Настоящая заявка заявляет приоритет согласно §119 35 U.S.C. заявки на патент США № 60/702499, поданной 25 июля 2005 г., заявки на патент США № 60/798344, поданной 16 мая 2006 г., каждая из которых в полном объеме включена ссылкой.
Область техники, к которой относится изобретение
Настоящее изобретение в основном относится к способам снижения количества В-клеток у индивидуума с использованием CD37-специфических связывающих молекул. В частности, изобретение относится к способам снижения количества В-клеток с использованием только одних CD37-специфических связывающих молекул или комбинации CD37-специфических связывающих молекул и CD20-специфических связывающих молекул, в некоторых случаях синергической комбинации. Изобретение дополнительно относится к веществам и способам лечения заболеваний, включающих аномальную активность В-клеток.
Уровень техники
В обычной роли иммунная система человека защищает организм от повреждений в результате воздействия чужеродных веществ и патогенов. Один путь, посредством которого иммунная система защищает организм, заключается в продукции специализированных клеток, называемых В-лимфоцитами, или В-клетками. В-клетки продуцируют антитела, которые связываются с чужеродным веществом или патогеном и в некоторых случаях опосредуют его разрушение.
В то же время в некоторых случаях иммунная система человека и, в частности, В-лимфоциты иммунной системы человека становятся поврежденными и приводят к развитию заболевания. Существуют многочисленные злокачественные опухоли, патогенез которых включает неконтролируемую пролиферацию В-клеток. Также имеются многочисленные аутоиммунные заболевания, в развитии которых принимает участие продукция В-клетками антител, которые вместо связывания с чужеродными веществами и патогенами связываются с системами организма. Такие антитела в некоторых случаях называют аутоантителами. Кроме того, имеются многочисленные аутоиммунные и воспалительные заболевания, которые включают патологию В-клеток, например, в виде неадекватной презентации антигена В-клетками для Т-клеток, или других путей, в которых принимают участие В-клетки. Например, у мышей, склонных к аутоиммунным заболеваниям с дефицитом В-клеток, не происходит развития аутоиммунного заболевания почек, васкулита или продукции аутоантител. Смотри Shlomchic et al., J. Exp. Med., 180: 1295-306 (1994). Интересно, что у тех же мышей, склонных к аутоиммунным заболеваниям, которые имеют В-клетки, но с недостаточностью продукции иммуноглобулинов, происходит развитие заболеваний в эксперименте, как описано Chan et al., J. Exp. Med., 189: 1639-48 (1999), этот факт указывает на то, что В-клетки играют существенную роль в развитии аутоиммунных заболеваний.
В-клетки можно идентифицировать по молекулам на их клеточной поверхности. CD20 была первой человеческой дифференцировочной, специфической для В-клеток поверхностной молекулой, идентифицированной с помощью моноклональных антител. Она представляет собой негликозилированный, гидрофобный В-клеточный трансмембранный фосфопротеин массой 35 кДа, у которого оба - амино- и карбоксиконцы - расположены внутри клетки. Смотри Einfeld et al., EMBO J., 7: 711-17 (1998). CD20 экспрессируется всеми нормальными зрелыми В-клетками, но не экспрессируется предшественниками В-клеток или плазматическими клетками. Природные лиганды для CD20 не установлены, и полностью еще не понятна функция CD20 в биологии В-клеток.
Другой дифференцировочной, специфической для В-клеток поверхностной молекулой является CD37. CD37 представляет сильно гликозилированный белок массой 40-52 кДа, который относится к четырехмерному трансмембранному семейству клеточных поверхностных антигенов. Он пересекает клеточную мембрану четыре раза, образуя две внеклеточные петли и располагая свои амино- и карбоксиконцы в цитоплазме. CD37 интенсивно экспрессируется на нормальных продуцирующих антитела (slg+) В-клетках, но не экспрессируется на предшественниках В-клеток или плазматических клетках. Экспрессия CD37 на покоящихся и активированных Т-клетках, моноцитах и гранулоцитах является низкой, и отсутствует сколько-нибудь детектируемая экспрессия CD37 на природных клетках-киллерах, тромбоцитах или эритроцитах. Смотри Belov et al., Cancer Res., 61(11): 4483-4489 (2001); Schwartz-Albiez et al., J. Immunol., 140(3): 905-914 (1988) и Link et al., J. Immunol., 137(9): 3013-3018 (1988). Помимо нормальных В-клеток, почти все злокачественные клетки, производные В-клеток, являются позитивными на экспрессию CD37, включая CLL, NHL и волосатоклеточной лейкемии [Moore et al., Journal of Pathology, 152: 13-21 (1987); Merson and Brochier, Immunology Letters, 19: 269-272 (1988) и Faure et al., American Journal of Dermatopathology, 12(3) (1990)]. CD37 принимает участие в регуляции функции В-клеток, поскольку было установлено, что у мышей с отсутствием CD37 имеются низкие уровни IgG1 в сыворотке крови и нарушения у них гуморального ответа на вирусные антигены и модельные антигены. Оказалось, что он функционирует как неклассическая костимуляторная молекула или при непосредственном воздействии на презентацию антигена посредством образования комплекса с молекулами МНС класса II. Смотри Knobeloch et al., Mol. Cell Biol., 20(15): 5363-5369 (2000). Также вероятно, что CD37 играет роль в передаче сигналов TCR. Смотри Van Spriel et al., J. Immunol., 172: 2953-2961 (2004).
Проводили исследования и разрабатывали лекарственные препараты на основе концепции, что дифференцировочные, специфические для В-клеток поверхностные молекулы, такие как CD37 или CD20, могут сами представлять мишени для антител, которые будут связываться с ними и опосредовать разрушение В-клеток, приводящих к развитию злокачественных или аутоиммунных заболеваний, которые имеют на их поверхности CD37 или CD20. Названные средствами «иммунотерапии» антитела, полученные (или на основе полученных антител) на животных, отличных от человека, которые связываются с CD37 или CD20, вводили пациентам для истощения В-клеток, приводящих к развитию злокачественных или аутоиммунных заболеваний.
Одно антитело против CD37 метили 135I и проводили его тестирование в клинических испытаниях для лечения NHL. Смотри Press et al., J. Clin. Oncol., 7(3): 1027-1038 (1989); Bernstein et al., Cancer Res. (Suppl.), 50: 1017-1021 (1990); Press et al., Front. Radiat. Ther. Oncol., 24: 204-213 (1990); Press et al., Adv. Exp. Med. Biol., 303: 91-96 (1991) и Brown et al., Nucl. Med. Biol., 24: 657-663 (1997). Антитело, МВ-1, представляет собой мышиное моноклональное IgG1-антитело, у которого отсутствуют опосредуемые Fс эффекторные функции, такие как антителозависимая клеточная цитотоксичность (ADCC), и МВ-1 не подавляет рост опухолей на модели ксенотрансплантатов в условиях in vivo, если только в него не введена изотопная метка (Buchsbaum et al., Cancer Res., 52(83): 6476-6481 (1992)). Наблюдали благоприятное распределение 135I-МВ-1 у пациентов с лимфомой при низкой опухолевой нагрузке (<1 кг), и лечение данных пациентов приводило к полной ремиссии опухолей в течение 4-11 месяцев (Press et al., 1989 и Bernstein et al., 1990).
Кроме того, исследовали иммуноконъюгат, состоящий из лекарственного препарата адриамицина, связанного с G28-1, другим антителом против CD37, на мышах и наблюдали эффекты, проявляющиеся через интернализацию и внутриклеточное высвобождение препарата. Смотри Braslawsky et al., Cancer Immunol. Immunother., 33(6): 367-374 (1991).
Различные научно-исследовательские группы применяли антитела против CD20 для лечения связанных с В-клетками заболеваний. Один способ лечения состоит из применения антител против CD20, полученных в виде радионуклиодов, для лечения В-клеточной лимфомы (например, антител против CD20, меченных 131I), а также в виде меченных 89Sr для облегчения болей в костях, вызванных метастазами злокачественной опухоли простаты и молочной железы [Endo, Gan To Kagaku Ryoho, 26: 744-748 (1999)].
Другая группа исследователей разработала химерное моноклональное антитело против CD20, состоящее из вариабельных областей тяжелой и легкой цепей мышиного происхождения, слитых с константными областями тяжелой цепи человеческого IgG1 и человеческой легкой цепи каппа. Сообщалось, что химерное антитело сохраняло способность к связыванию с CD20 и способность опосредовать ADCC и фиксировать комплемент. Смотри Liu et al., J. Immunol., 139: 3521-26 (1987). Еще одно химерное анти-CD20-антитело было получено из IDEC гибридомы C2B8, и оно было названо ритуксимабом. Полагают, что механизм противоопухолевого действия ритуксимаба заключается в комбинации нескольких активностей, включая ADCC, фиксацию комплемента и «запуск» сигналов, способствующих апоптозу злокачественных В-клеток, хотя большой размер химерного антитела препятствует оптимальной диффузии молекулы в лимфоидные ткани, которые содержат злокачественные В-клетки, что ограничивает его противоопухолевую активность. ADCC представляет собой опосредуемую клетками реакцию, в которой неспецифические цитотоксические клетки, которые экспрессируют Fc-рецепторы (FcR) (например, природные клетки-киллеры (NK), нейтрофилы и макрофаги) распознают связанное антитело на клетке-мишени и затем вызывают лизис клетки-мишени. Фиксация комплемента или комплемент-зависимая цитотоксичность (CDC) представляет способность молекулы лизировать мишень в присутствии комплемента. Путь активации комплемента инициируется связыванием вначале первого компонента системы комплемента (C1q) c молекулой (например, антителом, связанным со «своим» антигеном). Большой размер молекулы ритуксимаба препятствует оптимальной диффузии молекулы в лимфоидные ткани, которые содержат злокачественные В-клетки, что ограничивает его противоопухолевую активность.
Как правило, ритуксимаб, вводимый в виде курса инфузий раз в 4 недели, в настоящее время применяют для лечения низких стадий лимфомы или фолликулярной не-ходжкинской лимфомы [McLaughlin et al., Oncology, 12: 1763-1777 (1998); Leget et al., Curr. Opin. Oncol., 10: 548-551 (1998)] и на стадиях рецидивирования III/IV фолликулярной лимфомы [White et al., Pharm. Sci. Technol. Today, 2: 95-101 (1999)]. Другие заболевания, поддающиеся лечению ритуксимабом, включают фолликулярную центральноклеточную лимфому (FCC), покровноклеточную лимфому (MCL), диффузную крупноклеточную лимфому (DLCL) и мелкоклеточную лимфоцитарную лимфому (SLL) [Nguyen et al., Eur. J. Haematol., 62: 76-82 (1999)]. Ритуксимаб, вводимый курсом инфузий раз в неделю, также применяют для лечения СLL [Lin et al., Sem. Oncol., 30: 483-92 (2003)].
Также антитела против CD20 использовали для лечения пациентов, страдающих аутоиммунными заболеваниями, ассоциированными с продукцией В-клетками аутоантител. Например, было показано, что ритуксимаб обладает высокой лечебной эффективностью в снижении количества CD20+ В-клеток у пациентов с аутоиммунными/воспалительными заболеваниями, включая RА [Edwards, N. Engl. J. Med., 350: 2546-2548 (2004); Cambridge et al., Arthritis Rheum., 48: 2146-54 (2003)]. Пациенты с RА в течение продолжительного времени получали метотрексат (МТХ) и курс инфузий ритуксимаба из 4 введений (Edwards, смотри выше). У данных пациентов отмечали повышенные реакции по оценочной шкале American College of Rheumatology (ACR) по сравнению с контрольными группами.
В испытаниях по оценке лечения системной красной волчанки (SLE) [Leandro et al., Arthritis Rheum., 46: 2673-2677 (2002)] пациентам вводили две инфузии с высокой дозой ритуксимаба, и было показано снижение числа В-клеток и улучшение состояния пациентов. Во втором исследовании для уменьшения числа В-клеток при SLE [Looney et al., Arthritis Rheum., 50: 2580-2589 (2004)] пациентам вводили одну инфузию в дозе 100 мг/м2 (низкая доза), одну инфузию в дозе 375 мг/м2 (промежуточная доза) или в виде 4 инфузий (с интервалом 1 неделя) в дозе ритуксимаба 375 мг/м2 (высокая доза). У данных пациентов наблюдали снижение количества В-клеток и улучшенную оценку состояния в баллах, но лечение не приводило к изменению уровня аутоантител. Были проведены испытания ритуксимаба при макроглобулинемии Вальденстрема [Treon et al., Immunotherap., 24: 272-279 (2000)], и у пациентов наблюдали повышение уровня гематокрита (НСТ) и количества тромбоцитов (PLT) после 4 инфузий ритуксимаба.
Недавние сообщения по оценке лечения ритуксимаба у пациентов, страдающих рассеянным склерозом, аутоиммунным заболеванием, поражающим центральную нервную систему, показали, что курс лечения приводит к истощению периферических В-клеток, но оказывает слабое действие на уровень В-клеток в спинномозговой жидкости. Смотри Monson et al., Arch. Neurol., 62: 258-264 (2005).
Дополнительные публикации, касающиеся применения ритуксимаба, включают: Stashi et al. «Rituximab chimeric anti-CD20 monoclonal antibody treatment for adults with chronic idiopathic thrombocytopenic purpura» Blood 98: 952-957 (2001); Matthews, R. «Medical Heretics» New Scientist (7 Apr., 2001); Leandro et al. «Clinical outcome in 22 patients with rheumatoid arthritis treated with В lymphocyte depletion» Ann. Rheum. Dis. 61: 833-888 (2002); Leandro et al. «Lymphocyte depletion in rheumatoid arthritis: early evidence for safety, efficacy and dose response. Arthritis and Rheumatism 44(9): S370 (2001); Leandro et al. «An open study of B lymphocyte depletion in systemic lupus erythematosus», Arthritis Rheum. 46: 2673-2677 (2002); Edwards et al. «Sustained improvement in rheumatoid arthritis following a protocol designed to deplete B lymphocytes» Rheumatology 40: 205-211 (2001); Edwards et al. «B-lymphocyte depletion in rheumatoid arthritis and other autoimmune disorders» Biochem. Soc. Trans. 30(4): 824-828 (2002); Edwards et al. «Efficacy and safety of rituximab, B-cell targeted chimeric monoclonal antibody: a randomised, placebo controlled trial in patients with rheumatoid arthritis» Arthritis Rheum. 46: S197 (2002); Levine et al. «IgM antibody-related polyneuropathies: B-cell depletion chemotherapy using rituximab» Neurology 52: 1701-1704 (1999); DeVita et al. «Efficacy of selective B-cell blockade in treatment of rheumatoid arthritis» Arthritis Rheum. 46: 2029-2033 (2002); Hidashida et al. «Treatment of DMARD-refractory rheumatoid arthritis with rituximab». Presented at Annual Scientific Meeting of the American College of Rheumatology; October 24-29; New Orleans, La. 2002; Tusca № et al. «Successful treatment of infliximab-refractory rheumatoid arthritis with rituximab». Presented at Annual Scientific Meeting of the American College of Rheumatology; October 24-29; New Orleans, La. 2002.
Проблемы, связанные с лечением ритуксимабом, остаются. Например, у большинства пациентов со злокачественными опухолями после лечения ритуксимабом возникают рецидивы, как правило, в течение 6-12 месяцев, и сообщалось о реакциях на инфузии препарата со смертельным исходом. Данные фатальные реакции развивались после проявления комплекса реакций на инфузию, включающего гипоксию, инфильтраты в легких, острый респираторный дистресс-синдром, инфаркт миокарда, фибрилляцию желудочков или кардиогенный шок. Также сообщалось об острой почечной недостаточности, для купирования которой требовался диализ, со случаями смертельного исхода при использовании синдрома лизиса опухоли после лечения ритуксимабом, с мукокожными реакциями, в некоторых случаях с фатальным исходом. Кроме того, для внутривенной инъекции требуются высокие дозы ритуксимаба, поскольку молекула является крупной, примерно 150 кДа, и, как уже отмечалось выше, проникновение в лимфоидные ткани, где находится большое количество опухолевых клеток, ограничено.
В результате того, что нормальные зрелые В-клетки также экспрессируют CD37 и CD20, количество нормальных В-клеток снижается под действием лечения антителами против CD37 (Press et al., 1989) или антителами против CD20 [Reff et al., Blood, 83: 435-445 (1994)]. Однако после окончания лечения нормальные В-клетки могут регенерироваться из предшественников В-клеток, негативных на CD37 и CD20; следовательно, у пациентов после лечения антителами против CD37 и CD20 отсутствует значительная иммуносупрессия.
Технология моноклональных антител и методы генной инженерии позволили получать молекулы иммуноглобулинов для диагностики и лечения заболеваний у человека. Инженерию белков применяли для повышения аффинности антитела для «своего» антигена со сведением до минимума проблем, связанных с иммуногенностью, и для изменения эффекторных функций антител. Структура домена иммуноглобулинов подвергается методам генной инженерии в том плане, что антигенсвязывающие домены и домены, ответственные за эффекторные функции, можно менять между иммуноглобулинами различных классов и подклассов. Обзор по структуре и функциям иммуноглобулинов приведен, например, у Harlow et al., Eds., Antibodies: a laboratory manual, chapter 14, Cold Spring Harbor Laboratory, Cold Spring Harbor (1988). Глубокое введение в эту проблему, а также подробную информацию почти обо всех аспектах рекомбинантной технологии антител можно найти в книге «Recombinant Antibodies» (John Wiley & Sons, NY, 1999). Исчерпывающее обобщение подробных лабораторных протоколов генной инженерии антител можно найти у R. Kontermann and S. Dubel (eds.) «The antibody engineering lab manual» (Springer Verlag, Heidelberg/New York, 2000).
Недавно были сконструированы более мелкие молекулы иммуноглобулинов для преодоления проблем, связанных с терапией целыми молекулами иммуноглобулинов. Одноцепочечные фрагменты Fv (scFv) содержат вариабельную область тяжелой цепи антитела, связанную коротким пептидом-линкером с вариабельной областью легкой цепи антитела [Huston et al., Proc. Natl. Acad. Sci. USA, 85: 5879-5883 (1988)]. В дополнение к вариабельным областям каждая из цепей антитела имеет одну или более константных областей. Легкие цепи имеют одну константную область. Таким образом, легкие цепи имеют одну вариабельную область и одну константную область. Тяжелые цепи имеют несколько константных областей. Тяжелые цепи в IgG-, IgA- и IgD-антителах имеют три константные области, обозначаемые CH1, CH2 и CH3, и тяжелые цепи в IgМ- и IgЕ-антителах имеют четыре константные области, обозначаемые CH1, CH2, CH3 и СН4. Таким образом, тяжелые цепи имеют одну вариабельную область и три или четыре константные области.
Также тяжелые цепи иммуноглобулинов можно разделить на три функциональные области: Fd-область (фрагмент, включающий VH и СН1, т.е. два N-концевых домена тяжелой цепи), шарнирную область и Fс-область («кристаллизуемый фрагмент», полученный из константных областей и образовавшийся после расщепления пепсином). Fd-область в комбинации с легкой цепью образует Fab («антигенсвязывающий фрагмент»). Поскольку антиген будет стереохимически взаимодействовать с антигенсвязывающей областью на аминоконце каждого Fab, молекула IgG является двухвалентной, т.е. она может связываться с двумя молекулами антигена. Fс содержит домены, которые взаимодействуют с рецепторами иммуноглобулина на клетках и с первоначальными элементами каскада комплемента. Таким образом, как правило, считается, что Fс-фрагмент ответственен за эффекторные функции иммуноглобулина, такие как связывание комплемента и связывание с Fс-рецепторами.
За счет небольшого размера молекул scFv они имеют очень высокий клиренс из плазмы крови и тканей и обладают более эффективным проникновением в ткани по сравнению с целыми молекулами иммуноглобулинов. У scFv с противоопухолевым действием имело место более быстрое проникновение в опухоль и более равномерное распределение в массе опухоли по сравнению с соответствующим химерным антителом [Yokota et al., Cancer Res., 52, 3402-3408 (1992)]. Слияние scFv с другой молекулой, такой как токсин, обладает преимуществом способности к специфическому связыванию с антигеном, а небольшой размер scFv способствует доставке токсина в ткань-мишень [Chaudary et al., Nature, 339: 394 (1989); Batra et al., Mol. Cell. Biol., 11: 2200 (1991)].
Несмотря на преимущества молекул scFv существует несколько недостатков при их применении. Несмотря на то что высокий клиренс scFv может привести к снижению токсических эффектов в нормальных клетках, такой высокий клиренс может препятствовать доставке минимально эффективной дозы в ткань-мишень. Ставится под сомнение производство адекватных количеств scFv для введения пациентам за счет трудностей, возникающих при экспрессии и выделении scFv, которые оказывают отрицательное влияние на выход продукта. Во время экспрессии молекулы scFv теряют стабильность и часто подвергаются агрегации за счет связывания вариабельных областей из разных молекул. Кроме того, уровни получения молекул scFv в экспрессирующих системах млекопитающих являются низкими, что ограничивает эффективное получение молекул scFv для применения в терапии [Davis et al., J. Biochem. Chem., 256: 10410-10418 (1990); Traunecker et al., EMBO J., 10: 3655-3659 (1991)]. Разрабатывали стратегии для повышения продукции, включая введение сайтов гликозилирования в вариабельные области [Jost, C.R., патент США № 5888773; Jost et al., J. Biol. Chem., 69: 26267-26273 (1994)].
Другим недостатком применения scFv для терапии является отсутствие у них эффекторной функции. scFv без проявления цитолитических функций, ADCC и комплемент-зависимой цитотоксичности (CDC), связанных с константной областью иммуноглобулина, могут быть не эффективными для лечения заболеваний. Несмотря на то что разработка технологии применения scFv началась более чем 12 лет тому назад, в настоящее время отсутствуют продукты на основе scFv, разрешенные для лечения.
Альтернативно было высказано предположение, что слияние scFv с другой молекулой, такой как токсин, может иметь преимущество специфической антигенсвязывающей активности, а небольшого размера scFv способствует доставке токсина к ткани-мишени. Chaudary et al., Nature, 339: 394 (1989); Batra et al., Mol. Cell. Biol., 11: 2200 (1991). Таким образом, конъюгация или слияние токсинов с scFv были предложены в качестве альтернативной стратегии для обеспечения эффективных, антиген-специфических молекул, но введение таких конъюгатов или химерных молекул может ограничиваться ввиду явления высокой и/или неспецифической токсичности за счет наличия группы токсина в таких препаратах. Токсические эффекты могут включать повышение активности ферментов печени выше физиологического уровня и сосудистый синдром, а также проявление других нежелательных эффектов. Кроме того, иммунотоксины сами по себе являются иммуногенными при введении хозяину, и антитела хозяина, продуцированные против иммунотоксинов, также будут ограничивать потенциальную применимость при повторных терапевтических курсах лечения.
Другие генно-инженерные рекомбинантные белки, называемые малыми модуляторными иммунофармацевтическими (SMIPTM) продуктами, описаны в совместных патентных публикациях США 2003/133939, 2003/0118592 и 2005/0136049 и совместных международных патентных публикациях WO02/056910, WO2005/037989 и WO2005/017148, которые включены здесь ссылкой. Продукты SMIPTM представляют собой новые, на основе связывающего домена иммуноглобулинов рекомбинантные белки, которые содержат связывающий домен для «своей» структуры, такой как антиген, противорецептор или тому подобное; шарнирную область полипептида IgG1, IgA или IgЕ или мутантную шарнирную область полипептида IgG1, содержащую ноль, один или два остатка цистеина; и области СН2 и СН3 иммуноглобулина. Продукты SMIPTM способны к проявлению ADCC и/или CDC.
Несмотря на то что проводились интенсивные исследования по терапии, основанной на применении антител, в данной области остается потребность в усовершенствованных способах лечения заболеваний, ассоциированных с аномальной активностью В-клеток. Описанные и заявленные здесь способы по настоящему изобретению обеспечивают такие усовершенствованные способы, а также их преимущества.
Сущность изобретения
Настоящее изобретение относится к способам снижения количества В-клеток с использованием CD37-специфических связывающих молекул. В некоторых способах по изобретению применение комбинаций CD37-специфических связывающих молекул (одной или более CD37-специфических связывающих молекул) и CD20-специфических связывающих молекул (одной или более CD20-специфических связывающих молекул) приводит к повышенному снижению количества В-клеток. В некоторых из данных способов комбинации являются синергическими. В близком аспекте изобретение относится к способу лечения индивидуума, имеющего заболевание или с подозрением на наличие заболевания, ассоциированного с аномальной активностью В-клеток.
Также настоящее изобретение относится к гуманизированным CD37-специфическим связывающим молекулам (например, гуманизированным конструкциям TRU-016) и способам снижения количества В-клеток с использованием таких молекул. В некоторых вариантах осуществления способов по изобретению предусматривается применение комбинаций гуманизированных конструкций TRU-016 с одной или более CD37-специфическими связывающими молекулами. В другом аспекте изобретение относится к способам лечения индивидуумов, имеющих заболевание или с подозрением на наличие заболевания, ассоциированного с аномальной активностью В-клеток. Связанные аспекты изобретения относятся к способам профилактики любого такого заболевания и способам ослабления симптома, связанного с заболеванием, включающим введение дозы гуманизированной CD37-специфической связывающей молекулы, эффективной для лечения и профилактики такого заболевания или ослабления симптома такого заболевания.
«Аномальная активность В-клеток» относится к активности В-клеток, которая отличается от нормального, правильного или предполагаемого характера. Например, аномальная активность В-клеток может включать неадекватную пролиферацию клеток, ДНК или другие клеточные компоненты которых стали поврежденными или дефектными. Аномальная активность В-клеток может включать пролиферацию клеток, характеристики которой связаны с заболеванием, вызванным, опосредуемым или возникшим в результате неадекватно высоких уровней деления клеток, неадекватно низких уровней апоптоза или обоих факторов вместе. Для таких заболеваний может быть характерна, например, единичная или множественная локальная аномальная пролиферация клеток, групп клеток или ткани(й), независимо от того, являются они раковыми или не раковыми, доброкачественными или злокачественными. Аномальная активность В-клеток также может включать аномальную продукцию антител, такую как продукция аутоантител, или очень повышенную продукцию антител, как правило, желаемых при нормальных уровнях. Предполагается, что аномальная активность В-клеток может иметь место в некоторых субпопуляциях В-клеток и отсутствовать в других субпопуляциях. Также аномальная активность В-клеток может включать неадекватную стимуляцию Т-клеток, возникающую в результате неадекватной презентации В-клетками антигена для Т-клеток, или осуществляемую другими путями с участием В-клеток.
«Лечение» или «проведение лечения» относится к терапевтическому лечению или профилактическому/превентивному лечению. В результате терапевтического лечения может ослабиться, по меньшей мере, один симптом заболевания у индивидуума, получающего лечение, или может замедлиться прогрессирование заболевания у индивидуума, или оно может привести к предупреждению начала развития дополнительных связанных заболеваний.
«Терапевтически эффективная доза» или «эффективная доза» CD20-специфической связывающей молекулы относится к количеству соединения, достаточному для того, чтобы привести к ослаблению одного или более симптомов заболевания, которое лечится. По отношению к отдельному активному ингредиенту, вводимому одному, терапевтически эффективная доза относится к данному одному ингредиенту. По отношению к комбинации терапевтически эффективная доза относится к объединенным количествам активных ингредиентов, которые приводят к проявлению терапевтического эффекта, независимо от того, вводят их последовательно или одновременно. В изобретении, в частности, предусматривается, что одну или более специфических связывающих молекул можно вводить согласно способам по изобретению, каждую в эффективной дозе.
«Индивидуум, имеющий заболевание или с подозрением на наличие заболевания, ассоциированного с аномальной активностью В-клеток», представляет собой индивидуума, у которого заболевание или симптом нарушения могут быть вызваны аномальной активностью В-клеток, могут обостриться в результате аномальной активности В-клеток или могут ослабиться при соответствующей регуляции активности В-клеток. Примерами таких заболеваний является В-клеточное злокачественное заболевание (например, В-клеточная лимфома, В-клеточная лейкемия или В-клеточная миелома), заболевание, для которого характерна продукция аутоантител, или заболевание, характеризующееся несоответствующей презентацией антигена В-клетками для Т-клеток, или вызванное другими путями с участием В-клеток.
В одном примерном аспекте у индивидуума, подвергающегося лечению способами по изобретению, отмечается ответная реакция на лечение, которая сильнее или выше по сравнению с реакцией в ответ на лечение ритуксимабом. Реакция, которая является сильнее по сравнению с таковой, наблюдаемой при лечении ритуксимабом, относится к проявлению клинической ответной реакции, когда лечение способом по изобретению приводит к клинической ответной реакции у пациента, которая сильнее по сравнению с клинической ответной реакции у пациента, получающего лечение ритуксимабом, таким как ритуксан. Более высокая ответная реакция оценивается при сравнении клинических показателей, хорошо известных в данной области и описанных здесь. Примерные показатели включают, но не ограничиваются этим, продолжительность снижения количества В-клеток, снижение количества В-клеток в целом, снижение количества В-клеток в биологической пробе, уменьшение размера опухолей, уменьшение количества опухолей уже имеющихся и/или появляющихся после лечения, и более высокая ответная реакция по оценке самих пациентов и врачей, например, с использованием международного прогностического индекса. Улучшение может иметь место по одному или более чем одному клиническому показателю. Более сильная реакция при применении способа по изобретению может быть за счет неадекватной реакции на предшествующее или настоящее лечение ритуксимабом, например, в результате проявления токсичности и/или неадекватной эффективности лечения ритуксимабом.
В-клеточные злокачественные заболевания включают В-клеточные лимфомы [такие как различные формы болезни Ходжкина, не-ходжкинская лимфома (NHL) или лимфомы центральной нервной системы], лейкемии [такие как острая лимфобластная лейкемия (ALL), хроническая лимфоцитарная лейкемия (СLL), волосистоклеточная лейкемия и хроническая миелобластная лейкемия] и миеломы (такие как множественная миелома). Дополнительные В-клеточные злокачественные заболевания включают мелкоклеточную лимфоцитарную лимфому, В-клеточную пролимфоцитарную лейкемию, лимфоплазмоцитарную лимфому, лимфому маргинальной зоны селезенки, плазмоцитарную миелому, солитарную плазмоцитому костей, внекостную плазмоцитому, экстранодулярную В-клеточную лимфому маргинальной зоны, связанной со слизистой лимфоидной ткани (MALT), нодулярную В-клеточную лимфому маргинальной зоны, фолликулярную лимфому, покровноклеточную лимфому, диффузную крупноклеточную В-клеточную лимфому, крупноклеточную В-клеточную лимфому средостения (тимуса), внутрисосудистую крупноклеточную В-клеточную лимфому, первичную эффузионную лимфому, лимфому Беркитта/лейкемию, пролиферации В-клеток неопределенного злокачественного значения, лимфоматоидный грануломатоз и лимфопролиферативное нарушение после трансплантации.
Нарушения, характеризующиеся продукцией аутоантител, часто расценивают в качестве аутоиммунных заболеваний. Аутоиммунные заболевания включают, но не ограничиваются этим: артрит, ревматоидный артрит, юношеский ревматоидный артрит, остеоартрит, полихондрит, псориатический артрит, псориаз, дерматит, полимиозит/дерматомиозит, миозит с включениями, воспалительный миозит, токсический эпидермальный некролиз, системную склеродерму и склероз, синдром CREST, реакции, связанные с воспалительным заболеванием кишечника, болезнь Крона, язвенный колит, респираторный дистресс-синдром, респираторный дистресс-синдром взрослых (ARDS), менингит, энцефалит, увеит, колит, гломерулонефрит, аллергические состояния, экзему, астму, состояние с инфильтрацией Т-клеток и хронические воспалительные реакции, атеросклероз, аутоиммунный миокардит, недостаточность адгезии лейкоцитов, системную красную волчанку (SLE), подострую кожную красную волчанку, дискоидную волчанку, миелитную волчанку, энцефалитную волчанку, диабет с началом в юношеском возрасте, рассеянный склероз, аллергический энцефаломиелит, нейромиелит зрительного нерва, ревматическую лихорадку, хорею Сиденгама, иммунные реакции, связанные с гиперчувствительностью немедленного или замедленного типа, опосредуемые цитокинами и Т-лимфоцитами, туберкулез, саркоидоз, грануломатоз, включая грануломатоз Вегенера и болезнь Хуга-Штраусса, агранулоцитоз, васкулит (включая васкулит/ангиит при гиперчувствительности, ANCA и ревматоидный васкулит), апластическую анемию, анемию Диамонда-Блэкфана, иммунную гемолитическую анемию, включая аутоиммунную гемолитическую анемию (AIHA), пернициозную анемию, чистую аплазию эритроцитов (PRCA), недостаточность фактора VIII, гемофилию А, аутоиммунную нейтропению, панцитопению, лейкопению, заболевания с диапедезом лейкоцитов, воспалительные заболевания центральной нервной системы (ЦНС), синдром множественного повреждения органов, тяжелую псевдопаралитическую миастению, заболевания, опосредуемые комплексом антиген-антитело, антигломерулярное базальное мембранное заболевание, синдром антител против фосфолипидов, аллергический нейрит, болезнь Бехчета, синдром Кастельмана, синдром Гудпасчера, миастенический синдром Ламберта-Итона, синдром Рейно, синдром Шегрена, синдром Стивенса-Джонсона, отторжение трансплантата солидного органа, болезнь трансплантат против хозяина (GVHD), пемфигоид буллезный, пузырчатку, аутоиммунные полиэндокринопатии, серонегативные спондилоартропатии, болезнь Рейтера, синдром негнущегося человека, гигантоклеточный артериит, иммунный комплексный нефрит, опосредуемые IgА нефропатии, опосредуемые IgМ полинейропатии или опосредуемую IgМ нейропатию, идиопатическую тромбоцитопеническую пурпуру (ITP), тромбоцитопеническую тромбогемолитическую пурпуру (ТTP), пурпуру Геноха-Шенлейна, аутоиммунную тромбоцитопению, аутоиммунное заболевание семенников и яичников, включая аутоиммунный орхит и оофорит, первичный гипотиреоидизм, аутоиммунные эндокринные заболевания, включая аутоиммунный тиреоидит, хронический тиреоидит (тиреоидит Хашимото), подострый тиреоидит, болезнь Аддисона, болезнь Граве, аутоиммунные полижелезистые синдромы (или полижелезистые эндокринопатические синдромы), диабет типа I, также называемый сахарным инсулинозависимым диабетом (IDDM) и синдром Шихана, аутоиммунный гепатит, лимфоидную интерстициальную пневмонию (HIV), бронхолит (нетрансплантантный) против NSIP, синдром Гийена-Барре, васкулит крупных сосудов (включая ревматическую полимиалгию и гигантоклеточный артериит (Такасу), васкулит средних сосудов (включая болезнь Кавасаки и полиартеритнодозный), полиартерит (PAN), анкилозирующий спондилит, болезнь Бергера (опосредуемая IgА нефропатия), быстропрогрессирующий гломерулонефрит, первичный цирроз желчных протоков, брюшную спру (глютеновую энтеропатию), криоглобулинемию, криоглобулинемию, связанную с гепатитом, амиотрофический боковой склероз (ALS), заболевание коронарной артерии, семейную средиземноморскую лихорадку, микроскопический полиангиит, синдром Когана, синдром Вискотта-Олдрича и облитерирующий тромбоангиит.
Ревматоидный артрит (RA) представляет собой хроническое заболевание, характеризующееся воспалением суставов, приводящее к развитию отека, боли и потери функции. У пациентов с RA, как правило, в течение продолжительного периода времени имеет место прогрессирующая деструкция суставов, деформация, отсутствие трудоспособности и преждевременная смерть.
