Область техники, к которой относится изобретение
Настоящее изобретение относятся к композициям, включающим антитела и их антигенсвязывающие фрагменты, в которых отсутствует сульфатирование тирозина, а также к способам очистки для получения композиций.
Предпосылки создания изобретения
Сульфатирование тирозина представляет собой пост-трансляционную модификацию (PTM), где группа сульфат-триоксида (SO3) ковалентно связана с гидроксильной группой на боковой цепи тирозиновой аминокислотной группы. Эта PTM происходит в сети транс-Гольджи и катализируется двумя ферментами тирозилпротеинсульфотрансферазами (TPST). Молекулярный механизм включает перенос активированного сульфата из 3'-фосфоаденозин-5'-фосфосульфата к тирозину, и был обнаружен на различных белках и пептидах. Полученные в последнее время данные показывают, что тирозилпротеинсульфотрансфераза 2 распознает тирозины, фланкированные кислотными остатками, для сульфатирования. Эта PTM является ответственной за усиление взаимодействий между белками и происходит на секретированных и трансмембранных белках. Было показано что некоторые хемокиновые рецепторы являются тирозинсульфатированными, например, в N-концевом внеклеточном домене CCR5, основном рецепторе для ВИЧ-1 и некоторых рецепторах гликопротеиновых гормонов. Например, нативная форма происходящего из пиявки ингибирующего тромбин пептида гирудина является тирозинсульфатированной. Интересно то, что, две рекомбинантные формы гирудина (Revasc и Refludan), используемые для лечения различных расстройств свертывания крови, не являются сульфатированными. Сульфатирование увеличивает массу биомолекулы на 80 Да, что является такой же разницей в массе, как фосфатная группа (PO3). В отличие от PO3, которая образует достаточно стабильную P-O связь, SO3 является очень лабильной и легко разлагается при высокой температуре и низком pH.
Присутствие разных PTM вариантов в терапевтическом препарате антител приводит к гетерогенности, что, в зависимости от локализации модификации, может привести к изменениям активности, биодоступности или иммуногенности антитела. Такие проблемы также создают проблемы для органов государственного регулирования. Хотя сульфатирование тирозина было описано в хемокиновых рецепторах и других белках, необходимо определить, происходят ли такие модификации в препаратах антител, и, если они будут выявлены, удалить их.
Сущность изобретения
Настоящее изобретение обеспечивает композицию, включающую анти-LAG3 антитело или его антигенсвязывающий фрагмент (например, Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9), которое, например, включает:
вариабельный домен легкой цепи, включающий:
CDR-L1, которая включает аминокислотную последовательность: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2, которая включает аминокислотную последовательность: GASNLES (SEQ ID NO: 39); и
CDR-L3, которая включает аминокислотную последовательность: QQSTEDPRT (SEQ ID NO: 40); и/или
вариабельный домен тяжелой цепи, включающий:
CDR-H1, которая включает аминокислотную последовательность: DYNVD (SEQ ID NO: 33);
CDR-H2, которая включает аминокислотную последовательность:
DINPNNGGTIYAQKFQE (SEQ ID NO: 59);
DINPNSGGTIYAQKFQE (SEQ ID NO: 60);
DINPNDGGTIYAQKFQE (SEQ ID NO: 61);
DINPNQGGTIYAQKFQE (SEQ ID NO: 62);
DINPNGGGTIYAQKFQE (SEQ ID NO: 63); или
DINPNX1GGTIYX2QKFX3X4 (SEQ ID NO: 64) где, X1= D,N,S или Q, X2= A или S, X3= Q или K, и X4= E или G; и CDR-H3: NYRWFGAMDH (SEQ ID NO: 35); в котором отсутствуют определяемые уровни сульфатированного тирозина на CDR-L1. Например, в одном варианте осуществления изобретения антитела или фрагменты в композиции также не содержат определяемых уровней сульфатированного тирозина в одном или нескольких членах, выбранных из группы, включающей FR-L1, FR-L2, CDR-L2, FR-L3, CDR-L3, FR-L4, FR-H1, CDR-H1, FR-H2,CDR-H2, FR-H3,CDR-H3, FR-H4 и константный домен. В одном варианте осуществления изобретения антитело или фрагмент включает N-связанные гликаны рекомбинантных дрожжей или CHO. В одном варианте осуществления изобретения композиция, содержащая анти-LAG3 антитело (например, Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9), включает одну или несколько разновидностей антитела, не имеющих сульфатирования тирозина и имеющих молекулярные массы около 148590 Да, 148752 Да и/или 148914 Да (например, которые содержат G0F и/или G1F виды гликанов, например, как показано в Таблице 1, и в которых N-концевой глутамин в тяжелой цепи преобразован в пироглутамат и/или C-концевой лизин в тяжелой цепи удален).
Настоящее изобретение также обеспечивает способ для удаления содержащих сульфатированный тирозин антител или их антигенсвязывающих фрагментов (например, Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9) из водной смеси, включающей антитела или антигенсвязывающие фрагменты, которые включают один или несколько сульфатированных тирозинов (например, на CDR-L1), и антитела или антигенсвязывающие фрагменты, не содержащие сульфатированного тирозина, включающий доведение pH смеси до около 6,5 - около 7,0 или до около 6,5 - около 7,5, контактирование смеси с анионообменной смолой и удаление и сохранение не связанной со смолой водной фракции смеси из смолы. В одном варианте осуществления изобретения способ включает промывку колонки водной композицией, например, в изократических условиях, и удаление и сохранение промывочной композиции из смолы. В одном варианте осуществления изобретения смола находится в колонке, и способ включает добавление указанной смеси в колонку и сбор проточной фракции из колонки. В одном варианте осуществления изобретения способ включает уравновешивание хроматографической смолы, включающей диметиламинопропильный анионообменный лиганд, в хроматографической колонке раствором 25 мМ фосфата натрия pH 6,5, доведение pH смеси до около 6,5, нанесение смеси на колонку, сбор проточной фракции из колонки, промывку смолы в колонке раствором 25 мМ фосфата натрия pH 6,5 и сбор проточной фракции из промывки. В одном варианте осуществления изобретения способ включает уравновешивание хроматографической смолы, включающей кватернизированный полиэтилениминовый анионообменный лиганд, в хроматографической колонке раствором 25 мМ фосфата натрия pH 7,0; необязательно, 5 мМ NaCl, доведение pH смеси до около 7,0, нанесение смеси на колонку, сбор проточной фракции из колонки, промывку смолы в колонке раствором 25 мМ фосфата натрия pH 7,0; необязательно 5 мМ NaCl, и сбор проточной фракции из промывки. В одном варианте осуществления изобретения измеряют A280 поглощение проточной фракции анионообменной хроматографии и ее собирают и сохраняют, когда A280 сначала достигает по меньшей мере около 2,5 единиц оптической плотности/см; и не собирают и не сохраняют, когда A280 падает ниже около 1,0 единиц оптической плотности/см. В одном варианте осуществления изобретения способы по настоящему изобретению дополнительно включают очистку антитела или антигенсвязывающего фрагмента катионообменной хроматографией, далее анионообменной хроматографией в режиме связывания-элюирования, хроматографией гидрофобных взаимодействий, хроматографией с протеином-A, хроматографией с протеином-L, хроматографией с протеином-G, хроматографией на гидроксиапатите, эксклюзионной хроматографией, фракционированным осаждением, фильтрованием, центрифугированием или вирусной инактивацией. В одном варианте осуществления изобретения легкие цепи и/или тяжелые цепи иммуноглобулинов антитела или антигенсвязывающего фрагмента экспрессируются в клетках яичников китайского хомячка. В одном варианте осуществления изобретения антитело или антигенсвязывающий фрагмент включает:
вариабельный домен легкой цепи, включающий:
CDR-L1, которая включает аминокислотную последовательность: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2, которая включает аминокислотную последовательность: GASNLES (SEQ ID NO: 39); и
CDR-L3, которая включает аминокислотную последовательность: QQSTEDPRT (SEQ ID NO: 40); и/или
вариабельный домен тяжелой цепи, включающий:
CDR-H1, которая включает аминокислотную последовательность: DYNVD (SEQ ID NO: 33);
CDR-H2, которая включает аминокислотную последовательность:
DINPNNGGTIYAQKFQE (SEQ ID NO: 59);
DINPNSGGTIYAQKFQE (SEQ ID NO: 60);
DINPNDGGTIYAQKFQE (SEQ ID NO: 61);
DINPNQGGTIYAQKFQE (SEQ ID NO: 62);
DINPNGGGTIYAQKFQE (SEQ ID NO: 63); или
DINPNX1GGTIYX2QKFX3X4 (SEQ ID NO: 64), где X1= D,N,S или Q, X2= A или S, X3= Q или K, и X4= E или G; и CDR-H3: NYRWFGAMDH (SEQ ID NO: 35). Композиции, которые являются продуктом такого способа, также являются частью настоящего изобретения. В одном варианте осуществления изобретения анти-LAG3 антитело (например, Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9) очищают AEX хроматографией, где антитела, не содержащие сульфатированный тирозин, имеют молекулярные массы около 148590 Да, 148752 Да и/или 148914 Да (например, содержат G0F и/или G1F виды гликанов, например, как показано в Таблице 1, N-концевой глутамин в тяжелой цепи преобразованный в пироглутамат и/или удаленный лизин на C-конце тяжелой цепи).
Краткое описание чертежей
Фиг. 1. Наложение IEX-ВЭЖХ УФ профиля исходного образца (жирная линия), десорбированной (тонкая линия) и объединенной фракции (пунктирная линия) AEX.
Фиг. 2. Спектр интактных масс AEX исходного, объединенного и десорбированного образцов.
Фиг. 3. Масс-спектр восстановленной легкой цепи объединенного и десорбированного образцов AEX.
Фиг. 4A-B. УФ-спектр, полученный картированием LysC восстановленного пептида объединенной и десорбированной AEX фракций.
Фиг. 5A-C. (A)Спектр СИД фрагментации легкой цепи AA25-43+80 Да в масштабе 400-1800 m/z (B) Измененный в масштабе m/z 300-1100 (C) Измененный в масштабе m/z 1200-2000.
Фиг. 6. ДПЭ фрагментация пептида легкой цепи AA25-43+80 Да. 80Да присоединенные фрагментарные ионы помечены.
Фиг. 7A-B. (A) Спектры интактной массы после деконволюции AEX десорбированной фракции с обработкой и без обработки щелочной фосфатазой. (B) Спектры интактной массы после деконволюции куриного овальбумина с обработкой и без обработки щелочной фосфатазой.
Фиг. 8A-B. (A) Нормализованные концентрации AEX объединенной и десорбированной фракций mAb подвергали SDS-PAGE в восстановительных условиях, зондировали на человеческие тяжелые (HC) и легкие цепи (LC) гибридизацией с использованием вестерн-блоттинга (верхняя панель), затем десорбировали и снова зондировали на антисульфотирозин (нижняя панель). См. указания HC и LC справа. (B) Нормализованные концентрации различных CHO-происходящих mAb в дополнение к AEX десорбированной и объединенной фракции подвергали SDS-PAGE в восстановительных условиях, зондировали на человеческие HC и LC гибридизацией с использованием вестерн-блоттинга, затем десорбировали и снова зондировали на антисульфотирозин. Для обоих (A) и (B) MagicMark XP использовали в качестве стандарта молекулярной массы белка и равные количества HEK293 и EGF-обработанных A431 клеточных экстрактов анализировали в качестве контролей.
Фиг. 9A-C. SIC (A) LC25-43+80 Да из Десорбированной Фракции AEX, (B) Синтетический Пептид XSXSXDYEGDSDXXXXXXX (SEQ ID NO: 65)+Фосфорилирование и (C) Синтетический Пептид XSXSXDYEGDSDXXXXXXX (SEQ ID NO: 65)+Сульфатирование.
Фиг. 10. Тирозиновый (Y31) участок mAb, показывающий CDR петли в ленточной диаграмме как для тяжелой, так и легкой цепи.
Фиг. 11. Преимущественные N-связанные гликаны для моноклональных антител, которые были получены в клетках яичников китайского хомячка (CHO N-связанные гликаны) и в генно-инженерных дрожжевых клетках (генно-инженерные дрожжевые N-связанные гликаны): квадраты: N-ацетилглюкозамин (GlcNac); кружки: манноза (Man); ромбы: галактоза (Gal); треугольники: фукоза (Fuc).
Подробное описание изобретения
Некоторые антитела и другие белки, экспрессированные в клетках яичников китайского хомячка(CHO), загрязнены вариантами, содержащими сульфатированный тирозин. Масс-спектрографический анализ таких вариантов характеризуется аддуктом около +80 Да, что соответствует массе добавленной сульфатной группы. Такие аддукты также резистентны к щелочной фосфатазе и способны реагировать с антителами против сульфатированного тирозина. Настоящее изобретение обеспечивает способ для очистки композиции, включающей такие загрязняющие включающие сульфатированный тирозин варианты, а также композиции антител, по существу не содержащие таких вариантов.
В соответствии с настоящим изобретением, можно использовать традиционные методы молекулярной биологии, микробиологии и рекомбинантной ДНК, известные специалистам в данной области. Если не определено иначе, научные и технические термины, используемые в связи с настоящим изобретением, должны иметь значения, которые хорошо известны средним специалистам в данной области. Кроме того, если контекст не диктует иное, термины в единственном числе должны включать множественное число, а термины во множественном числе должны включать единственное число. Как правило, номенклатуры, используемые в связи с методами биохимии, энзимологии, молекулярной и клеточной биологии, микробиологии, генетики и химии белков и нуклеиновых кислот и гибридизации, которые описаны в настоящей заявке, являются хорошо известными и широко используемыми в данной области техники. Способы и процедуры настоящего изобретения обычно осуществляют в соответствии с традиционными способами, хорошо известными в данной области техники, и как описано в различных общих и более конкретных ссылках, которые цитируются и обсуждаются в настоящем описании, если не указано иное. См., например, James M. Cregg (Editor), Pichia Protocols (Methods in Molecular Biology), Humana Press (2010), Sambrook et al. Molecular Cloning: A Laboratory Manual, 2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates (1992, and Supplements to 2002); Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1990); Taylor and Drickamer, Introduction to Glycobiology, Oxford Univ. Press (2003); Worthington Enzyme Manual, Worthington Biochemical Corp., Freehold, N.J.; Handbook of Biochemistry: Section A Proteins, Vol I, CRC Press (1976); Handbook of Biochemistry: Section A Proteins, Vol II, CRC Press (1976); Essentials of Glycobiology, Cold Spring Harbor Laboratory Press (1999), Animal Cell Culture (R.I. Freshney, ed. (1986)); Immobilized Cells And Enzymes (IRL Press, (1986)); B. Perbal, A Practical Guide To Molecular Cloning (1984).
Сульфатированный тирозин включает тирозин, содержащий присоединенную сульфатную группу, например, имеющий структуру:
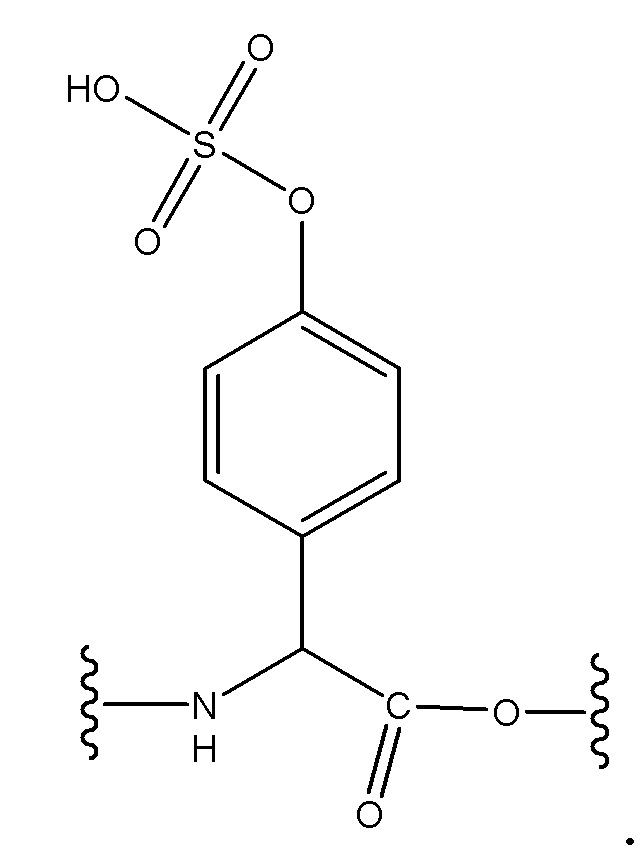
Хроматография
Настоящее изобретение обеспечивает способ для удаления загрязняющих вариантов антител или их антигенсвязывающих фрагментов (например, Ab1-Ab9), которые включают сульфатированный тирозин, из композиции, например, композиции, которая включает смесь антител или фрагментов, некоторые из которых содержат сульфатированный тирозин, а некоторые из которых не содержат сульфатированного тирозина, для получения композиции, включающей неопределяемые уровни сульфатированных вариантов тирозина (например, CDR-L1 с сульфатированным тирозином, например, из Ab1 или Ab6). В одном варианте осуществления изобретения композицию очищают анионообменной (AEX) хроматографией в проточном режиме для удаления сульфатированных вариантов тирозина. В одном варианте осуществления изобретения AEX смола содержит диметиламинопропильный лиганд (т.е. лиганд, который включает диметиламинопропильную группу). Например, в одном варианте осуществления изобретения композиция, которую подвергают AEX хроматографии, является продуктом предварительной очистки хроматографией с протеином-A. В одном варианте осуществления изобретения pH композиции доводят до pH около 6,5, например, при помощи Tris (например, 0,5M, 0,725M или 1M), перед AEX очисткой (например, содержащей диметиламинопропильный лиганд). В одном варианте осуществления изобретения AEX колонку (например, содержащую диметиламинопропильный лиганд) уравновешивают, например, фосфатом натрия, например, 25 мМ, например, фосфатом натрия pH 5, 6,2 или 6,5. Колонку (например, содержащую диметиламинопропильный лиганд) можно, в одном варианте осуществления изобретения, промыть буфером (например, фосфатом натрия, например, 25 мМ, например, фосфатом натрия pH 6,5) для выделения антитела или фрагмента, находящегося в колонке, но не сильно связанного с AEX смолой. Элюат, не сильно связанный с AEX смолой, собирают (например, в виде отдельных фракций) и, например, объединяют. В одном варианте осуществления изобретения после использования в колонке осуществляют десорбцию, например, раствором 1M NaCl.
Масс-спектрометрический анализ AEX элюата выявил некоторые гликозилированные разновидности Ab6 с отсутствием сульфатирования тирозина на CDR-L1. Эти разновидности кратко описаны ниже в Таблице 1. Эти теоретические массы относятся к рассчитанной массе молекулы Ab6, где N-концевой глутамин на тяжелой цепи преобразован в N-концевую пироглутаминовую кислоту (pE1) и C-концевой лизином на тяжелой цепи удален (-K).
/G0F, pE1, -K
/G0F, pE1, -K
/G1F, pE1, -K
Настоящее изобретение включает композицию, включающую анти-LAG3 антитела (например, Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9; предпочтительно Ab6), не содержащие определяемых уровней сульфатирования тирозина, например, на CDR-L1, включающие разновидности, имеющие одну или несколько из молекулярных масс около 148590, 148749 и/или 148915; и/или включающие виды гликанов G0F и/или G1F.
Проточный режим относится к очистке полипептида с использованием хроматографической смолы способом, который не включает стадию элюирования для выделения полипептида. В таком способе полипептид, представляющий интерес, не связывается сильно со смолой, но загрязняющие вещества, подлежащие удалению из полипептидов, представляющих интерес, сильно связываются со смолой. Например, AEX смолу используют в проточном режиме в способе, включающем загрузку композиции, которая включает загрязненные варианты антител с сульфатированием тирозина и антитела с отсутствием сульфатирования тирозина, на колонку, содержащую AEX смолу, и сбор и удерживание антитела или фрагмента в проточной фракции колонки. Несвязанное антитело с отсутствием сульфатирования можно вымывать из колонки (и сохранять) в условиях, которые не приводят к элюированию, например, изократические условия. В таком способе загрязняющая примесь остается связанной с колонкой, а антитело с отсутствием сульфатирования тирозина будет оставаться в проточной фракции.
Режим связывания/элюирования относится к очистке полипептида с использованием хроматографической смолы способом, который включает стадию элюирования. В таком способе полипептид, представляющий интерес, сильно связывается со смолой, а загрязняющие вещества, подлежащие удалению из полипептидов, представляющих интерес, не связываются сильно со смолой. В хроматографической колонке загрязняющие примеси протекают через колонку и остаются в основном несвязанными со смолой. Связанные антитела, после необязательной промывки, высвобождают и собирают и сохраняют при воздействии элюирующим буфером, который вызывает их высвобождение из смолы.
Лиганд хроматографической смолы представляет собой вещество, которое прикрепляется к частице неподвижной фазы (например, частице сефарозы), которая обратимо связывается с желаемой молекулой (например, антителом или примесью), присутствующей в многокомпонентной подвижной фазе.
