Изобретение относится к области биотехнологии, а именно к созданию и производству комплексных ферментных препаратов (ФП), эффективно деградирующих некрахмальные полисахариды зерна. Изобретение может быть использовано в кормовой отрасли в качестве кормовой добавки в корм сельскохозяйственных животных и птиц.
Зерно, являющееся основой любых комбикормов, содержит набор некрахмальных полисахаридов (НПС), которые являются антипитательным фактором и затрудняют процесс переваривания комбикорма. Важнейшими НПС злаков являются β-глюкан и ксилан. Именно вязкие растворы β-глюканов и арабиноксиланов, не гидролизуемых в желудочно-кишечном тракте моногастричных животных, являются основным источником проблем при скармливании ячменя и ржи, и, отчасти, пшеницы (в зависимости от ее вязкости). НПС, в первую очередь ксиланы, сорбируют на себя значительную часть питательных веществ и жидкости, которые в неизменном виде выводятся из организма животных. Это приводит к перерасходу кормов и снижению показателей продуктивности у с/х животных и птицы. Следует учесть, что затраты на корма составляют до 70% от себестоимости конечной продукции животноводства.
Ячмень и рожь содержат значительный процент (до 15%) НПС [Бутейкис Г., Блажинскас Д. Ферменты – гарантия ощутимой выгоды сегодня и в будущем. Комбикорма. №6. 2012. С. 105-106]. Тем не менее, в России, особенно в Центральном и Северо-Западном районе, цена на рожь, овес и ячмень значительно ниже, чем на пшеницу, что определяет экономическую целесообразность использования этих злаков в рационе с/х животных. Влияние антипитательных факторов в виде арабиноксилана и β-глюкана корректируют введением соответствующих ФП, а именно ксиланаз и эндоглюканаз, в рацион животных. Поэтому одной из основных составляющих производства современных комбикормов, обеспечивающих высокие показатели продуктивности животноводства и птицеводства, является использование в рецептурах ФП разного целлюлазно/эндоглюканазного/ксиланазного состава для коррекции антипитательных факторов и улучшения усвояемости компонентов кормов.
Не менее важным фактором, определяющим эффективность применения ферментов (ксиланаз, эндоглюканаз, целлюлаз) для гидролиза природных НПС, является низкая степень или отсутствие ингибирования этих ферментов под действием белков-ингибиторов, входящих в состав большинства злаков. Наличие у зерновых белков-ингибиторов является эволюционным фактором, призванным защитить зерно от поражения микроорганизмами, расщепляющими полисахариды. Водорастворимые зерновые ингибиторы карбогидраз (семейства TAXI, XYP и др.) оказывают значительное действие на эндо-1,4-β-ксиланазы, относящиеся к 11 семье гликозилгидролаз, в то время как ксиланазы, относящиеся к 10 семье, менее подвержены ингибирующему воздействию [Гусаков А.В. // Биохимия. 2010. Т. 75. № 10. С. 1331].
Мицелиальный гриб Penicillium canescens относится к почвенным грибам, основным питательным субстратом которого является ксилано-богатая растительная биомасса, поэтому комплекс секретируемых ферментов гриба представлен, в основном, ксиланазами (КсилА, КсилЕ, КсилF и др.) и арабинофуранозидазами (АБФА, АБФБ и др). Ксиланазы А и Е относятся к 10 семейству гликозилгидролаз, при этом КсилЕ практически не ингибируется белками ржи. Разница в уменьшении приведенной вязкости для нативного и термоинактивированного экстрактов составляет лишь 5%. В случае КсилА эта разница выражена намного более заметно (~30%) [Денисенко Ю.А. с соавт // Сравнительная характеристика ксиланаз XylA и XylE из гриба Penicillium canescens. Вестник московского университета. Серия 2. Химия. 2015, т. 56, №6, с. 348-353].
Мицелиальный гриб Penicillium verruculosum PV2007 (ВКМ F-3972D) обладает собственным гидролитическим комплексом ферментов для биодеградации целлюлозосодержащих растительных материалов. В этот комплекс входят целлобиогидролазы различной специфичности, набор эндоглюканаз и β-глюкозидаза. Для реципиентного штамма P. verruculosum 537 (ΔniaD) разработана трансформационная и экспрессионная системы [патент RU 2378372 С2, опубл. 10.01.2010, бюл. №1], что позволяет использовать данный гриб как основу для получения рекомбинантных штаммов различного промышленного применения [Мерзлов Д.А. с соавт // Свойства ферментных препаратов и гомогенных ферментов эндоглюканазы EG2 Penicillium verruculosum и эндоглюканазы LAM Myceliophtora thermophile. Биохимия, 2015, том 20, вып.4, с.556-567; Синицын А.П. с соавт // Оптимизация состава целлюлазного ферментного комплекса Penicillium verruculosum: увеличение гидролитической способности с помощью методов генетической инженерии. Кат в пром. 2015, Т. 15, №6, с.78-83; заявка на патент №2015151483 от 02.12.2015; патент RU 2550044 C2, опубл. 10.05.2015, бюл. №13; патент RU 2532840 C2, опубл. 19.11.2014, бюл. №31].
