Предпосылки изобретения
Настоящая заявка относится к способам и композицям для лечения атеросклероза и других сердечно-сосудистых и воспалительных заболеваний.
Адгезия лейкоцитов с эндотелием является причиной различных воспалительных заболеваний, включая атеросклероз, аутоиммунные заболевания и бактериальные и вирусные инфекции. Рекрутинг лейкоцитов эндотелием начинается, когда индуцирующиеся адгезией молекулы рецепторов на поверхности эндотелиальных клеток взаимодействуют с контррецептором на иммунных клетках. Эндотелиальные клетки сосуда определяют, какой тип лейкоцитов (моноциты, лимфоциты или нейтрофилы) обновляется под действием факторов экспрессии специфической адгезии, таких как фактор васкулярной клеточной адгезии 1 (VCAM-1), фактор внутриклеточной адгезии 1 (ICAM-1) и Е-селектин. На самой ранней стадии атеросклеротического повреждения происходит локализованная эндотелиальная экспрессия VСАМ-1 и избирательный рекрутинг моноядерных лейкоцитов, которые экспрессируют контррецептор интегрина VLA-4. Вследствие избирательной экспрессии VLA-4 на моноцитах и лимфоцитах, но не на нейтрофилах, VCAM-1 играет важную роль в опосредовании избирательной адгезии моноядерных лейкоцитов. Последующее превращение лейкоцитов в пенистые макрофаги приводит к синтезу широкого разнообразия воспалительных цитокинов, факторов роста и хемоаттрактантов, которые способствуют увеличению лейкоцитов и рекрутингу, пролиферации гладкомышечных клеток, активации эндотелиальных клеток и синтезу характерного межклеточного матрикса атеросклеротических бляшек.
VCAM-1 экспрессируется в культивируемых васкулярных эндотелиальных клетках человека после активации под действием липополисахарида (LPS) и цитокинов, таких как интерлейкин-1 (IL-1) и фактор некроза опухолей (TNF-α).
Данные факторы не являются избирательными в активации экспресирующего фактора клеточной адгезии. Фиг.1 иллюстрирует процесс активации цитокином активации генной экспрессии VCAM-1 в васкулярных эндотелиальных клетках.
На фиг.1 регуляторные схемы активации цитокином экспрессии гена фактора васкулярной клеточной адгезии-1, опосредованного чувствительными к восстановлению-окислению факторами, такими как NF-kВ, в васкулярных эндотелиальных клетках (IkB является ингибирующей субъединицей; NF-kB является ядерным фактором кВ; NН3 относится к аминоконцу белка и РНК Pоl II является РНК полимеразой П).
1. Цитокин связывается со своим рецептором.
2. Активированный рецептор генерирует внутриклеточный сигнал.
3. Активация транскрипционного регуляторного белка.
4. Трансляция на ядро.
5. Связывание с энхансерным элементом специфической ДНК на промоторе VCAM-1.
6. Активация транскрипции РНК.
7. Процессинг РНК.
8. Трансляция в белок.
9. Пост-трансляционный процессинг.
10. Инсерция VCAM-1 в мембрану.
Молекулярный анализ регуляторных элементов гена VСАМ-1 человека, которые контролируют его экспрессию, подтверждают важную роль ядерного фактора-кВ (NF-kВ), фактора регуляции транскрипции, или подобного NF-kВ связывающего белка в чувствительной к окислению-восстановлению регуляции экспрессии гена VCAM-1. Факторы транскрипции являются белками, которые активируют (или подавляют) экспрессию гена в ядре клетки путем связывания со специфической последовательностью ДНК, называемой "энхансерными элементами", которые расположены обычно рядом с областью гена, называемой "промотером", с которого начинается синтез РНК. Ядерный фактор-кВ является повсеместно экспрессируемым мультисубъединичным фактором транскрипции, активируемым некоторыми типами клеток при помощи большой и различающейся группы агентов воспаления, таких как TNF-α, IL-1β, бактериальный эндотоксин и РНК вирусы. Он играет ключевую роль в опосредовании воспаления и других сигналов стресса между регуляторным аппаратом ядра. Хотя непосредственные биохимические сигналы, которые активируют NF-kB, неизвестны, данный фактор транскрипции может включаться в общий молекулярный путь многих факторов риска и "причинных" сигналов атеросклероза, таких как гиперлипидимия, курение, гипертония и сахарный диабет.
Важно, что активация NF-kB в эндотелиальных клетках сосуда под действием различных сигналов может быть избирательно ингибирована под действием антиоксидантов, таких как N-ацетилцистеин и пирролидин дитиокарбамат (см. USS № 07/969934, ныне разрешенный). Это приводит к гипотезе, что радикалы кислорода играют важную роль в активации NF-kB посредством неидентифицированного окислительно-восстановительного механизма. Поскольку NF-kB-подобный энхансерный элемент также регулирует транскрипцию промотера VCAM-1 чувствительным к окислению-восстановлению образом, окислительный стресс при атеросклеротическом поражении может играть роль в регуляции гена экспрессии VCAM-1 посредством данного чувствительного к окислению-восстановлению регуляторного белка транскрипции.
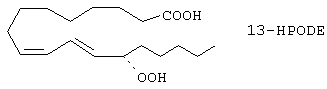
Была высказана гипотеза, что модификация липопротеина низкой плотности (ЛНП-LDL) в окисленный модифицированный ЛНП (ox-LDL) при помощи активного кислорода является основным в инициировании и распространении атеросклероза. Steinberg, et al., N. Engl. J. Med., 1989; 320-915-924. Окисленный ЛНП представляет сложную структуру, состоящую, по крайней мере, из нескольких химических различных окисленных материалов, каждый из которых, сам по себе или в сочетании, может модулировать активированную цитокином экспрессию гена молекулы адгезии. Гидропероксиды жирных кислот, таких как линолеил гидропероксид (13-НРОDЕ), получают из свободных жирных кислот под действием липоксигеназ, и они являются важным компонентом окисленного ЛНП.
Было предложено, что генерация окисленных липидов осуществляется под действием системы липоксигеназ клетки и что окисленные липиды последовательно переносятся в ЛНП. Это представляет, тем самым, реакцию распространения по ЛНП в среде, катализируемую переходными металлами и/или сульфгидрильными соединениями. Предшествующие исследования показали, что модификация жирной кислоты культивируемых эндотелиальных клеток может изменить их восприимчивость к воздействию окислителя. Добавление насыщенных или мононенасыщенных жирных кислот к культивируемым эндотелиальным клеткам понижает их восприимчивость к воздействию окислителя, тогда как добавление полиненасыщенных жирных кислот (PUFA) ее увеличивает.
Используя анализ ЖХВР с обращенной фазой природных и омыленных экстрактов липидов, было показано, что 13-HPODЕ является доминирующей окисленной жирной кислотой в липидах, окисленных под действием активированных моноцитов человека. Длительное выдерживание с окисленным ЛНП проводит окислительный сигнал к сосудистой эндотелиальной клетке, возможно посредством специфической гидроперекиси жирной кислоты, которая избирательно усиливает индуцированную цитокином экспрессию гена VCAM-1.
Области сосудистой стенки, предрасположенной к атеросклерозу, предпочтительно захватывает циркулирующие ЛНП посредством механизма, который не достаточно хорошо определен. Затем, посредством плохо понятного пути, эндотелий, гладкая мышца и/или воспалительные клетки превращают ЛНП в окисленный ЛНП. В отличие от ЛНП, которые захватываются посредством рецептора ЛНП, моноциты жадно захватывают окисленные ЛНП посредством рецептора-мусорщика, чья экспрессия, в отличие от рецептора ЛНП, не ингибируется при увеличении содержания внутриклеточного липида. Таким образом, моноциты продолжают захватывать окисленные ЛНП и становиться переполненными липидами, пенистыми макрофагальными клетками, которые образуют жировую прослойку.
Поскольку сердечно-сосудистые заболевания в настоящее время являются главной причиной смертности в Соединенных Штатах и девяносто процентов сердечно-сосудистых заболеваний представляет собой атеросклероз, существует необходимость в разработке новых способов и фармацевтических агентов для его лечения. В достижении этого важным является идентификация и управление специфическими окисленными биологическими соединениями, которые действуют в качестве избирательных регуляторов экспрессии медиаторов воспалительного процесса, в частности VCAM-1. Более общей целью является идентификация избирательных методов для подавления экспрессии чувствительных к окислению-восстановлению генов или активация подавленных, чувствительных к окислению-восстановлению генов.
Таким образом, предмет настоящего изобретения описывает лечение атеросклероза и других сердечно-сосудистых и воспалительных заболеваний.
Другой предмет настоящего изобретения описывает способ для избирательного ингибирования VСАМ-1.
Другой предмет настоящего изобретения еще описывает способ лечения заболевания человека или нарушения, которое опосредовано путем экспрессии или подавления генов, чувствительных к окислительно-восстановительному потенциалу.
Другой предмет настоящего изобретения описывает фармацевтические композиции для лечения атеросклероза и других сердечно-сосудистых и воспалительных заболеваний.
Сущность изобретения
Было обнаружено, что полиненасыщенные жирные кислоты (РUFАS) и их гидропероксиды (ox-PUFAS), которые являются важными компонентами окислительно модифицированных липопротеинов низкой плотности (ЛНП), индуцируют экспрессию VCAM-1, а не фактор внутриклеточной адгезии-1 (ICAM-1) или Е-селектин в эндотелиальных клетках аорты человека, посредством механизма, который не опосредован цитокинами или другими моноцитокинными сигналами. Это является фундаментальным открытием важного, ранее не известного биологического пути, в опосредованном VCAM-1 иммунном ответе.
В качестве примеров, линолевая кислота, линоленовая кислота, арахидоновая кислота, линолеил гидропероксид (13-НРОDЕ) и арахидоновый гидропероксид (15-НРЕТЕ) вызывают экспрессию на клеточной поверхности гена VCAM-1, но не ICAM-1 или Е-селектина. Ненасыщенные жирные кислоты (такие как стеариновая кислота) и ненасыщенные жирные кислоты (такие как олеиновая кислота) не вызывают экспрессию VCAM-1, ICAM-1 или Е-селектина.
Индукция VCAM-1 под действием РUFAS и их гидроперекисей жирных кислот подавляется антиоксидантом пирролидин дитиокарбаматом (PDTC). Это показывает, что индукция опосредована оксиленной сигнальной молекулой и что индукция предотвращается, когда окисление молекулы заблокировано (т.е. окисление не наблюдается), обращено (т.е. сигнальная молекула восстановлена) или когда модифицированный окислительно-восстановительный сигнал так или иначе заблокирован для взаимодействия со своей регуляторной мишенью.
Клетки, длительное время выдерживаемые с повышенным уровнем полиненасыщенных жирных кислот или их окисленных производных могут инициировать иммунный ответ, который не является нормальным, и которые, вне части, находящейся под имеющейся угрозой, приводят к болезни. Например, сверхчувствительность эндотелиальных клеток сосуда к PUFAS и ox-PUFАS может ускорять образование атеросклеротических бляшек.
Основываясь на этих открытиях, описывается способ для лечения атеросклероза, постангиопластического рестеноза, заболеваний коронарных артерий, грудной жабы и других сердечно-сосудистых заболеваний, также как и не сердечно-сосудистых воспалительных заболеваний, которые опосредованы VCAM-1, включающий удаление, снижение концентрации или предотвращение образования окисленных полиненасыщенных жирных кислот, включая, но не ограничиваясь, окисленную линолевую (C18, Δ9, 12), линоленовую (C18, Δ6, 9, 12), арахидоновую (C20, Δ5, 8, 11, 14) и эйкозотриеноевую (С20, Δ8, 11, 14) кислоты.
Неограничивающие примеры не сердечно-сосудистых воспалительных заболеваний, которые опосредуются VCAM-1, включают ревматоидный и остеоартрит, астму, дерматит и сложный склероз.
Данный способ имеет явное преимущество при лечении сердечно-сосудистого заболевания путем предотвращения развития новых поражений и регрессии имеющихся поражений и, при правильном применении, помимо текущей терапии, предназначенной только для ингибирования прогресирования заболевания, обеспечивает возможность медицинского "лечения" атеросклероза.
В альтернативном воплощении описывается способ для подавления экспрессии чувствительного к окислению-восстановлению гена или активирование гена, который супрессирован посредством чувствительного к окислению-восстановлению пути, который включает введение эффективного количества вещества, которое предотвращает окисление окисляемого сигнала, обычно окисление полиненасыщенной жирной кислоты. Характерные, чувствительные к окислению-восстановлению гены, которые участвуют в проявлении иммунного ответа, включают, но не ограничиваются, те экспрессируемые цитокины, участвующие в инициировании иммунного ответа (например, 1L-1β), хемоаттрактанты, которые стимулируют миграцию клеток воспаления к точке повреждения (например, МСР-1), Факторы роста (например, IL-6 и рецептор тромбина) и факторы адгезии (например, VCAM-1 и Е-селектин).
Картина заболеваний, опосредованных VCAM-1 или геном, чувствительным к окислению-восстановлению, предусматривает также включение суррогатных маркеров заболевания. В одном воплощении уровень окисленных полиненасыщенных жирных кислот или других подходящих маркеров в ткани или крови хозяина определяется, например, как среднее из оценки "окислительного окружения" хозяина и восприимчивости хозяина к VCAM-1 или гену, чувствительному к окислению-восстановлению, опосредующих заболевание.
В другом воплощении подсчитывается уровень VCAM-1, циркулирующего или на поверхности клетки, или другого подходящего маркера, и влияние на этот уровень введения подходящего антиоксиданта.
С помощью другого анализа оценивается чувствительность эндотелиальных клеток сосуда хозяина к полиненасыщенным жирным кислотам или их окисленным производным. Это может быть выполнено, например, путем индуктирования хозяина РUFА или ох-РUFА и сравнения конечной концентрации циркулирующего VCAM-1 или на поверхности клетки или других суррогатных маркеров с нормальной популяцией.
В другом воплощении на модели атеросклероза in vivo или других сердечных или воспалительных заболеваний, которые опосредуются VCAM-1, для индукции заболевания может быть предусмотрено введение животному-хозяину избыточного количества PUFA или окисленных полиненасыщенных жирных кислот. Данные животные могут быть использованы в клинических исследованиях для дальнейшего анализа этих заболеваний.
В другом воплощении изобретения соединения могут быть оценены еще по их способности лечить заболевания, опосредованные VCAM-1, на основании их способности ингибировать окисление полиненасыщенных жирных кислот, или по взаимодействию РUFA или ox-PUFA с белковой мишенью.
Это может быть выполнено путем стимулирования хозяина, например человека или животного, такого как мышь, высоким уровнем PUFA или ox-PUFA, и затем определения терапевтической эффективности исследуемого соединения на основании его способности снижать концентрацию циркулирующего VCAM-1 или на поверхности клетки. Альтернативно может быть использован отбор in vitro, который основывается на способности исследуемого соединения предотвращать окисление PUFA или взаимодействие PUFA или ох-РUFА с белковой мишенью в присутствии окисляющего соединения, например меди или фермента, такого как пероксидаза, липоксигеназа, циклооксигеназа или цитохром Р450.
В другом воплощении эндотелиальные клетки сосуда выдерживают подходящее время с TNF-α или другим индуцирующим VCAM-1 материалом, затем разрушают при помощи, например, звука или методом замораживания-оттаивания. Выделяют цитозольные и мембранные фракции. Для определения их количества добавляют радиоактивномеченую PUFA. Исследуется способность жидкости превращать PUFA в ox-PUFA в присутствии или отсутствие исследуемого соединения. Вместо разрушенных клеточных систем могут быть использованы интактные клетки.
Пирролидин дитиокарбамат (РDТС), при оральном введении в дозе 25-50 мг/кг/день, игбириует образование атерогенной жировой полоски, моноцит-макрофагавое артериальное воспаление, экспрессию эндотелием VCAM-1 и существенно нормализует функцию эндотелийзависимого расслабления у гиперхолестеринемических кроликов, находящихся на диете, вызывающей уровень холестерина выше 1000 мг/дл. В тех же самых дозах другие естественные терапевтические агенты, такие как антиоксиданты пробукол и витамин Е, не оказывали такого действия на образование поражений на этой модели.
При экспериментальном атеросклерозе эндотелийзависимое артериальное расслабление восстанавливается при введении РDТС. При диете индуцированная гиперхолестеринемичная модель кроликов при оральном введении РDТС (25-50 мг/кг/день) восстанавливает зависимую от эндотелия реактивность сосуда. Это определяли при помощи изучения сокращения кольца аорты, вырезанной у контрольных и тестируемых животных. У пациентов с атеросклерозом это проявляется само в виде нормализации периферической сосудистой реактивности в ответ на гиперемию, что определяли путем неинвазивных исследований при помощи эффекта Допплера в потоке. Это является стандартизованным, широко применимым и легко проводимым тестом, которым может быть использован для титрования функционального уровня препарата при оральном введении дозы. РDTС действует при противоишемической терапии путем быстрой нормализации нарушений при патологическом эндотелий-зависимом артериальном сокращении сосудов при сердечно-сосудистом заболевании и атеросклерозе. Данное улучшение тока крови в сосуде проявляется в виде улучшения симтоматики и осуществления ограниченной ишемией функции и представляет неинвазивную оценку защиты сосуда. Другие клинические показания при нарушениях в эндотелий-зависимом сосудистом сокращении включают импотенцию.
Молекулярный регуляторный фактор, который подавляет транскрипцию гена VCAM-1, представляет новый сложный регуляторный фактор транскрипции, состоящий из р65 и р50 субъединиц NF-kB/Rel семейства, перекрестно-связанного с с-fos и с-jun субъединицами АР-1 семейства. При помощи структурного и функционального исследования было установлено, что данные АР-1 факторы играют роль в регуляции промотера VСАМ-1, который, по-видимому, является центральным в терапевтической регуляции экспрессии гена VCAM-1. Это является первым свидетельством функциональной роли данного перекрестно-связанного комплекса транскрипции в регуляции эндогенного гена.
Краткое описание чертежей
Фиг.2 представляет график экспрессии VСАМ-1 на поверхности клетки (О.П. 450 нм) в виде функции количества часов в эндотелиальных клетках аорты человека, подверженных воздействию цитокина TNF-α (закрашенные кружки); линолевой кислоты (закрашенные треугольники); линолеил гидропероксида (13-HPODE, закрашенные квадраты); в отсутствие воздействия данными веществами (контроль, незакрашенные квадраты).
Фиг.3 представляет график экспрессии VCAM-1 на поверхности клетки (О.П. 450 нм) в эндотелиальных клетках аорты человека при взаимодействии с линолевой кислотой (закрашенные треугольники) и линолеил гидропероксидом (13-НРODЕ, закрашенные квадраты) как функция концентрации жирной кислоты (мкМ).
Фиг.4 представляет графическую диаграмму экспрессии (О.П. 450 нм) на поверхности клетки VCAM-1, ICAM-1 и Е-селектина эндотелиальными клетками аорты человека под воздействием цитокина TNF-α, стеариновой кислоты, олеиновой кислоты, линолевой кислоты, линоленовой кислоты и арахидоновой кислоты.
Фиг.5 представляет графическую диаграмму экспрессии VCAM-1 (О.П. 450 нм) на поверхности эндотелиальных клеток аорты человека под действием линолевой кислоты, 13-НРODЕ, арахидоновой кислоты и гидропероксида арахидоновой кислоты (15-НРЕТЕ), с (закрашено черным) или без (пунктирная линия) антиоксидантом пирролидин дитиокарбаматом.
