Область, к которой относится изобретение
Настоящее изобретение относится к способу изменения субстратной специфичности липолитического фермента путем модификации аминокислотной последовательности и к вариантам липолитических ферментов, полученным посредством такой модификации. Настоящее изобретение также относится к способу скрининга липолитических ферментов.
Предпосылки создания изобретения
Липолитические ферменты (такие как липазы и фосфолипазы) обладают способностью гидролизовать связи сложных эфиров карбоновых кислот в субстрате с высвобождением этих карбоновых кислот. Гидролитическая активность в отношении различных сложноэфирных связей имеет важное значение для использования липолитического фермента в различных промышленных целях.
Таким образом, ферменты с высокой фосфолипазной активностью могут иметь широкое применение, такое как хлебопекарное производство (патент США 4567046), фильтрация гидролизата пшеничного крахмала (патент США 5264367) и обработка растительного масла для снижения содержания в нем фосфолипида (патент США 5264367). Для обработки растительного масла этот фермент должен иметь низкую липазную активность, то есть низкую гидролитическую активность в отношении сложно-эфирных связей в триглицеридах.
В WO 98/45453 показано, что фермент с высокой гидролитической активностью по отношению к дигалактозилдиглицериду (ДГДГ) может быть использован в хлебопекарном производстве.
Хорошо известно, что добавление липазы в моющие средства для стирки способствует удалению жирных пятен (например, ЕР 258068).
Высвобождение короткоцепочечных жирных кислот в качестве свободных жирных кислот (СЖК) может оказаться желательным для формирования нужного запаха в пищевых продуктах, например при созревании сыра (M.Hanson, ZFL, 41 (10), 664-666 (1990)).
Трехмерная структура (3D) некоторых липолитических ферментов известна, и также известно, что некоторые структуры содержат так называемую "крышку", которая может находиться в открытом или закрытом состоянии, покрывая активный центр. Brady et al., Nature, 343, 767-770 (1990). Brzozowski A.M. et al., Nature, 351, 491 (1991). Derewenda et al., Biochemistry, 31(5), 1532-1541 (1992).
F.Hara et al., JAOCS, 74(9), 1129-32 (1997) указывают, что некоторые липазы обладают определенной фосфолипазной активностью, тогда как большинство липаз обладают слабой активностью в отношении фосфолипидов, либо вообще не обладают такой активностью. Так, например, была описана фосфолипазная активность для липаз поджелудочной железы морских свинок, Fusarium oxysporum и Staphylococcus hyicus, и были предприняты попытки установить взаимосвязь фосфолипазной активности со структурой этой липазы. WO 98/26057; M.D.van Kampen et al., Chemistry and Physics of Lipids, 93 (1998), 39-45; A.Hjorth et al.. Biochemistry 1993, 32, 4702-4707.
В предшествующих работах было описано влияние замен аминокислот в липазе от Rhizopus delemar на селективность этих липаз в отношении длины цепи кислот. Таким образом, R.D. Joerger et al., Lipids, 29(6), 377-384 (1994) показал, что варианты F95D, F112W и V209W имеют модифицированную предпочтительность в пользу С4- и С8-кислот. R.R. Klein et al., JAOCS, 74 (11), 1401-1407 (1997) показал, что вариант V206T+F95D имеет более высокую селективность по отношению к С8-кислоте. R.R.Klein et al., Lipids, 32(2), 123-130 (1997) показал, что варианты V209W+F112W, V94W и F95D+F214R обладают более высокой гидролитической активностью в отношении С4- и С8-кислот, и высказал предположение, что структурные детерминанты для специфичности в отношении средней длины цепи могут находиться на удаленном конце ацил-связывающего "желобка".
Краткое описание изобретения
Авторами настоящего изобретения было обнаружено, что субстратная специфичность липолитического фермента может быть модифицирована путем внесения модификаций в аминокислотную последовательность на определенном участке этого липолитического фермента в целях увеличения уровня нужной активности или в целях снижения уровня нежелательной активности. Таким образом, авторы настоящего изобретения получили липолитические ферменты с модифицированной аминокислотной последовательностью (далее называемые вариантами липолитических ферментов или, вкратце, вариантами), с определенной специфичностью к субстрату, и которые могут быть использованы для конкретных целей.
В соответствии с этим настоящее изобретение относится к способу продуцирования варианта липолитического фермента и вариантов липолитических ферментов, полученных данным способом. Указанный способ предусматривает:
a) выбор субстрата и нужной сложноэфирной связи,
b) выбор исходного липолитического фермента,
c) выбор, по крайней мере, одного аминокислотного остатка в области возле активного сайта, в области возле С-конца или в области "крышки" исходного липолитического фермента, описанного ниже,
d) создание модификаций, каждая из которых представляет собой инсерцию, делецию или замену аминокислотного остатка,
e) необязательно, создание модификаций, каждая из которых представляет собой инсерцию, делецию или замену аминокислотного остатка в одном или нескольких положениях, отличающихся от положений, указанных в с),
f) получение сконструированного варианта,
g) тестирование на активность данного варианта по отношению к сложноэфирной связи в данном субстрате, и
h) отбор варианта, обладающего модифицированной активностью по отношению к сложноэфирной связи.
Таким образом, в одном из аспектов настоящего изобретения, исходный липолитический фермент имеет спирт-связывающий сайт, содержащий глицериновую часть в положении sn2, а аминокислотная модификация находится в пределах 10 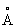 от С-атома в положении sn2 глицериновой части триглицеридного субстрата.
от С-атома в положении sn2 глицериновой части триглицеридного субстрата.
В другом аспекте настоящего изобретения исходный липолитический фермент имеет структуру, содержащую каталитическую триаду, состоящую из активного остатка Ser, активного остатка Asp и активного остатка His, а модифицируемая аминокислота расположена либо между активным остатком His каталитического остатка и С-концом, либо принадлежит к серии Е, определенной нижеследующими стадиями:
i) сопоставление первичной структуры липолитического фермента со структурой липазы 4TGL Rhizomucor miehei, содержащей каталитическую триаду и атом фосфора ингибитора (4TGL-inhP), для минимизации суммы квадратов отклонения между атомами каталитических триад двух структур,
ii) определение серии А, состоящей из атомов липолитического фермента внутри сферы радиусом 18 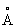 с центром в 4TGL-inhP,
с центром в 4TGL-inhP,
iii) формирование первой плоскости, определенной 4TGL-inhP, атомом Сα активного остатка Ser исходного липолитического фермента и атомом Сα активного остатка Asp исходного липолитического фермента и определение серии В как субсерии серии А, состоящей из атомов на той же самой стороне первой плоскости, на которой расположен атом Сα активного остатка His исходного липолитического фермента,
iv) формирование второй плоскости, определенной 4TGL-inhP, атомом Сα активного остатка Ser исходного липолитического фермента и атомом Сα активного остатка His исходного липолитического фермента и определение серии С как субсерии серии А, состоящей из атомов на противоположной стороне второй плоскости, на которой расположен атом Сα активного остатка Asp исходного липолитического фермента,
v) формирование набора D, состоящего из атомов, принадлежащих к объединенным наборам В и С, и имеющего доступность к растворителю 15 или выше, и
vi) формирование набора Е, состоящего из аминокислотных остатков в структуре, который содержит атом, принадлежащий к набору D, или атом, принадлежащий к объединенным наборам В и С и расположенный на расстоянии, менее чем 3,5 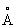 от атома, принадлежащего к набору D.
от атома, принадлежащего к набору D.
В третьем аспекте настоящего изобретения данный липолитический фермент имеет активный центр, содержащий активный остаток His, а модификацию в аминокислотной последовательности создают между активным остатком His и С-концом.
В еще одном аспекте настоящего изобретения аминокислотную модификацию создают в пределах 10 аминокислотных остатков у С-конца.
В другом аспекте настоящего изобретения исходный липолитический фермент имеет "крышку", и данную модификацию создают в области этой крышки.
Настоящее изобретение также относится к ДНК-последовательности, кодирующей данный вариант, к экспрессирующему вектору, содержащему эту ДНК-последовательность, к трансформированной клетке-хозяину, содержащей указанную ДНК-последовательность или указанный экспрессирующий вектор, и к способу получения данного варианта путем культивирования указанной трансформированной клетки-хозяина так, чтобы эта клетка продуцировала данный вариант, с последующим выделением данного варианта из полученной питательной среды. Кроме того, настоящее изобретение относится к использованию указанных вариантов.
Авторами настоящего изобретения было также обнаружено, что липолитический фермент, обладающий липазной и фосфолипазной активностью, а также активностью по отношению к дигалактозилдиглицериду, может быть с успехом использован в хлебопекарном производстве, и ими был разработан способ скрининга липолитических ферментов путем их тестирования на указанные активности.
Краткое описание чертежа
На чертеже проиллюстрировано сопоставление последовательностей липаз.
Подробное описание изобретения
Модифицированная активность по отношению к выбранной сложноэфирной связи в субстрате
Целью настоящего изобретения является изменение, по сравнению с исходным липолитическим ферментом, активности по отношению, по крайней мере, к одной выбранной сложноэфирной связи, по крайней мере, в одном субстрате, то есть увеличение нужной активности, снижение нежелательной активности или изменение специфичности к субстрату путем уменьшения отношения нежелательной активности к желательной активности.
Таким образом, фермент с повышенной фосфолипазной активностью может быть использован, например, в хлебопечении или для очистки растительного масла. Может оказаться желательным увеличение гидролитической активности фермента по отношению к дигалактозилдиглицериду (ДГДГ) в целях его использования в хлебопекарном производстве.
Увеличение липазной активности может оказаться желательным для любого промышленного производства, в котором используются липазы. Для использования в моющих средствах или при выпечке хлеба может оказаться желательным увеличение активности по отношению к длинноцепочечным (C16-C20)триглицеридам, и может оказаться желательным увеличение специфичности к длинноцепочечным жирным кислотам путем снижения отношения активности, направленной на короткоцепочечные или среднецепочечные (С4-С8)жирные кислоты, к активности, направленной на длинноцепочечные жирные кислоты.
Увеличение липазной активности в отношении короткоцепочечных или среднецепочечных (С4-С8)триглицеридов может оказаться желательным для использования в целях формирования запаха в пищевых продуктах (например, при созревании сыра).
При использовании в качестве фосфолипаз для очистки растительного масла может оказаться желательным снижение отношения липазной активности, направленной на длинноцепочечные (C16-C20)триглицериды, к фосфолипазной активности.
Исходный липолитический фермент
Липолитическим ферментом, используемым в настоящем изобретении, является фермент, который может гидролизовать сложноэфирные связи. Такими ферментами являются, например, липазы, такие как триацилглицеринлипаза (ЕС 3.1.1.3), липопротеинлипаза (ЕС 3.1.1.34), моноглицеридлипаза (ЕС 3.1.1.23), лизофосфолипаза, (феруловая кислота)-эстераза и эстераза (ЕС 3.1.1.1, ЕС 3.1.1.2). Цифры в скобках означают систематические номера, присвоенные Комиссией по ферментам Международного биохимического союза в соответствии с типом ферментативной реакционной способности данного фермента.
Исходный липолитический фермент может быть прокариотическим, а в частности, бактериальным ферментом, например, происходящим от Pseudomonas. Примерами являются липазы Pseudomonas, например P.cepacia (патент США № 5290694, pdb file 10IL), P.glumae (N.Frenken et al. (1992), Appl. Envir. Microbiol. 58 3787-3791, pdb files 1TAH and 1QGE), P.pseudoalcaligenes (EP 334462) и штамм SD705 вида Pseudomonas sp. (FERM BP-4772) (WO 95/06720, EP 721981, WO 96/27002, EP 812910). Последовательность липазы P.glumae идентична аминокислотной последовательности Chromobacterium viscosum (DE 3908131 A1). Другими примерами являются бактериальные кутиназы, происходящие, например, от Pseudomonas, таких как P.mendocina (US 5389536) или P.putida (WO 88/09367).
Альтернативно, исходным липолитическим ферментом может быть эукариотический, например, липолитический фермент грибов, такой как липолитические ферменты семейства Humicola и семейства Zygomycetes и кутиназы грибов.
Примерами кутиназ грибов являются кутиназы Fusarium solani pisi (S. Longhi et al., Journal of Molecular Biology, 268 (4), 779-799 (1997)) и Humicola insolens (США 5827719).
Семейство липолитических ферментов, происходящих от Humicola, состоит из липазы штамма DSM 4109 Н.lanuginosa и липаз, имеющих более чем 50%-ную гомологию с указанной липазой. Липаза Н.lanuginosa (синоним Thermomyces lanuginosus) описана в ЕР 258068 и ЕР 305216 и имеет аминокислотную последовательность, показанную в положениях 1-269 SEQ ID NO: 2 в патенте США 5869438.
Семейство Humicola также включает следующие липолитические ферменты: липазу от Penicillinum camembertii (P25234), липазу/фосфолипазу от Fusarium oxysporum (EP 130064, WO 98/26057), липазу от F.heterosporum (R87979), лизофосфолипазу от Aspergillus foetidus (W33009), фосфолипазу Al от А.oryzae (JP-A 10-155493), липазу от A.oryzae (D85895), липазу/(феруловая кислота)-эстеразу от A.niger (Y09330), липазу/(феруловая кислота)-эстеразу от A.tubingensis (Y09331), липазу от A.tubingensis (WO 98/45453), лизофосфолипазу от A.niger (WO 98/31790), липазу от F.solanii, имеющую изоэлектрическую точку 6,9 и очевидно молекулярную массу 30 кДа (WO 96/18729).
Семейство Zygomycetes включает липазы, имеющие, по крайней мере, 50% гомологию с липазой Rhizomucor miehei (Р19515). Это семейство также включает липазы от Absidia reflexa, A.sporofhora, A.corymbifera, A.blakesleeana, A.griseola (все они описаны в WO 96/13578 и WO 97/27276) и Rhizopus oryzae (P21811). Цифры в скобках указывают на публикацию или номер доступа к базам данных EMBL, GenBank, GeneSeqp или Swiss-Prot.
Особый интерес представляет получение варианта с фосфолипазной активностью из исходного липолитического фермента, не обладающего или обладающего очень незначительной фосфолипазной активностью, например, соответствующей отношению фосфолипазной активности к липазной активности, равному менее 0,1 PHLU/LU или менее 50 PHLU/мг.
Модификация, созданная возле спирт-связывающего сайта
Как было уже установлено, аминокислотная последовательность исходного липолитического фермента может быть модифицирована в положении, которое находится возле глицериновой части триглицеридного субстрата. Эта область будет далее называться "спирт-связывающим участком" липазы, и она описана в Brzozowski A.M. et al., Nature, 351: 491 (1991); Uppenberg et al., Biochemistry, 1995, 34, 16838-16851; A. Svendsen, Inform, 5(5), 619-623 (1994).
Для липазы Rhizomucor miehei протяженность спирт-связывающего сайта может быть определена из файла PDB "5tgl.pdb", доступного в "Структурной классификации белков" (SCOP) в Интернете, в сайте http://www.rcsb.org/pdb/, где показан комплекс с ингибиторным, этиловым сложным эфиром н-гексилфосфоновой кислоты, имитирующим субстрат. Он описан у Derewenda et al. (см. выше), Brzozowski et al. (см. выше) и Brady et al. (см. выше). Положением sn2 этой модели является атом СЕ2.
Указанный вариант обычно содержит не более 10 модификаций в спирт-связывающем сайте, например 1, 2, 3, 4, 5 или 6 модификаций.
Эта модификация может, в частности, находиться в той части спирт-связывающего сайта, которая входит в 20 положений (например, в 10 положений) С-конца.
