Настоящее изобретение относится к способу получения непротеиногенных L-аминокислот путем ферментативной биотрансформации.
Непротеиногенные аминокислоты представляют собой аминокислоты, которые в естественных условиях не участвуют в качестве компонентов в биосинтезе протеинов и в этом существенно отличаются от 20 протеиногенных аминокислот. В контексте настоящего изобретения к непротеиногенным аминокислотам относят также очень редкую аминокислоту L-селеноцистеин, которая всегда присутствует в протеинах.
Непротеиногенные аминокислоты являются соединениями, которые, например, представляют интерес при получении действующих веществ в фармацевтической промышленности и сельском хозяйстве. В качестве действующего вещества или компонента действующего вещества благодаря молекулярному сходству они могут имитировать структуру аминокислот, встречающихся в естественных условиях, и модулировать таким путем, например, происходящие в естественных условиях реакции при взаимодействии с рецептором. Кроме того, такие аминокислоты, поскольку они представляют собой хиральные соединения, в целом могут служить в качестве компонентов синтеза в рамках "хирального пула".
Существующие в настоящее время методы получения непротеиногенных аминокислот в виде чистых энантиомеров преимущественно основаны на сложных в осуществлении методах синтеза, которые помимо этого позволяют получать только определенные типы соединений. Лишь некоторые из этих методов позволяют получать различные соединения за счет простой замены эдукта.
В большинстве случаев применяют методы химического синтеза, которые в свою очередь главным образом уже основаны на применении хиральных компонентов. Другие методы основаны на сочетании химического синтеза рацематов и разделения рацематов, которое часто осуществляют ферментативным путем.
Наряду с этим известен также ряд ферментативных методов, которые основаны на использовании прохиральных соединений и позволяют осуществлять стереоселективный синтез непротеиногенных аминокислот. Так, в частности, с помощью трансаминаз из α-кетокислот с использованием L-глутаминовой кислоты в качестве донора аминогруппы можно получать различные непротеиногенные аминокислоты (Taylor и др., TIBTECH 16: 412-418 (1998)). Другим примером является синтез L-трет.-лейцина с помощью лейцин-дегидрогеназы (Drauz, Chimia 51:310-314 (1997)).
О-ацетил-L-серин служит предшественником L-цистеина. Последнее соединение образуется в результате замещения ацетатной группы в β-положении тиольным остатком. Такая реакция, которую называют β-замещением, катализируется ферментом из класса О-ацетил-L-серинсульфгидрилаз (КФ 4.2.99.8). При добавлении в процессе ферментации нуклеофильных соединений, относящихся к определенным классам веществ (тиолы, азолы или изоксазолиноны), они вступают в реакцию β-замещения, в результате чего образуются непротеиногенные L-аминокислоты. При этом структура соответствующего фрагмента образующихся аминокислот определяется добавляемым нуклеофильным соединением.
Проблема, возникающая при осуществлении этого метода, состоит в том, что добавляемые нуклеофильные соединения нельзя применять в слишком высоких количествах, поскольку в противном случае либо само соединение, либо образующаяся аминокислота оказывают токсическое действие на метаболизм, осуществляемый с помощью микроорганизмов. Это относится прежде всего к многочисленным тиольным соединениям, поскольку они в качестве соединений с высокой окислительно-восстановительной активностью в высоких концентрациях характеризуются токсическим действием. Кроме того, применение тиольных соединений в ферментативных методах связано с особыми проблемами, поскольку при интенсивной аэрации ферментера они проявляют тенденцию к окислению, что без принятия специальных технических мер приводит к значительной эмиссии зловонных веществ. Добавление азида или цианида, которые, как известно, вступают в реакцию β-замещения в присутствии О-ацетил-L-серинсульфгидрилаз (Flint и др., J. Biol. Chem., 271: 16053-16067 (1996)), также не представляется возможным вследствие их высокой токсичности.
Исходя из вышеизложенного, в основу настоящего изобретения была положена задача разработать способ получения непротеиногенных L-аминокислот, который допускал бы добавление больших количеств нуклеофильных соединений, в том числе, в частности, и токсичных соединений.
Указанная задача решается с помощью способа ферментативной биотрансформации, заключающегося в том, что О-ацетил-L-серин и нуклеофильное соединение при катализе O-ацетил-L-серинсульфгидрилазой превращают в непротеиногенную L-аминокислоту, и отличающегося тем, что взаимодействие проводят при значении рН в диапазоне от 5,0 до 7,4.
Такой способ позволяет осуществлять синтез многочисленных непротеиногенных энантиомерно чистых и отчасти новых L-аминокислот в промышленном масштабе.
О-ацетил-L-серинсульфгидрилазы являются известными соединениями. До настоящего времени их выделяли из различных видов растений и микроорганизмов. Наиболее изученными являются соответствующие ферменты из бактерий вида Salmonella typhimurium. В этом организме присутствуют два фермента из класса О-ацетил-L-серинсульфгидрилаз, которые обозначают как OASS-A и OASS-B соответственно. Известны также соответствующие гены, которые обозначены как cysK и cysM. Хотя оба фермента обладают исключительно сходным механизмом действия, их аминокислотные последовательности идентичны только на 45%.
Известно, что OASS-B (CysM) в отличие от OASS-A (CysK) обладает способностью катализировать взаимодействие О-ацетил-L-серина с тиосульфатом с образованием S-сульфоцистеина. Эта реакция имеет важное значение для роста бактерий при использовании тиосульфата в качестве единственного источника серы.
Сравнительный анализ последовательностей генов О-ацетил-L-серинсульфгидрилазы из различных организмов свидетельствует также о существовании двух филогенетических групп (Kitabatake и др., J. Bacteriol., 182:143-145 (2000)). CysK из Salmonella typhimurium и Escherichia coli вместе с O-ацетил-L-серинсульфгидрилазами из других микроорганизмов, относящихся к Eubacteria, метаногенным Archaea, и из растений образуют одну большую группу. В отличие от этого CysM из Salmonella typhimurium и Escherichia coli относятся к очень небольшой родственной группе, в которую входят О-ацетил-L-серинсульфгидрилазы из гипертермофильных видов Archaea (например, Pyrococcus, Sulfblobus, Thermoplasma).
Согласно настоящему изобретению О-ацетил-L-серинсульфгидрилазы отличаются тем, что они обладают способностью катализировать синтез L-цистеина из О-ацетил-L-серина и сульфида. В контексте настоящего изобретения ферменты, родственные как CysM, так и CysK, относятся поэтому к О-ацетил-L-серинсульфгидрилазам.