Таким образом, разработка новых высокоэффективных и стабильных ФП для кормопроизводства, которые не будут подвержены ингибирующему действию растительных белков-ингибиторов карбогидраз, а также будут эффективны при кормлении сельскохозяйственных животных и птицы, является важной и актуальной задачей современного агропромышленного комплекса.
Техническая задача, на решение которой направлено данное изобретение, состоит в получении комплексных ФП на основе новых рекомбинантных штаммов Penicillium verruculosum, являющихся продуцентами гомологичной высокоэффективной эндоглюканазы II, принадлежащей 5 семейству гликозил-гидролаз, и гетерологичной неингибируемой ксиланазы Е из Penicillium canescens, относящейся к 10 семейству гликозил-гидролаз для применения в кормопроизводстве при обработке рационов, содержащих зерновые, в частности рожь и ячмень.
Технический результат от предлагаемого изобретения состоит в снижении вязкости корма, содержащего НПС ячменя или ржи, и повышении кормовой ценности рационов для сельскохозяйственных моногастричных животных и птицы при обработке корма новыми комплексными ФП, содержащими высокоактивную эндоглюканазу, неингибируемую ксиланазу и комплекс сопутствующих карбогидраз.
Сущность изобретения заключается в получении новых штаммов-продуцентов:
- Penicillium verruculosum EX13 (ВКМ F-4765D), который при выращивании на ферментационных средах на основе микрокристаллической целлюлозы и гидролизата куриного белка обеспечивает получение ФП комплексного действия, включающего неингибируемую эндо-1,4-β-ксиланазу (ксиланазу Е), высокоактивную эндо-1,4-β-глюканазу (эндоглюканазу II) с активностями в КЖ 550 ед./мл (по ксилану, рН 5,0, 50°С) и 500 ед./мл (Na-КМЦ, рН 5,0, 50°С) соответственно, и комплекс сопутствующих карбогидраз: целлобиогидролаза I, целлобиогидролаза II, β-глюкозидаза;
- Penicillium verruculosum EX35 (ВКМ F-4766D), который при выращивании на ферментационных средах на основе микрокристаллической целлюлозы и гидролизата куриного белка обеспечивает получение ФП комплексного действия, включающего неингибируемую эндо-1,4-β-ксиланазу (ксиланазу Е), высокоактивную эндо-1,4-β-глюканазу (эндоглюканазу II) с активностями в КЖ 800 ед./мл (по ксилану, рН 5,0, 50°С) и 300 ед./мл (Na-КМЦ, рН 5,0, 50°С) соответственно, и комплекс сопутствующих карбогидраз: целлобиогидролаза I, целлобиогидролаза II, β-глюкозидаза.
Способ получения ФП предусматривает глубинное культивирование штаммов–продуцентов P. verruculosum EX13 (BKM F-4765D) и P. verruculosum EX35 (ВКМ F-4766D) на удешевленной среде с заменой дрожжевого экстракта на гидролизат куриного белка с последующей распылительной сушкой культуральной жидкости. Изобретение позволяет получать ФП с высокой активностью целевых эндоглюканазы II и неингибируемой ксиланазы Е. Полученный ФП характеризуется высокой стабильностью, эффективностью при обработке кормовых смесей, содержащих ячмень и/или рожь, по сравнению с коммерческими ФП. Применение нового комплексного ФП позволит повысить кормовую ценность рационов на основе зерновых культур.
Изобретение реализуется следующим образом.
Штаммы P. verruculosum EX13 (BKM F-4765D) и P. verruculosum EX35 (ВКМ F-4766D) получены из исходного штамма Penicillium verruculosum PV2007 (ВКМ F-3972D) путем трансформации плазмидой pEGII-XylE с последующей селекцией на агаризованной среде с 10 мМ NaNO3.
Культурально-морфологические и микроскопические особенности штаммов (общие для P. verruculosum EX13 и P. verruculosum EX35):
Растут на агаризованных средах (среда Чапека с дрожжевым автолизатом, Мальц-агар, глюкозокартофельный агар, сусло-агар) при t 26-30°С в течение 7-10 суток, рН 4.5-5.0.