В одном варианте осуществления изобретения AEX смола содержит лиганд кватернизированный полиэтиленимин (т.е. лиганд, который включает кватернизированную полиэтилениминовую группу). В одном варианте осуществления изобретения смолу (например, содержащую кватернизированный полиэтилениминовый лиганд) предварительно уравновешивают раствором 1M NaCl. В одном варианте осуществления изобретения смолу (например, содержащую кватернизированный полиэтилениминовый лиганд) уравновешивают фосфатом натрия, например 25 мМ, и NaCl, например, 5 мМ; pH около 7,0. В одном варианте осуществления изобретения колонку (например, содержащую кватернизированный полиэтилениминовый лиганд) загружают исходной смесью и промывают фосфатом натрия, например 25 мМ, и NaCl, например 5 мМ; pH около 7,0; и вытекающий из колонки раствор собирают, например в виде отдельных фракций, и например объединяют. В другом варианте осуществления изобретения способ по изобретению включает уравновешивание хроматографической смолы, включающей анионообменный лиганд, в хроматографической колонке при помощи около 10-50 мМ фосфата натрия; pH около 6,5-7,5, доведение pH смеси до около 6,5-7,5, нанесение смеси на колонку, сбор проточной фракции из колонки, промывку смолы в колонке раствором фосфата натрия около 10-50 мМ; pH около 6,5-7,5 и сбор проточной фракции из промывки. В другом варианте осуществления изобретения способ по изобретению включает уравновешивание хроматографической смолы, включающей анионообменный лиганд, в хроматографической колонке раствором фосфата натрия около 10-50 мМ; pH около 6,5-7,0, доведение pH смеси до около 6,5-7,0, нанесение смеси на колонку, сбор проточной фракции из колонки, промывку смолы в колонке раствором фосфата натрия около 10-50 мМ; pH около 6,5-7,0, и сбор проточной фракции из промывки.
Любое подходящее количество антитела или антигенсвязывающего фрагмента можно загружать на хроматографическую смолу, например, хроматографическую колонку (например, AEX, содержащую кватернизированный полиэтилениминовый лиганд или диметиламинопропильный лиганд). Например, в одном варианте осуществления изобретения около 100, 110, 120, 130, 140, 150, 100-150, 160, 170, 180, 190, 200, 300, 150-200, 100-200, 250-350, или 280-320 граммов вещества, например антитела или фрагмента, загружают на литр смолы (например, AEX, содержащей кватернизированный полиэтилениминовый лиганд или диметиламинопропильный лиганд).
Если используют хроматографическую колонку (например, содержащую AEX смолу, содержащую кватернизированный полиэтилениминовый лиганд или диметиламинопропильный лиганд), можно использовать любые приемлемые размеры. Например, в одном варианте осуществления изобретения диаметр или высота колонки составляет около 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 см.
Скорость потока относится к объему подвижной фазы, проходящему через колонку (например, содержащую AEX смолу, содержащую кватернизированный полиэтилениминовый лиганд или диметиламинопропильный лиганд) за определенный период времени. В одном варианте осуществления изобретения скорость потока составляет около 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 105, 110, 115, 120, 125, 130, 135, 140, 145, 150, 155, 160, 165, 170, 175, 180, 185, 190, 195, 200, 205, 210, 215 литров в час.
В одном варианте осуществления изобретения измеряют поглощение при 280 нм (A280) проточной фракции колонки (например, содержащей AEX смолу, содержащую кватернизированный полиэтилениминовый лиганд или диметиламинопропильный лиганд). В одном варианте осуществления изобретения продукт антитела или фрагмента в основном A280 пике проточной фракции собирают и сохраняют. В одном варианте осуществления изобретения проточную фракцию собирают, когда A280 достигает около 1,0, 1,5, 2,0, 2,5 или 3,0 A280 оптических единиц на см (длина оптического пути), и сбор прекращается, когда A280 падает ниже около 1,0, 1,5, 2,0, 2,5 или 3,0 A280 оптических единиц на см (длина оптического пути).
Для защиты хроматографических колонок (например, содержащих AEX смолу, содержащую кватернизированный полиэтилениминовый лиганд или диметиламинопропильный лиганд) от забивания из-за частиц вещества в подвижной фазе можно использовать предколоночный фильтр. В одном варианте осуществления изобретения фильтр представляет собой мембрану из полиэфирсульфона. Также можно использовать постколончный фильтр, чтобы отфильтровать любый твердые частицы из проточной жидкости. В одном варианте осуществления изобретения фильтр имеет размер пор 0,2 или 0,5 мм.
Присутстви варианта, содержащего сульфатированный тирозин, можно подтвердить, например, масс-спектрографическим анализом проточных фракций. Сульфатированные варианты будут иметь бóльшую массу, чем у несульфатированных вариантов. Например, в одном варианте осуществления изобретения сульфатированный вариант на около 80 Да тяжелее, чем варианты с отсутствием сульфатирования. В одном варианте осуществления изобретения сульфатирование является резистентным к расщеплению фосфатазой, и сульфатированный пептид имеет другой паттерн фрагментации, вызванный диссоциацией в результате переноса электрона (ETD), по сравнению с фосфорилированными пептидами.
В одном варианте осуществления изобретения композиция, включающая антитела (например, Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9; предпочтительно Ab6) с отсутствием сульфатирования тирозина относится к композиции, в которой отсутствует определяемое сульфатирование тирозина (например, на CDR-L1). Композиция, включающая неопределяемые уровни сульфатирования тирозина (например, на CDR-L1), включает уровень, который не может наблюдаться масс-спектрометрическим анализом композиции. Например, в одном варианте осуществления изобретения масс-спектрометрический анализ композиции осуществляют путем измерения интактной и восстановленной массы и картирования восстановленных пептидов иммуноглобулиновых пептидов композиции. В одном варианте осуществления изобретения картирование восстановленных пептидов включает денатурацию и восстановление дисульфидных связей в молекуле антитела и алкилирование свободных цистеинов, с последующим ферментативным расщеплением (например, с использованием LysC, Трипсина или GluC). Ферментативно расщепленные пептиды анализировали масс-спектрометрией. В одном варианте осуществления изобретения ʺнеопределяемыйʺ уровень относится к меньше чем около 0,5% (меньше чем около 0,4, 0,3, 0,2, 0,1%) видов с сульфатированным тирозином (например, на CDR-L1) по сравнению с немодифицированными молекулами в композиции.
Молекулярную массу полипептида можно рассчитать, например, на основе известной массы аминокислот (модифицированных или немодифицированных/сульфатированных или несульфатированных) и известных модификаций (например, окисления, дезамидирования, гликозилирования, C- и N-концевой модификации). Молекулярную массу можно измерить масс-спектрометрическим анализом, например, в сочетании с жидкостной хроматографией. В одном варианте осуществления изобретения масс-спектрометрия представляет собой квадрупольную масс-спектрометрию по времени пролета (Q-TOF) или масс-спектрометрию Orbitrap.
Термин "хроматография" относится к процессу, посредством которого растворенное вещество, представляющее интерес, например, вещество в композиции, отделяют от других веществ в композиции путем контактирования веществ со смолой, которая действует как адсорбент. Адсорбент представляет собой вещество, которое адсорбирует или удерживает вещество более или менее сильно, например, из-за свойств растворенного вещества, таких как pI, гидрофобность, размер и структура, в определенных буферных условиях процесса. Хроматографию можно осуществлять традиционными способами перколяции композиции через слой хроматографической смолы, например, через колонку, содержащую смолу. Периодический режим хроматографической очистки включает приготовление суспензии смолы и контактирование композиции, содержащей антитело или фрагмент, с суспензией для адсорбции вещества, которое должно быть отделено от смолы. Раствор, включающий вещество, не связанное со смолой, отделяют от суспензии, например, давая суспензии осадиться и удаляя супернатант, и несвязанное вещество можно сохранить или выбросить. Суспензию необязательно подвергают одной или нескольким стадиям промывки. Если желательно, можно обеспечить контактирование суспензии с подходящим элюирующим буфером для десорбции связанных со смолой веществ из смолы. Десорбированное вещество можно сохранить или выбросить. В одном варианте осуществления изобретения варианты антител с сульфатированным тирозином в композиции связаны с анионообменной смолой, в то время как варианты с несульфатированным тирозином не связываются существенным образом со смолой.
В одном варианте осуществления изобретения антитело или его антигенсвязывающий фрагмент очищают хроматографией с протеином А или протеином G. Протеин-G и протеин-A являются бактериальными белками из Группы G Streptococci и Staphylococcus aureus, соответственно. Аффинность протеина G и протеина A к Fc области IgG-типа антител служит основой для очистки IgG, фрагментов IgG, содержащих Fc область, и подклассов IgG. Протеин-А или протеин-G могут быть связаны с твердой фазой, такой как сефароза, которая может использоваться для хроматографии с протеином-А или протеином-G. Настоящее изобретение включает способы для получения композиции, включающей антитело или его антигенсвязывающий фрагмент с отсутствием определяемых уровней сульфатированного варианта тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим AEX-хроматографию в проточном режиме и на протеине-A и/или протеине-G.
В одном варианте осуществления изобретения антитело или его антигенсвязывающий фрагмент очищают мультимодальной хроматографией (смешанный режим). Мультимодальная или смешанная хроматография белков основана на смолах, которые были функционализированы лигандами, способными к множественным способам взаимодействия, например, на основе ионного обмена, гидроксиапатита, аффинности, исключения по размеру и/или гидрофобным взаимодействиям. Настоящее изобретение включает способы для получения композиции, включающей антитело или его антигенсвязывающий фрагмент, в которой отсутствуют обнаруживаемые уровни сульфатированных вариантов тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления сульфатированных вариантов тирозина способом, включающим AEX-хроматографию в проточном режиме и хроматографию в смешанном режиме.
В одном варианте осуществления изобретения антитело или его антигенсвязывающий фрагмент очищают хроматографией с протеином-L. Протеин L представляет собой белок Peptostreptococcus magnus, который связывает иммуноглобулины через легкую цепь иммуноглобулина. Протеин L связывается с представителями всех классов антител, включая IgG, IgM, IgA, IgE и IgD. Рекомбинантный протеин L связывается с вариабельной областью легкой цепи каппа иммуноглобулинов и фрагментов иммуноглобулинов. Протеин L связывается с тремя из четырех подтипов легкой цепи каппа у людей (1, 3 и 4) и каппа 1 у мышей. Настоящее изобретение включает способы для получения композиции, включающей антитело или его антигенсвязывающий фрагмент с отсутствием определяемых уровней сульфатированного варианта тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим AEX-хроматографию в проточном режиме и хроматографию с протеином-L.
В одном варианте осуществления изобретения антитело или его антигенсвязывающий фрагмент очищают хроматографией гидрофобных взаимодействий (HIC). HIC разделяет белки с различиями в гидрофобности. Разделение основано на обратимом взаимодействии между белком и гидрофобной поверхностью хроматографической среды. Настоящее изобретение включает способы для получения композиции, включающей антитело или его антигенсвязывающий фрагмент с отсутствием определяемых уровней сульфатированного варианта тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим AEX-хроматографию в проточном режиме и HIC.
В одном варианте осуществления изобретения антитело или его антигенсвязывающий фрагмент очищают эксклюзионной хроматографией (SEC). SEC разделяет белки с разным размером молекул. Настоящее изобретение включает способы получения композиции, включающей антитело или его антигенсвязывающий фрагмент, в которой отсутствуют обнаруживаемые уровни сульфатированного варианта тирозина, или очистку антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим AEX в проточном режиме и SEC хроматографию.
В одном варианте осуществления изобретения антитело или его антигенсвязывающий фрагмент подвергают вирусной инактивации. Например, в одном варианте осуществления изобретения вирусную инактивацию осуществляют путем pH обработки композиций, включающих антитело или его антигенсвязывающий фрагмент. В частности, для очистки от вирусов можно использовать предельные значения рН для непосредственного воздействия на композицию. Например, рН обработка в одном варианте осуществления изобретения представляет собой обработку с низким рН (например, рН 3,0-3,6). В одном варианте осуществления изобретения антитела или антигенсвязывающие фрагменты подвергают обработке при высоком уровне pH. В одном варианте осуществления изобретения вирусную инактивацию композиций, включающих антитело или его антигенсвязывающий фрагмент, осуществляют с использованием растворителя или детергента. Настоящее изобретение включает способы для получения композиции, включающей антитело или его антигенсвязывающий фрагмент с отсутствием определяемых уровней сульфатированного варианта тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим AEX-хроматографию в проточном режиме и вирусную инактивацию.
"Ионный обмен" разделяет молекулы на основе различий в их суммарном поверхностном заряде. Молекулы значительно различаются по своим зарядным свойствам и будут проявлять различные степени взаимодействия с заряженными хроматографическими смолами в соответствии с различиями в их общем заряде, плотности заряда и распределении поверхностного заряда. В одном варианте осуществления изобретения антитело или его антигенсвязывающий фрагмент очищают ионообменной хроматографией. "Ионообменная хроматография" включает катионообменную, анионообменную хроматографию и хроматографию в смешанном режиме.
Фраза "ионообменная" смола относится к твердой фазе, которая отрицательно заряжена (то есть катионообменная) или положительно заряжена (то есть анионообменная).
В одном варианте осуществления изобретения антитело или его антигенсвязывающий фрагмент очищают катионообменной хроматографией. "Катионообменная" смола относится к твердой фазе, которая отрицательно заряжена и которая имеет свободные катионы для обмена с катионами в водном растворе, проходящем над твердой фазой или через нее. Можно использовать любой отрицательно заряженный лиганд, присоединенный к твердой фазе, подходящий для образования катионообменной смолы. Катионообменные материалы включают, но не ограничиваются этим, материалы, имеющие лиганд: сульфопропил (SP) -CH2-CH2-CH2-SO3-, метилсульфонат (S) -CH2-SO3- или карбоксиметил (CM) -CH2-COO-. Настоящее изобретение включает способы для получения композиции, включающей антитело или его антигенсвязывающий фрагмент с отсутствием определяемых уровней сульфатированного варианта тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим AEX хроматографию в проточном режиме и катионообменную хроматографию.
В одном варианте осуществления изобретения антитело или его антигенсвязывающий фрагмент очищают анионообменной хроматографией. "Анионообменная" смола относится к твердой фазе, которая положительно заряжена, таким образом, к ней присоединены один или несколько положительно заряженных лигандов. Можно использовать любой положительно заряженный лиганд, присоединенный к твердой фазе, подходящий для образования анионообменной смолы. Анионообменные материалы включают, но не ограничиваются этим, материалы, имеющие лиганд: четвертичный аммоний (Q) -CH2-N+-(CH3)3; диэтиламиноэтил (DEAE) -CH2-CH2-N+-(CH2-CH3)2; или диэтиламинопропил (ANX) -CH2-CHOH-CH2-N+-(CH2-CH3)2. Колонка GoPure D 50 мкм содержит диметиламинопропильную функциональную группу. Настоящее изобретение включает способы для получения композиции, включающей антитело или его антигенсвязывающий фрагмент с отсутствием определяемых уровней сульфатированного варианта тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим AEX хроматографию в проточном режиме и AEX хроматографию (в режиме связывания/элюирования).
Термин "твердая фаза" или "неподвижная фаза" используется для обозначения любой неводной матрицы, к которой может присоединиться один или несколько лигандов (например, анионообменных лигандов или катионообменных лигандов), или, альтернативно, в случае эксклюзионной хроматографии это может относиться к гелевой структуре смолы. Подвижная фаза представляет собой жидкость, например водное вещество, которое переносит антитело или антигенсвязывающий фрагмент над твердой фазой для хроматографической очистки. Подвижная фаза может включать загрузочный буфер, который вводят в колонку. Примеры материалов, которые можно использовать для образования твердой фазы, включают полисахариды (такие как агароза и целлюлоза) и другие механически стабильные матрицы, такие как диоксид кремния (например, стекло с контролируемыми порами), поли(стиролдивинил) бензол, полиакриламид, керамические частицы и производные любого из этих веществ.
"Уравновешивающий" буфер или раствор используют для регулирования рН и проводимости хроматографической смолы перед загрузкой смесью, содержащей антитело или антигенсвязывающий фрагмент, для очистки. Подходящие буферы или растворы, которые можно использовать для этой цели, хорошо известны в данной области техники, например, буферы, описанные выше, и включают любой буфер при рН, который совместим с выбранной смолой, используемой на стадии хроматографии для очистки представляющего интерес белка.
"Загрузочный" буфер или раствор используют для загрузки смеси, содержащей антитело или антигенсвязывающий фрагмент, на очищающую смолу (например, анионообменную смолу или катионообменную смолу). Любой подходящий раствор можно использовать в качестве загрузочного буфера. В одном варианте осуществления изобретения загрузочный буфер получают из забуференной смеси с предыдущей стадии очистки, такой как элюирующий буфер.
Термин "промывочный" буфер или раствор означает композицию, используемую для элюирования одной или нескольких примесей из очищающей смолы (например, анионообменной смолы или катионообменной смолы) перед элюированием антитела или антигенсвязывающего фрагмента. Термин "промывка" описывает пропускание соответствующей композиции через хроматографическую смолу или над ней. В одном варианте осуществления изобретения промывка является изократической. В условиях изократической промывки подвижная фаза хроматографии остается по существу одинаковой.
Хотя антитела и антигенсвязывающие фрагменты с сульфатированны вариантом тирозина являются загрязняющими веществами, настоящее изобретение включает композиции, содержащие такие варианты, например, связанные с AEX хроматографической смолой или несвязанные в отсутствие вариантов без сульфатированного тирозина. Несвязанные варианты могут быть получены путем элюции из AEX колонки после удаления из антител и фрагментов, не содержащих сульфированный тирозин.
"Элюирующий" буфер диссоциирует молекулу (например, антитело или его антигенсвязывающий фрагмент), связанную с хроматографической смолой.
Предыдущие этапы обработки
Антитела и антигенсвязывающие фрагменты, которые необходимо очистить от загрязняющих вариантов с сульфатированным тирозином, можно получить путем экспрессии в клетке-хозяине. Например, способ по настоящему изобретению включает, в одном варианте осуществления, до удаления таких вариантов экспрессию тяжелых и/или легких цепей иммуноглобулинов в клетке-хозяине в культуральной среде в условиях, благоприятных для такой экспрессии, и выделения антител или антигенсвязывающих фрагментов из клетки-хозяина и/или культуральной среды. Настоящее изобретение включает способы получения композиции, включающей антитело или его антигенсвязывающий фрагмент, в которой отсутствуют определяемые уровни сульфатированных вариантов тирозина, или очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов, включающих сульфатированный тирозин, способом, включающим экспрессию в клетке-хозяине и AEX хроматографию в проточном режиме.
В объем настоящего изобретения входят способы получения композиции, включающей антитела или антигенсвязывающие фрагменты, в которых отсутствует сульфатирование тирозина (например, на их CDR-L1), включающие (i)введение полинуклеотида, кодирующего легкие и/или тяжелые цепи иммуноглобулинов указанных антител или фрагментов, в клетку-хозяин (например, клетку СНО) и (ii) культивирование клетки-хозяина в условиях, благоприятных для экспрессии цепей иммуноглобулина в клетке, например, где антитело или антигенсвязывающий фрагмент, имеющее цепь (цепи) иммуноглобулина, секретируется из клетки-хозяина в культуральную среду, и (iii) выделение полипептида(полипептидов) цепи иммуноглобулина из клетки-хозяина и/или культуральной среды способом, который включает анионообменную хроматографию в проточном режиме, как обсуждается выше.
Например, антитела или фрагменты могут высвобождаться из клетки-хозяина путем лизиса, например, такими способами, как измельчение/истирание (например, со стеклянными шариками), лизис клеток с использованием пресса Френча, ферментативное расщепление или обработка ультразвуком. Лизированные клетки, включая растворимые и нерастворимые материалы из них, образуют клеточный лизат. Настоящее изобретение включает способы для получения антитела или его антигенсвязывающего фрагмента, в котором отсутствует сульфатированный вариант тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим лизис клеток и AEX хроматографию в проточном режиме.
В одном варианте осуществления изобретения антитела или антигенсвязывающие фрагменты очищают способами, включающими центрифугирование. Центрифугирование клеточного лизата или другой суспензии удаляет большинство твердых частиц, таких как клеточный дебрис, из водной фракции, содержащей антитело или фрагмент. Например, в одном варианте осуществления изобретения центрифугирование осуществляют (например, на клеточном лизате, включая удаление твердой фракции лизата) при около 40000-50000 g в течение 15-30 минут. В одном варианте осуществления изобретения клетки удаляют из жидкой культуральной среды центрифугированием. Например, центрифугирование с использованием гравитационной силы в диапазоне от около 8000×g до около 15000×g (например, около 8000, 9000, 10000, 11000, 12000, 13000, 14000 или 15000), например, характеризуется Q/SIGMA отношением в пределах от около 0,9×10-8 до 2,8×109. В одном варианте осуществления изобретения жидкость из центрифуги подвергают глубинному фильтрованию (например, с размером пор от 0,1 до около 0,2 мкм). Настоящее изобретение включает способы для получения антитела или его антигенсвязывающего фрагмента, в котором отсутствует сульфатированный вариант тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим центрифугирование и AEX хроматографию в проточном режиме.
В одном варианте осуществления изобретения тяжелые и легкие цепи иммуноглобулина экспрессируются в клетке-хозяине, слитой с сигнальной последовательностью секреции, и секретируются из клеток-хозяев в культуральную среду клеток-хозяев.