Фиг.6 иллюстрирует авторадиограммы, показывающие резкую индукцию мРНК VCAM-1 под действием линолевой кислоты и 13-НРОDЕ. НАЕС (эндотелиальные клетки аорты человека) выдерживали с олеиновой кислотой (7,5 мкМ), 13-НРОDЕ (7,5 мкМ) или ТNF-α (100 единиц/мл) или без них. Общую РНК выделяли и 20 мкг фракционировали по размеру путем денатурирующего электрофореза на 1,0% агарозаформальдегидном геле, переносили на нитроцеллюлозу и гибридизовали с или 32Р-меченым человеческим: А) VСАМ-1, или с В) β-актинспецифической кДНК. После отмывания фильтры облучали с ренгеновской пленкой при -70°С с одним интенсифицирующим экраном в течение 24 часов. Идентификация линий:
1) контроль;
2) линолевая кислота (резкий ответ, 8 часов воздействия);
3) линолевая кислота (резкий ответ, 48 часов воздействия);
4) 13-НРОDЕ (резкий ответ, 8 часов воздействия); и
5) TNF-α (100 единиц/мл), 4 часа воздействия).
Фиг.7 представляет иллюстрацию авторадиограммы, которая показывает, что индукция мРНК VCAM-1 под действием полиненасыщенных жирных кислот является независимой от синтеза клеточного белка. НАЕС выдерживали с линолевой (7,5 мкМ) или арахидоновой (7,5 мкМ) кислотой в присутствии или в отсутствии циклогексимида (10 мкг/мл) в течение 4-часового периода, и затем обрабатывали, как описано для фиг.5.
Фиг.8 представляет иллюстрацию авторадиограммы, которая показывает, что линолевая кислота индуцирует транскрипционную активацию промотера VCAM-1 при помощи чувствительного к окислению-восстановлению NF-kB-подобного фактора. НАЕС трансфецировали либо 30 мкг р288 VCAMCAT, р85 VCAMCAT, либо pSV2CAT плазмидой методом соосаждения фосфатом кальция, используя обычную технику. После 24-часового восстановительного периода НАЕС предварительно обрабатывали, или нет, 50 мкМ PDTC, и после 30 минут выдерживания с линолевой кислотой (7,5 мкМ) или ТNF-α (100 ед/мл) непосредственно наносили на чашки. Через 18 часов готовили экстракты клеток методом быстрого замораживания-оттаивания в 0,25 М Трис, рН 8,0. Для определения активности хлорамфениколацетил-трансферазы (CAT), как ранее описано [Ausubel, 1989], исследовали белок каждого клеточного экстракта (Ас-ацетилированный; N-неацетилированный хлорамфеникол).
Фиг.9 иллюстрирует полоску акриламидного геля, на которой показано, что полиненасыщенные жирные кислоты активируют NF-kВ-подобные ДНК связывающие активности, которые подавляются под действием антиоксиданта PDTC. Конфлюентные НАЕС в среде, содержащей 4% FBS (как описано на фиг.1), предварительно обрабатывали с или без РDТС (50 мкМ) в течение тридцати минут и затем выдерживали в течение трех часов с линолевой кислотой (7,5 мкМ), олеиновой кислотой (7,5 мкМ) или TNF-α (100 ед/мл) соответственно. Пять микрограмм экстракта ядер инкубировали с двухнитевым 32P-меченым wt VCAM, фракционировали по размеру в 4% чистом полиакриламидном геле и выдерживали с авторадиографической пленкой при -70°С в течение 18 часов. Выделяли две полосы А и С, представляющие NF-kB-подобную связывающую активность. Слабая полоса В наблюдалась в контроле (необработанные) клетки.
Фиг.10А и 10B представляют графические столбчатые диаграммы сравнительной реактивности тиобарбитуровой кислоты (О.П. 532 нм) и соединений арахидоновой кислоты и 15-НРЕТЕ в присутствии или отсутствии РDТС. Исследование реактивности тиобарбитуровой кислоты (TBARS) определяет окислительную способность материала в бесклеточном, свободном от среды окружении.
Фиг.11 представляет иллюстрацию авторадиограммы мРНК, полученную, как описано ниже, гибридизованную с 32Р-меченой специфической кДНК VCAM-1 (Панель А), специфической кДНК Е-селектина (ELAM-1) (Панель В) или специфической кДНК ICAM-1 (Панель С). После обработки в течение 30 минут 50 мкМ натрий пирролидин дитикарбаматом (РDТС) клетки HUVЕ (аллантоисная вена человека) выдерживали с IL-1b (10 ед./мл) при постоянном присутствии 50 мкМ РDТС. Идентично проводили параллельный контроль, в отсутствие PDTC. В определенное время выделяли общую РНК и фракционировали по размерам 20 мкг материала путем денатурирующего электрофореза в 1,0% агарозаформальдегидном геле, переносили на нитроцеллюлозу, гибридизовали, как описано выше, и визуализировали путем авторадиографии. Линия 1 - 0 часов; Линии 2, 4, 6, 8 - только OL - 1 для 2, 4, 8 и 24 часов соответственно; Линии 3, 5, 7, 9 - 1L-1 с РDТС для 2, 4, 8 и 24 часов, соответственно.
Фиг.12 представляет авторадиограмму мРНК, полученную, как описано ниже, гибридизованную с 32Р-меченым специфическим VCAM-1 человека (Панель А), специфической кДНК-Е-селектина (ELAM-1) (Панель В) или специфической кДНК ICAM-1 (Панель С). Клетки HUVE предварительно обрабатывали определенными концентрациями РDТС, затем выдерживали с 1L-1b в присутствии РDТС в течение четырех часов и определяли аккумуляцию мРНК для VCAM-1 Northern-анализом путем гибридизации на фильтре. Линия 1 - контроль, линия 2 - IL-1 (10 ед/мл), линия 3 - IL-1b+PDTC (0,05 мкМ), линия 4 - IL-1 LB+PDTC (0,5 мкМ), линия 5 - IL-1b+PDTC (5,0 мкМ), линия 6 - IL-1b+РDТС (50,0 мкМ), линия 7 - IL-1b+РDТС (100 мкМ).
Фиг.13 представляет иллюстрацию авторадиограммы мРНК, полученную, как описано ниже, гибридизованную с 32Р-меченой специфической кДНК VCAM-1 человека (Панель А), специфической кДНК Е-селектина (ELAM-1) (Панель В) или специфической кДНК (ICAM-1) (Панель С). Клетки HUVE предварительно обрабатывали, как описано на фиг.10, 50 мкМ PDTC, выдерживали четыре часа с агентами, отмеченными ниже, и исследовали на аккумуляцию мРНК VCAM-1 (Панель А) и ICAM-1 (Панель В). Линия 1 - ТNF-α (100 ед/мл), линия 2 - TNF-α+PDTC, линия 3 - липополисахарид (LPS) (100 нг/мл), линия 4 - LPS+PDTC, линия 5 - поли(I:С) (100 мг/мл), линия 6 - поли(I:С) + PDTC.
Фиг.14 представляет график экспрессии VCAM-1 и ICAM-1 на поверхности клетки в присутствии (темные столбцы) или отсутствие (белые столбцы) РDТС и в присутствии множества типов индуцирующих стимулов. Конфлюэнтные клетки НUVЕС предварительно обрабатывали или не обрабатывали (только CTL) в течение 30 минут с 50 мкМ PDTC, затем выдерживали указанное время с указанным агентом в присутствии или отсутствие (только CTL) PDTC. Экспрессию на клеточной поверхности определяли путем первичного связывания со специфическими моноклональными антителами к VCAM-1 (4B9) и ICAM-1 (84H10) и последующим вторичным связыванием с пероксидазой хрена, привязанной к антимышиному (IgG) козла. Количественный подсчет проводили путем определения калориметрической конверсии ТМВ при 450 нм. Фиг.14 показывает, что множественные регуляторные сигналы индуцируют VСАМ-1, но не ICAM-1, посредством общего, чувствительного к дитиокарбамату пути и эндотелиальных клетках сосуда человека.
Фиг.15 представляет график относительной экспрессии VСАМ-1 на поверхности клетки (О.П. 595 нм) эндотелиальных клеток аллантоисной вены человека, активированных TNF-α, по отношению к концентрациям различных антиоксидантов. РDТС представляет натрий N-пирролидин дитиокарбамат; DETC представляет натрий N,N-диэтил-N-карбодитиолат, также называемый как натрий диэтилдитиокарбамат; NАС представляет N-ацетилцистеин и DF представляет десферроксимин).
Фиг.16 представляет график относительной экспрессии VCAM-1 на поверхности клетки (О.П. 595 нм) в эндотелиальных клетках аллантоисной вены человека, активированных ТNF-α в присутствии специфического количества антиоксиданта. (РDТС представляет натрий N-пирролидин дитиокарбамат; DIDТС представляет натрий N,N-диэтил-N-карбодитиоат; SarDTC представляет натрий N-метил-N-карбоксиметил-N-карбодитиоат; IDADC представляет тринатрий N,N-ди(карбоксиметил)-N-карбодитиоат; MGDTC представляет натрий N-метил-D-глюкамин-N-карбодитиоат; МеОВGDТС представляет натрий N-(4-метоксибензил)-D-глюкамин-N-карбодитиоат; DEDTC представляет натрий N,N-диэтил-N-карботиоат; DiPDTC представляет натрий N,N-диизопропил-N-карбодитиоат; NАС представляет N-ацетилцистеин).
Фиг.17 представляет график процента связывания Mo1t-4 клеток с клетками НUVЕ, стимулированных TNF-α (100 ед./мл) или не стимулированных, в течение шести часов в присутствии или отсутствие РDТС.
Фиг.18 представляет химические структуры следующих активных дитиокарбаматов: натрий пирролидин-N-дитиокарбамат, натрий N-метил-N-карбоксиметил-N-карбодитиоат, тринатрий N,N-ди(карбоксиметил)-N-карбодитиоат, натрий N-метил-D-глюкамин-N-карбодитиоат, натрий N,N-диэтил-N-карбодитиоат (натрий диэтилдитиокарбамат) и натрий N,N-диизопропил-N-карбодитиоат.
Фиг.19 представляет графическую столбчатую диаграмму влияния РDТС на образование флуоресцентных продуктов присоединения БСА и 13-НОРОDЕ, измеряемое в единицах флуоресценции, по отношению к концентрации РDТС. Один микромоль 13-НРОDЕ инкубировали с 200 микрограммами БСА в присутствии PDTC в течение шести дней. Флуоресценцию измеряли при 430-460 нм, возбуждение при 330-360 нм.
Фиг.20 представляет графическое отображение влияния концентрации РDTС на образование флуоресцентных продуктов присоединения БСА и ох-РUFА как функция длины волны (нм) и концнтрации РDТС. При увеличении концентрации РDТС количество флуоресцентных продуктов присоединения снижается.
Фиг.21 представляет графическое отображение влияния РDTС на окисление ЛНП под действием пероксидазы хрена (ПХ), измеряемого по увеличению О.П. (234 нм), относительно времени (минуты) при различных концентрациях РDТС. Было обнаружено, что после инкубационного периода PDTC ингибирует окисление ЛНП под действием ПХ по концентрационно-зависимому типу.
Фиг.22 представляет диаграмму действия РDТС на образование ох-РUFA, индуцированного цитокином в эндотелиальных клетках аорты человека. Как отмечено, и TNF-α, и IL-1b вызывают окисление линолевой кислоты в окисленную линолевую кислоту. Окисление существенно предотвращается PDTC.
Подробное описание изобретения
1. Определения
Использующийся термин полиненасыщенная жирная кислота (также указывается здесь как "ПНЖК") относится к жирной кислоте (обычно С8-С24) которая имеет, по меньшей мере, две алкенильные связи и включает, но не ограничивается ими, линолевую (C18 Δ9, 12), линолиновую (C18 Δ6, 9, 12), арахидоновую (С20 Δ5, 8, 11, 14) и экозатриеновую (С20 Δ8, 11, 14) кислоты.
Термин окисленная полиненасыщенная жирная кислота относится к ненасыщенной жирной кислоте, в которой, по меньшей мере, одна из алкенильных связей преобразована в гидропероксид. Неограничивающими примерами являются

Термин алкил, как здесь используется, за исключением особых случаев относится к насыщенному прямому, разветвленному или циклическому (в случае С5 или более) углеводороду C1-10 (или низший алкил, то есть С1-С5), который, в частности, включает метил, этил, пропил, изопропил, бутил, изобутил, трет-бутил, пентил, циклопентил, изопентил, неопентил, гексил, изогексил, циклогексил, циклогексилметил, 3-метилпентил, 2,2-диметилбутил и 2,3-диметилбутил. Алкильная группа может быть не обязательно замещена по любому из углеродов одним или несколькими радикалами, выбранными из группы, содержащей гидроксил, амино или моно- или дизамещенный амино, где замещающая группа представляет собой, независимую, алкил, арил, алкарил или аралкил; арил, алкокси, арилокси, нитро, циано, сульфоновая кислота, сульфат, фосфоновая кислота, фосфат или фосфонат, либо незащищенные, либо защищенные, как необходимо, как известно специалисту в данной области, например, как описано Greene et at.; "Protective Groups in Organic Sunthesis", John Wiley and Sons. Second edition, 1991.
Термин алкенил, как здесь используется и если не указано особо, относится к прямому, разветвленному или циклическому углеводороду C2-C10 с, по меньшей мере, одной двойной связью.
Термин алкинил, как здесь используется и если не указано особо, относится к C2-C10 прямому или разветвленному углеводороду с, по меньшей мере, одной тройной связью.
Термин аралкил относится к арильной группе с, по меньшей мере, одним алкильным заместителем.
Термин алкарил относится к алкильной группе, которая имеет, по меньшей мере, один арильный заместитель.
Термин галоген (алкил, алкенил или алкинил) относится к алкильной, алкенильной или алкинильной группе, в которой, по меньшей мере, один из водородов в группе замещен атомом галогена.
Термин арил, как здесь используется, и если не указано особо, относится к фенилу, бифенилу или нафтилу, предпочтительно к фенилу. Арильная группа может быть необязательно замещена одним или несколькими радикалами, выбранными из группы, содержащей алкил, гидроксил, амино, алкиламино, ариламино, алкокси, арилокси, нитро, циано, сульфоновую кислоту, сульфат, фосфоновую кислоту, фосфат или фосфонат, СО2Н или его фармацевтически приемлемую соль, СО2(алкил, арил, алкарил или аралкил), или глюкамин, либо незащищенные, либо защищенные, как необходимо, как известно специалисту в данной области, например, как описано Jreene et al. "Protective Jroups in Organic Synthesis", John Wiley and Sons. Second edition. 1991.
Термин алкокси, как здесь используется и если не указано особо, относится к радикалу структуры -O-алкил.
Термин ацил, как здесь используется, относится к группе формулы С(О)R', где R' обозначает алкильную, арильную, алкарильную или аралкильную группу.
Термин гетероарил или гетероароматический, как здесь используется, относится к ароматическому радикалу, который включает, по меньшей мере, один атом серы, кислорода или азота в ароматическом кольце. Неограничивающими примерами являются феназин, фенотиазин, фурил, пиридил, пиримидил, тиенил, изотиазолил, имидазолил, тетразолил, пиразинил, бензофуранил, бензотиофенил, хинолил, изохинолил, бензотиенил, изобензофурил, пиразолил, индолил, изоиндолил, бензимидазолил, пуринил, морфолинил, карбазолил, оскалил, тиазолил, изотиазолил, 1,2,4-тиадиазолил, изооксазолил, пирролил, пиразолил, хиназолинил, пиридазинил, пиразинил, циннолинил, фталазинил, хиноксалинил, ксантинил, гипоксантинил, птеридинил, 5-азацитидинил, 5-азауроцилил, триазолопиридинил, имидазолопиридинил, пирролопиримидинил, пиразолопиримидинил, аденин, N6-алкилпурины, N6-бензилпурин, N6-галопурин, N6-винилпурин, N6-ацетиленовый пурин, N6-ацилпурин, N6-гидроксиалкилпурин, N6-тиоалкилпурин, тимин, цитозин, 6-азапиримидин, 2-меркаптопиримидин, урацил, N5-алкилпиримидины, N5-бензилпиримидины, N5-галогенпиримидины, N5-винилпиримидин, N5-ацетиленовый пиримидин, N5-ацилпиримидин, N5-гидроксиалкилпурин и N6-тиоалкилпурин и изоксазолил. Гетероароматическая группа может быть не обязательно замещена, как описано выше для арила. Гетероароматическая группа может быть частично или полностью гидрирована, как желательно. В качестве неограничивающего примера, вместо пиридина может быть использован дигидропиридин. Функциональные кислородные и азотные группы у гетероциклической основы могут быть защищены, если необходимо, в ходе реакции. Подходящими защитными группами, хорошо известными специалистам в данной области, являются 3-метилсилил, диметилгексилсилил, трет-бутилдиметилсилил и трет-бутилдифенилсилил, тритилметил, алкильные группы, ацильные группы, такие как ацетил и пропионил, метилсульфонил и п-толуилсульфонил.
Термин гидроксиалкил, как здесь используется, относится к C1-C6алкильной группе, в которой, по меньшей мере, один из водородов, присоединенных к любому из атомов углерода, замещен гидрокси-группой.
Термин тиольный антиоксидант относится к серосодержащему соединению, которое замедляет окисление.
Термин фармацевтически приемлемое производное относится к производному активного соединения, которое при введении реципиенту способно создать, прямо или опосредованно, основное соединение или которое проявляет его активность.
Термин "фармацевтически приемлемый катион" относится к органическому или неорганическому радикалу, который несет положительный заряд и который может быть введен в ассоциации с фармацевтическим агентом, например, в качестве противокатиона в соли. Фармацевтически приемлемые катионы известны специалистам в данной области и включают, но не ограничиваются ими, натрий, калий и четвертичный амин.
Термин "физиологически отщепляемая уходящая группа" относится к радикалу, который может быть отщеплен in vivo от молекулы, к которой он присоединен, и включает, но не ограничивается ими, органический или неорганический анион, фармацевтически приемлемый катион, ацил (включая, но не ограничиваясь ими, (алкил)С(О), включая ацетил, пропионил и бутирил), алкил, фосфат, сульфат и сульфонат.
Термин "энантиомерно обогащенная композиция или соединение" относится к композиции или соединению, которое включает, по меньшей мере 95%, и предпочтительно, по меньшей мере, 97, 98, 99 или 100% по массе индивидуального энантиомера соединения.
Термин аминокислота включает синтетические или встречающиеся в природе аминокислоты, включая, но не ограничиваясь ими, например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, трептофан, метионин, глицин, серин, треонин, цистеин, тирозин, аспарагин, глутамин, аспартил, глутаоил, лизин, аргенин и гистидин.
"Связующий радикал", как здесь используется, является любой двухвалентной группой, которая связывает два химических остатка, включая, но не ограничиваясь ими, алкил, алкенил, алкинил, арил, полиалкиленокси (например, -[(СН2)nO-]n-), -С1-6алкокси-С1-10алкил-, -С1-6алкилтио-С1-10алкил-, NR3- и -(СНОН)nСН2ОН, где n независимо, равно 0, 1, 2, 3, 4, 5 или 6.
II. Идентификация окисленных и неокисленных полиненасыщенных жирных кислот в качестве непосредственных медиаторов экспрессии VCAM-1
Для установления, действуют ли PUFA или окисленная РUFА в качестве непосредственного иммуномодулятора экспрессии гена в эндотелиальных клетках, ранние пересевы эндотелиальных клеток аорты человека (НАЕС) культивировали в течение восьми часов в среде с сывороткой и выдерживали с насыщенной (стеариновая), мононенасыщенной (олеиновая) и полиненасыщенными (линолевая и арахидоновая) жирными кислотами; а также с гидропероксидами линолевой (13-HPODE) или арахидоновой (15-НРЕТЕ) кислот. НАЕС альтернативно выдерживали также с цитокином - фактором некроза опухолей α.