Как указывалось выше, аминокислотная последовательность исходного липолитического фермента может быть модифицирована в положении, которое находится в пределах 10  (например, в пределах 8
(например, в пределах 8 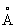 , а в частности в пределах 6
, а в частности в пределах 6 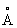 ) от С-атома в положении sn2 глицериновой части триглицеридного субстрата. В липазе Rhizomucor miehei в пределах 10
) от С-атома в положении sn2 глицериновой части триглицеридного субстрата. В липазе Rhizomucor miehei в пределах 10 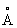 от положения sn2 находятся следующие аминокислотные положения: 25, 28, 80-84, 88, 143-146, 175, 203, 205, 254-255, 257-259, 264-267. В пределах 8
от положения sn2 находятся следующие аминокислотные положения: 25, 28, 80-84, 88, 143-146, 175, 203, 205, 254-255, 257-259, 264-267. В пределах 8  находятся следующие аминокислотные положения: 81-83, 144, 257-258, 265-267, а в пределах 6
находятся следующие аминокислотные положения: 81-83, 144, 257-258, 265-267, а в пределах 6 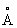 находятся следующие аминокислотные положения: 82, 144, 257, 266.
находятся следующие аминокислотные положения: 82, 144, 257, 266.
В липазе Humicola lanuginosa в пределах 10 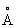 от положения sn2 находятся следующие аминокислотные положения: 18, 21, 81-85, 89, 145-148, 172, 201, 203, 255-256, 258-260, 264-267. В пределах 8
от положения sn2 находятся следующие аминокислотные положения: 18, 21, 81-85, 89, 145-148, 172, 201, 203, 255-256, 258-260, 264-267. В пределах 8 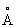 находятся следующие аминокислотные положения: 82-84, 89, 146, 258-259, 265-267, а в пределах 6
находятся следующие аминокислотные положения: 82-84, 89, 146, 258-259, 265-267, а в пределах 6 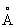 находятся следующие аминокислотные положения: 83, 146, 258, 266.
находятся следующие аминокислотные положения: 83, 146, 258, 266.
Модификация возле каталитической триады
Как уже указывалось в одном из аспектов настоящего изобретения, исходный липолитический фермент имеет структуру, содержащую каталитическую триаду, состоящую из активного остатка Ser, активного остатка Asp и активного остатка His, a модифицируемая аминокислота принадлежит к набору, определенному конкретным способом, описанным выше. Данная структура может быть открытой или замкнутой структурой, и она может включать, а может и не включать субстрат или ингибитор.
Эту процедуру обычно проводят с использованием программного обеспечения, такого как MSI's Insight II. Она предусматривает сопоставление с 4TGL, кристаллической структурой липазы от Rhizomucor miehei, обратимо ингибируемой диэтил-п-нитрофенилфосфатом. Эта структура доступна в "Структурной классификации белков" (SCOP) в Интернете, в сайте http://www.rcsb.org/pdb/, и описана у Derewenda et аl. (см.выше). Липаза от Rhizomucor miehei включает каталитическую триаду, состоящую из аминокислотных остатков S144, D203 и Н257.
Для липазы Humicola lanuginosa, может быть использована структура 1tib; она доступна в "Структурной классификации белков" (SCOP) в Интернете. С использованием этой структуры набор, определенный с помощью указанной процедуры, включает следующие положения: 10-23, 26, 40, 55-64, 80-87, 116-117, 119, 145-149, 151, 168, 170, 194, 196-201, 220-222, 224-227 и 254-269.
Модификация между С-концом и активным остатком His
Как указывалось выше, одна или несколько модификаций в аминокислотной последовательности могут быть созданы между активным остатком His и концом, а в частности, в 12-ти аминокислотах со стороны С-конца от активного остатка His.
Липаза Humicola lanuginosa имеет активный His в Н258 и С-концевой остаток в L269, так что эта область включает положения 259-269. Липаза P.cepacia имеет активный остаток в Н286 и С-концевой остаток в 297, так что эта область включает остатки 287-297.
Модификация возле С-конца
Как указывалось выше, одна или несколько модификаций могут быть сделаны в 10 аминокислотных положениях от С-конца зрелого белка или в положениях, соответствующих указанным положениям в липазе Н.lanuginosa, то есть в положениях 260-269 липазы H.lanuginosa. Соответствующие положения могут быть обнаружены путем сопоставления двух последовательностей, как описано далее в настоящей заявке.
Вариант липолитического фермента может быть усечен путем делеции аминокислотных остатков, соответствующих первым положениям 1, 2, 3, 4, 5 или 6 у С-конца. Усеченный вариант может иметь улучшенную термостабильность.
Альтернативно, данный вариант может иметь пептидное удлинение у С-конца и/или у N-конца. С-концевое удлинение может состоять из 1-10 аминокислотных остатков, например А, Р, AG, DG, PG, AGG, PVGF, AGRF, PRGF, AGGF или AGGFS; либо оно может состоять из 40-50 остатков, например из 48 С-концевых остатков AGGFSWRRYRSAESVDKRATMTDAELEKKLNSYVQMDKEYVKNNQARS липазы Fusarium oxysporum. С-концевое удлинение может повышать фосфолипазную активность.
Ниже описаны некоторые модификации в области, перекрывающейся со спирт-связывающим участком.
Конкретной модификацией является замена в положении, соответствующем G266 в липазе Humicola lanuginosa, в частности, на аминокислоту промежуточного размера, например А, С, D, N, L, I, S, Т, Р или V. Было обнаружено, что одной такой модификации достаточно для повышения фосфолипазной активности.
Другими конкретными модификациями являются такие модификации, которые изменяют третичную структуру, например, путем введения объемных боковых цепей или путем разрушения углов связи, например путем введения Pro. Такие модификации могут быть введены в положения, соответствующие положениям G263, L264, I265, Т267 или L269 в липазе Humicola lanuginosa. Некоторыми конкретными заменами являются G263A, Е, Q, R; L264A, С, Р, Q; I265L, N, Т; Т267А, Q, или L269N.
Модификация в "крышке"
Как было уже установлено выше, аминокислотная последовательность исходного липолитического фермента может быть модифицирована в области "крышки" исходного липолитического фермента. Эта область описана Brady et al., Nature 343, 1990, pp. 767-770 и Brzozowski A.M. et al., Nature, 351: 491 (1991). В липазе Humicola lanuginosa "крышка" расположена в положениях 80-100, и указанная модификация может быть введена в положения 82-98, например 91-98.
Этот вариант обычно содержит не более 5 модификаций в области "крышки"; он может содержать 0, 1, 2 или 3 модификации. Конкретной модификацией является замена аминокислоты, соответствующей G91, L93, N94, D96, К98, L97 и/или Е99 в липазе Humicola lanuginosa на нейтральную или положительно заряженную аминокислоту, например замена, соответствующая G91A,T, L93K, N94D, D96S,W,G, L97Q, K98D,F,E и/или E99K,D.
В частности, вариант с модификацией в области "крышки" также содержит одну или несколько модификаций возле каталитической триады, возле субстрат-связываюшего сайта или возле С-конца.
Варианты липолитического фермента
Вариант липолитического фермента настоящего изобретения содержит одну или несколько модификаций аминокислотного остатка в любой из областей, описанных выше. Каждая модификация может представлять собой делецию или замену аминокислотного остатка, либо она может представлять собой инсерцию перед данным аминокислотным остатком или после него. Если данный аминокислотный остаток находится у С-конца, то указанная инсерция может представлять собой С-концевое удлинение. Инсерция обычно состоит из 1-5 аминокислотных остатков, например 1-2 аминокислотных остатка, а С-концевое удлинение может состоять из 1-50 или 2-10 аминокислотных остатков.
Общее число модификаций в вышеуказанных областях обычно не превышает 20, например не превышает 10 или не превышает 5, и может составлять не более 1 или 2 модификаций в вышеуказанных областях.
Кроме того, указанный вариант липолитического фермента настоящего изобретения может, но необязательно, включать другие модификации исходного фермента, обычно не более 10, например не более 5 таких модификаций.
Указанный вариант обычно имеет гомологию с исходным липолитическим ферментом, составляющую, по крайней мере, 80%, например, по крайней мере, 85%, а обычно, по крайней мере, 90%, или по крайней мере, 95%.
Вариант настоящего изобретения может, кроме того, содержать пептидное удлинение у N-конца, например, состоящее из 1-15 (в частности, 4-10) аминокислотных остатков, а в частности, содержащих 1, 2 или 3 положительно заряженных аминокислоты. Некоторыми конкретными N-концевыми пептидными удлинениями являются AS, SPIRR, E1RP, E1SPIRPRP, E1SPPRRP и E1SPIRPRP. Кроме того, может быть использовано любое пептидное удлинение, описанное в WO 97/04079 и WO 97/07202.
Конкретные варианты
Для получения вариантов липолитического фермента, происходящего от семейства Humicola, могут быть, в частности, созданы модификации в положениях, соответствующих 20-25, 56-64, 81-85 или 255-269 в липазе Humicola lanuginosa. Так, например, такой модификацией может быть замена, делеция или инсерция в положении, соответствующем А20, Y21, G23, К24, N25, V63, R81, G82, R84, А257, W260, Y261, F262 или G266 (например, исключая G23C, К24С, R81C), замена аминокислоты в положении, соответствующем С268 или L269.
Некоторыми конкретными модификациями являются замены, соответствующие нижеследующим положениям в липазе Н. lanuginosa:
Y21V/I/L/A/G/M/W/P/F/N/Q/S/T, V60V/I/L/A/G/M/W/P/F/N/Q/S/T, G61V/I/L/A/G/M/W/P/F/N/Q/S/T, D62E/A/V, S83T, R84K/L/W, Р256А, G263E,Q,R,F, L264A,С,Р,F,G,I, I265L,N,F, G266D/E или T267A,Q,P,S,E, или вставка, соответствующая T267GS или T267GL.
Для изменения активности по отношению к короткоцепочечным (C4-C8)жирным кислотам в триглицеридах могут быть сделаны модификации в положениях, соответствующих Y21, Е56, D57, V60, G61, D62, R81, S83, R84, L259, Y261 или G266, например, замена, соответствующая Y21V/I, V60G, D62E/A/V, S83T, R84K/L/W или G266D/E.
Для повышения активности ДГДГ могут быть сделаны модификации в положениях, соответствующих Y21, G23, N26, D57, D62, R81, S83, R84, S85, G266, Т267 или L269; например, могут быть сделаны две или несколько таких модификаций, например, вместе с одной или несколькими модификациями в области "крышки". Для повышения фосфолипазной активности могут быть сделаны модификации в положениях, соответствующих R81, R84, S85 или 263-267, например G266 или Т267.
Для получения вариантов липазы Pseudomonas могут быть сделаны аминокислотные модификации в положениях, соответствующих 12-13, 16-34, 45-52, 59-66, 68, 86-87, 107-109, 111, 143-153, 155, 157-158, 207-212, 228, 230, 242-249, 264, 279-280, 282-297, 301-302, 304-305, 307-308 в липазе P.cepacia, а в частности, L17/L17, Т18/А18, Y29/Y29, L287/L286, Е289/Е288, I290/I289, Q292/Q291 или L293/L292 в липазе Р.cepacia/P.glumae.
Конкретные варианты липазы Н.lanuginosa описаны в примерах. Соответствующие модификации могут быть сделаны в других исходных липолитических ферментах. Из них могут быть получены другие варианты путем исключения аминокислотных модификаций в положениях 1, 106, 186, 225, 232, 237, 239 или 274. Варианты с 274S могут, но не обязательно, иметь дополнительное С-концевое удлинение WRRYRSAESVDKRATMTDAELEKKLNSYVQMDKEYVKNNQARS (соответствующее С-концу липазы F.oxysporum) в полной или усеченной форме.
Номенклатура для аминокислотных модификаций
Используемая здесь номенклатура для определения мутаций является, в основном, такой, как она была описана в WO 92/05249. Так, например, G91A означает замену G в положении 91 на A. T267A,Q означает замену Т в положении 267 на А или Q. E1E,D,A означает, что Е1 остается неизмененным или замену на D или А.
T267stop означает стоп-кодон, т.е. делецию Т267 и всех последующих аминокислот (то есть С268 и L269). 270Р, 271V означают С-концевое удлинение PV (то есть в новых положениях 270 и 271). -G266 обозначает делецию G в положении 266. Скобки указывают на то, что эта модификация является необязательной или, например, что данная модификация является неточной. SPIRR означает N-концевое удлинение. D266 может означать положение или замену на любую аминокислоту (за исключением D).
E1SPPCGRRP или SPPCGRRP (-Е) означают замену Е1 на SPPCGRRP, то есть присоединение пептида к N-концу. T267GS означает замену Т267 на GS или, другими словами, замену T267G и инсерцию S между G267 и С268.
Гомология и сопоставление
В целях настоящего изобретения степень гомологии может быть соответствующим образом определена с помощью известных компьютерных программ, таких как программа GAP, имеющаяся в программном пакете GCG (Program Manual for the Wisconsin Package, Version 8, August 1994, Genetics Computer Group, 575 Science Drive, Madison, Wisconsin, USA 53711) (Needleman, S.B. & Wunsch C.D.(1970), Journal of Molecular Biology, 48, 443-45), где программа GAP предусматривает нижеследующую установку параметров для сравнения полипептидных последовательностей: введения "штрафа на брешь-пропуск" 3,0 и "штрафа на брешь-удлинение" 0,1.
В настоящем изобретении соответствующие (или гомологичные) положения в липазных последовательностях Rhizomucor miehei (rhiml), Rhizopus delemar (rhidi), Thermomyces lanuginosa (первая; Humicola lanuginosa) (SP400), Penicillium camembertii (Pcl) и Fusarium oxysporum (FoLnp11) определены путем сопоставления, показанного на чертеже.
Для обнаружения гомологичных положений в липазных последовательностях, не показанных в сопоставительном анализе, нужную последовательность сравнивают с первичными последовательностями, показанными на чертеже. Новую последовательность сравнивают в соответствии с сопоставлением, показанным на чертеже, с использованием GAP-сопоставления для большей части гомологичной последовательности, найденной с помощью программы GAP. GAP имеется в программном пакете GCG (Program Manual for the Wisconsin Package, Version 8, August 1994, Genetics Computer Group, 575 Science Drive, Madison, Wisconsin, USA 53711) (Needleman, S.B. & Wunsch C.D. (1970), Journal of Molecular Biology, 48, 443-45). Программа GAP предусматривает нижеследующую установку параметров для сравнения полипептидных последовательностей: введения "штрафа на брешь-пропуск" 3,0 и "штрафа на брешь-удлинение" 0,1.
Варианты с фосфолипазной активностью
Как описано выше, вариант настоящего изобретения может иметь более высокую фосфолипазную активность, чем исходный липолитический фермент. Благодаря методу с использованием монослоя, описанному ниже в данной заявке, этот вариант может иметь фосфолипазную активность, составляющую, по крайней мере, 0,1 нмоль/мин при рН 5.
Благодаря PHLU-методу, описанному ниже в данной заявке, этот вариант может иметь фосфолипазную активность, составляющую, по крайней мере, 100 PHLU/мг (мг чистого ферментного белка), а в частности, по крайней мере, 500 PHLU/мг. Этот вариант имеет отношение фосфолипазной активности к липазной активности (обе активности измерены при рН 7), составляющее, по крайней мере, 0,1 PHLU/LU, например, по крайней мере, 0,5, а в частности, по крайней мере, 2.
Варианты настоящего изобретения могут обладать способностью гидролизовать интактный фосфолипид, как было продемонстрировано с помощью PHLU-метода. Они могут обладать A1- и/или А2-активностью, а поэтому они могут обладать способностью гидролизовать одну или обе ацильные группы жирных кислот в фосфолипиде.
Оптимальный рН
Многие варианты липазы Humicola lanuginosa имеют щелочной рН, оптимальный для липазной активности, и кислотный рН, оптимальный для фосфолипазной активности (например, рН 9-10 для липазы и рН 4-6 для фосфолипазы). Такие варианты могут быть использованы при кислотном рН (например, при рафинировании масла, описанном ниже) в качестве фосфолипаз с сопутствующей очень низкой липазной активностью.