Несмотря на наличие отдельных публикаций, в которых говорится, что O-ацетил-L-серинсульфгидрилазы из CysK-группы характеризуются относительно широким спектром субстратов (Ikegami и Murakoshi, Phytochemistry, 35: 1089-1104 (1994); Flint и др., J. Biol. Chem., 271:16053-16067 (1996)), возможность технического применения О-ацетил-L-серинсульфгидрилаз для производства непротеиногенных аминокислот до настоящего времени не рассматривалась.
До настоящего времени основной причиной, препятствующей технической реализации ферментативного метода получения непротеиногенных аминокислот с использованием О-ацетил-L-серинсульфгидрилаз, являлась нестабильность O-ацетил-L-серина именно в том диапазоне значений рН, который соответствует интервалу, в котором проявляется активность О-ацетил-L-серинсульфгидрилаз.
В зависимости от значения рН О-ацетил-L-серин подвергается реакции изомеризации с образованием N-ацетил-L-серина. Эта реакция является необратимой и при этом протекает с высокой скоростью, например при значении рН 7,6 со скоростью 1%/мин. При понижении значения рН скорость реакции уменьшается, и поэтому, например, при рН 4,0 соединение является стабильным. Механизм такой реакции заключается во внутримолекулярном захвате нуклеофильной группой депротонированной аминогруппы на атоме углерода карбонильной группы ацильного фрагмента (Tai и др., Biochemistry, 34: 12311-12322 (1995)).
Оптимальное значение рН для О-ацетил-L-серинсульфгидрилаз составляет приблизительно 8,0 (Ikegami и Murakoshi, Phytochemistry, 35: 1089-1104 (1994); Tai и др., Biochemistry, 34: 12311-12322 (1995)), т.е. находится в диапазоне, который является весьма неблагоприятным с точки зрения изомеризации O-ацетил-L-серина.
Поэтому у Ikegami и Murakoshi был предложен метод биомиметического синтеза непротеиногенных аминокислот с использованием О-ацетил-L-серина, нуклеофила, пиридоксалфосфата и ионов металла (предпочтительно Ga2+). Эту реакцию можно осуществлять в диапазоне значений рН 3,5-5,5, обеспечивая тем самым стабильность О-ацетил-L-серина. Однако эффективность этого метода является не очень высокой при максимальном выходе продукта менее 45%. Существенный недостаток при этом по сравнению с ферментативным синтезом состоит в отсутствии энантиоселективности.
Еще одной задачей, на решение которой направлен предлагаемый в изобретении способ, состояла в разработке метода ферментативного получения чистых энантиомеров непротеиногенных аминокислот, который несмотря на невозможность одновременного достижения стабильности О-ацетил-L-серина и оптимальной ферментативной активности обеспечивал бы эффективное ферментативное превращение (>>45%) при умеренной степени изомеризации.
Эту задачу позволяет решить предлагаемый в изобретении способ преимущественно благодаря тому, что
- реакцию осуществляют при значении рН, которое лежит ниже оптимального значения рН для О-ацетил-L-серинсульфгидрилаз, и
- фермент применяют в достаточно высоких концентрациях.
Предпочтительным диапазоном значений рН для получения непротеиногенных аминокислот согласно настоящему изобретению является интервал от 5,0 до 7,4.
Особенно предпочтительным является диапазон значений рН от 6,0 до 7,1.
Наиболее предпочтительным является диапазон значений рН от 6,0 до 6,99.
Значение рН предпочтительно поддерживать на постоянном уровне с помощью активного рН-контроля с целью воспрепятствовать образованию стехиометрического количества уксусной кислоты. Активный рН-контроль предпочтительно осуществлять с помощью измерительного и регулировочного устройства, которое при отклонении значения рН от заданного значения позволяет восстановить требуемое значение рН путем дозированного добавления щелочи или кислоты.
Известные в настоящее время из литературы реакции синтеза непротеиногенных аминокислот с использованием О-ацетил-L-серинсульфгидрилаз в отличие от предлагаемого в изобретении способа осуществляют при значении рН в диапазоне его оптимальных значений для O-ацетил-L-серинсульфгидрилаз без активного регулирования рН и только в масштабе, пригодном для аналитических целей. Кроме того, применяют только фермент из филогенетической группы CysK-ферментов.
Изобретение относится также к способу получения непротеиногенной L-аминокислоты путем катализируемого О-ацетил-L-серинсульфгидрилазой взаимодействия О-ацетил-L-серина с нуклеофильным соединением, отличающимся тем, что в качестве О-ацетил-L-серинсульфгидрилазы используют CysM.
Применение фермента в достаточно высокой концентрации позволяет обеспечивать достаточно высокую степень превращения в ходе ферментативной реакции даже при значениях рН, которые не являются оптимальными. Такую концентрацию фермента предпочтительно использовать при осуществлении способа в том случае, когда объемная активность О-ацетил-L-серинсульфгидрилазы ACys в реакционной смеси составляет по меньшей мере 2 ед./мл. Наиболее предпочтительно активность составляет 2-200 ед./мл. Активность определяют с помощью теста, описанного в примере 3.
При создании изобретения было установлено, что для получения непротеиногенных аминокислот в качестве предпочтительного фермента можно применять также О-ацетил-L-серинсульфгидрилазу из филогенетической группы CysM-ферментов. Спектр нуклеофильных соединений, которые можно использовать в качестве субстрата, является неожиданно даже еще более широким, чем для CysK-ферментов.
Согласно наиболее предпочтительному варианту осуществления изобретения ферментативную реакцию проводят в непрерывном режиме. Для этого в ходе реакции постоянно добавляют О-ацетил-L-серин, О-ацетил-L-серинсульфгидрилазу и нуклеофильное соединение и одновременно из реакционной смеси отбирают раствор, содержащий непротеиногенные L-аминокислоты (раствор, содержащий продукт). Последнюю операцию предпочтительно осуществлять таким образом, чтобы объем реакционной смеси оставался постоянным. Особое преимущество такого подхода состоит в том, что при этом достигается динамическое равновесие, и поэтому О-ацетил-L-серин присутствует в реакционной смеси в постоянно низкой концентрации, и в результате этого минимизируется изомеризация. Концентрацию О-ацетил-L-серина в реакционной смеси предпочтительно поддерживать на уровне менее 1,0 г/л. Это значение можно регулировать варьированием средней продолжительности пребывания раствора в реакционной смеси. С целью обеспечить достаточно высокую стабильность необходимо, чтобы в процессе непрерывного превращения О-ацетил-L-серин находился в условиях, соответствующих кислым значениям рН, предпочтительно при рН 4-5.
