Изобретение в области биотехнологии относится к созданию новой системы трансформации для генетической инженерии грибных продуцентов промышленных ферментов. Изобретение имеет отношение к микробиологической и текстильной промышленности и включает в себя: а) - мицелиальный гриб Penicillium canescens в качестве хозяина; б) - фрагмент ДНК мицелиального гриба Penicillium verruculosum, кодирующий синтез секретируемого фермента эндоглюканазы III; в) - промоторные районы и сигнальные пептиды генов бета-галактозидазы и ксиланазы Penicillium canescens, использованные для реализации высокого регуляторного транскрипционного потенциала этих генов в экспрессии гетерологичного гена эндоглюканазы III Penicillium verruculosum в штамме-хозяине Penicillium canescens и г) - группа штаммов Penicillium canescens, сконструированных методами трансформации и генетической инженерии на основе этого фрагмента ДНК и синтезирующих эндоглюканазу III Penicillium verruculosum в промышленных количествах.
Природный продуцент секретируемой кислой бета-галактозидазы мицелиальный гриб Penicillium canescens (ВКПМ F-178) в процессе роста секретирует в культуральную жидкость группу белков с ферментативной активностью. Среди секретируемых ферментов, которые накапливаются в культуральной жидкости при глубинной ферментации этого гриба бета-галактозидаза и ксиланаза занимают около половины всего секретируемого белка (Николаев И.В. и др., 1992. Биохимия, т.57: 873-879).
Геном Penicillium canescens содержит одну копию гена секретируемой бета-галактозидазы (Николаев И.В. и др. Молекулярная биология. 1992, т.26: 869-875). Гены бета-галактозидазы и ксиланазы включают в себя нуклеотидную последовательность ДНК, кодирующую структурную часть гена, промотор (5'-нетранслируемую область ДНК), являющийся местом прикрепления и функционирования регуляторных белков и белков механизма транскрипции, 3'-нетранслируемую область терминатора. Структурные части генов состоят из последовательности, кодирующей лидерный пептид из 39 аминокислот у бета-галактозидазы и 25 аминокислот у ксиланазы, и последовательности, кодирующей зрелый белок. Регуляция активности как бета-галактозидазы, так и ксиланазы осуществляется путем индукции или репрессии ее синтеза на транскрипционном уровне. Индуцирующим началом является наличие в среде арабинозы. В промышленных условиях источником арабинозы является свекловичный жом, который входит в состав индустриальной среды как основной углеводный компонент (Николаев И.В., Винецкий Ю.П., 1998. Биохимия, т.63: 1523-1528)
Промоторы генов бета-галактозидазы и ксиланазы Penicillium canescens относятся к немногочисленной группе сильных грибных промоторов, которые могут быть использованы в экспрессионных конструкциях для синтеза гетерологичных белков. Под контролем промотора гена бета-галактозидазы в условиях индукции в клетках гриба Aspergillus nidulans в следовых количествах была синтезирована люцифераза светлячка и на транскрипционном уровне была обнаружена экспрессия гена интерлейкина-6 человека (Zakharova E.S. et al,1992. Int. Conf. AIDS, cancer and human retroviruses, St.-Petersburg, p.36; Zakharova E.S., 1994. 2nh Europ.Conf.Fung. Genet., abstr.AS, Lunteren, The Netherlands).
Эндо-β-1,4-глюканазы (эндо-β-1,4-глюкан-D-гликозилгидролазы, КФ 3.2.1.4) или целлюлазы - это ферменты, обладающие гидролитической активностью по отношению к полиглюканам, имеющим β-1,4-гликозидные связи, к которым относятся природная нерастворимая кристаллическая целлюлоза - например, хлопковая и древесная целлюлоза; растворимые производные целлюлозы - например, карбоксиметилцеллюлоза и метилцеллюлоза; β-глюканы - например, β-глюкан овса и лихенин, а также некоторые другие полисахариды, например - ксиланы (Синицын А.П. и др. Биоконверсия лигноцеллюлозных материалов. М.: Изд-во Моск. ун-та, 1995). Под эндо-β-1,4-глюканазной активностью понимается способность фермента осуществлять гидролиз внутренних β-1,4-глюкозидных связей, удаленных от концов полимерных молекул целлюлозы с образованием целлоолигосахаридов (Синицын А.П. и др. Биоконверсия лигноцеллюлозных материалов. М.: Изд-во Моск. ун-та, 1995). По гомологиям последовательностей аминокислот эндоглюканазы, а также другие целлюлазы и гликозил-гидролазы подразделяются на более чем 70 семейств (SWISS-POT Protein Sequence Data Bank, http://www.expasy.ch/cgi-bin/lists.glycosid).
Грибные секретируемые эндоглюканазы используются в текстильной промышленности, например, для частичного удаления индиго из джинсовой ткани (для получения так называемых "вареных" джинсов"), биополировки хлопчатобумажной ткани и предотвращения пиллинга (Gusakov A.V. et al, 2000. Appl. Biochem. Biotechnol. v.88: 119-126).
Эндоглюканаза III Penicillium verruculosum относится к 12 семейству гликозил-гидролаз. Молекулярная масса эндоглюканазы III Penicillium verruculosum составляет 24,213 кДа, pI=5,4, рН-оптимум 3,5-4,5 (при 50°С) температурный оптимум 45-55°С (при рН 4,0). Фермент представлен единой полипептидной цепью из 222 аминокислотных остатков, состоит только из каталитического домена и не имеет целлюлозо-связывающего домена.