Однако многие варианты липазы Humicola lanuginosa, которые включают замену G266D,E, имеют оптимальный рН как для липазной, так и для фосфолипазной активности, составляющий приблизительно рН 5-6. Такие варианты могут быть использованы при кислотном рН, в случае, когда желательны как липазная, так и фосфолипазная активность, например при выпечке хлеба.
Термостабильность
Термостабильность данного варианта может быть оценена стандартными методами дифференциальной сканирующей калориметрии (ДСК). В зависимости от конкретных мутаций, варианты настоящего изобретения обычно имеют ту же самую или слегка пониженную термостабильность по сравнению с исходным липолитическим ферментом.
Температура на вершине пика денатурации (Тd) липазы Humicola lanuginosa при ее нагревании при 90°С/час и при рН 5 составляет чуть выше 70°С (=Td). Тd для вариантов настоящего изобретения обычно на 5-10 градусов ниже.
Использование варианта
В зависимости от специфичности к субстрату, варианты настоящего изобретения могут быть использованы, например, для улучшения фильтрации, при обработке растительного масла, в хлебопекарном производстве, в моющих средствах или для получения лизофосфолипидов.
Улучшение фильтрации
Вариант с лизофосфолипазной активностью может быть использован для улучшения фильтруемости водного раствора или суспензии углеводного источника путем его обработки указанным вариантом. Этот метод может быть, в частности, применен к раствору или к суспензии, содержащей гидролизат крахмала, а в частности, гидролизат пшеничного крахмала, поскольку его фильтрация представляет определенные трудности и дает мутные фильтраты. Такая обработка может быть выполнена аналогично обработке, описанной в ЕР 219269 (Международный СРС).
Обработка растительного масла
Вариант с фосфолипазной активностью может быть использован в способе снижения содержания фосфолипида в пищевых маслах, предусматривающем обработку данного масла указанным вариантом для гидролиза большей части фосфолипида и отделение водной фазы, содержащей гидролизованный фосфолипид, от масла. Этот способ может быть применен для очистки любого пищевого масла, содержащего фосфолипид, например растительного масла, такого как соевое масло, рапсовое масло и подсолнечное масло. Эта обработка может быть осуществлена при кислотном рН, например рН 3-5. Конкретный вариант может быть, предпочтительно, выбран так, чтобы он имел высокую фосфолипазную активность и низкую липазную активность при низком рН, что обусловлено различием оптимальных рН для этих двух активностей.
Данный способ обработки масла может быть проведен в соответствии с методикой, известной специалистам, например по аналогии с методикой, описанной в патенте США 5264367 (Metallgesellschaft,  ); K.Dahеke & H.Buchold, INFORM, 6(12), 1284-91 (1995); H.Buchold, Fat Sci. Technol., 95(8), 300-304 (1993); JP-A 2-153997 (Showa Sangyo) или ЕР 654527 (Metallgesellschaft,
); K.Dahеke & H.Buchold, INFORM, 6(12), 1284-91 (1995); H.Buchold, Fat Sci. Technol., 95(8), 300-304 (1993); JP-A 2-153997 (Showa Sangyo) или ЕР 654527 (Metallgesellschaft,  ).
).
Использование фосфолипазы в различных целях
Вариант с фосфолипазной активностью может быть использован для получения лизофосфолипида (например, лизолецитина) путем обработки соответствующего фосфолипида указанным вариантом, например, как описано в ЕР 870840, JP-A 10-42884, JP-А 4-135456 или JP-A 2-49593. Этот вариант может быть использован для получения майонеза, например, как описано в ЕР 628256, ЕР 398666 или ЕР 319064.
Вариант с фосфолипазной активностью может быть использован в приготовлении молочных и других пищевых продуктов, например, как описано в ЕР 567662 (Nestle), EP 426211 (Unilever), EP 166284 (Nestle), JP-A 57-189638 (Yakult) или US 4119564 (Unilever).
Данный вариант может быть использован при обработке кожи, как описано в JP-A 7-177884 (Као).
Выпечка хлеба
Вариант с фосфолипазной и/или ДГДГазной активностью может быть использован для приготовления теста, хлеба и пирожных, например для повышения стабильности теста и его пригодности для обработки, либо для улучшения эластичности хлеба или пирожного. Так, например, данный вариант может быть использован в способе приготовления хлеба, предусматривающем добавление указанного варианта в ингредиенты теста, замешивание теста и выпечки хлеба из этого теста. Данный способ может быть осуществлен по аналогии со способом, описанным в патенте США 4567046 (Kyowa Hakko), JP-A 60-78529 (QP Corp.), JP-A 62-111629 (QP Corp.), JP-A 63-258528 (QP Corp.), EP 426211 (Unilever) или в WO 99/53769 (Novo Nordisk).
Особенно предпочтительно использовать данный вариант вместе с эндоамилазой, препятствующей черствению, и также можно, но необязательно, добавлять фосфолипид для уменьшения черствения хлеба, а в частности, для увеличения мягкости хлеба в течение первых 24 часов после его выпечки. Эта эндоамилаза может представлять собой мальтозообразующую α-амилазу (например, от Bacillus sp., такую как Novamyl® от Novo Nordisk), либо грибковую или бактериальную α-амилазу, например, от Aspergillus или Bacillus, а в частности, А.oryzae, В.lichenformis или В.amyloliquefaciens.
При использовании в хлебопечении данный вариант может обладать низкой активностью по отношению к короткоцепочечным или среднецепочечным (С4-С8)жирным кислотам, например активностью, соответствующей отношению SLU/LU выше 3. Использование такого варианта позволяет предотвращать или подавлять возникновение нежелательного запаха, обусловленное высвобождением короткоцепочечных жирных кислот. Этот вариант может обладать активностью по отношению к триглицеридам и фосфолипиду, а также к ДГДГ.
Запах сыра
Вариант фермента с активностью, направленной на ацильные группы короткоцепочечных жирных кислот, может быть использован для высвобождения свободных жирных кислот (СЖК) в целях формирования определенного запаха в пищевых продуктах, например, при созревании сыра, например, как описано Hanson, ZFL, 41 (10), 664-666 (1990)).
Варианты липолитического фермента с повышенной способностью к высвобождению короткоцепочечных жирных кислот, в отличие от высвобождения длинноцепочечных жирных кислот из жирного молока, могут быть использованы при производстве сыра, например, для усиления запаха или для ускорения времени созревания для созревающих сыров, таких как чеддер или пармезан. Другим применением таких вариантов липолитического фермента является их использование для получения ферментативно модифицированного сыра (ЕМС) и в качестве отдушки для различных пищевых продуктов, включая сыры, заправки и легкие завтраки.
Высвобождение короткоцепочечных жирных кислот, таких как масляная кислота, является главным фактором для продуцирования сырного запаха, тогда как высвобождение длинноцепочечных жирных кислот, таких как олеиновая кислота, приводит к изменению запаха. Варианты липолитического фермента, используемые при изготовлении сыров, включая ЕМС, должны иметь отношение SLU/LU менее чем 0,5, например, менее чем 0,25, а наиболее предпочтительно менее чем 0,1.
Использование в моющих средствах
Данный вариант может быть использован в качестве моющей добавки, например, при концентрации (выражаемой в количестве чистого ферментного белка) 0,001-10 (например, 0,01-1) мг на грамм моющего средства или 0,001-100 (например, 0,01-10) мг на литр моющей жидкости.
При использовании в моющих средствах для облегчения удаления жирных пятен данный вариант может иметь высокую активность по отношению к длинноцепочечным (C16-C20)триглицеридам. Данный вариант может обладать фосфолипазной активностью. Данный вариант может обладать низкой активностью по отношению к короткоцепочечным (C4-C8)жирным кислотам в триглицеридах, например, соответствующей отношению SLU/LU, превышающему 10. Использование такого варианта позволяет предотвращать или подавлять возникновение нежелательного запаха, обусловленное высвобождением короткоцепочечных жирных кислот.
В моющих средствах могут быть использованы варианты, обладающие как липазной, так и фосфолипазной активностью, при щелочном рН.
Композиция моющего средства
Композиция моющего средства настоящего изобретения может быть, например, приготовлена в виде композиции моющего средства как для ручной стирки, так и для стирки в стиральной машине, включающего вспомогательную композицию, подходящую для предварительной обработки окрашенных тканей, и вспомогательную композицию-мягчитель для полоскания тканей, либо она может быть приготовлена в виде композиции моющего средства для использования в домашнем хозяйстве для чистки твердых поверхностей. В моющем средстве для стирки данный вариант может быть эффективным для удаления жирных пятен, для сохранения белизны изделия и для очистки от грязи. Композиция моющего средства для стирки может быть изготовлена, как описано в WO 97/04079, WO 97/07202, WO 97/41212, PCT/DK WO 98/08939 и WO 97/43375.
Композиция моющего средства настоящего изобретения может быть, в частности, изготовлена для мытья посуды вручную и в посудомоечной машине, например, как описано в патенте GB 2247025 (Unilever) или WO 99/01531 (Procter & Gamble). В композиции для мытья посуды данный вариант может быть эффективным для удаления жирных/масляных пятен, для предупреждения окрашивания/обесцвечивания посуды и пластиковых посудомоечных компонентов интенсивно окрашенными компонентами и для предотвращения осаждения извести и мыла на посуду.
Композиция моющего средства настоящего изобретения может быть изготовлена в любой стандартной форме, например в виде куска, таблетки, порошка, гранул, пасты или жидкости. Жидкое моющее средство может быть водным раствором, обычно содержащим вплоть до 70% воды и 0-30% органического растворителя, или безводным раствором.
Указанная композиция моющего средства содержит одно или несколько поверхностно-активных веществ, которые могут быть неионогенными, включая полуполярные поверхностно-активные вещества, и/или анионогенными, и/или катионогенными, и/или цвиттерионными поверхностно-активными веществами. Эти поверхностно-активные вещества обычно присутствуют в количестве от 0,1 до 60 мас.%, например 0,5-40%, а именно 1-30%, а обычно 1,5-20%.
Если моющее средство включает указанное поверхностно-активное вещество, то это моющее средство содержит от около 1% до около 40% анионогенного поверхностно-активного вещества, такого как линейный алкилбензолсульфонат, альфа-олефинсульфонат, алкилсульфат (сульфат жирного спирта), этоксисульфат спирта, вторичный алкансульфонат, метиловый эфир альфа-сульфожирной кислоты, алкил- или алкенилянтарную кислоту или мыло.
Если моющее средство включает указанное поверхностно-активное вещество, то это моющее средство обычно содержит от около 0,2% до около 40% неионогенного поверхностно-активного вещества, такого как этоксилированный спирт, этоксилированный нонил-фенол, алкилполигликозид, оксид алкилдиметиламина, этоксилированный моноэтаноламид жирной кислоты, моноэтаноламид жирной кислоты, амид полигидроксиалкилжирной кислоты или N-ацил-N-алкиловые производные глюкозамина ("глюкамиды").
Настоящее изобретение также относится к моющим добавкам, включающим вариант настоящего изобретения. Такая моющая добавка, а также композиция моющего средства могут включать один или несколько других ферментов, таких как протеаза, липаза, кутиназа, амилаза, карбогидраза, целлюлаза, пектиназа, маннаназа, арабиназа, галактаназа, ксиланаза, оксидаза, например лаксаза и/или пероксидаза.
В основном, свойства выбранного(ых) фермента(ов) должны быть совместимыми с выбранным моющим средством (т.е. оптимальный рН, совместимость с другими ферментными и неферментными ингредиентами и т.п.), и этот фермент(ы) должен присутствовать в эффективных количествах.
Протеазы: подходящими протеазами являются протеазы животного, растительного или микробного происхождения. Предпочтительными являются протеазы микробного происхождения. Подходящими также являются химически модифицированные и сконструированные белки-мутанты. Указанной протеазой может быть сериновая протеаза или металлопротеаза, например щелочная микробная протеаза или трипсин-подобная протеаза. Примерами щелочных протеаз являются субтилизины, а в частности, субтилизины, происходящие от Bacillus, например субтилизин Novo, субтилизин Carlsberg, субтилизин 309, субтилизин 147 и субтилизин 168 (описанные в WO 89/06279). Примерами трипсин-подобных протеаз являются трипсин (например, происходящий от свиньи или быка) и протеаза Fusarium, описанная в WO 89/06270 и WO 94/25583.
Примерами используемых протеаз являются варианты, описанные в WO 92/19729, WO 98/20115, WO 98/20116 и WO 98/34946, а в частности, варианты с заменами в одном или в нескольких из следующих положений: 27, 36, 57, 76, 87, 97, 101, 104, 120, 123, 167, 170, 194, 206, 218, 222, 224, 235 и 274.
Конкретными коммерчески доступными протеазными ферментами являются Алкалаза® (Alcalase®), Савиназа® (Savinase®), Примаза® (Primase®), Дуралаза® (Duralase®), Эспераза® Esperase®) и Канназа® (Kannase®) (Novo Nordisk A/S), Максатаза® (Maxatase®), Максакал® (Maxacal®), Максапем® (Maxapem®), Пропераза® (Properase®), Пурафект® (Purafect®), Пурафект ОхР® (Purafect OxP®), FN2™ и FN3™ (Genencor International Inc.).
Целлюлазы: подходящими целлюлазами являются целлюлазы бактериального или грибкового происхождения. Подходящими также являются химически модифицированные и сконструированные белки-мутанты. Подходящими целлюлазами являются целлюлазы, происходящие от бактерий рода Bacillus, Pseudomonas, Humicola, Fusarium, Thielavia, Acremonium, например грибковые целлюлазы, происходящие от Humicola insolens, Myceliophthora thermophila и Fusarium oxysporum, описанные в патентах США 4435307, 5648263, 5691178, 5776757 и WO 89/09259.
Особенно подходящими целлюлазами являются щелочные или нейтральные целлюлазы, обеспечивающие цветостойкость. Примерами таких целлюлаз являются целлюлазы, описанные в ЕР 0495257, ЕР 0531372, WO 96/11262, WO 96/29397, WO 98/08940. Другими примерами являются варианты целлюлаз, описанные в WO 94/07998, ЕР 0531315, патентах США 5457046, 5686593, 5763254, WO 95/24471, WO 98/12307 и PCT/DK 98/00299.
Коммерчески доступными целлюлазами являются Целлузим® (Celluzyme®) и Карезим® (Carezyme®) (Novo Nordisk A/S), Клазиназа® (Clazinase®) и Пурадакс НА® (Puradax НА®) (Genencor International Inc.), и КАС-500(В)® (Као Corporation).
Пероксидазы/оксидазы: подходящими пероксидазами/оксидазами являются пероксидазы/оксидазы растительного, бактериального или грибкового происхождения. Подходящими также являются химически модифицированные и сконструированные белки-мутанты. Примерами подходящих пероксидаз являются пероксидазы, происходящие от Coprinus, например С. cinereus, и их варианты, описанные в WO 93/24618, WO 95/10602 и WO 98/152257.
Коммерчески доступными пероксидазами являются Гвардзим® (Guardzyme®) (Novo Nordisk A/S).
Ферменты, используемые в моющих средствах, могут быть включены в композицию моющего средства путем добавления отдельных добавок, содержащих один или несколько ферментов, либо путем добавления комбинированной добавки, содержащей все указанные ферменты. Моющая добавка настоящего изобретения, то есть отдельная добавка или комбинированная добавка, может быть изготовлена, например, в виде гранул, жидкости, суспензии и т.п. Конкретными композициями, содержащими моющие добавки, являются грануляты, а в частности, непудрообразные грануляты, жидкости, а в частности, стабилизированные жидкости, или суспензии.
