ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Дифференциация клеток многоклеточных организмов регулируется гормонами и полипептидными факторами роста. Эти диффундируемые молекулы позволяют клеткам осуществлять коммуникацию друг с другом и действовать совместно для образования тканей и органов и для репарации и регенерации поврежденной ткани. Примеры гормонов и факторов роста включают в себя, среди прочих, стероидные гормоны, паратиреоидный гормон, фолликулостимулирующий гормон, интерфероны, интерлейкины, тромбоцитарный фактор роста, эпидермальный фактор роста, гранулоцитарно-макрофагальный колониестимулирующий фактор.
Гормоны и факторы роста влияют на клеточный метаболизм посредством связывания с рецепторными белками. Некоторые рецепторы являются интегральными мембранными белками, которые связываются с гормоном или фактором роста вне клетки и которые связаны с путями передачи сигналов внутри клетки, такими как системы вторичных мессенджеров. Другие классы рецепторов являются растворимыми внутриклеточными молекулами.
Цитокины обычно стимулируют пролиферацию и/или дифференцировку клеток гемопоэтического направления дифференцировки или участвуют в механизмах иммунных и воспалительных реакций организма. Примеры цитокинов, которые влияют на гемопоэз, включают в себя эритропоэтин (ЕРО), который стимулирует развитие эритроцитов; тромбопоэтин (ТРО), который стимулирует развитие клеток мегакариоцитарной линии дифференцировки; и гранулоцитарный колониестимулирующий фактор (G-CSF), который стимулирует развитие нейтрофилов. Эти цитокины применимы в восстановлении нормальных уровней клеток крови в пациентах, страдающих от анемии, тромбоцитопении и нейтропении, или получающих химиотерапию по поводу рака. Цитокины играют важные роли в регуляции гемопоэза и иммунных реакций и могут влиять на развитие лимфоцитов.
Класс II семейства цитокинов человека включает в себя подтипы интерферона-α (IFN-α), интерферон-β (IFN-β), интерферон-γ (IFN-γ), IL-10, IL-19 (патент США 5 985 614), MDA-7 (Jiang et al., Oncogene 11, 2477-2486, (1995)), IL-20 (Jiang et al., Oncogene 11, 2477-2486, (1995)), IL-22 (Xie et al., J.Biol. Chem. 275, 31335-31339, (2000)) и АК-155 (Knappe et al., J.Virol. 74, 3881-3887, (2000)). Большинство цитокинов связывают и передают сигналы через рецепторы цитокинов Класса I или Класса II. Члены семейства рецепторов цитокинов класса II человека включают в себя интерферон-αR1 (IFN-α R1), интерферон-γ-R2 (IFN-γ-R2), интерферон-γR1 (IFN-γR1), интерферон-γR2 (IFN-γR2), IL-10R (Liu et al., J.Immunol. 152, 1821-1829, (1994)), CRF2-4 (Lutfalla et al., Genomics 16, 366-373, (1993)), IL-20Rβ (Blumberg et al., Cell 104, 9-19, (2001)) (так же известный как zcytor7 (патент США 5 945 511) и CRF2-8 (Kotenko et al., Oncogene 19, 2557-2565, (2000)), IL-20Rβ (Blumberg et al., ibid, (2001)) (так же известный как DIRS1 (РСТ WO 99/46379)), IL-21 (IL-22 рецептор-α1, представленный в HUGO для одобрения) (так же известный как IL-22R (Xie et al., J.Biol. Chem. 275, 31335-31339, (2000)), zcytor11 (патент США 5 965 704) и CRF2-9 (Kotenko et al, Oncogene 19, 2557-2565, (2000)) и тканевой фактор.
Рецепторы цитокинов класса II являются обычно гетеродимерами, состоящими из двух различных цепей рецептора, α- и β-субъединиц рецептора (Stahl et al., Cell 74, 587-590, (1993)). Обычно α-субъединицы являются первичными цитокинсвязывающими белками, а β-субъединицы требуются для образования сайтов связывания высокой аффинности, а также для трансдукции сигнала. Исключением является рецептор IL-20, в котором обе субъединицы необходимы для связывания IL-20 (Blumberg et al., ibid, (2001)).
Рецепторы цитокинов класса II идентифицируют по консервативному цитокинсвязывающему домену из приблизительно 200 аминокислот (D200) во внеклеточной части этого рецептора. Этот цитокинсвязывающий домен состоит из двух доменов фибронектина типа III (Fnlll), из приблизительно 100 аминокислот каждый (Bazan J.F. Proc. Natl. Acad. Sci. USA 87, 6934-6938, (1990); Thoreau et al., FEBS Lett. 282, 16-31, (1991)). Каждый домен FnIII содержит консервативные остатки Cys, Pro и Trp, которые определяют характерную конфигурацию укладки семи β-тяжей, сходную с константным доменом иммуноглобулинов (Uze et al., J. Interferon Cytokine Res. 15, 3-26 (1995)). Эти консервативные структурные элементы семейства рецепторов цитокинов класса II позволяют идентифицировать новые члены этого семейства на основе гомологии первичной аминокислотной последовательности. Ранее авторы данного изобретения успешно идентифицировали два новых члена семейства рецепторов цитокинов класса II, zytor7 (патент США 5 945 511) (так же известный как IL-20R α (Blumberg et al., ibid, (2001)) и zcytor11 (патент США 5 965 704) (так же известный как IL-22R (Blumberg et al., ibid, (2001)) с использованием этого подхода. Идентификация дополнительных новых членов семейства рецепторов цитокинов класса II представляет интерес, так как цитокины играют первостепенную роль в регуляции биологических реакций.
IL-22, так же известный как IL-TIF (IL-10-родственный происходящий из Т-клеток индуцируемый фактор) (Dumoutier et al., J.Immunology 164, 1814-1819, (2000)), является недавно описанным гомологом IL-10. Мышиный IL-22 первоначально идентифицировали как ген, индуцируемый IL-9 в Т-клетках и тучных клетках in vitro (Dumoutier et al., J.Immunology 164, 1814-1819, (2000)). Активность индукции реагентом острой фазы наблюдали в печени мыши при инъекции IL-22, и экспрессия IL-22 быстро индуцировалась после инъекции липополисахарида (LPS), что предполагает, что IL-22 способствует воспалительной реакции in vivo (Dumoutier et al., Proc. Natl. Acad. Sci. USA 97, 10144-10149, (2000)).
Интерлейкины являются семейством цитокинов, которые опосредуют иммунологические реакции, в том числе воспаление. Интерлейкины опосредуют разнообразные воспалительные патологии. Центральной для иммунной реакции является Т-клетка, которая продуцирует многие цитокины и создает искусственный (приобретенный) иммунитет к антигенам. Цитокины, продуцируемые Т-клеткой, были классифицированы как цитокины Типа 1 и Типа 2 (Kelso, A. Immun. Cell Biol. 76:300-317, 1998). Цитокины Типа 1 включают в себя IL-2, IFN-γ, LT-α и участвуют в воспалительных реакциях, противовирусном иммунитете, иммунитете против внутриклеточных паразитов и отторжении аллотрансплантата. Цитокины Типа 2 включают в себя IL-4, IL-5, IL-6, IL-10 и IL-13 и участвуют в гуморальных реакциях, иммунитете против гельминтов и аллергической реакции. Имеются некоторые доказательства для предположения, что продуцирующие цитокины Типа 1 и Типа 2 популяции Т-клеток преимущественно мигрируют в различные типы воспаленной ткани.
Особый интерес, с терапевтической точки зрения, представляют собой интерфероны (обзоры по интерферонам обеспечены De Maeyer and De Maeyer-Guignard, "Interferons," in The Cytokine Handbook, 3rd Edition, Thompson (ed.), pages 491-516 (Academic Press Ltd. 1998) и Walsh, Biopharmaceuticals: Biochemistry and Biotechnology, pages 158-188 (John Wiley & Sons 1998)). Интерфероны проявляют различные биологические активности и применимы для лечения некоторых аутоиммунных заболеваний, в частности, раков, и усиления иммунной реакции против инфекционных агентов, в том числе вирусов, бактерий, грибков и простейших. К настоящему времени были идентифицированы шесть форм интерферонов, которые были классифицированы на две основные группы. Так называемые интерфероны «типа I» включают в себя интерферон-α, интерферон-β, интерферон-ω, интерферон-δ и интерферон-τ. В настоящее время интерферон-γ и один подкласс интерферона-α являются единственными интерферонами типа II.
Интерфероны типа I, которые, как считается, происходят из одного и того же предкового гена, сохранили достаточно сходную структуру, чтобы действовать посредством одного и того же рецептора поверхности клетки, α-цепь рецептора интерферона-α/β человека содержит внеклеточный N-концевой домен, который имеет характеристики рецептора цитокинов класса II. Интерферон-γ не имеет значимой общей гомологии с интерферонами типа 1 или с подтипом интерферона-α типа II, но имеет ряд общих биологических активностей с интерферонами типа I.
У людей по меньшей мере 16 неаллельных генов кодируют различные подтипы интерферона-α, тогда как интерфероны-β и ω кодируются единственными генами. Гены интерферонов типа I собраны в кластер в коротком плече хромосомы 9. В противоположность типичным структурным генам человека, гены интерферона-α, интерферона-β и интерферона-ω не имеют интронов. Единственный ген для интерферона-γ локализован на хромосоме 12 и содержит три интрона. К настоящему времени интерферон-τ был описан только в крупном рогатом скоте и овцах, тогда как интерферон-δ был описан только в свиньях.
Клиницисты используют преимущество множественных активностей интерферонов посредством использования этих белков для лечения большого диапазона состояний. Например, одна форма интерферона-α была одобрена для применения в более, чем 50 странах, для лечения таких медицинских состояний, как ретикулоэндотелиоз, почечно-клеточный рак (гипернефрома), базально-клеточный рак (базалиома), злокачественная меланома, связанная со СПИДом саркома Капоши, множественная миелома, хронический миелогенный лейкоз, не-Ходжкинская лимфома, папилломатоз гортани, грибовидный микоз, остроконечная кондилома, хронический гепатит В, гепатит С, хронический гепатит D и хронические не-А, не-В/С гепатиты. Департамент по контролю за качеством пищевых продуктов, медикаментов и косметических средств США одобрил применение интерферона-β для лечения множественного склероза, хронического заболевания нервной системы, интерферон-γ используют для лечения хронических гранулематозных заболеваний, в которых интерферон усиливает иммунную реакцию пациента для разрушения инфекционных бактериальных, грибковых и принадлежащих к простейшим патогенов. Клинические исследования показывают также, что интерферон-γ может быть применим в лечении СПИДа, лейшманиоза и лепроматозной проказы.
Продемонстрированные in vivo активности этого семейства цитокинов иллюстрируют огромный клинический потенциал этих цитокинов и потребность в других цитокинах, агонистах цитокинов и антагонистах цитокинов. Данное изобретение удовлетворяет эти потребности обеспечением нового цитокина, стимулирующего клетки гемопоэтической линии дифференцировки, а также связанных с этим цитокином композиций и способов.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКОГО МАТЕРИАЛА
На фиг.1-5 представлен интерферон-подобный белок Zcyto21.
На фиг.6 представлен профиль гидрофильности Hoop/Woods последовательности белка Zcyto21, показанной в SEQ ID NO:2. Этот профиль основан на скользящем окне из шести остатков. Скрытые остатки G, S и Т и открытые остатки Н, Y и W не принимались во внимание. Эти остатки указаны на фиг.6 строчными буквами.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Перед подробным изложением данного изобретения может быть полезным для его понимания определение следующих терминов:
Термин «аффинная метка» используется здесь для обозначения полипептидного сегмента, который может быть присоединен ко второму полипептиду для обеспечения очистки или детектирования второго полипептида или обеспечения сайтов для присоединения второго полипептида к субстрату. В принципе, любой пептид или белок, для которого доступны антитело или другой агент специфического связывания, может быть использован в качестве аффинной метки. Аффинные метки включают в себя полигистидиновый участок (тракт), белок A (Nilsson et al., EMBO J. 4:1075, 1985; Nilsson et al., Methods Enzymol. 198:3, 1991), глутатион S-трансферазу (Smith and Johnson, Gene 67:31, 1988), аффинную метку Glu-Glu (Grussenmeyer et al., Proc. Natl. Acad. Sci. USA 82:7952-4, 1985), вещество Р, пептид Flag™ (Hopp et al., Biotechnology 6:1204-10, 1988), стрептавидинсвязывающий пептид или другой антигенный эпитоп или связывающий домен. См., в общем, Ford et al., Protein Expression and Purification 2:95-107, 1991. ДНК, кодирующие аффинные метки, доступны от коммерческих поставщиков (например, Pharmacia Biotech, Piscataway, NJ).
Термин "аллельный вариант" используется здесь для обозначения любой из двух или нескольких альтернативных форм гена, занимающих один и тот же хромосомный локус. Аллельная вариация возникает природно через мутацию и может приводить к фенотипическому полиморфизму в популяциях. Мутации генов могут быть молчащими (без изменения в кодируемом полипептиде) или могут кодировать полипептиды, имеющие измененную аминокислотную последовательность. Термин аллельный вариант используется здесь также для обозначения белка, кодируемого аллельным вариантом гена.
Термины "амино-концевой" и "карбоксил-концевой" используются здесь для обозначения положений внутри полипептидов. Там, где позволяет контекст, эти термины используются со ссылкой на конкретную последовательность или часть полипептида для обозначения близости или относительного положения. Например, некоторая последовательность, расположенная карбоксил-терминально относительно ссылочной последовательности в полипептиде, расположена проксимально относительно карбоксил-конца ссылочной последовательности, но не обязательно находится при карбоксил-конце полного полипептида.
Термин "пара комплемент/антикомплемент" обозначает неидентичные части молекулы, которые образуют нековалентно связанную, стабильную пару при подходящих условиях. Например, биотин и авидин (или стрептавидин) являются членами-прототипами пары комплемент/антикомплемент. Другие примеры пар комплемент/антикомплемент включают в себя пары рецептор/лиганд, пары антитело/антиген (или гаптен или эпитоп), пары смысловой/антисмысловой полинуклеотид и т.п. В случае, когда желательна последующая диссоциация пары комплемент/антикомплемент, пара комплемент/антикомплемент предпочтительно имеет аффинность связывания <109 М-1.
Термин «комплементы полинуклеотидной молекулы" обозначает полинуклеотидную молекулу, имеющую комплементарную последовательность оснований и обращенную ориентацию в сравнении со ссылочной последовательностью. Например, последовательность 5′-ATGCACGGG-3′ комплементарна 5′-CCCGTGCAT-3′.
Термин «вырожденная нуклеотидная последовательность» обозначает последовательность нуклеотидов, которая включает в себя один или более вырожденных кодонов (в сравнении со ссылочной полинуклеотидной молекулой, которая кодирует полипептид). Вырожденные кодоны содержат различные триплеты нуклеотидов, но кодируют один и тот же аминокислотный остаток (т.е. триплеты GAU и GAC, каждый, кодируют Asp).
Термин «экспрессирующий вектор» используется для обозначения молекулы ДНК, линейной или кольцевой, которая содержит сегмент, кодирующий представляющий интерес полипептид, функционально связанный с дополнительными сегментами, которые обеспечивают его транскрипцию. Такие дополнительные сегменты включают в себя промоторную и терминаторную последовательности и могут также включать в себя одну или несколько точек начала репликации, один или несколько селектируемых маркеров, энхансер, сигнал полиаденилирования и т.д. Экспрессирующие векторы обычно произведены из плазмидной или вирусной ДНК или могут содержать элементы обеих.
Термин "выделенный", в применении к полинуклеотиду, обозначает, что этот полинуклеотид был удален из его природной генетической среды и, следовательно, не содержит других посторонних или нежелательных кодирующих последовательностей и находится в форме, подходящей для применения в генетически сконструированных системах продуцирования белков. Такие выделенные молекулы являются молекулами, которые выделены из их природного окружения, и включают в себя кДНК-клоны и геномные клоны. Выделенные молекулы ДНК данного изобретения не содержат других генов, с которыми они обычно связаны, но могут включать в себя природно встречающиеся 5′- и 3′-нетранслируемые районы, такие как промоторы и терминаторы. Идентификация связанных районов будет очевидной лицу с обычной квалификацией в данной области (см., например, Dynan and Tijan, Nature 316:774-78, 1985).
«Выделенным» полипептидом или белком является полипептид или белок, который обнаруживается в условиях, иных, чем его природное окружение, например, отдельно от крови и ткани животного. В предпочтительной форме, выделенный полипептид является по существу не содержащим других полипептидов, в частности, других полипептидов животного происхождения. Предпочтительно обеспечение полипептидов в высокоочищенной форме, т.е. имеющих чистоту более 95%, более предпочтительно чистоту более 99%. При использовании в этом контексте, термин «выделенный» не исключает присутствия того же самого полипептида в альтернативных физических формах, таких как димеры или альтернативно гликозилированные или дериватизованные формы.
Термин «неопластический», при упоминании в отношении клеток, обозначает клетки, подвергающиеся новой и отклоняющейся от нормы (аномальной) пролиферации, в частности, в ткани, где эта пролиферация является неконтролируемой и прогрессирующей, приводящей к неоплазме. Неопластические клетки могут быть либо злокачественными, т.е. инвазивными и метастатическими, либо доброкачественными.
Термин «функционально (операбельно) связанные», при ссылке на ДНК- сегменты, указывает, что эти сегменты расположены таким образом, что они функционируют совместно для их предполагаемых целей, например, транскрипция инициируется в промоторе и протекает через кодирующий сегмент до терминатора.
Термин «ортолог» обозначает полипептид или белок, полученный из одного вида, являющийся функциональной копией полипептида или белка из другого вида. Различия последовательностей среди ортологов являются результатом видообразования.
"Паралоги" являются отличающимися, но структурно родственными белками, производимыми организмом. Считается, что паралоги возникают в результате дупликации генов. Например, α-глобин, β-глобин и миоглобин являются паралогами друг друга.
Термин "полинуклеотид" обозначает одно- или двухцепочечный полимер дезоксирибонуклеотидных или рибонуклеотидных оснований, считываемых от 5′-конца к 3′-концу. Полинуклеотиды включают в себя РНК и ДНК и могут быть изолированы из природных источников, синтезированы in vitro или получены из комбинации природных и синтетических молекул. Размеры полинуклеотидов выражаются как пары нуклеотидов (сокращенно "п.н."), нуклеотиды ("нт") или тысячи пар нуклеотидов ("т.п.н."). Там, где позволяет контекст, два последние термина могут описывать полинуклеотиды, которые являются одноцепочечными или двухцепочечными. При применении этого термина к двухцепочечным молекулам его используют для обозначения общей длины, и должно быть понятно, что он является эквивалентным термину "пары нуклеотидов". Специалистам в данной области будет понятно, что две цепи двухцепочечного полинуклеотида могут слегка отличаться по длине и что их концы могут быть расположены ступенчато в результате ферментативного расщепления; таким образом, не все нуклеотиды в двухцепочечной полинуклеотидной молекуле могут быть спаренными.
"Полипептид" является полимером аминокислотных остатков, соединенных пептидными связями, получен ли он природным путем или синтетическим путем. Полипептиды, имеющие менее приблизительно 10 аминокислотных остатков, обычно называют "пептидами".
Термин "промотор" используется здесь в его признанном в данной области значении для обозначения части гена, содержащей последовательности ДНК, которые обеспечивают связывание РНК-полимеразы и инициацию транскрипции. Промоторные последовательности обнаруживаются обычно, но не всегда, в 5′-некодирующих районах генов.
"Белок" обозначает макромолекулу, содержащую одну или несколько полипептидных цепей. Белок может включать в себя также непептидные компоненты, такие как углеводные группы. Углеводы и другие непептидные компоненты могут быть присоединены к белку клеткой, в которой продуцируется данный белок, и будут варьироваться в зависимости от типа клетки. Белки определены здесь в терминах их аминокислотных каркасных структур; заместители, такие как углеводные группы, обычно не указаны, но тем не менее они могут присутствовать.
Термин "рецептор" используется здесь для обозначения связанного с клеткой белка, который связывается с биоактивной молекулой (т.е. "лигандом") и опосредует действие этого лиганда на клетку. Мембраносвязанные рецепторы характеризуются мультипептидной структурой, содержащей внеклеточный лигандсвязывающий домен и внутриклеточный эффекторный домен, который обычно участвует в трансдукции сигнала. Связывание лиганда с рецептором приводит к конформационному изменению в рецепторе, которое вызывает взаимодействие между эффекторным доменом и другой молекулой (молекулами) в клетке. Это взаимодействие, в свою очередь, приводит к изменению в метаболизме клетки. Метаболические события, связанные с взаимодействиями рецептор-лиганд, включают в себя транскрипцию генов, фосфорилирование, дефосфорилирование, увеличение продуцирования циклического АМФ, мобилизацию клеточного кальция, мобилизацию мембранных липидов, клеточную адгезию, гидролиз инозитоллипидов и гидролиз фосфолипидов. Обычно рецепторы могут быть мембраносвязанными, цитозольными или ядерными; мономерными (например, рецептор тиреоидстимулирующего гормона, бета-адренергический рецептор) или мультимерными (например, рецептор PDGF, рецептор гормона роста, рецептор IL-3, рецептор GM-CSF, рецептор G-CSF, рецептор эритропоэтина и рецептор IL-6).
Термин «секреторная сигнальная последовательность» обозначает последовательность ДНК, которая кодирует полипептид («секреторный пептид»), который, как компонент большего полипептида, направляет этот больший полипептид через секреторный путь клетки, в которой он синтезируется. Этот больший полипептид обычно расщепляется с удалением секреторного пептида во время прохождения через этот секреторный путь.
Термин "сплайсинговый вариант" используется здесь для обозначения альтернативных форм РНК, транскрибируемых из гена. Сплайсинговая вариация возникает природно посредством использования альтернативных сайтов сплайсинга в транскрибируемой молекуле РНК, или менее обычно между раздельно транскрибируемыми молекулами РНК, и может приводить к нескольким мРНК, транскрибируемым из одного и того же гена. Сплайсинговые варианты могут кодировать полипептиды, имеющие измененную аминокислотную последовательность. Термин сплайсинговый вариант используют здесь также для обозначения белка, кодируемого сплайсинговым вариантом мРНК, транскрибируемым из гена.
Должно быть понятно, что молекулярные массы и длины полимеров, определяемые непрецизионными аналитическими методами (например, гель-электрофорезом), являются приблизительными величинами. Когда такая величина выражается как "около" Х или "приблизительно" X, указанная величина Х должна пониматься как величина, определенная с точностью до ±10%.
Все цитируемые здесь ссылки включены в качестве ссылки в их полном виде.
Ген Zcyto21 кодирует полипептид из 200 аминокислот, показанный в SEQ ID NO:2. Сигнальная последовательность для Zcyto21 может быть предсказана как содержащая аминокислотный остаток 1 (Met) - аминокислотный остаток 19 (Ala) SEQ ID NO:2. Зрелый белок для Zcyto21 начинается при аминокислотном остатке 20 (Gly).
Ген Zcyto21 содержится в ВАС-последовательностях АС 011445 и АС 018477, которые картированы в хромосоме человека 19q13.13. Этот район хромосомы 19 может также содержать кластер интерферон-подобных генов. Консенсусная кДНК, показывающая полинуклеотидную последовательность Zcyto21, показана в SEQ ID NO:6, а кодируемый ею полипептид показан в SEQ ID NO:7.