Болезнь Крона и близкое заболевание, язвенный колит, представляют два основных вида заболеваний, которые относятся к группе заболеваний, называемых воспалительным заболеванием кишечника (IBD). Болезнь Крона представляет хроническое заболевание, которое вызывает воспаление пищеварительного или желудочно-кишечного тракта (ЖКТ). Несмотря на то что оно может охватывать площадь ЖКТ от ротовой полости до ануса, чаще всего оно поражает тонкий кишечник и/или ободочную кишку. При язвенном колите поражение ЖКТ ограничивается ободочной кишкой.
Для болезни Крона могут быть характерны антитела против антигенов нейтрофилов, т.е. «перинуклеарные антитела против нейтрофилов» (pANCA), и Saccharomyces cervisiae, т.е. «антитела против Saccharomyces cervisiae» (ASCA). У многих пациентов с язвенным колитом имеются антитела pANCA в крови, но не антитела ASCA, в то время как у многих других пациентов с болезнью Крона имеются антитела ASCA, но антитела pANCA отсутствуют. Одним методом оценки болезни Крона является индекс активности болезни Крона (CDAI), основанный на 18 переменных баллах, собранных врачами. Значения CDAI, равные 150 и ниже, ассоциированы с заболеванием на стадии покоя; значения выше указывают на активную форму заболевания, и значения выше 450 имеют место при тяжелой форме заболевания [Best et al., «Development of a Crohn's disease activity index», Gastroenterology 70: 439-444 (1976)]. Однако в первоначальном исследовании некоторые исследователи применяют «субъективный показатель», равный 200-250, в качестве показателя состояния здоровья.
Системная красная волчанка (SLE) представляет собой аутоиммунное заболевание, вызванное рецидивирующими повреждениями кровеносных сосудов во многих органах, включая почки, кожу и суставы. У пациентов с SLE нарушенное взаимодействие Т-клеток и В-клеток приводит к продукции аутоантител, которые атакуют клеточное ядро. Существует общее мнение, что аутоантитела ответственны за развитие SLE, и таким образом, новые виды лечения, приводящие к снижению количества В-клеток, позволяющие иммунной системе восстанавливать новые В-клетки из предшественников, будут давать надежду на обеспечение длительного лечебного действия у пациентов с SLE.
Рассеянный склероз (MS) также представляет собой аутоиммунное заболевание. Оно характеризуется воспалением центральной нервной системы и разрушением миелина, что приводит к изоляции нервных волокон в головном мозге, спинном мозге и организме в целом. Несмотря на то что причина MS остается неизвестной, существует широко распространенное мнение, что аутоиммунные Т-клетки являются основными клетками, вносящими свой вклад в патогенез заболевания. Однако у пациентов с MS антитела присутствуют в высоких концентрациях в спинно-мозговой жидкости, и благодаря некоторым теориям предполагается, что ответная реакция В-клеток, приводящая к продукции антител, является важной для опосредования заболевания.
Аутоиммунное заболевание щитовидной железы возникает в результате продукции аутоантител, которые стимулируют щитовидную железу с развитием гипертиреоидизма (болезнь Граве) или разрушают щитовидную железу с развитием гипотиреоидизма (тиреоидит Хашимото). Стимуляция щитовидной железы вызывается антителами, которые связываются и активируют рецептор стимулирующего гормона щитовидной железы (TSH). Причиной разрушения щитовидной железы являются аутоантитела, которые взаимодействуют с другими антигенами щитовидной железы.
Синдром Шегрена представляет собой аутоиммунное заболевание, характеризующееся разрушением продуцирующих влагу желез организма.
Причиной иммунной тромбоцитопенической пурпуры (ITP) являются аутоантитела, которые связываются с тромбоцитами крови и вызывают их разрушение.
Тяжелая псевдопаралитическая миастения (MG) представляет собой хроническое аутоиммунное нейромышечное заболевание, характеризующееся аутоантителами, которые связываются с ацетилхолиновыми рецепторами в нейромышечных соединениях, приводя к слабости произвольных групп мышц.
Псориаз характеризуется аутоиммунным воспалением в коже и также связан с артритом в 30% случаев, склеродермой, воспалительным заболеванием кишечника, включая болезнь Крона и язвенный колит.
Также предусматривается лечение идиопатической воспалительной миопатии (IIM), включая дерматомиозит (DM) и полимиозит (РМ). Воспалительные миопатии подразделяют на категории с использованием ряда классификационных систем. Согласно системе классификации Miller (Miller, Rheum. Dis. Clin. North Am., 20: 811-826, 1994) идентифицируется 2 идиопатические воспалительные миопатии (IIM), полимиозит (РМ) и дерматомиозит (DM).
Полимиозит и дерматомиозит являются хроническими, разрушительными воспалительными заболеваниями, при которых поражаются мышцы и в случае DM - кожа. Данные заболевания являются редкими, по некоторым сообщениям, их частота в США составляет примерно 5-10 случаев на миллион взрослых и 0,6-3,2 случаев на миллион детей (Targoff, Curr. Probl. Dermatol. 1991, 3: 131-180). Для идиопатической воспалительной миопатии характерно тяжелое течение и смертность, при том что у половины заболевших взрослых имеются существенные повреждения (Gottdiener et al., Am. J. Cardiol. 1978, 41: 1141-49). Miller (Rheum. Dis. Clin. North Am., 1994, 20: 811-826 и Arthritis and Allied Conditions, Ch. 75, Eds. Koopman and Moreland, Lippincott Williams and Wilkins, 2005) выделяет пять групп критериев, используемых для диагностики IIM, т.е. критериев оценки идиопатической воспалительной миопатии (IIM), которые включают мышечную слабость, доказательства наличия дегенерации мышц, полученные при биопсии, повышенную активность в сыворотке крови связанных с мышцами ферментов, наличие электромагнитной триады миопатии, кожные высыпания при дерматомиозите, и также включаются данные о наличии аутоантител в качестве вторичного критерия.
Ассоциированные с IIM факторы, в том числе связанные с мышцами ферменты и аутоиммунные антитела, включают, но не ограничиваются этим, креатининкиназу (СК), лактатдегидрогеназу, альдолазу, С-реактивный белок, аспартатаминотрансферазу (АСТ), аланинаминотрансферазу (АЛТ) и антинуклерные антитела (ANA), специфические для миозита антитела (MSA) и антитела против экстрагируемых нуклеарных антигенов.
«Связывающая молекула» по изобретению может представлять, например, белок («белок» может быть полипептидом или пептидом), нуклеиновую кислоту, углевод, липид или соединение с малой молекулой, которое связывается с мишенью. Тип белковой связывающей молекулы, которая подразумевается в изобретении, представляет собой антитело или фрагмент антитела, который сохраняет способность к связыванию. Связывающую молекулу можно модифицировать по изобретению стандартными в данной области способами для повышения ее аффинности связывания, снижения иммуногенности, изменения ее эффекторных функций и/или повышения ее доступности в организме индивидуума. Такие модификации молекул могут включать, например, модификации аминокислотной последовательности или экспрессии в виде рекомбинантного белка. Такие рекомбинантные белки также являются связывающими молекулами по изобретению. Примером связывающей молекулы по изобретению является малый модуляторный иммунофармацевтический препарат (SMIPTM).
Связывающая молекула, которая является «специфической» для мишени, связывается с такой мишенью с более высокой аффинностью по сравнению с любой другой мишенью. Например, CD37-специфическая связывающая молекула связывается с CD37 с большей аффинностью по сравнению с любой другой мишенью, и CD20-специфическая связывающая молекула связывается с CD20 с большей аффинностью по сравнению с любой другой мишенью. Связывающие молекулы по изобретению могут обладать аффинностью для их мишеней со значениями Ка выше чем или на уровне примерно 104 М-1, предпочтительно выше чем или на уровне примерно 105 М-1, более предпочтительно выше чем или на уровне примерно 106 М-1 и еще более предпочтительно выше чем или на уровне примерно 107 М-1. Значения аффинности даже выше чем примерно 107 М-1 более предпочтительны, такие как аффинности на уровне или выше чем примерно 107 M-1, примерно 108 М-1 и примерно 109 М-1, и примерно 1010 М-1. Аффинность связывающих молекул по настоящему изобретению можно легко определить с использованием обычных методик, например, описанных Scatchard et al., Ann. N.Y. Acad. Sci. 51: 660 (1949).
Некоторые CD37-специфические связывающие молекулы, предусмотренные изобретением, обладают аффинностью по отношению к CD37 на уровне примерно от 0,5 до примерно 10 нМ. Некоторые CD20-специфические связывающие молекулы, предусмотренные изобретением, обладают аффинностью по отношению к CD37 на уровне примерно от 1 до примерно 30 нМ.
Другим свойством некоторых CD37-специфических связывающих молекул, предусмотренных изобретением, является то, что они имеют период полураспада в пределах примерно от 7 до примерно 30 суток.
Антителами, специфичными к CD37, с помощью которых характеризовали антиген CD37 на семинаре Thrid HLDA Workshop, были HD28, G28-1, HH1, BI14, WR17 и F93G6. Смотри Ling and MacLennan, pp. 302-335 в Leucocyte Typing III. White Cell Differentiation Antigens, Oxford University Press (1987). Другие описанные антитела, специфичные по отношению к CD37, включают RFB-7, Y29/55, MB-1, M-B371, M-B372 и IPO-24. Смотри Moldenhaurer, J. Biol., Regul. Homeost. Agents, 14: 281-283 (2000), который утверждает, что все данные антитела распознают только один эпитоп CD37. Schwartz-Albeiz et al., 14: 905-914 (1988) указывают, что эпитоп расположен в углеводной группе CD37. Еще одним CD37-специфическим антителом является S-B3 (Biosys).
Патенты и публикации патентов, в которых описываются антитела против CD20, включают патенты США № 5776456, 5736137, 6399061 и 5843439, а также заявки на патент США № US 2002/0197255А1 и US 2003/0021781А1 (Anderson et al.); патент США № 6455043В1 и WO00/0960 (Grillo-Lopez, A.); WO00/27428 (Grillo-Lopez and White); WO00/27428 (Grillo-Lopez and Leonard); WO00/44788 (Braslawsky et al.); WO01/10462 (Rastetter W.); WO01/10461 (Rastetter and White); WO01/10460 (White and Grillo-Lopez); заявку на патент США № US 2002/0006404 и WO02/04021 (Hanna and Hariharan); заявку на патент США № US 2002/0012665A1 и WO01/74388 (Hanna N.); заявку на патент США № US 2002/0009444A1 и WO01/80884 (Grillo-Lopez, A.); WO01/97858 (White C.); заявку на патент США № US 2002/0128488A1 и WO02/34790 (Reff, M.); WO02/060955 (Braslawsky et al.); WO02/096948 (Braslawsky et al.); WO02/079255 (Reff and Davies); патент США № 6171586В1 и WO98/56418 (Lam et al.); WO98/58964 (Raju, S.); WO99/22764 (Raju, S.); WO99/51642; патент США № 6194551В1, патент США № 6242195В1, патент США № 6528624В1 и патент США № 6538124 (Idusogie et al.); WO00/42072 (Presta, L.); WO00/67796 (Curd et al.); WO01/03734 (Grillo-Lopez et al.); заявку на патент США № US 2002/0004587A1 и WO01/77342 (Miller and Presta); заявку на патент США № US 2002/0197256 (Grewal, I.); патенты США № 6090365B1, 6287537В1, 6015542, 5843398 и 5595721 (Kaminski et al.); патенты США № 5500362, 5677180, 5721108 и 6120767 (Robinson et al.); патент США № 6410391B1 (Raubitschek et al.); патент США № 6224866B1 и WO00/20864 (Barbera-Guillem, E.); WO01/13945 (Barbera-Guillem, E.); WO00/67795 (Goldenberg); WO00/74718 (Goldenberg and Hansen); WO00/76542 (Golay et al.); WO01/72333 (Wolin and Rosenblatt); патент США № 6368596B1 (Ghetie et al.); заявку на патент США № US 2002/0041847A1 (Goldenberg, D.); заявку на патент США № US 2003/0026801A1 (Weiner and Hartmann); WO02/102312 (Engleman, E.); каждый из этих источников включен здесь в виде ссылки. Смотри также патент США № 5849898 и заявку на европейский патент № 330191 (Seed et al.); патент США № 4861579 и EP332865A2 (Meyer and Weiss); и WO95/03770 (Bhat et al.).
Ритуксимаб был разрешен для применения в клинике у людей в виде ритуксана®. Ритуксан® считается специфической в отношении CD20 связывающей молекулой по изобретению.
Малые модуляторные иммунофармацевтические препараты (SMIP) рассматриваются в качестве одного типа связывающих молекул по изобретению. Способы получения SMIP были описаны ранее в совместной заявке на патент США № 10/627556 и публикации заявки на патент США 2003/0133939, 2003/0118592 и 2005/0136049, которые в полном объеме включены здесь в виде ссылки. SMIP являются новыми рекомбинантными белками со связывающим доменом иммуноглобулинов, которые, как правило, имеют связывающий домен для «своей» структуры, такой как антиген, противорецептор или тому подобное, полипептид шарнирной области IgG1, IGA и IGЕ или мутантный полипептид шарнирной области IgG1, содержащий ноль, один или два остатка цистеина, и области СН2 и СН3 иммуноглобулина. В одном варианте осуществления молекула связывающего домена имеет один или два остатка цистеина (Cys) в шарнирной области. В близком варианте осуществления, когда молекула связывающего домена содержит два остатка Cys, первый Cys, который принимает участие в связывании тяжелой и легкой цепей, не подвергается делеции или замене на другую аминокислоту.
Связывающие домены молекул, пригодных в способах по изобретению, предусмотрены как имеющие одну или более связывающих областей, таких как вариабельная связывающая область легкой цепи и вариабельная связывающая область тяжелой цепи, полученные из одного или более представителей надсемейства иммуноглобулинов, таких как иммуноглобулин. Кроме того, данные области, как правило, разделены пептидами-линкерами, которые могут представлять собой любой пептид-линкер, известный в данной области, совместимый с соединительным доменом или областью в связывающей молекуле. Примерами линкеров являются линкеры на основе мотива-линкера Gly4Ser, такого как (Gly4Ser)n, где n=1-5. Молекулы для применения в способах по изобретению также включают достаточную аминокислотную последовательность, полученную из константной области иммуноглобулина для обеспечения эффекторной функции, предпочтительно ADCC и/или CDC. Таким образом, молекулы будут содержать последовательность, полученную из области СН2 иммуноглобулина, или областей СН2 и СН3, полученных из одного или более иммуноглобулинов. SMIP способны к проявлению ADCC и/или CDC, но не обладают способностью к образованию связанных дисульфидными связями мультимеров.
Изобретение включает гуманизированные CD37-специфические полипептиды SMIP, которые обладают, по меньшей мере, 80%-ной идентичностью по отношению к полипептиду с последовательностью SEQ ID NO:2, где гуманизированные CD37-специфические полипептиды SMIP связываются с CD37. В одном аспекте гуманизированные CD37-специфические полипептиды SMIP содержат любую аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 80, 82, 84, 86 и 88. В другом аспекте гуманизированные CD37-специфические полипептиды SMIP содержат, по меньшей мере, одну аминокислотную модификацию в гипервариабельном участке (CDR), выбранную из группы, состоящей из: CDR1 легкой цепи, CDR1 тяжелой цепи, CDR2 легкой цепи, CDR2 тяжелой цепи, CDR3 легкой цепи и CDR3 тяжелой цепи.
В одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR1 легкой цепи имеет аминокислотную последовательность SEQ ID NO:61 (RASENVYSYLA). Изобретение также включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR1 легкой цепи имеет аминокислотную последовательность SEQ ID NO:62 (RTSENVYSYLA). Изобретение также включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR1 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:63 (GYMNM).
В другом варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR2 легкой цепи имеет аминокислотную последовательность SEQ ID NO:64 (FAKTLAE). Изобретение также включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR2 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:65 (NIDPYYGGTTTYNRKFKG).
В дополнительном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR3 легкой цепи имеет аминокислотную последовательность SEQ ID NO:66 (QHHSDNPWT). Изобретение дополнительно включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR3 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:67 (SVGPFDY). Изобретение дополнительно включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR3 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:68 (SVGPFDS). Изобретение также включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR3 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:69 (SVGPMDY).
В еще одном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, содержащий, по меньшей мере, одну, по меньшей мере, две или, по меньшей мере, три последовательности из аминокислотных последовательностей CDR легкой цепи, выбранных из группы, состоящей из последовательностей SEQ ID NO:61, 62, 64 и 66. В еще одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, содержащий аминокислотную последовательность CDR1 легкой цепи SEQ ID NO:61 или 62 или их вариант, в которых одна или две аминокислоты SEQ ID NO:61 или 62 заменены; аминокислотную последовательность CDR2 легкой цепи SEQ ID NO:64 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:64 заменены; и аминокислотную последовательность CDR3 легкой цепи SEQ ID NO:66 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:66 заменены.
В еще одном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, содержащий, по меньшей мере, одну, по меньшей мере, две или, по меньшей мере, три аминокислотные последовательности CDR тяжелой цепи, выбранные из группы, состоящей из последовательностей SEQ ID NO:63, 65, 67-69. В дополнительном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, содержащий аминокислотную последовательность CDR1 тяжелой цепи SEQ ID NO:63 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:63 заменены; аминокислотную последовательность CDR2 тяжелой цепи SEQ ID NO:65 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:65 заменены; и аминокислотную последовательность CDR3 тяжелой цепи, выбранную из группы, состоящей из последовательностей SEQ ID NO:67-69 или их варианта, в которых одна или две аминокислоты любой из последовательностей SEQ ID NO:67-69 заменены.
Изобретение включает гуманизированные CD37-специфические полипептиды SMIP, имеющие, по меньшей мере, одну аминокислотную модификацию в каркасной области (FR), выбранную из группы, состоящей из: FR1 легкой цепи, FR1 тяжелой цепи, FR2 легкой цепи, FR2 тяжелой цепи, FR3 легкой цепи, FR3 тяжелой цепи, FR4 легкой цепи, FR4 тяжелой цепи. В одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором первая каркасная область (FR1) легкой цепи имеет аминокислотную последовательность SEQ ID NO:70 (EIVLTQSPATLSLSPGERATLSC). В другом варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR1 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:71 (EVQLVQSGAEVKKPGESLKISCKGSGYSFT). В еще одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR2 легкой цепи имеет аминокислотную последовательность SEQ ID NO:72 (WYQQKPGQAPRLLIY). В дополнительном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR2 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:73 (WVRQMPGKGLEWMG). В еще одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR3 легкой цепи имеет аминокислотную последовательность SEQ ID NO:74 (GIPARFSGSGSGTDFTLTISSLEPEDFAVYYC). В еще одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR3 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:75 (QVTISADKSISTAYLQWSSLKASDTAMYYCAR). В еще одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR4 легкой цепи имеет аминокислотную последовательность SEQ ID NO:76 (FGQGTKVEIK). В еще одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR4 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:77 (WGQGTLVTVSS). В еще одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR4 тяжелой цепи имеет аминокислотную последовательность SEQ ID NO:78 (WGRGTLVTVSS).
Изобретение дополнительно включает гуманизированные CD37-специфические полипептиды SMIP, содержащие, по меньшей мере, одну, по меньшей мере, две или, по меньшей мере, три последовательности из аминокислотных последовательностей FR легкой цепи, выбранных из группы, состоящей из последовательностей SEQ ID NO:70, 72, 74 и 76. В еще одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, содержащий аминокислотную последовательность FR1 легкой цепи SEQ ID NO:70 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:70 заменены; аминокислотную последовательность FR2 легкой цепи SEQ ID NO:72 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:72 заменены; аминокислотную последовательность FR3 легкой цепи SEQ ID NO:74 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:74 заменены; и аминокислотную последовательность FR4 легкой цепи SEQ ID NO:76 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:76 заменены.
Кроме того, изобретение дополнительно включает гуманизированные CD37-специфические полипептиды SMIP, содержащие, по меньшей мере, одну, по меньшей мере, две или, по меньшей мере, три последовательности из аминокислотных последовательностей FR тяжелой цепи, выбранных из группы, состоящей из последовательностей SEQ ID NO:71, 73, 75, 77 и 78. В одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, содержащий аминокислотную последовательность FR1 тяжелой цепи SEQ ID NO:71 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:71 заменены; аминокислотную последовательность FR2 тяжелой цепи SEQ ID NO:73 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:73 заменены; аминокислотную последовательность FR3 тяжелой цепи SEQ ID NO:75 или ее вариант, в которой одна или две аминокислоты SEQ ID NO:75 заменены; и аминокислотную последовательность FR4 тяжелой цепи SEQ ID NO:77 или 78 или их вариант, в которых одна или две аминокислоты SEQ ID NO:77 или 78 заменены.
Изобретение также включает выделенную молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую гуманизированный CD37-специфический полипептид SMIP, который обладает, по меньшей мере, 80%-ной идентичностью по отношению к полипептиду с последовательностью SEQ ID NO:2, где гуманизированный CD37-специфический полипептид SMIP связывается с CD37. Такая выделенная молекула нуклеиновой кислоты может содержать нуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 79, 81, 83, 85 и 87. В одном варианте осуществления изобретение включает векторы, которые содержат данные молекулы нуклеиновой кислоты, и клетки-хозяева, содержащие векторы.
Изобретение также включает способы получения полипептидов, описанных здесь, включающие культивирование клеток-хозяев в подходящих условиях для экспрессии полипептидов, и необязательно выделение полипептидов из культуры.
В еще одном аспекте изобретение включает композиции, содержащие гуманизированные CD37-специфические полипептиды SMIP по изобретению и фармацевтически приемлемый носитель.
Изобретение дополнительно включает применение CD37-специфических SMIP и CD37-специфических связывающих молекул, описанных здесь, в любом из способов по изобретению. Такие способы включают применение любого из CD37-специфических SMIP и CD37-специфических связывающих молекул, содержащих аминокислотную последовательность, выбранную из последовательностей SEQ ID NO:6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 80, 82, 84, 86 и 88.
В еще одном аспекте изобретение включает наборы для снижения количества В-клеток, содержащие композиции по изобретению; и протоколы по применению наборов для снижения количества В-клеток. Такие наборы могут дополнительно содержать одну или более CD20-специфических связывающих молекул. В изобретении предусматривается, что такой CD20-специфической связывающей молекулой является TRU-015.
Изобретение также включает CD37-специфические полипептиды SMIP, содержащие CDR1, CDR2 и CDR3, которые обладают, по меньшей мере, 80%-ной идентичностью по отношению к полипептиду с последовательностью SEQ ID NO:2. Такие CD37-специфические полипептиды SMIP могут дополнительно включать человеческую каркасную область, разделяющую каждый из CDR1, CDR2 и CDR3.
В другом аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, который обладает, по меньшей мере, 80%-ной идентичностью по отношению к полипептиду с последовательностью SEQ ID NO:2, где гуманизированный CD37-специфический полипептид SMIP связывается с CD37 и содержит полипептид шарнирной области, имеющий аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:90, 92, 94, 96, 98, 100, 102, 104, 106, 108, 110, 112, 114, 115, 116, 118, 120, 122, 124, 126 и 127.
В изобретении также предусматривается гуманизированный CD37-специфический полипептид SMIP, который обладает, по меньшей мере, 80%-ной идентичностью по отношению к полипептиду с последовательностью SEQ ID NO:2, где гуманизированный CD37-специфический полипептид SMIP связывается с CD37 и содержит линкер, содержащий (Gly4Ser)n, где n равно 1, 2, 3, 4, 5 или 6.
В еще одном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR1 легкой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:128 (RTSQNVYSYLA), 129 (RTSESVYSYLA), 130 (RASQSVYSYLA), 131 (RASQSVSSYLA) и 132 (RASQSVSYYLA). В другом варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR1 тяжелой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:133 (SYMNM) и 134 (SYWIG). В дополнительном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR2 легкой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:135 (AASSLQS), 136 (GASTRAT) и 137 (DASNRAT). В еще одном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR2 тяжелой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:138 (IIYPGDSDTRYSPSFQG) и 139 (RIDPSDSYTNYSPSFQG).
В дополнительном варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR3 легкой цепи имеет аминокислотную последовательность SEQ ID NO:220 (QHHSDNPWT). В другом варианте осуществления изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором CDR3 тяжелой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:211 (SVGPMDY), 212 (SVGPFDY), 213 (SVGPMDV), 214 (SVGPFDS), 215 (SVGPFDP), 217 (SVGPFDV), 218 (SVGPFDI) и 219 (SVGPFDL).
В еще одном аспекте изобретение включает гуманизированные CD37-специфические полипептиды SMIP с альтернативными каркасными областями. В одном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR1 легкой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:170-181. В другом аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR1 тяжелой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:140-146. В еще одном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR2 легкой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:182-193. В еще одном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR2 тяжелой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:147-153. В дополнительном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR3 легкой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:194-205. В еще одном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR3 тяжелой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:154-160. В дополнительном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR4 легкой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:206-210. В еще одном аспекте изобретение включает гуманизированный CD37-специфический полипептид SMIP, в котором FR4 тяжелой цепи имеет аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:161-169.
Примеры CD37-специфических SMIP, пригодных в изобретении, включают, но не ограничиваются этим, G28-1 scFv (SSS-S)H WCH2 WCH3, который состоит из одноцепочечного Fv-фрагмента G28-1, в котором все три остатка цистеина в соединении или шарнирных областях подвергались мутации в остатки серина, и области СН2 и СН3 дикого типа; G28-1 scFv IgAH WCH2 WCH3, содержащий шарнирную область IgA и области WT IgG1; G28-1 scFv VHL11S (SSS-S)H WCH2 СН3, в котором все три остатка цистеина в соединении или шарнирных областях подвергались мутации в остатки серина и лейцин в положении 11 вариабельной области тяжелой цепи замещен на серин; G28-1 scFv VHL11S (СSS-S)H WCH2 СН3, в котором остатки цистеина замещены во втором и третьем положениях на серин; G28-1 scFv VHL11S (СSС-S)H WCH2 СН3, в котором остатки цистеина замещены во втором положении на серин; G28-1 scFv VH11S (SSC-P)Н WCH2 WСН3 (называемый здесь TRU-016), в котором первый и второй остатки цистеина в соединении или шарнирных областях подвергались мутации с заменой остатков серина и лейцина в положении 11 вариабельной области тяжелой цепи на серин; G28-1 scFv VH11S (SСS-S)H WCH2 WСН3, в котором первый и третий остатки цистеина в шарнирных областях подвергались мутации в остатки серина; G28-1 scFv VH11S (ССS-P)Н WCH2 WСН3, в котором третий остаток цистеина в шарнирной области замещен на серин; G28-1 scFv VHL11S (SСС-P)Н WCH2 WСН3, в котором первый остаток цистеина замещен на серин; G28-1 scFv VHL11S mlgE CH2 СН3 CH4, содержащий области СН 2-4 мышиного IgE, в котором лейцин в положении 11 вариабельной области тяжелой цепи замещен на серин; G28-1 scFv VHL11S mlgA WlgACH2 T4CH3, содержащий шарнирную область мышиного IgA с СН2 IgA дикого типа и усеченную область СН3 IgA без 4 аминокислот на карбоксильном конце GTCY; G28-1 scFv VHL11S hlgE CH2 CH3 CH4, содержащий области СН IgЕ, в которых лейцин в положении 11 вариабельной области тяжелой цепи замещен на серин; и G28-1 scFv VHL11S hlgАН WlgACH2 ТCH3, содержащий шарнирную область IgA, СН2 IgA дикого типа и усеченную СН2 IgA, и усеченную область СН3 IgA без 4 аминокислот на карбоксильном конце GTCY.
Примеры CD20-специфических SMIP, пригодных в изобретении, включают SMIP, полученные из моноклонального антитела против C20 2Н7, описанного в публикации патента США №№ 2003/133939 и 2003/0118592. SMIP включают 2H7 scFv-Ig или его производное. Производные включают CytoxB-MHWTG1C, который имеет Fc-область человеческого IgG1 и мутантную шарнирную область IgG1; CytoxB-MHМG1C, который содержит мутантную Fc-область; MG1H/MG1C, который содержит Fc-рецептор с подвергшимся мутации остатком лейцина 234; CytoxB-IgAHWTHG1C, который содержит участок шарнирной области человеческого IgА, гибридизованный с человеческой Fc-областью дикого типа; 2H7 scFv-IgG1 ламы, содержащий шарнирную область и области СН2СН3 IgG1 ламы; 2H7 scFv-IgG2 ламы, содержащий шарнирную область и области СН2СН3 IgG2 ламы; 2H7 scFv-IgG3 ламы, содержащий шарнирную область и области СН2СН3 IgG3 ламы.
2H7 scFv MTH (SSS) WTCH2CH3, в котором все три остатка цистеина в соединении или шарнирных областях подверглись мутации в остатки серина, и области СН2 и СН3 дикого типа; 2H7 scFv MTH (SSС), в котором первые два остатка цистеина замещены на остатки серина; 2H7 scFv MTH (SСS), в котором первый и третий остатки цистеина замещены на остатки серина; 2H7 scFv MTH (СSS), WTCH2CH3, в котором остатки цистеина замещены во втором и третьем положениях на серин; 2H7 scFv VH11SER IgG MTH (SSS) WTCH2CH3, в котором лейцин в положении 11 в вариабельной области тяжелой цепи замещен на серин; 2H7 scFv IgA шарнирная область-IgG1 CH2-CH3, содержащий шарнирную область IgА и области WT IgG1; 2H7 scFv IgA шарнирная область-CH2-CH3, содержащий шарнирную область IgА и области СН2-3; 2H7 IgAWH IgACH2-T4CH3, содержащий шарнирную область IgA, СН2 IgA дикого типа и усеченную область СН3 IgA без 4 аминокислот на карбоксильном конце GTCY.
Производные с мутациями в области СН3 IgG включают 2H7 scFv MTH WTCH2 MTCH3 Y405, в котором остаток фенилаланина в положении 405 (система нумерации по Kabat et al., выше) замещен на тирозин; 2H7 scFv MTH WTCH2 MTCH3 А405, в котором фенилаланин в положении 405 замещен на аланин; scFv MTH WTCH2 MTCH3 А407, в котором остаток тирозина в положении 407 замещен на аланин; scFv MTH WTCH2 MTCH3 Y405А407, содержащий две мутации; и scFv MTH WTCH2 MTCH3 А405А407, содержащий две мутации.
2H7 scFv MTH (СSS) WTCH2CH3 представляет конструкцию с третьим остатком цистеина в шарнирной области IgG1, замещенным на остаток серина. 2H7 scFv IgG MTH (SSS) MTCH2WTCH3 SMIP содержит мутантную шарнирную область (MH (SSS)) и мутантную область СН2, в которой пролин в положении 238 (по Ward et al.) замещен на серин.
Производные 2H7scFv-Ig также включают мутанты 2H7 scFv с точечными мутациями в вариабельной области тяжелой цепи. Все последующие конструкции включают мутации, в которых лейцин в положении 11 в вариабельной области тяжелой цепи замещен на серин: 2H7 scFv VH11SER IgG MTH (SSS-S) WTCH2CH3, 2H7 scFv VHL11SER (CSS-S) H WTCH2 WCH3, содержащие подвергшуюся мутации шарнирную область, представленную выше; 2H7 scFv VHL11SER (СSС-S) Н WTCH2 WCH3, содержащий подвергшуюся мутации шарнирную область, представленную выше; 2H7 scFv VHL11S IgAH IgACH2 T4CH3, содержащий шарнирную область IgA, WT IgA CH2 и усеченную CH3 IgA; 2H7 scFv VHL11S IgECH2 СН3 СН4, содержащий области СН 2-4 IgE; 2H7 scFv VHL11S (SSS-S) IgЕCH3СН4, содержащий мутантную шарнирную область и области СН3 и СН4 IgЕ; 2H7 scFv VHL11S mlgE CH2 CH3 CH4, содержащий области мышиного IgЕ; 2H7 scFv VHL11S mlgАН WIGACH2 T4CH3, содержащий мутации, описанные выше, и константную область мышиного IgA, состоящую из области СН2 дикого типа и мутантную область СН3; 2H7 scFv VHL11S (SSS-S) H K322S CH2 WCH3, содержащий мутацию в области СН2 человеческого IgG1 в остатке 322, где лизин замещен на серин; 2H7 scFv VHL11S (СSS-S) H K322S CH2 WCH3, содержащий мутантную шарнирную область, представленную выше, и мутантную область СН2, описанную ранее; 2H7 scFv VHL11S (SSS-S) H Р331S CH2 WCH3, содержащий мутантную шарнирную область, представленную выше, и мутантную область СН2, в которой пролин в остатке 331 замещен на серин; 2H7 scFv VHL11S (СSS-S) H Р331S CH2 WCH3, содержащий мутантную шарнирную область и мутацию с заменой пролина на серин в положении 331 в области СН2; 2H7 scFv VHL11S (SSS-S) H T256N CH2 WCH3, содержащий мутантную шарнирную область и мутацию с заменой треонина на аспарагин в положении 256 в области СН2; 2H7 scFv VHL11S (SSS-S) H RTPE/QNAK (255-258) CH2 WCH3, содержащий мутантную шарнирную область и ряд мутаций, в которых остатки 255-258 подверглись мутации с заменой аргинина, треонина, пролина, глутаминовой кислоты соответственно на глутамин, аспарагин, аланин и лизин; 2H7 scFv VHL11S (SSS-S) H K290Q CH2 WCH3, содержащий мутантные шарнирные области и замену лизина на глутамин в положении 290; 2H7 scFv VHL11S (SSS-S) H А339Р CH2 WCH3, содержащий мутантную шарнирную область и замену аланина на пролин в положении 339; SMIP 2H7 scFv (SSS-S) H P238S CH2 WCH3, содержащий мутантную шарнирную область и замену пролина на серин в положении 238 в области СН2, которая является такой же, как в 2H7 scFv IgG MTH (SSS) MTCH2 WTCH3. 2H7 scFv IgGАH IGAHCH2 T18CH3 содержит шарнирную область IgА дикого типа и область СН2 и область СН3 с усеченным участком длиной 18 аминокислот на карбоксильном конце.
Связывающая молекула по изобретению может содержать нативный и рекомбинантный внеклеточный домен из другого белка, который приводит к повышению связывающей способности молекулы. В одном варианте осуществления внеклеточный домен выбран из группы, состоящей из CD154 и CTLA4.
«Синергическая комбинация» CD37-специфических связывающих молекул и CD20-специфических связывающих молекул представляет комбинацию, которая обладает эффектом, который выше по сравнению с суммой эффектов связывающих молекул при введении их одних.
В одном аспекте изобретения связывающие молекулы вводят в одной или более фармацевтических композициях. Для введения связывающих молекул человеку или опытным животным предпочтительно формулировать связывающие молекулы в композиции, содержащей один или более фармацевтически приемлемых носителей. Выражение «фармацевтически или фармакологически приемлемый» относится к молекулам или композициям, которые не приводят к развитию аллергических или других побочных реакций при введении с использованием путей, известных в данной области, описанных ниже. «Фармацевтически приемлемые носители» включают любые или все пригодные для применения в клинике растворители, дисперсионные среды, оболочки, антибактериальные и противогрибковые препараты, изотонические вещества и вещества для замедления всасывания и тому подобное.
Кроме того, соединения могут образовывать сольваты с водой или обычными органическими растворителями. Такие сольваты также предусматриваются.
Композиции связывающих молекул можно вводить перорально, местно, трансдермально, парентерально, ингаляционно, интравагинально, ректально или внутричерепной инъекцией. Термин парентарально, в том смысле, в котором он здесь используется, включает подкожные инъекции, внутривенную, внутримышечную, интрацистенальную инъекцию или инфузии. Также предусматривается введение внутривенной, внутрикожной, внутримышечной инъекцией, инъекций в молочную железу, внутрибрюшинной, внутриоболочечной, ретробульбарной, внутрилегочной инъекцией или хирургической имплантацией в определенное место. Как правило, композиции в основном свободны от пирогенов, а также других загрязнителей, которые могут быть опасными для реципиента. Предпочтительной является инъекция, в частности, внутривенная.
