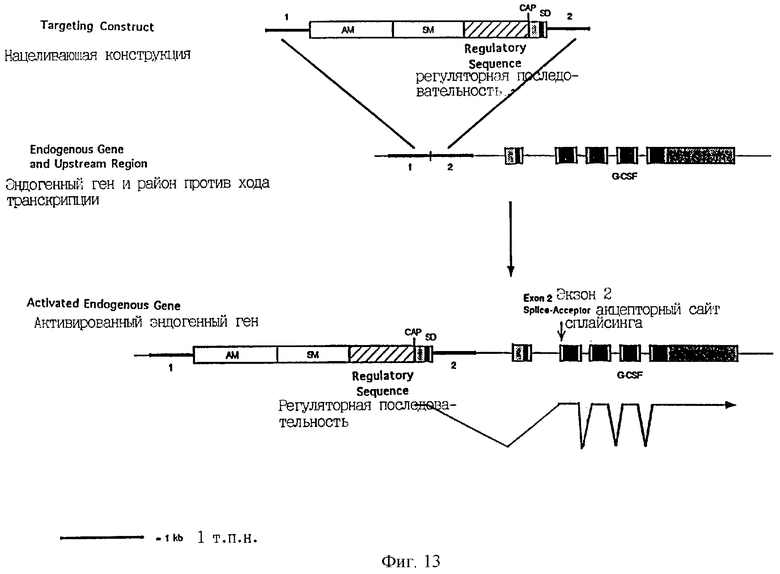
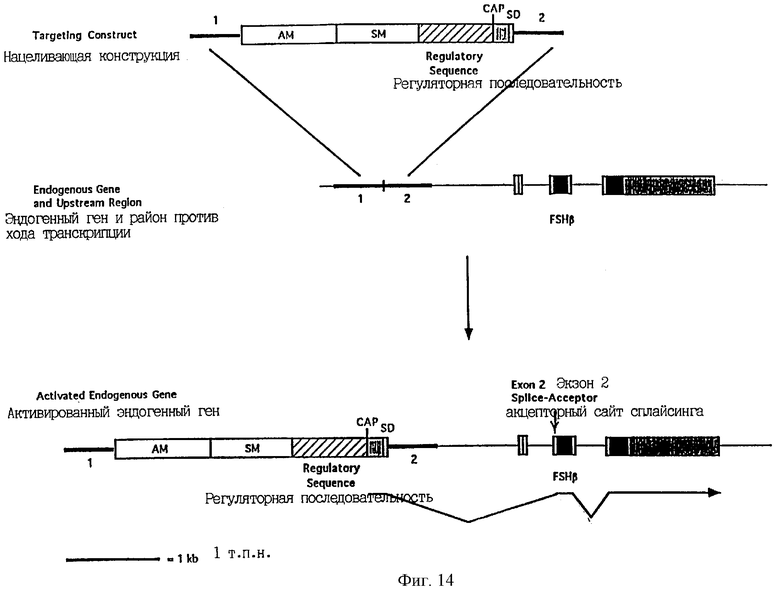
Изобретение относится к области генной инженерии и может быть использовано в медико-биологической промышленности для получения белков терапевтического назначения, а также в медицине как средство для генной терапии. Предложены варианты конструкции ДНК, предназначенной для интегрирования в определенный сайт (сайт-мишень) хромосомной ДНК клетки, располагающийся перед интересующим эндогенным геном, за счет гомологичной рекомбинации между последовательностью указанного сайта-мишени и «направляющей» последовательностью, входящей в структуру данной конструкции. ДНК-конструкция также включает экзогенный промотор и экзон с неспаренным донорным сайтом сплайсинга или экзон с донорным сайтом и интрон с акцепторным сайтом сплайсинга. В результате встраивания такой конструкции в геном клетки образуются новые (не существующие в природе) транскрипционные единицы и обеспечивается возможность направленного изменения экспрессии любого эндогенного гена. 59 з.п. ф-лы, 15 ил., 7 табл.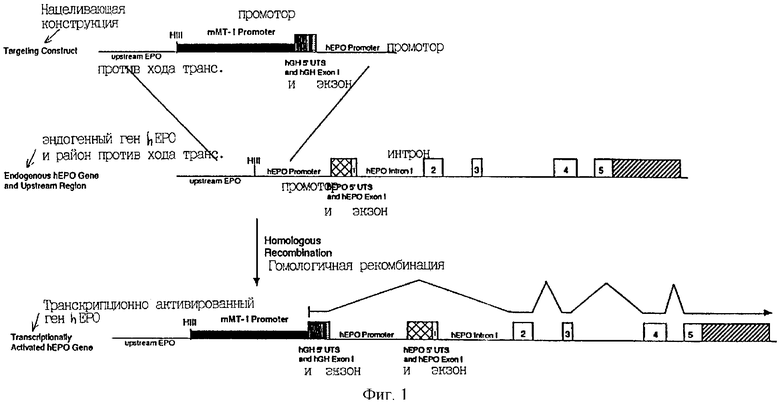
(a) направляющую последовательность, гомологичную сайту-мишени;
(b) экзогенный промотор;
(c) функционально связанный с указанным промотором экзон с неспаренным донорным сайтом сплайсинга на 3'-конце экзона;
где в результате гомологичной рекомбинации направляющей последовательности с сайтом-мишенью хромосомная ДНК клетки включает производный от конструкции экзон в дополнение ко всем эндогенным экзонам гена-мишени и где ген-мишень кодирует белок, выбираемый из группы, включающей α-интерферон, β-интерферон, γ-интерферон, интерлейкин-1, интерлейкин-2, интерлейкин-3, интерлейкин-6, интерлейкин-11, интерлейкин-12, колониестимулирующий фактор гранулоцитов (G-CSF), колониестимулирующий фактор макрофагов (M-CSF), колониестимулирующий фактор гранулоцитов/макрофагов (GM-CSF), FSHβ, TGF-β, фактор некроза опухолей, глюкагон, фактор-2 роста костей, фактор-7 роста костей, TSH-β, протеинкиназу-С, урокиназу, антитромбин-III, ДНКазу, α-галактозидазу, тирозингидроксилазу, фактор V свертывания крови, фактор VII свертывания крови, фактор Х свертывания крови, фактор XIII свертывания крови, антагонист IL-2 и растворимый CD4.
(a) направляющую последовательность, гомологичную сайту-мишени;
(b) экзогенный промотор;
(c) функционально связанный с указанным промотором экзон с неспаренным донорным сайтом сплайсинга на 3'-конце экзона;