Как описано ниже, данное изобретение обеспечивает выделенные полипептиды, имеющие аминокислотную последовательность, которая является по меньшей мере на 70%, по меньшей мере на 80% или по меньшей мере на 90%, 95%, 96%, 97%, 98% или 99% идентичной либо аминокислотным остаткам 20-200 SEQ ID NO:2, либо аминокислотным остаткам 1-200 SEQ ID NO:2. Данное изобретение обеспечивает также выделенные полипептиды, имеющие аминокислотную последовательность, которая является по меньшей мере на 70%, по меньшей мере на 80% или по меньшей мере на 90%, 95%, 96%, 97%, 98% или 99% идентичной либо аминокислотным остаткам 20-219 SEQ ID NO:9, либо аминокислотным остаткам 1-219 SEQ ID NO:9. Данное изобретение обеспечивает также выделенные полипептиды, имеющие аминокислотную последовательность, которая является по меньшей мере на 70%, по меньшей мере на 80% или по меньшей мере на 90%, 95%, 96%, 97%, 98% или 99% идентичной либо аминокислотным остаткам 20-203 SEQ ID NO:12, либо аминокислотным остаткам 1-203 SEQ ID NO:12. Данное изобретение включает в себя также полипептид, который дополнительно содержит сигнальную секреторную последовательность, которая находится в амино-концевом положении относительно первой аминокислотной последовательности, причем эта сигнальная секреторная последовательность содержит аминокислотные остатки 1-19 аминокислотной последовательности SEQ ID NO:2.
В общем предсказывается, что цитокины имеют четырех-α-спиральную структуру, причем спирали А, С и D являются наиболее важными в лиганд-рецепторных взаимодействиях и являются более высококонсервативными среди других членов этого семейства. Однако, интерфероны (INF) и, в частности, интерферон-альфа и интерферон-тау, характеризуются как шестиспиральные пучки. Спираль А интерферона эквивалентна спирали A Zcyto21; спираль В интерферона эквивалентна спирали С Zcyto21; спираль С интерферона эквивалентна спирали D Zcyto21 и спираль D интерферона эквивалентна спирали F Zcyto 21. Таким образом, петля между АВ-петлей и CD-петлей интерферона расширена в Zcyto21 таким образом, чтобы содержать короткие спирали В и Е Zcyto21.
Предсказывается, что спирали Zcyto21 являются следующими: спираль А определяется аминокислотными остатками 49 (Ser) - 63 (Leu); спираль В - аминокислотными остатками 76 (Asn) - 84 (Val); спираль С - аминокислотными остатками 89 (Val) - 104 (Ala); спираль D - аминокислотными остатками 111 (Glu) - 133 (Gln); спираль Е - аминокислотными остатками 137 (Thr) - 158 (Lys) и спираль F - аминокислотными остатками 163 (Gly) - 189 (Leu); как показано в SEQ ID NO:2. Остатки цистеина являются консервативными между Zcyto21 и IFN-α и могут образовывать межмолекулярную дисульфидную связь, в частности, для образования гомодимеров с дополнительными молекулами Zcyto21. Дополнительный анализ Zcyto21 на основе множественных сопоставлений предсказывает, что цистеины в положениях аминокислотных остатков 34 и 131 и 68 и 164 (как показано в SEQ ID NO:2) будут образовывать внутримолекулярные дисульфидные связи. Цистеин в остатке 190 является свободным и может образовывать межмолекулярную дисульфидную связь. Соответствующие полинуклеотиды, кодирующие районы, домены, мотивы, остатки и последовательности Zcyto21, описанные здесь, показаны в SEQ ID NO:1. Вырожденная полинуклеотидная последовательность SEQ ID NO:2 показана в SEQ ID NO:3. Вырожденная полинуклеотидная последовательность SEQ ID NO:9 показана в SEQ ID NO:10. Вырожденная полинуклеотидная последовательность SEQ ID NO:12 показана в SEQ ID NO:13.
Подробный мутационный анализ мышиного IL-2 (Zurawski et al., EMBO J. 12:5113-5119, 1993) показывает, что остатки в спиралях А и С являются важными для связывания с IL-2Rβ; критическими остатками являются Asp34, Asn99 и Asn103, Множественные остатки в петле А/В и спирали В мышиного IL-2 являются важными для связывания с IL-2Rα, тогда как только единственный остаток, Gln141 в спирали D, является первостепенным для связывания с IL-2Rα. Подобным образом, спирали А и С являются сайтами взаимодействия между IL-4 и IL-4Rα (структурно сходного с IL-2Rα), и остатки внутри спирали D являются критическими для взаимодействия IL-2Rα (Wang et al., Proc. Natl. Acad. Sci. USA 94:1657-1662, 1997; Kruse et al., EMBO J. 11:3237-3244, 1992). В частности, мутация Tyr124 в Asp в IL-4 человека создает антагонист, который связывается с IL-4Rα, но не с IL-2Rα и, следовательно, не может передавать сигнал (Kruse et al. ibid., 1992).
Цитокины с четырехспиральными пучками группируются также по длине из компонентов-спиралей. Цитокины «длинноспиральной» формы состоят из спиралей с 24-30 остатками и включают в себя IL-6, цилиарный нейтрофический фактор (CNTF), ингибирующий лейкоз фактор (LIF) и гормон роста человека (hGH). Цитокины «короткоспиральной» формы обычно состоят из спиралей с 18-21 остатками и включают в себя IL-2, IL-4 и GM-CSF. Исследования с использованием CNTF и IL-6 демонстрируют, что спираль CNTF может заменять эквивалентную спираль в IL-6, придавая свойства связывания CNTF полученной химере. Таким образом, по-видимому, функциональные домены четырехспиральных цитокинов определяются на основе структурной гомологии, независимо от идентичности последовательностей, и могут сохранять функциональную целостность в химере (Kallen et al., J.Biol. Chem. 274:11859-11867, 1999). Таким образом, спиральные домены Zcyto 21 будут применимы для получения химерных слитых молекул, в частности, с другими интерферонами, для определения и модуляции специфичности связывания рецептора. Особый интерес представляют слитые белки, которые объединяют спиральные и петлевые домены из интерферонов и цитокинов, таких как IFN-α, IL-10, гормон роста человека.
мРНК Zcyto21 была идентифицирована в тканях головного мозга, островков, предстательной железы, яичка, гипофиза, плаценты, опухоли яичника, опухоли легкого, ректальной опухоли и опухоли яичника, а также активированной линии иммунных клеток (CD3+) и линии эпителиальных клеток предстательной железы, которые были трансформированы вирусом папилломы человека IV (HPVS).
Данное изобретение обеспечивает полинуклеотидные молекулы, в том числе молекулы ДНК и РНК, которые кодируют описанные здесь полипептиды Zcyto21. Специалистам в данной области будет понятно, что, в связи с вырожденностью генетического кода, возможна значительная вариация среди этих полинуклеотидных молекул. SEQ ID NO:3, SEQ ID NO:10 и SEQ ID NO:13 являются вырожденными последовательностями ДНК, которые включают в себя все ДНК, которые кодируют полипептид Zcyto21 SEQ ID NO:2, 9 и 12, соответственно. Специалистам в данной области будет понятно, что вырожденная последовательность SEQ ID NO:3, например, обеспечивает все последовательности РНК, кодирующие SEQ ID NO:2, путем замены U (урацилом) Т (тимина). Таким образом, кодирующие полипептид Zcyto21 полинуклеотиды, содержащие район от нуклеотида 1 или 58 до нуклеотида 603 SEQ ID NO:3, и их РНК-эквиваленты рассматриваются данным изобретением. Таблица 1 дает однобуквенные коды, используемые в SEQ ID NO:3 для обозначения положений вырожденных нуклеотидов. "Разрешения" представляют собой нуклеотиды, обозначенные кодовой буквой. "Комплемент" указывает код для комплементарного нуклеотида (комплементарных нуклеотидов). Например, код Y обозначает либо С, либо Т, а его комплемент R обозначает А или G, причем А является комплементарным Т, а G является комплементарным С.
Вырожденные кодоны, используемые в SEQ ID NO:3, 10 и 13, включающие в себя все возможные кодоны для конкретной аминокислоты, представлены в таблице 2.
Специалисту с обычной квалификацией в данной области будет понятно, что некоторая двусмысленность вводится в определение вырожденного кодона, представляющего все возможные кодоны, кодирующие каждую аминокислоту. Например, вырожденный кодон для серина (WSN) может, в некоторых обстоятельствах, кодировать аргинин (AGR), а вырожденный кодон для аргинина (MGN) может, в некоторых обстоятельствах, кодировать серин (AGY). Сходная взаимосвязь существует между кодонами, кодирующими фенилаланин и лейцин. Таким образом, некоторые полинуклеотиды, охватываемые вырожденной последовательностью, могут кодировать вариантные аминокислотные последовательности, но специалист с обычной квалификацией в данной области сможет легко идентифицировать такие вариантные последовательности сравнением с аминокислотной последовательностью SEQ ID NO:2. Вариантные последовательности могут быть легко тестированы на функциональность, как описано здесь.
Специалисту с обычной квалификацией в данной области будет также понятно, что различные виды могут проявлять "предпочтительное (преферентивное) использование кодонов". В общем, см., Grantham, et al., Nuc. Acids Res. 8:1893-912, 1980; Haas, et al, Curr. Biol. 6:315-24,1996; Wain-Hobson, et al., Gene 13:355-64, 1981; Grosjean and Fiers, Gene 18:199-209, 1982; Holm, Nuc. Acids Res. 14:3075-87, 1986; Ikemura, J.Mol. Biol. 158:573-97, 1982. В применении здесь, термин "предпочтительное (преферентивное) использование кодонов" или "предпочтительные (преферентивные) кодоны" является термином данной области, относящимся к кодонам трансляции белка, которые наиболее часто используются в клетках определенных видов, отдавая, следовательно, предпочтение одному или немногим представителям возможных кодонов, кодирующих каждую аминокислоту (См. таблицу 3). Например, аминокислота треонин (Thr) может кодироваться АСА, АСС, ACG или ACT, но в клетках млекопитающих наиболее обычно используемым кодоном является АСС; в других видах, например, в клетках насекомых, дрожжей, вирусов или бактерий, могут быть предпочтительными другие кодоны Tnr. Предпочтительные кодоны для конкретных видов могут быть введены в полинуклеотиды данного изобретения различными способами, известными в этой области. Введение предпочтительных последовательностей кодонов в рекомбинантную ДНК может, например, усиливать продуцирование этого белка, делая трансляцию белка более эффективной в конкретном типе клеток или виде. Таким образом, вырожденная последовательность кодонов, описанная в SEQ ID NO:3, служит в качестве матрицы для оптимизации экспрессии полинуклеотидов в различных типах клеток и видах, обычно используемых в данной области и описанных здесь. Последовательности, содержащие предпочтительные кодоны, могут быть тестированы и оптимизированы для экспрессии в различных видах и испытаны на функциональность, как описано здесь.
Как отмечалось ранее, выделенные полинуклеотиды данного изобретения включают в себя ДНК и РНК. Способы получения ДНК и РНК хорошо известны в данной области. Обычно РНК выделяют из ткани или клетки, которая продуцирует большие количества РНК Zcyto21. Такие ткани и клетки идентифицируют Нозерн-блоттингом (Thomas, Proc. Natl. Acad. Sci. USA 77:5201, 1980) или скринингом кондиционированной среды из различных типов клеток на активность на клетках-мишенях или ткани-мишени. После идентификации активности или продуцирующей РНК клетки или ткани, тотальная РНК может быть получена с использованием экстракции изотиоцианатом гуанидиния с последующим выделением центрифугированием в градиенте CsCl (Chirgwin et al., Biochemistry 18:52-94, 1979). Поли (А)+ РНК получают из тотальной РНК с использованием способа Aviv and Leder, Proc. Natl. Acad. Sci. USA 69:1408-1412 (1972). Комплементарную ДНК (кДНК) получают из поли (A)+ РНК при помощи известных способов. Альтернативно, может быть выделена геномная ДНК. Затем полинуклеотиды, кодирующие полипептиды Zcyto21, идентифицируют и выделяют, например, с использованием гибридизации или полимеразной цепной реакции (ПЦР).
Более длинный клон, кодирующий Zcyto21, может быть получен общепринятыми процедурами клонирования. Клоны комплементарной ДНК (кДНК) являются предпочтительными, хотя для некоторых применений (например, экспрессиии в трансгенных животных) могут быть предпочтительными использование геномного клона или модификация клона кДНК с целью включения по меньшей мере одного геномного интрона. Способы получения клонов кДНК и геномных клонов хорошо известны и находятся в пределах обычной квалификации в данной области и включают в себя использование описанной здесь последовательности или ее частей для зондирования или праймирования библиотеки. Экспрессионные библиотеки могут быть зондированы антителами к фрагментам рецептора Zcyto21 или другими партнерами специфического связывания.
Далее, данное изобретение обеспечивает копии полипептидов и полинуклеотидов из других видов (ортологи). Эти виды включают в себя, но не ограничиваются ими, виды млекопитающих, птиц, земноводных, пресмыкающихся, рыб, насекомых и другие виды позвоночных и беспозвоночных. Особый интерес представляют собой полипептиды Zcyto21 из других видов млекопитающих, в том числе полипептиды мышей, свиней, овец, коров, псовых, кошачьих, лошадиных видов и полипептиды других приматов. Ортологи Zcyto21 человека могут быть клонированы с использованием информации и композиций, обеспеченных данным изобретением, в сочетании с общепринятыми способами клонирования. Например, кДНК может быть клонирована с использованием мРНК, полученной из типа ткани или клеток, которые экспрессируют Zcyto21, как описано здесь. Подходящие источники мРНК могут быть идентифицированы зондированием Нозерн-блотов зондами, сконструированными из описанных здесь последовательностей. Затем готовят библиотеку из мРНК позитивной ткани или линии клеток. Затем кодирующая полипептид Zcyto21 кДНК может быть выделена различными способами, такими как зондирование полной или частичной кДНК человека или одним или несколькими наборами вырожденных зондов, основанных на описанных последовательностях. Эта кДНК может быть также клонирована с использованием полимеразной цепной реакции, или ПЦР (Mullis, патент США №4 683 202), с рименением праймеров, сконструированных из описанной здесь репрезентативной последовательности Zcyto21 человека. В дополнительном способе, эта библиотека кДНК может быть использована для трансформации или трансфекции клеток-хозяев, и экспрессия представляющей интерес кДНК может быть детектирована антителом к полипептиду Zcyto21, исследованиями связывания или анализами активности. Подобные способы могут быть применены также для выделения геномных клонов.
Специалистам в данной области будет понятно, что последовательность, описанная в SEQ ID NO:1, представляет отдельный аллель ДНК Zcyto21 человека и что ожидается существование аллельной вариации и альтернативного сплайсинга. Аллельные варианты этой последовательности могут быть клонированы зондированием библиотек кДНК или геномных библиотек из различных индивидуумов в соответствии со стандартными процедурами. Аллельные варианты последовательности ДНК, показанной в SEQ ID NO:1, в том числе варианты, содержащие молчащие мутации, и варианты, в которых мутации приводят к изменениям в последовательности аминокислот, находятся в объеме данного изобретения, так же как и белки, которые являются аллельными вариантами SEQ ID NO:2. кДНК, генерируемые из образованных альтернативным сплайсингом мРНК, которые сохраняют свойства полипептида Zcyto21, включены в объем данного изобретения, так же как полипептиды, кодируемые такими кДНК и мРНК. Аллельные варианты и сплайсинговые варианты этих последовательностей могут быть клонированы зондированием библиотек кДНК или геномных библиотек из различных индивидуумов или тканей в соответствии со стандартными процедурами, известными в данной области. Примеры альтернативно сплайсированных вариантов показаны в SEQ ID NO:8 (SEQ ID NO:9 для соответствующего полипептида) и в SEQ ID NO:11 (SEQ ID NO:12 для соответствующего полипептида). Пример аллельного варианта показан в SEQ ID NO:4, которая соответствует полипептидной последовательности, показанной в SEQ ID NO:5. Существует полиморфизм между полипептидной последовательностью, показанной в SEQ ID NO:1, и последовательностью, показанной в SEQ ID NO:4, при номере нуклеотида 572. Этот полиморфизм может создавать антагонист Zcyto21 или молекулу пониженной или измененной функции, которая может приводить к более высокой вероятности восприимчивости заболевания.
Данное изобретение обеспечивает также реагенты, которые найдут применение в диагностических приложениях. Например, ген Zcyto21, зонд, содержащий ДНК или РНК Zcyto21 или их субпоследовательность, могут быть использованы для определения, присутствует ли ген Zcyto21 на хромосоме человека, например, хромосоме 19, или имела ли место мутация гена. Zcyto21 локализован в районе q13.13 хромосомы 19. Детектируемые хромосомные аберрации в локусе гена Zcyto21 включают в себя, но не ограничиваются ими, анеуплоидию, изменения копийности гена, потерю гетерогенности (LOH), транслокации, инсерции, делеции, изменения сайтов рестрикции и реаранжировки. Такие аберрации могут быть детектированы с использованием полинуклеотидов данного изобретения путем применения молекулярно-генетических способов, таких как анализ полиморфизма длины фрагментов рестрикции (RELP), анализ коротких тандемных повторов (STR) с использованием способов ПЦР и других способов анализа генетического сцепления, известных в данной области (Sambrook et al., ibid; Ausubel et al., ibid; Marian, Chest 108:255-65, 1995).
Точное знание положения гена может быть полезным для ряда целей, в том числе: 1) определения, является ли последовательность частью существующего контига, и получения дополнительных окружающих генетических последовательностей в различных формах, таких как YAC, ВАС или кДНК-клоны; 2) обеспечения возможного гена-кандидата для наследственного заболевания, который обнаруживает сцепление с тем же самым хромосомным районом; и 3) перекрестно-ссылочных модельных организмов, таких как мышь, которые могут способствовать определению, какую функцию может иметь конкретный ген.
Например, Delague et al. (Am. J.Hum. Genet. 67:236-243, 2000) идентифицировали, что болезнь Шарко-Мари-Тута (наследственная невральная амиотрофия) локализована в 19q13.1-13.3 (Delague et al., Am. J.Hum. Genet. 67:236-243, 2000).
Диагностика может помочь врачам в определении типа заболевания и подходящей связанной с ним терапии или помощи в генетической консультации. Сами по себе антитела против Zcyto21, полинуклеотиды и полипептиды Zcyto21 могут быть использованы для обнаружения полипептида, мРНК Zcyto21 или антител против Zcyto21, служа в качестве маркеров, и могут быть непосредственно использованы для обнаружения генетических болезней или раков, как описано здесь, с использованием способов, известных в данной области и описанных здесь. Далее, полинуклеотидные зонды Zcyto21 могут быть использованы для обнаружения отклонений от нормы или генотипов, связанных с делециями и транслокациями хромосомы 19, связанными с болезнями человека, или других транслокаций, участвующих в злокачественном прогрессировании опухолей, или других мутаций 19q13.13, которые, как ожидается, участвуют в хромосомных реаранжировках в злокачественности; или в других раках. Подобным образом, полинуклеотидные зонды Zcyto21 могут быть использованы для обнаружения отклонений от нормы или генотипов, связанных с трисомией хромосомы 19q13.13 и потерей хромосомы, связанной с болезнями человека или самопроизвольным абортом. Таким образом, полинуклеотидные зонды Zcyto21 могут быть использованы для обнаружения аномалий или генотипов, связанных с этими дефектами.
В общем, диагностические способы, используемые в анализах генетического сцепления, для обнаружения генетического нарушения или отклонения от нормы в пациенте, известны в данной области. Аналитические зонды будут обычно иметь длину по меньшей мере 20 п.н., хотя могут быть использованы несколько более короткие зонды (например, 14-17 п.н.). ПЦР праймеры имеют длину по меньшей мере 5 нт, предпочтительно 15 или более, еще более предпочтительно 20-30 нт. Для макроскопического анализа генов или хромосомной ДНК полинуклеотидный зонд Zcyto21 может содержать полный экзон или большую часть гена. Экзоны легко определяются специалистом с квалификацией в данной области сравнением последовательностей Zcyto21 (SEQ ID NO:1) с геномной ДНК для Zcyto21. Обычно методы диагностики, применяемые в анализе гинетического сцепления для определения отклонений от нормы или абберраций у пациента, известны специалистам в данной области. В общем, эти диагностические способы включают в себя стадии (а) получения генетической пробы из потенциально заболевшего пациента, заболевшего пациента или потенциально не имеющего заболевания носителя рецессивного аллеля заболевания; (b) получения первого продукта реакции инкубированием этой генетической пробы с полинуклеотидным зондом Zcyto21, причем этот полинуклеотид будет гибридизоваться с комплементарной полинуклеотидной последовательностью, например, в RFLP-анализе, или инкубированием этой генетической пробы со смысловым или антисмысловым праймерами в подходящих условиях реакции ПЦР; (iii) визуализации этого первого продукта реакции с использованием гель-электрофореза и/или другого известного способа, например, визуализации первого продукта реакции с полинуклеотидным зондом Zcyto21, где этот полинуклеотид будет гибридизоваться с комплементарной последовательностью первой реакции; и iv) сравнения визуализированного первого продукта реакции со вторым продуктом контрольной реакции генетической пробы из пациента дикого типа. Различие между первым продуктом реакции и продуктом контрольной реакции указывает на генетическое отклонение от нормы в заболевшем или потенциально заболевшем пациенте или присутствие гетерозиготного рецессивного фенотипа-носителя для незаболевшего пациента или присутствия генетического дефекта в опухоли из заболевшего пациента или присутствие генетического отклонения от нормы в плоде или преимплантационном зародыше. Например, различие в картине рестрикционных фрагментов, длине ПЦР-продуктов, длине повторяющихся последовательностей в генетическом локусе Zcyto21 и т.п. являются показателями генетического отклонения от нормы, генетической аберрации или аллельного различия в сравнении с нормальным контролем дикого типа. Контролями могут быть непораженные члены семьи или неродственные индивидуумы, в зависимости от теста и доступности проб. Генетические пробы для применения в данном изобретении включают в себя геномные ДНК, мРНК и кДНК, выделенные из любой ткани или другой биологической пробы из пациента, такие как, но не только, кровь, слюна, сперма, эмбриональные клетки, амниотическая жидкость и т.п. Полинуклеотидным зондом или праймером может быть РНК или ДНК, и он будет содержать часть SEQ ID NO:1, комплемент SEQ ID NO:1 или их РНК-эквивалент. Такие способы анализа результатов генетического сцепления относительно фенотипов заболеваний человека хорошо известны в данной области. В отношении ссылки на способы в диагностике, основанные на ПЦР, см., в общем, Mathew (ed.) Protocols in Human Molecular Genetics (Humana Press, Inc. 1991), White (ed.), PCR Protocols: Current Methods and Applications (Humana Press, Inc. 1993), Cotter (ed.) Molecular Diagnosis of Cancer (Humana Press, Inc. 1996), Hanausek and Walaszek (eds.), Tumor Marker Protocols (Humana Press, Inc. 1998), Lo (ed.) Clinical Applications of PCR (Humana Press, Inc. 1998) и Meltzer (ed.) PCR in Bioanalysis (Humana Press, Inc. 1998)).