До настоящего времени О-ацетил-L-серин получали только путем химического синтеза ацетилированием L-серина, что является дорогим в осуществлении методом вследствие высокой стоимости L-серина. В заявке DE 10107002, поданной 15 февраля 2001 г., описан способ ферментативного получения О-ацетил-L-серина. Несмотря на то, что в этом случае можно использовать экономичную технологическую систему, выделение продукта из ферментационного бульона связано с определенными проблемами из-за нестабильности О-ацетил-L-серина.
Преимущество настоящего изобретения заключается в том, что содержащий О-ацетил-L-серин ферментационный бульон, который получают, например, путем ферментации согласно DE 10107002, можно использовать в способе по настоящему изобретению непосредственно в качестве источника О-ацетил-L-серина. Такой подход является исключительно экономичным и не требует выделения нестабильного соединения.
Получение О-ацетил-L-серинсульфгидрилаз для синтеза непротеиногенных аминокислот осуществляют предпочтительно с помощью обычных методов рекомбинантной ДНК, которые хорошо известны специалистам в данной области.
Для этой цели ген, кодирующий О-ацетил-L-серинсульфгидрилазу, клонируют в пригодном векторе и затем им трансформируют соответствующий штамм-хозяин. В качестве штамма-хозяина можно использовать любой микроорганизм, который можно подвергать обработке с помощью методов рекомбинантной ДНК и который пригоден для ферментативного получения рекомбинантных протеинов.
Предпочтительным микроорганизмом для получения О-ацетил-L-серинсульфгидрилаз является Escherichia coli.
В принципе можно также осуществлять интеграцию рекомбинантного гена О-ацетил-L-серинсульфгидрилазы в хромосому, а также применять самореплицирующийся плазмидный вектор.
Для клонирования предпочтительно использовать векторы, которые уже содержат генетические элементы (например, регулируемые промоторы, терминаторы), позволяющие осуществлять контролируемую, обладающую высокой индуцибельностью экспрессию гена О-ацетил-L-серинсульфгидрилазы. Наиболее предпочтительными являются плазмидные векторы, несущие большое количество копий, такие, например, как основанные на Escherichia coli векторы pUC18, pBR322, pACYC184 и их производные. В качестве обладающих высокой индуцибельностью промоторов можно применять, например, lac-, tac-, trc-, lambda PL-, ara- или tet-промоторы.
О-ацетил-L-серинсульфгидрилазы получают, например, ферментативным путем культивированием рекомбинантного штамма микроорганизма. Для этой цели применяют известные специалистам методы культивирования, при этом параметры технологического процесса необходимо согласовывать для каждого конкретного штамма микроорганизма. В качестве питательных сред можно применять как полные среды, так и минимальные среды. Процесс можно проводить как в периодическом, так и в полунепрерывном режиме. При использовании индуцибельных промоторных систем в необходимый момент времени вызывают экспрессию гена О-ацетил-L-серинсульфгидрилазы путем добавления соответствующего индуктора. После получения достаточного количества продукта клетки, содержащие О-ацетил-L-серинсульфгидрилазу, собирают известными методами (например, центрифугированием).
Полученный таким путем фермент О-ацетил-L-серинсульфгидрилазу можно выделять обычными методами, которые используются для очистки протеинов. Для этого можно использовать как классические методы (например, преципитацию, ионообменную хроматографию, хроматографию на основе гидрофобного взаимодействия, изоэлектрическое фокусирование), так и современную аффинную хроматографию с использованием "аффинных меток". Последовательности, которые кодируют такую аффинную метку, могут быть слиты с кодирующей областью уже при клонировании гена, в результате чего образуются слитые протеины, несущие соответствующую аффинную метку. Затем их выделяют путем одностадийной очистки. В качестве примера пригодной для этой цели комбинации аффинной метки и метода выделения на основе аффинности является последовательность StrepTag, которая используется для аффинной хроматографии, основанной на применении стрептавидина, и производится фирмой IBA, Геттинген, Германия.
Согласно изобретению О-ацетил-L-серинсульфгидрилазу можно применять в качестве добавки в очищенной форме. Наряду с проведением реакции в растворе фермент можно также иммобилизовать на носителе. Соответствующие методы известны из уровня техники.
Однако для получения непротеиногенных аминокислот выделение O-ацетил-L-серинсульфгидрилазы не является абсолютно необходимым. В соответствии с предлагаемым в изобретении способом существует также возможность использовать непосредственно клетки микроорганизмов, обладающих активностью О-ацетил-L-серинсульфгидрилазы. Примерами таких штаммов микроорганизмов являются штаммы Escherichia coli DH5α/pFL145 и BLR21(DE3)/pLE4. Эти штаммы описаны в примерах 1 и 2 и депонированы в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ) в Брауншвейге под регистрационными номерами DSM 14088, соответственно DSM 14089.
В этом варианте осуществления предлагаемого в изобретении способа O-ацетил-L-серин подвергают биотрансформации в непротеиногенную L-аминокислоту с использованием иммобилизованных клеток. При этом происходит проникновение О-ацетил-L-серина и нуклеофильного соединения в клетки, а также высвобождение клетками продукта реакции, т.е. непротеиногенной L-аминокислоты.
При необходимости можно интенсифицировать обмен веществ между внутренней областью клеток и реакционной средой за счет обработки клеток веществами, которые повышают проницаемость клеточной мембраны. Такими веществами являются, например, хлороформ или толуол, и их применение известно специалистам в данной области.
В предпочтительном варианте непротеиногенную аминокислоту получают в результате взаимодействия О-ацетил-L-серина с нуклеофильным соединением при катализе О-ацетил-L-серинсульфгидрилазой, при этом О-ацетил-L-серинсульфгидрилаза присутствует внутри клетки микроорганизма.
Наиболее предпочтительным является получение непротеиногенных L-аминокислот в результате взаимодействия полученного ферментативным путем неочищенного О-ацетил-L-серина с нуклеофильным соединением при катализе О-ацетил-L-серинсульфгидрилазой, при этом О-ацетил-L-серинсульфгидрилаза присутствует внутри клетки микроорганизма.
В качестве нуклеофильных соединений, которые можно применять при осуществлении предлагаемого в изобретении способа для катализируемого O-ацетил-L-серинсульфгидрилазами β-замещения, предпочтительно использовать соединения, которые содержат фрагмент, выбранный из группы, включающей
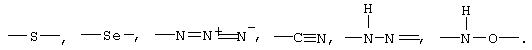
Особенно предпочтительно в качестве компонента реакционной смеси использовать нуклеофильное соединение, выбранное из группы, включающей следующие соединения:
- тиосульфаты,
- тиолы общей формулы (1):

