ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет согласно предварительной заявке на патент США №62/062,351, поданной 10 октября 2014 г., описание которой включено в настоящий документ посредством ссылки.
ГОСУДАРСТВЕННАЯ ПОДДЕРЖКА
Изобретение, описанное в настоящем документе, выполнено, по меньшей мере частично, при государственной поддержке в рамках грантов Национального института здравоохранения США № U01AI082174 и R01AI080653. Соответственно, правительство США обладает определенными правами на данное изобретение.
ОБЛАСТЬ ТЕХНИКИ
В настоящем изобретении предложены новые праймеры и зонды на основе низкоселективных молекулярных маяков (sloppy molecular beacons, SMB) и молекулярных маяков (molecular beacons, MB) для амплификации и обнаружения сегментов различных генов Mycobacterium tuberculosis (М. tb) с целью выявления наличия ДНК М. tb и выявления устойчивости к лекарственным средствам против туберкулеза.
УРОВЕНЬ ТЕХНИКИ
Туберкулез (ТВ) был объявлен глобальной угрозой приблизительно двадцать лет назад (Глобальный отчет ВОЗ по туберкулезу-2013). Хотя частота новых случаев ТВ снижается во всем мире, достижение цели развития, сформулированной в "Декларации тысячелетия" - снижения заболеваемости на 50% к 2015 г. является маловероятным (Глобальный отчет ВОЗ по туберкулезу-2013). Увеличение частоты множественной лекарственной устойчивости (МЛУ) и широкой лекарственной устойчивости (ШЛУ) ТВ представляет собой серьезную проблему с точки зрения достижения указанных целей снижения заболеваемости (Глобальный отчет ВОЗ по туберкулезу-2013). МЛУ ТВ определяют как ТВ, устойчивый к лечению с применением по меньшей мере рифампицина и изониазида, а ШЛУ ТВ определяют как МЛУ ТВ, дополнительно устойчивый к лечению с применением фторхинолоновых антибиотиков и лекарственных средств амикацин, канамицин и капреомицин, вводимых посредством инъекций. Пациентов с туберкулезом, устойчивым к лекарственным средствам, лучше всего выявлять как можно быстрее, чтобы можно было быстро инициировать надлежащий контроль и лечение инфекции. Boehme, СС, et. al,, 2011, Lancet 377: 1495-1505.
Полное определение картины лекарственной устойчивости Mycobacterium tuberculosis (М. tb) обычными фенотипическими способами может занять от нескольких недель до нескольких месяцев из-за очень медленного роста этой бактерии (Heifets, L., et al., J Clin Microbiol 38:1227-1230; Kim, S.J., 2005, Eur Respir J 25:564-569; и PT, K., and K. GP., 1985, Public health Mycobacteriology: A guide for level III laboratory. Center for Disease Control, U.S Department of Health and Human Services, Atltanta, Georgia). Молекулярный анализ создает перспективы более быстрого обнаружения лекарственной устойчивости. М. tb в естественном состоянии не содержит плазмид лекарственной устойчивости; таким образом, молекулярный анализ направлен по отношению к хромосомной ДНК. Генотипические анализы относительно просты в разработке, поскольку геном М. tb характеризуется очень высокой степенью консервативности последовательности. Практически все клинические изоляты М. tb, чувствительные к лекарственным средствам, содержат идентичные последовательности ДНК мишеней лекарственной устойчивости, за исключением нескольких легко идентифицируемых «естественных полиморфизмов». Из этого следует, что любое отклонение от последовательности дикого типа гена-мишени лекарственной устойчивости указывает на наличие лекарственной устойчивости к соответствующему лекарственному средству. Генотипические анализы более быстры и чувствительны, чем фенотипические анализы, поскольку ДНК-мишени можно амплифицировать с помощью ПЦР. Биологические угрозы можно минимизировать путем уничтожения инфекционных организмов на ранних стадиях.
Генетические мишени, обуславливающие большинство случаев устойчивости к лекарственным средствам при ТВ, хорошо известны в настоящее время. ПЦР в реальном времени остается самым чувствительным, быстрым и надежным способом обнаружения мутаций в бактериях. Практически все другие способы обнаружения мутаций, включая ПЦР-МС, микрочипы, миничипы и секвенирование нового поколения, требуют амплификации нуклеиновых кислот в качестве первого этапа процесса обнаружения. В отличие от них, ПЦР в реальном времени позволяет выполнять амплификацию, обнаружение и анализ образца в одной лунке. Не требуется открывать пробирки, не нужны сложные жидкостные системы. Тем не менее, до сих пор не разработана широкая методология анализа лекарственной устойчивости, достаточно простая и надежная для выполнения вне референтных лабораторий. Таким образом, существует потребность в новых праймерах и зондах для обнаружения М. tb и для обнаружения лекарственной устойчивости М. tb к наиболее распространенным лекарственным средствам первой и второй линии.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к праймерам, зондам и связанным с ними вариантам применения для обнаружения М. tb, а также устойчивости М. tb к лекарственным средствам (лекарственной устойчивости).
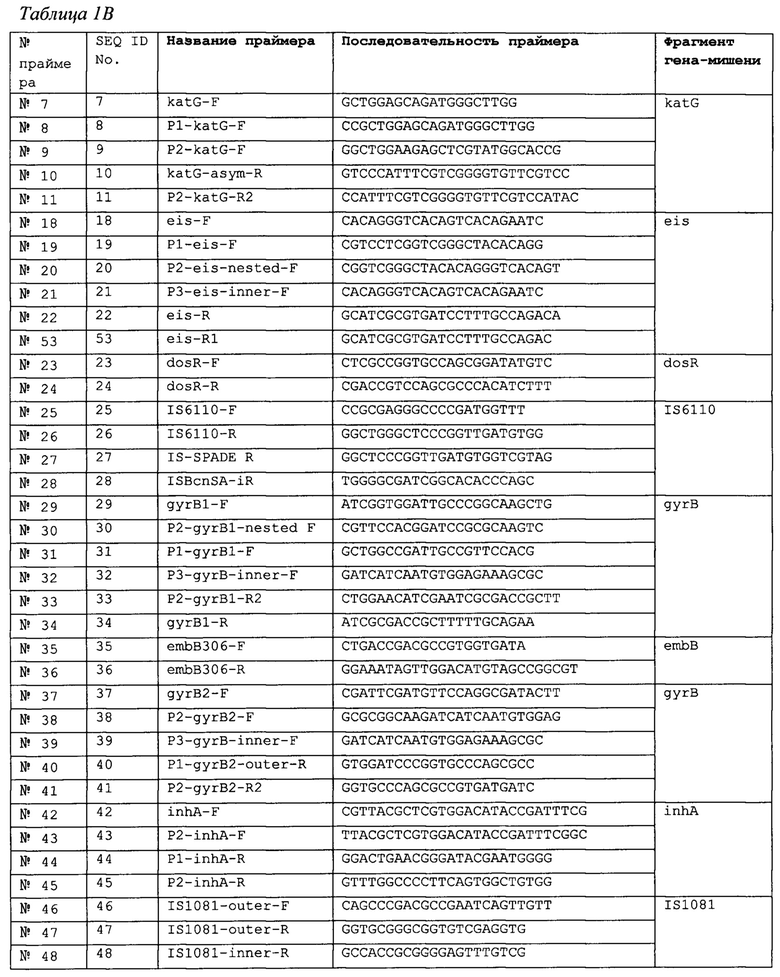
В одном аспекте настоящее изобретение относится к выделенному набору олигонуклеотидов или набору праймеров для амплификации части области М tuberculosis, выбранной из группы, состоящей из гена rpoB, гена gyrA, гена gyrB, промотора inhA, гена rrs, промотора eis, гена embB, гена katG, гена dosR, гена IS6110, гена IS1081. Набор включает пару из прямого и обратного праймеров, специфичных по отношению к указанному фрагменту, где каждый праймер обладает последовательностью, по существу идентичной последовательности олигонуклеотида, выбранного из олигонуклеотидов, перечисленных ниже в Таблицах 1А и 1В. Соответственно, каждый праймер обладает последовательностью, по существу комплементарной комплементарной цепи последовательности олигонуклеотида, выбранного из олигонуклеотидов, перечисленных в указанных Таблицах. В некоторых вариантах реализации последовательность праймера идентична последовательности олигонуклеотида, выбранного из олигонуклеотидов, перечисленных в Таблицах 1А и 1В.
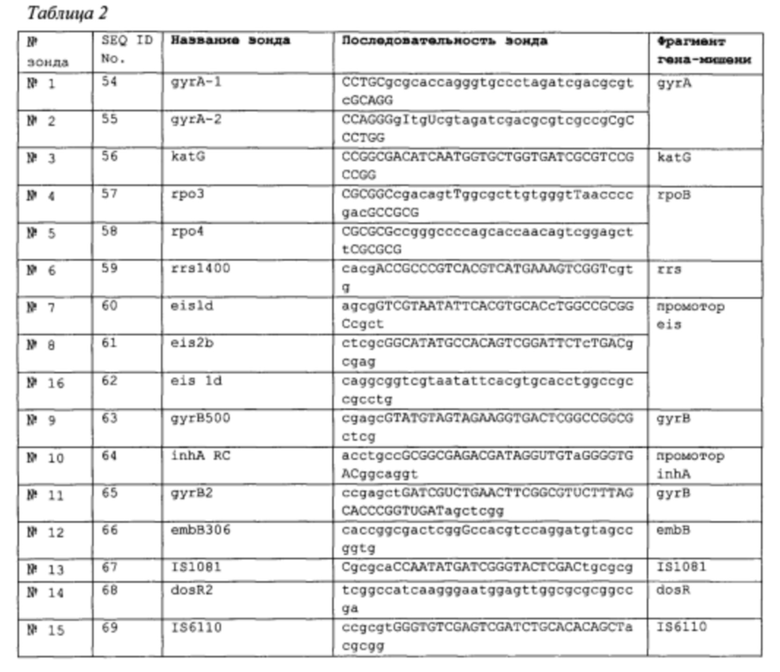
Во втором аспекте настоящее изобретение относится к выделенной нуклеиновой кислоте, обладающей последовательностью, по существу идентичной последовательности, выбранной из последовательностей, перечисленных в Таблице 2. В некоторых вариантах реализации нуклеиновая кислота содержит последовательность, выбранную из последовательностей, перечисленных в Таблице 2. Нуклеиновую кислоту можно метить, например, флуорофором и гасителем, присоединенными к ее двум концам, соответственно, или флуорофором, связанным с внутренним нуклеотидом в составе зонда. Примеры флуорофоров включают флуоресцеин, цианин 5 или TexasRed и TAMRA. Примеры гасителей включают BHQ1, BHQ2 и DABCYL.
В настоящем изобретении предложен набор, содержащий один или более из описанных выше наборов олигонуклеотидов и нуклеиновую кислоту. Указанный набор может дополнительно содержать ДНК-полимеразу, нуклеотиды для удлинения последовательности и буфер.
В третьем аспекте настоящего изобретения предложен способ обнаружения лекарственной устойчивости у М. tuberculosis. Указанный способ включает этапы амплификации первой нуклеотидной последовательности-мишени с использованием первой пары праймеров и получением первого ампликона, где (i) первая пара праймеров специфична по отношению к фрагменту области, выбранной из группы, состоящей из гена rpoB, гена gyrA, гена gyrB, промотора inhA, гена rrs, промотора eis, гена embB и гена katG, и (ii) каждый праймер обладает последовательностью, по существу идентичной последовательности олигонуклеотида, выбранного из олигонуклеотидов, перечисленных в Таблицах 1А и 1В, и обнаружения мутации в первом ампликоне. Наличие мутации свидетельствует о лекарственной устойчивости. В указанном способе этап обнаружения можно выполнять с использованием различных способов обнаружения нуклеиновых кислот, известных в данной области техники, включая, например, способы на основе секвенирования и методики на основе гибридизации нуклеотидных или пептидо-нуклеиновых зондов.
В одном варианте реализации этап обнаружения выполняют с помощью процесса, включающего (i) приведение в контакт первого ампликона с первым зондом, специфичным по отношению к мутации, в условиях, благоприятствующих гибридизации, с образованием гибрида зонд-мишень; (ii) проведение анализа температуры плавления (Tm) для определения тестового значения Tm для гибрида зонд-мишень; и (iii) сравнение тестового значения Tm с заранее определенным значением Tm. Тестовое значение Tm, отличающееся от заранее определенного значения Tm, указывает на наличие мутации. Например, сдвиг тестового значения Tm по меньшей мере на 3 (например, 3, 4 или 5) стандартных отклонения от эталонного значения Tm указывает на наличие мутации. И наоборот, сдвиг тестового значения Tm менее чем на 3 стандартных отклонения от эталонного значения Tm указывает на отсутствие мутации. В настоящем документе заранее определенное эталонное значение Tm может представлять собой среднее значение Tm дикого типа. В одном примере ситуация, при которой тестовое значение Tm меньше заранее определенного значения Tm, например, на по меньшей мере 3 стандартных отклонения, указывает на наличие мутации. В других случаях ситуация, при которой тестовое значение Tm не меньше заранее определенного значения Tm, например, на 3 стандартных отклонения, указывает на отсутствие мутации.
Указанный способ может дополнительно включать амплификацию второй нуклеотидной последовательности-мишени с использованием второй пары праймеров и получением второго ампликона, причем указанная вторая пара праймеров специфична по отношению к фрагменту второй области, выбранной из группы, состоящей из гена rpoB, гена gyrA, гена gyrB, промотора inhA, гена rrs, промотора eis, гена embB и гена katG. В некоторых вариантах реализации первая область представляет собой ген rrs или промотор eis. Например, первая область может представлять собой ген rrs, а вторая область может представлять собой промотор eis. Указанные две области можно амплифицировать независимо или в одной той же реакционной системе с помощью таких методик, как гнездовая ПЦР. В этом случае мутация может представлять собой мутацию A1401G или С1402Т в гене rrs. Мутация может находиться в пределах области промотора eis, опрашиваемой последовательностями праймеров eis.