НАЕС выдерживали с линолевой кислотой или 13-НРОDE различное время, до 48 часов, и затем определяли экспрессию VCAM-1 на клеточной поверхности при помощи анализа ELISA. Результаты сравнивали с НАЕС, выдержанных с цитокином TNF-α (100 ед./мл) в течение такого же периода. Экспрессия VCAM-1 в НАЕС, выдерженных с либо линолевой кислотой, либо 13-НРОDE, кратковременно индуцировалась. Наблюдались пики экспрессии на приблизительно 8-9 часы со значительной экспрессией, на 24 часа и затем снижались к 48 часам. Кинетика индукции VCAM-1 под действием и линолевой кислоты, и 13-НРОDЕ отражает таковую для TNF-α, и, тем самым, механизм, по которому полиненасыщенные жирные кислоты индуцируют VCAM-1, следовательно, представляется подобным таковому для ТNF-α.
Было проведено также дозозависимое исследование экспрессии гена VCAM-1 под действием линолевой кислоты и 13-НРОDЕ. Было обнаружено, что 7,5 мкМ является самой низкой пиковой дозой, при которой линолевая кислота и 13-НРОDЕ существенно индуцирует экспрессию гена VCAM-1.
Было проведено исследование, чтобы выяснить, вызывает ли кратковременная инкубация эндотелиальных клеток с полиненасыщенными жирными кислотами индукцию экспрессии как ICAM-1, так и Е-селектина. Было определено, что полиненасыщенные жирные кислоты линолевая и арахидоновая кислоты индуцируют экспрессию гена на поверхности клетки до 59% от THF-индуцированной экспрессии гена VCAM-1. Как оказалось, ни IСАМ-1, ни Е-селектин не индуцировались данными жирными кислотами. Наоборот, насыщенная жирная кислота - стеариновая кислота и мононенасыщенная жирная кислота - олеиновая кислота не индуцируют экспрессию VCAM-1, ICAM-1 или Е-селектина. Экспрессия гена VCAM-1 также наблюдалась при инкубации НАЕС с окисленными метаболитами линолевой кислоты (13-HPODE) и арахидоновой кислоты (15-НРЕТЕ).
Для исследования, действительно ли окислительный стресс у эндотелиальных клеток, вызванный полиненасыщенными жирными кислотами и их окисленными метаболитами, индуцирует VCAM-1 посредством чувствительного к окислению-восстановлению механизма, НАЕС предварительно обработали антиоксидантом пирролидин дитиокарбаматом (РDТС, 50 мкМ) в течение 30 минут, и затем клетки независимо инкубировали с линолевой кислотой, арахидоновой кислотой, 13-НРОDЕ и 15-НРЕТЕ (все 7,5 мкМ) в течение 8 часов. Было определено, что PDTC подавляет экспрессию гена VСАМ-1, индуцированную полиненасыщенными жирными кислотами и их окисленными производными. Это показывает, что индукция опосредуется при помощи окисленной сигнальной молекулы, и что индукция предотвращается, когда окисление молекулы заблокировано (т.е. отсутствие окисления), обращается (т.е. сигнальная молекула является восстановленной), или ее взаимодействие с белковой мишенью предотвращено, возможно, посредством окислительно-восстановительного комплекса.
Для определения, действительно ли избирательная индукция VCAM-1 под действием РUFАS и ее окисленных метаболитов наблюдается на уровне мРНК, НАЕС инкубировали с линолевой кислотой или 13-HPODE. Аккумуляция мРНК VCAM-1, индуцированная линолевой кислотой и 13-НРОDЕ, была близка к уровням, которые индуцировал TNF-α. Напротив, индукции экспрессии гена VCAM-1 и Е-селектина на уровне мРНК и НАЕС, инкубированных с линолевой килотой или 13-НРОDЕ, не наблюдалось. Полученные данные соответствуют данным, полученным при исследовании клеточной поверхности. Результаты показывают, что претрансляционные регуляторные механизмы опосредуют индукцию экспрессии гена VCAM-1 под действием полиненасыщенных жирных кислот и их окисленных метаболитов.
Было также необходимо определить, действительно ли полиненасыщенные жирные кислоты действуют в качестве первичного сигнала или действуют посредством регуляторного белка, участвующего в индукции экспрессии гена VCAM-1 под действием IL-4. Для исследования, действительно ли недавно синтезированные белки, такие как IL-4, участвуют в синтезе и экспрессии VCAM-1, индуцированным PUFAS, такими как линолевая кислота, НАЕС инкубировали с 13-HPODЕ (7,5 мкМ) и выдерживали с ингибитором белкового синтеза циклогексимидом. Ингибирования аккумуляции мРНК VCAM-1 под действием циклогексимида в НАЕС, инкубированных с 13-НРОDЕ, не наблюдалось. Определяли также количество продуцирующегося IL-4 в НАЕС, инкубированных с линолевой кислотой или арахидоновой кислотой и их окисленными метаболитами, при помощи метода ELISA. Увеличение выхода IL-4 из НАЕС, инкубированных с данными РUFAS и их окисленными метаболитами, не наблюдалось.
Ранее было обнаружено, посредством изучения делении и гетерологического промотера, что цитокины и нецитокины активируют экспресссию гена VCAM-1 в эндотелиальных клетках, по крайней мере, частично на уровне транскрипции, посредством двух NF-kB-подобных ДНК связывающих элементов. Было также показано, что РDTС ингибирует экспрессию гена VCAM-1 посредством чувствительного к окислению-восстановлению NF-kB подобного фактора. Для определения, действительно ли полиненасыщенные жирные кислоты индуцируют транскрипционную активацию промотера VCAM-1 человека по такому же механизму, НАЕС временно трансфецировали химерным репортерным геном р288 V CAM-CAT, содержащим координаты от -288 до +22 промотера VCAM-1 человека. Добавление линолевой кислоты (7,5 мкМ) индуцировало активность промотера VCAM-1, которая была более чем двукратной, чем в контроле, и приблизительно составляла 60% от максимального сигнала, индуцируемого TNF-α. Подобные результаты были получены с минимальным цитокин-индуцируемым промотером гена VCAM-1 (p85 VCAM-CAT), содержащим -77 и -63 пары оснований NF-kB подобных участков. Ни линолевая кислота, ни ТНF-α не обладали никаким эффектом на активность при использовании конструкции органически экспрессируемого рSV2 CAT. РDТС ингибирует транскрипционную активацию обеих конструкционных промотеров VCAM-1, индуцированных линолевой кислотой. Эти данные отмечают, что, аналогично ТНF-α, полиненасыщенные жирные кислоты, такие как линолевая кислота, индуцируют транскрипционную активацию VСАМ-1 посредством NF-kB подобного чувствительного к окислению-восстановлению механизма.
Для определения, действительно ли полиненасыщенные жирные кислоты и их окисленные метаболиты регулируют активность промотера VCAM-1 посредством NF-kB подобного транскрипционного регуляторного фактора, в ядерных экстрактах НАЕС была исследована активность связывания ДНК с двухнитевым нуклеотидом, содержащим элементы NF-кВ подобного промотера VCAM-1, расположенного в положении -77 и -63. Как показано на фиг.8, две полосы А и С, представляющие NF-kB подобную активность, были индуцированы в ответ на трехчасовое выдерживание с линолевой кислотой (7,5 мкМ). Подобное обнаружение наблюдали при выдерживании с цитокином TNF-α (100 ед./мл). Слабую полосу наблюдали в контроле (необработанных) клетках. Не наблюдали никакого NF-кВ подобного связывания с олеиновой кислотой - мононенасыщенной жирной кислотой. Предварительная обработка клеток в течение тридцати минут с PDTC ингибировала активность связывания ДНК с А и С, после активации линолевой кислотой. Данные обнаружения близки к полученным ранее данным, показывающим, что РDТС блокирует активацию экспрессии гена VCAM-1 в HUVEC путем ингибирования активации данных NF-kВ-подобных ДНК-связывающих белков.
Пример 1.
Влияние окисленных и неокисленных полиненасыщенных жирных кислот на кинетику активации экспрессии гена VCAM-1
Эндотелиальные клетки аорты человека (НАЕС) наносили на микротитрационное плато с 96 ячейками и инкубировали с линолевой кислотой (7,5 мкМ), 13-НРОDЕ (7,5 мкМ) или ТNF-α (100 ед/мл) в течение пяти различных временных промежутков, до 48 часов. НАЕС, полученные от Clonetics (Boston, MA), культивировали в среде 199, дополненной 20% эмбриональной бычьей сывороткой (FBS), 16 ед/мл гепарином, 10 ед/мл эпидермальным фактором роста, 50 мкг/мл добавкой для роста эндотелиальных клеток, 2 мМ L-глютамина, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина. За один день до эксперимента клетки поместили в среду, содержащую 4% FBS. Конфлюентные НАЕС инкубировали до 48 часов с ТNF-α (100 ед/мл) или стеариновой, олеиновой, линолевой, линоленовой или арахидоновой кислотами (7,5 мкМ). Подобные исследования проводили с различными дозами линолевой кислоты или 13-HPODE в течение 8 часового периода (1-60 мкМ) (фиг.3). Подсчет проводили путем определения калориметрического перехода окраски ТМВ при 450 нм. Исследования проводили трижды (n=4, для каждого экспериментального значения) * - значение отличается от контроля (р<0,05).
Как показано на фиг.2, и линолевая кислота, и 13-НРОDЕ индуцировали экспрессию VCAM-1. Через десять часов после выдерживания количество поверхностно-клеточного VCAM-1, индуцированного линолевой кислотой и 13-НРОDЕ, составляло более чем половину от того, которое индуцирует цитокин TNF-α.
Как показано на фиг.3, индукция VCAM-1 под действием линолевой кислоты и 13-НРОDЕ является чувствительной к концентрации. При концентрации данных соединений между 2 и 10 мкМ наблюдается резкое увеличение количества индуцированного VCAM-1 на поверхности клетки, которое затем остается приблизительно постоянным, вплоть до концентрации, по крайней мере, 100 мкМ. Необходимо отметить, что концентрация PUFA, отмеченная на фиг.3, является дополнительной к той, которая эндогенно обнаружена в НАЕС.
Пример 2
Полиненасыщенные жирные кислоты индуцируют экспрессию гена VCAM-1, но не ICAM-1 или Е-селектина
Экспрессию VСАМ-1, IСAМ-1 и Е-селектина на клеточной поверхности НАЕС определяли методом ELISA. НАЕС, полученные от Clonetics (California), культивировали в среде 199, дополненной 20% эмбриональной бычьей сывороткой (FВS), 16 ед/мл гепарином, 10 ед/мл эпидермальным фактором роста, 50 мкг/мл добавкой для роста эндотелиальных клеток, 2 мМ L-глютамина, 100 ед/мл пенициллина и 100 мкг/мл стрептомицина. На один день до эксперимента клетки поместили в среду, содержащую 4% FBS. Конфлюентные НАЕС инкубировали или нет 8 часов с TNF-α (100 ед/мл) или стеариновой, олеиновой, линолевой, линоленовой или арахидоновой кислотами (7,5 мкМ). Экспрессию A) VCAM-1, В) ICAM-1 и С) Е-селектина на поверхности клеток определяли при помощи первичного связывания с антителами мыши, специфическими к VCAM-1, специфическими к ICAM-1 и специфическими к Е-селектину, затем вторичного связывания с козьим антимышиными (IgG), связанным с меченой пероксидазой хрена. Подсчет проводили при помощи определения калориметрического перехода ТМВ при 450 нм. Исследование проводили трижды (n=4, для каждого экспериментального значения) * - значение отличается от контроля (р<0,05).
Как показано на фиг.4, линолевая кислота, линоленовая кислота и арахидоновая кислота значительно индуцировали экспрессию VCAM-1, но не экспрессию ICAM-1 и Е-селектина на поверхности клетки. Ни стеариновая кислота, ни олеиновая кислота не индуцировали экспрессию VCAM-1, ICAM-1 и Е-селектина. TNF-α сильно индуцировал экспрессию всех трех молекул на поверхности клетки.
Пример 3
Антиоксидант РDТС подавляет индукцию VCAM-1 под действием полиненасыщенных жирных кислот и их окисленных метаболитов
Конфлюентные НАЕС предварительно обрабатывали в присутствии или отсутствие РDТС (натрий пирролидин дитиокарбамата, 50 мкМ) в течение тридцати минут. Затем клетки инкубировали в течение восьми часов с TNF-α (100 ед/мл), линолевой или арахидоновой кислотой (7,5 мкМ) или гидропероксидами жирных кислот 13-НРОDЕ (7,5 мкМ) или 15-НРЕТЕ (7,5 мкМ). Экспрессию VСАМ у НАЕС на поверхности клеток определяли методом ELISA, как описано в примере 1. Исследования проводили в трипликатах (n=4, для каждого экспериментального значения), * - значение отличается от контроля (р<0,05).
Как отмечено на фиг.5, РDТС подавляет индукцию VCAM-1 под действием линолевой кислоты, 13-HPODЕ, арахидоновой кислоты и 15-НРЕТЕ.
Пример 4
Быстрая индукция мРНК VCAM-1 под действием линолевой кислоты и 13-HPODE
НАЕС выдерживали с линолевой кислотой (7,5 мкМ) или 13-НРОDЕ (7,5 мкМ). Выделяли общую РНК и 20 мкг фракционировали по размеру путем денатурирующего электрофореза в 1,0% агарозаформальдегидном геле, переносили на нитроцелюлозу и гибридизовали с 32Р-меченой ДНК человека, специфической или к А) VCAM-1, или специфической к В) β-актину, и визуализировали путем авторадиографии. После промывания фильтры выдерживали с рентгеновской пленкой при -70°С с одним интенсифицирующим экраном в течение 24 часов. Идентификация линий:
1) контоль;
2) линолевая кислота (острая, 8 часов выдержки);
3) линолевая кислота (48 часов выдержки);
4) 13-НРODЕ (острая, 8 часов выдержки); и
5) TNF-α (100 ед/мл, 4 часа выдержки).
Как показано на фиг.6, и линолевая кислота, и 13-НРОDЕ индуцируют продукцию мРНК для VCAM-1 через 8 часов. Через 48 часов линолевая кислота больше не вызывала индукцию мРНК VСАМ-1.
Пример 5
Индукция мРНК VCAM-1 под действием PUFA является независимой от синтеза клеточного белка
НАЕС выдерживали с либо линолевой, либо арахидоновой кислотой (7,5 мкМ) в присутствии или отсутствие циклогексимида (10 мкг/мл) в течение 4-часового периода. Общую РНК выделяли и 20 мкг фракционировали по размеру путем денатурирующего электрофореза в 1,08 агарозаформальдегидном геле, переносили на нитроцеллюлозу и гибридизовали с специфической кДНК А) 32P-меченой VCAM-1 или B) β-актина и визуализировали путем авторадиографии. После промывания фильтры выдерживали с рентгеновской пленкой при -70°С с одним интенсифицирующим экраном в течение 24 часов.
Как отмечено на фиг.7, индукция VCAM-1 под действием линолевой и арахидоновой кислот является независимой от синтеза клеточного белка.
Пример 6
Линолевая кислота индуцирует транскрипционную активацию промотера VCAM-1 через NF-kB подобный, чувствительный к окислению-восстановлению фактор
НАЕС нанесли на 100-мм чашки для выращивания в соотношении, необходимом для получения 60% конфлюентости. НАЕС трансфецировали с либо 30 мкг р288 VCAMCAT, р85 VСАМСАТ, либо с плазмидой pSV2 CAT путем способа соосаждения с кальцийфосфатом, используя обычную технику. После 24-часового восстановительного периода НАЕС предварительно обрабатывали 50 мкМ РDТС и после 30 минут выдерживания с линолевой кислотой (7,5 мкМ) или TNF-α (100 ед/мл), непосредственно нанося на чашки. Через 18 часов приготовили экстракты клеток при помощи быстрого замораживания-оттаивания в 0,25 М Трис, рН 8,0. Для определения активности хлорамфениколацетилтрансферазы (CAT), как ранее описано [Ausubel, 1989], определяли белок для каждого клеточного экстракта (Ас, ацетилированный; N, неацетилированный хлорамфеникол).
Фиг.8 иллюстрирует результаты данного эксперимента. Линолевая кислота индуцирует транскрипционную активацию промотера VСАМ-1 посредством NF-kB подобного чувствительного к окислению-восстановлению фактора. Данные результаты близки к тем, которые получены при активации промотера VCAM-1 под действием цитокинов, таких как ТNF-α. Это подтверждает, что PUFAS действуют через окисленные промежуточные продукты, которые также опосредуют активацию цитокином VCAM-1.
Пример 7
Полиненасыщенные жирные кислоты активируют NF-kB подобную, ДНК связывающую активность, которая подавляется под действием антиоксиданта РDTС
Конфлюентные НАЕС в среде, содержащей 4% FBS (как описано в примере 1), предварительно обрабатывали PDTC (50 мкМ) в течение 30 минут и затем выдерживали в течение 3 часов с линолевой кислотой, или олеиновой кислотой (7,5 мкМ), или ТNF-α (100 ед/мл). Пять микрограмм экстракта ядер инкубировали с двухнитевой 32Р-меченой wtVCAM, фракционировали по размеру на 4% акриламидном геле без добавок и выдерживали с авторадиографической пленкой при -70°С в течение 18 часов. Выделяли две полосы А и В, представляющие NF-kB подобную связывающую активность. Наблюдали слабую полосу В в контроле (необработанные) клетки).
Фиг.9 иллюстрирует, что линолевая кислота индуцирует NF-kB связывающую активность с VCAM-1 промотером, чувствительным к окислению-восстановлению образом. Это, аналогично цитокину ТNF-α, подтверждает близость механизма действия. TNF-α индуцирует VCAM-1, по-видимому, посредством механизма, который опосредуется при действии ox-PUFA.
Пример 8
Окисление в свободных от клеток, свободных от среды условиях, под действием неокисленной и окисленной (15-НРЕТЕ) арахидоновой кислоты
Фиг.10А и 10В представляют графические диаграммы реактивности соединений (О.П. 532 нм) арахидоновой кислоты и 15-НРЕТЕ, по отношению к тиобарбитуровой кислоте, в присутствии или отсутствие РDТС. Исследование реактивности тиобарбитуровой кислоты (TBARS) позволяет определять окислительную способность материала в бесклеточной среде, в свободном от среды окружении. Как видно из фигур, и арахидоновая кислота, и 15-НРЕТЕ проявляют заметную активность TBARS, которая ингибируется под действием РDТС.
III. Способ лечения нарушений, опосредованных VCAM-1
Обнаружение того что полиненасыщенные жирные кислоты и их окисленные метаболиты являются избирательными, чувствительными к окислению-восстановлению иммуномодуляторами, предлагает основу для терапии заболеваний, которые опосредуются VCAM-1 или генами, чувствительными к окислению-восстановлению.
Описывается способ лечения атеросклероза, постангиопластического рестеноза, заболеваний коронарных артерий, грудной жабы и других сердечно-сосудистых заболеваний, а также не сердечно-сосудистых воспалительных заболеваний, которые опосредуются VCAM-1, который включает устранение, уменьшение концентрации или предотвращение образования окисленных полиненасыщенных жирных кислот, включая, но не ограничиваясь, окисленную линолевую, линоленовую и арахидоновую кислоты. В альтернативном воплощении описывается способ лечения данных заболеваний, который включает предотвращение взаимодействие PUFA или ох-РUFА с белками или пептидами, которые опосредуют экспрессию VCAM-1.
