Родственные заявки
Настоящая заявка является частичным продолжением заявки на патент США №08/956323, поданной 23 октября 1998 г., заявки на патент США №08/956564, поданной 23 октября 1998 г., и заявки на патент США №08/956324, поданной 23 октября 1998 г., которые полностью включены в это описание изобретения в качестве ссылки.
Предпосылки изобретения
Сериновые протеазы входят в подгруппу карбонильных гидролаз. Они образуют класс разнообразных ферментов, обладающих различными специфичностями и биологическими функциями. Stroud R.Sci.Amer., 131:74-88. Несмотря на функциональное разнообразие, каталитический механизм сериновых протеаз присущ, по крайней мере, двум генетически различным семействам ферментов: 1) субтилизинам и 2) родственным химотрипсину человека и гомологичным бактериальным сериновым протеазам (таким как, трипсин и трипсин S.gresius). Эти два семейства сериновых протеаз обладают чрезвычайно похожими механизмами катализа. Kraut J., (1977), Annu.Rev.Biochem., 46:331-358. Кроме того, хотя первичные структуры ферментов этих двух семейств не являются родственными, их третичные структуры имеют консервативную каталитическую триаду аминокислот, состоящую из серина, гистидина и аспартата.
Субтилизины являются сериновыми протеазами (примерная молекулярная масса 27500), которые в больших количествах секретируются различными видами Bacillus и другими микроорганизмами. Белковая последовательность субтилизина определена, по крайней мере, у девяти разных видов Bacillus. Markland. F.S., et al. (1983), Hoppe-Seyler’s Z.Physiol. Chem., 364:1537-1540. В научной литературе описана трехмерная кристаллографическая структура субтилизинов, выделенных из Bacillus amyloliquefaciens, Bacillus licheniformis и нескольких природных вариантов B.lentus. Эти исследования показывают, что, хотя у субтилизина нет генетического родства с сериновыми протеазами млекопитающих, он имеет сходную структуру активного сайта. Рентгеновские кристаллические структуры субтилизина, содержащие ковалентно связанные ингибиторы пептида (Robertus J.D., et al. (1972), Biochemistry, 11:2439-2449) или комплексы продуктов (Robertus J.D., et al. (1976), J.Biol.Chem., 251:1097-1103), также