Область изобретения
Данное изобретение относится к диагностическому способу для обнаружения мутации цитохрома b в грибах, которая приводит к устойчивости к аналогам стробилурина или соединениям в той же самой группе перекрестной устойчивости, с использованием способа детектирования полиморфизма любого (или одного) единственного нуклеотида, предпочтительно с использованием системы амплификации рефракторной (устойчивой) мутации (ARMS). Данное изобретение относится также к специфическим праймерам с мутациями для применения в данном способе и к диагностическим наборам, содержащим эти праймеры. Кроме того, данное изобретение относится к идентификации специфической мутации в гене цитохрома b грибов, которая приводит к устойчивости грибов, содержащих указанную мутацию, к аналогам стробилурина или соединениям в той же самой группе перекрестной устойчивости.
Предпосылки изобретения
Широко распространенное применение фунгицидов в сельском хозяйстве является относительно недавним явлением, и большинство основных разработок имели место во время последних 40 лет. Ранее фермеры часто игнорировали или не распознавали действие, которое грибковые патогены оказывают на урожай и качество их урожаев. Однако в настоящее время эти потери являются неприемлемыми, и фермеры полагаются на применение фунгицидных химикалиев для борьбы с грибными болезнями. В результате коммерческие фунгициды стали важным компонентом общей агрохимической практики с распространенными по всему миру продажами в 1996 году около 5,9 биллионов долларов, что эквивалентно 18,9% общего агрохимического рынка (Wood Mackenzie, 1997a ‘Agcheiu products - The key agrochemical product groups’, in Agrochemical Service, Update of the Products Section, May 1997, 1-74). Большое количество фунгицидов уже доступны для фермера; недавнее издание The Pesticide Manual (Tomlin, 1994 10th Edition, British Crop Protection Council, Farnham, UK, and the Royal Society of Chemistry, Cambridge, UK) содержит 158 различных фунгицидных активных ингредиентов для применения в настоящее время. Тем не менее дальнейшее промышленное исследование, нацеленное на обнаружение и развитие новых соединений, является чрезвычайно интенсивным и процедуры управления по продукту являются крайне важными в гарантировании наилучшей и наиболее продолжительной производительности от применения фунгицидов с конкретным способом действия и/или принадлежащих к конкретным сериям соединений. В частности жизненно важной является разработка стратегий борьбы с устойчивостью при введении фунгицидов с новыми механизмами действия (Fungicide Resistance Management: Into The Next Millenium (Russel) 1999, in Pesticide Outlook, October 1999 (213-215).
Аналоги стробилурина составляют основную новую серию сельскохозяйственных фунгицидов, которые считаются наиболее захватывающим развитием на сцене сельскохозяйственных фунгицидов со времени обнаружения 1,2,4-триазолов в 1970-х годах.
Фунгицидная активность аналогов стробилурина является результатом их способности ингибировать митохондриальное дыхание в грибах. Более конкретно, было установлено, что эти соединения имеют новый односайтный способ действия, проявляя свое действие на грибы блокированием комплекса убихинол : цитохром с-оксидоредуктазы (цитохрома bc1), уменьшая, таким образом, генерирование богатого энергией АТФ в грибной клетке (Becker et al. FEBS Letts. 132:329-33). Это семейство ингибиторов предотвращает транспорт электронов в окислительно-восстановительном сайте убихинона Q0 на мультимерном белке цитохрома b (Esposti et al. 1993 Biochem. et Biophys. Acta 243-271). В противоположность многим митохондриальным белкам белок цитохрома b является митохондриально кодируемым.
Сообщения в литературе показывают, что специфические замены аминокислот в сайте-мишени цитохрома b могут влиять на активность аналогов стробилурина. Глубокие исследования мутагенеза проводились на Saccharomyces cerevisiae (далее назваемом S. cerevisiae) (JP Rago et al. 1989 J. Biol. Chem. 264, 14543-14548), мыши (Howell et al. 1988 J. Mol. Biol. 203, 607-618), Chlamydomonas reinhardtii (Bennoun et al. 1991 Genetics 127, 335-343) и Rhodobacter spp (Daldal et al., 1989, EMBO J. 3951-3961). Относящаяся к данной проблеме информация была также собрана из исследования природной основы для устойчивости к аналогам стробилурина у морского ежа Paracentrotus lividus (Esposti et al., 1990, FEBS 263, 245-247) и грибов семейства базидиомицетов Mycena galopoda и Strobilurus tenacellus (Kraiczy et al., 1996 Eur. J. Biochem. 235, 54-63), которые продуцируют природные варианты аналогов стробилурина. Имеются два различающихся района гена цитохрома b, где аминокислотные замены оказывают драматическое действие на активность аналогов стробилурина. Эти зоны охватывают аминокислотные остатки 125-148 и 250-295 (на основе системы нумерации остатков S. cerevisiae). Было показано более точно, что аминокислотные замены остатков 126, 129, 132, 133, 137, 142, 143, 147, 148, 256, 275 и 295 приводят к появлению устойчивости к аналогам стробилурина (Brasseur et al., 1996, Biochim. Biophys. Acta, 1275, 61-69 и Esposti et al. (1993), Biochimica et Biophysica Acta, 1143, 243-271).
Данное изобретение идентифицирует впервые ключевую важность одной из этих мутаций в гене цитохрома b полевых изолятов важных фитопатогенных грибов, обнаруживающих устойчивость к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости.
Сущность изобретения
Согласно первому аспекту данного изобретения авторы обеспечивают способ для обнаружения мутации в грибной нуклеиновой кислоте, где присутствие указанной мутации вызывает появление грибной устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает идентификацию присутствия или отсутствия указанной мутации в грибной нуклеиновой кислоте с использованием способа детектирования полиморфизма любого (или одного) единственного нуклеотида.
В данном изобретении авторы в настоящее время придумали новые диагностические способы для обнаружения точковой мутации в грибном гене цитохрома b на основе способов детектирования полиморфизма единственного нуклеотида, включающих в себя аллель-специфическую амплификацию. Специалисту с квалификацией в данной области будет очевидным, что имеется большое число аналитических процедур, которые могут быть использованы для обнаружения присутствия или отсутствия вариантных нуклеотидов в одном или нескольких полиморфных положениях в соответствии с данным изобретением. Обычно обнаружение аллельной вариации требует способа дискриминации мутаций, необязательно реакции амплификации и необязательно системы генерирования сигнала. Многие современные способы для обнаружения аллельной вариации обсуждаются в обзоре Nollau et al. Clin. Chem. 43, 1114-1120, 1997 и в стандартных руководствах, например, ’Laboratory Protocols for Mutation Detection’, Ed. by U. Landegren, Oxford University Press, 1996 и ’PCR’ 2nd Edition by Mewton and Graham, BIOS Scientific Publishers limited, 1997. Аллель-специфические реакции амплификации включают способы на основе праймеров, в том числе способы на основе полимеразной цепной реакции (ПЦР) и, более конкретно, развитие аллель-специфической полимеразной цепной реакции (ASPCR) и конкретно ARMS (системы амплификации рефракторного мутагенеза (Amplification Refractory Mutagenesis System)), где мутация вызывает появление устойчивости к аналогу стробилурина, и эти способы являются особенно предпочтительными для использования в способах данного изобретения. Способы данного изобретения включают в себя также не дискриминирующие ПЦР с последующим специфическим зондированием генерируемого ампликона (продукта амплификации). Эти способы пригодны для обнаружения специфических аллелей, которые могут придавать устойчивость к любому из аналогов стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости. Надежные тесты были разработаны для детектирования этой точковой мутации в диапазоне грибных фитопатогенов. Соединения могут считаться находящимися в одной и той же группе перекрестной устойчивости, когда механизм устойчивости к одному соединению сообщает также устойчивость к другому соединению, даже в том случае, когда способы их действия не являются одними и теми же. Способ ASPCR описан в патенте США с номером 5639611, а способ ARMS описан подробно в Европейском патенте с номером ЕР 332435.
Другие способы обнаружения единственного полиморфизма, которые могут быть использованы для обнаружения мутаций, включают в себя, например, полиморфизм длины рестрикционных фрагментов (RFLP), полиморфизм конформации одной цепи, множественный клональный анализ, аллель-специфическая гибридизация олигонуклеотидов, праймерное удлинение единственного нуклеотида (Juvonen et al. (1994) Hum Genet 93 16-20; Huoponen et al., (1994) Hum Mutat 3 29-36; Mashima et al., (1995), Invest Opthelmol. Vision. Sci 36, 1714-20; Howell et al. (1994) Am. J. Hum Genet. 55 203-206; Koyabashi et al. (1994) Am. J. Hum. Genet. 55 206-209; Johns and Neufeld (1993) Am. J. Hum Genet 53 916-920; Chomyn et al. (1992) Proc. Natl. Acad. Sci. USA 89 4221-4225) и способ Invader™ (доступный из Third Wave Technologies Inc. 502 South Rosa Road, Madison, WI 53719 USA).
Предпочтительным является применение систем детектирования на основе ПЦР.
Согласно предпочтительному варианту первого аспекта данного изобретения авторы обеспечивают способ обнаружения мутации в грибной нуклеиновой кислоте, где присутствие указанной мутации вызывает появление грибной устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает детектирование присутствия ампликона, генерируемого во время ПЦР-реакции, где указанная ПНР-реакция предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с праймером в присутствии подходящих нуклеотидтрифосфатов и агента для полимеризации, причем детектирование указанного ампликона непосредственно связано с присутствием или отсутствием указанной мутации в указанной нуклеиновой кислоте.
Обнаружение ампликона, генерируемого во время ПЦР-реакции, может быть непосредственно зависимым от удлинения праймера, специфического в отношении присутствия этой мутации, т.е. когда удлинение праймера зависит от присутствия данной мутации и, следовательно, ампликон генерируется только в том случае, когда этот праймер связывается и/или удлиняется, когда данная мутация присутствует (как это имеет место в случае технологии ARMS), подобным образом оно может непосредственно зависеть от удлинения праймера, специфического в отношении отсутствия этой мутации, например, последовательности дикого типа, или может быть непосредственно связано с продуктом удлинения ПЦР, содержащим мутантную последовательность ДНК, т.е. когда это обнаружение относится к ампликону, содержащему мутантную ДНК-последовательность. Первая альтернатива является особенно предпочтительной.
Ампликон может быть ампликоном из любого цикла ПЦР и он включает в себя первый продукт удлинения аллель-специфического праймера.
В дополнительном предпочтительном варианте данное изобретение обеспечивает способ обнаружения мутации в грибной нуклеиновой кислоте, где присутствие указанной мутации вызывает появление грибной устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с подходящим диагностическим праймером в присутствии подходящих нуклеотидтрифосфатов и агента для полимеризации, так что этот диагностический праймер удлиняется либо когда указанная мутация присутствует в пробе, либо когда присутствует последовательность дикого типа; и детектирование присутствия или отсутствия указанной мутации по присутствию или отсутствию продукта удлинения диагностического праймера.
В дополнительном предпочтительном варианте данное изобретение обеспечивает способ обнаружения мутации в грибной нуклеиновой кислоте, где присутствие указанной мутации вызывает появление грибной устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером для специфической мутации в присутствии подходящих нуклеотидтрифосфатов и агента для полимеризации, так что этот диагностический праймер удлиняется, когда указанная мутация присутствует в пробе, и детектирование присутствия или отсутствия указанной мутации по присутствию или отсутствию продукта удлинения диагностического праймера.
Согласно особенно предпочтительному варианту первого аспекта данного изобретения авторы обеспечивают способ обнаружения мутации в грибной нуклеиновой кислоте, где присутствие указанной мутации вызывает появление грибной устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером для специфической мутации в присутствии подходящих нуклеотидтрифосфатов и агента для полимеризации, так что этот диагностический праймер удлиняется только в том случае, когда указанная мутация присутствует в пробе, и детектирование присутствия или отсутствия указанной мутации по присутствию или отсутствию продукта удлинения диагностического праймера.
В применении здесь термин диагностический праймер используют для обозначения праймера, который используется специфически для идентификации присутствия или отсутствия мутации или последовательности дикого типа, а термин обычный праймер используют для обозначения праймера, связывающегося с противоположной цепью ДНК относительно диагностического праймера и 3’ относительно района, узнаваемого этим диагностическим праймером, и который, действуя с указанным диагностическим праймером, позволяет амплификацию лежащего между ними участка ДНК во время ПЦР. Если диагностический праймер является праймером ARMS, он может иметь 3’ дефект спаривания по сравнению с мутантной последовательностью или последовательностью дикого типа.
В этом и во всех последующих аспектах и вариантах данного изобретения предпочтительно детектировать удлинение продукта праймерного удлинения с использованием системы детектирования, которая является интегральной частью либо диагностического праймера, либо обычного праймера на противоположной цепи. Это описано более подробно здесь.
Способы данного изобретения особенно пригодны для обнаружения мутаций в митохондриальном гене, который кодирует белок, являющийся мишенью для фунгицида, особенно для обнаружения мутаций в грибном гене цитохрома b, где указанные мутации приводят к ингибированию фунгицидной активности белка цитохрома b, но все еще позволяют осуществление генерирования АТФ и наиболее предпочтительно, где указанная мутация в грибном гене цитохрома b приводит к одной из следующих аминокислотных замен: A126T, F129L, Y132C, C133Y, G137R/S/E/V, W142T/K, G143A, I147F, T148M, N256Y/K/I, I275F/S/T или L295F, где первая аминокислота заменяется второй аминокислотой в положении в последовательности, обозначенном номером, присутствие которой вызывает появление грибной устойчивости к аналогам стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем идентификация остатков основана на системе нумерации остатков цитохрома в S. cerevisiae.
Аналоги стробилурина и соединения в той же самой группе перекрестной устойчивости включают в себя, например, азоксистробин, пикоксистробин, крезоксим-метил, трифлоксистробин, фамоксадон и фенамидон.
Авторы изобретения нашли, что положение в нуклеиновой кислоте грибного цитохрома b, соответствующее 143-ему кодону/аминокислоте в последовательности цитохрома b S. cerevisiae, является ключевым определяющим фактором грибной устойчивости к аналогам стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости в полевых изолятах, устойчивых к аналогам стробилурина фитопатогенных грибов. Способы данного изобретения, описанные здесь, пригодны, в частности, для обнаружения мутации в положении, соответствующем остатку 143 цитохрома b Saccharomyces cerevisiae, где остаток глицина заменен другой аминокислотой, которая ингибирует активность аналогов стробилурина или любого другого соединения в той же самой группе перекрестной устойчивости и приводит к устойчивому фенотипу в грибах, несущих мутантный ген цитохрома b, вызывая тем самым появление грибной устойчивости к аналогам стробилурина или любого другого соединения в той же самой группе перекрестной устойчивости.
Этот способ предпочтительно используют для обнаружения мутации, приводящей к замене указанного остатка глицина в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, аминокислотой, выбранной из группы аргинин, серин, цистеин, валин, аспарагиновая кислота, глутаминовая кислота, триптофан и, наиболее предпочтительно, аланин.
Еще в одном предпочтительном варианте первого аспекта данного изобретения авторы изобретения обеспечивают способ обнаружения мутации в грибном гене цитохрома b, приводящей к замене глицина на аланин в положении, соответствующем остатку 143 цитохрома b S. cerevisiae (G143A), в кодируемом белке, приводящей, таким образом, к появлению грибной устойчивости к аналогам стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает идентификацию присутствия или отсутствия указанной мутации в грибной нуклеиновой кислоте с использованием способа детектирования полиморфизма любого (или одного) единственного нуклеотида.
Мутация в грибном гене цитохрома b, приводящая к замене G143A в кодируемом белке, обычно представляет собой замену основания гуанина на основание цитозин во втором положении (основании) этого кодона, и обнаружение этого полиморфизма единственного основания является предпочтительным для всех аспектов и вариантов описанного здесь изобретения.
Еще в одном предпочтительном варианте первого аспекта данного изобретения авторы обеспечивают диагностический способ обнаружения мутации в грибном гене цитохрома b, приводящей к замене G143A в кодируемом белке, причем указанный способ предусматривает детектирование присутствия ампликона, генерируемого во время ПЦР-реакции, причем указанная ПЦР-реакция предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, где детектирование указанного ампликона прямо связано с присутствием или отсутствием указанной мутации в указанной нуклеиновой кислоте.
В особенно предпочтительном варианте первого аспекта данного изобретения авторы изобретения теперь обеспечивают диагностический способ обнаружения мутации в грибном гене цитохрома b, приводящей к замене G143A в кодируемом белке, причем указанный способ предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером для мутации, приводящей к замене G143A в кодируемом белке, в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что этот диагностический праймер удлиняется, когда мутация присутствует в пробе, приводя к замене G143A в кодируемом белке; и детектирование присутствия или отсутствия указанной мутации по присутствию или отсутствию продукта удлинения диагностического праймера.
В следующем особенно предпочтительном варианте первого аспекта данного изобретения авторы теперь обеспечивают диагностический способ обнаружения мутации в грибном гене цитохрома b, приводящей к замене G143A в кодируемом белке, причем указанный способ предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером для мутации, приводящей к замене G143A в кодируемом белке, в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что этот диагностический праймер удлиняется только в том случае, когда мутация, приводящая к замене С143А в кодируемом белке, присутствует в пробе; и детектирование присутствия или отсутствия указанной мутации по присутствию или отсутствию продукта удлинения диагностического праймера.
В применении здесь термин G143A используется для обозначения замены остатка глицина остатком аланина в последовательности грибного цитохрома b в эквивалентном положении 143 его кодона/аминокислоты последовательности цитохрома b S. cerevisiae. Эта номенклатура используется для всех других замен остатков, цитируемых здесь, т.е. все положения цитируются относительно белковой последовательности цитохрома b S. cerevisiae. Последовательности гена и белка цитохрома b S. cerevisiae доступны на базах данных EMBL и SWISSPROT (см. EMBL ACCESSION NO. X84042 и SWISSPROT ACCESSION NO. P00163). Специалисту с квалификацией в данной области будет понятно, что точная длина и спектр эквивалентных белков из различных видов могут варьироваться как результат амино- или карбоксиконцевых и/или одной или нескольких внутренних делений или инсерций. Поскольку аминокислотный участок, содержащий остаток, соответствующий 6143 в S. cerevisiae, является достаточно консервативным (Widger et al. Proc. Natl. Acad. Sci. USA 81 (1984) 674-678), нетрудно идентифицировать точно соответствующий остаток во вновь полученной последовательности грибного цитохрома b либо визуальным исследованием, либо с использованием одной из нескольких программ сопоставления последовательностей, в том числе Megalign или Macaw. Хотя и обозначенное как G143 в данной заявке, вследствие позиционной и функциональной эквивалентности точное положение этого глицина в новом цитохроме b может не быть 143-м остатком от его амино-конца. Консенсусная последовательность цитохрома b S. cerevisiae обеспечена в SWISSPROT ACCESSION NO. P00163. Во всех аспектах и вариантах данного изобретения, описанных здесь, положения в последовательности цитохрома b являются предпочтительно определенными относительно последовательности цитохрома b S. cerevisiae, обеспеченной в EMBL ACCESSION NO. Х84042. Альтернативно, во всех аспектах и вариантах данного изобретения, описанных здесь, положения в последовательности цитохрома b являются предпочтительно определенными относительно консенсусной последовательности цитохрома b S. cerevisiae, обеспеченной в SWISSPROT ACCESSION NO. P00163.
Согласно одному аспекту данного изобретения обеспечен способ для диагностики полиморфизма единственного нуклеотида в грибном гене цитохрома b, причем этот способ предусматривает определение последовательности грибной нуклеиновой кислоты в положении, соответствующем одному или нескольким основаниям в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке цитохрома b, и определение состояния устойчивости указанных грибов в отношении аналога стробилурина или соединения в той же самой группе перекрестной устойчивости по полиморфизму в гене цитохрома b.
Во всех аспектах и вариантах данного изобретения, описанных здесь, предпочтительно, чтобы только одно основание в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке цитохрома b, обнаруживало мутацию, т.е. где имеется полиморфизм единственного нуклеотида, имеющий место только в одном положении и, кроме того, предпочтительно, чтобы мутация находилась в первом или втором основании этого триплета и наиболее предпочтительно, чтобы она находилась во втором основании этого триплета.
Согласно предпочтительному варианту этого аспекта данного изобретения, обеспечен способ для диагностики полиморфизма единственного нуклеотида в грибном гене цитохрома b, причем этот способ предусматривает определение последовательности грибной нуклеиновой кислоты в положении, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке цитохрома b, и определение состояния устойчивости указанных грибов в отношении аналога стробилурина или соединения в той же самой группе перекрестной устойчивости по полиморфизму в гене цитохрома b.
В варианте вышеописанного аспекта данного изобретения способ для диагностики, описанный здесь, является способом, в котором полиморфизм единственного нуклеотида в положении ДНК, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке цитохрома b, представляет собой присутствие G и/или С.
Использование кодонов
Точковая мутация глицина в аланин требует замены G на С во втором основании этого кодона. Другие мутации могут также возникать в 3-м положении в этом кодоне вследствие вырожденности генетического кода в отношении аланина и глицина (см. таблицу 1), но это легко учесть при конструировании диагностического праймера. Диагностический праймер является предпочтительно праймером ARMS (Концепция праймеров ARMS в полном виде описана в Newton et al. Nucleic Acid Research 17(7) 2503-2516 1989). В результате праймеры ARMS могут быть сконструированы для обнаружения точковой мутации G143A при условии информации только о последовательности чувствительного к аналогу стробилурина гена цитохрома b дикого типа. Нет необходимости иметь доступный устойчивый изолят в новых представляющих интерес грибах, происходящий из мутации G143A. Некоторые примеры релевантных фитопатогенных грибов приведены в списке в таблице 2. Этот список не рассматривается как каким-либо образом ограничивающий. Квалифицированный фитопатолог будет способен легко идентифицировать те грибы, к которым могут быть применены способы данного изобретения.
Пример видов, в которых может анализироваться G143A
Способы данного изобретения, описанные здесь, особенно применимы в связи с фитопатогенными грибами, в частности со следующими видами грибов: Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Venturia inaequalis, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Magnaporthe grisea, Phytophthora infestans, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola.
В дополнительном аспекте данное изобретение обеспечивает способ обнаружения грибной устойчивости к аналогу стробилурина или любого другого соединения в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает идентификацию присутствия или отсутствия мутации в грибной нуклеиновой кислоте, причем присутствие указанной мутации вызывает появление устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает идентификацию присутствия или отсутствия полиморфизма единственного нуклеотида, имеющего место в положении, соответствующем одному или нескольким основаниям в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке цитохрома b.
В дополнительном предпочтительном варианте этого аспекта данное изобретение обеспечивает способ обнаружения грибной устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает идентификацию присутствия или отсутствия мутации в грибной нуклеиновой кислоте, причем присутствие указанной мутации вызывает появление устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает идентификацию присутствия или отсутствия полиморфизма единственного нуклеотида, имеющего место в положении, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке цитохрома b.
В предпочтительном варианте этого аспекта данного изобретения присутствие или отсутствие полиморфизма единственного нуклеотида в положении, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в гене цитохрома b в грибной нуклеиновой кислоте, идентифицируют с использованием способов детектирования полиморфизма любого (или одного) единственного нуклеотида.
Далее, данное изобретение обеспечивает грибную последовательность ДНК, кодирующую весь белок цитохрома b дикого типа или часть этого белка, причем указанная последовательность ДНК кодирует остаток глицина в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке дикого типа, причем указанная последовательность может быть получена или получена из гриба, выбранного из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctania solani, Mycosphaerella musicola и Cercospora arachidola.
Грибные последовательности ДНК в соответствии с вышеописанными аспектами данного изобретения предпочтительно содержат около 30 нуклеотидов на любой или на обеих сторонах от положения в этой ДНК, соответствующего одному или более основаниям в триплете, предпочтительно соответствующего второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке, так как эта степень нуклеиновой кислоты обеспечивает специалиста всей информацией, необходимой для конструирования видо- и мутация-специфических реагентов и/или способов для использования во всех способах детектирования полиморфизма единственного нуклеотида. В применении здесь термин около 30 означает, что эта последовательность может содержать до 30 нуклеотидов, например 5, до 10, 15, 20 или 25 нуклеотидов, или может содержать более 30 нуклеотидов.
В применении здесь в связи со всеми ДНК- и белковыми последовательностями термин "вся или часть" используется для обозначения ДНК-последовательности или белковой последовательности или их фрагмента. Фрагмент ДНК или белка может составлять, например, 10, 20, 25, 30, 40, 50, 60, 70, 75, 80, 85, 90 или 95% длины всей последовательности.
Специалисту с квалификацией в данной области будет очевидно, что пробы, содержащие как геномную (митохондриальную) ДНК, так и кДНК, могут анализироваться в соответствии с данным изобретением. Если проба содержит геномную ДНК, следует учитывать интронную организацию при использовании информации последовательности. Примеры грибных последовательностей ДНК дикого типа, содержащих часть последовательности гена цитохрома b дикого типа, в соответствии с вышеописанным аспектом данного изобретения, обеспечены в таблице 3 и указанные последовательности образуют следующий аспект данного изобретения.
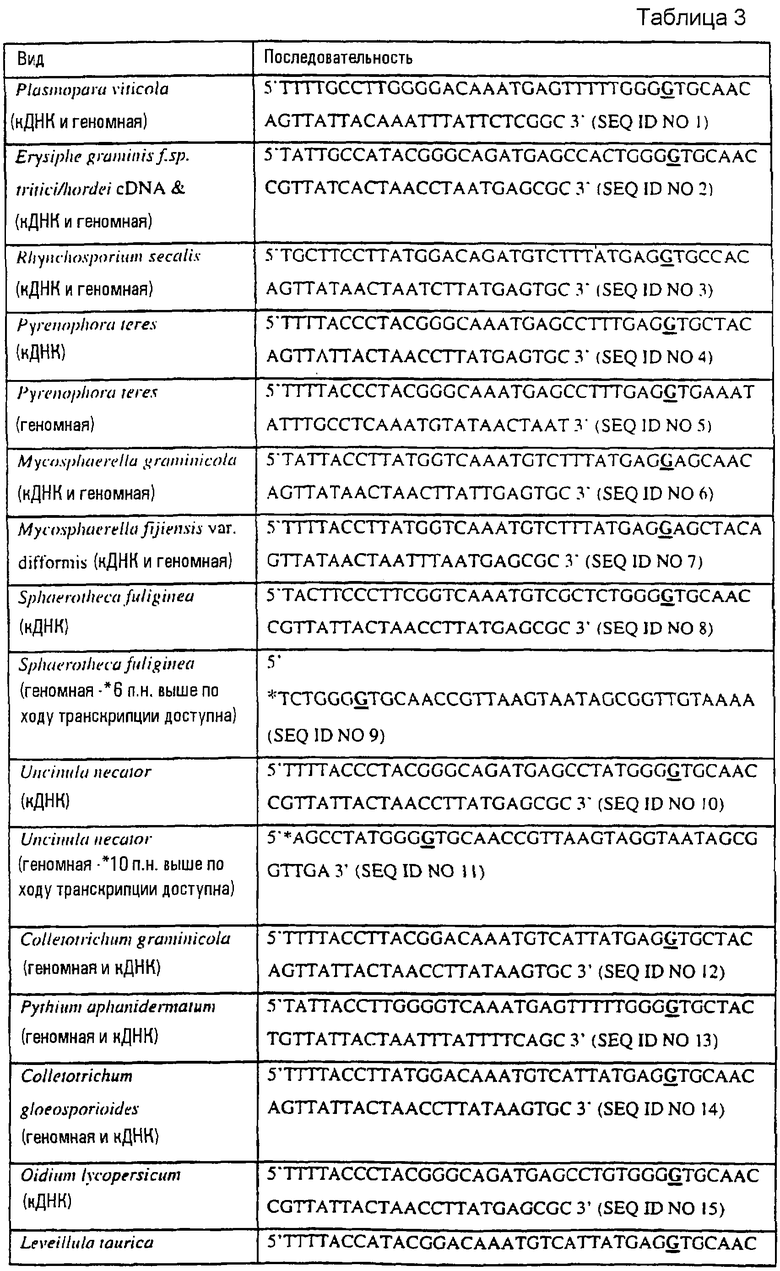

В таблице 3 второе основание в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, которое приводит к замене обычного остатка глицина альтернативной аминокислотой, причем указанная замена сообщает устойчивость к аналогам стробилурина или соединению в той же самой группе перекрестной устойчивости, выделено жирным шрифтом и подчеркнуто.
