ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет предварительной заявки США № 61/392333 от 12 октября 2010, раскрытие которой, таким образом, включено путем ссылки во всей своей полноте.
ЗАЯВЛЕНИЕ ПРАВ НА ИЗОБРЕТЕНИЯ НА БАЗЕ ИССЛЕДОВАНИЙ, СПОНСИРУЕМЫХ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА
[0002] Не применимо.
ОБЛАСТЬ ТЕХНИКИ
[0003] Настоящее раскрытие относится к области генной модификации и лечения гемофилии.
ПРЕДПОСЫЛКИ
[0004] Гемофилия B является генетическим нарушением системы свертывания крови, характеризуемым кровотечением в суставы и мягкие ткани и чрезмерным кровотечением в любом месте травмирования или подвергаемом хирургической операции. Хотя гемофилия B клинически неотличима от гемофилии A, при гемофилии A фактор VIII (FVIII) присутствует в недостаточном количестве или отсутствует, а у пациентов с гемофилией В фактор IX (FIX или F.IX) присутствует в недостаточном количестве или отсутствует. Фактор IX кодирует одну из сериновых протеаз, вовлеченных в систему свертывания крови, и было показано, что восстановление даже 3% от нормальных уровней циркулирующего белка фактора IX дикого типа может предотвратить спонтанное кровотечение.
[0005] Описана генная терапия, включающая протоколы печень-направленной генной терапии и прямые внутримышечные инъекции, касающиеся введения плазмиды и других векторов (например, AAV), кодирующих функциональный белок FIX, для лечения гемофилии B. См., например, патент США № 6936243; Lee и другие (2004) Pharm. Res. 7:1229-1232; Graham и другие (2008) Genet Vaccines Ther. 3:6-9. Однако в этих протоколах образование ингибирующих анти-фактор IX (анти-FIX) антител и антител против средства доставки остается главным осложнением для лечения гемофилии B, основанного на замещении белка FIX.
[0006] В патентной заявке США № 12/798749 описана целенаправленная интеграция функционального белка FIX в выделенные стволовые клетки и лечение гемофилии B посредством введения FIX-продуцирующих стволовых клеток пациентам, нуждающимся в лечении.
[0007] Однако сохраняется потребность в дополнительных композициях и способах лечения гемофилии у пациентов с этим заболеванием.
КРАТКОЕ СОДЕРЖАНИЕ
[0008] Здесь раскрыты способы и композиции для направленной интеграции последовательности, кодирующей функциональный белок FIX, с целью лечения гемофилии B. В частности, способы включают введение нуклеаз, которые опосредуют направленную вставку последовательности, кодирующей функциональный белок FIX, в геном клеток с целью облегчения болезни.
[0009] Один описанный здесь аспект касается ДНК-связывающего домена (например, белка «цинкового пальца» (ZFP) или белка TALE), который связывается с сайтом-мишенью в области, представляющей интерес (например, гене фактора IX), в геноме, где ZFP содержит один или более генно-инженерных связывающих доменов «цинковый палец», и TALE содержит один или более генно-инженерных ДНК-связывающих доменов TALE. В одном варианте ДНК-связывающий домен представляет собой нуклеазу, например, ZFP – цинк-пальцевую нуклеазу (ZFN) и TALE - нуклеазу TALE (TALEN), которая расщепляет представляющую интерес целевую геномную область, где ZFN или TALEN содержат один или более генно-инженерных ДНК-связывающих доменов и нуклеазный расщепляющий домен или расщепляющий полудомен. Расщепляющие домены и полудомены можно получить, например, из различных рестрикционных эндонуклеаз и/или хоминг-эндонуклеаз. В одном варианте расщепляющие полудомены являются производными рестрикционных эндонуклеаз типа IIS (например, FokI). В некоторых вариантах домен «цинковый палец» распознает целевой сайт в эндогенном гене FIX, например, домен «цинковый палец», как показано в таблице 1 (или домен "цинковый палец", который связан с целевым сайтом, как показано в таблице 1).
[0010] Другой описанный здесь аспект касается полинуклеотида, кодирующего одну или более описанных здесь ZFN и/или TALEN. Полинуклеотид может представлять собой, например, мРНК.
[0011] Другой описанный здесь аспект касается вектора экспрессии ZFN и/или TALEN, содержащего полинуклеотид, кодирующий одну или более описанных здесь ZFN или TALEN, функционально связанных с промотором. В одном варианте вектор экспрессии представляет собой вирусный вектор. В одном аспекте вирусный вектор демонстрирует тканеспецифический тропизм.
[0012] Другой описанный здесь аспект касается клетки-хозяина, содержащей один или более векторов экспрессии ZFN и/или TALEN. Клетки-хозяева можно стабильно трансформировать или временно трансфицировать (или то и другое) посредством одного или большего количества векторов экспрессии ZFP или TALEN. В одном варианте клетка-хозяин представляет собой эмбриональную стволовую клетку. В других вариантах один или более векторов экспрессии ZFP и/или TALEN экспрессируют один или более ZFN или TALEN в клетке-хозяине. В другом варианте клетка-хозяин может дополнительно включать экзогенные полинуклеотидные донорные последовательности. В любом из описанных здесь вариантов клетка-хозяин может включать клетку печени, мышечную клетку, стволовую клетку или эмбриональную клетку. Клетки могут быть из любого организма, например, клетки человека, примата, отличного от человека, мыши, крысы, кролика, кошки, собаки или клетки других млекопитающих.
[0013] Другой представленный здесь аспект касается способов лечения гемофилии B с применением нуклеаз для интеграции последовательности, кодирующей белок FIX, в клетке у нуждающегося в этом субъекта. В некоторых вариантах FIX-кодирующую последовательность доставляют с помощью вирусного вектора, невирусного вектора (например, плазмиды) и/или их комбинации. В некоторых вариантах вектор включает вектор AAV, например, AAV8. В некоторых вариантах нуклеазы и/или FIX-кодирующие последовательности доставляют посредством внутривенного (например, через воротную вену) введения в печень интактного животного.
[0014] В любом из описанных здесь способов нуклеаза может представлять собой одну или более из цинк-пальцевых нуклеаз «цинковый палец», одну или более из хоминг-эндонуклеаз (мегануклеаз) и/или одну или более из TAL-эффекторный домен нуклеаз ("TALEN"). Нуклеазы (например, ZFN или TALEN), которые здесь описаны, могут связывать и/или расщеплять представляющую интерес область в кодирующей или некодирующей области внутри или рядом с геном, например, лидерную последовательность, концевую последовательность или интрон, или внутри нетранскрибируемой области, выше или ниже кодирующей области. В некоторых вариантах ZFN связывает или расщепляет ген эндогенного фактора IΧ (мутант или дикий тип). В других вариантах ZFN или TALEN связывают или расщепляют ген «безопасную гавань» (например, любой ген, разрушение которого не является токсичным или разрушительным для клетки), например ген CCR5, ген PPP1R12C (также известный как AAV S1) или ген Rosa. См., например, патентные публикации США № 20080299580; 20080159996 и 201000218264.
[0015] Кроме того, любой из описанных здесь способов может также включать дополнительные стадии, в том числе частичную гепатэктомию или лечение вспомогательными агентами, которые усиливают трансдукцию и/или индуцируют деление клеток печени. Примеры вспомогательных агентов включают гамма-облучение, УФ-облучение, меченные тритием нуклеотиды, например, тимидин, цис-платину, этопозид, гидроксимочевину, афидиколин, преднизолон, четыреххлористый углерод или аденовирус.
[0016] Описанные здесь способы можно осуществить in vitro, ex vivo или in vivo. В некоторых вариантах композиции вводят живому, интактному млекопитающему. В момент введения млекопитающее может быть на любой стадии развития, например, эмбрион, плод, новорожденное, инфантильное, ювенильное или взрослое. Кроме того, клетки-мишени могут быть здоровыми или больными. В некоторых вариантах композиции (например, полинуклеотиды, кодирующие нуклеазу(ы) и/или FIX-кодирующие последовательности) вводят в печень живого животного, например, посредством инъекции в воротную вену. В других вариантах одну или более композиций вводят внутривенно (не через воротную вену, например, посредством инъекции в хвостовую вену), внутриартериально, внутрибрюшинно, в паренхиму печени (например, посредством инъекции), в печеночную артерию (например, посредством инъекции) и/или через билиарное дерево (например, посредством инъекции).
[0017] Для направленного воздействия композиций на клетки конкретного типа, например, гепатоциты, можно объединить одну или более из вводимых композиций с хоминг-агентом, который связывается именно с поверхностными рецепторами клетки. Например, вектор может быть соединен с лигандом (например, галактозой), для которого определенные клетки печеночной системы имеют рецепторы. Связывание может быть ковалентным, например, через сшивающий агент, такой как глутаровый альдегид, или нековалентным, например, привязка авидинированного лиганда к биотинилированному вектору. Другой вид ковалентной связи обеспечивают посредством конструирования хелперной плазмиды AAV, используемой для подготовки векторного штамма, так что один или несколько из кодированных белков оболочки представляют собой гибриды природного белка оболочки AAV и белкового или пептидного лиганда, таким образом, что данный лиганд открыт для воздействия на поверхности вирусной частицы.
[0018] Клетки-мишени могут представлять собой клетки человека или клетки других млекопитающих (в том числе ветеринарных животных), в частности приматов, отличных от человека, и млекопитающих отряда грызунов (мышей, крыс, хомяков), зайцеобразных (кроликов), плотоядных (кошек, собак) и парнокопытных (коров, свиней, овец, коз, лошадей). В некоторых аспектах клетки-мишени включают ткань (например, печень). В некоторых аспектах клетка-мишень представляет собой стволовую клетку (например, эмбриональную стволовую клетку, индуцированную плюрипотенциальную стволовую клетку, печеночную стволовую клетку и т.д.) или эмбрион животного при любом из описанных здесь способов, и в таком случае эмбрион имплантируют таким образом, что рождается живое животное. Далее животное развивается до половой зрелости, и ему позволяют произвести потомство, в котором, по меньшей мере, некоторые из потомков содержат геномную модификацию.
[0019] В свете раскрытия в целом эти и другие аспекты очевидны для специалиста в данной области. Таким образом, раскрытие охватывает следующие варианты:
[0020] 1. Белок, содержащий генно-инженерный ДНК-связывающий домен белка «цинковый палец», где ДНК-связывающий домен содержит четыре или пять областей распознавания «цинковый палец», обозначенных от F1 до F4 или от F1 до F5 от N-конца до C-конца, и где
(i) если ДНК-связывающий домен содержит пять областей распознавания «цинковый палец», то области с F1 по F5 включают следующие последовательности аминокислот:
F1: QSGDLTR (SEQ ID NO:4)
F2: RSDVLSE (SEQ ID NO:5)
F3: DRSNRIK (SEQ ID NO:6)
F4: RSDNLSE (SEQ ID NO:7)
F5: QNATRIN (SEQ ID NO:8)
(ii) если ДНК-связывающий домен содержит четыре области распознавания «цинковый палец», то области с F1 по F4 включают следующие последовательности аминокислот:
F1: RSDSLSV (SEQ ID NO:10)
F2: TSGHLSR (SEQ ID NO:11)
F3: RSDHLSQ (SEQ ID NO:12)
F4: HASTRHC (SEQ ID NO:13)
[0021] 2. Белок по п. 1, дополнительно содержащий расщепляющий домен или полудомен.
[0022] 3. Белок по п. 2, где расщепляющий полудомен представляет собой дикий тип или генно-инженерный расщепляющий полудомен FokI.
[0023] 4. Полинуклеотид, кодирующий белок по любому из пп. 1-3.
[0024] 5. Вектор для доставки генов, содержащий полинуклеотид по п. 4.
[0025] 6. Выделенная клетка, содержащая белок по любому из пп. 1-3 или полинуклеотид по п. 4.
[0026] 7. Выделенная клетка, содержащая белок по любому из пп. 1-3 или полинуклеотид по п. 4.
[0027] 8. Способ лечения гемофилии В у субъекта, причем указанный способ включает введение (например, посредством целенаправленной интеграции) последовательности, кодирующей функциональный белок фактор IX (FIX), в геном клетки, используя, по меньшей мере, одну нуклеазу, где субъект содержит данную клетку.
[0028] 9. Способ по п. 8, где последовательность интегрируют в эндогенный ген.
[0029] 10. Способ по п. 9, где эндогенный ген выбран из группы, включающей ген FIX и ген «безопасная гавань».
[0030] 11. Способ по любому из пп. 8-10, где последовательность и/или нуклеазу доставляют в клетку, используя вектор, выбранный из группы, включающей вирусный вектор, невирусный вектор и их комбинации.
[0031] 12. Способ по любому из пп. 8-11, где клетка представляет собой клетку печени, и последовательность доставляют в клетку путем внутривенного введения (например, в печень) интактному животному.
[0032] 13. Способ по любому из пп. 8-12, где, по меньшей мере, одна нуклеаза представляет собой цинк-пальцевую нуклеазу, TALEN или хоминг-эндонуклеазу.
[0033] 14. Способ по любому из пп. 8-13, дополнительно включающий стадию проведения частичной гепатэктомии у субъекта.
[0034] 15. Способ по любому из пп. 8-14, дополнительно включающий стадию лечения субъекта, по меньшей мере, одним вспомогательным агентом.
[0035] 16. Способ 15, где вспомогательный агент выбран из группы, включающей гамма-облучение, УФ-облучение, меченные тритием нуклеотиды, цис-платину, преднизолон, четыреххлористый углерод, этопозид, гидроксимочевину, афидиколин, аденовирус и их комбинации.
[0036] 17. Способ по любому из пп. 8-16, где клетка представляет собой выделенную клетку, и способ дополнительно включает введение выделенной клетки субъекту.
[0037] 18. Способ по любому из пп. 8-17, где субъект выбран из группы, включающей эмбрион, плод, новорожденного, инфантильного, ювенильного или взрослого субъекта.
[0038] 19. Способ по любому из пп. 8-18, дополнительно включающий связывание последовательности с хоминг-агентом, который специфически связывается с поверхностным рецептором клетки.
[0039] 20. Способ 19, где хоминг-агент содержит галактозу или гибрид белка оболочки AAV и галактозу.
[0040] 21. Способ по любому из пп. 8-20, дополнительно включающий связывание полинуклеотида, кодирующего, по меньшей мере, одну нуклеазу, с хоминг-агентом, который специфически связывается с поверхностным рецептором клетки.
[0041] 22. Способ по п. 21, где хоминг-агент содержит галактозу или гибрид белка оболочки AAV и галактозу.
[0042] 23. Способ по любому из пп. 8-22, где клетка выбрана из группы, включающей клетку человека, клетку примата, отличного от человека, клетку грызуна, клетку зайцеобразного, клетку плотоядного и клетку парнокопытного.
[0043] 24. Способ по любому из пп. 8-22, где клетка-мишень представляет собой стволовую клетку.
[0044] 25. Способ по п. 24, где стволовая клетка представляет собой эмбриональную стволовую клетку, индуцированную плюрипотенциальную стволовую клетку, гемопоэтическую стволовую клетку, гепатоцит или печеночную стволовую клетку.
[0045] 26. Способ по любому из пп. 8-25, где нуклеаза включает цинк-пальцевую нуклеазу по любому из пп. 1-3, полинуклеотид по п. 4 или средство для доставки генов по п. 5.
[0046] На фигуре 1, панели A-E, показано, что N2 ZFN эффективно расщепляют интрон 1 гена, кодирующего фактор IΧ (F9), и индуцируют гомологичную рекомбинацию в клетках человека. На фигуре 1 представлено схематическое изображение цели для пары N2 ZFN в интроне 1 гена человека F9. На фигуре 1B изображена бицистронная экспрессионная плазмида FLAG-меченая ZFN. На фигуре 1С показан гель с результатами Surveyor® исследования несоответствия (Transgenomics, "Cel-I") после трансфекции N2 ZFN экспрессионной плазмиды в клетки K562. Исследование демонстрирует результат NHEJ-репарации DSB, индуцированного посредством N2 ZFN на интроне 1 гена hF9, на 3 день после трансфекции. ZFN экспрессию подтверждают посредством иммуноблоттинга α-FLAG и оценивают загрузку белка, используя антитела α-NFkB Р65. На рисунке 1D показана схема исследования целенаправленной интеграции (TI), детализирующая течение во времени ZFN-опосредованного направленного воздействия сайта рестрикции NheI на ген hF9. На фигуре 1E показан гель, изображающий результаты RFLP-исследования, после котрансфекции ZFN экспрессионной плазмиды с возрастающими количествами NheI-меченой донорной плазмиды (0-4 мкг). Данные показывают повышение уровней направленного воздействия на ген в дни 3 и 10 после трансфекции, тогда как трансфекция только NheI-меченого донора (4 мкг, ‘(-)ZFN’) не дает детектируемого направленного воздействия на ген. Черные стрелки обозначают NheI-чувствительные продукты расщепления, получающиеся в результате TI, в день 3 и день 10. TI PCR осуществляют посредством PCR с использованием 32P-меченых нуклеотидов и оценивают количественно интенсивность полос, используя устройство для чтения пластин phosphorimager. ZFN экспрессию подтверждают посредством α-FLAG иммуноблоттинга и оценивают загрузку белков с помощью антитела α-NFkB Р65.