В одном варианте осуществления изобретения антитела или антигенсвязывающие фрагменты очищают фильтрацией (например, до или после AEX хроматографической очистки). Например, в одном варианте осуществления изобретения водную композицию, включающую антитело или антигенсвязывающий фрагмент, фильтруют для удаления материала в виде твердых частиц, например, через фильтр, имеющий размер пор около 1 мкм, 0,45 мкм или 0,22 мкм. В одном варианте осуществления изобретения фильтр изготовлен из ацетата целлюлозы или поливинилиденфторида (PVDF). Настоящее изобретение включает способы для получения антитела или его антигенсвязывающего фрагмента, в котором отсутствует сульфатированный вариант тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим AEX хроматографию в проточном режиме и фильтрацию.
В одном варианте осуществления изобретения антитела или антигенсвязывающие фрагменты очищают фракционным осаждением. Повышенная концентрация соли может улучшить гидрофобное взаимодействие между белками и привести к селективному осаждению. В одном варианте осуществления изобретения водную композицию, включающую антитело или фрагмент, осаждают в присутствии сульфата аммония, декстрансульфата, поливинилпирролидина, полиэтиленгликоля (PEG; например, PEG4000), ацетона, полиэтиленимина, сульфата протамина, сульфата стрептомицина или каприловой кислоты. Настоящее изобретение включает способы для получения антитела или его антигенсвязывающего фрагмента, в котором отсутствует сульфатированный вариант тирозина, или для очистки антитела или его антигенсвязывающего фрагмента для удаления вариантов с сульфатированным тирозином способом, включающим AEX хроматографию в проточном режиме и фракционное осаждение.
В одном варианте осуществления изобретения клетка-хозяин, в которой экспрессируется цепь иммуноглобулина, представляет собой клетку млекопитающего, такую как клетка яичника китайского хомячка (СНО), клетка миеломы мыши, клетка PER, клетка гибридомы или грибковая или дрожжевая клетка, например, Pichia, такая как Pichia pastoris, или Saccharomyces cerevisiae. В одном варианте осуществления изобретения клетка-хозяин, например, клетка СНО, не содержит глутаминсинтазу.
В одном варианте осуществления изобретения полинуклеотид(полинуклеотиды), кодирующий тяжелую и/или легкую цепь иммуноглобулина, функционально связан с одной или несколькими последовательностями контроля экспрессии, такими как промотор. Например, иммуноглобулин находится в векторе экспрессии. Для достижения высоких уровней экспрессии антител или антигенсвязывающих фрагментов можно использовать сильный промотор/энхансер, такой как цитомегаловирусный (CMV) промотор и/или промотор фактора элонгации альфа (EF1α), для стимуляции экспрессии тяжелой цепи и/или легкой цепи иммуноглобулина.
В одном варианте осуществления изобретения интронная последовательность в 5'-нетранслируемой области включена после промотора/энхансера для увеличения экспорта транскрибированной мРНК в цитоплазму из ядра, и одна или несколько сигнальных последовательностей 3'-полиаденилирования включены для максимизации уровней мРНК. В одном варианте осуществления изобретения сигнальная последовательность полиаденилирования представляет собой сигнальную последовательность позднего или раннего полиаденилирования SV40 или последовательность полиаденилирования бычьего гормона роста. В одном варианте осуществления изобретения создана консенсусная последовательность Козака путем помещения GCC GCC(A/G)CC (SEQ ID NO: 69) непосредственно перед первым кодоном инициации трансляции для усиления инициации трансляции. В одном варианте осуществления изобретения сигнальная пептидная последовательность помещена непосредственно перед цепью иммуноглобулина для направления секреции антитела или фрагмента.
Условия культивирования клеток можно контролировать и корректировать по мере необходимости. Например, такие условия, как рН, количество клеток, жизнеспособность клеток и температура, можно контролировать и регулировать. В одном варианте осуществления изобретения температуру клеточной культуры доводят, например, от 37 до 30-35°С через 48 часов после инокуляции. Растворенный кислород, в одном варианте осуществления изобретения, контролируют и/или доводят до заданного значения, такого как 20-50%. В одном варианте осуществления изобретения растворенный СО2 контролируют и/или доводят, например, до не более чем около 120-150 мм рт.ст. В одном варианте осуществления изобретения осмоляльность контролируют и/или доводят, например, до около 270-330 мОсм/кг.
Антитела
Настоящее изобретение обеспечивает композиции, включающие антитела и их антигенсвязывающие фрагменты, в которых отсутствуют определяемые уровни сульфатированного тирозина, а также способы для выделения композиций, включающих такие антитела и фрагменты. Например, в одном варианте осуществления изобретения антитело или фрагмент включает сульфатированный тирозин и связывается с антигеном, выбранным из: PD1, CD27, LAG3, CTLA4, BTLA, TIM3, ICOS, B7-H3, B7-H4, CD137, GITR, PD-L1, PD-L2, ILT1, ILT2 CEACAM1, CEACAM5, TIM3, TIGIT, VISTA, ILT3, ILT4, ILT5, ILT6, ILT7, ILT8, CD40, OX40, CD137, KIR2DL1, KIR2DL2, KIR2DL3, KIR2DL4, KIR2DL5A, KIR2DL5B, KIR3DL1, KIR3DL2, KIR3DL3, NKG2A, NKG2C, NKG2E, IL-10, IL-17 или TSLP.
Термин ʺLAG3ʺ, в отношении полипептида, с которым связываются антитела и антигенсвязывающие фрагменты по настоящему изобретению, относится к LAG3 человека и обезьяны cynomolgous, например, Macaca fascicularis или Macaca mulatta, а также их фрагментам, таким как их зрелый фрагмент, не содержащий сигнального пептида.
Примеры цепей иммуноглобулинов анти-LAG3 антител (например, Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9, которые раскрыты в WO2016028672) с отсутствием сульфатирования тирозина включают примеры, приведенные ниже. Например, где антитело или фрагмент включает одну или несколько из CDR и/или цепей иммуноглобулина, представленных ниже. В одном варианте осуществления изобретения загрязняющее антитело или антигенсвязывающий фрагмент включает CDR-L1, имеющую аминокислотную последовательность KASQSLDYEGDSDMN (SEQ ID NO: 38), где Y (выделен жирным шрифтом и подчеркнут) является сульфатированным.
В одном варианте осуществления изобретения анти-LAG3 антитело или антигенсвязывающий фрагмент включает тяжелые цепи иммуноглобулина и/или легкие цепи иммуноглобулина 4A10; VH и/или VL цепи или CDRs легкой цепи и/или CDRs тяжелой цепи (например, CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2 и CDR-H3 4A10).
В одном варианте осуществления изобретения, для любого из Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9, любой N-концевой глутамин в тяжелой цепи преобразован в пироглутамат и/или любой C-концевой лизин в тяжелой цепи удален.
4A10- VH последовательность
ATGAAATGCAGCTGGGTCATCTTCTTCCTGATGGCAGTGGTTATAGGAATCAATTCAGAGGTTCAGCTGCTCCAGTCTGGGGCAGAACTTGTGAGGTCAGGGGCCTCAGTCAAGTTGTCCTGCACAGCCTCTGGCTTCAACATTGAAGACTACTATATGCACTGGATGAAACAGAGGCCTGAACAGGGCCTGGAGTGGATTGGATGGATTGATCCTGTGAATGGTGATACTGAATATGCCCCGAAGTTCCAGGGCAAGGCCACTATGACTGCAGACACATCCTCCAACACAGCCTACCTACACCTCAACAGCCTGACATCTGAGGACACTGCCGTCTATTACTGTAATTTCTATGATGGTTACCTCTTTGCTTTCTGGGGCCAAGGGACCCTGGTCACTGTCTCTGCA
(SEQ ID NO: 1; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
MKCSWVIFFLMAVVIGINSEVQLLQSGAELVRSGASVKLSCTASGFNIEDYYMHWMKQRPEQGLEWIGWIDPVNGDTEYAPKFQGKATMTADTSSNTAYLHLNSLTSEDTAVYYCNFYDGYLFAFWGQGTLVTVSA
(SEQ ID NO: 2; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
CDR-H1: GFNIEDYYMH (SEQ ID NO: 3)
CDR-H2: WIDPVNGDTEYAPKFQG (SEQ ID NO: 4)
CDR-H3: YDGYLFAF (SEQ ID NO: 5)
4A10- VL последовательность
ATGAGGTGCCTAGCTGAGTTCCTGGGGCTGCTTGTGCTCTGGATCCCTGGAGCCATTGGGGATATTGTGCTGACTCAGGCTGCACCCTCTGTACCTGTCACTCCTGGAGAGTCAGTGTCCATCTCCTGCAGGTCTAGTAAGAGTCTCCTGCATAGTGATGGCAACACTTATCTGTATTGGCTCCTGCAGAGGCCAGGCCAGTCTCCTCAGCTCCTGATATATCGGATGTCCAACCTTGCCTCAGGGGTCCCAGACAGGTTCAGCGGCAGTGGGTCAGGAACTGTTTTCACACTGAGAATCAGCAGACTGGAGGCTGAGGATGTGGGTATTTATTACTGTATGCAACATCTAGAATATCCTTTCACGTTTGGAGGGGGGACCAAGCTGGAAATAAAA
(SEQ ID NO: 6; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
MRCLAEFLGLLVLWIPGAIGDIVLTQAAPSVPVTPGESVSISCRSSKSLLHSDGNTYLYWLLQRPGQSPQLLIYRMSNLASGVPDRFSGSGSGTVFTLRISRLEAEDVGIYYCMQHLEYPFTFGGGTKLEIK
(SEQ ID NO: 7; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
CDR-L1: RSSKSLLHSDGNTYLY (SEQ ID NO: 8)
CDR-L2: YRMSNLAS (SEQ ID NO: 9)
CDR-L3: MQHLEYPFT (SEQ ID NO: 10)
В одном варианте осуществления изобретения анти-LAG3 антитело или антигенсвязывающий фрагмент включает тяжелые цепи иммуноглобулинов и/или легкие цепи иммуноглобулинов 19E8; VH и/или VL цепи или CDRs легкой цепи и/или CDRs тяжелой цепи (например, CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2 и CDR-H3 19E8):
19E8- VH последовательность
ATGGGATGGAGCTGGATCTTTCTTTTCCTCCTGTCAGGAACTGCAGGTGTCCGTTGCCAGATCCGACTGCAGCAGTCTGGACCTGAGCTGGTGAAGCCTGGGGCTTCAGTGAAGATATCCTGCAAGGCTTCTGGGTCCTCCTTCACTGACTACTATATAAACTGGGTGAAGCAGAAGCCTGGACAGGGACTTGAGTGGATTGGATGGATTTATCCTGGAAGCGGTAATTCTATCTACAATGAGAACTTCAAGGCCAAGGCCACATTGACTGTAGACACATCCTCCAGCACAGCCTACATGCATCTCAGCAGCCTGACATCTGAGGACACTGCTGTCTATTTCTGTGCAAGAGAGGCTGATTACGACGATGCTTTGGACTACTGGGGTCAAGGAACCTCGGTCACCGTCTCCTCA
(SEQ ID NO: 11; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
MGWSWIFLFLLSGTAGVRCQIRLQQSGPELVKPGASVKISCKASGSSFTDYYINWVKQKPGQGLEWIGWIYPGSGNSIYNENFKAKATLTVDTSSSTAYMHLSSLTSEDTAVYFCAREADYDDALDYWGQGTSVTVSS
(SEQ ID NO: 12; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
CDR-H1: GSSFTDYYIN (SEQ ID NO: 13)
CDR-H2: WIYPGSGNSIYNENFKA (SEQ ID NO: 14)
CDR-H3: EADYDDALDY (SEQ ID NO: 15)
19E8- VL последовательность
ATGGTATCCACACCTCAGTTCCTTGTATTTTTGCTTTTCTGGATTCCAGCCTCCAGAGGTCACATCTTGCTGACTCAGTCTCCAGCCATTCTGTCTGTGAGTCCAGGAGAAAGAGTCAGTTTCTCCTGCAGGGCCAGTCAGAGCATTGGCACAAGCATACACTGGTATCAGCAAAGAACAAATGGTTCTCCAAGGCTTCTCATAAAGTATGCTTCTGAGTCTATCTCTGGGATCCCTTCCAGGTTTAGTGGCAGTGGATCAGGGACAGATTTTACTCTTAGCATCAACAGTGTGGAGTCAGAAGATATTGCAGATTATTACTGTCAACAAAGTAATAGCTGGCCAACGTACACGTTCGGAGGGGGGACCAAGCTGGAAATAAAA
(SEQ ID NO: 16; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
MVSTPQFLVFLLFWIPASRGHILLTQSPAILSVSPGERVSFSCRASQSIGTSIHWYQQRTNG SPRLLIKYASESISGIPSRFSGSGSGTDFTLSINSVESEDIADYYCQQSNS WPTYTFGGGTKLEIK
(SEQ ID NO: 17; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
CDR-L1: RASQSIGTSIH (SEQ ID NO: 18)
CDR-L2: YASESIS (SEQ ID NO: 19)
CDR-L3: QQSNSWPTYT (SEQ ID NO: 20)
В одном варианте осуществления изобретения анти-LAG3 антитело или антигенсвязывающий фрагмент включает тяжелые цепи иммуноглобулинов и/или легкие цепи иммуноглобулинов 11C9; VH и/или VL цепи или CDRs легкой цепи и/или CDRs тяжелой цепи (например, CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2 и CDR-H3 11C9):
11C9- VH последовательность
ATGAGATGGAGCTGTATCATCCTCTTCTTGGTAGCAACAGCTACAGGTGTCAACTCCCAGGTCCAACTGCAGCAGCCTGGGGCTGAGCTTGTGATGCCTGGGGCTTCAGCGAAGATGTCCTGCAAGGCTTCTGGCTACACACTCACTGACTACTGGATGCACTGGGTGAAGCAGAGGCCTGGACAAGGCCTTGAGTGGATCGGAGCGATTGATATTTCTGATAGTTATTCTAGCTACAATCAAAAGTTCAAGGGCAAGGCCACATTGACTGTAGACGAATCCTCCAGCACAGCCTACATGCAGCTCACCAGCCTGACATCTGAGGACTCTGCGGTCTATTACTGTGCAAGATCCCCTTTCTACAATAGTAGAGGGGGGAACTACTTTGACTACTGGGGCCAAGGCACCACTCTCACAGTCTCCTCA
(SEQ ID NO: 21; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
MRWSCIILFLVATATGVNSQVQLQQPGAELVMPGASAKMSCKASGYTLTDYW MHWVKQRPGQGLEWIGAIDISDSYSSYNQKFKGKATLTVDESSSTAYMQLTSLTSEDSAVYYCARSPFYNSRGGNYFDYWGQGTTLTVSS
(SEQ ID NO: 22; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
CDR-H1: GYTLTDYWMH (SEQ ID NO: 23)
CDR-H2: AIDISDSYSSYNQKFKG (SEQ ID NO: 24)
CDR-H3: SPFYNSRGGNYFDY (SEQ ID NO: 25)
11C9- VL последовательность
ATGATGTCCTCTGCTCAGTTCCTTGGTCTCCTGTTGCTCTGTTTTCAAGGTACCAGATGTGATATCCAGATGACACAGACTACATCCTCCCTGTCTGCCTCTCTGGGAGACAGAGTCACCATCAGTTGCAGGGCAAGTCAGGACATTAGCAATTATTTAAACTGGTATCAGCAGAAACCAGATGGAACTGTTAAACTCCTGATCTACTACACATCAAGATTACACTCAGGAGTCCCATCAAGGTTCAGTGGCAGTGGGTCTGGAACAGATTATTCTCTCACCATTAGCAACCTGGAGCAAGAAGATATTGCCACTTACTTTTGCCAACAGGGTGATACGCTTCCTCCGTGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAA
(SEQ ID NO: 26; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
MMSSAQFLGLLLLCFQGTRCDIQMTQTTSSLSASLGDRVTISCRASQDISNYLNWYQQKPDGTVKLLIYYTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGDTLPPWTFGGGTKLEIK
(SEQ ID NO: 27; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
CDR-L1: RASQDISNYLN (SEQ ID NO: 28)
CDR-L2: YTSRLHS (SEQ ID NO: 29)
CDR-L3: QQGDTLPPWT (SEQ ID NO: 30)
В одном варианте осуществления изобретения анти-LAG3 антитело или антигенсвязывающий фрагмент включает тяжелые цепи иммуноглобулинов и/или легкие цепи иммуноглобулинов 22D2; VH и/или VL цепи или CDRs легкой цепи и/или CDRs тяжелой цепи (например, CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2 и CDR-H3 22D2):
22D2- VH последовательность
ATGGGATGGACCTGGATCTTTCTCTTCTTCCTGTCAGGAACTGCAGGTGTCCTCTCTGAGGTCCTGCTGCTACAGTCTGGACCTGAACTGGTGAAGCCTGGGACTTCAGTGAAAATCCCCTGCAAGGCTTCTGGATACACATTCACTGACTACAACGTGGACTGGGTGAAGCAGCGCCATGGAAAGGGCCTTGAGTGGATTGGAGATATTAATCCAAACAATGGTGGTACTATCTACAGTCAGAAATTCAAGGGCAAGGCCACATTGACTGTTGACAAGTCCTCCAGCACAGCCTTCATGGAGCTCCGCAGCCTGACATCTGAGGACACTGCAGTCTATTTCTGTGCAAGGAACTATAGGTGGTTTGGTGCTATGGACCACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCAGCCAAAACAACAGCCCCATCGGTCTATCCACTG
(SEQ ID NO: 31; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
MGWTWIFLFFLSGTAGVLSEVLLLQSGPELVKPGTSVKIPCKASGYTFTDYNVDWVKQRHGKGLEWIGDINPN NGGTIYSQKFKGKATLTVDKSSSTAFMELRSLTSEDTAVYFCARNYRWFGAMDHWGQGTSVTVSS
(SEQ ID NO: 32; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
CDR-H1: DYNVD (SEQ ID NO: 33)
CDR-H2: DINPNNGGTIYSQKFKG (SEQ ID NO: 34)
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
22D2- VL последовательность
ATGGAGACAGACACAATCCTGCTATGGGTGCTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACATTGTGTTGACCCAATCTCCAGCTTCTTTGGCTGTGTCTCCAGGGCAGAGGGCCACCATTTCCTGCAAGGCCAGTCAAAGTCTTGATTATGAAGGTGATAGTGATATGAATTGGTACCAACAGAAACCAGGACAGCCACCCAGACTCCTCATCTCTGGTGCATCCAATCTAGAGTCTGGGATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTGTTAACATCCATCCTGTGGAGGAGGAGGATGCTGCAACCTATTACTGTCAGCAAAGTACTGAGGATCCTCGGACGTTCGGTGGAGGCACCAAGCTGGAAATCAAACGGGCTGATGCTGCACCAACTGTATCCATCTTCCCACCATCCAGTGAGCAGTTAACATCTGGAGGTGCCTCAGTCGTGTGCTTCTTGAACAACTTCTACCCCAAAGACATCAATGTCAAGTGGAAGATTGATGGCAGTGAACGACAAAATGGCG
(SEQ ID NO: 36; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
METDTILLWVLLLWVPGSTGDIVLTQSPASLAVSPGQRATISCKASQSLDYEGDSDMNWYQQKPGQPPRLLISGASNLESGIPARFSGSGSGTDFTVNIHPVEEEDAATYYCQQSTEDPRTFGGGTKLEIK
(SEQ ID NO: 37; где CDR подчеркнуты, и где сигнальная последовательность выделена жирным шрифтом)
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38)
CDR-L2: GASNLES (SEQ ID NO: 39)
CDR-L3: QQSTEDPRT (SEQ ID NO: 40).