Фармацевтические композиции по настоящему изобретению, содержащие связывающие молекулы, применяемые в способе по изобретению, могут содержать фармацевтически приемлемые носители или добавки в зависимости от пути введения. Примеры таких носителей или добавок включают воду, фармацевтически приемлемый органический растворитель, коллаген, поливиниловый спирт, поливинилпирролидон, карбоксивиниловый полимер, натриевую соль карбоксиметилцеллюлозы, полиакрилат натрия, альгинат натрия, водорастворимый декстран, карбоксиметил-крахмал натрий, пектин, метилцеллюлозу, этилцеллюлозу, ксантановую камедь, аравийскую камедь, казеин, желатин, агар, диглицерин, глицерин, пропиленгликоль, полиэтиленгликоль, вазелин, парафин, стеариловый спирт, стеариновую кислоту, человеческий сывороточный альбумин (HAS), маннит, сорбит, лактозу, фармацевтически приемлемое поверхностно-активное вещество и тому подобное. Используемые добавки выбраны, но не ограничиваются этим, из вышеуказанных добавок или их комбинаций, там где что подходит, в зависимости от лекарственной формы по настоящему изобретению.
Состав фармацевтической композиции будет варьировать в зависимости от выбранного пути введения (например, раствор, эмульсия). Подходящую композицию, содержащую антитело для введения, можно приготовить в физиологически приемлемом растворителе или носителе. Для растворов или эмульсий подходящие носители включают, например, водные или спиртовые/водные растворы, эмульсии или суспензии, включающие физиологический раствор или забуференную среду. Растворители для парентерального введения могут включать раствор хлорида натрия, раствор декстрозы Рингера, декстрозу или хлорид натрия, раствор Рингера с лактозой или нелетучие жирные масла. Растворы для внутривенного введения могут включать различные добавки, консерванты или жидкость, питательное вещество или восполнитель электролитов.
Разнообразные водные носители, например вода, забуференная вода, 0,4%-ный солевой раствор, 0,3%-ный глицин, или водные суспензии могут содержать активное соединение в смеси с наполнителями, подходящими для получения водных суспензий. Такие наполнители представляют собой суспендирующие вещества, например натриевую соль карбоксиметилцеллюлозы, метилцеллюлозу, гидроксипропилметилцеллюлозу, альгинат натрия, поливинилпирролидон, трагакантовую камедь или аравийскую камедь; диспергирующие или смачивающие вещества могут представлять собой природный фосфатид, например, лецитин, или продукты конденсации оксида алкилена с жирными кислотами, например полиоксиэтиленстеарат, или продукты конденсации оксида этилена с длинноцепочечными алифатическими спиртами, например гептадекаэтиленоксицетанол, или продукты конденсации оксида этилена с неполными эфирами, полученными из жирных кислот, и гексита, такие как полиоксиэтиленсорбит моноолеат, или продукты конденсации оксида этилена с неполными эфирами, полученными из жирных кислот, и ангидридами гексита, например полиэтиленсорбитан моноолеат. Водные суспензии могут также содержать один или более консервантов, например этил- или н-пропил-, п-гидроксибензоат.
Композиции связывающих молекул можно лиофилизировать для хранения или восстановления в подходящем носителе перед применением. Было показано, что данный метод является эффективным для обычных иммуноглобулинов. Можно использовать любые методы лиофилизирования и восстановления. Специалисты в данной области, очевидно, понимают, что лиофилизирование и восстановление могут привести к потере активности антитела в различной степени и что данные уровни применения могут быть компенсированы.
Диспергируемые порошки и гранулы, подходящие для приготовления водной суспензии добавлением воды, обеспечивают активное соединение в смеси с диспергирующим или смачивающим веществом, суспендирующим веществом или одним или более консервантами. Примерами подходящих диспергирующих или смачивающих веществ и суспендирующих веществ являются уже указанные выше.
Концентрация связывающей молекулы в данных композициях может варьировать в широких пределах, например, от менее чем примерно 0,5%, как правило, примерно на уровне или, по меньшей мере, примерно от 1% до 15 или 20% по массе, и она будет выбрана в основном с учетом объемов жидкостей, вязкости и т.д., в зависимости от выбранного способа введения. Так, типичная фармацевтическая композиция для парентеральной инъекции может быть приготовлена с содержанием 1 мл стерильной забуференной воды и 50 мг антитела. Типичная композиция для внутривенной инфузии может быть приготовлена с содержанием 250 мл стерильного раствора Рингера и 150 мг антитела. Фактические способы получения композиций для парентерального введения известны и очевидны специалистам в данной области и более подробно они описаны, например, в Remington's Pharmaceutical Science, 15th ed., Mack Publishing Company, Easton, Pa. (1980). Эффективная доза антитела находится в пределах от 0,01 мг до 1000 мг на кг массы тела на введение.
Фармацевтические композиции могут находиться в форме стерильной инъекционной водной, масляной суспензии, дисперсий или стерильных порошков для приготовления стерильных инъекционных растворов или дисперсий экстемпоро. Суспензию можно готовить известными в данной области методами с использованием подходящих диспергирующих или смачивающих веществ, и суспендирующих веществ, указанных выше. Также стерильный инъекционный препарат может представлять собой стерильный инъекционный раствор или суспензию в нетоксичном, приемлемом для парентерального введения разбавителе или растворителе, например, в виде раствора в 1,3-бутандиоле. Носитель может представлять растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и тому подобное), их подходящие смеси, растительные масла, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, в качестве растворителя или суспендирующей среды обычно применяют нелетучие жирные масла. Для данной цели можно использовать любое чистое, нелетучее жирное масло, включая синтетические моно- или диглицериды. В дополнение, при получении инъекционных препаратов находят свое применение жирные кислоты, такие как олеиновая кислота.
Во всех случаях форма должна быть стерильной и должна быть жидкой до такой степени, чтобы она легко набиралась шприцем. Необходимую текучесть можно сохранить, например, при использовании оболочки, такой как лецитин, при сохранении требуемого размера частиц в случае дисперсии и при использовании поверхностно-активного вещества. Форма должна быть стабильной в условиях производства и хранения и должна быть устойчивой к загрязняющему воздействию микроорганизмов, таких как бактерии и грибы. Предупредить действие микроорганизмов можно при использовании различных антибактериальных и противогрибковых препаратов, например парабенов, хлорбутанола, фенола, сорбиновой кислоты, тримерозала и тому подобное. Во многих случаях будет желательным включить изотонические вещества, например сахара или хлорид натрия. Пролонгированного всасывания инъекционных композиций можно достичь при использовании в композициях веществ, обеспечивающих замедленное всасывание, например, моностеарата алюминия и желатина.
Композиции, пригодные для введения, можно готовить вместе с веществами, усиливающими поглощение или всасывание, с целью повышения их эффективности. Такие усилители включают, например, салицилат, гликохолат/линолеат, глихолат, апротинин, бацитрацин, SDS, капрат и тому подобное. Смотри, например, Fix (J. Pharm. Sci., 85: 1282-1285, 1996) и Oliyai and Stella (Ann. Rev. Pharmacol. Toxicol., 32: 521-544, 1993).
Кроме того, предполагается баланс гидрофильных и гидрофобных свойств композиций, предусмотренных для использования в изобретении, посредством чего повышается их применимость в условиях in vitro и особенно в условиях in vivo, в то время как иные композиции с отсутствием такого баланса имеют меньшую применимость. В частности, композиции, предусмотренные для применения в изобретении, обладают соответствующей степенью растворимости в водной среде, что позволяет им всасываться и быть биодоступными в организме, в то же время они в определенной степени растворяются в липидах, что позволяет соединениям проникать через клеточную мембрану к предполагаемому месту действия. Таким образом, предусматриваемые композиции являются максимально эффективными, когда их можно доставить в место проявления активности антигена-мишени.
В одном аспекте способы по изобретению включают стадию введения композиции связывающей молекулы.
Способы по изобретению осуществляют с использованием любых медицинских приемлемых средств для введения терапевтического препарата прямо или опосредованно отдельному млекопитающему, включая, но не ограничиваясь этим, прием внутрь, интраназальное, местное, трансдермальное, парентеральное, ингаляционное с помощью спрея, интравагинальное или ректальное введение. Термин парентеральное, в том смысле, в котором он здесь используется, включает подкожную, внутривенную, внутримышечную и внутриполостную инъекции, а также введение с помощью катетера и инфузии. Также предусматривается внутрикожная инъекция, инъекция в молочную железу, внутрибрюшинная, внутриоболочечная, ретробульбарная, внутрилегочная инъекция или хирургическая имплантация в определенное место.
В одном варианте осуществления проводят введение в место локализации злокачественной опухоли или пораженную ткань, когда требуется лечение прямой инъекцией в место поражения посредством замедленной доставки или с помощью механизма замедленной доставки, которые приводят к доставке композиции внутрь. Например, биодеградируемые микросферы или капсулы, или другие конструкции из биодеградируемых полимеров, которые способны замедленно высвобождать композицию (например, растворимый полипептид, антитело или малую молекулу), можно включить в композиции по изобретению, имплантированные около места локализации злокачественной опухоли.
Терапевтические композиции также можно доставлять пациенту во многие места. Множественное введение можно проводить одновременно или вводить в течение определенного периода времени. В некоторых случаях полезно обеспечить непрерывный поток терапевтической композиции. Можно проводить дополнительную терапию на периодичной основе, например, выражающейся в часах, сутках, неделях или месяцах.
Композиции связывающих молекул по изобретению могут содержать один вид или могут содержать более чем один вид связывающих молекул. Также в настоящем изобретении предусматривается введение композиций связывающих молекул в сочетании со вторым средством. Вторые средства, предусмотренные для применения в изобретении, приведены в абзацах ниже.
Второе средство может представлять связанную с В-клетками молекулу. Другие связанные с В-клетками молекулы, предусмотренные для применения в изобретении, включают связывающие молекулы, которые связываются с поверхностными молекулами В-клеток, которые не являются CD37 или CD20. Связанные с В-клетками молекулы включают, но не ограничиваются этим, CD19 (антиген В-лимфоцитов CD19, также называемый поверхностным антигеном В-лимфоцитов В4 или Leu-12), CD21, CD22 (рецептор CD22 В-клеток, также называемый Leu-14, молекулой клеточной адгезии В-лимфоцитов или BL-CAM), CD23, CD40 (поверхностный антиген В-клеток CD40, также называемый членом 5 надсемейства рецепторов фактора некроза опухолей, рецептор CD40L или Вр50), CD80 (антиген активации Т-лимфоцитов CD80, также называемый антигеном активации В7-1, В7, В7-1 или ВВ1), CD86 (антиген активации Т-лимфоцитов CD86, также называемый антигеном активации В7-2, В70, FUN-1 или BU63), CD137 (также называемый членом 9 надсемейства рецепторов фактора некроза опухолей), CD152 (также называемый цитотоксическим белком 4 Т-лимфоцитов или CTLA-4), L6 (ассоциированный с опухолями антиген L6, также называемый членом 1 трансмембранного надсемейства 4, поверхностным маркером 1 мембранного компонента или M3S1), CD30 (антиген активации лимфоцитов CD30, также называемый членом 8 надсемейства рецепторов фактора некроза опухолей, рецептором CD30L или Ki-1), CD50 (также называемый молекулой-3 межклеточной адгезии (ICAM3) или ICAM-R), CD54 (также называемый молекулой-1 межклеточной адгезии (ICAM1) или рецептором главной группы риновируса), В7-Н1 (лиганд иммуноингибирующего рецептора, экспрессируемый активированными Т-клетками, В-клетками и миелоидными клетками, также называемый PD-L1; смотри Dong et al., «B7-H1, a third member of B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion», Nat. Med., 5: 1365-1369 (1999)), CD134 (также называемый членом 4 надсемейства рецепторов фактора некроза опухолей, OX40, рецептором OX40L, антигеном АСТ35 или рецептором активированного транскрипцией ТАХ гликопротеина 1), 41ВВ (рецептор лиганда 4-1ВВ, антиген Т-клеток 4-1ВВ или антиген Т-клеток ILA), CD153 (также называемый членом 8 надсемейства рецепторов фактора некроза опухолей, лигандом CD30 или CD 30-L), CD154 (также называемый членом 5 надсемейства рецепторов фактора некроза опухолей, белком активации, ассоциированным с TNF, TRAP или антигеном Т-клеток Gp39) и рецепторы Toll. Приведенный выше перечень конструкций-мишеней и/или антигенов-мишеней является только примерным и не претендует на то, чтобы быть исчерпывающим.
Цитокины и факторы роста представляют собой вторые средства, предусмотренные изобретением, и они включают, без ограничения, одно или более из TNF, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-13, IL-14, IL-15, IL-16, IL-17, IL-18, IFN, G-CSF, Meg-CSF, GM-CSF, тромбопоэтин, фактор стволовых клеток и эритропоэтин. Фармацевтические композиции по изобретению могут также включать другие известные ангиопоэтины, например Ang-1, Ang-2, Ang-4, Ang-Y и/или человеческий ангиопоэтин-подобный полипептид и/или васкулярный эндотелиальный фактор роста (VEGF). Факторы роста для применения в фармацевтических композициях по изобретению включают ангиогенин, костный морфогенный белок-1, костный морфогенный белок-2, костный морфогенный белок-3, костный морфогенный белок-4, костный морфогенный белок-5, костный морфогенный белок-6, костный морфогенный белок-7, костный морфогенный белок-8, костный морфогенный белок-9, костный морфогенный белок-10, костный морфогенный белок-11, костный морфогенный белок-12, костный морфогенный белок-13, костный морфогенный белок-14, костный морфогенный белок-15, рецептор IA костного морфогенного белка, рецептор IВ костного морфогенного белка, нейротрофический фактор мозга, цилиарный нейротрофический фактор роста, рецептор α цилиарного нейротрофического фактора роста, индуцированный цитокинами хемотаксический фактор 1 нейтрофилов, индуцированный цитокинами хемотаксический фактор 2α нейтрофилов, индуцированный цитокинами хемотаксический фактор 2β нейтрофилов, эндотелиальный клеточный фактор роста β, эндотелин 1, эпидермальный фактор роста, эпителиальный аттрактант нейтрофилов, фактор роста фибробластов 4, фактор роста фибробластов 5, фактор роста фибробластов 6, фактор роста фибробластов 7, фактор роста фибробластов 8, фактор роста фибробластов 8b, фактор роста фибробластов 8с, фактор роста фибробластов 9, фактор роста фибробластов 10, кислый фактор роста фибробластов, основный фактор роста фибробластов, рецептор нейротрофического фактора роста α1 глиальной клеточной линии, рецептор нейротрофического фактора роста α2 глиальной клеточной линии, связанный с ростом белок, связанный с ростом белок α, связанный с ростом белок β, связанный с ростом белок γ, связывающий гепарин эпидермальный фактор роста, фактор роста гепатоцитов, рецептор фактора роста гепатоцитов, инсулиноподобный фактор роста I, рецептор инсулиноподобного фактора роста, инсулиноподобный фактор роста II, связывающий белок инсулиноподобного фактора роста, фактор роста кератиноцитов, ингибирующий фактор лейкемии, рецептор α ингибирующего фактора лейкемии, фактор роста нервов, рецептор фактора роста нервов, нейротрофин-3, нейротрофин-4, плацентарный фактор роста, плацентарный фактор роста 2, эндотелиальный клеточный фактор роста тромбоцитов, фактор роста тромбоцитов, цепь А фактора роста тромбоцитов, фактор роста АА тромбоцитов, фактор роста АВ тромбоцитов, цепь В фактора роста тромбоцитов, фактор роста ВВ тромбоцитов, рецептор α фактора роста тромбоцитов, рецептор β фактора роста тромбоцитов, стимулирующий фактор роста предшественников В-клеток, фактор роста стволовых клеток, рецептор фактора роста стволовых клеток, трансформирующий фактор роста α, трансформирующий фактор роста β, трансформирующий фактор роста β1, трансформирующий фактор роста β1.2, трансформирующий фактор роста β2, трансформирующий фактор роста β3, трансформирующий фактор роста β5, поздний трансформирующий фактор роста β1, связывающий белок I трансформирующего фактора роста β, связывающий белок II трансформирующего фактора роста β, связывающий белок III трансформирующего фактора роста β, рецептор типа I фактора некроза опухолей, рецептор типа II фактора некроза опухолей, рецептор активатора плазминогена урокиназного типа, васкулярный эндотелиальный фактор роста и химерные белки и их биологически или иммунологически активные фрагменты.
Примеры химиотерапевтических препаратов, предусмотренных в качестве вторых средств, включают, но не ограничиваются этим, алкилирующие агенты, такие как азотистые иприты (например, меклоретамин, циклофосфамид, ифосфамид, мелфалан и хлорамбуцил); нитрозомочевины (например, кармустин (BCNU), ломустин (СCNU) и семустин (метил-СCNU)); этиленимины и метилмеламины (например, триэтиленмеламин (ТЕМ), триэтилентиофосфорамид (тиотепа) и гексаметилмеламин (НММ, алтретамин)); алкилсульфонаты (например, буслфан) и триазины (например, дакабазин (DTIC)); антиметаболиты, такие как аналоги фолиевой кислоты (например, метотрексат, триметрексат и пеметрексат (антифолат с множественными мишенями)); аналоги пиримидинов (такие как 5-фторурацил (5-FU), фтордезоксиуридин, гемцитабин, цитозин арабинозид (АраС, цитарабин), 5-азацитидин и 2,2'-дифтордезоксицитидин); и аналоги пуринов (например, 6-меркаптопурин, 6-тиоуанин, азатиоприн, 2'-дезоксикоформицин (пентостатин), эритрогидроксинониладенин (EHNA), флударабин фосфат, 2-хлордезоксиаденозин (кладрибин, 2-CdA)); ингибиторы топоизомеразы типа I, такие как камптотецин (СРТ), топотекан и иринотекан; природные продукты, такие как эпиподофилотоксины (например, этопозид и тенипозид); и алкалоиды растений барвинок розовый (например, винбластин, винкристин и винорелбин); противоопухолевые антибиотики, такие как актиномицин D, доксорубицин и блеомицин; радиосенсибилизаторы, такие как 5-бромдезоксиуридин, 5-йододезоксиуридин и бромдезоксицитидин; координационные комплексы платины, такие как цисплатин, карбоплатин и оксалиплатин; замещенные мочевины, такие как гидроксимочевина; и производные метилгидразина, такие как N-метилгидразин (MIH) и прокарбазин.
Не ограничивающие примеры химиотерапевтических препаратов, радиоактивных терапевтических препаратов и других активных и вспомогательных средств также приведены в таблице 1.
(антифолат для многих мишеней)
(AraC, цитарабин)
(пентостатин)
(кладрибин, 2-СdА)
антагонисты
ацетат
Вторые средства, предусмотренные изобретением для лечения аутоиммунных заболеваний, включают иммуносупрессоры, которые функционируют с подавлением или «маскированием» иммунной системы индивидуума, который подвергается лечению. Иммуносупрессоры включают, например, нестероидные противовоспалительные препараты (НПВС), анальгетики, глюкокортикоиды, противоревматические препараты, модифицирующие заболевание (DMARD), для лечения артрита или модификаторы биологических ответных реакций. Композиции при описании DMARD также являются пригодными для лечения многих других аутоиммунных заболеваний помимо RA.
Примеры НПВС выбраны из группы, состоящей из ибупрофена, напроксена, натриевой соли напроксена, ингибиторов Сох-2, таких как Vioxx и Celebrex, и сиалилатов. Примерные анальгетики выбраны из группы, состоящей из ацетаминофена, оксикодона, трамадола или пропоксифена гидрохлорида. Примерные глюкокортикоиды выбраны из группы, состоящей из кортизона, дексаметазона, гидрокортизона, метилпреднизолона, преднизолона или преднизона. Примеры модификаторов биологических ответных реакций включают, но не ограничиваются этим, молекулы с направленным действием против клеточных поверхностных маркеров (например, CD4, CD5 и т.д.), ингибиторы цитокинов, такие как антагонисты TNF (например, этанерцепт (Enbrel)), адалимумаб (Humira) и инфликсимаб (Remicade), ингибиторы хемокинов и ингибиторы молекул адгезии. Модификаторы биологических ответных реакций включают моноклональные антитела, а также рекомбинантные формы молекул. Примерные DMARD включают, но не ограничиваются этим, азатиоприн, циклофосфамид, циклоспорин, метотрексат, пеницилламин, лефлуномид, сульфасалазин, гидроксихлорхин, золото [для перорального (ауранофин) и внутримышечного введения] и миноциклин.
Предполагается, что композицию связывающей молекулы и второе средство можно вводить одновременно в одной лекарственной форме. Альтернативно препараты можно вводить в отдельных лекарственных формах, но одновременно, при том что одновременно означает, что препараты вводят в течение 30 мин по отношению друг к другу.
В другом аспекте второе средство вводят перед введением композиции связывающей молекулы. "Перед введением" означает, что второе средство вводят в период, исчисляемый от одной недели перед лечением антителом до 30 мин перед введением антитела. Также предусматривается, что второе средство вводят после введения композиции связывающей молекулы. Последующее введение означает, что введение проводят в период от 30 мин после лечения антителом до одной недели после введения антитела.
Дополнительно предусматривается, что, когда связывающую молекулу вводят в комбинации со вторым средством, где второе средство представляет цитокин или фактор роста, или химиотерапевтический препарат, введение также может включать применение радиоактивного терапевтического препарата или лучевой терапии. Лучевую терапию, назначенную в комбинации с композицией антитела, проводят согласно указаниям лечащего врача и в дозах, как правило, назначаемых для лечения пациентов со злокачественными опухолями.
Количества связывающей молекулы в данной дозе будут варьировать в зависимости от размеров индивидуума, которому проводят терапию, а также характеристик заболевания, которое подвергается лечению. В примерах лечения может быть необходимым вводить примерно 1 мг/сутки, примерно 5 мг/сутки, примерно 10 мг/сутки, примерно 20 мг/сутки, примерно 50 мг/сутки, примерно 75 мг/сутки, примерно 100 мг/сутки, примерно 150 мг/сутки, примерно 200 мг/сутки, примерно 250 мг/сутки, примерно 500 мг/сутки или примерно 1000 мг/сутки. Можно также вводить дозы, основываясь на массе пациента, в дозе от примерно 0,01 до примерно 50 мг/кг. В близком варианте осуществления связывающую молекулу можно вводить в дозе в пределах от примерно 0,015 до примерно 30 мг/кг. В дополнительном варианте осуществления связывающую молекулу вводят в дозе примерно 0,015, примерно 0,05, примерно 0,15, примерно 0,5, примерно 1,5, примерно 5, примерно 15 или примерно 30 мг/кг.
Данные композиции можно вводить в одной дозе или во многих дозах. В обычных исследованиях зависимости доза-эффект, вначале проводимых на моделях животных, а затем в клинических испытаниях, устанавливают оптимальные дозы для конкретных заболеваний и групп пациентов.
Введение композиции связывающей молекулы снижает популяцию В-клеток, по меньшей мере, на 20% после одной дозы лечения. В одном варианте осуществления популяция В-клеток снижается, по меньшей мере, примерно на 20, примерно на 30, примерно на 40, примерно на 50, примерно на 60, примерно на 70, примерно на 80, примерно на 90 или примерно на 100%. Снижение количества В-клеток определяется как уменьшение абсолютного количества В-клеток ниже нижнего предела нормы. Восстановление В-клеток определяется как восстановление абсолютного количества В-клеток до одного из следующих значений: 70% от нормального значения или нормальных пределов у субъекта.
Введение CD20-специфических связывающих молекул также приводит к усиленному апоптозу определенных субпопуляций В-клеток. Апоптоз означает индукцию запрограммированной гибели клеток, выражаемой и оцениваемой по фрагментации ДНК, сморщиванию клеток, образованию мембранных везикул или изменению липидного состава мембран, что определяют окрашиванием аннексином V.
Кроме того, введение композиций связывающих молекул по изобретению приводит к желаемым клиническим эффектам в отношении заболевания или нарушения, которое подвергается лечению. Например, у пациентов с ревматоидным артритом, в одном аспекте, введение приводит к улучшению состояния пациента, оцениваемому в клинических баллах [например, достигает показателя American College of Rheumatology Preliminary Detection of Improvement (ACR20)] и/или улучшению на 20% состояния связок и отечного сустава и улучшению на 20% в 3/5 случаях по остальным ACR (Felson et al., Arthritis Rheum., 1995, 38: 828-35). Биологические показатели улучшения состояния пациентов с RA после введения CD37-специфических и CD20-специфических связывающих молекул включают оценку изменений концентрации цитокинов по определению уровня белка или РНК. Интересующие цитокины включают, но не ограничиваются этим, TNF-α, IL-1, интерфероны, Blys и APRIL. Изменения цитокинов могут быть результатом снижения количества В-клеток или уменьшения количества активированных Т-клеток. У пациентов с RA определяют соответствующие маркеры обмена в костной ткани (резобция или эрозия костей) до и после введения CD20-специфических связывающих молекул. Соответствующие маркеры включают, но не ограничиваются этим, щелочную фосфатазу, остеокальцин, фрагменты разрушения коллагена, оксипролин, кислую фосфатазу, устойчивую к тартрату, и лиганд RANK (RANKL). Другие показатели, связанные с улучшением состояния пациентов с RA, включают определение концентрации С-реактивного белка (CRP), скорости оседания эритроцитов (СОЭ), ревматоидного фактора, антител против ССР (циклический, содержащий цитруллин пептид) и оценку количества В-клеток в крови и количества лимфоцитов проточной цитометрией. Можно также определить специфические факторы в синовиальной жидкости у пациентов с RA, включая оценку уровня В-клеток в синовиальной жидкости при ее биопсии, уровней RANKL и других факторов костной ткани и цитокинов, приведенных выше.
В близком аспекте эффекты комбинированного введения в отношении других заболеваний оценивают по показателям, известным в данной области. Например, предусматривается, что у пациентов с болезнью Крона после лечения по изобретению имеет место улучшение значения индекса активности болезни Крона (CDAI) в пределах от примерно 50 до примерно 70 единиц, в то время как ремиссия оценивается 150 единицами (Simonis et al., Scand. J. Gastroent., 1998, 33: 283-8). Число баллов 150 или 200 означает норму, в то время как значение 450 соответствует тяжелой форме заболевания. Также желательно, чтобы введение CD37-специфических и CD20-специфических связывающих молекул приводило к снижению уровня перинуклеарных антител против нейтрофилов (pANCA) и антител против Saccaromyces cervisiae (ASCA) у индивидуумов с воспалительным заболеванием кишечника.
Дополнительно предполагается, что у взрослых пациентов и пациентов юношеского возраста с миозитом, подвергшихся лечению по изобретению, наступает улучшение основных показателей при оценке, например, для 3 из 6, что означает улучшение примерно на 20%, при оценке не более чем 2 улучшение оценивается примерно на 25% по основным показателям (смотри Rider et al., Arthritis Rheum., 2004, 50: 2281-90).
Дополнительно предусматривается, что у пациентов с SLE, подвергшихся лечению по изобретению, наступает улучшение по шкале оценки активности системной красной волчанки (SLAM) или индексу активности системной красной волчанки (SLEDAI), по меньшей мере, на 1 значение (Gladman et al., J. Rheumatol. 1994, 21: 1468-71) (Tan et al., Arthritis Rheum., 1982, 25: 1271-7). Число баллов по SLAM >5 или по SLEDAI >2 рассматривается в качестве показателя клинически активного заболевания. Ответную реакцию на лечение можно трактовать в виде улучшения или стабилизации по 2 показателям активности заболевания (индексу активности системной красной волчанки [SLEDAI] и шкале активности системной красной волчанки и по 2 показателям качества жизни (оценка пациента в целом и шкала Krupp Fatigue Severity Scale) (Petri et al., Arthritis Rheum., 2004, 50: 2858-68). Также предусматривается, что введение связывающей молекулы пациентам с SLE приводит к снижению уровня антител против двухцепочечной ДНК. Альтернативно об улучшении можно судить по критериям оценочной системы British Assessment Group Criteria (BILAG).
Также предусматривается, что у пациентов с рассеянным склерозом, подвергшихся лечению по изобретению, наступает улучшение по значению клинических баллов оценочной шкалы Kurtzke Expanded Disability (EDSS) (Kurtzke, F., Neurology 1983, 33: 1444-52), по меньшей мере, на 0,5 или замедление клинического прогрессирования заболевания, по меньшей мере, на 1,0 по шкале Kurtzke (Rudick et al., Neurology 1997, 49: 358-63).
Дополнительно предполагается, что у пациентов с IIM, получающих CD37-специфические и CD20-специфические связывающие молекулы, имеет место снижение одного из пяти критериев по оценочной системе Idiopathic Inflammatory Myopathy Criteria (IIMC) (Miller F., выше). Также предусматривается, что введение пациентам с IIM приводит к снижению связанных с IIM факторов, выбранных из группы, состоящей из креатинкиназы (СК), лактатдегидрогеназы, альдолазы, С-реактивного белка, аспартатаминотрансферазы (АСТ), аланинаминотрансферазы (АЛТ) и антинуклерного аутоантитела (ANA), специфических для миозита антител (MSA) и антител против экстрагируемых ядерных антигенов. Альтернативно, у пациентов имеется 3 из 6 критериев, приведенных Rider et al., Arthritis Rheum., 50(7): 2281-2290 (2004), без ухудшения не более чем по 2 критериям.
В некоторых вариантах осуществления у пациентов с В-клеточной злокачественной опухолью, получающих лечение по изобретению, имеет место положительная в целом реакция на лечение при ее оценке по клиническим показателям, хорошо известным и обычно используемым в данной области, и описанным ниже, таким как уменьшение размеров опухолей, снижение числа опухолей и/или ослабление симптомов заболевания.
Примерные клинические критерии приведены Национальным институтом рака (NCI), который подразделяет некоторые виды группы злокачественных опухолей на клинические категории «медленно текущие» и «агрессивные» лимфомы. «Медленно текущие» лимфомы включают фолликулярные клеточные лимфомы, подразделяемые по их цитологии на «категории», диффузную мелкоклеточную лимфоцитарную лимфому/хроническую лифоцитарную лейкемию (CLL), лимфоплазмоцитоидную/макроглобулинемию Вальденстрема, лимфому маргинальной зоны и волосистоклеточную лейкемию. «Агрессивные» лимфомы включают диффузную смешанную и крупноклеточную лимфому, лимфому Беркитта/диффузную мелкоклеточную лимфому, лимфобластную лимфому, покровноклеточную лимфому и связанную со СПИДом лимфому. В некоторых случаях применяют международный прогностический индекс (IPI) для агрессивной и фолликулярной лимфомы. Факторы, которые следует учитывать при использовании IPI, включают возраст (возраст <60 лет против возраста >60 лет), активность лактатдегидрогеназы в сыворотке крови (нормальные уровни против повышенных), состояние активности (0 или 1 против 2-4) (смотри определение ниже), стадию заболевания (I или II против III или IV) и поражение областей помимо лимфатических узлов (0 или 1 против 2-4). Пациенты с наличием 2 или более факторов риска имеют 50%-ный шанс отсутствия рецидивов или выживания в целом в течение 5 лет.
Состояние активности IPI при агрессивной лимфоме определяют следующим образом: описание категории: 0 - полностью активен, способен к активности, сходной с таковой до начала заболевания, без ограничений; 1 - ограничен в физической, требующей усилий активности, но подвижен и способен выполнять легкую или сидячую, например, легкую домашнюю работу, работу в офисе; 2 - подвижен и способен ухаживать за собой, но не способен к какой-либо деятельности, до и примерно более чем 50% времени бодрствует; 3 - способен только к ограниченному уходу за собой, в 50% времени бодрствования проводит в постели или кресле; 4 - полностью беспомощен, не способен ухаживать за собой, полностью ограничен постелью или креслом; и 5 - смерть. (Смотри The International Non-Hodgkin's Lymphoma Prognostic Factors Project. A predictive model for aggressive non-Hodgkin's lymphoma. N. Engl. J. Med. 329: 987-94, 1993.)
Как правило, категорию лимфомы клинически оценивают с использованием положения о том, что лимфомы низкой категории, как правило, проявляются в виде заболевания лимфатических узлов и часто являются вялотекущими или медленно растущими. Обычно лимфомы промежуточной или высокой категории протекают в виде значительно более агрессивного заболевания с большой опухолевой нагрузкой помимо лимфатических узлов.
Классификационная система Ann Arbor также используется для оценки прогрессирования опухолей, в частности, не-ходжкинских лимфом. Согласно данной системе лимфомы стадий I, II, III и IV у взрослых пациентов с NHL можно классифицировать в категории А и B в зависимости от того, имеются ли у пациента хорошо выраженные, генерализованные симптомы (В) или нет (А). Обозначение В дается пациентам со следующими симптомами: не поддающаяся объяснению потеря более чем 10% массы тела в течение 6 месяцев до постановки диагноза, необъяснимая лихорадка с повышением температуры тела выше 38°С и интенсивное потоотделение ночью. Определение стадий является следующим: стадия I - поражение одного лимфатического узла или локализованное поражение одного органа или области вне лимфатического узла. Стадия II - поражение двух или более лимфатических узлов с одной стороны диафрагмы или локализованное поражение одного органа или области вне лимфатического узла и его региональных лимфатических узлов с или без других лимфатических узлов с одной стороны диафрагмы. Стадия III - поражение лимфатических узлов с обеих сторон диафрагмы, возможно сопровождаемое локализованным поражением органа или области вне лимфатических узлов, поражением селезенки или обоими видами поражений. Стадия IV - диссеминированное (мультифокальное) поражение одной или более областей вне лимфатических узлов с или без ассоциированного поражения лимфатических узлов или отдельное поражение органа вне лимфатических узлов с поражением отдаленных (нерегиональных) лимфатических узлов. Дополнительные детали смотри в "The International Non-Hodgkin's Lymphoma Prognostic Factors Project: A predictive model for aggressive non-Hodgkin's lymphoma", New England J. Med., (1993) 329: 987-994.
В одном аспекте терапевтический эффект способов по изобретению определяется по уровню ответной реакции, например частичная ответная реакция определяется как уменьшение опухоли до размера, составляющего менее половины от ее первоначального размера. Полная реакция определяется как полная элиминация заболевания, подтвержденная при клиническом или радиологическом обследовании. В одном варианте осуществления у индивидуума, подвергшегося лечению по изобретению, имеется, по меньшей мере, частичная ответная реакция на лечение.
Согласно критериям Cheson, используемым для оценки NHL, разработанным совместно с Национальным институтом рака (Cheson et al., J. Clin. Oncol. 1999, 17: 1244; Grillo-Lopez et al., Ann. Oncol. 2000, 11: 399-408), полную ответную реакцию получают, когда имеется полное исчезновение всех детектируемых клинических признаков, признаков заболевания, обнаруживаемых радиографическими методами, и связанных с заболеванием симптомов, все лимфатические узлы возвращаются к своему нормальному размеру, селезенка уменьшается в размере и в костном мозге отсутствует лимфома.
Неподтвержденную полную ответную реакцию получают, когда у пациента наблюдают полное исчезновение заболевания и уменьшение размера селезенки, и лимфатические узлы подвергаются регрессии более чем на 75%, и не определено состояние костного мозга. Неподтвержденная полная ответная реакция соответствует и превышает критерии для частичной ответной реакции. В целом, реакцию оценивают как снижение опухолевой нагрузки, по меньшей мере, на 50%.