[0047] На фигуре 2, панели A-E, показаны результаты AAV-опосредованной доставки N2 ZFN мышам, имеющим N2 «посадочную площадку» (LP), по эффективному расщеплению посадочной площадки (LP) интрона 1. На фигуре 2 изображена диаграмма, показывающая, как N2 ZFN целенаправленно воздействует на интрон 1 человеческого мини-гена F9 (LP), который симулирует опубликованную HB-вызывающую мутацию (Thompson и другие (1994) Hum. Genet. 94:299-302). На фигуре 2B показан гель из PCR-анализа, демонстрирующий поражение LP-конструкции по мышиному локусу ROSA26. На фигуре 2С показаны результаты ELISA по детектированию циркулирующего hFIX плазмы. Данные показывают, что LP-мыши не имеют циркулирующего hFIX плазмы, что определяют, применяя hFIX-специфический анализ ELISA, пока мышам не вводят вирусный вектор, экспрессирующий hFIX (1e10 вирусные геномы (v.g.) AAV-hFIX вводят в хвостовую вену). На фигуре 2D изображен бицистронный вектор экспрессии AAV8-N2 ZFN при регулировании экспрессии энхансером АроЕ и человеческим альфа-антитрипсиновым промотором. На фигуре 2E показаны результаты Cel-1 исследования, выполняемого после инъекции в хвостовую вену LP-мышей вектора экспрессии 1e11 v.g. AAV-N2, что приводит к расщеплению LP-интрона 1 в ДНК печени на 7 день после инъекции. Исследование Cel-1 выполняют с PCR-ампликоном, используя 32P-меченые нуклеотиды, и определяют интенсивность полос количественно на phosphorimager. Экспрессию ZFN подтверждают посредством α-FLAG иммуноблоттинга всех лизатов α-NFkB Р65.
[0048] На фигуре 3, панели A и B, показано, что N2 ZFN промотируют AAV-опосредованное направленное воздействие экзонов F9 дикого типа 2-8 на посадочную площадку интрон 1 in vivo. На фигуре 3A схематически показано, как можно обойти мутацию LP гена посредством TI экзонов hF9 2-8 в интрон 1. Целевые и нецелевые LP-аллели можно дифференцировать посредством PCR, используя праймеры P1, P2 и P3. На фигуре 3B изображен гель из PCR-анализа с парами праймеров P1/P2 и P1/P3, демонстрирующий успешное направленное воздействие на ген после I.P. совместной инъекции 5e10 v.g. AAV8-N2 и 2.5e11 v.g. AAV8-донор у мышей LP/HB на 2 день жизни, но не при инъекции только 5e10 v.g. AAV8-N2, или совместной инъекции 5e10 v.g. AAV8-Mock и 2.5e11 v.g. AAV8-донор. PCR проводят, используя 32P-меченые нуклеотиды, что позволяет количественно определять интенсивности полос продукта на устройстве phosphorimager для оценки частоты направленного воздействия. В целевых образцах праймеры P1 и P2 генерируют меньший продукт, что указывает на успешную амплификацию целевых экзонов F9 дикого типа 2-8, тогда как праймеры P1 и P3 генерируют больший продукт, чем нецелевой аллель.
[0049] На фигуре 4, панели A-F, показаны результаты in vivo коррекции гена печени при терапевтических уровнях циркулирующего FIX. На фигуре 4A приведен график, показывающий уровни hFIX в плазме LP-мышей после I.P.-инъекции на 2 день жизни при введении только 5e10 v.g. AAV-N2 (n=7 до и после частичной гепатэктомии (PHx)), 5e10 v.g. AAV-N2 и 2.5e11 v.g. AAV-донор (n=7 до и после PHx) или 5e10 v.g. AAV-Mock и 2.5e11 v.g. AAV-донор (n=6 до и после PHx). Сроки PHx указаны стрелкой. Интервалы погрешностей обозначают стандартную ошибку. На фигуре 4B приведен график, показывающий уровни hFIX в плазме мышей дикого типа (n=5) после инъекции в хвостовую вену 1e12 v.g. AAV-hFIX (преимущественно эписомного) с последующей PHx. Интервалы погрешностей обозначают стандартную ошибку. На фигуре 4С приведен график, показывающий уровни hFIX в плазме мышей дикого типа после I.P.-инъекции на 2 день жизни при введении только 5e10 v.g. AAV-N2 (n=8 до PHx, n=4 после PHx), 5e10 v.g. AAV-N2 и 2.5e11 v.g. AAV-донор (n=9 до PHx, n=5 после PHx) или 5e10 v.g. AAV-Mock и 2.5e11 v.g. AAV-донор (n=6 до PHx, n=5 после PHx). Интервалы погрешностей обозначают стандартную ошибку. На фигуре 4D приведен график уровней hFIX в плазме мышей LP/HB после внутрибрюшной (I.P.) инъекции на 2 день жизни при введении только 5e10 v.g. AAV-N2 (n=10 до PHx, n=1 после PHx), 5e10 v.g. AAV-N2 и 2.5e11 v.g. AAV-донор (n=9 до PHx, n=5 после PHx) или 5e10 v.g. AAV-Mock и 2.5e11 v.g. AAV-донор (n=9 до PHx, n=3 после PHx). Интервалы погрешностей обозначают стандартную ошибку. На фигуре 4E показан гель, демонстрирующий печень-специфическую экспрессию hFIX РНК, которую детектируют посредством RT-PCR на 20 неделе жизни у мышей LP/HB, принимающих на 2 день жизни I.P. инъекцию 5e10 v.g. AAV-N2 и 2.5e11 v.g. AAV-донор. На фигуре 4F приведен график, показывающий время формирования сгустка, которое определяют, проводя исследование активированного частичного тромбопластинового времени (aPTT) на мышах возраста 14 недель, принимающих на 2 день жизни I.P. инъекцию 5e10 v.g. AAV-N2 и 2.5e11 v.g. AAV-донор (n=5) или 5e10 v.g. AAV-Mock и 2.5e11 v.g. AAV-донор (n=3) (p-значение из 2-стороннего t теста Стьюдента). Для сравнения приведены aPTT мышей дикого типа (WT) и мышей с гемофилией B (HB).
[0050] На фигуре 5, панели A и B, показаны in vivo результаты коррекции гена печени при экспрессии терапевтических уровней циркулирующего FIX. На фигуре 5 приведен график, показывающий уровни hFIX в плазме взрослых мышей LP после I.V. инъекции в возрасте 6 недель и при введении только 1e11 v.g./мышь AAV-N2 ('ZFN только'), 1e11 v.g./мышь AAV-N2 и 5.5e11 v.g./мышь AAV-донор ('ZFN + донор') или 1e11 v.g./мышь AAV-Mock и 5.5e11 v.g. AAV-донор ('Mock+донор'). Представленные данные являются репрезентативными для 3 экспериментов при количестве примерно 20 мышей на группу. В этих экспериментах уровни hF.IX дикого типа приблизительно составляют 1000 нг/мл. На фигуре 5B представлен график, показывающий уровни hFD в плазме взрослых мышей LP после I.V. инъекции в возрасте 6 недель и введении только 1e11 v.g./мышь AAV-N2 ('только ZFN'), 1e11 v.g./мышь AAV-N2 и 5.5e11 v.g./мышь AAV-донор ('ZFN+донор') или 1e11 v.g./мышь AAV-Mock и 5.5e11 v.g. AAV-донор ('Mock+донор'). Через два дня после инъекции животным групп, указанных на фигуре 5B, производят частичную гепатэктомию. Приведенные данные являются репрезентативными для 3 экспериментов при количестве примерно 20 мышей на группу. В этих экспериментах уровни hF.IX дикого типа составляют примерно 1000 нг/мл. Данные демонстрируют, что экспрессия hF.IX является стабильной при введении взрослым мышам с (или без) последующей частичной гепатэктомией, и что возможно выполнять редактирование генома у взрослых животных.
ПОДРОБНОЕ ОПИСАНИЕ
[0051] Здесь раскрыты композиции и способы лечения субъектов с гемофилией B. В частности для введения последовательности, кодирующей FIX, в геном одной или нескольких клеток субъекта (in vivo или ex vivo) применяют нуклеаза-опосредованную целенаправленную интеграцию, чтобы клетки вырабатывали FIX in vivo. В некоторых вариантах способы дополнительно включают индуцирование пролиферации (введение клеточного цикла) клеток субъекта, в частности печеночных клеток, например, посредством частичной гепатэктомии и/или введения одного или нескольких соединений, которые индуцируют протекание клеточного цикла клеток печени. Субъекты включают, но не ограничены этим, людей, приматов, отличных от человека, ветеринарных животных, таких как кошки, собаки, кролики, крысы, мыши, морские свинки, коровы, свиньи, лошади, козы и подобные.
[0052] Описанные здесь способы обеспечивают в результате лечение гемофилии B. В отличие от описанных ранее способов нуклеаза-опосредованной коррекции гена на in vivo моделях с применением мегануклеаз (см. работу Gouble и другие (2006) J Gene Med., 8 мая (5):616-22) после нуклеаза-опосредованной интеграции гена FIX на животных моделях наблюдается низкая (или вообще не наблюдается) токсичность. Кроме того, способы и композиции по изобретению являются функциональными у новорожденных и взрослых животных, приводя к функциональной активности введенного трансгена фактора IΧ.
Общее
[0053] В практике применения способов, а также при получении и применении раскрытых здесь композиций используют, если не указано иное, обычные методы молекулярной биологии, биохимии, структуру и анализ хроматина, компьютерную химию, культивирование клеток, рекомбинантную ДНК и смежные области в пределах возможностей специалистов в данной области. Эти способы полностью описаны в литературе. См., например, работы Sambrook и другие MOLECULAR CLONING: A LABORATORY MANUAL, второе издание, Cold Spring Harbor Laboratory Press, 1989 и третье издание, 2001; Ausubel и другие, CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, New York, 1987 и периодические обновления; серии METHODS IN ENZYMOLOGY, Academic Press, San Diego; Wolffe, ХРОМАТИН STRUCTURE AND FUNCTION, третье издание, Academic Press, San Diego, 1998; METHODS IN ENZYMOLOGY, Vol. 304, "Chromatin" (под редакцией P.M. Wassarman и A. P. Wolffe), Academic Press, San Diego, 1999; и METHODS IN MOLECULAR BIOLOGY, Vol. 119, "Chromatin Protocols" (под редакцией P.B. Becker) Humana Press, Totowa, 1999.
Определения
[0054] Термины «нуклеиновая кислота», «полинуклеотид» и «олигонуклеотид» используются взаимозаменяемо и относятся к дезоксирибонуклеотиду или рибонуклеотидному полимеру с линейной или кольцевой конформацией и в любой форме одно- или двунитевой. Для целей настоящего раскрытия эти термины не следует толковать как ограничительные в отношении длины полимера. Данные термины могут охватывать известные аналоги природных нуклеотидов, а также нуклеотиды, которые модифицированы по основным, сахарным и/или фосфатным фрагментам (например, фосфоротиоатным основным цепям). Вообще, аналог конкретного нуклеотида имеет такую же специфичность спаривания оснований; то есть аналог А будет спарен с Т.
[0055] Термины «полипептид», «пептид» и «белок» используют взаимозаменяемо для обозначения полимера из аминокислотных остатков. Этот термин также относится к аминокислотным полимерам, в которых одна или более аминокислот представляют собой химические аналоги или модифицированные производные соответствующих аминокислот, встречающихся в природе.
[0056] Термин «связывание» относится к последовательность-специфическому нековалентному взаимодействию между макромолекулами (например, между белком и нуклеиновой кислотой). Не все компоненты связывающего взаимодействия должны быть последовательность-специфическими (например, контакты с фосфатными остатками в основной цепи ДНК), пока взаимодействие в целом является последовательность-специфическим. Такие взаимодействия обычно характеризуются константой диссоциации (Kd) 10-6 M-1 или ниже. Термин «аффинность» относится к прочности связывания: повышенная аффинность связывания коррелирует с меньшей Kd.
[0057] Выражение «связывающий белок» обозначает белок, способный нековалентно присоединяться к другой молекуле. Связывающий белок может присоединяться, например, к молекуле ДНК (ДНК-связывающий белок), молекуле РНК (РНК-связывающий белок) и/или молекуле белка (белок-связывающий белок). В случае белок-связывающего белка он может связываться сам с собой (в виде гомодимеров, гомотримеров и т.д.), и/или он может присоединяться к одной или несколькими молекулам другого белка или белков. Связывающий белок может иметь активности связывания более одного типа. Например, белки «цинковый палец» обладают ДНК-связывающей, РНК-связывающей и белок-связывающей активностью.
[0058] Выражение «ДНК-связывающий белок типа цинкового пальца» (или связывающий домен) обозначает белок или домен внутри большего белка, который связывает ДНК последовательность-специфическим образом посредством одного или нескольких «цинковых пальцев», которые представляют собой области аминокислотной последовательности внутри связывающего домена, структура которого стабилизирована координацией иона цинка. Выражение «ДНК-связывающий белок типа цинкового пальца» часто сокращенно обозначают белок «цинковый палец» или ZFP.
[0059] Выражение «ДНК-связывающий домен TALE» или «TALE» обозначает полипептид, содержащий один или более повторяющихся доменов/звеньев TALE. Повторяющиеся домены вовлечены в связывание TALE с родственной последовательностью целевой ДНК. Отдельная «повторяющаяся единица» (также обозначаемая «повтор») обычно имеет длину 33-35 аминокислот и демонстрирует, по меньшей мере, некоторую гомологию последовательности с другими повторяющимися последовательностями TALE в природном белке TALE.
[0060] Можно «сконструировать» связывающие домены «цинковый палец» и TALE для привязки к предварительно определенной нуклеотидной последовательности, например, посредством генной инженерии (изменяя одну или несколько аминокислот) спиральной области распознавания встречающегося в природе белка «цинковый палец» или TALE. Таким образом, генно-инженерные ДНК-связывающие белки («цинковые пальцы» или TALE) представляют собой белки, которые не являются природными. Неограничительными примерами способов конструирования ДНК-связывающих белков являются конструкция и отбор. Генно-инженерный ДНК-связывающий белок представляет собой белок, не встречающийся в природе, конструкция/состав которого являются результатом, главным образом, критериев рациональности. Рациональные критерии для конструирования включают применение правил замены и компьютерных алгоритмов для обработки информации в базе данных, хранящей информацию о существующих конструкциях ZFP и/или TALE, и данных по связыванию. См., например, патенты США 6140081; 6453242 и 6534261; а также WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496 и заявку США № 13/068735.
[0061] «Отобранный» белок «цинковый палец» или TALE представляет собой белок, не обнаруженный в природе, получение которого является результатом в основном эмпирического процесса, такого как фаговый дисплей, ограничитель взаимодействия или отбор гибридов. См., например, патенты США 5789538; США 5925523; США 6007988; США 6013453; США 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970 WO 01/88197 и WO 02/099084.
[0062] Термин «рекомбинация» относится к процессу обмена генетической информацией между двумя полинуклеотидами, включая, но не ограничиваясь этим, захват донора при негомологичном соединении концов (NHEJ) и гомологичную рекомбинацию. Для целей этого раскрытия выражение «гомологичная рекомбинация (HR)» относится к специальной форме такого обмена, который имеет место, например, во время репарации двунитевых разрывов в клетках посредством механизмов направляемой гомологией репарации. Этот процесс требует гомологии нуклеотидной последовательности, использует «донорную» молекулу для матричной репарации «целевой» молекулы (то есть, молекулы, которая претерпевает двунитевой разрыв) и по-разному известен как «некроссоверная конверсия гена» или «конверсия гена по короткому пути», так как это приводит к передаче генетической информации от донора к целевому объекту. Не желая связываться какой-либо конкретной теорией, такая передача может включать неудачную коррекцию гетеродуплексной ДНК, которая образуется между разрушенным целевым объектом и донором, и/или «синтез-зависимую ренатурацию цепей», где донор используют для ресинтеза генетической информации, которая станет частью целевого объекта, и/или родственные процессы. Такая специализированная HR часто дает в результате изменение последовательности целевой молекулы, так что часть или вся последовательность донорного полинуклеотида включается в целевой полинуклеотид.