В одном варианте осуществления изобретения анти-LAG3 антитело или антигенсвязывающий фрагмент включает тяжелые цепи иммуноглобулинов и/или легкие цепи иммуноглобулинов Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9; VH и/или VL цепи или CDRs легкой цепи и/или CDRs тяжелой цепи (например, CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2 и CDR-H3 Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 или Ab9):
Ab1: гуманизированная легкая цепь 45AGX Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 (VL3)) Каппа (PX) (или ее вариабельный домен) и гуманизированная тяжелая цепь 53AHH Гуманизированного x[LAG3_H] mAb (LB145.22D2.E1.D1 VH6) IgG1/Каппа (PX) (или ее вариабельный домен); например, включающие:
легкую цепь иммуноглобулина, включающую аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
(SEQ ID NO: 41); и
тяжелую цепь иммуноглобулина, включающую аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNNGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
(SEQ ID NO: 42); или
вариабельный домен легкой цепи иммуноглобулина, включающий аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIK
(аминокислоты 1-111 SEQ ID NO: 41 (CDR подчеркнуты)); и
вариабельный домен тяжелой цепи иммуноглобулина, включающий аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNNGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSS
(аминокислоты 1-119 SEQ ID NO: 42 (CDR подчеркнуты));
или включающие CDR:
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2: GASNLES (SEQ ID NO: 39);
CDR-L3: QQSTEDPRT (SEQ ID NO: 40);
CDR-H1: DYNVD (SEQ ID NO: 33);
CDR-H2: DINPNNGGTIYAQKFQE (SEQ ID NO: 59); и
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
Ab2: гуманизированная легкая цепь 45AGX Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 (VL3)) Каппа (PX) (или ее вариабельный домен) и гуманизированная тяжелая цепь 56AHH Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 VH6 N55S) IgG1/Каппа (PX) (или ее вариабельный домен); например: включающие:
легкую цепь иммуноглобулина, включающую аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
(SEQ ID NO: 43); и
тяжелую цепь иммуноглобулина, включающую аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNSGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
(SEQ ID NO: 44); или
вариабельный домен легкой цепи иммуноглобулина, включающий аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIK
(аминокислоты 1-111 SEQ ID NO: 43 (CDR подчеркнуты)); и
вариабельный домен тяжелой цепи иммуноглобулина, включающий аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNSGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSS
(аминокислоты 1-119 SEQ ID NO: 44 (CDR подчеркнуты));
или включающие CDR:
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2: GASNLES (SEQ ID NO: 39);
CDR-L3: QQSTEDPRT (SEQ ID NO: 40);
CDR-H1: DYNVD (SEQ ID NO: 33);
CDR-H2: DINPNSGGTIYAQKFQE (SEQ ID NO: 60); и
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
Ab3: гуманизированная легкая цепь 45AGX Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 (VL3)) Каппа (PX) (или ее вариабельный домен) и гуманизированная тяжелая цепь 54AHH Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 VH6 N55D) IgG1/Каппа (PX) (или ее вариабельный домен); например, включающие:
легкую цепь иммуноглобулина, включающую аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
(SEQ ID NO: 45)
тяжелую цепь иммуноглобулина, включающую аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNDGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
(SEQ ID NO: 46); или
вариабельный домен легкой цепи иммуноглобулина, включающий аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIK
(аминокислоты 1-111 SEQ ID NO: 45 (CDR подчеркнуты)); и
вариабельный домен тяжелой цепи иммуноглобулина, включающий аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNDGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSS
(аминокислоты 1-119 SEQ ID NO: 46 (CDR подчеркнуты));
или включающие CDR:
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2: GASNLES (SEQ ID NO: 39);
CDR-L3: QQSTEDPRT (SEQ ID NO: 40);
CDR-H1: DYNVD (SEQ ID NO: 33);
CDR-H2: DINPNDGGTIYAQKFQE (SEQ ID NO: 61); и
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
Ab4: гуманизированная легкая цепь 45AGX Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 (VL3)) Каппа (PX) (или ее вариабельный домен) и гуманизированная тяжелая цепь 52AHH Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 VH6 N55Q) IgG1/Каппа (PX) (или ее вариабельный домен); например, включающие:
легкую цепь иммуноглобулина, включающую аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
(SEQ ID NO: 47); и
тяжелую цепь иммуноглобулина, включающую аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNQGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
(SEQ ID NO: 48); или
вариабельный домен легкой цепи иммуноглобулина, включающий аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIK
(аминокислоты 1-111 SEQ ID NO: 47 (CDR подчеркнуты)); и
вариабельный домен тяжелой цепи иммуноглобулина, включающий аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNQGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSS
(аминокислоты 1-119 SEQ ID NO: 48 (CDR подчеркнуты));
или включающие CDR:
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2: GASNLES (SEQ ID NO: 39);
CDR-L3: QQSTEDPRT (SEQ ID NO: 40);
CDR-H1: DYNVD (SEQ ID NO: 33);
CDR-H2: DINPNQGGTIYAQKFQE (SEQ ID NO: 62); и
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
Ab5: гуманизированная легкая цепь 45AGX Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 (VL3)) Каппа (PX) (или ее вариабельный домен) и гуманизированная тяжелая цепь 57AHH Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 VH6) IgG4 S228P (PX) (или ее вариабельный домен); например, включающие:
легкую цепь иммуноглобулина, включающую аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
(SEQ ID NO: 49); и
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNNGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
(SEQ ID NO: 50); или
вариабельный домен легкой цепи иммуноглобулина, включающий аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIK
(аминокислоты 1-111 SEQ ID NO: 49 (CDR подчеркнуты)); и
вариабельный домен тяжелой цепи иммуноглобулина, включающий аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNNGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSS
(аминокислоты 1-119 SEQ ID NO: 50 (CDR подчеркнуты));
или включающие CDR:
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2: GASNLES (SEQ ID NO: 39);
CDR-L3: QQSTEDPRT (SEQ ID NO: 40);
CDR-H1: DYNVD (SEQ ID NO: 33);
CDR-H2: DINPNNGGTIYAQKFQE (SEQ ID NO: 59); и
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
Ab6: гуманизированная легкая цепь 45AGX Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 (VL3)) Каппа (PX) (или ее вариабельный домен) и гуманизированная тяжелая цепь 73AHD Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 VH6 N55D/VL3) IgG4 S228P/Каппа (PX) (или ее вариабельный домен); например, включающие:
легкую цепь иммуноглобулина, включающую аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
(SEQ ID NO: 51); и
тяжелую цепь иммуноглобулина, включающую аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNDGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
(SEQ ID NO: 52); или
вариабельный домен легкой цепи иммуноглобулина, включающий аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIK
(аминокислоты 1-111 SEQ ID NO: 51 (CDR подчеркнуты)); и
вариабельный домен тяжелой цепи иммуноглобулина, включающий аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNDGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSS
(аминокислоты 1-119 SEQ ID NO: 52 (CDR подчеркнуты));
или включающие CDR:
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2: GASNLES (SEQ ID NO: 39);
CDR-L3: QQSTEDPRT (SEQ ID NO: 40);
CDR-H1: DYNVD (SEQ ID NO: 33);
CDR-H2: DINPNDGGTIYAQKFQE (SEQ ID NO: 61); и
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
Ab7: гуманизированная легкая цепь 45AGX Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 (VL3)) Каппа (PX) (или ее вариабельный домен) и гуманизированная тяжелая цепь 21AHG Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 VH6 N55S/VL3) IgG4 S228P/Каппа (PX) (или ее вариабельный домен); например, включающие:
легкую цепь иммуноглобулина, включающую аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
(SEQ ID NO: 53); и
тяжелую цепь иммуноглобулина, включающую аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNSGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
(SEQ ID NO: 54); или
вариабельный домен легкой цепи иммуноглобулина, включающий аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIK
(аминокислоты 1-111 SEQ ID NO: 53 (CDR подчеркнуты)); и
вариабельный домен тяжелой цепи иммуноглобулина, включающий аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNSGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSS
(аминокислоты 1-119 SEQ ID NO: 54 (CDR подчеркнуты));
или включающие CDR:
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2: GASNLES (SEQ ID NO: 39);
CDR-L3: QQSTEDPRT (SEQ ID NO: 40);
CDR-H1: DYNVD (SEQ ID NO: 33);
CDR-H2: DINPNSGGTIYAQKFQE (SEQ ID NO: 60); и
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
Ab8: гуманизированная легкая цепь 45AGX Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 (VL3)) Каппа (PX) (или ее вариабельный домен) и гуманизированная тяжелая цепь 80AHG Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 VH6 N55Q/VL3) IgG4 S228P/Каппа (PX) (или ее вариабельный домен); например, включающие:
легкую цепь иммуноглобулина, включающую аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
(SEQ ID NO: 55); и
тяжелую цепь иммуноглобулина, включающую аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNQGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
(SEQ ID NO: 56); или
вариабельный домен легкой цепи иммуноглобулина, включающий аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIK
(аминокислоты 1-111 SEQ ID NO: 55 (CDR подчеркнуты)); и
вариабельный домен тяжелой цепи иммуноглобулина, включающий аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNQGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSS
(аминокислоты 1-119 SEQ ID NO: 56 (CDR подчеркнуты));
или включающие CDR:
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2: GASNLES (SEQ ID NO: 39);
CDR-L3: QQSTEDPRT (SEQ ID NO: 40);
CDR-H1: DYNVD (SEQ ID NO: 33);
CDR-H2: DINPNQGGTIYAQKFQE (SEQ ID NO: 62); и
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
или
Ab9: гуманизированная легкая цепь 45AGX Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 (VL3)) Каппа (PX) (или ее вариабельный домен) и гуманизированная тяжелая цепь 72AHD Гуманизированного x [LAG3_H] mAb (LB145.22D2.E1.D1 VH6 N55G/VL3) IgG4 S228P/Каппа (PX)) (или ее вариабельный домен); например, включающие:
легкую цепь иммуноглобулина, включающую аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
(SEQ ID NO: 57); и
тяжелую цепь иммуноглобулина, включающую аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNGGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
(SEQ ID NO: 58); или
вариабельный домен легкой цепи иммуноглобулина, включающий аминокислотную последовательность:
DIVMTQTPLSLSVTPGQPASISCKASQSLDYEGDSDMNWYLQKPGQPPQLLIYGASNLESGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCQQSTEDPRTFGGGTKVEIK
(аминокислоты 1-111 SEQ ID NO: 57 (CDR подчеркнуты)); и
вариабельный домен тяжелой цепи иммуноглобулина, включающий аминокислотную последовательность:
QMQLVQSGPEVKKPGTSVKVSCKASGYTFTDYNVDWVRQARGQRLEWIGDINPNGGGTIYAQKFQERVTITVDKSTSTAYMELSSLRSEDTAVYYCARNYRWFGAMDHWGQGTTVTVSS
(аминокислоты 1-119 SEQ ID NO: 58 (CDR подчеркнуты));
или включающие CDR:
CDR-L1: KASQSLDYEGDSDMN (SEQ ID NO: 38);
CDR-L2: GASNLES (SEQ ID NO: 39);
CDR-L3: QQSTEDPRT (SEQ ID NO: 40);
CDR-H1: DYNVD (SEQ ID NO: 33);
CDR-H2: DINPNGGGTIYAQKFQE (SEQ ID NO: 63); и
CDR-H3: NYRWFGAMDH (SEQ ID NO: 35)
В одном варианте осуществления изобретения CDR-H2 любого анти-LAG3 антитела или его антигенсвязывающего фрагмента по настоящему изобретению включает аминокислотную последовательность:
DINPNX1GGTIYX2QKFX3X4 (SEQ ID NO: 64)
где,
X1= D,N,S или Q
X2= A или S
X3= Q или K
X4= E или G
Настоящее изобретение включает антитела и их антигенсвязывающие фрагменты (например, 4A10, 19E8, 11C9 и/или 22D2; например, Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 и/или Ab9), включающие N-связанные гликаны, которые типично добавляются к иммуноглобулинам, продуцируемым в клетках яичников китайского хомячка (CHO N-связанные гликаны) или к генно-инженерным дрожжевым клеткам (N-связанные гликаны генно-инженерных дрожжей), таким как, например, Pichia pastoris. Например, в одном варианте осуществления изобретения антитело или антигенсвязывающий фрагмент включает один или несколько ʺN-связанных гликанов генно-инженерных дрожжейʺ или ʺN-связанных гликанов CHOʺ, которые показаны на Фиг. 11 (например, G0 и/или G0-F, и/или G1, и/или G1-F, и/или G2-F, и/или Man5). В одном варианте осуществления изобретения антитело или антигенсвязывающий фрагмент включает N-связанные гликаны генно-инженерных дрожжей, т.е. G0 и/или G1 и/или G2, необязательно дополнительно включающие Man5. В одном варианте осуществления изобретения антитело или антигенсвязывающий фрагмент включают N-связанные гликаны CHO, т.е. G0-F, G1-F и G2-F, необязательно дополнительно включающие G0 и/или G1, и/или G2, и/или Man5. В одном варианте осуществления изобретения от около 80% до около 95% (например, около 80-90%, около 85%, около 90% или около 95%) из всех N-связанных гликанов на цепи иммуноглобулина антитела или антигенсвязывающего фрагмента представляют собой N-связанные гликаны генно-инженерных дрожжей или N-связанные гликаны CHO. См. Nett et al. Yeast. 28(3): 237-252 (2011); Hamilton et al. Science. 313(5792): 1441-1443 (2006); Hamilton et al. Curr Opin Biotechnol. 18(5): 387-392 (2007). Например, в одном варианте осуществления изобретения генно-инженерная дрожжевая клетка представляет собой GFI5.0 или YGLY8316 или штаммы, описанные в Патенте США № 7795002 или Zha et al. Methods Mol Biol. 988:31-43 (2013). См. также публикацию международной патентной заявки № WO2013/066765.
Варианты анти-LAG3 антител с сульфатированием тирозина (например, Ab1, Ab2, Ab3, Ab4, Ab5, Ab6, Ab7, Ab8 и/или Ab9) включают молекулярные массы около 148670 Да, 148832 Да и/или 148994 Да. Варианты с отсутствием сульфатирования тирозина включают молекулярные массы около 148590 Да, 148752 Да и/или 148914 Да.
ʺВыделенныеʺ антитела или их антигенсвязывающие фрагменты, по меньшей мере частично, свободны от других биологических молекул из клеток или клеточной культуры, из которой они получены. Такие биологические молекулы включают нуклеиновые кислоты, белки, липиды, углеводы или другие материалы, такие как клеточный дебрис и питательная среда. Выделенное антитело или его антигенсвязывающий фрагмент также могут, по меньшей мере частично, быть свободными от компонентов системы экспрессии, таких как биологические молекулы из клетки-хозяина или ее питательной среды. Как правило, термин "выделенный" не предполагает полного отсутствия таких биологических молекул или отсутствия воды, буферов или солей или компонентов фармацевтической композиции, которая включает антитела или фрагменты.
Антигенсвязывающий фрагмент антитела представляет собой часть антитела, которая сохраняет способность специфически связываться с антигеном, с которым связывается полноразмерное антитело. Примеры антигенсвязывающих фрагментов включают, но не ограничиваются этим, Fab, Fab', F(ab')2 и Fv фрагменты; диатела; молекулы одноцепочечных антител, например sc-Fv; нанотела и мультиспецифические антитела, образованные из фрагментов антител.
Как правило, основная структурная единица антитела включает тетрамер. Каждый тетрамер включает две идентичные пары полипептидных цепей, каждая пара имеет одну "легкую" и одну "тяжелую" цепь. См. общее описание в Fundamental Immunology Ch. 7 (Paul, W., ed., 2nd ed. Raven Press, N.Y. (1989).
Моноклональные антитела являются по существу гомогенными антителами, т.е. молекулы антител, образующие популяцию, являются идентичными по аминокислотной последовательности, за исключением возможных природных мутаций, которые могут присутствовать в незначительных количествах. См. Kohler et al. (1975) Nature 256: 495; Патент США № 4816567; Clackson et al. (1991) Nature 352: 624-628; Marks et al. (1991) J. Mol. Biol. 222: 581-597; и Presta (2005) J. Allergy Clin. Immunol. 116:731.
Химерное антитело представляет собой антитело, содержащее вариабельный домен из первого антитела и константный домен из второго антитела, где первое и второе антитела происходят из разных видов. (Патент США № 4816567; и Morrison et al., (1984) Proc. Natl. Acad. Sci. USA 81: 6851-6855). Типично, вариабельные домены получают из антитела от экспериментального животного ("родительское антитело"), например, из вида грызунов, а последовательности константных доменов получают из человеческих антител, таким образом, полученное химерное антитело менее вероятно будет вызывать неблагоприятный иммунный ответ у человека по сравнению с родительским (например, мышиным) антителом.
Гуманизированное антитело содержит последовательности как из человеческого, так и нечеловеческого антитела (например, мыши или крысы). Как правило, гуманизированное антитело будет включать по существу все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют гипервариабельным петлям из нечеловеческого иммуноглобулина, и все или по существу все каркасные области (FR) происходят из последовательности человеческого иммуноглобулина. Гуманизированное антитело необязательно может включать по меньшей мере часть константной области человеческого иммуноглобулина (Fc).
Иммуноглобулины могут быть отнесены к разным классам в зависимости аминокислотных последовательностей константного домена их тяжелых цепей. Существует по меньшей мере пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно давлее подразделить на подклассы (изотипы), например, IgG-1, IgG-2, IgG-3 и IgG-4; IgA-1 и IgA-2. Изобретение включает антитела и антигенсвязывающие фрагменты (например, анти-LAG3) любого из этих классов или подклассов антител.
В одном варианте осуществления антитело или антигенсвязывающий фрагмент (например, анти-LAG3) включает константную область тяжелой цепи, например, человеческую константную область, такую как константная область человеческой тяжелой цепи γ1, γ2, γ3 или γ4 или ее вариант. В другом варианте осуществления антитело или антигенсвязывающий фрагмент (например, анти-LAG3) включает константную область легкой цепи, например, константную область человеческой легкой цепи, такую как область человеческой легкой цепи лямбда или каппа или ее вариант. В качестве примера, а не ограничения, константная область человеческой тяжелой цепи может быть γ4, а константная область человеческой легкой цепи может быть каппа. В альтернативном варианте осуществления Fc область антитела представляет собой γ4 с Ser228Pro мутацией (Schuurman, J et. al., Mol. Immunol. 38: 1-8, 2001).
Примеры
Эти примеры иллюстрируют настоящее изобретение и не предназначены для его ограничения.
Пример 1: Идентификация вариантов антител с сульфатированием тирозина и способы очистки. В этом примере выявляли присутствие имеющих сульфатирование тирозина вариантов антител Ab6, и был разработан способ очистки для удаления таких вариантов.
Материалы и методы
Щелочную фосфатазу приобретали у New England Biolabs (Ipswich, MA). Антитело против сульфатирования тирозина приобретали у Millipore (Billerica, MA). Синтетический пептид приобретали у AnaSpec (Fremont, CA).
Анионообменная (AEX) хроматография
AEX хроматографию осуществляли с использованием предупакованной колонки POROS GoPure D, 0,5 × 5 см, 1 мл в проточном режиме с использованием системы GE Akta Avant. Очищенное хроматографией с протеином-A mAb доводили до pH 6,5 раствором 1M Tris и загружали на колонку. Перед загрузкой белка колонку уравновешивали раствором 25 мМ фосфата натрия pH 6,5, после загрузки колонку промывали раствором 25 мМ фосфата натрия pH 6,5 и десорбировали раствором 1M NaCl. Поглощение при 280 нм контролировали на протяжении всего эксперимента. Фракции, объединенные и десорбированные, и AEX нагрузку собирали и анализировали.
Ионообменная ВЭЖХ
Ионообменную ВЭЖХ осуществляли на MabPac SCX-10 колонке (4 × 250 мм, 3,14 мл) при температуре окружающей среды с использованием системы серии Agilent 1600. Подвижная фаза B представляла собой 30 мМ фосфата натрия pH 8,0, а подвижная фаза A представляла собой 25 мМ MES pH 5,8. Колонку сначала уравновешивали при 14% подвижной фазы B при скорости потока 1,0 мл/мин в течение 10 мин. mAb белок затем элюировали из колонки с использованием градиента подвижной фазы B (14% до 80% в течение 18 мин). Колонку затем очищали с использованием 100% подвижной фазы B в течение 3 мин и снова уравновешивали при 14% подвижной фазы B для анализа следующего образца. Поглощение при 280 нм элюата контролировали на протяжении всего ЖХ анализа.
ЖХ/МС анализ интактного и восстановленного образца
20 мкг образца разбавляли до 0,5 мг/мл при помощи 50 мМ Tris буфера pH 8,0. Разделение методом ОФ-ВЭЖХ осуществляли с использованием Waters Acquity UPLC H-Class. Использовали колонку Acquity UPLC BEH300 C4, 1,7 мкм, 1,0 × 100 мм (Waters, Milford, MA; -O-(Si)(CH3)2-C4H9 лиганд). Подвижные фазы представляли собой 0,1% муравьиной кислоты (FA) в воде в качестве подвижной фазы A и 0,1% FA в ацетонитриле (ACN) в качестве подвижной фазы B. Скорость потока в ЖХ была 0,08 мл/мин, и температуру колонки поддерживали при 80°C. Антитело элюировали с использованием градиента 4-15 мин 30% - 90% B. МС-спектры получали на системе Waters Xevo G2 Q-TOF при сканировании в диапазоне m/z 800-4000.
20 мкг образца разбавляли восстанавливающим буфером (50 мМ Tris pH 8,0, содержащий 6 M Гуанидина HCl) до конечного объема 100 мкл. Два микролитра раствора 1M дитиотреитола (DTT) (Sigma-Aldrich, St.Louis, MO) добавляли к каждому из образцов с последующей инкубацией при 56°C в течение 20 минут. ОФ-СВЭЖХ разделение осуществляли на Waters Acquity UPLC H-Class. Использовали колонку Acquity UPLC, BEH300 C4, 2,1 × 100 мм, 1,7 мкм (Waters). МС-спектры получали на системе Waters Xevo G2 Q-TOF при сканировании в диапазоне m/z 600-3000. МС данные анализировали при помощи MaxEnt1 программы MassLynx 4.1.
Пептидное картирование с использованием ЖХ/МС
100 мкг образца подвергали буферному обмену с 100 мкл денатурирующего буфера, содержащего 50 мМ Tris pH 8,0, 6 M Гуанидина HCl и 5 мМ EDTA. Восстановительные реакции осуществляли при 56°C в течение 30 минут с 20 мМ DTT в растворе. Образцы алкилировали с использованием 50 мМ иодацетамида при комнатной температуре в течение 30 минут в темноте. Реакцию алкилирования останавливали путем добавления 1мкл 500 мМ раствора DTT. Восстановленные и алкилированные образцы разбавляли буфером для гидролиза (50 мМ Tris pH 8,0) до конечного объема 300 мкл перед добавлением Lys-C фермента (Wako, Richmond, VA) при соотношении фермент:субстрат 1:20 (масс:масс). Раствор инкубировали при 37°C в течение 4 часов. Пептиды разделяли методом ОФ-ВЭЖХ на Waters Acquity UPLC H-Class с использованием колонки HALO Peptide ES-C18, 2,1 × 150 нм, 2,7 мкм (MAC-MOD Analytical, Inc., Chadds Ford, PA). МС-спектры получали на системе Waters Xevo G2 Q-TOF при сканировании в диапазоне m/z 100-2000. МС данные анализировали при помощи BiopharmaLynx 1.3 (Waters).