где R1 обозначает одновалентную замещенную или незамещенную алкильную, алкокси-, арильную или гетероарильную группу, содержащую 1-15 атомов углерода,
- селениды,
- селенолы общей формулы (2)

где R1 имеет значения, указанные для формулы (1),
- азиды,
- цианиды,
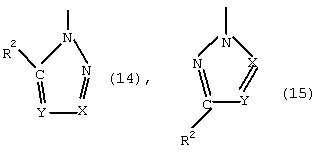
- азолы общей формулы (3) или (4)

в которой Х и Y могут иметь одинаковые или различные значения и обозначают CR4 или N, где R4 означает -Н, -СООН, -ОН, -NH2, -NO2, -SH, -SO
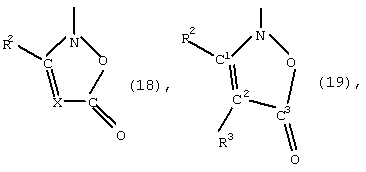
- изоксазолиноны общей формулы (5) или (6)

в которой X, R1, R2 и R3 имеют указанные выше значения,
при этом С1 и С2 в формуле (6) вместо заместителей R2 и R3 могут быть присоединены к кольцу с помощью мостика, указанного для формулы (4).
Примерами тиосульфатов являются тиосульфат натрия, тиосульфат калия или тиосульфат аммония.
В качестве примеров тиолов можно назвать соединения, выбранные из группы, включающей 2-меркаптоэтанол, 3-меркаптопропанол, 3-меркаптопропионовую кислоту, 3-меркапто-1-пропансульфоновую кислоту, меркаптоэтансульфоновую кислоту, 2-меркаптоэтиламин, тиогликолевую кислоту, тиомолочную кислоту, тиоуксусную кислоту, меркаптоянтарную кислоту, меркаптопировиноградную кислоту, дитиотреитол, дитиоэритритол, 1-тиоглицерин, тиофенол, 4-фтортиофенол, 4-хлортиофенол, 4-меркаптофенол, пара-тиокрезол, 5-тио-2-нитробензойную кислоту, 2-меркаптотиазол, 2-меркаптотиазолин, 2-меркаптоимидазол, 3-меркапто-1,2,4-триазол, 2-тиофентиол, 2-меркаптопиридин, 2-меркаптопиримидин, 2-тиоцитозин, 2-меркаптоникотиновую кислоту, 2-меркапто-1-метилимидазол, 2-меркаптобензотиазол, 2-меркаптобензоксазол, 6-меркаптопурин.
Примерами селенолов являются соединения, выбранные из группы, включающей метилселенол, этилселенол, пропилселенол, фенилселенол.
Примерами азидов являются азид натрия, азид калия или азид аммония.
Примерами цианидов являются цианид калия, цианид натрия или цианид аммония.
В качестве примеров азолов можно назвать соединения, выбранные из группы, включающей 1,2-пиразол, 3-метилпиразол, 4-метилпиразол, 3,5-диметилпиразол, 3-аминопиразол, 4-аминопиразол, пиразол-4-карбоновую кислоту, пиразол-3,5-дикарбоновую кислоту, 1,2,3-триазол, 1,2,4-триазол, 3-амино-1,2,4-триазол, 1,2,3,4-тетразол, индазол, индазол-3-карбоновую кислоту, индазол-5-карбоновую кислоту, 5-аминоиндазол, бензотриазол, бензотриазол-5-карбоновую кислоту, 5-аминобензотриазол, аминопиразолопиримидин, 8-азагуанин, 8-азааденин.
Примерами изоксазолинонов являются соединения, выбранные из группы, включающей изоксазолин-5-он, 3-метилизоксазолин-5-он, 4-метилизоксазолин-5-он, 4,5-диметилизоксазолин-2-он, 1,2,4-оксадиазолидин-3,5-дион.
Концентрацию нуклеофильного соединения в реакционной смеси предпочтительно выбирают таким образом, чтобы она была эквимолярна концентрации О-ацетил-L-серина.
Температуру реакционной смеси предпочтительно выбирают в диапазоне от 5 до 70°С. Наиболее предпочтительный диапазон температур составляет от 20 до 40°С.
В качестве растворителя для осуществления реакции предпочтительно используют воду.
В качестве продуктов реакции предпочтительно образуются L-аминокислоты общей формулы (7), имеющие L-конфигурацию

где Z обозначает одновалентный остаток, выбранный из группы формул (8)-(19)
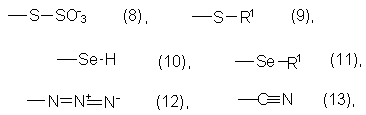


а также их простые эфиры, сложные эфиры или соли и
R1, R2, R3, R4, Х и Y имеют значения, указанные выше для формул (1)-(6).
По завершении предлагаемого в изобретении процесса и после разделения по известным методам предпочтительно водного раствора на биомассу и на супернатант культуры из этого супернатанта предпочтительно выделяют растворимые непротеиногенные L-аминокислоты. Такие методы выделения аминокислот также известны специалистам в данной области. К этим методам относятся, например, фильтрация, центрифугирование, экстракция, адсорбция, ионообменная хроматография, преципитация, кристаллизация. При наличии малорастворимых непротеиногенных аминокислот предпочтительно проводить сортировочное (классификационное) центрифугирование, как это известно в данной области, при этом биомасса по возможности в полном объеме сохраняется в супернатанте, получаемом путем центрифугирования. Отделенный продукт предпочтительно растворяют и переосаждают с помощью традиционных методов.
Ниже изобретение более подробно проиллюстрировано на примерах.
Пример 1: Клонирование гена cysK, соответственно гена cysM из Escherichia coli в виде гена, слитого с StrepTag-последовательностью
Для клонирования гена cysK, соответственно гена cysM из Escherichia coli сначала проводили полимеразную цепную реакцию в присутствии ДНК-полимеразы Pwo с использованием защищенных с помощью фосфортиоата следующих пар олигонуклеотидных праймеров:

При этом в последовательности праймеров встраивали сайты рестрикции, распознаваемые рестриктазой BsaI (подчеркнутые нуклеотиды). В качестве матрицы использовали хромосомную ДНК штамма Е. coli W3110 (АТТС 27325). Продукт, полученный в результате амплификации, расщепляли с помощью рестриктазы BsaI и затем клонировали в расщепленном с помощью Есо3 II векторе pASK-IBA3 (Institut  Bioanalytik, Геттинген, Германия). Этот вектор позволяет клонировать слитую генную конструкцию с расположенной на 3'-конце последовательностью, которая кодирует определяющий аффинность пептид (WSHPQFEK, SEQ ID NO: 5) и которую обозначили как StrepTagII. Экспрессия слитой генной конструкции, осуществляемая с помощью tet-промотора, приводит к получению протеина, на С-конце которого находится фрагмент (метка) StrepTagII. Последний можно применять для выделения протеина, поскольку StrepTagII обеспечивает связывание с колонками, заполненными стрептавидином.
Bioanalytik, Геттинген, Германия). Этот вектор позволяет клонировать слитую генную конструкцию с расположенной на 3'-конце последовательностью, которая кодирует определяющий аффинность пептид (WSHPQFEK, SEQ ID NO: 5) и которую обозначили как StrepTagII. Экспрессия слитой генной конструкции, осуществляемая с помощью tet-промотора, приводит к получению протеина, на С-конце которого находится фрагмент (метка) StrepTagII. Последний можно применять для выделения протеина, поскольку StrepTagII обеспечивает связывание с колонками, заполненными стрептавидином.
Пример 2: Очистка протеина CysK, соответственно CysM
С использованием конструкции cysM-StrepTag (pFL145) можно обеспечить исключительно эффективную экспрессию гена в штамме Е. coli DH5α (фирма Clontech, Гейдельберг, Германия). При этом культивирование и экспрессию осуществляли согласно инструкциям поставщика Institut  Bioanalytik, Геттинген, Германия. Штамм DH5α/pFL145 в соответствии с Будапештским договором депонирован в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ) под регистрационным номером DSMZ 14088. При использовании же конструкции cysK-StrepTag (pLEl) уровень экспрессии был очень низким Однако после повторного клонирования слитой генной конструкции в виде XbaI-HmdIII-фрагмента в векторе с использованием промотора фага Т7 (pRSET5a) (фирма Stratagene, Гейдельберг, Германия) и экспрессии в штамме BLR21 (DE3) (фирма Novagen, Дармштадт, Германия) оказалось возможным и в этом случае (pLE4) добиться высокого выхода генного продукта. Штамм BLR21 (DE3)/pLE4 в соответствии с Будапештским договором депонирован в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ) под регистрационным номером DSMZ 14089 Протеины CysK, соответственно CysM выделяли с помощью аффинной хроматографии на колонках со стрептавидином, в основном следуя рекомендациям поставщика Institut
Bioanalytik, Геттинген, Германия. Штамм DH5α/pFL145 в соответствии с Будапештским договором депонирован в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ) под регистрационным номером DSMZ 14088. При использовании же конструкции cysK-StrepTag (pLEl) уровень экспрессии был очень низким Однако после повторного клонирования слитой генной конструкции в виде XbaI-HmdIII-фрагмента в векторе с использованием промотора фага Т7 (pRSET5a) (фирма Stratagene, Гейдельберг, Германия) и экспрессии в штамме BLR21 (DE3) (фирма Novagen, Дармштадт, Германия) оказалось возможным и в этом случае (pLE4) добиться высокого выхода генного продукта. Штамм BLR21 (DE3)/pLE4 в соответствии с Будапештским договором депонирован в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ) под регистрационным номером DSMZ 14089 Протеины CysK, соответственно CysM выделяли с помощью аффинной хроматографии на колонках со стрептавидином, в основном следуя рекомендациям поставщика Institut  Bioanalytik, Геттинген, Германия. При использовании 250 мл культуры выход составил 3 мг очищенного протеина CysK, соответственно 4,5 мг очищенного протеина CysM. Оба протеина из-за присутствия кофактора пиридоксалфосфата имели выраженную желтую окраску, и для них был характерен максимум абсорбции при 420 нм.
Bioanalytik, Геттинген, Германия. При использовании 250 мл культуры выход составил 3 мг очищенного протеина CysK, соответственно 4,5 мг очищенного протеина CysM. Оба протеина из-за присутствия кофактора пиридоксалфосфата имели выраженную желтую окраску, и для них был характерен максимум абсорбции при 420 нм.
Пример 3: Определение ферментативной активности
Активность О-ацетил-L-серинсульфгидрилазы определяли путем выявления "естественного" продукта реакции L-цистеина (Cys) или, в случае CysM, также продукта, представляющего собой S-сульфо-L-цистеин (S-Cys). Присутствие обоих продуктов можно выявить с помощью обладающего высокой специфичностью и высокой чувствительностью теста, описанного у Gaitonde, Biochem. J., 104: 627-633(1967) Для обоих соединений необходимо иметь отдельные калибровочные кривые, поскольку окраска комплекса, содержащего сульфоцистеин, имеет более слабую интенсивность.
Типичная смесь для тестирования содержала 10 мМ O-ацетилсерин (полученный из 200 мМ маточного раствора добавлением в 500 мМ натрийсукцинатный буфер, рН 5,5), 10 мМ сульфид натрия или тиосульфат натрия, 100 мМ калий-фосфатный буфер, рН 7,0, 5 мкг/мл очищенного фермента, и эту смесь инкубировали при 37°С. Удельную активность А выражали в количестве единиц/мг протеина, т.е. в следующем виде количество мкмолей продукта × мин-1 × мг-1 протеина. ACys обозначает удельную активность образования цистеина при использовании сульфида. AS-Cys обозначает удельную активность образования S-сульфоцистеина при использовании тиосульфата.
Протеины, выделенные по методу, описанному в примере 2, имели следующие значения удельной активности:
Cys К: ACys 140 ед./мг,
Cys M: ACys 199 ед./мг,
Cys M: AS-Cys 145 ед./мг.
Для определения активности O-ацетилсеринсульфгидрилаз в иммобилизованных клетках сначала приготавливали клеточную суспензию с оптической плотностью (ОП) 20,0 (измерения проводили при длине волны 600 нм). Затем увеличивали проницаемость клеточных мембран добавлением хлороформа до 10 об.% и инкубировали в течение 5 мин при комнатной температуре. После этого для проведения теста по определению ферментативной активности оптическую плотность доводили до 1,0 и по описанному выше методу проводили реакцию с использованием клеток и О-ацетил-L-серина и сульфида, соответственно тиосульфата. При этом удельную активность А клеток выражали в мкмолях × мин-1 × (мл клеточной суспензии с оптической плотностью 1,0, измеренной при 600 нм)-1 или в сокращенном виде: кол-во ед./мл ОП.
Пример 4: Исследование каталитических свойств О-ацетил-L-серинсульфгидрилаз CysK, соответственно CysM
Для получения с помощью О-ацетил-L-серинсульфгидрилаз непротеиногенных аминокислот вместо сульфида и тиосульфата в реакции применяли другие нуклеофильные соединения. Для их выявления осуществляли предварительную дериватизацию аминокислот на колонке с помощью ортофталдиальдегида. Такой метод позволяет осуществлять широкомасштабный скрининг пригодных нуклеофильных субстратов.
Образцы реакционной смеси содержали 4 мМ О-ацетил-L-серин (полученный из 200 мМ маточного раствора путем добавления в 500 мМ натрийсукцинатный буфер, рН 5,5), 20 мМ нуклеофил, 100 мМ калийфосфатный буфер, рН 7,0, 5 мкг/мл очищенного фермента. Реакцию прекращали добавлением 1/100 объема уксусной кислоты (96 об.%) и продукт анализировали с помощью ЖХВР с использованием колонки типа HP Aminoquant (200×2,1 мм), следуя инструкциям фирмы-производителя Hewlett Packard, Вальдбронн, Германия.
Для определения характеристик полученного продукта оценивали его молекулярную массу с помощью ЖХВР-МС. Для этой цели использовали колонку Luna-CIS (фирма Phenomenex, Ашафенбург, Германия), используя в качестве подвижной фазы муравьиную кислоту (0,1 об.%). Ионизацию осуществляли в положительном режиме пучком электронов.
В приведенной ниже таблице представлены показатели реакционной способности обоих ферментов CysK и CysM при использовании различных нуклеофилов, а также значения массы, определенные с помощью ЖХВР- и МС-анализа (МН+).