[0063] В способах раскрытия, одна или более целевых нуклеаз, которые здесь описаны, создают двунитевой разрыв в рассматриваемой последовательности (например, клеточном хроматине) на предварительно определенном сайте, и «донорный» полинуклеотид, имеющий гомологию с нуклеотидной последовательностью в области разрыва, можно ввести в клетку. Как показано, наличие двунитевого разрыва облегчает интеграцию донорной последовательности. Донорную последовательность можно физически интегрировать, или, по-другому, используют донорный полинуклеотид как матрицу для репарации разрыва посредством гомологичной рекомбинации, получая в результате введение всей или части нуклеотидной последовательности как донора в клеточный хроматин. Таким образом, первую последовательность в клеточном хроматине можно изменить и в некоторых вариантах можно преобразовать в последовательность, присутствующую в донорном полинуклеотиде. Следовательно, выражения «заменить» или «замена» можно понимать как обозначающие замену одной нуклеотидной последовательности на другую (то есть, замену последовательности в информационном смысле) и не обязательно требующие другой физической или химической замены одного полинуклеотида на другой.
[0064] В любом из описанных здесь способов можно использовать дополнительные пары белков «цинковый палец» или TALEN для дополнительного двунитевого расщепления дополнительных целевых сайтов внутри клетки.
[0065] В некоторых вариантах способов целенаправленной рекомбинации и/или замены, и/или изменения последовательности в представляющей интерес области в клеточном хроматине изменяют хромосомную последовательность посредством гомологичной рекомбинации с помощью экзогенной «донорной» нуклеотидной последовательности. Такую гомологичную рекомбинацию стимулируют присутствием двунитевого разрыва в клеточном хроматине, если присутствуют последовательности, гомологичные области разрыва.
[0066] В любом из описанных здесь способов первая нуклеотидная последовательность («донорная последовательность») может содержать последовательности, которые гомологичны, но не идентичны геномным последовательностям в области, представляющей интерес, стимулируя тем самым гомологичную рекомбинацию для введения неидентичной последовательности в область, представляющую интерес. Таким образом, в некоторых вариантах части донорной последовательности, которые гомологичны последовательностям в области, представляющей интерес, демонстрируют примерно от 80 до 99% (или любое целочисленное значение между указанными) идентичности последовательности с геномной последовательностью, которая замещается. В других вариантах гомология между донором и геномной последовательностью выше 99%, например, если только 1 нуклеотид отличается между донором и геномными последовательностями, включающими более 100 соседних пар оснований. В некоторых случаях негомологичная часть донорной последовательности может содержать последовательности, не присутствующие в области, представляющей интерес, так что в область, представляющую интерес, вводят новые последовательности. В этих случаях негомологичная последовательность обычно фланкирована последовательностями из 50-1000 пар оснований (или любым целочисленным значением между указанными) или любым количеством пар оснований более 1000, которые гомологичны или идентичны последовательностям в области, представляющей интерес. В других вариантах донорная последовательность негомологична первой последовательности и вводится в геном посредством механизмов негомологичной рекомбинации.
[0067] Любой из описанных здесь способов можно использовать для частичной или полной инактивации одной или нескольких целевых последовательностей в клетке путем целенаправленной интеграции донорной последовательности, которая нарушает экспрессию представляющего интерес гена(ов). Получают также клеточные линии с частично или полностью инактивированными генами.
[0068] Кроме того, способы направленной интеграции, которые здесь описаны, также можно применять для интеграции одной или нескольких экзогенных последовательностей. Экзогенная последовательность нуклеиновой кислоты может содержать, например, один или более генов, или молекул кДНК, или кодирующую или некодирующую последовательность любого типа, а также один или более регулирующих элементов (например, промоторов). Кроме того, экзогенная последовательность нуклеиновой кислоты может продуцировать одну или более молекул РНК (например, малые РНК, образующие шпильки (мшРНК), ингибиторные РНК (РНКи), микроРНК (мкРНК) и другие.)
[0069] Термин «расщепление» относится к разрыву ковалентной основной цепи молекулы ДНК. Расщепление можно инициировать разнообразными способами, в том числе, но не ограничиваясь этим, ферментативным или химическим гидролизом Фосфодиэфирной связи. Возможно однонитевое расщепление и двунитевое расщепление, и двунитевое расщепление может происходить в результате двух отдельных случаев однонитевого расщепления. Расщепление ДНК может приводить к получению тупых концов или ступенчатых концов. В некоторых вариантах используют слитые полипептиды для целевого двунитевого расщепления ДНК.
[0070] «Расщепляющий полудомен» представляет собой полипептидную последовательность, которая в сочетании со вторым полипептидом (одинаковым или отличным) образует комплекс, обладающий расщепляющей активностью (предпочтительно активностью двунитевого расщепления). Выражения «первый и второй расщепляющие полудомены» «+ и - расщепляющие полудомены» и «правый и левый расщепляющие полудомены» используют взаимозаменяемо для обозначения пар расщепляющих полудоменов, которые димеризуются.
[0071] «Генно-инженерный расщепляющий полудомен» представляет собой расщепляющий полудомен, который модифицирован так, чтобы формировать облигатные гетеродимеры с другим расщепляющим полудоменом (например, другим генно-инженерным расщепляющим полудоменом). См. также публикации патентов США №№ 2005/0064474, 20070218528 и 2008/0131962, включенные здесь в виде ссылок во всей своей полноте.
[0072] Термин «последовательность» относится к нуклеотидной последовательности любой длины, которая может представлять собой ДНК или РНК; может быть линейной, кольцевой или разветвленной и может быть однонитевой или двунитевой. Выражение «донорная последовательность» относится к нуклеотидной последовательности, которую вводят в геном. Донорная последовательность может иметь любую длину, например, от 2 до 10000 нуклеотидов (или любое целочисленное значение между указанными или выше), предпочтительно длину примерно от 100 до 1000 нуклеотидов (или любое целочисленное значение между указанными), более предпочтительно длину примерно от 200 до 500 нуклеотидов.
[0073] «Хроматин» представляет собой нуклеопротеиновую структуру, содержащую клеточный геном. Клеточный хроматин содержит нуклеиновую кислоту, главным образом ДНК, и белок, в том числе гистоны и негистонные хромосомные белки. Большая часть эукариотического клеточного хроматина существует в виде нуклеосом, в которых ядро нуклеосомы содержит примерно 150 пар оснований ДНК, связанных с октамером, содержащим два гистона из следующих: H2A, H2B, H3 и H4; и линкерная ДНК (переменной длины в зависимости от организма) тянется между ядрами нуклеосом. Молекула гистона H1 обычно ассоциирована с линкером ДНК. Для целей настоящего раскрытия имеется в виду, что термин «хроматин» охватывает все типы клеточного нуклеопротеина, прокариотического и эукариотического. Клеточный хроматин включает хромосомный и эписомный хроматин.
[0074] Термин «хромосома» обозначает хроматиновый комплекс, содержащий весь геном клетки или его часть. Геном клетки часто характеризуют его кариотипом, который является коллекцией всех хромосом, которые составляют геном клетки. Геном клетки может содержать одну или более хромосом.
[0075] «Эписома» представляет собой репликативную нуклеиновую кислоту, нуклеопротеиновый комплекс или другую структуру, содержащую нуклеиновую кислоту, которая не является частью хромосомного кариотипа клетки. Примеры эписом включают плазмиды и некоторые вирусные геномы.
[0076] «Целевой сайт» или «целевая последовательность» представляет собой последовательность нуклеиновой кислоты, которая определяет часть нуклеиновой кислоты, с которой связывается присоединяющаяся молекула, при условии наличия условий, достаточных для связывания.
[0077] «Экзогенная» молекула представляет собой молекулу, которая обычно не присутствует в клетке, но может быть введена в клетку одним или несколькими генетическими, биохимическими или другими способами. «Нормальное присутствие в клетке» определяют в отношении конкретной стадии развития и условий среды клетки. Таким образом, например, молекула, которая присутствует только во время эмбрионального развития мышцы, является экзогенной молекулой в отношении взрослой мышечной клетки. Аналогично, молекула, индуцированная тепловым шоком, является экзогенной молекулой в отношении клетки, не подверженной воздействию тепловым шоком. Экзогенная молекула может включать, например, версию функционирования неправильно действующей эндогенной молекулы или неправильно функционирующую версию нормально действующей эндогенной молекулы.
[0078] Экзогенная молекула может представлять собой, среди прочего, малую молекулу, такую как генерируется способом комбинаторной химии, или макромолекулу, такую как белок, нуклеиновая кислота, углевод, липид, гликопротеин, липопротеид, полисахарид, любое модифицированное производное указанных выше молекул или любой комплекс, содержащий одну или более из указанных выше молекул. Нуклеиновые кислоты включают ДНК и РНК, могут быть одно- или двунитевыми; могут быть линейными, разветвленными или кольцевыми; и могут быть любой длины. Нуклеиновые кислоты включают соединения, способные образовывать дуплексы, а также триплекс-образующие нуклеиновые кислоты. См., например, патенты США №№ 5176996 и 5422251. Белки включают, но не ограничены этим, ДНК-связывающие белки, факторы транскрипции, хроматин-ремоделирующие факторы, метилированные ДНК-связывающие белки, полимеразы, метилазы, деметилазы, ацетилазы, деацетилазы, киназы, фосфатазы, интегразы, рекомбиназы, лигазы, топоизомеразы, гиразы и геликазы.
[0079] Экзогенная молекула может представлять собой молекулу того же типа, что и эндогенная молекула, например, молекулу экзогенного белка или нуклеиновой кислоты. Например, экзогенная нуклеиновая кислота может включать инфицирующий вирусный геном, плазмиду или эписому, вводимую в клетку или хромосому, которая обычно не присутствует в клетке. Способы введения экзогенных молекул в клетки известны специалистам в данной области и включают, но не ограничены этим, липид-опосредованный перенос (то есть, липосомы, включающие нейтральные и катионные липиды), электропорацию, прямую инъекцию, слияние клеток, бомбардировку частиц, соосаждение с фосфатом кальция, DEAE-декстран-опосредованный перенос и вирусный вектор-опосредованный перенос. Экзогенная молекула может представлять собой молекулу того же типа, что и эндогенная молекула, но происходить из вида, отличного от происхождения клетки. Например, последовательность нуклеиновой кислоты человека можно вводить в клеточную линию, изначально полученную от мыши или хомячка.
[0080] В противоположность этому «эндогенная» молекула является молекулой, которая обычно присутствует в конкретной клетке на определенной стадии развития в конкретных условиях среды. Например, эндогенная нуклеиновая кислота может содержать хромосому, геном митохондрии, хлоропласт или другую органеллу или встречающуюся в природе эписомную нуклеиновую кислоту. Дополнительные эндогенные молекулы могут включать белки, например, факторы транскрипции и ферменты.
[0081] «Слитая» молекула представляет собой молекулу, в которой связаны две или более субъединичных молекул, предпочтительно ковалентно. Субъединичные молекулы могут быть молекулами одного химического типа или могут быть молекулами различных химических типов. Примеры слитой молекулы первого типа включают, но не ограничены этим, слитые белки (например, слияние ДНК-связывающего домена ZFP или TALE и одного или нескольких активирующих доменов) и слитые нуклеиновые кислоты (например, нуклеиновую кислоту, кодирующую описанный выше слитой белок). Примеры слитой молекулы второго типа включают, но не ограничены этим, слияние триплекс-образующей нуклеиновой кислоты и полипептида и слияние связующего малой бороздки и нуклеиновой кислоты.
[0082] Экспрессия слитого белка в клетке может происходить в результате доставки слитого белка в клетку или доставки полинуклеотида, кодирующего слитой белок, в клетку, где полинуклеотид транскрибируется и транскрипт транслируется, генерируя слитой белок. В экспрессию белка в клетке также могут быть вовлечены транс-сплайсинг, расщепление полипептида и лигирование полипептида. Способы доставки полинуклеотидов и полипептидов в клетки представлены в другом месте этого раскрытия.
[0083] Термин «ген» для целей настоящего раскрытия включает область ДНК, кодирующую генный продукт (см. ниже), а также все области ДНК, которые регулируют производство генного продукта, являются ли или нет такие регуляторные последовательности соседними к кодирующим и/или транскрибируемым последовательностям. Соответственно, ген включает, но не обязательно ограничен этим, промоторные последовательности, терминаторы, трансляционные регуляторные последовательности, такие как сайты связывания рибосом и внутренние сайты входа рибосом, энхансеры, глушители, изоляторы, граничные элементы, точки инициации репликации, сайты присоединения матрикса и области контроля локуса.
[0084] Выражение «генная экспрессия» относится к конверсии информации, содержащейся в гене, в генный продукт. Генный продукт может представлять собой прямой транскрипционный продукт гена (например, мРНК, тРНК, рРНК, антисмысловую РНК, рибозим, структурную РНК или любой другой тип РНК) или белок, продуцируемый при трансляции мРНК. Генные продукты также включают РНК, которые модифицированы такими процессами как кеппинг, полиаденилирование, метилирование и редактирование, и белки, модифицированные, например, метилированием, ацетилированием, фосфорилированием, убиквитинированием, ADP-рибозилированием, миристилированием и гликозилированием.
[0085] Термин «модуляция» генной экспрессии касается изменения активности гена. Модуляция экспрессии может включать, но не ограничена этим, активацию гена и репрессию гена. Для модуляции экспрессии можно использовать редактирование генома (например, расщепление, изменение, инактивацию, случайную мутацию). Инактивация гена относится к любому снижению генной экспрессии по сравнению с клеткой, которая не включает ZFP, который здесь описан. Таким образом, инактивация гена может быть частичной или полной.
[0086] «Область, представляющая интерес» - это любая область клеточного хроматина, например, ген или некодирующая последовательность внутри или рядом с геном, в которой желательно присоединить экзогенную молекулу. Целью привязки может быть расщепление целевой ДНК и/или целенаправленная рекомбинация. Область, представляющая интерес, может присутствовать, например, в хромосоме, эписоме, геноме органелл (например, митохондриальном, хлоропластном) или инфицирующем вирусном геноме. Область, представляющая интерес, может находиться в кодирующей области гена, в пределах транскрибируемых некодирующих областей, таких как, например, лидерные последовательности, концевые последовательности или интроны, или внутри нетранскрибируемых областей, выше или ниже кодирующей области. Область, представляющая интерес, может быть небольшой, как отдельная пара нуклеотидов или иметь длину до 2000 пар нуклеотидов или любое целочисленное значение пар нуклеотидов.
[0087] «Эукариотические» клетки включают, но не ограничены этим, грибковые клетки (например, дрожжи), растительные клетки, клетки животных, клетки млекопитающих и клетки человека (например, Т-клетки).
[0088] Термины «оперативная связь» и «оперативно связанный» (или «функционально связанный») используют взаимозаменяемо в отношении сопоставления двух или более компонентов (например, последовательности элементов), в которых компоненты расположены таким образом, что оба компонента функционируют нормально и допускают возможность, что, по меньшей мере, один из компонентов может опосредовать функцию, которая проявляется, по меньшей мере, на одном из других компонентов. В качестве иллюстрации транскрипционная регуляторная последовательность, такая как промотор, оперативно связана с кодирующей последовательностью, если транскрипционная регуляторная последовательность регулирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или нескольких транскрипционных регуляторных факторов. Транскрипционная регуляторная последовательность обычно оперативно связана по цис-положению относительно кодирующей последовательности, но не обязательно должна непосредственно соседствовать с ней. Например, энхансер представляет собой транскрипционную регуляторную последовательность, которая оперативно связана с кодирующей последовательностью, несмотря на то, что они не являются смежными.
[0089] Что касается слитых полипептидов, выражение «оперативно связанный» может касаться того факта, что каждый из компонентов выполняет ту же функцию в связи с другим компонентом, какую бы он выполнял, если бы не был так связан. Например, в отношении слитого полипептида, в котором ДНК-связывающий домен ZFP является слитым с активирующим доменом, ДНК-связывающий домен ZFP и активирующий домен находятся в оперативной связи, если в слитом полипептиде часть ДНК-связывающего домена ZFP способна связать его целевой сайт и/или его сайт связывания, тогда как активирующий домен способен повышающе регулировать генную экспрессию. В слитом полипептиде, где ДНК-связывающий домен ZFP слит с расщепляющим доменом, ДНК-связывающий домен ZFP и расщепляющий домен находятся в оперативной связи, если в слитом полипептиде часть ДНК-связывающего домена ZFP способна связывать его целевой сайт и/или его сайт связывания, тогда как расщепляющий домен способен расщеплять ДНК около целевого сайта.
[0090] «Функциональный фрагмент» белка, полипептида или нуклеиновой кислоты представляет собой белок, полипептид или нуклеиновую кислоту, последовательность которой неидентична первичному белку, полипептиду или нуклеиновой кислоте, но сохраняет ту же функцию, как первичный белок, полипептид или нуклеиновая кислота. Функциональный фрагмент может иметь больше, меньше или равное количество остатков, что и соответствующая нативная молекула, и/или может содержать одно или более аминокислотных или нуклеотидных замещений. Способы определения функции нуклеиновой кислоты (например, кодирующей функции, способности гибридизации с другой нуклеиновой кислотой) хорошо известны в данной области. Аналогично, хорошо известны способы определения функции белка. Например, функцию полипептида по ДНК-связыванию можно определить, посредством исследований связывания на фильтрах, сдвига электрофоретической подвижности или иммунопреципитации. Расщепление ДНК можно исследовать методом гель-электрофореза. См. указанную выше работу Ausubel и другие. Способность белка взаимодействовать с другим белком можно определить, например, посредством исследований совместной иммунопреципитации, двугибридной системы или комплементационного анализа, генетического и биохимического. См., например, Fields и другие (1989) Nature 340: 245-246; патент США № 5585245 и РСТ WO 98/44350.