На среде Чапека с дрожжевым экстрактом при культивировании гриба при 25°С на 7 сутки колонии достигают 24-30 мм в диаметре, складчатые, поверхность сильно радиально плотная, тонкая, ростовая зона врастает в агар, имеет ширину 1.5-2.0 мм. Мицелий светло-желтоватый, шерстистый, центр колонии выпуклый, конидиогенез слабый, серо-зеленоватого оттенка. Экссудата и растворимого пигмента нет. Обратная сторона светлая, в центре колонии – палево-оранжевая. При температуре 37°С колонии диаметром 5 мм, мицелий светлый, конидиообразования нет. При температуре 5°С роста нет.
При росте на Мальц-агаре диаметр колонии 23-24 мм, поверхность сильно радиально складчатая, плотная, тонкая, ростовая зона врастает в агар, шириной 1,5-2,0 мм. Мицелий белый, шерстистый, прижатый, конидиогенез очень слабый, практически отсутствует. Эксудата и растворимого пигмента нет. Обратная сторона светлая.
При микроскопировании штаммы имеют конидиеносцы двухъярусные, терминальные, бивертициллятные, гладкие длиной около 150 мкм, шириной 2-3 мкм. Метулы расходящиеся размером 10-13х2.5-3.0 мкм, фиалиды ампуллиформные размером 7-8х2.8-3.0 мкм. Конидии округлые, шероховатые размером 3.0-3.5 мкм.
При культивировании в глубинных условиях с использованием растворимых субстратов (глюкоза, фруктоза, лактоза) штаммы образуют рыхлый разветвленный мицелий со слабой пеллетизацией, удельная культивирования 0,1 ч-1.
Физиолого-биохимические признаки штамма:
Мезофилен. Оптимальная температура роста мицелия 32°С (29-34°С), оптимум для образования целлюлаз 28°С (26-29°С). Оптимальные значения рН роста и секреции целлюлаз 3,5-5,0. Рост мицелия наблюдается и при рН 2,5, но при этом не наблюдается очень слабое образование целлюлаз и других карбогидраз.
Резистентность к нистатину хорошая. При поверхностном культивировании устойчив к концентрации до 0,5 мкг/мл, при концентрации 2,5 мкг/мл рост подавляется. При добавлении в среду дигитонина (3,5-4,0 мкг/мл) или бенгальского розового (30-50 мкг/мл) размер колоний уменьшается.
Является прототрофом. Способен быстро ассимилировать глюкозу, лактозу, глицерин, галактозу, ксилозу, D-маннозу, D-маннит, трегалозу, сорбозу и сорбит, медленнее - D-ксилозу, L- и D-арабинозу, L-рамнозу и рибозу. Слабо ассимилирует: D-глюкозамин, дезоксирибозу, дезоксигалактозу, 2-дезокси-D-глюкозу и 5-тио-D-глюкозу.
Использует неорганический и органический азот, хорошо ассимилирует нитратную и аммонийную форму азота.
Образует ферментные системы, позволяющие расти на соответствующих комплексных субстратах: целлюлозе, крахмале, ксилане, ламинарине, β-глюкане.
Штамм P. verruculosum EX13 (BKM F-4765D) отличается от исходного повышенной продукцией неингибируемой ксиланазы Е и высокоактивной эндоглюканазы II.
Штамм P. verruculosum EX35 (BKM F-4766D) отличается от исходного преимущественной продукцией неингибируемой ксиланазы Е и повышенной продукцией высокоактивной эндоглюканазы II.
В предлагаемом изобретении метод определения ксиланазной, β-глюканазной и КМЦ-азной активностей основан на измерении скорости образования восстанавливающих сахаров (ВС) методом Шомоди-Нельсона при гидролизе полисахаридных субстратов (ксилана из древесины березы, βглюкана ячменя и карбоксиметилцеллюлозы, натриевой соли соответственно). За единицу активности принимается такое количество фермента, которое приводит к образованию 1 мкмоль ВС в минуту при рН 5.0 и 50°C [Синицын А.П., Гусаков А.В., Черноглазов В.А. Биоконверсия лигноцеллюлозных материалов. – М.: МГУ, 1995. – 144 с.].
Новые комплексные ФП обладают улучшенными эксплуатационными характеристиками, включая устойчивость к ингибированию со стороны зерновых белковых ингибиторов и увеличенную операционную стабильность, что является преимуществом данных ферментных препаратов в сравнении с имеющимися на рынке коммерческими аналогами.
Применение новых комплексных ФП высокоактивной эндоглюканазы II и неингибируемой ксиланазы Е существенно повысит рентабельность применения комплексных препаратов в кормопроизводстве за счет обеспечения оптимального сочетания основных ферментативных активностей, необходимых для гидролиза НПС зерновой составляющей рациона корма с/х животных и птиц.
Возможность использования изобретения иллюстрируется примерами, которые не ограничивают объем и сущность притязаний, связанных с ними.