Пример 5: Ферментативное получение О-ацетил-L-серинсульфгидрилазы CysM
В качестве предварительной культуры для ферментации использовали 20 мл LB-среды (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl), которая дополнительно содержала 100 мг/л ампициллина и которую инфицировали штаммом DH5α/pFL145 (см. выше) и инкубировали в течение ночи в шейкере при 30°С и 150 об/мин. Затем всю смесь переносили в 100 мл SMl-среды (12 г/л К2НРO4, 3 г/л КН2РO4, 5 г/л (NH4)2SO4, 0,3 г/л MgSO4·7H2O, 0,015 г/л CaCl2·2H2O, 0,002 г/л FеSO4·7Н2О, 1 г/л Nа3-цитрата·2Н2О, 0,1 г/л NaCl, 1 мл/л раствора микроэлементов, содержащего 0,15 г/л Na2MoO4·2H2O, 2,5 г/л Na3BO3, 0,7 г/л CoCl2·6H2O, 0,25 г/л CuSO4·5H2O, 1,6 г/л MnCl2·4H2O, 0,3 г/л ZnSO4·7H2O), дополненной 5 г/л глюкозы, 0,5 мг/л витамина B1 и 100 мг/л ампициллина. Последующую инкубацию предварительной культуры проводили в течение 8 ч при 30°С и 150 об/мин.
В качестве ферментера использовали оборудование Biostat M фирмы Braun Biotech (Мельзунген, Германия) с максимальной вместимостью для культуры 2 л. Описанную выше предварительную культуру (оптическая плотность при 600 нм приблизительно 2) загружали в ферментер, содержащий 900 мл среды для ферментации (15 г/л глюкозы, 10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л (NH4)2SO4, 1,5 г/л КН2РO4, 0,5 г/л NaCl, 0,3 г/л MgSO4·7H2O, 0,015 г/л CaCl2·2H2O, 0,075 г/л FeSO4·7H2O, 1 г/л Na3-цитрата·2H2O и 1 мл/л описанного выше раствора микроэлементов, 5 мг/л витамина B1 и 100 мг/л ампициллина, значение рН доводили до 7,0 с помощью 25%-ного раствора аммиака). В процессе ферментации температуру поддерживали на уровне 32°С, а значение рН поддерживали на постоянном уровне 7,0 добавлением 25 об.%-ного раствора аммиака. Через культуру пропускали стерильный сжатый воздух с расходом 1,5 об./об/мин и перемешивали при скорости вращения мешалки 200 об/мин. При снижении насыщения кислородом до 50% скорость вращения увеличивали с помощью контрольного прибора до 1200 об/мин для поддержания степени насыщения кислородом на уровне 50% (что определяли с помощью рO2-зонда, откалиброванного на значение насыщения, равное 100%, при 900 об/мин). Как только содержание глюкозы в ферментере снижалось с исходного значения, составлявшего 15 г/л, приблизительно до 5-10 г/л, добавляли 56% (маc./об.) раствора глюкозы. Подпитку осуществляли с расходом 3-6 мл/ч, при этом концентрацию глюкозы в ферментере поддерживали на уровне 0,5-10 г/л. Концентрацию глюкозы определяли с помощью анализатора глюкозы фирмы YSI (Йеллоу Спрингс, шт. Огайо, США). По достижении оптической плотностью величины, равной 30, индуцировали продуцирование фермента добавлением 3 мг/л тетрациклина. Продолжительность индукции составляла 7 ч. По истечении указанного промежутка времени отбирали пробы, клетки отделяли от культуральной среды путем центрифугирования и промывали. Полученную суспензию клеток анализировали по методу, описанному в примере 3. При использовании тиосульфата активность AS-Cys O-ацетилсеринсульфгидрилазы составляла 12 ед./мл ОП.
Клетки собирали центрифугированием, быстро замораживали с помощью жидкого азота и хранили при -20°С.
Пример 6: Ферментативное получение О-ацетил-L-серинсульфгидрилазы CysK
Фермент CysK в целом получали аналогично методу, описанному в примере 5. Однако в данном случае применяли штамм BLR21(DE3)/pLE4. После израсходования глюкозы, предварительно внесенной в концентрации 15 г/л, осуществляли подпитку с помощью 60 об.%-ного раствора глицерина. Расход составлял 9,5 мл/ч. Одновременно с этим индуцировали продуцирование фермента путем добавления 0,4 мМ изопропил-β-тиогалактозида (ИПТГ). По завершении цикла индукции, длившегося 21 ч, отбирали пробы, клетки отделяли от культуральной среды путем центрифугирования и промывали. Полученную клеточную суспензию анализировали аналогично примеру 3. При использовании сульфида активность ACys O-ацетилсеринсульфгидрилазы составила 2 ед./мл ОП.
Клетки собирали центрифугированием, быстро замораживали с помощью жидкого азота и хранили при -20°С.
Пример 7: Ферментативное получение S-фенил-L-цистеина с использованием иммобилизованных клеток
Сначала в термостате, снабженном рН-зондом и регулятором рН, в 100 мл воды растворяли гидрохлорид О-ацетил-L-серина (фирма Sigma, Дейзенхофен, Германия) до получения конечной концентрации О-ацетил-L-серина, равной 200 мМ (29,4 г/л). Затем при перемешивании добавляли 200 мМ тиофенол. После этого значение рН реакционной смеси постепенно доводили до 6,7 путем титрования с помощью 5М NaOH и сразу же смешивали с клеточной суспензией штамма-продуцента CysM DH5α/pFL145. Клеточная суспензия обладала удельной активностью AS-Cys, равной 12 ед./мл ОП. Оптическая плотность клеток (измеренная при 600 нм) в реакционной смеси составляла 2,0 и, следовательно, активность AS-Cys О-ацетил-L-серинсульфгидрилазы составляла 24 ед./мл. В процессе реакции значение рН поддерживали с помощью системы регулирования значения рН на постоянном уровне добавлением 5М NaOH.
В процессе реакции наблюдалось образование осадка белого цвета. Через 30 мин отбирали пробы и смешивали с 1%-ной (по объему) уксусной кислотой. Осадок отделяли центрифугированием, растворяли в равном объеме 21%-ной (по объему) фосфорной кислоты и анализировали с помощью ЖХВР. Надосадочную жидкость, полученную при центрифугировании, также подвергали хроматографии. Анализ проводили с помощью ЖХВР с обращенной фазой на колонке типа LUNA 5 μ С 18(2) (фирма Phenomenex, Ашаффенбург, Германия). В качестве элюента использовали разбавленную фосфорную кислоту (0,1 мл конц. фосфорной кислоты/л) при скорости потока 0,5 мл/мин.
Содержание S-фенил-L-цистеина в надосадочной жидкости составляло 1,6 г/л, а в осадке - 24,8 г/л. В пересчете на О-ацетил-L-серин это соответствовало 67%-ному общему выходу продукта реакции. Реакцию также проводили с использованием уменьшенного на 12, соответственно на 2 ед./мл количества O-ацетил-L-серинсульфгидрилазы. Это приводило к более замедленной ферментативной реакции и вследствие изомеризации О-ацетил-L-серина - к уменьшенному выходу продукта, который составил 52%, соответственно 15%.
Пример 8: Ферментативное получение S-фенил-L-цистеина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В данном случае опыт проводили с использованием содержащего O-ацетил-L-серин ферментационного бульона вместо чистого О-ацетил-L-серина. Его получали путем ферментации по методу, описанному в заявке DE 10107002. В конце ферментации для стабилизации О-ацетил-L-серина значение рН доводили до 4,0 с помощью 21%-ной (по объему) фосфорной кислоты. Клетки отделяли центрифугированием и концентрацию О-ацетил-L-серина в ферментационном бульоне доводили путем упаривания до 30 г/л. Затем к 100 мл бульона добавляли 200 мМ тиофенол и после титрования до достижения значения рН 6,7 добавляли клетки штамма-продуцента CysM DH5α/pFL145. Активность AS-Cys составляла 24 ед./мл. Дальнейшую обработку проводили аналогично примеру 7. Выход в пересчете на О-ацетил-L-серин составил 65%.
Для идентичной реакционной смеси, содержащей клетки штамма-продуцента CysK BLR21(DE3)/pLE4, при величине ACys, равной 38 ед./мл, выход составил 72%.
Пример 9: Непрерывное ферментативное получение S-фенил-L-цистеина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
Целью проведения реакции являлось достижение максимально высокой степени превращения дорогостоящего и нестабильного эдукта О-ацетил-L-серина. С этой целью была разработана непрерывная технология производства. Для этого сначала, как описано в примере 8, величину активности AS-Cys CysM повышали до 48 ед./мл. По истечении 20 мин реакции с помощью дозаторов добавляли содержащий О-ацетил-L-серин ферментационный бульон (30 г/л), тиофенол и суспензию клеток штамма-продуцента CysM DH5α/pFL145 (величина ОП составляет 240 при 600 нм, в калий-фосфатном буфере с рН 7,0). Указанные компоненты подавали с расходом 150; 2,5 и 3 мл/ч соответственно. Одновременно с этим из реакционного сосуда начинали непрерывно отбирать раствор, содержащий продукт, с расходом 155,5 мл/ч. Расход подбирали с таким расчетом, чтобы объем реакционной смеси в реакторе оставался постоянным, а среднее время пребывания в реакторе составляло 30 мин. При проведении процесса в таком режиме в реакционной смеси устанавливалось динамическое равновесие при постоянной концентрации О-ацетил-L-серина, равной 1,0 г/л. Благодаря такой низкой концентрации, при которой фермент вследствие своего низкого показателя КМ еще обладал достаточно высокой активностью, оказалось возможным успешно минимизировать изомеризацию до N-ацетил-L-серина. Выход продукта реакции в пересчете на О-ацетил-L-серин составил 85%.
Пример 10: Ферментативное получение азидо-L-аланина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ азид натрия. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 72 ед./мл, выявленная степень превращения составляла 45%.
Пример 11: Ферментативное получение циано-L-аланина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ цианид калия. Этот цианид добавляли после доведения значения рН реакционной смеси до 6,7. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 72 ед./мл, выявленная степень превращения составила 65%.
Пример 12: Ферментативное получение S-сульфо-L-цистеина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ тиосульфат натрия. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 16 ед./мл, выявленная степень превращения составила 91%.
Пример 13: Ферментативное получение S-тиазол-2-ил-L-цистеина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ 2-меркаптотиазол. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 72 ед./мл, выявленная степень превращения составила 62%.
Пример 14: Ферментативное получение S-1,2,4-триазол-3-ил-L-цистеина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ 3-меркапто-1,2,4-триазол. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 72 ед./мл, выявленная степень превращения составила 74%.
Пример 15: Ферментативное получение селено-L-цистеина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ селенид натрия. Селенид натрия получали восстановлением селенита натрия с помощью борогидрида натрия. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 72 ед./мл, выявленная степень превращения составила 75%.
Пример 16: Ферментативное получение фенилселено-L-цистеина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ фенилселенол. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 72 ед./мл, выявленная степень превращения составила 62%.
Пример 17: Ферментативное получение 1,2,4-триазол-1-ил-L-аланина с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ 1,2,4-триазол. Для этой реакции проницаемость клеточных мембран повышали обработкой 10 об.% хлороформа. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 72 ед./мл, выявленная степень превращения составила 48%.
Пример 18: Ферментативное получение 5-карбокси-1,2,3-бензотриазол-2-ил-L-аланина с использованием ферментационного бульона, содержащего O-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ 5-карбокси-1,2,3-бензотриазол. Для этой реакции проницаемость клеточных мембран повышали обработкой 10 об.% хлороформа. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 72 ед./мл, выявленная степень превращения составила 54%.
Пример 19: Ферментативное получение 1,2,4-оксадиазолидин-3,5-дионил-L-аланина (хисквалевая кислота) с использованием ферментационного бульона, содержащего О-ацетил-L-серин, и иммобилизованных клеток
В этом примере реакцию проводили в основном аналогично примеру 8 с использованием клеток, содержащих CysM. Вместо тиофенола применяли 200 мМ 1,2,4-оксадиазолидин-2,5-дион. Для этой реакции проницаемость клеточных мембран повышали обработкой 10 об.% хлороформа. По истечении 30 мин реакции с использованием в реакционной смеси клеток, активность которых составляла 72 ед./мл, выявленная степень превращения составила 11%.