Ингибирование экспрессии VCAM-1 может быть осуществлено множеством путей, включая путь посредством введения антиоксиданта, который предотвращает окисление полиненасыщенной жирной кислоты, путем модификации in vivo метаболизма PUFAS в ox-PUFAS, как описано подробно ниже.
1. Введение антиоксидантов
Любое соединение, которое восстанавливает ox-PUFA или которое ингибирует окисление PUFA и которое является относительно нетоксичным и биодоступным или которое может быть модифицировано для превращения его в биодоступное, может быть использовано в данной терапии. Обыкновенный лаборант, используя обычную технику, может легко определить, способно ли соединение восстанавливать ох-РUFА или ингибировать окисление PUFA.
Дитиокарбоксилантые антиоксиданты
Было обнаружено, что дитиокарбоксилаты являются полезными при лечении атеросклероза и других сердечно-сосудистых и воспалительных заболеваний. Дитиокарбоксилаты, включая дитиокарбаматы, могут быть использованы для блокирования способности клеток, включая эндотелиальные клетки, к экспрессии VCAM-1 или для подавления экспрессии редокс-чувствительного гена или активации гена, который подавляется посредством редокс-чувствительного следа.
По крайней мере, одно из соединений, перролидиндитиокарбамат (ПДТК), ингибирует экспрессию VCAM-1 гена при концентрации менее чем 1,0 микромолярная. Эти соединения также проявляют предпочтительную токсичность к пролифиративным или патологично делящимся клеткам сосудов гладких мышц. Другой дитиокарбамат, N-метил-N-карбоксиметил-N-карботитиоат натрия, также ингибирует экспрессию VCAM-1, без заметного действия на ICAM-1, но не проявляет предпочтительной токсичности к патологично делящимся клеткам сосудов гладких мышц.
Было обнаружено, что пирролидиндитиокарбамат не блокирует заметно экспрессию ELAM-1 или ICAM-1, и поэтому обработка этим соединением не оказывает вредного действия на противовоспалительный ответ опосредованный ELAM-1 или ICAM-1. Таким образом, избегается генерализация иммуносупрессии. Это позволяет избежать системных осложнений, вызываемых генерализованным ингибированием адгезивных молекул во многих других клеточных типах, в которых известно экспрессирование. Другие фармакологические приемлемые соли ПДТК также являются эффективными средствами при лечении сердечно-сосудистых и воспалительных заболеваний.
Дитиокарбаматы являются хелаторами переходных металлов, используемыми в клинике при интоксикации тяжелыми металлами. Baselt, R.C., F.W.J. Sunderman, et al. (1997), "Comparisons of antidotal efficacy of sodium diethyldithiocarbamat, D-penicillamine and triethylenetetramine upon acute toxicity of nickel carbonyl in rats." Res Commun Chem Pathol Pharmacol 18(4): 677-88; Henne, Т. and К. Kaaber (1978), "Treatment of pompholyx due to nickel allergy with chelating agents." Contact Dermatitis 4(5): 289-90; Sunderman, F.V. (1978), "Clinical response to therepeutic agents in poisoning from mercury vapor". Ann clin lab sci 8(4): 259-69; Sunderman, F.V. (1979), "Efficacy of sodium diethyldithiocarbamate (dithiocarb) in acute nickel carbonil poisoning." Ann clin lab sci 9(1): 1-10; Gale, G.R., A.B. Smith, et al. (1981), "Diethilditiocarbamat in treatment of acute cadmium poisoning." Ann clin lab sci 11(6): 476-83; Jons, M.M. and M.G. Cheriman (1990), "The search for chelate antagonists for chronic cadmium intoxication." Toxicology 62(1): 1-25; Jones, S.G., M.A. Basinger, et al. (1982), "A comparison of diethyldithiocarbamate and EDTA as antidotes for acure cadmium intoxication." Res Commun Chem Pathol Pharmacol 38(2): 271-8; Pades, A., J.S. Casas, et al. (1985), "Diethyldithiocarbamates in heavy metal poisoning: complexis of N,N,-di(1-hydroxiethyl)dithiocarbamate with Zn (II), Cd (II), Hg (II), СН3Нg (II) and C6H5Hg (II), J. Inorg Biochem 25 (I): 35-42; Tandon S.K., N.S. Hashmi, et al. (1990), "The lead-chelatinq effects of substituted dithiocarbamates." Biomed Environ Sci 3(3): 299-305.
Дитиокарбаматы также использовались как добавочные средства в цис-платиновой химиотерапии для предотвращения почечной токсичности. Hacker, M.R., W.B. Ershler, et al. (1982). "Effect of disulfiram (tetraethylthiuram disulfide) and diethyldithiocarbamat on the bladder toxicity and antitumor activity of cyclophosphamide in mice." Cancer Res 42(11): 4490-4. Bodenner, 1986 #733; Saran, M. and Bors, W. (1990). "Radical reaction in vivo- -an overview." Radiat. Environ. Biophys. 29(4): 249-62.
Недавно использованным при лечении алкоголизма дитиокарбаматом является дисульфирам, димер диэтилдитиокарбамата. Дисульфирам ингибирует альдегиддегидрогеназу печени. Inoue, К. and Fukunada, et al., (1982). "Effect of diculfiram and its reduced metabolite, diethyldithiocarbamate on aldehyde dehydroqenase of human erythrocytes." Life Sci 30(5): 419-24.
Сообщалось, что дитиокарбаматы ингибируют репликацию вируса ВИЧ, а также ускоряют созревание субпопуляции специфических Т-клеток. Это было показано в клинических испытаниях диэтилдитиокарбамата у больных СПИДом. Reisinger, E., et al., (1990). "Inhibition of HIV progression by dithiocarb." Lancet 335: 679.
Дитиокарбоксилаты являются соединениями структуры А-SС(S)-В, которые являются членами общего класса соединений, известных как тиоловые антиоксиданты, и которые альтернативно указаны как карбодитиолы или карбодитиолаты. Кажется, что радикал -SС(S)- является существенным для проявления терапевтической активности и что А и В могут быть любой группой, которая не проявляет отрицательного действия на эффективность или токсичность соединения.
При альтернативном осуществлении один или оба атома серы дитиокарбамата замещены атомом селена. Замещение серы на селен может понизить токсичность молекулы в некоторых случаях и, таким образом, может быть более толерантным для пациента.
А и В могут быть выбраны специалистом в данной области таким образом, чтобы придать желаемые свойства соединению, включая размер, заряд, токсичность и степень стабильности (включая стабильность в кислой среде, такой как в желудке, или в основной среде, такой как в кишечном тракте). Выбор А и В также будет иметь большое значение для распространения в тканях и фармакокинетики соединения. Обычно для лечения сердечно-сосудистого заболевания желательно, чтобы соединение накапливалось или локализовалось в интиме артерии, содержащей эндотелиальные клетки сосудов. Соединение предпочтительно удаляется путем экскретирования почками.
Преимуществом фармацевтического использования дитиокарбоксилата является то, что оно, как кажется, не подвергается ферментному расщеплению in vivo и поэтому может проявлять пролонгированный период полувыведения in vivo.
При предпочтительном осуществлении А является водородом или фармацевтически приемлемым катионом, включая, но не ограничиваясь ими, натрий, калий, кальций, магний, алюминий, цинк, висмут, барий, медь, кобальт, никель или кадмий; соль образующей органической кислотой, обычно карбоновой кислотой, включая, но не ограничиваясь ими, уксусную кислоту, щавелевую кислоту, виноградную кислоту, янтарную кислоту, яблочную кислоту, аскорбиновую кислоту, бензойную кислоту, танниновую кислоту, памоевую кислоту, альгиновую кислоту, полиглутаминовую кислоту, нафталинсульфоновую кислоту, нафталиндисульфоновую кислоту или полигалактуроновую кислоту; или катионом, образованным аммонием или другим азотным основанием, включая, но не ограничиваясь ими, азотсодержащий гетероцикл или радикал формулы NR4R4R5R6R7, где R4, R5, R6 и R7, независимо, обозначают водород, C1-6линейный разветвленный или (в случае C4-6) циклический алкил, гидрокси(C1-6)алкил (где одна или несколько гидроксильных групп находятся у любого углеродного атома) или арил, N,N-дибензилэтилендиамин, D-глюкозамин, холин, тетраэтиламмоний или этилендиамин.
При другом осуществлении А может быть физиологически отщепляемой уходящей группой, которая может быть отщеплена in vivo от молекулы, к которой она присоединена, и включает, но не ограничивается ими, ацил (включая ацетил, пропионил и бутирил), алкил, фосфат, сульфат или сульфонат.
При одном осуществлении В представляет алкил, алкенил, алкинил, алкарил, аралкил, галогеналкил, галогеналкенил, галогеналкинил, арил, алкарил, водород, C1-6алкокси-C1-11алкил, С1-6алкилтио-С1-10алкил, NR2R3, -(CHOH)nСН2ОН, где n равно 0, 1, 2, 3, 4, 5 или 6, -(CH2)nСО2R1, включая алкилацетил, алкпропионил и алкилбутирил, или гидрокси(С1-6)алкил- (где одна или несколько гидроксильных групп расположены у любого из атомов углерода).
При другом осуществлении В обозначает NR2R3, где R2 и R3, независимо, обозначают алкил; -(СНОН)nСН2ОН, где n равно 0, 1, 2, 3, 4, 5 или 6; -(СН2)nCO2R1, -(СН2)nСO2R4, гидркоси(C1-6)алкил-; алкенил, включая, но не ограничиваясь ими, винил, аллил и СН3СН=CН-СН2СН2; алкил(СО2Н), алкенил(СО2Н), алкинил(СО2Н) или арил, где арильная группа может быть замещена, как описано выше, в частности, например, замещена NО2-, СН3, трет-бутилом, СО2Н, галогеном или п-ОН группой; или R2 и R3 могут вместе образовывать мостик, такой как -(CH2)m-, где m равно 3, 4, 5 или 6 и где R4 обозначает алкил, арил, алкарил или аралкил, включая ацетил, пропионил и бутирил.
При еще одном осуществлении изобретения В может быть гетероциклической или алкилгетероциклической группой. Гетероцикл может быть необязательно частично или полностью гидрированным. Неограничивающими примерами являются перечисленные выше, включая феназин, фенотиазин, пиридин и дитиопиридин.
При еще одном осуществлении В является остатком фармацевтически активного соединения или лекарственного средства. Термин лекарственное средство, как здесь используется, относится к любому веществу для внутреннего или наружного использования в качестве лечебного препарата для обработки, лечения или профилактики заболевания или поражения.
Неограничивающими примерами являются лекарственные средства для лечения или профилактики сердечно-сосудистого заболевания, включая антиоксиданты, такие как пробукол; никотиновая кислота; средства, которые предохраняют кровяные пластинки от слипания, такие как аспирин; антитромбоцитарные средства, такие как кумадин; блокаторы кальциевых каналов, такие как веропамил, дилтиазем и нифедипин; ингибиторы ангиотензинпревращающего фермента (АСЕ), такие как каптоприл и иналоприн, β-блокаторы, такие как пропанолол, тербуталол и лабеталол, нестероидные противовоспалительные препараты, такие как ибупрофен, индометацин, фенопрофен, мефенамовая кислота, флюфенамовая кислота, сулиндак, или кортикостероиды. Группа -С(S)SА может быть непосредственно присоединена к лекарственному средству либо присоединена через какой-нибудь подходящий связующий радикал.
При другом осуществлении дитиокарбамат является производным аминокислоты структуры AO2C-R9-NR10-С(S)SА, где R9 представляет двухвалентный радикал В, связующий радикал или внутренний остаток каких-либо природных аминокислот (например, СН3СН для аланина, СН2 для глицина, СH(СН2)4NH2 для лизина и т.д.) и R10 обозначает водород или низший алкил.
В также может быть полимером, к которому присоединены одна или несколько дитиокарбоматных групп либо непосредственно, либо через какой-либо подходящий связующий радикал. Дитиокарбамат высвобождается, предпочтительно, из полимера в условиях in vivo в течение подходящего периода времени с достижением терапевтического эффекта. При предпочтительном осуществлении полимер сам по себе также способен разрушаться in vivo. Термин биоразрушающийся или биодеградирующий, как здесь используется, относится к полимеру, который растворяется или разрушается в течение периода времени, которое приемлемо при желаемом использовании (обычно терапия in vivo), обычно менее чем за пять лет, и предпочтительно менее чем за один год, при выдерживании в физиологическом растворе с рН 6-8, имеющим температуру между 25 и 37°С. При предпочтительном осуществлении полимер разрушается в течение периода между одним часом и несколькими неделями, в соответствии со способом применения.
Известно множество способных разрушаться полимеров. Неограничивающими примерами являются пептиды, белки, нуклеопротеины, липопротеины, гликопротеины, синтетические и природные полипептиды и полиаминокислоты, включая, но не ограничивается ими, полимеры и сополимеры лизина, аргинина, аспарагина, аспартамовой кислоты, цистеина, цистина, глутамовой кислоты, глутамина, гидроксилизина, серина, треонина и тирозина; полиортоэфиры, включая поли(α-гидроксикислоты), например полимолочная кислота, полигликолевая кислота, поли(лактид-согликолид), полиангидриды, альбумин или коллаген, полисахаридсодержащие сахарные звенья, такие как лактоза, и поликапролактон.
Полимер может быть любым или блок-сополимером.
В также может быть группой, которая повышает растворимость дитиокарбамата в воде, например -низший алкил-O-R8, где R8 представляет -PO2(OH)-M+ или PO3(M+)2, где М+ обозначает фармацевтически приемлемый катион; -С(O)(СН2)
При альтернативном осуществлении может быть введен димер, такой как В-С(S)S-SC(S)-B. Неограничивающими примерами дитиокарбаматов являются дитиокарбаматы следующей структуры: 
Для применения при лечении атеросклероза и других сердечно-сосудистых и противовоспалительных заболеваний следует выбирать такие дитиокарбаматы, которые обладают необходимой липофильностью для того, чтобы локализоваться на участке действия. Соединение не должно проникать в области низкого обмена веществ, такие как жировые отложения. При предпочтительном осуществлении для лечения сердечно-сосудистого заболевания на фармакокинетику не должны существенным образом влиять хроническая сердечная недостаточность и почечная недостаточность.
Для наружного применения при лечении воспалительных заболеваний кожи выбранное соединение должно входить в состав препарата, адсорбируемого кожей в достаточном количестве для обеспечения терапевтического эффекта на пораженном участке.
Дитиокарбоксилат должен быть физиологически приемлемым. Обычно приемлемыми являются соединения с терапевтическим индексом, по меньшей мере, 2 и предпочтительно, по меньшей мере, 5 или 10. Терапевтический индекс определяется как EC50/IC50, где ЕС50 является концентрацией соединения, которая ингибирует экспрессию VCAM-1 на 50%, и IС50 является концентрацией соединения, которая токсична на 50% по отношению к клеткам-мишеням. Клеточная токсичность может быть измерена прямым обсчетом клеток, исключенных трепановым синим, или различными исследованиями метаболической активности, такими как включение 3Н-тимидина, известных специалистам в данной области. Терапевтический индекс ПДТК в культуре тканей выше 100, как определено клеточной токсичностью, деленной на способность ингибировать экспрессию VCAM-1, активированную TNFa, в клетках HUVE. Первоначальные исследования по быстро разрастающейся человеческой глиоме клеточного типа NT18 не демонстрирует токсичность при концентрациях, 100-кратно превышающих терапевтическую концентрацию.
Дисульфирам, перорально назначаемая форма диэтилдитиокарбамата, используемый при лечении алкоголизма, обычно не проявляет в основном клиническую токсичность при соответствующем введении.
Известно небольшое число дитиокарбаматов, которые генотоксичны, эти соединения не входят в объем настоящего изобретения, которое ограничено использованием физиологически приемлемых веществ. Примером генотоксичного дитиокарбамата является фунгицид диметилдитиокарбамат цинка. Кроме того, антихолинэстеразные свойства некоторых дитиокарбаматов могут привести к нейротоксическим эффектам. Miller D. (1982). Neurotoxisity of the pesticidal carbamates. Neurobehav. Toxicol. Teratol. 4(6): 779-87.
Термин дитиокарбоксилат, как здесь используется, особенно включает, но не ограничивается ими, дитиокарбаматы формул: R1SС(S)NR2R3 или R2R3N(S)СS-SС(S)NR2R3, где R1 обозначает H или фармацевтически приемлемый катион, включая, но не ограничивается ими, натрий, калий или NR4R5R6R7, где R4, R5, R6 и R7, независимо, обозначают водород, C1-6 линейный, разветвленный или циклический алкил, гидрокси(C1-6)алкил (где одна или несколько гидроксильных групп находятся у любого углеродного атома) или арил, и R2 и R3, независимо, обозначают С1-10 линейный, разветвленный или циклический алкил; -(СНОН)nСН2ОН, где n равно 0, 1, 2, 3, 4, 5 или 6; -(СН2)nСО2R1, -(CH2)n CO2R4, гидрокси(С1-6)алкил-; или R2 и R3 могут вместе образовывать мостик, такой как -(СН2)m-, где m равно 3-6 и где R4 обозначает алкил, арил, алкарил или аралкил, включая ацетил, пропионил и бутирил.
Конкретные примеры используемых дитиокарбаматов, показанные на фиг.16, включают, но не ограничиваются ими, пирролидин-N-карбодитиоат натрия, N-метил-N-карбоксиметил-N-карбодитиоат натрия, N,N-ди (карбоксиметил)-N-карбодитиоат тринатрия, N-метил-D-глюкамин-N-карбодитиоат натрия, N,N-диэтил-N-карбодитиоат натрия (диэтилдитиокарбамат натрия) и N,N-диизопропил-N-карбодитиоат натрия.
Активные дитиокарбоксилаты, и, в частности, дитиокарбаматы либо коммерчески доступны, либо могут быть получены известными способами.
II. Биологическая активность
Способность дитиокарбаматов ингибировать экспрессию VСAM-1 можно определить различными путями, включая способ, описанный в подробностях ниже, в примерах от 9 до 15. Для удобства, примеры 9-11 и 14-15 описывают оценку биологической активности натрий пирролидин-N-карбодитиоата (также обозначенного как РDТС). Данные примеры не ограничивают объем изобретения, но специфически включают использование вышеописанных соединений для лечения атеросклероза и других типов воспалительных и сердечно-сосудистых заболеваний, опосредованных VCAM-1. Любое из соединений, описанных выше, может быть заменено вместо PDTC и оценено аналогичным образом.
Примеры 12 и 13 описывают сравнительные данные по способности некоторых дитиокарбаматов ингибировать экспрессию гена VCAM-1. Примеры ниже устанавливают, что заявляемые дитиокарбаматы специфически подавляют способность VCAM-1 экспрессироваться эндотелиальными клетками сосуда в ответ на многие известные сигналы, активные при атеросклерозе и воспалительных заболеваниях.
Экспериментальные способы
Культуры клеток. Клетки HUVE выделяли из пупочных вен, в которые вводили канюли, перфузировали раствором Хенкса для удаления крови и затем инкубировали с 1% коллагеназой в течение 15 минут при 37°С. После удаления коллагеназы клетки культивировали в среде М199, дополненной 20% эмбриональной бычьей сывороткой (HyC1oпe), 16 мкг/мл гепарина (ESI Pharmaceuticals, Cherry Нill, nj), 50 мкг/мл добавки для роста эндотелиальных клеток (Collaborative Research Jncorporated, Bedford, MA), 25 мМ буфера Нереs, 2 мМ L-глютамина, 100 мкг/мл пенициллина и 100 мкг/мл стрептомицина, и выращивали при 37°С на чашках для выращивания клеток, покрытых 0,1% желатином. Клетки пересаживали в конфлюентом состоянии путем распределения 1:4. Использовали клетки первых 8 пересевов.