Вышеописанный способ позволяет обнаружить устойчивость к лекарственному средству, выбранному из группы, состоящей из изониазида, рифампицина, амикацина, канамицина, капреомицина, этамбутола и лекарственных средств класса фторхинолонов. Пара праймеров может быть парой праймеров, выбранных из праймеров, перечисленных в Таблицах 1А и 1В. Зонд может обладать последовательностью, по существу или полностью идентичной последовательности, выбранной из последовательностей, перечисленных в Таблице 2.
В четвертом аспекте настоящего изобретения предложен способ обнаружения наличия М. tuberculosis в тестовом образце, например, полученном у субъекта. Указанный способ включает приведение в контакт тестового образца с первой парой праймеров в условиях, благоприятствующих реакции амплификации с получением первого ампликона, и обнаружения присутствия ампликона, что позволяет обнаружить наличие Mycobacterium tuberculosis в тестовом образце. Первая пара праймеров может представлять собой набор олигонуклеотидов для амплификации части области М. tuberculosis, выбранной из группы, состоящей из гена gyrB, промотора inhA, промотора eis, гена embB, гена katG, гена dosR, гена IS6110, гена IS1081. Каждый праймер первой пары праймеров обладает последовательностью, которая может быть по существу идентична последовательности олигонуклеотида, выбранного из олигонуклеотидов, перечисленных в Таблице 1В. Способ может дополнительно включать приведение в контакт тестового образца или ампликона, полученного с помощью первой пары праймеров, со второй парой праймеров в условиях, благоприятствующих реакции амплификации, с получением второго ампликона, и обнаружение наличия второго ампликона. В этом случае наличие как первого, так и второго ампликона указывает на присутствие Mycobacterium tuberculosis в тестовом образце.
В пятом аспекте настоящего изобретения предложен еще один способ обнаружения наличия М. tuberculosis в тестовом образце. Указанный способ включает приведение в контакт тестового образца с первым зондом-молекулярным маяком в условиях, благоприятствующих реакции гибридизации с получением гибрида зонд-мишень, и обнаружения присутствия гибрида зонд-мишень, что позволяет обнаружить наличие Mycobacterium tuberculosis в тестовом образце. В данном способе первый зонд-молекулярный маяк обладает последовательностью, по существу идентичной последовательности, выбранной из последовательностей, перечисленных в Таблице 2. В примере первый зонд - молекулярный маяк выбирают из группы, состоящей из зонда IS1081, dosR2 и IS6110 (SEQ ID No. 67-69).
Подробности по меньшей мере одного варианта реализации настоящего изобретения изложены в описании, приведенном ниже. Другие свойства, цели и преимущества настоящего изобретения будут понятны из описания и рисунков, а также из формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 представляет собой диаграмму, на которой показано обнаружение устойчивости к АМК и/или KAN в 603 клинических образцах ДНК с использованием трехточечного профиля Tm, полученного с помощью SMB-зонда. Каждый из трех SMB, использованных при анализе, протестировали по отношению ко всем образцам ДНК М. tb в многолокусной ПЦР. Результаты для каждого образца показаны в виде трехточечного графика Tm на оси X, причем значение Tm каждого SMB указано на оси Y. Изоляты сортировали слева направо от фенотипически чувствительных к устойчивым. Для каждого устойчивого изолята видны четкие сдвиги Tm для по меньшей мере одного из трех зондов.
Фигуры 2А, 2В и 2С представляют собой диаграммы, на которых показаны первые производные профилей пиков плавления трех SMB-зондов. Профили пиков плавления образцов ДНК дикого типа, мутантных и смешанных образцов ДНК показаны для SMB-зонда rrs-1400 (2А), SMB-зонда eis1 (2В) и SMB-зонда eis2 (2С). Каждая кривая плавления представляет отдельный штамм.
Фигура 3 представляет собой диаграмму, на которой показаны значения MIC штаммов, мутантных по rrs и eis, и штаммов дикого типа. Показаны средние значения MIC АМК и KAN для мутантов по rrs и eis и штаммов дикого типа. Показатели ошибки означают ± одно стандартное отклонение значений MIC. eis-P означает промотор гена eis.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение по меньшей мере частично основано на неожиданном открытии новых праймеров, SMB-зондов и МВ-зондов для амплификации сегментов одиннадцати различных генов М. tb с целью определения наличия ДНК М. tb и устойчивости к противотуберкулезным лекарственным средствам, например, изониазиду, рифампицину, амикацину, канамицину, капреомицину, этамбутолу и лекарственным средствам класса фторхинолонов.
Праймеры и зонды
Праймеры, описанные в настоящем документе, используемые для амплификации генов rpoB, gyrA, gyrB, области промотора inhA, гена rrs, области промотора eis, генов embB и katG позволяют выполнять чувствительную амплификацию горячих точек мутагенеза, вызывающих лекарственную устойчивость М. tb. Соответствующие SMB-зонды специфически связываются и выявляют указанные мутации, приводящие к лекарственной устойчивости к наиболее распространенным лекарственным средствам первой и второй линии. Эти праймеры можно использовать с очень высокой эффективностью как при симметричном, так и асимметричном ПЦР-анализе. Авторы изобретения использовали последовательности праймеров и зондов, описанные в настоящем документе, для разработки быстрых и точных молекулярных анализов чувствительности М. tb к лекарственным средствам. Помимо очевидной применимости в анализах молекулярной диагностики М. tb, эти праймеры также должны найти применение при секвенировании генов-мишеней с целью выявления мутаций, вызывающих устойчивость, при анализах в рамках эпидемиологического надзора, и любых других анализах на основе зондов, целью которых является специфичное и чувствительное выявление мутаций, вызывающих устойчивость M. tb к распространенным лекарственным средствам. Последовательности праймеров для амплификации генов dosR, IS6110 и IS1081 позволяют выполнять высокочувствительное и специфичное выявление М. tb и могут использоваться при ПЦР-анализе любого формата, целью которого является высокоспецифичная и чувствительная молекулярная диагностика туберкулеза. В приведенных ниже Таблицах перечислены типичные праймеры и зонды согласно настоящему изобретению.

Одну или более из последовательностей зондов, приведенных в Таблице 2, можно получить в различных форматах обнаружения, например, зонды с двойной меткой, включая линейные зонды, зонды типа Taqman, зонды-молекулярные маяки и зонды-низкоселективные молекулярные маяки. Термин «низкоселективный» зонд относится к зонду, допускающему несоответствие. Зонды, допускающие несоответствие, гибридизуются с более чем одной последовательностью-мишенью и генерируют обнаружимый сигнал при температуре обнаружения в ходе анализа, и различные гибриды, образованные таким образом, характеризуются различными температурами плавления. Линейные или случайным образом спирализованные одноцепочечные зонды, как правило, допускают несоответствие. Примерами таких зондов являются шпилькообразные или линейные зонды с внутренней флуоресцентной группой, уровень флюоресценции которой возрастает после гибридизации с той или иной цепью-мишенью. См., например, патенты США №7662550 и 5925517. US 20130095479.
Низкоселективные зонды предпочтительно представляют собой шпилькообразные зонды с двойной меткой или зонды - молекулярные маяки, описанные в патентах США №7662550 и 5925517. Эти шпилькообразные зонды содержат последовательность, связывающую мишень, фланкированную парой плеч, комплементарных друг другу. Они могут представлять собой ДНК, РНК или ПНК или комбинацию всех трех видов нуклеиновых кислот. Кроме того, они могут содержать модифицированные нуклеотиды и модифицированные межнуклеотидные связи. Они могут содержать первый флуорофор в одном плече и второй флуорофор во втором плече, причем спектр поглощения второго флуорофора по существу перекрывает спектр испускания первого флуорофора. В наиболее предпочтительном случае такие шпилькообразные зонды представляют собой «зонды - молекулярные маяки», содержащие флуорофор в одном плече и гаситель в другом плече, так что зонды не светятся, находясь в свободном состоянии в растворе. Они также могут представлять собой зонды - молекулярные маяки, осуществляющие сдвиг спектра излучения и содержащие, например, несколько флуорофоров в одном плече, которые взаимодействуют посредством резонансного переноса энергии флуоресценции (FRET), и гаситель в другом плече. Длина последовательности, связывающей мишень, может составлять, например, 12-50 или 25-50 нуклеотидов, а длина гибридизирующихся плеч - 4-10 или 4-6 (например, 5 или 6) нуклеотидов. Зонды - молекулярные маяки можно связать с праймерами, как описано в патентах США №7662550 и 5925517 и WO 01/31062.
Таким образом, термин «зонды-низкоселективные молекулярные маяки» относится к такому классу шпилькообразных олигонуклеотидных зондов для гибридизации, меченых флуоресцентной меткой. Такие зонды генерируют обнаружимый сигнал при гомогенном анализе, т.е. анализе без необходимости разделения зондов, гибридизированных с мишенью, и несвязанных зондов. В силу их способности связывать более чем один вариант данной последовательности-мишени, указанные зонды можно использовать при анализах с целью выявления присутствия одного варианта исследуемого сегмента нуклеотидной последовательности среди ряда возможных вариантов или даже обнаружения присутствия двух или более вариантов. Таким образом, зонды можно использовать в комбинации по два или более в одном и том же анализе. Поскольку их последовательности, связывающие мишень, различаются, относительная авидность различается для различных вариантов. Например, первый зонд может сильно связываться с последовательностью дикого типа, в умеренной степени - с первым аллелем, слабо со вторым аллелем и вообще не связываться с третьим аллелем; в то время как второй зонд может слабо связываться с последовательностью дикого типа и первым вариантом и в умеренной степени - со вторым вариантом и третьим вариантом. Дополнительные низкоселективные зонды должны характеризоваться еще более отличающейся картиной связывания вследствие различия из последовательностей, связывающихся с мишенью. Таким образом, спектры испускания флуоресценции комбинации низкоселективных зондов определяют различные штаммы или виды микроорганизмов, а также аллельные варианты/мутации генов.
Поскольку низкоселективные зонды воспроизводимо флуоресцируют с переменной интенсивностью после связывания с различными последовательностями ДНК, их комбинации можно использовать, например, при простых, быстрых и чувствительных анализах на основе реакции амплификации нуклеиновых кислот (например, анализах на основе ПЦР), которые позволяют выявить множество патогенных организмов или вариантов в одном реакционном контейнере. В то же время следует понимать, что указанные анализы также можно выполнять на образцах, предположительно содержащих непосредственно обнаружимые количества неамплифицированных нуклеиновых кислот-мишеней. Этот анализ выявления основан на анализе спектров набора частично гибридизующихся низкоселективных сигнальных зондов, например, зондов-низкоселективных молекулярных маяков, каждый из которых мечен флуорофором, испускающим свет со своим волновым оптимумом с целью получения «спектра сигнатур» видоспецифичных или вариант-специфических последовательностей ДНК.
С помощью зондов можно достичь мультилокусности, например, путем разработки различных зондов-молекулярных маяков для дискриминации аллелей для каждой мишени и дифференциального мечения каждого зонда. (См., например, патенты США №7662550 и 5925517, WO 01/31062, и Tyagi et al. (2000) Nature Biotechnology 18: 1191-1196). Смеси зондов для дискриминации аллелей, каждая из которых содержит аликвоты различного цвета, расширяют количество сигнатур зондов. В связи с этим каждый гибрид молекулярный маяк-мишень с уникальной температурой плавления должен характеризоваться соответствующей уникальной интенсивностью сигнала при определенной температуре и концентрации зонда и ампликона. Таким образом, ограниченное количество низкоселективных зондов можно использовать в качестве зондов для выявления множества различных возможных последовательностей-мишеней посредством ПЦР в реальном времени. Зонды можно добавить в реакционную смесь для амплификации до, во время или после амплификации. См. патент США №7662550.
В настоящем изобретении дополнительно предложены наборы, содержащие реагенты для выполнения вышеописанных способов, включая ПЦР и/или реакции гибридизации зонд-мишень. С этой целью один или более из компонентов реакции, например, ПЦР-праймеров, полимеразы и зондов для способов, описанных в настоящем документе, можно поставлять в виде набора для варианта применения. Такой набор содержит соответствующее количество одного или более из компонентов реакции в одном или более контейнерах или на подложке.
Указанный набор также содержит дополнительные материалы для реализации вышеописанных способов. В некоторых вариантах реализации набор содержит некоторые или все из реагентов, материалов для выполнения способа, в котором используются праймеры и/или зонды согласно настоящему изобретению. Некоторые или все компоненты наборов могут содержаться в контейнерах, отделенных от контейнера(ов), содержащих праймеры и/или зонды согласно настоящему изобретению. Примеры дополнительных компонентов наборов включают одну или более из различных полимераз, один или более из контрольных реагентов (например, зондов или ПЦР-праймеров или контрольных матриц) и буферы для реакций (в 1X или концентрированной форме), но не ограничиваются ими. Набор также может содержать один или более из следующих компонентов: подложек, реагентов для остановки реакции, модификации или гидролиза, осмолитов и аппарат для обнаружения.
Используемые компоненты реакции могут быть представлены в различных формах. Например, компоненты (например, ферменты, зонды и/или праймеры) могут быть суспендированы в водном растворе или представлены в виде лиофилизированного порошка, осадка или гранул. В последнем случае компоненты при восстановлении образуют полноценную смесь компонентов для использования при анализе. Компоненты набора могут быть представлены при любой подходящей температуре. Например, для хранения наборов, содержащих белковые компоненты (например, фермент) в жидкости, предпочтительна доставка и хранение при температуре ниже 0°С, предпочтительно при -20°С или ниже, или иным образом в замороженном состоянии.