позволили получить информацию об активном сайте и предполагаемом субстратсвязывающем участке субтилизина. Помимо этого, было проведено большое количество исследований кинетических и химических модификаций субтилизина; Svendsen В. (1976), Carlsberg Res. Commun., 41:237-291; Markland F.S., там же), при этом, по крайней мере, в одной научной работе описывается превращение боковой цепи метионина в положении остатка 222 субтилизина в метионинсульфоксид под действием перекиси водорода (Stauffer D.C., et al. (1965), J.Biol.Chem., 244:5333-5338) и обширный сайтспецифический мутагенез (Wells and Estell (1988) TIBS 13:291-297).
Краткое изложение существа изобретения
Объектом этого изобретения является вариант протеазы с замещением аминокислоты в положениях одного или нескольких остатков, соответствующих положениям остатков, выбираемым из группы, включающей положения 62, 212, 230, 232, 252 и 257 субтилизина Bacillus amyloliquefaciens.
Хотя может иметь место любая комбинация вышеуказанных замещений аминокислот, у предпочтительных ферментов варианта протеазы по настоящему изобретению могут быть замещены аминокислотные остатки в следующих комбинациях положений. Все указанные положения остатков соответствуют положениям субтилизина Bacillus amyloliquefaciens:
(1) вариант протеазы с замещениями аминокислотных остатков в положении 62 и в одном или нескольких следующих положениях: 103, 104, 109, 159, 213, 232, 236, 245, 248 и 252;
(2) вариант протеазы с замещениями аминокислотных остатков в положении 212 и в одном или нескольких следующих положениях: 12, 98, 102, 103, 104, 159, 232, 236, 245, 248 и 252;
(3) вариант протеазы с замещениями аминокислотных остатков в положении 230 и в одном или нескольких следующих положениях: 68, 103, 104, 159, 232, 236 и 245;
(4) вариант протеазы с замещениями аминокислотных остатков в положении 232 и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 76, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 205, 209, 210, 212, 213, 217, 230, 236, 245, 248, 252, 257, 260, 270 и 275;
(5) вариант протеазы с замещениями аминокислотных остатков в положении 232 и в одном или нескольких следующих положениях: 103, 104, 236 и 245;