Данное изобретение относится также к грибной последовательности ДНК, обнаруживающей гомологию или идентичность последовательности относительно указанных последовательностей ДНК в таблице 3, и включает в себя, например, вариации в последовательностях ДНК, обнаруживаемые в различных пробах или изолятах одного и того же вида. Эти вариации могут, например, быть следствием использования альтернативного кодона, варьирующей митохондриальной организации интрон/экзон и замены аминокислот.
В следующем аспекте данное изобретение обеспечивает грибную последовательность ДНК, кодирующую весь белок цитохрома b или его часть, которая, при выстраивании против соответствующей последовательности ДНК дикого типа, кодирующей белок цитохрома b, содержит, как это видно, мутацию полиморфизма единственного нуклеотида в положении в ДНК, соответствующем одному или более основаниям в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в этом белке, что приводит к замене обычного остатка глицина альтернативной аминокислотой, при условии, что указанная последовательность ДНК не является последовательностью Mycena galopoda, кодирующей цитохром b.
В следующем предпочтительном варианте этого аспекта данное изобретение обеспечивает грибную последовательность ДНК, кодирующую весь белок цитохрома b или его часть, которая при выстраивании против соответствующей последовательности ДНК дикого типа, кодирующей белок цитохрома b, содержит, как это видно, мутацию полиморфизма единственного нуклеотида в положении в ДНК, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в этом белке, что приводит к замене обычного остатка глицина альтернативной аминокислотой, при условии, что указанная последовательность ДНК не является последовательностью Mycena galopoda, кодирующей цитохром b.
Грибная последовательность ДНК в соответствии с вышеописанным аспектом данного изобретения содержит предпочтительно около 30 нуклеотидов на любой стороне или на обеих сторонах от положения в этой ДНК, соответствующего одному или нескольким основаниям в триплете, предпочтительно соответствующего второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке, так как эта степень нуклеиновой кислоты обеспечивает специалиста всей информацией, необходимой для конструирования видо- и мутация-специфических реагентов и/или способов для использования во всех способах детектирования полиморфизма единственного нуклеотида. В применении здесь термин около 30 означает, что эта последовательность может содержать до 30 нуклеотидов, например 5, до 10, 15, 20 или 25 нуклеотидов, или может содержать более 30 нуклеотидов.
Далее, данное изобретение обеспечивает грибную последовательность ДНК, кодирующую весь мутантный белок цитохрома b или его часть, причем присутствие мутации в указанной ДНК сообщает устойчивость к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, причем указанная мутация имеет место в положении в ДНК, соответствующем одному или более основаниям в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке, при условии, что указанная последовательность ДНК не является последовательностью Мyсеnа galopoda, кодирующей цитохром b.
В предпочтительном варианте этого аспекта данное изобретение обеспечивает также грибную последовательность ДНК, кодирующую весь мутантный белок цитохрома b или его часть, причем присутствие мутации в указанной ДНК сообщает устойчивость к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, причем указанная мутация имеет место в положении в ДНК, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке, при условии, что указанная последовательность ДНК не является последовательностью Mycena galopoda, кодирующей цитохром b.
В приведенных выше аспектах данного изобретения мутация, происходящая в положении в ДНК, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке, является предпочтительно заменой основания гуанина на основание цитозин.
Грибная последовательность ДНК, кодирующая весь мутантный белок цитохрома b или его часть, причем присутствие мутации в указанной ДНК сообщает устойчивость к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, в соответствии с вышеописанными аспектами данного изобретения, предпочтительно может быть получена или предпочтительно получена из гриба, выбранного из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Venturia inaequalis, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum, gloeosporioides, Oidium lycopersicum, Magnaporthe grisea, Phytophthora infestans, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola.
Данное изобретение охватывает также последовательности ДНК, содержащие все последовательности или часть последовательностей, обеспеченных в таблице 3, где остаток в положении в ДНК, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке, представляет собой остаток цитозина (SEQ ID NO: 176 - SEQ ID NO: 196). Эти последовательности показаны ниже и образуют следующий аспект данного изобретения:

Данное изобретение относится также к грибной последовательности ДНК, обнаруживающей гомологию или идентичность последовательности относительно указанной последовательности ДНК, содержащей указанный полиморфизм, и охватывает, например, вариации в последовательностях ДНК, обнаруженные в различных пробах одного и того же вида. Эти вариации могут, например, быть следствием использования альтернативного кодона, варьирующей митохондриальной организации интрон/экзон и замены аминокислот.
Последовательности ДНК, кодирующие весь белок цитохрома b дикого типа или мутантного цитохрома b или их часть, как описано здесь, находятся предпочтительно в изолированной форме.
Например, они являются частично очищенными от любого вещества, с которым они встречаются в природе. Эта последовательность ДНК является изолируемой (получаемой) или изолированной (полученной) из описанных здесь грибов.
Далее, данное изобретение обеспечивает считываемый компьютером носитель, имеющий хранящуюся на нем любую из описанных и заявленных здесь последовательностей и включающий в себя всю последовательность ДНК или ее часть, кодирующую описанный здесь мутантный белок цитохрома b, или последовательность или ее часть описанного здесь мутантного белка цитохрома b, причем присутствие мутации вызывает появление грибной устойчивости к аналогу стробилурина или любому соединению в той же самой группе перекрестной устойчивости; всю последовательность ДНК или ее часть, кодирующую мутантный белок цитохрома b, или белковую последовательность или ее часть мутантного белка цитохрома b, причем указанный белок сообщает грибную устойчивость к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, из гриба, выбранного из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Venturia inaequalis, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythiuin aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Magnaporthe grisea, Phytophthora infestans, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola, при условии, что указанная ДНК или белковая последовательность не является последовательностью Mycena galopoda, кодирующей цитохром b; всю последовательность ДНК или ее часть, кодирующую последовательность цитохрома b дикого типа, или белковую последовательность цитохрома b данного типа, из гриба, выбранного из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola; или любой аллель-специфический олиго-нуклеотид; аллель-специфический олигонуклеотидный зонд, аллель-специфический праймер, обычный или диагностический праймер, описанные здесь.
Считываемый компьютером носитель может быть использован, например, в скрининге гомологии, картировании, гаплотипировании, генотипировании или любом другом биоинформативном анализе. Любой считываемый компьютером носитель может быть использован, например компакт-диск, лента, гибкий диск, винчестер или компьютерные чипы.
Полинуклеотидные последовательности данного изобретения, или их части, в частности последовательности, относящиеся к идентифицированным здесь полиморфизмам единственного нуклеотида и идентифицирующие идентифицированным здесь полиморфизмы единственного полинуклеотида, в частности замену G→С в гене грибного цитохрома b, вызывающую замену С143А в кодируемом белке, представляют собой ценнный источник информации. Применение этого источника информации наиболее легко облегчается хранением информации последовательностей в считываемом компьютером носителе и затем использованием этой информации в стандартных программах биоинформатики. Полинуклеотидные последовательности данного изобретения применимы, в частности, в качестве компонентов в базах данных для анализа идентичности последовательности и других анализах с применением поиска. В применении здесь хранение информации последовательностей в считываемом компьютером носителе и применение в базах данных относительно полинуклеотида или полинуклеотидной последовательности данного изобретения включает в себя любую детектируемую химическую или физическую характеристику полинуклеотида данного изобретения, которая может быть сокращена, преобразована или сохранена в материальном носителе, таком как компьютерный диск, предпочтительно в считываемой компьютером форме. Например, данные хроматографического сканирования или данные пиков, данные фотографического сканирования или данные пиков, масс-спектрографические данные, данные геля по секвенированию (или другие данные).
Способ на основе компьютера обеспечен также для выполнения идентификации последовательности, причем указанный способ предусматривает стадии обеспечения полинуклеотидной последовательности, содержащей полиморфизм данного изобретения в считываемой компьютером носителе, и сравнения указанной содержащей полиморфизм полинуклеотидной последовательности по меньшей мере с одной другой полинуклеотидной или полипептидной последовательностью для идентификации идентичности (гомологии), т.е. скрининга на присутствие этого полиморфизма.
Далее, данное изобретение обеспечивает грибной белок цитохрома b, который сообщает грибную устойчивость к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, причем в указанном белке обычный остаток глицина изменен вследствие присутствия мутации в ДНК, кодирующей указанный белок, причем указанная мутация имеет место в положении в ДНК, соответствующем одному или более основаниям в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке, при условии, что указанная последовательность не является последовательностью цитохрома b Mycena galopoda.
В предпочтительном варианте этого аспекта данное изобретение обеспечивает также грибной белок цитохрома b, который сообщает грибную устойчивость к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, причем в указанном белке обычный остаток глицина изменен вследствие присутствия мутации в ДНК, кодирующей указанный белок, причем указанная мутация имеет место в положении в ДНК, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке, при условии, что указанная последовательность не является последовательностью цитохрома b Mycena galopoda.
Последовательность цитохрома b Mycena galopoda описана Kraiczy et al. (Eur. J.. Biochem. 235, 54-63 (1996)), и эта последовательность ДНК находится в базе данных EMBL accession No. X87997.
Остаток глицина в белке согласно вышеописанному аспекту данного изобретения заменен предпочтительно альтернативной аминокислотой, и указанная замена приводит к указанным грибам, обнаруживающим устойчивость к аналогу стробилурина или любого другого соединения в той же самой группе перекрестной устойчивости.
Мутация в соответствии с вышеописанным аспектом данного изобретения предпочтительно приводит к замене указанного остатка глицина аминокислотой, выбранной из группы, состоящей из аргинина, серина, цистеина, валина, аспарагиновой кислоты, глутаминовой кислоты и наиболее предпочтительно аланина.
В следующем аспекте данное изобретение обеспечивает антитело, способное узнавать указанный мутантный белок цитохрома b.
В следующем аспекте данное изобретение обеспечивает способ обнаружения мутации в грибном гене цитохрома b, приводящей к замене в кодируемом белке остатка глицина в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, причем указанный способ предусматривает идентификацию присутствия и отсутствия указанной мутации в пробе грибной нуклеиновой кислоты, где способ детектирования полиморфизма любого (или одного) единственного нуклеотида основан на информации последовательности от около 30 до 90 нуклеотидов против хода транскрипции и/или по ходу транскрипции от положения, соответствующего одному или более основаниям в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, либо в белке дикого типа, либо в мутантном белке.
В следующем предпочтительном варианте этого аспекта данное изобретение обеспечивает способ обнаружения мутации в грибном гене цитохрома b, приводящей к замене G143A в кодируемом белке, причем указанный способ предусматривает идентификацию присутствия и отсутствия указанной мутации в пробе грибной нуклеиновой кислоты, где способ детектирования полиморфизма любого (или одного) единственного нуклеотида основан на информации последовательности от около 30 до 90 нуклеотидов против хода транскрипции и/или по ходу транскрипции от положения, соответствующего второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, либо в белке дикого типа, либо в мутантном белке.
В следующем предпочтительном варианте этого аспекта данное изобретение обеспечивает способ обнаружения мутации гуанина в цитозин в грибном гене цитохрома b, приводящей к замене G143A в кодируемом белке, причем указанный способ предусматривает идентификацию присутствия и отсутствия указанной мутации в пробе грибной нуклеиновой кислоты, где способ детектирования полиморфизма любого (или одного) единственного нуклеотида основан на информации последовательности от около 30 до 90 нуклеотидов против хода транскрипции и/или по ходу транскрипции от положения, соответствующего второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, либо в белке дикого типа, либо в мутантном белке.
Информация последовательности согласно приведенному выше аспекту данного изобретения предпочтительно получена из гриба, выбранного из группы Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Venturia inaequalis, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Magnaporthe grisea, Phytophthora infestans, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola и, более предпочтительно из группы Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctania solani, Mycosphaerella musicola и Cercospora arachidola.
В применении здесь термин около 30 означает, что эта последовательность может содержать до 30 нуклеотидов, например 5, до 10, 15, 20 или 25 нуклеотидов, или может содержать более 30 нуклеотидов. В приведенных выше аспектах данного изобретения предпочтительно, чтобы используемая информация последовательности составляла около 30, предпочтительно 30 нуклеотидов выше по ходу транскрипции и/или ниже по ходу транскрипции от положения, соответствующего второму основанию триплета, кодирующего аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, либо в белке дикого типа, либо в мутантном белке.
Нуклеиновая кислота в соответствии с данным изобретением представляет собой предпочтительно ДНК, тест-проба нуклеиновой кислоты представляет собой, что удобно, препарат тотальной ДНК из грибного материала, препарат кДНК из грибного материала или сам грибной материал или экстракты растения или семян, содержащие грибную нуклеиновую кислоту. В этом изобретении авторы описывают обнаружение мутации С143А с использованием препарата тотальной ДНК, препарата кДНК и с прямым использованием материала грибных спор в качестве матрицы в этих ПЦР. Должно быть понятно, что тест-проба может равным образом быть последовательностью нуклеиновой кислоты, соответствующей последовательности в данной тест-пробе. То есть можно сказать, что весь район или его часть в нуклеиновой кислоте пробы могут быть сначала выделены или амплифицированы с использованием любого подходящего способа, такого как ПЦР, перед применением в способе данного изобретения.
Данное изобретение обеспечивает средства анализа мутаций в ДНК сельскохозяйственных полевых проб, которые по самому их происхождению являются значительно менее хорошо определенными по сравнению с аналогичной ситуацией, имеющей дело с пробами человека. С сельскохозяйственными полевыми пробами значительно более трудно работать, и они предъявляют технически больше требований для обнаружения мутационного события, встречающегося с низкой частотой среди очень большого количества ДНК дикого типа и/или посторонней ДНК из других организмов, присутствующих в полевом изоляте, по сравнению с пробой человека, которая обычно содержит ДНК только от одного индивидуума.
Любой подходящий фермент для полимеризации может быть использован при условии, что он не имеет присущей ему способности дискриминировать между нормальной и мутантной матричными последовательностями в какой-либо значимой степени. Примеры подходящих ферментов включают в себя термостабильные ферменты, которые не имеют значимой 3’-5’-экзонуклеазной активности, например ДНК-полимераза Taq, в частности ДНК-полимераза ’Ampli Taq Gold’™ (РЕ Applied Biosystems), фрагмент Stoffel или другие подходящим образом делегированные на N-конце модификации ДНК-полимераз Taq (Thermus aquaticus) или Tth (Thermus thermophilus).
В следующем аспекте данное изобретение обеспечивает аллель-специфический олигонуклеотид, способный связываться со всей последовательностью или частью последовательности грибной нуклеиновой кислоты, кодирующей белок цитохрома b дикого типа, выбранной из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Venturia inaequalis, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Magnaporthe grisea, Phytophthora infestans, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola, причем указанный олигонуклеотид содержит последовательность, которая узнает последовательность нуклеиновой кислоты, кодирующую остаток глицина в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
В предпочтительном варианте этого аспекта данного изобретения указанная последовательность грибной нуклеиновой кислоты выбрана из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum. gloeosporioides, Oidium lycopersicum, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola.
Грибная нуклеиновая кислота в соответствии с приведенными выше аспектами данного изобретения является предпочтительно ДНК.
В следующем аспекте данного изобретения авторы обеспечивают аллель-специфический олигонуклеотид, способный связываться со всей последовательностью или частью последовательности грибной нуклеиновой кислоты, кодирующей весь белок или часть белка мутантного цитохрома b, причем указанный олигонуклеотид содержит последовательность, которая узнает последовательность нуклеиновой кислоты, кодирующую аминокислоту, выбранную из группы аргинин, серин, цистеин, валин, аспарагиновая кислота, глутаминовая кислота, триптофан, и наиболее предпочтительно аланин, в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
В предпочтительном варианте этого аспекта данного изобретения авторы обеспечивают аллель-специфический олигонуклеотид, способный связываться со всей последовательностью или частью последовательности грибной нуклеиновой кислоты, кодирующей весь белок или часть белка мутантного цитохрома b, выбранной из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Venturia inaequalis, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Magnaporthe grisea, Phytophthora infestans, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola, причем указанный олигонуклеотид содержит последовательность, которая узнает последовательность нуклеиновой кислоты, кодирующую аминокислоту, выбранную из группы аргинин, серин, цистеин, валин, аспарагиновая кислота, глутаминовая кислота, триптофан, и наиболее предпочтительно аланин, в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
Грибная нуклеиновая кислота в соответствии с приведенными выше аспектами данного изобретения является предпочтительно ДНК.
В следующем аспекте данное изобретение обеспечивает аллель-специфический олигонуклеотидный зонд, способный детектировать полиморфизм грибного гена цитохрома b в положении в ДНК, соответствующем одному или более основаниям в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке.
В следующем предпочтительном варианте этого аспекта данное изобретение обеспечивает аллель-специфический олигонуклеотидный зонд, способный детектировать полиморфизм грибного гена цитохрома b в положении в ДНК, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке.
В следующих предпочтительных вариантах вышеописанного аспекта данного изобретения указанный полиморфизм представляет собой замену основания гуанина на основание цитозин, эта мутация находится в грибе, выбранном из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Venturia inaequalis, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Magnaporthe grisea, Phytophthora infestans, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola.
Аллель-специфический зонд имеет предпочтительно длину 17-50 нуклеотидов, более предпочтительно около 17-35 нуклеотидов и наиболее предпочтительно около 17-30 нуклеотидов.
Конструкция таких зондов будет очевидной для молекулярных биологов с обычной квалификацией и может быть основана на информации последовательности ДНК или РНК. Такие зонды имеют любую удобную длину, например до 50 оснований, до 40 оснований, более удобно до 30 оснований, например 8-25 или 8-15 оснований. Обычно такие зонды будут содержать последовательности оснований, полностью комплементарные соответствующему локусу дикого типа или вариантному локусу в данном гене. Однако в случае необходимости могут быть введены одно или несколько дефектов спаривания при условии, что при этом не происходит чрезмерного влияния на дискриминирующую способность данного олигонуклеотидного зонда. Зонды данного изобретения могут нести одну или несколько меток для облегчения детектирования.
Далее, данное изобретение обеспечивает нуклеотидные праймеры, которые могут детектировать нуклеотидные полиморфизмы в соответствии с данным изобретением.
Согласно другому аспекту данного изобретения обеспечен аллель-специфический праймер, способный детектировать полиморфизм гена цитохрома b в положении в ДНК, соответствующем одному или более основаниям в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке.
Согласно предпочтительному варианту этого аспекта данного изобретения обеспечен аллель-специфический праймер, способный детектировать полиморфизм гена цитохрома b в положении в ДНК, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, в белке.
В приведенных выше аспектах указанная мутация в последовательности ДНК является предпочтительно мутацией замены основания гуанина на основание цитозин.
В следующем аспекте данное изобретение обеспечивает аллель-специфический праймер, способный детектировать грибную последовательность ДНК, кодирующую белок цитохрома b дикого типа, выбранную из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Venturia inaequalis, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Magnaporthe grisea, Phytophthora infestans, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola, причем указанный праймер способен детектировать последовательность ДНК, кодирующую остаток глицина в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
В предпочтительном варианте этого аспекта данного изобретения указанная грибная последовательность ДНК выбрана из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Mycosphaerella fijiensis var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graminicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola.
В следующем аспекте данного изобретения авторы обеспечивают аллель-специфический праймер, способный детектировать грибную последовательность ДНК, кодирующую весь белок или часть мутантного белка цитохрома b, причем указанный аллель-специфический праймер способен детектировать последовательность ДНК, кодирующую аминокислоту, выбранную из группы аргинин, серин, цистеин, валин, аспарагиновая кислота, глутаминовая кислота, триптофан, и наиболее предпочтительно аланин, в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
В предпочтительном варианте этого аспекта данного изобретения авторы обеспечивают аллель-специфический праймер, способный детектировать грибную последовательность ДНК, кодирующую весь белок или часть мутантного белка цитохрома b, выбранную из группы, состоящей из Plasmopara viticola, Erysiphe graminis f.sp. tritici/hordei, Rhynchosporium secalis, Pyrenophora teres, Mycosphaerella graminicola, Venturia inaequalis, Mycosphaerella fijiensis.var. difformis, Sphaerotheca fuliginea, Uncinula necator, Colletotrichum graininicola, Pythium aphanidermatum, Colletotrichum gloeosporioides, Oidium lycopersicum, Magnaporthe grisea, Phytophthora infestans, Leveillula taurica, Pseudoperonospora cubensis, Alternaria solani, Rhizoctonia solani, Mycosphaerella musicola и Cercospora arachidola, причем указанный праймер способен детектировать последовательность ДНК, кодирующую аминокислоту, выбранную из группы аргинин, серин, цистеин, валин, аспарагиновая кислота, глутаминовая кислота, триптофан, и наиболее предпочтительно аланин, в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
Аллель-специфический праймер используют обычно с обычным праймером в реакции амплификации, такой как ПЦР-реакция, что обеспечивает дискриминацию между аллелями через селективную амплификацию одного аллеля в конкретном положении последовательности, например, как это используется в анализе ARMS.
Авторы теперь изобрели праймеры для мутации G143A в перечисленных выше видах грибов, которые, как было показано, детектируют надежно и хорошо специфические мутации. Эти праймеры детектируют замену оснований гуанин-цитозин в положении в ДНК, соответствующем второму основанию в триплете, кодирующем аминокислоту в положении, соответствующем остатку 143 цитохрома b S. cerevisiae в белке. Аллель-специфические праймеры называют здесь диагностическими праймерами. Таким образом, в следующем аспекте данное изобретение обеспечивает диагностический праймер, способный связываться с матрицей, содержащей нуклеотидную последовательность мутантного грибного цитохрома b, где конечный 3’-нуклеотид этого праймера соответствует нуклеотиду, присутствующему в указанной мутантной форме грибного гена цитохрома b, и присутствие указанного нуклеотида вызывает появление грибной устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости.
Диагностический праймер данного изобретения имеет предпочтительно длину по меньшей мере 20 нуклеотидов, наиболее предпочтительно 26 нуклеотидов, но он может иметь длину между 15 и 20 нуклеотидами.
В предпочтительном варианте вышеуказанного аспекта данного изобретения предпоследний нуклеотид (-2) праймера не является таким же, какой присутствует в соответствующем положении в последовательности цитохрома b дикого типа.
В следующем предпочтительном варианте нуклеотид -3 этого праймера не является таким же, какой присутствует в соответствующем положении в последовательности цитохрома b дикого типа.
Другие дестабилизирующие компоненты могут быть включены вместе с нуклеотидом -2 или -3.
В следующем особенно предпочтительном варианте указанного выше аспекта данного изобретения авторы обеспечивают диагностические праймеры, способные связываться с матрицей, содержащей нуклеотидную последовательность грибного цитохрома b мутантного типа, где конечный 3’-нуклеотид этого праймера соответствует нуклеотиду, присутствующему в указанной мутантной форме грибного гена цитохрома b, и где до 10, например до 8, 6, 4, 2, 1, остальных нуклеотидов могут варьироваться относительно последовательности дикого типа без значимого влияния на свойства диагностического праймера.
В следующем особенно предпочтительном варианте указанного выше аспекта данного изобретения авторы обеспечивают диагностические праймеры, содержащие приведенные ниже последовательности и их производные, где конечный нуклеотид на 3’-конце является идентичным последовательностям, приведенным ниже, и где до 10, например до 8, 6, 4, 2, 1, остальных нуклеотидов могут варьироваться без значимого влияния на свойства диагностического праймера.
Удобно, чтобы последовательность диагностического праймера точно соответствовала указанным ниже последовательностям. Предпочтительно, чтобы праймеры ARMS во всех аспектах данного изобретения имели длину 26 нуклеотидов. В большинстве перечисленных ниже праймеров предпоследний нуклеотид был изменен относительно последовательности cyt b дикого типа для дестабилизации праймера, чтобы сделать его вследствие этого более селективным в отношении желательной матрицы, и эти праймеры особенно предпочтительны в соответствии с данным изобретением. Специалисту с квалификацией в области конструирования праймеров будет очевидно, что основания, альтернативные обсуждаемым выше, или в дополнение к обсуждаемым выше, могут также варьироваться без вредного влияния на способность праймера связываться с матрицей.
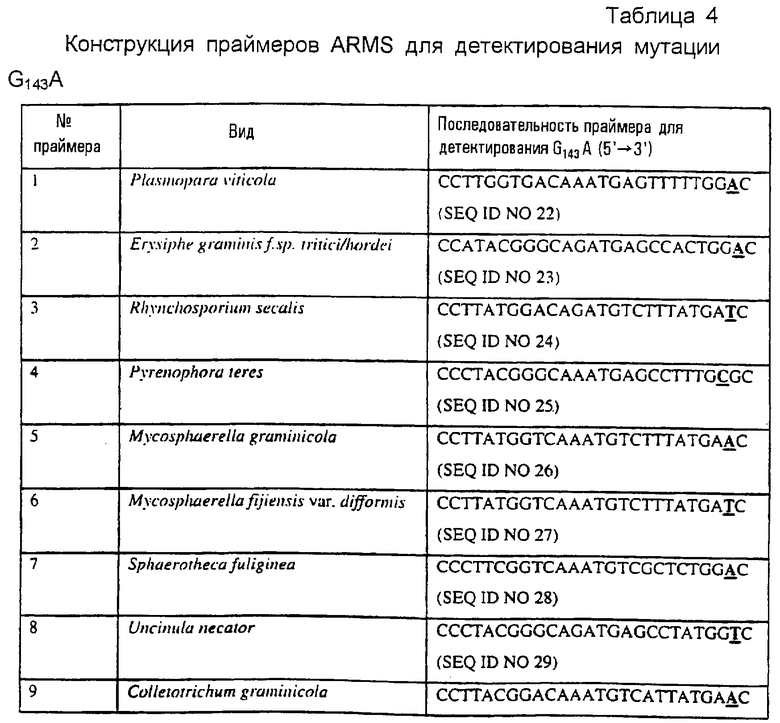

В качестве примеров праймеры, приведенные в таблице 4, включают в себя:
- ARMS-праймеры для Р. teres и V. ineaqualis, которые могут быть применимы эффективно либо к препаратам геномной ДНК, либо к биологическим пробам, в том числе грибным изолятам, грибным культурам, грибным спорам или инфицированному растительному материалу.