Набор или система согласно настоящему изобретению могут содержать любую комбинацию компонентов, описанных в настоящем документе, в количестве, достаточном для выполнения по меньшей мере одного анализа. В некоторых вариантах применения один или более из компонентов реакции может быть представлен в заранее измеренном количестве для однократного применения у индивида, обычно в одноразовых пробирках или эквивалентных контейнерах. При таком расположении ПЦР можно выполнить путем добавления нуклеиновой кислоты-мишени или образца/клетки, содержащих нуклеиновую кислоту-мишень, непосредственно в отдельные пробирки. Количество компонентов, поставляемых в наборе, может представлять собой любое подходящее количество и может зависеть от целевого рынка, на который направляется продукт. Контейнер(ы), в которых поставляются компоненты, могут представлять собой любой обычной контейнер, способный удерживать поставляемую форму, например, микроцентрифужные пробирки, ампулы, бутыли или интегральные устройства для анализа, например, жидкостные устройства, картриджи, устройства бокового потока или другие аналогичные устройства.
Наборы также могут содержать упаковочные материалы для удерживания контейнера или комбинации контейнеров. Типичные упаковочные материалы для таких наборов и систем включают твердые материалы (например, стекло, пластик, бумагу, фольгу, микрочастицы и т.п.), удерживающие компоненты реакции или зонды для обнаружения в любой из различных конфигураций (например, во флаконе, лунке титрационного микропланшета, микрочипе и т.п.). Наборы также могут дополнительно содержать инструкции, записанные в материальной форме для применения компонентов.
Определения
Нуклеиновая кислота относится к молекуле ДНК (например, кДНК или геномной ДНК, но не ограничиваясь ими), молекуле РНК (например, мРНК, но не ограничиваясь ею), или аналогу ДНК или РНК. Аналог ДНК или РНК можно синтезировать из аналогов нуклеотидов. Молекула нуклеиновой кислоты может быть двуцепочечной или одноцепочечной. «Выделенная» нуклеиновая кислота представляет собой нуклеиновую кислоту, структура которой не идентична любой природной нуклеиновой кислоте или любому фрагменту природной геномной нуклеиновой кислоты. Таким образом, данный термин охватывает, например, (а) ДНК, содержащую последовательность фрагмента природной молекулы геномной ДНК, но не фланкированную обеими кодирующими последовательностями, фланкирующими указанный фрагмент молекулы в геноме организма, в котором он встречается в природе; (b) нуклеиновую кислоту, включенную в вектор или в геномную ДНК прокариот или эукариот таким образом, что полученная молекула не является идентичной любому природному вектору или геномной ДНК; (с) отдельную молекулу, например, кДНК, фрагмент генома, фрагмент, полученный посредством ПЦР, или рестрикционный фрагмент; и (d) рекомбинантную нуклеотидную последовательность, являющуюся фрагментом гибридного гена, т.е. гена, кодирующего гибридный белок.
В настоящем документе термин «нуклеиновая кислота-мишень» или «мишень» относится к нуклеиновой кислоте, содержащей исследуемую нуклеотидную последовательность-мишень. Нуклеиновая кислота-мишень может быть одноцепочечной или двуцепочечной, и часто представляет собой двуцепочечную ДНК. «Нуклеотидная последовательность-мишень», «последовательность-мишень» или «область-мишень» означает определенную последовательность, полностью или частично содержащую последовательность одноцепочечной нуклеиновой кислоты. Последовательность-мишень может находиться в нуклеотидной матрице или в геноме клетки, которые могут представлять собой любую форму одноцепочечной или двуцепочечной нуклеиновой кислоты. Матрица может представлять собой очищенную или выделенную нуклеиновую кислоту, или может быть неочищенной или не выделенной.
"Комплементарные" последовательности в настоящем документе могут содержать или быть полностью образованными уотсон-криковскими парами оснований (например, A-T/U и C-G), не уотсон-криковскими парами оснований и/или парами оснований, образованными неприродными и модифицированными нуклеотидами, при условии соблюдения вышеприведенных требований касательно их способности к гибридизации. Полная комплементарная цепь или полностью комплементарный может означать 100% (полностью) комплементарное или по существу комплементарное спаривание оснований между нуклеотидов или аналогами нуклеотидов молекул нуклеиновой кислоты.
«В значительной мере комплементарный» означает, что нуклеиновая кислота или олигонуклеотид обладает последовательностью, содержащей по меньшей мере 10 смежных оснований, которые по меньшей мере на 80%, (например, 85%, 90%, 95%, 96%, 97%, 98%, 99% и 100%) комплементарны по меньшей мере 10 смежным основаниям в нуклеотидной последовательности-мишени, так что указанная нуклеиновая кислота или олигонуклеотид может гибридизоваться или отжигаться с нуклеотидной последовательностью-мишенью, например, при условиях отжига при ПЦР или условиях гибридизации зонда и мишени. Комплементарность между последовательностями можно выражать в виде количества несовпадающих оснований в каждом сравниваемом наборе из по меньшей мере 10 смежных оснований. Термин «по существу идентичный» означает, что первая нуклеиновая кислота по меньшей мере на 80%), (например, на 85%, 90%, 95%, 96%), 97%, 98%, 99% и 100%) комплементарна второй нуклеиновой кислоте, так что первая нуклеиновая кислота по существу комплементарна комплементарной цепи второй нуклеиновой кислоты и способна гибридизоваться с ней в условиях ПЦР-отжига или гибридизации зонда и мишени.
Термины «гибридизация» или «гибридизующийся» или «гибридизироваться» или «отжигать» относится к способности полностью или частично комплементарных цепей нуклеиновой кислоты соединяться друг с другом в указанных условиях гибридизации в параллельной или предпочтительно антипараллельной ориентации с образованием стабильной двуцепочечной структуры или области (иногда называемой «гибридом» или «дуплексом» или «стеблем»), в которой две составляющих цепи соединены водородными связями. Хотя водородные связи обычно образуются между аденином и тимином или урацилом (А и Т или U) или цитозином и гуанином (С и G), могут образовываться и другие пары оснований (например, Adams et al., The Biochemistry of the Nucleic Acids, 11th ed., 1992).
Термины "нуклеотидный дуплекс", "дуплекс", "стебель", "нуклеотидный гибрид" или "гибрид" относятся к стабильной нуклеотидной структуре, содержащей двуцепочечную нуклеотидную структуру с водородными связями, например, двуцепочечным молекулам РНК:РНК, РНК:ДНК и ДНК:ДНК и их аналогам. Такую структуру можно обнаружить любыми известными средствами, например, с использованием меченого зонда, оптически активной подложки, покрытой зондом, чувствительной к изменениям массы на ее поверхности (патент США №6060237) или связывающих агентов (патент США №5994056).
В настоящем документе термин «амплификация» и его варианты включает любой процесс получения множественных копий или комплементарных цепей по меньшей мере фрагмента полинуклеотида, причем указанный полинуклеотид обычно называют «матрицей». Матричный полинуклеотид может быть одноцепочечным или двуцепочечным. Матрица может представлять собой очищенную или выделенную нуклеиновую кислоту, или может быть неочищенной или не выделенной. Амплификация заданной матрицы может привести к получению популяции продуктов амплификации полинуклеотида, совместно называемых «ампликоном». Полинуклеотиды ампликона могут быть одноцепочечными или двуцепочечными или представлять собой смесь одноцепочечных и двуцепочечных полинуклеотидов. Как правило, матрица содержит последовательность-мишень, и полученный ампликон должен содержать полинуклеотиды, обладающие последовательностью, по существу идентичной или по существу комплементарной последовательности-мишени. В некоторых вариантах реализации полинуклеотиды конкретного ампликона по существу идентичны или по существу комплементарны друг другу; в качестве альтернативы, в некоторых вариантах реализации нуклеотидные последовательности полинуклеотидов в составе заданного ампликона могут отличаться друг от друга. Амплификация может происходить по линейному или экспоненциальному принципу и может включать повторяющуюся или последовательную репликацию заданной матрицы с образованием двух или более продуктов амплификации. Некоторые типичные реакции амплификации включают последовательные и повторяющиеся циклы синтеза нуклеиновой кислоты на основе матрицы с образованием множества дочерних полинуклеотидов, содержащих по меньшей мере часть нуклеотидной последовательности матрицы и характеризующихся по меньшей мере некоторой общей степенью идентичности (или комплементарности) нуклеотидной последовательности по сравнению с матрицей. В некоторых вариантах реализации каждый случай синтеза нуклеиновой кислоты, который можно называть «циклом» амплификации, включает создание свободного 3'-конца (например, путем никирования одной цепи дцДНК), что приводит к образованию праймера и этапам удлинения праймера; необязательно можно включить дополнительные этапы денатурации, при которых происходит полная или частичная денатурация матрицы. В некоторых вариантах реализации один запуск амплификации включает заданное количество повторов цикла амплификации. Например, запуск амплификации может включать 5, 10, 15, 20, 25, 30, 35, 40, 50 или более повторов конкретного цикла. В одном типичном варианте реализации амплификация включает любую реакцию, при которой конкретную полинуклеотидную матрицу подвергают двум последовательным циклам синтеза нуклеиновой кислоты. Синтез может включать матричный синтез нуклеиновой кислоты.
Амплификация согласно настоящему изобретению также может включать изотермическую амплификацию. Термин «изотермический» означает выполнение реакции при практически постоянной температуре, т.е. без изменения температуры реакции, при которой происходит реакция полимеризации нуклеиновых кислот. Изотермические температуры для реакций изотермической амплификации зависят от вытесняющей полимеразы нуклеиновых кислот, используемой при реакциях. Как правило, изотермические температуры ниже температуры плавления (Tm; температуры, при которой половина потенциально двуцепочечных молекул в смеси находятся в одноцепочечном, денатурированном состоянии) преобладающего продукта реакции, т.е., как правило, составляют 90°С или ниже, обычно составляют между приблизительно 20°С и 75°С, предпочтительно составляют между приблизительно 30°С и 60°С, и более предпочтительно составляют приблизительно 37°С.
Термин «праймер» или «праймерный олигонуклеотид» относится к нуклеотидной цепи или олигонуклеотиду, способным гибридизоваться с нуклеотидной матрицей и действующим в качестве сайта инициации включения нуклеотидов при удлинении цепи в соответствии с составом нуклеотидной матрицы для синтеза нуклеиновой кислоты. "Нуклеотиды для удлинения цепи" относятся к любым нуклеотидам (например, dNTP) и их аналогам, способным встраиваться в удлиняющийся продукт при амплификации, т.е. ДНК, РНК или производное ДНК или РНК, которые могут содержать метку.
В настоящем документе термин «олигонуклеотид» относится к короткому полинуклеотиду, длина которого обычно меньше или равна 300 нуклеотидам (например, находится в диапазоне от 5 до 150, предпочтительно в диапазоне от 10 до 100, более предпочтительно в диапазоне от 15 до 50 нуклеотидов). Однако в настоящем документе также подразумевается, что данный термин охватывает более длинные или более короткие полинуклеотидные цепи. «Олигонуклеотид» может гибридизоваться с другими полинуклеотидами, таким образом действуя в качестве зонда для обнаружения полинуклеотида или праймера для удлинения полинуклеотидной цепи.
Термин «зонд» в настоящем документе относится к олигонуклеотиду, способному связываться к нуклеиновой кислотой-мишенью с комплементарной последовательностью посредством химических связей одного или более типов, обычно посредством образования комплементарных пар оснований, обычно за счет образования водородных связей. Зонды могут связывать последовательности-мишени, не полностью комплементарные последовательности зонда, в зависимости от жесткости условий гибридизации. Может присутствовать любое количество несовпадающих пар оснований, которые должны мешать гибридизации между последовательностью-мишенью и одночепочечными нуклеиновыми кислотами, описанными в настоящем документе. Однако если количество мутаций настолько велико, что гибридизация невозможна даже при наименее строгих условиях гибридизации, указанная последовательность не комплементарна последовательности-мишени. Зонд может быть одноцепочечным или частично одно- и частично двуцепочечным. Количество цепей в составе зонда определяется структурой, составом и свойствами последовательности-мишени. Зонды могут быть непосредственно или опосредованно мечены меткой, например, биотином, с которым позже можно связать комплекс стрептавидина.
Термин «зонд для обнаружения» относится к олигонуклеотиду, обладающему последовательностью, по существу комплементарной последовательности-мишени, и способному образовывать стабильный для обнаружения гибрид зонд:мишень в жестких условиях гибридизации. Зонд обычно представляет собой синтетический олигомер, который может содержать основания, комплементарные последовательности, находящейся за пределами области мишени, что не препятствует гибридизации в строгих условиях гибридизации с нуклеотидной мишенью. Последовательность, не комплементарная мишени, может представлять собой гомополимерный участок (например, полиА или полиТ), последовательность промотора, последовательность, распознаваемую эндонуклеазой рестрикции, или последовательность, придающую желательную вторичную или третичную структуру (например, каталитический сайт или структуру шпильки), или маркерную область, которая может облегчать обнаружение и/или амплификацию. Термин «стабильный» или «стабильный для обнаружения» означает, что температура реакционной смеси по меньшей мере на 2°С ниже температуры плавления (Tm) нуклеотидного дуплекса, содержащегося в смеси, более предпочтительно по меньшей мере на 5°С ниже Tm, и еще более предпочтительно по меньшей мере на 10°С ниже Tm.
«Метка» или «репортерная молекула» представляет собой химический или биохимический объект, который можно использовать для мечения нуклеиновой кислоты (в том числе одиночного нуклеотида), полинуклеотида, олигонуклеотида или белкового лиганда, например, аминокислоты или антитела. Примеры включают флуоресцентные агенты, хемилюминесцентные агенты, хромогенные агенты, гасители, радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, магнитные частицы и другие химические объекты, известные в данной области техники. Метки или репортерные молекулы могут генерировать измеримый сигнал и могут ковалентно или нековалентно присоединяться к олигонуклеотиду или нуклеотиду (например, неприродному нуклеотиду) или лиганду.