Инкубация с цитокинами и другими реагентами
Конфлюентные клетки HUVE промывали фосфатным буфером на физиологическом растворе и затем добавляли свежую среду. Добавляли отмеченные концентрации РDТС в виде предварительной обработки перед добавлением цитокинов. Цитокины и другие индукторы непосредственно добавляли в среду в определенное время и в концентарциях, указанных для каждого эксперимента. Рекомбинантный IL-2b человека представлял любезный дар Upiohn Соmpany (Kalamazoo, Michigan). TNF-α получили от Boehringer Engelheim). Бактериальный липополисахарид (LPS), полиинозиновую кислоту:полицитидиновую кислоту (Поли I:С) и пирролидин дитиокарбамат (РDТС) получили от Sigma Chemical (St.Zouis, MО). Все другие реактивы представляли соответствующую степень чистоты для реактивов.
Выделение РНК. Общую клеточную РНК выделяли путем экстракции, используя подкисленную смесь гуанидинтиоцианата-фенола-хлороформа. Клетки промывали фосфатным буфером на физиологическом растворе и затем лизировали 2 мл гуанидинизотиоцианата. Раствор подкислили 0,2 мл ацетата натрия (рН 4,0) и затем экстрагировали 2 мл фенола и 0,4 мл смеси хлороформ:изоамиловый спирт (24:1). РНК подвергали двум осаждениям этанолом перед использованием для Northern blot анализа.
Northern blot анализ. Общую ДНК клетки (20 мкг) фракционировали по размеру, используя 1% агарозаформальдегидный гель в присутствии 1 мкг/мл этидиум бромида. РНК переносили на нитроцеллюлозный фильтр и ковалентно пришивали путем облучения ультрафиолетом, используя перекрестный сшиватель Stratlinker UV (Siratagene, Za Jolla, CA).
Гибридизацию проводили при 42°С в течение 18 часов в 5Х (кратной) SSС (1Х=150 мМ NaCl, 15 мМ цитрат Nа), 1% додецилсульфат натрия, 5Х раствор Denhardt, 50% формамид, 10% декстран сульфат и 100 мкг/мл порезанной денатурированной ДНК спермы лосося. На гибридизацию использовали приблизительно 1-2·106 срm/мл меченого зонда (специфическая активность >108 cpm/мкг ДНК). После гибридизации фильтры промыли конечным раствором ионной силы 0,2Х при 55°С. Нитоцеллюлозу отмыли, используя кипящую воду, перед повторной гибридизацией с другими пробами. Авторадиографию проводили с интесифицирующим экраном при -70°С.
32Зонды. 32Р меченые зонды ДНК приготовили, используя способ случайных олигонуклеотидных праймеров. Зонд ICAM-1 представлял фрагмент Eco R1 кДНК человека. Зонд ELAM-1 представлял фрагмент 1,85 кб Нind III кДНК человека. Зонд VCAM-1 представлял фрагмент Нind III-Xho I кДНК человека, включающий нуклеотиды от 132 до 1814.
Ферментный иммуносорбентный тест
(ELISA). Клетки HUVE помещали на микротитрационное плато с 96 ячейками на 48-72 часа до исследования. Добавляли первичное антитело в М199 с 5% FBS к каждой ячейке и инкубировали один час конъюгатом козьего антимышиного IqG с пероксидазой (Вio Rad), разбавленного 1/500 в М199 с 5% FBS. Затем ячейки промывали и определяли связывание антитела при добавлении 100 мкл раствора 10 мг/мл 3,3,5,5'-тетраметилбензидина (Sigma) в присутствии 0,003% Н2О2. Реакцию останавливали путем добавления 25 мкл 8 H серной кислоты. Ячейки считывали в считывающем устройстве (ридере) ЕLISА (Вiо Rad) при О.П. 450 нм, после определения бланка в ряду, прокрашенного только антителом на второй стадии. Данные представляют средние из трипликатов.
Антитела. Моноклональное антитело (Mab) 4B9, распознающее молекулу адгезии-1 клеток сосуда (VCAM-1), было поставлено Dr. John Harlan (University of Wаshington). Антитела, распознающие молекулу адгезии эндотелиальных клеток (ELAM-1), поставлялись Dr.Schwerlick (Emory University) Гибридомы, продуцирующие mАB 84Н10, распознающие внутриклеточную молекулу адгезии-1 (ICAM-1), обычно выращивались в лаборатории, и использовали супернатант клеточной культуры в качестве антитела.
Пример 9
РDТС блокирует опосредованную 1L-2b индукцию VCAM-1 в HUVEC, но не аккумуляцию мРНК ICAM-1 или ELAM-1.
Для определения, действительно ли окисленное состояние энотелиальных клеток может изменять базальную и индуцированную экспрессию гена молекулы клеточной адгезии, культивируемые эндотелиальные клетки сосудов человека выдерживали с цитокином индукции IL-1b (10 ед/мл) в присутствии или в отсутствие антиоксидантов, хелатирующих тиолированный металл, пирролидин дитиокарбамат (РDТС, 50 мкМ) вплоть до 24 часов. Как показано на фиг.11, один 1L-2b (линии 2, 4, 6, 8) вызывает ожидаемую быструю и преходящую индукцию мРНК VCAM-1 (Панель А), Е-селектина (ELAM-1, Панель В) и ICAM-1 (Панель С), все пики на четыре часа. Однако в присутствии РDТС индукция аккумуляции мРНК VCAM-1 под действием 1L-2b драматически ингибируется более чем на 90% (Панель А, линии 3, 5, 7, 9). Напротив, несмотря на то что 1L-2b опосредованная индукция ЕLAМ-1 немного ингибируется на 2 и 24 часа (сравните линию 2 и 3, 8 и 9, Панель В), РDТС не ингибирует индукцию на 4 и 8 часов (линия 5 и 7, Панель В), 1L-2b, опосредованная индукция аккумуляции мРНК ICAM-1 не изменяется (панель В, линии 3, 5, 7, 9). Несомненно, наблюдается умеренное угнетение индукции аккумуляции мРНК ICAM-1 под действием 1L-2b (-30%) (сравните линии 4 и 5, панель В). Эквивалентные количества перенесенной РНК на целлюлозу на линию подтверждается окрашиванием этидиум бромидом и визуализацией.
Для определения, действительно ли РDТС ингибирует индукцию экспрессии гена VCAM-1 под действием 1L-2b дозозависимым образом, был проведен анализ дозовой зависимости. Как показано на фиг.12, РDТС ингибирует 1L-2b опосредованную индукцию экспрессии гена VCAM-1 последовательно дозозависимой кривой (фиг.12, панель А), с рассчитанной ЕС50 приблизительно 10 мкМ, несмотря на то что РDТС не ингибирует 1L-2b опосредованную индукцию экспрессии ELAM-1 в тех же концентрациях (фиг.12, панель В). 1L-2b опосредованная индукция аккумуляции мРНК ICAM-1 усиливается под действием РDТС при концентрациях выше, чем 0,5 мкМ (фиг.13, сравните линию 2 и линию 4-7, панель С).
Эти данные показывают, что 1L-2b использует дитиокарбоксилат и, в частности, стадию, чувствительную к дитиокарбамату, в качестве своего сигнального механизма при индукции экспрессии гена VСАМ-1. Данные, по-видимому, также показывают, что данная дитиокарбаматчувствительная стадия не играет существенной роли в IL-2b опосредованной индукции эспрессии гена ELAM-1 или ICAM-1.
Пример 10
РDТС блокирует индукцию аккумуляции мРНК VCAM-1 HUVEC под действием множественных стимулов
Для определения, действительно ли другие хорошо описанные активаторы экспрессии гена VCAM-1 также используют чувствительную к РDТС стадию, были протестированы три различных класса активаторов: классический, опосредуемый рецептором другой индуцирующий агент (TNF-α), неопосредуемый рецептором индуктор (липополисахарид (LPS)) и недавно описанный новый индуктор (двухнитевая РНК, поли(I:С). Во всех трех случаях РDТС ингибировал индукцию аккумуляции мРНК VCAM-1 в HUVEC через четыре часа (фиг.13, панель А). Несмотря на то что опосредованная ТNF-α экспрессия гена VCAM-1 подавляется в той же степени (фиг.13, линия 1 и 2, панель В), опосредованная LРS и поли(I:С) аккумуляция мРНК VCAM-1 была неизменной (фиг.13, линия 3-6, панель В). Индукция аккумуляции мРНК ICAM-1 была неизменной (фиг.13, Панель С). Эти данные показывают, что структурно различные индуцируемые гены, действующие через различные пути, объединяет общая регуляторная стадия, специфическая для индукции экспрессии гена VCAM-1.
Пример 11
РDТС блокирует экспрессию VCAM-1 на клеточной поверхности HUVЕ, индуцированную множественными стимулами
Для определения, действительно ли, подобно своей мРНК, индукция экспрессии на поверхности эндотелиальной клетки белка VCAM-1 также может быть заингибирована РDТС, в определении ELISА были использованы моноклональные антитела для количественного определения индукции на поверхности клеток VСАМ-1 и ICAM-1 у клеток HUVE в культуре. Как показано на фиг.14, множественные классы активирующих агентов в отсутствие РDТС (-РDТС) индуцируют быструю и временную аккумуляцию VCAM-1 (вверху левой панели) на клеточной поверхности с максимумом на шесть часов. В присутствии РDТС (+РDТС, вверху правой панели) индукция экспресси VCAM-1 на поверхности клеток под действием всех тестируемых агентов драматически заингибирована (80-90%). Напротив, индуцированная экспрессия ICAM-1 на поверхности клеток не изменяется в идентичных условиях (внизу левая и правая панели).
Эти данные свидетельствуют, что подобно аккумуляции собственной мРНК экспрессия VCAM-1 на поверхности клетки селективно подавляется под действием дитиокарбаматов и что множественные классы активирующих агентов используют подобный, чувствительный к дитиокарбамату механизм индукции экспрессии гена VCAM-1.
Пример 12
Сравнительная эффективность антиоксидантов по блокированию индукции VCAM-1 под действием TNF-α
Для определения, действительно ли структурно подобные или не подобные антиоксиданты могли бы также ингибировать экспрессию гена VCAM-1 и с какой эффективностью, клетки HUVE выдерживали с TNF-α в течение шести часов в присутствии или отсутствии различных концентраций четырех различных антиоксидантов. Как показано на фиг.15, оба диэтилдитиокарбамата (DЕТС) и N-ацетилцистеин (NАС) ингибировали экспрессию VCAM-1 в концентрациях 5 мкМ и 30 мкМ соответственно. Напротив, PDTC (PDTC) был эффективен между 5 и 50 мкМ. Хелатор металла железа, десферроксимин, не обладал действием в исследованных концентрациях.
Пример 13
РDТС ингибирует индуцированную ТNF, опосредованную VCAM-1/VLA-4 адгезию
Способность различных антиоксидантов ингибировать индукцию VCAM-1 в клетках HUVE под действием ТNF-α определяли по способу, описанному в примере 12. Фиг.16 представляет график относительной экспрессии VСАМ-1 на поверхности клеток (О.П. 595 нм) у клеток HUVE, активированных ТNF-α, относительно концентраций РDТС (натрий N-пирролидин дитиокарбамат), DIDTC (натрий N,N-диэтил-N-карбодитиоат), SarDTC (натрий N-метил-N-карбоксиметил-N-карбодитиоат), IDADC (тринатрий N,N-ди(карбоксиметил)-N-карбодитиоат), MGDTC (натрий N-метил-D-глюкамин-N-карбодитиоат), MeOBGDTC (натрий N-(4-метоксибензил) -D-глюкамин-N-карбодитиоат), DЕDТС (натрий N,N-диэтил-N-карботиоат), Di-PDTC (натрий N,N-диизопропил-N-карбодитиоат), и NАС представляет (N-ацетилцистеин).
Пример 13
РDTC ингибирует индукцию под действием TNF опосредованной адгезии VCAM-1/VLA-4
Для того чтобы определить, действительно ли ингибирование регуляции VCAM-1 под действием РDТС связано с функциональной последовательностью событий, было исследовано связывание клеток Molt-4 с клетками HUVEC, либо не стимулированных, либо стимулированных TNFa (100 ед/мл) в течение шести часов в присутствии или отсутствие PDTC. Клетки Molt-4, как было ранее показано, связываются с активированными HUVEC посредством VСАМ-1 зависимого механизма. Как показано на фиг.17, процент связывания Molt-4 с клетками HUVEC снижался, когда PDTC присутствовал в среде.
Пример 14
РDТС ингибирует связывание моноцитов с брюшной аортой кроликов, находящихся на холестериновой диете
Эксперимент проводили на животных на экспериментальной модели для определения, действительно ли тиольный антиоксидант РDТС мог бы быть эффективным в блокировании связывания первичного компонента моноцитов при атеросклерозе. Взрослый новозеландский белый кролик (3,5 кг) получал внутривенные инъекции РDТС (20 мг/кг, в виде раствора, концентрации 20 мг/мл в РВS) ежедневно в течение 5 дней. Инъекции осуществляли через вживленную канюлю в краевую ушную вену, которую сохраняли незакупоренной путем промывания гепаринизированным физиологическим раствором. Раствор РDТС готовили или ежедневно или в предыдущий день (хранили защищенным от света при 4°С) и фильтровали (фильтр с порами 0,45 мм) непосредственно перед употреблением. Через вживленную канюлю, после первой инъекции, препарат вводили кролику в здоровом состоянии в отсутствие дискомфорта или другого болезненного эффекта. На второй день инъекций кролик получал питание, содержащее 1% холестерина по весу, которое продолжали давать в течение оставшегося периода эксперимента. На пятый день животное умерщвляли, вырезали и фиксировали брюшную аорту. После соответствующей подготовки образцы отображали на низкой стадии на электронном сканирующем микроскопе ISI-130, снабженным LaB эмиттером. Используя изображения на двух экранах и прозрачную сетку на экране СRT, исследовали 64 примыкающих поля при 620х увеличении, перекрывая область в 1,3 мм2. Внутри каждого поля подсчитывали и регистрировали количество слипшихся лейкоцитов (WBC) и эритроцитов (RBC).
Данные из образца свода следующие: 5 WBC и приблизительно 25 RBC на 1,3 мм2 поля. Данный уровень адгезии WBC является близким к контрольным животным, получающих обычное питание (было обнаружено около 7 случаев на поле в образцах свода и брюшного отдела из 2 ‘отрицательных контрольных’ экспериментов). ‘Положительные контрольные' кролики, получавшие 1% холестерина в течение 4 дней, но не получавшие антиоксидант, проявляли увеличение адгезии в 5 раз, до 38 WBS/1,3 мм2. Наблюдали, в основном, большое количество волокон размером с клетку, прилипших к каждому образцу свода. Неясно, действительно ли этот материал является артефактом подготовки или присутствовал in vivo и, если это так, имеет ли это отношение к введению РDТС. Данные исследования подтверждают, что введения РDТС могут эффективно блокировать начальное прилипание моноцитов к эндотелию аорты.
Пример 15
Ингибирование ВSА продуктов присоединения 13-НРОDЕ с РDТС
Фиг.19 представляет графическую диаграмму действия РDТС на образование флуоресцентных продуктов присоединения BSА с 13-HPODE, что определено в единицах флуоресценции в зависимости от микромолярной концентрации РDТС. Один микромоль 13-HPODE инкубировали с 200 микрограммами ВSА в присутствии РDТС в течение шести дней. Флуоресценцию измеряли при 430-460 нм при возбуждении при 330-360 нм. Для деталей определения см. Freebs, J., Parthasarathy, S., Steinberg, D., Prосееdings of the National Academy оf Sciences 89, 10588-10592, 1992. В ходе обычной реакции 100 нмолей LOOH (полученных при помощи липооксигеназы, катализирующей окисление линолевой кислоты) инкубировали с 100 мкг бычьего сывороточного альбумина в течение 48-72 часов и измеряли последующее образование флуоресцентных продуктов путем определения спектра флуоресценции при возбуждении при 360 нм и эмиссии между 390 и 500 нм.
Как отмечено, РDТС снижает концентрацию флуоресцентных продуктов присоединения BSA и 13-HPODE.
Фиг.20 представляет график влияния РDТС на образование флуоресцентных продуктов присоединения ВS А и ox-PUFA в виде функции длины волны (нм) и концентрации РDТС. С увеличением концентрации РDТС количество флуоресцентных продуктов присоединения снижается.
Пример 16
Влияние РDТС на окисление ЛНП под действием пероксидазы хрена
Фиг.21 представляет собой график действия РDТС на окисление ЛНП под действием пероксидазы хрена (HRP), измеряемое во времени (минуты) при различных концентрациях РDТС. Окисление ЛНП проводили путем измерения жирнокислотных компонентов ЛНП по определению увеличения оптической плотности при 234 нм. Когда окисляются полиненасыщенные жирные кислоты, наблюдается сдвиг в двойной связи, приводящий к образованию конъюгированных диенов, которые обладают поглощением при 234 нм. Задержка инициирования и развития кривой (лаг-фаза) подтверждает, что происходит измерение окислительной способности липидов. Более продолжительная лаг-фаза, следовательно, большая устойчивость ЛНП к окислению. Обычно инкубируют 100 мкг ЛНП человека с 5 мкМ Н2О2, и затем следует увеличение поглощения при 234 нм.
Было обнаружено, что после инкубационного периода РDТС ингибирует окисление ЛНП под действием HRP концентрационно-зависимым образом.
Пример 17
Влияние РDTС на индуцированное цитокином образование ох-РUFА
Фиг.22 представляет диаграмму влияния РDТС на индуцированное цитокином образование ох-РUFA в эндотелиальных клетках аорты человека. Как отмечается, и ТNF-α, и IL-2b вызывают окисление линолевой кислоты до окисленной линолевой кислоты. Окисление значительно приостанавливается под действием PDTC.
2. Модификация синтеза и метаболизм РUFА и ох-РUFА
Ингибирование экспрессии VCAM-1 может быть завершено путем модификации метаболизма PUFA в ох-РUFA. Многие известные ферменты окисляют ненасыщенные материалы, включая, например, пероксидазы, липооксигеназы, циклооксигеназы и цитохром Р450. Ингибирование данных ферментов может предотвращать окисление PUFA in vivo. PUFAS могут быть также окислены под действием металлозависимых неферментных материалов.
IV. Способ модификации экспресси генов, чувствительных к окислению-восстановлению
В альтернативном воплощении описывается способ подавления экспрессии генов, чувствительных к окислению-восстановлению, или активации гена, который подавляется посредством пути, чувствительного к окислению-восстановлению, который включает введение эффективного количества вещества, которое предотвращает окисление окисляемого сигнала, и обычно, окисление полиненасыщенной жирной кислоты. Характерные гены, чувствительные к окислению-восстановлению, которые участвуют в развитии иммунного ответа, включают, но не ограничиваются теми экспрессируемыми цитокинами, участвующими в инициации иммунного ответа (например, IL-1B), хемоаттрактантами, которые способствуют миграции клеток воспаления к точке повреждения (например, МСР-1), факторами роста (IL-6, рецептор тромбина) и молекулы адгезии (например, VCAM-1 и Е-селектин).