[0091] «Вектор» способен переносить генные последовательности в клетки-мишени. Обычно выражения «векторная конструкция», «вектор экспрессии» и «вектор генного переноса» обозначают любую конструкцию нуклеиновой кислоты, способную направлять экспрессию представляющего интерес гена, которая может переносить генные последовательности в клетки-мишени. Таким образом, этот термин включает носители для клонирования и экспрессии, а также векторы интеграции.
[0092] Выражение «репортерный ген» или «репортерная последовательность» относится к любой последовательности, продуцирующей белковый продукт, который легко определить, предпочтительно, хотя не обязательно, в рутинном исследовании. Подходящие репортерные гены включают, но не ограничены этим, последовательности, кодирующие белки, которые опосредуют резистентность к антибиотику (например, резистентность к ампициллину, резистентность к неомицину, резистентность к G418, резистентность к пуромицину), последовательности, кодирующие окрашенные, флуоресцентные или люминисцентные белки (например, зеленый флуоресцентный белок, усиленный зеленый флуоресцентный белок, красный флуоресцентный белок, люциферазу) и белки, которые опосредуют усиленный рост клеток и/или амплификацию генов (например, дигидрофолатредуктазу). Эпитопные теги включают, например, одну или более копий FLAG, His, myc, Tap, HA или любой детектируемой аминокислотной последовательности. «Экспрессионные теги» включают последовательности, кодирующие репортеры, которые могут быть оперативно связаны с желательной генной последовательностью, для того, чтобы контролировать экспрессию гена, представляющего интерес.
[0093] Локус «безопасная гавань» представляет собой место в геноме, куда можно ввести ген без каких-либо вредных влияний на клетку-хозяина. Наиболее выгодным является локус «безопасная гавань», где экспрессия введенной генной последовательности не нарушается какой-либо экспрессией с пропуском от соседних генов. Неограничительными примерами локусов «безопасная гавань» в клетках млекопитающих являются ген AAVS1 (см. публикацию США № 20080299580), ген CCR5 (см. публикацию США № 20080159996) и/или локус Rosa (см. WO 2010/065123).
Нуклеазы
[0094] Здесь описаны композиции, в частности нуклеазы, которые пригодны для интеграции последовательности, кодирующей функциональный белок FIX в геноме клетки от субъекта с гемофилией B. В некоторых вариантах нуклеаза является природной. В других вариантах нуклеаза является неприродной, то есть генно-инженерной в ДНК-связывающем домене и/или расщепляющем домене. Например, ДНК-связывающий домен встречающейся в природе нуклеазы можно изменить для привязки к выбранному целевому сайту (например, мегануклеаза, которая сконструирована для привязки к сайту, отличному от распознаваемого сайта связывания). В других вариантах нуклеаза включает гетерологичные ДНК-связывающие и расщепляющие домены (например, цинк-пальцевые нуклеазы; TAL-эффекторный домен ДНК-связывающие белки; ДНК-связывающие домены мегануклеазы с гетерологичными расщепляющими доменами).
А. ДНК-связывающие домены
[0095] В некоторых вариантах нуклеаза представляет собой мегануклеазу (хоминг-эндонуклеазу). Встречающиеся в природе мегануклеазы распознают сайты расщепления из 15-40 пар оснований и обычно сгруппированы в четыре семейства: семейство LAGLIDADG, семейство GIY-YIG, семейство His-Cyst box и семейство HNH. Типичные хоминг-эндонуклеазы включают I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. Известны их последовательности распознавания. См. также патент США № 5420032; патент США № 6833252; Belfort и другие (1997) Nucleic Acids Res. 25:3379-3388; Dujon и другие (1989) Gene 82:115-118; Perler и другие (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble и другие (1996) J. Mol. Biol. 263:163-180; Argast и другие (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs.
[0096] В некоторых вариантах нуклеаза включает генно-инженерную (не встречающуюся в природе) хоминг-эндонуклеазу (мегануклеазу). Известны последовательности распознавания хоминг-эндонуклеаз и мегануклеаз, такие как I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII. См. также патент США № 5420032; патент США № 6833252; Belfort и другие (1997) Nucleic Acids Res. 25:3379-3388; Dujon и другие (1989) Gene 82:115-118; Perler и другие (1994) Nucleic Acids Res. 22, 1125-1127; Jasin (1996) Trends Genet. 12:224-228; Gimble и другие (1996) J. Mol. Biol. 263:163-180; Argast и другие (1998) J. Mol. Biol. 280:345-353 и каталог New England Biolabs. Кроме того, для привязки неприродных целевых сайтов можно сконструировать ДНК-связывающую специфичность хоминг-эндонуклеаз и мегануклеаз. См., например, работы Chevalier и другие (2002) Molec. Cell 10:895-905; Epinat и другие (2003) Nucleic Acids Res. 31:2952-2962; Ashworth и другие (2006) Nature 441:656-659; Paques и другие Current Gene Therapy 7:49-66; публикацию патента США № 20070117128. ДНК-связывающие домены хоминг-эндонуклеаз и мегануклеаз могут быть изменены в контексте нуклеазы в целом (то есть так, чтобы нуклеаза включала родственный расщепляющий домен) или могут быть слиты с гетерологичным расщепляющим доменом.
[0097] В других вариантах ДНК-связывающий домен включает встречающийся в природе или генно-инженерный (не встречающийся в природе) TAL эффекторный ДНК-связывающий домен. См., например, патентную заявку США № 13/068735, включенную здесь в виде ссылки во всей своей полноте. Известно, что фитопатогенные бактерии рода Xanthomonas являются причиной многих заболеваний у важных сельскохозяйственных культур. Патогенность Xanthomonas зависит от системы секреции консервативного типа III (T3S), которая вводит в растительную клетку более 25 различных эффекторных белков. Среди этих вводимых белков есть эффекторы, подобные активаторам транскрипции (TAL), которые симулируют растительные транскрипционные активаторы и манипулируют транскриптоном растения (см. работу Kay и другие (2007) Science 318:648-651). Эти белки содержат ДНК-связывающий домен и транскрипционный активирующий домен. Одним из наиболее хорошо охарактеризованных TAL-эффекторов является AvrBs3 из Xanthomonas campestgris pv. Vesicatoria (см. Bonas и другие (1989) Mol Gen Genet 218: 127-136 и WO2010079430). TAL-эффекторы содержат централизованный домен тандемных повторов, причем каждый повтор содержит примерно 34 аминокислоты, которые являются ключевыми для ДНК-связывающей специфичности этих белков. Кроме того, они содержат последовательность ядерной локализации и кислотный транскрипционный активирующий домен (см. обзор в работе Schornack S, 163(3) и другие (2006) J Plant Physiol 163(3): 256-272). Кроме того, в фитопатогенных бактериях Ralstonia solanacearum были обнаружены два гена, обозначенные brg11 и hpx17, которые являются гомологами AvrBs3 семейства Xanthomonas в R.solanacearum, биовар 1 штамм GMI1000 и биовар 4 штамм RS1000 (см. работу Heuer и другие (2007) Appl and Envir Micro 73(13): 4379-4384). Эти гены на 98,9% идентичны друг другу по нуклеотидной последовательности, но отличаются делецией 1575 пар оснований в повторяющемся домене hpx17. Однако оба генных продукта имеют менее 40% идентичности последовательности с AvrBs3 семейством белков Xanthomonas. См., например, патентную заявку США № 13/068735, включенную здесь в виде ссылки во всей своей полноте.
[0098] Специфика этих TAL-эффекторов зависит от последовательностей, обнаруженных в повторах этого тандема. Повторяющаяся последовательность включает примерно 102 пары оснований, и повторы, как правило, гомологичны друг другу на 91-100% (Bonas и другие, там же). Полиморфизм повторов обычно находится в положениях 12 и 13 и, по-видимому, представляет однозначное соответствие идентичности гипервариабельных диостатков в положениях 12 и 13 при идентичности соседних нуклеотидов в целевых последовательностях TAL-эффекторов (см. работы Moscou и Bogdanove, Science 326:1501 и Boch и другие (2009) Science 326:1509-1512). Экспериментально определен природный код распознавания ДНК этих TAL-эффекторов, так что HD последовательность в положениях 12 и 13 ведет к связи с цитозином (C), NG связывается с T, NI с А, C, G или T, NN связывается с A или G, и ING связывается с Т. Эти ДНК-связывающие повторы собирают в белки с новыми комбинациями и количествами повторов, чтобы получить искусственные факторы транскрипции, способные взаимодействовать с новыми последовательностями и активировать экспрессию неэндогенного репортерного гена в клетках растений (Boch и другие, там же). Генно-инженерные белки TAL соединяют с полудоменом расщепления FokI, получая в результате слияние TAL-эффекторный домен нуклеаза (TALEN), демонстрирующее активность в репортерном исследовании дрожжей (на основе плазмиды). См., например, патентную заявку США № 13/068735; Christian и другие ((2010)<Genetics эл. публикацию 10.1534/genetics.110.120717).
[0099] В некоторых вариантах ДНК-связывающий домен включает белок «цинковый палец». Предпочтительно, если белок «цинковый палец» не является природным, в этом случае его конструируют для привязки к выбранному целевому сайту. См., например, работы Beerli и другие (2002) Nature Biotechnol. 20:135-141; Pabo и другие (2001) Ann. Rev. Biochem. 70:313-340; Isalan и другие (2001) Nature Biotechnol. 19:656-660; Segal и другие (2001) Curr. Opin. Biotechnol. 12:632-637; Choo и другие (2000) Curr. Opin. Struct. Biol. 10:411-416; патенты США №№ 6453242; 6534261; 6599692; 6503717; 6689558; 7030215; 6794136; 7067317; 7262054; 7070934; 7361635; 7253273; и публикации патентов США №№ 2005/0064474; 2007/0218528; 2005/0267061, все включены здесь в виде ссылок во всей своей полноте.
[0100] Генно-инженерный связывающий домен «цинковый палец» может иметь новую специфичность связывания по сравнению с встречающимся в природе белком «цинковый палец». Методы генной инженерии включают, но не ограничены этим, рациональную конструкцию и различные типы отбора. Рациональная конструкция включает, например, использование баз данных, содержащих триплетные (или квадруплетные) последовательности нуклеотидов и индивидуальные последовательности аминокислот «цинковый палец», в которых каждая триплетная или квадруплетная последовательность нуклеотидов ассоциирована с одной или несколькими последовательностями аминокислот «цинковый палец», которые связывают конкретную триплетную или квадруплетную последовательность. См., например, находящиеся в совместной собственности патенты США 6453242 и 6534261, включенные здесь в виде ссылок во всей своей полноте.
[0101] Типичные способы отбора, в том числе фаговый дисплей и двугибридные системы, раскрыты в патентах США 5789538; 5925523; 6007988; 6013453; 6410248; 6140466; 6200759 и 6242568; а также WO 98/37186; WO 98/53057; WO 00/27878; WO 01/88197 и GB 2338237. Кроме того, повышение специфичности связывания для связывающих доменов «цинковый палец» описано, например, в находящемся в совместной собственности WO 02/077227.
[0102] Кроме того, как раскрыто в этих и других ссылках, домены «цинковый палец» и/или многопальцевые белки «цинковый палец» можно соединить вместе с помощью любых подходящих линкерных последовательностей, включающих, например, линкеры длиной 5 или более аминокислот. См. также патенты США №№ 6479626; 6903185 и 7153949 по вопросу типичных линкерных последовательностей длиной 6 или более аминокислот. Описанные здесь белки могут включать любую комбинацию подходящих линкеров между индивидуальными «цинковыми пальцами» белка.
[0103] Отбор целевых сайтов; ZFP и способы проектирования и конструирования слитых белков (и кодирующих их полинуклеотидов) известны специалистам в данной области и подробно описаны в патентах США №№ 61400815; 789538; 6453242; 6534261; 5925523; 6007988; 6013453; 6200759; WO 95/19431; WO 96/06166; WO 98/53057; WO 98/54311; WO 00/27878; WO 01/60970 WO 01/88197; WO 02/099084; WO 98/53058; WO 98/53059; WO 98/53060; WO 02/016536 и WO 03/016496.
[0104] Кроме того, как раскрыто в этих и других ссылках, домены «цинковый палец» и/или многопальцевые белки «цинковый палец» можно соединять вместе с помощью любых подходящих линкерных последовательностей, включающих, например, линкеры длиной 5 или более аминокислот. См. также патенты США №№ 6479626; 6903185 и 7153949 по вопросу типичных линкерных последовательностей длиной 6 или более аминокислот. Описанные здесь белки могут включать любую комбинацию подходящих линкеров между индивидуальными «цинковыми пальцами» белка.
[0105] Таким образом, нуклеаза включает ДНК-связывающий домен для специфической привязки к целевому сайту в любом гене, в который желательно ввести последовательность, кодирующую белок FIX.
Б. Расщепляющие домены
[0106] Любой подходящий расщепляющий домен может быть оперативно связан с ДНК-связывающим доменом с образованием нуклеазы. Например, ZFP ДНК-связывающие домены сливают с нуклеазными доменами для создания ZFN-функционального объекта, который способен распознавать свою предполагаемую мишень - нуклеиновую кислоту, благодаря генно-инженерному (ZFP) ДНК-связывающему домену, и вызвать разрез ДНК рядом с сайтом связывания ZFP посредством нуклеазной активности. См., например, работу Kim и другие (1996) Proc Natl Acad Sci USA 93 (3): 1156-1160. Позже ZFN были использованы для модификации генома в различных организмах. См., например, публикации патентов США 20030232410; 20050208489; 20050026157; 20050064474; 20060188987; 20060063231; и международную публикацию WO 07/014275. Аналогично сливают TALE ДНК-связывающие домены с нуклеазными доменами для создания TALEN. См., например, заявку США № 13/068735.
[0107] Как отмечено выше, расщепляющий домен может быть гетерологичен относительно ДНК-связывающего домена, например, ДНК-связывающий домен «цинковый палец» и расщепляющий домен из нуклеазы или TALEN ДНК-связывающий домен и расщепляющий домен, или ДНК-связывающий домен мегануклеазы и расщепляющий домен из другой нуклеазы. Гетерологичные расщепляющие домены можно получить из любой эндонуклеазы или экзонуклеазы. Типичные эндонуклеазы, из которых можно получить расщепляющий домен, включают, но не ограничены этим, рестрикционные эндонуклеазы и хоминг-эндонуклеазы. См., например, каталог 2002-2003 New England Biolabs, Beverly, MA; и Belfort и другие (1997) Nucleic Acids Res. 25:3379-3388. Известны дополнительные ферменты, которые расщепляют ДНК (например, S1 нуклеаза; нуклеаза из золотистой фасоли; DNase I поджелудочной железы; микрококковая нуклеаза; дрожжевые HO эндонуклеазы; см. также работу под редакцией Linn и другие Nucleases, Cold Spring Harbor Laboratory Press, 1993). Один или более из этих ферментов (или их функциональных фрагментов) можно использовать в качестве источника расщепляющих доменов и расщепляющих полудоменов.
[0108] Аналогично, расщепляющий полудомен можно получить из любой нуклеазы или ее части, как указано выше, что требует димеризации для обеспечения расщепляющей активности. Вообще, для расщепления необходимы два слитых белка, если слитые белки содержат расщепляющие полудомены. В качестве альтернативы можно использовать один белок, содержащий два расщепляющих полудомена. Два расщепляющих полудомена можно получить из одной эндонуклеазы (или ее функциональных фрагментов) или каждый расщепляющий полудомен можно получить из разных эндонуклеаз (или их функциональных фрагментов). Кроме того, целевые сайты для двух слитых белков предпочтительно располагаются относительно друг друга таким образом, что привязка двух слитых белков к их соответствующим целевым сайтам располагает расщепляющие полудомены пространственно ориентированным образом относительно друг друга, что позволяет расщепляющим полудоменам формировать функциональный расщепляющий домен, например, посредством димеризации. Таким образом, в некоторых вариантах ближние концы целевых сайтов разделены 5-8 нуклеотидами или 15-18 нуклеотидами. Однако между двумя целевыми сайтами может вмещаться любое целое количество нуклеотидов или нуклеотидных пар (например, от 2 до 50 пар нуклеотидов или более). Обычно сайт расщепления лежит между целевыми сайтами.