В настоящем документе термин «приведение в контакт» и его варианты, используемые по отношению к любому набору компонентов, включает любой процесс, при котором компоненты, которые должны контактировать, смешивают в одну и ту же смесь (например, добавляют в один и тот же отсек или раствор) и не обязательно требует реального физического контакта между указанными компонентами. Указанные компоненты можно приводить в контакт в любом порядке или любой комбинации (или подкомбинации), включая ситуации, где один или некоторые из указанных компонентов впоследствии удаляют из смеси, необязательно до введения других указанных компонентов. Например, «приведение в контакт А с В и С» включает любые из следующих ситуаций: (i) А смешивают с С, затем В добавляют в смесь; (ii) А и В смешивают в смесь; В удаляют из смеси, а затем добавляют в смесь С; и (iii) А добавляют в смесь В и С. «Контакт» нуклеотидной мишени или клетки с одним или более компонентами реакции, например, полимеразой, набором праймеров или зондом, включает любую или все из следующих ситуаций: (i) мишень или клетка контактирует с первым компонентом реакционной смеси с образованием смеси; затем в смесь добавляют другие компоненты реакционной смеси в любом порядке или комбинации; и (ii) реакционную смесь полностью образуют до смешивания с мишенью или клеткой.
Термин «смесь» в настоящем документе относится к комбинации элементов, которые находятся вперемешку друг с другом, но не в определенном порядке. Смесь является разнородной и пространственно не разделяется на компоненты. Примеры смесей элементов включают ряд различных элементов, растворенных в одном и том же растворе, или ряд различных элементов, присоединенных к твердой подложке в случайном порядке или без какого-либо порядка, причем различные элементы не разделены пространственно. Другими словами, смесь не является адресуемой.
В настоящем документе термин «субъект» относится к любому организму, обладающему геномом, предпочтительно, живому животному, например, млекопитающему, являющемуся объектом диагностики, лечения, наблюдения или эксперимента. Примером субъекта может быть человек, продуктивное животное (мясной и молочный скот, овцы, птицы, свиньи и т.д.) или животное-компаньон (собаки, кошки, лошади и т.д.).
«Образец» в настоящем документе означает любые биологические жидкости или ткани, полученные из организма (например, пациента), или компоненты организма (например, кровь). Образец может представлять собой любую биологическую ткань, клетку(и) или жидкость. Образец может представлять собой «клинический образец», являющийся образцом, полученным от субъекта, например, пациента-человека или субъекта-животного. Используемые биологические образцы включают, без ограничения, цельную кровь, слюну, мочу, синовиальную жидкость, костный мозг, спинномозговую жидкость, вагинальную слизь, цервикальную слизь, выделения из носа, мокроту, сперму, амниотическую жидкость, бронхоальвеолярный лаваж и другие клеточные экссудаты пациента или субъекта. Такие образцы можно дополнительно разбавить раствором NaCl, буфером или физиологически приемлемым разбавителем. В качестве альтернативы, такие образцы концентрируют обычными средствами. Биологические образцы также могут включать срезы, например, замороженные срезы тканей для гистологических целей. Биологический образец также можно называть «образцами пациента». Биологический образец может также включать по существу очищенный или выделенный белок, мембранный препарат или культуру клеток.
Термины «определение», «измерения», «оценка» и «анализ» используются на равных основаниях и включают как количественные, так и качественные измерения и определение наличия или отсутствия характеристик, признаков или свойств. Оценка может быть относительной или абсолютной. Оценка наличия мишени включает определение количества присутствующей мишени, а также определение ее наличия или отсутствия.
В настоящем документе термин «эталонное» значение относится к значению, статистически коррелирующему с конкретным результатом при сравнении с результатом анализа. В предпочтительных вариантах реализации эталонное значение можно определить на основании статистического анализа, исследующего среднее значение дикого типа. Эталонное значение может представлять собой пороговое оценочное значение или предельное оценочное значение. Обычно эталонное значение является верхним (или нижним) пороговым значением, в зависимости от того, какой результат является более вероятным, и насколько вероятным является альтернативный результат менее указанного значения.
В настоящем документе различие этих значений свидетельствует о наличии или отсутствии возбудителя (например, Mycobacterium tuberculosis) или мутации. Фраза «различие» уровня или значения относится к различиям значений переменной (например, Tm) анализируемого соединения (например, гибрида зонд-мишень) в образце по сравнению с контрольным или эталонным уровнем или значением. В одном варианте реализации различие значения или уровня может представлять собой статистически значимое различие между количеством анализируемого соединения в образце по сравнению с контролем. Например, различие может быть статистически значимым, если измеренный уровень анализируемого соединения выходит за пределы 1,0, 2,0, 3,0, 4,0 или 5,0 стандартных отклонений от среднего значения любой контрольной или эталонной группы.
В настоящем документе представлен ряд диапазонов значений. Следует понимать, что любое промежуточное значение, вплоть до десятых долей величины нижнего предела (если иное не следует из контекста явным образом), располагающееся между верхним и нижним пределами указанного диапазона, также включено в описание изобретения. Каждый меньший диапазон, находящийся между любыми указанными значениями, или промежуточное значение в указанном диапазоне и любое другое указанное или промежуточное значение в указанном диапазоне входят в рамки настоящего изобретения. Верхний и нижний пределы этих меньших диапазонов могут независимо входить в эти диапазоны или не входить в них, и каждый диапазон где любое или оба предельных значения входят или не входят в меньшие диапазоны, также включены в рамки настоящего изобретения, за исключением любого явным образом исключенного предельного значения в указанном диапазоне. Если указанный диапазон включает одно или оба предельных значения, диапазоны, в которые не входит одно или оба из этих предельных значений, также входят в рамки изобретения. Термин "приблизительно" обычно означает плюс-минус 10% от указанного численного значения. Например, «приблизительно 10%» может означать диапазон от 9 до 11%, а «приблизительно 20» может означать 18-22%. Другие значения термина «приблизительно» (например, округление) могут быть очевидны из контекста, например, «приблизительно 1» может также означать от 0,5 до 1,4.
Применение
Недавно описаны два отдельных анализа на основе SMB, позволяющие быстро и надежно выявлять мутации М. tb, которые по большей части обуславливают устойчивость к рифампицину и фторхинолонам (FQ) (Chakravorty, S., В. et al., 2011, J Clin Microbiol 49:932-940; Chakravorty, S., H., et al., 2012, J Clin Microbiol 50:2194-2202). Преимущество анализов, описанных в настоящем документе, состоит в том, что они представлены в формате ПЦР в реальном масштабе, так что они просты в применении и не подвержены перекрестному загрязнению ампликонов. Кроме того, показано, что обнаружения мутации является надежным и пригодным для высокопроизводительных анализов. Приведенные ниже примеры представляют систему анализа на основе SMB для обнаружения лекарственной устойчивости ТВ с добавлением анализов, позволяющих обнаруживать устойчивость к АМК и KAN. Кроме того, изучены причины несовпадения результатов анализа, описанного в настоящем документе, и фенотипических способов анализа чувствительности. Результаты показывают, что некоторые из наиболее часто используемых фенотипических способов могут пропускать изоляты М. tb с мутациями, обуславливающими устойчивость, если эти мутации лишь умеренно увеличивают значения минимальной ингибиторной концентрации (MIC) KAN. Кроме того, обнаружены новые мутации в whiB7, ассоциированные с низким уровнем устойчивости к KAN.
Как описано в настоящем документе, многолокусная SMB-ПЦР и анализ плавления позволяли точно выявлять мутации в гене rrs гена и промоторе eis, связанные с устойчивостью к АМК и/или KAN. Анализ не выдает ложноположительных результатов устойчивости при тестировании на NTM, грамположительных и грамотрицательных бактериях. Большинство случаев гетерогенной устойчивости также обнаруживались посредством данного анализа (при их наличии). В отличие от платформы MTBDRsl, данный анализ можно выполнять в замкнутой системе ПЦР в реальном времени и легко адаптировать для высокопроизводительного анализа, поскольку все этапы анализа выполняются в 384-луночных планшетах. SMB-анализ также позволяет избежать потенциальных проблем, связанных с альтернативными способами обнаружения мутаций. Анализ кривой плавления с высоким разрешением требует возможности обнаруживать мелкие различия в кривых плавления (Yadav, R., S., et al., 2012, J Appl Microbiol 113:856-862). Другие молекулярные анализы плавления после ПЦР должны обнаруживать мутации при декодировании сложных контуров флуоресценции (Rice, J.Е., et al., 2012. Nucleic Acids Res 40:e164.) В отличие от этого, SMB-анализ позволяет получить четкие и легко различимые пики и характерные сдвиги Tm для выявления исследуемых мутаций. Отдельные значения Tm также можно использовать для формирования кластеров образцов, обладающих одним и тем же генотипом.
Как описано в настоящем документе, анализ тестировали на панели из 603 клинических образцов, представляющих как новые случаи ТВ, так и невылеченные случаи повторного лечения, и оценивали взаимосвязь целевых мутаций с картиной чувствительности клинических изолятов. Отмечено, что 100% изолятов с мутацией A1401G в гене rrs характеризовались сильной корреляцией с высоким уровнем устойчивости как к АМК, так и к KAN. Однако мутации промотора eis приводили лишь к умеренной или слабой устойчивости к KAN и не приводили к устойчивости к АМК, что согласуется с предыдущими исследованиями (Campbell, PJ, et al., 2011, Antimicrob Agents Chemother 55: 2032-2041, Zaunbrecher, MA, et al., 2009, Proc Natl Acad Sci USA 106: 20004-20009). Настоящее исследование также показало, что способ абсолютной концентрации LJ для проверки чувствительности не позволяет адекватно обнаруживать умеренную или слабую устойчивость к KAN. Фактически, почти две трети образцов с мутациями промотора eis обнаруживались как чувствительные к KAN при использовании среды LJ. Вместе с тем, все образцы с мутациями промотора eis, кроме одного, обнаруживались как устойчивые к KAN при использовании способа MGIT. Два таких изолята содержали С(-12)Т-мутацию eis. Эти мутанты также были устойчивы к KAN при тестировании с использованием MYCOTB, которое показало, что MIC KAN составляла 5 мкг/мл. Предыдущие исследования показали, что клинические изоляты с мутациями С(-12)Т не коррелируют (Zaunbrecher (2009) или плохо коррелируют (Campbell, 2011; Hoshide, М., L. et al., 2014, J Clin Microbiol 52: 1322 -1329.) с устойчивостью к KAN. Эти исследования, возможно, не принимали во внимание связь между этой мутацией и низким уровнем устойчивости к KAN из-за способа тестирования, использованного для установления фенотипической чувствительности. Эти результаты показывают, что способы MGIT или MYCOTB должны быть предпочтительными при тестировании фенотипической устойчивости к KAN. Кроме того, они особо выделяют эффективность тестов генотипической устойчивости, например, описанных в настоящем документе, при выявлении мутаций, обуславливающих слабую устойчивость, которые могут пропускаться при использовании только фенотипических тестов (Rigouts, L., М. et al., 2013, J Clin Microbiol 51: 2641-2645, Sirgel, FA, et al., 2012, Microb Drug Resist 18: 193-197, и Van Deun, A., et al., 2013, J Clin Microbiol 51: 2633-2640).
Приведенное в настоящем документе исследование включало один образец, который представлял собой смесь rrs дикого типа и rrs-мутантов С1402Т. Этот образец был чувствителен как к АМК, так и к KAN при использовании среды LJ. Сообщалось, что изоляты с мутациями С1402Т были чувствительны к АМК, но устойчивы к KAN (Maus, СЕ, et al., 2005, Antimicrob Agents Chemother 49: 3192-3197). В данном конкретном случае повторные анализы чувствительности с использованием среды LJ продемонстрировали восприимчивость к KAN, предположительно, из-за гетерогенной устойчивости образца. В настоящем изобретении молекулярный анализ являлся лучшим прогностическим фактором потенциальной устойчивости, чем фенотипический анализ, поскольку SMB-анализ позволял точно обнаружить присутствие как ДНК дикого типа, так и мутантной ДНК.
Частота мутаций 1484 rrs в клинических штаммах с устойчивостью к АМК или KAN была очень низкой (Georghiou, SB, et al., 2012, PLoS One 7: e33275), что делает ее клиническое значение спорным. Отдельная версия анализа, мишенью которой является кодон 1484 rrs, не обнаружила мутаций ни в одном из 603 исследованных изолятов, а также в дополнительных 259 изолятах из района Нью-Джерси - Нью-Йорк, в который вошли 33 изолята, устойчивые к АМК и KAN. Отсутствие мутаций 1484 rrs в этом расширенном исследуемом наборе подтвердили секвенированием по Сэнгеру (данные не показаны). В свете крайне низкой распространенности мутаций 1484 rrs маловероятно, что этот кодон обладает большой ценностью при прогнозировании устойчивости к аминогликозидам. Таким образом, при молекулярных анализах устойчивости к аминогликозидам рекомендуется использовать только кодоны 1401-1402 гена rrs.
Обнаружено, что 22 штамма, устойчивые к АМК или KAN, содержали последовательности дикого типа гена rrs и области промотора eis. В недавнем исследовании показана возможная связь между мутациями в 5'UTR whiB7 и устойчивостью к KAN путем выявления 5'UTR-мутации whiB7 в одном клиническом штамме с необъяснимой устойчивостью к KAN (Reeves, AZ, et al., 2013, Antimicrob Agents Chemother 57: 1857-1865). Кроме того, описаны несколько новых 5'UTR-мутаций, а также делеция в whiB7, которые, по-видимому, ассоциированы с устойчивостью к KAN. Подходящие универсальные биомаркеры, которые могут являться причиной устойчивости к KAN и АМК в остальных 15-20% клинических штаммов с rrs и областью промотора eis дикого типа, не выявлены. Образцы, содержащие ДНК гена rrs и области промотора eis дикого типа, смешанные с следовыми количествами мутантных мишеней из субпопуляции, устойчивой к KAN или АМК, могут также объяснять остальные несоответствия между тестами фенотипической устойчивости и SMB-анализом, описанным в настоящем документе. Однако дорогостоящее исследование гетерогенной устойчивости выходит за рамки настоящего исследования. В некоторых недавних исследованиях предполагалось, что гены РРЕ60 и Rv3168 могут быть вовлечены в механизмы необъяснимой устойчивости к KAN (Farhat, MR, et al., 2013). Nat Genet 45: 1183-1189; Zhang, H., et al., 2013, Nat Genet 45: 1255-1260), хотя это еще предстоит проверить в клинических условиях.