Мутации, связанные с локусом Zcyto21, могут быть обнаружены с использованием молекул нуклеиновых кислот данного изобретения посредством применения стандартных способов для прямого мутационного анализа, такого как анализ полиморфизма длин рестрикционных фрагментов, анализ коротких тандемных повторов с использованием способов ПЦР, анализ амплификационной системы идентификации мутаций, обнаружение полиморфизма одноцепочечной конформации, способы расщепления РНКазой, денатурирующий градиентный гель-электрофорез, флуоресцентный анализ ошибочного спаривания и другие генетические способы анализа, известные в данной области (см., например, Mathew (ed.) Protocols in Human Molecular Genetics (Humana Press, Inc. 1991), Marian, Chest 108:255 (1995). Coleman and Tsongalis, Molecular Diagnostics (Humana Press, Inc. 1996), Elles (ed.) Molecular Diagnosis of Genetic Diseases (Humana Press, Inc. 1996), Landegren (ed.) Laboratory Protocols for Mutation Detection (Oxford University Press 1996), Birren et al., (eds.), Genome Analysis, Vol.2: Detecting Genes (Cold Spring Harbor Laboratory Press 1998), Dracopoli et al., (eds.), Current Protocols in Human Genetics (John Wiley & Sons 1998) и Richards and Ward, "Molecular Diagnostic Testing," in Principles of Molecular Medicine, pages 83-88 (Humana Press, Inc. 1998)). Прямой анализ гена Zcyto21 на мутацию может быть выполнен с использованием геномной ДНК субъекта. Способы амплификации геномной ДНК, полученной, например, из лимфоцитов периферической крови, хорошо известны специалистам с квалификацией в данной области (см., например, Dracopoli et al., (eds.), Current Protocols in Human Genetics, pages 7.1.6-7.1.7 (John Wiley & Sons 1998)).
В вариантах данного изобретения, выделенные кодирующие Zcyto21 молекулы нуклеиновых кислот могут гибридизоваться в строгих условиях с молекулами нуклеиновых кислот, имеющими нуклеотидную последовательность SEQ ID NO:1, с молекулами нуклеиновых кислот, имеющими нуклеотидную последовательность нуклеотидов 58-603 SEQ ID NO:1, или молекулами нуклеиновых кислот, имеющими нуклеотидную последовательность, комплементарную SEQ ID NO:1. Обычно, строгие условия выбирают таким образом, чтобы температура была на приблизительно 5°С ниже, чем точка термического плавления (Тm) для конкретной последовательности при определенных ионной силе и рН. Тm является температурой (при определенных ионной силе и рН), при которой 50% последовательности-мишени гибридизуются с точно совместимым зондом.
Пара молекул нуклеиновых кислот, такая как ДНК-ДНК, РНК-РНК и ДНК-РНК, может гибридизоваться, если эти нуклеотидные последовательности имеют некоторую степень комплементарности. Гибриды могут выносить ошибочно спаренные пары оснований в двойной спирали, но степень ошибочного спаривания влияет на стабильность этого гибрида. Тm ошибочно спаренного гибрида уменьшается на 1°С для каждых 1-1,5% ошибочных спариваний оснований. Варьирование строгости условий гибридизации позволяет контролировать степень ошибочного спаривания, которая будет присутствовать в гибриде. Степень строгости увеличивается по мере увеличения температуры гибридизации и по мере уменьшения ионной силы буфера для гибридизации.
Специалист с квалификацией в данной области вполне сможет адаптировать эти условия для применения к конкретному полинуклеотидному гибриду. Тm для конкретной последовательности-мишени является температурой (при определенных условиях), при которой 50% последовательности-мишени будут гибридизоваться с точно совместимым зондом. Эти условия, которые влияют на Тm, включают в себя размер и содержание пар оснований полинуклеотидного зонда, ионную силу гибридизационного раствора и присутствие дестабилизирующих агентов в гибридизационном растворе. Многочисленные уравнения для расчета Тm известны в данной области и являются специфическими для ДНК, РНК и ДНК-РНК-гибридов и полинуклеотидных последовательностей-зондов варьирующей длины (см., например, Sambrook et al. (Molecular Cloning. A Laboratory Manual, Second Edition (Cold Spring Harbor Laboratory Press 1989); Ausubel et al., (eds.), Current Protocols in Molecular Biology (John Wiley and Sons, Inc., 1987); Berger and Kimmel (eds.), Guide to Molecular Cloning Techniques (Academic Press, Inc. 1987); и Wetmur, Crit. Rev. Biochem. Mol. Biol. 26:227 (1990)). Программное обеспечение для анализа последовательности, такое как OLIGO 6.0 (LSR; Long Lake, MN) и Primer Premier 4.0 (Premier Biosoft International; Palo Alto, CA), а также сайты в Интернете, являются доступными инструментами для анализа конкретной последовательности и расчета Тm на основе определенных пользователем критериев. Такие программы могут также анализировать конкретную последовательность при определенных условиях и идентифицировать подходящие последовательности зондов. Обычно, гибридизацию более длинных полинуклеотидных последовательностей, >50 пар оснований, выполняют при температурах на приблизительно 20-25°С ниже рассчитанной Тm. Для меньших зондов, <50 пар оснований, гибридизацию обычно проводят при Тm или на 5-10°С ниже рассчитанной Тm. Это позволяет достичь максимальной скорости гибридизации для гибридов ДНК-ДНК и ДНК-РНК.
После гибридизации молекулы нуклеиновых кислот могут быть промыты для удаления негибридизовавшихся молекул нуклеиновых кислот в строгих условиях или в условиях высокой строгости. Типичные строгие условия промывки включают в себя промывку в растворе 0,5х-2х SSC с 0,1% додецилсульфатом натрия (ДСН) при 55-65°С. То есть, молекулы нуклеиновых кислот, кодирующие вариантный пептид Zcyto21, гибридизуются с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплементом) в строгих условиях промывки, где строгость промывки эквивалентна 0,5х-2х SSC с 0,1% ДСН при 55-65°С, в том числе 0,5х SSC с 0,1% ДСН при 55°С, или 2х SSC с 0,1% ДСН при 65°С. Специалист с квалификацией в данной области сможет легко создать эквивалентные условия, например, заменой SSC с использованием SSPE в промывном растворе.
Типичные условия высокой строгости включают в себя промывку в растворе 0,1х-0,2х SSC с 0,1% додецилсульфатом натрия (ДСН) при 50-65°С. Другими словами, молекулы нуклеиновых кислот, кодирующие вариантный пептид Zcyto21, гибридизуются с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплементом) в условиях промывки высокой строгости, где строгость промывки эквивалентна 0,1х-0,2х SSC с 0,1% ДСН при 50-65°С, в том числе 0,1х SSC с 0,1% ДСН при 50°С, или 0,2х SSC с 0,1% ДСН при 65°С.
Данное изобретение обеспечивает также выделенные полипептиды Zcyto21, которые имеют по существу одинаковую идентичность последовательности с полипептидами SEQ ID NO:2 и их ортологами. Термин "по существу одинаковую идентичность последовательности" используется здесь для обозначения полипептидов, имеющих по меньшей мере 70%-ную, по меньшей мере 80%-ную, по меньшей мере 90%-ную, по меньшей мере 95%-ную или большую, чем 95%-ную, 96%-ную, 97%-ную, 98%-ную или 99%-ную, идентичность последовательности с последовательностями, показанными в SEQ ID NO:2, или их ортологами. Данное изобретение включает в себя также полипептиды, которые содержат аминокислотную последовательность, имеющую по меньшей мере 70%-ную, по меньшей мере 80%-ную, по меньшей мере 90%-ную, по меньшей мере 95%-ную или большую, чем 95%-ную, 96%-ную, 97%-ную, 98%-ную или 99%-ную, идентичность последовательности с последовательностью аминокислотных остатков 1-200 или 20-200 SEQ ID NO:2. Данное изобретение включает в себя дополнительно молекулы нуклеиновых кислот, которые кодируют такие полипептиды. Способы определения процентной идентичности описаны ниже.
Данное изобретение рассматривает также вариантные молекулы нуклеиновых кислот Zcyto21, которые могут быть идентифицированы с использованием двух критериев: определения сходства между кодируемым полипептидом и аминокислотной последовательностью SEQ ID NO:2 и/или гибридизационного анализа, описанного выше. Такие варианты Zcyto21 включают в себя молекулы нуклеиновых кислот: (1) которые гибридизуются с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент), при строгих условиях промывки, где строгость промывки эквивалентна 0,5х-2х SSC с 0,1% ДСН при 55-65°С; или (2) которые кодируют полипептид, имеющий по меньшей мере 70%-ную, по меньшей мере 80%-ную, по меньшей мере 90%-ную, по меньшей мере 95%-ную или большую, чем 95%-ную, 96%-ную, 97%-ную, 98%-ную или 99%-ную, идентичность последовательности с аминокислотной последовательностью SEQ ID NO:2. Альтернативно, варианты Zcyto21 могут быть охарактеризованы как молекулы нуклеиновых кислот: (1) которые гибридизуются с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1 (или ее комплемент), при условиях промывки высокой строгости, где строгость промывки эквивалентна 0,1х-0,2х SSC с 0,1% ДСН при 50-65°С; или (2) которые кодируют полипептид, имеющий по меньшей мере 70%-ную, по меньшей мере 80%-ную, по меньшей мере 90%-ную, по меньшей мере 95%-ную или большую, чем 95%-ную, идентичность последовательности с аминокислотной последовательностью SEQ ID NO:2.
Процентную идентичность последовательности определяют общепринятыми способами. См., например, Altschul et al., Bull. Math, Bio. 48:603 (1986) и Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA 89:10915 (1992). Вкратце, две последовательности аминокислот сопоставляют выстраиванием для оптимизации оценок сопоставления с использованием штрафа (penalty) открывания гэпа 10 и штрафа (penalty) удлинения гэпа 1 и оценочной матрицы "BLOSUM62" Henikoff and Henikoff (ibid.), показаной в таблице 3 (аминокислоты представлены с использованием стандартных однобуквенных кодов).

Специалистам в данной области должно быть понятно, что имеется много установленных алгоритмов для сопоставления двух аминокислотных последовательностей. Алгоритм поиска сходства "FASTA" Pearson и Lipman представляет собой подходящий способ сопоставления белков для исследования уровня идентичности, который является общим у описанной здесь аминокислотной последовательности и аминокислотной последовательности возможного варианта Zcyto21. Алгоритм FASTA описан Pearson and Lipman, Proc. Natl. Acad. Sci. USA 85:2444 (1988) и Pearson, Meth. Enzymol. 183:63 (1990).
Вкратце, FASTA сначала характеризует сходство последовательностей путем идентификации районов, общих у запрашиваемой (query) последовательности (например, SEQ ID NO:2) и тест-последовательности, которые имеют наивысшую плотность либо идентичностей (если переменная ktup равна 1), либо пар идентичностей (если ktup=2), без учета консервативных аминокислотных замен, инсерций или делеций. Затем десять районов с наиболее высокой плотностью идентичностей повторно оценивают сравнением сходства всех спаренных аминокислот с использованием матрицы аминокислотных замен и концы этих районов «подравнивают» для включения только тех остатков, которые способствуют наивысшей оценке. Если имеются несколько районов с оценками, большими, чем «предельная» величина (рассчитанная по предварительно определенной формуле на основе длины данной последовательности и величины ktup), то приведенные в порядок подравниванием первоначальные районы исследуют для определения, могут ли эти районы быть соединены с образованием приближенного сопоставления с гэпами. Наконец, районы с наивысшей оценкой двух аминокислотных последовательностей сопоставляют с использованием модификации алгоритма Needleman-Wunsch-Sellers (Needleman and Wunsch, J.Mol. Biol. 48:444 (1970); Sellers, SIAM J.Appl. Math. 26:787 (1974)), который делает возможными инсерции и делеции аминокислот. Предпочтительными параметрами для анализа FASTA являются: ktup=1, штраф (penalty) открывания гэпа=10, штраф (penalty) удлинения гэпа=1 и матрица замен=BLOSUM62. Эти параметры могут быть введены в программу FASTA модификацией файла оценочной матрицы ("SMATRIX"), как объясняется в Приложении 2 Pearson, Meth. Enzymol. 183:63 (1990).
Программа FASTA может быть использована для определения идентичности последовательности молекул нуклеиновых кислот с использованием приведенного выше отношения. Для сравнений нуклеотидных последовательностей величина ktup может быть в диапазоне между 1 и 6, предпочтительно от 3 до 6, наиболее предпочтительно 3, с другими параметрами, установленными по умолчанию.
Вариантные полипептиды Zcyto21 или полипептиды с по существу одинаковой идентичностью последовательностей характеризуются как имеющие одну или несколько аминокислотных замен, делеций или присоединений. Эти изменения предпочтительно имеют минорный характер, то есть, замены консервативных аминокислот (см. таблицу 4) и другие замены, которые не влияют значимо на укладку или активность полипептида; небольшие делеции, обычно от одной до ˜30 аминокислот; и небольшие амино- или карбоксил-концевые удлинения, такие как амино-концевой остаток метионина, небольшой линкерный пептид до приблизительно 20-25 остатков, или аффинная метка. Таким образом, данное изобретение включает в себя полипептиды из приблизительно от 149 до приблизительно 230 аминокислотных остатков, которые включают в себя последовательность, которая по меньшей мере на 70%, предпочтительно по меньшей мере на 90% и более предпочтительно на 95%, 96%, 97%, 98%, 99% или более идентична соответствующему району SEQ ID NO:2. Полипептиды, содержащие аффинные метки, могут дополнительно содержать сайт протеолитического расщепления между полипептидом Zcyto21 и этой аффинной меткой. Предпочтительные подобные сайты включают в себя сайты расщепления тромбином и сайты расщепления фактором Ха.
Могут быть произведены определения аминокислотных остатков, которые составляют районы или домены, являющиеся критическими для сохранения структурной целостности. В этих районах можно определить специфические остатки, которые будут более или менее устойчивыми к изменению и сохранению общей третичной структуры молекулы. Способы анализа структуры последовательности включают в себя, но не ограничиваются ими, сопоставление множественных последовательностей с высокой идентичностью аминокислот или нуклеотидов, склонности к образованию вторичной структуры, бинарные распределения, комплементарную упаковку и скрытые полярные взаимодействия (Barton, Current Opin. Struct. Biol. 5:372-376, 1995 и Cordes et al., Current Opin. Struct. Biol. 6:3-10, 1996). Обычно, при конструировании модификаций для молекул или идентификации специфических фрагментов определение структуры будет сопровождаться оценкой активности модифицированных молекул.
Изменения аминокислотной последовательности выполняют в полипептидах Zcyto21 таким образом, чтобы минимизировать разрушение структуры более высокого порядка, существенной для биологической активности. Например, когда полипептид Zcyto21 содержит одну или несколько спиралей, изменения в аминокислотных остатках будут выполняться таким образом, чтобы не нарушить геометрию спиралей и других компонентов молекулы, где изменения в конформации уменьшают некоторую критическую функцию, например, связывание данной молекулы с ее партнерами связывания. Эффекты изменений аминокислотной последовательности могут быть прогнозированы, например, компьютерным моделированием, как описано выше, или определены анализом структуры кристаллов (см., например, Lapthorn et al., Nat. Struct. Biol. 2:266-268, 1995). Другие способы, которые хорошо известны в данной области, сравнивают укладку вариантного белка со стандартной молекулой (например, нативным белком). Например, может быть сделано сравнение распределения цистеина в вариантной и стандартной молекулах. Масс-спектрометрия и химическая модификация с использованием восстановления и алкилирования обеспечивают способы для определения остатков цистеина, которые связаны дисульфидными связями или свободны от таких связей (Bean et al., Anal. Biochem. 201:216-226, 1992; Gray, Protein Sci. 2:1732-1748, 1993 и Patterson et al., Anal. Chem. 66:3727-3732, 1994). Обычно считается, что, если модифицированная молекула не имеет такого же распределения цистеинов, что и стандартная молекула, укладка может быть нарушенной. Другим хорошо известным и общепринятым способом измерения укладки является круговой дихроизм (КД). Измерение и сравнение КД-спектров, генерируемых модифицированной молекулой и стандартной молекулой, является рутиной (Johnson, Proteins 7:205-214, 1990). Кристаллография представляет собой другой хорошо известный способ для анализа укладки и структуры. Ядерный магнитный резонанс (ЯМР), ферментативное пептидное картирование и картирование эпитопов также являются известными способами для анализа сходства укладки и структуры между белками и полипептидами (Schaanan et al., Science 257:961-964, 1992).
Может быть получен профиль гидрофильности Hopp/Woods белковой последовательности Zcyto21, показанной в SEQ ID NO:2 (Hopp et al., Proc. Natl. Acad. Sci. 78:3824-3828, 1981; Hopp, J.Immun. Meth. 88:1-18, 1986 и Triquier et al., Protein Engineering 11:153-169, 1998). Этот профиль основан на скольжении окна для шести остатков. Скрытые остатки G, S и Т и экспонированные (открытые) остатки Н, Y и W не принимались во внимание. Например, в Zcyto21 гидрофильные районы включают в себя остатки 155 (Glu) - 160 (Glu); остатки 51 (Lys) - 56 (Ala); остатки 50 (Phe) - 55 (Asp); остатки 140 (Pro) - (145 (Arg); и остатки 154 (Gln) - 159 (Lys), как показано в SEQ ID NO:2.
Специалистам в данной области будет понятно, что гидрофильность или гидрофобность следует учитывать при конструировании модификаций в аминокислотной последовательности полипептида Zcyto21, так чтобы не нарушить общий структурный и биологический профиль. Особый интерес для замены представляют гидрофобные остатки, выбранные из группы, состоящей из Val, Leu и Ile, или из группы, состоящей из Met, Gly, Ser, Ala, Tyr и Trp.
Идентичности незаменимых аминокислот могут быть также выведены из анализа сходства последовательности между IFN-α и другими интерферонами. С использованием таких способов, как анализ "FASTA", описанный выше, районы высокого сходства идентифицируют в семействе белков и используют для анализа аминокислотной последовательности для консервативных районов. Альтернативным подходом к идентификации вариантного полинуклеотида Zcyto21 на основе структуры является определение, может ли молекула нуклеиновой кислоты, кодирующая потенциальный вариантный полинуклеотид Zcyto21, гибридизоваться с молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID NO:1, как обсуждалось выше.
Другими способами идентификации незаменимых аминокислот в полипептидах данного изобретения являются процедуры, известные в данной области, такие как сайт-направленный мутагенез или аланинсканирующий мутагенез (Cunningham and Wells, Science 244:1081 (1989); Bass et al., Proc. Natl. Acad. Sci. USA 88:4498 (1991), Coombs and Corey, "Site-Directed Mutagenesis and Protein Engineering" in Proteins: Analysis and Design, Angeletti (ed.), pages 259-311 (Academic Press, Inc. 1998)). В последнем способе отдельные мутации аланина вводят при каждом остатке в молекуле и полученные в результате мутантные молекулы тестируют на биологическую или биохимическую активность, как описано ниже, для идентификации аминокислотных остатков, которые являются критическими для активности данной молекулы. См. также Hilton et al., J.Biol. Chem. 271:4699 (1996).
Данное изобретение относится также к функциональным фрагментам полипептидов Zcyto21 и молекулам нуклеиновых кислот, кодирующим такие функциональные фрагменты. «Функциональный» Zcyto21 или его фрагмент, определенный здесь, отличается его пролиферативной и дифференцирующей активностью, его способностью индуцировать или ингибировать специализированные клеточные функции или его способностью специфически связываться с антителом против Zcyto21 или рецептором Zcyto21 (растворимым или иммобилизованным). Как описано ранее, Zcyto21 характеризуется структурой шестиспирального пучка, содержащей: спираль А, которая определяется аминокислотными остатками 49 (Ser) - 63 (Leu); спираль В - аминокислотными остатками 76 (Asn) - 84 (Val); спираль С - аминокислотными остатками 89 (Val) - 104 (Ala); спираль D - аминокислотными остатками 111 (Glu) - 133 (Gln); спираль Е - аминокислотными остатками 137 (Thr) - 158 (Lys); и спираль F - аминокислотными остатками 163 (Gly) - 189 (Leu); как показано в SEQ ID NO:2. Таким образом, данное изобретение дополнительно обеспечивает слитые (гибридные) белки, включающие в себя: (а) полипептидные молекулы, содержащие описанные выше одну или несколько спиралей; и (b) функциональные фрагменты, содержащие одну или несколько из этих спиралей. Другая полипептидная часть слитого белка может состоять из другого цитокина с пучком спиралей или интерферона, такого как IFN-α, или неприродного и/или неродственного секреторного сигнального пептида, который облегчает секрецию этого слитого белка.
Полипептиды Zcyto21 данного изобретения, в том числе полноразмерные полипептиды, биологически активные фрагменты и слитые полипептиды, могут продуцироваться в соответствии с общепринятыми способами с использованием клеток, в которые был введен экспрессирующий вектор, кодирующий данный полипептид. В применении здесь, «клетки, в которые был введен экспрессирующий вектор» включают в себя как клетки, которыми непосредственно манипулировали посредством введения экзогенных молекул ДНК, так и их потомство, которое содержит введенную ДНК. Подходящими клетками-хозяевами являются те типы клеток, которые могут быть трансформированы или трансфицированы экзогенной ДНК и выращены в культуре, и они включают в себя бактерии, грибковые клетки и культивируемые высшие эукариотические клетки. Способы манипулирования клонированными молекулами ДНК и введения экзогенной ДНК в различные клетки-хозяева описаны в Sambrook et al., Molecular Cloning: A Laboratory Manual, (2nd ed.) (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989 и Ausubel et al. (eds.) Current Protocols in Molecular Biology, John Wiley and Sons, Inc., NY, 1987.
В общих чертах, последовательность ДНК, кодирующую полипептид Zcyto21, функционально связывают с другими генетическими элементами, требующимися для ее экспрессии, обычно включающими в себя промотор и терминатор транскрипции, в экспрессирующем векторе. Этот вектор обычно будет содержать один или несколько селектируемых маркеров и одну или несколько точек инициации репликации, хотя специалистам в данной области будет понятно, что в некоторых системах селектируемые маркеры могут быть обеспечены на отдельных векторах, и репликация экзогенной ДНК может быть обеспечена интеграцией в геном клетки-хозяина. Выбор промоторов, терминаторов, селектируемых маркеров, векторов и других элементов является предметом рутинного конструирования в пределах обычной квалификации в данной области. Многие такие элементы описаны в литературе и являются доступными через коммерческих поставщиков.
Для направления полипептида Zcyto21 в секреторный путь клетки-хозяина, в экспрессирующем векторе обеспечивают секреторную сигнальную последовательность (так же известную как лидерная последовательность, пре-пропоследовательность или пре-последовательность). Секреторная сигнальная последовательность может быть сигнальной последовательностью Zcyto21 или может быть произведена из другого секретируемого белка (например, t-PA (тканевого активатора плазминогена, см. патент США №5 641 655) или синтезирована de novo. Секреторная сигнальная последовательность функционально связана с последовательностью ДНК Zcyto21, т.е. эти две последовательности соединены в правильной рамке считывания и правильно помещены для направления вновь синтезированного полипептида в секреторный путь клетки-хозяина. Секреторные сигнальные последовательности обычно расположены 5′ (слева) от последовательности ДНК, кодирующей представляющий интерес полипептид, хотя некоторые сигнальные последовательности могут быть расположены в другом месте в представляющей интерес последовательности ДНК (см., например, Welch et al., U.S. Patent No. 5 037 743; Holland et al., U.S. Patent No. 5 143 830).