Непудрообразные грануляты могут быть получены, например, как описано в патентах США 4106991 и 4661452, и на них может быть, но необязательно, нанесено покрытие известными методами. Примерами материалов для воскообразного покрытия являются полиэтиленоксидные продукты (полиэтиленгликоль, ПЭГ) со средней молярной массой от 1000 до 20000; этоксилированные нонилфенолы, имеющие от 16 до 50 этиленоксидных звеньев; этоксилированные жирные спирты, в которых спирт содержит от 12 до 20 атомов углерода и в которых присутствует 15-80 этиленоксидных звеньев; жирные спирты; жирные кислоты; и моно-, ди- и триглицериды жирных кислот. Примеры пленкообразующих материалов для покрытия, подходящих для применения в методах с использованием псевдоожиженного слоя, представлены в GB 1483591. Жидкие ферментные препараты могут быть, например, стабилизированы путем добавления полиола, такого как пропиленгликоль, сахара или спирта ряда сахаров, молочной кислоты или борной кислоты в соответствии с разработанными методами. Защищенные ферменты могут быть получены методом, описанным в ЕР 238216.
Данное моющее средство может содержать 0-65% моющего компонента или комплексообразующего агента, такого как цеолит, дифосфат, трифосфат, фосфонат, карбонат, цитрат, нитрилотриуксусная кислота, этилендиаминтетрауксусная кислота, диэтилентриаминопентауксусная кислота, алкил- или алкенилянтарная кислота, растворимые силикаты или слоистые силикаты (например, SKS-6 от Hoechst).
Данное моющее средство может содержать один или несколько полимеров. Примерами являются карбоксиметилцеллюлоза, поли(винилпирролидон), поли(этиленгликоль), поливиниловый спирт, поли(винилпиридин-N-оксид), поли(винилимидазол), поликарбоксилаты, такие как полиакрилаты, сополимеры малеиновой/акриловой кислоты и сополимеры лаурилметакрилата/акриловой кислоты.
Данное моющее средство может содержать отбеливающую систему, которая может включать источник H2O2, такой как перборат или перкарбонат, который может быть объединен с перкислотообразующим активатором-отбеливателем, таким как тетраацетилэтилендиамин или нонаноилоксибензолсульфонат. Альтернативно, данная отбеливающая система может содержать пероксикислоты, например, амидного, имидного или сульфонового типа.
Ферменты в композиции моющего средства настоящего изобретения могут быть стабилизированы с использованием стандартных стабилизирующих агентов, например полиола, такого как пропиленгликоль или глицерин, сахара или спирта ряда сахаров, молочной кислоты, борной кислоты или производного борной кислоты, например ароматического эфира борной кислоты, производного фенилбороновой кислоты, такое как 4-формилфенилбороновая кислота, и эта композиция может быть изготовлена, как описано, например, в WO 92/19709 и WO 92/19708.
Данное моющее средство может также содержать и другие стандартные ингредиенты, обычно используемые в моющих средствах, такие как, например, кондиционирующая добавка для тканей, включая глины, пенообразователи, ингибиторы мыльной пены, противокоррозионные агенты, агенты, суспендирующие загрязнения, добавки, предотвращающие ресорбцию загрязнений, красители, бактериоциды, оптические осветлители, гидротропы, ингибиторы потускнения или ароматизирующие добавки.
При этом считается, что в рассматриваемых композициях моющих средств любой фермент, а в частности, вариант настоящего изобретения, может быть добавлен в количестве, соответствующем 0,01-100 мг ферментного белка на литр моющей жидкости, например 0,05-5 мг ферментного белка на литр моющей жидкости, а в частности, 0,1-1 мг ферментного белка на литр моющей жидкости.
Вариант настоящего изобретения может быть дополнительно включен в композиции моющих средств, описанные в заявке WO 97/07202, которая вводится в настоящее описание посредством ссылки.
Способы получения вариантов ферментов
Вариант фермента настоящего изобретения может быть получен известными методами, например методами, описанными в WO 97/04079 (Novo Nordisk). Ниже описаны методы клонирования ДНК-последовательностей, кодирующих данный фермент, а также методы создания мутаций в конкретных сайтах последовательности, кодирующей данный фермент.
Клонирование ДНК-последовательности, кодирующей фермент
ДНК-последовательность, кодирующая исходный фермент, может быть выделена из любой клетки или микроорганизма, продуцирующих нужный фермент, с использованием различных способов, хорошо известных специалистам. Сначала должна быть сконструирована библиотека геномных ДНК и/или кДНК с использованием хромосомной ДНК или матричной РНК от организма, продуцирующего исследуемый фермент. Затем, в том случае, если аминокислотная последовательность этого фермента известна, то из геномной библиотеки, полученной от нужного организма, могут быть синтезированы меченные олигонуклеотидные зонды, которые могут быть использованы для идентификации клонов, кодирующих данный фермент. Альтернативно, меченный олигонуклеотидный зонд, содержащий последовательности, гомологичные гену другого известного фермента, может быть использован в качестве зонда для идентификации клонов, кодирующих данный фермент, с использованием условий гибридизации и промывки пониженной строгости.
Еще один метод идентификации клонов, кодирующих данный фермент, предусматривает встраивание фрагментов геномной ДНК в экспрессирующий вектор, такой как плазмида, трансформацию негативных по данному ферменту бактерий полученной библиотекой геномных ДНК, а затем высевание этих трансформированных бактерий на агар, содержащий субстрат для фермента (то есть мальтозу), что, тем самым, позволяет идентифицировать клоны, экспрессирующие данный фермент.
Альтернативно, ДНК-последовательность, кодирующая данный фермент, может быть получена синтетически с помощью разработанных стандартных методов, например фосфороамидитным методом, описаннным S.L. Beaucage & М.Н. Caruthers, (1981), Tetrahedron Letters 22, р. 1859-1869, или методом, описанным Matthes et al., (1984), EMBO J. 3, р.801-805. В фосфороамидитном методе олигонуклеотиды синтезируют, например, в автоматическом ДНК-синтезаторе, очищают, отжигают, лигируют и клонируют в соответствующих векторах.
И наконец, данная ДНК-последовательность может иметь смешанное геномное и синтетическое происхождение; смешанное синтетическое и кДНК происхождение, или смешанное геномное и кДНК происхождение, полученная путем лигирования фрагментов синтетического, геномного или кДНК происхождения (если это необходимо, то она может представлять собой фрагменты, соответствующие разным частям целой ДНК-последовательности) стандартными методами. Указанная ДНК-последовательность может быть также получена с помощью полимеразной цепной реакции (ПЦР) с использованием специфических праймеров, например, как описано в патенте США 4683202 или R.K. Saiki et al., (1988), Science 239, 1988, pp. 487-491.
Сайт-специфический мутагенез
После выделения ДНК-последовательности, кодирующей данный фермент, и идентификации нужных сайтов для введения мутаций эти мутации могут быть введены с использованием синтетических олигонуклеотидов. Эти олигонуклеотиды содержат нуклеотидные последовательности, фланкирующие нужные сайты мутации. В конкретном методе, в векторе, несущем ген данного фермента, создают одноцепочечную брешь ДНК, т.е. последовательности, кодирующей данный фермент. Затем синтетический нуклеотид, несущий нужную мутацию, подвергают отжигу с образованием гомологичной части одноцепочечной ДНК. Затем оставшуюся брешь достраивают ДНК-полимеразой I (фрагментом Кленова), и эту конструкцию лигируют с использованием лигазы Т4. Конкретный пример этого метода описан в работе Morinaga et al., (1984), Biotechnology 2, р.646-639. В патенте США 4760025 описано введение олигонуклеотидов, кодирующих множественные мутации, путем создания небольших модификаций в кластере. Однако с использованием метода Morinaga в одно и то же время можно вводить даже более широкий ряд мутаций, поскольку может быть введено множество олигонуклеотидов различной длины.
Другой метод введения мутаций в ДНК-последовательность, кодирующую фермент, описан в работе Nelson & Long (1989), Analytical Biochemistry 180, р.147-151. Этот метод предусматривает 3-стадийное генерирование ПЦР-фрагмента, содержащего нужную мутацию, введенную с использованием химически синтезированной ДНК-цепи в качестве одного из праймеров в ПЦР-реакциях. Из ПЦР-генерированного фрагмента ДНК-фрагмент, несущий указанную мутацию, может быть выделен путем расщепления рестриктирующими эндонуклеазами и снова встроен в экспрессирующую плазмиду.
Кроме того, в работах Sierks et al., (1989) "Site-directed mutagenesis at the active site Trpl20 of Aspergillus awamori glucoamilase. Protein Eng., 2, 621-625; Sierks et al., (1990) "Catalytic mechanism of fungal glucoamylase as defined by mutagenesis of Aspl76, Glul79 and Glu180 in the enzyme from Aspergillus awamori", Protein Eng., vol.3, 193-198 также описан сайт-направленный мутагенез в глюкоамилазе Aspergillus.
Экспрессия вариантов фермента
В соответствии с настоящим изобретением ДНК-последовательность, кодирующая данный вариант, продуцируемый методами, описанными выше или любыми известными альтернативными методами, может быть экспрессирована в виде фермента с использованием экспрессирующего вектора, который обычно содержит регуляторные последовательности, кодирующие промотор, оператор, сайт связывания с рибосомой, сигнал инициации трансляции, и, необязательно, ген-репрессор или различные гены-активаторы.
Экспрессирующий вектор
Рекомбинантным экспрессирующим вектором, несущим ДНК-последовательность, кодирующую вариант фермента настоящего изобретения, может быть любой вектор, который может быть подвергнут стандартной технике рекомбинантных ДНК, и выбор такого вектора часто зависит от клетки-хозяина, в которую он должен быть встроен. Данный вектор при его введении в клетку-хозяина может быть интегрирован в геном этой клетки-хозяина и реплицирован вместе с хромосомой(ами), в которую он был интегрирован. Примером подходящих экспрессирующих векторов является рМТ838.
Промотор
В данном векторе ДНК-последовательность должна быть функционально присоединена к подходящей промоторной последовательности. Таким промотором может быть любая ДНК-последовательность, которая обладает транскрипционной активностью в данной клетке-хозяине и может происходить от генов, кодирующих белки, либо гомологичные, либо гетерологичные белкам данной клетки-хозяина.
Примерами промоторов, подходящих для регуляции транскрипции ДНК-последовательности, кодирующей вариант фермента настоящего изобретения, в частности, в бактериальном хозяине, являются промотор laс-оперона E.coli, промоторы гена dagA агаразы Streptomyces coelicolor, промоторы гена α-амилазы Bacillus lichenformis (amyL), промоторы гена мальтозообразующей амилазы Bacillus stearothermophilus (аmyМ), промоторы α-амилазы Bacillus amyloliquefaciens (amyQ), промоторы генов хylА и xylB Bacillus subtilis и т.п. Для транскрипции в грибковом хозяине примерами подходящих промоторов являются промоторы, происходящие от гена, кодирующего амилазу ТАКА A.oryzae, промотор TPI (триозофосфат-изомеразы) от S.cerevisiae (Alber et al. (1982), J.Mol. Appl. Genet 1, p. 419-434), промоторы (аспарагиновая кислота)-протеиназы Rhizomucor miehei, нейтральной α-амилазы A.niger, кислотоустойчивой α-амилазы A.niger, глюкоамилазы A.niger, липазы Rhizomucor miehei, щелочной протеазы A.oryzae, триозофосфат-изомеразы A.oryzae или ацетамидазы A.nidulans.
Экспрессирующий вектор
Экспрессирующий вектор настоящего изобретения может также содержать подходящий терминатор транскрипции и в эукариотах последовательности полиаденилирования, функционально присоединенные к ДНК-последовательности, кодирующей вариант α-амилазы настоящего изобретения. Последовательности терминации и полиаденилирования могут быть соответствующим образом получены из тех же самых источников, что и промотор.
Данный вектор, кроме того, может содержать ДНК-последовательность, позволяющую этому вектору реплицироваться в нужной клетке-хозяине. Примерами таких последовательностей являются сайты инициации репликации плазмид pUC19, pACYC177, pUB110, pE194, pAMB1 и pIJ702.
Этот вектор может также содержать селективный маркер, например ген, продукт которого комплементирует дефект в клетке-хозяине, такой как гены dal от В.subtilis или В.licheniformis, либо гены, которые придают резистентность к антибиотику, такую как резистентность к ампициллину, канамицину, хлорамфениколу или тетрациклину. Кроме того, данный вектор может содержать селективные маркеры, Aspergillus, такие как amdS, argB, niaD и sC, маркеры, определяющие повышение резистентности к гигромицину, либо такой отбор может быть осуществлен путем ко-трансформации, например, как описано в WO 91/17243.
Процедуры, используемые для лигирования ДНК-конструкции настоящего изобретения, кодирующей вариант фермента, промотор, терминатор и другие элементы, соответственно, и для их встраивания в подходящие векторы, содержащие необходимую информацию для репликации, хорошо известны специалистам (см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor, 1989).
Клетки-хозяева
Клетка настоящего изобретения, содержащая либо ДНК-конструкцию, либо экспрессирующий вектор настоящего изобретения, описанный выше, преимущественно используется в качестве клетки-хозяина в рекомбинантном продуцировании варианта фермента настоящего изобретения. Эта клетка может быть трансформирована ДНК-конструкцией настоящего изобретения, кодирующей указанный вариант, обычно путем интегрирования этой ДНК-конструкции (в одной или нескольких копиях) в хромосоме хозяина. Обычно считается, что такая интеграция имеет определенное преимущество, поскольку, более вероятно, что эта ДНК-последовательность будет стабильно поддерживаться в клетке. Интегрирование ДНК-конструкций в хромосому хозяина может быть осуществлено стандартными методами, например путем гомологичной или гетерологичной рекомбинации. Альтернативно, эта клетка может быть трансформирована экспрессирующим вектором, описанным выше, в связи с клетками-хозяевами различных типов.
Клеткой настоящего изобретения может быть клетка высшего организма, такого как млекопитающее или насекомое, но она может быть и микробной клеткой, например бактериальной или грибковой клеткой (включая дрожжи).
Примерами подходящих бактерий являются грамм-положительные бактерии, такие как Bacillus subtilis, Bacillus licheniformis, Bacillus lentus, Bacillus brevis, Bacillus stearothermophilus, Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus coagulans, Bacillus circulans, Bacillus lautus, Bacillus megaterium, Bacillus thuringiensis, или Streptomyces lividans или Streptomyces murinus, либо грамм-отрицательные бактерии, такие как E.coli. Трансформация этих бактерий может быть проведена, например, путем трансформации протопластов или с использованием компетентных клеток способом, известным per se.
Дрожжевой микроорганизм может быть предпочтительно выбран из вида Saccharomyces или Schizosaccharomyces, например Saccharomyces cerevisiae.
Клеткой-хозяином могут быть также гифомицеты, например, штамм, принадлежащий к виду Aspergillus, такой как Aspergillus oryzae или Aspergillus niger, или штамм Fusarium, такой как штамм Fusarium oxysporium, Fusarium graminearum (в совершенной стадии называемый Gibberella zeae, ранее называемый Sphaeria zeae, синоним Gibberella гоsеum и Gibberella roseum f. sp. cerealis), или Fusarium sulphureum (в совершенной стадии называемый Gibberella puricaris, синоним Fusarium trichothecioides, Fusarium bacteridioides, Fusarium sambucium, Fusarium roseum и Fusarium roseum var. gramineanum), Fusarium cerealis (синоним Fusarium crokkwellnse) или Fusarium venenatum.
В конкретном варианте настоящего изобретения клеткой-хозяином является дефицитный по протеазе штамм "протеаза-минус".
Таким штаммом может быть дефицитный по протеазе штамм JaL 125 Aspergillus oryzae, у которого делегирован ген щелочной протеазы, называемый "alp". Этот штамм описан в WO 97/35956 (Novo Nordisk).
