Настоящая заявка связана по существу излагаемого материала с материалом, содержащимся в Международной Публикации WO 90/07572. Содержание указанной предыдущей заявки специально приводится по этой причине в настоящем изобретении в качестве ссылки.
Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к результатам, полученным при исследовании дезоксирибонуклеазы (ДНКазы)-фосфодиэстеразы, способной проводить гидролиз полидезоксирибонуклеиновой кислоты. Оно относится в основном к разделению нескольких форм упомянутой ДНКазы, к таким ее формам, на основе которых могут быть составлены полезные лекарственные композиции, а также к способам использования ДНКаз и созданных на их основе композиций.
Предпосылки создания изобретения
ДНКаза представляет собой фосфодиэстеразу, способную гидролизовать полидезоксирибонуклеиновую кислоту. ДНКаза была выделена из различных источников с различной степенью чистоты. Впервые полная аминокислотная последовательность для ДНКазы из млекопитающих была определена в 1973 г. См., Liao, et al., J. Biol. Chem.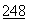 :1489 (1973).ДНКаза характеризуется множеством уже известных полезных свойств и применялась в связи с этим для лечебных целей. Основное терапевтическое применение ДНКазы связывалось со способностью снижать вязкоупругость легочных выделений при таких заболеваниях, как пневмония и муковисцидоз, что способствовало очищению дыхательных путей. См. Lourenco, et al., Arch. Intern. Med.
:1489 (1973).ДНКаза характеризуется множеством уже известных полезных свойств и применялась в связи с этим для лечебных целей. Основное терапевтическое применение ДНКазы связывалось со способностью снижать вязкоупругость легочных выделений при таких заболеваниях, как пневмония и муковисцидоз, что способствовало очищению дыхательных путей. См. Lourenco, et al., Arch. Intern. Med. 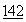 :2299 (1982); Shak, et al., Proc. Nat. Acad. Sci.
:2299 (1982); Shak, et al., Proc. Nat. Acad. Sci. 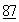 :9188 (1990); Hubbard, et al., New Engl. J. Med.
:9188 (1990); Hubbard, et al., New Engl. J. Med. 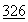 :812 (1992).
:812 (1992).
ДНК, кодирующая ДНКазу человека, была выделена и секвенирована, при этом была достигнута экспрессия ДНК в рекомбинантных клетках, что открывало возможности получения человеческой ДНКазы в коммерчески значимых количествах. См., Shak, et al., Proc. Nat. Acad. Sci. 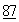 :9188-9192 (1990). Рекомбинантные ДНКазы человека оказались полезными в терапевтическом отношении, особенно при использовании в очищенном виде, когда ее освобождали от протеаз и других белков, обычно связанных с нею в естественном состояние. См., Hubbard, et al., New Engl. J. Med.
:9188-9192 (1990). Рекомбинантные ДНКазы человека оказались полезными в терапевтическом отношении, особенно при использовании в очищенном виде, когда ее освобождали от протеаз и других белков, обычно связанных с нею в естественном состояние. См., Hubbard, et al., New Engl. J. Med. 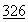 :812 (1992).
:812 (1992).
В цитированных выше заявках на патент описаны способы и методики получения человеческой ДНКазы в фармацевтически приемлемой форме. Существующий уровень техники располагает различными специфическими методами очистки ДНКазы. См., Khouw, et al., U.S. Patent No.4065355 (опубликован 27 декабря 1977); Markey, FEBS Letters 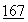 :155 (1984); Nefsky, et al., Eur. J. Biochem.
:155 (1984); Nefsky, et al., Eur. J. Biochem. 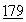 :215 (1989).
:215 (1989).
Хотя это не было известно на момент подачи вышеуказанной заявки на выдачу патента, как стало теперь ясно, ДНКазу-содержащий продукт, полученный из культуры рекомбинантных клеток-хозяев, как правило, включает в себя смесь дезамидированной и недезамидированной форм ДНКазы. Существование дезамидированных форм ДНКазы оставалось неизвестным, несмотря на то, что было известно, что в некоторых белках имеет место дезамидирование аспарагиновых и глутаминовых остатков. См. Eipper et al., Ann. Rev. Physiol. 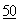 :333 (1988); Kossiakoff, Science
:333 (1988); Kossiakoff, Science 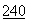 :191 (1988); Bradbury et al., Trends in Biochem, Sci.
:191 (1988); Bradbury et al., Trends in Biochem, Sci. 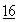 :112 (1991); и Wright, Protein Engineering
:112 (1991); и Wright, Protein Engineering 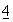 :283 (1991).
:283 (1991).
В настоящем изобретении утверждается на основании неотмечавшегося ранее факта, что рекомбинантная человеческая ДНКаза может существовать в виде смеси дезамидированной и недезамидированной форм. С использованием способов настоящего изобретения было показано, что дезамидированная человеческая ДНКаза отличается меньшей ферментативной активностью, чем недезамидированная человеческая ДНКаза. Таким образом, совместное присутствие в смеси дезамидированной и недезамидированной форм ДНКазы, а также возможность осуществления дальнейшего дезамидирования продукта, которое, как было показано, имеет место при хранении препаратов человеческой ДНКазы in vitro, может усложнить получение устойчивых однородных препаратов ДНКазы, пригодных для клинического применения. Исходя из этого, поскольку само существование, а тем более характеристики дезамидированной ДНКазы до настоящего изобретения оставались неизвестными, в данном изобретении приведены методы идентификации дезамидированности ДНКазы, а также способы отделения таких ДНКаз от препаратов ДНКазы, в которых дезамидированная ДНКаза могла быть найдена, однако ее присутствие неочевидно.
Сущность изобретения
Настоящее изобретение относится к способам разделения дезамидированной и недезамидированной форм человеческой ДНКазы из их смеси. В предпочтительном варианте этот процесс включает хроматографирование смеси с использованием смолы или другой поддерживающей среды, способной к связывание с катионным полимером, таким как гепарин или аналог негидролизованной дезоксирибонуклеиновой кислоты (ДНК), или хроматографирование с использованием так называемой ворсистой катионообменной смолы. Настоящее изобретение относится также к применению таких хроматографических методов, отличных от человеческой ДНКазы, в частности для бычьей ДНКазы.
Настоящее изобретение относится также к дезамидированной человеческой ДНКазе, как к очищенному продукту, по существу не содержащему недезамидированную человеческую ДНКазу.
Настоящее изобретение относится также к недезамидированной человеческой ДНКазе как к очищенному продукту, по существу, не содержащему дезамидированную человеческую ДНКазу. С помощью настоящего изобретения было показано, что очищенная недезамидированная человеческая ДНКаза обладает более полной ферментативной активностью в сравнении с дезамидированной ДНКазой.
Настоящее изобретение относится также к фармацевтическим композициям, содержащим в качестве активного ингредиента либо очищенную дезамидированную человеческую ДНКазу, либо очищенную недезамидированную человеческую ДНКазу, необязательно в сочетании с фармацевтически приемлемым наполнителем.
Настоящее изобретение относится также к способу, предусматривающему введение терапевтически эффективного количества очищенной дезамидированной ДНКазы или очищенной недезамидированной ДНКазы для лечения, например, таких больных, у которых отмечается накопление вязкого ДНК-содержащего материала. Предпочтительным способом введения таких очищенных ДНКаз является ингаляция препарата непосредственно в легкие.
Настоящее изобретение относится в особенности к способу лечения больных заболеваниями легких, такими как хронический бронхит, муковисцидоз или эмфизема, предусматривающему введение терапевтически эффективного количества очищенной недезамидированной ДНКазы, предпочтительно в дыхательные пути.
Настоящее изобретение относится также к фармацевтическим композициям, включающим недезамидированную человеческую ДНКазу, помещенную в пластиковый сосуд, необязательно в сочетании с фармацевтически приемлемым носителем.
Краткое описание фигур
На фигуре 1 представлена аминокислотная (SEQ ID No.1) и нуклеотидная последовательность ДНК (SEQ ID No. 2), соответствующие человеческой ДНКазе I. Нативная сигнальная последовательность подчеркнута, потенциальные инициирующие кодоны обведены кружками, а зрелая последовательность заключена в скобки.