Культивируемые клетки млекопитающих могут быть использованы в качестве хозяев в данном изобретении. Способы введения экзогенной ДНК в клетки-хозяева млекопитающих включают в себя опосредованную фосфатом кальция трансфекцию (Wigler et al., Cell 14:725, 1978; Corsaro and Pearson, Somatic Cell Genetics 7:603, 1981; Graham and Van der Eb, Virology 52:456, 1973), электропорацию (Neumann et al., EMBO J. 1:841-845, 1982), опосредованную ДЭАЭ-декстраном трансфекцию (Ausubel et al., ibid.) и опосредованную липосомами трансфекцию (Hawley-Nelson et al., Focus 15:73, 1993; Ciccarone et al., Focus 15:80, 1993). Получение рекомбинантных полипептидов в культивируемых клетках млекопитающих описано, например, Levinson et al., U.S. Patent No. 4 713 339; Hagen et al., U.S. Patent No. 4 784 950; Palmiter et al., U.S. Patent No. 4 579 821; и Ringold, U.S. Patent No. 4 656 134. Подходящие культвируемые клетки млекопитающих включают в себя клеточные линии COS-1 (ATCC No. CRL 1650), COS-7 (ATCC No. CRL 1651), BHK (ATCC No. CRL 1632), BHK 570 (ATCC No. CRL 10314), 293 (ATCC No. CRL 1573; Graham et al., J.Gen. Virol. 36:59-72 1977) и линии клеток яичника китайского хомячка (например, СНО-К1; ATCC No. CCL 61 или СНО DG44, Chasm et al., Som. Cell. Molec. Genet. 12:555, 1986). Дополнительные подходящие клеточные линии известны в данной области и доступны из публичных депозитариев, таких как Американская коллекция типовых культур (American Type Culture Collection, Manassas, VA). В общем, предпочтительны сильные промоторы транскрипции, такие как промоторы из SV-40 или цитомегаловируса. См., например, патент США No. 4 956 288. Другие подходящие промоторы включают в себя промоторы из генов металлотионеина (U.S. Patent No. 4 579 821 и U.S. Patent No. 4 601 978) и основной поздний промотор аденовируса. Экспрессирующие векторы для применения в клетках млекопитающих включают в себя pZP-1 и pZP-9, которые были депонированы Американской коллекцией типовых культур (American Type Culture Collection, Manassas, VA) под номерами доступа 98669 и 98668, соответственно, и их производные.
Отбор с лекарственным средством обычно применяют для отбора на культивируемые клетки млекопитающих, в которые была встроена чужеродная ДНК. Такие клетки обычно называют "трансфектантами". Клетки, которые культивировались в присутствии селективного агента и способны передавать представляющий интерес ген их потомству, называют "стабильными трансфектантами". Предпочтительным селектируемым маркером является ген, кодирующий устойчивость к антибиотику неомицину. Отбор проводят в присутствии лекарственного средства типа неомицина, такого как G-418 или т.п. Системы отбора могут быть также использованы для увеличения уровня экспрессии представляющего интерес гена, способа, называемого "амплификацией". Амплификацию проводят культивированием трансфектантов в присутствии низкого уровня селективного агента и затем увеличением количества селективного агента для отбора клеток, которые продуцируют высокие уровни продуктов введенных генов. Предпочтительным амплифицируемым селектируемым маркером является дигидрофолатредуктаза, которая сообщает клеткам устойчивость к метотрексату. Другие гены устойчивости к лекарственным средствам (например, устойчивости к гигромицину, множественной устойчивости к лекарственным средствам, пуромицинацетилтрансферазе) также могут быть использованы.
Аденовирусная система может быть также использована для получения белка in vitro. Посредством культивирования инфицированных аденовирусом клеток не-293 в условиях, в которых клетки не осуществляют быстрое деление, эти клетки могут продуцировать белки в течение продолжительных периодов времени. Например, клетки ВНК выращивают до конфлюентности в клеточных фабриках (кассетах биореакторов для крупномасштабного производства клеток) и затем подвергают действию аденовирусного вектора, кодирующего представляющий интерес секретируемый белок. Затем эти клетки выращивают в бессывороточных условиях, которые позволяют инфицированным клеткам выживать в течение нескольких недель без значимого деления клеток. В альтернативном способе инфицированные аденовирусным вектором клетки 293 могут быть выращены в виде прикрепленных клеток или в суспензионной культуре при относительно высокой плотности клеток для получения значительных количеств белка (См. Garnier et al., Cytotechnol. 15:145-55, 1994). С любым протоколом экспрессируемый, секретируемый гетерологичный белок может быть периодически выделен из супернатанта, лизата или мембранных фракций клеточной культуры в зависимости от размещения экспрессируемого белка в клетке. В протоколе продуцирования инфицированных клеток 293 могут быть также получены несекретируемые белки.
Клетки насекомых могут быть инфицированы рекомбинантным бакуловирусом, обычно произведенным из вируса ядерного полиэдроза Autographs californica (AcNPV) в соответствии со способами, известными в данной области. В предпочтительном способе рекомбинантный бакуловирус получают с использованием системы на основе транспозона, описанной Luckow et al. (J.Virol. 67:4566-4579, 1993). Эта система, которая использует векторы-переносчики, является коммерчески доступной в форме набора (Bac-to-Bac™ Kit; Life Technologies, Rockville, MD). Этот вектор-переносчик (например, pFastBac 1™; Life Technologies) содержит транспозон Тn7 для перемещения ДНК, кодирующей представляющий интерес белок, в геном бакуловируса, поддерживаемый в Е.coli, в виде большой плазмиды, названной "бакмидой". См. Hill-Perkins and Possee, J.Gen. Virol. 71:971-976. 1990; Bonning et al., J. Gen. Virol. 75:1551-1556, 1994 и Chazenbalk and Rapoport, J. Biol. Chem. 270:1543-1549, 1995. Кроме того, векторы-переносчики могут содержать в рамке считывания с ДНК, кодирующей полипептид, удлинение или аффинную метку, как описано выше. При помощи способов, известных в данной области, вектор-переносчик, содержащий кодирующую Zcyto21 последовательность, трансформируют в клетки-хозяева Е.coli и эти клетки подвергают скринингу на бакмиды, которые содержат прерванный lacZ ген, свидетельствующий о рекомбинантном бакуловирусе. Бакмидную ДНК, содержащую рекомбинантный геном бакуловируса, выделяют при помощи известных способов и используют для трансфекции клеток Spodoptera frugiperda, например, клеток Sf9. Затем получают рекомбинантный вирус, экспрессирующий белок Zcyto21. Исходные материалы рекомбинантного вируса готовят способами, обычно используемыми в данной области.
Для получения белка этот рекомбинантный вирус используют для инфицирования клеток-хозяев насекомых, обычно клеточной линии, полученной из осенних "походных (ратных) червей", Spodoptera frugiperda (например, клеток Sf9 или Sf21) или Trichoplusia ni (например, клеток High Five™, Invitrogen, Carlsbad, CA). См., например, патент США №5 300 435. Для выращивания и поддержания этих клеток используют бессывороточные среды. Подходящие композиции сред известны в данной области и могут быть получены от коммерческих поставщиков. Клетки выращивают от плотности инокуляции приблизительно 2-5×105 клеток до плотности 1-2×106 клеток, в этот момент добавляют исходный материал рекомбинантного вируса при множественности заражения (MOI) 0,1-10, более часто около 3. Используемые процедуры обычно известны в данной области.
Другие высшие зукариотические клетки могут быть также использованы в качестве хозяев, в том числе клетки растений и клетки птиц. Применение Agrobactehum rhizogenes в качестве вектора для экспрессии генов в клетках растений рассмотрено в обзоре Sinkar et al., J. Biosci. (Bangalore) 11:47-58, 1987.
Грибковые клетки, в том числе дрожжевые клетки, могут быть также использованы в данном изобретении. Виды дрожжей, представляющие особый интерес в этом отношении, включают в себя Saccharomyces cerevisiae, Pichia pastoris и Pichia methanolica. Способы трансформации клеток S.cerevisiae экзогенной ДНК и получения из них рекомбинантных полипептидов описаны, например, Kawasaki, U.S. Patent No. 4 599 311; Kawasaki et al., U.S. Patent No. 4 931 373; Brake, U.S. Patent No. 4 870 008; Welch et.al., U.S. Patent No. 5 037 743 и Murray et al., U.S. Patent No. 4 845 075. Трансформированные клетки отбирают по фенотипу, определяемому по селектируемому маркеру, обычно по устойчивости к лекарственному средству или по способности расти в отсутствие определенного питательного вещества (например, лейцина). Предпочтительной векторной системой для применения в Saccharomyces cerevisiae является векторная система РОТ1, описанная Kawasaki et al. (U.S. Patent No. 4 931 373), которая позволяет отбирать трансформированные клетки по росту в содержащей глюкозу среде.
Подходящие промоторы и терминаторы для применения в дрожжах включают в себя промоторы и терминаторы из генов гликолитических ферментов (см., например, Kawasaki, U.S. Patent No. 4 599 311; Kingsman et at., U.S. Patent No. 4 615 974 и Bitter, U.S. Patent No. 4 977 092) и генов алкогольдегидрогеназы. См. также Патенты США №№4 990 446, 5 063 154, 5 139 936 и 4 661 454. Системы трансформации для других дрожжей, в том числе Hansenula polymorpha, Schizosaccharomyces pombe, Kluyveromyces lactis, Kluyveromyces fragilis, Ustilago maydis, Pichia pastoris, Pichia methanolica, Pichia guillermondii и Candida maltosa, известны в данной области. См., например, Gleeson et al., J.Gen. Microbiol. 132:3459-3465, 1986; Cregg, U.S. Patent No. 4 882 279 и Raymond et al., Yeast 14, 11-23, 1998. Клетки Aspergillus могут быть использованы в соответствии со способами McKnight et al., U.S. Patent No. 4 935 349. Способы трансформации Acremonium chrysogenum описаны Sumino et al., U.S. Patent No. 5 162 228. Способы трансформации Neurospora описаны Lambowitz, U.S. Patent No. 4 486 533. Получение рекомбинантных белков в Pichia methanolica описано в патентах США №№5 716 808, 5 736 383, 5 854 039 и 5 888 768.
Прокариотические клетки-хозяева, в том числе штаммы бактерий Escherichia coli, Bacillus и других родов, также являются применимыми клетками-хозяевами в данном изобретении. Способы трансформации этих хозяев и экспрессии клонированных в них чужеродных ДНК хорошо известны в данной области (см, например, Sambrook et al., ibid.). При экспрессии полипептида Zcyto21 в бактериях, таких как Е.coli, этот полипептид может сохраняться в цитоплазме, обычно в виде нерастворимых гранул, или может быть направлен в периплазматическое пространство бактериальной секреторной последовательностью. В первом случае эти клетки лизируют и гранулы извлекают и денатурируют с использованием, например, гуанидинизотиоцианата или мочевины. Затем денатурированный полипептид может быть повторно уложен и димеризован путем разведения продукта денатурации, например, диализом против раствора мочевины и комбинации восстановленного и окисленного глутатиона, с последующим диализом против забуференного солевого раствора. В последнем случае этот полипептид может быть извлечен из периплазматического пространства в растворимой и функциональной форме путем разрушения этих клеток (например, обработкой ультразвуком или осмотическим шоком) для высвобождения содержимого периплазматического пространства и извлечения этого белка, посредством чего устраняется необходимость денатурации и повторной укладки.
Трансформированные и трансфицированные клетки-хозяева культивируют в соответствии с общепринятыми процедурами в культуральной среде, содержащей питательные вещества и другие компоненты, необходимые для роста выбранных клеток-хозяев. Различные подходящие среды, в том числе среды с определенным составом среды и комплексные среды, известны в данной области и обычно включают в себя источник углерода, источник азота, незаменимые аминокислоты, витамины и минеральные соединения. Среды могут также содержать такие компоненты, как факторы роста или сыворотка, если требуется. Среда для выращивания будет обычно производить отбор на клетки, содержащие экзогенно добавленную ДНК, посредством, например, отбора с использованием лекарственного средства или недостаточности незаменимого питательного элемента, который дополняется селектируемым маркером, несомым на экспрессирующем векторе или котрансфицируемым в клетку-хозяина. Жидкие культуры обеспечивают достаточной аэрацией с использованием общепринятых средств, таких как встряхивание небольших колб или барботирование ферментеров.
Предпочтительно очищать полипептиды данного изобретения до ≥80% чистоты, более предпочтительно до ≥90% чистоты, еще более предпочтительно до ≥95% чистоты и особенно предпочтительно до фармакологически чистого состояния, т.е. до чистоты более 99,9%, в отношении загрязняющих макромолекул, в частности, других белков и нуклеиновых кислот, и инфекционных и пирогенных агентов. Предпочтительно очищенный полипептид является по существу не содержащим других полипептидов, в частности, других полипептидов животного происхождения.
Экспрессируемые рекомбинантные полипептиды Zcyto21 (в том числе химерные полипептиды и мультимерные белки) очищают общепринятыми способами очистки белка, обычно комбинацией хроматографических способов. См., в общем виде, Affinity Chromatography: Principles & Methods, Pharmacia LKB Biotechnology, Uppsala, Sweden, 1988; и Scopes, Protein Purification: Principles and Practice, Springer-Verlag, New York, 1994. Белки, содержащие полигистидиновую аффинную метку (обычно около 6 остатков гистидина), очищают аффинной хроматографией на смоле с хелатом никеля. См., например, Houchuli et al., Bio/Technol. 6: 1321-1325, 1988. Белки, содержащие метку Glu-Glu, могут быть очищены иммуноаффинной хроматографией в соответствии с общепринятыми процедурами. См., например, Grussenmeyer et al., ibid. Мальтозусвязывающие белки очищают на амилозной колонке в соответствии со способами, известными в данной области.
Полипептиды Zcyto21 могут быть также получены при помощи химического синтеза в соответствии со способами, известными в данной области, в том числе с использованием эксклюзивного (т.е. только) твердофазного синтеза, частичных твердофазных способов, конденсации фрагментов или классического синтеза в растворе. См., например, Merrifield, J. Am. Chem. Soc. 85:2149, 1963; Stewart et al., Solid Phase Peptide Synthesis (2nd edition), Pierce Chemical Co., Rockford, IL, 1984; Bayer and Rapp, Chem. Pept. Prot. 3:3, 1986 и Atherton et al., Solid Phase Peptide Synthesis: A Practical Approach, IRL Press, Oxford, 1989. Синтез in vitro является особенно предпочтительным для получения небольших полипептидов.
С использованием способов, известных в данной области, белки Zcyto21 могут быть получены в виде мономеров или мультимеров; гликозилированных или негликозилированных; пэгилированных или непэгилированных и могут содержать или могут не содержать исходный остаток аминокислоты метионина.
Клетки-мишени для применения в анализах активности Zcyto21 включают в себя, без ограничения, васкулярные клетки (в частности, эндотелиальные клетки и клетки гладких мышц), гемопоэтические (миелоидные и лимфоидные) клетки, клетки печени (в том числе гепатоциты, имеющие многочисленные отверстия эндотелиальные клетки, клетки Купфера и клетки Ито), фибробласты (в том числе кожные фибробласты и легочные фибробласты человека), фетальные легочные клетки, суставные синовиоциты, перициты, хондроциты, остеобласты и эпителиальные клетки предстательной железы. Эндотелиальные клетки и гемопоэтические клетки получают из общей предковой клетки, гемангиобласта (Choi et al., Development 125:725-732, 1998).
Белки Zcyto21 данного изобретения характеризуются их активностью, т.е. модуляцией пролиферации, дифференцировки, миграции, адгезии или метаболизма соответствующих типов клеток. Биологическую активность белков Zcyto21 анализируют с использованием анализов in vitro или in vivo, предназначенных для обнаружения пролиферации, дифференцировки, миграции или адгезии клеток; или изменений в клеточном метаболизме (например, продуцировании других факторов роста или других макромолекул). Многочисленные подходящие анализы известны в данной области и репрезентативные анализы описаны здесь. Анализы с использованием культивируемых клеток являются наиболее подходящими для скрининга, например, для определения эффектов аминокислотных замен, делеций или инсерций. Однако, ввиду комплексного характера процессов развития (например, ангиогенеза, заживления ран) обычно должны использоваться анализы in vivo для подтверждения и дополнительной характеристики биологической активности. Некоторые модели in vitro, такие как модель трехмерного матрикса из геля коллагена Pepper et al. (Biochem. Biophys. Res. Comm. 189:824-831, 1992), являются достаточно сложными для анализа гистологических эффектов. Анализы могут выполняться с использованием экзогенно полученных белков или могут проводиться in vivo или in vitro с использованием клеток, экспрессирующих представляющие интерес полипептид или полипептиды. Анализы могут проводиться с использованием белков Zcyto21 отдельно или в комбинации с другими факторами роста, такими как члены семейства VEGF или гемопоэтические цитокины (например, ЕРО, ТРО, G-CSF, фактор стволовых клеток). Репрезентативные анализы описаны ниже.
Активность белков Zcyto21 может быть измерена in vitro с использованием культивируемых клеток или in vivo введением молекул данного изобретения подходящей модели животного. Анализы, измеряющие пролиферацию или дифференцировку клеток, хорошо известны в данной области. Например, анализы, измеряющие пролиферацию, включают в себя такие анализы, как анализ хемочувствительности к красителю нейтральному красному (Cavanaugh et al., Investigational New Drugs 8:347-354, 1990), включения радиоактивно меченых нуклеотидов (как описано, например, Raines and Ross, Methods Enzymol. 109:749-773, 1985; Wahl et al., Mol. Cell Biol. 8:5016-5025, 1988 и Cook et al., Analytical Biochem. 179:1-7, 1989), включение 5-бром-2′-дезоксиуридина (BrdU) в ДНК пролиферирующих клеток (Porstmann et al., J.Immunol. Methods 82:169-179) и использование солей тетразолия (Mosmann, J.Immunol. Methods 65:55-63, 1983; Alley et al., Cancer Res. 48:589-601, 1988; Marshall et al., Growth Reg. 5:69-84, 1995 и Scudiero et al., Cancer Res. 48:4827-4833, 1988). Дифференцировка может быть тестирована с использованием подходящих клеток-предшественников, которые могут быть индуцированы к дифференцировке в более зрелый фенотип. Анализы, измеряющие дифференцировку, включают в себя, например, измерение маркеров клеточной поверхности, связанных со стадиеспецифической экспрессией ткани, ферментативной активности, функциональной активности или морфологических изменений (Watt, FASEB, 5:281-284, 1991; Francis, Differentiation 57:63-75, 1994; Raes, Adv. Anim. Cell Biol. Technol. Bioprocesses, 161-171, 1989, все включены здесь в качестве ссылки).
Активность Zcyto21 может также детектироваться с использованием анализов, разработанных для измерения индуцируемого Zcyto21 продуцирования одного или нескольких дополнительных факторов роста или других макромолекул. Такие предпочтительные анализы включают в себя анализы для определения присутствия фактора роста гепатоцитов (HGH), эпидермального фактора роста (EGF), трансформирующего фактора роста альфа (TGFα), интерлейкина-6 (IL-6), VEGF, кислого фактора роста фибробластов (aFGF), ангиогенина и других макромолекул, продуцируемых печенью. Подходящие анализы включают в себя анализы митогенеза с использованием клеток-мишеней, отвечающих на представляющую интерес макромолекулу, анализы связывания рецептора, анализы конкурентного связывания, иммунологические анализы (например, ELISA) и другие форматы, известные в данной области. Измеряют секрецию металлопротеаз из обработанных первичных кожных фибробластов человека, синовиоцитов и хондроцитов. Относительные уровни коллагеназы, гелатиназы и стромализина, продуцируемых в ответ на культивирование в присутствии белка Zcyto21, измеряют с использованием зимограмных гелей (гелей для электрофореза, в которых положение фермента детектируется по реакции, которая зависит от его ферментативной активности) (Loita and Stetler-Stevenson, Cancer Biology 1:96-106, 1990)). Синтез проколлагена/коллагена кожными фибробластами и хондроцитами в ответ на тестируемый белок измеряют с использованием включения 3H-пролина в насцентный (возникающий) секретируемый коллаген, 3H-меченый коллаген визуализируют при помощи электрофореза в ДСН-ПААГ с последующей авторадиографией (Unemori and Amento, J.Biol. Chem. 265:10681-10685, 1999). Секрецию гликозаминогликана (GAG) из кожных фибробластов и хондроцитов измеряют с использованием анализа связывания с красителем 1,9-диметилметиленовым синим (Farndale et al., Biochim. Biophys. Acta 883:173-177, 1986). Анализы коллагена и GAG проводят также в присутствии IL-1α или TGF-α для испытания способности белка Zcyto 21 модифицировать установленные реакции на эти цитокины.
Анализы активации моноцитов проводят (1) для определения способности белков Zcyto21 дополнительно стимулировать активацию моноцитов и (2) испытания способности белков Zcyto21 модулировать индуцированную прикреплением или индуцируемую эндотоксином активацию моноцитов (Fuhlbrigge et al., J.Immunol. 138:3799-3802, 1987). Уровни IL-1α и TNFα, продуцируемые в ответ на активацию, измеряют с использованием ELISA (Biosource, Inc. Camarillo, СА). Моноциты/макрофаги, благодаря CD14 (рецептору LPS), являются чрезвычайно чувствительными к эндотоксину, и белки с умеренными уровнями эндотоксин-подобной активности будут активировать эти клетки.
Гемопоэтическая активность белков Zcyto21 может тестироваться на различных гемопоэтических клетках в культуре. Предпочтительные анализы включают в себя анализы первичных колоний костного мозга и анализы линиеспецифически рестриктированных колоний более поздней стадии дифференцировки, которые известны в данной области (например, Holly et al., WIPO Publication WO 95/21920). Клетки костного мозга, посеянные на подходящую полутвердую среду (например, 50% метилцеллюлозу, содержащую 15% фетальную телячью сыворотку, 10% бычий сывороточный альбумин и 0,6% смесь антибиотиков PSN), инкубируют в присутствии тест-полипептида, затем исследуют микроскопически на образование колоний. В качестве контролей используют известные гемопоэтические факторы. Митогенная активность полипептидов Zcyto21 на линиях гемопоэтических клеток может быть измерена, как описано выше.
Миграцию клеток анализируют по существу, как описано Kähler et al., (Arteriosclerosis, Thrombosis and Vascular Biology 17:932-939, 1997). Считается, что белок является хемотаксическим, если он индуцирует миграцию клеток из зоны низкой концентрации белка в зону высокой концентрации белка. Типичный анализ выполняют с использованием модифицированных камер Бойдена с полистироловой мембраной, разделяющей две камеры (Transwell; Corning Costar Corp.). Тест-пробу, разведенную в среде, содержащей 1% БСА, добавляют в нижнюю камеру 24-луночного планшета, содержащего Transwell. Затем клетки помещают на вкладыш Transwell, который предварительно был обработан 0,2% желатином. Миграцию клеток измеряют после 4 часов инкубации при 37°С. Немигрирующие клетки стирают с верхней поверхности мембраны Transwell, a клетки, прикрепленные к нижней поверхности мембраны, фиксируют и окрашивают 0,1% красителем кристаллическим фиолетовым. Затем окрашенные клетки экстрагируют 10% уксусной кислотой и поглощение измеряют при 600 нм. Затем миграцию рассчитывают из стандартной калибровочной кривой. Миграция клеток может быть также измерена с использованием способа с матригелем Grant et al. ("Angiogenesis as a component of epithelial-mesenchymal interactions" in Goldberg and Rosen, Epithelial-Mesenchymal Interaction in Cancer, Birkhäuser Verlag, 1995, 235-248; Baatout, Anticancer Research 17:451-456, 1997).