Клетки гифомицетов могут быть трансформированы способом, предусматривающим образование протопластов и трансформацию протопластов, с последующей регенерацией клеточных стенок методом, известным per se. Использование Aspergillus в качестве микроорганизма-хозяина описано в ЕР 238023 (Novo Nordisk A/S), содержание которого вводится в настоящее описание посредством ссылки.
Способ продуцирования варианта фермента настоящего изобретения
Вариант фермента настоящего изобретения может быть продуцирован методом, предусматривающим культивирование клетки-хозяина в условиях, способствующих продуцированию данного варианта, и выделение этого варианта из клетки и/или культуральной среды.
Средой, используемой для культивирования клеток, может быть любая стандартная среда, подходящая для культивирования нужной клетки-хозяина и осуществления экспрессии варианта фермента настоящего изобретения. Подходящими средами являются коммерчески доступные среды, либо эти среды могут быть получены известными способами (например, как описано в каталогах Американской коллекции типовых культур).
Вариант фермента, секретированный из клеток-хозяев, может быть легко выделен из культуральной среды хорошо известными методами, включая выделение клеток из данной среды путем центрифугирования или фильтрации и преципитацию белковых компонентов этой среды с использованием соли, такой как сульфат аммония, с последующим применением хроматографических методов, таких как ионообменная хроматография, аффинная хроматография или т.п.
Экспрессия варианта фермента в растениях
Настоящее изобретение также относится к трансгенному растению, части растения или клетки растения, которые были трансформированы ДНК-последовательностью, кодирующей вариант настоящего изобретения, так, чтобы они экспрессировали и продуцировали этот фермент в выделяемых количествах. Данный фермент может быть выделен из указанного растения или части растения. Альтернативно, может быть использовано само растение или его часть, содержащая рекомбинантный фермент.
Трансгенные растения могут быть двудольными или однодольными, короче говоря, содержащими две семядоли или одну семядолю. Примерами однодольных растений являются травы, такие как луговые травы (пырей, злаки (Роа)), кормовые травы, такие как овсянница (festuca), плевел (lolium), травы умеренного пояса, такие как злаковые (Agrostis) и зерновые культуры, например пшеница, овес, рожь, ячмень, рис, сорго и маис (кукуруза).
Примерами двудольных растений являются табак, бобовые, такие как люпин, картофель, сахарная свекла, горох, фасоль и соя, и крестоцветные (семейства Brassicaceae), такие как цветная капуста, масличный рапс и близкородственный модельный организм резушка Таля (Arabidopsis thaliana).
Примерами частей растений являются стебель, каллюс, листья, корни, плоды, семена и клубни. В контексте настоящего описания частями растения также считаются растительные ткани, такие как хлоропласт, апопласт, митохондрии, вакуоль, пероксисомы и цитоплазм. Кроме того, частью растения считается любая клетка растения независимо от происхождения ткани.
В объем настоящего изобретения также входит потомство таких растений, части растений и клетки растений.
Трансгенное растение или растительная клетка, экспрессирующие вариант фермента настоящего изобретения, могут быть сконструированы известными методами. Вкратце, растение или растительную клетку конструируют путем введения одной или нескольких экспрессирующих конструкций, кодирующих вариант настоящего изобретения, в геном растения-хозяина и размножения полученного модифицированного растения или клетки растения с продуцированием трансгенного растения или трансгенной растительной клетки.
Обычно, экспрессионная конструкция представляет собой ДНК-конструкцию, содержащую ген, кодирующий вариант настоящего изобретения и функционально присоединенный к соответствующим регуляторным последовательностям, необходимым для экспрессии данного гена в нужном растении или части этого растения. Кроме того, эта экспрессионная конструкция может содержать селективный маркер, подходящий для идентификации клеток-хозяев, в которые была интегрирована данная экспрессионная конструкция, и ДНК-последовательности, необходимые для введения этой конструкции в нужное растение (которое выбирают в зависимости от используемого метода введения ДНК).
Выбор регуляторных последовательностей, таких как промотор и терминатор, и необязательно сигнальных или транзиционных последовательностей осуществляют, например, в зависимости от того, когда, где и как экспрессируется нужный фермент. Так, например, экспрессия гена, кодирующего вариант настоящего изобретения, может быть конститутивной или индуцибельной, либо она может быть времяспецифической, стадиеспецифической или тканеспецифической, а данный генный продукт может быть нацелен на конкретную ткань или часть растения, такую как семена или листья. Регуляторные последовательности описаны, например, Tague et al.. Plant, Phys., 86, 506, 1988.
Для конститутивной экспрессии может быть использован промотор 353-CaMV (Franck et al., 1980, Cell 21:285-294).
Органспецифическими промоторами могут быть, например, промотор от тканей запасающего стока, таких как семена, клубни картофеля и плоды (Edwards & Coruzzi, 1990. Annu. Rev. Genet. 24:275-303), или от тканей метаболического стока, таких как меристемы (Ito et al., 1994, Plant Mol. Biol. 24: 863-878), семяспецифический промотор, такой как промотор глутелина, проламина, глобулина или альбумина риса (Wu et al., Plant and Cell Physiology Vol. 39, №8, pp. 885-889 (1998)), промотор Vicia faba бобов В4 и неизвестного гена белка семян от Vicia faba, описанного Conrad U. et al., Journal of Plant Physiology Vol. 152, № 6, pp. 708-711 (1998), промотор белка масляного тела семян (Chen et al., Plant and cell physiology vol. 39, № 9, pp. 935-941 (1998), промотор гена nарА запасаемого белка от Brassica napus, или какой-либо другой известный семяспецифический промотор, описанный, например, в WO 91/14772. Кроме того, этим промотором может быть листьеспецифический промотор, такой как промотор гена rbcs риса или томатов (Kyozuka et al., Plant Physiology Vol. 102, № 3, pp.991-1000 (1993)), промотор гена аденинметил-трансферазы вируса хлореллы (Mitra А. & Higgins, DW, Plant Molecular Biology Vol. 26, № 1, pp.85-93 (1994)) или промотор гена aldP риса (Kagaya et al., Molecular and General Genetics Vol.248, № 6, pp.668-674 (1995)), или индуцируемый повреждением промотор, такой как промотор гена pin2 картофеля (Xu et al., Plant Molecular Biology Vol. 22, № 4, pp.573-588 (1993)).
Элемент энхансера промотора может быть использован для осуществления более высокого уровня экспрессии данного фермента в растении. Так, например, таким элементом энхансера промотора может быть интрон, который расположен между промотором и нуклеотидной последовательностью, кодирующей данный фермент. Так, например, в работе Xu et al., op cit, описано использование первого интрона гена актина риса 1 для усиления экспрессии.
Ген селективного маркера и любые другие части экспрессионной конструкции могут быть выбраны из известных генов и других элементов.
ДНК-конструкцию встраивают в геном растения в соответствии со стандартными методами, включая опосредованную агробактерией трансформацию, вирус-опосредованную трансформацию, микроинъекцию, бомбардировку частицами, баллистическую трансформацию и электропорацию (Gasser et al., Science, 244, 1293; Potrykus, Bio/Techn. 8, 535, 1990; Shimamoto et al., Nature, 338, 274, 1989).
В настоящее время Agrobacterium tumefaciens-опосредованный перенос гена является одним из методов отбора для генерирования трансгенных двудольных растений (см. обзор Hooykas & Schilperoort, 1992, Plant Mol.Biol. 19: 15-38), однако этот метод может быть также использован для трансформации однодольных растений, хотя для этих растений обычно используют другие методы трансформации. В настоящем изобретении для генерирования трансгенных однодольных растений был выбран метод, предусматривающий бомбардировку частицами (микроскопическими золотыми или вольфрамовыми частицами, покрытыми трансформирующей ДНК) эмбрионных каллюсов или развивающихся эмбрионов (Christou, 1992, Plant J., 2: 275-281; Shimamoto, 1994, Curr. Opin. Biotechnol. 5: 158-162; Vasil et al., 1992, Bio/Technology 10: 667-674). Альтернативный метод трансформации однодольных растений основан на трансформации протопластов, описанной Omirulleh S. et al., Plant Molecular Biology Vol.21, No 3, pp. 415-428 (1993).
После трансформации трансформанты, содержащие встроенную экспрессионную конструкцию, отбирают и регенерируют с получением целых растений в соответствии с методами, хорошо известными специалистам.
Материалы и методы
Липазная активность по отношению к трибутирину (LU)
Субстрат для липазы получают путем эмульгирования трибутирина (трибутирата глицерина) с использованием аравийской камеди в качестве эмульгатора. Гидролиз трибутирина при 30°С и при рН 7 осуществляют в эксперименте по титрованию с использованием рН-стата. Одна единица липазной активности (1 LU) равна количеству фермента, способного высвобождать 1 мкмоль масляной кислоты/мин в стандартных условиях.
Липазная активность по отношению к триолеину (SLU)
Липолитическая активность может быть определена с использованием оливкового масла в качестве субстрата.
В этом SLU-методе липазную активность измеряют при 30°С и рН 9 с использованием стабилизированной эмульсии оливкового масла (Sigma catalog No 800-1) в качестве субстрата в 5 мМ Трис-буфере, содержащем 40 мМ NaCl и 5 мМ хлорида кальция. 2,5 мл субстрата смешивают с 12,5 мл буфера, рН доводят до 9, добавляют 0,5 мл разведенного образца липазы и получают определенное количество олеиновой кислоты с последующим титрованием с использованием рН-стата.
Одна SLU равна количеству липазы, которое высвобождает 1 моль титруемой олеиновой кислоты в минуту в стандартных условиях.
Фосфолипазная активность
Для качественного или количественного определения фосфолипазной активности были использованы нижеследующие аналитические методы.
Фосфолипазная активность (PHLU)
Фосфолипазную активность (PHLU) измеряли как степень высвобождения свободных жирных кислот из лецитина. В 50 мкл 4% L-альфа-фосфатидилхолина (растительного лецитина от Avanti), 4% Тритона Х-100, 5 мМ CaCl2 в 50 мМ HEPES, рН 7 добавляют 50 мкл раствора фермента, разведенного до соответствующей концентрации в 50 мМ HEPES, рН 7. Образцы инкубируют в течение 10 минут при 30°С и реакцию завершают при 95°С в течение 5 минут, а затем центрифугируют (5 минут при 7000 об/мин). Уровень свободных жирных кислот определяют с использованием набора NEFA С от Wako Chemicals GmbH; к 25 мкл реакционной смеси добавляют 250 мкл реагента А и инкубируют 10 минут при 37°С. Затем добавляют 500 мкл реагента В и образец снова инкубируют 10 минут при 37°С. Абсорбцию при 550 нм измеряют с использованием спектрофотометра на диодной матрице HP 8452A. Образцы измеряют, по крайней мере, в дубликатах. При этом используют смеси субстрата и фермента (предварительно обработанные образцы фермента (10 мин при 95°С) + субстрат). В качестве стандартной жирной кислоты используют олеиновую кислоту. 1 PHLU равна количеству фермента, способного высвобождать 1 мкмоль свободной жирной кислоты/мин в этих условиях.
Фосфолипазная активность (LEU)
Лецитин гидролизуют при постоянном рН и температуре, и фосфолипазную активность определяют как степень потребления титранта (0,1н NaOH) в процессе нейтрализации высвобожденной жирной кислоты.
Субстратом является лецитин сои (L-α-фосфотидил-холин), а реакцию проводят в условиях: рН 8,00, 40,0°С, реакционное время 2 минуты. Единицу активности определяют по отношению к стандарту.
Анализ с использованием монослоя фосфолипазы
На тщательно очищенную поверхность буферного раствора (либо 10 мМ глицина, рН 9,0, либо 10 мМ NaOAc, рН 5,0; 1 мМ CaCl2, 25°C) наносят монослой дидеканоилфосфатидилхолина (DDPC) из раствора хлороформа. После релаксации монослоя (выпаривания хлороформа) поверхностное давление доводят до 15 мН/м, соответствующее средней молекулярной площади DDPC, равной приблизительно 63 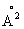 /молек. Раствор, содержащий приблизительно 60 мкг (микрограмм) фермента, впрыскивают через монослой в субфазу реакционной зоны (цилиндр с площадью поверхности 2230 мм2 и реакционным объемом 56570 мм3) в "кормушке нулевого порядка". Ферментативную активность выражают как скорость подвижного барьера, сжимающего монослой для поддержания постоянного поверхностного давления, по мере того как нерастворимые молекулы субстрата гидролизуются с образованием более водорастворимых продуктов реакции. После подтверждения того, что водорастворимость продуктов реакции (каприновой кислоты и MDPC) значительно превышает водорастворимость DDPC, оценивают число DDPC-молекул, гидролизованных данным ферментом в минуту, исходя из средней молекулярной площади (СМП) DDPC. Результаты вычисляют исходя из средней скорости барьера в течение первых 5 минут гидролиза.
/молек. Раствор, содержащий приблизительно 60 мкг (микрограмм) фермента, впрыскивают через монослой в субфазу реакционной зоны (цилиндр с площадью поверхности 2230 мм2 и реакционным объемом 56570 мм3) в "кормушке нулевого порядка". Ферментативную активность выражают как скорость подвижного барьера, сжимающего монослой для поддержания постоянного поверхностного давления, по мере того как нерастворимые молекулы субстрата гидролизуются с образованием более водорастворимых продуктов реакции. После подтверждения того, что водорастворимость продуктов реакции (каприновой кислоты и MDPC) значительно превышает водорастворимость DDPC, оценивают число DDPC-молекул, гидролизованных данным ферментом в минуту, исходя из средней молекулярной площади (СМП) DDPC. Результаты вычисляют исходя из средней скорости барьера в течение первых 5 минут гидролиза.
Результат считается положительным для фосфолипазы, если барьер перемещается со скоростью более, чем 2 мм/мин.
Анализ на чашках 1
A) 50 мл 2% агарозы в очищенной воде расплавляют/перемешивают в течение 5 минут и охлаждают до 60-63°С.
B) 50 мл 2%-ного растительного L-альфа-фосфатидилхолина, 95%, в 0,2М NaOAc, 10 мМ CaCl2, pH 5,5 при 60°С в течение 30 минут смешивают и центрифугируют 15 сек в центрифуге Ultrathorax.
Равные объемы 2% агарозы и 2% лецитина (А и В) смешивают и к этой смеси добавляют равный объем 1% Тритона Х-100.
Затем добавляют 250 мкл 4 мг/мл кристаллического фиолетового в очищенной воде в качестве индикатора. Эту смесь выливают в соответствующие чашки Петри (например, 30 мл в чашку диаметром 14 см) и в данном агаре делают соответствующие отверстия (3-5 мм) для нанесения ферментного раствора.
Образец фермента разводят до концентрации, соответствующей OD280=0,5, и 10 микролитров наносят на отверстия в агарозной/лецитиновой матрице. Чашки инкубируют при 30°С, и реакционные зоны в чашках идентифицируют приблизительно через 4-5 часов и/или приблизительно через 20 часов после инкубирования. Липазу Humicola lanuginosa используют в качестве контроля, а присутствие более обширной зоны осветления, чем у контроля, считают как положительный результат на фосфолипазную активность.
В варианте этого анализа добавление Тритона Х-100 не проводили.
Анализ на чашках 2
10 г агарозы расплавляют в 550 мл Н2О путем кипячения в микроволновой печи. После охлаждения до 60-70°С добавляют следующие ингредиенты:
250 мл 0,4М цитратного буфера (рН 4,5 или рН 7,1);
200 мл 3% лецитина (от Avanti) в 2% Тритона Х-100;
2 мл 2% кристаллического фиолетового;
30 мл смеси выливают в чашки Петри диаметром 14 см.
После нанесения образцов фермента эти чашки инкубируют и результаты обрабатывают так же, как и результаты анализа на чашках 1.