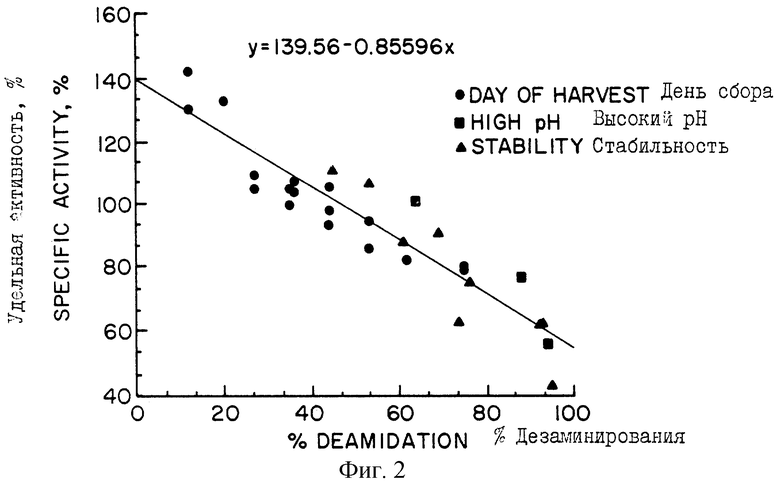
Фигура 2 отражает корреляцию между ферментативной активностью и уровнем дезамидирования образцов человеческой ДНКазы. Удельную активность определяли по нормализации ДНКазной активности от значений, полученных в тесте с метиленовым зеленым (MG) (в единицах концентрации, близких к стандартной кривой) до концентрации ДНКазы, измеряемой на основе иммуносорбентного теста по связыванию фермента (ELISA). Процент дезамидирования определяли по картированию с помощью трипсина. Образцы человеческой ДНКазы, соответствующие отметкам "День сбора клеток" ("Day of Harvest"), выделяли с последующей очисткой из культуры рекомбинантных клеток яичника китайского хомячка (СНО), способных к экспрессии ДНК, кодирующей человеческую ДНКазу I. Такие образцы отбирались на 3, 5, 7, 9, 11, 13 и 20 день от начала роста культуры. Образцы, обозначенные как "Высокий рН" ("Higt рН), представляют собой образцы очищенной ДНКазы 13 дня, которые инкубировали in vitro в течение двух дней при рН 8 и температуре 37°. Образцы, обозначенные как "Стабильность" ("Stability"), представляют собой образцы очищенной ДНКазы 13 дня, которые хранили in vitro при температуре 5°, 25° или 37°С в течение различных периодов времени.
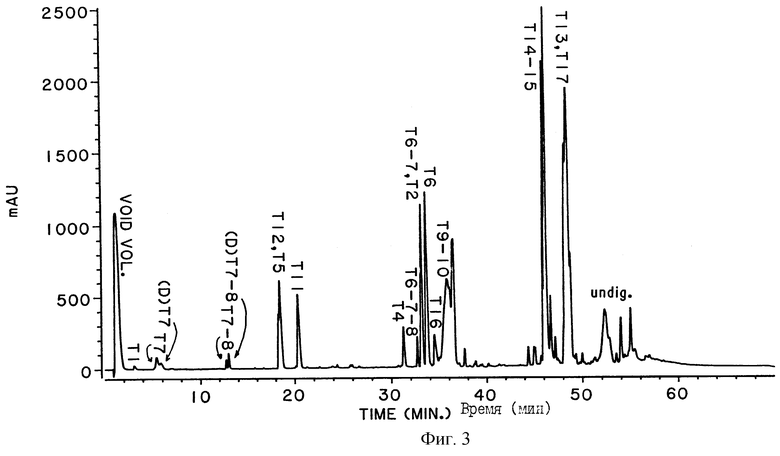
На фигуре 3 представлен пример триптического картирования ДНКазы, выполненного с целью определения уровня дезамидирования. На показанном примере приведена 65% дезамидированная ДНК. Аббревиатура "mAU" указывает миллиединицы поглощения при 214 нМ.
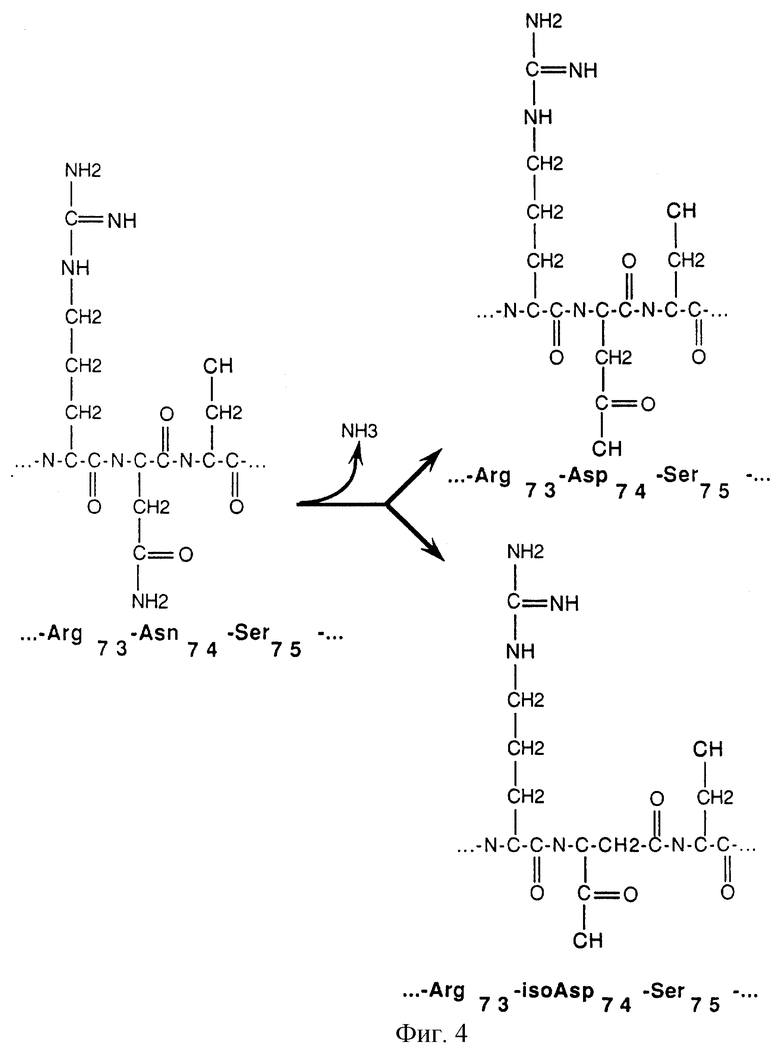
На фигуре 4 схематически представлено дезамидирование аспарагинового остатка в 74 положении аминокислотной цепи (Asn-74) в нативной человеческой ДНКазе. Дезамидирование превращает Asn-74 либо в аспарагиновую кислоту (Asp) либо в остаток изоаспарагиновой кислоты (iso-Asp). Каждая из этих трех форм ДНКаз дает при расщеплении трипсином пару пептидов, что указывает на идентичность указанных форм ДНКаз.
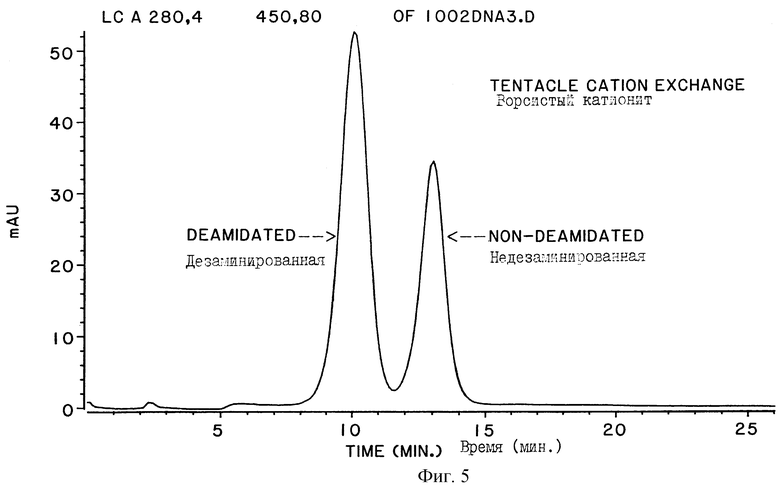
На фигуре 5 представлена хроматограмма образца человеческой ДНКазы, фракционированной на колонке с ворсистым катионообменником (ТСХ). Показанный образец представляет собой 67% дезамидированную ДНКазу.
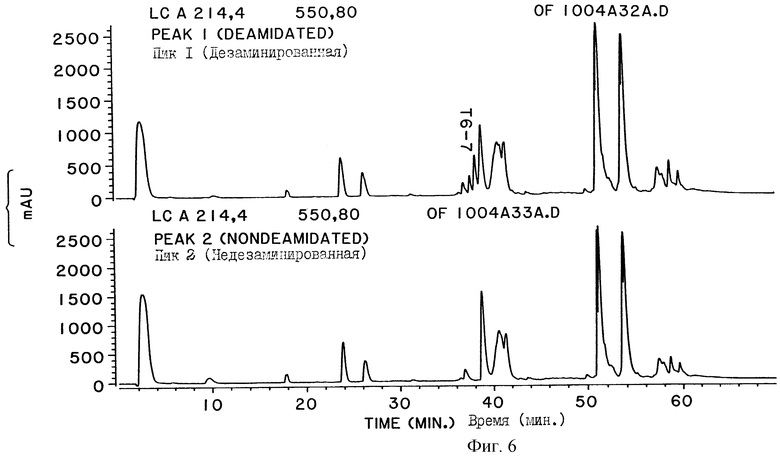
На фигуре 6 показаны карты расщепления трипсином фракций двух пиков, взятых в процессе разделения на ТСХ, показанном на фигуре 5. Отсутствие пептида Т6-7 на триптической карте расщепления пика 2 указывает на отсутствие в нем дезамидированной ДНКазы.