Имея настоящее изобретение, квалифицированный работник будет способен протестировать широкое разнообразие антиоксидантов по их способности подавлять экспрессию гена, чувствительного к окислению-восстановлению, или активацию гена, который подавляется через чувствительный к окислению-восстановлению путь. Все данные воплощения входят в объем настоящего изобретения.
Основываясь на результатах настоящего тестирования, можно идентифицировать молекулы нуклеиновых кислот, содержащие 5’-регуляторные последовательности генов, чувствительных к окислению-восстановлению, которые могут быть использованы для регуляции или ингибирования экспрессии генов in vivo, Векторы, включающие и плазмиду, и эукариотические вирусные векторы могут быть использованы для экспрессии конкретной конструкции 5'-фланкирующей области рекомбинантного гена в клетках в зависимости от предпочтения и суждения квалифицированного сотрудника (см., например, Sambrook et al., Chapter 16). Более того, были разработаны многие вирусные и невирусные векторы, которые способны к внедрению в последовательности нуклеиновой кислоты in vivo (см., например, Mulligan, 1993, Science, 260, 926-932; Патент США № 4980286; Патент США № 4868116, включенных здесь в качестве ссылки). Недавно была разработана система доставки, в которой нуклеиновая кислота энкапсулирована в катионные липосомы, которые можно внутривенно вводить млекопитающему. Данная система была использована для внедрения ДНК в клетки многих тканей взрослой мыши, включая эндотелий и костный мозг (см., Zhu еt аl., 1993, Sсiеnсе 261, 209-211, включена здесь в качестве ссылки).
5’-фланкирующие последовательности гена, чувствительного к окислению-восстановлению, могут быть использованы для ингибирования экспрессии гена, чувствительного к окислению-восстановлению. Например, антисенсная РНК всей или части 5'-фланкирующей регион последовательности гена, чувствительного к окислению-восстановлению, может быть использована для ингибирования экспрессии гена in vivo. Векторы экспрессии (например, ретровирусные векторы экспрессии) уже доступны в технике, можно использовать для получения антисенсной РНК, выбранной последовательности ДНК, которая экспрессируется в клетке (см., например, Патент США № 4868116; Патент США № 4980286). Соответственно, ДНК, содержащая всю часть последовательности 5’-фланкирующей области гена, может быть внедрена в подходящий вектор экспрессии так, чтобы при введении в клетку транскрипция внедренной ДНК давала антисенсную РНК, которая комплементарна к транскрипту мРНК гена обычно обнаруживаемого гена. Данный транскрипт антисенсной РНК по внедренной ДНК может затем образовать пары оснований с нормальным транскриптом мРНК, обнаруживаемым в клетках, и тем самым препятствовать подлежащей трансляции мРНК. Конечно, необходимо выбрать 5’-последовательность фланкирующего региона, которая располагается по направлению от транскрипционного начального сайта для гена, чувствительного к окислению-восстановлению, чтобы подтвердить, что антисенсная РНК содержит комплементарные последовательности, присутствующие в мРНК. Антисенсная РНК может быть получена также in vitro и, затем, внедрена в клетки. Олигонуклеотиды могут быть синтезированы на автоматическом синтезаторе (например, модель 8700 автоматического синтезатора Milligеn-Biosearch, МА или ABI Модель 380В). Помимо этого было показано, что антисенсные дезоксинуклеотиды являются эффективными при ингибировании транскрипции гена и вирусной репликации (см., например, Zamecnik et al., 1978, Proc. Natl. Acad. Sci. USA, 75, 280-284; Zamecnik еt al., 1986, Proc. Natl. Acad. Sci. USA, 83, 4143-4146; Wickstrom et al., 1988, Proc. Natl. Acad. Sci. USA, 85, 1028-1032; Crooke, 1993, FASEB J. 7, 533-539. Более того, в недавней работе показано, что возможно усиленное ингибирование экспрессии гена под воздействием антисенсных нуклеотидов, если антисенсные олигонуклеотиды содержат модифицированные нуклеотиды (см. Offensperger et al., 1993 ЕМВО J, 12, 1257-1262 (ингибирование вирусной репликации гепатита В и экспрессии гена in vivo посредством антисенсных фосфоротиоатных олигонуклеотидов); Rosenberg еt al., PST WO 93/01286 (синтез серотиоатных олигонуклеотидов); Agrawal et аl., 1988, Prос. Natl. Acad. Sci, USA 85, 7079-7083 (синтез антисенсных фосфорамидатных фосфоротиоатных нуклеотидов для ингибирования репликации вируса-1 иммунодефицита человека); Sarin et al., 1989, Proc. Natl. Acad. Sci. 85, 7448-7794 (синтез антисенсных метилфосфонатных олигонуклеотидов); Show et al., 1991, Nисleiс Acids Res, 19, 747-750 (синтез устойчивых к экзонуклеазе 3’-олигонуклеотидов, содержащих 3’-конечные фосфороамидатные модификации); включенные здесь в качестве ссылок).
Последовательность 5’-фланкирующей области гена, чувствительного к окислению-восстановлению, может быть также использована в трехспиральной (триплексной) генной терапии. Показано, что олигонуклеотиды, комплементарные к последовательности промотера гена одной из нитей ДНК, связывают промотер и регуляторную последовательность с образованием триплексной нуклеиновой кислоты, которая блокирует транскрипцию гена (см., например, 1989 Maher et al., Science 245, 725-730; Orson et al., 1991 Nucl. Asids Res. 19, 3435-3441; Postal et al., 1991 Proc. Natl. Acad. Sci. USA 88, 8227-8231; Cooney et al., 1988 Sciece 241, 456-459; Young et al., 1991 Proc. Natl. Acad. Sci. USA 88, 10023-10026; Duval-Valentin et al., 1992 Proc. Natl. Acad. Sci. USA 89, 504-508; 1992 Blume et al., Nucl. Acids Res. 20, 1777-1784; 1992, J. Bioi. Chem. 267, 3389-3395.
Недавно были доложены, в виде теоретических рассчетов и эмпирических обнаружений, которые дают руководство для конструирования олигонуклеотидов, тройная спираль которых ингибирует экспрессию гена. Например, преимущественными являются олигонуклеотиды, в общем длина которых больше, чем 14 нуклеотидов, для обеспечения специфичности мишеневой последовательности (см., например Maher et al., (1989); Grigoriev et al., (1992)). Многие клетки также жадно захватывают олигонуклеотиды, которые составляют менее чем 50 нуклеотидов по длине (см., например, Orson et al., (1991); Holt et al., 1988, Prос. Nаtl, Acad. Sci., USA, 85, 1028-1032). Для снижения восприимчивости к внутриклеточному разрушению, например, при воздействии 3’-экзонуклеаз, можно ввести свободный амин в 3’-концевую гидроксильную группу нуклеотидов, без нарушения специфического связывания последовательности (Orson еt аl., 1991). Кроме того, образуются более стабильные триплексы, если любые цитозины, которые могут находиться в олигонуклеотиде, являются метилированными, а также если интеркалирующий агент, такой как производное акридина, ковалентно связаны с 5’-концевым фосфатом (например, через пентаметиленовый мостик); снова без потери специфичности последовательностью (Maher et al., (1989); Grigoriev et al., (1992).
Способы получения или синтеза олигонуклеотидов хорошо известны в технике. Подобные способы можно расставить по порядку от обычного ферментативного расщепления с последующим выделением фрагментов нуклеотидов (см., например, Sambroоk, еt al., Части 5, 6) до чисто синтетических способов, например при помощи цианоэтилфосфорамидатного способа, используя системы Milligen или Весkman System 1 Plus синтезаторы ДНК (см. также Jkuta et al., in Ann. Rev. Bioсhеm. 1984, 53, 323-356 (фосфотриэфирный и фосфит-триэфирный способы); Nаrаng еt al., in Methods Enzymol., 65, 610-620 (1980) (фосфотриэфирный способ). Соответственно, последовательность ДНК 5’-фланкирующего региона гена, чувствительного к окислению-восстановлению, описанная здесь, может быть использована для конструирования олигонуклеотидов, включающих последовательность ДНК, состоящую из, по крайней мере, 15 последовательных нуклеотидов, с или без модифицированных оснований или производных интеркалирующих агентов, для получения тройных спиралей, специфически внутри 5’-фланкирующего участка гена, чувствительного к окислению-восстановлению, с целью ингибирования экспрессии гена.
В некоторых случаях может быть предпочтительным ввести энхансер или множественные копии регуляторных последовательностей для облегчения подбора способов и реактивов для управления экспрессией.
V. Модели и наборы
Для заболеваний, опосредованных VCAM-1 или геном, чувствительным к окислению-восстановлению, описываются также наборы, которые включает подсчет заменяющих маркеров заболевания. В одном воплощении, например, оценивается уровень окисленных полиненасыщенных жирных кислот или других подходящих маркеров в ткани или крови хозяина в виде среднего из оценок "окислительного состояния среды" хозяина и восприимчивости хозяина к опосредованным VСАМ-1 или геном, чувствительным к окислению-восстановлению, заболеваниям.
В другом воплощении количественно определяли уровень циркулирующего VCAM-1 на поверхности клетки или другого подходящего маркера и влияние введения на этот уровень необходимого антиоксиданта.
В ходе другого анализа оценивали чувствительность эндотелиальных клеток хозяина к полиненасыщенным жирным кислотам и их окисленным производным. Это может быть выполнено, например, путем замены у хозяина PUFA или ох-PUFA и сравнения конечной концентрации циркулирующего VCAM-1 на поверхности клетки или другого заменяющего маркера с нормальной популяцией.
В другом воплощении на модели атеросклероза in vivo или других сердечных или воспалительных заболеваний, которые опосредованы VCAM-1, для индукции заболевания может быть предусмотрено, посредством введения животному-хозяину, избыточного количества РUFА или полиненасыщенных жирных кислот. Данные животные могут быть использованы для клинических исследований для дальнейшего исследования данных заболеваний.
В другом воплощении изобретения соединения еще могут быть оценены по их способности лечить заболевания, опосредованные VCAM-1, на основании их способности ингибировать окисление полиненасыщенных жирных кислот, или взаимодействия PUFA или ох-РUFА с белковой мишенью.
Это может быть выполнено путем замены у хозяина, например человека или животного, такого как мышь, на высокий уровень PUFA или ох-РUFА, и затем определения терапевтической эффективности тестируемых соединений по их способности снижать концентрацию циркулирующего или клеточно-поверхностного VCAM-1. Альтернативно, можно использовать опробирование in vivo, которое основано на способности соединений предотвращать окисление PUFA, или взаимодействии PUFA или ox-PUFA с белковой мишенью, в присутствии окисляющих соединений, таких как металл, например медь, или ферменты, такие как пероксидаза, липооксигеназа, циклооксигеназа или цитохром Р450.
В другом воплощении эндотелиальные клетки сосуда выдерживали с TNF-α или другим, индуцирующим VCAM-1, материалом, в течение соответствующего времени, затем разрушали, например, под действием звука или методом замораживания-оттаивания. Выделяли цитозольную и мембранную фракции. Для определения количества фракций добавляли радиоактивно меченую РUFА. Исследовали способность жидкости превращать РUFА в ох-РUFА в присутствии или отсутствие тестируемого соединения. Можно использовать интактные клетки вместо разрушенной клеточной системы.
III. Фармацевтические композиции
Люди, лошади, собаки, крупный рогатый скот и другие животные, в частности млекопитающие, страдающие от сердечно-сосудистых и других воспалительных заболеваний, опосредованных VСАМ-1 и редокс-чувствительным геном, могут получить курс лечения путем введения пациенту эффективного количества соединения, что приводит к удалению, уменьшению концентрации или защите от образования окисленных полиненасыщенных жирных кислот, включая, но не ограничиваясь ими, линолевую (C18 Δ9, 12), линолиновую (C18, Δ6, 9, 12), арахидоновую (С20 Δ5, 8, 11, 14) и экозатриеновую (c20 Δ8, 11, 14) кислоты; другие сигналы окисления или их фармацевтически приемлемого производного или соли с фармацевтически приемлемым носителем или разбавителем. Активные вещества могут быть введены любым подходящим путем, например перорально, парентерально, внутривенно, интрадермально, подкожно или наружно.
Как здесь используется, термин фармацевтически приемлемые соли или комплексы относится к солям или комплексам, которые сохраняют желаемую биологическую активность вышеуказанных соединений и проявляют минимальные нежелательные токсические эффекты. Неограничивающими примерами таких солей являются:
(а) соли присоединения кислот, образованные неорганическими кислотами (например, хлористоводородная кислота, бромистоводородная кислота, серная кислота, фосфорная кислота, азотная кислота и тому подобное), и соли, образованные органическими кислотами, такими как уксусная кислота, щавелевая кислота, винная кислота, янтарная кислота, яблочная кислота, аскорбиновая кислота, бензойная кислота, танниновая кислота, памоевая кислота, альгиновая кислота, полиглутаминовая кислота, нафталинсульфоновая кислота, нафталиндисульфоновая кислота и полигалактуроновая кислота;
(б) соли прибавления оснований, образованные катионами поливалентных металлов, таких как цинк, кальций, висмут, барий, магний, алюминий, медь, кобальт, никель, кадмий, натрий, калий и тому подобное, или с органическим катионом, образованным из N,N-дибензилэтилендиамина, D-глюкозамина, аммиака, тетраэтиламмония или этилендиамина; или
(в) сочетания (а) и (б); например, цинктоннатная соль или тому подобное.
Активное соединение или смесь соединений вводят любым подходящим образом, включая, но не ограничиваясь пероральным или внутривенным путями. Обычные уровни доз для любого из вышеуказанных состояний будут от 0,5 до 500 мг/кг массы тела при режиме принятия доз, изменяющемся от 1 каждый другой день до нескольких раз в день. Предпочтительные дневные дозы находятся в промежутке от приблизительно 1 до 3000 мг/пациент/день, более предпочтительно между приблизительно от 5 до 500 мг/пациент/день, и еще более предпочтительно между приблизительно от 25 до 500 мг/пациент/день.
Активный ингредиент следует вводить до достижения пиковых концентраций активного вещества в плазме от около 0,1 до 100 мкМ, предпочтительно около 1-10 мкМ. Это может быть достигнуто, например, путем внутривенной инъекции раствора или препарата активного ингредиента, не обязательно, в физиологическом растворе или в водной среде, или введением активного ингредиента в виде болюса.
Соединение может быть также введено прямо в сосудистую стенку используя перфузионные балонные катетеры после или вместо коронарной или другой артериальной ангиопластики. В качестве примера, 2-5 мл физиологически приемлемого раствора, содержащего приблизительно 1-500 мкМ соединения или смесь соединений, вводят при 1-5 атмосферном давлении. С этого времени в течение курса следующих шести месяцев, в течении периода максимальной опасности рестеноза, активные соединения вводятся другими подходящими путями и схемами лечения.
Обработки в течение относительно короткого периода времени активным содинением используются для того, чтобы вызвать сокращение пораженных заболеванием участков коронарной артерии, которые не могут быть обработаны ни ангиопластически, ни хирургически. Неограничивающим примером обработки в течение короткого периода времени является от двух до шести месяцев лечение дозами от 0,5 до 500 мг/кг массы тела, даваемыми от одного раза каждый второй день до трех раз в день.
Обработки в течение более длительного периода времени могут быть использованы для профилактики развития прогрессирующих поражений у пациентов с повышенным риском заболевания. Обработка в течение длительного периода времени может представлять собой лечение в течение нескольких лет дозами от 0,5 до 500 мг/кг массы тела, вводимыми с интервалами от одного раза каждый второй день до трех раз в день.
Активное соединение может быть также введено сразу или после коронарной ангиопластики в качестве средства, которое уменьшает или устраняет аномальную пролиферативную и воспалительную реакцию, которые часто приводят к клинически значимому ре-стенозу.
Активные соединения могут назначаться в сочетании с другими лекарственными препаратами, используемыми при лечении сердечно-сосудистой болезни, включая средства, понижающие уровни липидов, такие как пробукол и никотиновая кислота; ингибиторы слипания тромбоцитов, такие как аспирин; антитромбоцитарные средства, такие как кумадин; блокаторы кальциевых каналов, такие как веропамил, дилтиазем и нифедипин; ингибиторы ангиотензин превращающего фермента (АСЕ), такие как каптоприл и иналоприн, и β-блокаторы, такие как пропанолол, тербуталол и лабеталол. Соединения также могут вводиться в сочетании с нестероидными противовоспалительными препаратами, такие как ибупрофен, индометацин, фенопрофен, мефенамовая кислота, флюфенамовая кислота, сулиндак. Соединения также могут вводиться в сочетании с кортикостероидами.
Концентрация активного соединения в лекарственной композиции будет зависеть от абсорбции, распределения, инактивации и путей экскреции лекарственного средства, а также от других факторов, известных специалистам в данной области. Необходимо также отметить, что размеры дозировок также будут изменяться в зависимости от серьезности состояния, которое необходимо облегчить. Далее следует учесть, что для каждого конкретного субъекта следует устанавливать конкретные схемы лечения лекарственными препаратами в течение периода времени, соответствующего индивидуальной потребности и в соответствии с профессиональным суждением лица, назначающего или наблюдающего за назначением композиций, и что указанные здесь области концентраций являются только примером и не предназначены для ограничения области практического применения заявленной композиции. Активный ингредиент может быть введен сразу или может быть разбит на более маленькие дозы для введения с различными интервалами времени.
Пероральные композиции обычно включают инертный разбавитель или пищевой носитель. Они могут быть заключены в желатиновые капсулы или спрессованы в таблетки. Для цели перорального терапевтического введения активное соединение может быть включено в наполнители и использовано в виде таблеток, пастилок или капсул. В качестве части композиции могут быть включены фармацевтически совместимые связующие агенты и/или вспомогательные вещества.
Таблетки, драже, капсулы, пастилки и тому подобное содержат любой из следующих ингредиентов или близкие по природе соединения: связующее, такое как микрокристаллическая целлюлоза, смола трагаканта или желатин; наполнитель, такой как крахмал или лактоза, разрыхляющий агент, такой как альгиновая кислота, примогель или кукурузный крахмал; смазывающий агент, такой как коллоидный диоксид кремния; подслащивающий агент, такой как сахароза или сахарин; или ароматизирующий агент, такой как перечная мята, метил салицилат или апельсиновый ароматизатор. Когда стандартной лекарственной формой является капсула, она может содержать, кроме веществ вышеуказанных видов, жидкий носитель, такой как жировое масло. Кроме того, стандартные лекарственные формы могут содержать различные другие вещества, которые модифицируют физическую форму лекарственной единицы, например покрытия сахаром, шеллаком или другими покрывающими агентами.
Активное соединение или его фармацевтически приемлемая соль или производное могут вводиться в виде компонента эликсира, суспензии, сиропа, облатки, жевательной резинки или тому подобное. Сироп может содержать, в дополнение к активному соединению, сахарозу в качестве подслащивающего агента и некоторые консерванты, краски и красители, ароматизаторы. Активное соединение или его фармацевтически приемлемые соли или производные также могут вводиться с другими активными веществами, которые не препятствуют желаемому действию, или с веществами, которые поддерживают желаемое действие, такие как антибиотики, противогрибковые, противовоспалительные или противовирусные соединения.
Растворы или суспензии, используемые для парентерального, интрадермального, подкожного или наружного применения, могут включать следующие компоненты: стерильный разбавитель, такой как вода для инъекций, физиологический раствор, нелетучие масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные агенты, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота; буферирующие агенты, такие как ацетаты, цитраты или фосфаты, и средства для придания изотоничности, такие как хлорид натрия или декстроза. Парентеральный препарат может быть помещен в ампулы, одноразовые шприцы или пузырьки с многократными дозами, изготовленными из стекла или пластмассы.