ДГДГазная активность, направленная на гидролиз дигалактозилдиглицерида
Анализ на монослое 1
На тщательно очищенную поверхность буферного раствора (10 мМ NaOAc, pH 5,5; 1 мМ CaCl2, 25°С, 10 мМ бета-циклодекстрина (Sigma С-4767)) наносят монослой ДГДГ (Sigma (D4651)) из раствора хлороформа. После релаксации монослоя (выпаривания хлороформа) поверхностное давление доводят до 15 мН/м. Раствор, содержащий приблизительно 60 мкг (микрограмм) фермента, впрыскивают через монослой в субфазу реакционной зоны (цилиндр с площадью поверхности 2230 мм2 и реакционный объем 56570 мм3) в "кормушке нулевого порядка". Ферментативная активность проявляется увеличением скорости подвижного барьера, сжимающего монослой для поддержания постоянного поверхностного давления по мере того, как нерастворимые молекулы субстрата гидролизуются с образованием более водорастворимых продуктов реакции (в присутствии бета-циклодекстрина).
Результат считается положительным на ДГДГазную активность, если барьер перемещается со скоростью более чем 1 мм/мин.
Монослой 2
На тщательно очищенную поверхность буферного раствора (прибл. 75 мл, 10 мМ NaOAc, pH 5,5; 1 мМ CaCl2, 25°C, 10 мМ бета-циклодекстрина (Sigma С-4767)) наносят монослой ДГДГ (Sigma (D4651)) из раствора хлороформа до поверхностного давления приблизительно 30 мН/м. После релаксации монослоя (выпаривания хлороформа) раствор, содержащий приблизительно 30 мкг (микрограмм) очищенного фермента, впрыскивают через монослой в 75 мл субфазы и при этом непрерывно измеряют поверхностное давление. Ферментативная активность проявляется увеличением скорости снижения поверхностного давления по мере того, как ДГДГ гидролизуется с образованием водорастворимых продуктов реакции (в присутствии бета-циклодекстрина).
Результат считается положительным на ДГДГазную активность, если максимальное падение поверхностного давления (dπ/dt) после добавления фермента превышает -0,5 мН/мин. После тестирования ряда вариантов липолазы было обнаружено, что они обладают ДГДГазной активностью, а исходный фермент (липолаза) обладал лишь очень ограниченной активностью (dπ/dt > -0,5 мН/мин).
Дрожжевой штамм
Штамм Saccharomyces cerevisiae YNG318: МАТа leu2-D2 ura3-52 his4-539 pep4-Dl[cir+] описан в WO 97/04079 и WO 97/07205.
Трансформация дрожжевого штамма
ДНК-фрагменты и разомкнутые векторы смешивают и трансформируют в дрожжевой штамм Saccharomyces cerevisiae YNG318 стандартными методами.
Вектор для трансформации дрожжей
pJS0026 (экспрессирующая плазмида S.cerevisiae) описана в WO 97/07205 и в работе J.S.Okkels, (1996) "A URA3-promoter deletion in a pYES vector increases the expression level of a fungal lipase in Saccharomyces cerevisiae. Recombinant DNA Biotechnology III: The Integration of Biological and Engineering Sciences", vol. 782 of the Annals of the New York Academy of Sciences). Эту плазмиду получали от pYES 2,0 путем замены индуцибельного GAL-1-промотора pYES 2,0 промотором конститутивно экспрессируемого гена TPI (триозофосфат-изомеразы) от Saccharomyces cerevisiae (Albert & Karwasaki, (1982), J.Mol.Appl. Genet., 1, 419-434) и делеции части промотора URA3.
Сайт-направленный мутагенез
Для конструирования вариантов липолитического фермента Н.lanuginosa может быть использован коммерческий набор Chameleon в целях проведения двухцепочечного сайт-направленного мутагенеза в соответствии с инструкциями производителей.
Ген, кодирующий нужный липолитический фермент, встраивают в плазмиду pHD414. В соответствии с инструкцией производителей Scal-сайт гена ампициллина pHD414 заменяют на Mlul-сайт с использованием следующего праймера:
Праймер 3: AGAAATCGGGTATCCTTTCAG.
Вектор pHD414, содержащий нужный ген липолитического фермента, затем используют в качестве матрицы для ДНК-полимеразы и олигонуклеотидов 7258 и 7770.
7258: 5'р gaa tga ctt ggt tga cgc gtc acc agt cac 3'
(Такая замена Scal-сайта присутствует в гене резистентности к ампициллину и используется для разрезания на Mlul-сайт).
Праймер № 7770 был использован в качестве селективного праймера.
7770: 5'р tct agc cca gaa tac tgg atc aaa tc 3' (замены Scal-сайта присутствуют в гене липазы Н.lanuginosa и не приводят к изменению аминокислотной последовательности).
Нужную мутацию (например, в N-конце гена липолитического фермента или введение цистеинового остатка) вводят в нужный ген липолитического фермента путем добавления соответствующих олигонуклеотидов, содержащих нужную мутацию.
ПЦР-реакции осуществляют в соответствии с рекомендациями производителей.
Метод скрининга
Дрожжевые библиотеки рассеивают на целлюлозные фильтры на чашках с агаром SC-ura и инкубируют в течение 3-4 дней при 30°С.
Затем эти фильтры переносят на чашки с лецитином и инкубируют при 37°С в течение 2-6 часов. Дрожжевые клетки, содержащие активные фосфолипазы, образуют белые зоны клиренса вокруг колоний. Затем положительные варианты могут быть дополнительно очищены и протестированы.
Среды
Среда SC-ura
Дрожжевой азот (без аминокислот) 7,5 г
Янтарная кислота 11,3 г
NaOH 6,8 г
Казаминокислота (без витамина) 5,6 г
Триптофан 0,1 г
Агар, Merck 20 г
Дистиллированная вода Доб. 1000 мл
Автоклавируют в течение 20 минут при 121°С. Из стерильного маточного раствора добавляют 4 мл 5% треонина до объема 900 мл вместе со 100 мл стерильной 20% глюкозы.
Примеры
Пример 1: Конструирование вариантов с остовом от липазы Humicola lanuginosa и С-концом от фосфолипазы Fusarium oxysporum с помощью ПЦР-реакции
Для остова от липазы Humicola lanuginosa в качестве матриц используют нижеследующие варианты: Е1А+G91A+D96W+Е99К+Q249R и SPIRR+G91A+D96W+Е99К+Q249R. Исходную липазу используют для генерирования фрагмента в С-конце без Q249R. В качестве матрицы для С-концевой фосфолипазы используют фосфолипазу Fusarium oxysporum, клонированную в том же самом векторе, в котором клонировали варианты липазы Humicola lanuginosa.
ПЦР-реакция 1: 4244 (SEQ ID NO:1) в качестве 5'-праймера и Н7 (SEQ ID NO: 6) в качестве 3'-праймера и одна из двух матриц, упомянутых выше.
ПЦР-реакция 2: FOL14 (SEQ ID NO:3) в качестве 5'-праймера и FOL15 (SEQ ID NO:4) в качестве 3'-праймера и липаза Humicola lanuginosa в качестве матрицы (мутация в положении 249 отсутствует).
ПЦР-реакция 3: FOL16 (SEQ ID NO:5) в качестве 5'-праймера и АР (SEQ ID NO:2) в качестве 3'-праймера, и фосфолипаза F. oxysporum в качестве матрицы.
ПЦР-реакцию 4 проводят для присоединения варианта липазы Humicola lanuginosa к С-концу от фосфолипазы с использованием FOL14 (SEQ ID NO:3) в качестве 5'-праймера и АР (SEQ ID NO:2) в качестве 3'-праймера, и продуктов ПЦР-реакций 2 и 3 в качестве матрицы.
Конечную ПЦР осуществляют с использованием 4244 (SEQ ID NO:1) в качестве 5'-праймера и KBoj14 (SEQ ID NO:7) в качестве 3'-праймера, и продуктов ПЦР-реакций 1 и 4 в качестве матрицы (при использовании липазы Humicola lanuginosa в качестве матрицы в реакции 2 имелась возможность исключить мутацию, которая была создана в положении 249).
Конечный ПЦР-фрагмент был использован в in vivo рекомбинации в дрожжах вместе с pJSО026, разрезанной рестриктирующими ферментами SmaI (или BamHI) и XbaI (для удаления кодирующей области и одновременно для создания перекрывания приблизительно в 75 нуклеотидов в каждом конце в целях стимуляции возможных событий рекомбинации). Эта конечная обработка также использовалась в последующих примерах.
Праймер FOL14 (SEQ ID NO:3) и праймер 15/16 представляют собой смешанные олигонуклеотиды, способствующие связыванию липазы Humicola lanuginosa и фосфолипазных матриц и в то же самое время способствующие введению аминокислот из обеих матриц в различные положения. В некоторые положения могут быть также введены новые аминокислоты.
Праймер FOL14 (SEQ ID NO:3)
Положение 205 в липазе Н.lanuginosa: 75% R, 25% S
Праймер FOL15 (SEQ ID NO:4)/FOL16 (SEQ ID NO:5)
Положение 256 в липазе Н.lanuginosa: 50% Р, 50% А
Положение 260 в липазе Н.lanuginosa: 25% R, 12,5% Q, 12,5% Н, 12,5% С, 12,5% Y, 12,5% W, 12,5% stop (стоп-кодон).
Были определены последовательности полученных вариантов и было обнаружено, что они соответствуют последовательностям липазы Humicola lanuginosa с нижеследующими модификациями. Модификации в скобках являются неточными
Е1А, G91A, D96W, Е99К, Р256А, W260H, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А, 271G, 272G, 273F, (274S)
Е1А, G91A, D96W, Е99К, Е239С, Q249R, Р256А, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А, 271G, 272G, 273F, (274S)
Е1А, G91A, D96W, Е99К, N248T, Q249R, W260Q, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А, 271G, 272G, 273F, (274S)
SPIRR, G91A, D96W, E99K, W260C, G263Q, L264A, I265T, G266D, T267A, L269N, 270A, 271G, 272, G273F, (274S)
SPIRR, G91A, D96W, E99K, G263Q, L264A, I265T, G266D, T267A, L269N, 270A, 271G, 272G, 273F, (274S)
E1A, G91A, D96W, E99K, G263Q, L264A, I265T, G266D, T267A, L269N, 270A, 271G, 272G, 273F, (274S)
Пример 2: Продуцирование усеченных последовательностей
Данные варианты были сделаны со стоп-кодоном после аминокислоты 269, 270, 271, 272 (273 и 274).
Нижеследующие реакции ПЦР были проведены с использованием следующей матрицы: E1A, G91A, D96W, E99K, Р256А, W260H, G263Q, L264A, I265T, G266D, T267A, L269N, 270A, 271G, 272G, 273F, (247S).
Реакция 1: 5'-праймер 4244 (SEQ ID NO:1) и 3'-праймер KBoj36 (стоп-кодон после 269).
Реакция 2: 5'-праймер 4244 (SEQ ID NO:1) и 3'-праймер KBoj37 (стоп-кодон после 270).
Реакция 3: 5'-праймер 4244 (SEQ ID NO:1) и 3'-праймер KBoj38 (стоп-кодон после 271).
Реакция 4: 5'-праймер 4244 (SEQ ID NO:1) и 3'-праймер KBoj39 (стоп-кодон после 272).
Были определены последовательности полученных вариантов и было обнаружено, что они соответствуют липазе Humicola lanuginosa с нижеследующими модификациями.
Е1А, G91A, D96W, Е99К, Р256А, W260H, G263Q, L264A, I265T, G266D, Т267А, L269N
Е1А, G91A, D96W, Е99К, Р256А, W260H, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А
Е1А, G91A, D96W, Е99К, Р256А, W260H, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А, 271G
Е1А, G91A, D96W, Е99К, Р256А, W260H, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А, 271G, 272G
Пример 3: Удаление мутаций в области "крышки"
G91A или Е99К могут быть удалены без потери фосфолипазной активности. Были определены последовательности полученных вариантов и было обнаружено, что они соответствуют липазе Humicola lanuginosa с нижеследующими модификациями.
Е1А, G91A, D96W, Р256А, W260H, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А, 271G, 272G, 273F, (274S)
SPIRR, D96W, Е99К, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А, 271G, 272G, 273F, (274S)
SPIRR, G91A, D96W, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А, 271G, 272G, 273F, (274S)
Е1А, G91A, D96W, Р256А, W260H, G263Q, L264A, I265T, G266D, Т267А, L269N, 270А, 271G, 272G, 273F, (274S)
Пример 4: "Допинг"-вставка в С-концевую область липазы Humicola lanuyinosa для сообщения фосфолипазной активности
Три различные библиотеки конструируют так, чтобы можно было внести мутации в положении 256 и положениях 263-269. При этом предусматривались возможности для удлинения С-конца 1, 2, 3 или 4 аминокислотами.
Допинг-вставки в последовательность дикого типа подчеркнуты:
256:  , А 3, Т 3
, А 3, Т 3
263:  , Е 4.8, А 3.8. R 3.6, Q 0.2, Р 0.2
, Е 4.8, А 3.8. R 3.6, Q 0.2, Р 0.2
264:  , Р 4.8. Q 3.8, V 3.6, А 0.2, Е 0.2
, Р 4.8. Q 3.8, V 3.6, А 0.2, Е 0.2
265:  , Т 5.6, L 2.2, S 1.6, N 1.5, F 1.4, R 0.4, К 0.4 А,Р 0.1, G,D,C,H,Y 0.03, Q,E 0.01, стоп-кодон 0.016
, Т 5.6, L 2.2, S 1.6, N 1.5, F 1.4, R 0.4, К 0.4 А,Р 0.1, G,D,C,H,Y 0.03, Q,E 0.01, стоп-кодон 0.016
266:  ,G 86. D 5.9, R 2, S 1.7, С 1.6, А 0.9, V 0.9, Е 0.7, W 0.2, H,Y 0.1, I,L,T,F,P 0.02, Q,K 0.01, стоп-кодон 0.014
,G 86. D 5.9, R 2, S 1.7, С 1.6, А 0.9, V 0.9, Е 0.7, W 0.2, H,Y 0.1, I,L,T,F,P 0.02, Q,K 0.01, стоп-кодон 0.014
267:  , А 6.6, S 1.9, R 0.9, N 0.9, I 0.9, К 0.9, М 0.9, Р 0.9, Р 0.9, G,V 0.14, D,E 0.07, L 0.03, C,Q,H,F,W,Y 0.01, стоп-кодон 0.01
, А 6.6, S 1.9, R 0.9, N 0.9, I 0.9, К 0.9, М 0.9, Р 0.9, Р 0.9, G,V 0.14, D,E 0.07, L 0.03, C,Q,H,F,W,Y 0.01, стоп-кодон 0.01
268:  , S 1.9, R 1.0, G 1.0, F 0.9, Y 0.9. L 0.04, A,N,D,H,I,P,T,V 0.01, стоп-кодон 2.8
, S 1.9, R 1.0, G 1.0, F 0.9, Y 0.9. L 0.04, A,N,D,H,I,P,T,V 0.01, стоп-кодон 2.8
269:  , стоп-кодон 8 (KBoj 32 (SEQ ID NO: 8) и KBoj33)/ N 86, К 2.7, D 1.8, Н 1.8, I 1.8. S 1.8, Т 1.9, Y 1.8, R 0.1, Q,M,E 0.06, A,C,G,L.F,P,V 0.04, стоп-кодон 0.06(KBoJ34)
, стоп-кодон 8 (KBoj 32 (SEQ ID NO: 8) и KBoj33)/ N 86, К 2.7, D 1.8, Н 1.8, I 1.8. S 1.8, Т 1.9, Y 1.8, R 0.1, Q,M,E 0.06, A,C,G,L.F,P,V 0.04, стоп-кодон 0.06(KBoJ34)
270: 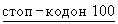 (KBoJ33)/A 44, Р 44, S 1.9, Т 1.8. R 1.5, L 1.5, G 1.4, V 1.4, D 0.7, Q 0.7, Е 0.7, Н 0.7, N,C,I,K,M,F,W,Y 0.03, стоп-кодон 0.03(КВоj 32 (SEQ ID NO: 8) и KBoj 34)
(KBoJ33)/A 44, Р 44, S 1.9, Т 1.8. R 1.5, L 1.5, G 1.4, V 1.4, D 0.7, Q 0.7, Е 0.7, Н 0.7, N,C,I,K,M,F,W,Y 0.03, стоп-кодон 0.03(КВоj 32 (SEQ ID NO: 8) и KBoj 34)
271: G 72, R 4.5, V 3.2, Е 3.0, С 2.9. А 1.6, S 1.2, D 1.0. L 0.5. I.K.Y 0.15. Q.T 0.08, N,P 0.05, стоп-кодон 9.2
272: G 72, R 4.5, V 3.2, Е 3.0, С 2.9, А 1.6, S 1.2, D 1.0, L 0.5, I,K,Y 0.15, Q,T 0.08, N,P 0.05, стоп-кодон 9.2
273: F 74, L 11, S 2.8, I 2.7, V 2.7, Y 2.5, С 2.5, A,R,T 0.1, N,D,H 0.08, Q,E,K 0.01, стоп-кодон 0.5
274 стоп-кодон
Библиотека А: ПЦР-реакция с использованием 4244 (SEQ ID NО:l) в качестве 5'-праймера и KBoj33 в качестве 3'-праймера и Е1А+G91A+D96W+Е99К+Q249R или Е1А+G225R в качестве матрицы. Варианты этой библиотеки не имеют удлинения.