- ARMS-праймеры для S. fulginea, O. lycopersicum, L. taurica, U. necator, Phytophthora infestans, Magnoporthe grisea, R. solani, которые могут быть эффективными только с кДНК.
- ARMS-праймеры для Р. viticola, E. graminis f.sp tritici или hordei, R. secalis, M. graminicola, M. fijiensis var. difformis, C. graminicola, P. aphanidermatum, C. gloesporoides, P. cubensis, C. arachidola, Mycosphaerella musicola и A. solani, которые могут быть использованы эффективно либо с препаратами геномной ДНК, препаратами кДНК, либо с биологическими пробами, в том числе грибными изолятами, грибными культурами, грибными спорами или инфицированным растительным материалом.
кДНК-материал рекомендуется для видов, в которых организация интрон / экзон является в настоящее время неохарактеризованной около представляющего интерес полиморфизма единственного нуклеотида (SNP).
Для приспособления показанных в таблице праймеров для применения в стандартной ASPCR-реакции последнее основание на 3’-конце должно соответствовать точковой мутации без дестабилизации введенного основания.
Такие праймеры могут быть приготовлены с использованием любого подходящего способа синтеза. Примеры таких способов могут быть найдены в стандартных руководствах, например, "Protocols For Oligonucleotides And Analogues: Synthesis And Properties;" Methods In Molecular Biology Series; Volume 20; Ed. Sudhir Agrawal, Humana ISBN: 0-89603-247-7; 1993; 1st Edition.
Должно быть понятно, что удлинение диагностического праймера может быть сконструировано для указания отсутствия мутации, приводящей к замене G143A в кодируемом белке. Предпочтительным является использование ARMS-праймеров для детектирования отсутствия мутации, приводящей к замене G143A в кодируемом белке. Праймеры, сконструированные для этой цели, описаны здесь.
Детектирование последовательности дикого типа применимо в качестве контроля относительно детектирования мутации, и оно также необходимо, когда желательно количественное определение аллеля дикого типа и мутантного аллеля в пробе.
Таким образом, в следующем аспекте данное изобретение обеспечивает диагностический праймер, способный связываться с матрицей, содержащей нуклеотидную последовательность грибного цитохрома b дикого типа, где конечный 3’-нуклеотид этого праймера соответствует нуклеотиду, присутствующему в грибном гене цитохрома b дикого типа гриба, обнаруживающего чувствительность к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости.
В предпочтительном варианте этого аспекта данного изобретения предпоследний нуклеотид (-2) этого праймера не является таким же, какой присутствует в соответствующем положении в последовательности цитохрома b дикого типа.
В следующем предпочтительном варианте нуклеотид -3 этого праймера не является таким же, какой присутствует в соответствующем положении в последовательности цитохрома b дикого типа.
Вместе с нуклеотидом -2 или -3 могут быть включены другие дестабилизирующие компоненты.
Диагностический праймер данного изобретения предпочтительно имеет длину по меньшей мере 20 нуклеотидов, наиболее предпочтительно 26 нуклеотидов, но он может иметь длину между 15 и 20 нуклеотидами. В следующем особенно предпочтительном варианте указанного выше аспекта данного изобретения авторы обеспечивают диагностические праймеры, способные связываться с матрицей, содержащей нуклеотидную последовательность грибного цитохрома b дикого типа, где конечный 3’-нуклеотид этого праймера соответствует нуклеотиду, присутствующему в грибном гене цитохрома b, и где до 10, например до 8, 6, 4, 2, 1, остальных нуклеотидов могут варьироваться относительно последовательности дикого типа без значимого влияния на свойства диагностического праймера.
В следующем особенно предпочтительном варианте указанного выше аспекта данного изобретения авторы обеспечивают диагностические праймеры, содержащие приведенные ниже последовательности и их производные, где конечный нуклеотид на 3’-конце является идентичным последовательностям, приведенным ниже и где до 10, например до 8, 6, 4, 2, 1, остальных нуклеотидов могут варьироваться без значимого влияния на свойства диагностического праймера. Удобно, чтобы последовательность диагностического праймера точно соответствовала указанным ниже последовательностям. В большинстве перечисленных ниже праймеров предпоследний нуклеотид был изменен относительно последовательности цитохрома b дикого типа для дестабилизации праймера, чтобы сделать его вследствие этого более селективным в отношении желательной матрицы. Специалисту с квалификацией в области конструирования праймеров будет очевидно, что основания, альтернативные обсуждаемым выше, или в дополнение к обсуждаемым выше, могут также варьироваться без вредного влияния на способность праймера связываться с матрицей.

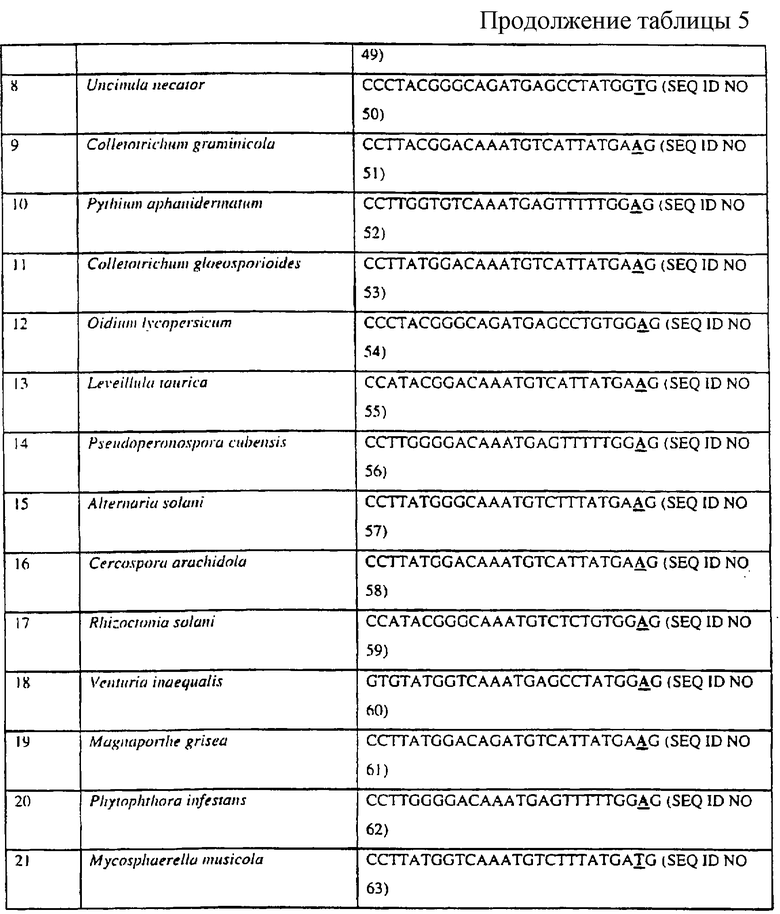
Для приспособления показанных в таблице праймеров для применения в стандартной ASPCR-реакции, последнее основание на 3’-конце должно соответствовать последовательности дикого типа без дестабилизации введенного основания.
Примеры, описанные выше, относятся к ARMS-праймерам на основе прямой (кодирующей) цепи ДНК. Применение ARMS-праймеров на основе прямой цепи ДНК является особенно предпочтительным.
АRМS-праймеры могут быть также на основе обратной цепи ДНК, если это желательно. Такие праймеры обратной цепи конструируют согласно тем же самым принципам, описанным выше для праймеров прямой цепи, а именно эти праймеры могут иметь длину по меньшей мере 20 нуклеотидов, наиболее предпочтительно 26 нуклеотидов, но они могут иметь длину между 15 и 20 нуклеотидами, и конечный 3’-нуклеотид этого праймера спаривается с релевантной матрицей, т.е. мутантной матрицей или матрицей дикого типа, и предпочтительно предпоследний остаток был изменен оптимально таким образом, чтобы он не образовывал пары с нуклеотидом релевантной матрицы. Кроме того, до 10, например до 8, 6, 4, 2, 1, остальных нуклеотидов в этом праймере могут варьироваться без значимого влияния на свойства диагностического праймера.
Во многих ситуациях будет удобным использование диагностического праймера данного изобретения с дополнительным праймером амплификации, называемым здесь обычным праймером, в одном или нескольких циклах ПЦР-амплификации. Подходящий пример этого аспекта представлен в Европейском патенте с номером ЕР-В1-0332435. Дополнительный праймер амплификации является либо прямым, либо обратным обычным праймером. Для каждого вида используемым праймером является праймер, указанный ниже. Праймеры, приведенные ниже, являются обратными праймерами.

В случае более длинных последовательностей, обеспеченных в таблице 6, специалист будет способен использовать эту информацию для конструирования подходящих праймеров.
Специалисту, квалифицированному в данной области, будет очевидно, что обычным праймером может быть любая подходящая патоген-специфическая последовательность, которая узнает комплементарную цепь гена цитохрома b (или другого представляющего интерес гена), лежащего 3’ от селективного праймера с мутацией.
Размер ПЦР-ампликона составляет преимущественно 50-400 п.н., но может быть иметь длину от 30 до 2500 п.н. или потенциально от 30 до 10000 п.н,
Может быть использован подходящий контрольный праймер, который сконструирован против хода транскрипции от положения G143A. Специалисту с квалификацией в данной области будет очевидно, что контрольный праймер может быть любым праймером, который не является специфическим для амплификации последовательностей мутации или последовательностей дикого типа. При использовании этих праймеров вместе с соответствующим обратным (’обычным’) праймером, описанным выше, амплицикация будет иметь место независимо от того, присутствует или не присутствует точковая мутация G143A.
Примеры конструкции возможных контрольных праймеров
Различные способы могут быть использованы для обнаружения присутствия или отсутствия продуктов удлинения диагностического праймера и/или продуктов амплификации. Эти способы будут очевидными для специалиста с квалификацией в области процедур детектирования нуклеиновых кислот. Предпочтительные способы не требуют применения радиоактивно меченых реагентов. Особенно предпочтительными способами детектирования являются способы на основе флуоресцентного детектирования присутствия и/или отсутствия продуктов удлинения диагностического праймера. Конкретные способы детектирования включают в себя анализ с использованием гель-электрофореза, детектирование продукта "Scorpions"™, описанное в заявке PCT GB 9803521, поданной от имени Zeneca Limited 25 ноября 1998 года, описания которой включены здесь в качестве ссылки, детектирование продукта "Таgman"™, например, описанное в патентах с номерами US A-5487972 и US A-5210015; детектирование продукта "Molecular Beacons"®, описанное в WO 9513399, и поверхностно усиленная резонансная спектроскопия Raman (SERRS), описанная в патентной заявке WO 9705280. Дополнительные предпочтительные способы детектирования включают в себя линейное удлинение ARMS (ALEX) и ПЦР с ALEX, описанные в опубликованной заявке РСТ с номером WO 99, 04037. Когда удобно используется детектирование реального времени. Использование детектирования продукта "Scorpions"™, описанное в заявке PCT GB 9803521 и опубликованной патентной заявке UK с номером GB 2338301, является особенно, предпочтительным для применения во всех аспектах данного изобретения, описанных здесь. Комбинирование технологии ARMS и Scorpion, описанное здесь, является особенно предпочтительным для применения во всех аспектах данного изобретения, описанных здесь, и предпочтительным способом детектирования является способ детектирования на основе флуоресценции. Многие из этих способов детектирования пригодны для количественной работы с использованием всех описанных выше праймеров. Эти различные ПЦР могут проводиться в различных пробирках или комплексно в одной пробирке. С использованием таких способов могут быть произведены оценки частоты молекул с точковыми мутациями, присутствующих в фоне молекул дикого типа.
Как приведено в качестве примеров здесь, авторы изобретения использовали ABMS-праймеры на основе прямой цепи ДНК в комбинации с детектированием Scorpion на основе обратной цепи ДНК, в качестве способа детектирования. Элементы детектирования Scorpion включают в себя предпочтительно обратные праймеры, показанные в таблице 6. Специалистам в данной области вполне понятно, что могут быть также использованы альтернативные комбинации ARMS-праймеров и элементов детектирования Scorpion. Например, праймер на основе прямой цепи ДНК может быть комбинацией ARMS-праймера и системы детектирования Scorpion, и это можно было бы использовать с обычным праймером на основе обратной цепи ДНК, или праймер на основе обратной цепи ДНК мог бы быть комбинацией ARMS-праймера с системой детектирования Scorpion, и эта комбинация могла бы быть использована с обычным праймером на основе прямой цепи ДНК.
В большинстве описанных здесь примеров элемент детектирования Scorpion находится на обычном праймере. ARMS-праймер, специфический для данной мутации и последовательности дикого типа, используют в комбинации с обычным флуоресцентно меченным праймером. Эти две реакции проводят в разных пробирках ПЦР и флуоресценция испускается, когда зонд связывается с генерированным ампликоном. Альтернативно, элемент Scorpion может быть помещен на ARMS-праймерах. В этом случае эти два ARMS-праймера могут быть меченными различными флуорофорами и использованы вместе с обычным праймером (в этот раз немеченым). Эти три праймера могут быть включены в одну и ту же реакцию, так как полученные ампликоны мутантного и дикого типа будут приводить к различной испускаемой флуоресценции.
Как описано в опубликованной патентной заявке Великобритании с номером GB 2338301, способ Scorpion может быть использован рядом различных путей, таких как вариант интеркаляции, где хвост праймера Scorpion несет интеркалированный краситель, который способен включаться между основаниями двухцепочечной молекулы нуклеиновой кислоты, в результате чего она становится высокофлуоресцентной; вариант FRET, где красители, включенные в праймер, образуют пару переноса электронов; вариант без гасителя, где флуорофор присоединен к хвосту праймера Scorpion; бимолекулярный вариант, где флуорофор и гаситель могут быть помещены на двух отдельных, но комплементарных молекулах; вариант с улавливающим зондом, где ампликоны могут специфически улавливаться и зондироваться с использованием одного и того же неамплифицируемого хвоста, и вариант Stem, где хвост праймера содержит самокомплементарные ножки. Эти варианты в полной форме описаны в опубликованной патентной заявке Великобритании с номером GB 2338301, описание которой включено здесь в качестве ссылки.
Способы данного изобретения, описанные здесь, надежно детектируют мутацию в грибном гене, где присутствие указанной мутации вызывает появление грибной устойчивости к фунгициду, белок-мишень которого кодируется митохондриальным геном, при уровне детектирования в диапазоне 1 мутированный аллель на 1000000 аллелей дикого типа - 1 мутированный аллель на 10000 аллелей дикого типа, и предпочтительно в диапазоне 1 мутированный аллель на 100000 аллелей дикого типа - 1 мутированный аллель на 10000 аллелей дикого типа. Способ данного изобретения может также детектировать мутации, встречающиеся при более высокой частоте, например 1 мутированный аллель на 100 аллелей дикого типа, 1 мутированный аллель на 10 аллелей дикого типа, или когда присутствуют только мутированные аллели. Подобным образом, способы данного изобретения могут быть использованы для детектирования частоты встречаемости аллеля дикого типа в фоне мутированных аллелей.
Комбинирование удлинения аллель-специфического праймера, сделанного более чувствительным с использованием технологии ARMS и способов количественного детектирования, которые используют в данном изобретении, делает этот способ крайне ценным способом для обнаружения грибных мутаций, встречающихся с низкой частотой.
Обнаружение аллелей, присутствующих в конкретных изолятах, позволяет связать результаты фенотипических биоанализов с профилем ДНК гена-мишени. Обнаружение единственной точковой мутации в качестве механизма устойчивости объясняет качественную природу этой устойчивости, а подтверждение последовательностей изолятов из единственной споры подтверждает точность скринингов в определении частот встречаемости устойчивых и чувствительных изолятов в испытуемых пробах.
Разработка способа комбинирования аллель-специфического удлинения праймера, специфичности ARMS и флуоресцентного детектирования реального времени, приводимых в качестве примеров здесь, с системой Scorpion позволяет анализировать большее число проб на присутствие мутации устойчивости, чем это было бы возможно в программе биоанализа. Большее число проб делает возможной идентификацию мутации устойчивости при частотах более низкого процента, чем можно было бы легко детектировать при помощи биоанализа. Это позволяет идентифицировать устойчивость в популяции раньше, чем она будет очевидной из полевых данных. Высокопроизводительный характер этого способа позволяет брать и тестировать пробы более широких площадей и больших участков, чем это было бы возможно с использованием биоанализа. Удлинение аллель-специфического праймера, например ARMS, связанное с флуоресцентным детектированиям реального времени, позволяет детектировать присутствие гена устойчивости в популяции до того, как эффекты этого гена могут оцениваться фенотипически с использованием биоанализа, в гетероплазматических и/или гетерокариотических клетках, что уменьшает ошибку классификации проб как чувствительных, когда они несут низкую частоту генотипа устойчивости. Результаты получают гораздо быстрее посредством одновременного считывания (реального времени) в сравнении с ожиданием развития болезни in planta, что позволяет быстро реагировать на полевые ситуации и рекоменовать более быстро предоставлять меры для преодоления устойчивости.
Один или несколько диагностических праймеров данного изобретения могут быть подходящим образом упакованы с инструкциями для применения в способах данного изобретения и подходящими упаковками и продаваться в виде набора. Эти наборы будут удобным образом включать в себя один или несколько из следующих компонентов: диагностический, дикого типа, контрольный и обычный олигонуклеотидные праймеры; подходящие нуклеотидтрифосфаты, например dATP, dCTP, dGTP и dTTP, подходящую полимеразу, описанную вьше, и буферный раствор.
В следующем аспекте данное изобретение обеспечивает способ обнаружения устойчивости фитопатогенного гриба к фунгициду, причем указанный способ предусматривает обнаружение мутации в грибном гене, где присутствие указанной мутации вызывает появление грибной устойчивости к аналогу стробилурина или любому другому соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает идентификацию присутствия или отсутствия указанной мутации в грибной нуклеиновой кислоте, с использованием способа детектирования полиморфизма любого (или одного) единственного нуклеотида.
В следующем варианте этого аспекта данное изобретение обеспечивает способ обнаружения устойчивости фитопатогенного гриба к фунгициду, причем указанный способ предусматривает детектирование присутствия ампликона, генерированного во время ПЦР-реакции, причем указанная ПЦР-реакция предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, причем детектирование указанного ампликона непосредственно связано с присутствием или отсутствием мутации в указанной нуклеиновой кислоте, где присутствие указанной мутации вызывает появление устойчивости к фунгициду, белок-мишень которого кодируется митохондриальыым геном.
В предпочтительном варианте этого аспекта данное изобретение обеспечивает способ обнаружения устойчивости фитопатогенного гриба к фунгициду, белок-мишень которого кодируется митохондриальным геном, предусматривающий контатктирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером для специфической мутации, присутствие которой вызывает появление устойчивости к указанному фунгициду, в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что диагностический праймер удлиняется, когда указанная мутация присутствует в пробе; и детектирование присутствия или отсутствия указанной мутации по присутствию или отсутствию продукта удлинения диагностического праймера.
В следующем предпочтительном варианте этого аспекта данное изобретение обеспечивает способ обнаружения устойчивости фитопатогенного гриба к фунгициду, белок-мишень которого кодируется митохондриальным геном, предусматривающий контатктирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером для специфической мутации, присутствие которой вызывает появление устойчивости к указанному фунгициду, в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что диагностический праймер удлиняется только в том случае, когда указанная мутация присутствует в пробе; и детектирование присутствия или отсутствия указанной мутации по присутствию или отсутствию продукта удлинения диагностического праймера.
Способы данного изобретения, описанные в указанных выше аспекте и вариантах, особенно пригодны для использования с фитопатогенными грибными штаммами, где присутствие мутации в гене цитохрома b вызывает появление устойчивости к фунгициду и, наиболее предпочтительно, устойчивости к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, и, наиболее предпочтительно, где мутация в грибной ДНК вызывает появление замены остатка глицина в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, более предпочтительно замены G143A в кодируемом белке, и особенно где мутация является заменой основания G на основание С во втором положении в кодоне, в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
В следующем аспекте данное изобретение обеспечивает способ обнаружения и количественного определения частоты мутации, вызывающей появление устойчивости фитопатогенного гриба к фунгициду, белок-мишень которого кодируется митохондриальным геном, причем указанный способ предусматривает обнаружение присутствия или отсутствия мутации в грибном гене, где присутствие указанной мутации вызывает появление грибной устойчивости к указанному фунгициду, причем указанный способ предусматривает идентификацию и количественное определение присутствия или отсутствия указанной мутации в грибной нуклеиновой кислоте с использованием способа детектирования полиморфизма любого (или одного) единственного нуклеотида.
В следующем предпочтительном варианте этого аспекта данное изобретение обеспечивает способ обнаружения и количественного определения частоты мутации, вызывающей появление устойчивости фитопатогенного гриба к фунгициду, белок-мишень которого кодируется митохондриальным геном, причем указанный способ предусматривает обнаружение присутствия или отсутствия мутации в грибном гене, где присутствие указанной мутации вызывает появление грибной устойчивости к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, причем указанный способ предусматривает идентификацию и количественное определение присутствия или отсутствия указанной мутации в грибной нуклеиновой кислоте с использованием способа детектирования полиморфизма любого (или одного) единственного нуклеотида.
В следующем варианте этого аспекта данное изобретение обеспечивает способ обнаружения и количественного определения частоты мутации, вызывающей появление устойчивости фитопатогенного гриба к фунгициду, белок-мишень которого кодируется митохондриальным геном, причем указанный способ предусматривает обнаружение присутствия ампликона, генерированного во время ПЦР-реакции, причем указанная ПЦР-реакция предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с подходящими праймерами в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, причем детектирование указанного ампликона непосредственно связано с присутствием или отсутствием мутации в указанной нуклеиновой кислоте, где присутствие указанной мутации вызывает появление устойчивости к фунгициду, белок-мишень которого кодируется митохондриальным геном, и детектирование и количественное определение относительного присутствия и отсутствия указанной мутации по присутствию или отсутствию ампликона, генерированного во время ПЦР-реакции.
В предпочтительном варианте этого аспекта данное изобретение обеспечивает способ обнаружения и количественного определения частоты мутации, вызывающей появление устойчивости фитопатогенного гриба к фунгициду, белок-мишень которого кодируется митохондриальным геном, предусматривающий контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическими праймерами для детектирования как присутствия, так и отсутствия специфической мутации в указанной нуклеиновой кислоте, присутствие которой вызывает появление устойчивости к указанному фунгициду, в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что диагностические праймеры, связанные с отстутствием и присутствием этой специфической мутации, удлиняются, когда подходящая грибная матрица присутствует в данной пробе; и детектирование и количественное определение относительного присутствия и отсутствия указанной мутации по присутствию или отсутствию продуктов удлинения диагностического праймера.
В следующем предпочтительном варианте этого аспекта данное изобретение обеспечивает способ обнаружения и количественного определения частоты мутации, вызывающей появление устойчивости фитопатогенного гриба к фунгициду, белок-мишень которого кодируется митохондриальным геном, предусматривающий контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическими праймерами для детектирования как присутствия, так и отсутствия специфической мутации в указанной нуклеиновой кислоте, присутствие которой вызывает появление устойчивости к указанному фунгициду, в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что диагностические праймеры, связанные с отсутствием и присутствием этой специфической мутации, удлиняются только в том случае, когда подходящая грибная матрица присутствует в данной пробе; и детектирование и количественное определение относительного присутствия и отсутствия указанной мутации по присутствию или отсутствию продуктов удлинения диагностического праймера.
Способы данного изобретения, описанные в указанных выше аспекте и вариантах, особенно пригодны для применения с фитопатогенными грибными штаммами, где присутствие мутации в гене цитохрома b вызывает появление устойчивости к фунгициду и, наиболее предпочтительно, устойчивости к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, и, наиболее предпочтительно, где мутация в грибной ДНК вызывает появление замены остатка глицина в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, более предпочтительно замены G143A в кодируемом белке, и особенно где мутация является заменой основания G на основание С во втором положении в кодоне, в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
Еще в одном аспекте данное изобретение обеспечивает способ выбора активного фунгицида и оптимальных уровней его применения к сельскохозяйственной культуре, предусматривающий анализ пробы гриба, способного инфицировать указанную культуру, и обнаружение и/или количественное определение присутствия и/или отсутствия мутации в гене из указанного гриба, где присутствие указанной мутации может вызывать появление устойчивости к фунгициду, белок-мишень которого кодируется митохондриальным геном, и затем выбора активного фунгицида и оптимальных уровней его применения.
В особенно предпочтительном варианте этого аспекта данного изобретения способ обнаружения предусматривает способ детектирования полиморфизма любого (или одного) единственного нуклеотида и более предпочтительно предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером для специфической мутации в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что этот диагностический праймер удлиняется, когда указанная мутация присутствует в пробе; и детектирование присутствия или отсутствия указанной мутации по присутствию или отсутствию продукта удлинения диагностического праймера, а количественное определение достигается контактированием тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическими праймерами для детектирования как присутствия, так и отсутствия специфической мутации в указанной нуклеиновой кислоте, присутствие которой вызывает появление устойчивости к фунгициду, белок-мишень которого кодируется митохондриальным геном, в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что эти диагностические праймеры, связанные с отсутствием и присутствием специфической мутации, удлиняются, когда подходящая грибная матрица присутствует в данной пробе; и детектирование и количественное определение относительного присутствия и отсутствия указанной мутации по присутствию или отсутствию продуктов удлинения диагностических праймеров.
В следующем особенно предпочтительном варианте этого аспекта данного изобретения способ обнаружения предусматривает контактирование тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическим праймером для специфической мутации в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что этот диагностический праймер удлиняется только в том случае, когда указанная мутация присутствует в пробе; и детектирование присутствия или отсутствия указанной мутации по присутствию или отсутствию продукта удлинения диагностического праймера, а количественное определение достигается контактированием тест-пробы, содержащей грибную нуклеиновую кислоту, с диагностическими праймерами для детектирования как присутствия, так и отсутствия специфической мутации в указанной нуклеиновой кислоте, присутствие которой вызывает появление устойчивости к фунгициду, белок-мишень которого кодируется митохондриальным геном, в присутствии подходящих нуклеотидтрифосфатов и агента полимеризации, так что эти диагностические праймеры, связанные с отсутствием и присутствием специфической мутации, удлиняются только в том случае, когда подходящая грибная матрица присутствует в данной пробе; и детектирование и количественное определение относительного присутствия и отсутствия указанной мутации по присутствию или отсутствию продуктов удлинения диагностических праймеров.