где в результате гомологичной рекомбинации направляющей последовательности с сайтом-мишенью хромосомная ДНК клетки включает производный от конструкции экзон в дополнение ко всем эндогенным экзонам гена-мишени и где экзогенный промотор является промотором гена аденовируса, промотором гена коллагена, промотором гена актина, промотором гена HMG-KoA-редуктазы или промотором гена EF-1α.
(a) получение клетки, геном которой включает
(i) ген-мишень, содержащий эндогенный регуляторный участок;
(ii) сайт-мишень, находящийся выше эндогенного сайта инициации транскрипции указанного гена-мишени;
(b) получение конструкции ДНК, включающей
(i) направляющую последовательность, гомологичную сайту-мишени;
(ii) экзогенный промотор;
(iii) функционально связанный с указанным промотором экзон с неспаренным, донорным сайтом сплайсинга на 3'-конце экзона;
(c) трансфекция клетки конструкцией ДНК с получением таким образом трансфицированной клетки;
(d) поддержание трансфицированной клетки в условиях, пригодных для гомологичной рекомбинации, с получением таким образом гомологично рекомбинированной клетки, геном которой включает экзогенный промотор, производный от конструкции экзон и производный от конструкции донорный сайт сплайсинга в дополнение ко всем эндогенным экзонам гена-мишени;
(e) поддержание гомологично рекомбинированной клетки в условиях, пригодных для транскрипции под контролем экзогенного промотора с целью получения транскрипта, производного от конструкции экзона, гена-мишени и любой последовательности, располагающейся между производным от конструкции экзоном и геном-мишенью, где РНК транскрипта, соответствующего производному от конструкции донорному сайту сплайсинга, направляет сплайсинг к акцепторному сайту сплайсинга в составе транскрипта, который соответствует сайту в пределах гена-мишени, и где ген-мишень кодирует белок, выбираемый из группы, включающей α-интерферон, β-интерферон, γ-интерферон, факторы роста нервной ткани, FSHβ, TGF-β, фактор некроза опухолей, глюкагон, фактор-2 роста костей, фактор-7 роста костей, TSH-β, интерлейкин-1, интерлейкин-2, интерлейкин-3, интерлейкин-6, интерлейкин-11, интерлейкин-12, G-CSF, M-CSF, GM-CSF, протеинкиназу-С, урокиназу, антитромбин-III, ДНКазу, α-галактозидазу, тирозингидроксилазу, фактор V свертывания крови, фактор VII свертывания крови, фактор Х свертывания крови, фактор XIII свертывания крови, антагонист IL-2 и растворимый CD4.