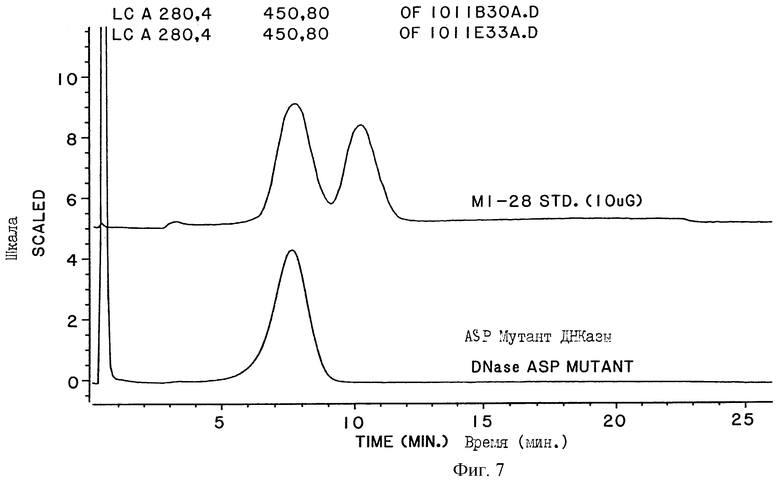
На фигуре 7 показаны хроматограммы нескольких образцов человеческой ДНКазы, фракционированной на ТСХ колонке. Образцы, обозначенные как "MI-28 STD", представляют собой препараты человеческой ДНКазы, полученные из культуры клеток яичника китайского хомячка (СНО), трансформированных с помощью ДНК, кодирующей нативную человеческую ДНКазу I. Образцы, обозначенные как "Asn-Мутант ДНКазы" ("Dnase ASP Mutant"), представляют собой ДНКазу, имевшую в положении 74 аминокислотной цепи остаток аспарагиновой кислоты (а не остаток аспарагина), и которая имеет таким образом ту же аминокислотную последовательность, что и Asp-форма дезамидированной ДНКазы, показанной на фигуре 4. Asp-Мутант ДНКазы был получен из культуры клеток, трансформированных ДНК, кодирующей мутантную форму человеческой ДНКазы. ДНК, кодирующая Asp-мутант ДНКазы, была получена с помощью сайт-направленного мутагенеза ДНК, кодирующей нативную человеческую ДНК. Сравнение хроматограмм показывает, что одна из форм человеческой ДНКазы представляет собой MI-28 STD, которая элюируется с ТСХ колонки в той же позиции, что и Asp-мутант ДНКазы.
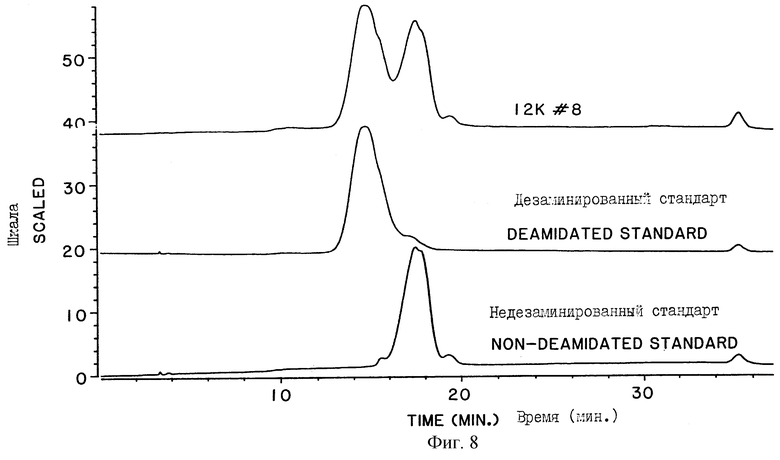
На фигуре 8 представлены хроматограммы нескольких образцов человеческой ДНКазы, фракционированных на TSK-гепариновой колонке (Toso Haas, Montgomeryville, Pennsylvania). Образец, обозначенный как "12К #8", представляет собой препарат человеческой ДНКазы, полученной из культуры клеток яичника китайского хомячка (СНО), трансформированной ДНК, кодирующей нативную человеческую ДНКазу I. Образец, обозначенный как "Дезамидированный Стандарт" ("Deamidated Standard"), представляет собой очищенную дезамидированную человеческую ДНКазу. Образец, обозначенный как "Недезамидированный Стандарт" ("Non-deamidated Standard"), относится к очищенной недезамидированной человеческой ДНКазе. Очищенную дезамидированную человеческую ДНКазу и очищенную недезамидированную человеческую ДНКазу получают с помощью ТСХ хроматографии.
На фигуре 9 представлены хроматограммы нескольких образцов человеческой ДНКазы, фракционированных на колонке с иммобилизованным аналогом ДНК. Образец, обозначенный как "MI-28", представляет собой человеческую ДНКазу, полученную из культуры клеток яичника китайского хомячка (СНО), трансформированной ДНК, кодирующей нативную человеческую ДНКазу. Образец, обозначенный как "Дезамидированный Стандарт" ("Deamidated Standard"), представляет собой очищенную дезамидированную человеческую ДНКазу. Образец, обозначенный как "Недезамидированный Стандарт" ("Non-deamidated Standard"), относится к очищенной недезамидированной человеческой ДНКазе. Очищенные дезамидированные человеческие ДНКазы и очищенные недезамидированные человеческие ДНКазы получают с помощью ТСХ хроматографии. Образец, обозначенный как "Asp-мутант ДНКазы" ("Dnase ASP Mutant"), представляет собой ДНКазу, несущую остаток аспарагиновой кислоты (а не остаток аспарагина) в позиции 74 аминокислотной последовательности.
Подробное описание
А. Определения
Под термином "человеческая ДНКаза" в настоящем описании подразумевается полипептид, имеющий такую же аминокислотную последовательность, как и в приведенной выше на фигуре 1 последовательности аминокислот человеческой зрелой ДНКазы I, то же самое относится к аминокислотной последовательности его вариантов (включая аллельные варианты), обладающих ферментативной гидролизующей активностью в отношении ДНК. Таким образом, в данном контексте термин "человеческая ДНКаза" обозначает широкий набор материалов, раскрытие или получение которых приведено в обозначенных выше заявках на патент.
Термин "человеческая ДНКаза" обязательно включает в себя нативную зрелую человеческую ДНКазу, имеющую остаток аспарагина (Asn) в 74 положении аминокислотной последовательности полипептида. Как показано в настоящем изобретении, этот остаток аспарагина является чувствительным к дезамидированию, которое может привести к получению смеси дезамидированной и недезамидированной форм человеческой ДНКазы. Вместо остатка (Asn) в положении 74 аминокислотной последовательности, дезамидированная ДНКаза содержит остаток аспарагиновой кислоты (Asp) или изо-аспарагиновой кислоты (iso-Asp) (См. фигуру 4).
Используемый в данном изобретении термин "дезамидированная человеческая ДНКаза" означает человеческую ДНКазу, дезамидированную по остатку аспарагина, при этом указанное дезамидирование имеет место в 74 положении аминокислотной последовательности нативной зрелой человеческой ДНКазы. Было показано, что дезамидированная ДНКаза может возникать при получении человеческой ДНКазы с применением рекомбинантных методов и может быть обнаружена в препаратах человеческой ДНКазы, полученных из рекомбинантных хозяйских клеток.
Кроме того, дезамидированная человеческая ДНКаза может возникать при хранении in vitro недезамидированной человеческой ДНКазы.
Несмотря на то, что остаток аспарагина в положении 7 аминокислотной последовательности, нативной зрелой человеческой ДНКазы тоже может быть дезамидирован (в дополнение к остатку аспарагина в 74 положении аминокислотной цепи), показано, что такое двойное дезамидирование приводит к получению ферментативно неактивной ДНКазы.
Используемый здесь термин "смесь" применительно к препаратам человеческой ДНКазы означает присутствие как дезамидированной, так и недезамидированной форм ДНКаз. Было показано, например, что в препаратах человеческой ДНКазы, полученных при экспрессии в рекомбинантных клетках, дезамидированию подвергается от 50 до 80%, а, возможно, и более высокий процент ДНКазы.
Термин "очищенная дезамидированная человеческая ДНКаза", используемый в настоящем изобретении, обозначает дезамидированную человеческую ДНКазу, которая по существу не содержит примеси недезамидированной человеческой ДНКазы. Иными словами, недезамидированная человеческая ДНКаза составляет примерно менее 10%, предпочтительно менее примерно 5%, а в наиболее предпочтительном варианте менее чем примерно 1% от общей массы ДНКазы в препарате очищенной дезамидированной человеческой ДНКазы.
Термин "очищенная недезамидированная человеческая ДНКаза", используемый в настоящем контексте, обозначает недезамидированную человеческую ДНКазу, которая по существу не содержит примесей дезамидированной человеческой ДНКазы. Иными словами, дезамидированная человеческая ДНКаза составляет примерно менее 25%, предпочтительно менее, чем около 5%, а в наиболее предпочтительном варианте менее примерно 1% от общей массы ДНКазы в препарате очищенной недезамидированной человеческой ДНКазы.
Применяемый в настоящем описании термин "наполнитель" означает фармацевтически приемлемый материал, используемый совместно с ДНКазой для приготовления подходящего лекарственного средства с целью улучшения действия его при введении ДНКазы пациентам. Подходящие для таких целей наполнители хорошо известны и описаны, например, в Physicians Desk Reference, the Merck Index и Remington's Phrmaceutical Sciences.