| название | год | авторы | номер документа |
|---|---|---|---|
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ ESCHERICHIA, - ПРОДУЦЕНТ L-ЦИСТЕИНА И СПОСОБ ПОЛУЧЕНИЯ L-ЦИСТЕИНА | 2003 |
|
RU2275425C2 |
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ СЕРУСОДЕРЖАЩИХ АМИНОКИСЛОТ | 2012 |
|
RU2651511C2 |
| МУТАНТНЫЕ О-ФОСФОСЕРИНСУЛЬФГИДРИЛАЗЫ И СПОСОБ ПОЛУЧЕНИЯ ЦИСТЕИНА С ИХ ПРИМЕНЕНИЕМ | 2011 |
|
RU2541782C2 |
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ АМИНОКИСЛОТ И ПРОИЗВОДНЫХ АМИНОКИСЛОТ ИЗ СЕМЕЙСТВА ФОСФОГЛИЦЕРАТОВ | 2003 |
|
RU2346038C2 |
| РЕКОМБИНАНТНЫЕ МИКРООРГАНИЗМЫ, ПРОДУЦИРУЮЩИЕ МЕТИОНИН | 2006 |
|
RU2447146C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ ИЗ ИХ РАЦЕМИЧЕСКИХ N-АЦЕТИЛ-D, L-ПРОИЗВОДНЫХ ПОСРЕДСТВОМ ФЕРМЕНТАТИВНОГО РАСЩЕПЛЕНИЯ | 2000 |
|
RU2270869C2 |
| МУТАНТНАЯ СЕРИНАЦЕТИЛТРАНСФЕРАЗА, ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ МУТАНТНУЮ СЕРИНАЦЕТИЛТРАНСФЕРАЗУ (ВАРИАНТЫ), БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ ESCHERICHIA, - ПРОДУЦЕНТ L-ЦИСТЕИНА, И СПОСОБ ПРОДУКЦИИ L-ЦИСТЕИНА | 2003 |
|
RU2279477C2 |
| СПОСОБ ПОЛУЧЕНИЯ МЕТИОНИНА | 2004 |
|
RU2376378C2 |
| Штамм бактерий RноDососсUS RноDоснRоUS - продуцент нитрилгидратазы | 1990 |
|
SU1731814A1 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ОРГАНИЧЕСКИХ СОЕДИНЕНИЙ С ИСПОЛЬЗОВАНИЕМ ДЕКСТРИНСОДЕРЖАЩИХ СРЕД | 2006 |
|
RU2429296C9 |
Изобретение относится к биотехнологии. Способ предусматривает ферментативную биотрансформацию путем катализируемого О-ацетил-L-серинсульфгидрилазой взаимодействия О-ацетил-L-серина с нуклеофильным соединением. Используют О-ацетил-L-серинсульфгидрилазу, обладающую активностью Acys, по меньшей мере, 2 ед/мл. Взаимодействия осуществляют при рН 5,0-7,4. Способ позволяет осуществить синтез непротеиногенных чистых L-аминокислот в промышленном масштабе. 4 с. и 13 з.п. ф-лы, 1 табл.