Библиотека В: ПЦР-реакция с использованием 4244 (SEQ ID NO:1) в качестве 5'-праймера и KBoj32 (SEQ ID NO: 8) в качестве 3'-праймера и Е1А+G91A+D96W+Е99К+Q249R или Е1А +G225R в качестве матрицы. Варианты этой библиотеки будут, по всей вероятности, содержать С-концевое удлинение, но они могут содержать стоп-кодоны перед удлинением.
Библиотека С: ПЦР-реакция с использованием 4244 (SEQ ID NO:1) в качестве 5'-праймера и KBoj34 в качестве 3'-праймера и Е1А+G91A+D96W+Е99К+Q249R или Е1А+G225R в качестве матрицы. Варианты этой библиотеки будут, по всей вероятности, содержать мутации в положении 269 и С-концевое удлинение, но они могут содержать стоп-кодоны перед удлинением.
Были получены нижеследующие варианты:
Библиотека А:
Е1А+G91A+D96W+Е99К+Q249R+G266D
Библиотека В:
E1A+G91A+D96W+E99K+(R232L)+Q249R+G266D+270A
E1A+G91A+D96W+E99K+Q249R+G266S+270D+271G
Е1A+G91A+D96W+E99K+Q249R+L264G+I265G+G266F+T267стоп-кодон
E1A+G91A+D96W+E99K+Q249R+G266A+270P+271G
E1A+G91A+D96W+E99K+Q249R+L264P+I265F+L269стоп-кодон
Библиотека С
Е1А+G91A+D96W+Е99К+Q249R+G263E+G266D+L269N+270Р+271V+272G+273F
Е1А+G91A+D96W+Е99К+Q249R+G263A+G266S+L269N+270А+271G+272R+273F
Е1А+G91A+D96W+Е99К+Q249R+L264P-G266+L269I+270Р+271R+272G+273F
Е1А+G91A+D96W+Е99К+Q249R+G266D+L269S+270А+271G+272G+273F
Е1А+D27G+G91A+D96W+Е99К+Q249R+G266S+L269N+270А+271G+272G+273F
Е1А+G91A+D96W+Е99К+Q249R+G266D+L269N+270А
Е1А+G91A+D96W+Е99К+Q249R+L264P+L267Q+L269N
Е1А+G91A+D96W+Е99К+Q249R+G263R+1265L+L269N+270Р
Пример 5: Для некоторых из вышеуказанных вариантов оптимальный рН для липазы и фосфолипазы определяли с использованием LU- и PHLU-методов при различных значениях рН. Результаты показали, что рН, оптимальный для фосфолипазной активности, составляет в пределах 4-6. рН, оптимальный для липазной активности, варьируется в пределах от около 6 до около 10.
8 вариантов, перечисленных в примере 5, были проанализированы на фосфолипазную активность с помощью вышеописанного анализа с использованием монослоя при рН 5 и 9. Результаты показали, что все варианты обладают фосфолипазной активностью при рН 5 и 9, тогда как исходная липаза (липаза Humicola lanuginosa) не обнаруживала какой-либо активности при рН 5 или 9. В зависимости от конкретного варианта активность при рН 5 была выше или ниже, чем активность при рН 9.
Было установлено, что вариант липазы Humicola lanuginosa, описанный в прототипе, SPIRR+N94K+F95L+D96H+N101S+F181L+D234Y+I252L+Р256Т+G263A+L264Q, не обнаруживал какой-либо активности при рН 5.
Пример 5: Варианты липазы Humicola с фосфолипазной активностью
Данные варианты были получены из исходной липазы Humicola lanuginosa и протестированы на фосфолипазную активность как описано выше. Тем же самым методом было обнаружено, что нижеследующие варианты обладали фосфолипазной активностью, в тех случаях, когда исходная липаза не обладала фосфолипазной активностью.





В вышеуказанной таблице (+274S) означает, что присутствие этого аминокислотного остатка в С-конце не является достоверным. Для одного из таких вариантов было установлено, что только малая фракция содержала этот остаток.
Несколько из вышеуказанных вариантов имели более высокое отношение фосфолипазы (PHLU) к липазе (LU), чем фермент прототипа от Fusarium oxysporum, который, как известно, обладает как липазной, так и фосфолипазной активностью.
Для некоторых из вышеуказанных вариантов рН, оптимальный для липазной и фосфолипазной активности, определяли с использованием LU- и PHLU-методов при различных значениях рН. Результаты показали, что рН, оптимальный для фосфолипазной активности, составляет в пределах 4-6. рН, оптимальный для липазной активности, варьируется в пределах от около 6 до около 10.
8 вариантов, перечисленных в примере 5, были проанализированы на фосфолипазную активность с помощью вышеописанного анализа с использованием монослоя при рН 5 и 9. Результаты показали, что все варианты обладают фосфолипазной активностью при рН 5 и 9, тогда как исходная липаза (липаза Humicola lanuginosa) не обнаруживала какой-либо активности при рН 5 или 9. В зависимости от конкретного варианта активность при рН 5 была выше или ниже, чем активность при рН 9.
Было установлено, что вариант липазы Humicola lanuginosa, описанный в прототипе, SPIRR+N94K+F95L+D96H+N101S+F181L+D234Y+1252L+Р256Т+G263A+L264Q, не обнаруживал какой-либо активности при рН 5.
Нижеследующие варианты, полученные от исходной липазы Humicola lanuginosa, могут также обладать фосфолипазной активностью:

Пример 6: Варианты липазы Rhizomucor с фосфолипазной активностью
Данные два варианта были получены из исходной липазы Rhizomucor miehei и протестированы на фосфолипазную активность, как описано выше. Тем же самым методом было обнаружено, что варианты обладали фосфолипазной активностью в тех случаях, когда исходная липаза не обладала фосфолипазной активностью.
G266N
G266V.
Пример 7: Варианты липазы Humicola с повышенной специфичностью по отношению к длинноцепочечным жирным кислотам
Данные варианты были получены из исходной липазы Humicola lanuginosa и протестированы на гидролитическую активность по отношению к двум триглицеридным субстратам с разной длиной цепи: трибутирину (С4:0) и триолеину (C18:1). Эти тесты проводили при рН 9 LU- и SLU-методами, описанными выше. Было обнаружено, что нижеследующие варианты обладают более высоким отношением активности, направленной на триолеин, к активности, направленной на трибутирин, чем исходный фермент (липаза Humicola lanuginosa):


Нижеследующие варианты исходной липазы Humicola lanuginosa могут также обладать повышенной специфичностью по отношению к длинноцепочечным жирным кислотам:

Пример 8: Варианты липазы Fusarium с повышенной специфичностью по отношению к длинноцепочечным жирным кислотам
Данные варианты были получены из исходной липазы Fusarium oxysporum и протестированы, как описано в предыдущем примере. Было обнаружено, что нижеследующие варианты обладают более высоким отношением активности, направленной на триолеин, к активности, направленной на трибутирин, чем исходный фермент:
Y23S
Y260L.
Нижеследующие варианты, полученные от липазы Fusarium oxysporum, могут также обладать повышенной специфичностью по отношению к длинноцепочечным жирным кислотам:
R80H+S82T
S82T+А129Т.
Пример 9: Варианты липазы Rhizomucor с повышенной специфичностью по отношению к длинноцепочечным жирным кислотам
Нижеследующие варианты, полученные от липазы Rhizomucor miehei, могут также обладать повышенной специфичностью по отношению к длинноцепочечным жирным кислотам:
Y260W
Y28L
Y28C+H217N.
Пример 10: Варианты липазы Humicola с повышенной специфичностью по отношению к короткоцепочечным жирным кислотам
Данные варианты были получены из исходной липазы Humicola lanuginosa и протестированы, как описано в предыдущем примере. Было обнаружено, что нижеследующие варианты обладают более высоким отношением активности, направленной на трибутирин, к активности, направленной на триолеин (более низкое отношение SLU/LU), чем исходный фермент:

Нижеследующие варианты исходной липазы от Humicola lanuginosa могут также иметь более высокое отношение активности, направленной на трибутирин, к активности, направленной на триолеин:

Пример 11: Варианты липазы Fusarium с повышенной специфичностью по отношению к короткоцепочечным жирным кислотам
Данные варианты были получены из исходной липазы Fusarium oxysporum и протестированы, как описано в предыдущем примере. Было обнаружено, что нижеследующие варианты обладают более высоким отношением активности, направленной на трибутирин, к активности, направленной на триолеин, чем исходный фермент:
Y23W
Y260D
Y260R
Y260C
Y260N
Пример 12: Варианты липазы Rhizomucor с повышенной специфичностью по отношению к короткоцепочечным жирным кислотам
Нижеследующие варианты исходной липазы Rhizomucor miehei могут также обладать повышенной специфичностью по отношению к короткоцепочечным жирным кислотам:
Y260C
Y260G
Y260V.
Пример 13: Варианты липазы Humicola с ДГДГазной активностью
Данные варианты были получены из исходной липазы Humicola lanuginosa и была определена гидролитическая активность по отношению к ДГДГ (дигалактозилдиглицерид)? как описано выше. Было обнаружено, что нижеследующие варианты обладают ДГДГазной активностью, тогда как исходная липаза дала отрицательный результат.

Пример 14: Варианты липазы Humicola с повышенным оптимальным рН
Данные варианты были получены из исходной липазы Humicola lanuginosa и протестированы на липазную активность, измеренную LU-методом при рН 7 и 9. Было обнаружено, что нижеследующие варианты обладают более высоким отношением активности при рН 9 к активности при рН 7, чем исходный фермент:
R84L
R84W
Y21I
Y21V
Y261I
Пример 15: Варианты липазы Humicola с пониженным оптимальным рН
Данные варианты были получены из исходной липазы Humicola lanuginosa и протестированы на липазную активность, измеренную LU-методом при рН 7 и 9. Было обнаружено, что нижеследующие варианты обладают более низким отношением активности при рН 9 к активности при рН 7, чем исходная липаза:
Y261D
G266D/E
Y261W.
Пример 16: Использование вариантов липазы Humicola при рафинировании растительного масла
Масло семян рапса обрабатывали двумя вариантами липазы от Humicola lanuginosa, в основном, как описано в примере 6 в WO 98/18912 (Novo Nordisk).
Один вариант был протестирован при ферментной дозе 0,6 мг ферментного белка на один килограмм масла. Результаты тестов при различных рН и температурах показали оптимальные характеристики при рН 5, 7, 35-45°С, где конечное содержание Р достигало 4 м.д. Отдельный эксперимент, проведенный при 45°С и рН 6, показал, что конечное содержание Р, составляющее 4 м.д., может быть достигнуто при дозе фермента, по крайней мере, 0,15 мг/кг.
Аналогичный эксперимент с другим вариантом липазы Humicola lanuginosa показал оптимальные характеристики при 40°С и рН 5,0-5,5. Доза фермента составляла 0,3 мг/кг.
Эксперимент по рафинированию масла был осуществлен с третьим вариантом липазы Humicola lanuginosa с использованием рапсового масла при 45°С, рН 5 и дозы 1,8 мг фермента/кг масла. Для сравнения был проведен аналогичный эксперимент с исходной липазой (липазой Humicola lanuginosa) при дозе 18 мг/кг. Результаты показали, что хорошее рафинирование (остаточное содержание Р<10 м.д.) с использованием этого варианта было достигнуто за 3,4 часа.
Было обнаружено, что исходная липаза (липаза Humicola lanuginosa) даже при 10-кратном увеличении дозы фермента давала очень низкий эффект рафинирования.
Пример 17: Использование вариантов липазы при выпечке хлеба
Вариант липазы от Humicola lanuginosa оценивали при проведении пробной выпечки хлеба следующим образом
Тесто приготавливали из муки Meneba в соответствии с Европейским безопарным способом приготовления теста (ABF-SP-1201.01) с использованием 40 м.д. аскорбиновой кислоты. Были использованы различные комбинации добавок при следующих дозах: вариант липазы при 0, 0,25, 0,5 или 1,5 мг/кг; фосфолипид (лецитин) при 0 или 10 г/кг; эндоамилаза при 0 или 750 MANU/кг.
Эндоамилаза представляет собой мальтозообразующую амилазу от В.stearothermophilus (торговый знак Novamyl®). Одну MANU (единицу мальтозообразующей амилазы, Maltogenic Amylase Novo Unit) определяли как количество фермента, необходимого для высвобождения одного мкмоль мальтозы/мин при концентрации 10 мг мальтотриозного субстрата на один мл 0,1М цитратного буфера, рН 5,0 при 37°С в течение 30 минут.
После выпечки хлеба батоны охлаждали и объем батона, плотность мякиша и мягкость хлеба оценивали приблизительно через 2 часа. Эту оценку повторяли на 1, 3 и 7 дни после хранения хлеба при 22°С, завернутого в двойные пластиковые пакеты.
Плотность мякиша измеряли с использованием анализатора текстуры ТА-ХТ2 от Stable Micro Systems (диаметр образца 40 мм).
Мягкость в граммах измеряли как силу, необходимую для впрессовывания образца 6,25 мм в мякиш нарезанных ломтиков хлеба толщиной 25 мм (проникновения 25%).
Результаты показали, что добавление 1,5 мг данного варианта приводило к увеличению объема батона. Результаты анализа на плотность и эластичность показали, что данный вариант давал значительно более мягкий мякиш и значительно более высокую эластичность, начиная со дня 0 и до дня 7.
Пример 18: Использование вариантов липазы для стабилизации теста при выпечке хлеба
Вариант липазы от Humicola lanuginosa тестировали при проведении пробной выпечки хлеба для оценки его толерантности в отношении удлинения времени расстойки теста.
Тесто приготавливали из муки Pelican в соответствии с Европейским безопарным способом приготовления теста (347-SP-1217) с использованием 30 м.д. аскорбиновой кислоты, грибковой α-амилазы (10 FAU of Fungamyl) и пентосаназы (100 FXU of Pentopan Mono). Дозы 0,2, 0,4 и 0,6 мг фермента белка/кг муки для данного варианта сравнивали с 1000 LU исходной липазы.