Предпочтительной фармацевтической композицией для человеческой ДНКазы является забуференный или незабуференный водный раствор, предпочтительно в изотоническом растворе соли, таком, как 150 мМ хлорида натрия, содержащем 1,0 мМ хлорида кальция при рН 7. Такие растворы особенно подходят для использования при коммерчески значимом производстве распылителей, включающих струйные распылители и ультразвуковые распылители, особенно применимые, в частности, при распылении непосредственно в дыхательные пути или легкие больного. Подробное описание, касающееся способов приготовления фармацевтических композиций, а также их введения для получения эффективного результата, можно найти в цитированных выше патентных заявках.
Под применяемым здесь термином "терапевтически эффективное количество" подразумеваются дозировки от 1 мкг до примерно 100 мг человеческой ДНКазы на 1 кг веса тела пациента, и вводимой в составе описываемых здесь фармацевтических композиций. Терапевтически эффективное количество человеческой ДНКазы зависит, например, от терапевтических целей, способа введения и от состояния больного. В этой связи лечащему врачу необходимо проводить титрование нужной дозировки и варьировать способ введения препарата для получения оптимального терапевтического эффекта. В связи с описанными выше различиями в ферментативной активности дезамидированной и недезамидированной форм ДНКаз может случиться так, что количество очищенной недезамидированной ДНКазы, требуемой для достижения терапевтического эффекта, будет меньше, чем количество очищенной дезамидированной человеческой ДНКазы или смеси этих двух форм, необходимых для достижения такого же эффекта при тех же условиях.
Очищенные ДНКазы, особенно недезамидированную ее форму, применяют для изменения вязкоупругости слизистых выделений. Такие очищенные препараты человеческой ДНКазы особенно применимы для лечения больных легочныии заболеваниями, для которых характерно наличие аномально вязких гнойных выделений, а также для лечения состояний, таких как острые или хронические бронхолегочные заболевания, включая инфекционную пневмонию, бронхит или трахеобронхит, бронхоэктаз, муковисцидоз, астму, туберкулез и микозы. Для такого лечения применяется закапывание в бронхи раствора хорошо размельченного сухого препарата или очищенной дезамидированной человеческой ДНКазы или же очищенной недезамидированной человеческой ДНКазы с применением традиционных способов, таких как аэрозольное распыление.
В. Предпочтительные варианты
В результате успешного клонирования и достижения экспрессии человеческой ДНКазы в рекомбинантных клетках-хозяевах было показано в ходе тщательного анализа, что получаемый при такой экспрессии в рекомбинантных клетках ДНКазный продукт представляет собой в типичном случае смесь к тому времени еще не определенных компонентов. В частности, исследование человеческой ДНКазы, выделенной из культуры клеток яичника китайского хомячка (СНО) и далее очищенных, с применением метода изоэлектрического фокусирования (ИЭФ), выявило сложный набор разных видов ДНКазы. Определяемые таким образом различные виды ДНКазы представляют собой результат нескольких пост-трансляционных модификаций ДНКазы, включая в том числе и дезамидирование.
Для определения наличия и уровня дезамидированных ДНКаз в таких препаратах применяли два метода анализа. Один из них включал расщепление исходных препаратов ДНКазы с помощью трипсина и дальнейший анализ полученных пептидов с помощью ВЭЖХ с обращением фаз. В этом методе количество дезамидированной ДНКазы в исходном препарате определяли по количеству шести характерных пептидов, полученных при расщеплении трипсином.
Другой метод включал хроматографирование исходного препарата ДНКазы на ионообменной колонке с ворсистым катионитом (ТСХ). Было показано, что применение ТСХ колонки дает возможность разрешения пиков дезамидированной человеческой ДНКазы и недезамидированной человеческой ДНКазы, так что каждая из имеющихся форм ДНКазы может быть эффективно отделена от другой и получена в чистом виде. В этом методе количества дезамидированной и недезамидированной ДНКаз в исходном препарате определяли за счет измерения зон хроматографических пиков, соответствующих отдельным формам ДНКаз.
Несмотря на то, что оба приведенных метода одинаково эффективны в обнаружении и количественном определение дезамидированной ДНКазы, ТСХ методу отдается предпочтение в связи с тем, что он является менее трудоемким, отнимающим меньше времени, т.е. в этом аспекте более эффективным, чем второй метод. Кроме того, метод хроматографии на ТСХ обеспечивает разделение дезамидированной и недезамидированной форм ДНК даже в тех случаях, когда традиционно применяемые катионообменные смолы, а также другого рода смолы, применяемые в хроматографической технике, не давали нужного разделения.
Основные принципы хроматографии на ТСХ были описаны, в частности, у Miller, J. Chromatography 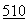 :133 (1990); Janzen et al., J. Chromatography
:133 (1990); Janzen et al., J. Chromatography  :77 (1990); и Hearn et al., J. Chromatography
:77 (1990); и Hearn et al., J. Chromatography 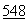 :117 (1991). Не желая ограничить изобретение предложением единственного механизма действия или теоретической основы функционирования, авторы полагают по аналогии с известной кристаллической структурой бычьей ДНКазы, что остаток Asn-74 в человеческой ДНКазе, чувствительный к дезамидированию, располагается в ферменте в сайте, удобном для связывания с бороздкой в спирали ДНК. Соответствующая ей бороздка в ферменте содержит основные аминокислотные остатки (для связывания с ДНК) и, по-видимому, именно она является чувствительной к лигандам ворсистой катионообменной смолы, но не к намного более коротким лигандам традиционных катионообменных смол. По-видимому, механизм действия основан на том, что лиганды ворсистой катионообменной смолы близки к натуральным субстратам на основе нуклеиновых кислот. В этой связи можно полагать, что ионообменная хроматография на ворсистой смоле будет полезна для очистки также других нуклеаз, таких как рибонуклеаза (РНКаза) или рестрикционные эндонуклеазы, а также белков, связывающихся с ДНК.
:117 (1991). Не желая ограничить изобретение предложением единственного механизма действия или теоретической основы функционирования, авторы полагают по аналогии с известной кристаллической структурой бычьей ДНКазы, что остаток Asn-74 в человеческой ДНКазе, чувствительный к дезамидированию, располагается в ферменте в сайте, удобном для связывания с бороздкой в спирали ДНК. Соответствующая ей бороздка в ферменте содержит основные аминокислотные остатки (для связывания с ДНК) и, по-видимому, именно она является чувствительной к лигандам ворсистой катионообменной смолы, но не к намного более коротким лигандам традиционных катионообменных смол. По-видимому, механизм действия основан на том, что лиганды ворсистой катионообменной смолы близки к натуральным субстратам на основе нуклеиновых кислот. В этой связи можно полагать, что ионообменная хроматография на ворсистой смоле будет полезна для очистки также других нуклеаз, таких как рибонуклеаза (РНКаза) или рестрикционные эндонуклеазы, а также белков, связывающихся с ДНК.
Альтернативно разделение дезамидированной и недезамидированной форм ДНКаз может быть достигнуто посредством хроматографии на основе смолы или другой несущей матрицы, ковалентно связанными с катионными полимерами, такими как гепарин или синтетический негидролизуемый аналог ДНК.
Колонки для хроматографии на основе иммобилизованного гепарина коммерчески доступны (например, от Toso Haas Co., Montgomeryville, Pennsylvania).
Негидролизуемые аналоги ДНК были описаны, в частности, у Spitzer et al., Nuc. Acid. Res. 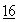 :11691 (1988). Иммобилизованный негидролизуемый аналог ДНК получают обычно синтезом аналога ДНК с аминокислотной группировкой на 3'-конце одной или обеих комплементарных нитей. Аминогруппа затем становится доступной для связывания с эпоксиактивированной колонкой, как это описано в материалах, опубликованных Рэйнин Байокемикел ЭлСи Продактс (Rainin Biochemical LC Products, Woburn, Massachusetts).
:11691 (1988). Иммобилизованный негидролизуемый аналог ДНК получают обычно синтезом аналога ДНК с аминокислотной группировкой на 3'-конце одной или обеих комплементарных нитей. Аминогруппа затем становится доступной для связывания с эпоксиактивированной колонкой, как это описано в материалах, опубликованных Рэйнин Байокемикел ЭлСи Продактс (Rainin Biochemical LC Products, Woburn, Massachusetts).
После успешного разделения дезамидированной и недезамидированной форм ДНКаз по методу данного изобретения было обнаружено с помощью исследования с метиленовым зеленым (MG), что дезамидированная человеческая ДНК имеет сниженную в сравнении с недезамидированной человеческой ДНКазой ферментативную активность. Kurnick, Arch. Biochem. 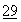 :41 (1950). Было показано, что дезамидированная человеческая ДНКаза обладает половинной ферментативной активностью в сравнении с недезамидированной человеческой ДНКазой. Таким образом, сочетая очищенные дезамидированные ДНКазы и очищенные недезамидированные ДНКазы настоящего изобретения в различных пропорциях, можно создавать фармацевтические композиции на основе человеческой ДНКазы, имеющие нужную удельную активность в широком диапазоне удельных активностей индивидуальных компонентов, с возможностью выбора оптимальных вариантов для лечения определенных заболеваний.
:41 (1950). Было показано, что дезамидированная человеческая ДНКаза обладает половинной ферментативной активностью в сравнении с недезамидированной человеческой ДНКазой. Таким образом, сочетая очищенные дезамидированные ДНКазы и очищенные недезамидированные ДНКазы настоящего изобретения в различных пропорциях, можно создавать фармацевтические композиции на основе человеческой ДНКазы, имеющие нужную удельную активность в широком диапазоне удельных активностей индивидуальных компонентов, с возможностью выбора оптимальных вариантов для лечения определенных заболеваний.