Эндоглюканаза III Penicillium verruculosum имеет специфическую активность по отношению к карбоксиметилцеллюлозе (КМЦ-азную активность), равную 25 ед/мг белка (КМЦ-азную активность определяли по начальной скорости образования восстанавливающих сахаров при гидролизе КМЦ при рН 4,5 и 50°С, восстанавливающие сахара определяли с помощью метода Нельсона-Шомоди, метод определения КМЦ-азной активности приведен в Синицын А.П.и др. Биоконверсия лигноцеллюлозных материалов. М.: Изд-во Моск. ун-та, 1995) и специфическую активность к β-глюкану, равную 15 ед/мг белка (β-глюканазную активность определяли по начальной скорости образования восстанавливающих сахаров при гидролизе β-глюкана овса при рН 4,5 и 50°С, восстанавливающие сахара определяли с помощью метода Нельсона-Шомоди, метод определения к β-глюкану активности приведен в Синицын А.П. и др. Биоконверсия лигноцеллюлозных материалов. М.: Изд-во Моск. ун-та, 1995). Эндоглюканаза III Penicillium verruculosum не активна по отношению к микрокристаллической целлюлозе, ксиланау, п-нитрофенил-β-D-глюкозиду и п-нитрофенил-β-D-целлобиозиду.
Целлюлазный комплекс Penicillium verruculosum обладает высокой "текстильной" активностью, пригодной для получения "вареной" джинсовой ткани и биополировки хлопчатобумажных изделий, причем эндоглюканаза III является ключевым для осуществления этих "текстильных" функций ферментом Penicillium verruculosum и обладает крайне высокой способностью к получению "вареной" джинсовой ткани (причем характеризуется низкой величиной ресорбции индиго) и к биополировке хлопчатобумажных изделий. Ключевая роль эндоглюканазы III объясняется особенностями тунельной структуры ее активного центра, отсутствием целлюлозо-связывающего домен, а также наличием особого кластера гидрофобных аминокислот, экспонированных на поверхности ее белковой глобулы (Gusakov A.V., et al, 2000. Enz. Microb. Technol. v.27: 664-671).
Собственная эндоглюканазная активность штамма Penicillium canescens ВКПМ F178 весьма мала, порядка менее единицы активности на 1 мл культуральной жидкости, что позволяет использовать этот штамм как реципиент для включения и экспрессии гена эндоглюканазы III Penicillium verruculosum под контролем сильных промоторов генов бета-галактозидазы и ксиланазы.
В качестве ближайшего аналога заявляемого изобретения может быть рассмотрен фрагмент ДНК PCG 2.6 кодирующий синтез бета-галактозидазы Penicillium canescens и сконструированный на его основе штамм-продуцент бета-галактозидазы Penicillium canescens F-725 (Патент РФ N 2126049, Бюллетень изобретений РФ N 4, от 10.02.1999).
Задачей заявляемой группы изобретений является достижение высокой продуктивности синтеза эндоглюканазы III Penicillium verruculosum в реципиентном штамме Penicillium canescens под контролем промотора гена бета-галактозидазы, под контролем промотора гена ксиланазы, а так же под контролем промоторов этих генов одновременно.
Для решения задачи получают фрагмент ДНК pREG B-1 размером 11.7 тысяч пар нуклеотидов (т.п.н.), несущий полную структурную часть гена эндоглюканазы III, и создают экспрессионную плазмидную конструкцию. Для получения этой конструкции из фрагмента ДНК pREG B-1 выделяют фрагмент с нуклеотидной последовательностью, кодирующий зрелый белок и терминатор гена эндоглюканазы III, соединяют этот фрагмент с фрагментом ДНК гена бета-галактозидазы Penicillium canescens, несущий 5'-промоторную область и нуклеотидную последовательность, кодирующую лидерный пептид бета-галактозидазы, получают гибридный фрагмент ДНК и включают полученную конструкцию в векторную плазмиду. Далее путем котрансформации реципиентного штамма Penicillium canescens создают мультикопийный штамм первого поколения. Затем создают плазмиду, содержащую маркерный ген замещения, и путем трансформации мультикопийного штамма первого поколения включают маркерный ген в природный сайт бета-галактозидазы, вследствие чего полученный трансформант более не производит этот фермент. Далее путем серии генно-инженерных манипуляций создают вторую плазмиду замещения, с помощью которой при котрансформации вследствие гомологичной рекомбинации в промоторной и терминаторной областях ген эндоглюканазы замещает маркерный ген и увеличивает копийность в штамме первого поколения. В результате создают мультикопийные штаммы Penicillium canescens R201 и R201-151 второго поколения. В качестве конечного этапа в создании новой системы трансформации на основе регуляторных элементов грибного хозяина Penicillium canescens получают серию штаммов третьего поколения путем включения в штамм R201-151 добавочных копий гена эндоглюканазы III под контролем промотора гена ксиланазы. Среди серии штаммов третьего поколения селекционируют штамм, способный за 96-120 час ферментации на среде со свекловичным жомом и пептоном накапливать в культуральной жидкости 160-180 ед./мл эндоглюканазы Ш.
Конструирование семейства штаммов включало в себя несколько этапов:
Этап 1 - выделение фрагмента ДНК Penicillium verruculosum, кодирующего ген секретируемой эндоглюканазы с полной регуляторной областью и включение его в составе фрагмента ДНК в структуру векторной молекулы.
Этап 2 - состыковка выделенного гена с промоторной областью и участком, кодирующим лидерный пептид гена бета-галактозидазы Penicillium canescens.