МС/МС целевого пептида
ЖХ/МС/МС анализ целевого пептида осуществляли на системе LTQ-Orbitrap MS (Thermo Fisher, Waltham, MA). Для получения МС/МС данных использовали разрешение 17500 с Фурье-преобразованием. Пептиды разделяли на Waters Acquity UPLC H-Class с использованием колонки HALO Peptide ES-C18, 2,1×150 мм, 2,7 мкм. МС/МС сканировали в m/z диапазонах в зависимости от m/z значений родительских ионов. Нормализованную энергию фрагментации устанавливали при 35% для СИД (столкновительно-индуцированная диссоциация) фрагментации и 35% для ДПЭ (диссоциация при переносе электрона) фрагментации. MS2 данные обрабатывали вручную.
Обработка щелочной фосфатазой
10 мкг mAb белка в AEX десорбированной фракции разбавляли в 50 мкл фосфатазного реакционного буфера. 1 мкл (10 единиц) щелочной фосфатазы из кишечника теленка (New England Biolabs, Ipswish, MA) добавляли для инкубации при 37°C в течение 1 часа. Куриный овальбумин (Sigma) обрабатывали одновременно в качестве положительного контроля. 10 мкл раствора вводили в систему ЖХ/МС для масс-анализа.
Вестерн-блоттинг
Magic Mark XP Western Standard (Invitrogen) и конкретные концентрации как моноклональных антител (mAb), так и контрольных клеточных экстрактов (HEK293 полный клеточный экстракт и EGF стимулированный A431 клеточный лизат (Millilpore)) восстанавливали при помощи β-меркаптоэтанола плюс нагревание при 95°C, затем разделяли методом SDS-PAGE на основе Tris-глицина с использованием 4-20% градиентного геля (Novex). Затем осуществляли электроперенос разделенных белков на нитроцеллюлозную мембрану и промывали в течение ночи в Tris-забуференном солевом растворе плюс 0,05% Tween20 (TBST) (Sigma) при взбалтывании при 4°C. Мембраны затем блокировали в течение 1 часа в Tris-забуференном солевом растворе плюс 1% BSA (TBS-BSA) (Sigma) при комнатной температуре при постоянном взбалтывании. Первичные антитела (против сульфотирозина/против сульфатирования тирозина (Millipore) или античеловеческое IgG (H+L) (Jackson ImmunoResearch Labratories Inc.)) разводили в TBS-BSA и инкубировали с мембраной в течение 2 часов при комнатной температуре. После промывки при помощи TBST HRP-конъюгированные вторичные антитела (козлиное-антимышиное или козлиное-антикроличье (Thermo Scientific)) разводили в 5% молочном белке из обезжиренного молока плюс 0,05% Tween20-фосфобуферный солевой раствор (Invitrogen) и инкубировали при комнатной температуре в течение 1 часа. После конечной промывки при помощи TBST использовали хемилюминесцентные субстраты (Thermo Scientific) для проявления; сигналы восстанавливали путем экспонирования на фотопленке (GE Healthcare Life Sciences) и последующей обработки. Очистку нитроцеллюлознной мембраны между первичными антителами осуществляли, как описано ранее (Kaufmann SH, E.C., Shaper JH., The erasable Western blot. Anal Biochem., 1987. 161(1): p. 89-95).
Результаты и обсуждение
Разделение молекул mAb
Анионообменную хроматографию (AEX) обычно используют в качестве стадии доочистки в процессе очистки моноклональных антител. Эта стадия типично работает в проточном режиме, где mAb протекает через колонку и выделяемые в ходе процесса примеси (белки клетки-хозяина, ДНК) связываются с колонкой. В ходе AEX было замечено, что есть фракция mAb, загруженных в колонку, связанная со смолой, которая влияет на выделение белка. Связанная фракция белка элюировала в десорбированной фракции AEX хроматографии. Чтобы охарактеризовать mAb, связанные с AEX колонкой, анализировали следующие фракции AEX хроматографии: ʺзагружаемаяʺ относится к образцу до AEX очистки; ʺобъединеннаяʺ относится к проточной части образца, и ʺдесорбированнаяʺ относится к связанной фракции образца. Загружаемую, объединенную и десорбированную фракции из AEX хроматографии сначала анализировали IEX-ВЭЖХ хроматографией. Фиг.1 показывает IEX-ВЭЖХ профиль mAb в загружаемой, объединенной и десорбированной фракции AEX. Как показано на Фиг. 1, десорбированная фракция имела значительно большее количество кислотных вариантов по сравнению с загружаемой и объединенной: ~65% кислотных вариантов в десорбированной фракции (тонкая линия) по сравнению с 23% в объединенной фракции (пунктирная линия) и 33% в загружаемой фракции (жирная линия). Дополнительное отличие было отмечено в кислотном пике, находящемся перед пиком главного компонента. Этот пик присутствовал в исходном образце AEX при более высоких уровнях, при этом в объединенной фракции AEX этот пик был минимальным. Десорбированная фракция была обогащена кислотным пиком, находящимся перед пиком главного компонента. AEX хроматографию также осуществляли при pH 7,0 или 7,5 с использованием таких же условий, что касается буферов и солей, как описано выше. При pH 7,0 подобное восстановление кислотного пика, находящегося перед пиком главного компонента, также наблюдали в объединенной фракции AEX.
Анализ интактного и восстановленного белка масс-спектрометрией
Чтобы охарактеризовать примеси, все три фракции AEX (исходная, объединенная и десорбированная) анализировали методом ЖХ/МС для определения интактного и восстановленного белка с использованием Q-TOF MS (времяпролетной масс-спектрометрии). Фиг. 2 показывает масс-спектры интактной молекулы после деконволюции. Три основные гликоформы наблюдали во всех трех фракциях: G0F/G0F, G0F/G1F и G1F/G1F с массой 148591 Да, 148751 Да и 148912 Да, соответственно. Рассчитанная интактная масса этой молекулы с G0F/G0F составляет 148590 Да. Погрешности в измерениях интактной массы все были в пределах 25 ч/млн. Дополнительные виды были обнаружены только в AEX десорбированной фракции. Эти виды сооветствуют увеличению массы на 80 Да (148668, 148830, 148991 Да) трех основных гликоформ (G0F/G0F, G1F/G0F, G1F/G1F). Для определения положения модификации массу легкой цепи и тяжелой цепи измеряли после расщепления дисульфидных связей восстанавливающим агентом DTT. Никакой разницы не было обнаружено в массе тяжелой цепи в десорбированной фракции и объединенной фракции, на основании чего можно предположить, что модификация не находится на тяжелой цепи (данные не показаны). Как показано на Фиг. 3, масса апо-формы легкой цепи (23674 Да) и масса гликированной легкой цепи (23836 Да) были обнаружены в обеих фракциях (десорбированной и объединенной фракции). Пик с 80 Да увеличением легкой цепи наблюдали только при 23754 Да в десорбированной фракции AEX. Погрешности в измерениях восстановленной массы были в пределах 20 ч/млн. Эти данные позволяют предположить, что 80 Да модификации присутствуют на легкой цепи Ab6.
Анализ mAb антитела методом пептидного картирования
Для дальнейшего уточнения места модификации десорбированную и объединенную фракции AEX восстанавливали, алкилировали и затем расщепляли LysC ферментом. Пептидные смеси анализировали методом масс-спектрометрического картирования с использованием масс-спектрометра Q-TOF MS. При сравнении УФ-спектров этих двух фракций было обнаружено два отличия. Как показано на Фиг. 4 (a) и (b), два новых пика были обнаружены при времени удерживания 37,6 мин и 65,2 мин в десорбированной фракции AEX. Наблюдаемые m/z в новых пиках были 1165,4796 (2+) через 37,6 мин и 1476,7372 (4+) Да через 65,2 мин. Наблюдаемые массы соответствуют пептиду легкой цепи AA25-43+80Da и AA25-78 +80 Да с погрешностью массы 6,4 ч/млн и 9,8 ч/млн, соответственно. Пептид легкой цепи AA25-78 содержит один сайт неправильного расщепления. Модифицированная и немодифицированная форма пептида легкой цепи AA25-43 и AA25-78 помечены на Фиг. 4. Уровень этого модифицированного пептида был оценен как 20,9% и 21,6% для AA25-43 и AA25-78 по сравнению с их апо-формами на основании площади пика на экстракционной ионной хроматограмме (SIC).
МС/МС фрагментация модифицированного пептида
Есть две возможности модификации с увеличением массы на 80 Да: фосфорилирование (+79,9663 Да) и сульфатирование (+79,9568 Да). Теоретически разница в массе между этими двумя модификациями только 0,0095 Да, что затрудняет установление различия только по массе. Сначала целевой пептид AA25-78 фрагментировали методом столкновительно-индуцированной диссоциации (СИД) и образовавшиеся фрагменты анализировали методом LTQ-Orbitrap MS. Как показано на Фиг. 5, наблюдали полную потерю группы модификации (80Da) из родительного иона. Были обнаружены только фрагменты из пептидного остова, что подтверждает пептидную последовательность LC25-43. При этом, никакой сайт-специфической информации не было получено из СИД фрагментации. Сообщалось о том, что сульфатированный тирозин (sY) очень лабилен и легко может потеряться в стандартных условиях СИД (Nemeth-Cawley JF1, K.S., Rouse JC., Analysis of sulfated peptides using positive electrospray ionization tandem mass spectrometry. J Mass Spectrom., 2001. 36(12): p. 1301-11). Было невозможно получить сайт-специфическую информацию о локализации сульфатных групп с использованием СИД/МС/МС в режиме положительных ионов, поскольку ни один из исходных родительских ионов на присутствовал во время фрагментации пептидного остова. В отличие от этого, фосфорилированные пептиды обычно сохраняются в условиях СИД, и фрагментация пептидного остова делает возможной сайт-специфическую идентификацию модификации (Nemeth-Cawley JF1, K.S., Rouse JC., Analysis of sulfated peptides using positive electrospray ionization tandem mass spectrometry. J Mass Spectrom., 2001. 36(12): p. 1301-11). На Фиг. 5 наблюдается нейтральная потеря 80 Да из родительского иона. Известно, что характерным ионом с нейтральной потерей для фосфорилирования является -H3PO4 (-98 Да), а характерным фрагментарным ионом является PO3- (-79 Да). Тогда как для сульфатирования характерными ионами с нейтральной потерей и фрагментарными ионами являются оба -SO3 иона с 80 Да (Monigatti F, H.B., Steen H., Protein sulfation analysis--A primer. Biochim Biophys Acta., 2006. 1764(12): p. 1904-13). СИД MS2 данные говорят о том, что 80 Да модификация представляет собой сульфатирование.
Другим широко используемым механизмом фрагментации является диссоциация при переносе электрона (ДПЭ). Она переносит электрон к многократно протонированному пептиду/белку, что может привести к расщеплению N-Cα связей в основной цепи и образованию c- и z-типа фрагментарных ионов без потери информации о локализации PTM (Mikesh LM, U.B., Chi A, Coon JJ, Syka JE, Shabanowitz J, Hunt DF., The utility of ETD mass spectrometry in proteomic analysis. Biochim Biophys Acta., 2006. 1764(12): p. 1811-22). ДПЭ может обеспечит комплементарную информацию с СИД: ДПЭ процесс позволяет сохранить SO3 группу и, таким образом, аминокислотную локализацию, тогда как СИД предпочтительно фрагментирует лабильные модификации. В данном случае, целевой пептид анализировали при помощи LTQ-Orbitrap с ДПЭ фрагментацией и масс-детекцией высокого разрешения. Как показано на Фиг. 6, частичную потерю 80 Да модификаций наблюдали на родительском ионе. Фрагментарные ионы с присоединенной SO3 группой (80 Да) являются мечеными. На основании детекции фрагментарных ионов с присоединенной SO3 группой (c9, c11, c12-16), сайт модификации был идентифицирован как тирозин 31 на легкой цепи, который находится в CDR1 области молекулы mAb.
Обработка щелочной фосфатазой
Поскольку фосфорилирование и сульфатирование тирозина являются изобарическими, использовали щелочную фосфатазу для различения этих двух модификаций (Yu Y, H.A., Moore KL, Leary JA., Determination of the sites of tyrosine O-sulfation in peptides and proteins. Nat Methods, 2007. 4(7): p. 583-8). Щелочная фосфатаза широко используется для удаления группы фосфорилирования из белков. Куриный альбумин использовали в качестве положительного контроля, так как этот белок широко известен своей формой фосфорилирования и гликозилирования. Куриный альбумин и mAb в десорбированной фракции AEX обрабатывали фосфатазой и инкубировали при 37°С одновременно. Фиг. 7 показывает измеренную интактную массу mAb и куриного овальбумина до и после обработки фосфатазой. Как показано на Фиг. 7(а), для mAb никакого изменения массы не наблюдали. Что касается куриного альбумина (Фиг. 7 (b)), то для всех основных гликоформ наблюдали явное изменение массы на 160 Да. Поскольку куриный альбумин содержит два сайта фосфорилирования, потеря 160 Да подтверждает активность щелочной фосфатазы. Поскольку никакого изменения массы не было обнаружено до и после обработки фосфатазой, это свидетельствует о том, что это mAb в десорбированной фракции AEX не является фосфорилированным.
Вестерн-блот
До настоящего времени ЖХ/МС анализ использовали для исследования природы 80 Да аддукта к тирозину 31 в CDR легкой цепи. MS2 анализ и анализ массы AEX десорбированной фракции mAb после обработки фосфатазой позволили предположить, что 80 Да аддукт представляет собой сульфатирование на тирозине 31. Однако способность этих основанных на масс-анализе методов устанавливать непосредственное различие между сульфатированием тирозина и фосфорилированием проблематична из-за одинаковой молекулярной массы этих двух групп. Чтобы приступить к решению этой проблемы, применяли вестерн-блоттинг с анти-сульфотирозин-специфическим моноклональным антителом для подтверждения присутствия сульфатирования тирозина в AEX десорбированной фракции mAb (Xu J, D.X., Tang M, Li L, Xiao L, Yang L, Zhong J, Bode AM, Dong Z, Tao Y, Cao Y., Tyrosylprotein sulfotransferase-1 and tyrosine sulfation of chemokine receptor 4 are induced by Epstein-Barr virus encoded latent membrane protein 1 and associated with the metastatic potential of human nasopharyngeal carcinoma. PLoS One., 2013. 8(3): p. e56114). На Фиг. 8а (верхняя панель) нормализованные концентрации mAb AEX объединенной и десорбированной фракций подвергали SDS-PAGE в восстановительных условиях, анализировали на человеческие тяжелые и легкие цепи гибридизацией с использованием вестерн-блоттинга. Повышенные концентрации тяжелых и легких цепей из объединенной и десорбированной фракции затем ʺочищалиʺ от первых обнаруживаемых антител и повторно анализировали с анти-сульфотирозин-специфическим моноклональным антителом (нижняя панель). Как показано на Фиг. 8а (нижняя панель), положительные сигналы были обнаружены только на легкой цепи десорбированных фракций, что свидетельствует о том, что она содержит сульфатирование тирозина. В качестве контроля для перекрестной реактивности с фосфорилированием, на полосу 1 загружали коммерческий источник EGF-обработанного А431клеточного экстракта, который обогащен фосфорилированными белками. Отсутствие положительного сигнала в полосе 1 нижней панели показывает, что анти-сульфотирозиновое моноклональное антитело не обладает сильной перекрестной реактивностью с фосфорилированием (Фиг. 8a, нижняя панель). Кроме того, экстракт HEK 293, который был предложен изготовителем в качестве положительного контроля для анти-сульфотирозинового моноклонального антитела, загружали на полосу 2. Положительный сигнал ниже нижнего значения 20 кДа согласуется с анализом изготовителя (Фиг. 8a, нижняя панель). На Фиг. 8b, нормализованные концентрации различных моноклональных антител, полученных из СНО (mAb1, 2 и 3), в дополнение к AEX десорбированной и объединенной фракции подвергали SDS-PAGE в восстановительных условиях, исследовали на человеческие HC и LC гибридизацией с использованием вестерн-блоттинга (верхняя панель), затем десорбировали и снова исследовали на антисульфотирозин (нижняя панель). В соответствии с Фиг. 8а, только AEX десорбированная фракция mAb показывает положительный сигнал в положении легкой цепи при зондировании анти-сульфотирозиновым антителом. Никакого положительного сигнала сульфатирования тирозина не наблюдали для трех других полученных из CHO Merck mAb при анализе аналогичных количеств белка. Это согласуется с наблюдением, что никакого повышенного уровня кислотного пика AEX или ʺгорячей точкиʺ сульфатирования тирозина не было обнаружено на этих трех mAb (данные не показаны).
Сравнение времени удерживания с использованием синтетического пептида с сульфатированием или фосфорилированием
Для дальнейшего разграничения фосфорилирования и сульфатирования синтетический пептид с идентичной последовательностью LC AA25-43 (XSXSXDYEGDSDXXXXXXX) (SEQ ID NO: 65), модифицированный либо фосфорилированием, либо сульфатированием по Y31, анализировали методом ЖХ/МС. Фиг. 9 показывает SIC синтетического пептида XSXSXDYEGDSDXXXXXXX (SEQ ID NO: 65)+фосфорилирование, XSXSXDYEGDSDXXXXXXX (SEQ ID NO: 65)+сульфатирование и AA25-43+80Da в десорбированной фракции AEX. Синтетический пептид с сульфатированием элюируется при таком же времени удерживания, как десорбированная фракция AEX, тогда как синтетический пептид с фосфорилированием элюируется раньше, чем десорбированная фракция AEX. Это является дополнительным подтверждением наблюдения, сделанного авторами настоящего изобретения, что Y31 на легкой цепи является сульфатированным.
Структура сайта сульфатирования тирозина
Реакция сульфатирования тирозина белка катализируется ферментом Гольджи, называемым тирозилпротеинсульфотрансферазой (TPST). Предыдущие исследования показали, что TPST распознают доступные тирозиновые остатки, которые обычно окружены несколькими кислотными остатками в пределах от -5 до +5 положений (Hortin G, F.R., Gordon JI, Strauss AW., Characterization of sites of tyrosine sulfation in proteins and criteria for predicting their occurrence. Biochem Biophys Res Commun., 1986. 141(1): p. 326-33; Rosenquist GL, N.H.J., Analysis of sequence requirements for protein tyrosine sulfation. Protein Sci., 1993. 2(2): p. 215-22; Teramoto T1, F.Y., Kawaguchi Y, Kurogi K, Soejima M, Adachi R, Nakanishi Y, Mishiro-Sato E, Liu MC, Sakakibara Y, Suiko M, Kimura M, Kakuta Y, Crystal structure of human tyrosylprotein sulfotransferase-2 reveals the mechanism of protein tyrosine sulfation reaction. Nat Commun., 2013. 4: p. 1572). Акцепторный тирозин должен иметь кислотные остатки поблизости, чтобы обеспечить распознавание положительно заряженных остатков в сайте связывания субстрата TPST2. Акцепторный тирозин также должен находиться в по существу гибкой области, чтобы вписаться в глубокую щель TPST2. Однако общая консенсусная последовательность для сайтов сульфатирования тирозина не была определена. Наиболее типичные признаки, описывающие окружение сульфатированного тирозина в последовательности, включают присутствие одной кислой аминокислоты в пределах двух остатков от тирозина; присутствие по меньшей мере трех кислых аминокислот в пределах 5 остатков и присутствие поблизости изгиб-индуцирующих аминокислот и т.д. (Monigatti F, H.B., Steen H., Protein sulfation analysis--A primer. Biochim Biophys Acta., 2006. 1764(12): p. 1904-13). Фиг. 10 показывает структуру тирозинового участка mAb в контексте петель CDR на ленточной диаграмме, созданной программой MOE (Chemical Computing Group, Montreal, Canada). Последовательность рядом с легкой цепью Y31 на этом mAb представляет собой: XSXSXDYEGDSDXXXXXXX (SEQ ID NO: 65). В этой последовательности соседние с Y31 остатки являются кислотными: аспарагиновая кислота (D) и глутаминовая кислота (E). В общей сложности четыре кислотных остатка находятся в пределах пяти остатков от Y31: три D и один E. Четыре изгиб-индуцирующих остатка находятся близко к Y31: три серина (S) и один глицин (G). Уникальная структура Y31 с соседними кислыми аминокислотами и элементами локальной вторичной структуры играет существенную роль, чтобы могла произойти эта модификация.
Авторы изобретения описывают в настоящей заявке доказательства, указывающие на присутствие неожиданного O-связанного сульфатирования тирозина в CHO-продуцированных антителах. Положение этой лабильной модификации было обнаружено в CDR1 области легкой цепи, что было идентифицировано масс-спектрометрией с ДПЭ фрагментацией. Это сульфатирование тирозина было дополнительно подтверждено методом обработки фосфатазой, вестерн-блоттингом с использованием антитела против сульфатирования тирозина и корреляцией времени удерживания с синтетическим сульфатированным пептидом. Структурный анализ тирозина CDR подтверждает влияние кислотных остатков на сульфатирование. Соседние кислые аминокислотные остатки и элементы локальной вторичной структуры могут играть существенную роль, чтобы сделать Y31 горячей точкой для сульфатирования.