Приведенные ниже примеры даны только для иллюстрации и никоим образом не претендуют на ограничение каким-либо образом настоящего изобретения.
С. Примеры
1. Картирование с помощью трипсина
Процедура, применяемая для триптического картирования человеческой ДНКазы, может быть суммирована следующим образом:
Стадия 1. Образец ДНК весом 1 мг доводится до концентрации 4 мг/мл посредством концентрации на устройстве Амикон Центрикон-10 (Amicon Centricon-10) или путем разбавления носителем. Конечный объем: 250 мкл.
Стадия 2. К образцу добавляют 250 мкл предварительно обработанного буфера (40 мМ БисТрис, 10 мМ ЭГТА, рН 6,0). Инкубацию проводят в течение 1 часа при 37°С.
Стадия 3. Меняют буфер, в котором находится образец, на расщепляющий буфер (100 мМ Трис, рН 8) с помощью колонки Фармация NAP-5. Конечный объем: 1 мл.
Стадия 4. К образцу добавляют 10 мкл трипсина (1 мг/мл трипсина, 1 мМ НСl) и инкубируют в течение 2 часов при 37°С.
Стадия 5. К образцу добавляют вторую аликвоту трипсинового раствора, равную 10 мкл, и инкубируют еще в течение 2 часов при 37°С.
Стадия 6. Останавливают процесс расщепления с помощью добавления 6 мкл трихлоруксусной кислоты (ТХУ). Вплоть до хроматографирования хранят образцы при температуре ниже 5°С.
Стадия 7. Разделяют смесь пептидов с применением метода ВЭЖХ при следующих условиях:
Колонка: Нуклеозил (Nucleosil) C18, 5 мкм, 100 A, 2,0×150 мм (Аллтек, Ко., Дирфилд, Иллинойс - Alltech, Co., Deerfield, Illinois).
Температура колонки: 40°С.
Элюент А: 0,12% ТХУ в воде
Элюент В: 0,10% ТХУ в ацетонитриле.
Градиентный профиль (см. табл. А):
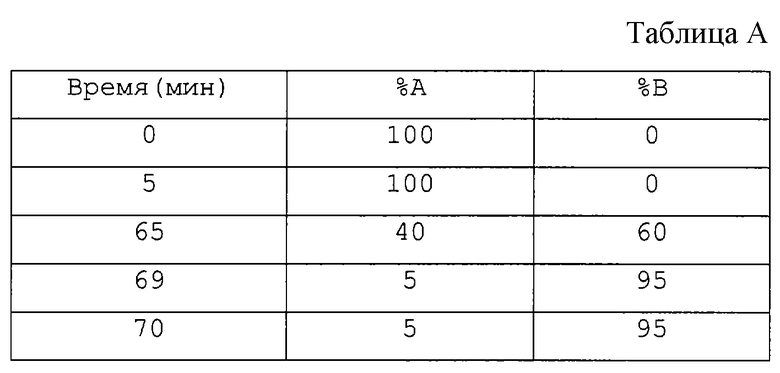
Скорость течения: 0,25 мл/мин. Объем наносимого образца: 250 мкл.
Время уравновешивания колонки после проведения разделения при 100% А: 20 мин.
Температура автоматического отборника проб: 5°.
Обнаружение: на основе поглощения при 214 и 280 нм.
Стадия 8. Определяют Т7, (D)T7, T7-8, (D)T7-8, T6-7-8 и Т6-7 триптических пептидов сравнением времени их удерживания со стандартом.
Стадия 9. Интегрируют пики хроматограммы, полученные при 280 нм. Проверяют качество хроматограммы по положению базовой линии и разделению рядом элюирующихся пиков. Особое внимание при этом стоит уделить рано элюируемым пептидам Т7 и (D)T7, которые могут не очень хорошо разойтись при элюции.
Стадия 10. Соотносят зоны пиков шести указанных выше пептидов с содержанием тирозина. Каждый из пептидов Т7, (D)T7, Т7-8 и (D)T7-8 содержат единственный остаток тирозина, тогда как Т6-7-8 и Т6-7 содержат три остатка тирозина. Вычисляют пропорцию дезамидированных видов на основе нормализованных, т.е. соотнесенных с содержанием тирозина зон пиков (D)T7, (D)T7-8, T6-7-8 и Т6-7, относительно общей суммы нормализованных зон пиков всех шести пептидов.
Для тщательного исполнения метода триптического картирования с целью определения дезамидированной ДНКазы в соответствии с описанной выше процедурой требуется 1 мг ДНКазы в объеме 250 мкл. В этой связи начальная подготовка к проведению этой процедуры включает либо концентрирование либо разбавление образца для получения нужного результата. ДНКаза в высшей степени устойчива к действию протеаз, включая трипсин, в присутствии кальция. По этой причине следующая стадия процедуры заключается в частичном удалении ионов кальция посредством обработки [этилен бис(оксиэтиленнитрило)]тетрауксусной кислотой (ЭГТА). Передозировка при обработке ЭГТА может привести к денатурации и аггрегации ДНКазы, так что эта стадия должна выполняться особенно тщательно. Для ЭГТА-обработанного образца в объеме 0,5 мл заменяют далее среду на расщепляющий буфер (1 мл), добавляют трипсин и инкубируют образец при температуре 37°С в течение 2 ч. Затем добавляют вторую аликвоту трипсина и инкубируют образец еще в течение 2 ч. Расщепление останавливают подкислением смеси, а образец либо оставляется для дальнейшего анализа либо загружается для исследования непосредственно на колонку для ВЗЖХ.
250 мкл (250 мкг) Смеси пептидов, полученных после расщепления трипсином, разделяют на колонке ВЭЖХ с обращением фаз с соблюдением описанных выше условий. Типичная карта триптического расщепления человеческой ДНКазы представлена на фигуре 3. ВЭЖХ проводили на модели Хьюдетт-Паккард (Hewlett Packard) 1090M ВЭЖХ. Элюируемый с колонки материал сразу же исследовался на наличие поглощения при 214 и 280 нм с помощью имеющегося в приборе диодного детектора. Поскольку пептидная карта, получаемая на основе ранней порции элюируемого материала, не дает возможности оценить количественно уровень дезамидированной ДНКазы, ниже описываются другие колонки для получения больших порций материала, элюируемого в более позднее время, а также для использования в исследованиях больших, необходимых для анализа объемов. Каждый анализ в соответствии с такой процедурой требует 70 мин для градиентного разделения и 20 мин для уравновешивания колонки для общего цикла ВЭЖХ, составляющего 90 мин. Теоретические основы и практические подходы для интегрирования пика с целью определения дезамидированной ДНКазы в образце представлены ниже.
Дезамидирование человеческой ДНКазы имеет место, по крайней мере, по аспарагиновому остатку в 74 положении аминокислотной цепи (Asn-74) нативной зрелой человеческой ДНК. Как видно из перечня ожидаемых пептидов при расщеплении трипсином человеческой ДНКазы, приведенного в таблице 1, Asn-74 находится на С-концевом участке сайта расщепления трипсином рядом с аргининовым остатком в 73 позиции аминокислотной цепи (Arg-73).

Вместо Asn (единственная обозначающая буква "N") остатка в положении 74 нативной недезамидированной человеческой ДНКазы, дезамидирование человеческой ДНКазы, может иметь место или по Asp или по iso-Asp остатку, как это показано на фигуре 4. Iso-Asp представляет собой изомерную, бета-аминокислотную форму аспарагиновой кислоты. Пептидная связь между Аrg-73 и iso-Asp устойчива к расщеплению трипсином, так что дезамидированная человеческая ДНКаза дает при расщеплении трипсином характерный пептид, содержащий остатки 51-77 и называемый Т6-7, поскольку представляет собой пептид, объединенный на основе пептидов Т6 и Т7. В условиях, используемых для триптического расщепления, пептидная связь Arg-73-Asn-74 в недезамидированной человеческой ДНКазе и пептидная связь Arg-73-Asn-74 в Asp-форме дезамидированной человеческой ДНКазы расщепляются трипсином. Отсюда на наличие недезамидированной ДНКазы указывает присутствие на триптической карте Т7 пептида, показанного в таблице 1, тогда как на наличие Asp-74 формы человеческой дезамидированной ДНКазы указывает присутствие на триптической карте дезамидированного Т7 пептида, называемого (D)T7. Эти три описанных пептида обозначены на фигуре 3. К сожалению, трипсин лишь частично расщепляет пептидную связь на С-концевом участке Т7, между остатками 77 и 78, так что каждый из описываемых пептидов Т7, (D)T7 и Т6-7 имеют Т7-коньюгат, Т7-8 и Т6-7-8 соответственно. Эти уже шесть репортерных пептидов должны приниматься во внимание для количественного определения дезамидированной человеческой ДНКазы с помощью метода триптического картирования.