Способы данного изобретения, описанные здесь, особенно пригодны для применения с фитопатогенными грибными штаммами, где присутствие мутации в гене цитохрома b вызывает появление устойчивости к фунгициду и, наиболее предпочтительно, устойчивости к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, и наиболее предпочтительно где мутация в грибной ДНК вызывает появление замены остатка глицина в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, более предпочтительно замены С143А в кодируемом белке, и особенно где мутация является заменой основания G на основание С во втором положении в кодоне, в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
Еще в одном аспекте данное изобретение обеспечивает способ борьбы с грибной инфекцией сельскохозяйственной культуры, предусматривающий применение фунгицида к этой культуре, где указанный фунгицид выбран в соответствии с любым из способов выбора данного изобретения, описанных выше.
Способ данного изобретения, описанный выше, особенно пригоден для применения с фитопатогенными грибными штаммами, где присутствие мутации в гене цитохрома b вызывает появление устойчивости к фунгициду и, наиболее предпочтительно, устойчивости к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости.
Еще в одном аспекте данное изобретение обеспечивает анализ для обнаружения фунгицидно активных соединений, предусматривающий скрининг этих соединений против штаммов грибов, которые были тестированы на присутствие или отсутствие мутации, вызывающей появление устойчивости к фунгициду, белок-мишень которого кодируется митохондриальным геном, в соответствии со способами данного изобретения, описанными здесь, и затем определение фунгицидной активности против указанных штаммов грибов.
Способы данного изобретения, описанные здесь, особенно пригодны для применения с фитопатогенными грибными штаммами, где присутствие мутации в гене цитохрома b вызывает появление устойчивости к фунгициду и, наиболее предпочтительно, устойчивости к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости, и наиболее предпочтительно где мутация в грибной ДНК вызывает появление замены остатка глицина в положении, соответствующем остатку 143 цитохрома b S. cerevisiae, более предпочтительно замены G143A в кодируемом белке, и особенно где мутация является заменой основания G на основание С во втором положении в кодоне, в положении, соответствующем остатку 143 цитохрома b S. cerevisiae.
Путем применения способов данного изобретения, описанных здесь, может быть определена подходящая норма применения фунгицидов и/или подходящая комбинация фунгицидов для применения к данной сельскохозяйственной культуре.
Способы данного изобретения, описанные здесь, особенно пригодны для мониторинга грибной устойчивости к аналогу стробилурина или соединению в той же самой группе перекрестной устойчивости в таких культурах, как зерновые, плодовые и овощные культуры, такие как рапс, подсолнечник, табак, сахарная свекла, хлопчатник, соя, кукуруза, пшеница, ячмень, рис, сорго, томат, манго, персики, яблони, груши, земляника, бананы, дыни, картофель, морковь, латук, капуста, лук, виноград и газонная трава.
Способы данного изобретения, описанные здесь, являются особенно чувствительными для определения низких частот встречаемости мутаций в митохондриально кодируемых генах, таких как ген цитохрома b, что делает их особенно ценным и коммерчески важным путем скрининга фитопатогенных грибов на появление устойчивости к фунгицидам, где эта устойчивость является следствием мутации в митохондриально кодируемом гене.
Краткое описание чертежей
Теперь изобретение будет иллюстрироваться со ссылкой на следующие неограничительные примеры и чертежи, в которых:
Фигура 1 показывает схематическое представление вторичной структуры Scorpion-праймера Р. viticola с использованием программы MFold Zucker.
Фигура 2а показывает графическую иллюстрацию ПЦР-амплификации серийного разведения мутантной ДНК Р. viticola в фоне ДНК дикого типа с использованием С-специфического праймера и Scorpion-праймера (в двух повторностях). Как указано на фигуре, ромбы, представляющие 50% C/G, = линия 1, ромбы, представляющие 100% G, = линия 7, кружки, представляющие 10% С, = линия 2, кружки, представляющие 1% С, = линия 3, кружки, представляющие 0,1% С, = линия 4, кружки, представляющие 0,01% С, = линия 5 и кружки, представляющие NTC,=линия 6.
Фигура 2b показывает график, иллюстрирующий комплексный эксперимент с ARMS-праймерами Р. viticola с двумя серийными разведениями мутантной ДНК в фоне ДНК дикого типа (в двух повторностях). Как указано на этой фигуре, линия, представленная ромбами, показывает результаты для 1:100 Gfam, линия, представленная треугольниками, показывает результаты для 1:500 Gfam, линия, представленная кружками, показывает результаты для 1:100 Ctet и линия, представленная квадратами, показывает результаты для 1:500 Ctet.
Фигура 3а показывает график, иллюстрирующий тотальную ДНК Е. graminis, амплифицированную с тремя парами праймеров (специфическими G/C и контрольными праймерами) (в двух повторностях). Как показано на этой фигуре, линия, представленная ромбами, показывает результаты для 1/100-стандарта, линия, представленная треугольниками, показывает результаты для 1/100 Gmix и линия, представленная крестиками, показывает результаты для 1/100 Cmix.
Фигура 3b показывает график, иллюстрирующий амплификацию чувствительного изолята Е. graminis с тремя парами праймеров. Как показано на этой фигуре, линия, представленная ромбами, показывает результаты для Qmix, 1/100 = линия 1, линия, представленная квадратами, показывает результаты для Qmix, 1/1000 = линия 2, линия, представленная треугольниками, показывает результаты для Gmix, 1/100 = линия 3, линия, представленная кружками, показывает результаты для Gmix, 1/1000 = линия 4, линия, представленная крестиками, показывает результаты для Cmix, 1/100 = линия 5, линия, представленная ромбами, показывает результаты для Cmix, 1/1000 = линия 6.
Фигура 4 показывает график, иллюстрирующий амплификацию устойчивого изолята Е. graminis с тремя парами праймеров. Как показано на этой фигуре, линия, представленная ромбами, показывает результаты для Qmix, 1/100 = линия 1, линия, представленная квадратами, показывает результаты для Qmix, 1/1000 = линия 2, линия, представленная треугольниками, показывает результаты для Gmix, 1/100 = линия 3, линия, представленная ромбами, показывает результаты для Gmix, 1/1000 = линия 4, линия, представленная крестиками, показывает результаты для Cmix, 1/100 = линия 5, линия, представленная кружками, показывает результаты для Cmix, 1/1000 = линия 6.
Фигура 5а показывает график, иллюстрирующий ПЦР-амплификацию серийного разведения плазмиды R. secalis дикого типа, амплифицированной с помощью пары праймеров, специфических для дикого типа (в двух повторностях). Как показано на этой фигуре, линия, представленная ромбами, показывает результаты для G-плазмиды 10^8, линия, представленная треугольниками, показывает результаты для G-плазмиды 10^6, линия, представленная квадратами, показывает результаты для G-плазмиды 10^4 и линия, представленная кружками, показывает результаты для G-плазмиды 10^2.
Фигура 5b показывает график, иллюстрирующий ПЦР-амплификацию наивысшей концентрации плазмиды дикого типа и мутантной плазмиды R. secalis, амплифицированных с помощью пары праймеров, специфических для дикого типа (в двух повторностях). Как можно видеть из этой фигуры, линия, представленная ромбами, показывает результаты для G-плазмиды 10^8, а линия, представленная треугольниками, показывает результаты для С-плазмиды 10^8.
Фигура 6а показывает график, иллюстрирующий ПЦР-амплификацию серийного разведения мутантной плазмиды R. secalis, амплифицированной с помощью пары мутант-специфических праймеров (в двух повторностях). Как показано на этой фигуре, линия, представленная ромбами, показывает результаты для С-плазмиды 10^8, линия, представленная треугольниками, показывает результаты для С-плазмиды 10^6, линия, представленная квадратами, показывает результаты для С-плазмиды 4 и линия, представленная кружками, показывает результаты для С-плазмиды 10^2.
Фигура 6b показывает график, иллюстрирующий ПЦР-амплификацию наивысшей концентрации плазмиды дикого типа и мутантной плазмиды R. secalis, амплифицированных с парой мутант-специфических праймеров (в двух повторностях). Как можно видеть из этой фигуры, линия, представленная ромбами, показывает результаты для G-плазмиды 10^8, а линия, представленная треугольниками, показывает результаты для с-плазмиды 10^8.
Фигуры 7а, b и с показывают графики, иллюстрирующие ПЦР-амплификацию ДНК- и кДНК-матриц R.secalis в трех разведениях с парой G-праймеров (в двух повторностях). Как можно видеть из этих фигур, линия, представленная ромбами, показывает тотальную ДНК, а линия, представленная треугольниками, показывает кДНК.
Фигуры 8а, b и с показывают графики, иллюстрирующие ПЦР-амплификацию ДНК- и кДНК-матриц R.secalis в трех разведениях с парой С-праймеров (в двух повторностях). Как можно видеть из этих фигур, линия, представленная ромбами, показывает тотальную ДНК, а линия, представленная треугольниками, показывает кДНК.
Фигуры 9а и b показывают графики, иллюстрирующие амплификацию изолятов Р13 и Р15 Р. teres в двух разведениях с тремя парами праймеров (в двух повторностях). Как показано на фигуре 9а, ромбы, представляющие S-mix = линия 1, треугольники, представляющие S-mix 1/10, = линия 2, квадраты, представляющие G-mix, = линия 3, кружки, представляющие G-mix 1/10, = линия 4, кружки, представляющие С-mix=линия 5 и квадраты, представляющие C-mix 1/10, = линия 6. На фигуре 9b ромбы, представляющие s-mix, = линия 1, треугольники, представляющие s-mix 1/10, = линия 2, квадраты, представляющие g-mix=линия 3, кружки, представляющие g-mix 1/10, = линия 4, кружки, представляющие c-mix, = линия 5 и квадраты, представляющие c-mix 1/10, = линия 6.
Подробное описание изобретения
ПРИМЕРЫ
В первых четырех примерах ниже в качестве системы детектирования продуктов использовали систему Scorpion™ (AstraZeneca Diagnostics). Эта система детектирования описана в полном виде в РСТ-заявке с номером PCT GB 9803521, поданной от имени Zeneca Limited 25 ноября 1998 года, описания которой включены здесь в качестве ссылки. Эта новая система детектирования использует праймер с хвостом и интегрированную систему передачи сигнала. Этот праймер имеет связывающий матрицу район и хвост, содержащий линкер и мишеньсвязывающий район. При использовании мишеньсвязывающий район в хвосте гибридизуется с комплементарной последовательностью в продукте удлинения праймера. Это мишень-специфическое событие гибридизации сопряжено с системой передачи сигнала, в которой гибридизация приводит к детектируемому изменению. Способ детектирования этой системы предоставляет ряд существенных преимуществ над другими системами. Требуется только один вид праймера/детектора. Это обеспечивает как повышенную простоту, так и повышенную специфичность на основе легкой доступности мишеньсвязывающего района для гибридизации с продуктом удлинения праймера. Вновь синтезированный продукт удлинения праймера является мишень-специфическим, так что полученный выходящий сигнал прямо связан с количеством удлиненного праймера. Он не зависит от дополнительных событий гибридизации или ферментативных стадий. Интра- и интерцепочечная конкуренция за этот сайт зонда является ограниченной, так что конструкция зонда становится упрощенной. Поскольку это взаимодействие является одномолекулярным, реакция передачи сигнала является очень быстрой, что делает возможными увеличенные скорости циклов, что является существенным признаком для экспериментальной эффективности.
Scorpion-праймеры, сконструированные в примерах, описанных ниже, имеют следующие общие модификации:
- Мономер гексэтиленгликоля (НEG) в качестве блокирующей части молекулы, которая находится между связывающим матрицу районом праймера и районом хвоста, причем эта часть молекулы предотвращает опосредованное полимеразой копирование цепи района хвоста праймерной матрицы.
- Флуоресцентная молекула FAM добавлена к 5’-концу праймера. FAM является одной из флуоресцентных молекул, которые могут, например, легко детектироваться лазером 488 нм прибора ABI PRISM 7700 (РЕ Biosystems).
- MR является нефлуорогенным гасителем, присоединенным к остатку урацила.
Другие флуоресцентные молекулы и гасящие механизмы могут быть также приспособлены в конструкции Scorpion-праймеров и могли бы быть пригодными для применения в данном изобретении.
В примере 4 в качестве механизма детектирования использовали также интеркалирующий краситель. Восемнадцать примеров вместе описывают характеристику частичных последовательностей гена цитохрома b из панели важных грибных изолятов дикого типа, тогда как примеры 1, 2 и 6 описывают также характеристику частичных последовательностей гена цитохрома b из грибных изолятов, которые являются устойчивыми к аналогам стробилурина и другим соединениям в той же самой группе перекрестной устойчивости. Эти способы пригодны, в частности, для применения с аналогами стробилурина азоксистробином и пикоксистробином.
ПРИМЕР 1
В примере 1 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Plasmopara viticola, характеристику полиморфизма единственного полинуклеотида (SNP), который вызывает появление устойчивости к аналогу стробилурина в Р. viticola, и оценку ПЦР-Scorpion-анализа реального времени для мониторинга этого SNP в Р. viticola. Описан также комплексный подход к проведению ПЦР-анализа реального времени. Р. viticola является возбудителем ложной мучнистой росы у винограда.
Изолят Р. viticola дикого типа ES2B, чувствительный к аналогу стробилурина, собирали в 1996 году в Испании. Этот изолят никогда не подвергался селекции в отношении аналога стробилурина. Инфицированные листься виноградной лозы собирали вручную и хранили в полиэтиленовом мешке и посылали на исследовательскую станцию Jealott’s Hill Research Station (Zeneca Agrochemicals). По прибытии листья помещали парами вместе со спорулирующими поверхностями, не имеющими жилок, на влажную абсорбирующую бумагу в пластиковые боксы и инкубировали в течение 24-48 часов при 21-24°С. Отдельные повреждения вырезали из листьев и спорангии суспендировали в 5 мл деионизованной воды, затем разбрызгивали на безжилочные поверхности 5-6-недельных саженцев винограда (var. Ohanez). Свежеинокулированные растения инкубировали в течение 24 часов в увлажненной камере (при температуре окружающей среды, относительной влажности 100%), затем перемещали в помещение для выращивания (день 24°С / ком. темп. 60%; ночь 18°С / ком. темп. 95%; длина дня 16 часов, 6000 люкс). Растения возвращали в увлажненную камеру после 6 дней на дополнительноый период 24 часа, после чего успешное инфицирование обнаруживали в виде спорулирующих повреждений на безжилочных поверхностях листьев. Последующее субкультивирование проводили, как описано выше, с доведением суспензии спорангиев до 5000-10000 спорангиев на мл.
ДНК-фрагмент, кодирующий значительную часть последовательности цитохрома b P. viticola дикого типа, амплифицировали с использованием праймеров на основе консервативных районов генов цитохрома b Phytophthora megasperma и Ashergillus nidulans (Cytbl2F (5’ TGAACATATTATGAGAGATGT 3’) (SEQ ID NO: 106) и Cyt10R (5’AATTGCATAAAAAGGTAAAAA 3’) (SEQ ID NO: 107), которые отражают последовательность, кодирующую район аминокислот 66-281 грибного цитохрома b, на основе системы нумерации S. cerevisiae). ДНК экстрагировали из чувствительного к аналогу стробилурина изолята с использованием протокола экстракции смесью фенол/хлороформ. Спорангии промывали в 30 мл бидистиллированной Н2О (ddH2O) из шести листьев с 90-100% поражением болезнью (происходящих из искусственно инокулированных шестинедельных саженцев винограда). Суспензию спорангиев фильтровали через Мираклотс (Calbiochem cat # 475855) и центрифугировали при 3600 об/мин в течение 10 минут при 4°С. Затем спорангии замораживали в жидком азоте и растирали до мелкого порошка при помощи пестика и ступки, которые предварительно очищали и стерилизовали промыванием кислотой и автоклавированием. 0,5 мл лизисного буфера (200 мМ Трис-НСl (рН 8,5), 250 мМ NaCl, 25 мМ ЭДТА и 0,5% ДСН) добавляли и суспензию переносили в стерильную пробирку Эппендорфа на 1,5 мл с завинчивающейся крышкой. Немедленно добавляли 0,5 мл смеси фенол/хлороформ, изоамиловый спирт (25:24:1) и смешивали перевертыванием несколько раз. Пробирки центрифугировали в течение 30 минут при 14000 об/мин и водную фазу переносили в свежие пробирки Эппедорфа. Затем экстракцию смесью фенол/хлороформ, изоамиловый спирт повторяли, но в этот раз пробирки центрифугировали при 14000 об/мин в течение только 15 минут. После перенесения водной фазы в новую пробирку Эппендорфа выполняли последнюю экстракцию хлороформом. Затем грибную ДНК осаждали добавлением 0,1 объема 3 М ацетата натрия (рН 5,5) и 0,6 объема изопропанола. После перевертывания несколько раз пробирки центрифугировали при 14000 об/мин в течение 20 минут при 4°С. Затем осадок ДНК промывали дважды 70% этанолом, сушили в вакууме и ресуспендировали в 50 мкл ddH2O. Выход и качество ДНК проверялли гель-электрофорезом и готовили серийное разведение (1:10, 1:100 и 1:1000) в ddH2O для применения в качестве матричного материала в последующих ПЦР. ПЦР проводили, как рекомендовано изготовителем фермента полимеразы Taq (Gibco) в конечном реакционном объеме 100 мкл. Праймеры добавляли к реакциям до конечной концентрации 1 пмоль/мкл. 10 мкл каждого разведения ДНК добавляли к соответствующим ПЦР. Стандартные процедуры проводили для ограничения риска загрязнения ПЦР. 30 циклов (94°С в течение 45 секунд, 42°С в течение 45 секунд и 72°С в течение 1 минуты 30 секунд) проводили в приборе для ПЦР Hybaid Omn-E. Выполняли также начальную стадию при 94°С в течение 3 минут и конечное удлинение при 72°С в течение 10 минут. Затем эффективность ПЦР оценивали анализом 18 мкл реакционных смесей при помощи гель-электрофореза. Затем пробу 2 мкл ПЦР-продуктов ~500 п.н. клонировали в клонирующем векторе pCR2.1 TA-PCR (Invitrogen Catalogue No. K4500-01) и трансформировали в клетки Escherichia coli (компетентные клетки ТОР10 One Shot™) в соответствии с рекомендациями изготовителя. Ряд полученных клонов проверяли на присутствие вставок (инсертов) проведением Wizard minipreps (Promega catalogue No. A7100) в соответствии с инструкциями изготовителя и рестрикционного анализа с использованием EcoRI.6 клонов с подходящими вставками (~500 п.н.) затем секвенировали с использованием прямого и обратного праймеров М13 (автоматический секвенатор ABI377XL). При анализе данных нуклеотидных последовательностей из этих исследований с использованием релевантного программного обеспечения биоинформатики (Seqman, Editseq and Macaw) было обнаружено, что полученная новая последовательность кодирует новый ген цитохрома b с близкой гомологией с другими известными последовательностями цитохрома b оомицетов, таких как Р. megasperma. Последовательность содержащего 61 нуклеотид отрезка полученного Р. viticola, кодирующая 30 п.н. выше по ходу транскрипции и ниже по ходу транскрипции от второго основания С143-кодона (согласно системе нумерации аминокислот S. cerevisiae), может быть найдена в таблице 3.
Р. viticola-специфическими праймерами на основе вышеописанных анализов, используемыми в последующих амплификациях представляющего интерес района цитохрома b, были PLAS17F (5’ AAATAACGGTTGGTTAATTCG 3’ (SEQ ID NO: 108) и PLAS15R (5’ TCTTAAAATTGCATAAAAAGG 3’ (SEQ ID NO: 109), отражающие район аминокислот 73-283 грибного цитохрома b согласно системе нумерации S. cerevisiae.
Устойчивый к аналогу стробилурина изолят Р. viticola идентифицировали в одном месте испытаний. Инфицированные листья винограда собирали и обрабатывали, по существу как описано выше, хотя пробы субкультивировали в виде массовых популяций и не выделяли в виде отдельных повреждений перед испытанием. Способом испытания для подтверждения устойчивости к аналогу стробилурина было (за 24 часа) опрыскивание на 4-недельных саженцев винограда. Серию химических разведений готовили растворением 5 мг аналога стробилурина (аналог стробилурина в применении во всех примерах здесь обозначает азоксистробин) (технический материал, чистота 97%) в 1 мл ацетона и проведением последующего разведения в деионизованной воде при комнатной температуре с получением дозы 10 м.д. (доза, которая, как известно, дает 100% уничтожение чувствительного к аналогу стробилурина базового изолята). Безжилковые поверхности листьев-мишеней опрыскивали с использованием пистолета для разбрызгивания DeVilbiss, 10 фунтов на кв. дюйм, до максимального удерживания. Контрольные растения опрыскивали только деионизованной водой. Обработанные растения оставляли высыхать в помещении для выращивания (условия, как описано выше) в течение ночи. Инокуляцию тест-пробой Р. viticola проводили при 5000 спорангиев на мл и свежеинокулированные растения инкубировали, как описано ранее. Спустя 7 дней после инокуляции любое потенциально устойчивое новообразование на обработанных листьях выделяли и субкультивировали для обеспечения достаточного материала для ПЦР-анализа. Этот изолят был назван Т5.
Частичную последовательность гена цитохрома b амплифицировали с праймерами PLAS17F и PLAS15R из устойчивого изолята (Т5). Тотальную геномную ДНК (ядерную и митохондриальную) из этого изолята экстрагировали с использованием протокола экстракции со смесью фенол/хлороформ, описанного выше. Опять присутствие и качество ДНК проверяли гель-электрофорезом и подходящие аликвоты разводили 1:10 и 1:100 в ddH2O для ПЦР-исследований. Затем 10 мкл каждого разведения ДНК добавляли к ПЦР-гранулам с полимераэой Taq Ready To Go™ (Amersham Pharmacia Biotech product number 27-9555-01) и доводили до 25 мкл растворами праймеров PLAS17F и PLAS15R, до конечной концентрации праймера 1 пмоль/мкл в каждом случае. Выполняли стандартные процедуры для ограничения риска загрязнения ПЦР. 30 циклов 94°С в течение 45 секунд, 52°С в течение 45 секунд и 72°С в течение 1 минуты 30 секунд проводили в приборе для ПЦР Hybaid Omn-E. Выполняли также начальную стадию при 94°С в течение 3 минут и конечное удлинение при 72°С в течение 10 минут. Эти ПЦР выполняли в двух повторностях. После анализа 10 мкл этих ПЦР при помощи гель-электрофореза на 0,8% ТВЕ-агарозном геле полученные ПЦР-продукты ~500 п.н. объединяли перед клонированием. Затем пробу 1 мкл объединенных ПЦР-продуктов клонировали в клонирующем векторе pCR2.1 ТА (Invitrogen) и трансформировали в клетки Е.coli (компетентные клетки ТОР10 One Shot™) в соответствии с рекомендациями изготовителя. Ряд полученных клонов проверяли на присутствие вставок проведением Wizard minipreps (Promega) в соответствии с инструкциями изготовителя и рестрикционного анализа с помощью EcoRI.10 клонов со вставками правильного размера (~500 п.н.) затем секвенировали с использованием прямого и обратного праймеров М13 (автоматический секвенатор ABI377XL).
Анализ данных нуклеотидных последовательностей с использованием релевантного программного обеспечения для биоинформатики (Seqman, Editseq and Macaw) во всех 10 случаях выявил точковую мутацию G→С в последовательности цитохрома b при сравнении с последовательностью дикого типа. Эта точковая мутация ДНК приводит к единственной замене глицина на аланин в положении 143 (согласно системе кодирования аминокислот S. cerevisiae).
Были сконструированы различные специфические ARMS-праймеры Р. viticola для обнаружения присутствия или отсутствия точковой мутации G143A:
два прямых ARMS-праймера на основе последовательности дикого типа:

и контрольный праймер, сконструированный выше по ходу транскрипции от этой точковой мутации:
STAND: GCCTTGGGGACAAATGAGTTTTTG (SEQ ID NO: 114)
Во всех приведенных выше ARMS-праймерах основание -1(3’-концевое основание праймерной последовательности) соответствует сайту-мишени точковой мутации. Основания, представленные жирным шрифтом, отличаются от последовательности цитохрома b P.viticola. Во всех ARMS-праймерах (не в контрольном праймере) основание -20 было изменено из основания G в основание Т. Это было сделано для разрушения непрерывно расположенных оснований G. В праймерах G-sp-f-2 и C-sp-f-2 положение -2 было изменено из основания G в основание А. В G-sp-f-1 положение -3 было изменено из основания G в основание Т. В праймере C-sp-f-1 основание -2 было изменено из основания G в основание С. Эти изменения в последовательности производили для дестабилизации праймера и придания любому удлинению праймера большей специфичности в отношении соответствующей матрицы. Примеры в литературе показали, что дестабилизация ARMS-праймера уменьшает риск ошибочного праймирования праймера на ошибочной матрице (Newton et al., Nucleic Acid Research 17 (7) 2503-2516 1989).
Систему детектирования продуктов Scorpion™ использовали в этом случае в качестве механизма детектирования, и эта система детектирования была включена на обратном праймере. Полученный ампликон имел длину 234 п.н. с ARMS-праймерами и длину 235 п.н. с контрольным праймером. Scorpion-праймер был сконструирован с использованием программ Oligo 5 и MFold (MFold предсказывает оптимальную и субоптимальную вторичные структуры для молекул РНК или ДНК с использованием способа минимизации энергии по Zucker (Zucker, M. (1989) Science 244, 48-52; SantaLucia, J.Jr. (1998), Proc. Natl. Acad. Sci. USA 95, 1460-1465). Последовательность Scorpion-праймера P. viticola была:

Подчеркнутые районы являются образующими шпильку частями, FAM обозначает краситель флуоресцеин, MR обозначает нефлуорогенный гаситель, присоединенный к остатку урацила, и HEG обозначает блокирующий репликацию мономер гексэтиленгликоля. Последовательность, изображенная курсивом, является последовательностью обратного праймера, а последовательность, изображенная жирным шрифтом, является Scorpion-последовательностью, которая связывается с продуктом удлинения обратного праймера.
Вторичная структура петли со стеблем этого Scorpion-праймера может быть визуализирована с использованием программы MFold (см. фигуру 1). Прогнозируется, что она имеет энергию -2,2 ккал/моль в ее инертной форме. Однако в присутствии продукта удлинения структура шпильки расходится, так как последовательность зонда Scorpion-праймера связывается с этим продуктом удлинения с предсказываемой энергией -6,1 кал/моль. Это отделяет краситель FAM от его гасителя, вызывая испускание флуоресценции, детектируемой, например, при помощи прибора ABI PRISM 7700. Отжиг Scorpion-элемента к вновь синтезированной цепи является, следовательно, энергетически выгодным по сравнению с конфигурацией петли со стеблем Scorpion в ее инертном состоянии.