Активность клеточной адгезии анализируют по существу, как описано LaFleur et al., (J.Biol. Chem. 272:32798-32803, 1997). Вкратце, микротитрационные планшеты покрывают тест-белком, неспецифические сайты блокируют БСА и клетки (например, клетки гладких мышц, лейкоциты или эндотелиальные клетки) высевают при плотности приблизительно 104-105 клеток на лунку. Лунки инкубируют при 37°С (обычно в течение приблизительно 60 минут), затем неприкрепленные клетки удаляют осторожным промыванием. Прикрепленные клетки определяют количественно общепринятыми способами (например, окрашиванием кристаллическим фиолетовым, лизисом клеток и определением оптической плотности лизата). Контрольные лунки покрывают известным адгезионным белком, таким как фибронектин или витронектин.
Активность белков Zcyto21 может быть измерена в биосенсорном микрофизиометре на основе кремния, который измеряет скорость внеклеточного подкисления или экскреции протона, связанную со связыванием рецептора и последующими физиологическими клеточными реакциями. Примером такого устройства является Микрофизиометр Cytosensor™, изготовляемый Molecular Devices, Sunnyvale, СА. При помощи этого способа могут быть измерены разнообразные клеточные реакции, такие как пролиферация клеток, транспорт ионов, продуцирование энергии, воспалительная реакция, регуляторная или рецепторная активация и т.п. См., например, McConnel et al., Science 257:1906-1912, 1992; Pitchford et al., Meth, Enzymol. 228:84-108, 1997; Armilli et al., J.Immunol. Meth. 212:49-59, 1998 и Van Liefde et al., Eur. J.Pharmacol. 346:87-95, 1998. Микрофизиометр может быть использован для анализа прикрепленных или неприкрепленных эукариотических или прокариотических клеток. Посредством измерения изменений внеклеточного подкисления в клеточной среде на протяжении времени микрофизиометр непосредственно измеряет клеточные реакции на разнообразные стимулы, в том числе белки Zcyto21, их агонисты и антагонисты. Предпочтительно, микрофизиометр используют для измерения реакций отвечающей на Zcyto21 эукариотической клетки в сравнении с контрольной эукариотической клеткой, которая не отвечает на полипептид Zcyto21. Отвечающие на Zcyto21 эукариотические клетки включают в себя клетки, в которые был трансфицирован рецептор для Zcyto21, посредством чего получали клетку, которая является отвечающей на Zcyto 21, а также клетки, природно отвечающие на Zcyto21. Различия, измеряемые по этому изменению, например, увеличение или уменьшение внеклеточного подкисления, в реакции клеток, подвергаемых действию полипептида Zcyto21, относительно контроля, не подвергаемого действию Zcyto21, являются прямым измерением модулируемых полипептидом Zcyto21 клеточных реакций. Кроме того, такие модулируемые Zcyto21 реакции могут анализироваться при различных стимулах. Таким образом, данное изобретение обеспечивает способы идентификации агонистов и антагонистов белков Zcyto21, предусматривающие обеспечение клеток, отвечающих на полипептид Zcyto21, культивирование первой порции этих клеток в отсутствие тест-соединения, культивирование второй порции этих клеток в присутствии тест-соединения и детектирование изменения, например, увеличения или уменьшения, в клеточной реакции второй порции клеток в сравнении с первой порцией клеток. Изменение в клеточной реакции представляют в виде измеряемого изменения скорости внеклеточного подкисления. Культивирование третьей порции этих клеток в присутствии белка Zcyto21 и в отсутствие тест-соединения обеспечивает положительный контроль на отвечающие на Zcyto21 клетки и контроль для сравнения агонистической активности тест-соединения с агонистической активностью полипептида Zcyto21. Антагонисты Zcyto21 могут быть идентифицированы подверганием клеток действию белка Zcyto21 в присутствии и в отсутствие тест-соединения, причем уменьшение стимулируемой Zcyto21 активности указывает на антагонистическую активность тест-соединения.
Экспрессия полинуклеотидов Zcyto21 в животных обеспечивает модели для дополнительного исследования биологических эффектов сверхпродуцирования или ингибирования активности белка in vivo. Кодирующие Zcyto21 полинуклеотиды и антисмысловые полинуклеотиды могут быть введены в тест-животных, таких как мыши, с использованием вирусных векторов или "голой" ДНК, или могут быть получены трансгенные животные.
Один из подходов in vivo анализа белков данного изобретения использует системы вирусной доставки. Примеры вирусов для этой цели включают в себя аденовирус, герпесвирус, ретровирусы, вирус коровьей оспы и аденоассоциированный вирус (AAV). Аденовирус, двухцепочечный ДНК-вирус, является в настоящее время наиболее исследованным вектором переноса генов для доставки гетерологичных нуклеиновых кислот. В отношении обзора следует смотреть Becker et al., Meth. Cell Biol. 43:161-89, 1994 и Douglas and Curiel, Science & Medicine 4:44-53, 1997. Аденовирусная система предоставляет несколько преимуществ. Аденовирус может (i) вместить относительно большие инсерционные сегменты (инсерты) ДНК; (ii) выращиваться до высокого титра; (iii) инфицировать широкий спектр типов клеток млекопитающих и (iv) использоваться с большим числом различных промоторов, в том числе вездесущих, тканеспецифических и регулируемых промоторов. Поскольку аденовирусы являются стабильными в кровотоке, они могут вводиться также внутривенной инъекцией.
Посредством делеции частей генома аденовируса могут быть предоставлены более крупные инсерты (до 7 т.п.н.) гетерологичной ДНК. Эти инсерты могут быть включены в вирусную ДНК прямым лигированием или гомологичной рекомбинацией с котрансфицируемой плазмидой. В приводимой в качестве примера системе, существенный для аденовируса ген Е1 делетируют из вирусного вектора, и вирус не будет реплицироваться, пока ген Е1 не обеспечивается клеткой-хозяином (например, клеточной линией 293 человека). При внутривенном введении интактным животным аденовирус прежде всего поражает печень. Если эта аденовирусная система доставки имеет делецию гена Е1, этот вирус не может реплицироваться в клетках-хозяевах. Однако, ткань хозяина (например, печень) будет экспрессировать и процессировать (и, если присутствует секреторная сигнальная последовательность, секретировать) этот гетерологичный белок. Секретированные белки будут входить в кровоток в высоковаскуляризованной печени, и могут быть определены эффекты на инфицированном животном.
Альтернативный способ доставки генов предусматривает взятие клеток из тела и включение вектора в эти клетки в виде "голой" ДНК-плазмиды. Затем эти трансформированные клетки повторно имплантируют в тело. "Голые" ДНК-векторы вводят в клетки-хозяева способами, известными в данной области, в том числе трансфекцией, электропорацией, микроинъекцией, трансдукцией, клеточным слиянием, ДЭАЭ-декстрановым способом, кальций-фосфатным осаждением, с использованием генного "дробовика" (шотган-способа) или с использованием вектора-переносчика ДНК. См., Wu et al., J.Biol. Chem. 263:14621-14624, 1988; Wu et al., J.Biol. Chem. 267:963-9677, 1992 и Johnston and Tang, Meth. Cell Biol. 43:353-365, 1994.
Могут быть также получены мыши, "сконструированные" для экспрессии гена Zcyto21, и мыши, которые обнаруживают полное отсутствие функции гена Zcyto21, называемые "мышами с нокаутом (гена)" (Snouwaert et al., Science 257:1083, 1992; Lowell et al., Nature 366:740-742, 1993). Эти мыши могут быть использованы для исследования гена Zcyto21 и белка, кодируемого им, в системе in vivo. Трансгенные мыши применимы, в частности, для исследования роли белков Zcyto21 в раннем развитии благодаря тому, что они позволяют идентифицировать отклонения развития от нормы или блокирования, происходящие из сверхэкспрессии или недостаточной экспрессии конкретного фактора. См. также Maisonpierre et al., Science 277:55-60, 1997 и Hanahan, Science 277:48-50, 1997. Предпочтительные промоторы для трансгенной экспрессии включают в себя промоторы из генов металлотионеина и альбумина.
Потеря нормального ингибиторного контроля мышечного сокращения была связана с повреждением или пертурбацией выбранных секретирующих гамма-аминомасляную кислоту нейронов. Например, синдром Stiff Man (оцепеневшего человека) включает в себя существенную ригидность мускулатуры, которая, как считают, опосредована нарушением функционирования продуцирующих гамма-аминомасляную кислоту (GABA) нейронов в таких пациентах. Другие родственные нейромышечные нарушения включают в себя миотонию, метаболические миопатии, синдром Исаака, дистонию и тетанические (столбнячные) спазмы (Valldeoriola, J.Neurol 246:423-431, 1999).
Подобным образом, прямое измерение полипептида Zcyto21 или потери его экспрессии в ткани может быть выполнено в ткани или клетках, когда они подвергаются прогрессированию опухоли. Увеличения инвазивности и подвижности клеток или увеличение или потеря экспрессии Zcyto21 в предраковом или раковом состоянии, в сравнении с нормальной тканью, могут служить в качестве диагностических показателей для перерождения опухоли, инвазии или метастазирования в прогрессировании опухоли. Само по себе знание стадии прогрессирования или метастазирования опухоли будет помогать врачу в выборе наиболее правильной терапии или агрессивности способа лечения для конкретного индивидуального ракового пациента. Способы измерения увеличения или потери экспрессии (мРНК или белка) хорошо известны в данной области и описаны здесь и могут применяться к экспрессии Zcyto21. Например, появление или исчезновение полипептидов, которые регулируют подвижность клеток, может быть использовано для помощи в диагностировании и прогнозировании рака предстательной железы (Banyard, J. and Zetter, B.R., Cancer and Metast. Rev. 17:449-458, 1999). В качестве эффектора подвижности клеток или в качестве специфического для печени маркера, увеличение или потеря экспрессии Zcyto21 может служить в качестве диагностического показателя для рака головного мозга и для других раков. Кроме того, аналоги специфического антигена предстательной железы (PSA), увеличенные уровни полипептидов Zcyto21 или антител против Zcyto21 в пациенте, относительно нормального контроля, могут быть показателями рака головного мозга и других раков (См., например, Mulders, TMT, et al., Eur. J.Surgical Oncol. 16:37-41, 1990). Сильная экспрессия Zcyto21 в ткани, которая обычно не обнаруживает экспрессии Zcyto21, могла бы служить диагностическим показателем отклонения от нормы в этом типе клеток или ткани, инвазии или метастазирования раковой печеночной ткани в непеченочную ткань и могла бы помогать врачу в направлении дальнейшего тестирования или исследования или помогать в выборе терапии.
Кроме того, полинуклеотидные зонды Zcyto21, антитела против Zcyto21 и детектирование присутствия полипептидов Zcyto21 в ткани могут быть использованы для оценки, присутствует ли ткань мозга или другая ткань, которая, как обнаружено, нормально экспрессирует Zcyto21, например, после хирургии по поводу иссечения патологической или раковой печеночной или нейронной ткани. Таким образом полинуклеотиды, полипептиды и антитела согласно данному изобретению могут быть использованы для определения, вся ли ткань иссечена после хирургии, например, хирургии головного мозга и другого рака. В таких случаях особенно важно удалить всю потенциально заболевшую ткань для максимизации излечения от рака и для минимизации рецидива. Предпочтительные варианты включают в себя флуоресцентные, радиоактивно меченые или калориметрически меченые антитела против Zcyto21 и партнеры специфического связывания Zcyto21, которые могут быть использованы гистологически in situ.
Кроме того, активность и действие Zcyto21 на прогрессирование опухоли и метастазирование может быть измерено in vivo. Были разработаны несколько сингенных мышиных моделей для исследования влияния полипептидов, соединений или других способов лечения на прогрессирование опухоли. В этих моделях опухолевые клетки, пассированные в культуре, имплантируют в мышей того же самого штамма, что и донор опухоли. Эти клетки будут развиваться в опухоли, имеющие сходные характеристики, в реципиентных мышах, и в некоторых из этих моделей будет также наблюдаться метастазирование. Подходящие модели опухолей для исследований авторов включают в себя рак легкого Lewis (АТСС No. CRL-1642) и меланому В16 (АТСС No. CRL-6323), среди прочих. Оба являются обычно используемыми опухолевыми линиями, сингенными относительно мыши C57BL6, которые легко культивируются и манипулируются in vitro. Опухоли, образующиеся в результате имплантации любой из этих клеточных линий, способны метастазироваться в легкое в мышах C57BL6. Модель рака легкого Lewis была недавно использована в мышах для идентификации ингибитора ангиогенеза (O′Reilly MS, et al., Cell 79:315-328, 1994). Мышей C57BL6/J обрабатывают экспериментальным агентом либо ежедневной инъекцией рекомбинантного белка, агониста или антагониста, либо единственной инъекцией рекомбинантного аденовируса. Спустя три дня после этой обработки 105-106 клеток имплантируют под кожу спинки. Альтернативно, сами эти клетки могут быть инфицированы рекомбинантным аденовирусом, например, экспрессирующим Zcyto21, перед имплантацией, так что этот белок синтезируется в участке опухоли или внутриклеточно, а не системно. Эти мыши обычно развивают видимые опухоли в пределах 5 дней. Опухолям позволяют расти в течение периода до 3 недель, причем во время этого периода времени они могут достигать размера 1500-1800 мм3 в обработанной контролем группе. Размер опухоли и вес тела подвергают точному наблюдению на протяжении всего эксперимента. Во время умерщвления опухоль извлекают и взвешивают вместе с легкими и печенью. Было показано, что вес легкого хорошо коррелировал с метастатической опухолевой нагрузкой. В качестве дополнительного критерия считали метастазы легочной поверхности. Иссеченные опухоль, легкие и печень готовили для гистопатологического исследования, иммуногистохимии и гибридизации in situ с использованием способов, известных в данной области и описанных здесь. Таким образом, можно было оценить влияние рассматриваемого экспрессируемого полипептида, например, Zcyto21, на способность опухоли рекрутировать сосудистую сеть и подвергаться метастазированию. Кроме того, наряду с использованием аденовируса, имплантированные клетки могут быть временно трансфицированы Zcyto21. Применение стабильных трансфектантов Zcyto21, а также использование индуцируемых промоторов для активации экспрессии Zcyto 21 in vivo известны в данной области, и они могут применяться в этой системе для оценки индукции метастазирования Zcyto21. Кроме того, очищенный Zcyto 21 или кондиционированные Zcyto 21 среды могут непосредственно инъецироваться в ту же самую модель мыши и, следовательно, использоваться в этой системе. В отношении общей ссылки см. O′Reilly MS, et al., Cell 79:315-328, 1994 и Rusciano D, et al., Murine Models of Liver Metastasis. Invasion Metastasis 14:349-361, 1995.
Антисмысловая методология может быть использована для ингибирования транскрипции гена Zcyto21 для исследования эффектов такого ингибирования in vivo. Конструируются полинуклеотиды, которые комплементарны сегменту кодирующего Zcyto21 полинуклеотида (например, полинуклеотида, представленного в SEQ ID NO:1) для связывания с кодирующей Zcyto21 мРНК и ингибирования трансляции такой мРНК. Такие антисмысловые олигонуклеотиды могут быть также использованы для ингибирования экспрессии кодирующих полипептид Zcyto21 генов в культуре клеток.
Большинство цитокинов, а также другие белки, продуцируемые активированными лимфоцитами, играют важную биологическую роль в дифференцировке клеток, активации, рекрутинге и гомеостазе клеток во всем теле. Ожидается, что Zcyto21 и ингибиторы активности Zcyto21 будут иметь разнообразные терапевтические применения. Эти терапевтические применения включают в себя лечение заболеваний, которые требуют иммунной регуляции, в том числе аутоиммунных заболеваний, таких как ревматоидный артрит, множественный склероз, тяжелая псевдопаралитическая миастения, системная красная волчанка и диабет. Zcyto21 может быть важным в регуляции воспаления и, следовательно, может быть полезным в лечении ревматоидного артрита, астмы и сепсиса. Возможна роль Zcyto21 в опосредовании онкогенеза, вследствие чего антагонист Zcyto21 был бы полезен в лечении рака. Zcyto21 может быть полезен в модуляции иммунной системы, вследствие чего Zcyto21 и антагонисты Zcyto21 могут быть использованы для уменьшения отторжения трансплантата, предупреждения реакции трансплантат против хозяина, бустинга иммунитета к инфекционным заболеваниям, лечения пациентов с ослабленным иммунитетом (например, ВИЧ-положительных пациентов) или в усовершенствовании вакцин.
В качестве интерферон-подобного полипептида в тканях головного мозга, островков, предстательной железы, яичка, гипофиза, плаценты, опухоли яичника, опухоли легкого, ректальной опухоли и опухоли яичника, а также линии CD3+ клеток и инфицированной вирусом линии эпителиальных клеток предстательной железы, Zcyto21 применим для модуляции вирусной инфекции, онкогенеза и метастазирования в этих и других тканях. В таких случаях эта интерферон-подобная молекула может высвобождаться клеткой в участке инфекции или отклоняющегося от нормы роста клеток, или, в виде секретированной молекулы, она может мигрировать к этому участку из отдаленной ткани.
Антивирусные свойства Zcyto21 особенно применимы в лечении инфекции вирусов папилломы in vitro и in vivo. Например, опухоли, вызываемые вирусами папилломы человека, индуцируют доброкачественные опухоли (т.е. генитальные остроконечные кондиломы), а также злокачественные опухоли, такие как плоскоклеточные карциномы. Лечением для таких состояний обычно является хирургия или деструкция ткани. Однако, в настоящее время показано, что некоторые антивирусные/иммуномодуляторные лекарственные средства, в том числе интерферон альфа, являются эффективными в снижении размера опухоли. См. Baker, G.E. et al., Dermatol. Clin. Apr. 15:331-340, 1997. Кроме того, как обсуждается Rockley, P.F. et al. (Pharmacol. Ther. 65 (2):265-287, 1995), иммунологическая терапия интерферонами может быть нацелена против всех участков инфекции, в том числе клинического, подклинического и латентного заболевания. В этом примере IFN-альфа, IFN-бета и IFN-гамма успешно использовали в качестве монотерапии, а также в комбинации с другими способами терапии для лечения аногенитальной остроконечной кондиломы. Zcyto21 будет полезным средством, сходным с IFN-альфа, IFN-бета и IFN-гамма в этом лечении. Кроме того, существовала тесная ассоциация между некоторыми типами вируса папилломы человека и раком шейки матки. Zcyto21 может быть использован для обнаружения, мониторинга и лечения раков шейки матки.
В качестве небольшого, секретируемого белка в островковых клетках Zcyto21 может модулировать рост и дифференцировку этих клеток. Кроме того, Zcyto21 может быть применим в лечении диабета и иммунологических состояний, связанных с ростом и дифференцировкой этих клеток.
Присутствие Zcyto21 в клетках головного мозга и гипофиза свидетельствует о том, что он может также найти применение в росте и дифференцировке этих клеток. Кроме того, молекулы данного изобретения могут быть ответственными за гомеостаз питания, в том числе поведенческие нарушения, связанные с питанием и снижением аппетита. Кроме того, молекулы Zcyto21 могут найти применение в лечении репродуктивных нарушений в целом.
Полипептиды Zcyto21 могут вводиться отдельно или в комбинации с другими васкулогенными или ангиогенными агентами, в том числе VEGF. При использовании Zcyto21 в комбинации с дополнительным агентом эти два соединения могут вводиться одновременно или последовательно в зависимости от конкретного подлежащего лечению состояния.
Zcyto21 будет применим в лечении онкогенеза и, следовательно, мог бы быть полезным в лечении рака. Ингибирование при помощи Zcyto21 анти-lgM-стимулированных нормальных В-клеток и сходный эффект, наблюдаемый в В-клеточных опухолевых линиях, предполагают, что может быть терапевтическая польза в лечении пациентов Zcyto21 для индукции клеток В-клеточных опухолей в менее пролиферативное состояние. Этот лиганд мог бы вводиться в комбинации с уже применяемыми другими агентами, в том числе, как общепринятыми химиотерапевтическими агентами, так и иммуномодуляторами, такими как интерферон альфа. Было показано, что интерфероны альфа/бета являются эффективными в лечении некоторых лейкозов и моделей заболеваний животных, и ингибирующие рост эффекты интерферона-альфа и Zcyto21 могут быть аддитивными для полученных из В-клеточной опухоли клеточных линий.
Данное изобретение обеспечивает способ снижения пролиферации неопластических В- или Т-клеток, предусматривающий введение млекопитающему с В- или Т-клеточной неоплазмой количества композиции Zcyto21, достаточного для снижения пролиферации неопластических В- или Т-клеток. Стимуляция при помощи Zcyto21 литических природных клеток-киллеров из клеток-предшественников костного мозга и пролиферации Т-клеток после активации рецепторов антигенов могла бы усиливать лечение пациентов, получающих аллогенные трансплантаты костного мозга, и, следовательно, Zcyto21 будет усиливать генерирование противоопухевых реакций с инфузией или без инфузии лимфоцитов доноров.
В другом аспекте данное изобретение обеспечивает способ снижения пролиферации неопластических В- или Т-клеток, предусматривающий введение млекопитающему с В- или Т-клеточной неоплазмой количества композиции антагониста Zcyto21, достаточного для снижения пролиферации неопластических В- или Т-клеток. Кроме того, антагонистом Zcyto21 может быть слитый белок лиганд/токсин.
Слитый токсин Zcyto21-сапорин может использоваться против сходного ряда лейкозов и лимфом, что расширяет диапазон лейкозов, которые могут лечиться Zcyto21. Опосредованная слитым токсином активация рецептора Zcyto21 обеспечивает два независимых способа ингибирования роста клеток-мишеней, из которых первый является идентичным эффектам, наблюдаемым с использованием одного лиганда, а второй обусловлен доставкой токсина посредством интернализации рецептора.
На основании описанного здесь, интерферон-подобные молекулы Zcyto21 данного изобретения будут применимы для обнаружения, мониторинга и лечения таких разнородных состояний как ретикулоэндотелиоз, почечно-клеточный рак (гипернефрома), базально-клеточный рак (базалиома), злокачественная меланома, связанная со СПИДом саркома Капоши, множественная миелома, хронический миелогенный лейкоз, не-Ходжкинская лимфома, папилломатоз гортани, грибовидный микоз, остроконечная кондилома, индуцируемая папиломавирусом бородавчатая епидермодисплазия, хронический гепатит В, гепатит С, хронический гепатит D и хронические не-А, не-В/С гепатиты.
Департамент по контролю за качеством пищевых продуктов, медикаментов и косметических средств США одобрил применение интерферона-β для лечения множественного склероза, хронического заболевания нервной системы. Интерферон-γ используют для лечения хронических гранулематозных заболеваний, в которых интерферон усиливает иммунную реакцию пациента для разрушения инфекционных бактериальных, грибковых и принадлежащих к простейшим патогенов. Клинические исследования показывают также, что интерферон-γ может быть применим в лечении СПИДа, лейшманиоза и лепроматозной проказы.