Аналогичные критерии были разработаны для различных других форм злокачественных заболеваний или гиперпролиферативных заболеваний, и их могут легко найти специалисты в данной области. Смотри, например, Cheson et al., Clin. Adv. Hematol. Oncol. 2006, 4: 4-5, где описываются критерии для оценки CLL; Cheson et al., J. Clin. Oncol. 2003, 21: 4642-9, где описываются критерии для оценки АМL; Cheson et al., Blood 2000, 96: 3671-4, где описываются критерии для оценки миелодиспластических синдромов.
В другом аспекте ответная реакция на лечение у пациентов с В-клеточным злокачественным заболеванием выражается в виде замедления прогрессирования заболевания по сравнению с пациентами, не получающими терапию. Определение замедленного прогрессирования заболевания или любого из указанных выше факторов можно провести с использованием методов, хорошо известных в данной области, включая сканирование костей, СТ-сканирование, сканирование с использованием галлия, лимфангиограмму, MRI, PET-сканирование, ультразвуковое исследование и тому подобное.
Также очевидно понятно, что схему лечения можно изменять, если применяются традиционные лекарственные препараты в комбинации с препаратами по изобретению.
В качестве дополнительного аспекта изобретение включает наборы, которые содержат одно или более соединений или композиций, пригодных для способов по изобретению, упакованных таким образом, чтобы облегчить их применение в практике способов по изобретению. В самом простом варианте осуществления такой набор содержит соединение или композицию, описанные здесь, в качестве пригодных для практического применения способа по изобретению, упакованный в контейнере, таком как запаянная бутыль или резервуар, с этикеткой, прикрепленной к контейнеру или вложенной в упаковку, на которой описано применение соединения или композиции для практического использования способа по изобретению. Предпочтительно соединение или композиция помещены в разовую лекарственную форму. Набор может дополнительно содержать устройство, подходящее для введения композиции предпочтительным путем введения или для постановки скринингового теста. Набор может включать этикетку, на которой описано применение композиции(й) связывающей молекулы в способе по изобретению.
Настоящее изобретение также включает промышленные изделия. Такие изделия включают CD37-специфические связывающие молекулы и CD20-специфические связывающие молекулы, необязательно вместе с фармацевтическим носителем или разбавителем и, по меньшей мере, одну этикетку, на которой описан способ применения связывающих молекул по изобретению. Такие промышленные изделия могут необязательно включать, по меньшей мере, одно второе средство для введения вместе со связывающими молекулами.
Настоящее изобретение также относится к применению композиции, содержащей CD37-специфическую связывающую молекулу или CD37-специфическую и CD20-специфическую связывающие молекулы в производстве лекарственного препарата для лечения или профилактики заболевания, связанного с аномальной активностью В-клеток.
Краткое описание чертежей
На фигуре 1А схематично приведена структура молекулы TRU-016; на фигуре 1В представлены результаты анализа SDS-PAGE, которые показывают, что экспрессированный белок мигрирует соответственно молекулярной массе, равной примерно 110 кДа в невосстанавливающих условиях и примерно 52 кДа в восстанавливающих условиях; и на фигуре 1С показано, что молекула TRU-016 проявляет высокий уровень специфического связывания с В-клетками периферической крови человека и существенно более низкий уровень связывания с другими клеточными субпопуляциями среди клеток, отличных от В-клеток (CD19 негативной популяции), при анализе проточной цитометрией.
На фигуре 2А-Е показано ингибирование связывания под действием различных реагентов с направленным действием против CD37.
На фигуре 3А показано связывание FITC C1q с молекулами TRU-016 при инкубации с В-клетками Ramos в нормальной сыворотке человека с и без фактора из яда кобры (CVF); на фигуре 3В показана CDC активность молекул TRU-016 при инкубации с В-клетками Ramos в нормальной сыворотке человека с и без CVF; и на фигуре 3С показана CDC активность молекул TRU-016 при инкубации с В-клетками Ramos и человеческим или кроличьим комплементом.
На фигуре 4 показаны хроматограммы гель-фильтрации (SEC) при высоком давлении (ВЭЖХ), полученные при GPC очистке TRU-016, при построении графика зависимости поглощения против времени удерживания для различных собранных фракций.
На фигуре 5А показаны связывающие свойства фракций, полученных при SEC; на фигуре 5В приведены данные по CDC активности фракций, полученных при SEC; и на фигуре 5С показана ADCC активность фракций, полученных при SEC.
На фигуре 6 приведены данные по CDC активности TRU-015, ритуксана, TRU-016 или их комбинаций на В-клетках Ramos.
На фигуре 7 показано влияние TRU-016 на CDC активность TRU-015 на В-клетках DHL-4.
На фигуре 8 показано влияние TRU-016 на CDC активность TRU-015 и ритуксана.
На фигуре 9 показано влияние TRU-016 на TRU-015 в тесте определения CDC активности.
На фигуре 10 показано влияние TRU-016 на ритуксан в тесте определения CDC активности.
На фигуре 11 показано взаимодействие TRU-015 и TRU-016 в тесте определения ADCC активности с использованием клеток BJAB.
На фигуре 12 показано взаимодействие TRU-015 и TRU-016 в тесте определения ADCC активности с использованием клеток Daudi.
На фигуре 13 показано взаимодействие TRU-015 и TRU-016 в тесте определения ADCC активности с использованием клеток Ramos.
На фигуре 14 показано влияние ритуксана, TRU-016 и их комбинации на специфическую гибель клеток BJAB.
На фигуре 15 показано влияние ритуксана, TRU-016 и их комбинации на специфическую гибель клеток BJAB.
На фигуре 16 показано влияние TRU-015, TRU-016 и их комбинации на специфическую гибель клеток BJAB.
На фигуре 17 показано влияние TRU-015, TRU-016 и их комбинации на специфическую гибель клеток BJAB.
На фигурах 18А-D показано, что димеры TRU-016 не опосредуют одну CDC, но усиливают CDC активность ритуксимаба в условиях in vitro.
На фигурах 19А-В показано, что очищенный на протеине А TRU-016 индуцирует апоптоз клеток Ramos и Daudi, в то время как для димеров необходимо перекрестное связывание.
На фигуре 20 показано, что TRU-016 предпочтительно снижает количество нормальных В-клеток из культур РВМС.
На фигуре 21 показана эффективность TRU-016 по сравнению с huIgG, ритуксаном и комбинированной обработкой TRU-016 и ритуксана в отношении объема опухолей у животных.
На фигурах 22А-В показано, что димеры TRU-016 проявляют высокую противоопухолевую активность по данным определения объема опухолей и процента выживаемости на модели ксенотрансплантатов опухолей.
На фигуре 23 показано, что димеры TRU-016 не усиливают CDC активность по результатам обработки специфическими реагентами для MHСII, CD19, CD80/86 или CD45.
На фигуре 24 представлены данные по проценту выживаемости мышей с опухолями Ramos (период наблюдений до 90 суток) после обработки TRU-016, ритуксимабом и их комбинацией.
На фигурах 25 и 26 представлен процент гибели мышей с опухолями Daudi (период наблюдений до 90 суток) после обработки TRU-016 или ритуксимабом.
На фигуре 27 показано, что TRU-016 эффективно снижал относительную жизнеспособность клеток, обработанных флударабином, тем самым потенцируя цитотоксическое действие одного флударабина.
На фигуре 28 показано, что TRU-016 индуцировал более высокую токсичность для клеток по сравнению с герцептином или ритуксимабом на резистентных к ритуксимабу клеточных линиях.
На фигуре 29 показано, что TRU-016 индуцировал фосфорилирование тирозина в CD19+ В-клетках первичной CLL.
На фигуре 30А приведена консенсусная аминокислотная последовательность гуманизированной конструкции TRU-016 No.019001 (SEQ ID NO:6) и TRU-016 (SEQ ID NO:2) по системе нумерации Kabat; на фигуре 30В показаны сопоставления аминокислотных последовательностей трех гуманизированных конструкций TRU-016 (019001, 019008 и 109009).
На фигуре 31 приведены сопоставления ДНК- и аминокислотных последовательностей трех гуманизированных конструкций TRU-016 (019001, 019041 и 019044).
На фигуре 32 представлены сопоставления последовательностей в формате FASTA тех же трех гуманизированных конструкций TRU-016 (019001, 019041 и 019044).
Примеры
Дополнительные аспекты и подробные детали изобретения станут очевидными из последующих примеров, которые предназначаются для иллюстрации, а не ограничения. В примере 1 описано получение CD37-специфической связывающей молекулы; в примере 2 показано, что TRU-016 и различные CD37-специфические антитела распознают одни и те же или перекрывающиеся эпитопы; в примере 3 показано, что TRU-016 мало активен в связывании C1q и активировании классического пути активации комплемента; в примере 4 показана активность и связывание мультимеров TRU-016; в примере 5 описано получение CD20-специфической связывающей молекулы; в примере 6 показано, что комбинации TRU-016 с TRU-015 или ритуксаном синергически усиливают апоптоз В-клеток; в примере 7 показано, что комбинации TRU-016 с CD20-специфическими антителами или SMIP синергически усиливают CDC; в примере 8 показано, что TRU-016 усиливает ADCC и CDC активность CD20-специфических антител и SMIP; в примере 9 показано, что TRU-016 индуцирует апоптоз В-клеток; в примере 10 показано, что комбинации CD37-специфических SMIP с CD20-специфическим антителом синергически уменьшают объем опухолей на модели опухолевых ксенотрансплантатов на мышах; в примере 11 показано, что CD37-специфические SMIP одни также уменьшают объем опухолей на модели опухолевых ксенотрансплантатов на мышах; в примере 12 показано, что TRU-016 не оказывает влияния на CDC активность в отношении других поверхностных рецепторов В-клеток; в примере 13 показано, что TRU-016 не усиливает CDC активность в отношении различных рецепторов-мишеней, включая MHCII, CD19, CD80/86 и CD40; в примере 14 представлены дополнительные данные, показывающие, что TRU-016 повышает выживаемость мышей с опухолями в условиях in vivo; в примере 15 показано, что TRU-016 потенцирует индуцированную флударабином гибель клеток CLL в условиях in vitro; в примере 16 показано, что TRU-016 индуцирует прямой цитотоксический эффект в отношении резистентных к ритуксимабу клеток; в примере 17 показано, что TRU-016 индуцирует фосфорилирование тирозина в CD19+ В-клетках первичной CLL; и в примере 18 приведены молекулы гуманизированных TRU-016.
Пример 1
Получение CD37-специфической связывающей молекулы
CD37-специфические SMIP описаны в совместной заявке на патент США № 10/627556 и патентных публикациях США № 2003/133939, 2003/0118592 и 2005/0136049. Получали примерный SMIP, TRU-016, как описано ниже.
TRU-016 [G28-1 scFv VH11S (SSC-P)H WCH2 WCH3] представляет рекомбинантный одноцепочечный белок, который связывается с антигеном CD37. Связывающий домен основан на последовательности антитела G28-1, ранее раскрытой в патентных публикациях, перечисленных в предыдущем абзаце, раскрытие которых включено здесь ссылкой. Связывающий домен связан с эффекторным доменом, СН2 и СН3 областями человеческого IgG1 посредством модифицированной шарнирной области. TRU-016 в растворе находится в виде димера, и димер имеет теоретическую молекулярную массу, равную примерно 106000 дальтон.
Выделяли общую фракцию РНК из гибридомы, продуцирующей G28-1, с использованием реагента на РНК Triazol (Gibco), следуя инструкциям изготовителя. Получали кДНК с использованием 5 мкг РНК, произвольных праймеров и обратной транскриптазы Superscript II (Gibco BRL). Вариабельные области клонировали с использованием пулов вырожденных праймеров для семейств генов различных мышиных VL или VН. Вариабельные области из гибридомы G28-1 клонировали в клонирующие векторы PCR 2.1 TOPO (Invitrogen) и секвенировали ДНК из трансформантов со вставками нужного размера. Затем вариабельные области тяжелой и легкой цепей из нужных клонов использовали в качестве матрицы для амплификации ПЦР G28-1 scFv, соединенного вместе в ориентации VL-VH, с линкером 15 аа (gly4ser3). scFv против CD37 присоединяли к модифицированной шарнирной области человеческого IgG1, областям СН2 и СН3 (смотри фигуру 1А). Для гарантии адекватной экспрессии клетками млекопитающих отбирали модификации вариабельных областей, позволяющие обеспечить существенное усиление экспрессии в клетках млекопитающих. В частности, лейцин заменяли на серин в положении 11 scFv. Предполагаемый зрелый пептид был длиной 473 аминокислоты.
Полинуклеотидная последовательность, кодирующая TRU-016, и аминокислотная последовательность TRU-016 соответственно представляют SEQ ID NO:1 и 2.
TRU-016 получали технологией рекомбинантной ДНК в экспрессирующей системе клеток млекопитающих, клетках яичников золотистого хомяка (СНО). Трансфицированные клетки СНО, которые продуцируют SMIP, культивировали в биореакторе с использованием подходящей среды.
TRU-016 SMIP выделяли из культуральных супернатантов СНО аффинной хроматографией на протеине А. С использованием dPBS колонку с 50 мл rПротеин А/сефароза FF (GE Healthcare rProtein A Sepharose FF, Catalog#17-0974-04) уравновешивали со скоростью потока 5,0 мл/мин (150 см/ч) 1,5 объемами колонки (CV). Культуральный супернатант наносили на колонку с rПротеин А/сефароза FF со скоростью проточной жидкости 1,7 мл/мин с использованием воздушного зонда AKTA 100 (GE Healthcare AKTA Explorer 100 Air, номер по каталогу #18-1403-00), с захватом рекомбинантного TRU-016. Колонку промывали dPBS в количестве 5 объемов колонки (CV), затем 1,0 М NaCl, 20 мМ фосфатом натрия, рН 6,0, и затем 25 мМ NaCl, 25 мМ NaOAc, рН 5,0. Данные стадии промывания приводили к удалению неспецифически связанных белков клеток-хозяев СНО с колонки с rПротеином А, которые вносят свой вклад в осаждение продукта после элюирования.
Рекомбинантный TRU-016 элюировали с колонки 100 мМ глицина, рН 3,5. Получали фракции элюированного продукта объемом 10 мл и затем элюированный продукт доводили до рН 5,0 0,5 М 2-(N-морфолино)этансульфоновой кислотой (MES), рН 6,0 в количестве 20% от элюированного объема. Данный элюированный продукт получали для очистки GPC концентрированием пробы примерно до 25 мг/мл TRU-016 и затем стерилизовали пропусканием через фильтр для очистки GPC.
Затем очищенный белок подвергали GPC гель-фильтрации (SEC) для дополнительной очистки молекулы TRU-016 (димера) от агрегатов с более высокой молекулярной массой. При использовании dPBS колонку ХК 50/100 (пустая хроматографическая колонка GE Healthcare XK 50/100, номер по каталогу #18-8753-01), содержащую 1 л сефарозы Superdex 200 FF, уравновешивали со скоростью потока 12,6 мл/мин (38 см/час) 1,5 объемами колонки. Максимальный объем 54 мл (3% CV) пробы наносили на колонку. Колонку продолжали элюировать со скоростью 12,6 мл/мин, и элюированный белок фракционировали на фракции по 40 мл. Каждую фракцию анализировали на качество продукта с использованием аналитической ВЭЖХ, и элюированные фракции TRU-016 с >95% POI (не подвергшиеся агрегации) объединяли. Данную полученную объединенную фракцию стерилизовали пропусканием через фильтр с размером пор 0,22 мкм. Затем вещество концентрировали и переводили в раствор 20 мМ фосфата натрия и 240 мМ сахарозы с полученным рН 6,0. Композицию фильтровали перед переносом в стеклянные флаконы с концентрацией 10 мг/мл. Каждый стеклянный флакон содержал 5 мл TRU-016 (50 мг/флакон).
Белок TRU-016 также подвергали анализу SDS-PAGE на гелях с 4-20% Трис-глицином Novex (Invitrogen, San Diego, CA). Пробы наносили с использованием буфера для образцов SDS Трис-глицин Novex (2×) в восстанавливающих условиях (добавление 1/10 объема восстановителя для проб NuPAGE) или невосстанавливащих условиях после нагревания при 95°С в течение 3 мин с последующим проведением электрофореза при 150 B в течение 90 мин. Электрофорез проводили с использованием буфера для электрофореза 1× Трис-глицин Novex SDS (Invitrogen). Гели после электрофореза окрашивали краской Кумасси SDS-PAGE R-250 в течение 30 мин при встряхивании, и краску удаляли, по меньшей мере, в течение 1 ч. Предполагаемое значение молекулярной массы зрелого пептида составляет 51,5 кДа. В восстанавливающих условиях рекомбинантный белок мигрирует соответственно предполагаемой молекулярной массе. В невосстанавливающих условиях молекула мигрирует соответственно массе, составляющей примерно 150 кДа (фигура 1В).
Также проводили опыты с целью определения того, что специфичность связывания исходного антитела с клеточным поверхностным рецептором CD37 сохраняется у TRU-016. Человеческие РВМС выделяли в градиенте плотности среды для разделения лимфоцитов и инкубировали с неконъюгированным TRU-016 и РЕ-конъюгированными антителами против человеческого CD19. Клетки промывали и инкубировали с FITC GAH IgG 1:100 (Fc-специфическим) в течение 45 мин на льду. Клетки промывали и анализировали двухцветной проточной цитометрией на цитометре FACsCalibur c использованием программного обеспечения CellQuest. Клетки анализировали с окном возбуждения на В-лимфоциты или отличные от В-лимфоцитов при окрашивании CD19.
При увеличении концентраций TRU-016 сигнал FITC на В-лимфоцитах (CD19 позитивное окно) быстро возрастал от 0,01-1,0 мкг/мл до тех пор, пока он не достигал насыщения примерно при 1 мкг/мл или среднего значения интенсивности флуоресценции (MFI), равного 1000. В противоположность окрашивание популяции клеток, отличных от В-лимфоцитов, подвергалось детектированию, но было очень слабым и медленно возрастало с повышением концентрации scFvlg. Таким образом, характер окрашивания мышиным моноклональным антителом G28-1 сохраняется у TRU-016 (фигура 1С).
CD37-связывающие молекулы по изобретению имеют структуры (связывающие области из антитела, варианты шарнирной области, где СН2СН3 области являются такими же или другими, и различных изотипов).
Пример 2
TRU-016 и различные CD37-специфические антитела связываются с одними и теми же или перекрывающимися эпитопами в CD37
Проводили опыты с целью идентификации эпитопа CD37, связывающегося с TRU-016 и другими описанными ранее CD37-специфическими антителами.
Неконъюгированные МВ371 (#555457) и FITC-конъюгированные МВ371 (#555456) получали от BD Pharmingen (San Jose, CA), FITC-конъюгированные BL14 (#0457) - от Immunotech/Beckman Coulter (Fullerton, CA), FITC-конъюгированные NMN46 (#RDI-CBL 136FT) и некоъюгированные NMN46 (#RDI-CBL 136) - от RDI (Flanders, NJ), FITC-конъюгированные IPO24 (#186-040) и неконъюгированные IPO24 (#186-020) - от Ancell Corporation (Bayport, MN), FITC-конъюгированные HHI (#3081) и неконъюгированные HHI (#3080) - от DiaTec. Com. (Oslo, Norway) и FITC-конъюгированные WR17 (YSRTMCA483F) и неконъюгированные WR17 (YSRTMCA483S) - от Accurate Chemical & Scientific (Westbury, NY). Белок TRU-016 получали, как описано в примере 1.
TRU-016 конъюгировали с FITC в Trubion с использованием набора для введения метки Molecular Probes Fluororeporter FITC (F6434), следуя инструкциям изготовителя, следующим образом: интересующий пик белка TRU-016 (POI) с концентрацией 13,5 мг/мл доводили до 5 мг/мл с использованием PBS. 1 мг (200 мкл) вносили в пробирки, имеющиеся в наборе, с перемешивающим стержнем и добавляли 1 М NaHCO3 (доводили до рН 8,5 6Н раствором NaOH) до конечной концентрации 0,1 М. Добавляли 50 мкл ДМСО к 370 мкг FITC и вносили в пробирки при молярном соотношении FITC:белок, равном 15, 20, 30 и 40, с использованием следующей формулы для определения мкл FITC для добавления: [мкл раствора FITC для внесения = 5 мг/мл белка × 0,2 мл × 389 × 100 × желаемое молярное соотношение/молекулярная масса TRU-016 (110000)].
Реакционные смеси ставили в темноту и непрерывно перемешивали в течение 75 мин при комнатной температуре. Реакционные смеси наносили на вращающиеся колонки, приготовленные, как описано в инструкции к набору, и центрифугировали при 1100 g в течение 5 мин с заменой буфера на PBS с азидом и удалением неконъюгированного FITC. Определяли оптическую плотность при 280 нм и 494 нм при добавлении 2 капель Nanodrop; экспериментально устанавливали для данного прибора коэффициент экстинкции для TRU-016 при замере показаний для разведений исходного неконъюгированного SMIP, концентрация каждого из конъюгатов составляла 4,25 мг/мл, и определяли значения при следующих соотношениях FITC:белок: 2,7 FITC:TRU-016 при соотношении 15; 3,7 FITC:TRU-016 при соотношении 20; 4,4 FITC:TRU-016 при соотношении 30 и 5,1 FITC:TRU-016 при соотношении 40.
Добавляли BSA до концентрации 3 мг/мл для стабилизации белка. Связывание каждой фракции оценивали при разведениях от 100-24300× на клетках Ramos и 3200-25600 на человеческих РВМС. Все связывались, но соотношение MR30 было выбрано для дальнейшего использования, поскольку оно давало высокое значение MFI, которое хорошо сохранялось в используемых пределах титрования, указывая на то, что в данной реакции оказывалось минимальное влияние на авидность связывания.
В первоначальном исследовании связывания конъюгаты меченного FITC антитела титровали в пределах от 10 нг/мл до 10 мкг/мл с целью установления оптимальных количеств для использования в опытах с блокированием. Выбранная концентрация была несколько ниже насыщающих количеств, и ее сохраняли постоянной в последующих опытах, в то время как концентрации блокирующего антитела увеличивали в 10-кратных пределах. По данным строили график зависимости процента максимального связывания от концентрации блокирующего антитела, при том что более высокие концентрации указывали на менее эффективное блокирование, в то время как более низкие концентрации указывали на имеющуюся более высокую активность блокирования. Все тестированные антитела проявили блокирующую активность при максимальном связывании, наблюдаемом без не меченных реагентов (фигура 2).
Затем клетки BJAB, В-лимфобластоидная линия В-клеток (любезно предоставленная Ed. Clark, University of Washington), окрашивали панелью различных клонов MAb против CD37, включая МВ371, BL14, NMN46, IPO24, HH1, WR17 и TRU-016 SMIP.
Для постановки опытов по конкурентному связыванию 2,5×105 клеток BJAB инкубировали в 96-луночных планшетах с U-образным дном в среде для окрашивания (PBS c 2% мышиной сыворотки) с FITC-конъюгированными MAb против CD37 в концентрации 1,25 мкг/мл в присутствии неконъюгированных MAb против CD37 в указанных концентрациях (2,5, 1,25, 0,6 или 0,3 мкг/мл) или в среде для окрашивания в течение 45 мин на льду в темноте. Блокирующие антитела и FITC-меченные конъюгаты антител вносили в реакционные смеси перед добавлением клеток. Затем клетки промывали 2,5 раза PBS и фиксировали в 1% параформальдегиде (#19943, USB, Cleveland, Ohio). Клетки анализировали проточной цитометрией с использованием цитометра FACsCalibur и программного обеспечения CellQuest (BD Biosciences, San Jose, CA).
Для постановки опытов по перекрестному блокированию с FACs 2,5×105 клеток BJAB инкубировали в 96-луночных планшетах с U-образным дном в среде для окрашивания (PBS c 2% мышиной сыворотки) в присутствии неконъюгированных MAb против CD37 в концентрации 5 мкг/мл среды для окрашивания в течение 45 мин при комнатной температуре на льду в темноте. Затем FITC-конъюгированные MAb против CD37 вносили в конечной концентрации 2 мкг/мл с разведением не меченных реагентов до концентрации 3,3 мкг/мл. Затем реакционные смеси дополнительно инкубировали в течение 45 мин при комнатной температуре в темноте. Реакционные смеси промывали 2,5 раза PBS и фиксировали в 1% параформальдегиде (#19943, USB, Cleveland, Ohio). Клетки анализировали проточной цитометрией с использованием цитометра FACsCalibur и программного обеспечения CellQuest (BD Biosciences, San Jose, CA).
Для постановки клеточных тестов по связыванию клетки суспендировали в PBS (#14040-133, Gibco/Invitrogen, Grand Island, NY), содержащем 2% FBS (#16140-071, Gibco/Invitrogen, Grand Island, NY) (среда для окрашивания) с получением титра примерно 4×106 клеток/мл. Затем клетки высевали в планшеты и затем вносили тестируемые пробы, разведенные в среде для окрашивания, в соотношении 1:1 до конечных указанных концентраций. Реакционные смеси инкубировали в течение 45 мин на льду. Пробы центрифугировали и промывали 2 раза PBS. Добавляли меченный FITC козий античеловеческий IgG (#H10501, CaITag, Burlingame CA) до конечного разведения 1:50 и инкубировали в течение 45 мин на льду. Пробы центрифугировали, промывали PBS, затем фиксировали в 1% параформальдегиде в PBS (#19943, USB, Cleveland, Ohio). Клетки анализировали проточной цитометрией с использованием цитометра FACsCalibur и программного обеспечения CellQuest (BD Biosciences, San Jose, CA).
Было показано, что каждое антитело проявляло зависимое от дозы ингибирование связывания, этот факт указывает на то, что все тестированные молекулы связываются с одним и тем же или очень близким эпитопом. Для каждого антитела наблюдали различную активность в ингибировании связывания. TRU-016 SMIP обладали наиболее высокой блокирующей активностью из всех тестированных молекул, в то время как НН1 проявляли промежуточный уровень блокирующей активности, и WR17, IPO24 блокировали сильнее, чем МВ371, но были менее эффективными в блокировании, чем две другие не меченные молекулы (фигура 2).
В дополнение к анализу блокирующей активности проводили аналогичную серию экспериментов, в которых различные антитела против CD37 тестировали на их способность конкурировать друг с другом за связывание с рецептором CD37. Результаты данных опытов, как и результаты, полученные в экспериментах по блокированию для всех тестированных молекул, указывали, что различные антитела против CD37 и TRU-016 имеют одни и те же или перекрывающиеся эпитопы.
Пример 3
TRU-016 мало активен в связывании C1q и активировании классического пути активации комплемента
Проводили эксперименты для исследования того, почему димер TRU-016 не способен опосредовать высокую зависимую от комплемента гибель В-клеток-мишеней. Одной возможностью является то, что димер TRU-016 проявляет низкое связывание с компонентами каскада комплемента по сравнению с нормальным человеческим IgG1-антителом. Таким образом, проводили опыты с целью определения того, активирует ли TRU-016 классический путь активации комплемента при оценке связывания TRU-016 с C1q. C1q представляет собой субъединицу ферментного комплекса С1, который активирует сывороточную комплементную систему и является компонентом распознавания классического пути активации комплемента.
Опыты по связыванию с C1q проводили, как описано ранее (Cragg et al., Blood 2004, 103: 2738-2743). Вкратце, В-клетки Ramos в среде Iscove (#12440-053, Gibco/Invitrogen, Grand Island, NY) без сыворотки высевали в 96-луночные планшеты с U-образным дном с титром 5×105 клеток/лунка в 100 мкл. Клетки инкубировали с реагентами в течение 15 мин при 37°С и затем вносили нормальную человеческую сыворотку (NHS, #A113, Quidel Corp., San Diego, CA), разведенную в среде Iscove, в объеме 50 мкл в каждую лунку до конечной концентрации 10, 5, 2,5 или 1,25% человеческой сыворотки. 50 мкл среды вносили в контрольную лунку. Для опытов с фактором из яда кобры CVF добавляли к пробам комплемента человеческой сыворотки из расчета 20 Ед. CVF/мл сыворотки на 90 мин при 37°С перед внесением сыворотки к пробам комплемента и с учетом разведения сыворотки CVF при приготовлении разведений проб.
Клетки плюс источник комплемента инкубировали еще в течение 5 мин при 37°С и промывали 2 раза холодным PBS (#14040-133, Gibco/Invitrogen, Grand Island, NY) при центрифугировании, и ресуспендировали в 100 мкл PBS. 50 мкл из каждой лунки переносили во второй планшет для проведения второй стадии контрольного окрашивания. Оба планшета окрашивали в течение 15 мин в темноте на льду либо меченным FITC анти-HU C1q (#C7850-06A, US Biological, Swampscott, Mass), или меченным FITC овечьим IgG (#11904-56P, US Biological, Swampscott, Mass). Пробы промывали, ресуспендировали в холодном PBS и сразу же снимали показания на проточном цитометре FACsCalibur, и их обрабатывали с использованием программного обеспечения CellQuest (BD Biosciences, San Jose, CA).
FITC C1q не связывался в значительной степени ни с одной субфракцией очищенного SEC TRU-016, несмотря на то, что фракция с более высокой молекулярной массой (HMW) или агрегаты А2 связывались сильнее по сравнению с другими формами (фигура 3А). В противоположность ритуксан проявлял более высокий уровень связывания с C1q, в частности, при более низких концентрациях NHS. Присутствие CVF не могло полностью блокировать данное связывание, хотя значения MFI достоверно уменьшались по сравнению с одной средой.
Затем ставили тесты по оценке CDC для сравнения способности различных субфракций очищенного TRU-016 и ритуксана опосредовать гибель клеток в присутствии или в отсутствие CVF и человеческого сывороточного комплемента (фигура 3В). Тесты по CDC проводили с использованием окрашивания пропидием йодидом для дифференциации живых и мертвых клеток после инкубации клеток-мишеней с антителом, рекомбинантными белками, асцитной жидкостью, молекулами TRU-016 или средой и источником комплемента, таким как человеческая сыворотка. Вкратце, 3×105 В-клеток Ramos предварительно инкубировали с тестируемыми реагентами в течение 30-45 мин при 37°С перед внесением комплемента. Предварительно связанные пробы центрифугировали, промывали и ресуспендировали в среде Iscove (#A113, Quidel, San Diego, CA) в указанных концентрациях, затем инкубировали в течение 90 мин при 37°С. Пробы промывали и добавляли пропидий йодид (#P-16063, Molecular Probes, Eugene, OR) до конечной концентрации 0,5 мкг/мл в PBS. Клетки инкубировали с пропидием йодидом в течение 15 мин при комнатной температуре в темноте и затем анализировали проточной цитометрией на цитометре FACsCalibur с использованием программного обеспечения CellQuest (BD Biosciences, San Jose, CA).
Гибель клеток, опосредованная фракцией А2 TRU-016 и ритуксаном, была достоверно ниже в присутствии CVF, несмотря на его неспособность полностью блокировать связывание C1q (фигура 3В).
Затем проводили сравнение человеческого и кроличьего комплемента по их способности к CDC в присутствии TRU-016. Оценивали активность к CDC молекул TRU-016, инкубированных с В-клетками Ramos и человеческим или кроличьим комплементом (фигура 3С). В-клетки Ramos вносили в лунки с бессывороточной средой. Ритуксан или димер, HMW A2 или рА фракции TRU-016 добавляли к клеткам с получением конечной концентрации 10 мкг/мл и инкубировали в течение 15 мин при 37°С перед промыванием 1,5× в бессывороточной среде и добавлением нормальной человеческой сыворотки (NHS) или кроличьего комплемента (Pelfreez) при концентрации 10, 5 или 2,5%. Клетки плюс источник комплемента инкубировали в течение 90 мин при 37°С. Клетки промывали один раз холодным PBS и вносили пропидий йодид (Molecular Probes, #P3566) до конечной концентрации 0,5 мкг/мл в холодном PBS. Клетки инкубировали с PI в темноте при комнатной температуре в течение 15 мин и затем анализировали проточной цитометрией.
Происхождение фракции комплемента оказывает влияние на полученные результаты по CDC (фигура 3С). Кроличий комплемент опосредовал более высокую CDC по сравнению с человеческим комплементом в присутствии молекул TRU-016. Интересно отметить, что димерная форма TRU-016 опосредовала высокую CDC с использованием кроличьего комплемента, но очень низкую активность CDC в присутствии человеческого комплемента.
Пример 4
Активность и связывание мультимеров TRU-016
Проводили эксперименты с целью оценки биологической активности мультимерных форм TRU-016 (мультимеры TRU-016) в растворе. Первое, для установления размера рекомбинантного белка TRU-016 очищенное на протеине А вещество анализировали SEC при высоком давлении (ВЭЖХ) и установили, что в растворе TRU-016 находится в виде мультимеров.
Хроматограммы гель-фильтрации (SEC) при высоком давлении (ВЭЖХ) получали очисткой GPC TRU-016, построив график зависимости поглощения от времени удерживания для различных собранных фракций (фигура 4). TRU-016 выделяли из клеточных культуральных супернатантов первоначально аффинной хроматографией с использованием протеин А/сефароза. Рекомбинантную молекулу элюировали с колонки 100 мМ глицином, рН 3,5. Собирали фракции элюированного продукта объемом 10 мл и затем рН элюированного продукта доводили до 5,0 0,5 М 2-(N-морфолино)этансульфоновой кислоты (MES), рН 6,0, в количестве 20% от элюированного объема. Элюат получали для очистки GPC концентрированием пробы примерно до концентрации 25 мг/мл TRU-016 и затем стерилизовали фильтрованием при приготовлении для очистки GPC. Проводили гель-фильтрацию c использованием воздушного зонда GE Healthcare AKTA Explorer 100 на колонке GE Healthcare ХК и препаративной Superdex 200 (GE Healthcare).
Фракции HMW или А2 показывали время удерживания, равное примерно 6,23 мин, в то время как наиболее выраженная форма имела время удерживания, составляющее 8,38 мин. Используемым в данном случае в качестве референс-стандарта было (рА стандарт или std) очищенное на протеине А вещество, содержащее димеры и мультимеры HMW, как представлено на первой панели на фигуре 4. Наиболее выраженная форма, мигрирующая со временем удерживания 8,38 мин, наиболее вероятно соответствовала молекуле димера в невосстанавливающих условиях SDS-PAGE и несколько минорных форм наиболее вероятно соответствовали мультимерам, которые образовывались посредством нековалентных взаимодействий, поскольку их не обнаруживали при SDS-PAGE в невосстанавливающих условиях. Для разделения данных различных форм TRU-016 вещество, полученное аффинной хроматографией культуральных супернатантов на протеин А/сефароза, подвергали дополнительной очистке GPC и фракционированию ВЭЖХ для отделения димера (обозначенного как «димеры» или «пик димеров») от мультимеров с более высокой молекулярной массой (обозначенных как фракция HMW или А2 agg). Затем каждую из данных трех субфракций анализировали по отдельности на функциональную активность в условиях in vitro с использованием тестов оценки связывания, ADCC и CDC.
Для исследования того, насколько фракции, выделенные SEC, проявляют различные связывающие свойства, каждую фракцию TRU-016 SEC анализировали на способность связываться с клетками Ramos. Для оценки связывающих свойств фракций, полученных с использованием SEC, клетки суспендировали в среде для окрашивания с титром 4×106 клеток/мл и затем высевали в планшеты из расчета 50 мкл/лунка (2×105 клеток/лунка) в среде для окрашивания. Затем серийные разведения фракций SEC вносили в последовательные лунки, инкубировали в течение 45 мин, промывали и активность связывания определяли с использованием FITC козьего античеловеческого IgG. Пробы фиксировали в 200 мкл 1% параформальдегида в PBS. Клетки анализировали проточной цитометрией на цитометре FACsCalibur с использованием программного обеспечения CellQuest (BD Biosciences, San Jose, CA).