Все праймеры были синтезированы Службой Oswell DNA Service (Lab 5005, Medical and Biological Sciences Building, Southhampton). Перед использованием праймеры разбавляли до 5 мкМ в общем объеме 500 мкл ddH2O каждый. Затем праймеры дополнительно разбавлялись до конечной концентраци 500 нМ в ПЦР.
Во всех случаях фермент AmpliTaq Gold (Perkin-Elmer/ABI) включали в реакционную смесь при 1 единице/25 мкл реакции. Реакционная смесь содержала также 1х буфер (10 мМ Трис-НСl (рН 8, 3), 50 мМ КСl, 3,5 мМ MgCl2, 0,01% желатин) и 100 мкМ dNTP. Амплификации проводили в приборе ABI PRISM 7700 для непрерывного мониторинга флуоресценции. Предварительный цикл 95°С в течение 10 минут проводили с последующими 50 циклами 95°С в течение 15 секунд и 60°С в течение 45 минут. Флуоресценцию подвергали мониторингу во время стадии отжига удлинения во всех циклах.
Праймеры сначала оценивали для использования в таких анализах с использованием плазмидной ДНК в качестве матрицы при различных концентрациях. Это проводили для проверки специфичности и чувствительности конструкций праймеров. Частичную последовательность гена цитохрома b дикого типа и соответствующий отрезок, содержащий мутацию G143A, клонировали в вектор ТА pCR2.1 (Invitrogen) для использования в этом процессе оценки. Праймеры C-sp-f-2 и G-sp-f-2 были более предпочтительными, чем праймеры C-sp-f-1 и G-sp-f-1, так как ПЦР в двух повторностях давали более согласующиеся результаты и были слегка более специфическими. Во всех случаях плазмидную ДНК разбавляли в 1 мг/мл бычьем сывороточном альбумине (БСА).
График, показанный на фигуре 2а, иллюстрирует ПЦР, где разведение мутантной плазмиды в фоне матрицы дикого-типа амплифицировали с использованием ABMS-праймера C-sp-f-2. Поскольку разведение С-плазмиды уменьшается в фоне плазмиды дикого типа, детектирование флуоресценции задерживается. Во всех случаях конечная концентрация плазмиды в ПЦР была 1×107 молекул/мкл. С каждым 10-кратным разведением С-плазмиды имеется задержка на 3-4 цикла в детектировании флуоресценции, Когда С-плазмида присутствует при только 1 на 10000 копиях в плазмиде-фоне дикого типа, она все еще детектируется этим специфическим ARMS-праймером. Праймер C-sp-f-2 является полностью специфическим в отношении его соответствующей матрицы, так как флуоресценция не детектируется в плазмидной пробе 100% дикого типа в этом эксперименте.
Были исследованы различные возможные материалы, которые могли бы использоваться в качестве исходного материала для анализов мониторинга устойчивости, с использованием Scorpion-технологии. Три различных изолята использовали в этом исследовании. Для каждого их этих изолятов спорангии собирали как непосредственно из пораженных болезнью листьев винограда на открытом воздухе, так и из искусственно инокулированных росших в оранжерее листьев (как описано ранее). В обоих случаях спорангии смывали с листьев при помощи ddH2O. Затем спорангии собирали центрифугированием и держали при -80°С до использования. Затем пробы спорангиев ресуспендировали в 1 мл ddH2O и разводили 1:10 и 1:100 опять в ddH2O. ПЦР-анализы реального времени проводили, как описано выше, за исключением того, что к каждой ПЦР добавляли также бычий сывороточный альбумин (0,25 мкг/мкл), с использованием 5 мкл каждого разведения в качестве матрицы. Каждое разведение выполняли в двух повторностях, с C-sp-f-2, G-sp-f-2 и контрольным прямым праймером (каждым), добавляемым вместе с обычным обратным Scorpion-праймером. Применение спорангиев, собранных из листьев винограда, росшего в оранжерее, в качестве исходного материала давало хорошие результаты с согласующимися пороговыми величинами циклов (Ct). Применение спорангиев, собранных непосредственно из листьев винограда на открытом воздухе, давало плохие результаты, так что следующие два подхода использовали для улучшения качества этих данных: для уменьшения ингибирования в ПЦР (полевой материал будет содержать гораздо больше возможных примесей по сравнению с листьями, выросших в оранжерее растений) и для того, чтобы сделать эту ДНК более доступной для амплификации.
Для уменьшения действия ингибиторных компонентов на ПЦР концентрацию БСА, добавляемого к реакциям, увеличивали и к ПЦР добавляли детергент Твин-20. Для того чтобы сделать ДНК более доступной для амплификации, проводили начальную ПЦР и ПЦР-продукт из этой реакции использовали в качестве матрицы в ПЦР реального времени, разведения спор кипятили в течение 10 минут перед добавлением к ПЦР реального времени для улучшения клеточного лизиса и ДНК экстрагировали из спор с использованием 3 различных протоколов (Прибор для выделения ДНК от Genosys Biotechnologies Inc., DNAzol от Helena Biosciences и набор Qiagen DNeasy plant mini kit). Каждый способ испытывали поочередно со спорангиями или ДНК, разведенной 1:10, 1:100 и 1:1000 в ddH2O в качестве матрицы. Каждое разведение тестировали в трех повторностях с ARMS-праймерами C-sp-f-2 и G-sp-f-2 и контрольным праймером; все с обратным обычным Scorpion-праймером. 5 мкл каждой матрицы добавляли к ПЦР-тестам и условия были по существу такими же, как описанные ранее, но с добавлением 0,25 мкг/мкл БСА (когда концентрация БСА не была переменной). ДНК, экстрагированная с использованием DNAzol или Qiagen DNA preps, давала хорошие и согласующиеся результаты.
Частоту встречаемости аллелей G143 и A143 оценивали в изоляте Р. viticola, собранном из полевого испытания (обозначенного 17А). Спорангии собирали в смыве ddH2O 120 листьев и осаждали центрифугированием. Эта проба содержала приблизительно 2400 повреждений и 6,4×107 спорангиев. 12×106 спорангиев хранили при -80°С для применения для экстракции ДНК. Экстракцию геномной ДНК с использованием набора DNeasy plant Minikit (Qiagen catalogue No. 69103) проводили и полученную ДНК разводили 1:10 и 1:100 в ddH2O для применения в качестве матрицы для ПЦР-анализа реального времени. ПЦР-условия были такими же, как описанные выше. Полученные пороговые величины циклов (Ct) были 20 для G-специфического праймера и 34 с С-специфическим праймером, давая Ct-различие 14 циклов и, следовательно, частоту встречаемости устойчивых аллелей приблизительно 1 на 10000.
Оценивали также комплексный подход. В этом случае элемент Scorpion-детектирования помещали на ARMS-праймерах для получения возможности амплификации с помощью ПЦР аллелей G143 и A143 в одной и той же ПЦР. Последовательности Scorpion/ARMS-праймеров были следующими:
G143-аллель-специфический праймер:

Детали элемента детектирования Scorpion являются такими же, как описанные выше. А143-аллель-специфический праймер был помечен ТЕТ для различения между двумя этими ампликонами. Использовали обычный обратный (немеченый) праймер:
Обратный 5’CATAACCAGTCAACAACTTCTTTTCC 3’ (SEQ ID NO: 121).
Ампликон, генерированный в обоих случаях, имел длину 95 п.н. Scorpion-праймеры опять были сконструированы с использованием программ Oligo 5 и MFold (как описано выше). Другие компоненты ПЦР реального времени были такими же, как описанные выше, причем варьировались только концентрации праймеров во время оценки этого комплексного подхода. Посредством уменьшения конечной концентрации G FAM Scorpion до 100 нМ и поддержания конечной концентрации С ТЕТ Scorpion и обратного праймера при 500 нМ можно надежно детектировать соотношения по меньшей мере 1:500 C:G с использованием плазмидной ДНК в качестве матрицы (см. фигуру 2b).
ПРИМЕР 2
В примере 2 авторы описывают характеристику частичных последовательностей генов цитохрома b Erysiphe graminis f.sp. tritici и hordei, характеристику SNP, который вызывает появление устойчивости к аналогам стробилурина или соединениям в той же самой группе перекрестной устойчивости, и описание Scorpion-анализа ПЦР реального времени для мониторинга этого SNP в Е. graminis f.sp. tritici и hordei.
Изоляты Е. graminis f.sp. tritici и f.sp. hordei (возбудителей настоящей мучнистой росы пшеницы и ячменя) собирали в Северной Франции, Германии, Ирландии и Соединенном Королевстве Великобритании и Северной Ирландии, Сбор выполняли одним из двух способов: ручной сбор полевых листьев или взятие проб воздушных спор при помощи смонтированной на автомобиле струйной ловушки для спор (Burkard Manufacturing Co. Ltd., Rickmansworth, UK).
Листья пшеницы, инфицированные спорулирующей мучнистой росой, собирали из мест, где эти популяции подвергались действию аналогов стробилурина во множественных испытаниях. При возвращении в лабораторию листья помещали в полиэтиленовые мешки и инкубировали при 21°С в течение ночи. На следующий день пустулы повторно образовывали споры. Все пустулы субкультивировали извлечением из инфицированных листьев выше участков свежих листьев (пшеница cv Rapier, 9-дневная), помещали на фильтровальную бумагу (Whatman No. 1) в чашки Петри 9 см, содержащие 1,2% агар в воде. Свежеинокулированные чашки инкубировали в течение 7 дней и затем полученные колонии тестировали на чувствительность к аналогам стробилурина.
Для извлечения спор листья пшеницы срезали из 9-дневных растений (cv. Rapier) и помещали на 1,8% агар в воде в пластиковых чашках и выдерживали при 5°С до применения.
Струйную ловушку для спор монтировали наверху автомобиля и этот автомобиль разъезжал при скорости до 90 км час по заранее определенным дорогам в каждой стране. Пластиковые чашки, содержащие кусочки листьев, помещали у основания колонки ловушки для спор, чтобы позволить переносимым по воздуху спорам входить в ловушку и оседать на листьях. Чашки заменяли приблизительно через каждые 80 км. Как только серию кусочков листьев использовали в ловушке для спор, их переносили в квадратные чашки Петри (10 см2), содержащие 60 мл 1,8% агара в воде и фильтровальную бумагу, и хранили при 5°С.
По возвращении в Jealott’s Hill, листья, подвергнутые действию спор в ловушке для спор, инкубировали при постоянной температуре (длина дня 16 часов, свет 4-5000 люкс, температура 21°С, относительная влажность такая же, как в окружающей среде). После 5-6 дней после выдерживания в ловушке для спор можно было видеть, что образовывались пустулы Е. graminis (небольшие зоны пожелтения листового материала с последующим появлением мучнистых спорулирующих повреждений). Их либо субкультивировали на кусочках листьев в чашках Петри 9 см в виде “популяций” (одна популяция на стадию взятия проб), либо вырезали в виде изолята единственной пустулы и инкубировали отдельно на кусочках листьев в чашках Петри 5 см на 15 мл 1,2% агара в воде, покрытого фильтровальной бумагой. Кусочки листьев, инокулированные популяциями Е. graminis, инкубировали в течение 7 дней, после чего споруляция была достаточной для инокуляции в фенотипическом анализе устойчивости. Изоляты единственной споры инкубировали в течение 7 дней, но субкультивировали еще один раз для обеспечения достаточного количества материала для тестирования. Если споруляция была слабой, вышеописанный процесс повторяли, пока не получали хорошее (60-70%) спорулирующее покрытие болезнью на всех кусочках листьев, чтобы генерировать достаточное количество конидий для анализа.
Тестирование и последующее поддержание устойчивых изолятов проводили на отделенных от проростков пшеницы листьях, обработанных за 24 часа перед инокуляцией водным раствором аналога стробилурина при 5 м.д. (дозе, которая, как известно, дает 100% уничтожение чувствительных к стробилурину фоновых изолятов) и увлажняющим агентом Твином 20. Изоляты тестировали либо в виде массовых популяций, либо в виде изолятов одной пустулы. Конидии инокулировали в сухом виде на обработанные кусочки листьев. Инфицированный материал инкубировали в регулируемой среде (как описано выше) в течение 7 дней перед оценкой.
Любое новообразование (рост) на кусочках листьев, обработанных аналогом стробилурина при 5 м.д., рассматривали как потенциально устойчивое. Материал из этих повреждений дополнительно субкультивировали на обработанных аналогом стробилурина листьях для подтверждения устойчивости in planta и анализировали с использованием молекулярного анализа, описанного в данной заявке. Частоты встречаемости фенотипической устойчивости приблизительно 1 на 100 и выше можно было детектировать скринингом массовой популяции, а более точные частоты оценивали сравнением изолятов из одной споры, где новообразование на обработанных листьях было сравнимо с контролями (устойчивые), с чашками, где обработанные листья давали 100% уничтожение болезни (чувствительные).
Частичную последовательность гена цитохрома b E. graminis f.sp. tritici амплифицировали с использованием праймеров на основе консервативных районов генов цитохрома b Aspergillus niger и Neurospora crassa (Cytb3F (5’ CAGCTTCAGCTTTCTTCT 3’) SEQ ID NO: 122) и Cytb9R (5’ ACTTAAAGGTCTAAATTG 3’) (SEQ ID NO: 122), которые определяют последовательность, кодирующую аминокислотный район 86-322 грибного цитохрома b на основе системы нумерации S. cerevisiae). Приблизительно 500 мг конидий из чувствительного к аналогу стробилурина изолята, который не подвергался отбору в отношении аналога стробилурина, собирали извлечением непосредственно из листьев со спорулирующей болезнью в пробирки Эппендорфа на 1,5 мл. ДНК экстрагировали из этой конидиальной пробы с использованием протокола экстракции смесью фенол/хлороформ (см. выше). Присутствие и качество ДНК анализировали гель-электрофорезом и делали серийное разведение (1:10, 1:100 и 1:1000) в ddH2O для применения в качестве матричного материала в ПЦР-реакциях. ПЦР устанавливали, как рекомендовано изготовителем фермента полимеразы Taq (Gibco) в конечном объеме 100 мкл и праймеры добавляли к реакциям до конечной концентрации 1 пмоль/мкл. 10 мкл каждого разведения ДНК добавляли к соответствующим ПЦР. Тщательные процедуры предпринимали для того, чтобы ограничить риск загрязнения ПЦР. 30 циклов (94°С в течение 45 секунд, 42°С в течение 45 секунд и 72°С в течение 1 минуты 30 секунд) проводили в приборе HyBaid Omn-E. Выполняли также начальную инкубацию при 94°С в течение 3 минут и конечное удлинение при 72°С в течение 10 минут. Затем эффективность ПЦР оценивали анализом 18 мкл реакционных смесей гель-электрофорезом. Пробу 2 мкл ПЦР-продуктов ~500 п.н. клонировали в клонирующем векторе pCR2.1 ТА (Invitrogen) и трансформировали в клетки Е.coli (компетентные клетки ТОР10 One Shot™) в соответствии с рекомендациями изготовителя. Ряд полученных клонов проверяли на присутствие вставок проведением Wizard minipreps (Promega) в соответствии с инструкциями изготовителя и рестрикционного анализа с помощью EcoRI.6 клонов со вставками правильного размера (~500 п.н.) затем секвенировали с использованием прямого и обратного праймеров М13 (автоматический секвенатор ABI377XL). При анализе данных нуклеотидных последовательностей из этих исследований с использованием релевантного программного обеспечения для биоинформатики было обнаружено, что полученная новая последовательность кодирует новый ген цитохрома b с близкой гомологией с другими известными последовательностями цитохрома b аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований выше по ходу транскрипции и 30 оснований ниже по ходу транскрипции от второго основания С143-кодона (согласно системе нумерации аминокислот S. cerevisiae), может быть найден в таблице 3.
Специфические праймеры для Е. graminis, сконструированные на основе этой новой последовательности, использовали в последующих амплификациях представляющего интерес района цитохрома b. Этими праймерами были ERY11F (5’ ATGAACAATTGGTACAGTAAT 3’) (SEQ ID NO: 124) и ERY12R (5’ GTTAGGTATAGATCTTAATAT 3’) (SEQ ID NO: 125). Вместе они представляют последовательность, кодирующую район аминокислот 114-287 грибного цитохрома b согласно системе кодирования S. cerevisiae.
Частичную последовательность цитохрома b амплифицировали с праймерами ERY11F и ERY12R из популяции Е. graminis f. sp. tritici, устойчивой к аналогу стробилурина. Конидии (~200 мг) суспендировали в 200 мкл ddH2O и разводили 1:10, 1:100 и 1:1000 в ddH2O. 10 мкл каждого разведения конидий добавляли к ПЦР-гранулам с полимеразой Taq Ready To Go™ (Amershara Pharmacia Biotech product number 27-9555-01) и доводили до 25 мкл растворами праймеров ERY11F и ERY12R до конечной концентрации праймера 1 пмоль/мкл. Выполняли стандартные процедуры для ограничения риска загрязнения ПЦР. 30 циклов 94°С в течение 45 секунд, 52°С в течение 45 секунд и 72°С в течение 1 минуты 30 секунд проводили в приборе для ПЦР Hybaid Omn-E. Включали также начальную стадию при 94°С в течение 3 минут и конечное удлинение при 72°С в течение 10 минут. Все ПЦР в этом случае выполняли в трех повторностях. После анализа 10 мкл этих ПЦР при помощи гель-электрофореза на 0,8% ТВЕ-агарозном геле полученные ПЦР-продукты ~500 п.н. объединяли перед клонированием. Затем пробу 2 мкл объединенных ПЦР-продуктов клонировали в клонирующем векторе pCR2.1 ТА (Invitrogen) и трансформировали в клетки Е.coli (компетентные клетки ТОР10 One Shot™) в соответствии с рекомендациями изготовителя. Ряд полученных клонов проверяли на присутствие вставок (инсертов) проведением Wizard minipreps (Promega) в соответствии с инструкциями изготовителя и рестрикционного анализа с помощью EcoRI.10 клонов со вставками ожидаемого размера (~500 п.н.) затем секвенировали с использованием прямого и обратного праймеров М13 (автоматический секвенатор ABI377XL). Анализ данных последовательностей, полученных с использованием подходящего программного обеспечения для биоинформатики во всех 10 случаях, выявил точковую мутацию G (С в последовательности цитохрома b при сравнении с ранее полученной последовательностью гена цитохрома b Е. graminis f.sp. tritici дикого типа. Эта точковая мутация ДНК приводит к единственной замене глицина на аланин в положении 143 (согласно системе кодирования аминокислот S. cerevisiae).
Частичную последовательность гена цитохрома b характеризовали также из двух изолятов Е. graminis f.sp. hordei. Небольшие пробы конидий (~100 мг) извлекали из инфицированных листьев ячменя (которые получали, как описано ранее для пшеницы) в стерильные пробирки Эппендорфа. Эти конидиальные пробы хранили при -80°С до использования. Затем каждую пробу суспендировали повторно в 200 мкл ddH2O, дополнительно разводили 1:10 и 1:100 и 10 мкл каждого разведения использовали в качестве матриц для амплификации. Частичные последовательности гена цитохрома b амплифицировали с праймерами ERY11F и ERY12R. Условия и компоненты ПЦР были такими, как описанные ранее. При анализе гель-электрофорезом был обнаружен ПЦР-продукт ожидаемого размера (~500 п.н.). Этот продукт клонировали в вектор ТА pCR2.1 и секвенировали, как описано ранее. При анализе последовательности было обнаружено, что последовательность гена цитохрома b, амплифицированная из Е. graminis f.sp. hordei, была идентична последовательности из Е. graminis f.sp. tritici кроме 1 п.н. (замена Т на А) 379 п.н. ниже по ходу транскрипции от второго основания кодона G143. Эта замена оснований не приводит к замене аминокислоты в конечном белке (т.е. является молчащей мутацией).
31 различных изолятов Е. graminis исследовали с использованием вышеописанного протокола и во всех случаях обнаруженная последовательность совпадала с мутацией G143A, являющейся причиной устойчивости к аналогам стробилурина в Е. graminis f.sp. tritici. Аллель устойчивости A143 не был обнаружен в пробах Е. graminis f.sp. hordei.
Были сконструированы специфические ARMS-праймеры Е. graminis на основе приведенной выше информации о точковой мутации G143A:
прямой ARMS-праймер на основе последовательности дикого типа:

и контрольный праймер, сконструированный выше по ходу транскрипции от этой точковой мутации:
STAND2: GCCATACGGGCAGATGAGCCACTG (SEQ ID NO: 128)
В обоих праймерах G-sp-1 и C-sp-1 основание -1(3’-концевое основание) соответствует второму нуклеотиду кодона G143/A143. Основания в праймерах, которые отличаются от последовательности цитохрома b E. graminis дикого типа, обозначены жирным шрифтом. Положение -2 было изменено из основания G в основание А. Это было сделано для дестабилизации праймера, что является нормальным в ARMS-реакциях.
Систему детектирования продуктов Scorpion™ использовали в этом случае в качестве механизма детектирования. Опять Scorpion-праймер был сконструирован с использованием программ Oligo 5 и MFold. (подробности см. в примере 1). Последовательность Scorpion-праймера Е. graminis была:
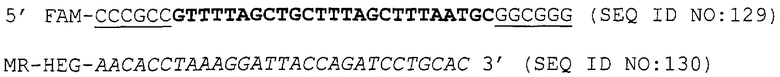
Подчеркнутые районы являются образующими шпильку частями, FAM обозначает краситель флуоресцеин, MR обозначает нефлуорогенный гаситель, присоединенный в остатку урацила, и HEG обозначает блокирующий репликацию мономер гексэтиленгликоля. Последовательность, изображенная курсивом, является последовательностью обратного праймера, а последовательность, изображенная жирным шрифтом, является Scorpion-последовательностью, которая связывается с продуктом удлинения обратного праймера.
Все праймеры были синтезированы Службой Oswell DNA Service. Перед использованием праймеры разбавляли до 5 мкМ в общем объеме 500 мкл каждый. Затем праймеры дополнительно разбавляли до конечной концентраци 500 нМ в ПЦР.
Праймеры сначала оценивали для использования в ARMS, Scorpion-анализах с использованием плазмидной ДНК и тотальной ДНК в качестве матрицы. Это проводили для проверки специфичности и чувствительности конструкций праймеров. ДНК-фрагменты, содержащие частичную последовательность гена цитохрома b дикого типа и соответствующую последовательность, содержащую мутацию G143A, клонировали в векторе ТА pCR2.1 для использования в процессе оценки. Плазмидную ДНК всегда разбавляли в 1 мг/мл БСА перед использованием в качестве матрицы в анализах ПЦР реального времени. Грибную ДНК для анализа экстрагировали из контрольного изолята Е. graminis f.sp. tritici, чувствительного к аналогу стробилурина, с использованием способа экстракции смесью фенол/хлороформ (как описано ранее). Затем конидиальную пробу Е. graminis f.sp. tritici из чувствительного к аналогу стробилурина изолята из Франции (F12C) и конидиальную пробу Е. graminis f.sp. tritici из устойчивого к аналогу стробилурина из Германии (11-8) тестировали с использованием проверенных праймеров при двух разведениях конидий. Все три грибных изолята происходили из отдельных пустул.
Во всех случаях фермент AmpliTaq Gold (Perkin-Elmer, ABI) включали в реакционную смесь при 1 единице/25 мкл реакции. Реакционная смесь содержала также 1х буфер (10 мМ Трис-НСl (рН 8,3), 50 мМ КСl, 3,5 мМ MgCl2, 0,01% желатин) и 100 мкМ dNTP. Амплификации проводили в приборе ABI PRISM 7700 для непрерывного мониторинга флуоресценции. Предварительную инкубацию 95°С в течение 10 минут проводили с последующими 50 циклами (95°С в течение 15 секунд и 60°С в течение 45 минут). Флуоресценцию подвергали мониторингу во время стадии отжига/удлинения во всех циклах.
При тестировании против контрольных матриц (плазмиды и ДНК из чувствительного к аналогу стробилурина контрольного изолята) ARMS/Scorpion-праймеры Е. graminis обнаружили хорошую специфичность без признаков ошибочного праймирования, происходящего из ошибочных матриц. На фигуре 3а показана амплификация чувствительной к аналогу стробилурина контрольной ДНК Е. graminis при разведении 1:100 с тремя смесями праймеров (Stand 2 + Scorpio Е. graminis, G-sp-1 + Scorpio E. graminis и C-sp-1 + Scorpio E. graminis). Каждую реакцию проводили в двух повторностях. Реакции с контрольным и G-специфическим праймерами испускают сильный сигнал флуоресценции, тогда как реакция с С-специфическим праймером не обнаруживает какого-либо увеличения флуоресценции. Контрольный и G-специфический ARMS-праймеры узнавали матрицу и связывались с ней, тогда как С-специфический праймер не связывался с матрицей, присутствующей в реакции. В этом случае анализ аллеля G143:A143 указывает присутствие только аллеля дикого типа.
Фигура 3b иллюстрирует ПЦР, где французский чувствительный изолят (F12C) анализировали с тремя смесями праймеров (Stand 2 + Scorpion E. graminis, G-sp-1 + Scorpion E. graminis и C-sp-1 + Scorpion E. graminis). В каждом случае ~200 мг конидий суспендировали в 200 мкл ddH2O и разводили 1:100 и 1:1000 в ddH2O. 5 мкл этих разведении добавляли к подходящим ПЦР. Опять при анализе реакции с контрольным и G-специфическим праймерами испускают сильный сигнал флуоресценции, тогда как реакция с С-специфическим праймером не обнаруживает какого-либо увеличения флуоресценции. Это указывает на то, что только аллель дикого типа детектировался в этой пробе. Существует определенная задержка во флуоресценции, продуцируемой при использовании конидий в качестве матрицы для этой реакции, по сравнению с использованием плазмидной ДНК в качестве матрицы. Это вызвано либо уменьшением копий молекул, которые могут быть использованы в качестве матрицы, присутствующих в этой реакции, либо ингибирующими компонентами, присутствующими в конидиальной пробе.
Фигура 4 иллюстрирует ПЦР, в которых устойчивый изолят из Германии в двух конидиальных разведениях амплифицировали с использованием трех смесей праймеров (Stand 2 + Scorpio E. graminis, G-sp-1 + Scorpio E. graminis и C-sp-1 + Scorpio E. graminis). В этом случае реакции с контрольным и С-специфическим праймерами испускают сильные сигналы, тогда как реакция с G-специфическим праймером не обнаруживает никакой флуоресценции. Это указывает на то, что только мутантный аллель G143A детектировался в данной пробе.
ПРИМЕР 3
В примере 3 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Rhynchosporium secalis и исследование ПЦР реального времени, в котором различные изоляты R. secalis подвергали скринингу на аллель устойчивости G143A. Это пример, в котором анализ G143A проводили на видах, в которых не была ранее идентифицирована эта точковая мутация.