Для фармацевтического применения белки Zcyto21 готовят для локальной или парентеральной, в частности, внутривенной или подкожной, доставки в соответствии с общепринятыми способами. Обычно фармацевтические композиции будут включать в себя полипептид Zcyto21 в комбинации с фармацевтически приемлемым носителем, таким как солевой раствор, забуференный солевой раствор, 5% декстроза в воде или т.п. Композиции могут дополнительно содержать один или несколько наполнителей, консервантов, солюбилизаторов, буферящих агентов, альбумин для предупреждения потери белка на поверхностях флаконов и т.д. Способы приготовления композиций хорошо известны в данной области и описаны, например, в Remington: The Science and Practice of Pharmacy, Gennaro, ed., Mack Publishing Co., Easton, PA, 19th ed., 1995. Zcyto21 будет предпочтительно использоваться в концентрации от приблизительно 10 до 100 мкг/мл общего объема, хотя могут быть использованы концентрации в диапазоне 1 нг/мл - 1000 мкг/мл. Для локального применения, например, для ускорения заживления ран, белок будет наноситься в диапазоне от 0,1 до 10 мкг/см2 зоны раны, причем точная доза должна определяться врачом в соответствии с принятыми стандартами, с учетом природы и тяжести подлежащего лечению состояния, особенностей пациента и т.д. Определение дозы находится в пределах обычной квалификации специалиста в данной области. Введение доз является ежедневным или периодическим на протяжении периода лечения. Внутривенное введение будет выполняться болюс-инъекцией или инфузией на протяжении обычного периода одного или нескольких часов. Могут также применяться композиции пролонгированного действия. Обычно, терапевтически эффективное количество Zcyto21 является количеством, достаточным для получения клинически значимого изменения в подвергающемся лечению состоянии, например, клинически значимого изменения в гемопоэтической или иммунной функции, значимого уменьшения болезненности или значимо увеличенного гистологического балла.
Белки, агонисты и антагонисты Zcyto21 применимы для модуляции растяжения, пролиферации, активации, дифференцировки, миграции или метаболизма отвечающих на них типов клеток, которые включают в себя как первичные клетки, так и культивируемые клеточные линии. Особый интерес в этом отношении представляют гемопоэтические клетки, мезенхимные клетки (в том числе стволовые клетки и зрелые миелоидные и лимфоидные клетки), эндотелиальные клетки, эпителиальные клетки, клетки гладких мышц, фибробласты, гепатоциты, невральные клетки и эмбриональные стволовые клетки. Полипептиды Zcyto21 добавляют в среду для культуры ткани для этих типов клеток в концентрации приблизительно 10 пг/мл - приблизительно 100 нг/мл. Специалистам с квалификацией в данной области будет понятно, что белки Zcyto21 могут быть выгодно комбинированы с другими факторами роста в культуральной среде.
В области лабораторных исследований белки Zcyto21 могут быть также использованы в качестве стандартов молекулярных масс или в качестве реагентов в анализах для определения уровней этого белка в кровотоке, например, в диагностике нарушений, характеризующихся сверхпродуцированием или недостаточным продуцированием белка Zcyto21 или в анализе клеточного фенотипа.
Белки Zcyto21 могут быть использованы также для идентификации ингибиторов их активности. Тест-соединения добавляют к описанным выше тестам для идентификации соединений, ингибирующих активность белка Zcyto21. Кроме этих описанных выше тестов, пробы могут быть тестированы на ингибирование активности Zcyto21 в многочисленных анализах, предназначенных для измерения связывания рецептора или стимуляции/ингибирования Zcyto21-зависимых клеточных ответных реакций. Например, зкспрессирующие Zcyto21 клеточные линии могут быть трансфицированы конструкцией с репортерным геном, которая отвечает на стимулируемый Zcyto21 клеточный путь. Конструкции с репортерным геном этого типа известны в данной области и обычно будут содержать Zcyto21-активируемый отвечающий на сыворотку элемент (SRE), функционально связанный с геном, кодирующим анализируемый белок, такой как люцифераза. Соединения-кандидаты, растворы, смеси или экстракты тестируют на способность ингибировать активность Zcyto21 на клетках-мишенях, определяемую уменьшением стимуляции Zcyto21 экспрессии репортерного гена. Тесты этого типа будут детектировать соединения, которые непосредственно блокируют связывание Zcyto21 с рецепторами клеточной поверхности, а также соединения, которые блокируют процессы в клеточном пути после связывания рецептор-лиганд. Альтернативно, соединения или другие пробы могут быть тестированы на прямое блокирование связывания Zcyto21 с рецептором с использованием Zcyto21, меченого детектируемой меткой (например, 125I, биотином, пероксидазой хрена, FITC и т.п.). В анализах этого типа способность тест-пробы ингибировать связывание меченого Zcyto21 с рецептором свидетельствует об ингибиторной активности, которая может быть подтверждена вторичными анализами. Рецепторы, используемые в анализах связывания, могут быть клеточными рецепторами или выделенными, иммобилизованными рецепторами.
В применении здесь, термин "антитела" включает в себя поликлональные антитела, моноклональные антитела, их антигенсвязывающие фрагменты, такие как F(ab′)2 и Fab-фрагменты, одноцепочечные антитела и т.п., в том числе генетически сконструированные антитела. Антитела не из человека могут быть "очеловечены" (гуманизированы) прививкой CDR не человека на каркасные и константные области иммуноглобулина человека или включением полных вариабельных доменов не человека (необязательно "окутыванием" их подобной человеческой поверхностью путем замены экспонированных остатков, результатом чего является "облицованное" антитело). В некоторых случаях "гуманизированные" антитела могут сохранять остатки иммуноглобулина не человека в каркасных доменах вариабельной области человека для усиления характеристик правильного связывания. Посредством "гуманизирования" антител может быть увеличен биологический полупериод существования в организме, и потенциал неблагоприятных иммунных реакций при введении людям уменьшается. Специалист в данной области может генерировать гуманизированные антитела со специфическими и различными константными доменами (т.е. различных подклассов Ig) для облегчения или ингибирования различных иммунных функций, связанных с конкретными константными доменами антител. Антитела по определению являются специфически связывающими, если они связываются с полипептидом или белком Zcyto21 с аффинностью по меньшей мере в 10 раз более высокой, чем аффинность связывания с контрольным (не-Zcyto21) полипептидом или белком. Аффинность моноклонального антитела может быть легко определена специалистом с обычной квалификацией в данной области (см., например, Scatchard, Ann. NY Acad. Sci., 51:660-672, 1949).
Способы получения поликлональных и моноклональных антител хорошо известны в данной области (см. например, статью Hurrel, J.G.R., Ed., Monoclonal Hybridoma Antibodies: Techniques and Applications, CRC Press, Inc., Boca Raton, FL, 1982, включенную здесь в качестве ссылки). Особый интерес представляет генерирование антител к гидрофильным антигенным сайтам, которые включают в себя, например, остатки 155 (Glu)- 160 (Glu); остатки 51 (Lys) - 56 (Ala); остатки 50 (Phe) - 55 (Asp); остатки 140 (Pro) - 145 (Arg) и остатки 154 (Gln) - 159 (Lys), как показано в SEQ ID NO:2. Как должно быть очевидно специалисту с обычной квалификацией в данной области, поликлональные антитела могут быть получены из различных теплокровных животных, таких как лошади, коровы, козы, овцы, собаки, куры, кролики, мыши и крысы. Иммуногенность полипептида Zcyto21 может быть увеличена путем применения адъюванта, такого как квасцы (гидроксид алюминия) или полный или неполный адъювант Фрейнда. Полипептиды, применимые для иммунизации, включают в себя также слитые (гибридные) полипептиды, такие как гибриды полипептида Zcyto21 или его части с полипептидом иммуноглобулина или с мальтозусвязывающим белком. Полипептидный иммуноген может быть полноразмерной молекулой или ее частью. Если эта часть полипептида является "гаптен-подобной", такая часть может быть предпочтительно соединена или связана с макромолекулярным носителем (таким как гемоцианин фиссуреллы (KLH), бычий сывороточный альбумин (БСА) или столбнячный токсоид) для иммунизации.
Альтернативные способы генерирования или отбора антител включают в себя подвергание in vitro лимфоцитов действию полипептидов Zcyto21 и отбор библиотек представления антител в фазовых или подобных векторах (например, посредством использования иммобилизованного или меченого полипептида Zcyto21). Антитела человека могут быть получены в трансгенных, не являющихся человеком животных, которые были «сконструированы» для получения генов иммуноглобулина человека, как описано в WIPO Publication WO 98/24893. Предпочтительно, чтобы эндогенные гены иммуноглобулинов в этих животных были инактивированы или элиминированы, например, гомологичной рекомбинацией.
Различные анализы, известные лицам с квалификацией в данной области, могут быть использованы для детектирования антител, которые специфически связывают полипептиды Zcyto21. Примеры анализов описаны подробно в Antibodies: A Laboratory Manual, Harlow and Lane (Eds.), Cold Spring Harbor Laboratory Press, 1988. Характерные примеры таких анализов включают в себя: конкурентный иммуноэлектрофорез, радиоиммуноанализы, радиоиммунопреципитации, твердофазные иммуноферментные анализы (ELISA), дот-блот-анализы, Вестерн-блот-анализы, ингибиторные или конкурентные анализы и сэндвич-анализы.
Антитела к Zcyto21 могут быть использованы для аффинной очистки белка, в диагностических анализах для определения уровней этого белка в кровотоке; для обнаружения или количественного определения растворимого полипептида Zcyto21 как маркера скрытых патологии или заболевания; для иммунолокализации в целых животных или срезах тканей, в том числе иммунодиагностических применениях; для иммунохимии и в качестве антагонистов для ингибирования активности белка in vitro и in vivo. Антитела к Zcyto21 могут быть также использованы для мечения клеток, экспрессирующих Zcyto21; для аффинной очистки полипептидов и белков Zcyto21; в аналитических способах с использованием FACS; для скрининга экспрессионных библиотек и для генерирования антиидиотипических антител. Антитела могут быть связаны с другими соединениями, в том числе терапевтическими и диагностическими агентами, с использованием известных способов для обеспечения нацеливания этих соединений в клетки, экспрессирующие рецепторы для Zcyto21. Для некоторых применений, в том числе диагностических применений in vitro и in vivo, предпочтительно использовать меченые антитела. Подходящие прямые тэги (маркеры) или метки включают в себя радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные маркеры, хемилюминесцентные маркеры, магнитные частицы и т.п.; непрямые тэги (маркеры) или метки могут выводить на первое место применение пары биотин-авидин или других пар комплемент/антикомплемент в качестве промежуточных соединений. Антитела данного изобретения могут быть также прямо или опосредованно конъюгированы с лекарственными средствами, токсинами, радионуклидами и т.п., и эти конъюгаты могут быть использованы для диагностических или терапевтических применений in vivo (например, ингибирования пролиферации клеток). См. в общем Ramakrishnan et al., Cancer Res. 56:1324-1330, 1996.
Полипептиды и белки данного изобретения могут быть использованы для идентификации и выделения рецепторов. Рецепторы Zcyto21 могут участвовать в регуляции роста в печени, образовании кровеносных сосудов и других связанных с развитием процессов. Например, белки и полипептиды Zcyto21 могут быть иммобилизованы на колонке и препараты мембран пропускают через эту колонку (как описано в общем в Immobilized Affinity Ligand Techniques, Hermanson et al., eds., Academic Press, San Diego, CA, 1992, pp.195-202). Белки и полипептиды могут быть радиоактивно мечеными (Methods Enzymol., vol.182, "Guide to Protein Purifications", M.Deutscher, ed., Academic Press, San Diego, 1990, 721-737) или фотоаффинно мечеными (Brunner et al., Ann. Rev. Biochem. 62:483-514, 1993 и Fedan et al., Biocem. Pharmacol. 33:1167-1180, 1984) и использованы с метка-специфическими белками клеточной поверхности. Подобным образом, радиоактивно меченые белки и полипептиды Zcyto21 могут быть использованы для клонирования родственного рецептора в анализах связывания с использованием клеток, трансфицированных экспрессионной библиотекой кДНК.
Полипептиды Zcyto21 могут быть также использованы для обучения аналитическому мастерству, такому как масс-спектрометрия, круговой дихроизм, для определения конформации, в частности, четырех альфа-спиралей, рентгеновская кристаллография для определения трехмерной структуры на уровне атомов, спектроскопия ядерного магнитного резонанса для выявления структуры белков в растворе. Например, набор, содержащий Zcyto21, может быть предоставлен студенту для анализа. Поскольку аминокислотная последовательность является известной инструктору, этот белок может быть дан студенту в качестве теста для определения его мастерства или для развития мастерства студента, при этом инструктор будет знать, правильно ли студент проанализировал данный полипептид. Поскольку каждый полипептид является уникальным, образовательная применимость Zcyto21 могла бы быть уникальной сама по себе.
Антитела, которые связываются специфически с Zcyto21, могут быть использованы в качестве инструмента обучения для инструктирования студентов, как готовить колонки для аффинной хроматографии для очистки Zcyto21, клонирования и секвенирования полинуклеотида, который кодирует антитело, и, следовательно, в качестве практикума для обучения студента, как конструировать гуманизированные антитела. Ген Zcyto21, полипептид или антитело могли бы тогда упаковываться продающими реагенты компаниями и продаваться образовательным учреждениям, чтобы студенты получали квалификацию в области молекулярной биологии. Поскольку каждый ген и белок является уникальным, каждый ген и белок создает уникальные задачи и опыт обучения для студентов в лабораторном практикуме. Такие наборы для обучения, содержащие ген, полипептид Zcyto21 или антитело, находятся в рамках данного изобретения. Данное изобретение, описанное таким образом в общем виде, будет более понятным со ссылкой на нижеследующие примеры, которые приведены для иллюстрации, а не для ограничения данного изобретения.
ПРИМЕРЫ
Пример 1
Экспрессирующую плазмиду, содержащую весь полинуклеотид, кодирующий Zcyto21, или его часть, конструируют с использованием гомологичной рекомбинации. Фрагмент кДНК Zcyto21 выделяют при помощи ПЦР с использованием полинуклеотидной последовательности SEQ ID NO:1 с фланкирующими районами при 5′- и 3′-концах, соответствующих последовательностям вектора, фланкирующим точку встраивания Zcyto21. Праймеры для ПЦР, каждый, включают в себя от 5′-конца к 3′-концу: 40 п.н. фланкирующей последовательности из этого вектора и 17 п.н. соответствующих амино-концу и карбоксил-концу из открытой рамки считывания Zcyto21.
Десять мкл из 100 мкл реакционной смеси ПЦР подвергают электрофорезу на 0,8% плавящемся при низкой температуре агарозном (SeaPlaque GTG®; FMC BioProducts, Rockland, ME) геле с 1 × ТБЭ-буфером для анализа. Остальные 90 мкл реакционной смеси осаждают добавлением 5 мкл 1 М NaCl и 250 мкл абсолютного этанола. Плазмиду pZMP6, которая была разрезана Smal, используют для рекомбинации с этим ПЦР-фрагментом. Плазмида pZMP6 является экспрессирующим вектором млекопитающих, содержащим экспрессионную кассету, имеющую немедленно ранний промотор цитомегаловируса, множественные сайты рестрикции для встраивания кодирующих последовательностей, стоп-кодон и терминатор гормона роста человека; точку начала репликации Е.coli; экспрессионную единицу селектируемого маркера млекопитающих, содержащую промотор, энхансер и точку начала репликации SV40, ген DHFR и терминатор SV40; и последовательности URA3 и CEN-ARS, требуемые для отбора и репликации в S. cerevisiae. Она была сконструирована из pZP9 (депонированной в American Type Culture Collection, 10801 University Boulevard, Manassas, VA 20110-2209 под номером доступа №98668) с дрожжевыми генетическими элементами, взятыми из pRS316 (депонированной в American Type Culture Collection, 10801 University Boulevard, Manassas, VA 20110-2209 под номером доступа №77145), элементом внутреннего сайта вхождения рибосомы (IRES) из полиовируса и внеклеточным доменом CD8, укороченным на С-концевой стороне трансмембранного домена,
Сто микролитров компетентных клеток дрожжей (S. cerevisiae) независимо объединяют с 10 мкл различных смесей ДНК из вышеописанного и переносят в кювету для электропорации 0,2 см. Смесь дрожжи/ДНК подвергают электроимпульсам с использованием установок источника мощности (BioRad Laboratories, Hercules, CA) 0,75 кВ (5 кВ/см), ∞ Ом, 25 мкФ. В каждую кювету добавляют 600 мкл 1,2 М сорбита и эти дрожжи высевают в виде двух аликвот по 300 мкл в два URA-D-планшета и инкубируют при 30°С. Спустя приблизительно 48 часов дрожжевые трансформанты Ura+ из одного планшета ресуспендируют в 1 мл H2O и откручивают быстро для осаждения дрожжевых клеток. Клеточный осадок ресуспендируют в 1 мл лизисного буфера (2% Тритон Х-100, 1% ДСН, 100 мМ NaCl, 10 мМ Трис, рН 8,0, 1 мМ ЭДТА). Пятьсот микролитров лизисной смеси добавляют в пробирку Эппендорфа, содержащую 300 мкл промытых кислотой стеклянных гранул и 200 мкл смеси фенол-хлороформ, встряхивают на вортексе в течение 1-минутных интервалов два или три раза и откручивают в течение 5 минут в центрифуге Эппендорфа при максимальной скорости. Триста микролитров водной фазы переносят в свежую пробирку и ДНК осаждают 600 мкл этанола (EtOH) с последующим центрифугированием в течение 10 минут при 4°С. Осадок ДНК ресуспендируют в 10 мкл Н2O.
Трансформацию электрокомпетентных клеток-хозяев Е.coli (клетки Electromax DH10B™; полученные из Life Technologies, Inc., Gaithersburg, MD) выполняют с 0,5-2 мл препарата ДНК дрожжей и 40 мкл клеток. Клетки подвергают электроимпульсам при 1,7 кВ, 25 мкФ и 400 Ом. После электропорации 1 мл SOC (2% Бакто™ Триптон (Difco, Detroit, Ml), 0,5% дрожжевой экстракт (Difco), 10 мМ NaCl, 2,5 мМ KCI, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза) высевают в виде аликвот 250 мкл на четырех планшетах с LB АМР (бульон LB (Lennox), 1,8% Бакто™ Агар (Difco), 100 мг/л Ампициллина).
Индивидуальные клоны, несущие правильную экспрессионную конструкцию для Zcyto21, идентифицируют расщеплением растриктазами для подтверждения присутствия инсерта Zcyto21 и для подтверждения, что различные последовательности ДНК были соединены правильно друг с другом. Инсерты положительных клонов подвергают анализу последовательности.
Крупномасштабное выделение плазмидной ДНК проводят с использованием коммерчески доступного набора (QIAGEN Plasmid Maxi Kit, Qiagen, Valencia, CA) в соответствии с инструкциями изготовителя. Правильная конструкция названа pZMP6/Zcyto21.
Пример 2
Клетки СНО DG44 (Chasin et al., Som. Cell, Molec. Genet. 12:555-666, 1986) высевают в чашки для культуры ткани 10 см и дают им расти до приблизительно 50-70% конфлюентности в течение ночи при 37°С, 5% CO2 в среде Ham F12/FBS (Ham′s F12 medium (Life Technologies), 5% фетальной телячьей сыворотке (Hycione, Logan, UT), 1% L-глутамине (JRH Biosciences, Lenexa, KS), 1% пирувате натрия (Life Technologies)). Затем эти клетки трансфицируют плазмидой Zcyto21/pZMP6 опосредованной липосомами трансфекцией с использованием 3:1 (масса/масса) липосомной композиции поликатионного липида 2,3-диолеоилокси-N-[2-(сперминкарбоксамидо)этил]-N,N-диметил-1-пропаниминий-трифторацетата и нейтрального липида диолеоилфосфатидилэтаноламина в отфильтрованной через мембрану воде (реагент Липофектамин™, Life Technologies) в композиции бессывороточной (SF)среды (Ham F12, 10 мг/мл трансферрина, 5 мг/мл инсулина, 2 мг/мл фетуина, 1% L-глутамин и 1% пируват натрия). Zcyto21/pZMP6 разбавляют в пробирки на 15 мл до общего конечного объема 640 мкл SF-средой. 35 мкл Липофектамина™ смешивают с 605 мкл SF-среды. Полученную смесь добавляют к ДНК-смеси и дают инкубироваться приблизительно 30 минут при комнатной температуре. Пять мл SF-среды добавляют к смеси ДНК:Липофектамин™. Клетки промывают один раз 5 мл SF-среды, отсасывают и добавляют смесь ДНК:Липофектамин™. Клетки инкубируют при 37°С в течение пяти часов, затем в каждую чашку добавляют 6,4 мл среды Ham F12/10% ФТС, 1% PSN. Чашки инкубируют при 37°С в течение ночи и смесь ДНК:Липофектамин™ заменяют свежей средой 5% ФТС/Ham на следующий день, В день 3 после трансфекции клетки распределяют в колбы Т-175 в среде для выращивания. В день 7 после трансфекции клетки окрашивают моноклональными антителами FITC-анти-СD8 (PharMingen, San Diego, CA) с последующим добавлением анти-FITC-конъюгированных магнитных гранул (Miltenyi Biotec). СD8-позитивные клетки отделяют с использованием коммерчески доступных колонок (мини-MACS-колонки; (Miltenyi Biotec) в соответствии с рекомендациями изготовителя и помещают в среду DMEM/Ham F12/5% ФТС без нуклеозидов, но с 50 нМ метотрексатом (селективную среду).
Клетки высевают для субклонирования при плотности 0,5, 1 и 5 клеток на лунку в 96-луночных планшетах в селективной среде и дают расти в течение приблизительно двух недель. Лунки проверяют на испарение среды и доводят опять до 200 мкл на лунку по мере необходимости во время этого процесса. Когда большой процент колоний в планшете становятся близкими к конфлюентности, 100 мкл среды берут из каждой лунки для анализа с использованием дот-блота и клеткам подают свежую селективную среду. Супернатант наносят на нитроцеллюлозный фильтр в устройстве для дот-блот-анализа и этот фильтр обрабатывают при 100°С в вакуумном термостате для денатурации белка. Фильтр инкубируют в 625 мМ Трис-глицине, рН 9,1, 5 мМ β-меркаптоэтаноле, при 65°С, 10 минут, затем в Вестерн-буфере А с 2,5% обезжиренным сухим молоком (0,25% желатин, 50 мМ Трис-HCl рН 7,4, 150 мМ NaCl, 5 мМ ЭДТА, 0,05% Igepal CA-630) в течение ночи при 4°С на ротационном шейкере. Фильтр инкубируют с конъюгатом антитело-HRP в Вестерн-буфере А с 2,5% обезжиренным сухим молоком в течение 1 часа при комнатной температуре на ротационном шейкере. Затем этот фильтр промывают три раза при комнатной температуре в ЗФР плюс 0,01% Твин 20, 15 минут на одну промывку. Фильтр проявляют хемилюминесцентными реагентами (набор для прямого мечения ECL™; Amersham Corp., Arlington Heigts, IL) в соответствии с рекомендациями изготовителя и экспонируют на пленке (Hyperfilm ECL, Amersham Corp.) в течение приблизительно 5 минут.Положительные клоны трипсинизируют из 96-луночного планшета и переносят в 6-луночные чашки в селективной среде для увеличения масштаба культуры и анализа при помощи Вестерн-блота.