Получение экспрессионной плазмиды pPrEG (фиг.2).
Этап 3 - трансформация штамма-реципиента Penicillium canescens PCA 10 (niaD-) экспрессионной плазмидой pPrEG с включенным геном эндоглюканазы и отбор трансформантов Penicillium canescens, секретирующих эндоглюканазу III Penicillium verruculosum. Селекция наиболее продуктивного штамма Penicillium canescens R201 первого поколения.
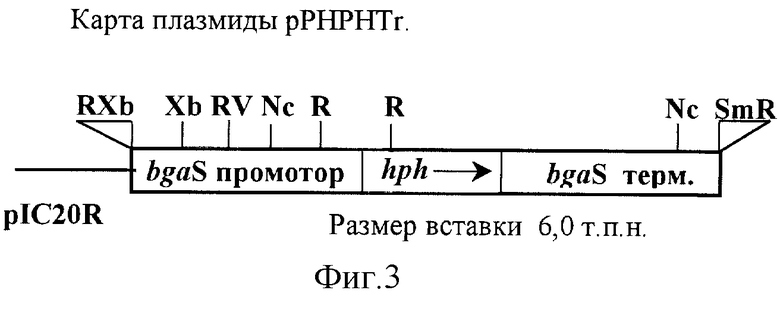
Этап 4 - конструкция плазмиды замещения рРrНРНТr, в которой структурную часть гена бета-галактозидазы заменяют геном устойчивости к гигромицину.
Этап 5 - трансформация штамма R201 плазмидой замещения рРrНРНТr и селекция штамма Penicillium canescens R201-55 с генотипом (bgaS-- hph), не синтезирующего бета-галактозидазу и устойчивого к гигромицину в условиях индукции.
Этап 6 - мутагенез клеток штамм Penicillium canescens R201-55 (bgaS-, hph) и селекция штамма - реципиента R201-55-15 (bgaS-, hph, niaD-).
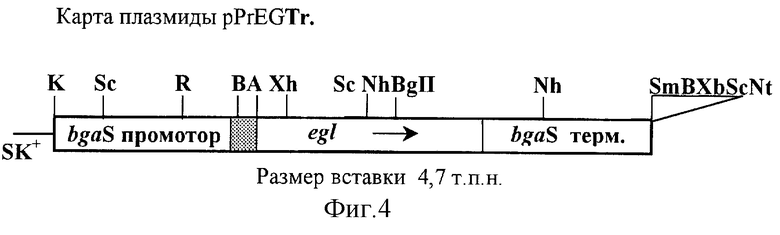
Этап 7 - включение дополнительных копий гена эндоглюканазы в природный сайт гена бета-галактозидазы в геноме Penicillium canescens R201-55-15 вместо гена устойчивости к гигромицину при котрансформации с помощью векторной плазмиды замещения pPrEGTr.
Этап 8 - селекция наиболее продуктивного штамма Penicillium canescens R201-151 (bgaS-, hph) второго поколения и его генотипический, фенотипический и физиолого-биохимический анализ.
Этап 9 - состыковка последовательности, кодирующей лидерный пептид гена бета-галактозидазы Penicillium canescens и гена эндоглюканазы III Penicillium verruculosum с промоторной областью гена ксиланазы Penicillium canescens.
Получение экспрессионной плазмиды pXEG (фиг.5).
Этап 10 - мутагенез клеток штамм Penicillium canescens R201-151 и селекция штамма - реципиента R201-151 (bgaS-, niaD-).
Этап 11 - включение дополнительных копий гена эндоглюканазы в штамм - реципиента R201-151 (bgaS--niaD-) при котрансформации с помощью экспрессионной плазмиды pXEG. Получение штамма третьего поколения R3020.
Этап 12 - выращивание штаммов R201 и R201-151 и R3020 на ферментационной среде в колбах и масштабирование процесса ферментации.
Полученные штаммы Penicillium canescens R201, R201-151 и R3020 характеризуются следующими культурально-морфологическими и физиолого-биохимическими признаками:
1. Культурально-морфологические признаки на различных питательных средах после 10 суток роста при 30°С:
а) Сусло-агар. Диаметр колоний 6,5-7,5 см. Обратная сторона оранжево-темно-коричневая. Конидиеносцы размером 430-450×3,0-3,2 мкм, конидии 2,0-2,2 мкм, шаровидные, в молодом возрасте гладкие, в старом - шероховатые, образуются в цепочках, соединяются в колонки, со временем распадающиеся.
б) Агаризованная среда Ролена-Тома с сахарозой. Диаметр колоний 38-48 мм. Колонии войлочные или слабо пушистые в центре, слабовыпуклые, с радиальными бороздками, белого цвета, по краям окрашены в серо-голубоватый тон, края неровные, лопастные, обратная сторона серо-бежевого тона.
в) Агаризованная среда Чапека с глюкозой. Диаметр колоний 2,8-4,0 см. Колонии серые, складчатые, с ровным краем, конидиеобразование интенсивное, цвет конидий светло-серый. Обратная сторона колоний с радиальными складками слабо-кремового тона.
По культурально-морфологическим признакам, перечисленным выше, не отмечено существенных отличий от природного штамма Penicillium canescens ВКПМ F-178 и от рекомбинантного штамма Penicillium canescens ВКПМ F-725 - продуцента бета-галактозидазы.
2. Физиолого-биохимические признаки.