В принципе (D)T7, (D)T7-8, T6-7 и Т-6-7-8 пептиды указывают на наличие дезамидированной человеческой ДНКазы, а Т7 и Т7-8 пептиды указывают на наличие недезамидированной человеческой ДНКазы и, зная количественное соотношение этих пептидов, можно прямо вычислить уровень дезамидирования, характерный для данного препарата ДНКазы. Для вычисления количества фракции дезамидированной ДНКазы необходимо располагать знанием молярных коэффициентов дезамидированной и недезамидированной видов ДНКаз. Однако в процедуре триптического картирования возникают две дополнительные проблемы, которые необходимо преодолеть: одна из них - хроматографическая проблема и вторая - проблема обнаружения. Хроматографическая проблема заключается в том, что Т2 пептид элюируется вместе с T6-7 и это затрудняет интеграцию четких зон пиков этого характерного для дезамидированного материала пептида. Указанная проблема может быть преодолена за счет интегрирования хроматограмм, полученных при 280 нм, поскольку все шесть релевантных пептидов имеют как минимум один тирозиновый остаток (Туr), и поэтому достаточно сильно поглощают при 280 нм, тогда как Т2 содержит Туr или триптофановый (Тrр) остаток, и поэтому его поглощение при этой длине волны пренебрежительно мало. Проблема обнаружения заключается в том, что Т6-7 и Т6-7-8 пептиды содержат каждый по три (Туr) остатка, тогда как каждый из оставшихся четырех пептидов содержат только по одному остатку Туr. Таким образом, Т6-содержащие пептиды имеют больший коэффициент полярной абсорбции, чем пептиды, содержащие только Т7, так что простое сравнение зон пиков приведет к завышению значения уровня дезамидированного вида в образце. Эта проблема может быть решена за счет нормализации, т.е. пересчета значений зон пиков шести пептидов с учетом числа Туr остатков в пептиде. Нормализация таким образом значений зон пиков подразумевает, что все тирозиновые остатки в каждом пептиде находятся в равном химическом окружении, которое, как можно предположить, достаточно хорошо подходит для относительно малых пептидов, в частности таких, какие рассматриваются здесь. При проведении нормализации скорректированные таким образом зоны пиков дезамидированных и недеэамидированных пептидов можно сравнивать для оценки содержания дезамидированной ДНКазы в образце.
2. Ионообменная хроматография на ворсистом катионите
Ворсистая катионитная смола (ТСХ) для ионообменной хроматографии в отличие от традиционных смол-катионитов имеет полиионные лиганды, связывающиеся с кремневой поверхностью. Используемая в приводимом примере колонка с лигандами ЛиКросфер® 1000 SО3 (LiChrospher®, EM Separations, Gibbstown, New Jersey) содержит, как представлено в рекламе, от 25 до 50 сульфопропильных групп вдоль полиэтиленового стержня, которые присоединяются со стороны кремневой поверхности.
ТСХ хроматограмма образца рекомбинантной человеческой ДНКазы проводится на ЛиКросфере® 1000 SО3 колонке, как показано на фигуре 5. Рекомбинантную человеческую ДНКазу выделяли с последующим очищением из культуры клеток яичника китайского хомячка (СНО), трансформированных ДНК, кодирующей человеческую ДНКазу, Shak, et al., Proc. Nat. Acad. Sci.  :9188-9192 (1990); Shak, et al., International Patent Application Publication No.WO 90/07572 (опубликована 12 июля 1990).
:9188-9192 (1990); Shak, et al., International Patent Application Publication No.WO 90/07572 (опубликована 12 июля 1990).
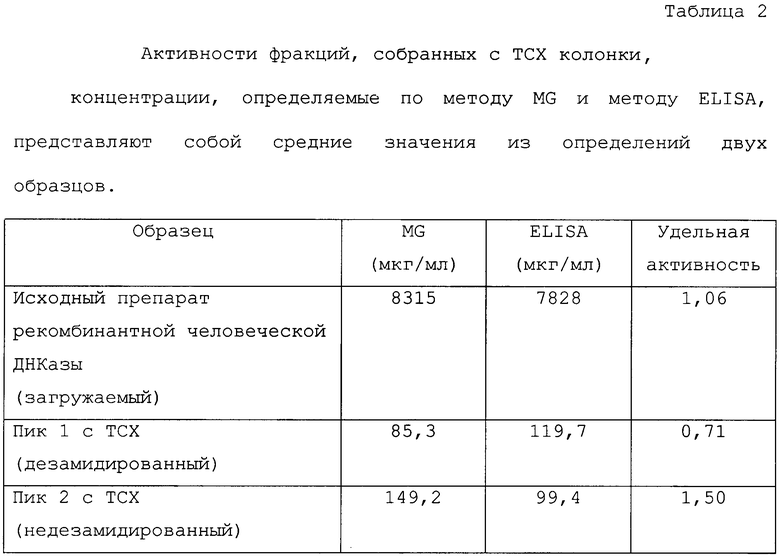
Оба полученных пика собирают и подвергают нескольким анализам для идентификации форм ДНКаз, отличающихся только остатком в 74 позиции аминокислотной последовательности. На фигуре 6 показаны карты триптического расщепления двух пиков, выделенных с помощью ТСХ колонки, которые подтверждают, что они являются соответственно дезамидированной и недезамидированной формами человеческой ДНКазы. Карта триптического расщепления также показывает, что обе формы дезамидированной ДНКазы (имеющие Asp и iso-Asp в позиции 74 аминокислотной последовательности) появляются уже в первом пике при разделении на ТСХ. В таблице 2 показано, что измеряемая для этих двух пиков удельная активность, подтверждающая связь между дезамидированием и удельной активностью, выведена на основе показанной на фигуре 2 корреляции, служащая, кроме того, для дополнительной идентификации ТСХ фракций. Активность фракции ДНКазы определялась с помощью метода исследования с метиловым зеленым (MG).
Мутантную форму человеческой ДНКазы, имеющей в позиции 74 аминокислотной последовательности Asp остаток, получают путем сайт-направленного мутагенеза ДНК, кодирующей нативную зрелую человеческую ДНК. Эта мутантная форма элюируется вместе с первым пиком, получаемым по описанной выше процедуре хроматографирования (см. фигуру 7).
Ниже приведена методика наполнения колонки ворсистой катионообменной смолой ЛиКросфер® (LiChrospher®) 1000 SО3. Другим близким к ней видом ворсистой катионобменной смолы, пригодной для разделения дезамидированной и недезамидированной форм ДНКазы, является ворсистая катионообменная смола Фрактогель (Fractogel®) (ЕМ Separations, Gibbstown, New Jersey). Смолы ЛиКросфер и Фрактогель являются зарегистрированными торговыми марками ЕМ Индастриз, Инк., Хауторн Н.Й. ЕМ (ЕМ Industries, Inc., Hawthorne, N.Y., или Е.Мерк, Дармштадт, Германия (Е. Merck, Darmstadt, West Germany). В настоящее время авторы полагают, что "сильные" формы ворсистой катионообменной смолы, имеющей SО3 функциональные группы (независимо от того, ЛиКросфер это или Фрактогель), способны дать наилучшие результаты разделения.
3. Методика наполнения колонки для ВЭЖХ смолой ЛиКросфер® 1000 SО3
а. Материалы и оборудование:
1. Стеклянный картридж с наилучшими для разрешения параметрами и размерами 1,0 см × 5,0 см.
2. Буфер для наполнения: 100 мМ ацетата натрия, 1 мМ CaCl2, рН до 4,5, с уксусной кислотой. Фильтруют через фильтр с порами 0,2 мкм.
3. Емкость для наполнения колонки в 20 мл. (Alltech part #9501 или эквивалентный вариант).
4. Пустая колонка из нержавеющей стали размером 4,6 мм × 50 мм с фриттой предельного размера 0,5 мкм.
5. Насос для проведения ВЭЖХ, способный поддерживать противодавление, равное 2000 фунт/дюйм2 (Модель Вотерс 510 (Watters) или эквивалентный вариант).
b. Процедура наполнения колонки
1. Подготовка смолы к наполнению:
а) Распаковать стеклянную колонку для высокоэффективного разделения размером 1,0 см × 5,0 см (чистый объем = 3,93 мл смолы). Ресуспендировать смолу в 20 мл чистом стакане, влить в сосуд буфер для наполнения (упаковочный буфер). Полученную суспензию взбалтывают до однородности и разделяют на две аликвоты: 2 × 10 мл. К каждой аликвоте добавляют по 10 мл упаковочного буфера до получения суспензий приблизительного состава - 1,95 мл смолы в 20 мл упаковочного буфера.
b) Смолу взбалтывают с целью достижения однородной суспензии. Затем позволяют пройти процессу осаждения до тех пор, пока частицы на дне сосуда не образуют твердый слой (2-4 ч). Осторожно сливают супернатант, содержащий мелкие частицы.
c) Добавляют к смоле 20 мл упаковочного буфера и повторяют стадию b). Эту процедуру повторяют не менее четырех раз для уверенности в том, что все мелкие частицы смолы удалены.
2. Наполнение колонки
a) Соединяют пустую ВЭЖХ колонку размером 4,6 мм × 50 мм с упаковочной емкостью. Снова взбалтывают смолу в упаковочном буфере.
b) Вливают перемешанную смолу в упаковочную емкость и быстро закрывают ее. Накачивают упаковочный буфер в таком режиме, чтобы давление не превышало 2000 фунтов/дюйм2. Устанавливают скорость течения такой, чтобы давление наполнения оставалось на уровне 2000 фунтов/дюйм2, а поток продолжался в течение 15 мин после того, как давление стабилизируется. Далее отсоединяют колонку и закрывают верхний конец. Колонка может быть использована сразу же или может храниться в 0,02% азиде натрия.