Таким образом, разработан чувствительный и специфичный анализ для обнаружения устойчивости к АМК и KAN у М tb, и выполнена его валидация на клинических изолятах с высокой распространенностью МЛУ и ШЛУ ТВ. Результаты показывают, что мутации A1401G в rrs кодируют высокоуровневую перекрестную устойчивость как к АМК, так и к KAN, и что мутации промотора eis кодируют умеренную или слабую устойчивость к KAN, что согласуется с предыдущими функционально-геномными исследованиями (Zaunbrecher 2009). При сравнении эффективности анализа, описанного в настоящем документе, с тремя различными способами анализа фенотипической чувствительности на твердых и жидких средах обнаружено, что анализы чувствительности на основе LJ по существу пропускают слабую или умеренную устойчивость к KAN, вызванную мутациями промотора eis. Эти результаты убедительно доказывают ценность генотипических тестов для обнаружения устойчивости к аминогликозидам, а также демонстрируют возможность специфического применения анализов на основе SMB, описанных в настоящем документе.
ПРИМЕРЫ
Пример 1
В данном примере описаны материалы и способы, используемые в примерах 2-7, представленных ниже.
Образцы ДНК
Тестовые образцы M. tb состояли из ДНК, выделенной из 603 последовательных изолятов М. tb, полученных от 503 пациентов, зачисленных в исследование естественной динамики МЛУ-туберкулеза (NCT00341601 на сайте cliniatialials.gov) в национальной больнице Масан в Чангвоне, Республика Корея. Протестировали две когорты. Когорта А состояла из наивных по отношению к лечению новых случаев ТВ (158 образцов), а когорта В состояла из случаев повторного лечения ТВ (445 образцов). Образцы свежей мокроты собирали у каждого пациента в начале лечения и культивировали для выделения М. tb. В подгруппе пациентов повторно собирали образцы мокроты в 1-й, 4-й и 6-й месяц лечения, и также культивировали их для выделения М. tb. Тестовые образцы микобактерий, не являющихся возбудителями туберкулеза (NTM), а также грамположительных и грамотрицательных бактерий были получены из репозитория ДНК медицинской школы Нью-Джерси (NJMS), как описано ранее (Chakravorty, S., 2012. J Clin Microbiol 50:2194-2202).
Анализ фенотипической чувствительности к лекарственным средствам
Анализ фенотипической чувствительности к лекарственным средствам выполняли для всех 603 изолятов согласно способу абсолютной концентрации на среде LJ с целью определения чувствительности к АМК и KAN с использованием критических концентраций 40 мкг/мл (стандартная концентрация, использовавшаяся в 2012 г., когда анализировали данные изоляты) обоих антибиотиков (Jnawali, HN, 2013, Diagn Microbiol Infect Dis 76: 187-196) в Международном научно-исследовательском центре туберкулеза (ITRC), Южная Корея. MIC АМК и KAN для 173/603 образцов также оценивали с использованием TREK Sensititre® MYCOTB MIC-планшетов («MYCOTB», TREK Diagnostic Systems, Кливленд, Огайо, США), как описано ранее (Lee, J., 2014, Antimicrob Agents Chemother 58: 11-18). Для 560/603 образцов также оценивали устойчивость к KAN с использованием системы индикаторных пробирок для роста микобактерий (Mycobacterial Growth Indicator Tube, MGIT) (Becton Dickinson, Франклин-Лейкз, штат Нью-Джерси, США) при критической концентрации 2,5 мкг/мл. Для образцов с результатами анализа фенотипической чувствительности, не согласующимися с результатами секвенирования генов-мишеней по Сэнгеру, анализ фенотипической чувствительности повторяли для подтверждения первоначальных результатов. В тех случаях, когда результаты анализов чувствительности на основе MGIT и LJ были не согласованы, оба анализа повторяли для подтверждения или исправления исходных результатов.
Подготовка и секвенирование ДНК
ДНК для SMB-анализа и секвенирования по Сэнгеру получали из культивируемых изолятов путем кипячения одной петли культуры в 200 мкл смолы Instagene Matrix (Bio-Rad Laboratories, Херкулиз, штат Калифорния, США) в присутствии 0,1% Triton X100 в течение 10-15 минут. Супернатант извлекали после центрифугирования и количественно анализировали с использованием спектрофотометра микрообъемных образцов Nanodrop (Thermo Fisher Scientific, Уолтем, штат Массачусетс, США). Для секвенирования по Сэнгеру два разных фрагмента гена rrs (нуклеотиды 420-980 и 1293-1537) и фрагмент выше кодирующей области eis со всей областью промотора eis амплифицировали с использованием 0,5 мкМ прямого и обратного праймеров, 1X буфера для ПЦР, 250 мМ dNTP, 2,5 мМ MgCl2 и 0,03 ед/мкл ДНК-полимеразы AmpliTaq Gold (Applied Biosystems, Фостер-Сити, Калифорния, США) в соответствии со следующими параметрами: начальная денатурация при 95°С в течение 10 мин, затем 40 циклов при 95°С в течение 10 с, 58 -60°С в течение 30 с и 72°С в течение 10-30 с в зависимости от размера ампликона. Промоторную область eis и фрагменты гена rrs амплифицировали, как описано ранее (10, 33). Для подгруппы образцов амплифицировали и секвенировали фрагмент гена whiB7 длиной 538 п.о., содержащий 412 п.о. 5'-нетранслируемой области и 126 п.о. ОРС, с использованием праймеров whiB7F 5'-aaacgcgcaggtcagaaaat-3' и whiB7R 5'-cagtgtcttggctacctcga-3' (SEQ ID NO: 70 и 71). Кроме того, амплифицировали фрагмент гена whiB7 длиной 275 п.о., содержащий почти всю ОРС whiB7, с использованием праймеров whiB7-ingene-F 5'-GTCGGTACTGACAGTCCCC-3' и whiB7-ingene-R 5'-ATGCAACAGCATCCTTGCG-3' (SEQ ID NO: 72 и 73). Продукты ПЦР подвергали двунаправленному секвенированию с использованием ген-специфических прямых и обратных праймеров в генетическом анализаторе 3130XL (Applied Bio-Systems, Фостер-Сити, штат Калифорния, США) с использованием набора для циклического секвенирования BigDye Terminator версии 3.1 (Applied Biosystems) в соответствии с инструкциями производителя.
Молекулярные маяки и праймеры для анализа
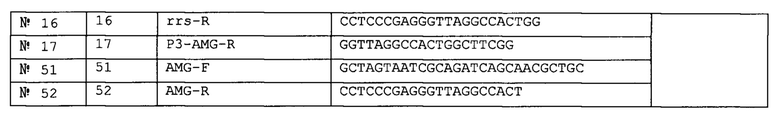
Мишенями SMB-анализов являлись мутации М. tb в ко донах 1401 и 1402 гена rrs и мутации области промотора гена eis. Фрагмент длиной 113 п. о. (нуклеотиды 1335-1451) гена rrs амплифицировали с использованием праймеров AMG-F (5'-GCTAGTAATCGCAGATCAGCAACGCTGC-3', SEQ ID No: 51) и AMG-R (5'-CCTCCCGAGGGTTAGGCCACT-3', SEQ ID No: 52), а фрагмент длиной 98 п. о., охватывающий область промотора и начальные пять кодонов гена eis (от -81 до 17 нуклеотида) амплифицировали с использованием праймеров eis-F (5'-CACAGGGTCACAGTCACAGAATC-3', SEQ ID No: 18) и eis-R (5'-GCATCGCGTGATCCTTTGCCAGAC-3', SEQ ID No: 53). Конструкция праймеров rrs предусматривала специфичность по отношению к роду Mycobacterium, а конструкция праймеров eis - к комплексу М. tb. Мишенями одного SMB-зонда rrs-1400 (5'-6-карбоксифлуоресцеин-cacgaccgcccgtcacgtcatgaaagtcggtcgtg-BHQ1-3,, SEQ ID No: 59) и двух SMB-зондов eis-1 (5'-цианин-5-caggcggtcgtaatattcacgtgcacctggccgccgcctg-BHQ2-3', SEQ ID No: 16) и eis-2 (5'-TexasRed-ctcgcggcatatgccacagtcggattctctgacgcgag-BHQ2-3', SEQ ID No: 61) (где подчеркнутые последовательности представляют собой основную часть SMB, а BHQ представляет собой гаситель Black Hole Quencher) являлись ген rrs и область промотора eis, соответственно. Зонд rrs был комплементарен антисмысловой цепи, а зонды eis - смысловой цепи SMB сконструировали с использованием программы фолдинга ДНК in silico на сайте http://mfold.rna.albany.edu/?q_mfold/dna-folding-form, а программы фолдинга гибрида зонд-мишень на сайте http://mfold.rna.albany.edu/?q_DiNAMelt/Two-state-melting использовали для прогнозирования возможных структур и температур плавления (Tm) гибридов зонд-мишень. Конструкция зондов предусматривала получение максимальной разности Tm между последовательностями дикого типа и мутантными последовательностями в соответствующих областях-мишенях для однозначного выявления мутаций. Праймеры получили от Sigma Aldrich (Сент-Луис, штат Миссури, США), а SMB-зонды - от Biosearch Technologies (Новато, штат Калифорния, США).
Процедура анализа
Все образцы независимо кодировали и распределяли случайным образом для обеспечения слепой валидации анализа. Анализ тестировали в ITRC в Масане, Южная Корея, и медицинской школе Нью-Джерси (NJMS), университет Рутгерса, Ньюарк, штат Нью-Джерси, США. После завершения тестирования всего набора из 603 образцов образцы декодировали и результаты ПЦР сравнивали с соответствующими результатами секвенирования и анализа фенотипической чувствительности к лекарственным средствам. Также сравнивались результаты, полученные в каждом центре. Результаты анализа не сообщали лечащим врачам и не использовали при принятии решений о лечении. ПЦР выполняли на 384-луночных планшетах с использованием системы ПЦР в реальном времени Roche Light Cycler 480 II (Roche Diagnostics Co., Индианаполис, штат Индиана, США) в 20-мкл объеме реакционной среды, содержащей 100 нм прямого праймера и 1 мкМ обратного праймера для гена rrs и 1 мкМ прямого праймера и 50 нМ обратного праймера для области промотора eis, 1 нг/мкл rrs-1400 и eis-1 и 0,8 нг/мкл eis-2, 4 мМ MgCl2, 250 мМ дезоксинуклеозидтрифосфатов (dNTP), 1X ПЦР-буфер, 8% глицерина, 0,06 ед/мкл ДНК-полимеразы Platinum® TfiExo(-) (Life Technologies, Гранд-Айленд, штат Нью-Йорк, США) и от 2 до 5 нг ДНК образца или эквивалентный объем воды. ПЦР выполняли в несколько следующих этапов: активация фермента в течение 2 мин при 95°С, затем 50 циклов денатурации при 95°С в течение 10 с и комбинированного отжига и удлинения при 67°С в течение 30 с. После выполнения циклов ПЦР выполняли анализ Tm после ПЦР путем денатурации при 95°С в течение 2 мин с последующим охлаждением до 45°С, а затем постепенным нагреванием до 85°С с непрерывным контролем флуоресценции с частотой 1 регистрация данных на градус Цельсия. Значения Tm определяли в конце реакции с использованием программного обеспечения для анализа Tm (Light Cycler 480). Вместе с тем, каждое значение Tm также проверял обученный наблюдатель, после чего выполняли окончательное определение значения Tm. Различающиеся двойные пики в образцах для каких-либо зондов, соответствующие Tm для дикого типа и мутантов, считались показателями гетерогенной устойчивости. Контроли, не содержащие матрицы, в которых вместо ДНК в качестве матрицы использовали стерильную воду, использовали в качестве ДНК-отрицательного контроля, и в каждый планшет для анализа включали ДНК-положительный контроль, в котором в качестве матрицы использовали 1 г геномной ДНК М. tb H37Rv.
Согласование участия субъектов-людей
Данное исследование было одобрено экспертными советами учреждений национальной больницы Масана, NIAID и университета Рутгерса (ранее UMDNJ), и все субъекты дали письменное информированное согласие (протокол ЭСУ университета Рутгерса №0120090104).
Пример 2. Определение значений Tm, ассоциированных с последовательностями дикого типа и мутантными последовательностями
Анализ на основе SMB, описанный в настоящем документе, обнаруживал устойчивость к АМК и KAN путем поиска мутаций в гене rrs и промоторе eis М. tb, заведомо ассоциированных с устойчивостью. Анализ состоял из этапа ПЦР с последующим анализом Tm в присутствии SMB-зондов, комплементарных фрагментам ампликонов-мишеней rrs и eis. Авторы изобретения вначале выполнили оценку способности анализа выявлять мутации-мишени в искусственных олигонуклеотидах и секвенированных ДНК-матрицах из выбранного штаммов М. tb дикого типа и мутантных штаммов (данные не показаны). Последовательности дикого типа выявляли по наличию значений Tm в пределах 1°С от известных средних значений для мишеней дикого типа. Мутантные последовательности выявляли по сдвигу значений Tm по меньшей мере на пять стандартных отклонений от средних значений Tm дикого типа. Затем оценивали способность анализа обнаруживать наиболее распространенные мутации, ассоциированные с устойчивостью к АМК и KAN, на клинических образцах ДНК. Анализы выполняли на панели из 603 клинических образцов, состоящей из 487 образцов с последовательностями дикого типа и 116 образцов с мутациями в мишенях анализа. Пять из этих образцов содержали смеси как ДНК дикого типа, так и мутантной ДНК, обнаруженные при секвенировании по Сэнгеру. SMB-анализ правильно определил 115/116 (99%) мутантных или смешанных образцов (гетерогенных образцов, содержащих как мутантную ДНК, так и ДНК дикого типа) как мутантные или смешанные образцы и 487/487 (100%) чистых образцов дикого типа как образцы дикого типа. Анализ, описанный в настоящем документе, определил один смешанный образец (согласно секвенированию по Сэнгеру) как образец дикого типа. Значения Tm, полученные для каждого SMB-зонда и мишеней дикого типа или мутантных мишеней, хорошо воспроизводились. Для мишеней дикого типа зонды rrs-1400, eis-1 и eis-2 демонстрировали средние значения Tm, равные 70,1±0,15°С, 63,9±0,19°С и 69±0,23°С, соответственно (Таблица 3). Мутации A1401G и С1402Т в мишенях приводили к уменьшению значений Tm для зонда rrs-1400 на 3,9°С (±0,17) и 5,6°С (±0,21), соответственно (Таблица 3). Аналогичным образом, зонды eis-1 и eis-2 зондов позволяли надежно обнаруживать ряд мутаций в области промотора eis как мутанты по снижению значений Tm на 4,3-6,5°С по сравнению с ожидаемыми значениями Tm дикого типа (Таблица 3). ПЦР-анализы, выполненные в двух разных лабораториях в университете Рутгерса и ITRC, полностью совпадали для всех образцов, определенных как образцы дикого типа и мутанты, а также смесей.