(6) вариант протеазы с замещениями аминокислотных остатков в положении 232 и 103, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 76, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 205, 209, 210, 212, 213, 217, 230, 236, 245, 248, 252, 257, 260, 270 и 275;
(7) вариант протеазы с замещениями аминокислотных остатков в положении 232 и 104, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 76, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 205, 209, 210, 212, 213, 217, 230, 236, 245, 248, 252, 257, 260, 270 и 275;
(8) вариант протеазы с замещениями аминокислотных остатков в положении 232 и 236, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 76, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 205, 209, 210, 212, 213, 217, 230, 236, 245, 248, 252, 257, 260, 270 и 275;
(9) вариант протеазы с замещениями аминокислотных остатков в положении 232 и 245, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 76, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 205, 209, 210, 212, 213, 217, 230, 236, 245, 248, 252, 257, 260, 270 и 275;
(10) вариант протеазы с замещениями аминокислотных остатков в положении 232, 103, 104, 236 и 245, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 76, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 205, 209, 210, 212, 213, 217, 230, 236, 245, 248, 252, 257, 260, 270 и 275;
(11) вариант протеазы с замещениями аминокислотных остатков в положении 252 и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 210, 212, 213, 217, 232, 236, 245, 248 и 270;
(12) вариант протеазы с замещениями аминокислотных остатков в положении 252 и в одном или нескольких следующих положениях: 103, 104, 236 и 245;
(13) вариант протеазы с замещениями аминокислотных остатков в положениях 252 и 103, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 210, 212, 213, 217, 232, 236, 245, 248 и 270;
(14) вариант протеазы с замещениями аминокислотных остатков в положениях 252 и 104, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 210, 212, 213, 217, 232, 236, 245, 248 и 270;
(15) вариант протеазы с замещениями аминокислотных остатков в положениях 252 и 236, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 210, 212, 213, 217, 232, 236, 245, 248 и 270;