Для большинства образцов, включая ДНКазу, приготовленную на 150 мМ NaCl, не требуется никакой подготовки образца перед нанесением на колонку. Колонку уравновешивают ацетатным буфером, рН 4,5, содержащим ионы кальция, вносят образец и затем проводят элюцию колонки с помощью солевого градиента. В случае небольших по масштабу процессов разделения дезамидированной и недезамидированной форм ДНКазы может быть использована следующая процедура. Соотношения зон пиков, вышедших на хроматограмме, соответствует соотношению дезамидированной и недезамидированной форм ДНКазы в образце.
Стадия 1. Загружают образец, содержащий до 150 мМ NaCl при рН 9 в автоматический пробоотборник. Образцы, содержащие жидкость с собранными клетками культуры, подвергают подведению рН до 4,5 и центрифугированию для удаления таких белков, которые нерастворимы в используемом по этой методике буфере.
Стадия 2. Разделяют две формы ДНКазы методом ВЭЖХ при следующих условиях:
Колонка: ТСХ ЛиКросфер® 1000 SО3, загруженный в колонку из нержавеющей стали. Размеры колонки 4,6 мм × 50 мм и 4,60 мм × 150 мм, уже наполненных и готовых в применению.
Температура колонки: соответствует температуре окружающей среды.
Элюент А: 10 мМ ацетата натрия, 1 мМ CaCl2, рН 4,5.
Элюент В: 1М NaCl в буфере А.
Градиентный профиль (см. табл. Б):
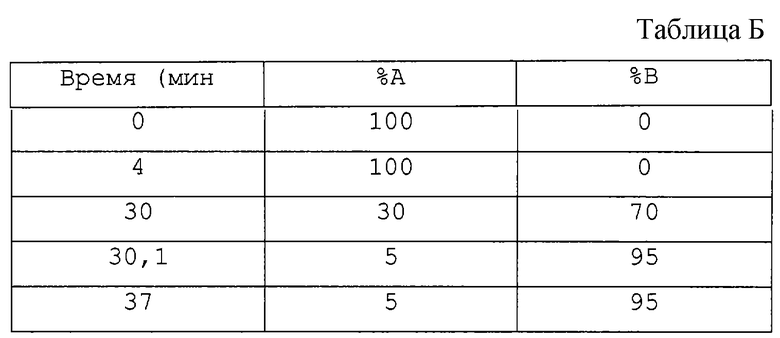
Скорость течения: 0,8 мл/мин (50 мм колонка), 0,5 мл/мин (150 мм колонка).
Объем наносимого образца: до 250 мкл.
Время уравновешивания колонки после отработки при 100% А: 20 мин.
Температура внутри автоматического пробоотборника: 5°.
Обнаружение: поглощение при 280 нм.
Стадия 3. Интегрируют хроматограмму. Вычисляют пропорцию дезамидированного вида на основе зоны пика элюированной раньше дезамидированной ДНКазы относительно общей зоны пиков обеих форм.
Хроматография на ворсистом катионообменнике дает также способ разделения в больших масштабах дезамидированной и недезамидированной форм человеческой ДНКазы. Широкомасштабные разделения наиболее удобно осуществлять с использованием более упрощенной схемы элюции, чем это описано для небольших по объему партий аналитического разделения двух форм ДНКаза. В этой связи крупномасштабное разделение производят с применением ворсистого катионита Фрактогеля на основе следующей процедуры рН - элюции.
Стадия 1. Заполняют 31,6 колонку (1,6 см внутр. диам. × 15,7 см высоты) Фрактогелем EMD SО3 650-М ворсистой катионообменной смолы (ЕМ Separations, Gibbstown, New Jersey).
Стадия 2. ДНКазу после диафильтрации загружают уравновешивающим буфером (30 мМ ацетата натрия (NaAc). 1 мМ Хлорида кальция (CaCl2), 50 мМ хлорида натрия (NaCl), рН 5). Концентрируют с помощью ультрафильтрации до объема 355 мл и концентрации 2,5 мг/мл.
Стадия 3. Промывают колонку 2,5 объемами колонки (CV) 2% раствором гидроксида натрия (NaOH).
Стадия 4. Промывают колонку 2,5 CV предварительно уравновешенного буфера (300 мМ NaAc, 1M NaCl, рН 5).
Стадия 5. Промывают колонну 2,5 CV уравновешивающего буфера.
Стадия 6. Загружают колонку 1-1,3 г диафильтрованной/ультрафильтрованной ДНКазы (от стадии 2).
Начинают сбор фракций вытекающего из колонки эффлюента с момента нанесения ДНКазы.
Стадия 7. Промывают колонку 5 CV уравновешивающего буфера.
Стадия 8. Промывают колонку 5 CV промывочного буфера, рН 5,3 (25 мМ сукцината, 1 мМ CaCl2, рН 5,3).
Стадия 9. Промывают колонку 10 CV промывочного буфера, рН 5,4 (25 мМ сукцината, 1 мМ CaCl2, рН 5,4).
Стадия 10. Промывают колонку 10 CV промывочного буфера, рН 6 (25 мМ MES, 1 мМ CaCl2, рН 6,0).
Стадия 11. Для получения пула, состоящего главным образом из дезамидированной ДНКазы, объединяют фракции, собранные на стадиях 6-8. Для получения пула, состоящего из недезамидированной ДНКазы, объединяют фракции, собранные в течение стадии 10. Фракции, собранные на стадии 9, содержат смесь двух форм ДНКазы и могут быть вновь подвергнуты разделению.
Описанная выше процедура представляет собой всего один пример использования ворсистой ионообменной смолы-катионита для разделения двух форм рекомбинантной человеческой ДНКазы в условиях, пригодных для крупномасштабного выделения очищенной дезамидированной и очищенной недезамидированной ДНКаз.
4. Хроматография с применением гепарина и иммобилизованного аналога ДНК
На фигуре 8 приведены данные по хроматографии на TSK-гепариновых колонках (Toso Haas, Montgomeryville, Pennsylvania) образцов, содержащих либо смесь дезамидированной и недезамидированной форм либо очищенную дезамидированную человеческую ДНКазу, либо очищенную недезамидированную человеческую ДНКазу. Хроматография на TSK-гепариновой колонке проводилась при тех же самых условиях, что были описаны для аналитического варианта на ТСХ колонке. Выровненные на фиг.8 хроматограммы демонстрируют, что колонка с иммобилизованным гепарином хорошо разрешает дезамидированную и недезамидированную формы ДНКазы.
Как было отмечено выше, другим способом разделения дезамидированной и недезамидированной форм ДНКаз является применение колонки, содержащей иммобилизованный аналог ДНК, устойчивый к гидролизу ДНКазой. Один из примеров такого подхода к использованию колонки с иммобилизованным аналогом ДНК включает синтез фосфоротиоатного олигонуклеотида: 5’-GCGCGCGCGCGCGCGCGCGCGC-NН3-3'. Эта самокомплементарная последовательность может быть превращена в двухцепочечную форму и далее введена в Рейнин-Гидропор-ЕР Колонку (Rainin Co., Woburn, Massachussetts). На фигуре 9 представлены данные по хроматографированию на этой колонке образцов, содержащих либо смесь дезамидированной и недезамидированной форм человеческой ДНКазы, очищенную дезамидированную человеческую ДНКазу, очищенную недезамидированную человеческую ДНКазу или очищенную мутантную человеческую ДНКазу, имеющую остаток аспарагиновой кислоты (а не остаток аспарагина) в позиции 74 аминокислотной последовательности. Колонка работала с целью такого анализа в буфере, содержащем 1 мМ хлорида кальция, 5 мМ/MES при рН 6, элюировали градиентом с концентрацией соли до 1 М хлорида натрия свыше 20 мин при скорости течения 1 мл/мин. Как показано на фигуре 9, при этих условиях дезамидированная и недезамидированная формы ДНКазы частично отделялись друг от друга. Кроме того, 2 изомерных формы дезамидированной ДНКазы, которые отличаются по позиции 74 аминокислотной последовательности тем, что имеют либо аспарагиновую кислоту, либо изо-аспарагиновую кислоту, также разрешаются этой колонкой. Таким образом, дополнительным плюсом данного хроматографического метода является то, что он позволяет проводить разделение двух изомеров, возникающих при дезамидировании человеческой ДНКазы.
5. Ферментативная активность дезамидированной человеческой ДНКазы и недезамидированной человеческой ДНКазы
Использовали несколько методов для выявления и изучения влияния дезамидирования на ферментативную активность человеческой ДНКазы.
Очищенную дезамидированную человеческую ДНКазу и очищенную недезамидированную человеческую ДНКазу для использования в приведенных далее тестах получали с помощью хроматографии на ТСХ по описанному выше методу.
В одном из методов для определения ферментативной активности ДНКазы использовали синтезированную двухцепочечную ДНК-25 пар оснований в длину, которую метили на одном конце динитрофенолом (ДНФ), а на другом конце - биотином. Гидролиз субстрата ДНКазой определяли по связыванию реакционных продуктов на лунках микротитрационного плато, покрытых антителом к ДНФ, а также количественно с помощью реакции интактной пробы со стрептавидин-пероксидазой хрена. Удельная активность обладающих стабильностью образцов коррелировала (r2=0,613: n=5) с уровнем дезамидированности ДНКазы (в пределах от 27 до 93%). Экстраполяция линейного уравнения дает оценку удельной активности дезамидированной человеческой ДНКазы примерно на 77% ниже, чем соответствующий показатель для недезамидированной человеческой ДНКазы.