Объем настоящего изобретения не должен ограничиваться конкретными вариантами осуществления, описанными в настоящей заявке. Действительно, объем настоящего изобретения включает варианты осуществления, конкретно изложенные в настоящей заявке, и другие варианты осуществления, не изложенные конкретно в настоящей заявке; варианты осуществления, конкретно изложенные в настоящей заявке, не обязательно должны быть исчерпывающими. Различные модификации изобретения в дополнение к тем, которые описаны в настоящей заявке, станут очевидными для специалистов в данной области техники из предшествующего описания. Такие модификации предусматриваются как охватываемые объемом формулы изобретения.
В настоящей заявке повсеместно присутствуют ссылки на патенты, патентные заявки, публикации, описания продуктов и протоколы, раскрытия которых включены в настоящую заявку посредством ссылки во всей их полноте для всех целей. Настоящая заявка испрашивает приоритет по предварительной заявке США № 62/414209, включенной в настоящую заявку посредством ссылки во всей полноте. Все ссылочные документы, цитируемые в настоящей заявке, включены посредством ссылки в той степени, как если бы каждая отдельная публикация, запись в базе данных (например, последовательности Genbank или GeneID запись), патентная заявка или патент были специально и индивидуально указаны для включения в качестве ссылки. Заявление о включении посредством ссылки, в соответствии с 37 C.F.R. 1.57 (b) (1), как подразумевают заявители, относится к каждой отдельной публикации, записи в базе данных (например, последовательности Genbank или GeneID записи), патентной заявке или патенту, каждая из которых четко идентифицирована в соответствии с 37 C.F.R. 1.57 (b) (2), даже если при указании такой ссылки непосредственно рядом нет специального указания о включении посредством ссылки. Включение специальных указаний о включении посредством ссылки, если таковые имеются, в описание изобретения никоим образом не ослабляет это общее заявление о включении посредством ссылки. Цитирование ссылочных документов в настоящей заявке не предназначено для признания того, что ссылочный документ относится к предшествующему уровню техники, и не является никаким признанием, что касается содержания или даты этих публикаций или документов. В случае, если ссылочные документы обеспечивают определение заявленного термина, которое противоречит определениям, приведенным в настоящем описании, должны использоваться определения, представленные в настоящем описании, для интерпретации заявленного изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Merck Sharp & Dohme Corp.
Zhao, Jia
Rios, Sandra
Dukleska Schussler, Svetlana
<120> Способ очистки для удаления вариантов антител с сульфатированием
тирозина; очищенные композиции
<130> 24380
<150> 62/414,209
<151> 2016-10-28
<160> 65
<170> PatentIn version 3.5
<210> 1
<211> 408
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 1
atgaaatgca gctgggtcat cttcttcctg atggcagtgg ttataggaat caattcagag
60
gttcagctgc tccagtctgg ggcagaactt gtgaggtcag gggcctcagt caagttgtcc
120
tgcacagcct ctggcttcaa cattgaagac tactatatgc actggatgaa acagaggcct
180
gaacagggcc tggagtggat tggatggatt gatcctgtga atggtgatac tgaatatgcc
240
ccgaagttcc agggcaaggc cactatgact gcagacacat cctccaacac agcctaccta
300
cacctcaaca gcctgacatc tgaggacact gccgtctatt actgtaattt ctatgatggt
360
tacctctttg ctttctgggg ccaagggacc ctggtcactg tctctgca
408
<210> 2
<211> 136
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 2
Met Lys Cys Ser Trp Val Ile Phe Phe Leu Met Ala Val Val Ile Gly
1 5 10 15
Ile Asn Ser Glu Val Gln Leu Leu Gln Ser Gly Ala Glu Leu Val Arg
20 25 30
Ser Gly Ala Ser Val Lys Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile
35 40 45
Glu Asp Tyr Tyr Met His Trp Met Lys Gln Arg Pro Glu Gln Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Asp Pro Val Asn Gly Asp Thr Glu Tyr Ala
65 70 75 80
Pro Lys Phe Gln Gly Lys Ala Thr Met Thr Ala Asp Thr Ser Ser Asn
85 90 95
Thr Ala Tyr Leu His Leu Asn Ser Leu Thr Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Asn Phe Tyr Asp Gly Tyr Leu Phe Ala Phe Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ala
130 135
<210> 3
<211> 10
<212> Белок
<213> Mus
<400> 3
Gly Phe Asn Ile Glu Asp Tyr Tyr Met His
1 5 10
<210> 4
<211> 17
<212> Белок
<213> Mus
<400> 4
Trp Ile Asp Pro Val Asn Gly Asp Thr Glu Tyr Ala Pro Lys Phe Gln
1 5 10 15
Gly
<210> 5
<211> 8
<212> Белок
<213> Mus
<400> 5
Tyr Asp Gly Tyr Leu Phe Ala Phe
1 5
<210> 6
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 6
atgaggtgcc tagctgagtt cctggggctg cttgtgctct ggatccctgg agccattggg
60
gatattgtgc tgactcaggc tgcaccctct gtacctgtca ctcctggaga gtcagtgtcc
120
atctcctgca ggtctagtaa gagtctcctg catagtgatg gcaacactta tctgtattgg
180
ctcctgcaga ggccaggcca gtctcctcag ctcctgatat atcggatgtc caaccttgcc
240
tcaggggtcc cagacaggtt cagcggcagt gggtcaggaa ctgttttcac actgagaatc
300
agcagactgg aggctgagga tgtgggtatt tattactgta tgcaacatct agaatatcct
360
ttcacgtttg gaggggggac caagctggaa ataaaa
396
<210> 7
<211> 132
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 7
Met Arg Cys Leu Ala Glu Phe Leu Gly Leu Leu Val Leu Trp Ile Pro
1 5 10 15
Gly Ala Ile Gly Asp Ile Val Leu Thr Gln Ala Ala Pro Ser Val Pro
20 25 30
Val Thr Pro Gly Glu Ser Val Ser Ile Ser Cys Arg Ser Ser Lys Ser
35 40 45
Leu Leu His Ser Asp Gly Asn Thr Tyr Leu Tyr Trp Leu Leu Gln Arg
50 55 60
Pro Gly Gln Ser Pro Gln Leu Leu Ile Tyr Arg Met Ser Asn Leu Ala
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Val Phe
85 90 95
Thr Leu Arg Ile Ser Arg Leu Glu Ala Glu Asp Val Gly Ile Tyr Tyr
100 105 110
Cys Met Gln His Leu Glu Tyr Pro Phe Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys
130
<210> 8
<211> 16
<212> Белок
<213> Mus
<400> 8
Arg Ser Ser Lys Ser Leu Leu His Ser Asp Gly Asn Thr Tyr Leu Tyr
1 5 10 15
<210> 9
<211> 8
<212> Белок
<213> Mus
<400> 9
Tyr Arg Met Ser Asn Leu Ala Ser
1 5
<210> 10
<211> 9
<212> Белок
<213> Mus
<400> 10
Met Gln His Leu Glu Tyr Pro Phe Thr
1 5
<210> 11
<211> 414
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 11
atgggatgga gctggatctt tcttttcctc ctgtcaggaa ctgcaggtgt ccgttgccag
60
atccgactgc agcagtctgg acctgagctg gtgaagcctg gggcttcagt gaagatatcc
120
tgcaaggctt ctgggtcctc cttcactgac tactatataa actgggtgaa gcagaagcct
180
ggacagggac ttgagtggat tggatggatt tatcctggaa gcggtaattc tatctacaat
240
gagaacttca aggccaaggc cacattgact gtagacacat cctccagcac agcctacatg
300
catctcagca gcctgacatc tgaggacact gctgtctatt tctgtgcaag agaggctgat
360
tacgacgatg ctttggacta ctggggtcaa ggaacctcgg tcaccgtctc ctca
414
<210> 12
<211> 138
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 12
Met Gly Trp Ser Trp Ile Phe Leu Phe Leu Leu Ser Gly Thr Ala Gly
1 5 10 15
Val Arg Cys Gln Ile Arg Leu Gln Gln Ser Gly Pro Glu Leu Val Lys
20 25 30
Pro Gly Ala Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Ser Ser Phe
35 40 45
Thr Asp Tyr Tyr Ile Asn Trp Val Lys Gln Lys Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Ser Gly Asn Ser Ile Tyr Asn
65 70 75 80
Glu Asn Phe Lys Ala Lys Ala Thr Leu Thr Val Asp Thr Ser Ser Ser
85 90 95
Thr Ala Tyr Met His Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val
100 105 110
Tyr Phe Cys Ala Arg Glu Ala Asp Tyr Asp Asp Ala Leu Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
130 135
<210> 13
<211> 10
<212> Белок
<213> Mus
<400> 13
Gly Ser Ser Phe Thr Asp Tyr Tyr Ile Asn
1 5 10
<210> 14
<211> 17
<212> Белок
<213> Mus
<400> 14
Trp Ile Tyr Pro Gly Ser Gly Asn Ser Ile Tyr Asn Glu Asn Phe Lys
1 5 10 15
Ala
<210> 15
<211> 10
<212> Белок
<213> Mus
<400> 15
Glu Ala Asp Tyr Asp Asp Ala Leu Asp Tyr
1 5 10
<210> 16
<211> 384
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 16
atggtatcca cacctcagtt ccttgtattt ttgcttttct ggattccagc ctccagaggt
60
cacatcttgc tgactcagtc tccagccatt ctgtctgtga gtccaggaga aagagtcagt
120
ttctcctgca gggccagtca gagcattggc acaagcatac actggtatca gcaaagaaca
180
aatggttctc caaggcttct cataaagtat gcttctgagt ctatctctgg gatcccttcc
240
aggtttagtg gcagtggatc agggacagat tttactctta gcatcaacag tgtggagtca
300
gaagatattg cagattatta ctgtcaacaa agtaatagct ggccaacgta cacgttcgga
360
ggggggacca agctggaaat aaaa
384
<210> 17
<211> 128
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 17
Met Val Ser Thr Pro Gln Phe Leu Val Phe Leu Leu Phe Trp Ile Pro
1 5 10 15
Ala Ser Arg Gly His Ile Leu Leu Thr Gln Ser Pro Ala Ile Leu Ser
20 25 30
Val Ser Pro Gly Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser
35 40 45
Ile Gly Thr Ser Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro
50 55 60
Arg Leu Leu Ile Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn
85 90 95
Ser Val Glu Ser Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Ser Asn
100 105 110
Ser Trp Pro Thr Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
115 120 125
<210> 18
<211> 11
<212> Белок
<213> Mus
<400> 18
Arg Ala Ser Gln Ser Ile Gly Thr Ser Ile His
1 5 10
<210> 19
<211> 7
<212> Белок
<213> Mus
<400> 19
Tyr Ala Ser Glu Ser Ile Ser
1 5
<210> 20
<211> 10
<212> Белок
<213> Mus
<400> 20
Gln Gln Ser Asn Ser Trp Pro Thr Tyr Thr
1 5 10
<210> 21
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 21
atgagatgga gctgtatcat cctcttcttg gtagcaacag ctacaggtgt caactcccag
60
gtccaactgc agcagcctgg ggctgagctt gtgatgcctg gggcttcagc gaagatgtcc
120
tgcaaggctt ctggctacac actcactgac tactggatgc actgggtgaa gcagaggcct
180
ggacaaggcc ttgagtggat cggagcgatt gatatttctg atagttattc tagctacaat
240
caaaagttca agggcaaggc cacattgact gtagacgaat cctccagcac agcctacatg
300
cagctcacca gcctgacatc tgaggactct gcggtctatt actgtgcaag atcccctttc
360
tacaatagta gaggggggaa ctactttgac tactggggcc aaggcaccac tctcacagtc
420