(16) вариант протеазы с замещениями аминокислотных остатков в положениях 252 и 245, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 210, 212, 213, 217, 232, 236, 245, 248 и 270;
(17) вариант протеазы с замещениями аминокислотных остатков в положениях 252, 103, 104, 236 и 245, и в одном или нескольких следующих положениях: 1, 9, 12, 61, 62, 68, 97, 98, 101, 102, 103, 104, 109, 130, 131, 159, 183, 185, 210, 212, 213, 217, 232, 236, 245, 248 и 270; и
(18) вариант протеазы с замещениями аминокислотных остатков в положении 257 и в одном или нескольких следующих положениях 68, 103, 104, 205, 209, 210, 232, 236, 245 и 275.
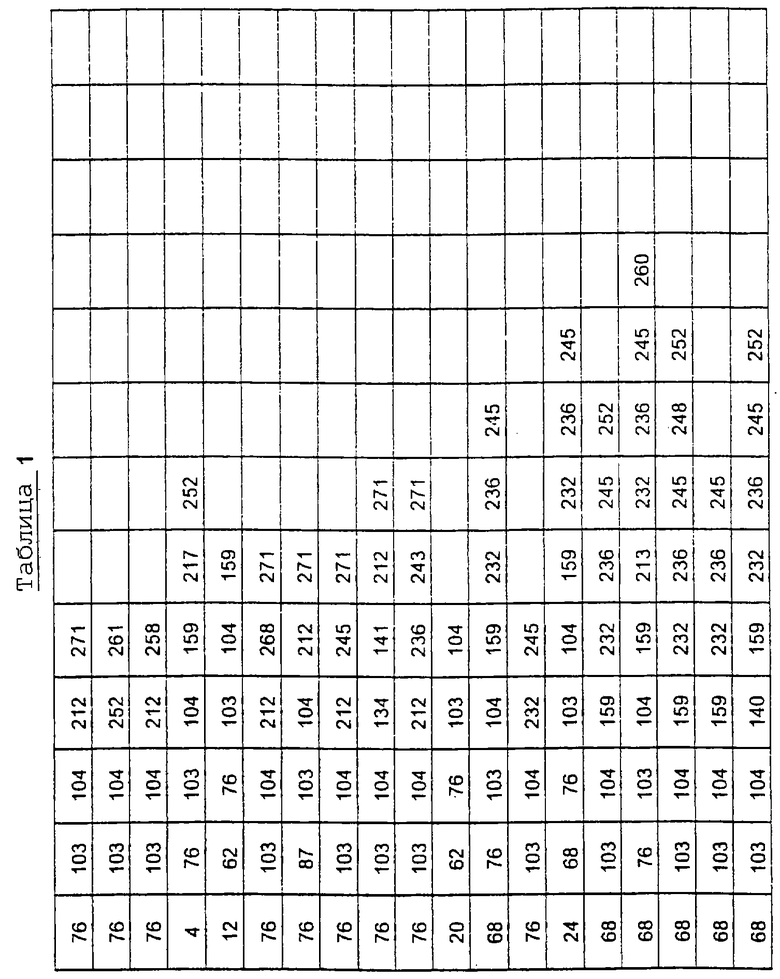
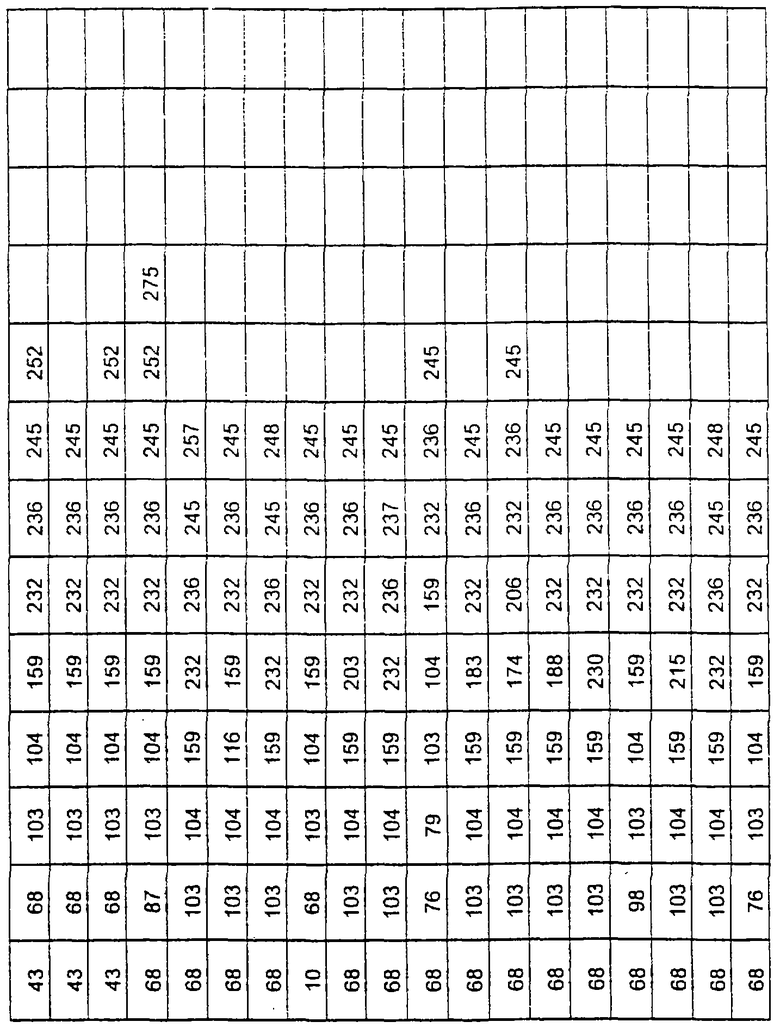
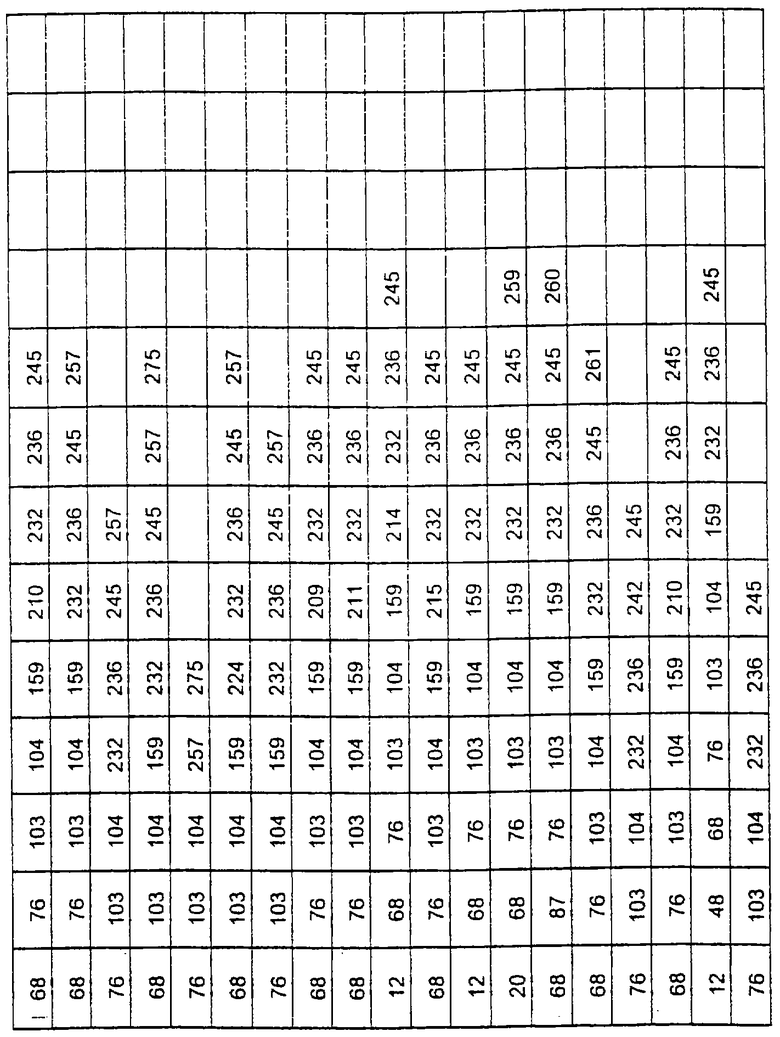
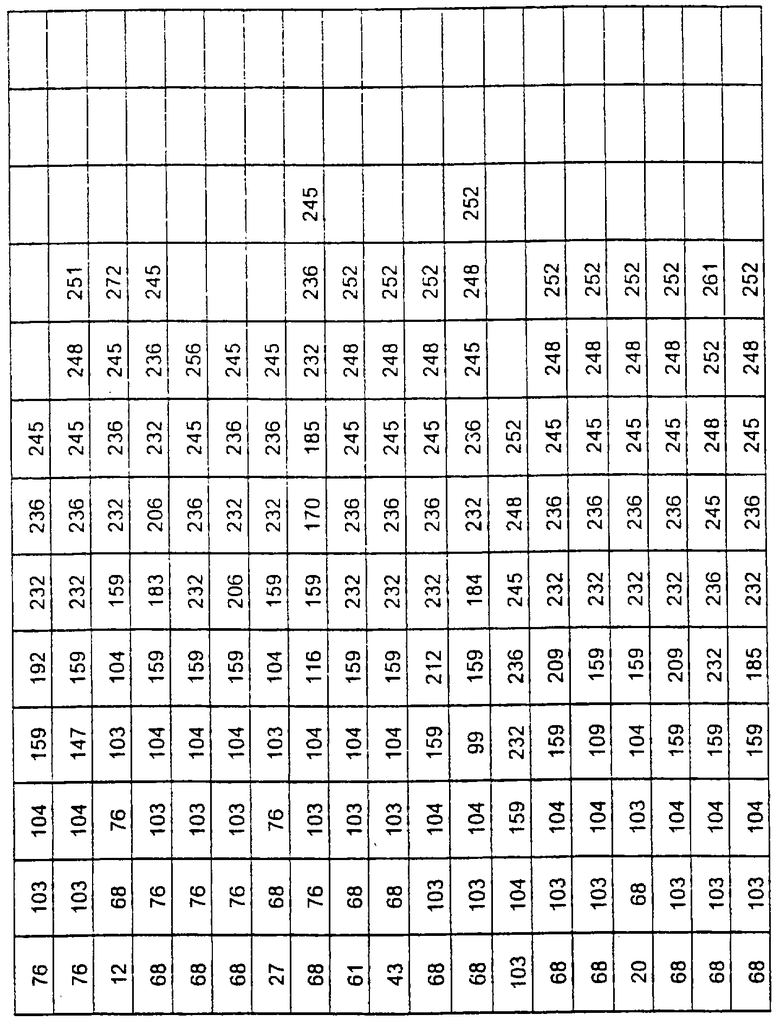
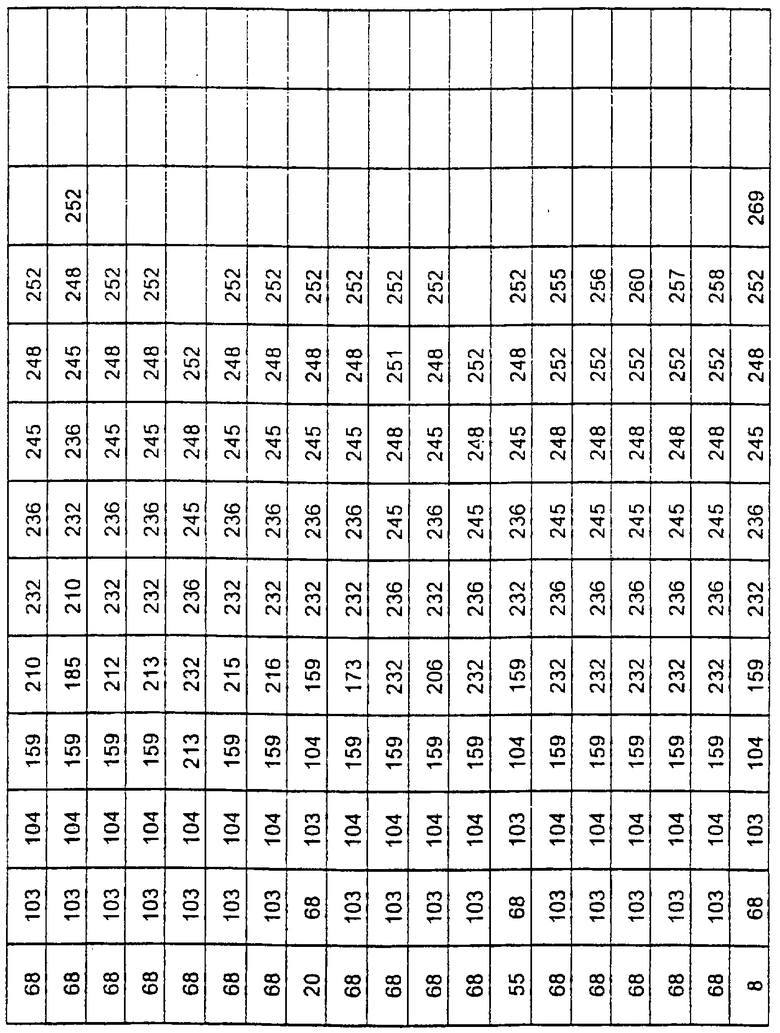
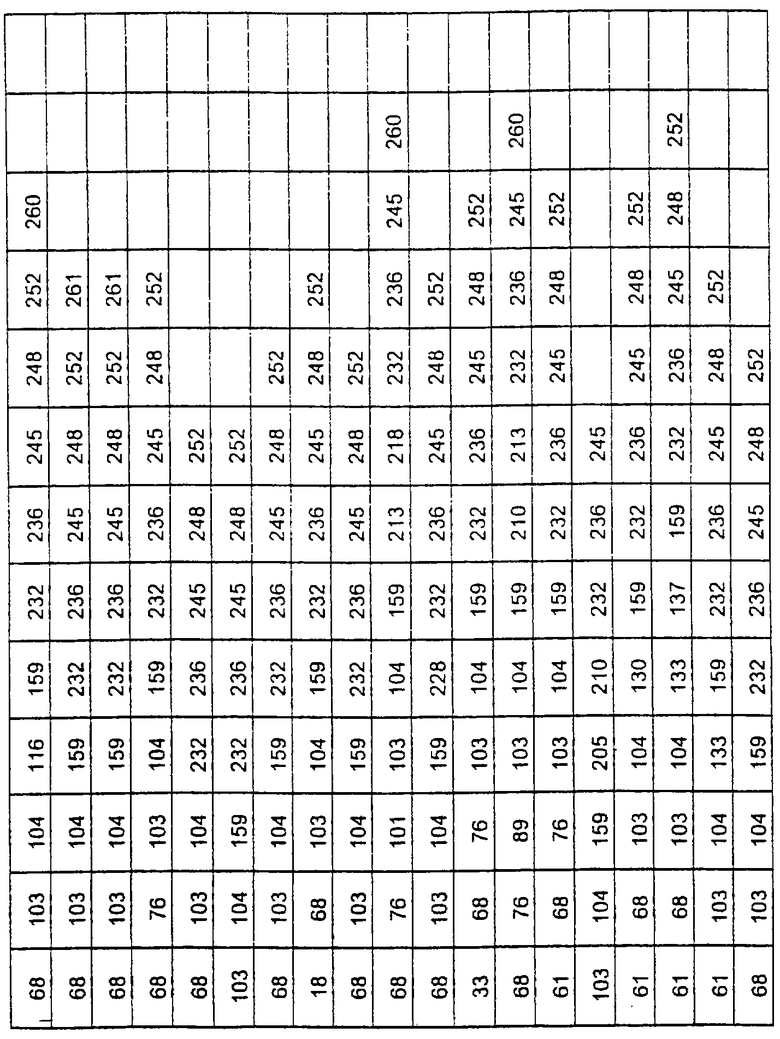
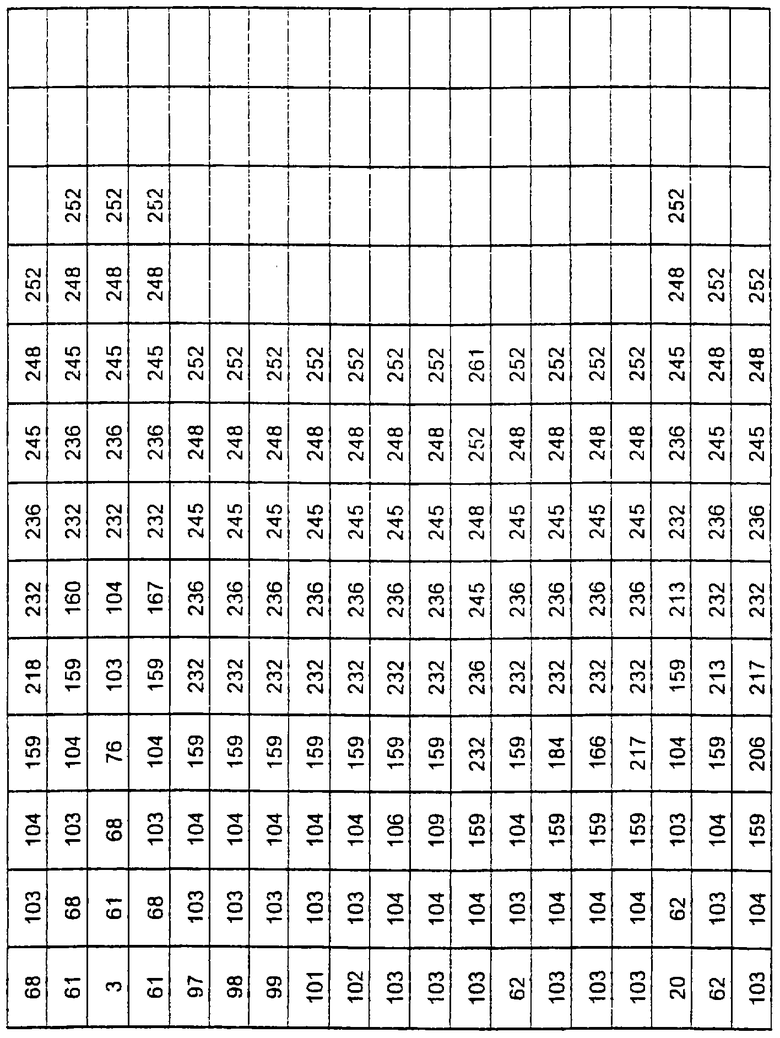
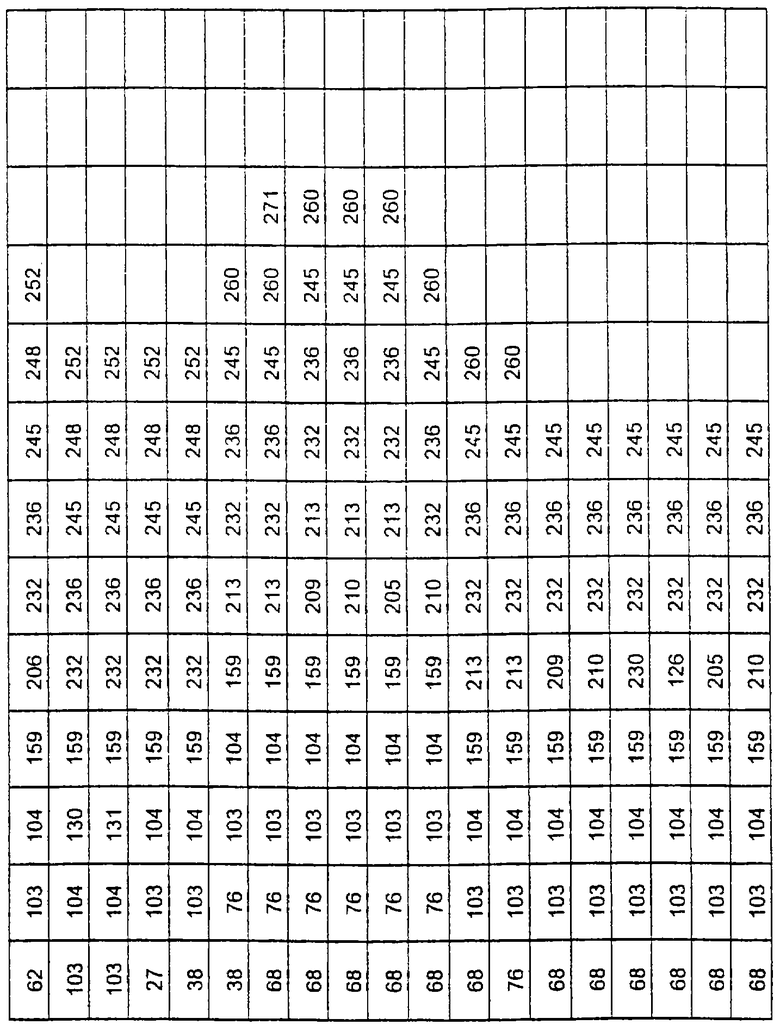
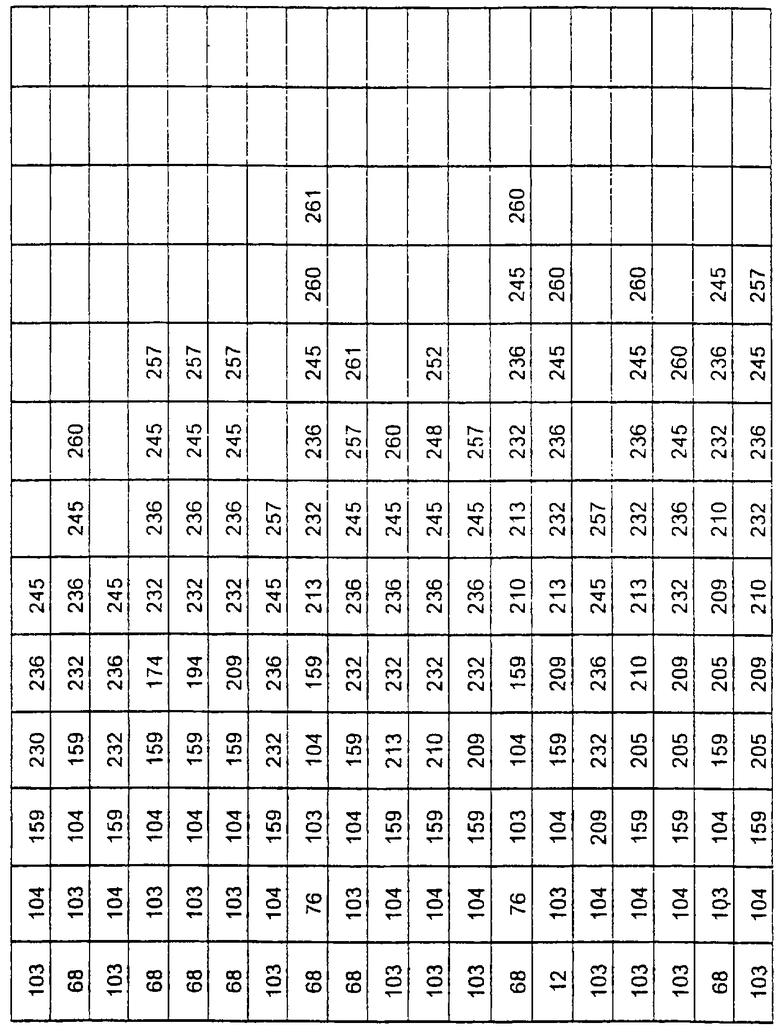
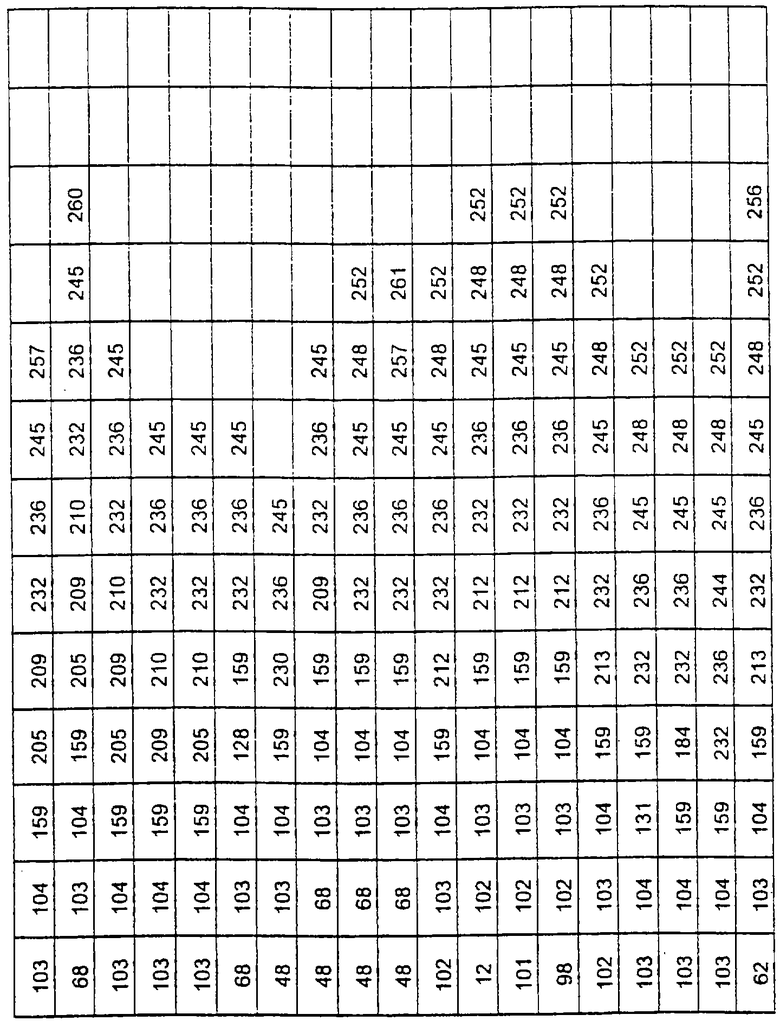