Подходящие растворители или носители для наружного применения известны и включают лосьоны, суспензии, мази, кремы, гели, настойки, спреи, порошки, пасты, подкожные лепешки с медленными высвобождением, аэрозоли для астмы и суппозитории для нанесения на ректальную, вагинальную, назальную или пероральные слизистые.
Для получения композиций для наружного применения могут быть использованы загустители, смягчители и стабилизаторы. Примеры загустителей включают вазелин, пчелиный воск, ксантановую смолу или полиэтиленгликоль, увлажнители, такие как сорбитол, смягчители, такие как минеральное масло, ланолин и его производные или сквален. Большое число растворов и мазей являются коммерчески доступными.
Природные и синтетические ароматизаторы или подслащивающие средства могут быть добавлены для улучшения вкуса наружных препаратов, применяемых для местного действия на поверхностях слизистых. Могут быть добавлены инертные краски или красители, в частности, в случае препаратов предназначенных для нанесения на поверхности пероральной слизистой.
Активные соединения могут быть получены с носителями, которые предохраняют соединения от быстрого высвобождения, такие как состав с контролируемым высвобождением, включая имплантаты и микроинкапсулированные системы доставки. Могут быть использованы биоразрушаемые биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Многие методы получения таких составов запатентованы или обычно известны специалистам в данной области.
Для внутривенного введения предпочтительными носителями являются физиологический раствор или фосфат-буферированный физиологический раствор (PBS).
Активное соединение также может быть введено посредством трансдермальной пластинки. Способы получения трансдермальной пластинки известны специалистам в данной области. Например, см. Brown, Z. and Zanger, R., Transdermal Delivery of Drugs, Annual Review of Mеdicine, 39:221-229 (1988), включенное здесь в качестве ссылки.
При другом осуществлении активные соединения получают с носителями, которые предохраняют соединения от быстрого вывода из организма, такие как состав с контролируемым высвобождением, включая имплантанты и микроинкапсулированные системы доставки. Могут быть использованы биоразрушаемые биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Способы получения таких составов известны специалистам в данной области. Вещества могут быть также коммерчески получены от Alza Corporacion and Nova Pharmaceuticals, Jnс.