tcctca
426
<210> 22
<211> 142
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 22
Met Arg Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val Asn Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Met
20 25 30
Pro Gly Ala Ser Ala Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Leu
35 40 45
Thr Asp Tyr Trp Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Ala Ile Asp Ile Ser Asp Ser Tyr Ser Ser Tyr Asn
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Glu Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Thr Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Ser Pro Phe Tyr Asn Ser Arg Gly Gly Asn Tyr
115 120 125
Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
130 135 140
<210> 23
<211> 10
<212> Белок
<213> Mus
<400> 23
Gly Tyr Thr Leu Thr Asp Tyr Trp Met His
1 5 10
<210> 24
<211> 17
<212> Белок
<213> Mus
<400> 24
Ala Ile Asp Ile Ser Asp Ser Tyr Ser Ser Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 25
<211> 14
<212> Белок
<213> Mus
<400> 25
Ser Pro Phe Tyr Asn Ser Arg Gly Gly Asn Tyr Phe Asp Tyr
1 5 10
<210> 26
<211> 384
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 26
atgatgtcct ctgctcagtt ccttggtctc ctgttgctct gttttcaagg taccagatgt
60
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagagtcacc
120
atcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca
180
gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca
240
aggttcagtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa
300
gaagatattg ccacttactt ttgccaacag ggtgatacgc ttcctccgtg gacgttcggt
360
ggaggcacca agctggaaat caaa
384
<210> 27
<211> 128
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 27
Met Met Ser Ser Ala Gln Phe Leu Gly Leu Leu Leu Leu Cys Phe Gln
1 5 10 15
Gly Thr Arg Cys Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser
20 25 30
Ala Ser Leu Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp
35 40 45
Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val
50 55 60
Lys Leu Leu Ile Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser
85 90 95
Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asp
100 105 110
Thr Leu Pro Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
115 120 125
<210> 28
<211> 11
<212> Белок
<213> Mus
<400> 28
Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 29
<211> 7
<212> Белок
<213> Mus
<400> 29
Tyr Thr Ser Arg Leu His Ser
1 5
<210> 30
<211> 10
<212> Белок
<213> Mus
<400> 30
Gln Gln Gly Asp Thr Leu Pro Pro Trp Thr
1 5 10
<210> 31
<211> 447
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 31
atgggatgga cctggatctt tctcttcttc ctgtcaggaa ctgcaggtgt cctctctgag
60
gtcctgctgc tacagtctgg acctgaactg gtgaagcctg ggacttcagt gaaaatcccc
120
tgcaaggctt ctggatacac attcactgac tacaacgtgg actgggtgaa gcagcgccat
180
ggaaagggcc ttgagtggat tggagatatt aatccaaaca atggtggtac tatctacagt
240
cagaaattca agggcaaggc cacattgact gttgacaagt cctccagcac agccttcatg
300
gagctccgca gcctgacatc tgaggacact gcagtctatt tctgtgcaag gaactatagg
360
tggtttggtg ctatggacca ctggggtcaa ggaacctcag tcaccgtctc ctcagccaaa
420
acaacagccc catcggtcta tccactg
447
<210> 32
<211> 138
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 32
Met Gly Trp Thr Trp Ile Phe Leu Phe Phe Leu Ser Gly Thr Ala Gly
1 5 10 15
Val Leu Ser Glu Val Leu Leu Leu Gln Ser Gly Pro Glu Leu Val Lys
20 25 30
Pro Gly Thr Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asp Tyr Asn Val Asp Trp Val Lys Gln Arg His Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Asp Ile Asn Pro Asn Asn Gly Gly Thr Ile Tyr Ser
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Phe Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val
100 105 110
Tyr Phe Cys Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp
115 120 125
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
130 135
<210> 33
<211> 5
<212> Белок
<213> Mus
<400> 33
Asp Tyr Asn Val Asp
1 5
<210> 34
<211> 17
<212> Белок
<213> Mus
<400> 34
Asp Ile Asn Pro Asn Asn Gly Gly Thr Ile Tyr Ser Gln Lys Phe Lys
1 5 10 15
Gly
<210> 35
<211> 10
<212> Белок
<213> Mus
<400> 35
Asn Tyr Arg Trp Phe Gly Ala Met Asp His
1 5 10
<210> 36
<211> 547
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 36
atggagacag acacaatcct gctatgggtg ctgctgctct gggttccagg ttccactggt
60
gacattgtgt tgacccaatc tccagcttct ttggctgtgt ctccagggca gagggccacc
120
atttcctgca aggccagtca aagtcttgat tatgaaggtg atagtgatat gaattggtac
180
caacagaaac caggacagcc acccagactc ctcatctctg gtgcatccaa tctagagtct
240
gggatcccag ccaggttcag tggcagtggg tctgggacag acttcactgt taacatccat
300
cctgtggagg aggaggatgc tgcaacctat tactgtcagc aaagtactga ggatcctcgg
360
acgttcggtg gaggcaccaa gctggaaatc aaacgggctg atgctgcacc aactgtatcc
420
atcttcccac catccagtga gcagttaaca tctggaggtg cctcagtcgt gtgcttcttg
480
aacaacttct accccaaaga catcaatgtc aagtggaaga ttgatggcag tgaacgacaa
540
aatggcg
547
<210> 37
<211> 131
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 37
Met Glu Thr Asp Thr Ile Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala
20 25 30
Val Ser Pro Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser
35 40 45
Leu Asp Tyr Glu Gly Asp Ser Asp Met Asn Trp Tyr Gln Gln Lys Pro
50 55 60
Gly Gln Pro Pro Arg Leu Leu Ile Ser Gly Ala Ser Asn Leu Glu Ser
65 70 75 80
Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
85 90 95
Val Asn Ile His Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys
100 105 110
Gln Gln Ser Thr Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Leu
115 120 125
Glu Ile Lys
130
<210> 38
<211> 15
<212> Белок
<213> Mus
<400> 38
Lys Ala Ser Gln Ser Leu Asp Tyr Glu Gly Asp Ser Asp Met Asn
1 5 10 15
<210> 39
<211> 7
<212> Белок
<213> Mus
<400> 39
Gly Ala Ser Asn Leu Glu Ser
1 5
<210> 40
<211> 9
<212> Белок
<213> Mus
<400> 40
Gln Gln Ser Thr Glu Asp Pro Arg Thr
1 5
<210> 41
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 41
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Ser Leu Asp Tyr Glu
20 25 30
Gly Asp Ser Asp Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Pro Pro
35 40 45
Gln Leu Leu Ile Tyr Gly Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 42
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 42
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Asn Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 43
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 43
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Ser Leu Asp Tyr Glu
20 25 30
Gly Asp Ser Asp Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Pro Pro
35 40 45
Gln Leu Leu Ile Tyr Gly Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 44
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 44
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Ser Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 45
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 45
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Ser Leu Asp Tyr Glu
20 25 30
Gly Asp Ser Asp Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Pro Pro
35 40 45
Gln Leu Leu Ile Tyr Gly Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 46
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 46
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Asp Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 47
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 47
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Ser Leu Asp Tyr Glu
20 25 30
Gly Asp Ser Asp Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Pro Pro
35 40 45
Gln Leu Leu Ile Tyr Gly Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 48
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 48
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Gln Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 49
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 49
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Ser Leu Asp Tyr Glu
20 25 30
Gly Asp Ser Asp Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Pro Pro
35 40 45
Gln Leu Leu Ile Tyr Gly Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 50
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 50
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Asn Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 51
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 51
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Ser Leu Asp Tyr Glu
20 25 30
Gly Asp Ser Asp Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Pro Pro
35 40 45
Gln Leu Leu Ile Tyr Gly Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 52
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 52
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Asp Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 53
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 53
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Ser Leu Asp Tyr Glu
20 25 30
Gly Asp Ser Asp Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Pro Pro
35 40 45
Gln Leu Leu Ile Tyr Gly Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 54
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 54
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Ser Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 55
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 55
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Ser Leu Asp Tyr Glu
20 25 30
Gly Asp Ser Asp Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Pro Pro
35 40 45
Gln Leu Leu Ile Tyr Gly Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 56
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 56
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Gln Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 57
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 57