Для оценки активности фракций, полученных SEC, в тесте CDC клетки суспендировали при титре 5×105 клеток/лунка в 75 мкл IMDM. Фракции TRU-016 SEC (75 мкл) добавляли к клеткам в двойной указанной концентрации. Давали возможность протекать реакциям связывания в течение 45 мин перед центрифугированием и промыванием в бессывороточной среде Iscove. Клетки ресуспендировали в среде Iscove с человеческой сывороткой (#A113, Quidel, San Diego, CA) в указанных концентрациях. Клетки инкубировали в течение 60 мин при 37°С, промывали и ресуспендировали в среде для окрашивания с 0,5 мкг/мл пропидия йодида (PI, #P-16063, Molecular Probes, Eugene OR). Пробы инкубировали в течение 15 мин при комнатной температуре в темноте перед анализом проточной цитометрией с использованием цитометра FACsCalibur и программного обеспечения CellQuest (Becton Dickinson)(фигура 5В).
Для оценки активности фракций, полученных SEC, в тесте ADCC лимфобластоидные В-клетки BJAB, Ramos и Daudi (107) метили 500 мккюри/мл 51Cr-хромата натрия в течение 2 ч при 37°С в среде IMDM/10% FBS. Выделяли РВМС из гепанизированной человеческой крови фракционированием в градиенте среды для разделения лимфоцитов (LSM, ICN Biomedical). Пробы реагентов добавляли к среде RPMI с 10% FBS и готовили пять серийных разведений для каждого реагента. Для получения комбинаций реагенты предварительно смешивали и разводили перед внесением в лунки. Меченные 51Cr BJAB вносили с титром 2×104 клеток/лунка. Затем добавляли РВМС (5×105 клеток/лунка) с конечным соотношением эффекторные клетки (РВМС):клетки мишеней (BJAB), равным 25:1. Реакционные смеси вносили в четыре параллельные лунки 96-луночного планшета. Фракции TRU-016 SEC вносили в лунки с конечной концентрацией в пределах от 10 нг/мл до 20 мкг/мл, как указано на графиках. Каждый ряд данных представляет график для различной фракции, полученной SEC, при описанных пределах титрования. Реакциям давали протекать в течение 6 ч при 37°С в атмосфере 5% СО2 перед сбором и подсчетом. Определяли высвобожденное количество имп/мин на счетчике Packard TopCountNXT из 50 мкл высушенного культурального супернатанта. Процент специфической гибели клеток рассчитывали вычитанием (имп/мин [среднее значение четырех параллельных проб] проб - имп/мин спонтанное высвобождение)/(имп/мин максимальное высвобождение - имп/мин спонтанное высвобождение) × 100 (фигура 5С).
На фигуре 5А представлены кривые титрования связывания различных фракций SEC с клетками Ramos. Все фракционированные молекулы связывались с CD37 при наличии аналогичных кривых связывания, за исключением варианта тестирования при самой высокой концентрации, когда в этом случае вещество HMW показывало лучшее связывание (более высокая интенсивность флуоресценции) по сравнению с рА стандартом и пиком димерных форм.
Также проводили эксперименты с целью определения того, проявляют ли фракции TRU-016 SEC различную функциональную активность, такую как опосредованная CDC и ADCC направленная гибель клеток. На графике, представленном на фигуре 5В, показано, что только очищенная фракция мультимеров HMW опосредовала высокую CDC активность в отношении В-клеток Ramos при использовании человеческого комплемента. Стандарт рА проявлял некоторую CDC активность в более высоких концентрациях, в то время как пик димера показывал очень низкую активность или отсутствие CDC активности при всех тестированных концентрациях.
Проводили тесты по оценке ADCC с серийными разведениями фракций TRU-016 различной массы с использованием меченых В-клеток BJAB в качестве мишени и человеческих РВМС в качестве эффекторных клеток. Фракции TRU-016 SEC находились в лунках в конечной концентрации в пределах от 10 нг/мл до 20 мкг/мл, как указано на графике на фигуре 5С. Каждый ряд данных представляет график для различных фракций, полученных SEC, при описанных пределах титрования. Строили график в виде функции % специфической гибели от концентрации белка. Все субфракции SEC, включая рА стандарт, фракцию HMW или А2 и пик димера, опосредовали высокую, зависимую от дозы ADCC в отношении клеток-мишеней BJAB. Также аналогичные результаты получали с использованием клеток Ramos в качестве меченых мишеней (данные не представлены).
Пример 5
Получение CD20-специфической связывающей молекулы
CD20-специфические SMIP описаны в совместных патентных публикациях США № 2003/133939, 2003/0118592 и 2005/0136049. Получение примерного CD20-специфического SMIP, TRU-015, описано ниже.
TRU-015 представляет рекомбинантный (мышиный/человеческий) одноцепочечный белок, который связывается с антигеном CD20. Связывающий домен основан на известной последовательности человеческого антитела против CD20. Связывающий домен связан с эффекторным доменом, СН2 и СН3 областями человеческого IgG1 посредством модифицированной шарнирной области CSS. TRU-015 в растворе находится в виде димера, и димер имеет теоретическую молекулярную массу, равную примерно 106000 дальтон. Нуклеотидная последовательность, кодирующая TRU-016, и аминокислотная последовательность TRU-015 соответственно представляют SEQ ID NO:3 и 4.
В отношении аминокислотной последовательности, представленной в SEQ ID NO:4, TRU-015 содержит лидерный пептид 2е12, клонированную последовательность из аминокислот 1-23; вариабельную область легкой цепи мышиного антитела 2Н7 против человеческого CD20 с заменой лизина на серин (VHL11S) в остатке 11 в вариабельной области, который указан в положении 34; линкер asp-gly3-ser-(gly4ser)2, начинающийся в остатке 129, при том что линкер имеет дополнительный серин в конце с включением сайта рестрикции Sacl для кассетной перестановки; вариабельную область тяжелой цепи мышиного антитела 2Н7 против человеческого CD20, в которой отсутствует остаток серина в конце области тяжелой цепи, т.е. измененной с VTVSS на VTVS; Fc-область человеческого IgG1, включающую модифицированную шарнирную область, содержащую последовательность (CSS), и области СН2 и СН3 дикого типа.
Клетки СНО, которые продуцируют TRU-015, культивировали в биореакторе с использованием подходящей среды. TRU-015 очищали с использованием ряда стадий хроматографии и фильтрования, включая фильтрование для элиминации вирусов. Затем вещество концентрировали и готовили раствор 20 мМ фосфата натрия и 240 мМ сахарозы с получением рН, равного 6,0. Композицию фильтровали перед переносом в стеклянные флаконы с концентрацией 10 мг/мл. Каждый стеклянный флакон содержал 5 мл TRU-015 (50 мг/флакон).
Пример 6
Комбинации TRU-016 с TRU-015 или ритуксаном синергически усиливают апоптоз В-клеток
Проводили опыты с целью исследования влияния SMIP c направленным действием против В-клеток на апоптоз В-клеток. Каждый SMIP тестировали индивидуально и затем в комбинации. Пробы анализировали через 24 и 48 ч после начала реакций инкубации. Анализ с использованием аннексина/PI проводили следующим образом: клетки BJAB (любезно представленные Ed. Clark, University of Washington), Ramos (ATCC# CRL-1596) и Daudi инкубировали в течение 24 и 48 ч при 37°С в атмосфере 5% СО2 в полной среде Iscove (Gibco) с 10% FBS при титре 3×105 клеток/мл и концентрации белка SMIP 20 мкг/мл. Кроме того, в реакционные смеси добавляли 20 мкг/мл козьего античеловеческого IgG для перекрестного связывания реагентов на клеточной поверхности. Затем клетки окрашивали аннексином V-FITC и пропидием йодидом с использованием набора I для детектирования апоптоза BD Pharmigen (#556547) и обрабатывали, следуя инструкциям, имеющимся в наборе. Вкратце, клетки промывали дважды холодным PBS и ресуспендировали в «связывающем буфере» при титре 1×106 клеток/мл. Затем 100 мкл клеток в «связывающем буфере» окрашивали 5 мкл смеси аннексина V-FITC и 5 мкл пропидия йодида. Клетки мягко обрабатывали на вортексе и инкубировали в темноте при комнатной температуре в течение 15 мин. К каждой пробе добавляли 400 мкл «связывающего буфера». Затем снимали показания и анализировали на цитометре FACsCalibur с использованием программного обеспечения CellQuest (Becton Dickinson).
В таблице 2, ниже, показано, что в присутствии реагента для перекрестного связывания обработка TRU-016 оказывала более существенное влияние в отношении проявления апоптоза на клеточные линии по сравнению с одним TRU-015, хотя обе молекулы при использовании их одних не индуцировали какой-либо апоптоз. Усиление апоптоза было различным в зависимости от клеточной линии.
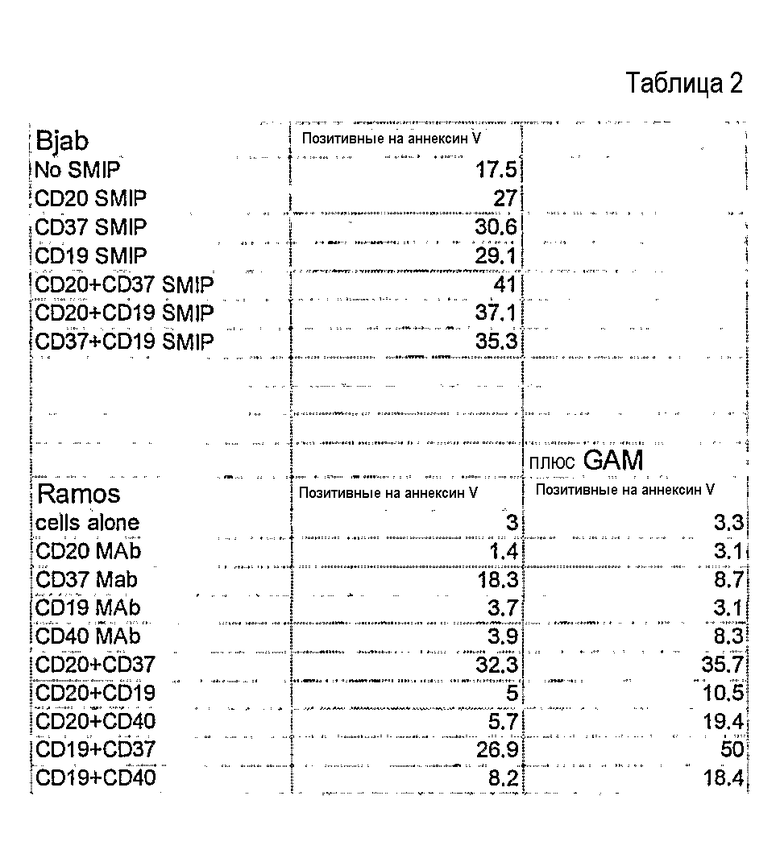
Пример 7
Комбинации TRU-016 с CD20-специфическими антителами или SMIP синергически усиливают CDC
Проводили эксперименты с целью оценки CDC активности комбинаций TRU-016 с CD20-специфическими антителами или SMIP в отношении В-клеток. Концентрации реагентов, выбранные для опытов с комбинациями, составляли 0,5 мкг/мл для TRU-016, для TRU-015 оно также равнялось 0,5 мкг/мл. Как правило, концентрация ритуксана составляла 0,04-0,06 мкг/мл ввиду его более высокой активности в опытах по CDC с одним этим реагентом. В некоторых опытах концентрацию реагента против CD20 сохраняли постоянной на субоптимальном уровне, в то время как концентрацию TRU-016 изменяли для установления минимальных уровней реагента против CD20, необходимых для усиления CDC.
Клетки суспендировали в среде Iscove (#12440-053, Gibco/Invitrogen, Grand Island, NY) при титре 5×105 клеток/лунка в 75 мкл. TRU-016 (75 мкл), TRU-015, ритуксан или комбинации данных реагентов добавляли к клеткам в двойных указанных концентрациях. Давали возможность протекать реакциям связывания в течение 45 мин перед центрифугированием и промыванием в бессывороточной среде Iscove. Клетки ресуспендировали в среде Iscove с человеческой сывороткой (#A113, Quidel, San Diego, CA) в указанных концентрациях. Клетки инкубировали в течение 60 мин при 37°С. Клетки промывали и ресуспендировали в 125 мкл PBS с 2% FBS (#16140-071, Gibco, Invitrogen, Grand Island, NY), среде для окрашивания. Клетки переносили в пробирки для FAСS (#4410, CoStar, Corning, NY) и вносили 125 мкл среды для окрашивания с 5 мкл пропидия йодида (PI, #P-16063, Molecular Probes, Eugene OR). Пробы инкубировали в течение 15 мин при комнатной температуре в темноте перед анализом проточной цитометрией с использованием цитометра FACsCalibur и программного обеспечения CellQuest (Becton Dickinson).
На фигуре 6 показано, что в субоптимальных концентрациях, обеспечивающих гибель клеток при использовании одних этих реагентов, TRU-015 и ритуксан проявляли более высокую активность в CDC в комбинации с TRU-016. Один TRU-016 не мог опосредовать CDC, если только не присутствовали агрегаты. Удаление C1q из реакционных смесей приводило к полной элиминации активности CDC.
На фигуре 7 показаны результаты опытов по комбинированию реагентов, проведенных на В-клетках DHL-4. Внесение TRU-016 в реакционные смеси для оценки CDC приводило к смещению вниз к кривой гибели клеток под действием TRU-015, что указывает на более высокую гибель в каждой тестированной концентрации, хотя один TRU-016 проявлял низкую активность или имело место отсутствие активности.
На фигуре 8 приведены данные другого опыта по оценке CDC, в котором реагенты смешивали в следующих количествах: 0,5 мкг/мл TRU-015, 0,5 мкг/мл TRU-016 и 0,06 мкг/мл ритуксана. Вновь одни реагенты применяли в субоптимальных концентрациях для того, чтобы детектировать эффект усиления в присутствии TRU-016. TRU-016 для обоих, TRU-015 и ритуксана, повышал гибель клеток в результате проявления CDC при его внесении в тестах.
На фигурах 9 и 10 приведены графики по данным тестов оценки CDC, в которых концентрацию TRU-015 или ритуксана сохраняли постоянной, а концентрацию TRU-016 увеличивали. Вновь активность CDC повышалась при добавлении в реакционные смеси TRU-016, но увеличение концентрации TRU-016 в пределах от 0,5 мкг/мл до 2,5 мкг/мл достоверно не увеличивало опосредованную CDC гибель клеток в данных опытах.
Пример 8
TRU-016 усиливает ADCC и CDC активность CD20-специфических антител и SMIP
Проводили опыты с целью определения того, насколько комбинации TRU-016 SMIP с CD37-специфическими антителами или SMIP могут усиливать ADCC и CDC активность в отношении В-клеток-мишеней.
Лимфобластоидные В-клетки BJAB, Ramos и Daudi (107) метили 500 мккюри/мл 51Cr-хромата натрия в течение 2 ч при 37°С в среде IMDM/10% FBS. Меченые клетки BJAB промывали три раза в среде RPMI/10% FBS и ресуспендировали при титре 4×105 клеток/мл в среде RPMI. Гепаринизированную человеческую цельную кровь получали от анонимных доноров и РВМС выделяли фракционированием в градиенте среды для разделения лимфоцитов (LSM, ICN Biomedical). Собирали светлый слой кровяного сгустка и дважды промывали в среде RPMI/10% FBS перед ресуспендированием в RPMI/10% FBS с конечным титром 3×106 клеток/мл. Подсчитывали количество клеток по вытеснению трипанового синего с использованием гемацитометра перед использованием в последующих тестах. К среде RPMI с 10% FBS добавляли пробы реагентов в четырехкратной конечной концентрации и готовили пять серийных разведений для каждого реагента. Для приготовления комбинаций реагенты смешивали и разбавляли перед внесением в лунки. Затем данные реагенты вносили в 96-луночные планшеты с U-образным дном из расчета 50 мкл/лунка для указанных конечных концентраций. Меченные 51Cr клетки BJAB вносили в планшеты из расчета 50 мкл/лунка (2×104 клеток/лунка). Затем добавляли РВМС в планшеты из расчета 100 мкл/лунка (3×105 клеток/лунка) до конечного соотношения эффекторные клетки (РВМС):(BJAB) клетки-мишени, равного 15:1.
Эффекторные клетки и клетки-мишени вносили в одну среду для определения уровня фоновой гибели. Меченные 51Cr клетки BJAB добавляли к одной среде для определения спонтанного высвобождения 51Cr и к среде с 5% NP40 (#28324, Pierce, Rockford, IL) для определения максимального высвобождения 51Cr. Реакции проводили в четырех параллельных лунках в 96-луночном планшете. SMIP вносили в лунки в конечной концентрации в пределах от 12 нг/мл до 10 мкг/мл, как указано на графиках. Для приготовления комбинаций SMIP реагенты смешивали перед внесением в лунки. Каждый ряд данных представляет график для разных одних SMIP или их комбинаций при описанных пределах титрования. Реакциям давали протекать в течение 6 ч при 37°С в атмосфере 5% СО2 перед сбором и подсчетом. 50 мкл супернатанта из каждой лунки переносили в планшет Luma Plate 96 (#6006633, Perkin Elmer, Boston, Mass) и высушивали в течение ночи при комнатной температуре. Определяли значение высвобожденного количества имп/мин на счетчике Packard TopCountNXT. Процент специфической гибели клеток рассчитывали вычитанием (имп/мин [среднее значение для четырех параллельных проб] пробы - имп/мин спонтанное высвобождение)/(имп/мин максимальное высвобождение - имп/мин спонтанное высвобождение) × 100.
Строили график в виде функции % гибели клеток от концентрации SMIP. На каждом чертеже указано соотношение эффекторные клетки:клетки-мишени, и также указана клеточная линия-мишень. На фигурах 11, 12 и 13 приведены результаты опытов, проведенных на различных клеточных линиях (BJAB, Ramos и Daudi), при этом использовали одного и того же донора.
На фигурах 14 и 15 (ритуксан+TRU-016) и на фигурах 16 и 17 (TRU-015+TRU-016) приведены данные экспериментов, в которых в качестве клеточной линии-мишени использовали BJAB. Специфическая гибель клеток, наблюдаемая для каждой комбинации, была выше по сравнению с каждым одним реагентом в такой же концентрации, этот факт указывает на то, что направленные против CD20 и CD37 SMIP усиливают гибель, опосредуемую каждым другим реагентом, хотя эффект повышения не был полностью аддитивным.
Таким образом, TRU-016 может повышать опосредуемую ADCC гибель В-клеток под действием CD20-специфических SMIP или CD20-специфических антител.
Первоначальные опыты, цель которых заключалась в исследовании действия комбинаций TRU-016 и антител против CD20, планировали провести для определения относительных количеств каждого реагента для применения таким образом, чтобы детектировался синергический эффект в CDC. Клетки Ramos суспендировали в среде IMDM и TRU-016, ритуксан или комбинации данных реагентов добавляли к клеткам до конечных концентраций, указанных на фигуре 18. Реакциям связывания давали протекать в течение 45 мин перед центрифугированием и промыванием в бессывороточной среде Iscove. Клетки ресуспендировали в среде Iscove с 10% NHS. Клетки инкубировали в течение 60 мин при 37°С. В опытах, представленных на фигурах 18А-С, клетки промывали центрифугированием и ресуспендировали в среде для окрашивания, содержащей 0,5 мкг/мл пропидия йодида (PI, #P-16063, Molecular Probes, Eugene OR). Пробы инкубировали в течение 15 мин при комнатной температуре в темноте перед анализом проточной цитометрией с использованием цитометра FACsCalibur и программного обеспечения CellQuest (Becton Dickinson).
Более высокоочищенный пик димера TRU-016 является слабым медиатором CDC, когда используется один, что показано на фигуре 18А ровным характером кривой зависимости доза-эффект даже в высоких концентрациях. В результате того что реагенты с действием против CD20 представляют собой эффективные индукторы активности CDC, в опытах с комбинациями желательными были ненасыщающие количества реагентов против CD20, так чтобы в результате было возможным детектировать у реагентов синергизм. По данным этих первоначальных опытов обычная концентрация реагента, выбранная для опытов с комбинациями, составляла 0,5 мкг/мл или 2 мкг/мл TRU-016. Как правило, концентрация ритуксана составляла 0,04-0,06 мкг/мл ввиду его более высокой активности, проявленной в опытах по CDC с использованием одного реагента. В некоторых экспериментах концентрацию реагента CD20 сохраняли постоянной на уровне субоптимальной концентрации, в то время как концентрацию TRU-016 изменяли для установления минимальных количеств реагента с действием против CD37, необходимых для усиления CDC. Таким образом, один TRU-016 не мог опосредовать CDC, если только не присутствовали агрегаты.
На фигуре 18В приведен график процента живых клеток (PI негативных), установленного в указанных пределах титрования (0,06-0,5 мкг/мл), при использовании одного ритуксана или в комбинации с TRU-016 в концентрации 2,5 мкг/мл. Ритуксан при использовании в пределах субоптимальных концентраций для обеспечения гибели в качестве одного реагента проявлял более высокую активность к CDC в любой концентрации в комбинации с TRU-016 (фигура 18В). Удаление C1q из реакционных смесей приводило к полной элиминации активности CDC (фигура 3В).
Как показано на фигуре 18С, пробы также инкубировали с FITC анти-C1q в течение 45 мин на льду перед анализом проточной цитометрией. Окно возбуждения для лимфоцитов было на компрометированных клетках. Процент клеток в данном канале увеличивался с повышением концентрации ритуксана, и графически представлено относительное значение MFI для данной популяции клеток. На фигуре 18С приведены результаты эксперимента по CDC, в котором реагенты смешивали в следующих количествах: 0,5 мкг/мл TRU-016 и концентрации ритуксана находились в пределах от 0,06 мкг/мл до 0,5 мкг/мл, и клетки окрашивали PI перед проведением проточной цитометрии. Результаты показывают зависимое от дозы увеличение значения MFI с повышением концентраций ритуксана. Добавление димера TRU-016 приводило к дополнительному увеличению MFI при каждой концентрации ритуксана. Проводили аналогичную серию опытов, сохраняя концентрацию ритуксана постоянной и увеличивая концентрацию TRU-016. Вновь CDC активность была выше при внесении TRU-016 в реакционные смеси с ритуксаном, но увеличение концентрации TRU-016 до 2,5 мкг/мл с 0,5 мкг/мл не приводило к достоверному повышению опосредованной CDC гибели клеток в данных опытах (данные не представлены).
Белки ритуксана и TRU-016, использованные одни и в комбинации друг с другом, сравнивали по их ADCC активности в условиях in vitro с использованием таких же пределов концентраций, какие были в опытах по CDC. На фигуре 18D показаны результаты теста оценки ADCC с мечеными клетками-мишенями Ramos и человеческими эффекторными клетками РВМС при соотношении эффекторных клеток и клеток-мишеней, составляющем 25:1, с использованием TRU-016 или ритуксана одних или в комбинации друг с другом в указанных пределах концентраций. Аналогичные данные получали при соотношении эффекторных клеток и клеток-мишеней, составляющем 12,5:1. Оба, димер TRU-016 и ритуксан, опосредовали высокую ADCC в отношении клеток Ramos, экспрессирующих антигены-мишени CD20 и CD37; однако объединение двух реагентов не приводило к достоверному усилению гибели клеток.
Пример 9
TRU-016 индуцирует апоптоз В-клеток
Проводили опыты с целью изучения влияния TRU-016 на апоптоз В-клеток. Проводили первоначальные тесты с целью оценки влияния молекул TRU-016 на различные рецепторы В-клеток с использованием очищенного на протеине А вещества, которое по-прежнему содержало агрегаты более высокого порядка. После обработки в течение 24 ч антителами против CD37 или рекомбинантными молекулами TRU-016 наблюдали аналогичную картину усиленного апоптоза во многих опытах с использованием в качестве показателя активности апоптоза процент позитивных на аннексин V клеток и при их проведении на клетках Ramos и BJAB в качестве связывающих мишеней (данные не представлены).
На фигуре 19А показано, что апоптоз достоверно повышался после инкубации В-клеток с нефракционированным TRU-016. На фигуре 19А приведен точечный график распределения окрашивания клеток Ramos аннексином V-PI после инкубации в течение 24 ч c TRU-016 (10 мкг/мл). Процент дважды позитивных клеток на аннексин V-PI повышался с 11,3% от общей популяции до 32,8%, и процент негативных на аннексин V-PI клеток увеличивался с 8,5% до 19,7%, указывая на то, что апоптоз индуцируется после воздействия TRU-016. Аналогичные данные получали независимо от того, использовали клетки Ramos или BJAB в качестве связывающих мишеней в данных тестах.
Проводили дальнейшие опыты с целью изучения влияния TRU-016 на апоптоз В-клеток с использованием более высокоочищенного димера TRU-016 (фигура 19В). Пробы анализировали через 24 и 48 ч после начала реакций инкубации. Анализ окрашивания аннексином/PI проводили на нескольких типах клеток с использованием 20 мкг/мл белка TRU-016. В результате того что апоптоз снижался при использовании димера TRU-016, в реакционные смеси вносили 20 мкг/мл козьего античеловеческого IgG для перекрестного связывания реагентов на клеточной поверхности. Затем клетки окрашивали аннексином V-FITC и пропидием йодидом. Данные, приведенные на фигуре 19В, показывают, что пик димера TRU-016 индуцирует апоптоз клеток Daudi через 24-48 ч, но присутствие перекрестно-сшивающего реагента, такого как античеловеческий IgG, приводит к достоверному повышению интенсивности опосредуемого CD37 апоптоза.
Также проводили опыты с целью определения влияния TRU-016 на нормальные человеческие В-клетки в культуре с использованием РВМС. На фигурах 20А и 20В приведены результаты одного такого эксперимента, на них представлены графики в колонках, отражающие процент CD19 и CD40 позитивных лимфоцитов (В-клеток) в культурах РВМС, обработанных в течение 48-72 ч одной средой, TRU-016 или ритуксаном.
Человеческие РВМС выделяли из цельной крови центрифугированием в градиенте плотности среды для разделения лимфоцитов. Клетки инкубировали в течение 48 или 72 ч с ритуксаном или TRU-016 в концентрации 1 мкг/мл. Порцию инкубационной среды отбирали через 48 ч и вновь через 72 ч после начала опыта. РВМС промывали и инкубировали с меченными FITC антителами против CD19, меченными FITC антителами против CD40 или меченными FITC антителами против CD3 в течение 45 мин на льду. Затем регистрировали процент общего окрашивания лимфоцитов данными реагентами и сравнивали с пробами РВМС, подвергшимися инкубации в аналогичных условиях, но без тестируемых реагентов, и окрашенными как испытуемые пробы. На фигурах 20А и 20В приведены графики в колонках, показывающие фракцию от общей популяции лимфоцитов (%), которая давала позитивный FAC сигнал через 48 и 72 ч инкубации с указанными реагентами. На фигуре 20С представлены данные аналогичного опыта в графической форме, показывающие процентное уменьшение исходного количества лимфоцитов, экспрессирующих указанный антиген CD (т.е. позитивных на CD19, CD40 или CD3) после инкубации РВМС с TRU-016 (при концентрации 1 мкг/мл) в течение 24 и 72 ч.
В присутствии перекрестно-сшивающего реагента обработка димером TRU-016 или ритуксаном приводила к уменьшению процента В-лимфоцитов в культурах РВМС по данным позитивного окрашивания на CD19 и CD40. Несмотря на то что процент В-лимфоцитов в культуре был низким в начале эксперимента, совместное культивирование с ритуксаном или TRU-016 приводило к снижению количества CD19 и CD40 позитивных лимфоцитов в культуре РВМС примерно в 1,5-2 раза через 48 ч и более чем в 3 раза через 72 ч. Данная общая тенденция к истощению В-клеток через 48-72 ч была воспроизводимой во всех нормальных тестированных культурах РВМС, независимо от исходного первоначального процента В-лимфоцитов в данных культурах, который находился в пределах примерно от 3% до 7% от общего количества лимфоцитов, в зависимости от пробы.
На фигуре 20С приведен график в колонках, показывающий процентное снижение В-лимфоцитов по сравнению с Т-лимфоцитами в короткоживущих культурах РВМС, инкубированных с TRU-016 в течение 24-72 ч. Эти данные свидетельствуют о том, что TRU-016 способен приводить к специфическому истощению CD37 позитивных В-лимфоцитов из нормальных культур периферической крови, и что низкий уровень связывания TRU-016 с клетками, отличными от В-лимфоцитов (фигура 1С), недостаточен для опосредования эффективного истощения данных лимфоцитов из клеточной популяции.
Пример 10
Комбинации TRU-016 и ритуксимаба синергически уменьшают объем опухолей на модели опухолевых ксенотрансплантатов на мышах.
Проводили опыты на модели ксенотрансплантатов опухолей на мышах с целью оценки влияния комбинированной терапии с использованием голых мышей (Harlan) или клеток человеческих опухолевых линий Ramos и Daudi. Опухолевые клетки Ramos и Daudi культивировали в матрасах Т150 на среде IMDM/10% FBS до тех пор, пока они не достигали 80% слияния. Использовали 5 млн (5×106) клеток на мышь в качестве дозы при прививке опухолей. Клетки вводили подкожно в правый бок с использованием РBS в общем объеме 0,1 мл или 5×107 клеток/мл. Опухолям давали вырасти у голых мышей, и животных разделяли на группы, основываясь на размере/объеме опухолей. Для каждой опытной группы использовали 12 мышей со средним объемом опухолей, равным примерно 222 мм3 (пределы = 152-296 мм3). Также использовали мышей со значениями среднего объема опухолей в пределах 237-251 мм3. На 0, 2, 4, 6 и 8 сутки животным вводили внутривенно (в/в) один из следующих реагентов: TRU-016 GPC POI (интересующий пик), 200 мкг/мышь; ритуксан, 200 мкг/мышь; или человеческий IgG (контроль), 200 или 400 мкг/мышь, в виде одних реагентов или в виде следующих комбинаций реагентов: ритуксан+TRU-016 в дозе 100 мкг на мышь или ритуксан+TRU-016 в дозе 200 мкг на мышь. Ежедневно определяли объем опухолей с использованием штангенциркуля до окончания эксперимента (убоя животных или регрессии заболевания). Строили график зависимости объема опухолей в виде функции времени обработки для каждого животного и также вычисляли среднее значение для каждой группы.
Аналогичные опыты также проводили с использованием опухолей меньшего размера на мышах, распределенных на группы, с меньшим средним объемом опухолей в пределах 153-158 мм3 и с опухолями большего размера, но с использованием в основном клеток Daudi, а не Ramos. Данные исследования проводили в аккредитованной лаборатории AAALAC и в рамках правил работы на животных согласно указаниям Institutional Animal Care and Use Committee (IACUC).
На фигуре 21 приведены графики с данными по эффективности TRU-016 в сравнении с huIgG, ритуксаном и комбинациями в дозах 100 и 200 мкг на мышь, представленными в виде средних значений по группе из 12 животных. Строили график зависимости объема опухолей в виде функции времени после обработки в/в инъекцией(и). Среднее значение объема опухолей после обработки TRU-016 было ниже по сравнению с наблюдаемым в группе отрицательного контроля (huIgG). В случае построения графика % выживаемости или % животных без опухолей комбинированная терапия в более высокой дозе проявляла более высокую противоопухолевую активность на данной модели опухолей в условиях in vivo. Однако комбинированная терапия в более низкой дозе (100 мкг на мышь) не была столь эффективной, как каждый один реагент в более высокой дозе.
Эти данные свидетельствуют о том, что лечение TRU-016 при использовании в комбинации с ритуксаном в соответствующих дозах будет более эффективным при лечении пациентов с опухолями по сравнению с терапией одним ритуксаном.
Пример 11
TRU-016 уменьшает объем опухолей и повышает выживаемость на модели опухолевых ксенотрансплантатов на мышах
Проводили опыты на модели ксенотрансплантатов опухолей с использованием голых мышей (Harlan) или клеток человеческих опухолевых линий Ramos и Daudi. Проводили три разных опыта, основываясь на типе опухолей и размере опухолей на время обработки TRU-016 или другим тестируемым реагентом. Опухолевые клетки Ramos и Daudi культивировали, и 5×106 клеток вводили подкожно в правый бок для прививки опухолей каждой опытной мыши. Опухолям давали вырасти у голых мышей, и животных разделяли на группы, основываясь на размере/объеме опухолей. В первом опыте для каждой опытной группы использовали 12 мышей со средним объемом опухолей, равным 155-237 мм3. На 0, 2, 4, 6 и 8 сутки животным вводили внутривенно (в/в) один из следующих реагентов: ритуксимаб, 200 мкг/мышь; TRU-016 GPC пик димера, 200 мкг/мышь; или человеческий IgG (контроль), 400 мкг/мышь. Ежедневно определяли объем опухолей с использованием штангенциркуля до окончания эксперимента (убоя животных или регрессии заболевания). Строили график зависимости объема опухолей в виде функции времени обработки для каждого животного и также вычисляли среднее значение для каждой группы. Средние значения по группе приведены на фигуре 22А, в то время как на фигуре 22В приведен сравнительный анализ данных по проценту выживаемости для каждой группы мышей в виде функции времени.
На фигуре 22А приведены данные по эффективности TRU-016 в сравнении с huIgG и ритуксаном на модели опухолей Ramos в виде средних значений по каждой группе из 12 животных. Строили график зависимости объема опухолей в виде функции времени после обработки в/в инъекцией(и). Среднее значение объема опухолей после обработки TRU-016 было ниже по сравнению с наблюдаемым в группе отрицательного контроля (huIgG). На фигуре 22В приведены графические данные в виде кривых выживаемости для различных опытных групп при сравнении huIgG, ритуксана и TRU-016. Введение TRU-016, при использовании более агрессивной модели опухолей Ramos с повышенным фоновым объемом опухолей, приводило к подавлению скорости роста опухолей по сравнению с человеческим IgG (данные не представлены). Введение TRU-016 мышам с более мелкими опухолями Ramos приводило к подавлению роста опухолей и повышенной средней выживаемости мышей.
Пример 12
TRU-016 не оказывает влияния на CDC активность в отношении других поверхностных рецепторов В-клеток
Для определения того, усиливает ли молекула TRU-016 активность CDC после обработки антителами к другим поверхностным рецепторам В-клеток, в дополнение к CD20, таким как MHCII, CD19, CD80/86 и CD40, проводили серию экспериментов, аналогичную только что описанной для комбинаций с действием против CD20-CD37.
Клетки Ramos вносили в лунки с полной средой Iscove с 10% FBS. Добавляли MAb (реагент В: антитела против CD19 HD37, реагент С: антитела против CD45 9.4), рекомбинантный белок (реагент D: CTLA-4 muIg-IgG2a, Ancell #501-820) и асцитную жидкость (реагент А: антитела против MHCII НВ10а) в указанных разведениях (смотри фигуру 23) и проводили две параллельные реакции с и без ритуксимаба (в концентрации 0,05 мкг/мл) или TRU-016 (в концентрации 2 мкг/мл). Реакционные смеси инкубировали в течение 30 мин при 37°С. Клетки промывали и вносили NHS до конечной концентрации 10% в бессывороточной среде. Клетки инкубировали в течение 90 мин при 37°C с источником комплемента. Клетки промывали, добавляли пропидий йодид до конечной концентрации 0,5 мкг/мл в PBS; клетки инкубировали в темноте при комнатной температуре в течение 15 мин; и затем клетки анализировали проточной цитометрией. Каждый график на панели A-D фигуры 23 отражает % позитивных на PI клеток в указанных пределах титрования.
В целом, данные свидетельствуют о том, что достоверное различие в уровне CDC активности отсутствовало как при использовании одних антител против данных рецепторов, так и в комбинации с TRU-016 (фигура 23A-D). Возможно, было небольшое повышение уровня CDC для реагентов с действием против CD19 и CD45 при использовании TRU-016 в субоптимальных концентрациях. Однако различия в значениях показателей CDC не были достоверными, как наблюдаемые в случае комбинации CD20-CD37. В дополнение к усилению CDC при использовании реагентов против CD20 и CD37 в комбинации, имело место усиление гибели клеток при использовании комбинаций антитела против classII (НВ10а), антитела против CD19, антитела против CD45 (9.4) или CTLA4Ig с ритуксаном в субоптимальной концентрации.