Пример 1. Конструирование экспрессионной плазмиды pEGII-XylE состояло из амплификации генов eglII P. verruculosum [патент РФ № 2378372], xylE P. canescens [AN: FJ860894], промоторной и терминаторной области гена cbhI P.verruculosum [патент РФ № 2378372], и финальной амплификации линейного продукта, состоящего из состыкованных генов, соответствующих промоторной области гена cbhI, гена eglII и гена xylE, соединенных через синтетический линкер, и терминаторной области гена cbhI P. verruculosum.
Для индивидуальной амплификации каждой из частей плазмиды использовались следующие олигонуклеотиды:
Далее с помощью полимеразной цепной реакции (ПЦР) и праймеров (1 и 3) синтезируют отдельно промоторную область гена cbhI размером 1303 п.н. В качестве матрицы для ПЦР используют геномную ДНК штамма P. verruculosum, выделенную из мицелия. Нуклеотидную последовательность полученного фрагмента определяют методом Сэнгера по обеим цепям.
Далее с помощью ПЦР и праймеров (2 и 5) синтезируют отдельно ген eglI I размером 1367 п.н., из которых 1343 п.н. составляет ген eglII, 24 п.н. относятся к синтетическому линкерному участку, соединяющему ген eglII c 3`-конца гена и ген xylE с 5`-конца гена. В качестве матрицы для ПЦР используют геномную ДНК штамма P. verruculosum, выделенную из мицелия. Нуклеотидную последовательность полученного фрагмента определяют методом Сэнгера по обеим цепям.
Далее с помощью ПЦР и праймеров (4 и 7) синтезируют отдельно ген xylE размером 1148 п.н. В качестве матрицы для ПЦР используют геномную ДНК штамма P. canescens, выделенную из мицелия. Нуклеотидную последовательность полученного фрагмента также определяют методом Сэнгера по обеим цепям.
Далее с помощью ПЦР и праймеров (6 и 8) синтезируют ген cbhI и его терминаторную область размером 570 п.н. В качестве матрицы для ПЦР используют геномную ДНК штамма P. verruculosum, выделенную из мицелия. Нуклеотидную последовательность полученного фрагмента определяют методом Сэнгера по обеим цепям.
Полученные ПЦР-фрагменты смешивают в эквимолярном соотношении и используют в качестве матрицы для финальной ПЦР-реакции при помощи праймеров (1) и (8). Синтезированный линейный фьюжн-фрагмент имеет размер 4388 п.н. Нуклеотидную последовательность полученного фрагмента определяют методом секвенирования по Сэнгеру по обеим цепям. Далее, полученный фрагмент клонируют в лабораторный вектор pUC-LIC с образованием плазмиды pEGII-XylE. Структура плазмиды pEGII-XylE представлена на фиг.1. На фиг. 1 темно-серым обозначены промоторная и терминаторная области гена cbhI, светло-серым гены eglII и xylE, между генами eglII и xylE обозначен линкер.
Плазмида pEGII-XylE была трансформирована в реципиентный штамм P.verruculosum 537 (ΔniaD) совместно с трансформирующей плазмидой pSTA10 по стандартной методике [Sambrook, J., and Russell, D.W. (2001) Molecular cloning:a laboratory manual, Cold Spring Harbor Laboratory Press, N.Y., A.Y. Aleksenko, N.A. Makarova, I.V. Nikolaev, A.J. Clutterbuck, Integrative and replicative transformation of Penicillium canescens with a heterologous nitrate-reductase gene, Curr. Genet. 28 (1995) 474-478]. В результате трансформации было получено более 200 рекомбинантных штаммов серии P. verruculosum EX, из которых в результате первичного скрининга на стандартной среде культивирования были отобраны 10 клонов (Таблица 1). Стандартная среда культивирования имела следующий состав (г/л): KH2PO4 - 15, (NH4)2SO4 - 5, MgSO4×7H2O - 0,3, CaCl2×2H2O - 0,3, дрожжевой экстракт - 10, целлюлоза – 20, пшеничные отруби - 10.
Анализ удельных активностей в культуральной жидкости 10 рекомбинантных штаммов (Таблица 1) показал наличие клонов с различными комбинациями ксиланазной и КМЦ-азной активностей, что позволило отобрать два штамма для дальнейших прикладных испытаний - Penicillium verruculosum EX13 с приблизительно равными целевыми ферментативными активностями ксиланазы и эндоглюканазы и Penicillium verruculosum EX35 с превалирующей ксиланазной активностью.