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ala Ser Gln Ser Leu Asp Tyr Glu
20 25 30
Gly Asp Ser Asp Met Asn Trp Tyr Leu Gln Lys Pro Gly Gln Pro Pro
35 40 45
Gln Leu Leu Ile Tyr Gly Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile Ser
65 70 75 80
Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 58
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированная мышь
<400> 58
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Gly Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Tyr Arg Trp Phe Gly Ala Met Asp His Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 59
<211> 17
<212> Белок
<213> Mus
<400> 59
Asp Ile Asn Pro Asn Asn Gly Gly Thr Ile Tyr Ala Gln Lys Phe Gln
1 5 10 15
Glu
<210> 60
<211> 17
<212> Белок
<213> Mus
<400> 60
Asp Ile Asn Pro Asn Ser Gly Gly Thr Ile Tyr Ala Gln Lys Phe Gln
1 5 10 15
Glu
<210> 61
<211> 17
<212> Белок
<213> Mus
<400> 61
Asp Ile Asn Pro Asn Asp Gly Gly Thr Ile Tyr Ala Gln Lys Phe Gln
1 5 10 15
Glu
<210> 62
<211> 17
<212> Белок
<213> Mus
<400> 62
Asp Ile Asn Pro Asn Gln Gly Gly Thr Ile Tyr Ala Gln Lys Phe Gln
1 5 10 15
Glu
<210> 63
<211> 17
<212> Белок
<213> Mus
<400> 63
Asp Ile Asn Pro Asn Gly Gly Gly Thr Ile Tyr Ala Gln Lys Phe Gln
1 5 10 15
Glu
<210> 64
<211> 17
<212> Белок
<213> Mus
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (6)..(6)
<223> X представляет собой D,N,S или Q
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (12)..(12)
<223> X представляет собой A или S
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (16)..(16)
<223> X представляет собой Q или K
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (17)..(17)
<223> X представляет собой E или G
<400> 64
Asp Ile Asn Pro Asn Xaa Gly Gly Thr Ile Tyr Xaa Gln Lys Phe Xaa
1 5 10 15
Xaa
<210> 65
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<220>
<221> прочий_признак
<222> (1)..(1)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> прочий_признак
<222> (3)..(3)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> прочий_признак
<222> (5)..(5)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> прочий_признак
<222> (13)..(19)
<223> Xaa может быть любой природной аминокислотой
<400> 65
Xaa Ser Xaa Ser Xaa Asp Tyr Glu Gly Asp Ser Asp Xaa Xaa Xaa Xaa
1 5 10 15
Xaa Xaa Xaa
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ LILRB1 И ЕГО ПРИМЕНЕНИЯ | 2021 |
|
RU2813373C1 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| ВЫСОКОАФФИННЫЕ АНТИ-PD-1 И АНТИ-LAG-3 АНТИТЕЛА И ПОЛУЧЕННЫЕ ИЗ НИХ БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2782381C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| CD131-СВЯЗЫВАЮЩИЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773927C2 |
| АНТИ-LILRB1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2801535C1 |
| ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2018 |
|
RU2810094C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2822200C2 |
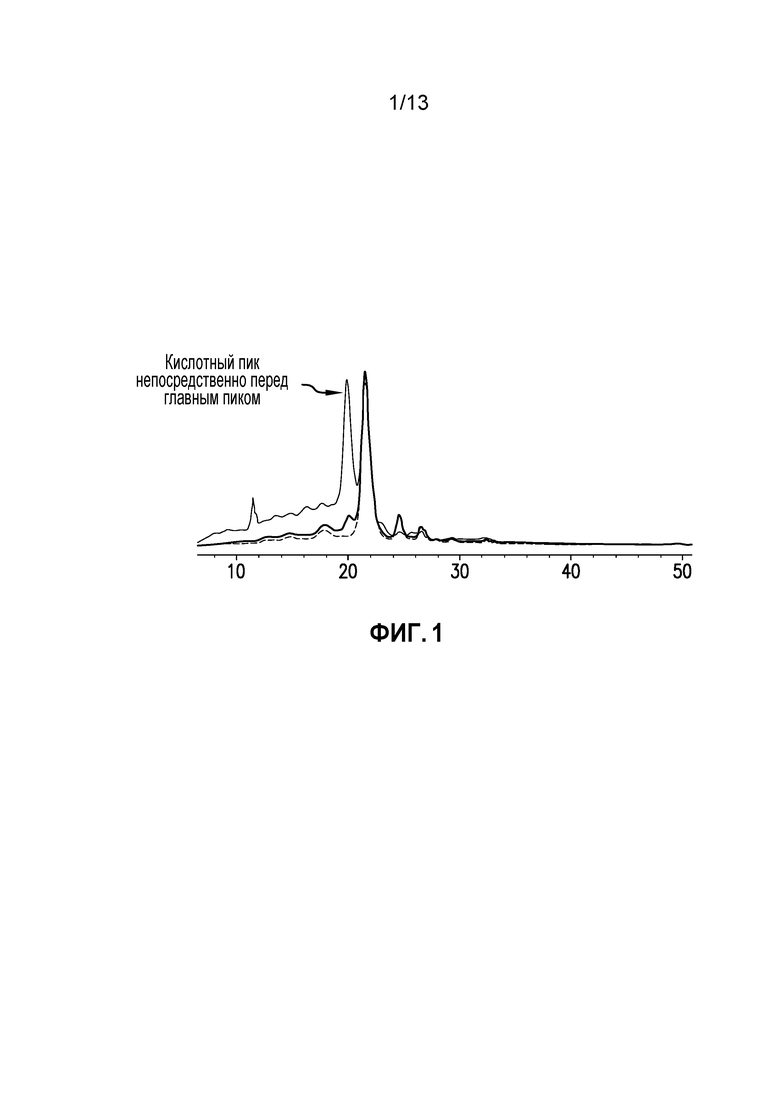
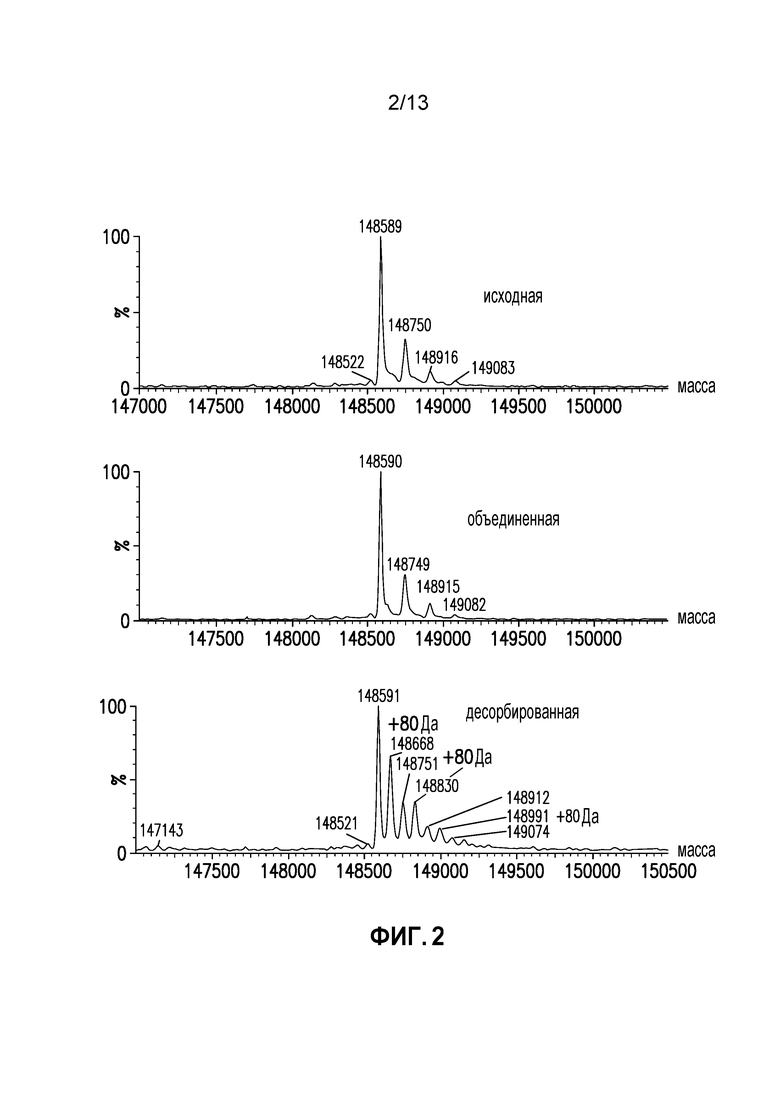
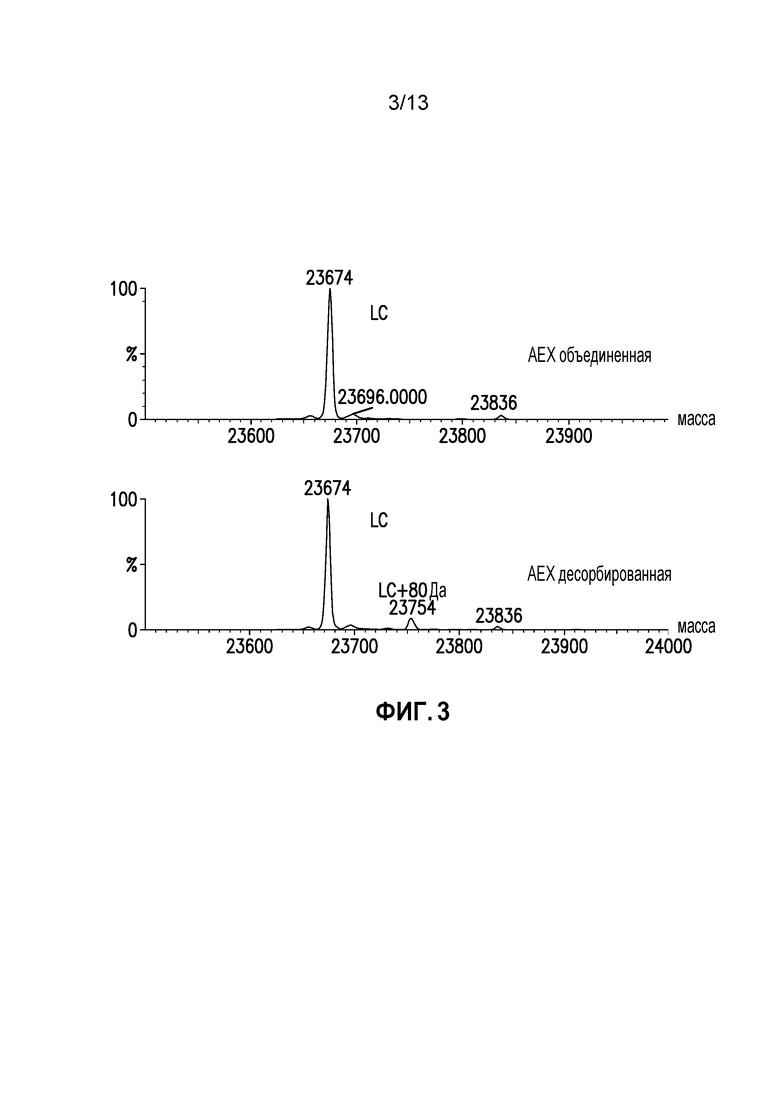
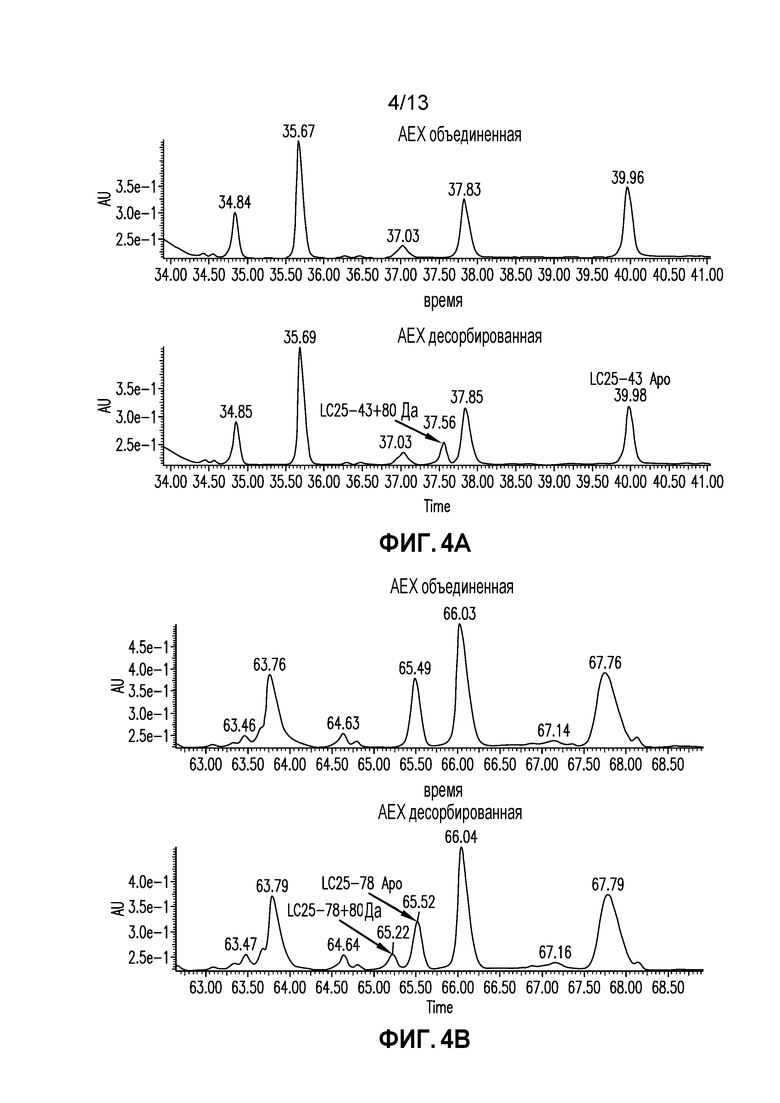
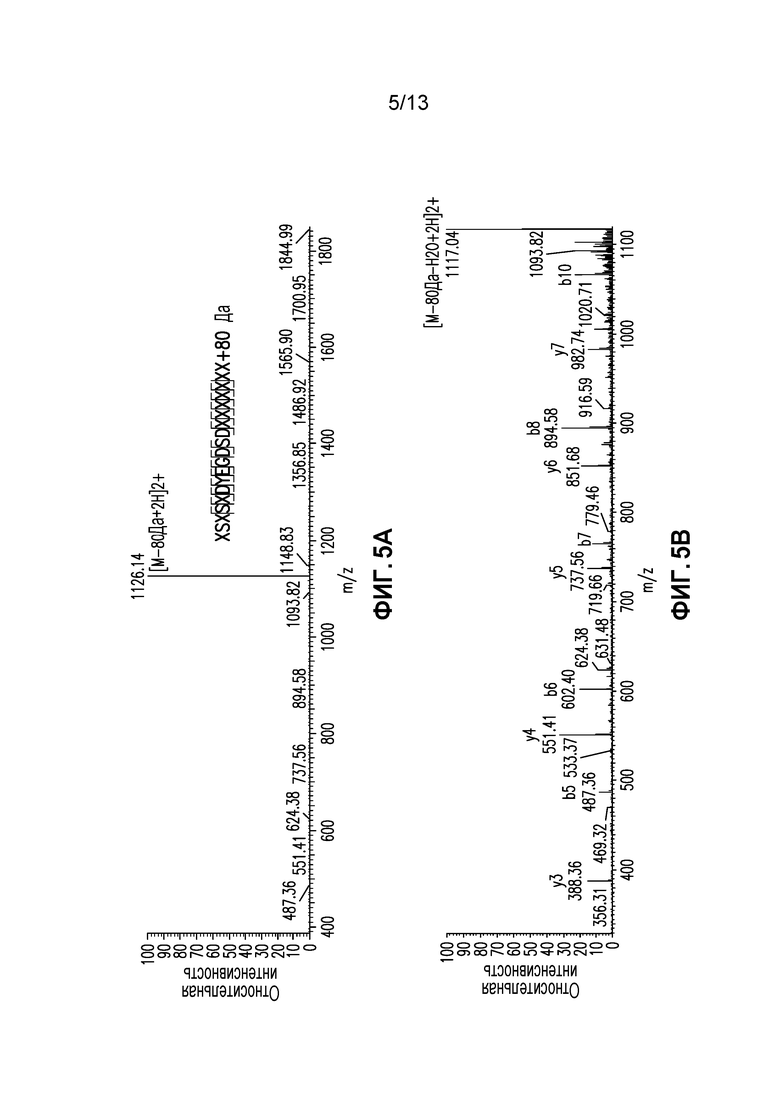
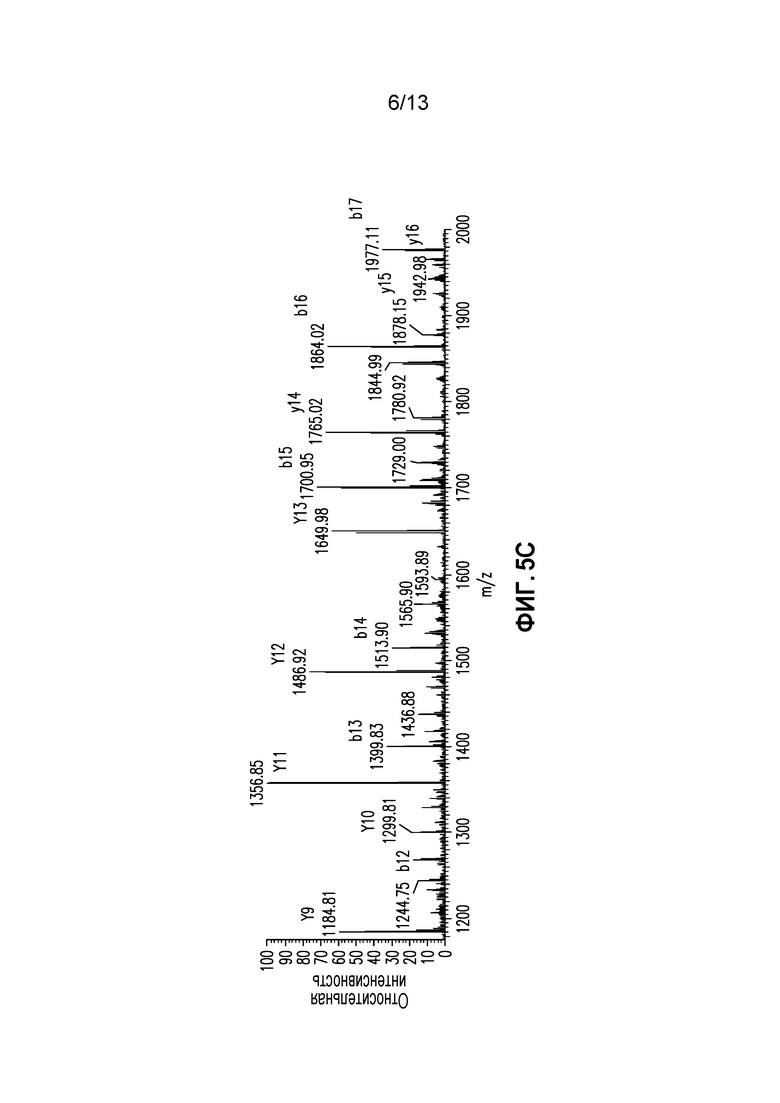
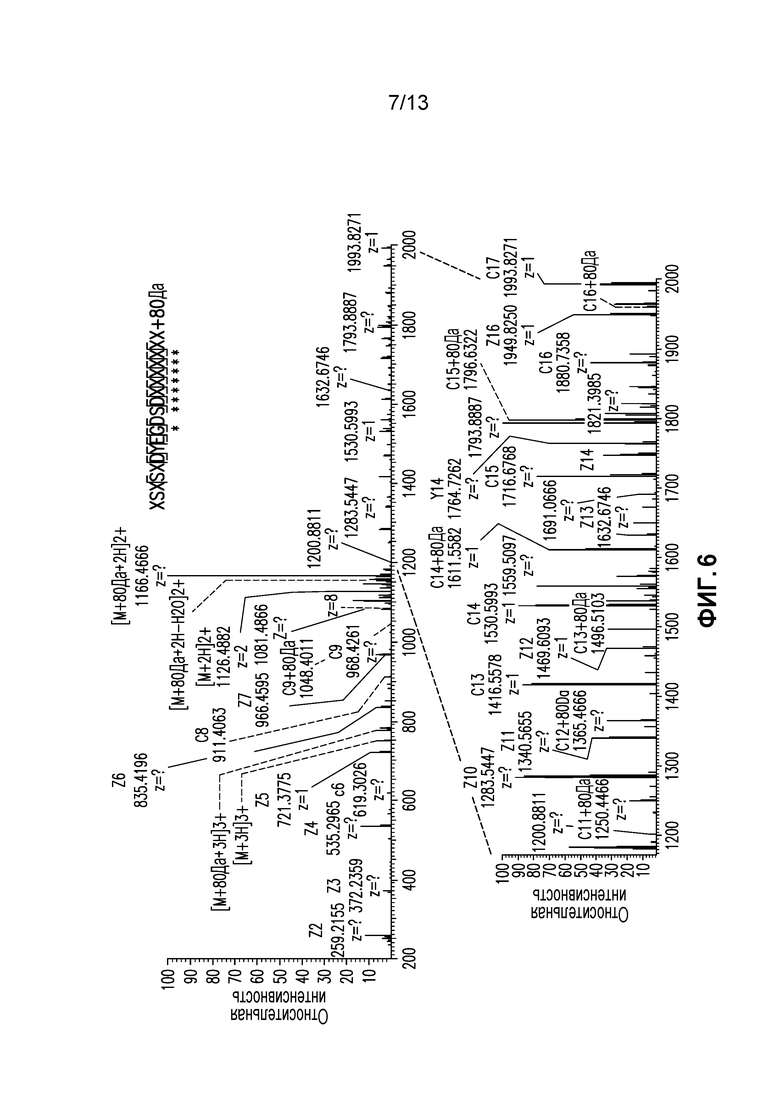
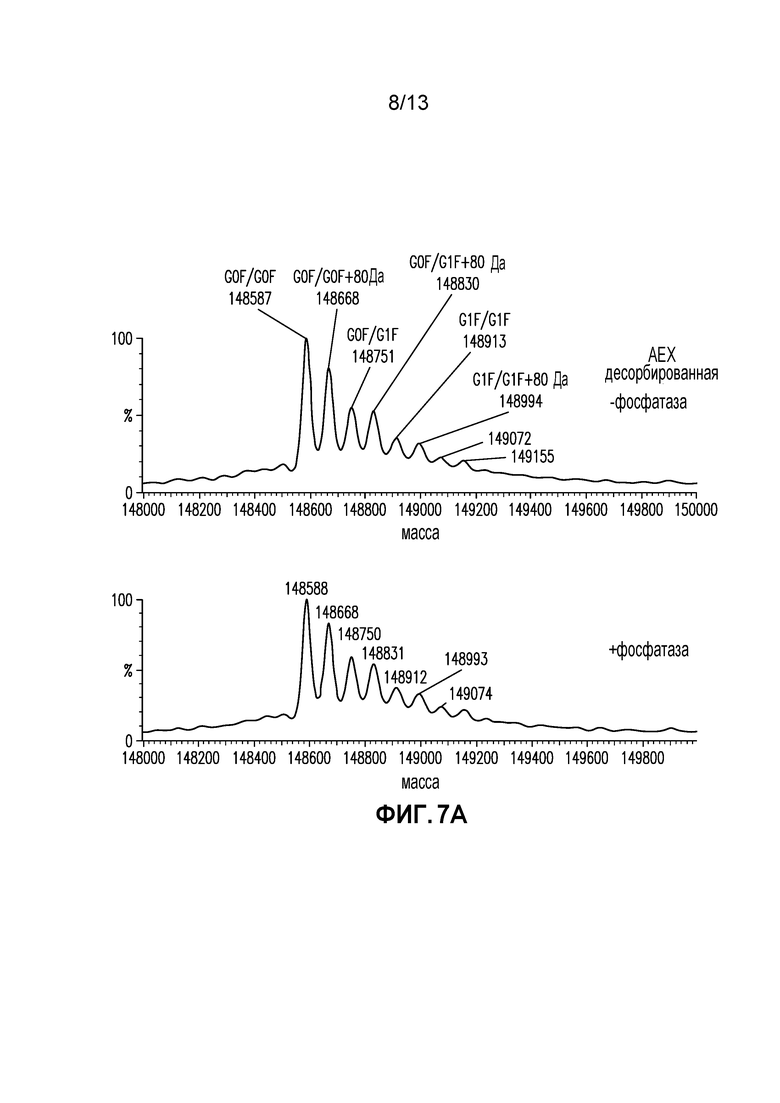
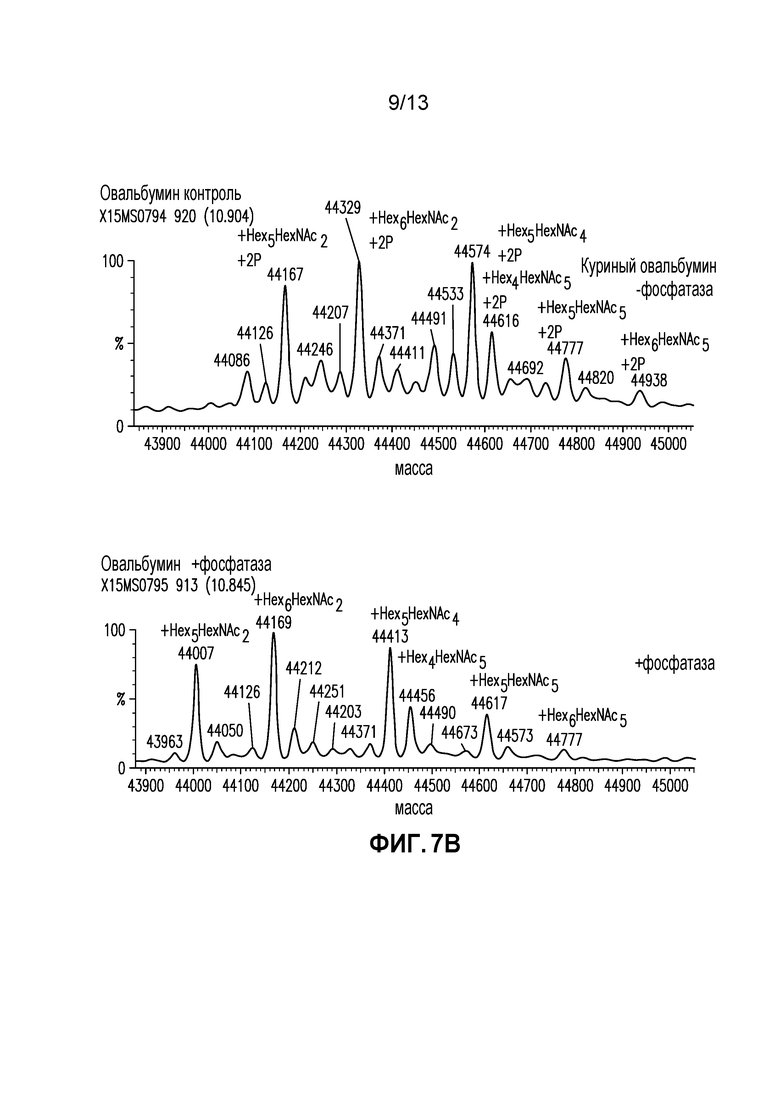
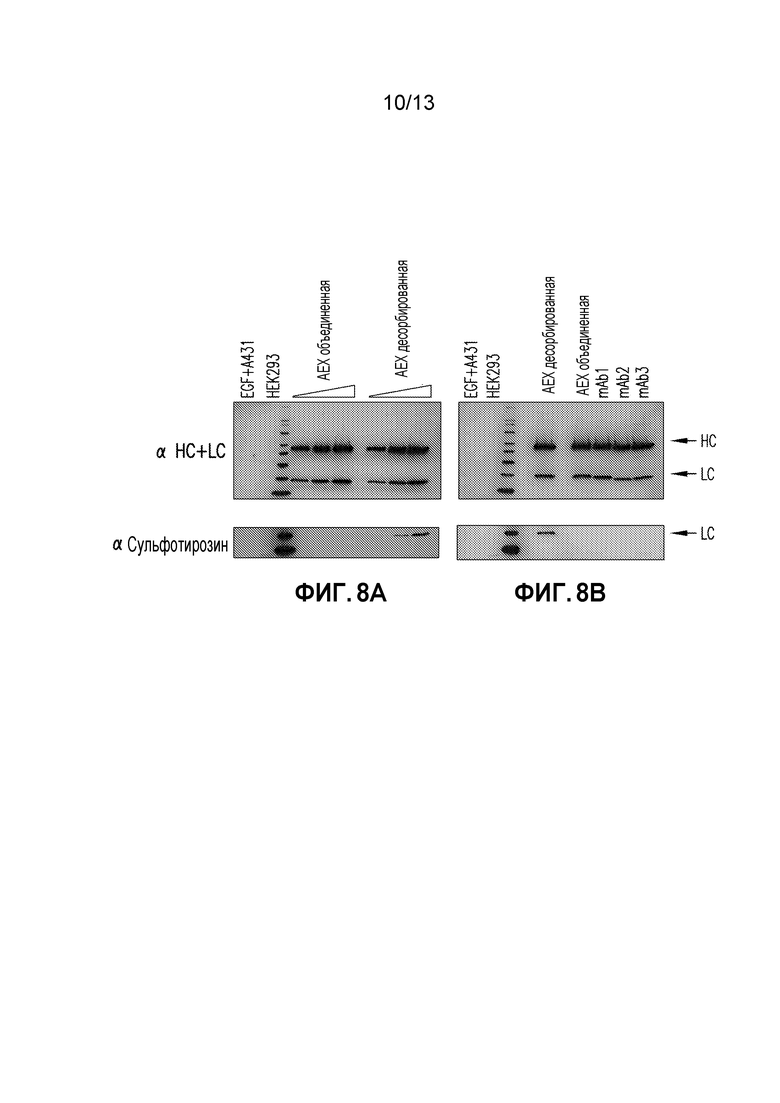
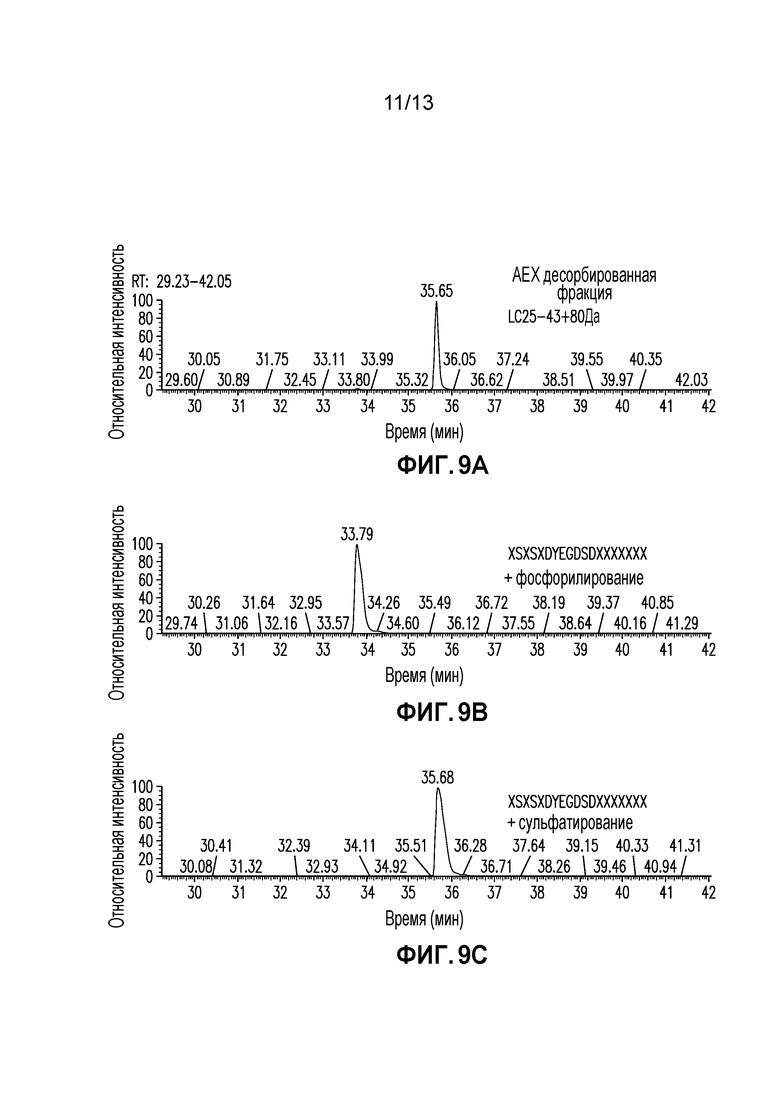
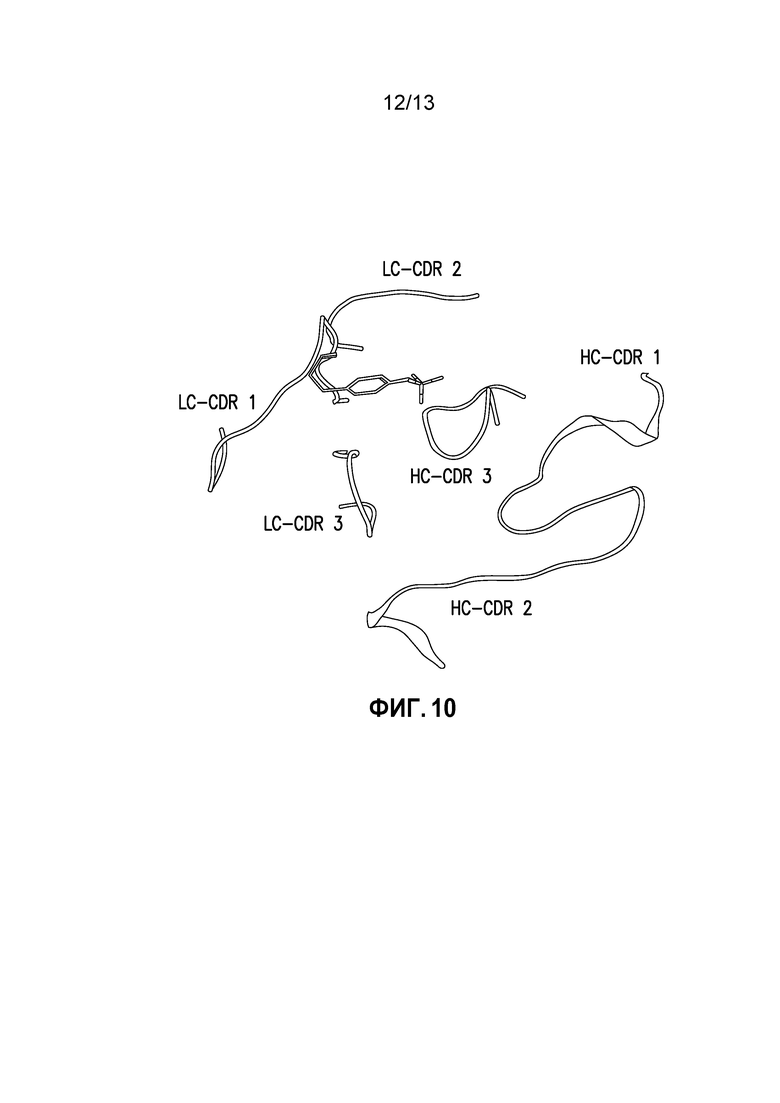
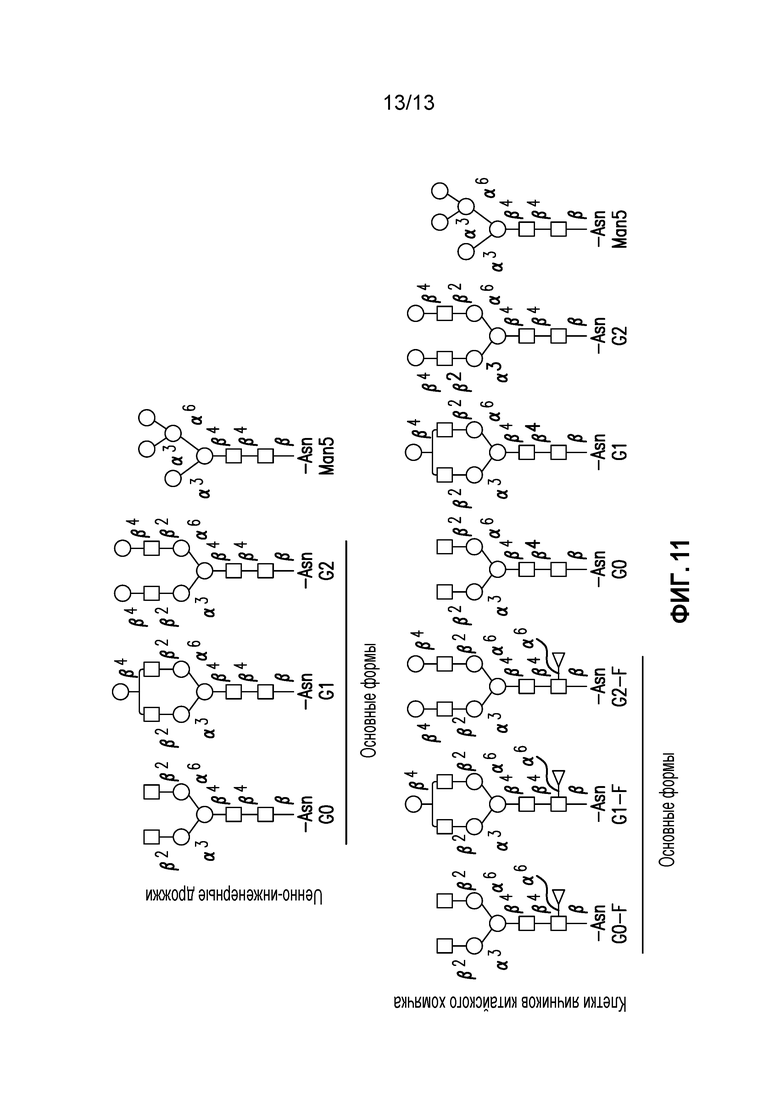
Изобретение относится к области биотехнологии. Предложен способ удаления антитела с сульфатированным тирозином из водной смеси, включающей антитело, в котором тирозин является сульфатированным, и антитело, в котором тирозин не является сульфатированным. Указанные антитела представляют собой антитела против LAG3 человека. Сульфатирование тирозина имеет место в CDR-L1 области. Также предложена композиция для терапии, включающая антитела против LAG3 человека. Композиция имеет менее чем 0,5% анти-hLAG3 антител, которые содержат сульфатированный тирозин на CDR-L1. Изобретение обеспечивает способ для очистки композиции антител от загрязняющих ее вариантов антитела, включающих сульфатированный тирозин. 2 н. и 12 з.п. ф-лы, 18 ил., 1 табл., 1 пр.
1. Способ для удаления антитела с сульфатированным тирозином из водной смеси, включающей антитело, в котором тирозин является сульфатированным, и антитело, в котором тирозин не является сульфатированным, включающий доведение pH смеси до 6,5 - 7,5, контактирование водной смеси с анионообменной смолой и сохранение не связанной со смолой водной фракции смеси из смолы, где антитело, в котором тирозин является сульфатированным, или антитело, в котором тирозин не является сульфатированным, представляют собой антитела против LAG3 человека, которые содержат аминокислотную последовательность тяжелой цепи SEQ ID NO: 52 и аминокислотную последовательность легкой цепи SEQ ID NO: 51; или аминокислотную последовательность тяжелой цепи SEQ ID NO: 52 и аминокислотную последовательность легкой цепи SEQ ID NO: 51 с N-концевой глутамином в тяжелой цепи, преобразованным в пироглутамат, и/или удаленным C-концевым лизином в тяжелой цепи; и где сульфатирование тирозина имеет место в CDR-L1 области.
2. Способ по п. 1, где стадия доведения pH смеси включает доведение рН смеси до 6,5 - 7,0.
3. Способ по п. 1, включающий уравновешивание анионообменной хроматографической смолы, которая включает анионообменный лиганд, в хроматографической колонке раствором фосфата натрия 10-50 мМ pH 6,5-7,5, доведение pH водной смеси до 6,5-7,5, нанесение смеси на колонку, сбор проточной фракции из колонки, промывку смолы в колонке раствором фосфата натрия 10-50 мМ pH 6,5-7,5 и сбор проточной фракции из промывки.
4. Способ по п. 1 или 2, который дополнительно включает промывку смолы водной композицией в изократических условиях и удаление и сохранение промывочной композиции из смолы.
5. Способ по п. 2, включающий уравновешивание анионообменной хроматографической смолы, которая включает диметиламинопропильный анионообменный лиганд, в хроматографической колонке раствором 25 мМ фосфата натрия pH 6,5, доведение pH водной смеси до 6,5, нанесение смеси на колонку, сбор проточной фракции из колонки, промывку смолы в колонке раствором 25 мМ фосфата натрия pH 6,5 и сбор проточной фракции из промывки.
6. Способ по п. 2, включающий уравновешивание анионообменной хроматографической смолы, которая включает кватернизированный полиэтилениминовый анионообменный лиганд, в хроматографической колонке раствором 25 мМ фосфата натрия и 5 мМ NaCl pH 7,0, доведение pH водной смеси до 7,0, нанесение смеси на колонку, сбор проточной фракции из колонки, промывку смолы в колонке раствором 25 мМ фосфата натрия и 5 мМ NaCl pH 7,0 и сбор проточной фракции из промывки.
7. Способ по любому из пп. 1-6, где осуществляют сбор проточной фракции анионообменной хроматографии при A280 поглощении, когда A280 сначала достигает по меньшей мере 2,5 единиц оптической плотности/см, и продолжают до тех пор, пока A280 не падает ниже 1,0 единиц оптической плотности/см.
8. Способ по любому из пп. 1-7, дополнительно включающий очистку антитела или антигенсвязывающего фрагмента катионообменной хроматографией, анионообменной хроматографией в режиме связывания-элюирования, хроматографией гидрофобных взаимодействий, хроматографией с протеином-A, хроматографией с протеином-L, хроматографией с протеином-G, хроматографией на гидроксиапатите, эксклюзионной хроматографией, фракционированным осаждением, фильтрованием, центрифугированием или вирусной инактивацией.
9. Способ по любому из пп. 1-8, где легкие цепи и тяжелые цепи иммуноглобулинов антитела фрагментов экспрессированы в клетке яичника китайского хомячка.
10. Способ по любому из пп. 1-8, где антитело, в котором тирозин не является сульфатированным, представляет собой антитело, имеющее молекулярную массу 148590 Да, 148752 Да и/или 148914 Да.
11. Композиция для терапии, включающая антитело, которое специфически связывается с геном 3 активации лимфоцитов человека (hLAG3) (анти-hLAG3), включающее: аминокислотную последовательность тяжелой цепи SEQ ID NO: 52 и аминокислотную последовательность легкой цепи SEQ ID NO: 51, где анти-hLAG3 антитело экспрессировано в клетках яичников китайского хомячка; и композиция имеет менее чем 0,5% анти-hLAG3 антител, которые содержат сульфатированный тирозин на CDR-L1, по сравнению с анти-hLAG3 антителами, в которых тирозин не является сульфатированным.
12. Композиция по п. 11, где анти-hLAG3 антитело, которое не содержит сульфатированный тирозин, представляет собой разновидности гликозилированного антитела, имеющие молекулярные массы около 148590 Да, 148752 Да или 148914 Да.
13. Композиция по п. 11, где анти-hLAG3 антитело имеет N-концевой глутамином в тяжелой цепи, преобразованный в пироглутамат, и/или удаленный C-концевой лизин в тяжелой цепи.
14. Композиция по любому из пп. 11-13, где анти-hLAG3 антитела, которые содержат сульфатированный тирозин на CDR-L1, или анти-hLAG3 антитела, которые не содержат сульфатированный тирозин, определяют масс-спектроскопией.
| WO 2016028672 A1, 25.02.2016 | |||
| HONGCHENG LIU et al., In vitro and in vivo modifications of recombinant and human IgG antibodies, MABS, 2014, vol | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| WO 2016118707 A1, 28.07.2016 | |||
| УСТРОЙСТВО ДЛЯ ИСПАРИТЕЛЬНОГО ОХЛАЖДЕНИЯ ЦИЛИНДРОВ ДВИГАТЕЛЕЙ ВНУТРЕННЕГО ГОРЕНИЯ | 1930 |
|
SU23032A1 |
| РАСТВОРИМЫЕ ПОЛИПЕПТИДНЫЕ ФРАКЦИИ ПРОТЕИНА LAG-3, СПОСОБ ПОЛУЧЕНИЯ, ТЕРАПЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, АНТИТЕЛО | 1995 |
|
RU2178306C2 |