Фармацевтически приемлемыми могут также быть липосомальные суспензии. Они могут быть получены в соответствии с методами, известными специалистам в данной области, например как описано в патенте США 4522811 (который включен здесь в качестве ссылки в своем полном объеме). Например, липосомные составы могут быть получены растворением соответствующего(их) липида(ов) (такие как стеароилфосфатидилэтаноламин, стеароилфосфатидилхолин, арахадоилфосфатидилхолин) и холестерин в неорганическом растворителе, который затем упаривают с получением тонкой пленки сухого липида на поверхности контейнера. В этот контейнер затем помещают водный раствор активного соединения или его монофосфатного, дифосфатного и/или трифосфатного производного. Затем контейнер подвергают ручному взбалтыванию для отделения липидного вещества от стенок контейнера и для диспергирования липидных агрегатов, получают таким образом липосомальную суспензию. Специалисту в данной области будут ясны модификации и варианты настоящего изобретения из предшествующего подробного описания изобретения. Такие модификации и вариации входят в объем прилагаемой формулы изобретения.

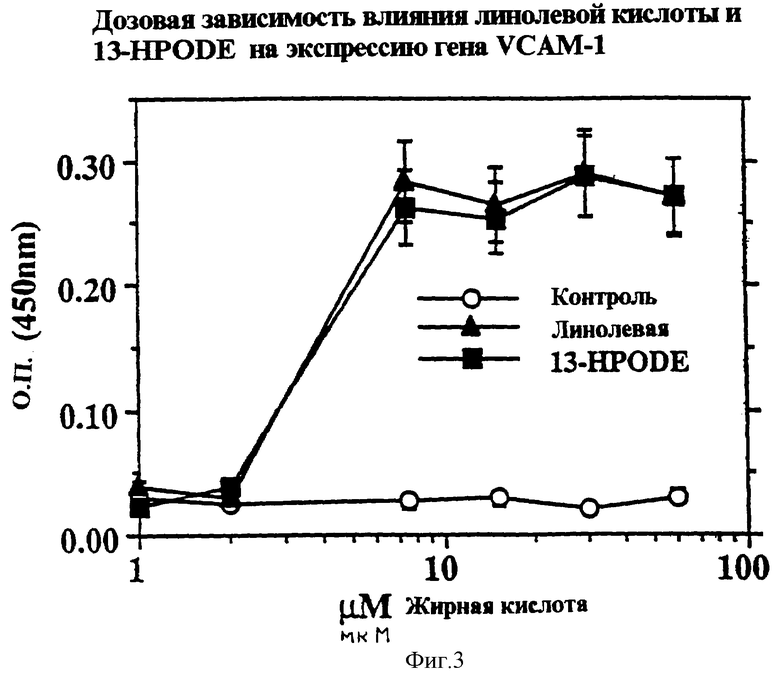
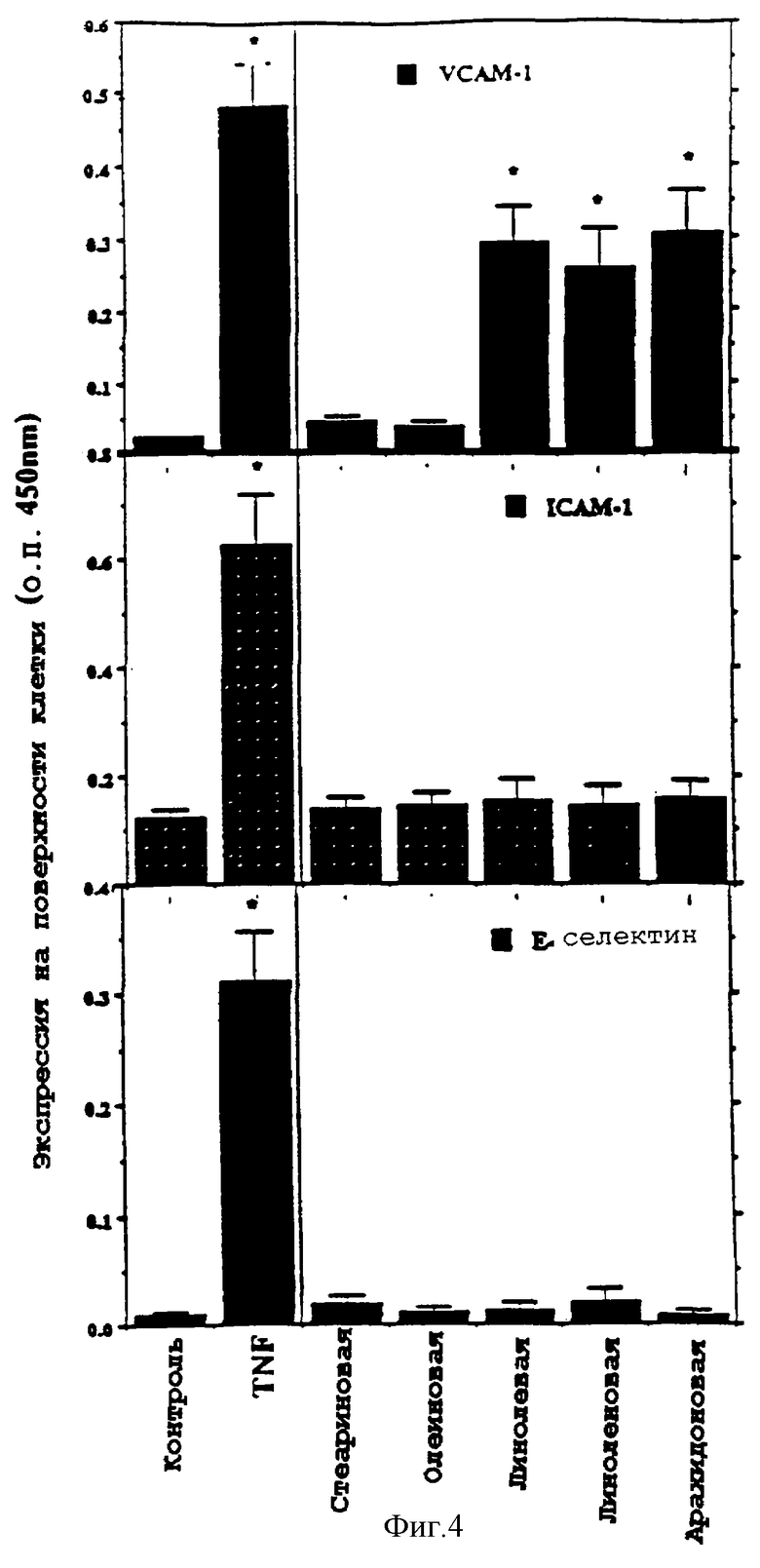
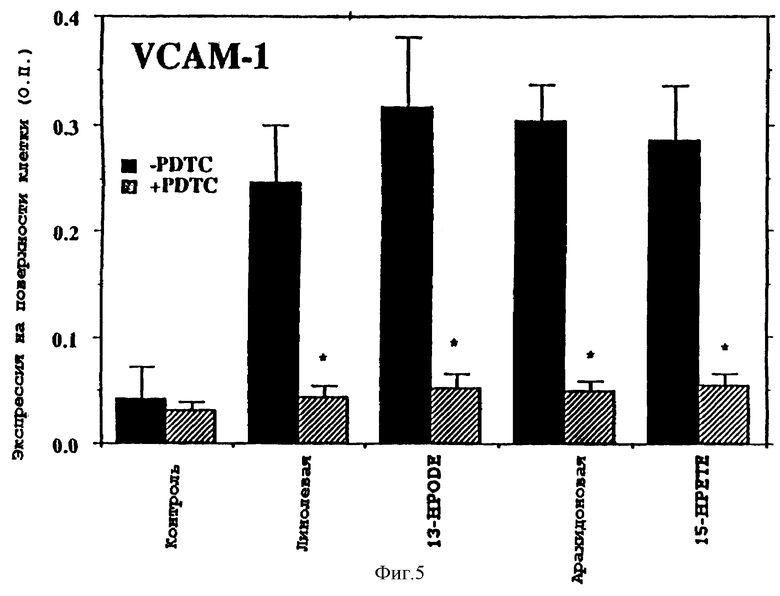


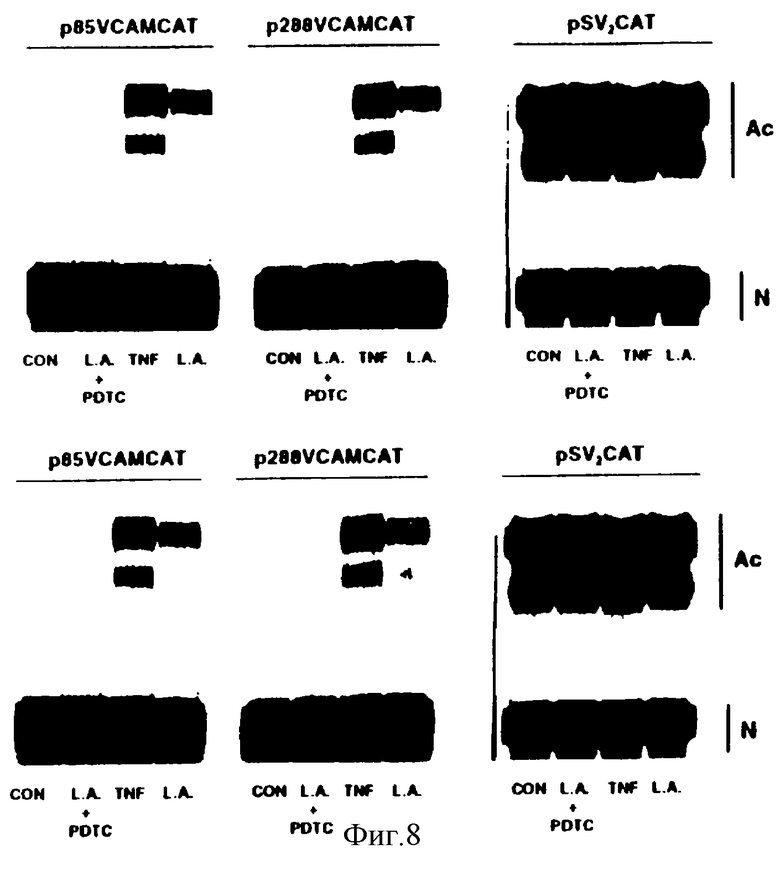
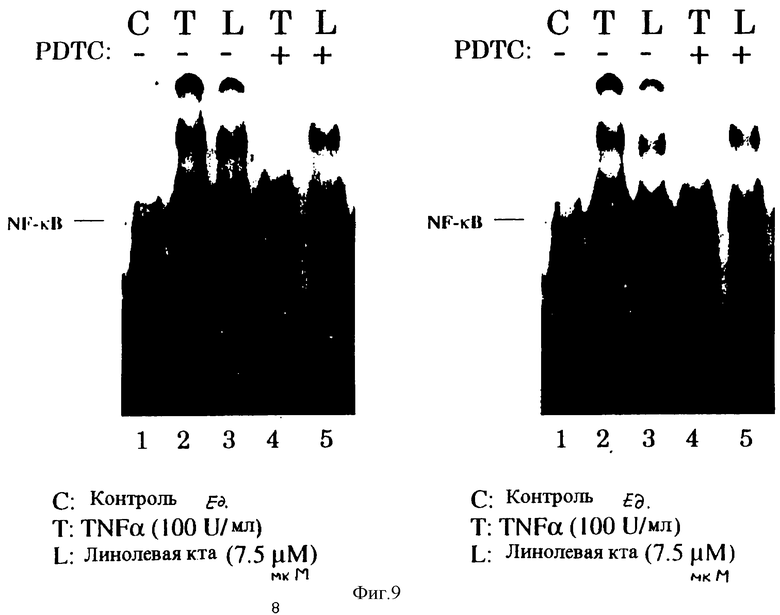
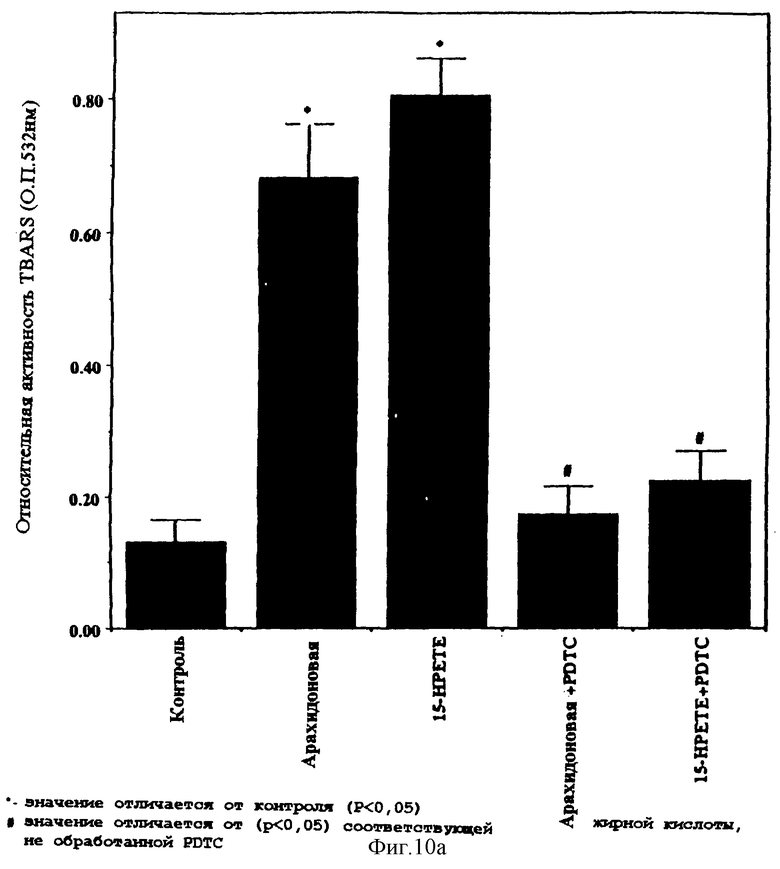
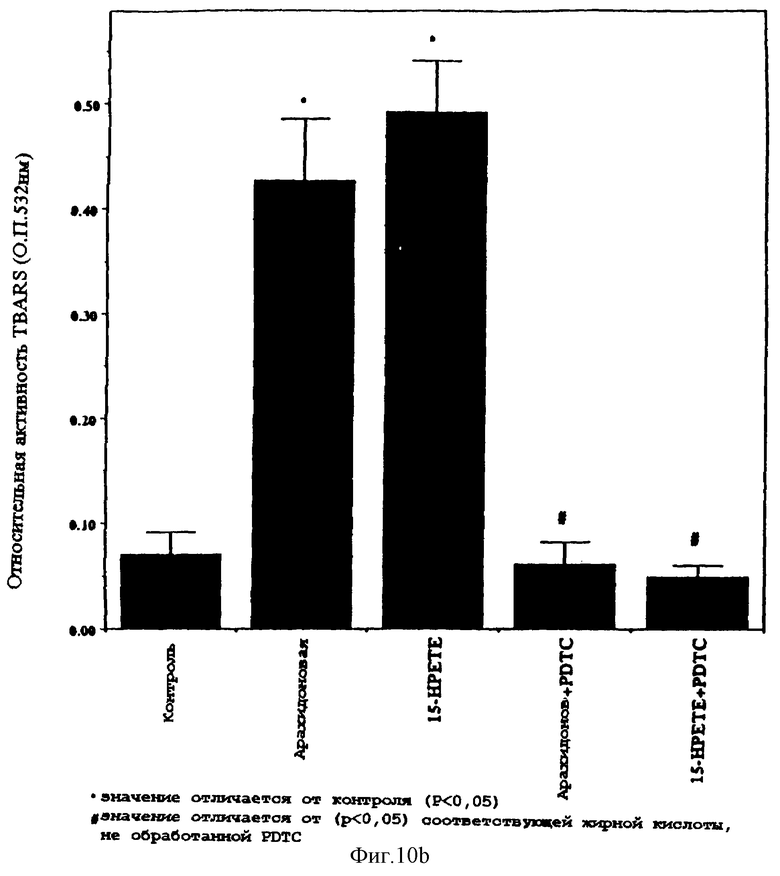
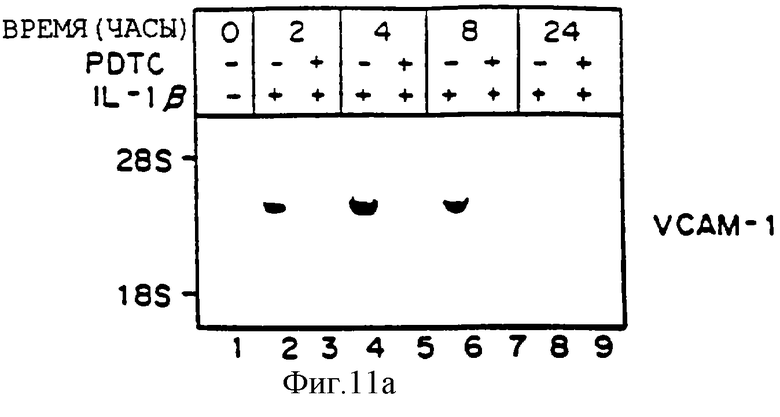
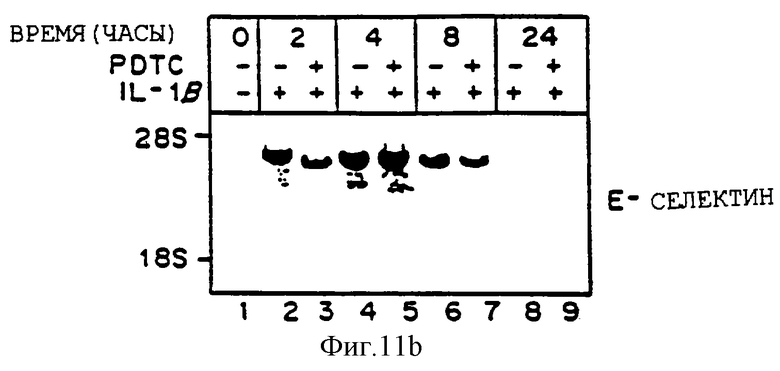
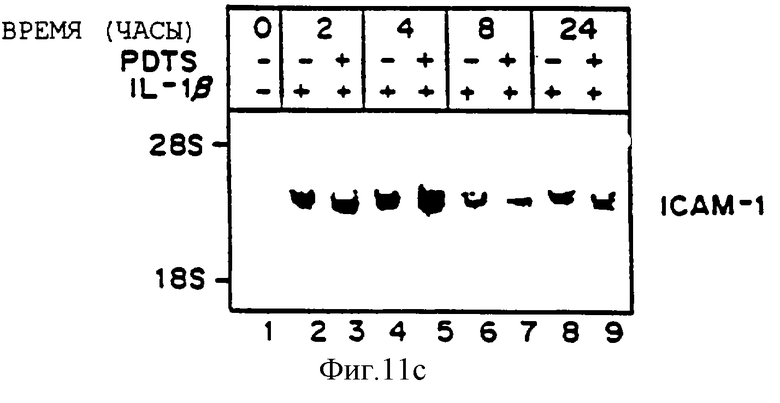
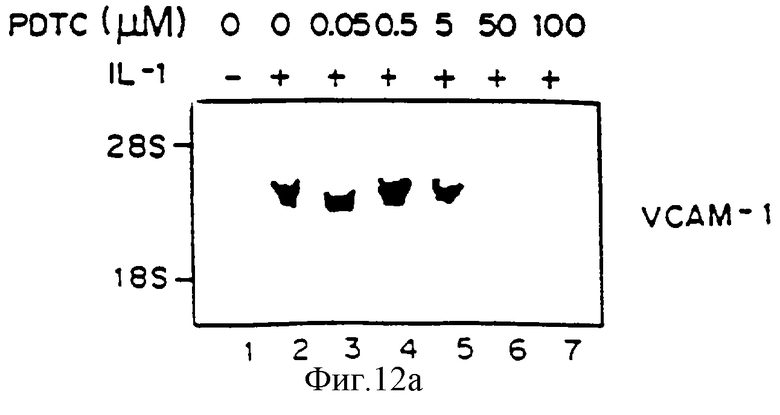

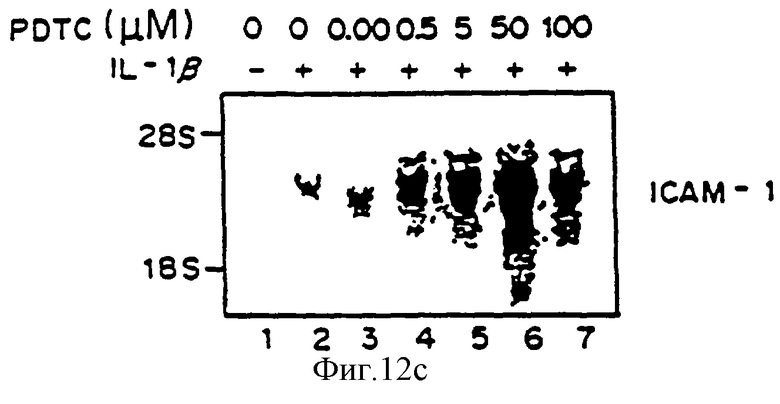
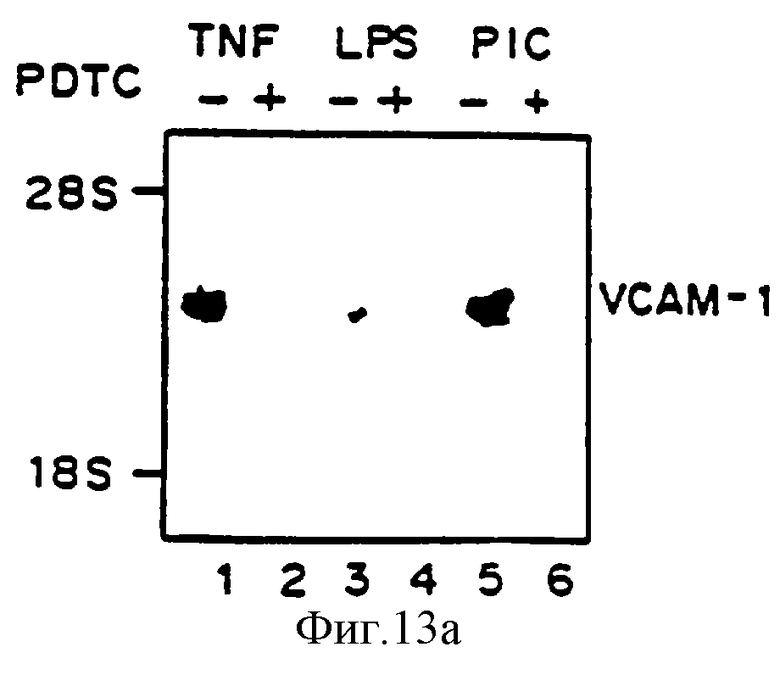
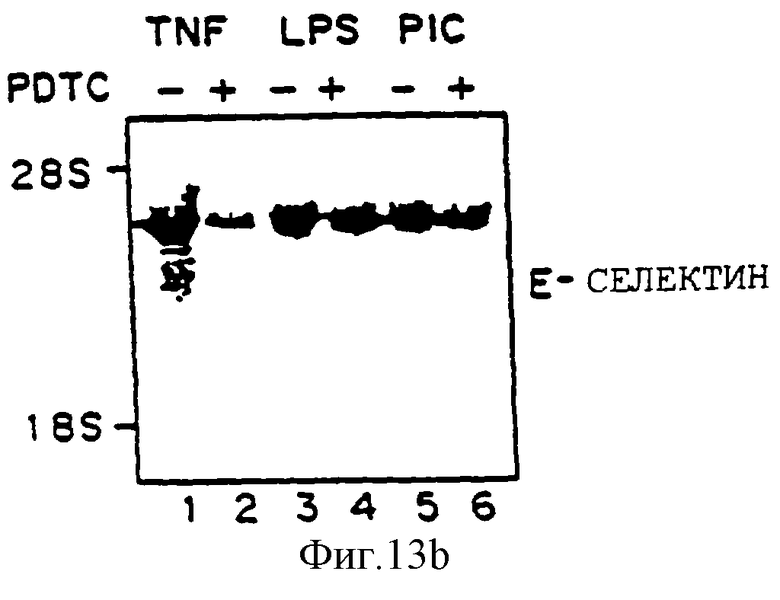
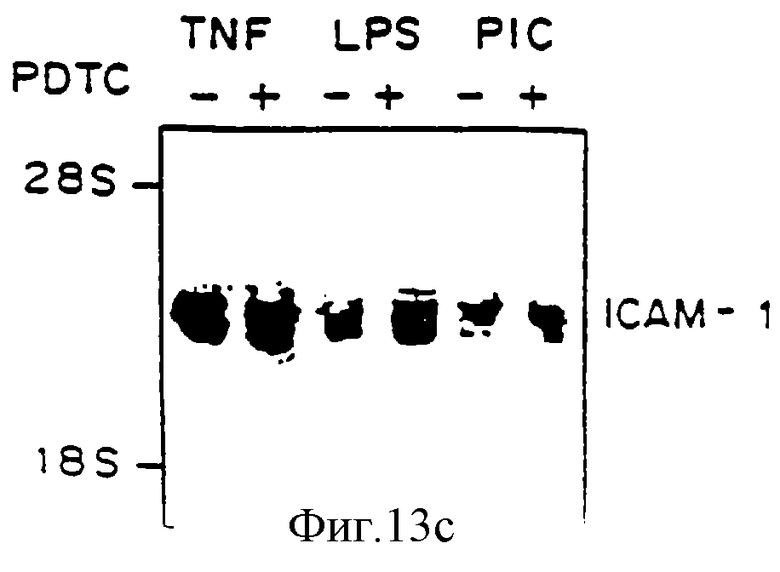
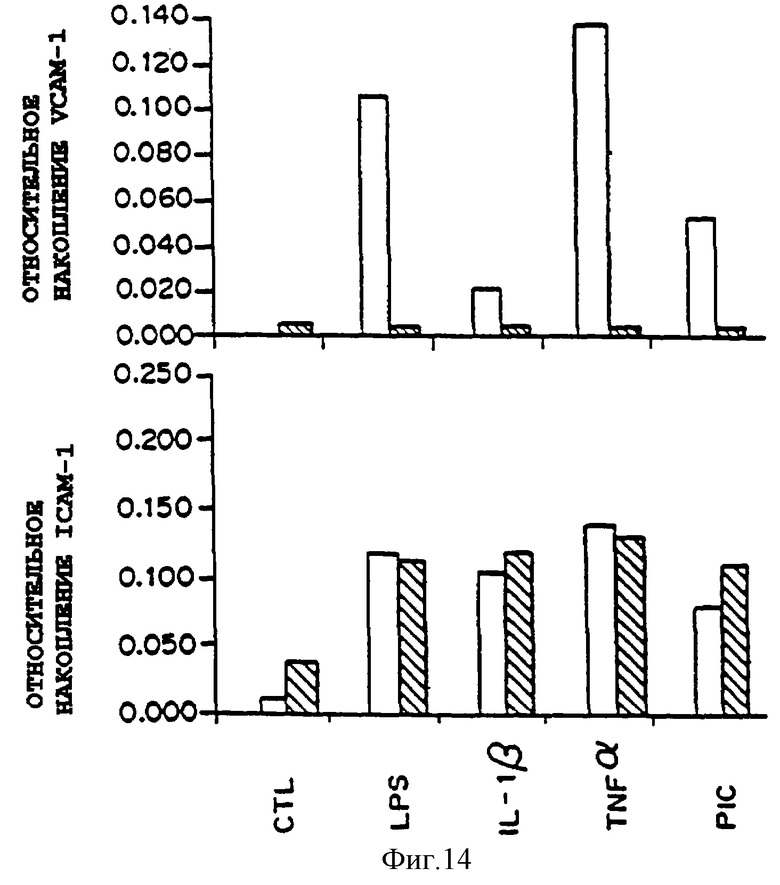

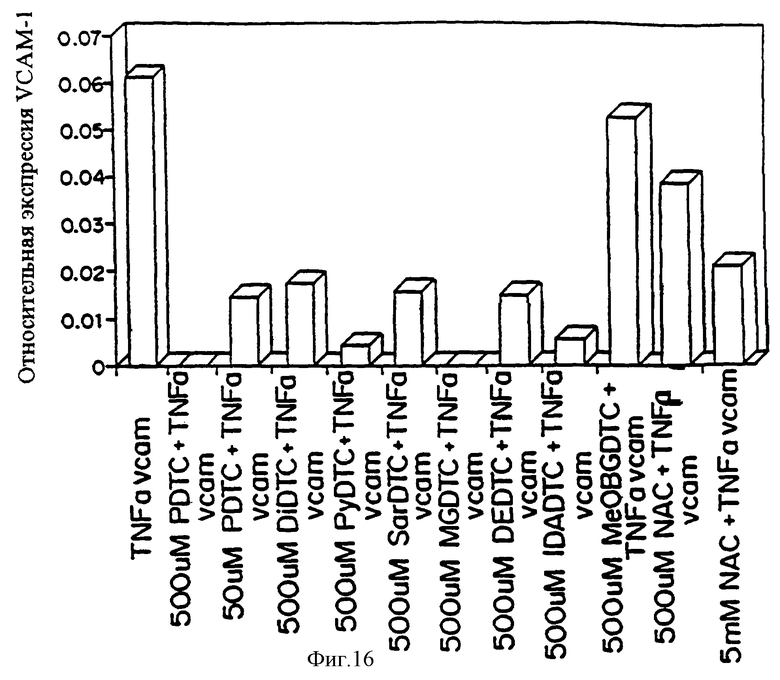

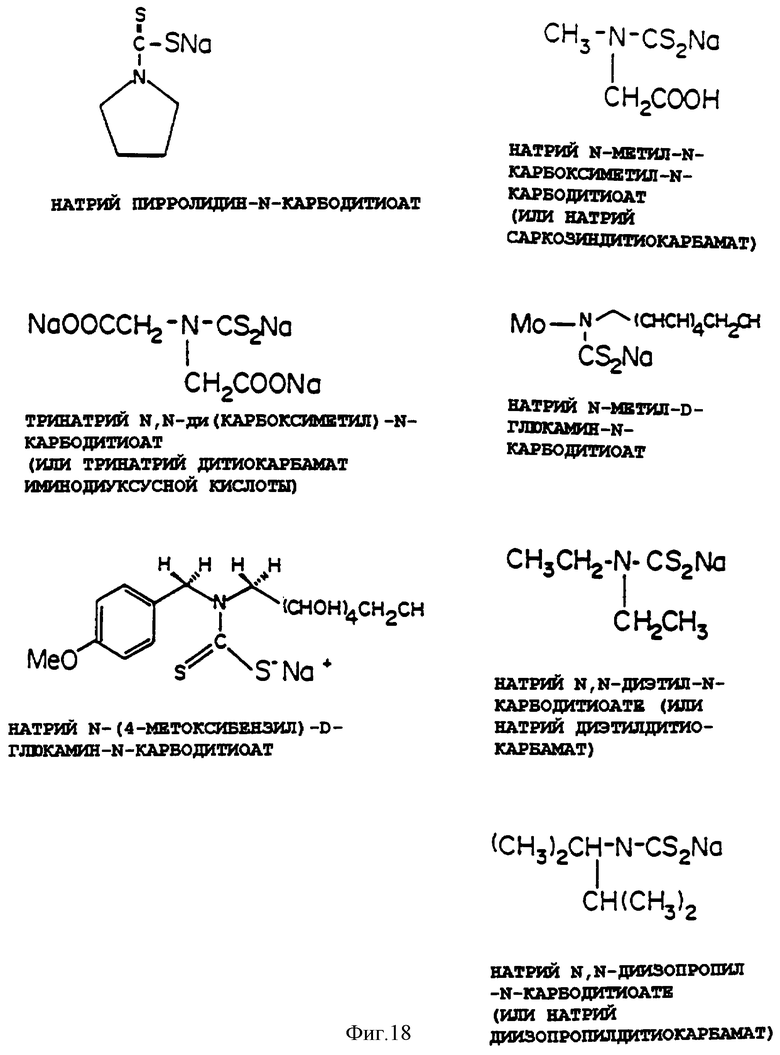
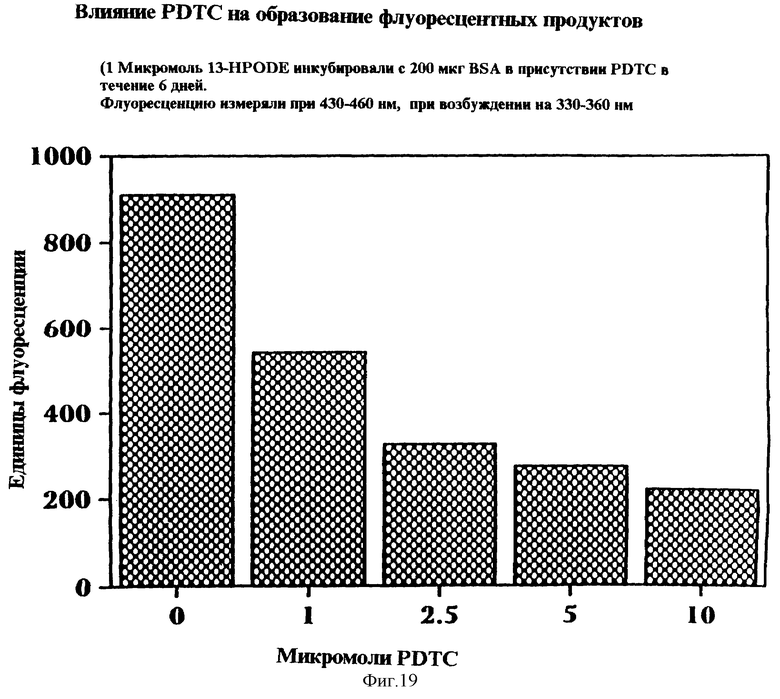
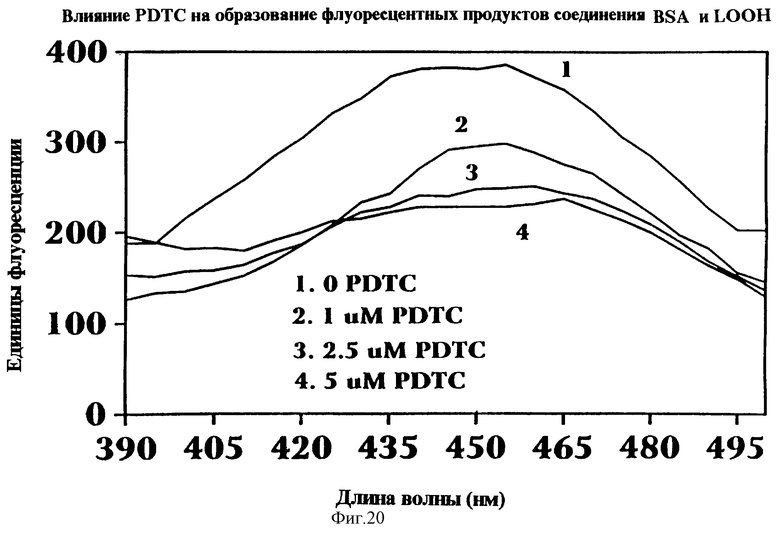


Изобретение относится к медицине, в частности к терапии, и касается лечения сердечно-сосудистых и воспалительных заболеваний, опосредованных VCAM-1 (фактором васкулярной клеточной адгезии 1) или экспрессией редокс-чувствительного гена. Для этого вводят вещество, которое предотвращает или минимизирует окисление полиненасыщенной жирной кислоты. Изобретение касается также тестирования соединения на его способность к лечению заболевания, опосредованного VCAM-1, и оценки сенсибилизации сосудистых эндотелиальных клеток к полиненасыщенным жирным кислотам или их окисленным производным. Изобретение обеспечивает идентификацию избирательных методов лечения указанных заболеваний и адекватный подбор необходимых терапевтических средств. 6 н. и 31 з.п. ф-лы, 29 ил.

| US 5061722 А, 29.10.1991 | |||
| СПОСОБ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ АТЕРОСКЛЕРОЗА | 1993 |
|
RU2095059C1 |
| RU 95103436 A1, 20.01.1997 | |||
| US 4900757 А, 13.02.1990 | |||
| ЕР 0461548 А2, 18.12.1991 | |||
| US 4870101 А, 26.09.1989. | |||
| CORKE et al | |||
| - in Int | |||
| J | |||
| of Immunopharmacol | |||
| Колосниковая решетка с чередующимися неподвижными и движущимися возвратно-поступательно колосниками | 1917 |
|
SU1984A1 |
| LANG et al | |||
| - in Jhe Lancet, 1988, v.2, №8613, p.702-706. | |||