Результаты анализа позволили четко разделить 603 образца на Tm-кластеры дикого типа и мутанты на основании индивидуальных трехточечных моделей Tm (Фиг. 1). Анализ правильно определил мутации во всех 75 образцах, которые содержали только мутацию A1401G (Таблица 3, Фиг. 2, график А). Три из четырех образцов, содержащих смеси A1401G и последовательностей дикого типа, также были обнаружены как смешанные образцы дикого типа/мутанты на основании наличия двойного пика Tm. Один образец, содержавший смесь ДНК с мутацией С1402Т и ДНК дикого типа, также был обнаружен по наличию двойных пиков Tm в образце с мутантной Tm, специфической для мутации С1402Т (Таблица 3, Фиг. 2, график А). 32 образца с мутациями в области промотора eis содержали пять различных полиморфизмов (в положениях -8, -10, -12, -14 и -37). Все эти мутации успешно обнаруживались каким-либо из SMB eis (Таблица 3, Фиг. 2, графики В и С). Четыре образца, содержавшие мутации как в гене rrs, так и в области промотора eis, также были правильно определены как двойные мутанты (Таблица 3). Секвенирование гена rrs не выявило образцов с мутацией кодона 1484, независимо от картины чувствительности к лекарственным средствам.
Пример 3. Выявление устойчивости к амикацину
В данном примере выполняли анализы с целью оценки эффективности молекулярного анализа по сравнению с результатами анализа фенотипической чувствительности к лекарственным средствам. Кажущаяся эффективность анализа генотипической чувствительности к лекарственным средствам может меняться в зависимости от мутаций, выбранных для включения в анализ, и фенотипического анализа, который используется в качестве «золотого стандарта» (Kim, SJ 2005 Eur Respir J 25: 564-569; Rigouts, L., et al., 2013, J Clin Microbiol 51: 2641-2645 и Van Deun, A., et al., 2013, J Clin Microbiol 51: 2633-2640). Принимая способ анализа чувствительности к лекарственным средствам на основе LJ в качестве «золотого стандарта» (для всех 603 исследуемых образцов), Tm SMB rrs, характерные для мутации A1401G, позволили классифицировать 82/90 образцов, устойчивых к АМК, как устойчивые (чувствительность 91,1%; 95% ДИ от 82,8% до 96,8%). Tm, характерные для дикого типа, позволили классифицировать 512/513 АМК-чувствительных образцов как чувствительные. С помощью SMB-анализа, описанного в настоящем документе, один изолят среди 513 АМК-чувствительных изолятов определили как смесь ДНК дикого типа и С1402Т-мутантной ДНК из-за наличия четкого двойного пика, образованного за счет зонда rrs, соответствующего Tm дикого типа и Tm, специфической для мутанта С1402Т (Фигура 2, график А). Этот результат подтвердили с помощью секвенирования по Сэнгеру. Поскольку предыдущие исследования показали, что мутация С1402Т не кодирует устойчивости к АМК (24), специфическая Tm, соответствующая мутации С1402Т, не может считаться показателем чувствительности к АМК. Это соображение позволило сделать вывод, что анализ, описанный в настоящем документе, позволял правильно определить все 513/513 АМК-чувствительных образцов, что привело к 100% специфичности (95% ДИ от 99 до 100%). Включение значений Tm, характерных для мутаций в области промотора eis, в анализ не повысило чувствительность по отношению к обнаружению устойчивости к АМК, но снизило специфичность со 100% до 93,8% (95% ДИ от 91,2 до 95,6%). Эти результаты согласуются с предыдущими отчетами, которые показывают, что мутации промотора eis не ассоциированы с устойчивостью к АМК, согласно LJ-анализу чувствительности к лекарственным средствам (Campbell 2011 и Zaunbrecher 2009).
Пример 4. Выявление устойчивости к канамицину
Производительность анализа при обнаружении устойчивости к KAN также оценивали с использованием анализа чувствительности к лекарственным средствам на основе LJ в качестве «золотого стандарта» для всех 603 образцов. При использовании значений Tm, полученных для SMB rrs, описанных в настоящем документе, типичных для мутации A1401G или С1402Т, с целью определения устойчивости анализ определил 82/106 образцов как устойчивые к KAN (чувствительность 77,4%, 95% ДИ от 68,0 до 84,7%). И наоборот, при использовании Tm SMB rrs, характерных для мишени дикого типа, для определения чувствительности, 496/497 KAN-чувствительных образцов определили как чувствительные (Таблица 4) (специфичность 99,8%, 95% ДИ от 98,7 до 100%). Добавление значений Tm для двух SMB eis, характерных для мутаций в области промотора eis, для определения устойчивости повысило чувствительность по отношению к обнаружению устойчивости к KAN с 77,4% до 87,7% (95% ДИ от 79,5 до 93%), поскольку 11 дополнительных устойчивых к KAN образцов были классифицированы как устойчивые. Однако специфичность снизилась с 99,8% до 95,6% (95% ДИ от 93,3 до 97,1%), поскольку 21 KAN-чувствительных образцов с мутациями промотора eis после этого «ложно» определялись как устойчивые к KAN (Таблица 4).
Затем выполнили аналогичный анализ с использованием результатов анализа чувствительности к лекарственным средствам на основе MGIT в качестве «золотого стандарта» для 560 образцов, для которых был получен результат MGIT. Эта подгруппа включала все образцы, содержащие только мутации промотора eis. Сравнение результатов анализа с «золотым стандартом» на основе MGIT позволило выявить мутанты eis с несогласующимися результатами анализа устойчивости к KAN в среде LJ. При использовании MGIT в качестве «золотого стандарта» и значений Tm SMB rrs, характерных только для мутаций A1401G или С1402Т, для определения устойчивости к KAN, только 63/113 KAN-устойчивых образцов были определены как устойчивые при SMB-анализе (чувствительность 55,8%, 95% ДИ от 46,1 до 65%). И наоборот, при использовании Tm SMB rrs, характерных для мишени дикого типа, для определения чувствительности, 445/447 образцов определили как чувствительные к KAN (специфичность 99,8%; 95% ДИ от 98,5 до 100%; Таблица 4). В отличие от случая с анализом чувствительности на основе LJ, включение значений Tm, характерных для мутаций промотора eis в данном случае повысило чувствительность по отношению к анализу устойчивости с 55,8% до 82,3%, при этом специфичность анализа устойчивости к KAN оставалась очень высокой и составляла 99,5% (95% ДИ от 98,2 до 100%). Таким образом, при использовании анализа чувствительности на основе MGIT анализ eis позволил обнаружить 29 дополнительных образцов, устойчивых к KAN, без влияния на специфичность (Таблица 4).
Пример 5. Связь между мутациями, обнаруженными с помощью анализа, и MIC
Несоответствие между устойчивостью, определяемой посредством анализа, описанного в настоящем документе, и устойчивостью, определяемой посредством двух способов анализа фенотипической чувствительности, описанных в настоящем документе, в основном относится к изолятам с мутациями промотора eis. Предыдущие исследования показали, что мутации промотора eis приводят к относительно низкому уровню устойчивости к KAN, тогда как мутации гена rrs приводят к высокоуровневой устойчивости к АМК, KAN и CAP (Campbell 2011; Du, Q., et al., 2013, Diagn Microbiol Infect Dis 77: 138-142, Georghiou 2012 и Zaunbrecher 2009). Дополнительным результатом являлось расхождение между результатами анализа чувствительности на основе LJ и MGIT. Для более тщательного изучения взаимосвязи между мутациями в гене rrs и промоторе eis и дифференциальными моделями их чувствительности в системах на основе LJ и MGIT выполнили анализ MIC. Образцы, чувствительные как к АМК, так и к KAN (и содержащие обе области-мишени дикого типа) или выбранные в качестве типичных представителей наиболее распространенных типов мутаций в двух генах-мишенях (A1401G в rrs и G(-10)A, С(-14)Т и G(-37)T) в eis, анализировали способом MYCOTB с целью определения их MIC. Дополнительные изоляты, заведомо содержащих обе мишени дикого типа, также анализировали в качестве контрольных образцов. Обнаружено, что MIC АМК для изолятов, содержавших только мутации промотора eis (без мутаций rrs) находились в диапазоне от 0,25 мкг/мл до 2 мкг/мл, причем для большинства образцов MIC составляли от 0,5 мкг/мл до 1 мкг/мл (Фиг. 3). Только у одного мутанта по промотору eis MIC АМК составляла 4 мкг/мл. Контрольные изоляты без мутаций промотора eis или гена rrs характеризовались MIC от 0,25 мкг/мл до 0,5 мкг/мл. Таким образом, MIC АМК для изолятов с мутантным промотором или промотором eis дикого типа по существу перекрывались. Напротив, MIC KAN для большинства мутантов промотора eis находились в диапазоне от 5 мкг/мл до 20 мкг/мл, причем у одного изолята MIC составляла 40 мкг/мл (Фиг. 3). Только у двух мутантов по промотору eis MIC были низкими и составляли 2,5 мкг/мл. Изоляты с последовательностями промотора eis дикого типа демонстрировали MIC в диапазоне от 0,6 мкг/мл до 2,5 мкг/мл, что в 2-30 раз меньше, чем средняя MIC у мутантов по промотору eis (Фиг. 3). Таким образом, в отличие от ситуации с АМК, MIC KAN для изолятов дикого типа в очень малой степени перекрывались с MIC KAN для мутантов по промотору eis. Эти результаты ясно показывают, что следует считать, что мутанты по промотору eis обладают слабой и умеренной устойчивостью к KAN, даже если устойчивость не обнаруживается при анализах чувствительности на основе LJ или даже MGIT.
Пример 6. Специфичность анализа по отношению к бактериям, не являющимся М. tb.
Чувствительность анализа проверяли по отношению к панели из 18 видов Mycobacteria, не являющихся возбудителями туберкулеза (NTM), полученными из хранилища АТСС (Манассас, штат Виргиния, США), 121 клинических штаммов NTM, представленных 26 видами, и 18 видов грамположительных и грамотрицательных бактерий. Область rrs, представлящая собой мишень анализа, описанного в настоящем изобретении, является высококонсервативной среди различных видов NTM. Таким образом, анализ rrs позволил получить Tm, равную 70°С (что идентично Tm, получаемой в присутствии ДНК М. tb дикого типа), для всех протестированных NTM, как и ожидалось на основании гомологии последовательностей, за исключением М. xenopi, для которой не было получено значения Tm. Получение значений Tm, идентичных значениям для М. tb,, чувствительных к аминогликозидам, для видов NTM не ожидалось, за исключением ложных результатов устойчивости. При смешивании ДНК штаммов М. tb, устойчивых к АМК и KAN и мутантных по rrs, с 10-20-кратным избытком ДНК NTM анализ позволил получить четкий двойной пик Tm, соответствующий мутантному значению Tm для мишени М. tb, и значению Tm дикого типа для последовательности NTM (данные не показаны), что указывало на возможность обнаружения мутаций rrs, связанных с устойчивостью, в М. tb посредством анализа, описанного в настоящем документе, даже в присутствии большого количества фоновой ДНК NTM. Видимой кривой плавления для зондов eis в присутствии любых протестированных видов NTM не получили даже при добавлении 107 геномных эквивалентов ДНК при ПЦР-анализе. Ни для одной из грамположительных и грамотрицательных бактерий не получили значений Tm при использовании любого из SMB rrs или eis; таким образом, они не приводят к получению ложных результатов устойчивости при данном анализе.