Пример 2. Культивирование штаммов Penicillium verruculosum EX13 (ВКМ F-4765D) и Penicillium verruculosum EX35 (ВКМ F-4766D) проводили в ферментерах объемом 3 л, оснащенных барботерами для подачи воздуха в аппарат и турбинной мешалкой на средах 1 и 2 следующего состава:
Среда 1. Стандартная среда (г/л):
Среда 2. Экспериментальная среда (г/л):
В экспериментальной среде вместо французского дрожжевого экстракта («Lessafre», Франция) использовался отечественный гидролизат куриного белка (ООО «Симбио», Россия) в том же процентном соотношении, что и дрожжевой экстракт в стандартной среде.
Культивирование проводили 144 ч, при рН не ниже 4,5 и 32°С. Образцы культуральной жидкости отбирали каждые сутки, начиная с 72 ч культивирования, центрифугировали и измеряли активности (Таблица 2).
Анализируя динамику накопления белка и биосинтез ксиланазы Е и эндоглюканазы II в культуральных жидкостях рекомбинантных штаммов P. verruculosum EX13 и P. verruculosum EX35 в процессе проведения ферментации, следует заключить, что замена дорогостоящего импортного дрожжевого экстракта на дешевый отечественный гидролизат куриного белка не влияет на биосинтез целевых ферментов в процессе культивирования рекомбинантных штаммов. Это означает что замена данного компонента целесообразна.
Культуральные жидкости рекомбинантных штаммов P. verruculosum EX13 и P. verruculosum EX35, полученные на Среде 2, были сепарированы с отделением биомассы гриба и высушены на распылительной сушке. Таким образом, были получены комплексные ферментные препараты (ФП) EX13 и EX35 c удельными ферментативными активностями по ксилану березы - 22500 и 30000 ед./г ФП, и по КМЦ - 19500 и 9000 ед./г ФП соответственно.
Сухие ферментные препараты были упакованы в двойные пластиковые мешки и хранились при температуре +6+3°С.
Пример 3. Водный экстракт из зерен ржи готовили следующим образом.
Зерно ржи очищали от посторонних включений и размалывали на лабораторной мельнице. На сите отбирали фракцию менее 0,5 мм.
Взвешивали 20,0 г размолотых зерен, прибавляли 100 мл натрий-ацетатного буфера (0,1 М, рН 5,0) и проводили экстракцию с использованием микробиологической качалки при температуре 40+1°С и 250 об/мин в течение 3 часов. Центрифугированием отделяли осадок от экстракта, после чего экстракт фильтровали через ткань типа ФП с размером пор около 10 мкм. Полученный нативный экстракт, содержащий водорастворимые белковые ингибиторы, укупоривали и хранили во льду в течение рабочего дня.
Экстракт, свободный от белковых ингибиторов, готовили путем нагревания части нативного экстракта в емкости на водяной бане при температуре 100°С в течение 10 минут с последующей фильтрацией через ткань типа ФП для отделения термоденатурированных белковых ингибиторов. Получали термоинактивированный экстракт, свободный от белковых ингибиторов, который укупоривали и хранили во льду в течение рабочего дня.
Капиллярный вискозиметр Оствальда подготавливали согласно инструкции по эксплуатации. Помещали вискозиметр в водяную баню при температуре 40+0,1°С, перед началом измерений все образцы инкубировали в течение 6-7 минут. Проводили определение времени истечения натрий-ацетатного буфера (0,1 М, рН 5,0), нативного и термоинактивированных экстрактов. Все измерения проводили с точностью до 0,1 с в трех повторностях.
К нативному и термоинактивированому экстракту прибавляли раствор ферментного препарата, содержащий 1 единицу ксиланазной активности, объемом, равным 1% от объема экстракта, и определяли динамику измерения времени истечения через равные промежутки времени на протяжении 25 минут. Время начала реакции отсчитывали от момента добавления к экстракту раствора ФП. Результаты представляли в виде зависимости отношения приведенной вязкости экстракта после добавления раствора ФП к приведенной вязкости исходного экстракта от времени ферментативной обработки.
Вязкость растворов ηi в каждой точке рассчитывали по формуле ηi = К⋅ρ⋅t К – константа вискозиметра, ρ – плотность раствора, t – время истечения раствора. η0, ηбуф – начальная вязкости экстракта и буфера, определенные вискозиметрическим методом. Относительную вязкость экстракта рассчитывали как ηотн = η/ηбуф; удельную вязкость рассчитывали, как ηуд = (η – ηбуф)/ηбуф = ηотн-1, приведенную вязкость рассчитывали как ηприв=ηуд/С, где С – весовая концентрация полимера. ηiуд/η0уд=ηiприв/η0прив, где η0уд и η0прив – удельная и приведенная вязкости экстракта до добавления раствора ферментного препарата; ηiуд и ηiприв после добавления раствора ферментного препарата в момент времени ферментативной обработки в точке i. Время ферментативной обработки i = время начала измерения времени истечения + ½⋅время истечения в точке i (за исключением начальной точки). Снижение удельной вязкости Δηiуд относительно вязкости экстракта в начальной точке рассчитывали, как (1-ηiуд/η0уд)⋅100% = (1-ηiприв/η0прив)⋅100%.