Мезофилы, растут при 25-37°С, оптимальная температура роста 29°С. Оптимум роста при значениях рН среды 4,1-5,8. Желатину разжижают слабо. Отношение к источникам углерода. Хорошо усваивают моно-, ди- и полисахариды, за исключением арабинозы, которую утилизируют слабо. Хорошо усваивают аммонийный и нитратный азот, пептон, мочевину. В процессе роста на среде, содержащей пептон, свекловичный жом и фосфорнокислый калий, активность секретируемой бета-галактозидазы у штамма Penicillium canescens R201 втрое меньше, чем у исходного штамма F-178 и составляет около 20 ед./мл, а активность штамма Penicillium canescens R201-151 в 20 раз меньше и составляет за то же время ферментации не более 1 ед./мл. Протеазная активность выражена в малой степени.
В результате включения гена эндоглюканазы Penicillium verruculosum у заявляемого штамма Penicillium canescens R201 эндоглюканазная активность при оптимальных условиях ферментации составляет 120 ед./мл культуральной жидкости, у штамма R201-151 и R3020 эндоглюканазная активность при тех же условиях составила 160 ед./мл и 180 ед./мл соответственно. Доля эндоглюканазы составляла 40-60% от суммарного секретируемого белка.
Изобретение иллюстрируется следующими примерами.
Пример 1. Получение фрагмента ДНК Penicillium verruculosum, кодирующего синтез эндоглюканазы III.
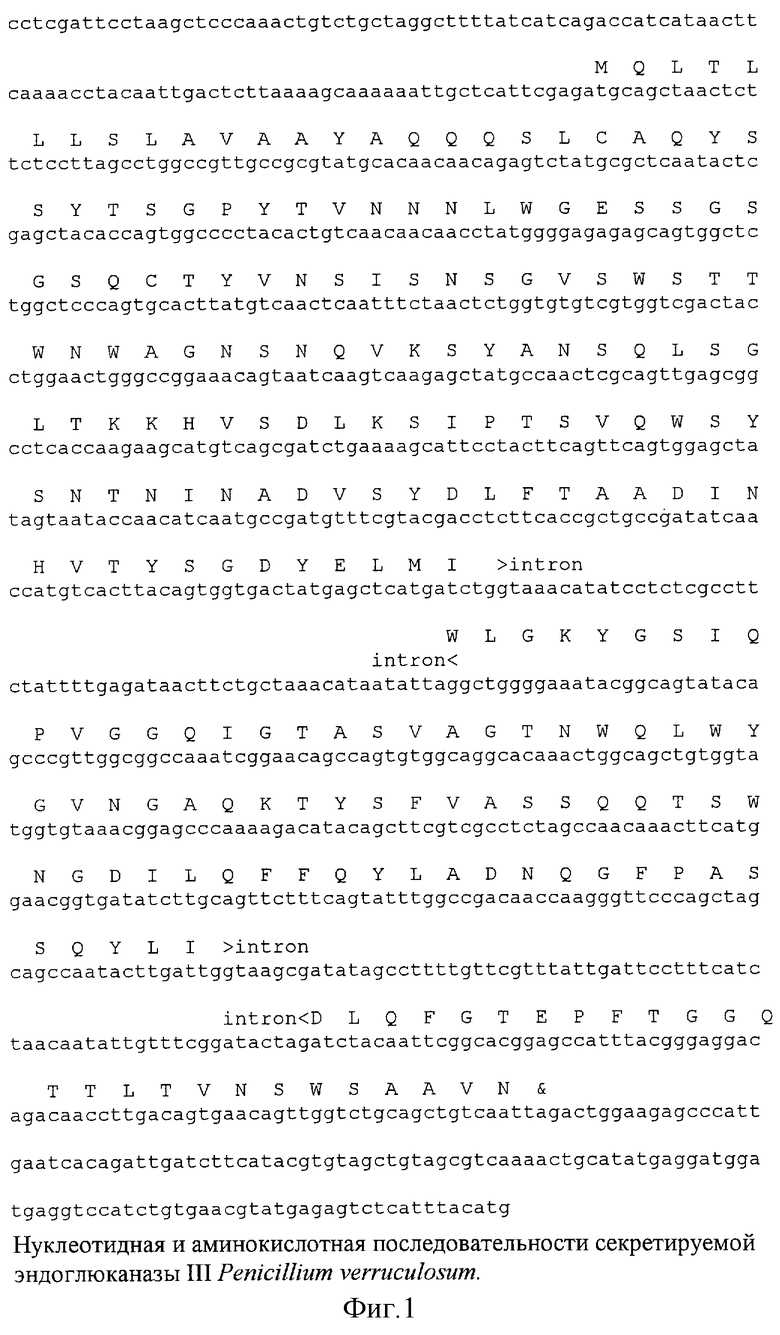
Для получения геномного банка ДНК этого штамма обрабатывают рестриктазой Sau3A в условиях неполного расщепления, фракционируют путем центрифугирования в градиенте плотности сахарозы, выделяя фрагменты ДНК размером 10-20 т.п.н., и включают путем лигирования в ДНК фагового вектора λGM12. Далее готовят упаковочную смесь (белки фаговых головок и белки хвостов фага лямбда по отдельности), проводят упаковку при оптимальных соотношениях компонентов смеси и полученные фаговые частицы с рекомбинантными ДНК рассевают на чашки Петри с питательной средой и индикаторными бактериями (Т. Маниатис, Э. Фрич, Дж.Самбрук. Молекулярное клонирование. М.: Мир, 1984). Полученные негативные колонии (независимые клоны) в количестве 6×104 в совокупности представляют собой банк генов пеницилла. Далее из этого банка генов выделяют фаговый клон, несущий фрагмент ДНК с геном эндоглюканазы с промоторным районом, несущим все сигнальные последовательности для регуляции ее синтеза.