(f) поддержание гомологично рекомбинированной клетки в условиях, пригодных для сплайсинга и трансляции транскрипта и
(g) подтверждение того, что был получен продукт трансляции транскрипта.
(a) получение клетки, геном которой включает
(i) ген-мишень, содержащий эндогенный регуляторный участок;
(ii) сайт-мишень, находящийся выше эндогенного сайта инициации транскрипции указанного гена-мишени;
(b) получение конструкции ДНК, включающей
(i) направляющую последовательность, гомологичную сайту-мишени;
(ii) экзогенный промотор;
(iii) функционально связанный с указанным промотором экзон с неспаренным донорным сайтом сплайсинга на 3'-конце экзона;
(c) трансфекция клетки конструкцией ДНК с получением таким образом трансфицированной клетки;
(d) поддержание трансфицированной клетки в условиях, пригодных для гомологичной рекомбинации, с получением таким образом гомологично рекомбинированной клетки, геном которой включает экзогенный промотор, производный от конструкции экзон и производный от конструкции донорный сайт сплайсинга так, что все они расположены выше эндогенного сайта инициации транскрипции гена-мишени;
(e) поддержание гомологично рекомбинированной клетки в условиях, пригодных для транскрипции под контролем экзогенного промотора с целью получения транскрипта производного от конструкции экзона, гена-мишени и любой последовательности, располагающейся между производным от конструкции экзоном и геном-мишенью, где РНК транскрипта, соответствующего производному от конструкции донорному сайту сплайсинга, направляет сплайсинг к акцепторному сайту сплайсинга в составе транскрипта, который соответствует сайту в пределах гена-мишени, и где экзогенный промотор является промотором гена аденовируса, промотором гена коллагена, промотором гена актина, промотором гена иммуноглобулина, промотором гена HMG-KoA-редуктазы или промотором гена EF-lα.
(a) трансфекция клетки конструкцией ДНК, включающей
(i) направляющую последовательность, гомологичную последовательности, находящейся выше эндогенного сайта инициации транскрипции указанного гена-мишени;
(ii) экзогенный промотор;
(iii) функционально связанные с указанным промотором экзон с донорным сайтом сплайсинга в составе его 3'-конца и следующий за ним интрон с акцепторным сайтом сплайсинга;
с получением в результате трансфицированной клетки;
(b) поддержание трансфицированной клетки в условиях, пригодных для гомологичной рекомбинации с получением таким образом гомологично рекомбинированной клетки, в которой экзогенный промотор помещен так, чтобы регулировать транскрипцию последовательности, включающей (iii) вместе с кодирующей последовательностью гена-мишени;
(с) поддержание гомологично рекомбинированной клетки в условиях, пригодных для транскрипции под контролем экзогенного промотора, с получением таким образом транскрипта.
(f) поддержание гомологично рекомбинированной клетки в условиях, пригодных для сплайсинга и трансляции транскрипта; (g) подтверждение того, что был получен продукт трансляции транскрипта.
(a) получение клетки, геном которой включает
(i) ген-мишень, содержащий эндогенный регуляторный участок; и
(ii) сайт-мишень, находящийся выше эндогенного сайта инициации транскрипции указанного гена-мишени;
(b) получение конструкции ДНК, включающей
(i) направляющую последовательность, гомологичную сайту-мишени;
(ii) экзогенный промотор;
(iii) функционально связанные с указанным промотором экзон с донорным сайтом сплайсинга в составе его 3'-конца и следующий за ним интрон с акцепторным сайтом сплайсинга;
(c) трансфекция клетки конструкцией ДНК с получением таким образом трансфицированной клетки;
(d) поддержание трансфицированной клетки в условиях, пригодных для гомологичной рекомбинации, с получением таким образом гомологично рекомбинированной клетки, геном которой включает (ii)-(iii) конструкции ДНК в дополнение ко всем эндогенным экзонам гена-мишени.
(a) трансфекция клетки конструкцией ДНК, включающей
(i) направляющую последовательность, гомологичную геномной ДНК выше кодирующего сегмента гена-мишени;
(ii) промотор актина или коллагена;
(iii) функционально связанный с указанным промотором экзон с неспаренным донорным сайтом сплайсинга на 3'-конце экзона;
с получением таким образом трансфицированной клетки;
(b) поддержание трансфицированной клетки в условиях, пригодных для гомологичной рекомбинации с получением таким образом гомологично рекомбинированной клетки; и
(c) поддержание гомологично рекомбинированной клетки в условиях, пригодных для экспрессии гена-мишени под контролем промотора.
| Огнетушитель | 0 |
|
SU91A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| Экономайзер | 0 |
|
SU94A1 |