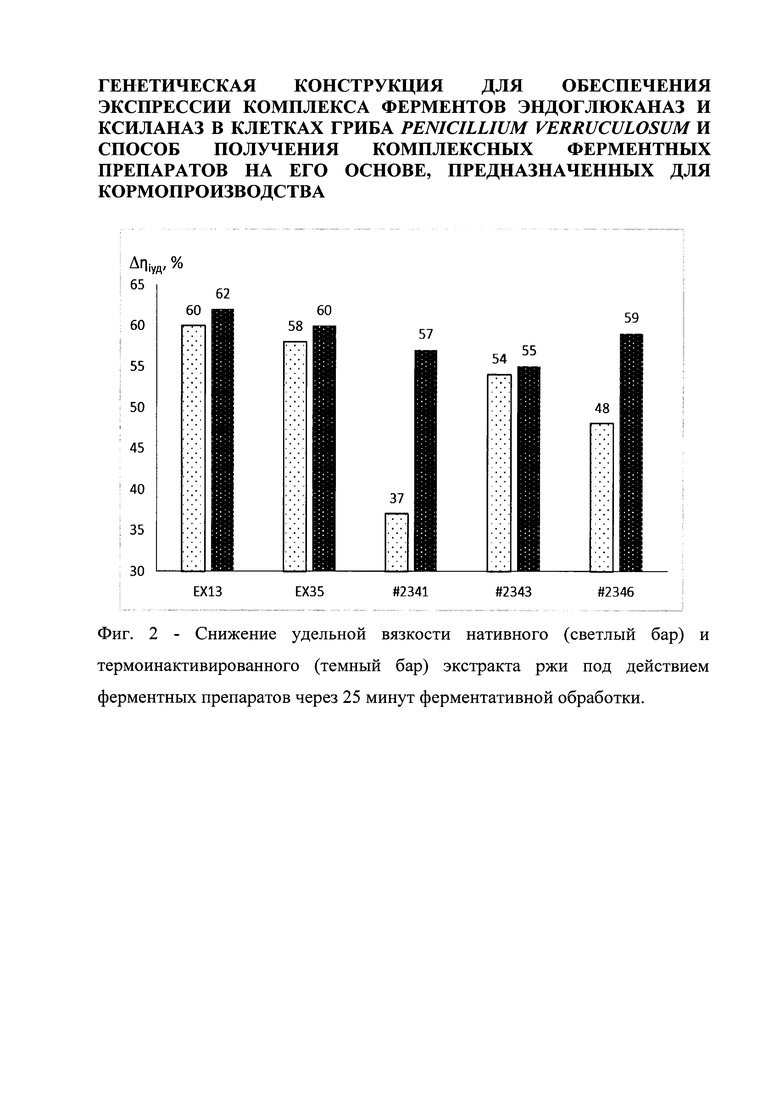
Среднее время эффективной работы внесенных с кормом ферментов в организме курицы составляет около 20-30 минут, после чего происходит их расщепление в желудочно-кишечном тракте. Результаты уменьшения вязкости нативного (светлые столбики на Фиг.2) и термоинактивированного (темные столбики на Фиг 2) экстракта после 25 минут ферментативной обработки представлены на Фиг. 2. В качестве контроля использовался ряд коммерческих ферментных препаратов, широко используемых в качестве кормовых добавок в составе современных рационов кормления с/х животных и птицы. К ним относятся: #2341 – Ronozyme WX (CT), #2343 – Natugrain TS, #2346 – Rovabio XL. Все препараты были дозированы по 1 Ед активности ксиланазы в реакционной смеси. Как видно из Фиг.2 максимальное значение снижения вязкости как термоинактивированного, так и нативного экстракта ржи наблюдалось в случае использования новых ферментных препаратов EX13 и EX35, что говорит о потенциальной эффективной применимости этих препаратов в рационах кормления с/х животных и птицы.
Таблица 1. Удельные активности ферментов (ед./мг) и концентрация белка (мг/мл) в культуральных жидкостях полученных рекомбинантных штаммов по сравнению с контрольным образцом (нетрансформированный штамм, Контроль)
Таблица 2. Динамика накопления белка (мг/мл) и биосинтез ксиланазы Е и эндоглюканазы II в культуральных жидкостях рекомбинантных штаммов P. verruculosum EX13 и P. verruculosum EX35 в процессе проведения ферментации в 1-л ферментерах на Средах 1 и 2.