Для решения этой задачи после исследования гомологии аминокислотных и нуклеотидных последовательностей эндоглюканаз грибов семейства 12 глюкозил гидролаз синтезируют два олигонуклеотидных праймера:
(1) CC(G/C)GTGAAIGGCTC(G/A)GTICC(A/G)AATTG
(2) GGIGACTA(T/C)GA(A/G)CT(T/C)ATGATITGG, где I - инозин.
Далее с помощью ПЦР (полимеразной цепной реакции) синтезируют внутренний фрагмент ДНК гена эндоглюканазы размером 0,4 т.п.н. Этот фрагмент клонируют в плазмидном векторе Bluescript SK и определяют частичную нуклеотидную последовательность. ПЦР-фрагмент ДНК используют для селекции фагового клона с геном эндоглюканазы. Для этой задачи проводят молекулярную гибридизацию на нейлоновых фильтрах с Р32 -меченым ПЦР-фрагментом. Среди 30 фаговых клонов, обнаруживавших позитивный сигнал с радиоактивным зондом, выделяют клон, в клонированном фрагменте которого были обнаружены два XhoI и PstI субфрагмента с геном эндоглюканазы. Эти фрагменты субклонируют в плазмидном векторе Bluescript SK+. Далее определяют нуклеотидную последовательность гена эндоглюканазы III Penicillium verruculosum по обеим цепям ДНК методом Сэнгера (фиг.1). Кодирующий район гена эндоглюканазы имел открытую рамку считывания протяженностью 711 п.н., в котором было два интрона 56 п.н. и 66 п.н. соответственно. Положение интронов и последовательность их 5’ и 3’ границ консервативна для структуры генов других грибных эндоглюканаз. Вычисленная последовательность аминокислот в белке совпадала с последовательностью аминокислот в пептидах зрелого белка, определенной прямым секвенированием пептидов.
Пример 2. Получение плазмидной конструкции pPrEG для экспрессии и секреции гена эндоглюканазы III Penicillium verruculosum в клетках Penicillium canescens.
Для введения сайта рестрикции АраI непосредственно после последовательности, кодирующей лидерный пептид гена эндоглюканазы III, проводят ПЦР фрагмента, кодирующего структурную часть эндоглюканазы III Penicillium verruculosum. Для этого используют праймеры:
EG3ApaI праймер:
5` -GTT-GCC-GCG-TGG-GCC-CAA-CAA-CAG-AG - 3`
M13/pUC обратный праймер для сиквенирования:
5` - GAG-CGG-ATA-ACA-ATT-TCA-CAC-AGC - 3`
Продукт ПЦР, расщепленный рестриктазами АраI и SalI, клонируют в плазмиду pEG5, содержащую часть гена egl (ген эндоглюканазы III Penicillium verruculosum), расщепленную теми же рестриктазами (получают плазмиду pEGApa).
Для проведения последующих манипуляций EcoRV-BglII фрагмент плазмиды рРr2.6 размером 3,1 т.п.н., содержащий промотор гена bgaS (ген секретируемой бета-галактозидазы Penicillium canescens) и 5`-транслируемую область гена bgaS переклонируют в плазмидный вектор pIC20R, расщепленный рестриктазами BglII и SmaI (получают плазмиду pPrBg-V).
KpnI-АрaI фрагмент плазмиды pPrBg-V длинной 2,0 т.п.н., содержащий промотор гена bgaS и последовательность, кодирующую лидерный пептид секретируемой бета-галактозидазы Penicillium canescens, лигируют в плазмиду pEGApa, расщепленную теми же рестриктазами, и получают плазмиду pPrEG.
Пример 3. Котрансформация штамма Penicillium canescens PCA10 (niaD) плазмидами pSTA-10 и pPrEG и селекция штамма Penicillium canescens R201 - продуцента эндоглюканазы III Penicillium verruculosum.
Штамм-реципиент Penicillium canescens PCA10 (niaD-) (Aleksenko A.Y. et al, 1995. Curr Genet. 28:474-477) выращивают в течение 16 ч при 30°С на полной питательной среде с NH4Cl в качестве источника азота, переносят мицелий в раствор 1,2 М MgSO4 и 10 mM NaH2PO4, pH 5,8 и добавляют лизирующий фермент Trichoderma harzianum до концентрации 5 мг/мл. Протопластирование проводят в течение 2 ч при 30°С в условиях перемешивания. Суспензию переносят в центрифужную пробирку и наслаивают 1-2 см раствора 0,6 М сорбитол, 10 mM CaCl, 10 mM Трис-HCl. После центрифугирования при 3000 об./мин, 4°С в течение 10 мин отбирают интерфазу, которая содержит протопласты. Протопласты промывают 2 раза в стабилизирующем растворе, содержащем 1,2 М сорбитола, 10 мМ Трис, pH 7,5, 10 мМ CaCl2 и ресуспендируют в нем же до концентрации 108 протопластов/мл. Трансформацию проводят следующим образом: к 200 мкл суспензии протопластов добавляют 1 мкг ДНК плазмиды pSTA-10, несущей в качестве селективного маркера ген нитрат редуктазы Aspergillus niger, и 10 мкг ДНК плазмиды pPrEG с геном эндоглюканазы Penicillium verruculosum, инкубируют в ледяной бане 20 мин, после чего проводят осмотический шок в течение 5 мин в 50% полиэтиленгликоле в присутствии 10 mM CaCl, 10 mM Трис-HCl и высевают протопласты в верхнем слое на агаризованную минимальную среду, содержащую 1,2 М сорбитол, 10 mM NaNO3. Трансформанты проверяют путем ферментации, как описано в примере 9.