Другой метод определения ферментативной активности ДНКазы включает гидролиз хромогенного субстрата п-нитрофенилфенилфосфоната (ПНФФ) по методу Лиао с соавт. (Liao, et al., Biochem. J.  :781-787 (1988)). Кинетика гидролиза ПНФФ человеческой ДНКазой представлена сигмоидной кривой и описывается уравнением нелинейной регрессии Хилла. С применением этого метода было показано, что Vmax полностью дезамидированной человеческой ДНКазы на 77% ниже, чем у недезамидированной человеческой ДНКазы. Концентрация субстрата для активности, равной половине максимальной (S0,5), существенно не отличалась для дезамидированного и недезамидированного образцов ДНКазы.
:781-787 (1988)). Кинетика гидролиза ПНФФ человеческой ДНКазой представлена сигмоидной кривой и описывается уравнением нелинейной регрессии Хилла. С применением этого метода было показано, что Vmax полностью дезамидированной человеческой ДНКазы на 77% ниже, чем у недезамидированной человеческой ДНКазы. Концентрация субстрата для активности, равной половине максимальной (S0,5), существенно не отличалась для дезамидированного и недезамидированного образцов ДНКазы.
Следующий метод определения ферментативной активности представляет собой тест, описанный Кунитцем (Kunitz, J. Gen. Physiol. 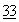 :349 (1950), в значительной степени модифицированный так, что ферментативная реакция проводится при рН примерно 7,0-7,5. Этот метод также показал, что ферментативная активность дезамидированной человеческой ДНКазы ниже, чем таковая у недезамидированной человеческой ДНКазы.
:349 (1950), в значительной степени модифицированный так, что ферментативная реакция проводится при рН примерно 7,0-7,5. Этот метод также показал, что ферментативная активность дезамидированной человеческой ДНКазы ниже, чем таковая у недезамидированной человеческой ДНКазы.
6. Хранение человеческой ДНКазы in vitro
Очищенную человеческую ДНКазу из рекомбинантных СНО клеток растворяют до концентрации 4 мг/мл в незабуференном водном растворе 150 мМ NaCl и 1 мМ СаСl2. Затем образцы полученного раствора ДНКазы помещают в стеклянные и пластиковые ампулы. При этом используют два типа пластиковых ампул: один из них приготовлен на основе пластиковой смолы Дюпон 20 (Dupont 20) (производство E.I. du Font de Nemours & Co., Inc., Wilmington, Delaware USA), а другой сделан на основе пластиковой смолы Эскорен (Escorene) (производство Exxon Corp.). Оба вида пластиковых материалов представляют собой полиэтилен низкой плотности, однако, могут также использоваться контейнеры, приготовленные на основе других пластиков, таких как полипропилен, полистирол или другие полиолефины. Ампулы, содержащие раствор ДНКазы, хранятся при температурах: -70, 2-8 или 25°С. Первоначально растворы содержат до 60-65% ДНКазы в дезамидированной форме.
Растворы в ампулах исследовались неоднократно после начала хранения с целью определения уровня дезамидированности ДНКазы.
Результаты исследований представлены в таблице 3.
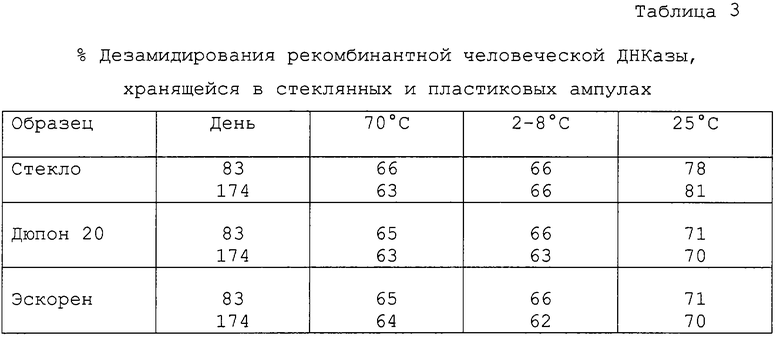
После 83 и 174 дней хранения при -70 или 2-8°С не было обнаружено различий в количестве дезамидированной ДНКазы как в пластиковых, так и в стеклянных ампулах. В каждом таком случае примерно 64% (±2%) ДНКазы в ампулах было представлено дезамидированной ДНКазой.
Однако неожиданно было отмечено различие при хранении препарата в течение 83 или 174 дней при 25°С в количестве дезамидированной ДНКазы в пластиковых и в стеклянных ампулах. В пластиковых ампулах присутствовало значительно меньшее количество дезамидированной ДНКазы. В частности, через 83 дня хранения при 25°С 78% ДНКазы в стеклянных ампулах было представлено дезамидированной ДНКазой, тогда как в пластиковых ампулах - только около 70% ДНКазы составляла дезамидированная ДНКаза. Через 174 дня хранения при 25°С 81% ДНКазы в стеклянных ампулах было представлено дезамидированной ДНКазой, тогда как только около 71% ДНКазы в пластиковых ампулах было представлено дезамидированной ДНКазой.
Не ограничивая изобретение определенным механизмом или определенной теоретической основой действия, мы тем не менее предположили, что различия в величине дезамидированной ДНКазы в пластиковых и стеклянных ампулах может быть следствием различий рН растворов в разных ампулах. Изначально рН раствора ДНКазы в стеклянной ампуле был несколько выше, чем в пластиковой (около рН 6,7 и около рН 6,5 соответственно), рН раствора ДНКазы в стеклянных ампулах продолжал с течением времени незначительно возрастать (примерно до рН 6,9 через 83 дня хранения при 25°С и примерно до 7,0 через 174 дня хранения при 25°С), возможно, как следствие растворения силикатов или ионов с поверхности стекла. При более высоком рН скорость дезамидирования человеческой ДНКазы увеличивается. Поскольку ранее не был оценен тот факт, что при повышении рН имеет место дезамидирование ДНКазы, одним из вариантов настоящего изобретения является разработка способа приготовления и/или хранения человеческой ДНКазы в растворах, имеющих кислый рН, в типичном случае при рН 4,5-6,8 и в наиболее предпочтительных случаях при рН около 5,0-6,8.
Таким образом, было достигнуто значительно улучшение свойств стабильности раствора человеческой ДНКазы при помещении такого раствора в пластиковые, а не стеклянные ампулы, поскольку при такой замене происходит значительно меньшее дезамидирование ДНКазы с течением времени. Это открытие существенно для выбора упаковки для препарата человеческой ДНКазы, используемой в терапевтических целях, особенно когда желательно, чтобы препарат человеческой ДНКазы был способен к хранению в течение длительного времени без значительного снижения ферментативной активности. Разумеется, стеклянные ампулы с нестеклянным выстилающим слоем будут одинаково полезны для такой цели. Важно, однако, избегать при хранении ДНКазы контакта со стеклом, особенно при хранении в течение срока свыше 15-30 дней.
Широко известно, что ДНКаза I является белком, применяемым для снижения вязкости гноя путем гидролитического расщепления ДНК в указанном гное. Под торговым наименованием Pulmozyme® ДНКаза I одобрена для такого применения при муковисцидозе.
ОБЩИЕ ЗАМЕЧАНИЯ
Ниже приведено описание специфических деталей, имеющих отношение к применению настоящего изобретения. Имея подробные специфические методы для идентификации, характеристики, разделения и использования очищенных дезамидированной и недезамидированной человеческих ДНКаз, равно как и знание специфических модельных систем их поддержания в активном состоянии, каждый, имеющий средний уровень знаний в данной области, может самостоятельно разработать собственные альтернативные способы использования плодов настоящего изобретения на основе имеющейся в нем информации. Несмотря на то, что ниже в тексте может встретиться излишняя детализация, приводимые примеры не следует истолковывать в плане какого-либо ограничения области настоящего изобретения: границы настоящего изобретения будут закономерно определены в формуле изобретения.


Изобретение относится к биотехнологии, касается идентификации и характеристики двух компонентов рекомбинантного препарата ДНКазы. Эти компоненты представляют собой очищенную дезамидированную и очищенную недезамидированную человеческую ДНКазу. Способы разделения этих ДНКаз осуществляют с помощью полидентатной катионообменной смолы или с помощью смолы с иммобилизованным на ней гепарином или с помощью смолы с иммобилизованным на ней негидролизуемым аналогом ДНК. Использование недезамидированной ДНКазы в составе фармацевтической композиции эффективно для введения пациентам, страдающим от нарушений легочного характера. 13 н.п. ф-лы, 10 ил., 5 табл.

| WO 9007572 А, 12.07.1990. | |||
| US 4065355, 27.12.1977. | |||
| М.Д | |||
| Машковский | |||
| Лекарственные средства, ч.2 | |||
| - М.: Медицина, 1965, с.54. | |||
| JOHN FRENZ et al | |||
| Characterization of human growth hormone by capillary electrophoresis, Journal OF CHROMATOGRAPHY., 1989, v.480, p.379-391. |