[0109] Рестрикционные эндонуклеазы (ферменты рестрикции) присутствуют во многих видах и способны к последовательность-специфическому связыванию с ДНК (по сайту распознавания) и расщеплению ДНК по сайту или вблизи сайта связывания. Некоторые рестрикционные ферменты (например, тип IIS) расщепляют ДНК по сайтам, удаленным от сайта распознавания и имеют отделимые связывающие и расщепляющие домены. Например, фермент типа IIS FokI катализирует двунитевой разрыв ДНК на расстоянии 9 нуклеотидов от своего сайта распознавания на одной нити и на расстоянии 13 нуклеотидов от своего сайта распознавания на другой нити. См., например, патенты США 5356802; 5436150 и 5487994; а также работы Li и другие (1992) Proc. Natl. Acad. Sci. USA 89:4275-4279; Li и другие (1993) Proc. Natl. Acad. Sci. USA 90:2764-2768; Kim и другие (1994a) Proc. Natl. Acad. Sci. USA 91:883-887; Kim и другие (1994b) J. Biol. Chem. 269:31,978-31, 982. Таким образом, в одном варианте слитые белки содержат расщепляющий домен (или расщепляющий полудомен), по меньшей мере, из одного рестрикционного фермента типа IIS и одного или нескольких связывающих доменов «цинковый палец», которые могут быть или не быть генно-инженерными.
[0110] Типичный рестрикционный фермент типа IIS, расщепляющий домен которого является отделимым от связывающего домена, представляет собой FokI. Этот особый фермент активен как димер. Bitinaite и другие (1998) Proc. Natl. Acad. Sci. USA 95: 10570-10575. Таким образом, для целей настоящего раскрытия часть фермента FokI, используемого в раскрытых слитых белках, считают расщепляющим полудоменом. Таким образом, для целевого двунитевого разрыва и/или целевого замещения клеточных последовательностей с использованием слияния «цинковый палец»-FokI: можно использовать два слитых белка, каждый содержащий FokI расщепляющий полудомен, с целью восстановления каталитически активного расщепляющего домена. По-другому, также можно использовать одну молекулу полипептида, содержащую связывающий домен «цинковый палец» и два расщепляющих полудомена FokI. Параметры для целевого расщепления и целевого изменения последовательности с использованием слияний цинковый палец-FokI приведены также в других местах данного раскрытия.
[0111] Расщепляющим доменом или расщепляющим полудоменом может быть любая часть белка, которая сохраняет расщепляющую активность или которая сохраняет способность мультимеризоваться (например, димеризоваться) с образованием функционального расщепляющего домена.
[0112] Типичные рестрикционные ферменты типа IIS описаны в международной публикации WO 07/014275, включенной здесь во всей своей полноте. Дополнительные рестрикционные ферменты также содержат отделимые связывающие и расщепляющие домены, и они рассматриваются в настоящем раскрытии. См., например, работу Roberts и другие (2003) Nucleic Acids Res. 31:418-420.
[0113] В некоторых вариантах расщепляющий домен содержит один или более генно-инженерных расщепляющих полудоменов (также обозначены как димеризационные мутанты доменов), которые минимизируют или предотвращают гомодимеризацию, как описано, например, в публикациях патентов США №№ 20050064474; 20060188987; 20070305346 и 20080131962, раскрытия всех их включены здесь в виде ссылки во всей своей полноте. Аминокислотные остатки в положениях 446, 447, 479, 483, 484, 486, 487, 490, 491, 496, 498, 499, 500, 531, 534, 537 и 538 FokI все являются мишенями для вызывающей изменения димеризации расщепляющих полудоменов FokI.
[0114] Типичные генно-инженерные расщепляющие полудомены из FokI, которые образуют облигатные гетеродимеры, включают пару, в которой первый расщепляющий полудомен включает мутации аминокислотных остатков в положениях 490 и 538 FokI, и второй расщепляющий полудомен включает мутации аминокислотных остатков 486 и 499.
[0115] Таким образом, в одном варианте мутация в положении 490 заменяет Glu (E) остатком Lys (K); мутация на 538 заменяет Iso (I) остатком Lys (K); мутация на 486 заменяет Gln (Q) остатком Glu (E); и мутация в положении 499 заменяет Iso (I) остатком Lys (K). В частности, описанные здесь генно-инженерные расщепляющие полудомены получены посредством мутации положений 490 (E→K) и 538 (I→K) в одном расщепляющем полудомене с образованием генно-инженерного расщепляющего полудомена, обозначенного «E490K:I538K», и посредством мутации положений 486 (Q→E) и 499 (I→L) в другом расщепляющем полудомене с образованием генно-инженерного расщепляющего полудомена, обозначенного «Q486E:I499L». Описанные здесь генно-инженерные расщепляющие полудомены являются облигатными гетеродимерными мутантами, в которых аберрантное расщепление минимизировано или аннулировано. См., например, публикацию патента США № 2008/0131962, раскрытие которого включено в виде ссылки во всей своей полноте для всех целей. В некоторых вариантах генно-инженерный расщепляющий полудомен содержит мутации в положениях 486, 499 и 496 (нумерация относительно FokI дикого типа), например, мутации, которые заменяют остаток Gin (Q дикого типа) в положении 486 остатком Glu (E), остаток Iso (I) дикого типа в положении 499 остатком Leu (L) и остаток Asn (N) дикого типа в положении 496 остатком Asp (D) или остатком Glu (E) (также обозначаемыми как домены «ELD» и «ELE», соответственно). В других вариантах генно-инженерный расщепляющий полудомен содержит мутации в положения 490, 538 и 537 (нумерация относительно FokI дикого типа), например, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 остатком Lys (K), остаток Iso (I) дикого типа в положении 538 остатком Lys (K) и остаток His (H) дикого типа в положении 537 остатком Lys (K) или Arg (R) (также обозначаемыми как домены «KKK» и «KKR», соответственно). В других вариантах генно-инженерный расщепляющий полудомен содержит мутации в положениях 490 и 537 (нумерация относительно FokI дикого типа), например, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 остатком Lys (K) и остаток His (H) дикого типа в положении 537 остатком Lys (K) или остатком Arg (R) (также обозначаемыми как домен «ΚIΚ» и «KIR», соответственно). (См. публикацию патента США № 20110201055). В других вариантах генно-инженерный расщепляющий полудомен содержит мутации «Sharkey» и/или «Sharkey'» (см. работу Guo и другие (2010) J. Mol. Biol. 400(1):96-107).
[0116] Описанные здесь генно-инженерные расщепляющие полудомены можно получить, применяя любой подходящий способ, например, способом сайт-направленного мутагенеза расщепляющего полудомена (FokI) дикого типа, как описано в публикациях патентов США №№ 20050064474; 20080131962 и 20110201055.
[0117] По-другому, нуклеазы можно собрать in vivo на целевом сайте нуклеиновой кислоты, используя так называемую методику «сплит-фермент» (см., например, публикацию патента США № 20090068164). Компоненты таких сплит-ферментов можно экспрессировать на отдельных экспрессионных конструкциях или можно соединить в одной открытой рамке считывания, где индивидуальные компоненты разделены, например, саморасщепляющимся пептидом 2A или последовательностью IRES. Компоненты могут представлять собой индивидуальные связывающие домены «цинковый палец» или домены из нуклеиновая кислота-связывающих доменов мегануклеазы.
[0118] Можно проводить скрининг нуклеаз на активность до их использования, например, в хромосомной системе на основе дрожжей, как описано в WO 2009/042163 и 20090068164. Нуклеазные экспрессионные конструкции можно легко конструировать, применяя способы, известные в данной области. См., например, публикации патентов США 20030232410; 20050208489; 20050026157; 20050064474; 20060188987; 20060063231 и международную публикацию WO 07/014275. Экспрессия нуклеазы может происходить под контролем конститутивного промотора или индуцибельного промотора, например, галактокиназного промотора, который активируется (дерепрессируется) в присутствии раффинозы и/или галактозы и репрессируется в присутствии глюкозы.
Целевые сайты
[0119] Как подробно описано выше, можно сконструировать домены ДНК для привязки к любой выбранной последовательности, например, в эндогенном гене FIX или эндогенном или введенном гене «безопасная гавань». Генно-инженерный ДНК связывающий домен может иметь новую специфичность связывания по сравнению с встречающимся в природе ДНК-связывающим доменом. Способы генной инженерии включают, но не ограничены этим, рациональную конструкцию и различные типы отбора. Рациональная конструкция включает, например, использование баз данных, содержащих триплетные (или квадруплетные) последовательности нуклеотидов и индивидуальные аминокислотные последовательности «цинковый палец», в которых каждая триплетная или квадруплетная нуклеотидная последовательность ассоциирована с одной или несколькими аминокислотными последовательностями «цинковых пальцев», которые связывают конкретную триплетную или квадруплетную последовательность. См., например, находящиеся в совместной собственности патенты США 6453242 и 6534261, включенные здесь в виде ссылок во всей своей полноте. Также можно получить рациональную конструкцию TAL-эффекторных доменов. См., например, предварительные заявки США №№ 61/395836 и 61/401429 от 17 мая 2010 и 21 августа 2010, соответственно.
[0120] Типичные способы отбора, применимые для ДНК-связывающих доменов и включающие фаговый дисплей и двугибридные системы, раскрыты в патентах США 5789538; 5925523; 6007988; 6013453; 6410248; 6140466; 6200759 и 6242568; а также WO 98/37186; WO 98/53057; WO 00/27878; WO 01/88197 и GB 2338237. Кроме того, повышение специфичности связывания для связывающих доменов «цинковый палец» описано, например, в находящемся в совместной собственности WO 02/077227.
[0121] Отбор целевых сайтов; нуклеазы и способы проектирования и конструирования слитых белков (и кодирующих их полинуклеотидов) известны специалистам в данной области и подробно описаны в публикациях патентных заявок США №№ 20050064474 и 20060188987, включенных здесь в виде ссылок во всей своей полноте.
[0122] Кроме того, как указано в этих и других ссылках, ДНК-связывающие домены (например, многопальцевые белки «цинковый палец») могут быть соединены вместе с помощью любых подходящих линкерных последовательностей, включающих, например, линкеры из 5 или большего количества аминокислот. См., например, патенты США №№ 6479626; 6903185 и 7153949 на предмет типичных линкерных последовательностей длиной 6 или более аминокислот. Описанные здесь белки могут включать любую комбинацию подходящих линкеров между индивидуальными ДНК-связывающими доменами белков. См. также заявку США № 13/066735.
[0123] Для лечения гемофилии B посредством целенаправленного введения последовательности, кодирующей функциональный белок FIX, любой желательный сайт вставки в геноме субъекта расщепляют нуклеазой, которая стимулирует целевое введение донорного полинуклеотида, несущего FIX-кодирующую последовательность. ДНК-связывающие домены нуклеаз могут быть нацелены на любой желательный сайт в геноме.
[0124] В некоторых вариантах ДНК-связывающий домен нуклеазы нацелен на эндогенный FIX (F9) ген, как описано, например, в патентной заявке США № 12/798749. Целевые сайты могут быть в любом месте кодирующей последовательности или выше или ниже кодирующей последовательности. В некоторых вариантах, целевой сайт(ы) находится около 3'-конца кодирующей последовательности.
[0125] В других вариантах нуклеаза (компонент ДНК-связывающего домена) предназначена для локуса «безопасная гавань», который включает (только в качестве примера) ген AAVS1 (см. публикацию США № 20080299580), ген CCR5 (см. публикацию США № 20080159996) и/или локус Rosa (см. WO 2010/065123).
Донорные последовательности
[0100] Для лечения гемофилии донорная последовательность включает последовательность, кодирующую функциональный белок FIX или его часть, что дает в результате последовательность, кодирующую и экспрессирующую функциональный белок FIX после интеграции донора. Донорную молекулу можно ввести в эндогенный ген таким образом, чтобы экспрессировались все, некоторые или никакие эндогенные гены. Например, можно ввести, как здесь описано, трансген, содержащий функциональные FIX последовательности, в эндогенный локус FIX, таким образом, чтобы экспрессировались некоторые или никакие из эндогенных FIX трансгеном FIX (например, донор может корректировать мутацию таким образом, чтобы экспрессировались эндогенные последовательности дикого типа). В других вариантах трансген FIX интегрируют в какой-либо эндогенный локус, например, локус «безопасная гавань» (эндогенный или введенный). См., например, публикации патентов США 20080299580; 20080159996 и 201000218264.
[0126] Донорную последовательность FIX можно ввести в клетку до, одновременно или после экспрессии слитого белка(ов). Донорный полинуклеотид FIX обычно обладает достаточной гомологией с геномной последовательностью, чтобы поддерживать гомологичную рекомбинацию (или направляемую гомологией репарацию) между ним и геномной последовательностью, на которую он переносит гомологию. См., например, публикации патентов США №№ 2005/0064474; 2007/0134796 и 2009/0263900. Легко понять, что донорная последовательность обычно неидентична геномной последовательности, которую она замещает. Например, последовательность донорного полинуклеотида может содержать одно или более изменений, введений, удалений, инверсий или перестановок отдельного основания относительно геномной последовательности, до тех пор, пока имеется достаточная гомология с хромосомными последовательностями. По-другому, донорная последовательность может содержать негомологичную последовательность, фланкированную двумя областями гомологии. По-другому, донорные последовательности могут включать векторную молекулу, содержащую последовательности, которые негомологичны области, представляющей интерес, в клеточном хроматине. Донорная молекула может содержать несколько перемежающихся областей гомологии с клеточным хроматином. Например, для целевого введения последовательностей, обычно не присутствующих в области, представляющей интерес, указанные последовательности могут присутствовать в молекуле донорной нуклеиновой кислоты и быть фланкированы областями гомологии с последовательностью в области, представляющей интерес.
[0127] Донорный полинуклеотид FIX может представлять собой ДНК или РНК, однонитевую или двунитевую и может быть введен в клетку в линейной или кольцевой форме. При введении в линейной форме концы донорной последовательности можно защитить (например, от экзонуклеотической деградации) способами, известными специалистам в данной области. Например, добавляют один или несколько дидезоксинуклеотидных остатков к 3'-концу линейной молекулы и/или лигируют самокомплементарные олигонуклеотиды по одному или обоим концам. См., например, работы Chang и другие (1987) Proc. Natl. Acad. Sci. USA 84:4959-4963; Nehls и другие (1996) Science 272:886-889. Дополнительные способы защиты экзогенных полинуклеотидов от деградации включают, но не ограничены этим, добавление терминальной аминогруппы(групп) и использование модифицированных межнуклеотидных связей, таких как, например, фосфоротиоаты, фосфорамидаты и O-метилрибозные или дезоксирибозные остатки. Полинуклеотид можно ввести в клетку как часть векторной молекулы, имеющей дополнительные последовательности, например, репликаторы, промоторы и гены, кодирующие устойчивость к антибиотику. Кроме того, донорные полинуклеотиды можно ввести как «голую» нуклеиновую кислоту, как нуклеиновую кислоту в виде комплекса с таким агентом как липосома или полоксамер или можно доставить при помощи вирусов (например, аденовируса, ААВ, вируса герпеса, ретровируса, лентивируса).
[0128] Донор FIX обычно вводят таким образом, что его экспрессия управляется эндогенным промотором на сайте интеграции (например, эндогенным FIX промотором, если донор интегрирован в дефектный FIX (F9) локус пациента). Однако очевидно, что донор может включать промотор и/или энхансер, например, конститутивный промотор или индуцибельный или ткань-специфический (например, печень-специфический) промотор, который управляет экспрессией функционального белка FIX после интеграции.
[0129] Донорную последовательность FIX можно интегрировать специально в любой выбранный целевой сайт, устраняя тем самым результаты, связанные со случайной интеграцией при традиционной генной терапии. В некоторых вариантах донорную последовательность интегрируют в эндогенный локус FIX для коррекции дефицита у пациента с гемофилией B. В других вариантах донорную последовательность FIX интегрируют в локус «безопасная гавань», например локус CCR5, локус AAVS1 или подобный.
[0130] Кроме того, хотя для экспрессии не требуется, экзогенные последовательности могут также содержать транскрипционные или трансляционные регуляторные последовательности, например, промоторы, энхансеры, изоляторы, внутренние сайты входа рибосомы, последовательности, кодирующие 2A пептиды и/или сигналы полиаденилирования.
Доставка
[0131] Нуклеазы, полинуклеотиды, кодирующие эти нуклеазы, донорные полинуклеотиды и композиции, содержащие описанные здесь белки и/или полинуклеотиды, можно доставлять in vivo или ex vivo любыми подходящими способами.
[0132] Способы доставки описанных здесь нуклеаз описаны, например, в патентах США №№ 6453242; 6503717; 6534261; 6599692; 6607882; 6689558; 6824978; 6933113; 6979539; 7013219 и 7163824, раскрытия которых включены здесь в виде ссылок во всей своей полноте.