Изоляты дикого типа R. secalis собирали в Соединенном Королевстве и Франции (см. таблицу 8: подробности для изолятов R. secalis). Изоляты К1124 и К3327 могут рассматриваться как “фон” (собранные до применения аналогов стробилурина в поле), а другие изоляты получали из мест испытаний, которые подвергали нескольким опрыскиваниям аналогами стробилурина на протяжении ряда сезонов. Инфицированные листья ячменя собирали вручную и хранили в пластиковом мешке и посылали в Jealott, s Hill Research Station (Zeneca Agrochemicals). По прибытии в лабораторию отдельные повреждения вырезали из листьев, поверхностно стерилизовали в этаноле (30 секунд) с последующей промывкой 0,1% раствором гипохлорита натрия (2 минуты) и затем помещали на агар-фасоль лима (см. таблицу 9) и инкубировали при цикле чередующегося 12 часов теплового излучения (365 нм Philips TLD 18W/08 - Philips Light Ltd., Croydon, UK)/12 часов без света при постоянной температуре 19°С в течение 4-5 дней. Колонии, выросшие из этих повреждений, субкультивировали в виде суспензии неподсчитанных спор на агаре-фасоли лима и инкубировали, как описано выше, в течение приблизительно 7 дней до спорообразования. Полученные споры удаляли и хранили в жидком азоте до использования изолятов. Восстановление изолятов достигалось посевом суспензии спор на агар-фасоль лима и инкубированием, как описано выше, в течение приблизительно 7 дней до получения спорообразования.
Подробности в отношении изолятов R. secalis
Частичную последовательность гена цитохрома b получали из двух изолятов R. secalis (K1124 и К3327). Эти изоляты были выращены из суспензии 100000 спор на мл, инокулированной в среду с неферментируемым источником углерода, встряхиваемую при 85 об/мин в течение 21 дня при 19°С (12 часов свет/12 часов темнота) и мицелии собирали фильтрованием через Мираклотс и замораживали при -20°С до применения. ДНК получали из этих мицелиев с использованием протокола экстракции смесью фенол/хлороформ (см. пример 1). Выход и качество ДНК проверяли гель-электрофорезом и затем готовили исходные серийные разведения (1:10, 1:100 и 1:1000 в ddH2O) для применения в качестве матриц в ПЦР-амплификациях. ПЦР проводили, как описано в предыдущих примерах. Использовали либо вырожденные праймеры (deg4F (5’ AGGTYTRTAYTRYGGDTCWTA 3’) (SEQ ID NO: 131) и deg3R (5’ AGCDATAACWCCTAATAATTT 3’) (SEQ ID NO: 132), сконструированные из гомологичных районов грибных генов цитохрома b (определяющие последовательность, кодирующую район аминокислот 100-294 согласно системе кодирования S. cerevisiae), либо Е. graminis-специфические праймеры ERY11F и ERY12R (детали, как в примере 2). Полосу ожидаемого размера (~500 п.н. амплифицировали с использованием обеих пар праймеров из каждого изолята и каждый ПЦР-продукт клонировали в векторе ТА pCR2.1 (Invitrogen). Wizard-minipreps осуществляли для идентификации клонов со вставками ожидаемого размера (~500 п.н.) и 5 клонов предоставляли для секвенирования из каждого клонирующего события с использованием прямого и обратного праймеров М13. При анализе с подходящим программным обеспечением по биоинформатике было обнаружено, что была идентифицирована новая последовательность гена цитохрома b, которая была близкородственной относительно других последовательностей генов цитохрома b аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
Различные специфические ARMS-праймеры R. secalis были сконструированы около местоположения точковой мутации G143А:
два прямых ARMS-праймера на основе последовательности дикого типа:

контрольный праймер, сконструированный против хода транскрипции от точковой мутации:
STAND3: TCCTTATGGACAGATGTCTTTATG (SEQ ID NO: 137)
В ARMS-праймерах основание -1(основание 3’-конца) соответствует второму нуклеотиду кодона С143/А143. Основания в праймерах, которые отличаются от последовательности цитохрома b R. secalis дикого типа, обозначены жирным шрифтом. Положение -2 изменено из основания G в основание А или Т. Это сделано для дестабилизации праймера, что является обычным для ARMS-праймеров.
Систему детектирования продуктов Scorpion™ использовали в этом случае в качестве механизма детектирования и опять обратный Scorpion-праймер был сконструирован с использованием программ Oligo 5 и MFold (подробности см. в примере 1). Последовательность Scorpion-праймера R. secalis была:

(См. предыдущие примеры в отношении подробностей).
Все праймеры были синтезированы Службой Oswell DNA Service. Перед использованием праймеры разбавляли до 5 мкМ в общем объеме 500 мкл каждый. Затем праймеры дополнительно разбавлялись до конечной концентрации 500 нМ в ПЦР.
Опять праймеры сначала оценивали для использования в ARMS, Scorpion-анализах с использованием плазмидной ДНК в качестве матрицы при различных концентрациях. Это проводили для проверки специфичности и чувствительности конструкций праймеров. Частичную последовательность гена цитохрома b дикого типа и соответствующую последовательность, содержащую мутацию G143A, клонировали в векторе ТА pCR2.1 для использования в процессе проверки. Поскольку эта мутация еще не была обнаружена в ДНК R. secalis, точковую мутацию A143 включали в последовательность с использованием стратегии сайт-направленного мутагенеза: точковую мутацию включали в конструкцию праймера и этот праймер использовали для амплификации представляющего интерес района с использованием клона дикого типа в качестве матрицы. ПЦР устанавливали с использованием стандартных способов, как описано ранее, и 30 циклов (94°С в течение 45 секунд, 56°С в течение 45 секунд и 72°С в течение 1 минуты 30 секунд) проводили в приборе для ПЦР Hybaid. Omn-E. Включали также начальную инкубацию при 94°С в течение 3 минут и инкубацию для конечного удлинения при 72°С в течение 10 минут. ПЦР-продукт ~370 п.н. клонировали в вектор ТА pCR2.1 и полученные клоны секвенировали для проверки на какую-либо индуцированную ПЦР ошибку перед применением в этом эксперименте.
Было рассчитано, что неразведенные препараты плазмидной ДНК имеют концентрации приблизительно 2×1011 молекул на мкл. Таким образом, два исходных препарата плазмид разводили до 2×107, 105, 103 и 101 молекул/мкл в 1 мг/мл БСА и аликвоты 5 мкл использовали из каждого разведения, что давало ~1×108, 106, 104 и 102 молекул плазмиды в соответствующих ПЦР. Условия и компоненты ПЦР были такими, как описанные ранее. На фигуре 5а, которая показывает серийное разведение G-плазмиды, детектируемой смесью G-праймера, детектирование флуоресценции задерживалось на ~4 цикла с каждым 10-кратным разведением плазмиды. Фигура 5b показывает наивысшую концентрацию G (дикого типа (wt)) и С (мутантной) кассеты, амплифицированной с помощью смеси G-sp-2-праймера. G-праймер не обнаруживает ошибочного праймирования С-матрицы до очень позднего срока в этой ПЦР, даже хотя концентрация ДНК-матрицы является высокой (~108 молекул матрицы в реакции). Таким образом, существенная задержка перед событием G-ошибочного праймирования демонстрирует, что этот праймер имеет хорошее окно специфичности. Набор C-sp-2 праймеров также обнаруживает хорошую специфичность при всех специфических и неспецифических разведениях плазмиды (фигуры 6а и 6b). Смеси праймеров G-sp-2 и C-sp-2 использовали в последующих экспериментах вместо смесей праймеров G-sp-3 и C-sp-3.
Вторая часть этого исследования заключалась в сравнении использования тотальной (геномной) ДНК и кДНК в качестве матрицы для ПЦР. Материал тотальной ДНК получали из изолятов R. secalis с использованием способа экстракции смесью фенол/хлороформ (как описано ранее). Тотальную РНК экстрагировали из 100 мг растертого мицелия с использованием мининабора PNeasy Plant minikit (Qiagen catalogue No. 74903) в соответствии с рекомендациями изготовителя. Затем синтез кДНК первой цепи проводили с 1 мкг тотальной РНК с использованием ОТ-ПЦР с набором Advantage RT-PCR kit (Clontech catalogue No. K1402-1) в соответствии с рекомендациями изготовителя. Получали пулы тотальной ДНК и кДНК из трех изолятов (К3278, К3274 и К3276). Пул тотальной ДНК использовали в качестве матрицы в разведениях 1:100, 1:1000 и 1:10000, а кДНК использовали в качестве матрицы в неразбавленном виде и в виде разведении 1:5 и 1:10. В каждом случае 5 мкл матрицы добавляли в реакции ПЦР. Условия ПЦР реального времени, описанные в примерах 1 и 2, использовали также здесь за исключением того, что в этом случае проводили 40 циклов ПЦР вместо 50. Фигуры 7а, b и с иллюстрируют результаты, полученные с матрицами тотальной ДНК и кДНК при трех разведениях (разведение 1: тотальная ДНК (1:100) и кДНК (неразбавленная); разведение 2: тотальная ДНК (1:1000) и кДНК (1:5); разведение 3: тотальная ДНК (1:10000) и кДНК (1:10)), амплифицированными с использованием смеси G-праймеров. Фигуры 8а, b и с иллюстрируют матрицы тотальной ДНК и кДНК, амплифицированные с использованием смеси С-праймеров. Использование тотальной ДНК вместо кДНК в качестве матрицы давало более чувствительные результаты с более ранними пороговыми величиными цикла. Для предоставления наилучшего шанса детектирования любой С-мутации, введения тотальной ДНК при разведениях 1:10 и 1:1000 были выбраны для последующих анализов. Не смогли обнаружить изменений флуоресценции при использовании смеси С-специфических праймеров в этой ПЦР из 40 циклов, что указывает на то, что в этих изолятах был обнаружен только аллель G143 дикого типа.
ПРИМЕР 4
В примере 4 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Pyrenophora teres и исследование, в котором анализ детектирования аллеля устойчивости G143A проводили на множестве изолятов Р. teres. Этот вид является видом, в котором не была обнаружена ранее мутация G143A. Этот пример разделен на два раздела: один, описывающий исследование ПЦР реального времени с использованием способа детекции с интеркалирующим красителем с применением кДНК-препаратов изолятов Р. teres, и другой, описывающий исследование ПЦР реального времени с использованием анализа детектирования Scorpion с препаратами геномной ДНК изолятов Р. teres.
Исследование ПЦР реального времени с использованием интеркалирующего красителя
Изоляты дикого типа Р. teres (возбудителя сетчатой пятнистости ячменя) собирали в Соединенном Королевстве и Франции во время 1994, 1996 и 1998 годов (см. таблицу 10 в отношении деталей изолятов Р.teres). Изоляты 1994 и 1996 могут рассматриваться как “фон” (собранные до применения аналогов стробилурина в поле), а изоляты 1998 получали из мест испытаний, которые подвергали нескольким опрыскиваниям аналогами стробилурина на протяжении ряда сезонов. Инфицированные листья ячменя собирали вручную и посылали в Jealott’s Hill Research Station (Zeneca Agrochemicals). По прибытии в лабораторию листья инкубировали во влажной среде при 21°С в течение 24-48 часов. Отдельные повреждения вырезали из листьев, поверхностно стерилизовали в этаноле (30 секунд) с последующей промывкой 0,1% раствором гипохлорита натрия (2 минуты) и затем помещали на агар-роза чайная китайская (см. таблицу 9) и инкубировали при режиме чередовании 12 часов теплового излучения, 12 часов без света при постоянной температуре 22°С в течение 4-5 дней. Все среды, описанные в таблице 9, стерилизовали автоклавированием при 121°С в течение 15 минут при 15 фунтах на кв. дюйм. Колонии, выросшие из этих повреждений, субкультивировали с использованием пробки мицелия на V8+стрептомицин-агаре (см. Таблицу 9) и инкубировали, как описано выше, в течение 14 дней.
Рецепты сред
Карбонат кальция (Fisher)
Агар №3 (Oxoid)
Деионизованная вода
3 г
20 г
800 мл
Полученный материал мицелия и спор удаляли и хранили в жидком азоте до момента использования изолятов. Восстановление изолятов достигалось посевом хранящихся грибных проб на V8-arap и инкубированием, как описано выше, в течение 14 дней до получения спорообразования.
Подробности для изолятов Р. teres
Частичную последовательность цитохрома b идентифицировали из двух изолятов (К1056 и К1916). Исходную последовательность дикого типа получали из биологической пробы, полученной инокуляцией картофельно-декстрозного бульона (см. таблицу 9) суспензией мацерированного мицелия. Колбу инкубировали при 85 об/мин на орбитальном шейкере при режиме 12 часов белого света, 12 часов без света при постоянной температуре 19°С в течение 21 дня, пока не получали достаточно материала мицелия для протокола экстракции ДНК. Затем мицелий собирали фильтрованием через Мираклотс и замораживали при -20°С до использования. Тотальную ДНК получали с использованием протокола экстракции смесью фенол/хлороформ (см. пример 1). После проверки выхода и качества ДНК гель-электрофорезом делали серийное разведение (1:10, 1:100 и 1:1000 в ddH2O) для применения в качестве ПЦР-матриц. Многочисленные комбинации праймеров (специфических и вырожденных) были подвергнуты испытанию и в большинстве случаев не получали амплифицированного продукта. Однако пара праймеров, которые были сконструированы к консервативным последовательностям цитохрома b Aspergillus niger и Neurospora crassa (Cytb3F и Cytb9R, см. пример 2 в отношении деталей) действительно дали ПЦР-продукт ~500 п.н. Этот продукт клонировали в векторе ТА pCR2.1 (Invitrogen) и 6 клонов секвенировали с использованием прямого и обратного праймеров М13. При анализе результатов было обнаружено, что этот обратный праймер правильно связывался с последовательностью цитохрома b, тогда как прямой праймер ошибочно праймировал в ДНК-последовательности с интрон-подобными признаками. Р. teres-специфический праймер (Pt2R: 5’ СТТ АСА ТСТ GTA ATA GGT ААТ 3’} (SEQ ID NO: 140) был сконструирован в новом отрезке гена цитохрома b и его использовали с прямым праймером, который конструировали на основе последовательности гена цитохрома b Venturia inaequalis (Pt5F:5’’ TGT TAC TTT AGC ААТ GCA СТА 3’) (SEQ ID NO: 141) на кДНК матрице Р. teres. кДНК получали из мицелиальных проб этих двух изолятов. Тотальную РНК экстрагировали из 100 мкг растертого мицелия с использованием набора PNeasy Plant kit (Qiagen) в соответствии с рекомендациями изготовителя. Синтез кДНК первой цепи проводили с 1 мкг тотальной РНК с использованием ОТ-ПЦР с набором Advantage RT-PCR Clontech (в соответствии с рекомендациями изготовителя). 5 мкл полученной кДНК использовали затем в ПЦР. Компоненты и условия ПЦР были такими, как описанные ранее. ПЦР-продукт ~800 п.н. амплифицировали для обоих изолятов (соответствующий последовательности, кодирующей район аминокислот 48-310 грибного цитохрома b согласно системе нумерации S. cerevisiae). В обоих случаях ПЦР-продукт клонировали в векторе ТА pCR2.1 (Invitrogen) и 4 клона, которые содержали вставки ожидаемого размера (~800 п.н,), секвенировали с использованием прямого и обратного праймеров М13 для каждого изолята. Анализ данных последовательностей выявил, что была изолирована новая последовательность гена цитохрома b, которая является близкородственной другим известным последовательностям цитохрома b аскомицетов. Отрезок кДНК из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
Специфические ARMS-праймеры Р. teres были сконструированы около местоположения точковой мутации G143A:
прямой ARMS-праймер на основе последовательности дикого типа:
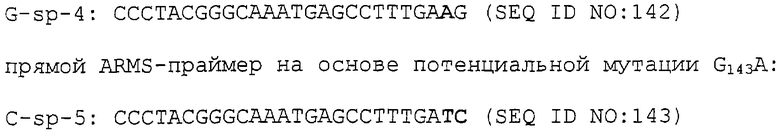
и контрольный праймер, сконструированный против хода транскрипции от точковой мутации:
STAND4: ACCCTACGGGCAAATGAGCCTTTG (SEQ ID NO: 144)
обратный праймер был:
UNLS4: TACACCTAAAGGATTTCCTGACCCTGCAA (SEQ ID NO: 145)
Опять в ARMS-праймерах основание -1 (основание 3’-конца) соответствует второму нуклеотиду кодона С143/А143. Основания в праймерах, которые отличаются от последовательности цитохрома b P. teres дикого типа, обозначены жирным шрифтом. Положение -2 изменено из основания G в основание А или Т. Это было сделано для дестабилизации праймера.
Все праймеры были синтезированы Службой Oswell DNA Service. Перед использованием праймеры разбавляли до 5 мкМ в общем объеме 500 мкл ddH2O каждый. Затем праймеры дополнительно разбавлялись до конечной концентрации 500 нМ в ПЦР.
В этом случае использовали интеркалирующий краситель для детектирования ПЦР-продукта. Использованным красителем был краситель YO-PRO-1 (Molecular Probes, Seattle Washington, USA), который связывается с двухцепочечной ДНК и испускает флуоресценцию, которая может детектироваться прибором ABI Prism 7700.
Праймеры сначала оценивали на валидность с использованием плазмидной ДНК в качестве матрицы при различных концентрациях. Это выполняли для проверки специфичности и чувствительности конструкций праймеров. Частичную последовательность гена цитохрома b Р. teres дикого типа и соответствующую последовательность, содержащую мутацию G143A, введенную сайт-направленным мутагенезом, клонировали в векторе ТА pCR2.1 для использования в процессе проверки (как описано в примере 3).
Было рассчитано, что неразведенные препараты плазмидной ДНК имеют концентрации приблизительно 2×1011 молекул на мкл. Эти две плазмиды разводили до 2×107, 105, 103 и 101 молекул/мкл и аликвоты 5 мкл использовали из каждого разведения, что давало ~1×108, 106, 104 и 102 молекул плазмиды в соответствующих ПЦР (см. пример 1 в отношении условий ПЦР). Единственное различие заключалось в том, что к реакционной смеси добавляли краситель YO-PRO-1. Генерирование димерного продукта праймеров интерферировало в сигнал после цикла 35. Этот способ детектирования является менее чувствительным, чем система детектирования Scorpion, так как на нее больше влияет фоновый шум (например, испускание флуоресценции из образования димера праймеров). Тем не менее, был сделан вывод, что ценная информация могла бы быть получена из использования данного способа, но следует проявлять повышенную осторожность в интерпретации результатов.
В следующем эксперименте описанные выше изоляты проверяли на присутствие аллеля G143A. Изоляты выращивали в среде с неферментируемым источником углерода для получения материала для ARMS, Scorpion-анализов. Исходную суспензию спор инокулировали (1 мл при 100000 спор/мл) в конические колбы, содержащие бульон. Культуры инкубировали на орбитальном шейкере, как описано ранее, и полученные мицелии собирали после этого для применения в получении кДНК. Материал кДНК получали (как описано ранее) из каждого из изолятов. Это выполняли, чтобы избежать конструирования праймеров в комплексной организации интрон/экзон последовательности цитохрома b P. teres.
Все ПЦР устанавливали, как описано выше, и проводили 50 циклов. Единственным различием было то, что краситель YO-PRO-1 добавляли к реакционной смеси. кДНК, неразведенную и разведенную 1:10, использовали в качестве матрицы, во всех случаях 5 мкл матрицы добавляли к ПЦР-реакции.
Фигуры 9а и b иллюстрируют результаты ПЦР-амплификации двух из одиннадцати изолятов, испытанных в двух разведениях, в двух повторностях с тремя парами праймеров. Ни в одном случае не были обнаружены детектируемые уровни аллеля G143A, присутствующего в испытанных пробах.
Исследование ПЦР реального времени с использованием системы детектирования Scorpion
Диагностический Scorpion-анализ С143А конструировали также на основе геномной организации, которая была определена для гена цитохрома b P. teres, кодирующего представляющий интерес район аминокислот.
Организацию интрон/экзон около представляющего интерес основания выясняли проведением ПЦР-амплификации на препаратах геномной ДНК с рядом праймеров, сконструированных к известной кодирующей последовательности и к идентифицированным затем последовательностям интронов. Добавку Taq Extender™ Additive и ПЦР-усилитель Perfect Match® PCR Enhancer (оба из Stratagene catalogue No. 600148 и 600129 соответственно) использовали в соответствии с инструкциями изготовителя в амплификации больших ПЦР-фрагментов с применением множества различных комбинаций праймеров. После начальной инкубации при 94°С в течение 3 минут проводили 30 циклов (94°С в течение 45 секунд, 52°С в течение 45 секунд и 72°С в течение 3 минут) на приборе Hybaid Omn-E. Включали также конечную инкубацию при 72°С в течение 10 минут. При анализе ПЦР-продуктов с использованием гель-электрофореза ПЦР-продукт 2,7 т.п.н. был обнаружен с парой праймеров pter23F (кодирующий район 5’ АСА TAG ТАА ТАС TGC TTC AGC 3’) (SEQ ID NO: 146) - pter 25R (район интрона 5’ ТАС АТТ TGA GGC ААА ТАТ TTC 3’ (SEQ ID NO: 147) и ПЦР-продукт 7,5 т.п.н. был обнаружен с парой праймеров pter7F (кодирующий район 5’СТА CGG GCA ААТ GAG CCT TTG 3’) (SEQ ID NO: 148) - pter6R (кодирующий район 5’ СТС TGG AAC TAT CGC TGC AGG 3’) (SEQ ID NO: 149). Эти ПЦР-продукты клонировали в векторе pCR-XL-TOPO (Invitrogen Catalogue No. K4700-10) в соответствии с инструкциями изготовителя и 2 клона для каждого клонирующего события секвенировали, как описано ранее. Было обнаружено, что второе основание кодона G143 расположено при 3’-конце экзона 36 п.н. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
Ряд ARMS-праймеров 31 п.н. и контрольный праймер (для амплификации как мутантного аллеля, так и аллеля дикого типа) конструировали на прямой цепи, тогда как обычный Scorpion-праймер конструировали на обратной цепи в последовательности интрона. Полученный ампликон имел длину 121 п.н. (с ARMS-праймерами) и длину 123 п.н. (с контрольным праймером). В этом случае второе основание кодона Gi43 находится на экзоне 36 п.н., на расстоянии только 1 п.н. от 3’-сайта сплайсинга. Опять конструировали Scorpion-праймер с использованием программ Oligo 5 и MFold (см. подробности в примере 1). Последовательность Scorpion-праймера Р. teres была:

Описание Scorpion-праймеров, синтез праймеров и условия ПЦР реального времени были такими же, как описанные в предыдущих примерах.
В оптимизации Scorpion-анализа большее разнообразие праймеров, чем в предыдущих примерах тестировали на специфичность, каждый с различными дефектными по спариванию основаниями на их 3’-конце для индукции дестабилизации (выделенными жирным шрифтом), как показано ниже.

Начальную оценку праймеров проводили на плазмидной матрице дикого типа и мутантной плазмидной матрице при 1×107 молекул на реакцию. ПЦР реального времени на приборе ABI Prism 7700 проводили в двух повторностях для каждой из плазмидных матриц с использованием как G-, так и С-специфических ARMS-праймеров и контрольного праймера (который амплифицирует как мутантный аллель, так и аллель дикого типа) с обычным обратным Scorpion-праймером.
Как видно из таблицы 12, различные конструкции ARMS-праймеров дали различные окна специфичности при тестировании против мутантной плазмидной матрицы и против плазмидной матрицы дикого типа.
При сравнении с другими тестированными праймерами PT-G-1 и РТ-С-5 дали самое широкое окно специфичности. Поэтому они были выбраны в качестве предпочтительной пары праймеров для этого анализа. Праймер РТ-С-5 имеет величину Ct 16 на правильной матрице и праймер PT-G-1 также имеет величину Ct 16 на правильной матрице. Праймер РТ-С-5 ошибочно праймирует на ошибочной матрице при цикле 34, давая окно специфичности 18 циклов, а праймер PT-G-1 ошибочно праймирует при цикле 32, давая окно специфичности 16 циклов, что равно приблизительно 10-кратному различию в чувствительности этих двух праймеров.
Оценка ARMS-праймеров
ARMS-праймеры PT-G-1 и РТ-С-5 тестировали в эксперименте введения плазмиды, где мутантную плазмиду “вводили” в фон плазмиды дикого типа при частотах 1:1, 1:10, 1:100, 1:1000, 1:10000 и 1:100000, все в 1 мг/мл БСА. Во всех случаях общая концентрация плазмиды каждой частоты введения в ПЦР-анализ была 1×107 молекул на реакцию. Условия и компоненты ПЦР-реакции реального времени были такими, как описанные ранее.
Полученные результаты суммированы в таблице 13.
Поскольку С-плазмида снижает фон плазмиды дикого типа, детектирование флуоресценции задерживается. С каждым 10-кратным разведением С-плазмиды имеется задержка 4 цикла в детектировании флуоресценции. Пороговая величина цикла РТ-С-5 на одной мутантной молекуле в фоне 10000 молекул дикого типа может быть ясно видна, но при 1:100000 не была различима, так как праймер РТ-С-5 ошибочно праймирует из 100% (G)-кассеты 100% дикого типа, маскируя, таким образом, более низкую частоту C:G. Это свидетельствует о том, что ARMS-переключение в этом случае будет с уверенностью допускать детектирование частоты ≤1:10000. Ряд проб Р. teres, собранных в г. Корк в Ирландии (Ir 5-8, Ir 9-13, Ir 14-17, Ir 18-21, Ir 30-34), тестировали с использованием этого заново оптимизированного Scorpion-ПЦР-анализа реального времени. Эти группы из 4 или 5 изолятов инокулировали на чашки с V8-агаром и инкубировали до достижения споруляции на полученной колонии.
Мицелии собирали из чашек с агаром в 10 мл стерильной ddH2O. Мицелиальную суспензию переносили в фальконовую пробирку на 15 мл и центрифугировали при 3200 об/мин в течение 10 минут. Воду удаляли и массу мицелия делили на две стерильные пробирки Эппендорфа. Затем мицелии осаждали центрифугированием при 13000 об/мин в течение 5 минут. Супернатант удаляли пипеткой и один из мицелиальных осадков использовали для экстракции ДНК, другой хранили при -80°С до использования. Экстракцию ДНК проводили с использованием протокола мининабора Qiagen DNeasy Plant Mini Kit для выделения ДНК из ткани растений, как описано в протоколе изготовителя. ДНК разводили 10-кратно и 100-кратно для применения в этом анализе и аликвоты 5 мкл ДНК использовали в каждом ПЦР-анализе. Все изоляты тестировали с каждой парой праймеров (РТ-С-5, PT-G-1 и контрольным праймером; каждую с обычным обратным праймером, содержащим систему детектирования Scorpion) в трех повторностях и в двух разведениях.