EX13/EX35*
рН 5,0, 50°С
EX13/EX35*
рН 5,0, 50°С
EX13/EX35*
*- Первая цифра соответствует параметрам биосинтеза ферментов в штамме P. verruculosum EX13, вторая цифра соответствует параметрам биосинтеза ферментов в штамме P. verruculosum EX35.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM MX-73 ПРОДУЦЕНТ МОДИФИЦИРОВАННОЙ КСИЛАНАЗЫ Е С ПОВЫШЕННОЙ ТЕРМОСТАБИЛЬНОСТЬЮ, ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В ПИЩЕВОЙ И КОРМОВОЙ ПРОМЫШЛЕННОСТИ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2018 |
|
RU2711578C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ПРОДУЦЕНТ КОМПЛЕКСА ФИТАЗЫ А И ЭНДО-1,4-β-ГЛЮКАНАЗЫ II И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ ДОБАВКИ В КОРМАХ | 2022 |
|
RU2810538C2 |
| ГЕНЕТИЧЕСКАЯ ФЬЮЖН-КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ МУЛЬТИФЕРМЕНТНОГО КОМПЛЕКСА КАРБОГИДРАЗ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА Penicillium verruculosum, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ШТАММА ГРИБА Penicillium verruculosum И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА НА ЕГО ОСНОВЕ | 2013 |
|
RU2550044C2 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ПРОДУЦЕНТ КОМПЛЕКСА ФИТАЗЫ А И ЭНДО-1,4-β-КСИЛАНАЗЫ Е И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ ДОБАВКИ В КОРМАХ | 2023 |
|
RU2819918C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ЕЕ-105 ПРОДУЦЕНТ КОМПЛЕКСА ВЫСОКОЭФФЕКТИВНЫХ ЭНДОГЛЮКАНАЗ И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ КОРМОВОЙ ДОБАВКИ В ЗЕРНОВЫХ КОРМАХ | 2018 |
|
RU2696036C1 |
| ШТАММ ГРИБА PENICILLIUM CANESCENS mtCBHI ПРОДУЦЕНТ ТЕРМОСТАБИЛЬНОЙ ЦЕЛЛОБИОГИДРОЛАЗЫ I И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ БИОКОНВЕРСИИ ВОЗОБНОВЛЯЕМОГО РАСТИТЕЛЬНОГО СЫРЬЯ В САХАРА | 2019 |
|
RU2741078C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM B6/PMO-BG-25 И СПОСОБ ПОЛУЧЕНИЯ КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА β-ГЛЮКОЗИДАЗЫ, ЦЕЛЛОБИОГИДРОЛАЗЫ И ПОЛИСАХАРИДМОНООКСИГЕНАЗЫ НА ЕГО ОСНОВЕ ДЛЯ БИОКОНВЕРСИИ ЦЕЛЛЮЛОЗОСОДЕРЖАЩИХ МАТЕРИАЛОВ | 2024 |
|
RU2836771C1 |
| ШТАММ ГРИБА Penicillium verruculosum B10 EGII ПРОДУЦЕНТ ЭНДО-1.3/1.4-β-ГЛЮКАНАЗЫ, ЦЕЛЛЮЛАЗЫ, β-ГЛЮКОЗИДАЗЫ И КСИЛАНАЗЫ И СПОСОБ ПОЛУЧЕНИЯ КОРМОВОГО КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА | 2012 |
|
RU2532840C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM ( ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА С ЕГО ИСПОЛЬЗОВАНИЕМ (ВАРИАНТЫ) | 2015 |
|
RU2646136C2 |
| Штамм мицелиального гриба TRICHODERMA LONGIBRACHIATUM TW-14-220 - продуцент целлюлаз, бета - глюканаз и ксиланаз для кормопроизводства и способ получения кормового комплексного ферментного препарата | 2017 |
|
RU2654564C1 |

Изобретение относится к области биохимии, генной инженерии и биотехнологии, в частности к генетической конструкции для обеспечения экспрессии целевых гомологичного и гетерологичного генов в клетках реципиентного гриба Penicillium verruculosum. Настоящая конструкция содержит целевую кодирующую последовательность, включающую последовательно соединенные ген eglII, кодирующий высокоактивную эндо-1,4-β-глюканазу II, и ген xyIE Penicillium canescens, кодирующий неингибируемую эндо-1,4-β-ксиланазу Е. Изобретение также относится к рекомбинантным штаммам Penicillium verruculosum ЕХ13, ВКМ F-4765D, и Penicillium verruculosum EX35, ВКМ F-4766D. Указанные штаммы предназначены для продукции гомологичной высокоактивной эндо-1,4-β-глюканазы II и неингибируемой эндо-1,4-β-ксиланазы Е Penicillium canescens. Настоящее изобретение позволяет получать неингибируемую эндо-1,4-β-ксиланазу Е Penicillium canescens и высокоактивную эндо-1,4-β-глюканазу II Penicillium verruculosum. 4 н.п. ф-лы, 2 ил., 2 табл., 3 пр.