Пример 4. Конструирование плазмид замещения рРrНРНТr и pPrEGTr. Плазмида замещения рРrНРНТr (фиг.3).
Для введения сайта рестрикции HindIII непосредственно после ATG кодона гена hph Escherichia coli устойчивости к гигромицину В, проводят ПЦР фрагмента, кодирующего структурную часть гена hph и терминатор гена trpC Aspergillus nidulans. В качестве матрицы используют экспрессионный вектор PAN7-1, содержащий ген hph под контролем промотора гена gpdA Aspergillus nidulans и терминатор гена trpC. Для этого используют праймеры:
HYG-I праймер:
5` - CATCACCAAGCTTGAACTCACCGC - 3`
pUC/m13 обратный праймер:
5`- CAGGAAACAGCTATGAC - 3`
ПЦР-фрагмент клонируют в векторе pBluescript KS+, разрезанном в сайте рестрикции SmaI (получают плазмиду рНРН).
Для удаления второго сайта рестрикции HindIII плазмиды рНРН, HindIII-SphI фрагмент плазмиды рНРН длинной 1,9 т.п.н., содержащий ген hph и терминатор гена trpC A.nidulans, переклонируют в вектор pIC20R, рестрицированный теми же эндонуклеазами (получают плазмиду pICH-HS).
XhoI-HindIII фрагмент длиной 193 п.н. промотора гена bgaS с ATG кодоном из плазмиды рРr2.6 лигируют в плазмиду pICH-HS, сохраняя рамку считывания гена hph (получают плазмиду pICH-XS).
Оставшуюся BglII-XhoI часть промотора bgaS из плазмиды рРr2.6 вставляют в плазмиду pICH-XS, рестрицированую теми же эндонуклеазами (получают плазмиду рРrНРН).
BglII-BglII фрагмент 3'-области гена bgaS длинной 2,5 т.п.н. из плазмиды pBG12, содержащей ген bgaS с укороченным промотором, субклонируют в плазмиду рРrНРН по сайту рестрикции BamHI (получают плазмиду рРrНРНТr).
Плазмида замещения pPrEGTr (фиг.4). BglII-BglII фрагмент 3'-области гена bgaS длиной 2,5 т.п.н. из плазмиды pCG12 субклонируют в плазмиду pPrEG по сайту рестрикции SmaI (получают плазмиду pPrEGTr). Для этого липкие концы BglII-BglII фрагмента предварительно достраивают, используя фрагмент Кленова ДНК-полимеразы I, а тупые концы рестрицированой плазмиды pPrEG дефосфорилируют с помощью щелочной фосфотазы.
Пример 5. Трансформация штамма Penicillium canescens R201 плазмидой замещения рРrНРНТr и селекция штамма Penicillium canescens R201-55, не синтезирующего бета-галактозидазу вследствие замещения структурной части гена бета-галактозидазы на маркерный ген устойчивости к гигромицину.
Штамм Penicillium canescens R201 выращивают в течение 16 ч. при 30°С на полной питательной среде с NaNO3 в качестве источника азота и далее осуществляют процедуру протопластирования по методике, описанной в примере 3.
Трансформацию проводят, добавляя к 200 мкл суспензии протопластов 10 мкг ДНК плазмиды рРrНРНТr, несущей в качестве селективного маркера ген устойчивости к гигромицину под контролем промотора гена бета-галактозидазы. Далее проводят осмотический шок в течение 5 мин в 50% полиэтиленгликоле в присутствии 10 mМ СаСl и 10 mМ Трис-НСl рН 7,5. Далее высевают протопласты в жидком слое на агаризованную минимальную среду, содержащую 720 mM KCl, 50 mМ фруктозу, 1 mM арабинозу, 10 mМ NaNO3 и гигромицин (0,8 мг/мл). Трансформанты тестируют на способность синтезировать секретируемую бета-галактозидазу.
Селекционируют штаммы, не синтезирующие бета-галактозидазу, и тестируют эти штаммы на продуктивность синтеза эндоглюканазы III Penicillium verruculosum.
Отбирают штамм с наилучшими показателями синтеза эндоглюканазы и наименьшей остаточной бета-галактозидазной активностью Penicillium canescens R201 -55.
Пример 6. Мутагенез клеток штамм Pemcillium canescens R20l(bgaS-, hph) и селекция штамма-реципиента R201-55-15 (bgaS-, hph, niaD-).
Конидии штамма Penicillium canescens R201 обрабатывают нитрозогуанидином (0,4 мг/мл) и рассевают на чашки с питательной средой, содержащей хлорат (0,45 М). Чашки инкубируют 4-5 суток при 30°С и выросшие колонии тестируют на чашках с добавками в качестве единственного источника азота: а) - 10 мМ NaNO3; б) - хлората (0,45М); в) - гипоксантина (5 мМ); г) - NH4Cl. Отбирают колонии, способные к росту на чашках с хлоратом и гипоксантином и не способные к росту на чашках с NaNO3. Отобранные штаммы выращивают на питательной среде с пептоном и свекловичным жомом и селекционируют штамм-реципиент R201-55-15 с продуктивностью эндоглюканазы не ниже исходного штамма Penicillium canescens R201-55.