[0133] Нуклеазы и/или донорные конструкции, которые здесь описаны, также можно доставлять, используя векторы, содержащие последовательности, кодирующие один или более белков «цинковый палец». Можно использовать любую векторную систему, включая, но не ограничиваясь этим, плазмидные векторы, ретровирусные векторы, лентивирусные векторы, аденовирусные векторы, поксивирусные векторы; векторы герпеса и векторы аденоассоциированного вируса и другие. См. также патенты США №№ 6534261; 6607882; 6824978; 6933113; 6979539; 7013219 и 7163824, включенные здесь в виде ссылок во всей своей полноте. Кроме того, очевидно, что любой из этих векторов может содержать одну или более последовательностей, необходимых для лечения. Таким образом, когда вводят в клетку одну или более нуклеаз и донорную конструкцию, то нуклеазы и/или донорный полинуклеотид можно переносить на одном векторе или на разных векторах. Когда используют несколько векторов, то каждый вектор может содержать последовательность, кодирующую одну или несколько нуклеаз и/или донорных конструкций.
[0134] Для введения нуклеиновых кислот, кодирующих нуклеазы, и донорных конструкций в клетки (например, клетки млекопитающих) и ткани можно применять обычные способы переноса генов на вирусной и невирусной основе. Невирусные векторные системы доставки включают ДНК плазмиды, «голую» нуклеиновую кислоту и нуклеиновую кислоту в виде комплекса со средством доставки, таким как липосома или полоксамер. Системы вирусной векторной доставки включают ДНК и РНК вирусы, которые после доставки в клетку имеют эписомные или интегрированные геномы. Обзоры по in vivo доставке генно-инженерных ДНК-связывающих белков и слитых белков, включающих эти связывающие белки, см., например, в работах Rebar (2004) Expert Opinion Invest. Drugs 13(7):829-839; Rossi и другие (2007) Nature Biotech. 25(12):1444-1454, а также в основных ссылках по доставке генов: Anderson, Science 256:808-813 (1992); Nabel & Feigner, TIBTECH 11:211-217 (1993); Mitani & Caskey, TIBTECH 11:162-166 (1993); Dillon, TIBTECH 11:167-175 (1993); Miller, Nature 357:455-460 (1992); Van Brunt, Biotechnology 6(10):1149-1154 (1988); Vigne, Restorative Neurology and Neuroscience 8:35-36 (1995); Kremer & Perricaudet, British Medical Bulletin 51(l):31-44 (1995); Haddada и другие, в работе Current Topics in Microbiology and Immunology, Doerfier и Bohm (редакторы) (1995); и Yu и другие, Gene Therapy 1:13-26 (1994).
[0135] Способы невирусной доставки нуклеиновых кислот включают электропорацию, липофекцию, микроинъекцию, биолистику, использование виросом, липосом, иммунолипосом, поликатионов или конъюгатов липид:нуклеиновая кислота, «голой» ДНК, использование искусственных вирионов и агент-усиленный захват ДНК. Для доставки нуклеиновых кислот можно также применять сонопорацию с использованием, например, системы Sonitron 2000 (Rich-Мар).
[0136] Дополнительные типичные системы доставки нуклеиновых кислот включают системы, поставляемые Amaxa Biosystems (Cologne, Germany), Maxcyte, Inc. (Rockville, Maryland), BTX Molecular Delivery Systems (Holliston, MA) и Copernicus Therapeutics Inc, (см., например, US6008336). Липофекция описана, например, в патентах США №№ 5049386; 4946787 и 4897355), и реагенты липофекции продаются коммерчески (например, от Transfectam™ и Lipofectin™). Катионные и нейтральные липиды, которые пригодны для эффективной рецептор-распознающей липофекции полинуклеотидов, включают липиды от Feigner, WO 91/17424, WO 91/16024.
[0137] Получение комплексов липид:нуклеиновая кислота, включающих целевые липосомы, таких как иммунолипидные комплексы, хорошо известно специалистам в данной области (см., например, работы Crystal, Science 270:404-410 (1995); Blaese и другие, Cancer Gene Ther. 2:291-297 (1995); Behr и другие, Bioconjugate Chem. 5:382-389 (1994); Remy и другие, Bioconjugate Chem. 5:647-654 (1994); Gao и другие, Gene Therapy 2:710-722 (1995); Ahmad и другие, Cancer Res. 52:4817-4820 (1992); патенты США №№ 4186183, 4217344, 4235871, 4261975, 4485054, 4501728, 4774085, 4837028 и 4946787).
[0138] Дополнительные способы доставки включают использование упаковки нуклеиновых кислот, подлежащих доставке, в EnGeneIC средства доставки (EDV). Эти EDV специально доставляются в ткани-мишени с использованием биспецифических антител, где одно плечо антитела обладает специфичностью относительно ткани-мишени и другое специфичностью относительно EDV. Антитело приносит EDV к поверхности клетки-мишени, и затем EDV доставляется в клетку посредством эндоцитоза. После этого содержимое высвобождается в клетке (см. работу MacDiarmid и другие (2009) Nature Biotechnology 27(7):643).
[0139] Применение РНК или ДНК вирусных систем для доставки нуклеиновых кислот, кодирующих генно-инженерные ZFP, имеет преимущество высоко развитых процессов направленного действия вируса на конкретные клетки в организме и переноса вирусной нагрузки в ядро. Вирусные векторы можно вводить пациентам непосредственно (in vivo) или можно использовать для обработки клеток in vitro и вводить пациентам модифицированные клетки (ex vivo). Обычные системы на основе вирусов для доставки ZFP включают, но не ограничены этим, следующие вирусные векторы для переноса генов: антиретровирусный, лентивирусный, аденовирусный, аденоассоциированный, вектор вируса коровьей оспы и вектор вируса простого герпеса. Интеграция в хозяйский геном возможна в способах переноса генов посредством ретровируса, лентивируса и аденоассоциированного вируса, часто приводя к длительной экспрессии введенного трансгена. Кроме того, в клетках и тканях многих различных типов наблюдается высокая эффективность трансдукции.
[0140] Тропизм ретровируса можно изменить путем включения чужеродных белков оболочки, расширяя потенциальную целевую группу клеток-мишеней. Лентивирусные векторы являются ретровирусными векторами, которые способны к трансдукции или инфицированию неделящихся клеток и обычно дают высокие вирусные титры. Выбор системы переноса ретровирусного гена зависит от целевой ткани. Ретровирусные векторы состоят из цис-активных длинных концевых повторов с вместимостью упаковки до 6-10 kb чужеродной последовательности. Минимальные цис-активные LTR достаточны для репликации и упаковки векторов, которые затем используют для интеграции терапевтического гена в клетку-мишень с целью обеспечения постоянной трансгенной экспрессии. Широко применяемые ретровирусные векторы включают векторы на основе вируса мышиной лейкемии (MuLV), вируса лейкемии обезьян гиббонов (GaLV), вируса иммунодефицита обезьян (SIV), вируса иммунодефицита человека (HIV) и их комбинации (см., например, работы Buchscher и другие, J. Virol. 66:2731-2739 (1992); Johann и другие, J. Virol. 66:1635-1640 (1992); Sommerfelt и другие, Virol. 176:58-59 (1990); Wilson и другие, J.Virol. 63:2374-2378 (1989); Miller и другие, J. Virol. 65:2220-2224 (1991); РСТ US94/05700).
[0141] В приложениях, где предпочтительна временная экспрессия, можно применять аденовирусную систему. Векторы на основе аденовирусов способны к очень высокой эффективности трансдукции во многих типах клеток и не требуют деления клеток. С такими векторами были получены высокий титр и высокие уровни экспрессии. Этот вектор можно получить в больших количествах в относительно простой системе. Векторы на основе аденоассоциированных вирусов («AAV») используют также для трансдукции клеток целевыми нуклеиновыми кислотами, например, при in vitro получении нуклеиновых кислот и пептидов и для процедур in vivo и ex vivo генной терапии (см., например, West и другие, Virology 160:38-47 (1987); патент США № 4797368; WO 93/24641; Kotin, Human Gene Therapy 5:793-801 (1994); Muzyczka, J. Clin. Invest. 94:1351 (1994). Конструкция рекомбинантных векторов AAV описана в ряде публикаций, включая патент США № 5173414; Tratschin и другие, Mol Cell. Biol. 5:3251-3260 (1985); Tratschin, и другие, Mol. Cell. Biol. 4:2072-2081 (1984); Hermonat и Muzyczka, PNAS 81:6466-6470 (1984); и Samulski и другие, J. Virol. 63:03822-3828 (1989).
[0142] В настоящее время доступны, по меньшей мере, шесть подходов с использованием вирусных векторов для переноса генов в клинических испытаниях, где применяют способы, которые включают комплементацию дефектных векторов генами, вводимыми в хелперные клеточные линии для генерации трансдуцирующего агента.
[0143] pLASN и MFG-S являются примерами ретровирусных векторов, которые используют в клинических испытаниях (Dunbar и другие, Blood 85:3048-305 (1995); Kohn и другие, Nat. Med. 1:1017-102 (1995); Malech и другие, PNAS 94:22 12133-12138 (1997)). PA317/pLASN является первым терапевтическим вектором, использованным в генно-терапевтическом исследовании. (Blaese и другие, Science 270:475-480 (1995)). Эффективность трансдукции 50% или выше наблюдают для упакованных векторов MFG-S. (Ellem и другие, Immunol Immunother. 44(l):10-20 (1997); Dranoff и другие, Hum. Gene Ther. 1:111-2 (1997).
[0144] Рекомбинантные аденоассоциированные вирусные векторы (rAAV) представляют собой многообещающие альтернативные системы доставки генов на базе дефектного и непатогенного парвовируса аденоассоциированного вируса типа 2. Все векторы являются производными плазмиды, которая сохраняет только AAV 145 bp инвертированные терминальные повторы, фланкирующие трансген-экспрессирующую кассету. Ключевыми характеристиками для этой векторной системы являются эффективный перенос гена и стабильная доставка трансгена, благодаря интеграции в геномы трансдуцированных клеток. (Wagner и другие, Lancet 351:9117 1702-3 (1998), Kearns и другие, Gene Ther. 9:748-55 (1996)). Согласно настоящему изобретению также можно использовать другие AAV серотипы, в том числе AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9 и AAVrh.10 и любой новый AAV серотип.
[0145] Можно получить репликативно-дефицитные рекомбинантные аденовирусные векторы (Ad) с высоким титром и легко инфицировать ряд различных типов клеток. Большинство аденовирусных векторов сконструированы таким образом, что трансген замещает Ad E1a, E1b и/или E3 гены; затем репликативно-дефектный вектор репродуцируется в человеческих клетках 293, которые снабжают удаленную функцию гена в транс. Ad векторы могут трансдуцировать многие типы тканей in vivo, в том числе неделящиеся, дифференцированные клетки, такие как клетки, обнаруживаемые в печени, почках и мышцах. Обычные векторы Ad имеют большую несущую способность. Пример использования вектора Ad в клиническом испытании включает полинуклеотидную терапию для противоопухолевой иммунизации посредством внутримышечной инъекции (Sterman и другие, Hum. Gene Ther. 7:1083-9 (1998)). Дополнительные примеры использования аденовирусных векторов для переноса генов в клинических испытаниях включают работы Rosenecker и другие, Infection 24:1 5-10 (1996); Sterman и другие, Hum. Gene Ther. 9:7 1083-1089 (1998); Welsh и другие, Hum. Gene Ther. 2:205-18 (1995); Alvarez и другие, Hum. Gene Ther. 5:597-613 (1997); Topf и другие, Gene Ther. 5:507-513 (1998); Sterman и другие, Hum. Gene Ther. 7:1083-1089 (1998).
[0146] Для получения вирусных частиц, которые способны инфицировать клетки-хозяев используют упаковывающие клетки. Такие клетки включают клетки 293, которые упаковывают аденовирус, и клетки ψ2 или клетки PA317, которые упаковывают ретровирус. Вирусные векторы, применяемые в генной терапии, обычно генерируются клеточной линией-продуцентом, которая упаковывает нуклеиновокислотный вектор в вирусную частицу. Векторы обычно содержат минимальные вирусные последовательности, необходимые для упаковки и последующей интеграции в хозяина (если применимо), другие вирусные последовательности заменяются экспрессионной кассетой, кодирующей белок, подвергаемый экспрессии. Отсутствующие вирусные функции подают в транс-положение посредством упаковочной линии клеток. Например, векторы AAV, обычно применяемые в генной терапии, имеют только последовательности инвертированных концевых повторов (ITR) из генома AAV, необходимые для упаковки и интеграции в геном хозяина. Вирусную ДНК упаковывают в клеточной линии, которая содержит хелперную плазмиду, кодирующую другие гены AAV, а именно rep и cap, но не содержит ITR последовательности. Клеточную линию также инфицируют аденовирусом в качестве хелпера. Хелперный вирус промотирует репликацию вектора AAV и экспрессию генов AAV из хелперной плазмиды. Хелперная плазмида не упаковывается в значительных количествах из-за отсутствия ITR последовательностей. Заражение аденовирусом можно снизить, например, посредством термообработки, к которой аденовирус более чувствителен, чем AAV.
[0147] Во многих генно-терапевтических приложениях желательно, чтобы генно-терапевтический вектор доставлялся в конкретный тип ткани с высокой степенью специфичности. Таким образом, вирусный вектор можно модифицировать таким образом, чтобы он имел специфичность относительно данного типа клеток, посредством экспрессии лиганда как слитого белка с белком вирусной оболочки на внешней поверхности вируса. Выбирают лиганд, обладающий аффинностью относительно рецептора, который, как известно, присутствует на клетках представляющего интерес типа. Например, в работе Han и другие, Proc. Natl. Acad. Sci. USA 92:9747-9751 (1995) сообщается, что вирус мышиной лейкемии Молони можно модифицировать для экспрессии человеческого херегулина, слитого с gp70, и рекомбинантный вирус инфицирует определенные раковые клетки молочной железы человека, экспрессирующие рецептор человеческого эпидермального фактора роста. Этот принцип можно распространить на другие вирус-меченые пары клеток, в которых клетка-мишень экспрессирует рецептор, и вирус экспрессирует слитой белок, включающий лиганд для рецептора на поверхности клетки. Например, можно сконструировать нитевидный фаг для отображения фрагментов антител (например, FAB или Fv), обладающий специфической аффинностью связывания в отношении практически любого выбранного клеточного рецептора. Хотя приведенное выше описание применяется главным образом к вирусным векторам, те же принципы можно применять к невирусным векторам. Можно сконструировать такие векторы, содержащие последовательности для специфического внедрения, которые способствуют захвату конкретными клетками-мишенями.
[0148] Векторы для генной терапии можно доставить in vivo посредством введения индивидуальному пациенту, обычно посредством системного введения (например, внутривенного, внутрибрюшного, внутримышечного, подкожного или внутричерепного введения) или локального применения, как описано ниже. Кроме того векторы можно доставлять в клетки ex vivo, например, в эксплантируемые клетки от индивидуального пациента (например, лимфоциты, пунктат костного мозга, биопсию ткани) или универсальные донорные гемопоэтические стволовые клетки с последующей реплантацией клеток пациенту, обычно после отбора клеток, которые имеют включенный вектор.
[0149] Векторы (например, ретровирусы, аденовирусы, липосомы и т.д.), содержащие нуклеазы и/или донорные конструкции, также можно вводить непосредственно в организм для трансдукции клеток in vivo. По-другому, можно вводить «голую» ДНК. Введение осуществляют любым из способов, обычно применяемых для приведения молекулы в окончательный контакт с клетками крови или ткани и включающих, но не ограниченных этим, инъекцию, инфузию, локальное применение и электропорацию. Подходящие способы введения таких нуклеиновых кислот доступны и хорошо известны специалистам в данной области, и хотя для введения конкретной композиции можно применять более одного способа, конкретный способ часто может обеспечить более быструю и более эффективную реакцию, чем другой способ.
[0150] Описанные здесь векторы, пригодные для внедрения полинуклеотидов (например, нуклеаза-кодирующих и/или FIX-кодирующих), включают неинтегрирующие лентивирусные векторы (IDLV). См., например, работы Ory и другие (1996) Proc. Natl. Acad. Sci. USA 93:11382-11388; Dull и другие (1998) J. Virol. 72:8463-8471; Zuffery и другие (1998) J. Virol. 72:9873-9880; Follenzi и другие (2000) Nature Genetics 25:217-222; публикацию патента США № 2009/054985.
[0151] Фармацевтически приемлемые носители определяются отчасти составом подлежащей введению композиции, а также конкретным способом, применяемым для введения композиции. Таким образом, имеется широкое разнообразие подходящих препаратов доступных фармацевтических композиций, которые описаны ниже (см., например, Remington's Pharmaceutical Sciences, 17 изд., 1989).
[0152] Очевидно, что нуклеаза-кодирующие последовательности и донорные конструкции можно доставлять, используя одну или разные системы. Например, донорный полинуклеотид можно переносить посредством плазмиды, тогда как одну или более нуклеаз можно переносить посредством вектора AAV. Кроме того, различные векторы можно вводить одинаковыми или разными способами (внутримышечно, посредством инъекции в хвостовую вену, другой внутривенной инъекции, посредством внутрибрюшинного введения или внутримышечной инъекции). Векторы можно доставлять одновременно или в любой последовательности.