1. Генетическая конструкция для обеспечения экспрессии целевых гомологичного и гетерологичного генов в клетках реципиентного гриба Penicillium verruculosum 537 (ΔniaD), используемого в качестве хозяина, содержащая целевую кодирующую последовательность, включающую последовательно соединенные ген eglII, кодирующий высокоактивную эндо-1,4-β-глюканазу II (ЭГII), и ген xyIE Penicillium canescens, кодирующий неингибируемую эндо-1,4-β-ксиланазу Е (КсилЕ) посредством синтетического линкера, и функционально связанную с регуляторными элементами гена целлобиогидролазы I Penicillium verruculosum, которые представляют собой промотор, сигнальный пептид и терминатор гена целлобиогидролазы I.
2. Рекомбинантный штамм Penicillium verruculosum ЕХ13, депонированный во Всероссийской коллекции микроорганизмов под номером ВКМ F-4765D, полученный путем трансформации клеток гриба Penicillium verruculosum 537 (ΔniaD) генетической конструкцией по п. 1 и предназначенный для продукции гомологичной высокоактивной эндо-1,4-β-глюканазы II и неингибируемой эндо-1,4-β-ксиланазы Е Penicillium canescens.
3. Рекомбинантный штамм Penicillium verruculosum EX35, депонированный во Всероссийской коллекции микроорганизмов под номером ВКМ F-4766D, полученный путем трансформации клеток гриба Penicillium verruculosum 537 (ΔniaD) генетической конструкцией по п. 1 и предназначенный для продукции преимущественно неингибируемой эндо-1,4-β-ксиланазы Е Penicillium canescens и высокоактивной эндо-1,4-β-глюканазы II.
4. Способ получения ферментного препарата ЕХ13, полученного на основе рекомбинантного штамма Penicillium verruculosum ЕХ13 по п. 2, при выращивании на ферментационных средах, обеспечивающий получение ферментного препарата комплексного действия, включающего неингибируемую эндо-1,4-β-ксиланазу Е Penicillium canescens и высокоактивную эндо-1,4-β-глюканазу II, и ферментного препарата ЕХ35, полученного на основе рекомбинантного штамма Penicillium verruculosum ЕХ35 по п. 3, при выращивании на ферментационных средах, обеспечивающий получение ферментного препарата преимущественно неингибируемой эндо-1,4-β-ксиланазы Е Penicillium canescens и высокоактивной эндо-1,4-β-глюканазы II и состоящий в культивировании рекомбинантных штаммов по пп. 2 и 3, раздельно, на ферментационной среде, состоящей из (г/л): глюкоза - 40, микрокристаллическая целлюлоза - 20, гидролизат куриного белка - 10, пшеничные отруби - 10, дигидрофосфат калия - 15, сульфат аммония - 5, хлорид кальция - 0,3, семиводный сульфат магния - 0,3, отличающейся от стандартной среды наличием гидролизата куриного белка вместо дрожжевого экстракта.
| ГЕНЕТИЧЕСКАЯ ФЬЮЖН-КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ МУЛЬТИФЕРМЕНТНОГО КОМПЛЕКСА КАРБОГИДРАЗ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА Penicillium verruculosum, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ШТАММА ГРИБА Penicillium verruculosum И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА НА ЕГО ОСНОВЕ | 2013 |
|
RU2550044C2 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ ЦЕЛЕВЫХ ГОМОЛОГИЧНЫХ И ГЕТЕРОЛОГИЧНЫХ ГЕНОВ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ ШТАММА ГРИБА PENICILLIUM VERRUCULOSUM И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА | 2008 |
|
RU2378372C2 |
| MERZLOV DA., et al., New enzyme preparation for animal feed production based on Penicillium recombinant strains, Микробные биотехнологии: фундаментальные и прикладные аспекты, тезисы докладов IX Международной научной конференции, 2015 | |||
| MERZLOV DA., et al., Properties of enzyme preparations and homogeneous enzymes -Endoglucanases EG2 Penicillium verruculosum and LAM Myceliophthora thermophila, Biochemistry, 2015 | |||
| MOROZOVA VV., et al., Cellulases of Penicillium verruculosum, Biotechnology journal, 2010 | |||
| КОВАЛЕНКО ЕА., и др., Изучение возможности переработки вторичного сырья убоя птицы в гидролизаты микробиологических сред, / Хранение и переработка сельхозсырья, 2015 | |||
| СПОСОБ ДИАГНОСТИКИ ДЕРМАТОМИКОЗОВ И ПИТАТЕЛЬНАЯ СРЕДА ДЛЯ ДЕРМАТОФИТОВ | 2004 |
|
RU2275631C1 |