Пример 3
Полноразмерный белок Zcyto21 получают в ВНК-клетках трансфицированных pZMP6/Zcyto21 (пример 1). Клетки ВНК570 (АТСС CRL-10314) высевают в чашки для культуры ткани 10 см и дают им расти до приблизительно 50-70% конфлюентности в течение ночи при 37°С, 5% CO2 в среде DMEM/ФТС (DMEM, Gibco/BRL High Glucose; Life Technologies), 5% фетальной телячьей сыворотке (Hyclone, Logan, UT), 1 мМ L-глутамине (JRH Biosciences, Lenexa, KS), 1 мМ пирувате натрия (Life Technologies). Затем эти клетки трансфицируют плазмидой pZMP6/Zcyto21 опосредованной липосомами трансфекцией (с использованием Липофектамина™; Life Technologies), в бессывороточной (SF) среде (DMEM, дополненной 10 мг/мл трансферрина, 5 мг/мл инсулина, 2 мг/мл фетуина, 1% L-глутамином и 1% пируватом натрия). Плазмиду разбавляют в пробирки на 15 мл до общего конечного объема 640 мкл SF-средой. 35 мкл липидной смеси смешивают с 605 мкл SF-среды и полученной смеси дают инкубироваться приблизительно 30 минут при комнатной температуре. Пять мл SF-среды добавляют к смеси ДНК:липид. Клетки промывают один раз 5 мл SF-среды, отсасывают и добавляют смесь ДНК:липид. Клетки инкубируют при 37°С в течение пяти часов, затем в каждую чашку добавляют 6,4 мл среды DMEM/10% ФТС, 1% PSN. Чашки инкубируют при 37°С в течение ночи и смесь ДНК:липид заменяют свежей средой 5% ФТС/DMEM на следующий день. В день 5 после трансфекции клетки распределяют в колбы Т-162 в селективной среде (DMEM+5% ФТС, 1% L-Gln, 1% пируват натрия, 1 мкМ метотрексат). Спустя приблизительно 10 дней после трансфекции две культуральные чашки 150 мм устойчивых к метотрексату колоний из каждой трансфекции трипсинизируют и эти клетки объединяют и высевают в колбу Т-162 и переносят в крупномасштабную культуру.
Пример 4
Для конструирования аденовирусных векторов кодирующий белок район Zcyto21 человека амплифицируют при помощи ПЦР с использованием праймеров, которые добавляют сайты рестрикции Pmel и Ascl на 5′- и 3'-концах, соответственно. Амплификацию выполняют с кДНК-матрицей полноразмерного Zcyto21 в реакции ПЦР следующим образом: один цикл при 95°С в течение 5 минут; затем 15 циклов при 95°С в течение 1 мин, 61°С в течение 1 мин и 72°С в течение 1,5 мин; затем 72°С в течение 7 мин, затем пропитывание при 4°С. Продукт реакции ПЦР наносят на 1,2% плавящийся при низкой температуре агарозный гель в буфере ТАЭ (0,04 М Трис-ацетат, 0,001 М ЭДТА). ПЦР-продукт Zcyto21 вырезают из геля и очищают с использованием коммерчески доступного набора, содержащего колонку с силикагелем с мембраной (набор для очистки для ПЦР QIAquick® PCR Purificaton Kit и набор для очистки геля; Qiagen, Inc.), в соответствии с инструкциями набора. Затем ПЦР-продукт расщепляют Pmel и AscI, экстрагируют смесью фенол/хлороформ, осаждают EtOH и регидратируют в 20 мл ТЭ (Трис/ЭДТА, рН 8). Затем фрагмент Zcyto21 лигируют в сайты Pmel-AscI трансгенного вектора pTG12-8 и трансформируют в компетентные клетки Е. coli DH10B™ электропорацией. Вектор pTG12-8 получали из р2999В4 (Palmiter et al., Mol. Cell Biol. 13:5266-5275, 1993) инсертированием интрона инсулина II крысы (приблизительно 200 п.н.) и полилинкера (Fsel/Pmel/Ascl) в сайт Nrul. Этот вектор содержит промотор мышиного металлотионеина (МТ-1) (приблизительно 750 п.н.) и нетранслируемый район и сигнал полиаденилирования гормона роста человека (hGH) (приблизительно 650 п.н.), фланкированные 10 т.п.н. 5′-фланкирующей последовательности МТ-1 и 7 т.п.н. 3′-фланкирующей последовательности МТ-1. Эту кДНК встраивают между последовательностями инсулина II и hGH. Клоны, содержащие Zcyto21, идентифицируют с использованием минипрепарата плазмидной ДНК с последующим расщеплением Pmel и AscI. Положительный клон секвенируют для гарантии того, что не произошли делеции или другие аномалии в данной конструкции.
ДНК получают с использованием коммерчески доступного набора (Maxi Kit, Qiagen, Inc.) и кДНК Zcyto21 выделяют из вектора pTG12-8 с использованием ферментов Pmel и Ascl. кДНК выделяют на 1% плавящемся при низкой температуре агарозном геле и вырезают из геля. Вырезанный гель расплавляют при 70°С и ДНК экстрагируют дважды равным объемом забуференного Трисом фенола, осаждают EtOH и ресуспендируют в 10 мкл Н2О.
кДНК Zcyto21 клонируют в сайты EcoRV-AscI модифицированной плазмиды pAdTrack-CMV (He, T-C. et al., Proc. Natl. Acad. Sci. USA 95:2509-2514, 1998). Эта конструкция содержит маркерный ген зеленого флуоресцентного белка (GFP). Промотор CMV, запускающий экспрессию GFP, заменяют промотором SV40, а сигнал полиаденилирования SV40 заменяют сигналом полиаденилирования гормона роста человека. Кроме того, природный линкер заменяют сайтами Fsel, EcoRV и Ascl. Эта модифицированная форма pAdTrack-CMV названа pZyTrack. Лигирование выполняют с использованием коммерчески доступного набора для лигирования и скрининга ДНК (Fast-Link® kit; Epicentre Technologies, Madison, Wl). Клоны, содержащие Zcyto21, идентифицируют расщеплением минипрепарата ДНК Fsel и Ascl. Для линеаризации этой плазмиды приблизительно 5 мкг полученной плазмиды pZyTrack Zcyto21 расщепляют Pmel. Приблизительно 1 мкг линеаризованной плазмиды котрансформируют с 200 нг суперспиральной pAdEasy (Не et al., ibid.) в клетки Е. coli BJ5183 (Не et al., ibid.). Котрансформацию выполняют с использованием генератора импульсов генов Bio-Rad при 2,5 кВ, 200 Ом и 25 мкФ. Всю смесь для котрансформации высевают на 4 LB-чашки, содержащие 25 мкг/мл канамицина. Самые маленькие колонии выскребают и размножают в LB/канамицине и рекомбинантную аденовирусную ДНК идентифицируют стандартными процедурами получения минипрепарата ДНК. Минипрепарат ДНК рекомбинантного аденовируса трансформируют в компетентные клетки Е.coli DH10B™ и ДНК получают с использованием набора Maxi Kit (Qiagen, Inc.) в соответствии с инструкциями набора.
Приблизительно 5 мкг рекомбинантной аденовирусной ДНК расщепляют ферментом Pad (New England Biolabs) в течение 3 часов при 37°С в реакционном объеме 100 мкл, содержащем 20-30 Е Pacl. Расщепленную ДНК экстрагируют дважды равным объемом смеси фенол/хлороформ и осаждают этанолом. Осадок ДНК ресуспендируют в 10 мкл дистиллированной воды. Колбу Т25 клеток QBI-293A (Quantum Biotechnologies, Inc. Montreal, Qc. Canada), инокулированную на день раньше и выращенную до 60-70% конфлюентности, трансфицируют расщепленной Pacl ДНК. Расщепленную Pacl ДНК разбавляют до общего объема 50 мкл стерильным HBS (150 мМ NaCl, 20 мМ HEPES). В отдельной пробирке 20 мкл 1 мг/мл солей N-[1-(2,3-диолеоилокси)пропил]-N,N,N-триметиламмония (DOTAP) (Boehringer Mannheim, Indianapolis, IN) разбавляют до общего объема 100 мкл HBS. ДНК добавляют к DOTAP, смешивают осторожно пипетированием вверх и вниз и оставляют при комнатной температуре на 15 минут. Среду удаляют из клеток 293А и промывают их 5 мл бессывороточной минимальной эссенциальной среды (MEM) альфа, содержащей 1 мМ пируват натрия, 0,1 мМ не незаменимые аминокислоты MEM и 25 мМ HEPES-буфер (реагенты получали из Life Technologies, Gaithersburg, MD). 5 мл бессывороточной MEM добавляют к клеткам 293А и выдерживают их при 37°С. Смесь ДНК/липид добавляют по каплям в колбу Т25 с клетками 293А, осторожно смешивают и инкубируют при 37°С в течение 4 часов. Спустя 4 часа среду, содержащую смесь ДНК/липид, отсасывают и заменяют 5 мл полной MEM, содержащей 5% фетальную телячью сыворотку. Трансфицированные клетки подвергают мониторингу на экспрессию GFP и образование очагов (вирусных бляшек).
Спустя семь дней после трансфекции клеток 293А рекомбинантной вирусной ДНК эти клетки экспрессируют белок GFP (зеленый флуоресцентный белок) и начинают образовывать очаги (вирусные «бляшки»). Неочищенный вирусный лизат собирают при помощи клеточного скрепера (скребка) для сбора всех клеток 293А. Этот лизат переносят в коническую колбу на 50 мл. Для высвобождения большинства вирусных частиц из этих клеток выполняют три замораживания/оттаивания в бане со смесью этанола и сухого льда и в водяной бане при 37°С.
Неочищенный лизат амплифицируют (первичная (1°С) амплификация) для получения рабочего "запаса" лизата Zcyto21-rAdV. Десять чашек 10 см почти конфлюентных (80-90%) клеток 293А готовят за 20 часов перед этим, 200 мл неочищенного лизата rAdV добавляют в каждую чашку 10 см и эти чашки наблюдают в течение 48-72 часов на цитопатический эффект (СРЕ) под световым микроскопом (белый свет) и на экспрессию GFP под флуоресцентным микроскопом. Когда все клетки 293А обнаруживают СРЕ, этот исходный лизат собирают и выполняют циклы замораживания/оттаивания, как описано выше.
Затем выполняют вторичную (2°С) амплификацию Zcyto21-rAdV. Двадцать чашек для культуры ткани 15 см с клетками 293А готовят таким образом, чтобы клетки имели 80-90% конфлюентность. Весь объем, кроме 20 мл 5% среды MEM, удаляют и каждую чашку инокулируют 300-500 мл амплифицированного при 1°С лизата rAdv. Спустя 48 часов клетки 293А лизируют из продуцированного вируса, лизат собирают в полипропиленовые центрифужные флаконы на 250 мл и очищают rAdV.
Добавляют детергент NP-40 до конечной концентрации 0,5% во флаконы неочищенного лизата для лизиса всех клеток. Флаконы помещают на вращающуюся платформу на 10 минут с по возможности быстрым взбалтыванием без опрокидывания флаконов. Дебрис осаждают центрифугированием при 20000×g в течение 15 минут. Супернатант переносят в поликарбонатные центрифужные флаконы на 250 мл и добавляют 0,5 объема раствора 20% ПЭГ8000/2.5 М NaCI. Флаконы встряхивают в течение ночи на льду. Флаконы центрифугируют при 20000×g в течение 15 минут и супернатант выбрасывают в обесцвечивающий раствор. С использованием стерильного клеточного скрепера (скребка) белый осадок вируса/ПЭГ из двух флаконов ресуспендируют в 2,5 мл ЗФР. Полученный раствор вируса помещают в микроцентрифужные пробирки на 2 мл и центрифугируют при 14000×g в микроцентрифуге в течение 10 минут для удаления дополнительного количества клеточного дебриса. Супернатант из микроцентрифужных пробирок на 2 мл переносят в полипропиленовую пробирку на 15 мл с защелкивающейся крышкой и доводят до плотности 1,34 г/мл при помощи CsCl. Этот раствор переносят в поликарбонатные толстостенные центрифужные пробирки на 3,2 мл и откручивают при 348000×g в течение 3-4 часов при 25°С. Вирус образует белую полосу. Полосу вируса извлекают кончиками пипеток с широким отверстием.
Для обессоливания препарата вируса используют коммерчески доступные ионообменные колонки (например, колонки PD-10, заранее упакованные Сефадексом® G-25M (Pharmacia Biotech, Piscataway, NJ). Колонку уравновешивают 20 мл ЗФР. Наносят вирус и дают ему пройти в колонку. В колонку добавляют 5 мл ЗФР и собирают фракции из 8-10 капель. Оптические плотности 1:50-разведений каждой фракции определяют при 260 нм на спектрофотометре. Фракции пиков объединяют и определяют оптическую плотность (OD) разведения 1:25. Величины OD преобразуют в концентрацию вируса с использованием формулы: (OD при 260 нм)(25)(1,1×1012)=вирионы/мл.
Для хранения вируса к очищенному вирусу добавляют глицерин до конечной концентрации 15%, осторожно, но эффективно перемешивают и хранят в виде аликвот при -80°С.
Протокол, разработанный Quantum Biotechnologies, Inc. (Montreal, Canada), используют для измерения инфективности рекомбинантного вируса. Вкратце, два 96-луночных планшета для культуры ткани засевают 1×104 клеток 293А на лунку в MEM, содержащей 2% фетальную телячью сыворотку, для каждого тестируемого рекомбинантного вируса. Спустя 24 часа готовят 10-кратные разведения каждого вируса от 1×10-2 до 1×10-14 в MEM, содержащей 2% фетальную телячью сыворотку. 100 мкл каждого разведения помещают в каждую из 20 лунок. После 5 дней при 37°С лунки считывают как либо положительные, либо отрицательные в отношении СРЕ и рассчитывают величину "бляшкообразующих единиц/мл" (БОЕ).
Пример 5
Трансгенных животных, экспрессирующих гены Zcyto21, получают с использованием взрослых фертильных самцов (пригодных для случки) (B6C3f1, 2-8-месячного возраста (Taconic Farms, Germantown, HY)), вазэктомированных самцов (непригодных для случки) (CD1, 2-8-месячного возраста (Taconic Farms), препубертатных фертильных самок (доноров) (B6C3f1, 4-5-недельного возраста (Taconic Farms) и взрослых фертильных самок (реципиентов) (CD1, 2-4-месячного возраста, (Taconic Farms)).
Доноров акклиматизируют в течение 1 недели и затем инъецируют приблизительно 8 МЕ/мышь сывороточным гонадотропином жеребой кобылы (Sigma, St. Louis, МО) I.P. и через 46-47 часов 8 МЕ/мышь хориотическим гонадотропином человека (hCG (Sigma)) для индукции суперовуляции. Доноров спаривают с пригодными для случки животными после инъекции гормонов. Овуляция обычно происходит в пределах 13 часов после инъекции hCG. Копуляция подтверждают присутствием слизистой пробки утром после спаривания.
Оплодотворенные яйцеклетки собирают под хирургическим микроскопом (Leika MZ12 Stereo Microscope, Leica, Wetzlar, DE). Яйцеводы собирают и яйцеклетки высвобождают в предметные стекла для анализа мочи, содержащие гиалуронидазу (Sigma). Яйцеклетки промывают один раз в гиалуронидазе и два раза в среде Виттена W640 (таблица 4), которая инкубировалась с 5% CO2, 5% O2 и 90% N2 при 37°С. Затем яйцеклетки выдерживают в термостате с 37°С/5% CO2 до микроинъекции.
10-20 микрограммов плазмидной ДНК, содержащей кДНК гена Zcyto21, линеаризуют, очищают из геля и ресуспендируют в 10 мМ Трисе рН 7,4, 0,25 мМ ЭДТА рН 8,0 при конечной концентрации 5-10 нанограммов на микролитр для микроинъекции.
Плазмидную ДНК микроинъецируют в собранные яйцеклетки, содержащиеся в капле среды W640, покрытой теплым СО2-уравновешенным минеральным маслом. ДНК оттягивают в иглу для инъекций (с использованием капилляра из боросиликатного стекла 0,75 вн. диаметр, 1 мм OD) и инъецируют в отдельные яйцеклетки. Каждая яйцеклетка проникает с иглой для инъекций, в один или в оба гаплоидных пронуклеуса.
ДНК в количестве пиколитров инъецируют в эти пронуклеусы и иглу для инъекций оттягивают без приведения ее в контакт с ядрышками. Эту процедуру повторяют, пока не будут инъецированы все яйцеклетки. Успешно микроинъецированные яйцеклетки переносят в чашку для культуры тканей органов с предварительно газированной средой W640 для выдерживания в течение ночи в термостате с 37°С/5% CO2.
На следующий день 12-17 здоровых 2-клеточных эмбрионов из инъекции предыдущего дня переносят в реципиента. Определяют положение набухшей ампулы и держат яйцевод между ампулой и бурсой, производят разрыв яйцевода иглой 28 G вблизи бурсы, стараясь не разорвать ампулу или бурсу. Эмбрионы имплантируют через этот разрыв и посредством удерживания на перитонеальной стенке репродуктивные органы направляют обратно в брюшную полость.
Реципиентов возвращают в клетки парами и дают проходить 19-21 дням беременности. После родов позволяют 19-20-дневный послеродовой период перед отъемом детенышей. У отъемышей определяют пол и помещают их в раздельные клетки по полу и берут 0,5 см-биопсию (используемую для генотипирования) хвоста с использованием чистых ножниц.
Геномную ДНК получают из кусочков хвоста с использованием набора Qiagen Dneasy в соответствии с инструкциями изготовителя. Геномную ДНК анализируют при помощи ПЦР с использованием праймеров, сконструированных относительно 3′-UTR-части гормона роста человека (hGH) этого трансгенного вектора. Район, уникальный для последовательности человека, идентифицировали из сопоставления 3′-UTR последовательностей ДНК гормона роста человека и мыши, обеспечивая условие, что ПЦР-реакция не амплифицирует мышиную последовательность. Праймеры, которые амплифицируют фрагмент hGH из 368 п.н., и праймеры, которые гибридизуются с векторными последовательностями и амплифицируют этот кДНК-инсерт, часто используют вместе с праймерами hGH. В этих экспериментах ДНК из животных, положительных в отношении этого трансгена, будет генерировать две полосы, полосу 368 п.н., соответствующую 3′-UTR-фрагменту hGH, и полосу вариабельного размера, соответствующую кДНК-инсерту.
После подтверждения, что животные являются трансгенными (ТГ), они могут обратно-скрещиваться в инбредную линию помещением ТГ-самки с самцом дикого типа или ТГ-самца с одной или двумя самками дикого типа. После рождения детенышей их разделяют по полу и от их хвостов отрезают кусочки для генотипирования.
Анализ уровня экспрессии мРНК каждого трансгенного животного выполняют с использованием анализа гибридизации раствора РНК или ПЦР реального времени на приборе ABI Prism 7700 (РЕ Applied Biosystems, Inc., Foster City, CA) в соответствии с инструкциями изготовителя.
Все реагенты доступны из Sigma.
Пример 6
1. Стимуляция экспрессии от интерферон-чувствительного промотора
В одной серии экспериментов кондиционированную среду (СМ), содержащую белок Zcyto21, генерируют инфицированием клеток 293А рекомбинантным аденовирусом, содержащим кДНК для Zcyto21 (AdZy-Zcyto21), при множественности заражения 400 частиц на клетку. СМ собирают во временных точках в пределах 40 часов после инфекции и хранят при -20°С. СМ получают также из инфицирования рекомбинантным аденовирусом, не имеющим кДНК (AdZy-исходным). Перед использованием порцию СМ концентрируют в 14 раз в центрифужном фильтре Millipore Ultrafree-15 (номинальный предел («отсечение») молекулярной массы равен 5000)) и затем фильтруют через центрифужный фильтр Millipore Ultrafree-15 (номинальный предел («отсечение») молекулярной массы равен 100000)) для уменьшения количества вирусных частиц, присутствующих в среде, и наконец фильтруют через шприцевой фильтр Millipore 0,2 мкм для стерилизации СМ. Пробы концентрированной СМ разводят 1:2 в буфере для связывания и инкубируют с клетками из мышиной клеточной линии в течение 5 часов при 37°С.
2. Антивирусная активность Zcyto21
Другая серия экспериментов испытывает антивирусную активность Zcyto21. В этих исследованиях антивирусный анализ выполняют посевом клеток L929 (АТСС No. CCL-1) в среде для выращивания RPMI 1640, содержащей 10% фетальную телячью сыворотку, пенициллин, стрептомицин и L-глутамин в 96-луночном формате при 50000 клеток на лунку. Аденовирусную СМ из клеток 293А, инфицированную либо AdZy-Zcyto21, либо AdZy-исходным, как описано выше, инкубируют с клетками в течение ночи. Положительный контроль в этом анализе обеспечивают с использованием мышиного интерферона-α, серийно разведенного 1:10, начиная с 100 нг/мл. Клетки L929 только со средой для выращивания обеспечивали отрицательный контроль. Обработанные клетки инкубируют в течение 24 часов. Среды выбрасывают, добавляют свежую среду и вводят вирус энцефаломиокардита (АТСС No. vr129b) при множественности заражения 0,1 (т.е. одна вирусная частица на каждые десять клеток L929). Клетки инкубируют в присутствии этого вируса в течение 24 часов и затем лунки оценивают на цитопатический эффект в процентах (СРЕ).
3. Анализ антипролиферативной активности с использованием клеточной линии BaF3
BaF3 используют для определения, имеет ли Zcyto21 антипролиферативные свойства. Клетки почки детеныша китайского хомячка (ВНК) стабильно трансфицировали экспрессирующим вектором, содержащим промотор CMV плюс интрон А слева (против хода транскрипции) от кДНК Zcyto21 или неродственной кДНК, названной Zα30, с использованием липофектамина BRL. Стабильно трансфицированные клетки высевают в клеточную фабрику в бессывороточной среде и позволяют им расти в течение трех дней перед сбором кондиционированной среды и концентрированием ее в фильтре 5К до 10-кратного концентрирования. Пробы концентрированной кондиционированной среды хранили при 4°С.
Следующий анализ используют для тестирования на антипролиферацию BaF3. В 96-луночном планшете выполняют 1:2 серийные разведения только средой для выращивания (RPMI 1640, 10% фетальная телячья сыворотка, 1 мМ пируват натрия, 2 мМ L-глутамин) или мышиным IL-3 (начиная с 50 пг/мл в среде для выращивания) с конечным объемом 100 мкл. Пятьдесят микролитров следующего состава добавляют как к разведенной только средой для выращивания, так и к разведенной mlL-3 дорожкам: интерферон-α человека (100 нг/мл, 10 нг/мл или 1 нг/мл, разведенные в среде для выращивания), интерферон-β человека (100 нг/мл, 10 нг/мл или 1 нг/мл, разведенные в среде для выращивания), мышиный интерферон-β (100 нг/мл, 10 нг/мл или 1 нг/мл, разведенные в среде для выращивания), мышиный интерферон-β (100 нг/мл, 10 нг/мл или 1 нг/мл, разведенные в среде для выращивания), Zcyto21 (при 2,5х, 0,5х или 0,1х) и мышиный Zα30 (при 2,5х, 0,5х или 0,1х).