Пример 13
TRU-016 не усиливает CDC активность в отношении других рецепторов-мишеней, включая MHCII, CD19, CD80/86 и CD40
Для определения того, усиливает ли молекула TRU-016 активность CDC после обработки антителами к другим поверхностным рецепторам В-клеток, в дополнение к CD20, проводили серию экспериментов, аналогичную описанной для комбинаций с действием против CD20-CD37 (смотри пример 8). Результаты данных опытов представлены на фигуре 23. В целом, достоверное различие в CDC активности отсутствовало при использовании одних антител против данных рецепторов или в комбинации с TRU-016. Уровень CDC увеличивался в ответ на реагенты с направленным действием против CD19 и CD45 при использовании TRU-016 в субоптимальных концентрациях. Однако различия в значениях показателей CDC не были достоверными, как наблюдаемые в случае комбинации CD20-CD37 (смотри пример 8). В дополнение к усилению CDC при использовании реагентов против CD20 и CD37 в комбинации, имело место усиление гибели клеток при использовании комбинаций антитела против МНСII (НВ10а), антитела против CD19, антитела против CD45 (9.4) или CTLA4Ig с ритуксаном в субоптимальной концентрации.
Пример 14
TRU-016 увеличивает выживаемость на модели опухолевых ксенотрансплантатов на мышах
Помимо опытов на модели ксенотрансплантатов опухолей, описанных в примере 11, проводили исследования с целью оценки эффективности TRU-016 в длительном увеличении выживаемости голых мышей (Harlan) или клеток человеческих опухолевых линий Ramos и Daudi.
Опухолевые клетки Ramos и Daudi культивировали, и 5×106 клеток вводили подкожно в правый бок мышей для инициации развития опухолевых ксенотрансплантатов у мышей. После развития опухолей мышей разделяли на группы, основываясь на размере/объеме опухолей (0 сутки). На 0, 2, 4, 6 и 8 сутки животным вводили внутривенно (в/в) один из следующих реагентов: ритуксимаб, 200 мкг/мышь; TRU-016, 200 мкг/мышь; ритуксимаб+TRU-016 в дозах 100 или 200 мкг/мышь; или человеческий IgG (контроль), 400 мкг/мышь. Объем опухолей определяли «вслепую» три раза в неделю с помощью штангенциркуля до окончания эксперимента (убоя животных или регрессии заболевания). Строили график зависимости объема опухолей в виде функции времени обработки для каждого животного и также вычисляли среднее значение для каждой группы. На фигуре 24 показан процент выживаемости мышей с опухолями Ramos (период наблюдений до 90 суток) после обработки TRU-016, ритуксимабом или их комбинацией. Комбинированная обработка TRU-016+ритуксимаб приводила к увеличению среднего значения времени выживания по сравнению с обработкой одним реагентом. На фигурах 25 и 26 представлен процент выживания мышей с опухолями Daudi (период наблюдений до 90 суток) после обработки TRU-016 или ритуксимабом. Обработка TRU-016 увеличивала среднее время выживания мышей с развившимися опухолями Daudi (фигура 25). TRU-016 был более эффективным по сравнению с ритуксимабом в отношении увеличения выживаемости мышей с опухолями Daudi (фигура 26).
При введении TRU-016 в виде одного реагента мышам с развившимися опухолями Ramos наблюдали подавление роста опухолей и увеличение времени выживаемости, аналогичные показатели отмечали при введении одного ритуксимаба, и они были выше по сравнению с контрольными мышами, обработанными huIgG. Объединенные данные 3 опытов показали, что комбинированная терапия TRU-016 и ритуксимабом приводила к статистически достоверному увеличению выживаемости по сравнению с монотерапией TRU-016 (р=0,028) или ритуксимабом (р=0,045). Полную регрессию опухолей также наблюдали в группах с комбинированным введением TRU-016 и ритуксимаба. У 45% мышей в группе, получавшей комбинированное лечение TRU-016+ритуксимаб в дозе 200 мкг, наблюдали длительную полную регрессию опухолей по сравнению с показателем 20% регрессии опухолей у мышей, обработанных одним TRU-016 или ритуксимабом (смотри таблица 3 и фигура 24).
Выживаемость после лечения развившихся опухолей Ramos
(200 мкг)
(100 мкг)
Подавление роста опухолей и увеличение выживаемости было установлено после обработки TRU-016 на модели опухолевых ксенотрансплантатов Daudi (смотри таблицу 4 и фигуры 25 и 26). Введение TRU-016 достоверно увеличивало выживаемость по сравнению с контрольной группой. При лечении SMIP TRU-016 также наблюдали повышение процента мышей без опухолей на данной модели по сравнению с животными контрольной группы и обработанными ритуксимабом.
Выживаемость после лечения развившихся опухолей Daudi
Лечение SMIP с действием против CD37 (TRU-016) было таким же эффективным, как и монотерапия ритуксимабом в отношении уменьшения объема опухолей и увеличения выживаемости на модели опухолевых ксенотрансплантатов Ramos. Было показано, что комбинированная обработка TRU-016+ритуксимаб оказывала повышенный лечебный эффект, выражающийся в уменьшении объема опухолей и достоверном увеличении выживаемости по сравнению с монотерапией ритуксимабом или TRU-016 на модели опухолевых ксенотрансплантатов Ramos. На модели опухолевых ксенотрансплантатов Daudi у мышей, обработанных TRU-016, наблюдали статистически достоверное увеличение среднего времени выживания по сравнению с контрольными животными после обработки HuIgG. Обработка ритуксимабом не повышала показатель выживаемости по сравнению с контрольными мышами. Эти данные подчеркивают эффективность терапии с направленным действием против CD37 на модели данных ксенотрансплантатов NHL.
Пример 15
TRU-016 усиливает индуцированную флударабином клеточную гибель на клетках CLL в условиях in vitro
Флударабин представляет собой химиотерапевтический препарат, используемый для лечения злокачественных заболеваний крови. Флударабин представляет собой аналог пуринов, который подавляет синтез ДНК, оказывая отрицательное влияние на рибонуклеотидредуктазу и ДНК-полимеразу. Флударабин является активным в отношении делящихся и покоящихся клеток. Флударабин является высокоэффективным для лечения хронической лимфоцитарной лейкемии (CLL), приводя к проявлению более сильных ответных реакций по сравнению с алкилирующими агентами, такими как один хлорамбуцил (Rai et al., N. Engl. J. Med. 343: 1750-1757, 2000). Флударабин применяется в различных комбинациях с циклофосфамидом, митоксантроном, дексаметазоном и ритуксимабом при лечении вялотекущей лимфомы и не-ходжкинской лимфомы. Однако при лечении флударабином наблюдается развитие резистентности. Флударабин вызывает зависимый от каспазы апоптоз клеток CLL, и оказалось, что апоптоз, опосредуемый TRU-016, не зависит от активации каспазы. Настоящее исследование было предпринято с целью изучения влияния TRU-016 в сочетании с флударабином на клетки CLL.
Клетки обрабатывали TRU-016 в концентрациях в пределах 0,1-100 мкг/мл и флударабином в концентрациях в пределах 0-20 мкМ (смотри фигуру 27). TRU-016 был предоставлен Trubion Pharmaceuticals (Seattle, WA). Флударабин (F-araA) получали от Sigma (St. Lois, MO). Среда RPMI 1640 была производства Invitrogen (Carlsbad, CA). Меченный флуоресцеином изотиоцианатом (FITC) аннексин V и пропидий йодид (PI) получали от BD Pharmingen, San Diego, CA. [3-(4,5-Диметилтиазол-2-ил)2,5-дифенилтетразолий бромид] (МТТ) получали от Sigma (St. Lois, MO). Клетки В-CLL выделяли сразу же после отбора с использованием центрифугирования в градиенте плотности фиколла (Ficoll-Paque Plus, Amersham Biosciences, Piscataway, NJ). Выделенные мононуклеарные клетки инкубировали в среде RPMI 1640 с добавлением 10% инактивированной нагреванием фетальной бычьей сыворотки (FBS, Hyclone Laboratories, Logan, UT), 2 мМ L-глутамина (Invitrogen, Carlsbad, CA) и пенициллина (100 Ед./мл)/стрептомицина (100 мкг/мл; Sigma-Aldrich, St. Louis) при 37°С в атмосфере 5% СО2. Только что выделенные клетки В-CLL использовали во всех описанных здесь опытах за исключением поверхностного окрашивания. Для проб с содержанием менее чем 90% В-клеток применяли негативный отбор с истощением клеток, отличных от В-клеток, с использованием набора для выделения В-клеток II (Miltenyi Biotec, Auburn, CA) или набора «Rosette-Sep» производства Stem Cell Technologies (Vancouver, British Columbia, Canada), следуя предложенному изготовителем протоколу. Клеточную линию Raji (линия клеток человеческой лимфомы Беркитта) получали из АТСС и поддерживали в среде RPMI 1640, содержащей 10% FBS, с добавлением пенициллина, стрептомицина и глутамина. Клетки подвергались расщеплению в соотношении 1:3, когда плотность клеток достигала значения 1×106 клеток/мл. Среду заменяли за ночь до каждого исследования для гарантии использования свежих клеток.
Клетки обрабатывали в условиях in vitro, как здесь описано. Готовили серийные разведения флударабина 1:4 (44, 11, 2,8, 0,7, 0,17 и 0,04 мкМ) в 6-луночном планшете при переносе 2 мл содержащей препарат среды в каждую следующую лунку, содержащую 6 мл одной среды. В отдельном 6-луночном планшете готовили серийные разведения TRU-016 1:4 (44, 11, 2,8, 0,7, 0,17 и 0,04 мкг/мл) на среде с использованием того же метода разведения. Из каждого планшета переносили по 0,45 мл среды в соответствующую лунку в 48-луночном планшете для получения смешанного раствора препаратов в среде (в целом 0,9 мл в каждой лунке). Затем к 0,9 мл среды в каждой лунке добавляли клетки CLL с титром 1×107 клеток/мл (0,1 мл) с получением конечного титра, равного 1×106 клеток/мл. Для клеток Raji конечный титр составил 5×104 клеток/мл. Таким образом, используемая клеточная суспензия имела титр 5×105 клеток/мл. Для постановки МТТ тестов готовили серийные разведения в 96-луночных планшетах и переносили в другие 96-луночные планшеты для инкубации с клетками. Общий объем при инкубации составлял 200 мкл (90 мкл раствора флударабина, 90 мкл раствора TRU-016 и 20 мкл клеточной суспензии). Жизнеспособность клеток оценивали с использованием МТТ теста при экспозиции 48 ч, и апоптоз оценивали с использованием окрашивания аннексином V/PI - на 24 ч.
МТТ тест проводили для оценки жизнеспособности клеток, как здесь описано. Вкратце, 106 клеток CLL высевали в 96-луночные планшеты. Клетки инкубировали в течение 48 ч. В каждую лунку вносили 50 мкл рабочего раствора для постановки МТТ теста (2 мг/мл, приготовлен из реагента для МТТ с концентрацией 5 мг/мл при смешивании со средой RPMI 1640 в соотношении 2:3 об./об.) и клетки инкубировали в течение 8 ч. Планшеты центрифугировали и супернатант удаляли, и остаток растворяли в 100 мкл лизирующего раствора. Определяли поглощение проб на ридере для планшетов при 540 нм. Жизнеспособность клеток выражали в виде процента жизнеспособности по сравнению с контрольной средой.
Апоптоз клеток CLL после инкубации с антителами оценивали с использованием окрашивания аннексином V-FITC/пропидием йодидом (PI) при анализе FACS. 5×105 клеток в 200 мкл 1× буфера для связывания (BD Pharmingen) окрашивали 5 мкл аннексина V (BD Pharmingen) и 5 мкл PI (BD Pharmingen) и выдержали в темноте при комнатной температуре в течение 15 мин перед приготовлением суспензии в 300 мкл 1× буфера и анализировали проточной цитометрией. Готовили клетки без окрашивания, клетки, окрашенные только аннексином V, и клетки, окрашенные только PI. При проведении всех экспериментов с проточной цитометрией проводили анализ FACs с использованием цитометра Beckman-Coulter EPICS XL (Beckman-Coulter, Miami, FL). Флуорофоры подвергались возбуждению при 488 нм. Интенсивность FITC-флуоресценции определяли с FL1, в то время как флуоресценцию PI и РЕ определяли с FL3. Использовали программное обеспечение системы II (Beckman-Coulter) для обработки данных. Определенное количество клеток выражали в пересчете на 10000 для каждой пробы.
Синергическое действие определяли при использовании изоболограмного метода. Для установления синергического эффекта проводили сравнение действия комбинации препаратов с действием каждого одного препарата. В этих целях использовали уравнение: Са/Са,b+Cb/Cb,a=CI, где Са и Cb представляют, соответственно, концентрации лекарственного препарата А и лекарственного препарата В, при которых воспроизводится желаемый эффект (например, 50% гибель клеток). Са,b и Cb,a представляют, соответственно, концентрации лекарственного препарата А и лекарственного препарата B в комбинации, при которых воспроизводится желаемый эффект. CI представляет индекс комбинации. Определяли концентрации флударабина и TRU-016, которые вызывали 50% гибель клеток (IC50), и они приведены на фигуре 27С [IC50 флударабина (I) и IC50 TRU-016 (II)]. Прямая линия между двумя данными точками на осях представляет собой линию аддитивного эффекта. Затем были определены различные комбинации флударабина и TRU-016, которые обеспечивали 50% гибель клеток, по оценке жизнеспособности, и строили такой же график зависимости. В том случае, когда точки были ниже линии аддитивного действия, это указывало на наличие синергического эффекта. Когда точки находились выше линии, то это указывало на наличие антагонизма. Когда точки находились на линии, это указывало на аддитивное действие.
На фигуре 27 показано, что TRU-016 эффективно снижает относительную жизнеспособность клеток, обработанных флударабином, т.е. потенцирует цитотоксическое действие одного флударабина. Таким образом, результаты данного опыта свидетельствуют, что TRU-016 можно вводить совместно с флударабином, приводя к повышению эффективности (т.е. синергическому снижению клеток CLL) при лечении злокачественных заболеваний крови.
Пример 16
TRU-016 индуцирует прямой цитотоксический эффект для резистентных к ритуксимабу клеток
Как здесь уже указывалось, ритуксимаб представляет собой моноклональное антитело, используемое для лечения NHL, FCC, MCL, DLCL, SLL и CLL. Настоящее исследование было проведено с целью оценки эффективности TRU-016 в индукции прямого цитотоксического эффекта для клеток, резистентных к ритуксимабу.
Резистентные к ритуксимабу клетки (1×106 клеток) (Raji 4RH RL 4RH, любезно предоставленные Dr. S. Czuczman, Roswell Park Cancer Institute, Buffalo, NY) обрабатывали герцептином (10 мкг/мл), ритуксимабом (10 мкг/мл) или TRU-016 (5 мкг/мл) в присутствии 5-кратного избытка козьего античеловеческого IgG в течение 24 ч. Прямой цитотоксический эффект оценивали окрашиванием аннексином/PI и определяли жизнеспособность (в процентах) по отношению к контрольным клетками (клеткам, обработанным герцептином).
TRU-016 индуцировал более высокую токсичность для клеток по сравнению с ритуксимабом на резистентных к ритуксимабу клеточных линиях (смотри фигуру 28). Таким образом, TRU-016 представляет собой эффективное средство для индукции цитотоксичности для резистентных к ритуксимабу клеток, что делает его пригодным для применения в качестве лекарственного препарата для лечения заболеваний, для которых характерны или которые связаны с резистентными к ритуксимабу клетками, такими как В-клетки.
Пример 17
TRU-016 индуцирует фосфорилирование тирозина в CD19+ В-клеток первичной CLL
Для определения того, как TRU-016 индуцирует передачу сигналов в В-клетках, проводили эксперименты с целью исследования влияния TRU-016 на фосфорилирование тирозина.
Только что выделенные CD19+ клетки (≈50-100×106) от пациентов с CLL суспендировали при титре 5×106 клеток/мл PBS. Затем клетки инкубировали в течение 10 мин при 37°С в атмосфере 5% СО2 с контролем, трастузумабом (герцептином) или TRU-016 с конечной концентрацией 5 мкг/мл. Клетки центрифугировали, супернатант удаляли и клетки ресуспендировали в свежей порции PBS в первоначальном объеме. Добавляли перекрестно-связывающий линкер козьего античеловеческого Fc-фрагмента (25 мкг/мл), и клетки инкубировали еще в течение 5 мин. Клетки вновь центрифугировали, супернатант удаляли и клетки лизировали в 1 мл лизирующего буфера RIPA с ингибиторами протеазы и фосфатазы (10 мМ Трис, рН 7,4, 150 мМ NaCl, 1% Тритона Х-100, 1% дезоксихолевой кислоты, 0,1% SDS и 5 мМ ЭДТА, все реагенты - в конечных концентрациях). Использовали смесь ингибиторов протеазы производства Sigma, номер по каталогу #Р-8340; смесь ингибиторов фосфатазы производства Sigma: смесь ингибиторов серин/треонинфосфатазы, номер по каталогу #Р-2850; и ингибитор тирозинфосфатазы, номер по каталогу #Р-5726; PMSF (100 мМ). Ингибиторы добавляли в лизирующий буфер непосредственно перед применением в разведении 1:100. Концентрацию белка в лизатах количественно определяли методом с бицинхониновой кислотой (BCA) (Pierce, Rockford, IL). Контрольные и опытные пробы белков (50 мкг общего белка) разделяли двумерным электрофорезом в геле (рН в пределах 3-10) (1 направление) и 10% SDS-PAGE (2 направление). Белок переносили на нитроцеллюлозные мембраны 0,2 Nm (Schleicher & Schuell, Keene, NH) и подвергали иммуноблоттингу с использованием клона 4G10 антител против фосфотирозина (Upstate Biotechnology), следуя стандартному протоколу. В качестве вторичных антител использовали конъюгированный с пероксидазой из хрена (HRP) козий антикроличий IgG. Детектирование фосфорилированного белка проводили с использованием хемилюминесцентного субстрата (SuperSignal, Pierce Inc. Rockford, IL).
TRU-016 индуцировал фосфорилирование тирозина в CD19+ В-клетках первичной CLL, как это было показано двумерным электрофоретическим анализом в геле (смотри фигуру 29). Таким образом, данные опыты показывают, что на одном пути TRU-016 функционирует посредством фосфорилирования тирозина.
Пример 18
Гуманизированные молекулы TRU-016
Как представлено в примере 1, CD37-специфические SMIP (такие как TRU-016) описаны в совместной заявке на патент США № 10/627556 и публикациях заявок на патенты США №№ 2003/133939, 2003/0118592 и 2005/0136049. Эти документы включены здесь ссылкой. Примерный CD37-специфический SMIP, полипептид TRU-016 (SEQ ID NO:2) получен и описан здесь. В настоящем примере описывается гуманизированный TRU-016 SMIP.
Гуманизированные антитела известны в данной области, и они обсуждаются в публикации заявки на патент США № 2006/0153837. В настоящей заявке применяются способы, использованные при получении гуманизированного антитела (обсуждается ниже) для гуманизации SMIP, и в частности для гуманизации TRU-016.
Полагается, что «гуманизация» обеспечивает антитело, которое в меньшей степени является иммуногенным, с полным сохранением антигенсвязывающих свойств исходной молекулы. Для сохранения всех антигенсвязывающих свойств исходного антитела структура его антигенсвязывающего сайта должна быть воспроизведена в «гуманизированном» варианте. Этого можно достичь «пересадкой» только CDR, отличных от человеческих, в человеческие вариабельные каркасные области и константные области, с или без сохранения критических остатков каркасной области (Jones et al., Nature 321: 522 (1986); Verhoeyen et al., Science 239: 1539 (1988)) или рекомбинацией целых вариабельных областей, отличных от человеческих (для сохранения лигандсвязывающих свойств), но «маскировкой» их поверхностью, подобной человеческой, посредством разумной замены подвергающихся воздействию остатков (для снижения антигенности) (Padlan, Molec. Immunol. 28: 489 (1991)).
В основном гуманизация «пересадкой» CDR включает рекомбинацию только CDR антитела, отличного от человеческого, в человеческую вариабельную каркасную область и человеческую константную область. Теоретически это должно привести к существенному снижению или элиминации иммуногенности (за исключением случаев, когда имеются аллотипические или идиотипические различия). Однако сообщалось, что также может быть необходимым сохранить некоторые остатки каркасной области исходного антитела (Reichmann et al., Nature, 332: 323 (1988); Queen et al., Proc. Natl. Acad. Sci. USA, 86: 10029 (1989)).
Остатки каркасной области, которые необходимо сохранить, можно установить при использовании компьютерного моделирования. Альтернативно критические остатки каркасной области можно потенциально идентифицировать при сравнении структур известных антигенсвязывающих сайтов (Padlan, Molec. Immunol. 31(3): 169-217 (1994)), этот источник включен здесь ссылкой.
Остатки, которые потенциально оказывают влияние на связывание антигена, можно подразделить на несколько групп. В первую группу входят остатки, расположенные смежно с поверхностью сайта антигена, которые, следовательно, могут прямо контактировать с антигенами. Данные остатки включают аминоконцевые остатки и смежные с CDR. Вторая группа включает остатки, которые могут изменять структуру или относительное расположение CDR посредством контактирования с CDR или другой пептидной цепью в антителе. Третья группа включает аминокислоты со «скрытыми» боковыми цепями, которые могут оказывать влияние на структурную целостность вариабельных областей. Остатки в данных группах, как правило, находятся в одних и тех же положениях (Padlan, 1994, выше), хотя их положения при идентификации могут различаться в зависимости от системы нумерации (смотри Kabat et al., «Sequences of proteins of immunological interest», 5th ed., Pub. No. 91-3242, US Dept. Health & Human Services, NIH, Bethesda, Md., 1991).
Несмотря на то что настоящее изобретение относится к гуманизации SMIP, а не антител, сведения о гуманизированных антителах в данной области применимы к SMIP по изобретению. Некоторые примеры гуманизированных молекул TRU-016 представлены в таблице 5 ниже.
Для получения гуманизированных конструкций TRU-016 по изобретению мышиные каркасные области TRU-016 сопоставляли с остатками человеческих VH1 и VH5 каркасной области тяжелой цепи и VК1 и VК3 легкой цепи. Наилучшие совпадения анализировали на совместимость каркасной области с CDR мышиных вариабельных областей. Несмотря на наличие нескольких в равной степени совместимых комбинаций, из которых можно было сделать выбор, авторы с успехом использовали ранее комбинацию VК3 (Х01668), VН5-5(Z12373), и таким образом гуманизированные анти-CD37-SMIP конструировали с использованием данных человеческих каркасных областей, соединенных линкером 15aa Gly4Ser((g4s)3) scFv. Конструкцию VК3 конструировали с JK1 в качестве предпочтительного совпадения FR4, и VH5 конструировали с JН1, кодирующей FR4, в виде ранее описанных конструкций. SMIP конструировали de novo с использованием перекрывающейся олигонуклеотидной ПЦР. Полноразмерные продукты клонировали в экспрессирующий SMIP вектор в рамке считывания с шарнирной областью человеческого IgG1, СН2 и СН3. Данные клоны секвенировали, переносили в клетки COS-7, и 3-суточную кондиционированную среду тестировали на связывание с В-клетками лимфомы Ramos. В целях повышения эффективности гуманизации были введены изменения в CDR1 легкой цепи в положениях L25, L27 и L28, и они хорошо переносились, проявляя равную связывающую активность с исходной гуманизированной молекулой 019001. Были получены дополнительные конструкции ДНК аналогичным способом с изменением CDR3 области VH включением аминокислот зародышевой линии, Н100-Н102, кодированной различными человеческими областями JH. Конструкции тестировали в отношении уровня экспрессии и степени связывания с CD37 на клетках Ramos.
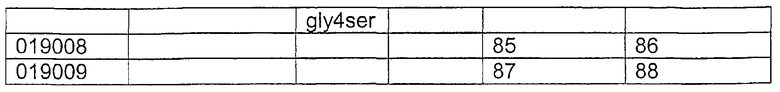
Аминокислотные консенсусные последовательности гуманизированной конструкции TRU-016 No.019001 (SEQ ID NO:6; Н016-019001) и негуманизированного TRU-016 (SEQ ID NO:2; 016-G28-1) представлены с использованием системы нумерации Kabat на фигуре 30А. На фигуре 30В приведено сопоставление аминокислотных последовательностей гуманизированной конструкции TRU-016 No.019001 (SEQ ID NO:6), 019008 (SEQ ID NO:86) и 019009 (SEQ ID NO:88).
Сопоставление последовательностей ДНК и аминокислотных последовательностей трех гуманизированных конструкций TRU-016 (019001, 019041 и 019044) с демонстрацией высокого CD37-специфического связывания с В-клетками Ramos приведено на фигуре 31.
Сопоставление последовательностей ДНК и аминокислотных последовательностей в формате FASTA тех же трех гуманизированных конструкций TRU-016 (019001, 019041 и 019044) приведено на фигуре 32.
Дополнительные шарнирные области (таблица 6) и каркасные области (таблица 7), которые можно использовать в гуманизированных молекулах TRU-016 по изобретению, приведены ниже.

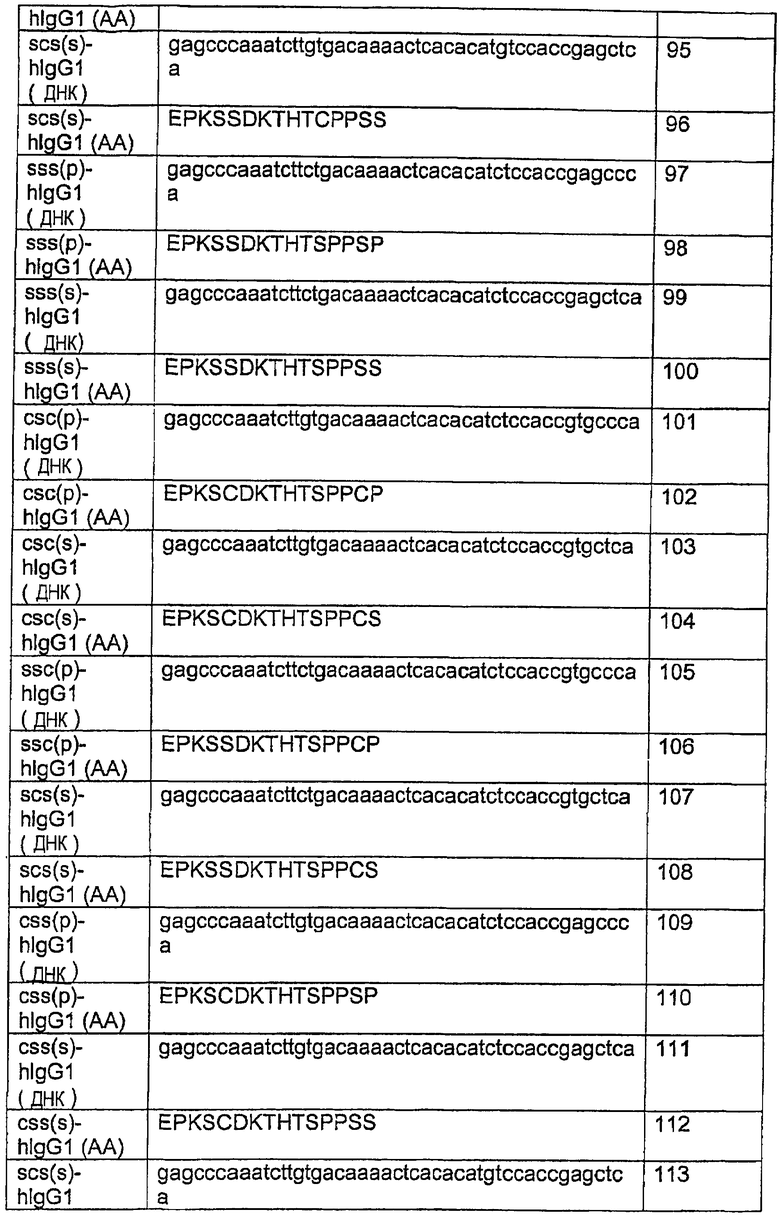

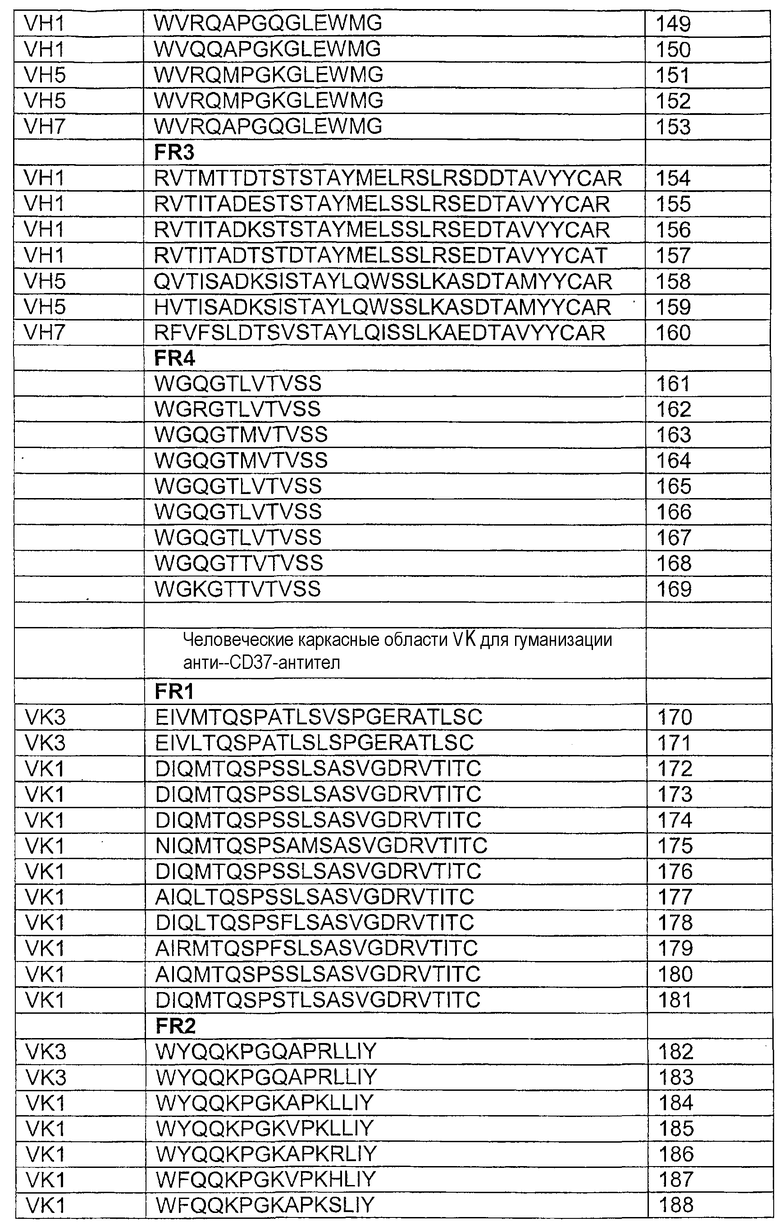

Специалистам в данной области очевидно понятно, что предполагаются многочисленные модификации и вариации в изобретении, как приведенные выше в иллюстративных примерах. Следовательно, в отношении изобретения могут быть поставлены только ограничения, приведенные в прилагаемой формуле изобретения.