Пример 7. Включение дополнительных копий гена эндоглюканазы в природный сайт гена бета-галактозидазы в геноме Penicillium canescens R201-55-15 при трансформации с помощью векторной плазмиды замещения pPrEGTr.
Штамм-реципиент Penicillium canescens R201-55-15 (bgaS- hph, niaD-) котрансформируют плазмидами pSTA-10 и pPrEGTr по методике, изложенной в примере 3. Отбирают 180 котрансформантов и среди них селекционируют 21 штамм, утративший устойчивость к гигромицину вследствие замещения гена гигромицина дополнительными копиями гена эндоглюканазы. Все отобранные штаммы ферментируют на питательной среде со свекловичным жомом и пептоном. Отбирают штамм Penicillium canescens R201-151 с активностью эндоглюканазы, превышающей активность штамма Penicillium canescens R201.
Пример 8. Конструкция плазмиды pXEG.
Для введения сайта рестрикции HindIII непосредственно после ATG кодона гена xylA Penicillium canescens ксиланазы проводят ПЦР фрагмента, содержащего промотор гена xylA. В качестве матрицы используют плазмиду pPCXYLA, содержащую полный ген xylA. Для этого используют праймеры:
XYLH праймер:
5` - CAGTCTTGAGAAGCTTCATGATTGTGAATGTG - 3`
pUC/m13 праймер для сиквенса:
5` -GCCAGGGTTTTCCCAGTCACGA - 3`
ПЦР-фрагмент длиной 2 т.п.н., разрезанный в сайтах рестрикции KpnI и HindIII, содержащий промотор гена xylA и ATG-кодон, клонируют в плазмиду pPrEG, разрезанную теми же рестриктазами, вместо промотора гена bgaS (получают плазмиду pXEG).
Пример 9. Мутагенез клеток штамм Penicillium canescens R201-151 и селекция штамма-реципиента R201-151 (niaD-).
Конидии штамма Penicillium canescens R201 обрабатывают нитрозогуанидином (0,4 мг/мл) и рассевают на чашки с питательной средой, содержащей хлорат (0,45 М). Чашки инкубируют и отбирают мутанты, как в примере 6. Отобранные штаммы выращивают на питательной среде с пептоном и свекловичным жомом и селекционируют штамм-реципиент R201-151 (niaD-) с продуктивностью эндоглюканазы не ниже исходного штамма Penicillium canescens R201-151.
Пример 10. Включение дополнительных копий гена эндоглюканазы под контролем промотора ксиланазного гена в геном Penicillium canescens R201-151(niaD-) при трансформации с помощью плазмиды pXEG.
Штамм-реципиент Penicillium canescens R201-151 (niaD-) котрансформируют плазмидами pSTA-10 и pXEG по методике, изложенной в примере 3. Отбирают 130 котрансформантов и среди них селекционируют штамм Penicillium canescens R3020 с активностью эндоглюканазы, превышающей активность штамма Penicillium canescens R201-151.
Пример 11. Выращивание штаммов Penicillium canescens R201, R201-151 и R3020 на ферментационной среде в колбах.
Для получения посевного материала культуру штамма Penicillium canescens R201, R201-151 иди R3020 выращивают на агаризованной минимальной среде при 30°С в течение 5-7 суток. Водной суспензией конидий (107 конидий/мл) инокулируют 100 мл среды (свекловичный жом - 30 г/л; пептон - 50 г/л; КН2РO4 - 25 г/л, рН 4,5) и инкубируют в качалочной колбе при 30°С в течение 120 час на круговой качалке при 240 об./мин. По окончании ферментации культуральную жидкость отделяют от мицелия и твердых остатков среды центрифугированием (5000g, 15 мин) и в супернатанте определяют активности эндоглюканазы III и бета-галактозидазы.
Определение активности эндоглюканазы:
Раствор хромогенного субстрата, 1% раствор ОС-41 (КМС окрашенная оранжевым красителем) в 0,1 М Na-ацетатном буфере рН 4,5 смешивают с разведенным ферментом, инкубируют в течение 10 мин при 40°С и останавливают реакцию добавлением 1 М CaCl2 в 80% этаноле. Смесь центрифугируют и активность фермента определяют по оптической плотности супернанта при 490 нм. Активность эндоглюканазы количественно вычисляют согласно руководству (А.П. Синицин и др. Биоконверсия лигноцеллюлезных материалов. М.: Изд-во Моск. ун-та, 1995).
Определение активности бета-галактозидазы.
Реакционную смесь, содержащую 3 мг о-нитрофенил-β-галактопиранозида (ОНФГ) в 1,9 мл, 50 mM Na-ацетатного буфера рН 4,5 и 100 мкл культуральной жидкости соответствующего разведения инкубируют 15 мин при 30°С, затем добавляют 1 мл 1 М Na2CO3 и измеряют оптическую плотность раствора при 420 нм. За единицу активности бета-галактозидазы принимают количество фермента, которое освобождает из субстрата ОНФГ 1 мкмоль о-нитрофенола за 1 мин при данных условиях реакции.