Пример 7. Дополнительные генетические причины устойчивости к АМК и KAN
Исследование в данном примере включало 22 образца, устойчивых к АМК и/или KAN, однако содержавших последовательности гена rrs и промотора eis дикого типа. Недавние исследования показали, что мутации в 5'-нетранслируемой области (UTR) гена whiB7 могут приводить к устойчивости к аминогликозидам в М. tb (27). Чтобы определить, могут ли мутации в whiB7 обуславливать фенотипическую устойчивость при чувствительности, выявляемой посредством данного анализа, у некоторых изолятов, для всех 22 образцов секвенировали область длиной 412 и.о., расположенную выше начального сайта гена whiB7, и включающую фрагмент открытой рамки считывания whiB7. В качестве контрольного набора также секвенировали 30 случайно выбранных полностью чувствительных изолята. Шесть из 22 несогласующихся изолятов от трех пациентов продемонстрировали мутации в 5'-UTR области whiB7. Один образец содержал делецию цитозина в положении +138 в 5'-UTR, два образца от одного пациента содержали мутацию замены А на G в положении +237, а остальные три образца от одного пациента продемонстрировали мутацию замены А на С в положении +273 (Таблица 5), считая, что сайт инициации транскрипции находится в положении +1 (Reeves, A. Z., 2013, 57:1857-1865). Для трех образцов от одного пациента не удалось получить амплификацию с 5'-UTR после неоднократных попыток ПЦР, несмотря на работающий положительный контроль ПЦР. Это указывало на наличие крупной делеции в 5'-UTR области, поскольку фрагмент длиной 275 п. о. легко амплифицировался с сайтов в пределах ОРС whiB7 для всех трех образцов. Все образцы с мутациями в whiB7 были устойчивы только к KAN, что согласуется с предполагаемым механизмом действия whiB7 путем активации гена eis (Reeves 2013). MIC KAN для этих изолятов также были низкими и составляли 5 мкг/мл, что было аналогично MIC, наблюдавшимся для мутантов по промотору eis. Ни один из 30 контрольных образцов, чувствительных к аминогликозидам, не содержал мутаций в 5'-UTR гена whiB7. Необходимы дальнейшие исследования для подтверждения взаимосвязи этих мутаций и делеции в 5'-UTR гена whiB7 с устойчивостью к аминогликозидам. Однако отсутствие таких мутаций в чувствительных штаммах подразумевает, что они могут играть определенную роль в устойчивости к аминогликозидам, и эти мутации могут быть мишенями анализа в будущем для улучшения чувствительности при обнаружении слабой устойчивости к KAN.
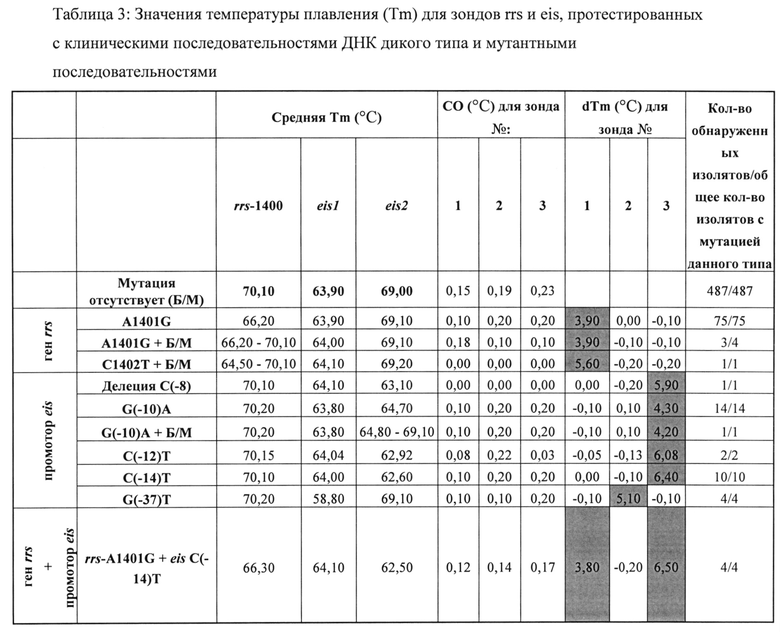
СО представляет +/- стандартное отклонение значения Tm для каждого из зондов и различных клинических образцов, a dTm представляет собой разность Tm для мутантных последовательностей и для последовательностей дикого типа при использовании каждого зонда.
СО: стандартное отклонение, dTM: дельта Tm
Зонды №1, 2 и 3 соответствуют зонам rrs-1400, eis-1 и eis-2, соответственно.
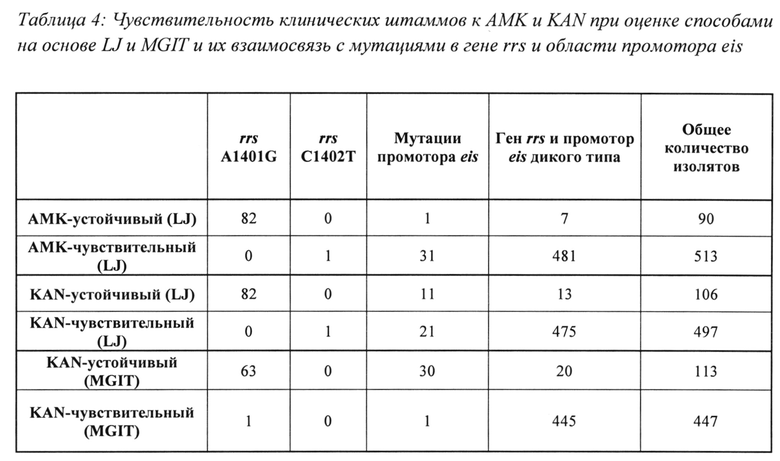
LJ и MGIT означают анализ чувствительности по долям LJ и способом MGIT, соответственно.
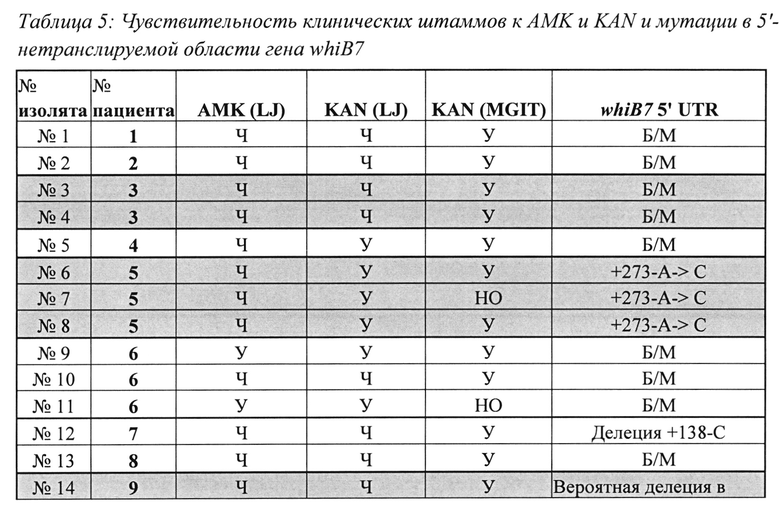
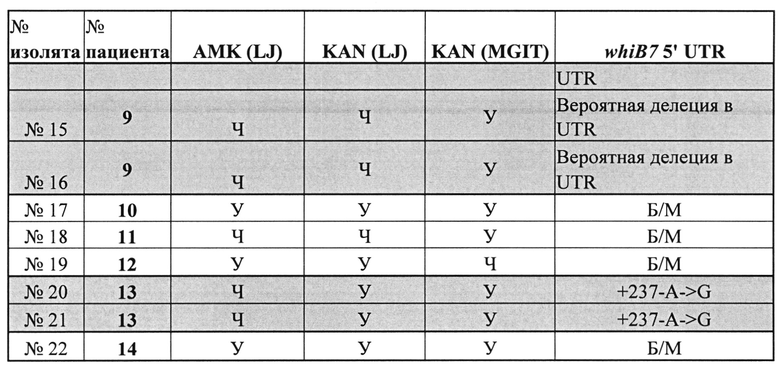
LJ и MGIT означают анализ чувствительности по долям LJ и способом MGIT, соответственно.
НО: не определяли, Б/М: без мутации, У: устойчивый, Ч; чувствительный.
Вышеупомянутые примеры и описание предпочтительных вариантов реализации следует воспринимать в качестве иллюстрации, а не в качестве ограничения настоящего изобретения, определяемого формулой изобретения. Следует принимать во внимание, что можно использовать многочисленные изменения и комбинации вышеизложенных функций без выхода за рамки настоящего изобретения, установленные формулой изобретения. Такие изменения не следует считать отходом от рамок настоящего изобретения, и все подобные изменения должны входить в рамки следующей формулы изобретения. Все ссылки, цитируемые в настоящем документе, полностью включены в настоящий документ посредством ссылки.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Alland, David
Chakravorty, Soumitesh
<120> Праймеры и зонды для полимеразной цепной реакции для обнаружения Mycobacterium Tuberculosis
<130> 096747.00312/ S2015-028
<150> US 62/062,351
<151> 10.10.2014
<160> 73
<170> Версия PatentIn 3.5
<210> 1
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 1
ccggtcggtt gccgagacc 19
<210> 2
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 2
cggtcggttg ccgagaccat gg 22
<210> 3
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 3
gtcggttgcc gagaccatgg gc 22
<210> 4
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 4
agcgggtagc gcagcgacca g 21
<210> 5
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 5
cgggtagcgc agcgaccagg gc 22
<210> 6
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 6
ccagcgggta gcgcagcgac cag 23
<210> 7
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 7
gctggagcag atgggcttgg 20
<210> 8
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 8
ccgctggagc agatgggctt gg 22
<210> 9
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 9
ggctggaaga gctcgtatgg caccg 25
<210> 10
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 10
gtcccatttc gtcggggtgt tcgtcc 26
<210> 11
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 11
ccatttcgtc ggggtgttcg tccatac 27
<210> 12
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 12
cgtcgcggac ctccagcccg gca 23
<210> 13
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 13
tcacgtgaca gaccgccggg c 21
<210> 14
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 14
gctcacgtga cagaccgccg ggc 23
<210> 15
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 15
gctagtaatc gcagatcagc aacgctgc 28
<210> 16
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 16
cctcccgagg gttaggccac tgg 23
<210> 17
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 17
ggttaggcca ctggcttcgg 20
<210> 18
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 18
cacagggtca cagtcacaga atc 23
<210> 19
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 19
cgtcctcggt cgggctacac agg 23
<210> 20
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 20
cggtcgggct acacagggtc acagt 25
<210> 21
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 21
cacagggtca cagtcacaga atc 23
<210> 22
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 22
gcatcgcgtg atcctttgcc agaca 25
<210> 23
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 23
ctcgccggtg ccagcggata tgtc 24
<210> 24
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 24
cgaccgtcca gcgcccacat cttt 24
<210> 25
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 25
ccgcgagggc cccgatggtt t 21
<210> 26
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 26
ggctgggctc ccggttgatg tgg 23
<210> 27
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 27
ggctcccggt tgatgtggtc gtag 24
<210> 28
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 28
tggggcgatc ggcacaccca gc 22
<210> 29
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 29
atcggtggat tgcccggcaa gctg 24
<210> 30
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 30
cgttccacgg atccgcgcaa gtc 23
<210> 31
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 31
gctggccgat tgccgttcca cg 22
<210> 32
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 32
gatcatcaat gtggagaaag cgc 23
<210> 33
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 33
ctggaacatc gaatcgcgac cgctt 25
<210> 34
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 34
atcgcgaccg ctttttgcag aa 22
<210> 35
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 35
ctgaccgacg ccgtggtgat a 21
<210> 36
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 36
ggaaatagtt ggacatgtag ccggcgt 27
<210> 37
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 37
cgattcgatg ttccaggcga tactt 25
<210> 38
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 38
gcgcggcaag atcatcaatg tggag 25
<210> 39
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 39
gatcatcaat gtggagaaag cgc 23
<210> 40
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 40
gtggatcccg gtgcccagcg cc 22
<210> 41
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 41
ggtgcccagc gccgtgatga tc 22
<210> 42
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 42
cgttacgctc gtggacatac cgatttcg 28
<210> 43
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 43
ttacgctcgt ggacataccg atttcggc 28
<210> 44
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 44
ggactgaacg ggatacgaat gggg 24
<210> 45
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 45
gtttggcccc ttcagtggct gtgg 24
<210> 46
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 46
cagcccgacg ccgaatcagt tgtt 24
<210> 47
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 47
ggtgcgggcg gtgtcgaggt g 21
<210> 48
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 48
gccaccgcgg ggagtttgtc g 21
<210> 49