Последовательности ДНК и аминокислотные последовательности SEQ ID NO:79-88





| название | год | авторы | номер документа |
|---|---|---|---|
| CD37-СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ИММУНОКОНЪЮГАТЫ | 2011 |
|
RU2610662C9 |
| ПРИМЕНЕНИЕ ОДНОКРАТНОЙ ДОЗЫ CD20-СПЕЦИФИЧЕСКИХ СВЯЗЫВАЮЩИХ МОЛЕКУЛ | 2006 |
|
RU2421242C2 |
| СВЯЗЫВАЮЩЕЕСЯ С CD37 ИММУНОТЕРАПЕВТИЧЕСКОЕ СРЕДСТВО И ЕГО КОМБИНАЦИЯ С БИФУНКЦИОНАЛЬНЫМ ХИМИОТЕРАПЕВТИЧЕСКИМ СРЕДСТВОМ | 2009 |
|
RU2531754C2 |
| ОДНОЦЕПОЧЕЧНЫЕ МУЛЬТИВАЛЕНТНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ЭФФЕКТОРНОЙ ФУНКЦИЕЙ | 2007 |
|
RU2487888C2 |
| CD37-ИММУНОТЕРАПЕВТИЧЕСКАЯ КОМБИНИРОВАННАЯ ТЕРАПИЯ И ЕЕ ПРИМЕНЕНИЯ | 2009 |
|
RU2526156C2 |
| CD37-СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ИММУНОКОНЪЮГАТЫ | 2011 |
|
RU2742743C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ СПЕЦИФИЧЕСКИЙ МЕМБРАННЫЙ АНТИГЕН ПРОСТАТЫ, И СООТВЕТСТВУЮЩИЕ КОМПОЗИЦИИ И СПОСОБЫ | 2012 |
|
RU2632647C2 |
| ХИМЕРНОЕ ТЕРАПЕВТИЧЕСКОЕ АНТИТЕЛО НН1 ПРОТИВ CD-37 | 2012 |
|
RU2658438C2 |
| КОМБИНАЦИЯ СЛИТОГО БЕЛКА АНТИТЕЛО ПРОТИВ EDb ФИБРОНЕКТИНА-IL-2 И МОЛЕКУЛЫ, СВЯЗЫВАЮЩЕЙСЯ С В-КЛЕТКАМИ, ПРЕДШЕСТВЕННИКАМИ В-КЛЕТОК И/ИЛИ ИХ ЗЛОКАЧЕСТВЕННЫМ АНАЛОГОМ | 2008 |
|
RU2484845C2 |
| СПОСОБ ЛЕЧЕНИЯ ВАСКУЛИТА | 2005 |
|
RU2411956C2 |






































































































































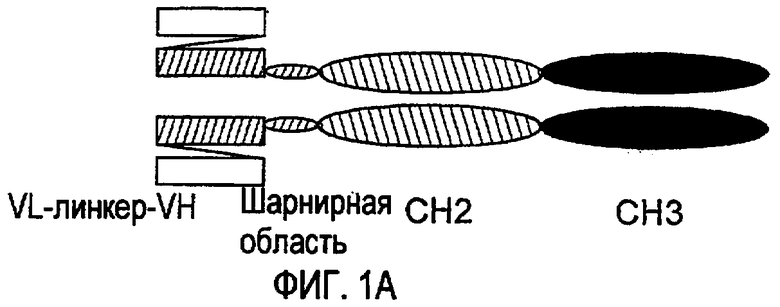
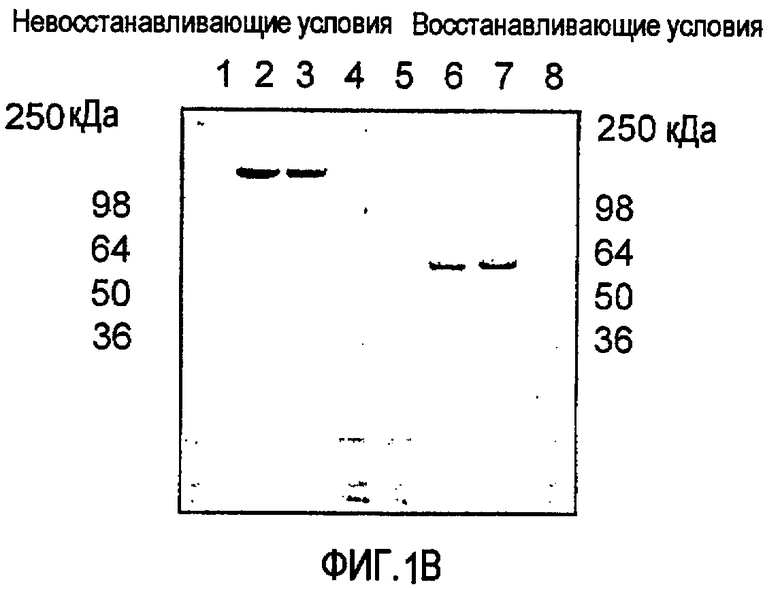
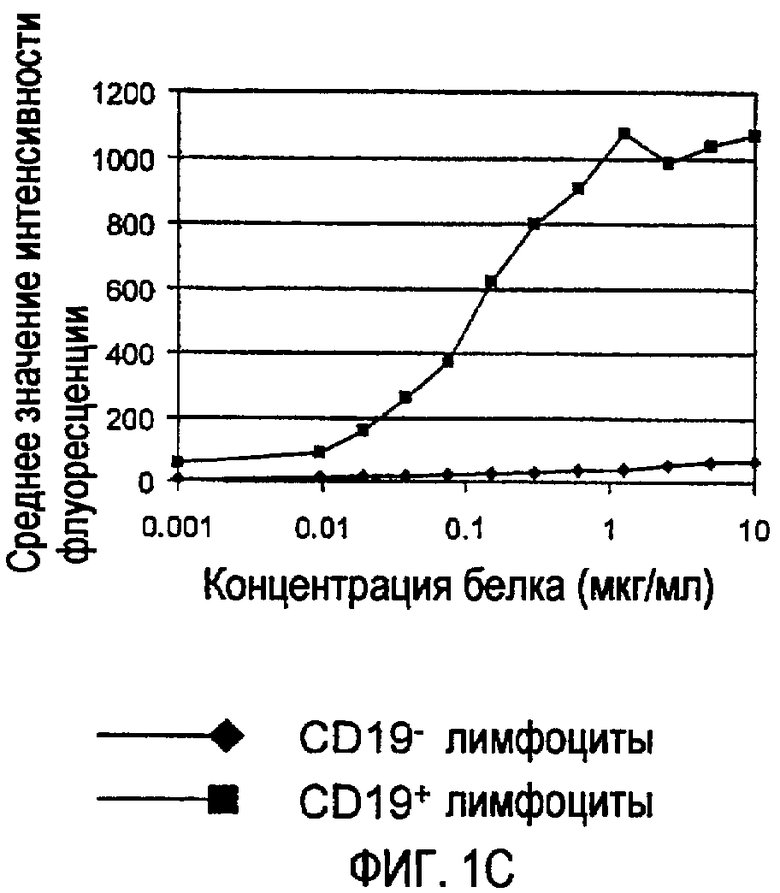
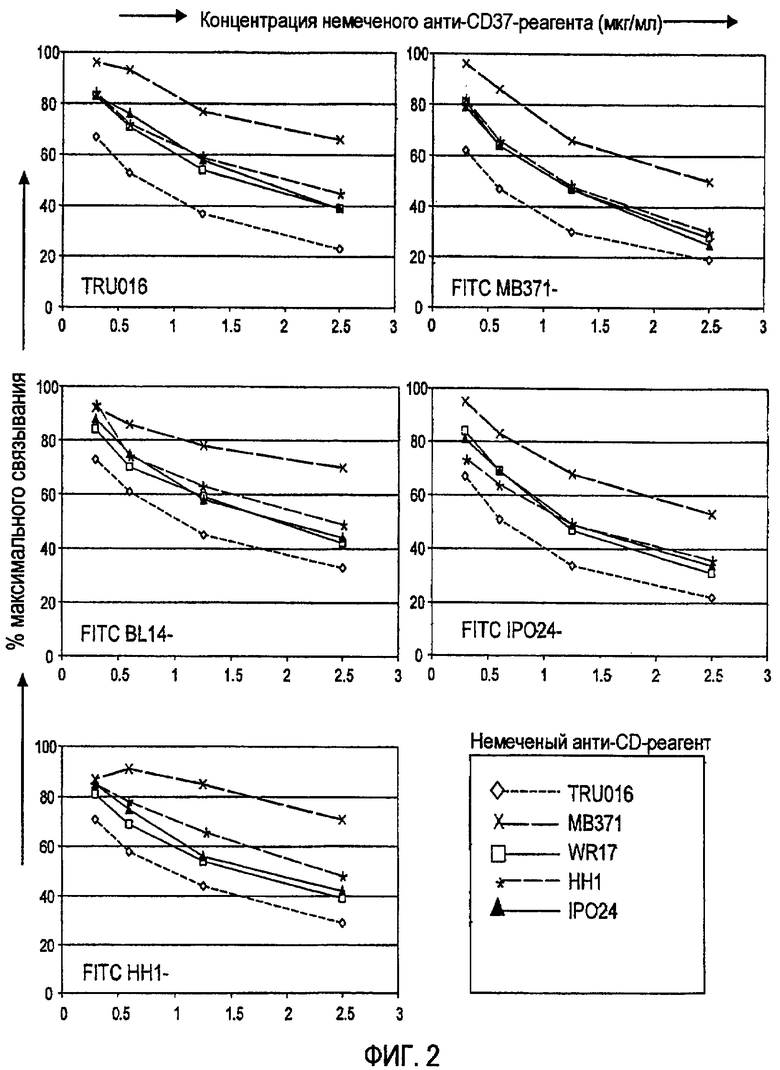
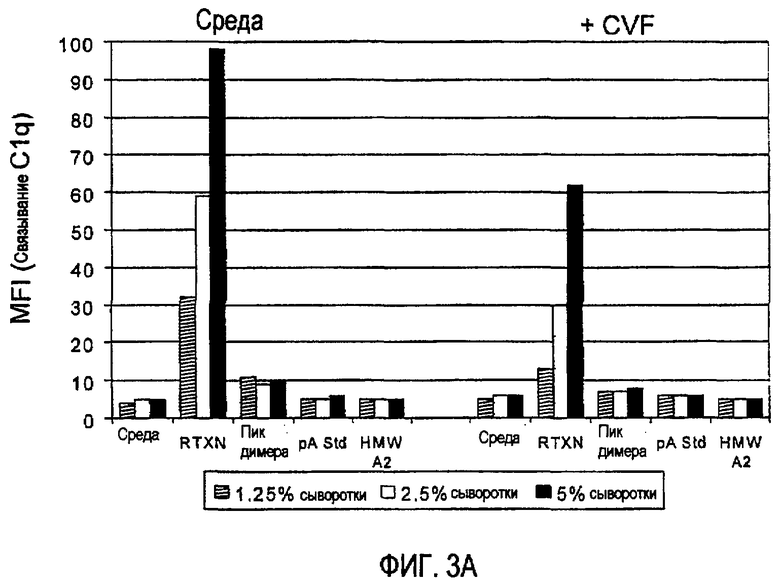
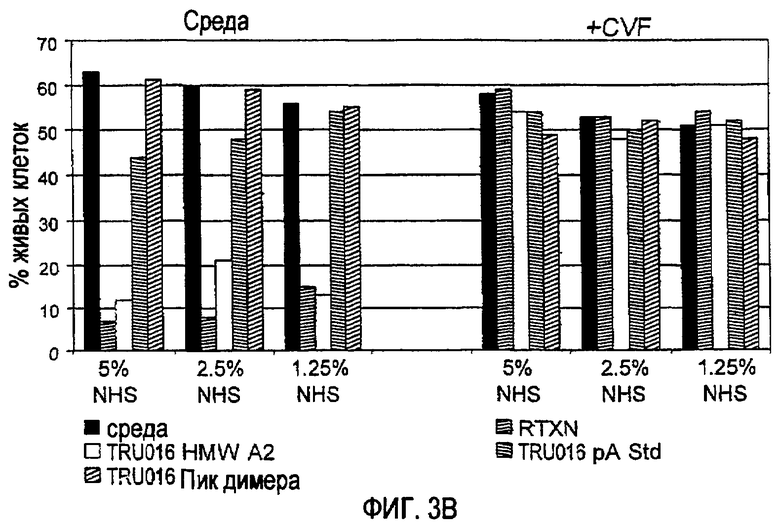
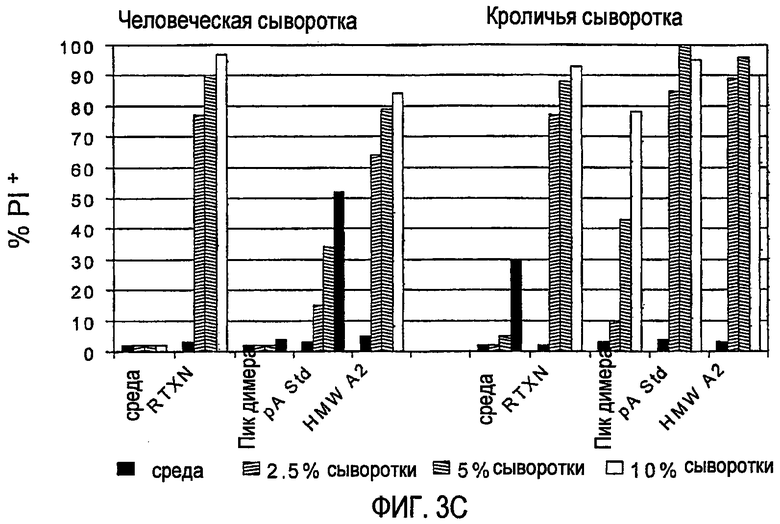
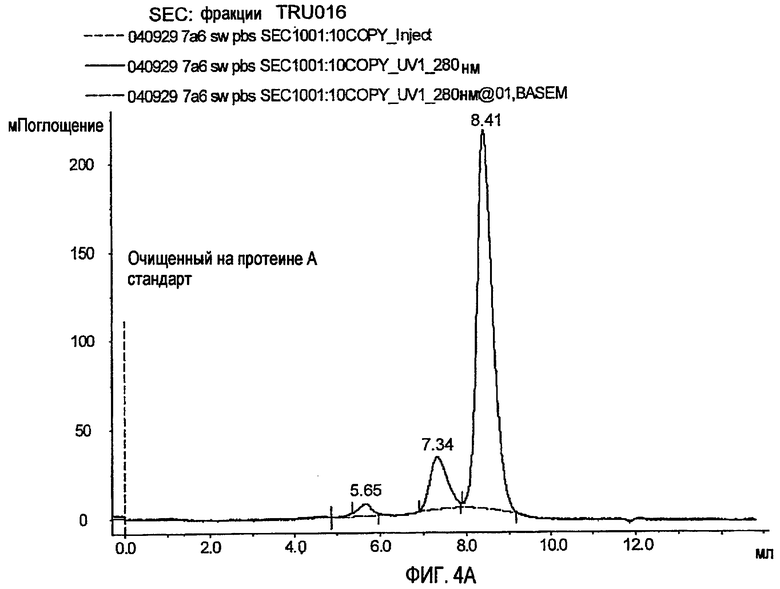
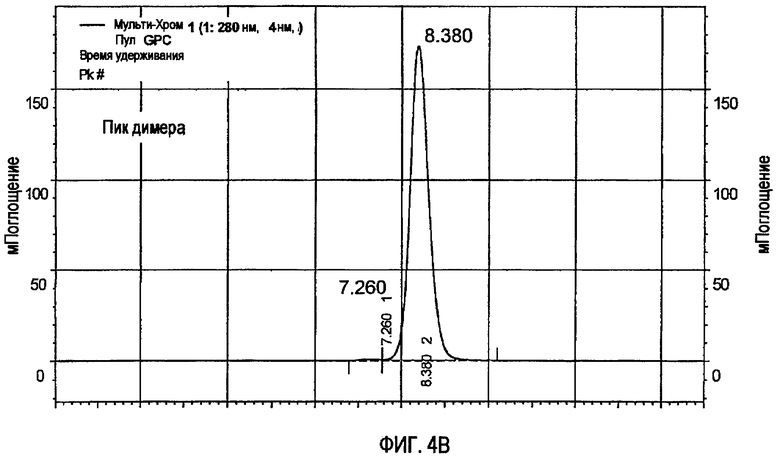
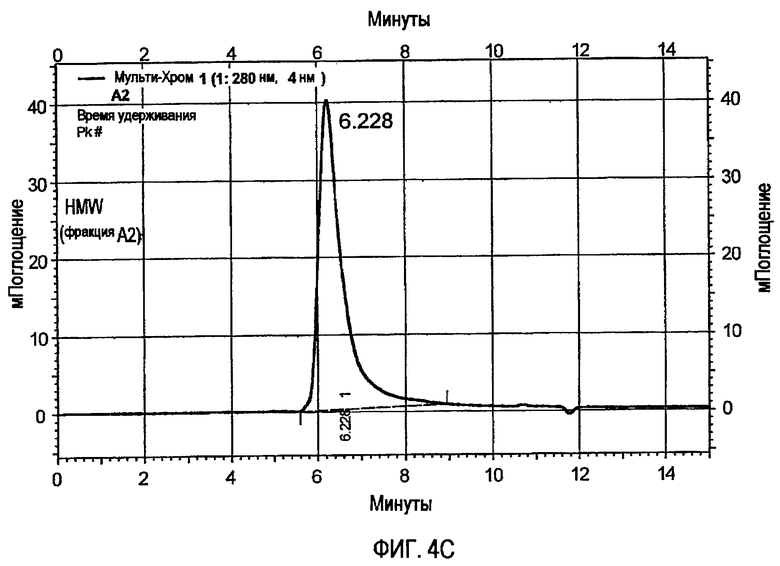

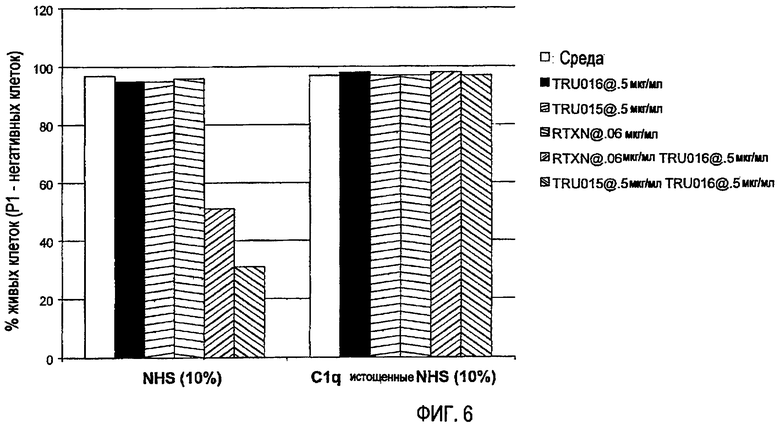
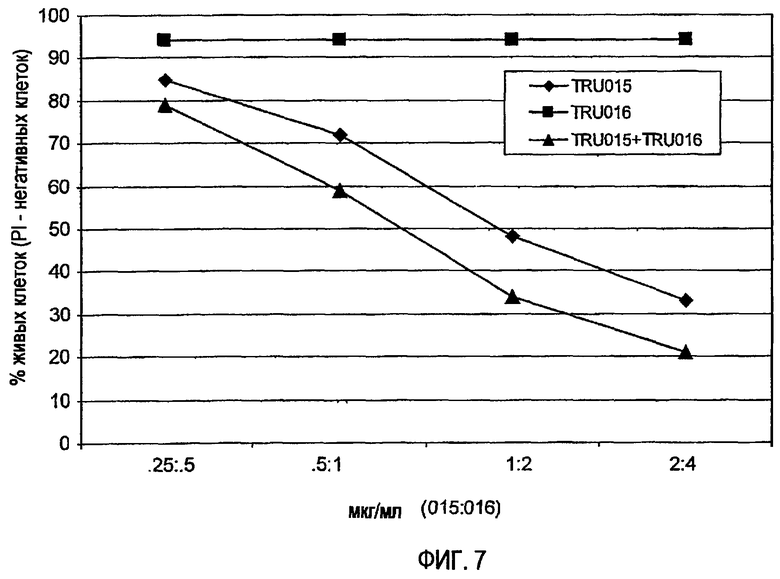
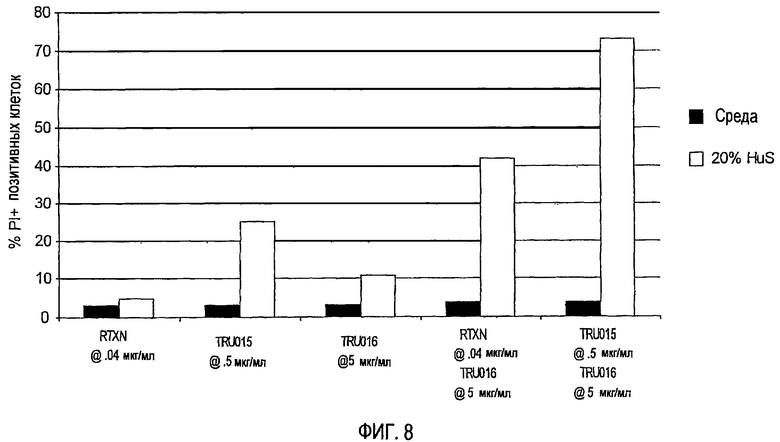
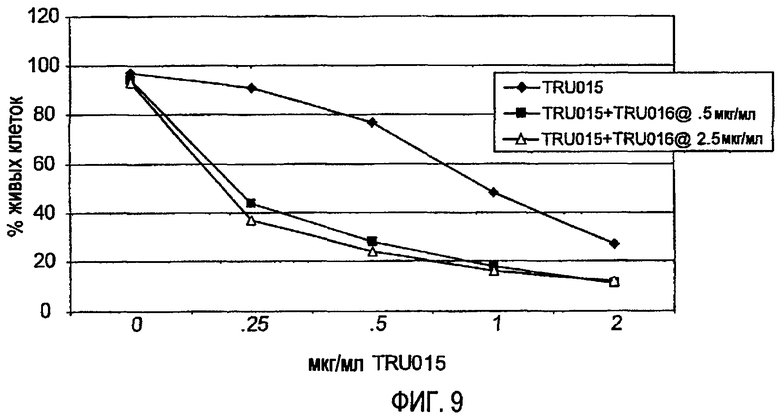
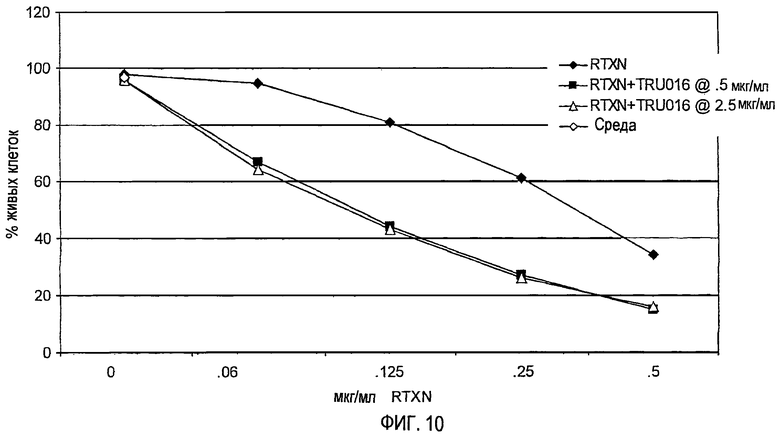
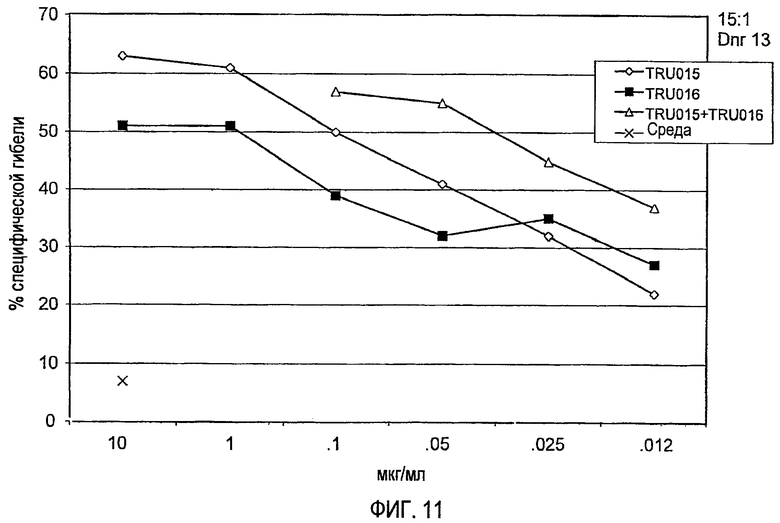

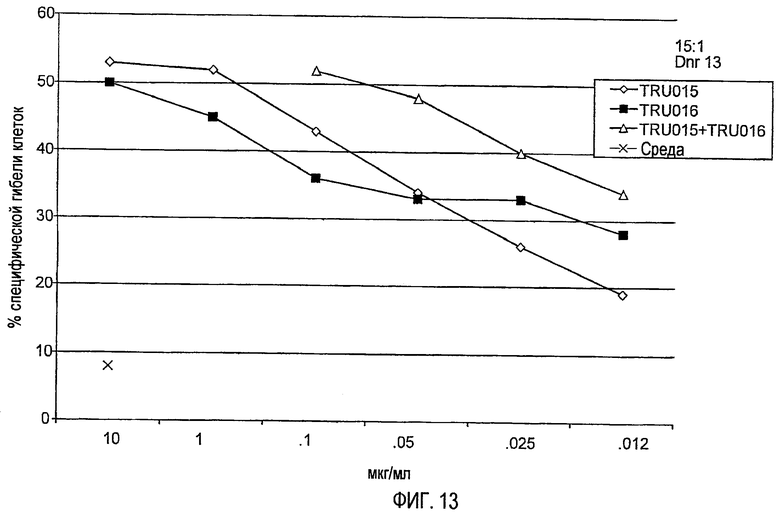
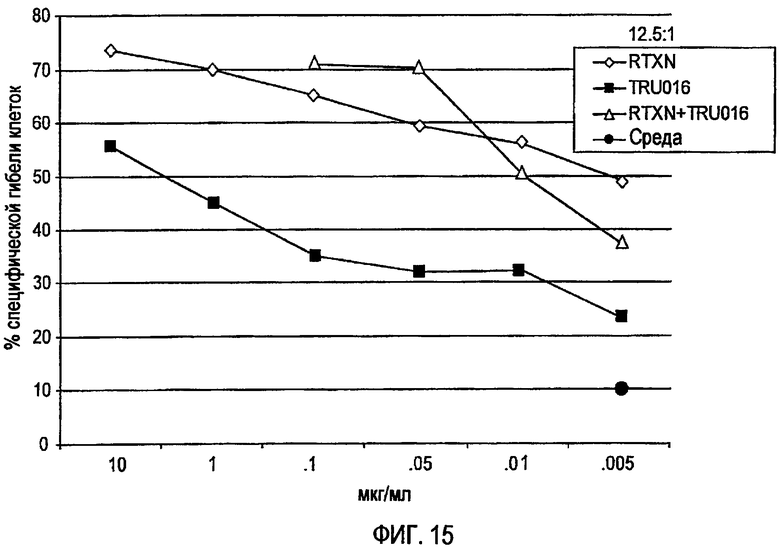
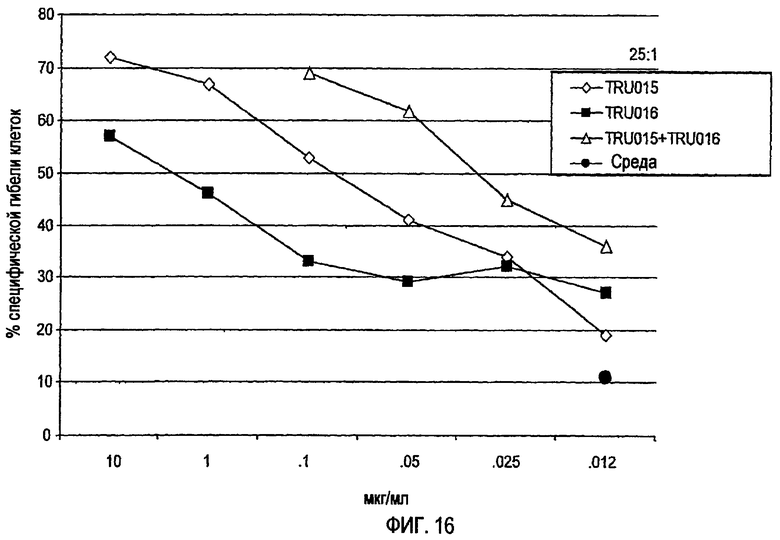
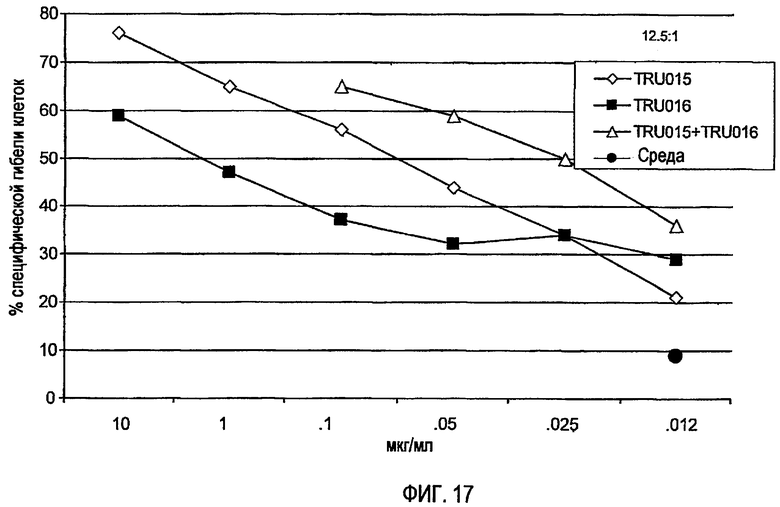
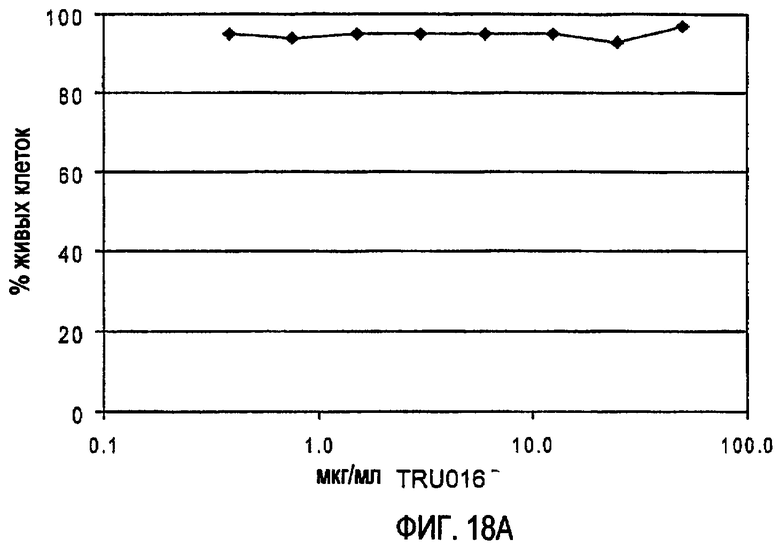
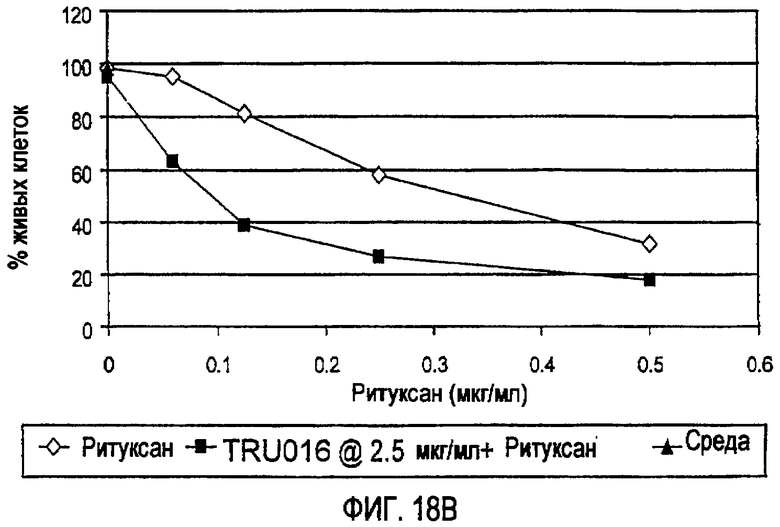
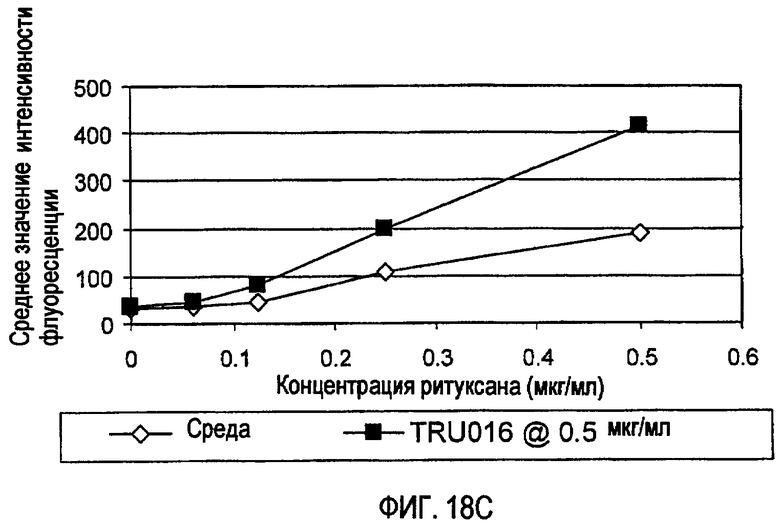
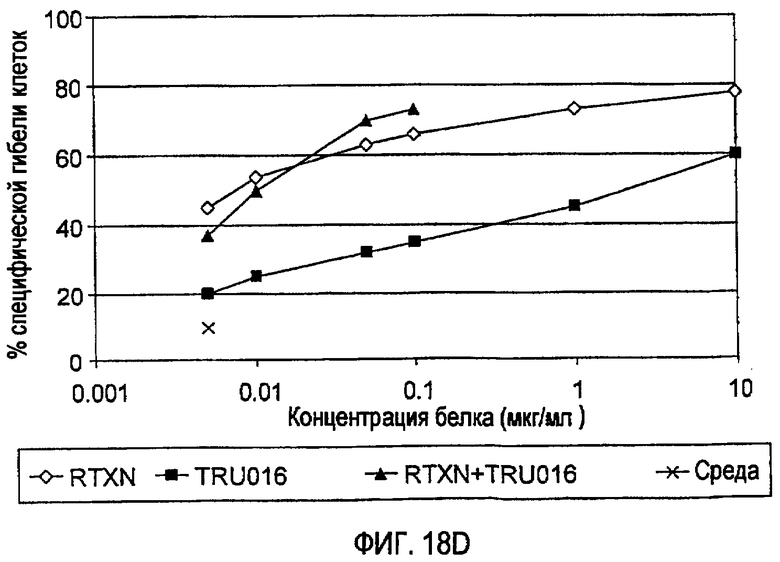
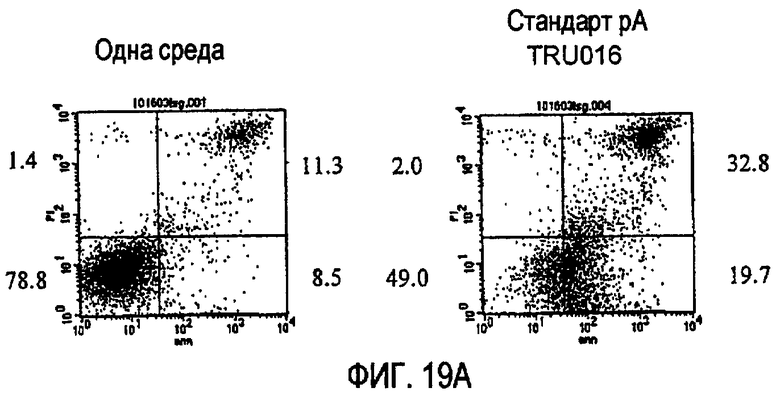
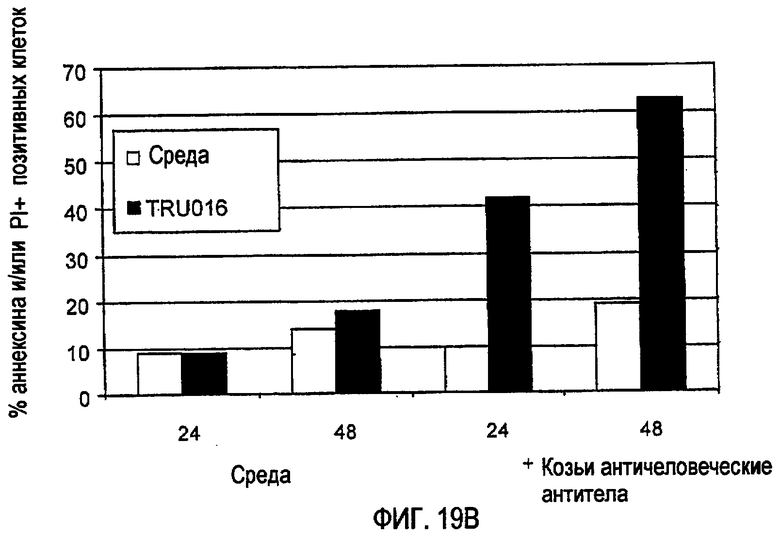
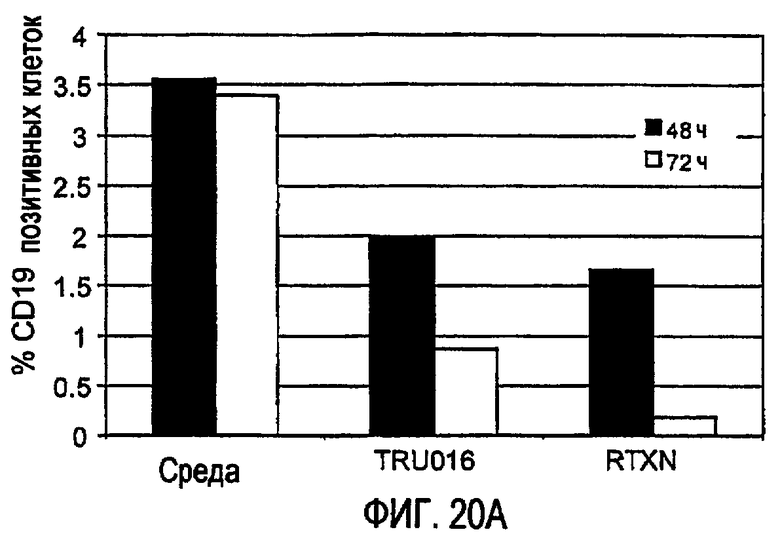
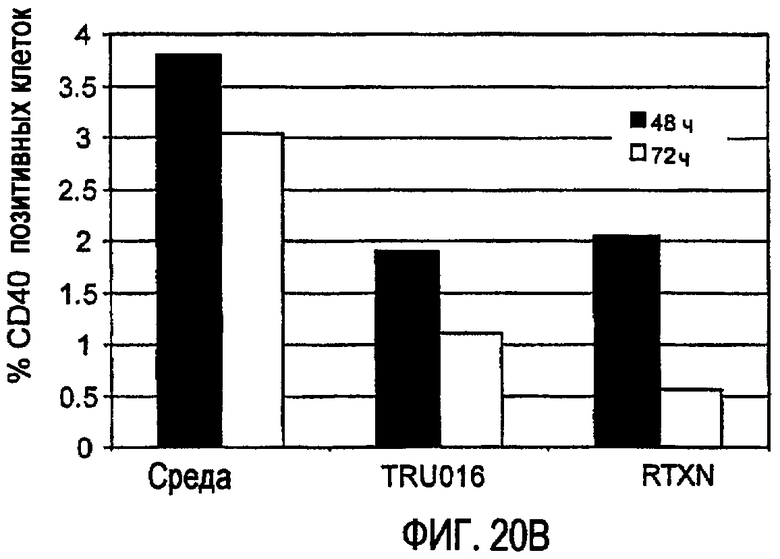
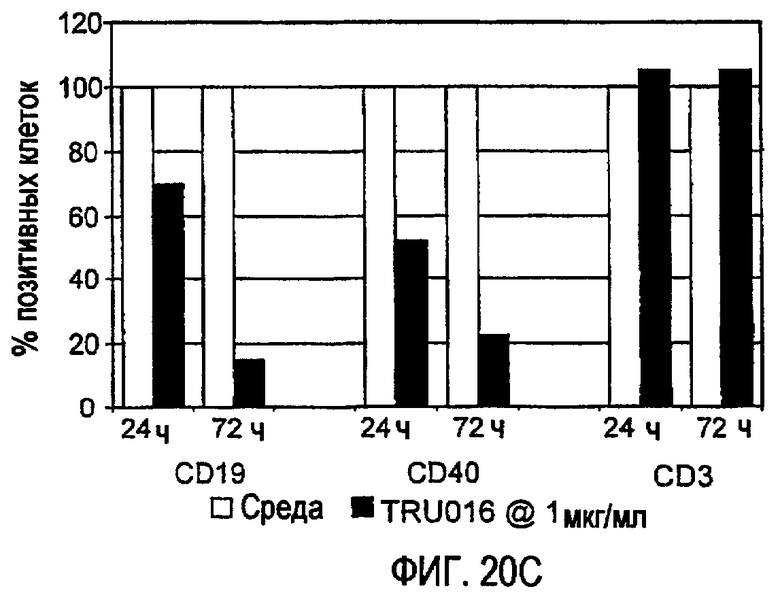
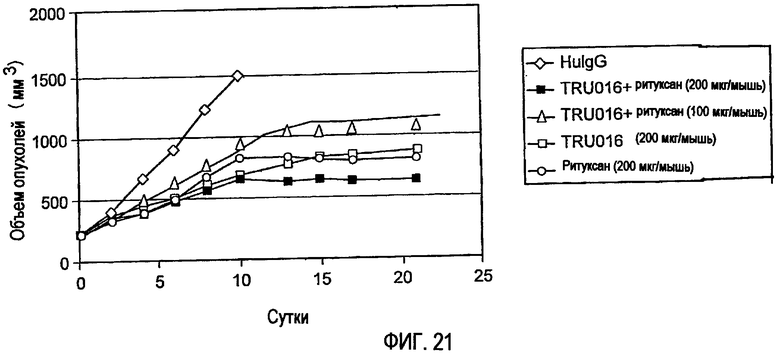
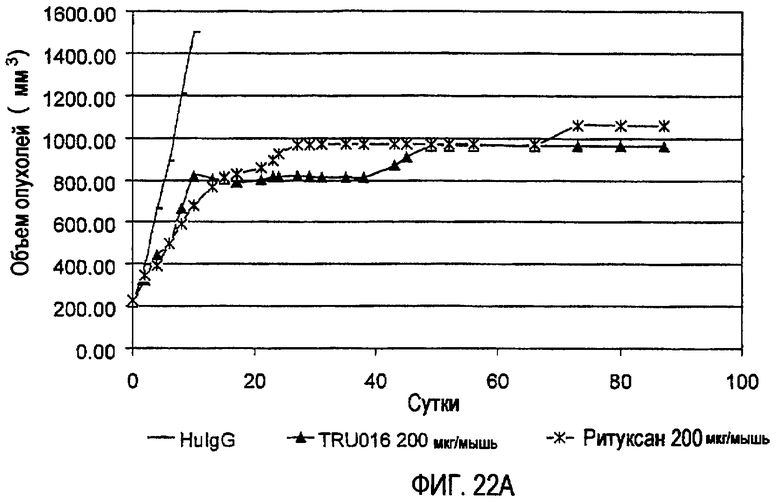
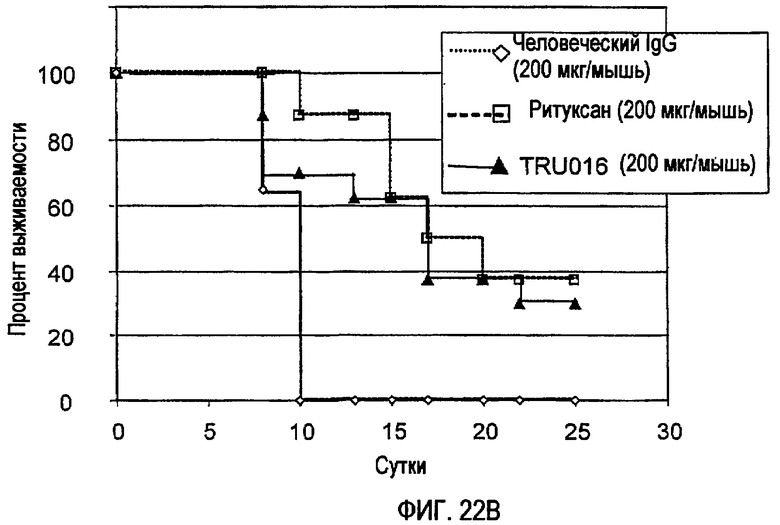
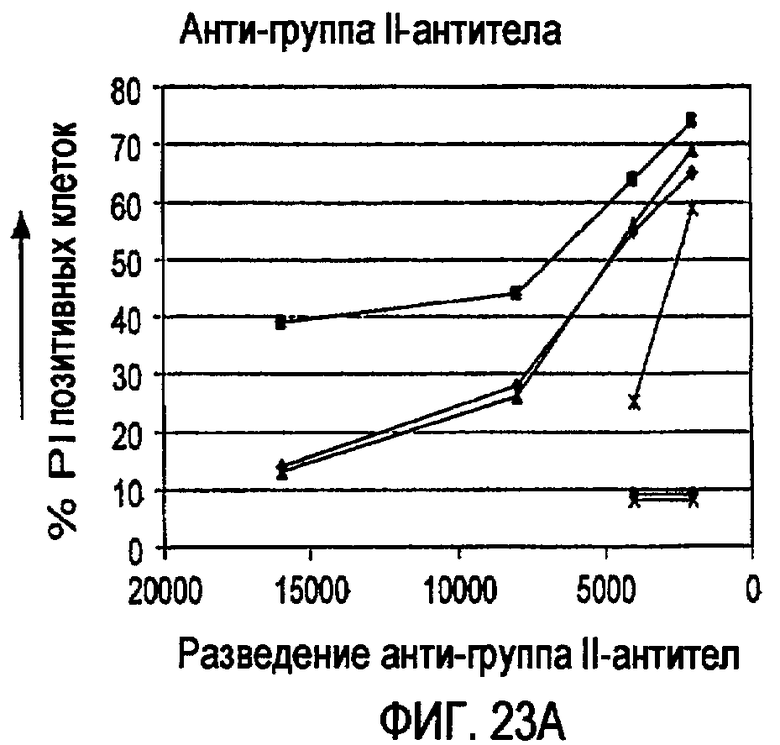
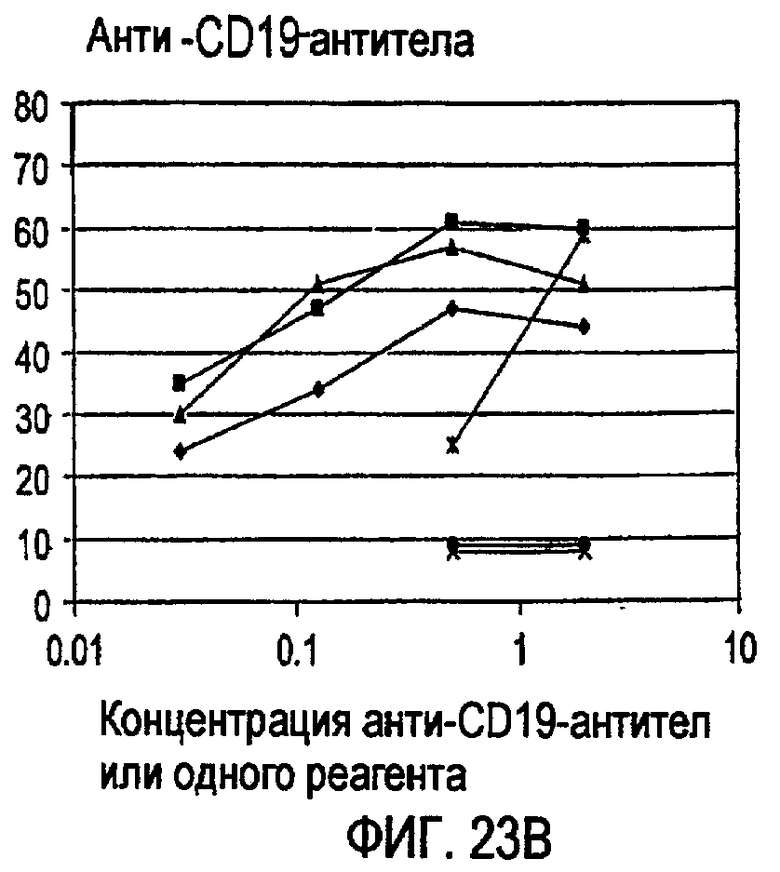
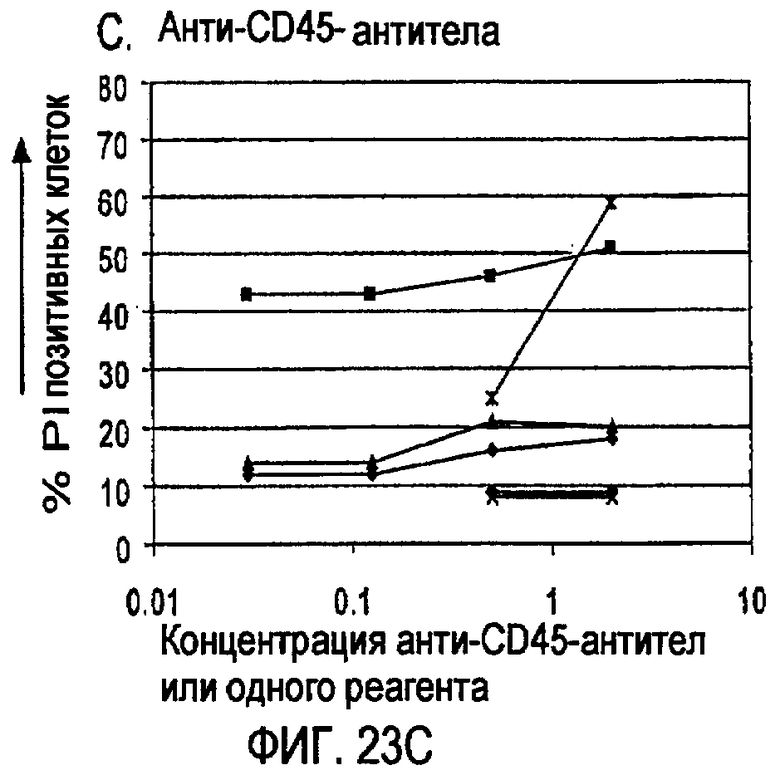
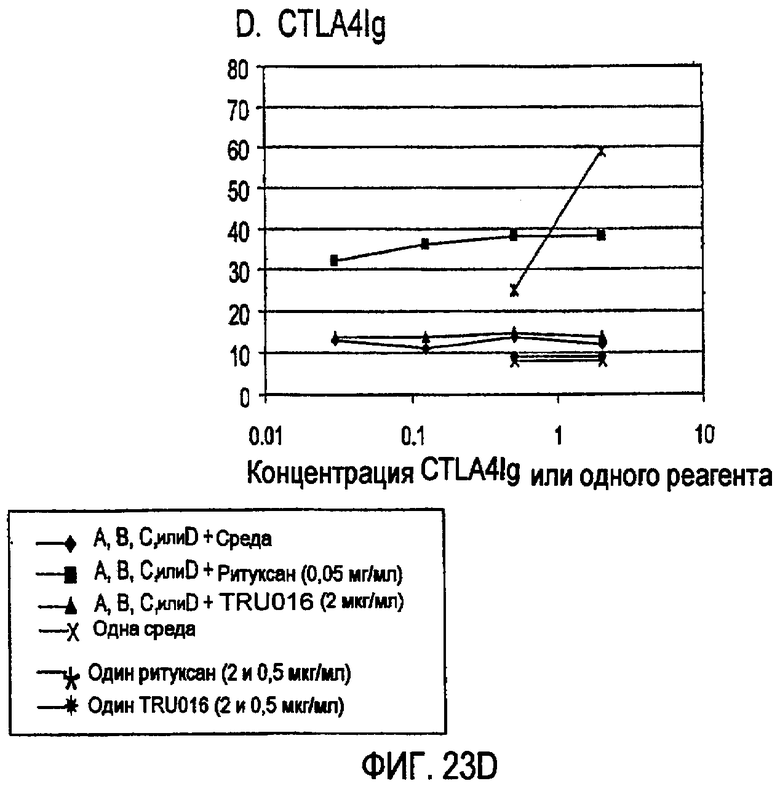
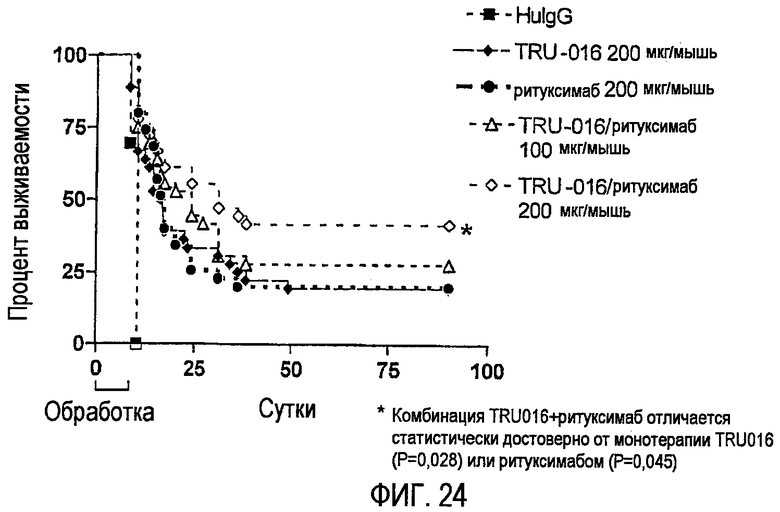
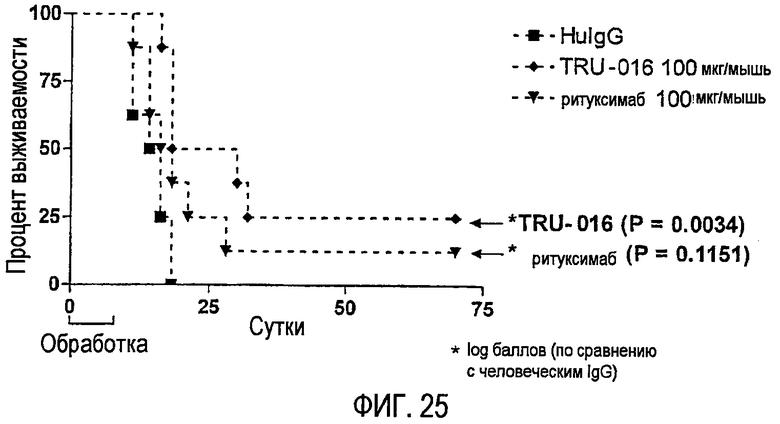
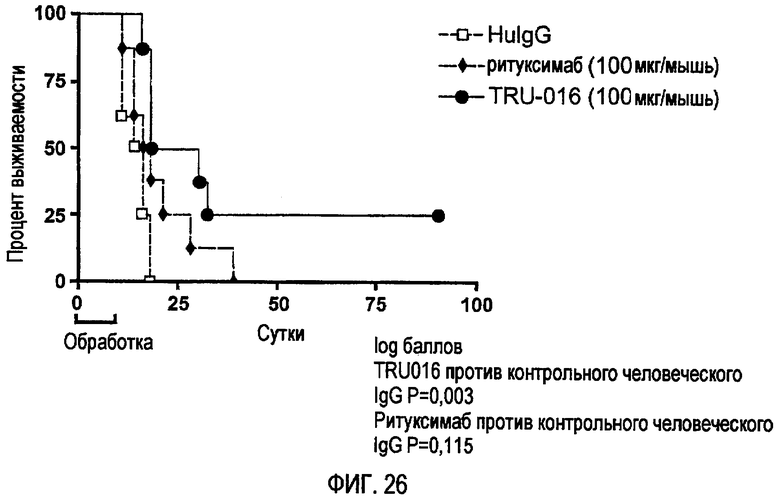
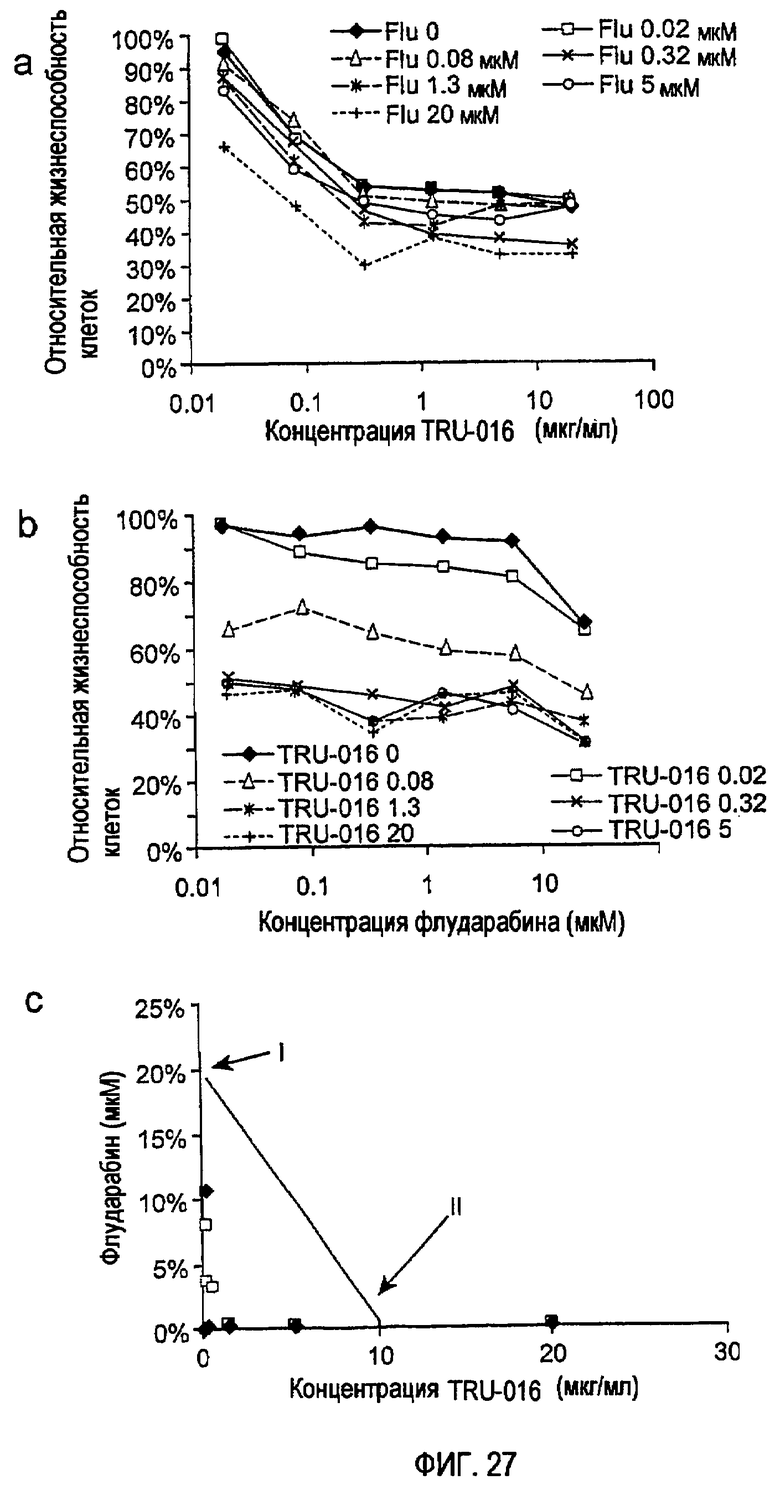
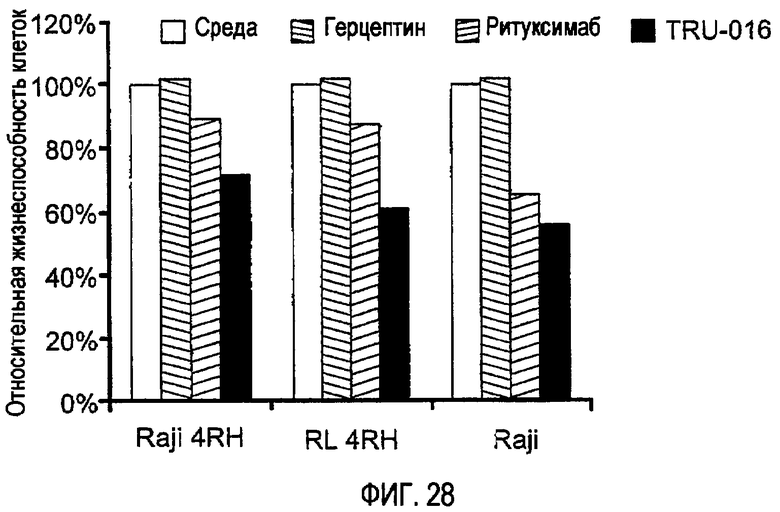
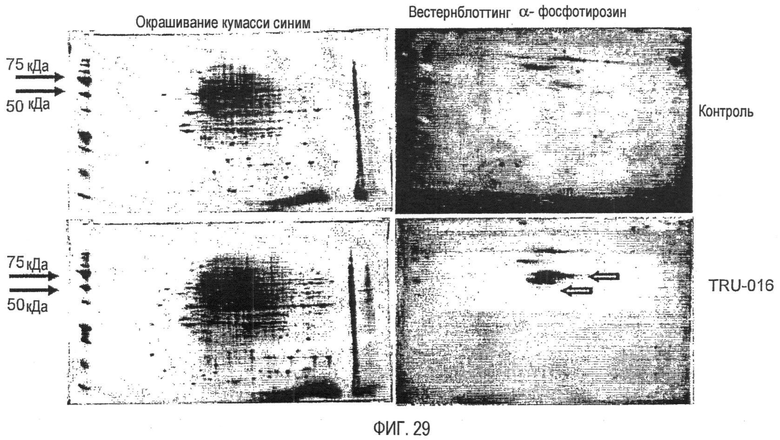


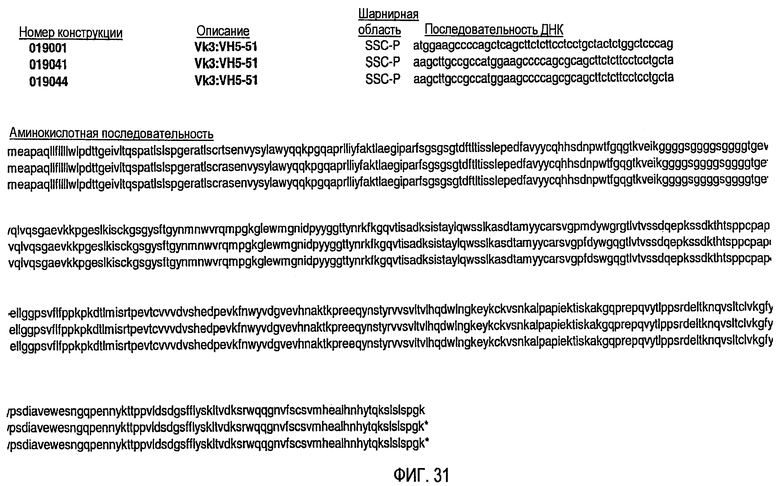



Изобретение относится к области биотехнологии и иммунологии. Предложены варианты антитела, специфичного к CD37 человека, каждый из которых содержит вариабельный участок легкой и тяжелой цепи. Описана кодирующая нуклеиновая кислота и вектор экспрессии на ее основе. Раскрыты: клетка-хозяин, содержащая вектор, способ получения антитела с использованием клетки, а также композиция для снижения количества В-клеток и способ лечения заболеваний, связанных с аберрантной активностью В-клеток, с использованием СD37-специфических антител. Использование изобретения обеспечивает специфическую гибель 80% клеток BJAB при концентрации антитела 10 мкг/мл, а также увеличивает выживаемость мышей Daudi, по сравнению с обработкой ритуксимабом. Это может найти применение для лечения различных опухолей. 7 н. и 32 з.п. ф-лы, 52 ил., 7 табл.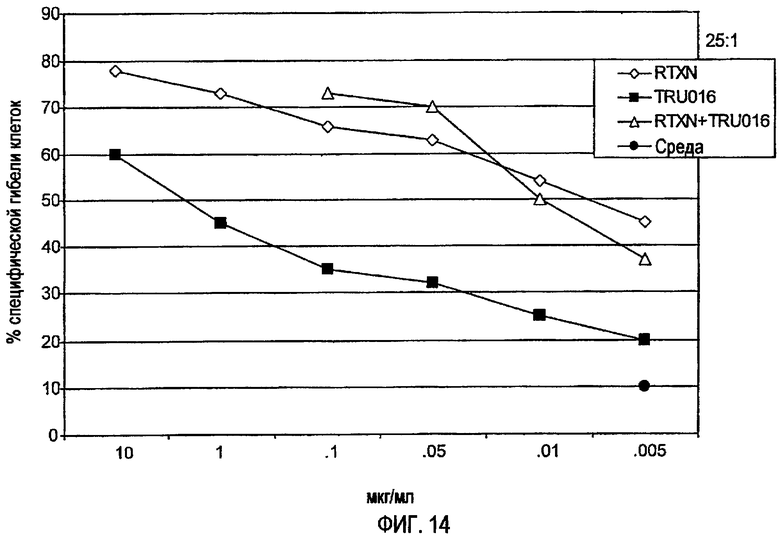
1. Специфичная к CD37 человека связывающая молекула иммуноглобулина, содержащая вариабельный участок легкой цепи и вариабельный участок тяжелой цепи, где
(i) связывающая молекула связывает CD37 человека, и
(ii) (а) вариабельный участок легкой цепи включает CDR1, CDR2 и CDR3 легкой цепи, приведенные в SEQ ID NO:6 или 48, и
(b) вариабельный участок тяжелой цепи включает CDR1, CDR2 и CDR3 тяжелой цепи, приведенные в SEQ ID NO:48 или 52.
2. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает вариабельный участок легкой цепи, приведенный в SEQ ID NO:6, и вариабельный участок тяжелой цепи включает CDR1, CDR2 и CDR3 тяжелой цепи, приведенные в SEQ ID NO:48.
3. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает вариабельный участок легкой цепи, приведенный в SEQ ID NO:48, и вариабельный участок тяжелой цепи включает CDR1, CDR2 и CDR3 тяжелой цепи, приведенные в SEQ ID NO:48.
4. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает вариабельный участок легкой цепи, приведенный в SEQ ID NO:6, и вариабельный участок тяжелой цепи включает CDR1, CDR2 и CDR3 тяжелой цепи, приведенные в SEQ ID NO:52.
5. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает вариабельный участок легкой цепи, приведенный в SEQ ID NO:48, и вариабельный участок тяжелой цепи включает CDR1, CDR2 и CDR3 тяжелой цепи, приведенные в SEQ ID NO:52.
6. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает CDR1, CDR2 и CDR3 легкой цепи, приведенные в SEQ ID NO:6, и вариабельный участок тяжелой цепи включает вариабельный участок тяжелой цепи, приведенный в SEQ ID NO:48.
7. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает CDR1, CDR2 и CDR3 легкой цепи, приведенные в SEQ ID NO:48, и вариабельный участок тяжелой цепи включает вариабельный участок тяжелой цепи, приведенный в SEQ ID NO:48.
8. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает CDR1, CDR2 и CDR3 легкой цепи, приведенные в SEQ ID NO:6, и вариабельный участок тяжелой цепи включает вариабельный участок тяжелой цепи, приведенный в SEQ ID NO:52.
9. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает CDR1, CDR2 и CDR3 легкой цепи, приведенные в SEQ ID NO:48, и вариабельный участок тяжелой цепи включает вариабельный участок тяжелой цепи, приведенный в SEQ ID NO:52.
10. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает вариабельный участок легкой цепи, приведенный в SEQ ID NO:6, и вариабельный участок тяжелой цепи включает вариабельный участок тяжелой цепи, приведенный в SEQ ID NO:48.
11. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает вариабельный участок легкой цепи, приведенный в SEQ ID NO:48, и вариабельный участок тяжелой цепи включает вариабельный участок тяжелой цепи, приведенный в SEQ ID NO:48.
12. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает вариабельный участок легкой цепи, приведенный в SEQ ID NO:6, и вариабельный участок тяжелой цепи включает вариабельный участок тяжелой цепи, приведенный в SEQ ID NO:52.
13. Специфичная к CD37 человека связывающая молекула по п.1, где вариабельный участок легкой цепи включает вариабельный участок легкой цепи, приведенный в SEQ ID NO:48, и вариабельный участок тяжелой цепи включает вариабельный участок тяжелой цепи, приведенный в SEQ ID NO:52.
14. Специфичная к CD37 человека связывающая молекула по п.1, где связывающая молекула представляет собой одноцепочечный полипептид Fv (scFv), антитело или его связывающий фрагмент или малый модуляторный иммунофармацевтический полипептид.
15. Специфичная к CD37 человека связывающая молекула по п.14, где связывающая молекула является химерной или гуманизированной.
16. Специфичная к CD37 человека связывающая молекула по п.1, где связывающая молекула представляет собой SMIP-полипептид, и вариабельный участок легкой цепи присоединен к вариабельному участку тяжелой цепи посредством линкерного пептида, содержащего (Gly4Ser)n, где n равно 1, 2, 3, 4, 5 или 6.
17. Специфичная к CD37 человека связывающая молекула по п.1, где связанные вариабельные участки легкой и тяжелой цепей присоединены к эффекторному домену посредством шарнира, состоящего из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:90, 92, 94, 96, 98, 100, 102, 104, 106, 108, 110, 112, 115, 116, 118, 120, 122, 124, 126 и 127.
18. Специфичная к CD37 человека связывающая молекула по п.17, где шарнир включает аминокислотную последовательность SEQ ID NO:92 или SEQ ID NO:106.
19. Специфичная к CD37 человека связывающая молекула по п.17, где эффекторный домен представляет собой домен IgG1.
20. Специфичная к CD37 человека связывающая молекула по п.1, где связывающая молекула представляет собой SMIP-полипептид, содержащий аминокислотную последовательность SEQ ID NO:84.
21. Специфичная к CD37 человека связывающая молекула по п.1, где связывающая молекула представляет собой SMIP-полипептид, содержащий аминокислотную последовательность SEQ ID NO:48 или SEQ ID NO:52.
22. Специфичная к CD37 человека связывающая молекула по п.1, где связывающая молекула обладает сродством связывания к CD37 человека от примерно 0,5 нМ до примерно 10 нМ.
23. Выделенная молекула нуклеиновой кислоты для экспрессирования специфичной к CD37 человека связывающей молекулы иммуноглобулина, содержащая нуклеотидную последовательность, кодирующую специфичную к CD37 человека связывающую молекулу иммуноглобулина, которая связывает CD37 человека, по любому из пп.1-22.
24. Вектор для экспрессирования специфичной к CD37 человека связывающей молекулы иммуноглобулина, содержащий выделенную молекулу нуклеиновой кислоты по п.23.
25. Клетка-хозяин, содержащая вектор по п.24.
26. Способ получения специфичной к CD37 человека связывающей молекулы иммуноглобулина, включающий культивирование клетки-хозяина по п.25 для экспрессии специфичной к CD37 человека связывающей молекулы иммуноглобулина и необязательное выделение специфичной к CD37 человека связывающей молекулы иммуноглобулина из культуры.
27. Композиция для снижения количества В-клеток, содержащая эффективное количество специфичной к CD37 человека связывающей молекулы иммуноглобулина, которая связывает CD37 человека, по любому из пп.1-22 и фармацевтически приемлемый носитель.
28. Способ лечения заболевания, связанного с аберрантной активностью В-клеток, включающий введение пациенту-человеку, нуждающемуся в этом, эффективного количества специфичной к CD37 человека связывающей молекулы иммуноглобулина по любому из пп.1-22.
29. Способ по п.28, где заболевание, связанное с аберрантной активностью В-клеток, представляет собой В-клеточный рак или аутоиммунное заболевание.
30. Способ по п.29, где аутоиммунное заболевание представляет собой ревматоидный артрит, рассеянный склероз, системную красную волчанку, болезнь Крона, синдром Шегрена, болезнь Граве, сахарный диабет типа I, псориаз, иммунную тромбоцитопеническую пурпуру, пузырчатку, идиопатическую воспалительную миопатию или макроглобулинемию Вальденстрема.
31. Способ по п.29, где В-клеточный рак представляет собой неходжкинскую лимфому или хроническую лимфоцитарную лейкемию.
32. Способ по п.28, где связывающую молекулу вводят в дозовом диапазоне от примерно 0,01 мг/кг до примерно 50 мг/кг.
33. Способ по п.28, где связывающую молекулу вводят в дозовом диапазоне от примерно 5 мг/кг до примерно 30 мг/кг.
34. Способ по п.28, дополнительно включающий введение пациенту-человеку эффективного количества специфичной к CD20 человека связывающей молекулы.
35. Способ по п.34, где специфичная к CD20 человека связывающая молекула представляет собой антитело или его связывающий фрагмент или малый модуляторный иммунофармацевтический полипептид.
36. Способ по п.35, где специфичная к CD20 связывающая молекула представляет собой моноклональное антитело ритуксимаб или малый модуляторный иммунофармацевтический полипептид, как приведено в SEQ ID NO:4.
37. Способ по п.28, дополнительно включающий введение пациенту-человеку эффективного количества специфичного к CD37 человека антитела или его связывающего фрагмента.
38. Способ по п.37, где специфичное к CD37 человека антитело обладает гуманизированным связывающим доменом антитела, выбранного из G28-1, МВ371, BL14, NMN46, IPO24, НН1 и WR17.
39. Способ по п.28, дополнительно включающий введение пациенту-человеку эффективного количества цитокина, хемокина, фактора роста, химиотерапевтического средства, радиотерапевтического средства или иммуносупрессивного средства.
| Ledbetter et al., "Monoclonal antibodies to a new gp40-45 (CD37) В cell associated cluster group modulate В cell proliferation" | |||
| In: McMichael AJ., Beverley P.C, Cobbold S., et al., editors | |||
| Leukocyte typing III: white cell differentiation antigens | |||
| Oxford, England: Oxford University Press; 1987 | |||
| Ручной ткацкий станок | 1922 |
|
SU339A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| KIESEL et | |||