Тесто приготавливали в виде рулетов. Половину рулетов оставляли на расстойку на 45 минут (нормальная расстойка), а другую половину оставляли на 70 минут (чрезмерно продолжительная расстойка).
После выпечки хлеб охлаждали и примерно через 2 часа оценивали объем и внешний вид рулетов. Внешний вид рулетов представляет собой критерий формы рулетов и определяется как высота в 10 рулетов, деленная на ширину в 10 рулетов, что означает, что хорошие круглые батоны имеют высокое значение оценки внешнего вида, тогда как плоские батоны имеют низкое значение оценки внешнего вида.
Результаты показали, что при нормальном времени расстойки объем 0,4 и 0,6 мг данного варианта давал лучшие результаты, чем тот же объем исходной липазы, и вид батонов с использованием этого варианта при всех дозах был лучше, чем вид батона для исходной липазы. При чрезмерно длительной расстойке рулетов объем и форма для этого варианта при всех дозах были лучше, чем объем и форма батона для исходной липазы.
Пример 19: Действие вариантов липазы на возникновение постороннего запаха
Развитие постороннего запаха в результате действия липаз с различной специфичностью по отношению к длине цепи жирных кислот оценивали в цельном молоке. Оценку на появление запаха масляной кислоты/кислого запаха проводили путем обнюханивая образцов после нагревания.
25 мл цельного молока помещали в 100 мл закрывающиеся синие колбы (с крышками) в водяную баню при 32°С. В эти колбы добавляли каждую из нижеперечисленных липаз в количестве 0,2 мг ферментного белка на один литр молока. Температуру повышали до 45°С и через 15 и 105 минут проводили оценку.
Тестируемыми липазами были липаза Humicola lanuginosa и ее варианты. Для каждой липазы специфичность в отношении длины цепи выражали как отношение активностей, направленных на триолеин (SLU) и трибутирин (LU).
Образцы оценивались тремя специалистами, которые руководствовались следующей шкалой оценок запаха:
+ Выявляемый запах,
++ Явный и характерный запах масляной кислоты и/или кислый запах,
+++ Сильный запах масляной кислоты и/или кислый запах.
Было обнаружено, что три варианта липазы Humicola lanuginosa, имеющих более высокое отношение SLU/LU, обладали меньшей способностью вызывать неприятный запах, чем исходная липаза.
Пример 20: Действие вариантов липазы на возникновение неприятного запаха у текстильных изделий после стирки
Загрязнение
Хлопчатобумажную ткань загрязняли молочным продуктом, как описано ниже. 50 мг сливочного масла наносили на площадь приблизительно 30 см2 в виде однородных пятен. Загрязненную ткань оставляли на 24 часа в условиях окружающей среды.
Процедура стирки
Стирку загрязненного текстильного материала осуществляли на приборе Terg-O-tometer с использованием коммерческого моющего средства (5 г/л), содержащего и несодержащего липазу (1250 и 5000 LU/I). Промывку осуществляли при 30°С в течение 20 минут при 100 м.д. После стирки образцы ткани оставляли на ночь для сушки в условиях окружающей среды.
Сенсорный анализ
На следующий день группой специалистов, состоящей, по крайней мере, из 10 опытных экспертов, был проведен анализ на неприятный запах. Образцы помещали в плотно закрытые стеклянные сосуды и между каждой оценкой оставляли, по крайней мере, на 30 минут для накопления неприятного запаха. Затем образцы ткани вынимали и оценивали на неприятный запах.
Неприятный запах масляной кислоты оценивали по нижепривиденной шкале. В качестве эталона использовали образец, выстиранный без применения липазы.
0. Более слабый запах по сравнению с эталоном.
1. Такой же запах, как и запах эталона.
2. Более сильный запах по сравнению с эталоном.
3. Отчетливо более сильный запах по сравнению с эталоном.
4. Более сильный запах, чем запах 3.
Было обнаружено, что варианты липазы Humicola lanuginosa с повышенным отношением активностей, направленных на триолеин/трибутирин (повышенное отношение SLU/LU), дают более слабый запах при расщеплении масляных пятен, чем исходный фермент (липаза Humicola lanuginosa). Отдельно проведенный эксперимент по стирке показал, что данные варианты так же, как и исходный фермент, оказались эффективными в удалении жирных пятен.
Альтернативные методы
Интенсивность запаха масляной кислоты от молочных пятен на ткани может быть оценена с помощью инструментальных анализов:
1. Парафазной газовой хроматографией или
2. Путем извлечения запахов из ткани с помощью газовой хроматографии.
Пример 21: Действие липазных вариантов на запах испеченного хлеба с маслом
Получали шесть вариантов липазы Humicola lanuginosa и оценивали их действие в хлебе, выпеченном в соответствии с Европейским безопарным способом приготовления теста (347-SP-1217) с добавлением 3% масла. Для каждого варианта использовали 0,2 мг ферментного белка/кг муки.
Также определяли специфичность этих вариантов в отношении длины цепи субстрата путем измерения отношения активности, направленной на триолеин/трибутирин (SLU/LU, как описано выше). Для сравнения также тестировали исходную липазу Humicola lanuginosa и липазу прототипа, происходящую от Fusarium oxysporum и обладающую фосфолипазной активностью.
Результаты систематизированы ниже:
+ Выявленный запах,
++ Явный и характерный запах масляной кислоты и/или кислый запах,
+++ Сильный запах масляной кислоты и/или кислый запах.
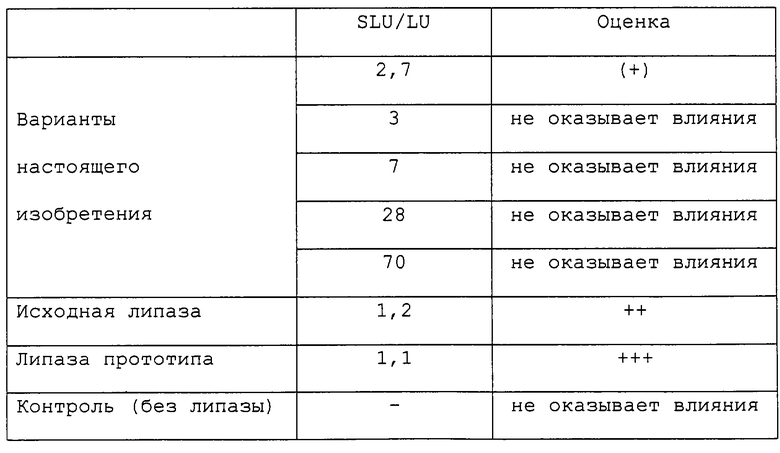
Результаты показали, что варианты липазы с отношением SLU/LU, равным 3 или выше (то есть обладающие высокой специфичностью по отношению к длинноцепочечным жирным кислотам), не продуцируют неприятного запаха при выпекании хлеба, даже если в рецептуре предусмотрено использование масла.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА В ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ | 2005 |
|
RU2406759C2 |
| ЛИПОЛИТИЧЕСКИЙ ФЕРМЕНТ ДЛЯ ПРИМЕНЕНИЯ В ХЛЕБОПЕЧЕНИИ | 2018 |
|
RU2763469C2 |
| ПОЛУЧЕНИЕ ЛИЗОГЛИКОЛИПИДА И БИОКОНВЕРСИЯ ГЛИКОЛИПИДОВ С ИСПОЛЬЗОВАНИЕМ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА | 2005 |
|
RU2548805C2 |
| ВАРИАНТЫ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА | 2007 |
|
RU2474611C2 |
| ЛИПОЛИТИЧЕСКИЕ ФЕРМЕНТЫ, ИХ ПРИМЕНЕНИЕ ПРИ ИСПОЛЬЗОВАНИИ ПИЩЕВЫХ ПРОДУКТОВ | 2010 |
|
RU2538144C2 |
| Композиции, содержащие липазы, и способы обработки поверхности | 2013 |
|
RU2612215C2 |
| БЕЛОК | 2005 |
|
RU2380414C2 |
| СПОСОБЫ УСИЛЕНИЯ ДЕГРАДАЦИИ ИЛИ ПРЕВРАЩЕНИЯ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА | 2006 |
|
RU2441912C2 |
| КЛОНИРОВАНИЕ, ЭКСПРЕССИЯ И ПРИМЕНЕНИЕ КИСЛЫХ ФОСФОЛИПАЗ | 2010 |
|
RU2567659C2 |
| ВАРИАНТЫ ЛИПАЗЫ И КОМПОЗИЦИИ В ВИДЕ МИКРОКАПСУЛ, СОДЕРЖАЩИЕ ТАКИЕ ВАРИАНТЫ ЛИПАЗЫ | 2018 |
|
RU2779301C2 |
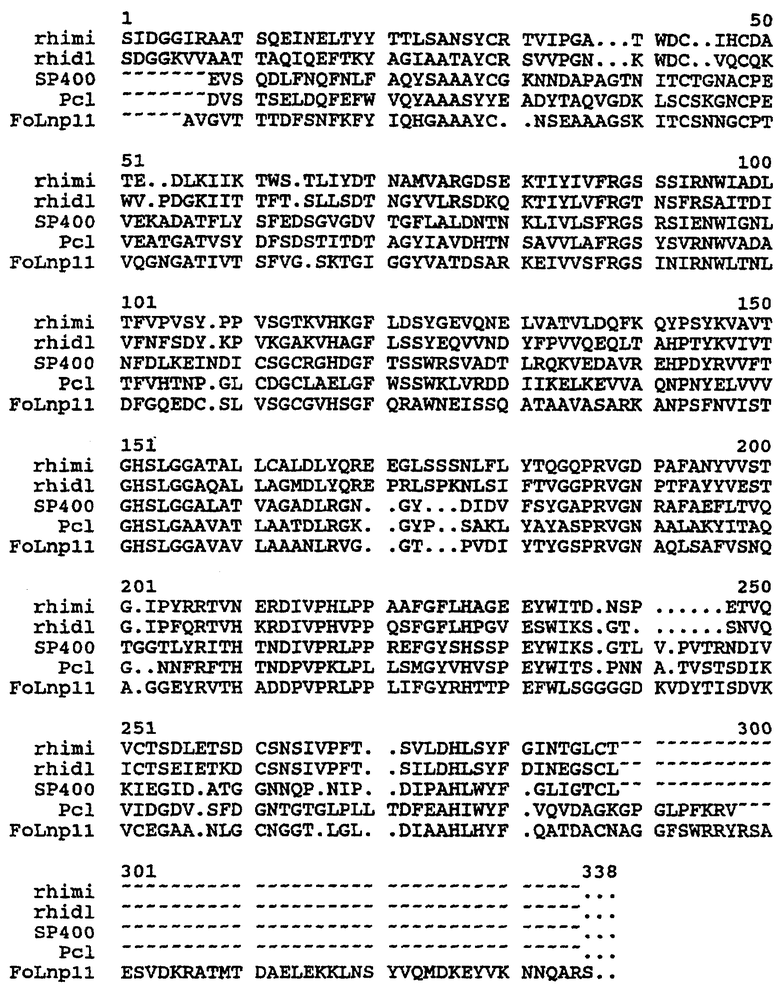









Изобретение относится к биотехнологии и представляет собой способ получения варианта фермента путем создания модификаций в аминокислотной последовательности в определенной области липолитического фермента. Данное изобретение включает также варианты липолитического фермента с модифицированной аминокислотной последовательностью и с модифицированной специфичностью к субстрату. Данное изобретение позволяет увеличить уровень желательной активности или снизить уровень нежелательной активности липолитического фермента. 3 с. и 41 з.п. ф-лы,1 ил., 8 табл.
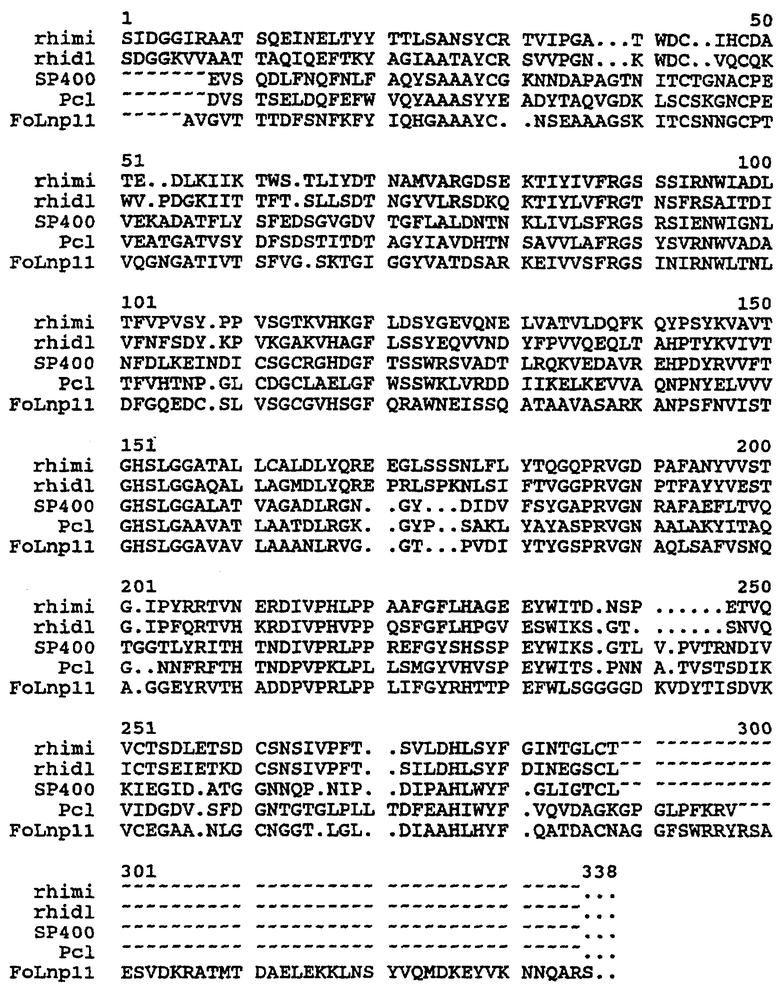
b) необязательно включает одну или более другую аминокислотную модификацию в положениях 21, 23, 26, 56, 57, 60, 61, 62, 83, 259 и 261, или пептидное удлинение в С-конце, содержащее 1-5 аминокислотных остатков; и c) имеет модифицированную специфичность к субстрату, которая представляет собой более высокую фосфолипазную активность.
(ii) замену или делецию аминокислоты, соответствующей С268 или L269 в липазе штамма DSM 4109 Humicola lanuginosa; (iii) замену, соответствующую V60G, D62E, L93K, L97Q, K98E,F, E99D, Р256А, G263E,Q,R,F,N, L264A,С,P,F,G,V,I, I265L,N,F, или T267A,Q,P,S,V,E в липазе штамма DSM 4109 Humicola lanuginosa: iv) вставку, соответствующую T267GS или T267GL в липазе штамма DSM 4109 Humicola lanuginosa; v) пептидное удлинение у С-конца, которое представляет собой А, P, MD, СР, AG, DG, PG, AGG, PVGF, AGRF, PRGF, AGGF или AGGFS; vi) пептидное удлинение у С-конца из 40-50 аминокислот или vii) усечение в 1, 2, 3, 4, 5 или 6 аминокислот у С-конца; b) необязательно включает одну или более другую аминокислотную модификацию в положениях 22, 56-59, 61, 62, 64, 83, 85, 91, 94, 96, 98, 99, 249, 255, 256, 259, 263 и 265, или пептидное удлинение в С-конце, содержащее 1-5 аминокислотных остатков, или пептидное удлинение у N-конца, содержащее 1-15 аминокислотных остатков; и c) имеет специфичность к субстрату, отличающуюся от специфичности известного липолитического фермента.
| WO 9205249 A1, 02.04.1992.WO 9401541 A1, 20.01.1994.RU 93042492 A , 10.01.1997. |