[0153] Таким образом, настоящее раскрытие включает in vivo или ex vivo лечение гемофилии B посредством нуклеаза-опосредованной интеграции FIX-кодирующей последовательности. Композиции вводят пациенту в количестве, эффективном для получения желательной концентрации терапевтического полипептида FIX в сыворотке, печени или клетках-мишенях. Введение можно осуществлять любыми средствами, когда полинуклеотиды доставляют к желательным клеткам-мишеням. Например, рассматривают оба способа, in vivo и ex vivo. Предпочтительным способом введения является внутривенная инъекция в воротную вену. Другие способы in vivo введения включают, например, прямую инъекцию в доли печени или желчный проток и внутривенную дистальную инъекцию в печень, в том числе через печеночную артерию, прямую инъекцию в паренхиму печени, инъекции через печеночную артерию и/или ретроградное заполнение через билиарное дерево. Ex vivo модели введения включают трансдукцию in vitro резецированных гепатоцитов или других клеток печени с последующей инфузией трансдуцированных резецированных гепатоцитов обратно в воротную зону сосудистой системы, печеночную паренхиму или билиарное дерево пациента-человека (см., например, работу Grossman и другие (1994) Nature Genetics (1994), 6:335-341.
[0154] Эффективное количество нуклеазы(нуклеаз) и FIX донора, подлежащих введению, варьируется от пациента к пациенту и в соответствии с терапевтическим полипептидом, представляющим интерес. Таким образом, эффективные количества лучше всего определяет лечащий врач, вводящий композиции, и соответствующие дозы может легко определить рядовой специалист в данной области. По прошествии времени, достаточного для интеграции и экспрессии (обычно, например, 4-15 дней), проводят анализ уровней терапевтического полипептида в сыворотке или других тканях и сравнение с начальным уровнем до введения, определяя, является ли подлежащее введению количество слишком низким, в правильном диапазоне или слишком высокими. Подходящие схемы начального и последующих введений также являются вариабельными, но типичны в том, что за начальным введением, если необходимо, следуют последующие введения. Последующие введения можно производить с переменными интервалами в диапазоне от ежедневного введения, до ежегодного, до одного введения каждые несколько лет. Специалисту в данной области понятно, что подходящие иммуносупрессивные способы можно рекомендовать, чтобы избежать ингибирования или блокады трансдукции при иммуносупрессии векторов доставки, см., например, работу Vilquin и другие (1995) Human Gene Ther., 6:1391-1401.
[0155] Препараты для введений ex vivo и in vivo включают суспензии в жидкости или эмульгированные жидкости. Активные ингредиенты часто смешивают с наполнителями, которые фармацевтически приемлемы и совместимы с активным ингредиентом. Подходящие наполнители включают, например, воду, физиологический раствор, декстрозу, глицерин, этанол или подобные и их комбинации. Кроме того, композиция может содержать незначительные количества вспомогательных веществ, таких как смачивающие агенты или эмульгаторы, pH буферные агенты, стабилизаторы или другие реагенты, которые повышают эффективность фармацевтической композиции.
[0156] Следующие примеры касаются типичных вариантов настоящего раскрытия, в которых нуклеаза включает нуклеазу «цинковый палец» (ZFN). Понятно, что этот пример приведен только для иллюстративных целей, и что можно использовать другие нуклеазы, например, хоминг-эндонуклеазы (мегануклеазы) с генно-инженерными ДНК-связывающими доменами и/или слияния ДНК-связывающих доменов природных или генно-инженерных хоминг-эндонуклеаз (мегануклеаз) и гетерологичных расщепляющих доменов или TALEN.
ПРИМЕРЫ
Пример 1: FIX-специфические ZFN и их применение для целенаправленной интеграции
[0157] Перенос генов как стратегия лечения генетического заболевания успешно выполнен на различных животных моделях заболевания и совсем недавно применен к человеку (см., например, работы Aiuti и другие (2009 N. Engl. J. Med. 358: 2240-2248; Carrier и другие (2009) Science 326: 818-823). Применяют направленное воздействие на ген с целью корректировки ex vivo культивируемых ES-подобных индуцированных плюрипотентных стволовых клеток (Hanna и другие (2007) Science 318: 1920-1923), но большинство генетических заболеваний затрагивают системы органов, где в настоящее время невыполнимы ex vivo манипуляции. Одним таким органом является печень, главное место синтеза белков плазмы, в том числе факторов свертывания крови. Модельным генетическим заболеванием для генной терапии печени является гемофилия B, вызываемая дефицитом фактора свертывания крови IX (FIX), кодированного геном F9. Целенаправленная интеграция (TI) экзонов дикого типа 2-8 в F9 интрон 1 допускает сплайсинг кодирующей последовательности дикого типа с экзоном 1 (фигура 1А), приводя к экспрессии FIX дикого типа и избавлению от дефекта, обусловленного большинством мутаций F9. Таким образом, пытаются исследовать, может ли ZFN в сочетании с целенаправленным вектором, несущим F9 экзоны 2-8 дикого типа, индуцировать направленное воздействие на ген in vivo для корректировки мутированного гена F9 in situ в геноме гепатоцитов.
[0158] Для тестирования способности этих ZFN индуцировать DSB на конкретном целевом сайте используют ZFN пары, оказывающие направленное воздействие на человеческий F9 интрон 1. Проводят исследование Cel-I (Surveyor™, Transgenomics. Perez и другие (2008) Nat. Biotechnol. 26: 808-816 и Guschin и другие (2010) Methods Mol Biol. 649:247-56), где после PCR-амплификации целевого сайта следует количественная оценка введений и удалений (делеций) с использованием фермента, детектирующего несоответствия Cel-I (Yang и другие (2000) Biochemistry (2000) 39, 3533-3541), который обеспечивает оценку нижнего предела частоты DSB. Через три дня после трансфекции ZFN вектора экспрессии выделяют геномную ДНК из K562 клеток, используя набор DNeasy (Qiagen). Праймерами для Cel-I анализа hF9 интрона 1 являются N2 For (TCGGTGAGTGATTTGCTGAG, SEQ ID №:1) и N2 Rev (AACCTCTCACCTGGCCTCAT, SEQ ID NO:2). ZFN пара с наивысшей активностью, обозначенная N2, нацеленная на интрон 1 гена hF9, показана ниже в таблице 1 (см. также публикацию США № 20110027235):
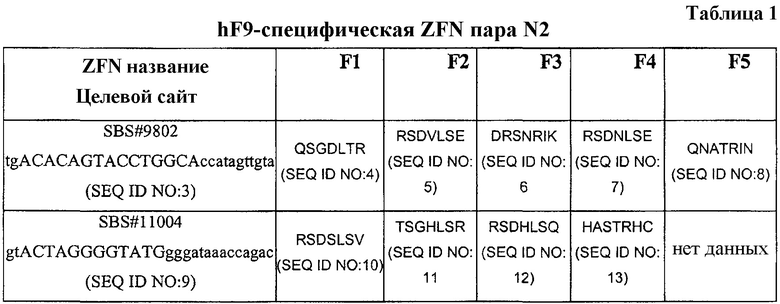
[0159] Экспрессионная плазмида N2 ZFN для трансфекции клеточной культуры сконструирована посредством введения 2A линкера от вируса Thosea asigna между кодирующими последовательностями для обеих ZFN и введения этой кассеты ниже CMV промотора. После трансфекции N2 ZFN пары в человеческие клетки K562 обнаружено, что расщепляется 30% N2 аллелей с целевыми сайтами (фигура 1С). Для тестирования способности N2 ZFN стимулировать гомологичную рекомбинацию посредством индуцирования DSB котрансфицируют N2 ZFN в клетки K562 посредством целенаправленного вектора, который вводит сайт рестрикции NheI (фигура 1D). Конструируют донорную плазмиду NheI посредством увеличения способом PCR левого и правого плеч гомологии из K562 геномной ДНК. Затем вводят короткую последовательность, содержащую сайт рестрикции NheI, между левым и правым плечом гомологии. Обнаружено, что в дни 3 и 10 после трансфекции 14% и 12% аллелей, соответственно, чувствительны к NheI поглощению (фигура 1E), что свидетельствует об эффективных степенях целенаправленной интеграции (TI) посредством гомологичной рекомбинации.
Пример 2: In vivo мышиная модель гемофилии человека
[0160] N2 целевой сайт присутствует в интроне 1 hF9, но отсутствует в мышином гене F9. Таким образом, для тестирования N2 ZFN in vivo создают гуманизированную модель мышиной гемофилии B (HB). Конструируют hF9 миниген (фигура 2А) под контролем печень-специфичного промотора (Shen и другие (1989) DNA 8:101-108 и Miao и другие (2000) Mol. Ther. 1:522-532). Праймерами для TI интрона 1 hF9 являются N2 TI For (GGCCTTATTTACACAAAAAGTCTG, SEQ ID NO:14) и N2 TI Rev (TTTGCTCTAACTCCTGTTATCCATC, SEQ ID NO:15).
[0161] Интрон 1 из этой конструкции содержит человеческий N2 целевой сайт (посадочную площадку, LP). Остальная часть последовательности F9 в конструкции LP симулирует идентифицированную ранее нонсенс-мутацию (Y155stop) (Thompson и другие (1989) Hum. Genet. 94: 299-302), в которой преждевременный терминирующий кодон перед экзонами, кодирующими каталитический домен FIX, дает в результате отсутствие циркулирующего белка FIX. Конструкцию LP получают генным синтезом (Genscript) и лигируют в плазмиду pUC57. Затем конструкцию LP удаляют посредством SwaI расщепления и лигируют в SwaI сайт собственной плазмиды между сайтами FLP рекомбиназы, пригодными для рекомбиназа-опосредованного кассетного обмена (RCME) (Taconic-Artemis) с целью создания плазмиды LP KI. Плазмиду LP KI и плазмиду, экспрессирующую FLP рекомбиназу (Taconic-Artemis), трансфицируют в эмбриональные стволовые (ES) клетки B6S6F1, содержащие сайты FLP рекомбиназы, пригодные для RCME на локусе ROSA26 (Zambrowicz и другие (1997) Proc Natl Acad Sci 94:3789-3794). Правильно меченые клоны клеток B6S6F1-LP ES идентифицируют посредством Саузерн-блоттинга и вводят в бластоцисты B6D2F1. Чистые ES клетки от B6S6F1-LP мышей (G0) получают при естественном рождении и выполняют возвратное скрещивание химерных детенышей с мышами C57BL/6J дикого типа (Jackson Laboratories): 5 поколений (in vivo эксперименты по расщеплению) или 7-10 поколений (in vivo эксперименты по TI).
[0162] LP мышей генотипируют, используя праймеры LP Oligo 1 (ACTGTCCTCTCATGCGTTGG, SEQ ID NO:16), LP Oligo 2 (GATGTTGGAGGTGGCATGG, SEQ ID NO:17), wtROSA Oligo 1 (CATGTCTTTAATCTACCTCGATGG, SEQ ID NO:18) и wtROSA Oligo2 (CTCCCTCGTGATCTGCAACTCC, SEQ ID NO:19) (фигура 2В). Также скрещивают LP мышей с существующей мышиной моделью, которая имеет делецию мышиного гена F9 (Lin и другие (1997) Blood 90, 3962-3966), для получения LP/HB мышей, у которых можно проверить N2 ZFN активность in vivo.
[0163] Проводят возвратное скрещивание HB мышей с мышами C57BL/6J (Jackson Laboratories), получая >10 поколений. Мышей C57Bl/6J (Jackson Laboratories) используют для LP-негативных TI-экспериментов. Как ожидалось, LP мыши не имеют детектируемого циркулирующего hFIX (рис. 2С). Количественное определение объема hFIX в плазме проводят, используя набор hFIX ELISA (Affinity Biologicals) со стандартной кривой, получаемой для объединенной нормальной человеческой плазмы (Trinity Biotech). Всем значениям ниже последнего значения для стандартной кривой (15 нг/мл) произвольно присвоено значение 15 нг/мл, что является пределом обнаружения. Плазму для hFDC ELISA получают посредством ретроорбитального кровотечения в гепаринизированные капиллярные пробирки.
Пример 3: целенаправленная доставка FIX-специфических ZFN in vivo
[0164] Для доставки N2 ZFN в печень, обычное место производства FIX, получают гепатотропный аденоассоциированный вирусный вектор, серотип 8 (AAV8-N2), экспрессирующий N2 ZFN из печень-специфического энхансера и промотора (Shen и другие, там же и Miao и другие, там же) (фигура 2D).
[0165] Для тестирования расщепляющей активности N2 ZFN in vivo делают инъекции в хвостовую вену LP мышей, используя 1e11 v.g. AAV8-N2 вектор экспрессии, и выделяют ДНК печени на 7 день после инъекции. PCR-амплификация LP и Cel-I исследования демонстрируют, что расщепляются 34-47% LP аллелей (фигура 2E). Праймерами для Cel-I конструкции LP являются LP N2 For (CTAGTAGCTGACAGTACC, SEQ ID NO:20) и LP N2 Rev (GAAGAACAGAAGCCTAATTATG, SEQ ID NO:21).
Пример 4: In vivo совместная доставка донора нуклеиновой кислоты и FIX-специфических ZFN
[0166] Введение экзонов дикого типа 2-8 перед акцепторным сайтом сплайсинга (SA) в интрон 1 конструкции LP допускает сплайсинг кодирующей последовательности дикого типа с экзоном 1 (фигура 3а). Для in situ коррекции мутировавшего гена F9 у LP мышей создают донорный матричный вектор AAV (AAV-донор) направленного воздействия на ген с плечами гомологии, фланкирующими кассету «SA - hF9 экзонов 2-8 дикого типа" (фигура 3а). Конструируют плазмиды, производящие донорный вектор, посредством увеличения методом PCR левого и правого плеч гомологии из LP мышиной геномной ДНК. Получают кассету «акцепторный сайт сплайсинга - кодирующая последовательность экзонов 2-8 - сигнал polyA гормона роста крупного рогатого скота» посредством PCR амплификации из плазмиды pAAV-hFIX16 (Manno и другие (2006) Nat. Med. 12:342-347) и лигируют между левым и правым плечами гомологии. Поскольку HR является преимущественной в течение S/G2 фаз клеточного цикла, N2 и донорные векторы доставляют новорожденным мышам, где быстро пролиферирующие гепатоциты вводят S/G2 в течение развития клеточного цикла.
[0167] На 2-й день жизни мышам LP/HB делают инъекцию либо только AAV-N2 (5e10 v.g.) (n=1), либо AAV-N2 (5e10 v.g.) + AAV-донор (2.5e11 v.g.) (n=5), либо AAV-Mock (5e10 v.g.) + AAV-донор (2.5e11 v.g.) (n=5). В Mock векторе N2 ZFN замещены люциферазой Renilla.
[0168] На 10 неделе жизни мышей умерщвляют и выделяют ДНК печени для исследования на TI донора с применением двух отдельных PCR стратегий. При первой стратегии используют праймеры, которые генерируют меньший ампликон для целевого LP аллеля и больший ампликон для нецелевого LP аллеля (фигура 3a, праймеры P1/P2). Вторая стратегия включает использование праймеров, которые генерируют больший ампликон для целевого LP аллеля и меньший ампликон для нецелевого LP аллеля (фигура 3a, праймеры P1/P3). Праймерами для TI конструкции LP являются P1 (ACGGTATCGATAAGCTTGATATCGAATTCTAG, SEQ ID NO:22), P2 (CACTGATCTCCATCAACATACTGC, SEQ ID №:23) и P3 (GAATAATTCTTTAGTTTTAGCAA, SEQ ID NO:24).
[0169] При проведении обоих PCR анализов обнаружено доказательство TI только у мышей, принимающих N2 + донор (фигура 3b). Количественная оценка интенсивностей полос предполагает частоту TI 1-7% (фигура 3b).
[0170] Для определения, происходит ли в результате геномной коррекции производство циркулирующего hFIX, вводят LP мышам на 2-й день жизни только AAV-N2 (n=7), AAV-Mock + AAV-донор (n=6) или AAV-N2 + AAV-донор (n=7) (такие же векторные дозы, как указано выше). Уровни hFIX в плазме мышей, принимающих только N2 или Mock+донор, в среднем ниже 15 нг/мл (нижний предел детектирования в исследовании), тогда как для мышей, принимающих N2+донор, уровень составляет в среднем 116-121 нг/мл (что соответствует 2-3% от нормального) (фигура 4a), значительно выше, чем для мышей, принимающих только N2 и Mock+донор (p≤0,006 во всех временных точках, 2-сторонний T-тест).