Результаты скрининга изолятов с использованием разведения 1:10 геномной ДНК в качестве матрицы показаны таблице 14.
Результаты ПЦР реального времени Р. teres
В большинстве случаев праймер РТ-С-5 обнаруживает пороговую величину цикла (Ct) при цикле 34 или позднее, при котором как показано, имеет место ошибочное праймирование на плазмидной матрице. В некоторых случаях праймер РТ-С-5 вообще не обнаруживает ошибочного праймирования. Эти результаты показывают, что не было доказательства присутствия мутации G143A в исследованных пробах.
ПРИМЕР 5
В примере 5 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Uncinula necator. U. necator является возбудителем настоящей мучнистой росы винограда.
Инфицированные листья и плоды винограда собирали из мест испытаний и коммерческих виноградников во Франции и Италии во время 1999 года и посылали на Jealott’s Hill Research Station (Zeneca Agrochemicals). По прибытии в лабораторию мицелий и споры переносили при помощи небольшой кисточки для краски на свежие поверхностно стерилизованные листья, отделенные от имеющих 6-7 листьев саженцев (var. Ohanez). U. necator из каждого места сбора обрабатывали в виде отдельной популяции.
После получения в лаборатории инокулированные листья помещали в помещение с постоянной температурой 21°С и инкубировали в течение 2-3 недель. Как только спорулирующее заболевание было обнаружено, инфицированные листья использовали для инокуляции имеющих 3-4 листа саженцев винограда, посаженных непосредственно в пластиковый рассадник для растений. Находящуюся под давлением воздушную струю использовали для сдувания конидий с листьев-источников на подходящие листья-мишени до 40 саженцев винограда на один изолят. Крышку опять помещали на рассадник, который затем инкубировали в помещении для выращивания с регулируемой окружающей средой в течение 2 недель. Конидии, образовавшиеся на этих растениях, собирали и замораживали при -80°С до использования.
Тотальную РНК экстрагировали из 100 мг измельченных спор U. necator, происходящих из одного изолята, с использованием набора PNeasy от Qiagen (в соответствии с рекомендациями изготовителя). Синтез кДНК первой цепи проводили с 1 мкг тотальной РНК с использованием ОТ-ПЦР с набором Advantage RT-PCR Clontech (в соответствии с рекомендациями изготовителя). 5 мкл кДНК использовали в качестве матрицы для ПЦР-амплификации с использованием ERY 11F (5’ ATGAACAATTGGTACAGTAAT 3’) (SEQ ID NO: 163) и ERY 4R (5’ AAATCTGTTAAAGGCATAGCC 3’) (SEQ ID NO: 164), которые представляют аминокислоты 114-309 грибного цитохрома b согласно системе нумерации S. cerevisiae. ДНК-фрагмент ожидаемого размера (~500 п.н.) амплифицировали и ПЦР-продукт клонировали в векторе ТА pCR2.1 (Invitrogen) в соответствии с рекомендациями изготовителя. Wizard minipreps проводили для идентификации клонов с подходящими вставками и 5 клонов предоставляли для секвенирования с использованием прямого и обратного праймеров М13. При анализе с использованием релевантного программного обеспечения для биоинформатики (Seqman and Macaw) было обнаружено, что была идентифицирована новая последовательность гена цитохрома b, которая является близкородственной другим последовательностям цитохрома b аскомицетов. Отрезок из 61 нуклеотида кДНК-последовательности, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3. Специфические праймеры U. necator были сконструированы на основе этой новой последовательности для применения непосредственно на материале спор. Предполагали присутствие интронных последовательностей в представляющем интерес районе (500 п.н. около SNP G143), так как прежняя ПЦР-амплификация на биологическом материале не была успешной. Добавку Taq Extender™ PCR Additive и ПЦР-усилитель Perfect Match® PCR Enhancer (оба от Stratagene) использовали (в соответствии с инструкциями изготовителя) в амплификации больших ПЦР-фрагментов с применением множества различных комбинаций праймеров. После начальной инкубации при 94°С в течение 3 минут проводили 30 циклов 94°С в течение 45 секунд, 52°С в течение 45 секунд и 72°С в течение 3 минут на приборе Hybaid Omn-E. Включали также конечную инкубацию при 72°С в течение 10 минут. Комбинации праймеров 2F (5’ GTT ТТА ССС ТАС GGG CAG ATG 34 (SEQ ID NO: 165) - 5R (5’ААА GAA TCT GTT TAA GGT TGC 3’) (SEQ ID NO: 166), 2F6R (5’ AAA ССА ССТ САА AGA ААС ТСС 3’) (SEQ ID NО: 167) и 4F (5’ CAT GAA TAG GAC AAG ATA TCG 3’) (SEQ ID NO: 168) успешно амплифицировали ПЦР-продукты в диапазоне длины 1,6 т.п.н. - 3 т.п.н. Эти ПЦР-продукты клонировали в векторе ТА pCR2.1 (Invitrogen) и последующие клоны использовали для секвенирования, как описано ранее. Стратегию праймерной “прогулки” использовали для одного клона, соответствующего каждому из различных ПЦР-продуктов, и анализ последовательности этих трех клонов показал присутствие двух интронов (с длиной 1,6 т.п.н. и 1,1 т.п.н.) в этих ПЦР-фрагментах. Второе основание кодона G143 расположено на расстоянии 10 п.н. выше по ходу транскрипции от сайта сплайсинга интрона. Отрезок из 41 нуклеотида, кодирующий 10 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 6
В примере 6 авторы описывают характеристику частичной последовательности гена цитохрома b Sphaerotheca fuliginea и полиморфизм единственного основания, который вызывает устойчивость к аналогам стробилурина и соединениям в той же самой группе перекрестной устойчивости. S. fuliginea является возбудителем настоящей мучнистой росы тыквы.
Инфицированные S. fuliginea листья огурца и дыни собирали в поле и конидии в сухом виде инокулировали в лаборатории на свежий материал листьев (огурца и дыни) при помощи небольшой кисточки для краски. Моноконидиальные изоляты субкультивировали и тестировали in planta с использованием 24-часового превентивного дискриминирующего дозового анализа (до доз 100 м.д., которые, как это известно, дают 100% уничтожение штаммов дикого типа). Конидии из предполагаемых устойчивых изолятов удаляли аспирацией при помощи вакуумного насоса в подходящий контейнер. Часть этой пробы анализировали с использованием ПЦР-анализа, а остаток повторно тестировали для подтверждения фенотипической устойчивости.
Стратегию ОТ-ПЦР использовали (как описано в примере 3) на ~100 мг конидиальной чувствительной к аналогу стробилурина пробы. Полученную кДНК использовали в качестве матрицы в реакции ПЦР-амплификации с использованием праймеров Ery2F (5’ TCACCTAGAACATTAACATGA 3’) (SEQ ID NO: 169) и 4R (5’ AAATCTGTTAAAGGCATAGCC 3’) (SEQ ID NO: 170), которые представляют аминокислоты 108-309 грибного цитохрома b на основе системы нумерации S. cerevisiae. Другие компоненты и условия ПЦР были такими, как описанные ранее. ПЦР-продукт ожидаемого размера (~600 п.н.) был обнаружен во время анализа гель-электрофорезом этих ПЦР-продуктов и этот продукт затем клонировали в векторе ТА pCR2.1 (Invitrogen) и 5 клонов со вставками правильного размера (~600 п.н.) секвенировали с использованием прямого и обратного праймеров М13, как описано ранее. При анализе с использованием релевантного программного обеспечения для биоинформатики (Seqman and Macaw) было обнаружено, что была идентифицирована новая последовательность гена цитохрома b, которая является близкородственной другим последовательностям цитохрома b аскомицетов. Затем были сконструированы специфические ПЦР-праймеры Sphaerotheca fuliginea для амплификации района G143 из геномной ДНК. Пары праймеров SF1 (5’ TTCCCTTCGGTCAAATGTCGC 3’) (SEQ ID NO: 171) - SF8 (5’ AAACCCCCTCAGAGAAACTCC 3’) (SEQ ID NО: 172) и SF1 - SF10 (5’ GACCCCGCGCTATCATGTAAG 3’) (SEQ ID NO: 173) использовали в ПЦР-амплификациях с использованием проб спор, ресуспендированных в ddH2O, в качестве матрицы. ПЦР-компоненты и условия ПЦР были такими, как описанные в предыдущих примерах. ПЦР-продукты анализировали гель-электрофорезом и продукт ~2 т.п.н. был обнаружен с парой праймеров SF1, 8 и полоса ~2,1 т.п.н. была обнаружена с парой праймеров SF1, 10. Оба ПЦР-продукта клонировали в клонирующем векторе ТА pCR2.1 (Invitrogen) и 5 клонов секвенировали, как описано ранее. Два клона из обоих событий клонирования полностью секвенировали с использованием стратегии праймерной “прогулки”. При анализе данных секвенирования с использованием релевантного программного обеспечения для биоинформатики было обнаружено, что интрон 1917 п.н. присутствовал на 9 п.н. ниже по ходу транскрипции от второго основания кодона G143. Последовательность кДНК и геномная последовательность (с учетом организации интрон, экзон) отрезка из 37 нуклеотидов, кодирующая 6 оснований выше по ходу транскрипции и 30 оснований ниже по ходу транскрипции от кодона G143 (в соответствии с системой нумерации S. cerevisiae), может быть найдена в таблице 3.
Частичную последовательность гена цитохрома b амплифицировали также из конидиальных устойчивых к аналогу стробилурина проб. Конидиальные пробы (<50 мг) ресуспендировали в 200 мкл ddH2O и разводили 1:10, 1:100 в ddH2O. 10 мкл каждого разведения использовали в качестве матрицы для ПЦР-амплификации с использованием SF1, SF8 специфических праймеров S. fuliginea. Добавку Taq Extender™ PCR Additive и ПЦР-усилитель Perfect Match® PCR Enhancer (оба от Stratagene) использовали (в соответствии с инструкциями изготовителя) в амплификации больших ПЦР-фрагментов. После начальной инкубации при 94°С в течение 3 минут проводили 30 циклов (94°С в течение 45 секунд, 50°С в течение 45 секунд и 72°С в течение 5 минут) на приборе Hybaid Omn-E. Включали также конечную инкубацию при 72°С в течение 10 минут. ПЦР-продукты анализировали при помощи гель-электрофореза и обнаружили полосу ожидаемого размера (~2 т.п.н.). Ее клонировали в векторе ТА pCR2.1 (Invitrogen) и 10 клонов, которые имели вставки ожидаемого размера, секвенировали, как описано в предыдущих примерах. Анализ данных последовательности с использованием подходящего программного обеспечения для биоинформатики (Seqman, Editseq and Macaw) выявил точковую мутацию G→C в последовательности цитохрома b в сравнении с последовательностью дикого типа во всех 10 случаях. Эта точковая мутация приводит к единственной замене глицина на аланин в положении 143 (в соответствии с с системой кодирования аминокислот S. cerevisiae).
ПРИМЕР 7
В примере 7 авторы описывают характеристику частичной последовательности гена цитохрома b Mycosphaerella filjiensis var. difformis. M. filjiensis является возбудителем черной "сигатоки" (церкоспороза) банана.
Инфицированные листья банана собирали в условиях открытого воздуха и аскоспоры инокулировали из листьев непосредственно на искусственную среду в чашку Петри. Моноаскоспоровые изоляты поддерживали на искусственной среде и готовили для ПЦР-анализа с использованием встряхиваемой культуры в бульонной среде. Мицелии собирали фильтрованием через Мираклотс и растирали до мелкого порошка с использованием промытых кислотой и автоклавированных стерильных пестика и ступки. 100 мг растертого мицелия использовали для экстракции геномной ДНК и синтеза кДНК первой цепи (как описано в предыдущих примерах). Геномную ДНК разводили 1:10, 1:100 и 1:1000 в ddH2O перед использованием в качестве ПЦР-матрицы. 5 мкл кДНК и 10 мкл каждого разведения геномной ДНК использовали в качестве матрицы для ПЦР-амплификации с использованием пары вырожденных праймеров, как описано в примере 3. Условия и компоненты ПЦР были такими, как описанные в предыдущих примерах. При анализе гель-электрофорезом ПЦР-продуктов была обнаружена полоса ожидаемого размера (~500 п.н.) при использовании кДНК в качестве матрицы и большая полоса (~1,6 т.п.н.) была обнаружена при применении в качестве матрицы геномной ДНК. Эти два ПЦР-продукта клонировали в векторе ТА pCR2.1 (Invitrogen) и пять полученных клонов со вставками правильного размера (~500 п.н. и 1,6 т.п.н.) секвенировали с использованием прямого и обратного праймеров М13. При анализе данных последовательности с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что больший ПЦР-продукт содержит интрон размером 1064 п.н., расположенный на расстоянии 78 п.н. ниже по ходу транскрипции от второго основания кодона G143. Кроме присутствия интрона в большем ПЦР-продукте эти ДНК-последовательности были идентичными. Эта последовательность гена цитохрома b была идентифицирована как новая и она была близкородственной другим последовательностям цитохрома b аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 8
В примере 8 авторы описывают характеристику частичной последовательности гена цитохрома b Pseudoperonospora cubensis. P. cubensis является возбудителем ложной мучнистой росы тыквы.
Спорангии смывали с инфицированного материала листьев в деионизованную воду. Полученную суспензию спорангиев инокулировали при помощи пистолета для опрыскивания на свежий материал листьев в концентрации 10000 спор на мл. После развития болезни пробу суспензии спор, собранную в ddH2O, откручивали в центрифуге с получением осадка и хранили при -80°С до использования. Затем спорангии Р. cubensis ресуспендировали в 200 мкл ddH2O и разводили 1:10 и 1:100 в ddH2O. 10 мкл каждого разведения спорангиев использовали в качестве матрицы для ПЦР-амплификации с использованием праймеров PLAS17F и PLAS15R (подробности см. в примере 1). Условия и компоненты ПЦР были такими, как описанные в предыдущих примерах. При анализе гель-электрофорезом ПЦР-продуктов была обнаружена полоса ожидаемого размера (~500 п.н.). Затем этот ПЦР-продукт клонировали в векторе ТА pCR2.1 и пять полученных клонов со вставками правильного размера (~500 п.н.) секвенировали с использованием прямого и обратного праймеров М13. При анализе данных секвенирования с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что была идентифицирована новая последовательность гена цитохрома b, которая является близкородственной другим последовательностям ци-тохрома b оомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 9
В примере 9 авторы описывают характеристику частичной последовательности гена цитохрома b Mycosphaerella graminicola. M. graminicola является возбудителем пятнистости листьев пшеницы.
Инфицированные листья пшеницы собирали в поле и инкубировали во влажных условиях для ускорения спорообразования. Усики удаляли из листьев и распределяли на чашки с Czapek Dox VS-агаром (см. таблицу 9) и инкубировали в регулируемой среде при 19°С в течение 6 дней. Изоляты отдельных колоний дополнительно субкультивировали и накапливали с использованием культуры во встряхиваемой колбе в подходящей среде. Грибной материал удаляли и хранили при -80°С до использования. Получение геномной ДНК проводили, как описано в примере 1, на 200 мг растертого мицелия. Выход и качество геномной ДНК проверяли гель-электрофорезом и разводили 1:10 и 1:100 в ddH2O. 10 мкл каждого разведения спорангиев использовали в качестве матрицы для ПЦР-амплификации с использованием праймеров Cyt3F и Cyt9R (подробности см. в примере 2). Компоненты и условия ПЦР были такими, как описанные в примере 2. ПЦР-продукты анализировали гель-электрофорезом, и была обнаружена полоса ожидаемого размера (~500 п.н.). Этот ПЦР-продукт клонировали в векторе ТА pCR2.1 и пять полученных клонов со вставками правильного размера секвенировали, как описано ранее. При анализе данных секвенирования с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что этот ПЦР-фрагмент кодирует часть новой последовательности гена цитохрома b, которая обнаруживала близкую гомологию с последовательностями цитохрома b других аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 10
В примере 10 авторы описывают характеристику частичной последовательности гена цитохрома b Colletotrichum graminicola. С. graminicola является возбудителем антракноза зерновых культур и злаковых трав.
Инфицированный материал листьев (дерн или “мох”) собирали в поле и грибной материал удаляли и субкультивировали на искусственных средах. Грибной материал накапливали с использованием культуры во встряхиваемой колбе, собирали и хранили при -80°С до использования. Получение геномной ДНК проводили, как описано в примере 1, на 200 мг растертого мицелия. Выход и качество геномной ДНК оценивали гель-электрофорезом и готовили развеления 1:10 и 1:100 в ddH2O. 10 мкл каждого разведения использовали в качестве матрицы для ПЦР-амплификации с использованием праймеров deg4F и deg3R (подробности см. в примере 3). Компоненты и условия ПЦР были такими, как описанные в примере 3. ПЦР-продукты анализировали гель-электрофорезом и была обнаружена полоса ожидаемого размера (~500 п.н.). Этот ПЦР-продукт клонировали в векторе ТА pCR2.1 и пять полученных клонов со вставками правильного размера секвенировали, как описано ранее. При анализе данных секвенирования с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что была идентифицирована новая последовательность гена цитохрома b, которая была близкородственной другим последовательностям цитохрома b аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 11
В примере 11 авторы описывают характеристику частичной последовательности гена цитохрома b Colletotrichum. gloeosporioides. С. gloeosporioides является возбудителем антракноза плодов (например, перца, авокадо и манго).
Инфицированный растительный материал (манго или перца стручкового) собирали в условиях открытого воздуха и грибной материал удаляли и субкультивировали на искусственных средах. Мицелии накапливали в культуре во встряхиваемой колбе и собирали фильтрованием через Мираклотс. Мицелии хранили при -80°С до использования. Мицелии растирали с использованием промытых кислотой стерильных пестика и ступки и 100 мг использовали для получения геномной ДНК и синтеза кДНК первой цепи (процедуры описаны в предыдущих примерах). 10 мкл каждого разведения геномной ДНК и 5 мкл неразведенной кДНК использовали в качестве матрицы для ПЦР-амплификации с использованием праймеров deg4F и deg3R (детали праймеров в примере 3). Компоненты и условия ПЦР были такими, как описанные в примерах. ПЦР-продукты анализировали гель-электрофорезом и были обнаружены полосы ожидаемого размера (~500 п.н.). ПЦР продукт был одного и того же размера при применении геномной ДНК или кДНК в качестве матрицы, что указывает на отсутствие интронов в амплифицированном районе ДНК. ПЦР-продукты клонировали в векторе ТА pCR2.1 и пять полученных клонов со вставками правильного размера для каждого события клонирования секвенировали, как описано в предыдущих.примерах. При анализе данных секвенирования с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что ПЦР-продукты были идентичными во всех случаях и они кодируют новую последовательность гена цитохрома b с близкой гомологией относительно других последовательностей аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 12
В примере 12 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Oidium lycopersicum. О. lycopersicum является возбудителем настоящей мучнистой росы томата.
Пораженные болезнью листья томата собирали в поле и конидии субкультивировали на свежем материале листьев инокуляцией в сухом виде в башенном отстойнике. Конидии, растущие в результате полученной инфекции, удаляли аспирацией при помощи вакуумного насоса в стерильную пробирку Эппендорфа и хранили при -80°С до использования. Синтез первой цепи кДНК проводили на РНК, выделенной из 100 мг спор, как описано ранее, и 5 мкл этой кДНК использовали для ПЦР-амплификации с использованием праймеров Еry2-4 (подробности в примере 6). Компоненты и условия ПЦР были такими, как описанные ранее. При анализе ПЦР-продуктов гель-электрофорезом был обнаружен ПЦР-продукт ожидаемого размера (~500 п.н.). Этот ПЦР-продукт клонировали и пять клонов, содержащих вставку правильного размера (~500 п.н.), секвенировали, как описано ранее. При анализе с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что этот ПЦР-фрагмент кодирует новую последовательность гена цитохрома b, которая является близкородственной другим последовательностям цитохрома b аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 13
В примере 13 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Leveillula taurica. L. taurica является возбудителем настоящей мучнистой росы томата.
Пораженные болезнью листья перца собирали в поле и конидии инокулировали в сухом виде на свежий материал листьев на целых растениях. Конидии из полученных инфекций удаляли аспирацией при помощи вакуумного насоса в подходящий контейнер и анализировали при помощи ПЦР. Пораженные болезнью листья перца и томата собирали в поле и в оранжерее при Jealott’s Hill и инфицированный материал листьев использовали непосредственно в ПЦР-анализе.
Синтез первой цепи кДНК проводили на РНК, выделенной из 100 мг спор или пораженного болезнью растительного материала, как описано ранее. При использовании пораженного болезнью растительного материала в качестве исходного материала грибные повреждения обогащали удалением не пораженной болезнью растительной ткани из препарата. Аликвоты кДНК 5 мкл использовали для ПЦР-амплификации с использованием праймеров Eryll-12 (подробности в примере 2). Компоненты и условия ПЦР были такими, как описанные ранее. При анализе ПЦР-продуктов гель-электрофорезом был обнаружен ПЦР-продукт ожидаемого размера (~500 п.н.). Этот ПЦР-продукт клонировали и пять клонов, содержащих вставку правильного размера (~500 п.н.), секвениро-вали, как описано ранее. При анализе с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что этот ПЦР-продукт кодирует новую последовательность гена цитохрома b, которая является близкородственной другим последовательностям цитохрома b аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 14
В примере 14 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Alternaria solani. A. solani является возбудителем ранней гнили у томата и картофеля.
Пораженный болезнью материал листьев (томата или картофеля) собирали в поле и грибной материал удаляли и субкультивировали на искусственной среде. Грибные пробы накапливали в культуре во встряхиваемой колбе. Изоляты коллекции культур хранили в жидком азоте и периодически субкультивировали на искусственной среде или субкультивировали через живой материал хозяина перед повторным хранением при -80°С. Мицелии, выращенные во встряхиваемой колбе, растирали с использованием промытых кислотой стерильных пестика и ступки и 100 мг использовали для получения геномной ДНК и синтеза кДНК первой цепи (процедуры такие же, как описанные в предыдущих примерах). 10 мкл каждого разведения геномной ДНК и 5 мкл неразведенной кДНК использовали в качестве матрицы для ПЦР-амплификации с использованием праймеров deg4F и deg3R (подробности в примере 3). Компоненты и условия ПЦР были такими, как описанные в примере 3. ПЦР-продукты анализировали гель-электрофорезом и были обнаружены полосы ожидаемого размера (~500 п.н.). ПЦР-продукт был одного и того же размера при применении геномной ДНК или кДНК в качестве матрицы, что указывает на отсутствие интронов в амплифицированном районе ДНК. ПЦР-продукты клонировали в векторе ТА pCR2.1 и пять полученных клонов со вставками правильного размера для каждого события клонирования секвенировали, как описано ранее. При анализе данных секвенирования с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что ПЦР-продукты были идентичными во всех случаях и они кодируют новую последовательность гена цитохрома b, которая обнаруживает близкую гомологию относительно других последовательностей аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
Последовательность, отличающуюся только на 12 п.н. от выделенной ранее последовательности A. solani, также амплифицировали при использовании геномной ДНК, выделенной непосредственно из инфицированных в полевых условиях листьев томатов, в качестве матрицы для ПЦР-амплификации с использованием праймеров Eryll-12 (см. выше в отношении подробностей). Хотя эта последовательность ДНК содержала 12 нуклеотидных различий, она не приводила к какому-либо различию на уровне аминокислот при трансляции этих двух последовательностей. Значимым различием между этими двумя последовательностями было присутствие интрона размером ~1,2 т.п.н., на расстоянии 62 п.н. по ходу транскрипции от второго основания в кодоне G143. Эти различия в организации интрон, экзон и использовании кодонов являются доказательством того, что такие вариации возможны в одном виде.
ПРИМЕР 15
В примере 15 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Cercospora arachidola. С. arachidola является возбудителем пятнистости листьев земляного ореха.
Инфицированный материал листьев (земляного ореха) собирали в поле и грибной материал удаляли и субкультивировали на искусственной среде. Культуры хранили в жидком азоте до использования. Материал извлекали из хранения на агар и полученные колонии накапливали в культуре во встряхиваемой колбе и хранили при -80°С до использования. Мицелии растирали с использованием промытых кислотой стерильных пестика и ступки и 100 мг использовали для получения геномной ДНК и синтеза кДНК первой цепи (процедуры такие же, как описанные в предыдущих примерах). 10 мкл каждого разведения геномной ДНК и 5 мкл неразведенной кДНК использовали в качестве матрицы для ПЦР-амплификации с использованием праймеров deg4F и deg3R (подробности в примере 3). Компоненты и условия ПЦР были такими, как описанные в примере 3. ПЦР-продукты анализировали гель-электрофорезом и были обнаружены полосы ожидаемого размера (~500 п.н.). Опять ПЦР продукт был одного и того же размера при применении геномной ДНК или кДНК в качестве матрицы, что указывает на отсутствие интронов в амплифицированном районе ДНК. ПЦР-продукты клонировали в векторе ТА pCR2.1 и пять полученных клонов со вставками правильного размера для каждого события клонирования секвенировали, как описано ранее. При анализе данных секвенирования с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что ПЦР-продукты были идентичными во всех случаях и они кодируют новую последовательность гена цитохрома b, которая обнаруживает близкую гомологию относительно других последовательностей гена цитохрома b. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 16
В примере 16 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Rhizoctonia solani. R. solani является возбудителем корневой и стеблевой гнили или полегания (черной ножки).
Инфицированный материал листьев (риса) собирали в поле и грибной материал удаляли и субкультивировали на искусственной среде. Культуры хранили в жидком азоте до использования. Материал извлекали из хранения на агар и полученные колонии накапливали в культуре во встряхиваемой колбе и хранили при -80°С до использования. Синтез кДНК первой цепи проводили на РНК, выделенной из 100 мг растертых мицелиев, как описано ранее, и 5 мкл использовали для ПЦР-амплификации с использованием вырожденных праймеров базидиомицетов 1F (5’ WYTRGTAYTAATGATGGCTATHGG 3’) (SEQ ID NO: 174) и 1R (5’ TCTTARWATWGCATAGAAWGG 3’) (SEQ ID NO: 175), которые соответствуют аминокислотам 121-283 грибного цитохрома b согласно системе нумерации S. cerevisiae. Компоненты и условия ПЦР были такими, как описанные ранее. При анализе ПЦР-продуктов гель-электрофорезом был обнаружен ПЦР-продукт ожидаемого размера (~500 п.н.). Этот ПЦР-продукт клонировали и пять клонов, содержащих вставку правильного размера (~500 п.н.), секвенировали, как описано ранее. При анализе данных секвенирования с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что этот ПЦР-фрагмент кодирует новую последовательность гена цитохрома b, которая обнаруживает близкую гомологию относительно других последовательностей базидиомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 17
В примере 17 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Pythium. aphanidermatum. P. aphanidermatum является возбудителем полегания.