в которой R1 обозначает одновалентную замещенную или незамещенную алкильную, алкокси-, арильную или гетероарильную группу, содержащую 1-15 атомов углерода,
селениды, селенолы общей формулы (2)

в которой R1 имеет значения, указанные для формулы (1),
азиды, цианиды, азолы общей формулы (3) или (4)


в которой Х и Y могут иметь одинаковые или различные значения и обозначают CR4 или N, где R4 представляет собой –H, –СООН, –ОН, –NН2, –NO
изоксазолиноны общей формулы (5) или (6)
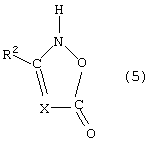
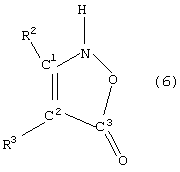
в которой Х, R1, R2 и R3 имеют указанные выше значения, при этом С1 и С2 в формуле (6) вместо заместителей R2 и R3 могут быть присоединены к кольцу с помощью мостика, указанного для формулы (4).

где Z обозначает одновалентный фрагмент одной из формул (8)-(19)

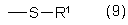
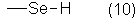




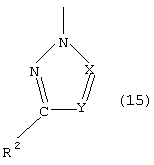




а также их сложные эфиры, простые эфиры и соли и
R1, R2, R3, R4, Х и Y имеют значения, указанные для формул (1)-(6).
| SAITO et al | |||
| Production of plant non-protein amino aciols by recombinant enzymes of sequential biosynthetic reactions in bacteria, Biologicaland pharmaceuntical bulletin, Tokyo | |||
| JP, 20, №1, Januar 1997, 47-53 | |||
| RU 97117473 A, 10.10.1999. |