[0171] Для подтверждения стабильной геномной коррекции проводят частичные гепатэктомии (PHx), что вызывает разведение и потерю экстрахромосомных эписом, так как во время регенерации печени пролиферируют гепатоциты (Nakai и другие (2001), J. Virol. 75:6969-6976) (фигура 4 a, b). Частичные гепатэктомии выполняют, как описано выше (Mitchell и Willenbring, (2008) Nat. Prot. 3:1167-1170), и все процедуры на животных утверждены в Children's Hospital of Philadelphia IACUC. Измерение уровней hFIX у мышей, обработанных N2+донор, остаются неизменными после регенерации печени после гепатэктомии, показывая стойкую коррекцию. Контрольные мыши, принимающие только N2 или Mock+донор, сохраняют в среднем менее 15 нг/мл (фигура 4a) после гепатэктомии, значительно ниже, чем мыши, принимающие N2+донор (p≤0,004 во всех временных точках, 2-сторонний T-тест). Для гарантии LP-специфичной экспрессии hFIX, а не случайной интеграции донора в геном, мышам дикого типа, не имеющим LP трансгена, вводят на 2-й день жизни только AAV-N2 (n=8), AAV-Mock + AAV-донор (n=6) или AAV-N2 + AAV-донор (n=9) (такие же векторные дозы, как указано выше). Уровни hFIX в плазме мышей, принимающих только N2, Mock+доноров и N2+доноров, в среднем ниже 15, 19 и 27 нг/мл, соответственно, это указывает, что большая экспрессия hFIX у LP мышей, принимающих N2+донор, идет от LP-специфической коррекции (фигура 4c).
[0172] Для определения, достаточны ли уровни hFIX для коррекции HB фенотипа, вводят LP/HB мышам на 2-й день жизни только AAV-N2 (n=10), AAV-Mock + AAV-донор (n=9) или AAV-N2 + AAV-донор (n=9) (такие же векторные дозы, как указано выше). Уровни HFIX в плазме для мышей, принимающих только N2, снова в среднем ниже 15 нг/мл. Мыши, принимающие Mock+донор, имеют в среднем ниже 25 нг/мл, и мыши, принимающие N2+донор имеют значительно более высокие уровни hFIX (p≤0,04 во всех временных точках по сравнению с Mock+донор, 2-сторонний T-тест), в среднем 166-354 нг/мл (3-7% от нормального циркулирующего уровня) (фигура 4d). Печень-специфическую экспрессию hFIX подтверждают посредством RT-PCR для hFIX мРНК (фигура 4е). Выделяют РНК из замороженной ткани мышей, используя набор RNeasy (Qiagen) и набор без РНазы RNase-free DNase (Qiagen). Синтез кДНК выполняют, используя набор iSCRIPT (Bio-Rad). Выполняют RT-PCR для hFIX транскрипта, используя праймеры hFIX Gen1 For (ACCAGCAGTGCCATTTCCA, SEQ ID NO:25) и hFIX Gen1 Rev (GAATTGACCTGGTTTGGCATCT, SEQ ID NO:26).
[0173] Для исследования, корректируется ли HB фенотип, определяют активированное частичное тромбопластиновое время (aPTT), меру кинетики формирования фибринового сгустка, которое заметно пролонгировано при гемофилии. Анализ aPTT выполняют, смешивая образец плазмы 1:1:1 с объединенной HB человеческой плазмой и aPTT реагентом. Образование тромба инициируют, добавляя 25 мМ хлорида кальция. Плазму для aPTT получают при хвостовом кровотечении 9:1 в цитрат натрия. Средние aPTT для мышей дикого типа (n=5) и HB мышей (n=12) составляют 36 сек и 67 сек, соответственно (фигура 4f). Мыши, принимающие Mock+донор (n=3), имеют в среднем 60 сек, тогда как мыши, принимающие N2+донор (n=5) имеют значительно меньшее aPTT, в среднем 44 сек. (p=0,0014, по сравнению с МОК+донор, 2-сторонний T-тест) (фигура 4f).
Пример 5: in vivo совместная доставка генно-инженерных нуклеаз и донора у взрослых животных
[0174] Затем проводят редактирование генома на взрослых животных по человеческому FIX LP, как описано выше для новорожденных. Взрослых LP мышей в возрасте 6 недель обрабатывают посредством I.V.-инъекции только 1e11 v.g./мышь AAV-N2 ('только ZFN'), 1e11 v.g./мышь AAV-N2 и 5.5e11 v.g./мышь AAV-донор ('ZFN+донор') или 1e11 v.g./мышь AAV-Mock и 5.5e11 v.g. AAV-донор ('Mock+донор'). Представленные на фигуре 5А данные являются репрезентативными для 3 экспериментов с участием примерно 20 мышей на группу. В этих экспериментах уровни hFIX дикого типа составляют приблизительно 1000 нг/мл. Аналогично, на фигуре 5B представлен график, показывающий уровни hFIX в плазме взрослых LP мышей в возрасте 6 недель после I.V.-инъекции либо только 1е11 v.g./мышь AAV-N2 ('только ZFN'), либо 1e11 v.g./мышь AAV-N2 и 5.5e11 v.g./мышь AAV-донор ('ZFN+донор'), либо 1e11 v.g./мышь AAV-Mock и 5.5e11 v.g. AAV-донор ('Mock+донор'). Через два дня после инъекции животным из групп, указанных на фигуре 5B, производят частичную гепатэктомию. Представленные данные являются репрезентативными для 3 экспериментов с участием примерно 20 мышей на группу. В этих экспериментах уровни hFIX дикого типа составляют примерно 1000 нг/мл. Данные демонстрируют, что экспрессия hFIX стабильна при введении взрослым мышам с (или без) последующей частичной гепатэктомией, и что возможно выполнять редактирование генома у взрослых животных.
[0175] Все упоминаемые здесь патенты, патентные заявки и публикации настоящим включены в виде ссылок во всей своей полноте.
[0176] Хотя для ясности понимания раскрытие представлено с некоторыми подробностями посредством иллюстраций и примеров, специалистам в данной области очевидно, что на практике можно применять различные изменения и модификации, не отклоняясь от сущности или объема раскрытия. Таким образом, приведенные выше описания и примеры не следует рассматривать как ограничение.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2017 |
|
RU2788133C2 |
| МОДИФИКАЦИЯ B-КЛЕТОК | 2018 |
|
RU2783116C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2691102C2 |
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2020 |
|
RU2822369C2 |
| МОДУЛЯТОРЫ ТАУ И СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ИХ ДОСТАВКИ | 2017 |
|
RU2789459C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2014 |
|
RU2693891C1 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОПОСРЕДОВАННОЙ НУКЛЕАЗОЙ НАПРАВЛЕННОЙ ИНТЕГРАЦИИ ТРАНСГЕНОВ | 2013 |
|
RU2650819C2 |
| ПЕЧЕНЬ-СПЕЦИФИЧНЫЕ КОНСТРУКЦИИ, ЭКСПРЕССИОННЫЕ КАССЕТЫ ФАКТОРА VIII И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2774874C2 |







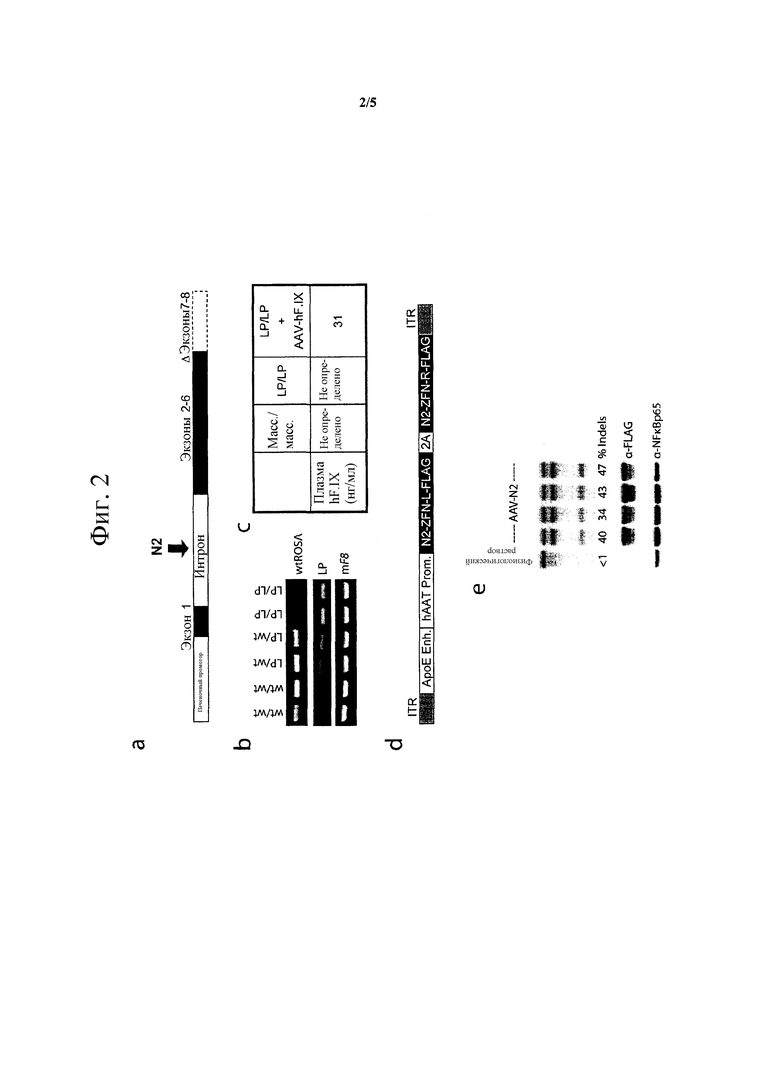

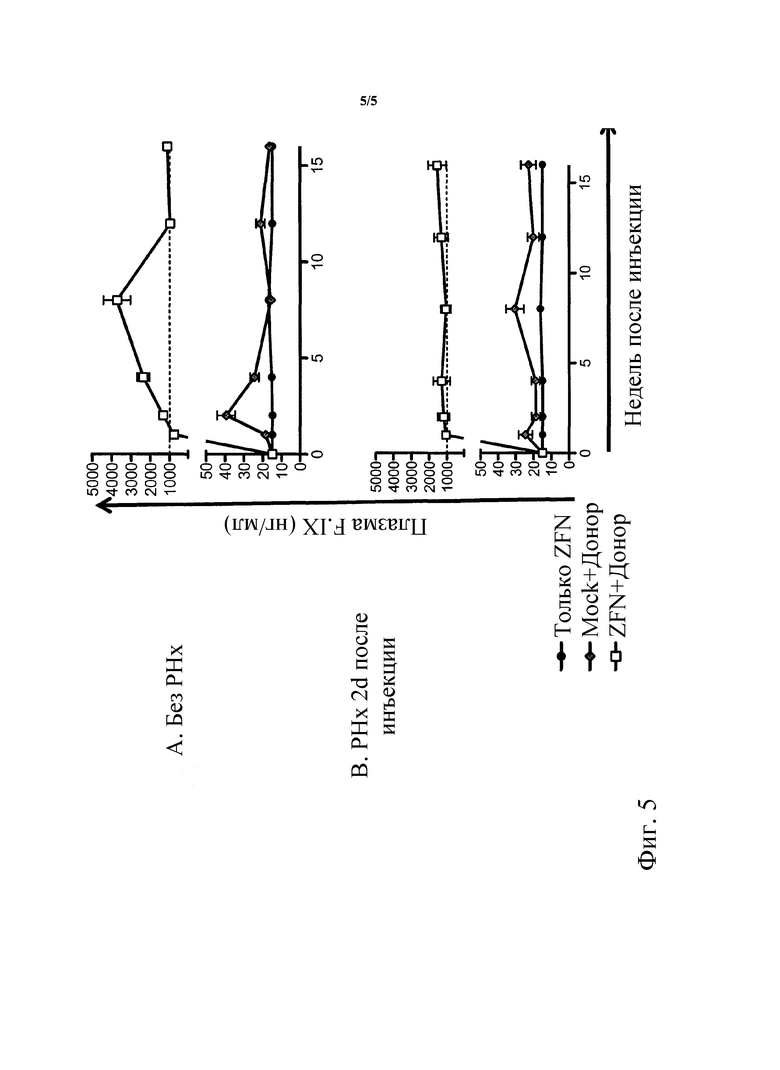
Описаны белок, который связывается с геном фактора IX (FIX), полинуклеотид, кодирующий такой белок, выделенная клетка-хозяин, экспрессирующая белок, и способ экспрессии белка. Представленный белок включает генно-инженерный ДНК-связывающий домен белка типа «цинкового пальца», где ДНК-связывающий домен содержит четыре или пять областей распознавания «цинковый палец». Представленные изобретения могут быть использованы в клинической практике у пациентов с гемофилией В. 4 н. и 11 з.п. ф-лы, 5 ил., 1 табл., 5 пр.
1. Белок, который связывается с геном фактора IX (FIX), где указанный белок включает генно-инженерный ДНК-связывающий домен белка типа «цинкового пальца», где ДНК-связывающий домен содержит четыре или пять областей распознавания «цинковый палец», обозначенных от F1 до F4 или от F1 до F5 от N-конца к С-концу, и где
(i) если ДНК-связывающий домен содержит пять областей распознавания «цинковый палец», то F1-F5 содержат следующие аминокислотные последовательности:
F1: QSGDLTR (SEQ ID NO: 4)
F2: RSDVLSE (SEQ ID NO: 5)
F3: DRSNRIK (SEQ ID NO: 6)
F4: RSDNLSE (SEQ ID NO: 7)
F5: QNATRIN (SEQ ID NO: 8)
(ii) если ДНК-связывающий домен содержит четыре области распознавания «цинковый палец», то F1-F4 содержат следующие аминокислотные последовательности:
F1: RSDSLSV (SEQ ID NO: 10)
F2: TSGHLSR (SEQ ID NO: 11)
F3: RSDHLSQ (SEQ ID NO: 12)
F4: HASTRHC (SEQ ID NO: 13)
2. Белок по п. 1, дополнительно содержащий расщепляющий домен или расщепляющий полудомен дикого типа или генно-инженерный.
3. Полинуклеотид, кодирующий белок по п. 1 или 2.
4. Выделенная клетка-хозяин, которая экспрессирует белок, который связывается с геном фактора IX, где указанная клетка содержит полинуклеотид по п. 3, которая экспрессирует белок по п. 1 или 2, где белок, экспрессированный в клетке-хозяине, модифицирует эндогенный FIX ген.
5. Способ экспрессии белка фактора IX у субъекта с гемофилией В, причем указанный способ включает интеграцию последовательности, кодирующей функциональный белок фактор IX (FIX), в геном клетки, используя по меньшей мере одну цинк-пальцевую нуклеазу, содержащую белок по п. 1 или 2 или кодируемую последовательностью, содержащей полинуклеотид по п. 3, где субъект содержит данную клетку.
6. Способ по п. 5, где последовательность интегрируют в эндогенный ген.
7. Способ по п. 5 или 6, где эндогенный ген выбран из группы, включающей FIX ген и ген «безопасную гавань».
8. Способ по п. 5, где клетка является печеночной клеткой и последовательность доставляют в клетку посредством внутривенного введения в печень интактного животного, внутрибрюшинного введения, прямой инъекции в паренхиму печени, инъекции в печеночную артерию или ретроградной инъекции через билиарное дерево.
9. Способ по п. 5, дополнительно включающий стадию проведения частичной гепатэктомии у субъекта и/или дополнительно включающий стадию лечения субъекта по меньшей мере одним вспомогательным агентом.
10. Способ по п. 9, где вспомогательный агент выбран из группы, включающей гамма-облучение, УФ-облучение, меченные тритием нуклеотиды, цис-платину, этопозид, гидроксимочевину, афидиколин, преднизолон, аденовирус и их комбинации.
11. Способ по п. 5, где клетка представляет собой выделенную клетку и способ дополнительно включает введение субъекту выделенной клетки.
12. Способ по п. 5, где субъект выбран из группы, включающей эмбрион, не являющийся человеческим, плод, не являющийся человеческим, новорожденного, инфантильного, ювенильного или взрослого субъекта.
13. Способ по п. 5, дополнительно включающий объединение последовательности, цинк-пальцевой нуклеазы, полинуклеотида, кодирующего цинк-пальцевую нуклеазу, или их комбинации с хоминг-агентом, который специфически связывается с поверхностным рецептором клетки.
14. Способ по п. 13, где хоминг-агент включает галактозу или гибрид белка оболочки AAV и галактозу.
15. Способ по любому из пп. 8-14, где клетка выбрана из группы, включающей человеческую клетку, клетку примата, отличного от человека, клетку грызуна, клетку зайцеобразного животного, клетку плотоядного животного и клетку парнокопытного животного и стволовую клетку, например нечеловеческую эмбриональную стволовую клетку, гемопоэтическую стволовую клетку, индуцированную плюрипотенциальную стволовую клетку, гепатоцит или печеночную стволовую клетку.
| US 0007732196 B2, 08.06.2010 | |||
| US 0007253273 B2, 07.08.2007 | |||
| WO 2009009086 A2, 15.01.2009 | |||
| KATHERINE A | |||
| HIGH, Update on Progress and Hurdles in Novel Genetic Therapies for Hemophilia, American Society of Hematology, 2007, pp.466-472 | |||
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ ПОЛИПЕПТИДОВ ФАКТОРА VII И ПОЛИПЕПТИДОВ ФАКТОРА IX | 2002 |
|
RU2292909C2 |