Инфицированный материал листьев (дерн) собирали в поле и грибной материал удаляли и субкультивировали на искусственной среде. Культуры хранили в жидком азоте до использования. Материал извлекали из хранения на агар и полученные колонии накапливали в культуре во встряхиваемой колбе и хранили при -80°С до использования. Получение геномной ДНК проводили, как описано в примере 1, на 200 мг растертых мицелиев. Полученную геномную ДНК анализировали гель-электрофорезом и исходные препараты разводили 1:10 и 1:100 в ddH2O. 10 мкл каждого разведения использовали в качестве матрицы для ПЦР-амплификации с использованием праймеров PLAS17F и PLAS15R (подробности в примере 1). Компоненты и условия ПЦР были такими, как описанные в примере 1. ПЦР-продукты анализировали гель-электрофорезом и была обнаружена полоса правильного размера (~500 п.н.). Этот ПЦР-продукт клонировали в векторе ТА Invitrogen pCR2.1 и пять клонов, содержащих вставки правильного размера, секвенировали, как описано ранее. При анализе данных секвенирования с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что этот ПЦР-фрагмент кодирует новую последовательность гена цитохрома b, которая обнаруживает близкую гомологию с другими последовательностями оомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона 6143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
ПРИМЕР 18
В примере 18 авторы изобретения описывают характеристику частичной последовательности гена цитохрома b Mycosphaerella musicola. M. musicola является возбудителем желтой “сигатоки” банана.
Инфицированные листья банана собирали в условиях открытого воздуха и аскоспоры инокулировали из листьев непосредственно на искусственную среду в чашку Петри. Моноаскоспоровые изоляты поддерживали на искусственной среде и готовили для ПЦР-анализа с использованием культуры во встряхиваемой колбе в бульонной среде. Мицелии собирали фильтрованием через Мираклотс и растирали до мелкого порошка с использованием промытых кислотой стерильных пестика и ступки. 100 мг растертых мицелиев использовали для экстракции ДНК и синтеза кДНК первой цепи (как описано в примере 7). Геномную ДНК разводили 1:10, 1:100 и 1:1000 в ddH2O перед использованием в качестве ПЦР-матрицы. 5 мкл кДНК и 10 мкл каждого разведения геномной ДНК использовали в качестве матрицы для ПЦР-амплификации с применением пары вырожденных праймеров F4, R3 (как описано в примере 3). Условия и компоненты ПЦР были такими же, как описанные в предыдущих примерах. При анализе с использованием гель-электрофореза ПЦР-продуктов была обнаружена полоса ожидаемого размера (~500 п.н.) при использовании как геномной ДНК, так и кДНК в качестве матрицы. Эти два продукта клонировали в векторе ТА pCR2.1 и пять клонов, содержащих вставки правильного размера, секвенировали с использованием прямого и обратного праймеров М13. При анализе данных секвенирования с использованием подходящего программного обеспечения по биоинформатике было обнаружено, что эти ПЦР-фрагменты кодирует новую последовательность гена цитохрома b с близкой гомологией с другими последовательностями гена цитохрома b аскомицетов. Отрезок из 61 нуклеотида, кодирующий 30 оснований против хода транскрипции и 30 оснований по ходу транскрипции от второго основания кодона G143 (согласно системе нумерации аминокислот S. cerevisiae), можно найти в таблице 3.
































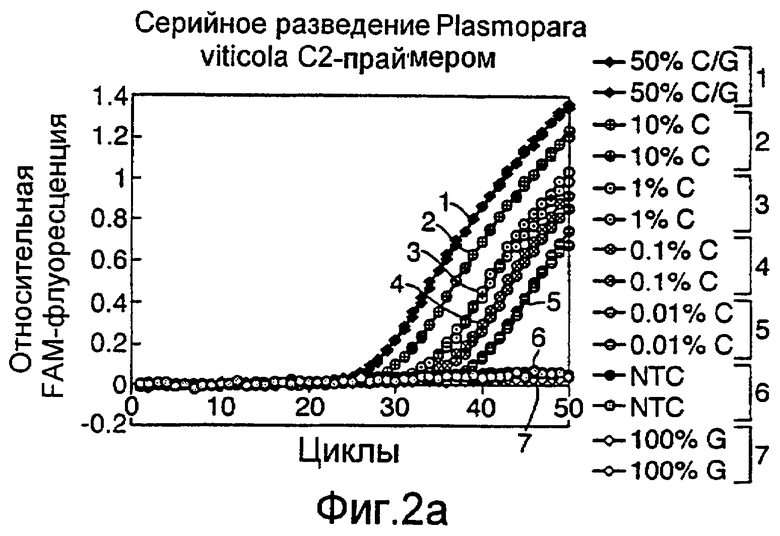
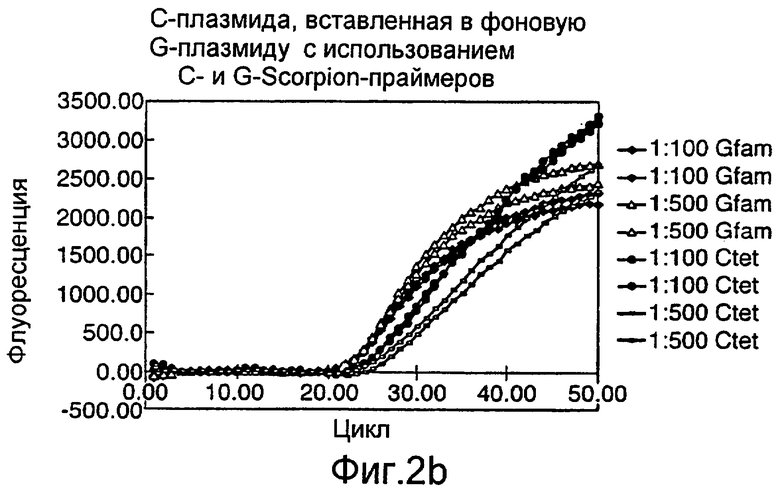
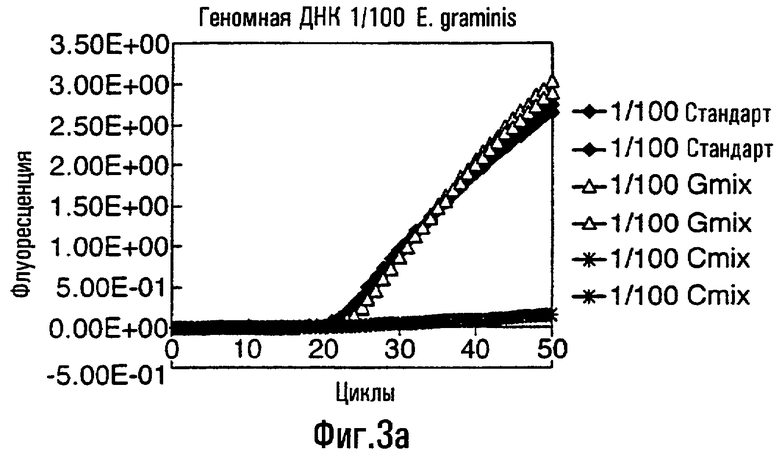
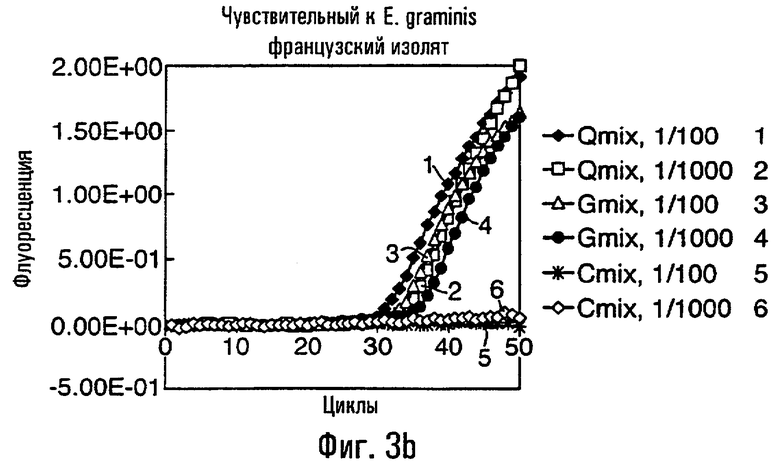
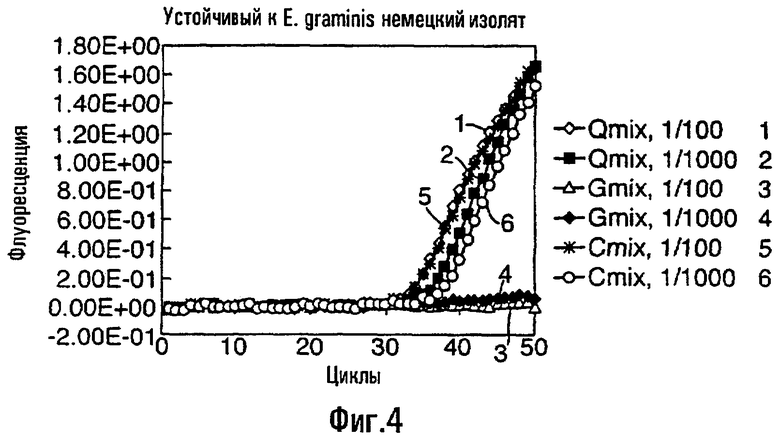
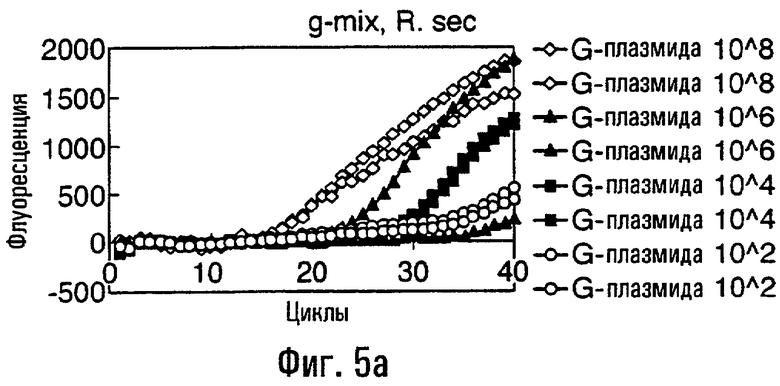
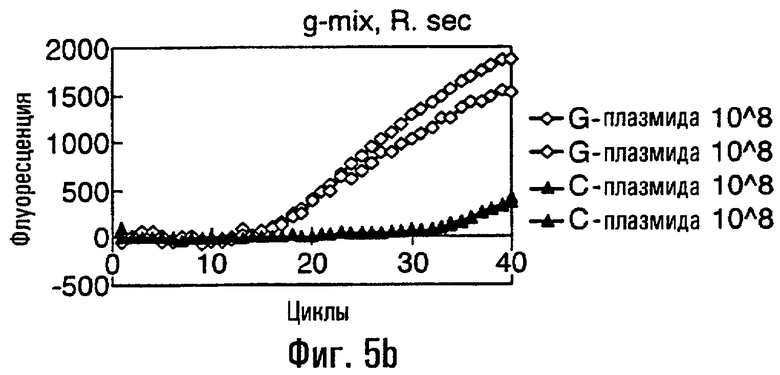
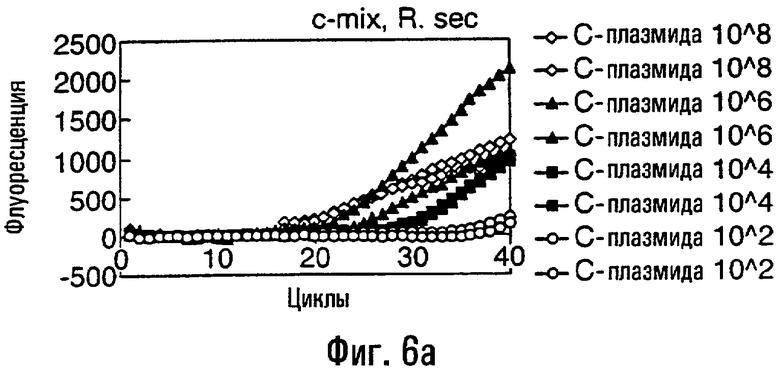
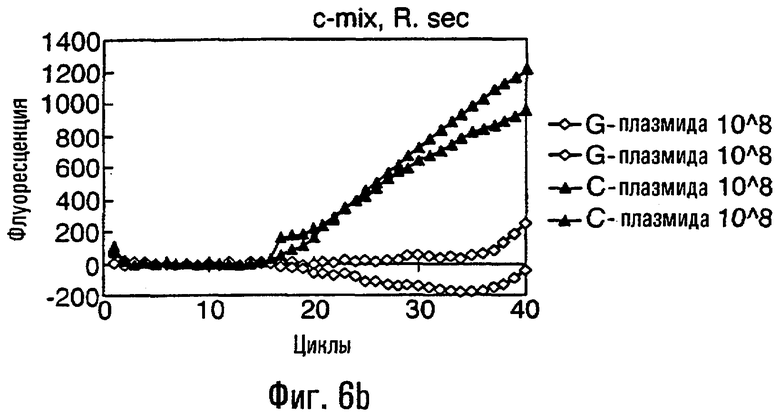
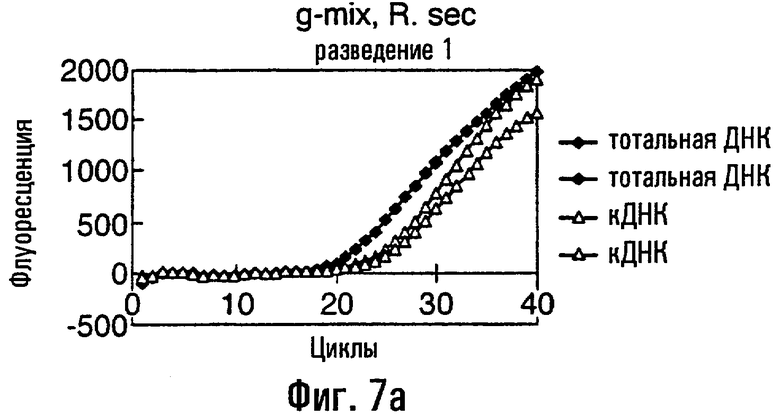
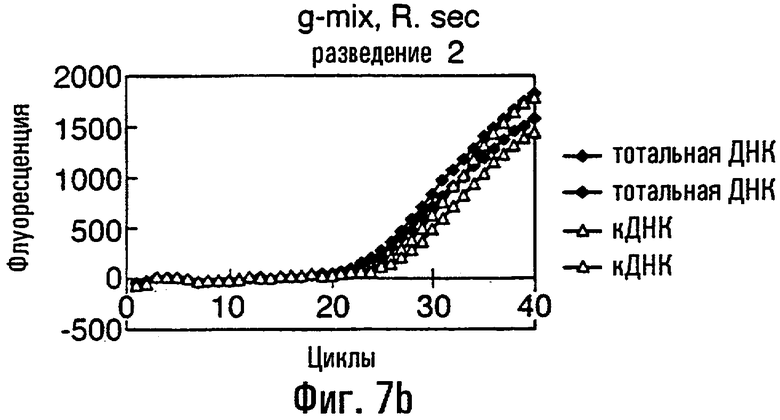
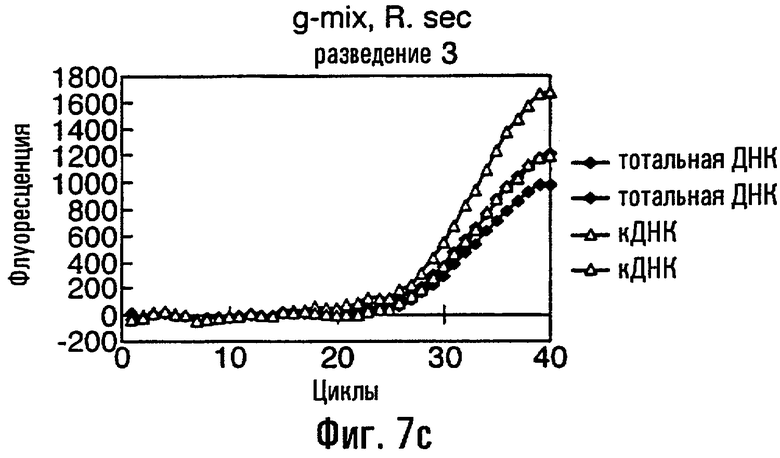
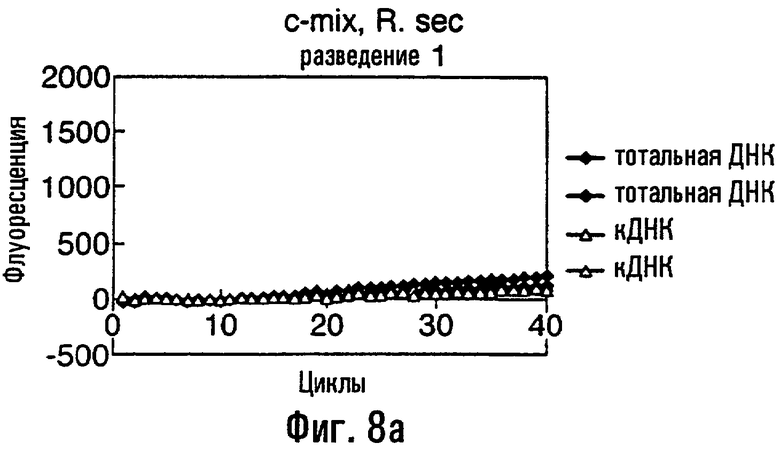
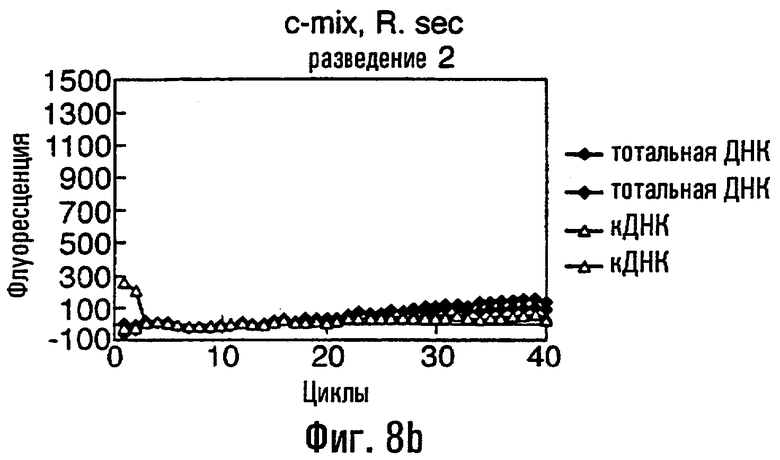
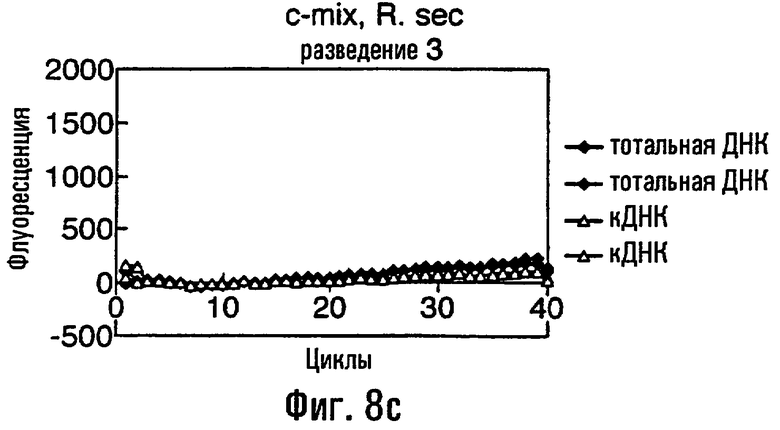
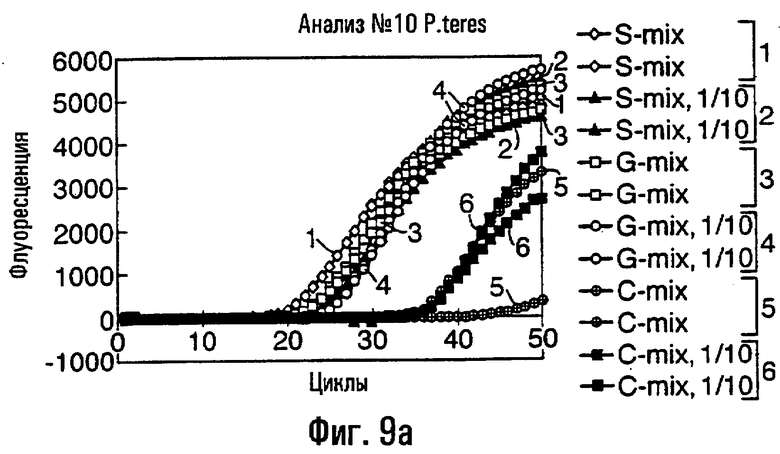
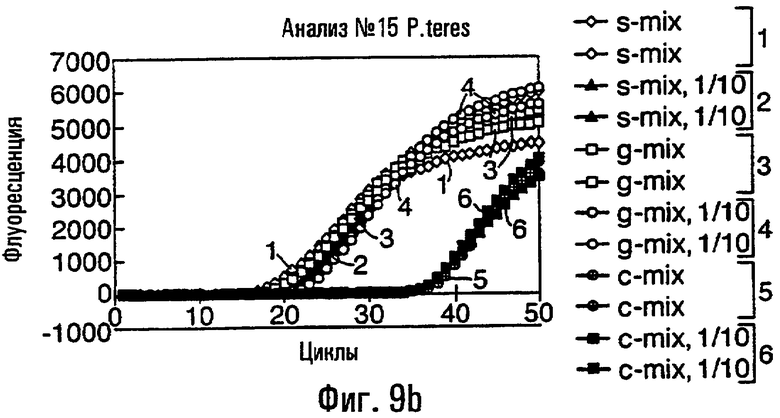
Изобретение относится к области биотехнологии, в частности к способам обнаружения низких частот встречаемости мутаций в гене цитохрома b. Выделяют ДНК из заданных грибов для конструирования олигонуклеотидного зонда или праймера. Проводят ПЦР-реакцию, в результате которой определяют связывание олигонуклеотидного зонда с ампликоном, генерированным в процессе данной реакции, или обнаруживают присутствие ампликона, который генерировался в результате ПЦР-реакции с использованием указанных праймеров. Изобретение позволяет быстро и точно определить мутации, придающие устойчивость гриба к фунгициду. 7 с. и 17 з.п. ф-лы, 9 ил.,14 табл.
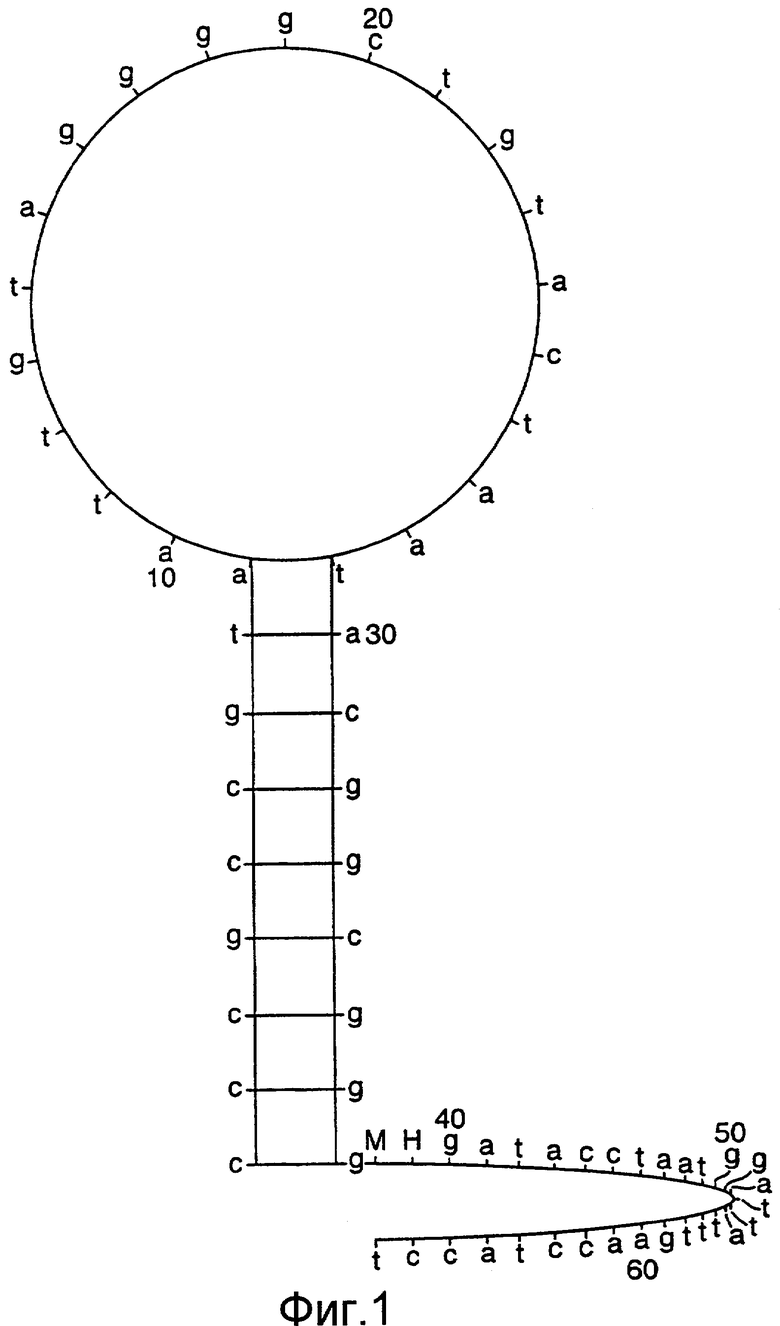
(i) генерирование ампликона и контактирование указанного ампликона с олигонуклеотидным зондом, который является специфичным для указанной точечной мутации, так что связывание олигонуклеотидного зонда с указанным ампликоном указывает на присутствие указанной мутации, или
(ii) обнаружение присутствия ампликона, где, когда один из указанных праймеров является специфичным для точечной мутации, присутствие указанного ампликона указывает на присутствие мутации, и где, когда один из указанных праймеров является специфичным для гена немутантного цитохрома b, присутствие указанного ампликона указывает на отсутствие мутации.
Приоритет по пунктам:
| ZHENG D ET AL: “Characterization of mitochondrial cytochrome b gene from Venturia inegualis ” CURRENT GENETICS, vol.32, November 1997, pages 361-66 | |||
| DI RAGO J-P AT AL: “Molecular basis for resistance to Myxothiazol, Mucidin (Strobilurin A), and Stigmatellin” THE JOURNAL OF BIOLOGICAL CHEMISTRY, vol.264, no.24, AUGUST 1989, pages 14543-548 | |||
| СПОСОБ ОПРЕДЕЛЕНИЯ ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕОТИДОВ | 1989 |
|
RU2099426C1 |