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 49
atcaacatcc ggccggtggt cgcc 24
<210> 50
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 50
agctccagcc cggcacgctc acgt 24
<210> 51
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 51
gctagtaatc gcagatcagc aacgctgc 28
<210> 52
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 52
cctcccgagg gttaggccac t 21
<210> 53
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 53
gcatcgcgtg atcctttgcc agac 24
<210> 54
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 54
cctgcgcgca ccagggtgcc ctagatcgac gcgtcgcagg 40
<210> 55
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 55
ccaggggtgu cgtagatcga cgcgtcgccg cgccctgg 38
<210> 56
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 56
ccggcgacat caatggtgct ggtgatcgcg tccgccgg 38
<210> 57
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 57
cgcggccgac agttggcgct tgtgggttaa ccccgacgcc gcg 43
<210> 58
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 58
cgcgcgccgg gccccagcac caacagtcgg agcttcgcgc g 41
<210> 59
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 59
cacgaccgcc cgtcacgtca tgaaagtcgg tcgtg 35
<210> 60
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 60
agcggtcgta atattcacgt gcacctggcc gcggccgct 39
<210> 61
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 61
ctcgcggcat atgccacagt cggattctct gacgcgag 38
<210> 62
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 62
caggcggtcg taatattcac gtgcacctgg ccgccgcctg 40
<210> 63
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 63
cgagcgtatg tagtagaagg tgactcggcc ggcgctcg 38
<210> 64
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 64
acctgccgcg gcgagacgat aggutgtagg ggtgacggca ggt 43
<210> 65
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 65
ccgagctgat cguctgaact tcggcgtuct ttagcacccg gtugatagct cgg 53
<210> 66
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 66
caccggcgac tcgggccacg tccaggatgt agccggtg 38
<210> 67
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 67
cgcgcaccaa tatgatcggg tactcgactg cgcg 34
<210> 68
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 68
tcggccatca agggaatgga gttggcgcgc ggccga 36
<210> 69
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 69
ccgcgtgggt gtcgagtcga tctgcacaca gctacgcgg 39
<210> 70
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 70
aaacgcgcag gtcagaaaat 20
<210> 71
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 71
cagtgtcttg gctacctcga 20
<210> 72
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 72
gtcggtactg acagtcccc 19
<210> 73
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезированный олигонуклеотид
<400> 73
atgcaacagc atccttgcg 19
<---
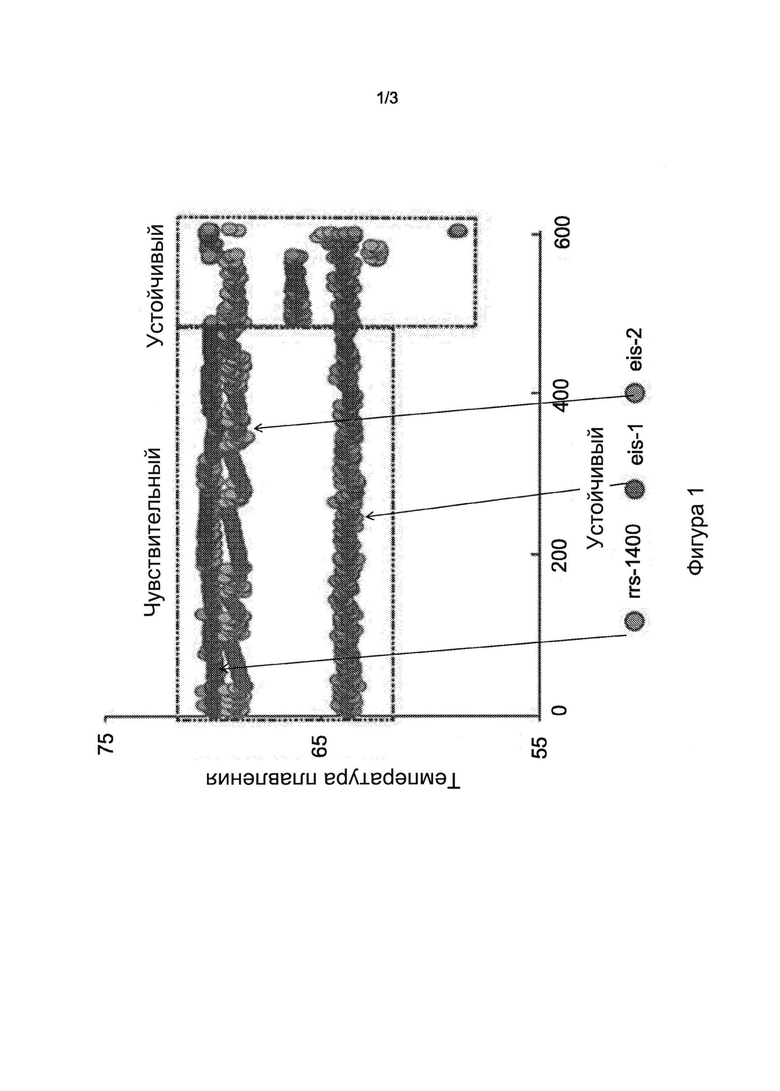
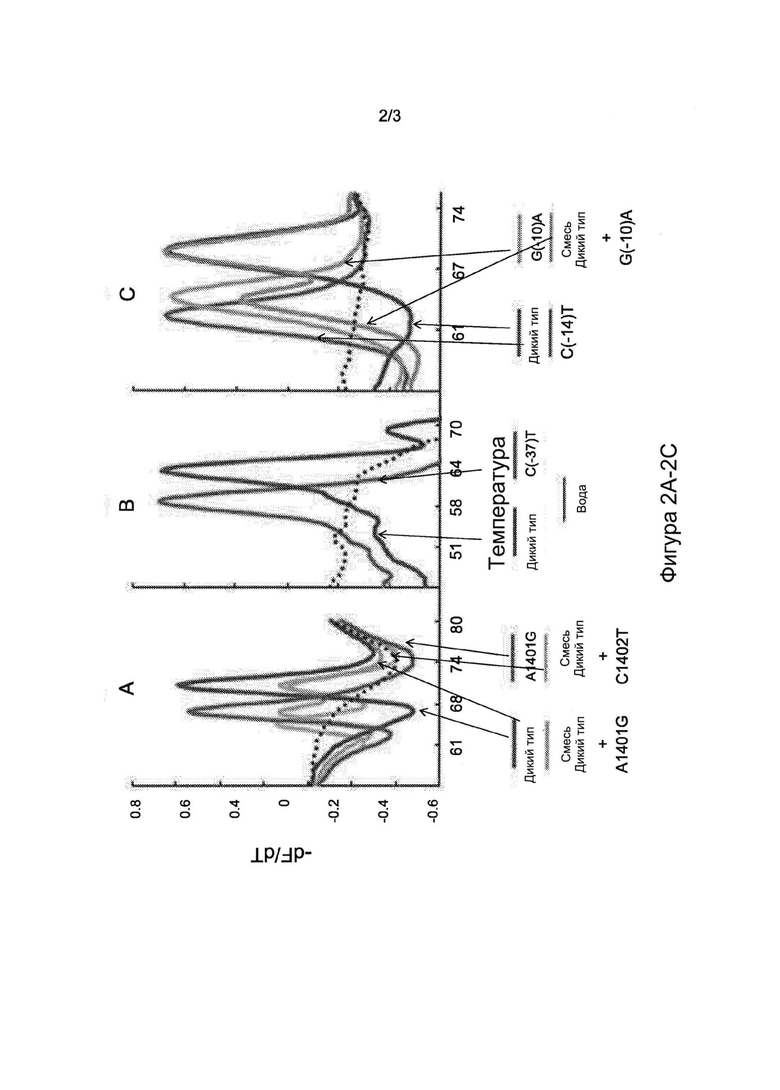
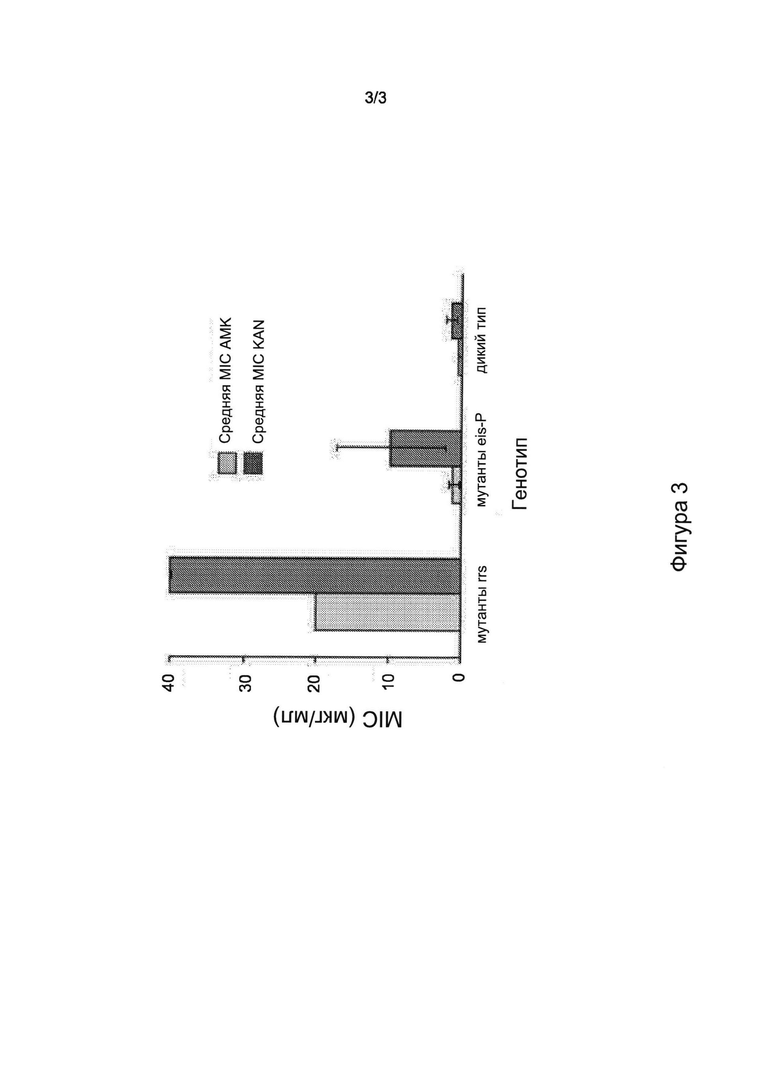
Изобретение относится к области биотехнологии. Предложен зонд, состоящий из выделенной нуклеиновой кислоты, для обнаружения M. tuberculosis, обладающей лекарственной устойчивостью, содержащий последовательность зонда, которая по меньшей мере на 85% идентична последовательности, выбранной из перечисленных в Таблице 2, а также соответствующие набор и способ обнаружения M. tuberculosis, обладающей лекарственной устойчивостью. Изобретение может быть использовано в медицине. 3 н. и 18 з.п. ф-лы, 3 ил., 5 табл., 7 пр.
1. Зонд, состоящий из выделенной нуклеиновой кислоты, для обнаружения M. tuberculosis, обладающей лекарственной устойчивостью, содержащий последовательность зонда, которая по меньшей мере на 85% идентична последовательности, выбранной из перечисленных в Таблице 2.
2. Зонд, состоящий из нуклеиновой кислоты, по п. 1, отличающийся тем, что последовательность зонда выбрана из последовательностей, перечисленных в Таблице 2.
3. Зонд, состоящий из нуклеиновой кислоты, по п. 1 или 2, отличающийся тем, что указанный зонд, состоящий из нуклеиновой кислоты, является меченым.
4. Зонд, состоящий из нуклеиновой кислоты, по п. 3, отличающийся тем, что указанный зонд, состоящий из нуклеиновой кислоты, мечен флуорофором и гасителем по своим двум концам, соответственно.
5. Зонд, состоящий из нуклеиновой кислоты, по п. 4, отличающийся тем, что указанный флуорофор представляет собой флуоресцеин, цианин 5, TexasRed или TAMRA.
6. Зонд, состоящий из нуклеиновой кислоты, по п. 4 или 5, отличающийся тем, что указанный гаситель представляет собой BHQ1, BHQ2 или DABCYL.
7. Набор для обнаружения M. tuberculosis, обладающей лекарственной устойчивостью, содержащий (i) зонд, состоящий из нуклеиновой кислоты, по любому из пп. 1-6 и (ii) один или более компонент, выбранный из группы, состоящей из набора праймеров, ДНК-полимеразы, нуклеотидов для удлинения последовательности и буфера.
8. Набор по п. 7, отличающийся тем, что указанный набор содержит указанную ДНК-полимеразу, указанные нуклеотиды для удлинения последовательности и указанный буфер.
9. Набор по п. 7 или 8, отличающийся тем, что указанный набор праймеров содержит первую пару прямого и обратного праймеров, выбранных из праймеров, которые перечислены в Таблицах 1А и 1В.
10. Набор по п. 9, отличающийся тем, что указанный набор праймеров дополнительно содержит вторую пару прямого и обратного праймеров, выбранных из праймеров, которые перечислены в Таблицах 1А и 1В.
11. Способ обнаружения M. tuberculosis, обладающей лекарственной устойчивостью, включающий амплификацию первой нуклеотидной последовательности-мишени с использованием первой пары прямого и обратного праймеров с получением первого ампликона, причем указанная первая пара праймеров специфична по отношению к фрагменту первой области, выбранной из группы, состоящей из гена rpoB, гена gyrA, гена gyrB, промотора inhA, гена rrs, промотора eis, гена embB и гена katG, и обнаружение мутации в указанном первом ампликоне при помощи зонда, состоящего из нуклеиновой кислоты, по любому из пп. 1-6, причем наличие мутации указывает на лекарственную устойчивость.
12. Способ по п. 11, отличающийся тем, что каждый из указанных праймеров обладает последовательностью, выбранной из перечисленных в Таблице 1А и Таблице 1В последовательностей.
13. Способ по п. 11, отличающийся тем, что указанный этап обнаружения осуществляют посредством способа, включающего:
приведение в контакт указанного первого ампликона с указанным первым зондом по любому из пп. 1-6, специфичным по отношению к указанной мутации в условиях, благоприятствующих гибридизации, с образованием гибрида зонд-мишень;
выполнение анализа температуры плавления (Tm) с целью определения тестового значения Tm для указанного гибрида зонд-мишень; и
сравнение указанного тестового значения Tm с заранее определенным референсным значением Tm,
при этом тестовое значение Tm, отличающееся от указанного заранее определенного значения Tm, указывает на наличие мутации.
14. Способ по п. 13, отличающийся тем, что указанное тестовое значение Tm, которое ниже, чем указанное заранее определенное референсное значение Tm, указывает на наличие мутации.
15. Способ по любому из пп. 11-14, дополнительно включающий амплификацию второй нуклеотидной последовательности-мишени с использованием второй пары прямого и обратного праймеров с получением второго ампликона, причем указанная вторая пара праймеров специфична по отношению к фрагменту второй области, выбранной из группы, состоящей из гена rpoB, гена gyrA, гена gyrB, промотора inhA, гена rrs, промотора eis, гена embB и гена katG.
16. Способ по любому из пп. 11-15, отличающийся тем, что указанная первая область представляет собой ген katG, ген rrs или промотор eis.
17. Способ по п. 16, отличающийся тем, что указанная первая область представляет собой ген katG или ген rrs.
18. Способ по любому из пп. 15-17, отличающийся тем, что указанная вторая область представляет собой промотор eis.
19. Способ по любому из пп. 11-17, отличающийся тем, что указанная мутация представляет собой мутацию A1401G или C1402T в гене rrs.
20. Способ по п. 18, отличающийся тем, что указанная мутация находится в пределах области промотора eis, на исследование которой направлены праймеры, используемые для амплификации области промотора eis.
21. Способ по любому из пп. 11-20, отличающийся тем, что указанная устойчивость представляет собой устойчивость к лекарственному средству, выбранному из группы, состоящей из изониазида, рифампицина, лекарственных средств класса фторхинолонов, амикацина, канамицина, капреомицина и этамбутола.
| SOUMITESH CHAKRAVORTY | |||
| Rapid Detection of Fluoroquinolone-Resistant and Heteroresistant Mycobacterium tuberculosis by Use of Sloppy Molecular Beacons and Dual Melting-Temperature Codes in a Real-Time PCR Assay | |||
| J Clin Microbiol | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| WO 2013155189 A1, 17.10.2013 | |||
| Счетно-решающее устройство на матричных сетках | 1959 |
|
SU127074A1 |