Клеточную линию BaF3 промывают три раза в среде для выращивания, осадки ресуспендируют в среде для выращивания, клетки считают и разбавляют в среде для выращивания до 5000 клеток/50 мкл. Затем пятьдесят микролитров разбавленных клеток добавляют к каждому разведению проб. Тест-планшеты ингубируют в термостате на 37°С в течение трех-четырех дней. Затем двадцать микролитров красителя Alomar blue добавляют в каждую лунку и планшет инкубируют в течение ночи при 37°С. Планшеты считывают на флуоресцентном планшет-ридере при волне возбуждения 544 нм и волне испускания 590 нм.
Пример 7
Тканевое распределение Zcyto21 человека в панелях тканей с использованием
ПЦР
Панель проб кДНК из тканей человека подвергали скринингу на экспрессию Zcyto21 с использованием ПЦР. Панель готовили в лаборатории и она содержала 77 проб марафон-кДНК (marathon cDNA) и кДНК из различных здоровых и раковых тканей человека и клеточных линий, как показано в таблице 6. Пробы кДНК происходили из библиотек, имеющихся в лаборатории, или препаратов марафон-кДНК из РНК, которые получали в лаборатории, или были получены от коммерческого поставщика, например, Clontech (Palo Alto, СА) или Invitrogen (Carlsbad, CA). Марафон-кДНК получали с использованием набора для амплификации марафон-кДНК (Clontech). Для гарантии качества проб панели проводили три теста на контроль качества (QC): (1) Для оценки качества РНК, используемых для библиотек, полученные в лаборатории кДНК тестировали на средний размер инсерта при помощи ПЦР с векторными олиго, которые были специфическими в отношении векторных последовательностей для индивидуальной библиотеки кДНК; (2) Стандартизацию концентрации кДНК в пробах панели получали с использованием стандартных способов ПЦР для амплификации полноразмерной кДНК альфа-тубулина или G3PDH; и (3) пробу посылали для секвенирования для проверки на возможное загрязнение рибосомной или митохондриальной ДНК. Эту панель устанавливали в 96-луночном формате, который включал в себя положительную контрольную пробу геномной ДНК человека (Clontech). Каждая лунка содержала приблизительно 0,2-100 пг/мкл кДНК. Первые ПЦР-реакции устанавливали с использованием олиго ZC39270 (SEQ ID NO:14) и ZC39272 (SEQ ID NO:15), смеси ДНК-полимеразы Advantage 2 (Clontech) и красителя Rediload (Research Genetics, Inc., Huntsville, AL). Амплификацию проводили следующим образом: 1 цикл при 94°С в течение 1 минуты, затем 35 циклов 94°С, 10 секунд; 67°С, 45 секунд и заканчивали 3-минутным конечным удлинением при 72°С. Правильный размер ДНК-фрагмента наблюдали в головном мозгу, островковых клетках, предстательной железе, яичке, гипофизе, плаценте, опухоли яичника, опухоли легкого, CD3+клетках и HPVS. Другую ПЦР-реакцию устанавливали с использованием олиго ZC39270 (SEQ ID NO:14) и ZC39271 (SEQ ID NO:16), смеси ДНК-полимеразы Advantage 2 (Clontech) и красителя Rediload (Research Genetics). Амплификацию проводили следующим образом: 1 цикл при 94°С в течение 1 минуты, затем 35 циклов 94°С, 10 секунд; 65°С, 30 секунд, 72°С, 30 секунд и заканчивали 3-минутным конечным удлинением при 72°С. Правильный размер ДНК-фрагмента наблюдали в гипофизе, ректальном раке и опухоли яичника.
Из вышесказанного должно быть понятно, что, хотя здесь были описаны конкретные варианты данного изобретения с целью иллюстрации, могут быть произведены различные модификации без отклонения от идеи и объема данного изобретения. Таким образом, данное изобретение не ограничивается ничем, за исключением прилагаемой формулы изобретения.












| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИПЕПТИД С АНТИВИРУСНОЙ АКТИВНОСТЬЮ, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2004 |
|
RU2372356C2 |
| ГОМОГЕННЫЕ ПРЕПАРАТЫ IL-28 И IL-29 | 2004 |
|
RU2518324C2 |
| НОВЫЙ ЛИГАНД РЕЦЕПТОРА ЦИТОКИНА ZCYTOR17 | 2003 |
|
RU2490276C2 |
| ГЕТЕРОДИМЕРНЫЕ ПОЛИПЕПТИДЫ IL-17 A/F И ВОЗМОЖНОСТИ ИХ ЛЕЧЕБНОГО ПРИМЕНЕНИЯ | 2007 |
|
RU2440134C2 |
| ВЫДЕЛЕННЫЙ РАСТВОРИМЫЙ IL-20 РЕЦЕПТОР (ВАРИАНТЫ) | 2000 |
|
RU2279441C2 |
| ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ЛИГАНД CD86 КОШКИ, ДИАГНОСТИЧЕСКИЙ ОЛИГОНУКЛЕОТИД, КЛОНИРУЮЩИЙ ВЕКТОР, ВАКЦИНА, СПОСОБЫ ИНДУКЦИИ ИЛИ ПОДАВЛЕНИЯ ИММУНИТЕТА У КОШКИ | 1999 |
|
RU2263145C2 |
| НОВЫЙ ЦИТОКИН ZALPHA11-ЛИГАНД | 2000 |
|
RU2258710C2 |
| НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ РЕЦЕПТОР CTLA-4 КОШКИ, ВЕКТОР, КЛЕТКИ-ХОЗЯЕВА, ВАКЦИНЫ, ОЛИГОНУКЛЕОТИДЫ, ПОЛИПЕПТИДЫ CTLA-4 КОШКИ И СПОСОБЫ ИНДУКЦИИ И ПОДАВЛЕНИЯ ИММУННОГО ОТВЕТА У КОШКИ | 1999 |
|
RU2377302C2 |
| СОБАЧИЙ ТИМУСНЫЙ СТРОМАЛЬНЫЙ ЛИМФОПОЭТИЧЕСКИЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2007 |
|
RU2457217C2 |
| НОВЫЕ ЦИТОКИНЫ ПТИЦ И КОДИРУЮЩИЕ ИХ ГЕНЕТИЧЕСКИЕ ПОСЛЕДОВАТЕЛЬНОСТИ | 2008 |
|
RU2562108C2 |
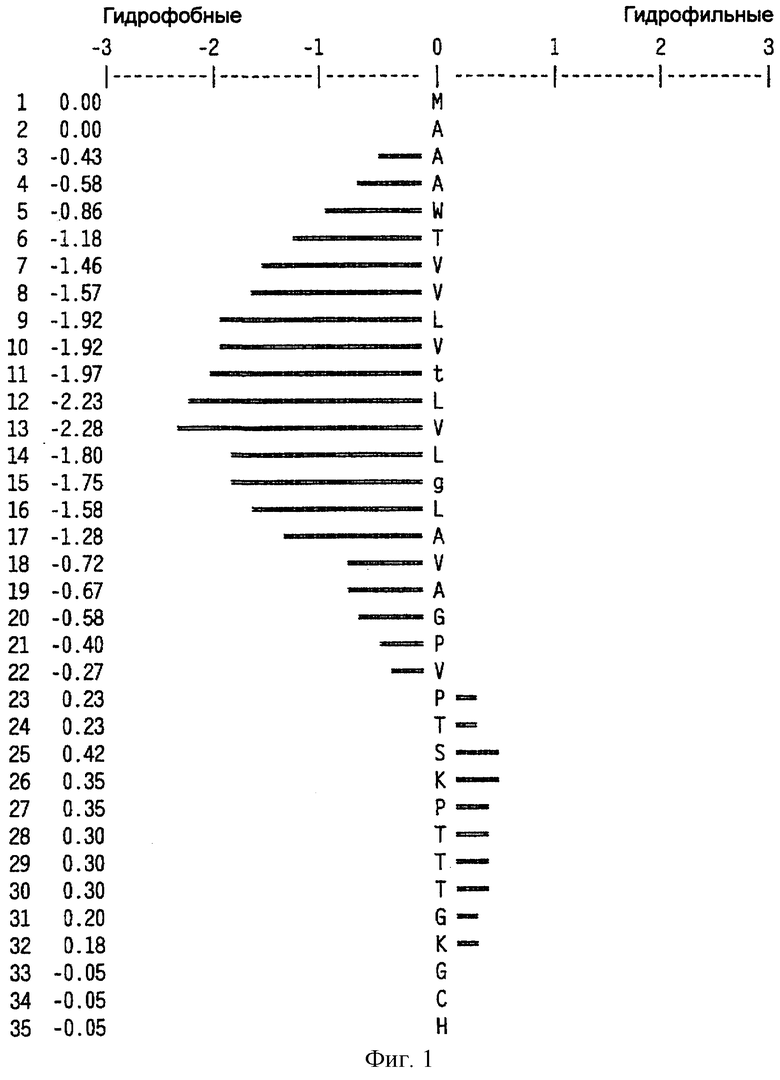
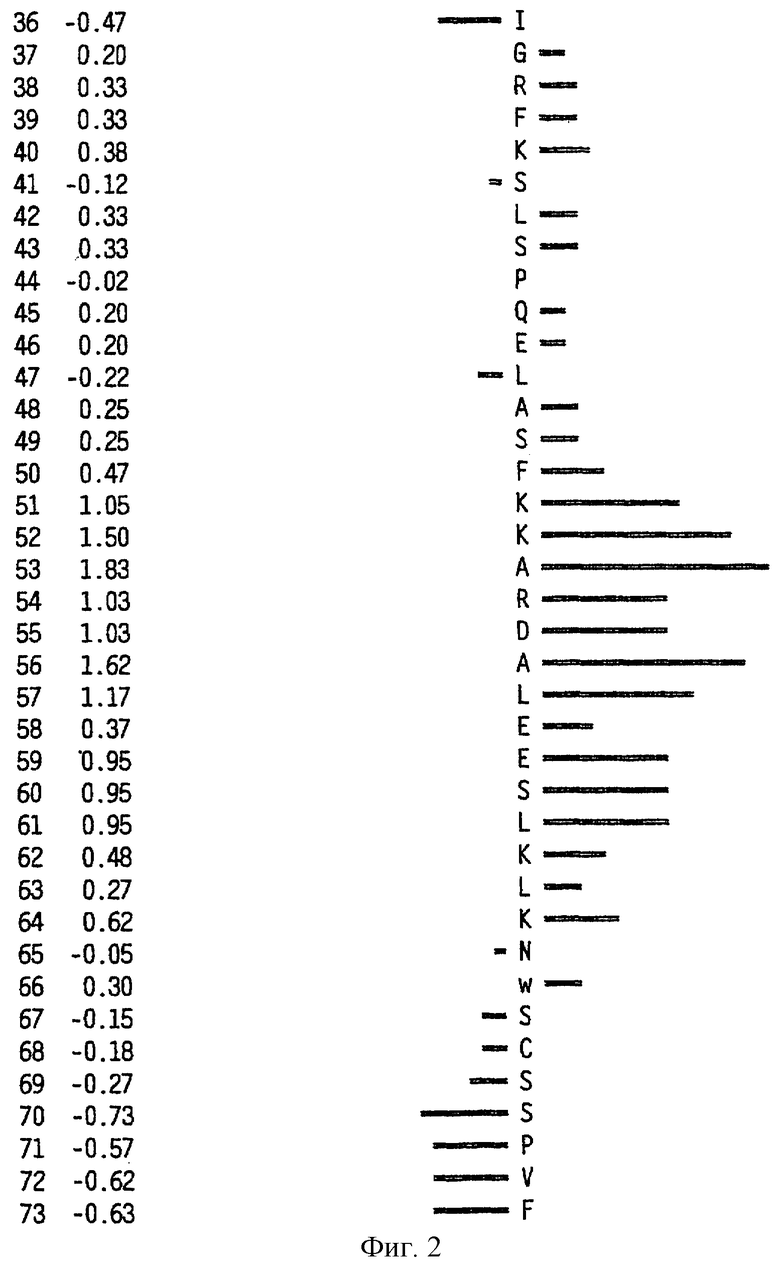
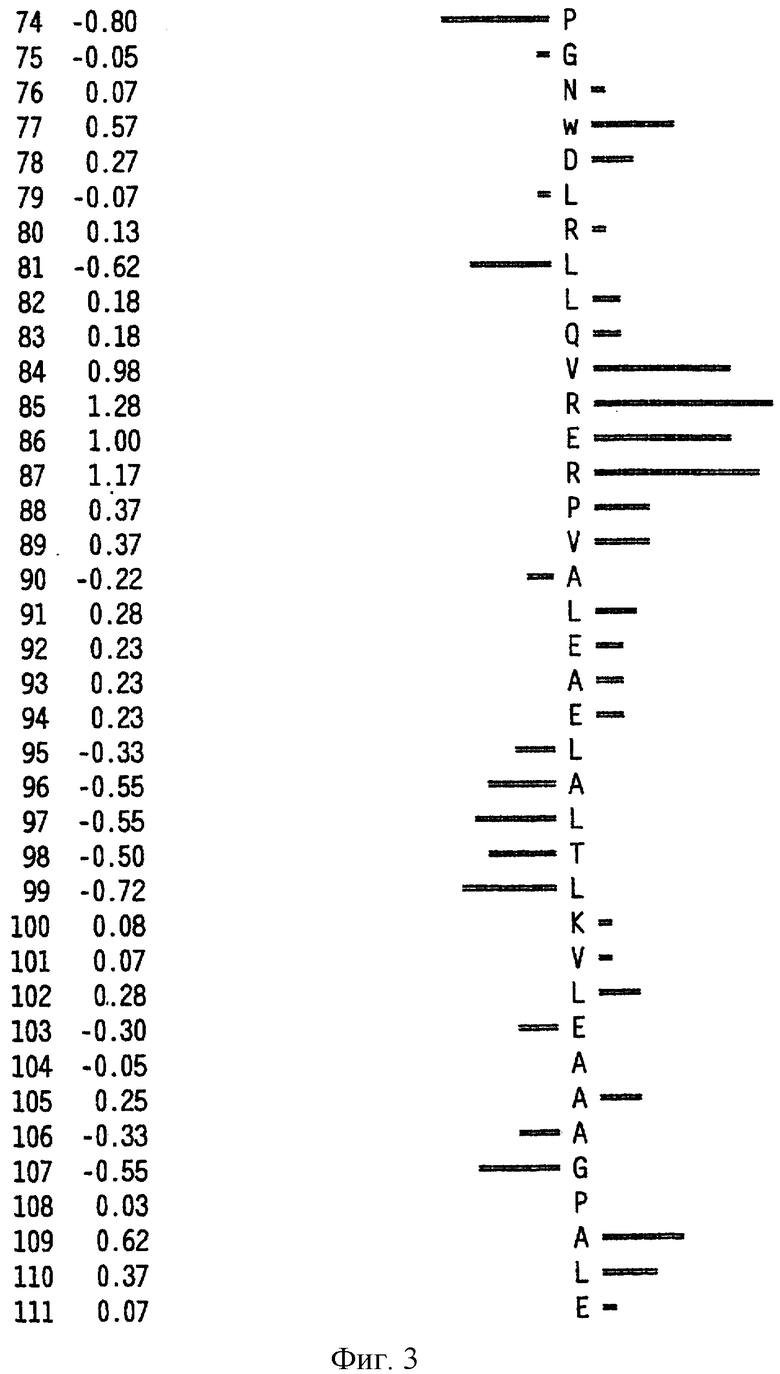
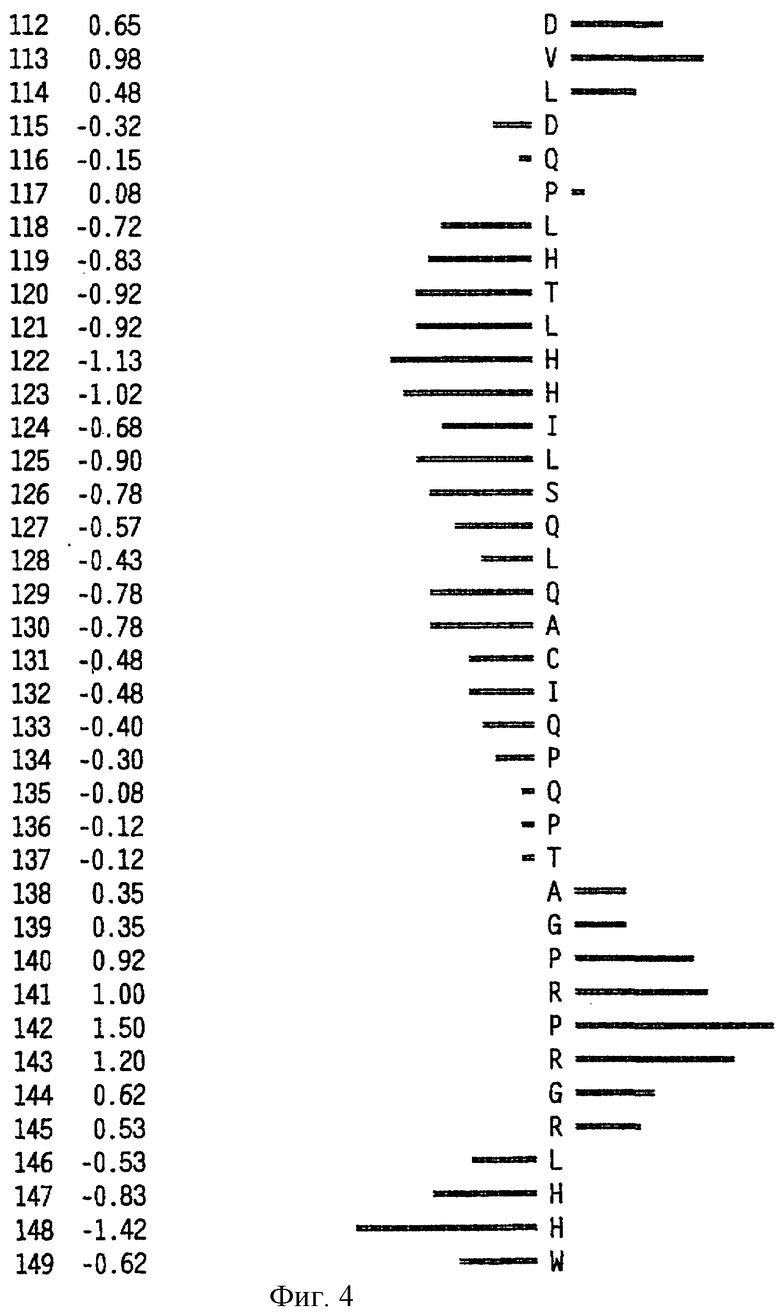
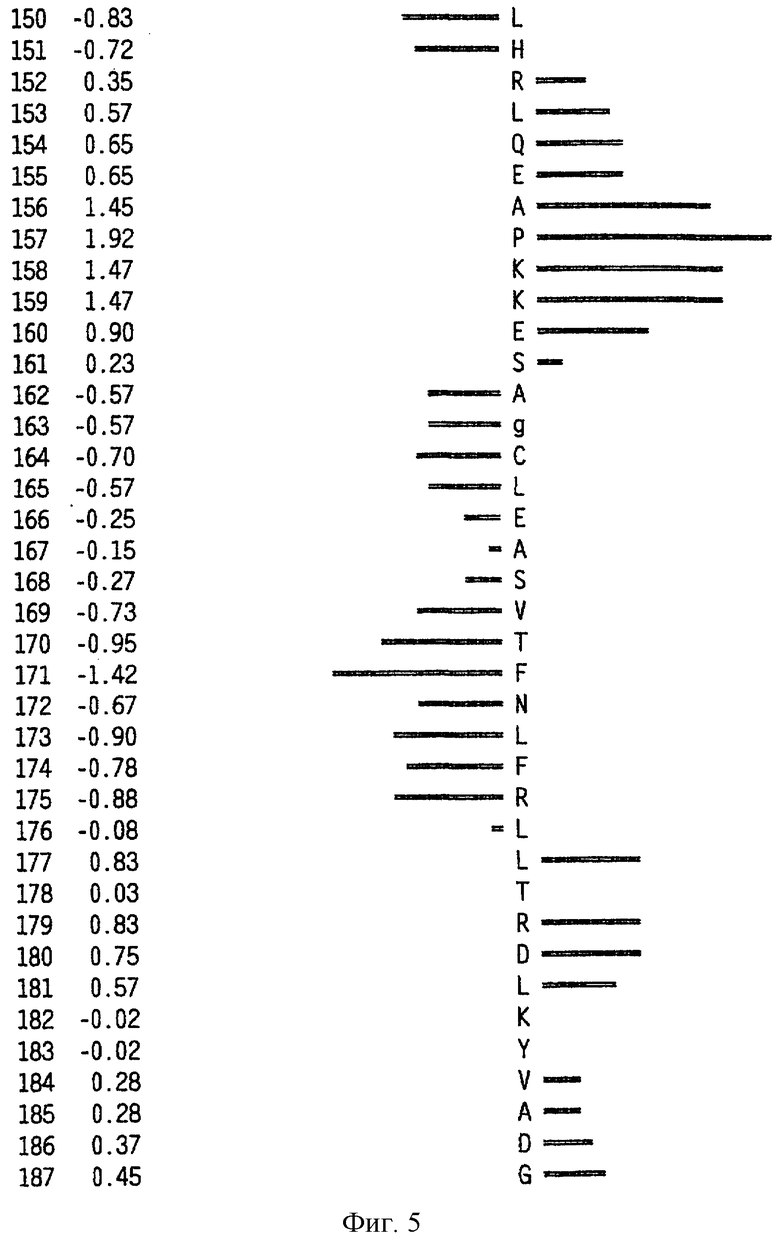
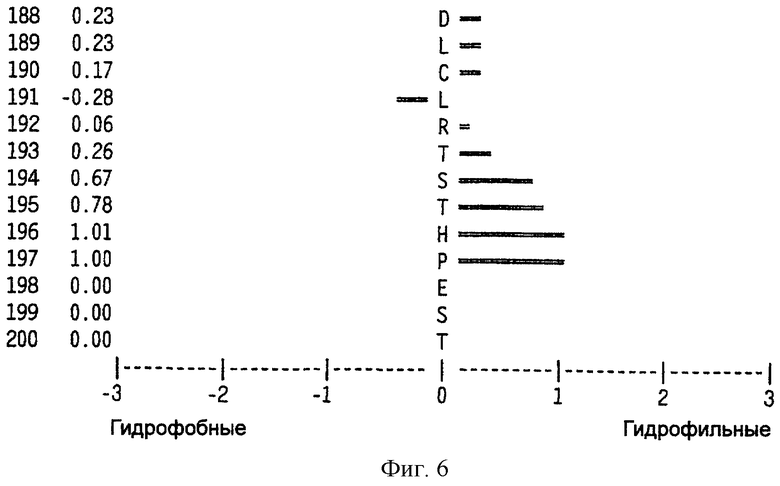
Изобретение относится к генной инженерии и может использоваться для лечения вирусных инфекций. Белок ZCYTO21 на уровне аминокислотной последовательности является близкородственным интерферону-α. Белок и антитела к этому белку обладают антивирусной активностью и могут быть эффективны при лечении гепатитов В и С, и других заболеваний. 14 н. и 57 з.п. ф-лы, 6 ил., 6 табл.
Приоритет по пунктам:
| Datebase EMBL | |||
| Приспособление для установки и поднятия забирного всасывающего клапана | 1928 |
|
SU11445A1 |
| Datebase EMBL | |||
| Способ поглощения паров бензола из воздушной, хлористо-водородной и т.п. газообразной среды помощью органических растворителей | 1928 |
|
SU18477A1 |