Через 96 часов ферментации штамм Penicillium canescens R201 накапливает в культуральной жидкости 120 ед./мл эндоглюканазной активности и 20 ед./мл бета-галактозидазной активности. Штамм Penicillium canescens R201-151 в тех же условиях продуцировал 160-170 ед./мл и 0,8-1,0 ед./мл активности, а штамм R3020 продуцировал 180-190 и менее 1 ед./мл активности соответственно.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ ЦЕЛЕВЫХ ГОМОЛОГИЧНЫХ И ГЕТЕРОЛОГИЧНЫХ ГЕНОВ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ ШТАММА ГРИБА PENICILLIUM VERRUCULOSUM И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА | 2008 |
|
RU2378372C2 |
| НОВЫЙ РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM CANESCENS CS15, ПРОДУЦИРУЮЩИЙ ЦЕЛЛЮЛАЗУ CLOSTRIDIUM THERMOCELLUM, И СПОСОБ ЕГО КУЛЬТИВИРОВАНИЯ | 2015 |
|
RU2612158C1 |
| КЛЕТКА МИЦЕЛИАЛЬНОГО ГРИБА Penicillium canescens - ПРОДУЦЕНТ КСИЛАНАЗЫ И ЛАККАЗЫ, СПОСОБ ПОЛУЧЕНИЯ КОМБИНИРОВАННОГО ФЕРМЕНТНОГО ПРЕПАРАТА КСИЛАНАЗЫ И ЛАККАЗЫ | 2012 |
|
RU2538149C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM CANESCENS CL14, ПРОДУЦИРУЮЩИЙ КОМПОНЕНТ ЦЕЛЛЛЮЛОСОМЫ CLOSTRIDIUM THERMOCELLUM, И СПОСОБ ЕГО КУЛЬТИВИРОВАНИЯ | 2016 |
|
RU2646132C1 |
| НОВАЯ ЭНДО-(1-4)-β-D-КСИЛАНАЗА ИЗ PENICILLIUM CANESCENS (ВАРИАНТЫ), ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ СЕКРЕТИРУЕМУЮ ЭНДО-(1-4)-β-D-КСИЛАНАЗУ ИЗ PENICILLIUM CANESCENS (ВАРИАНТЫ), И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2009 |
|
RU2412246C2 |
| СПОСОБ ПОЛУЧЕНИЯ КОРМОВОГО КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА (ВАРИАНТЫ) И ШТАММ PENICILLIUM CANESCENS (ВАРИАНТЫ) | 2004 |
|
RU2288267C2 |
| ГЕНЕТИЧЕСКАЯ ФЬЮЖН-КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ МУЛЬТИФЕРМЕНТНОГО КОМПЛЕКСА КАРБОГИДРАЗ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА Penicillium verruculosum, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ШТАММА ГРИБА Penicillium verruculosum И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА НА ЕГО ОСНОВЕ | 2013 |
|
RU2550044C2 |
| СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА ДЛЯ РАСЩЕПЛЕНИЯ ГЕМИЦЕЛЛЮЛОЗНЫХ ГЕТЕРОПОЛИСАХАРИДОВ КЛЕТОЧНОЙ СТЕНКИ РАСТЕНИЙ И ФЕРМЕНТНЫЙ ПРЕПАРАТ (ВАРИАНТЫ) | 2007 |
|
RU2358756C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ КОМПЛЕКСА ФЕРМЕНТОВ ЭНДОГЛЮКАНАЗ И КСИЛАНАЗ В КЛЕТКАХ ГРИБА PENICILLIUM VERRUCULOSUM И СПОСОБ ПОЛУЧЕНИЯ КОМПЛЕКСНЫХ ФЕРМЕНТНЫХ ПРЕПАРАТОВ НА ЕГО ОСНОВЕ, ПРЕДНАЗНАЧЕННЫХ ДЛЯ КОРМОПРОИЗВОДСТВА | 2017 |
|
RU2653429C1 |
| ФРАГМЕНТ ДНК PCG 2,6, КОДИРУЮЩИЙ СИНТЕЗ СЕКРЕТИРУЕМОЙ БЕТА-ГАЛАКТОЗИДАЗЫ PENICILLIUM CANESCENS, И ШТАММ ГРИБА PENICILLIUM CANESCENS - ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ БЕТА-ГАЛАКТОЗИДАЗЫ | 1997 |
|
RU2126049C1 |


Изобретение относится к микробиологии и биотехнологии. Фрагмент ДНК мицелиального гриба Penicillium verruculosum, кодирующий синтез секретируемой эндоглюканазы III, был включен в состав плазмид pPrEG и pXEG, в которых структурная часть гена эндоглюканазы III, кодирующая зрелый белок, состыкована с нуклеотидной последовательностью, кодирующей сигнальный пептид бета-галактозидазы и промоторными областями генов бета-галактозидазы и ксиланазы Penicillium canescens соответственно. На основе этих плазмид были получены штаммы Penicillium canescens R201 и R201-151, сконструированные методами трансформации и генетической инженерии, синтезирующие эндоглюканазу III Penicillium verruculosum под контролем промотора гена бета-галактозидазы Penicillium canescens и штамм Penicillium canescens R3020, синтезирующий эндоглюканазу III Penicillium verruculosum под контролем промоторов генов бета-галактозидазы и ксиланазы Penicillium canescens одновременно. Данное изобретение позволяет достичь более высокой продуктивности синтеза эндоглюканазы III Penicillium verruculosum. 7 с.п. ф-лы, 5 ил.
| ФРАГМЕНТ ДНК PCG 2,6, КОДИРУЮЩИЙ СИНТЕЗ СЕКРЕТИРУЕМОЙ БЕТА-ГАЛАКТОЗИДАЗЫ PENICILLIUM CANESCENS, И ШТАММ ГРИБА PENICILLIUM CANESCENS - ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ БЕТА-ГАЛАКТОЗИДАЗЫ | 1997 |
|
RU2126049C1 |
| Штамм гриба РеNIсILLIUм caNeSceNS - продуцент @ -галактозидазы | 1989 |
|
SU1717632A1 |