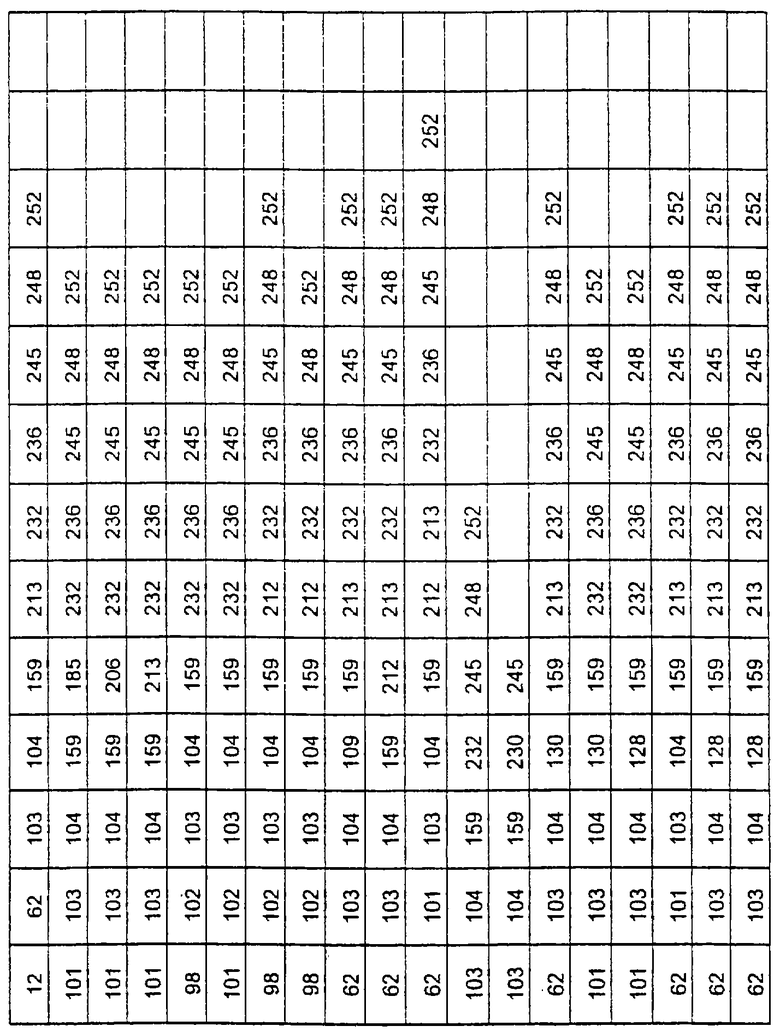
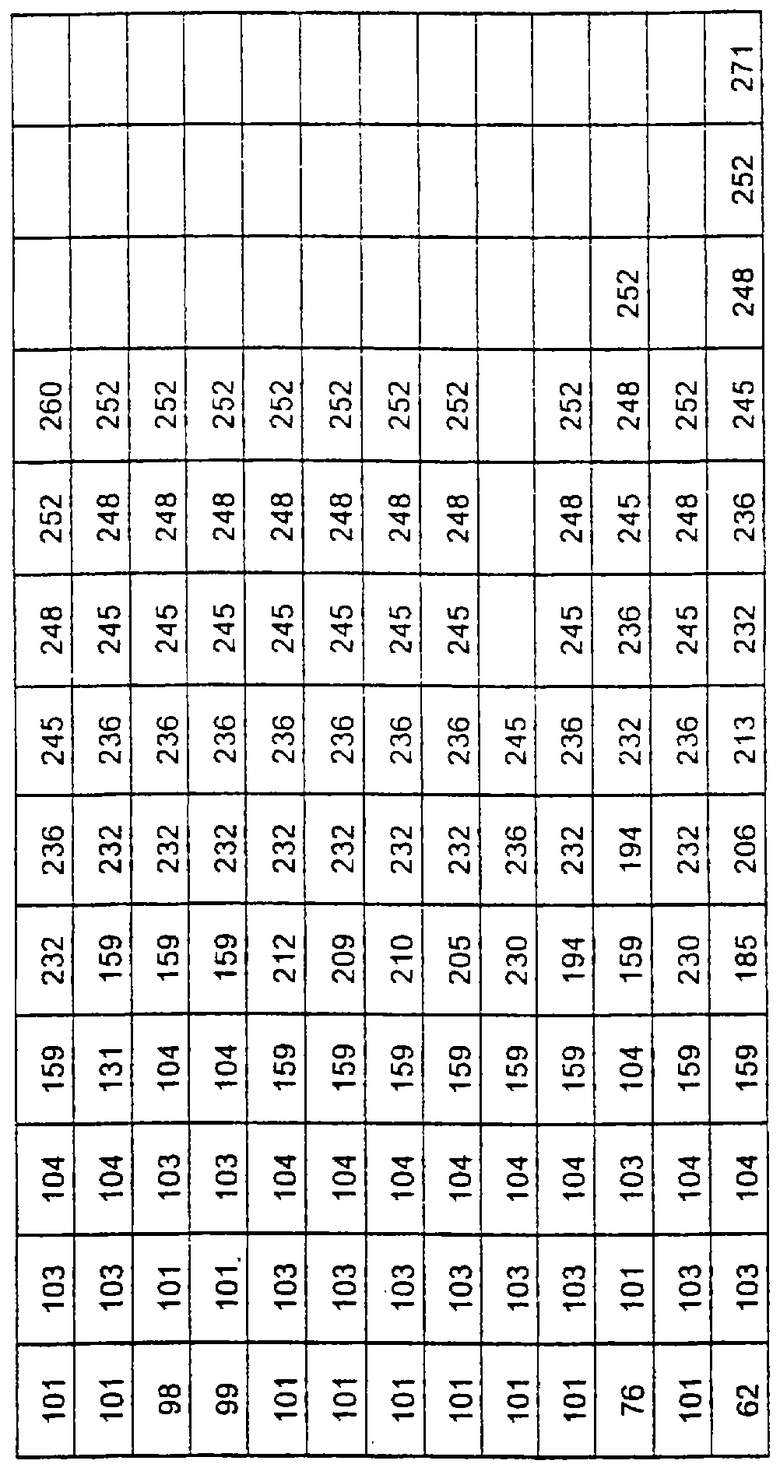
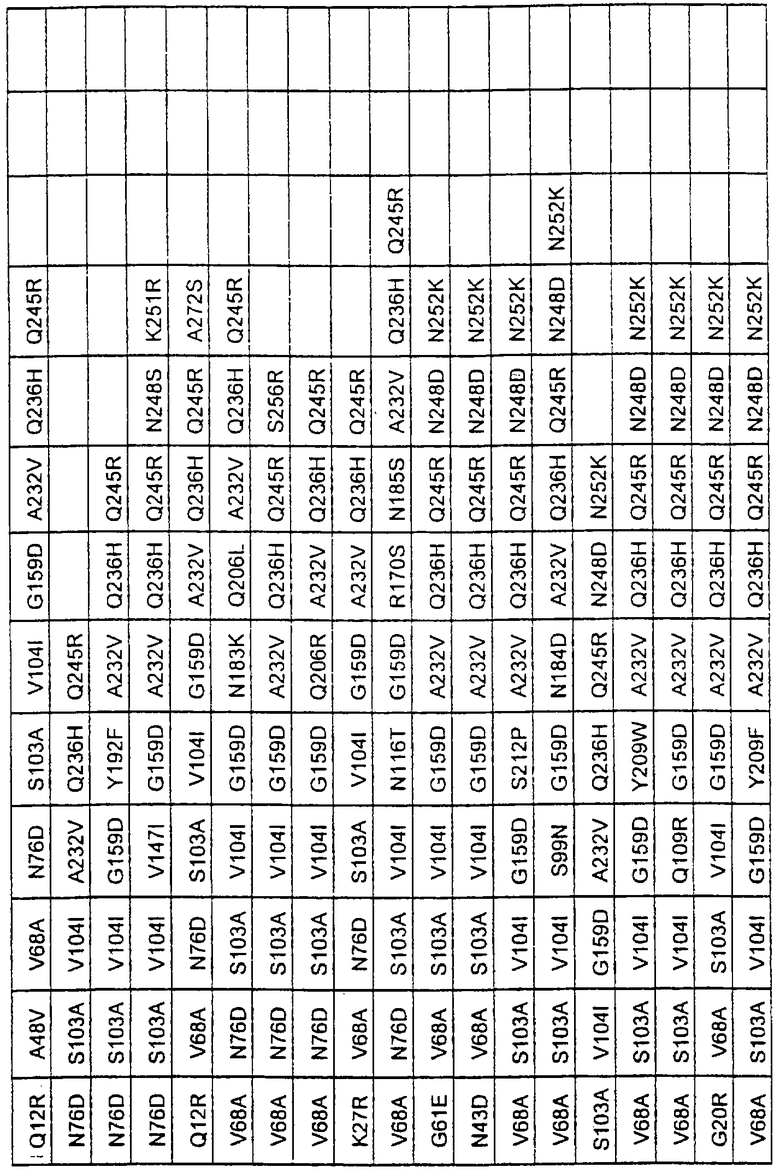
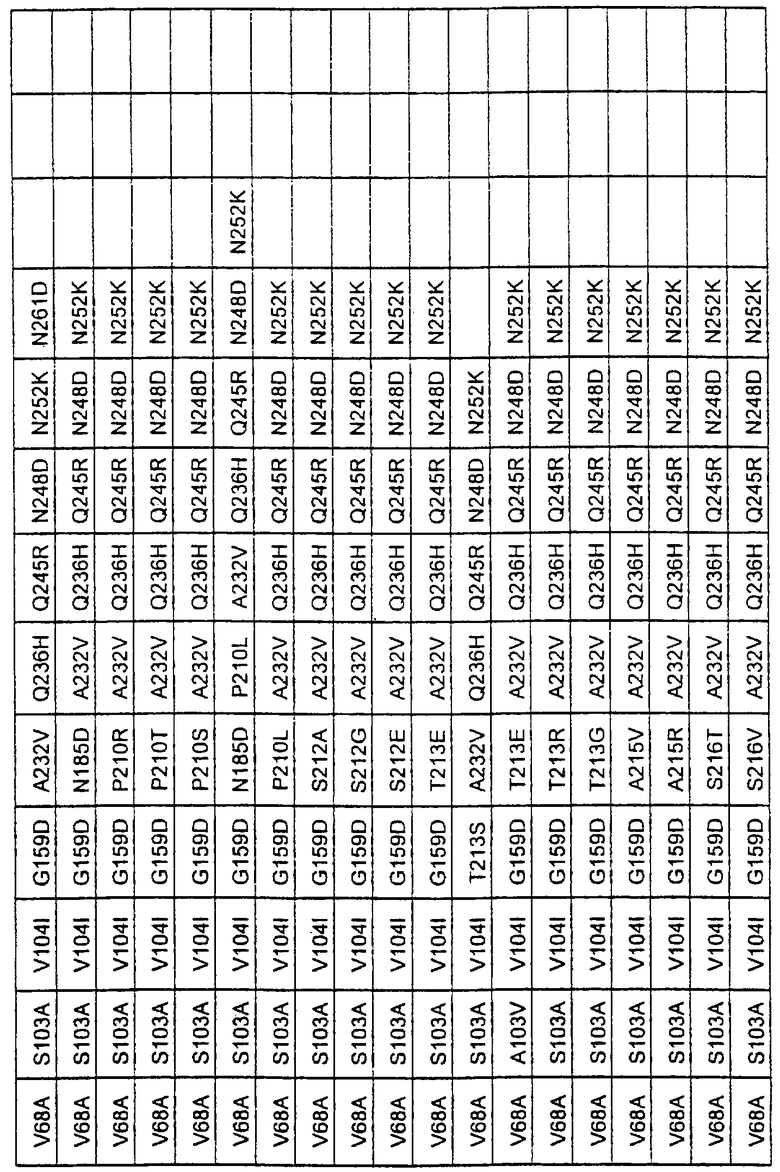
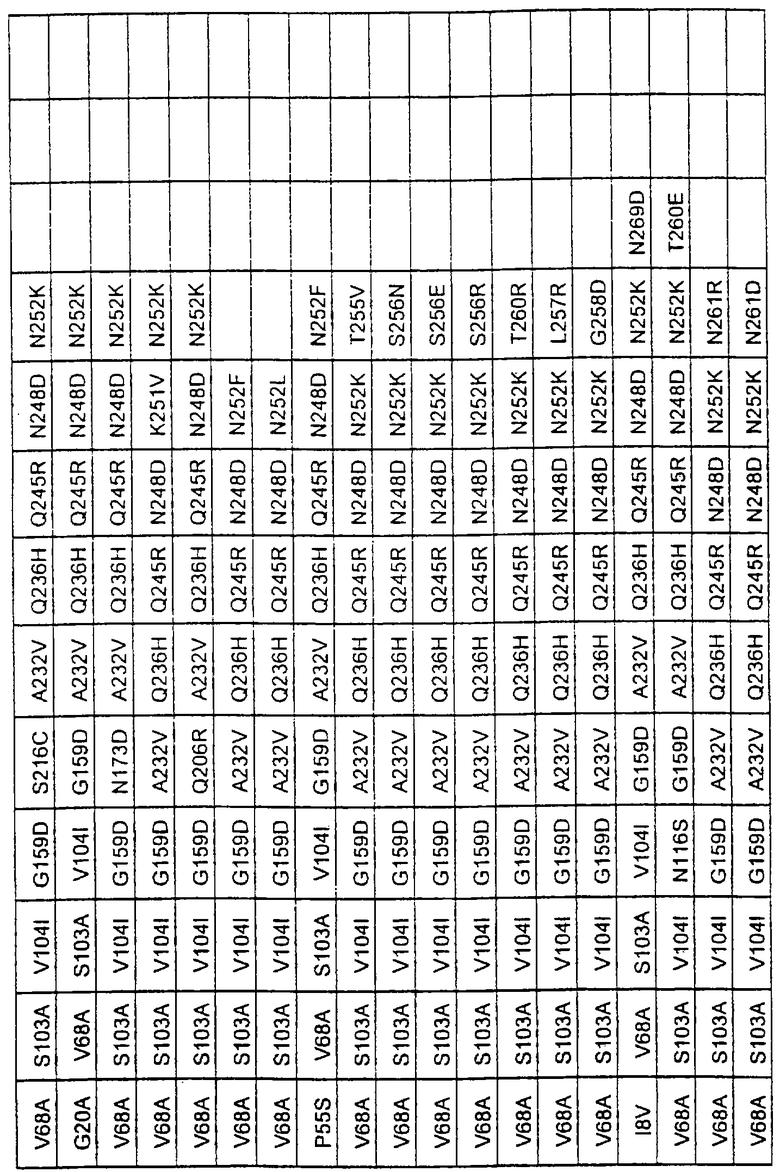
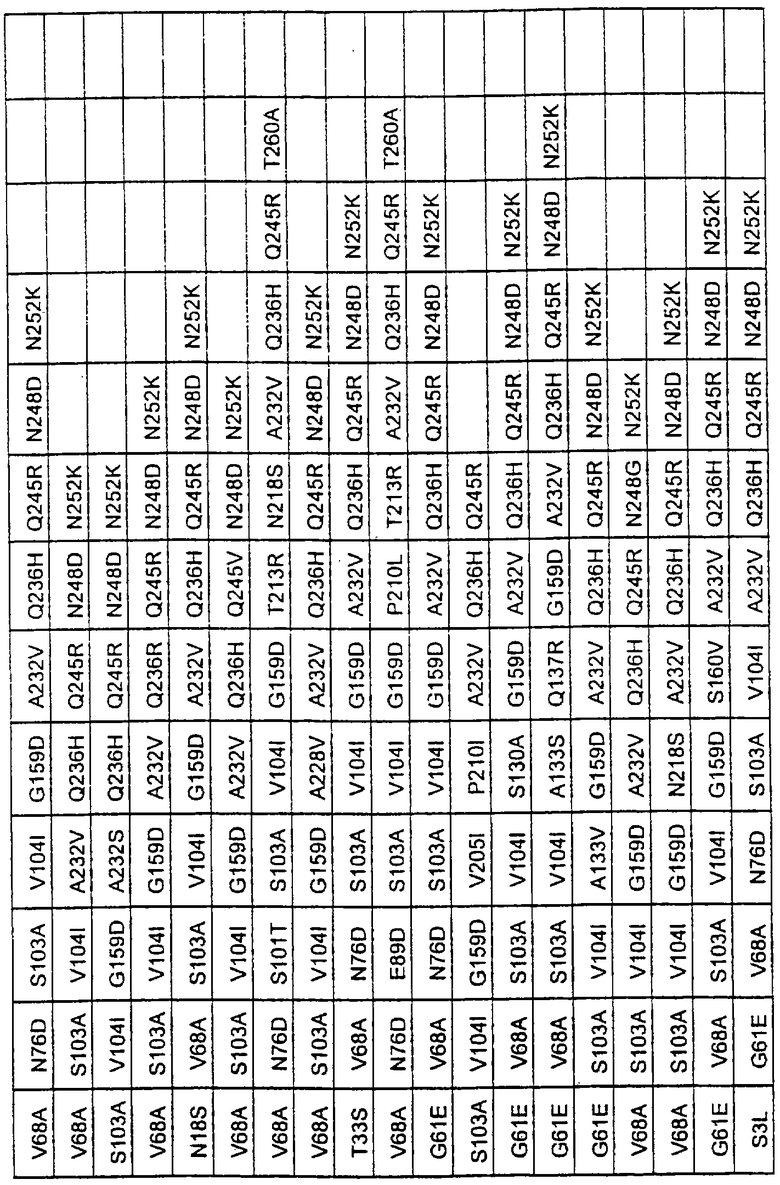
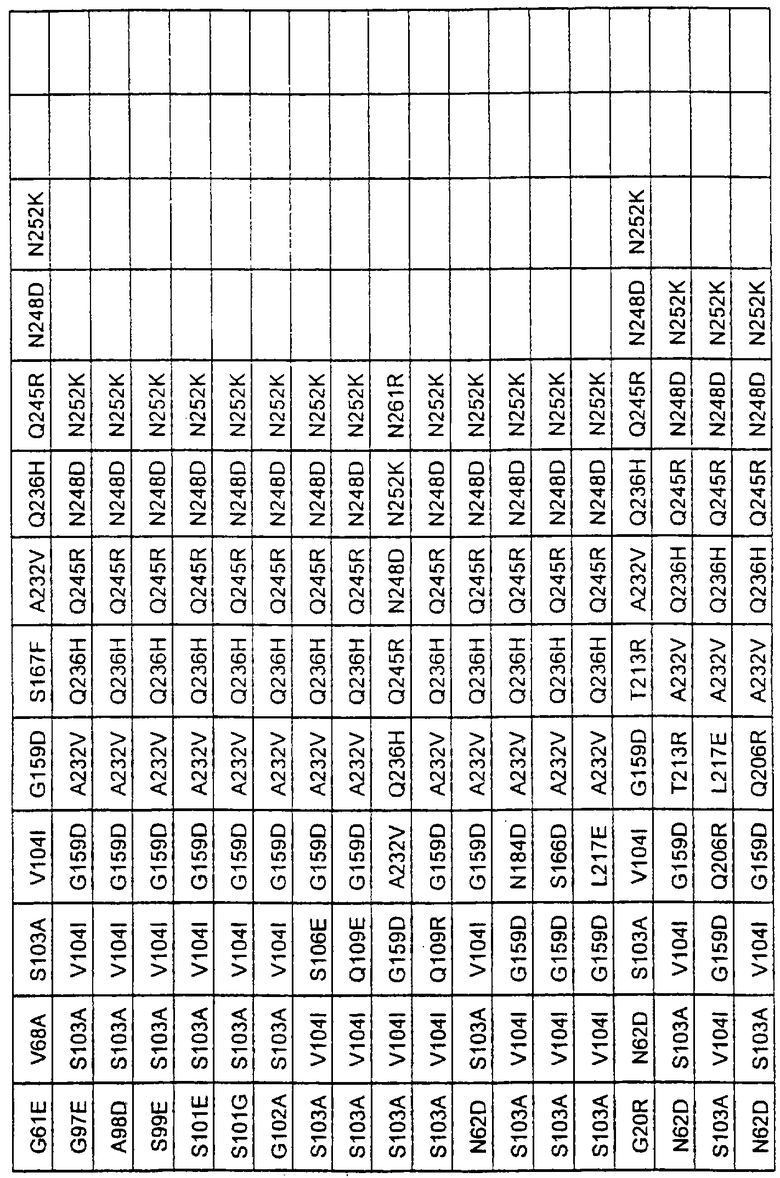
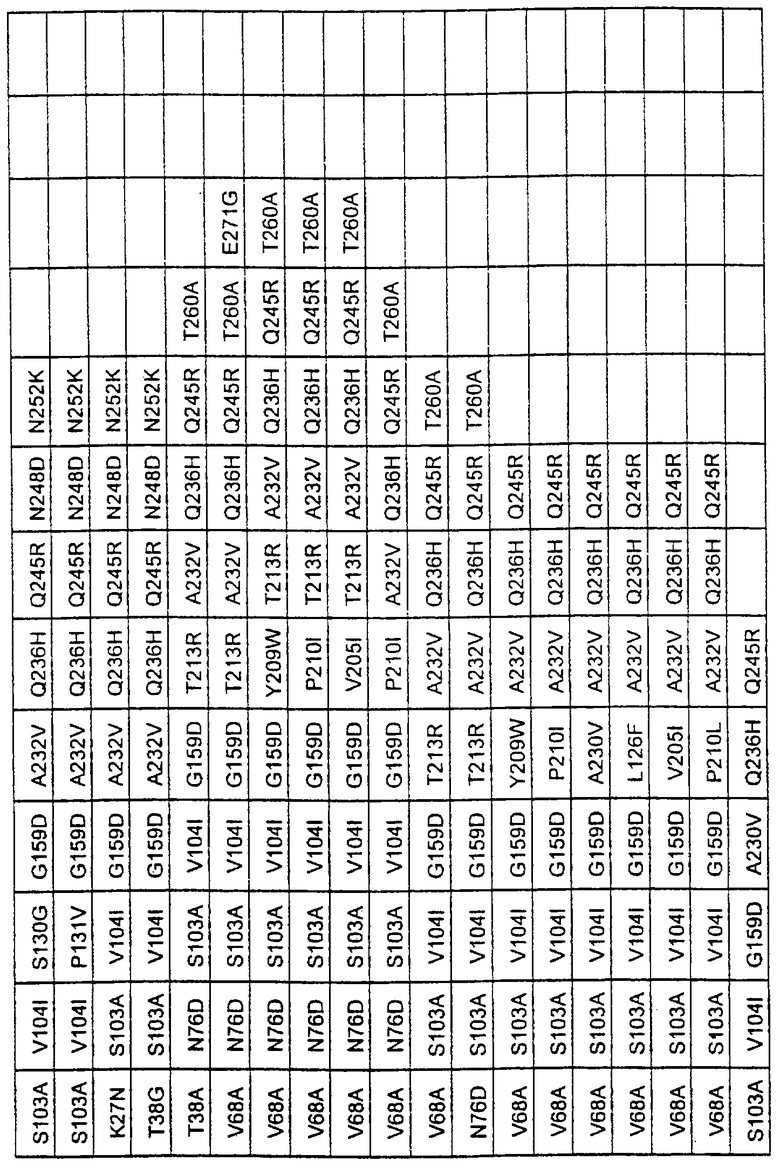
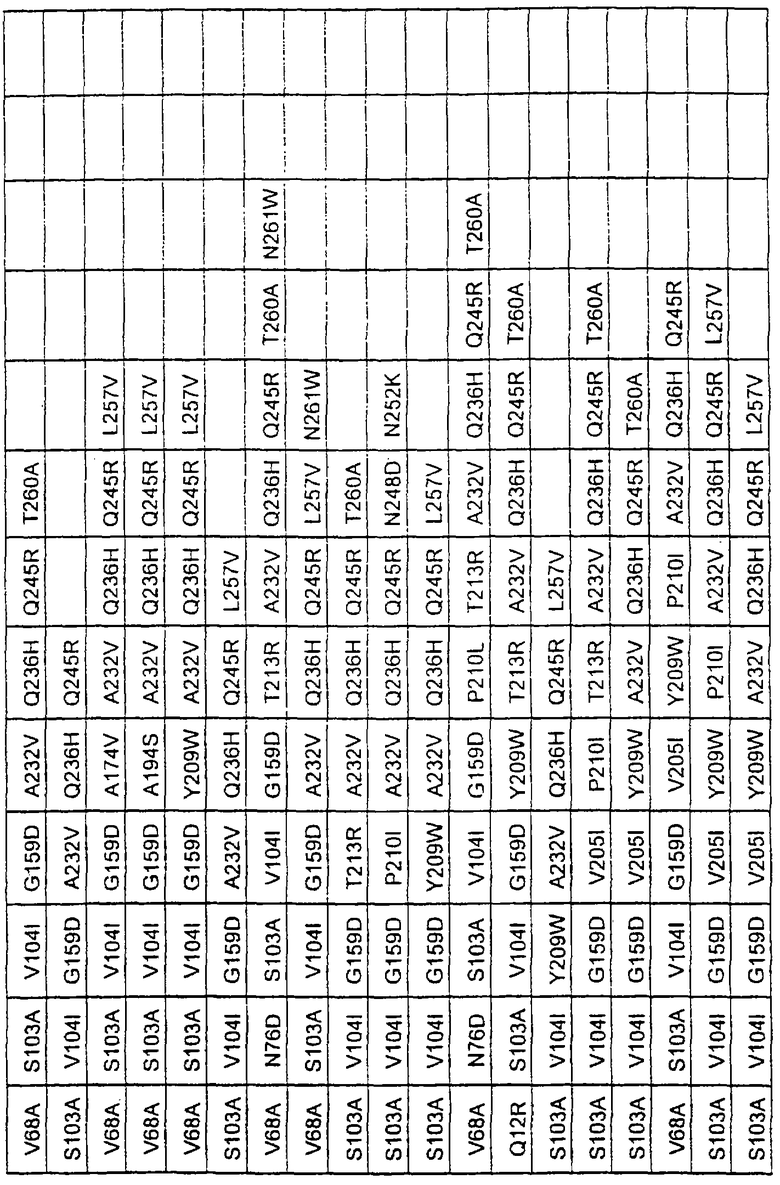
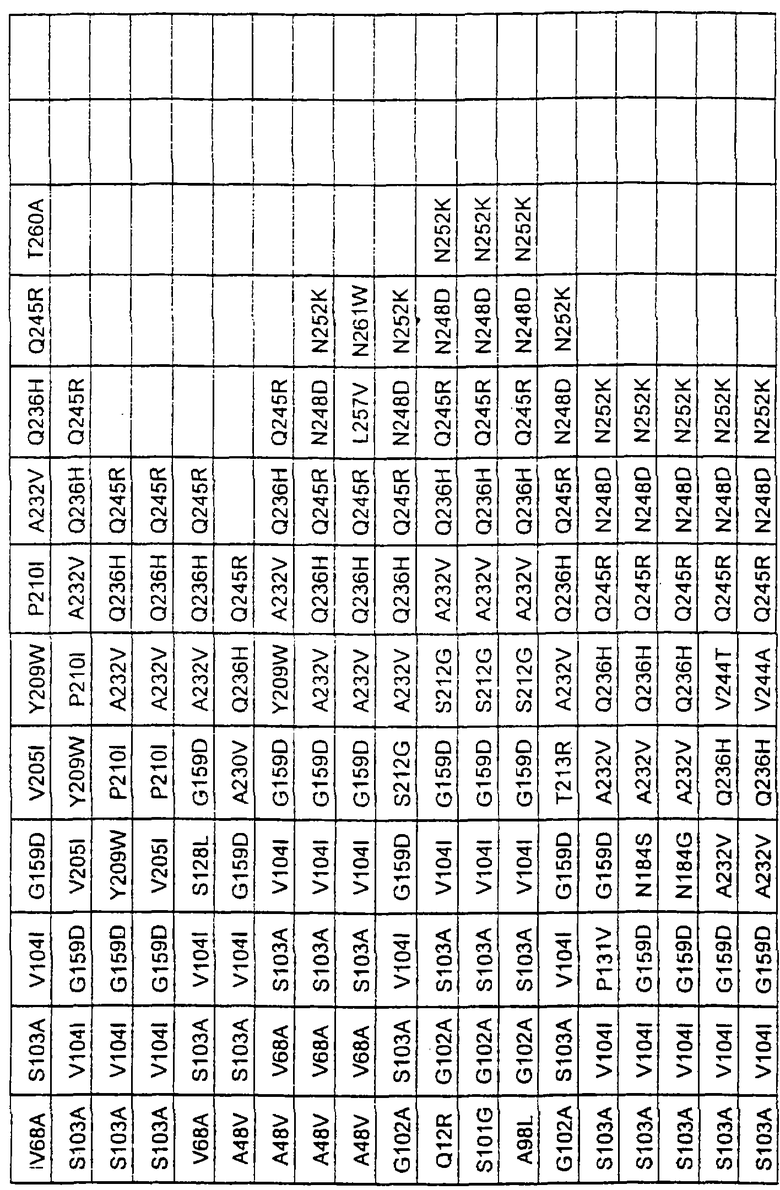
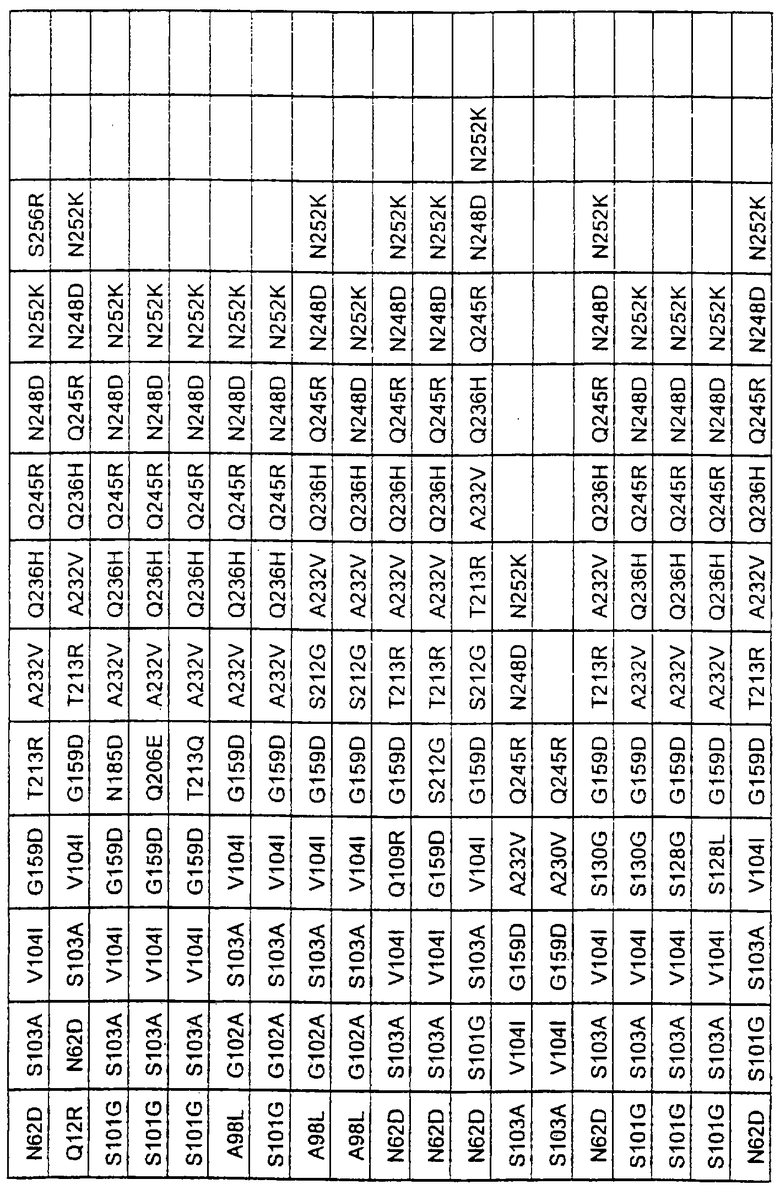
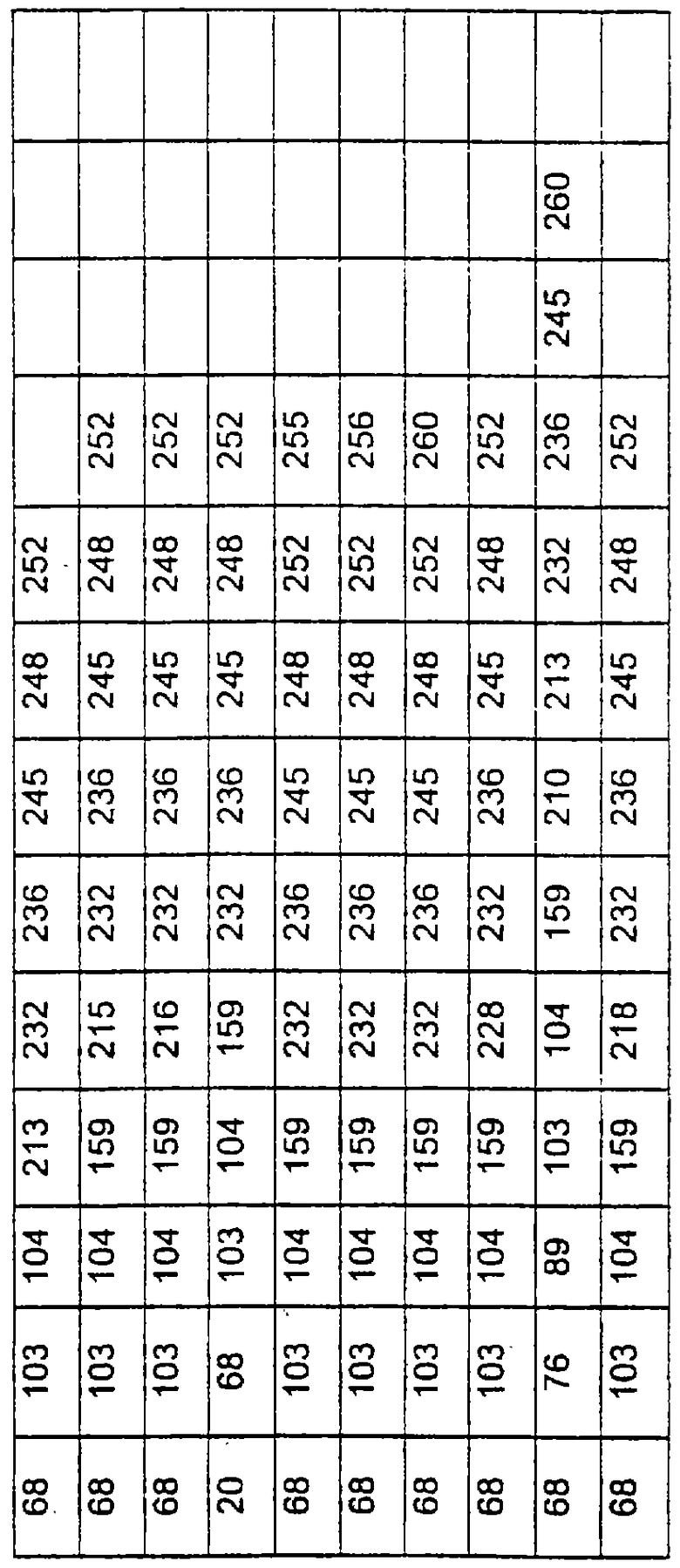
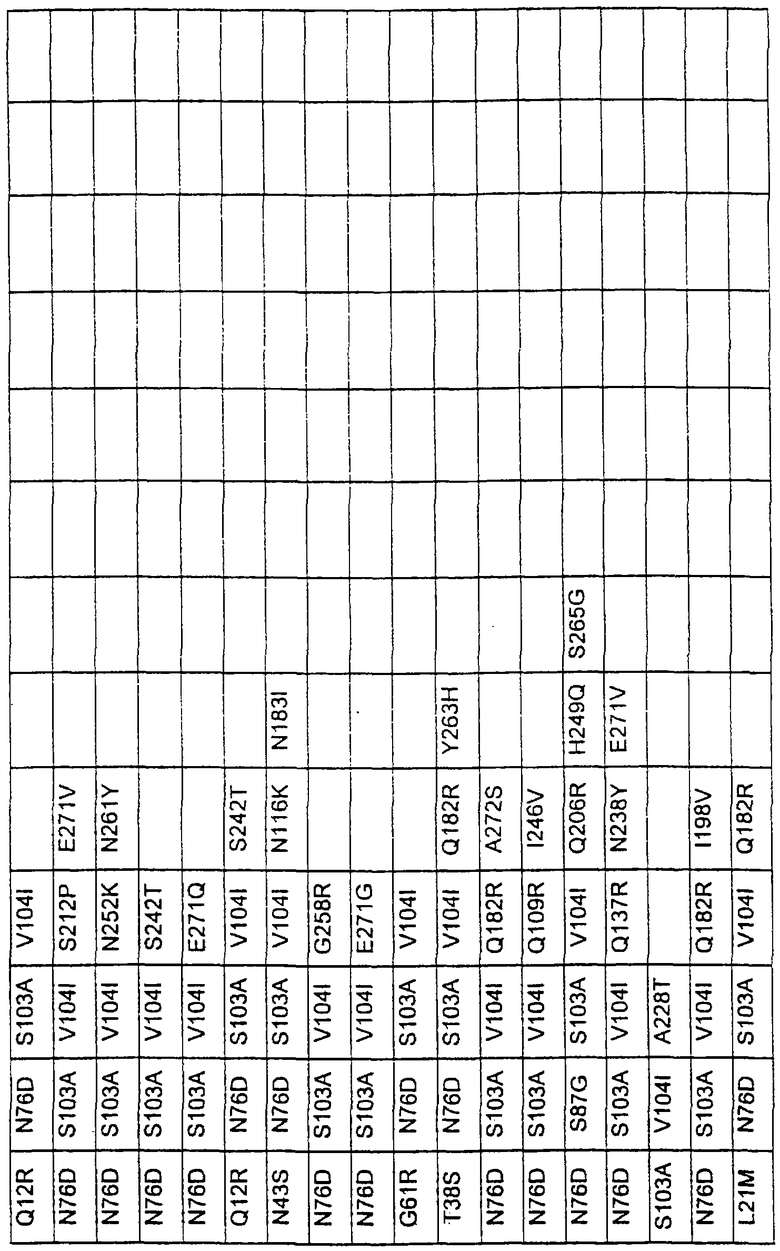
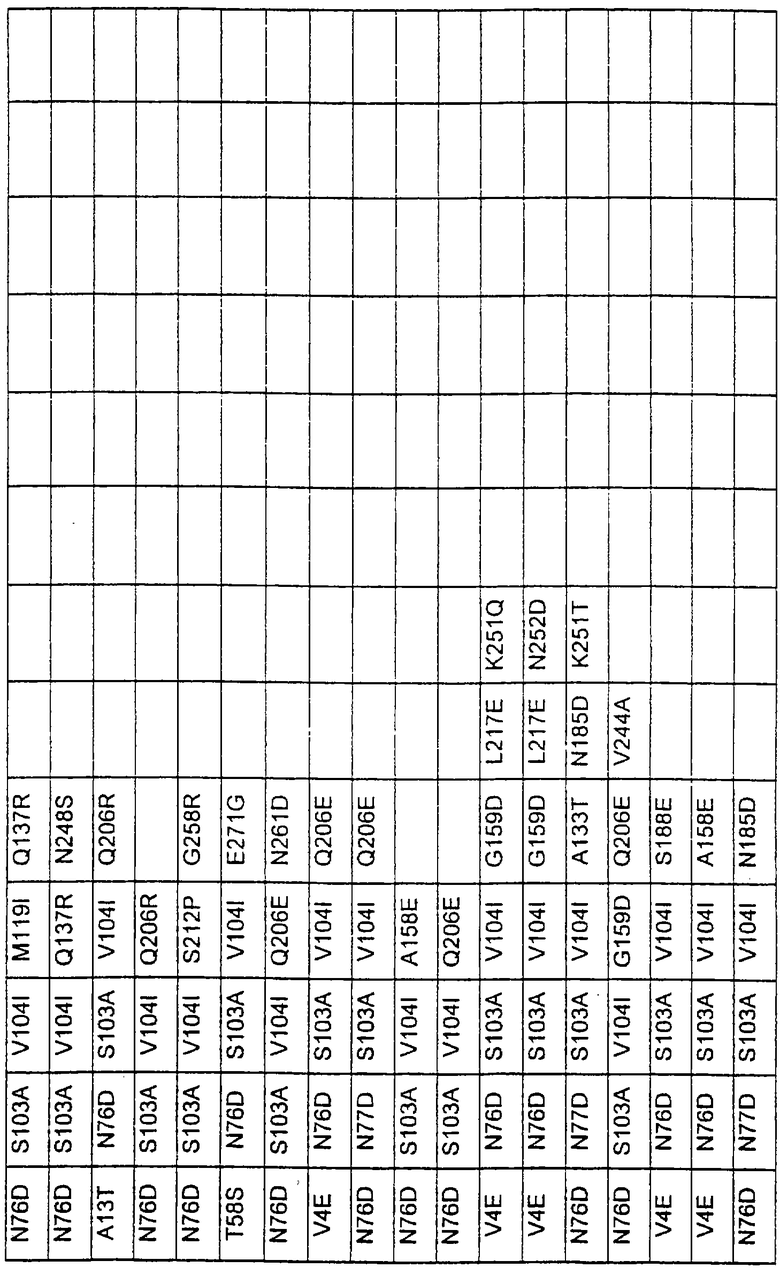
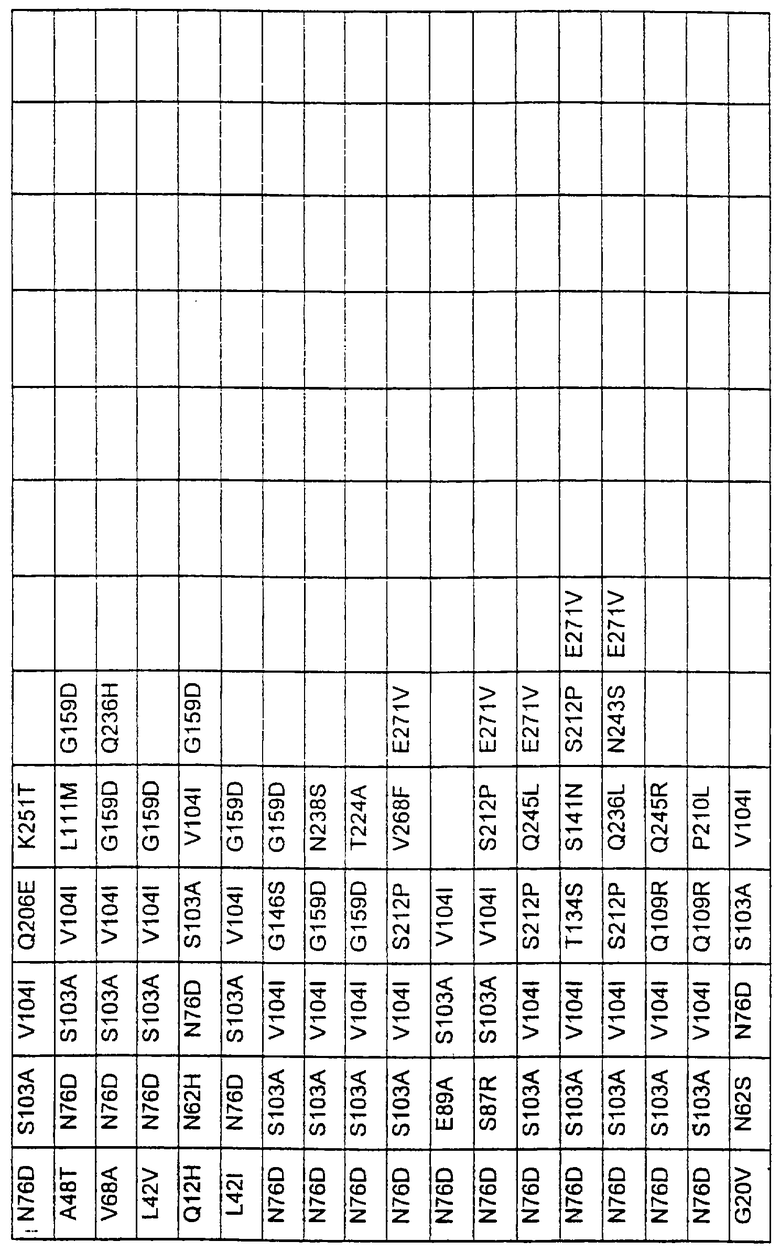
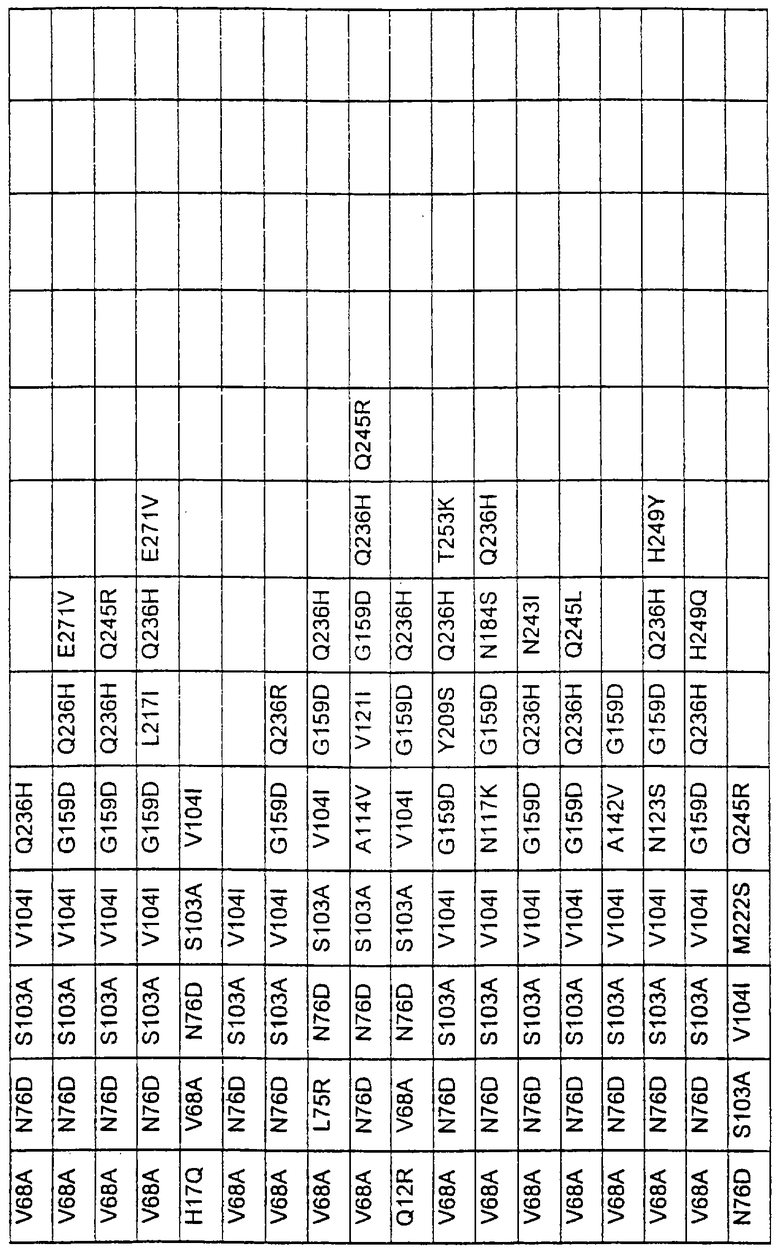
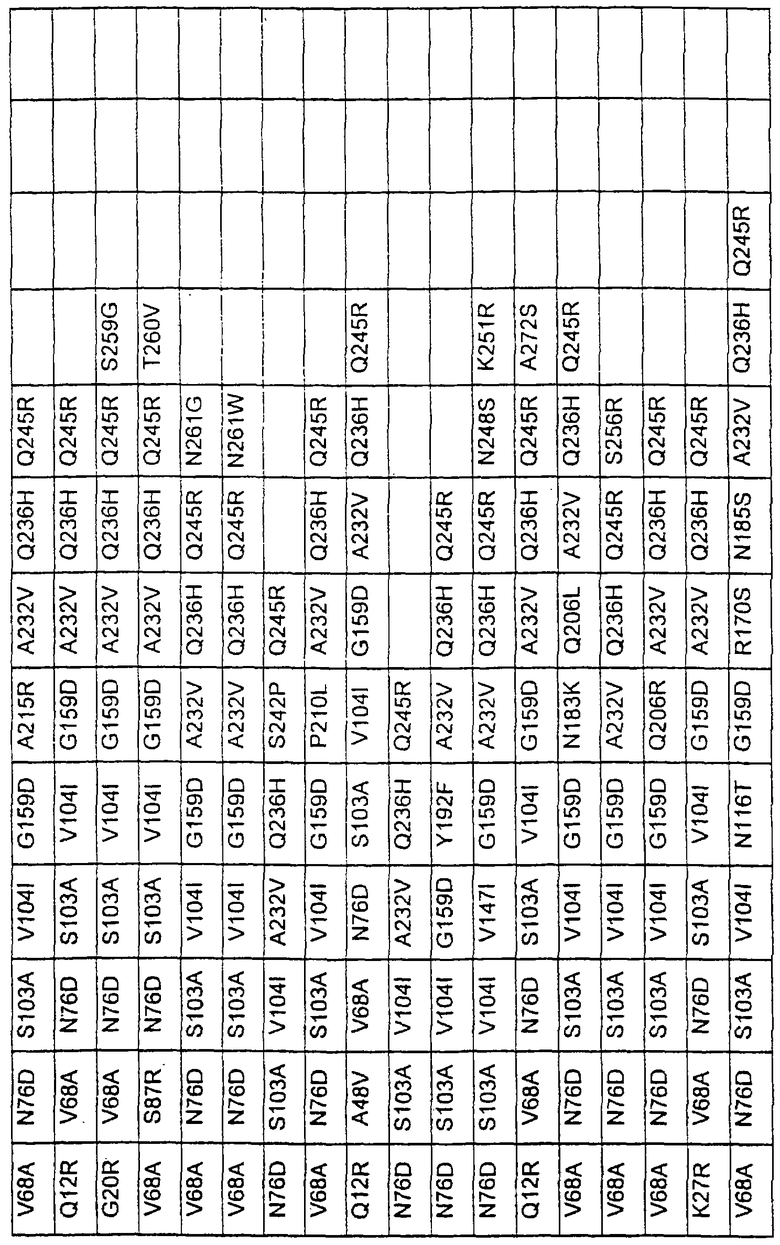
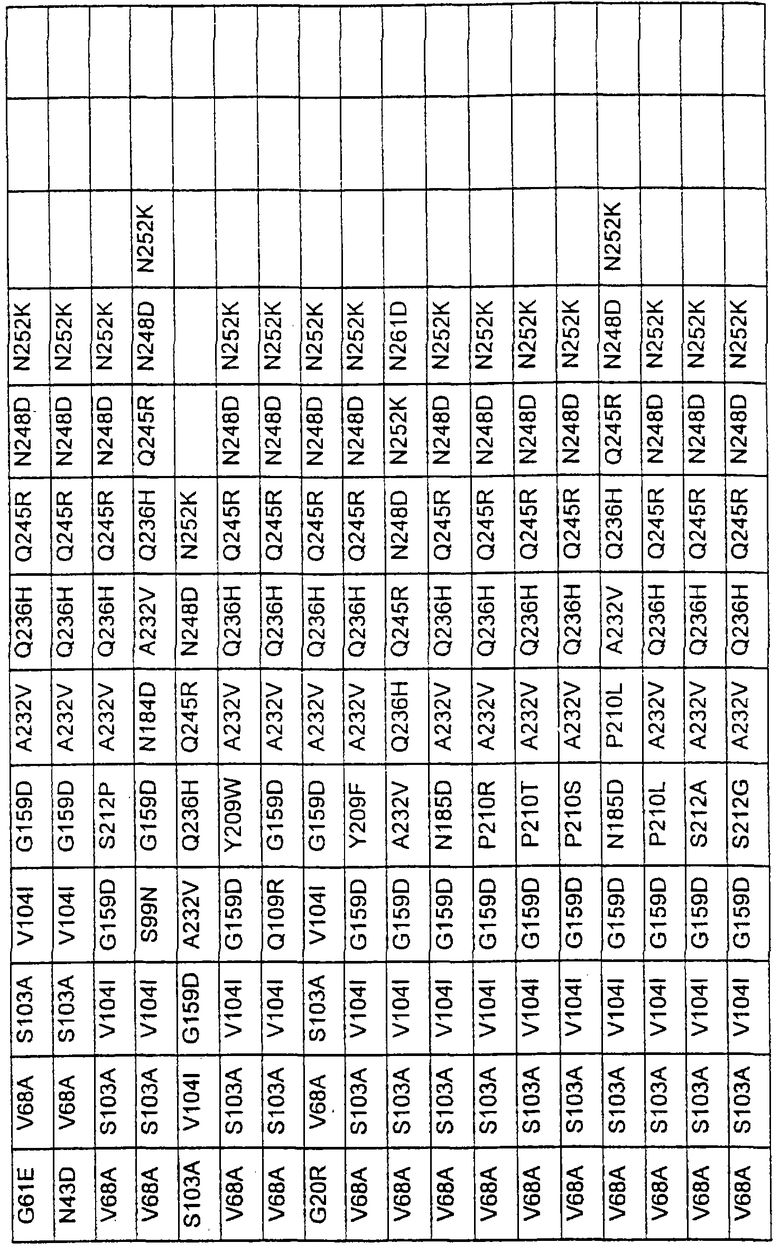
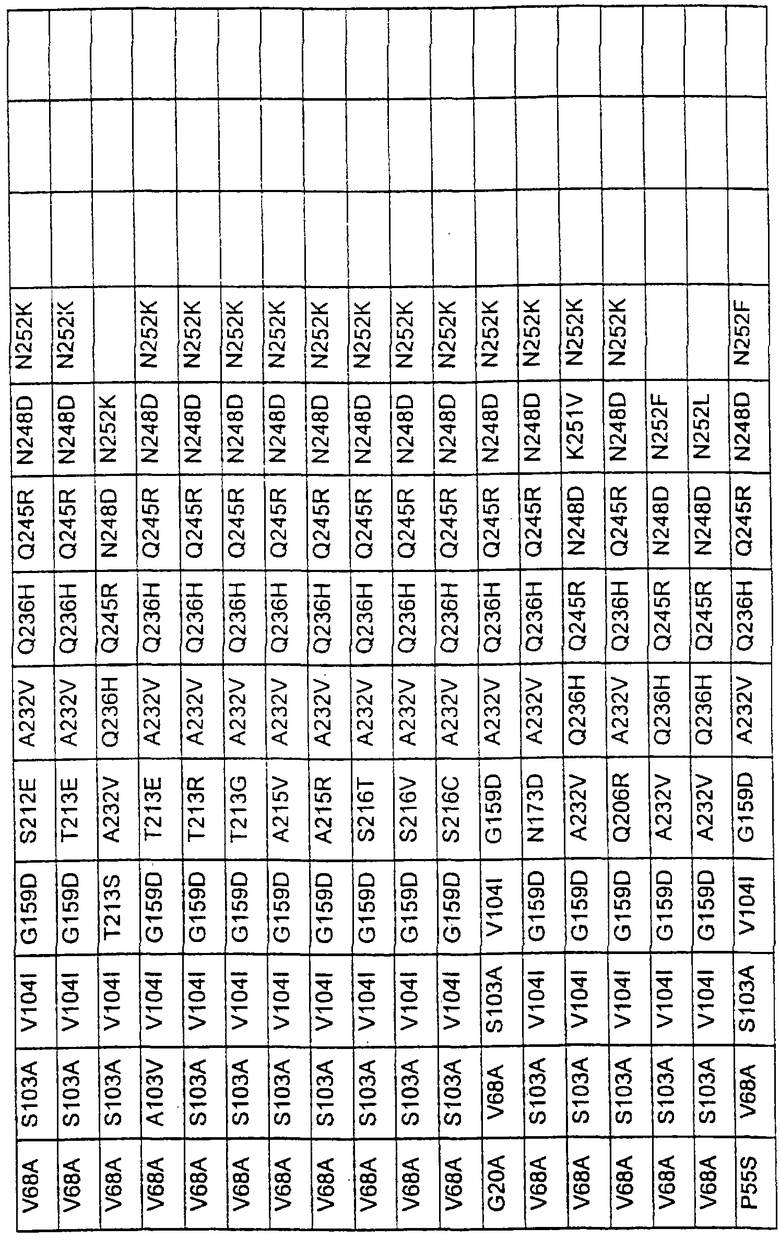
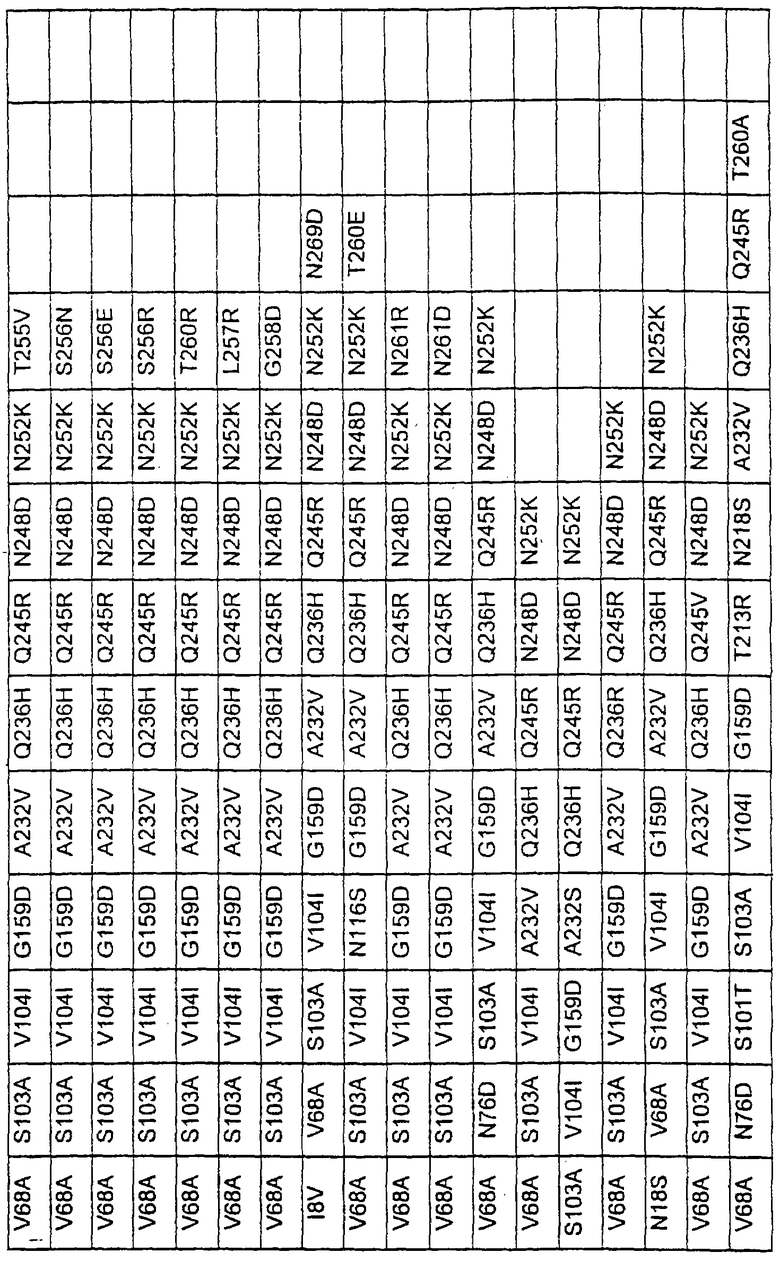
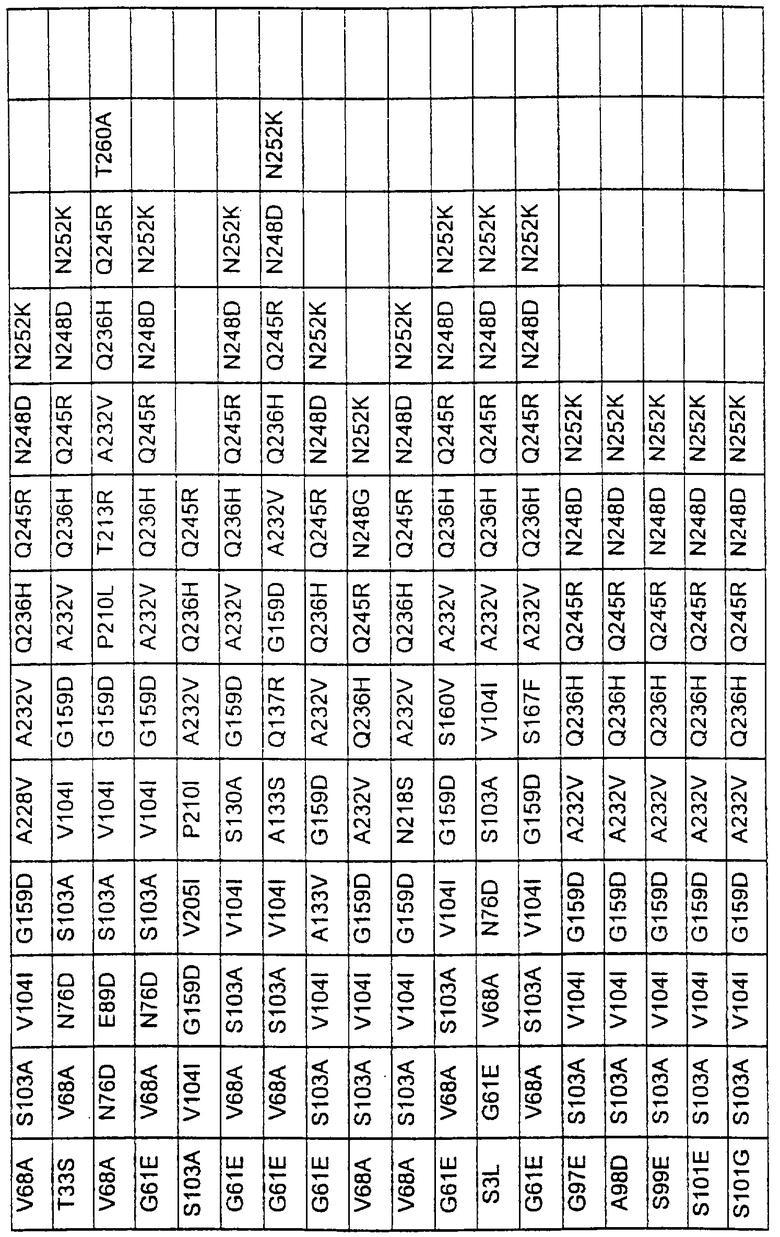
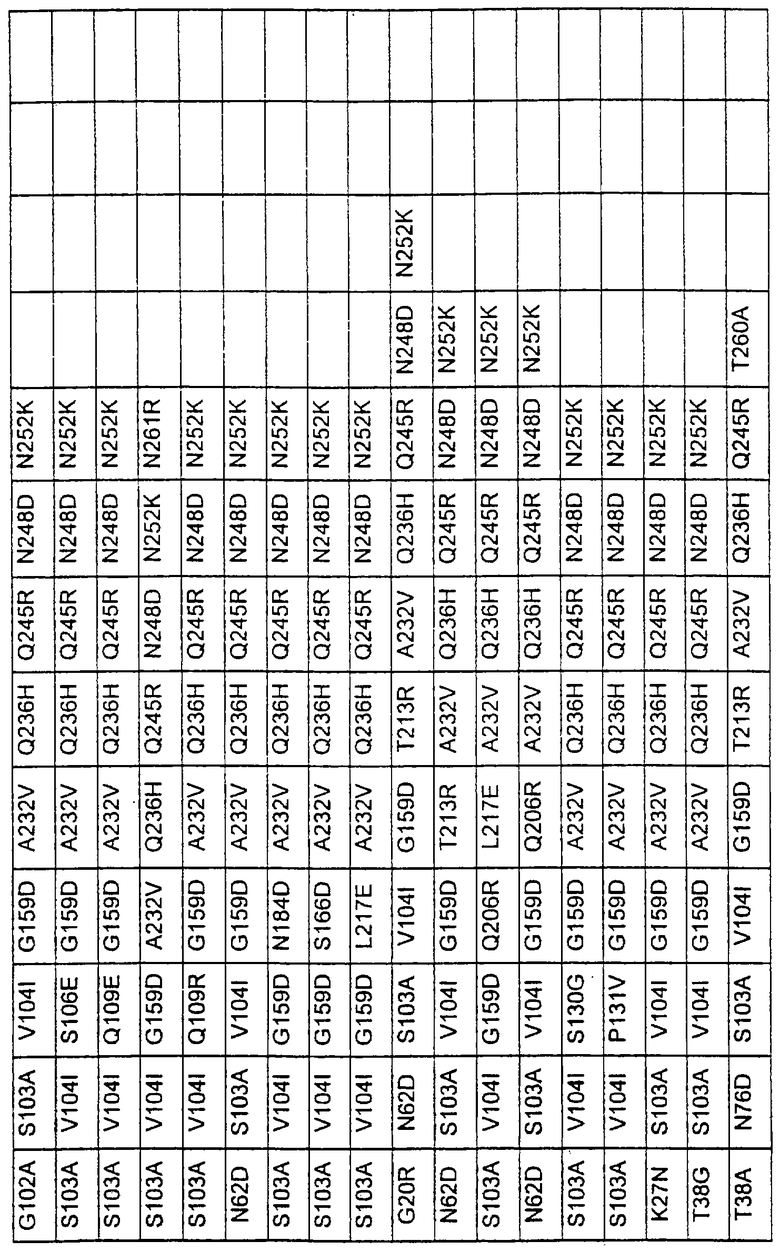
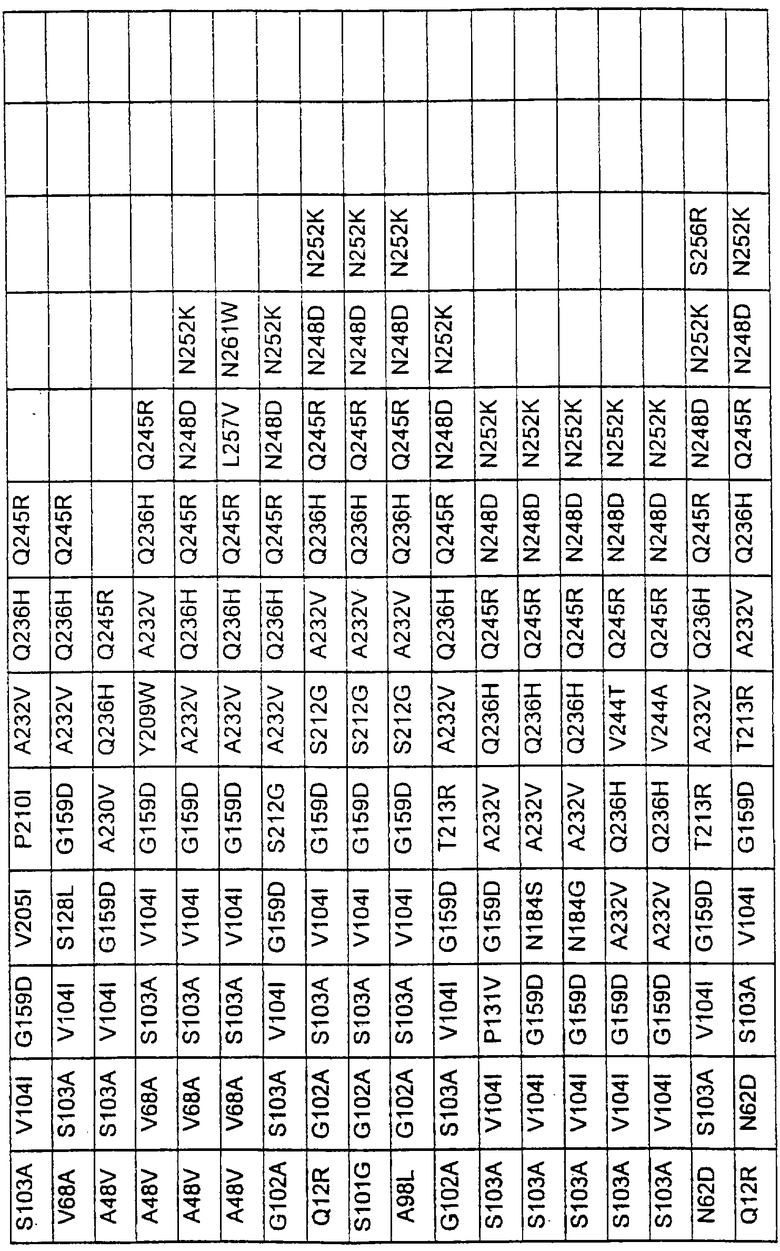
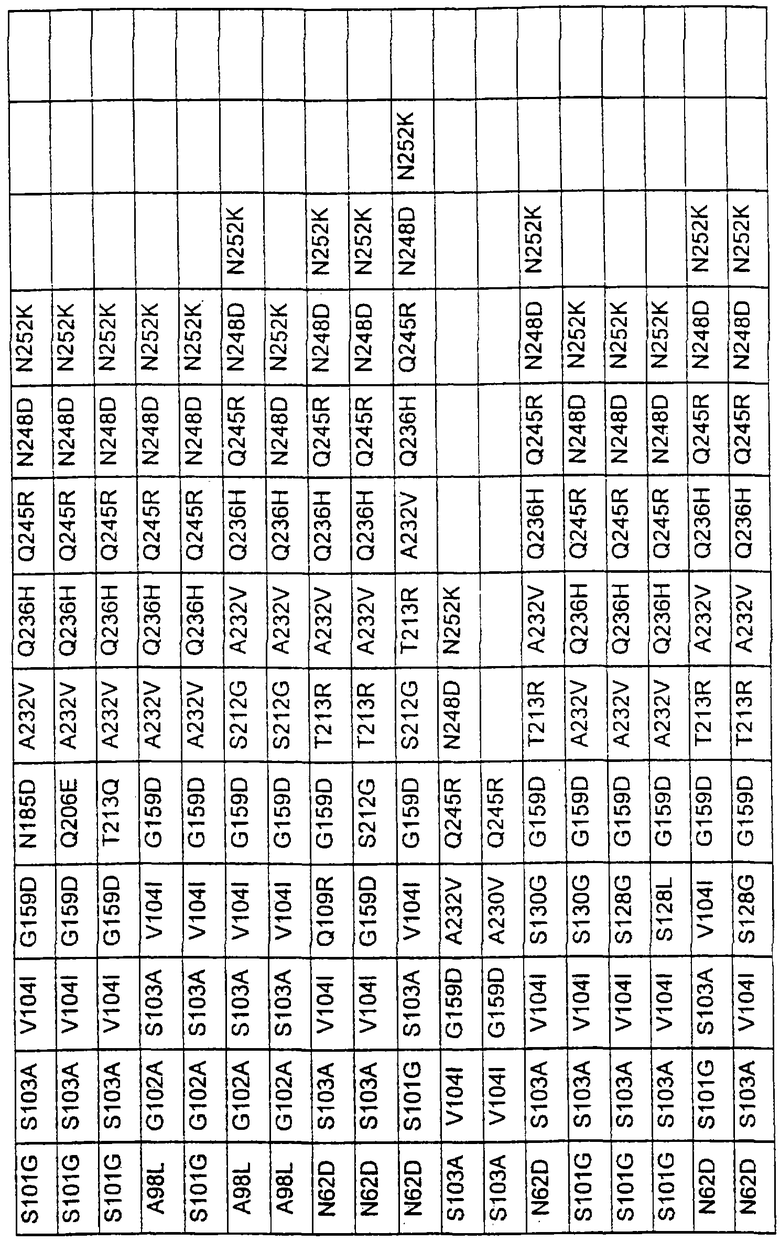
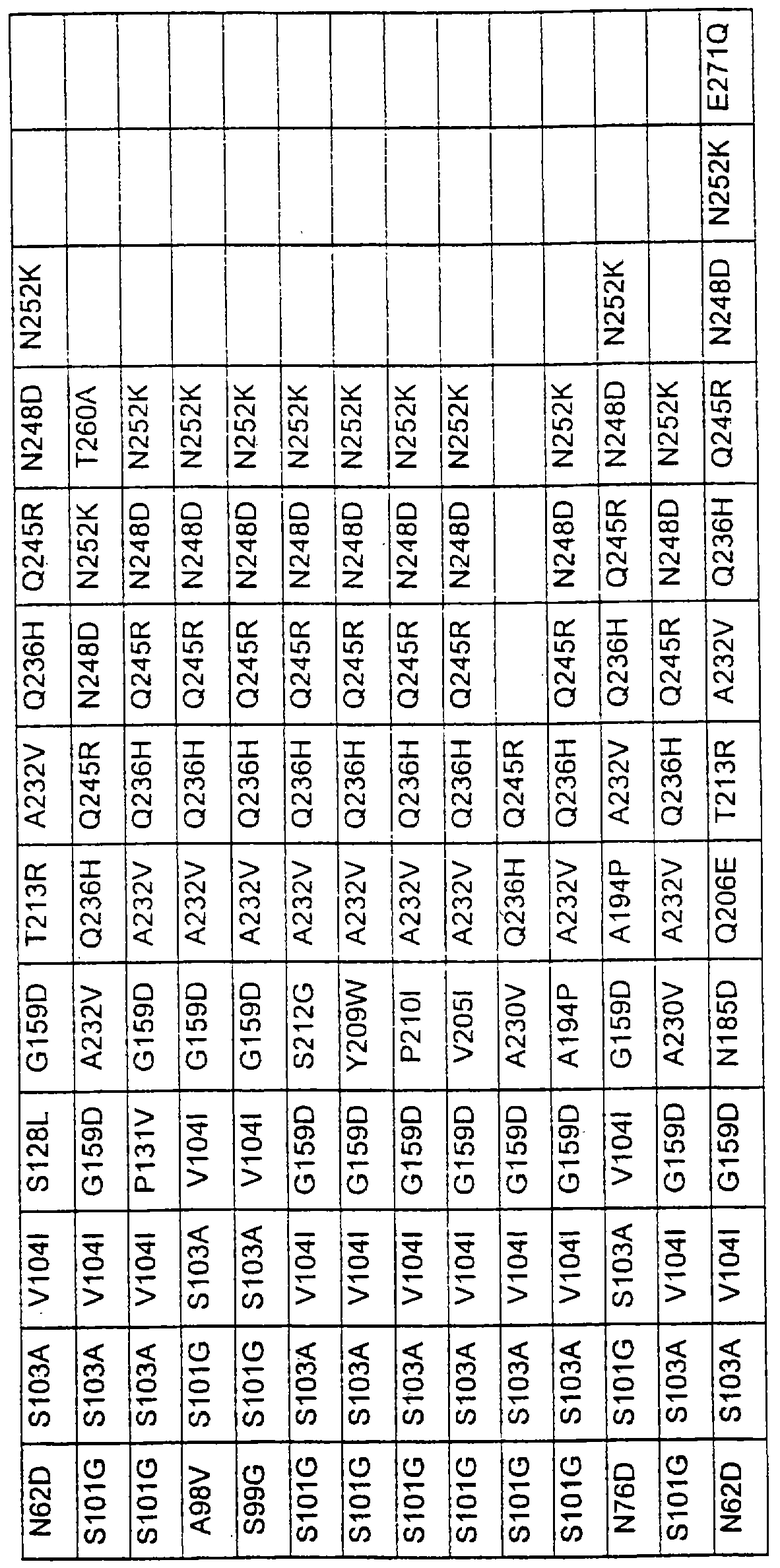
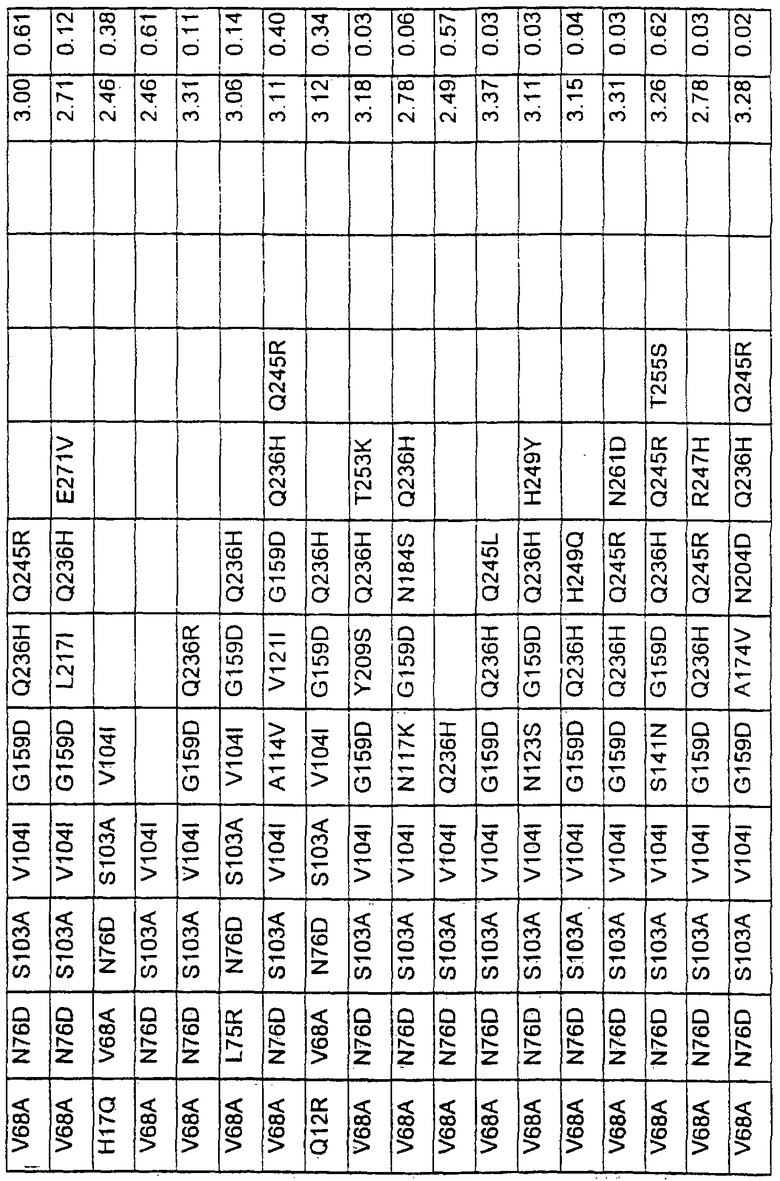
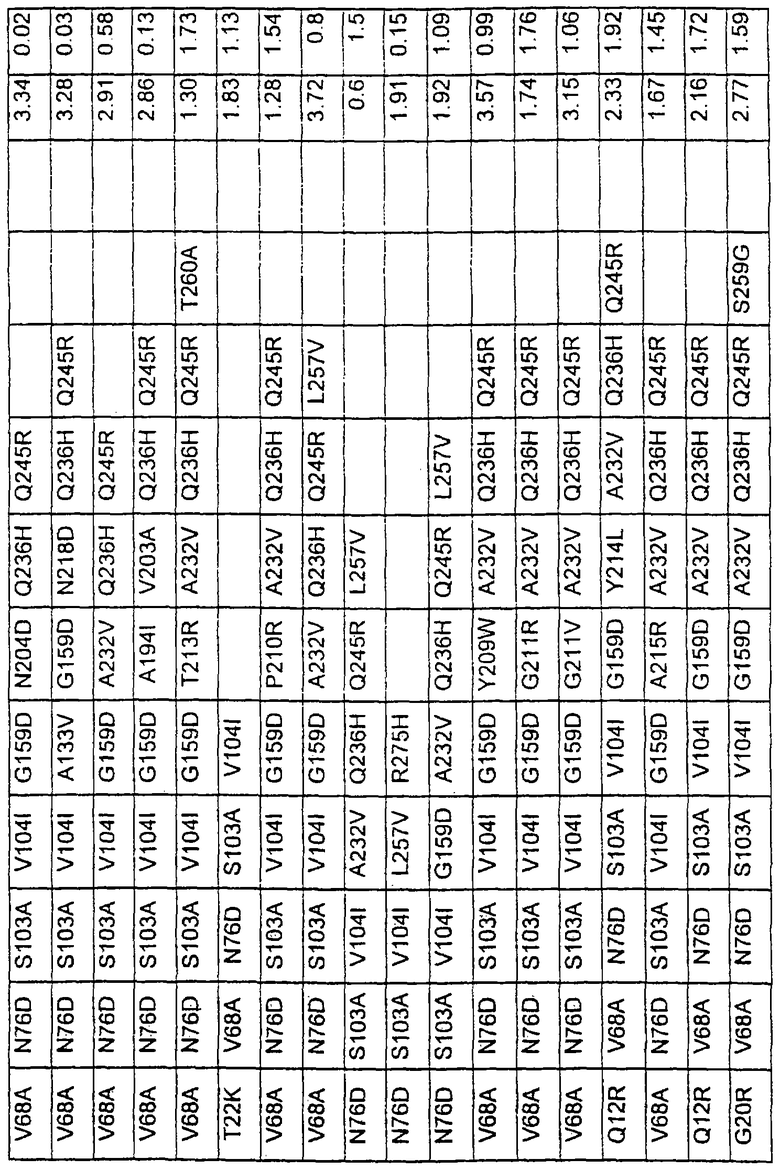
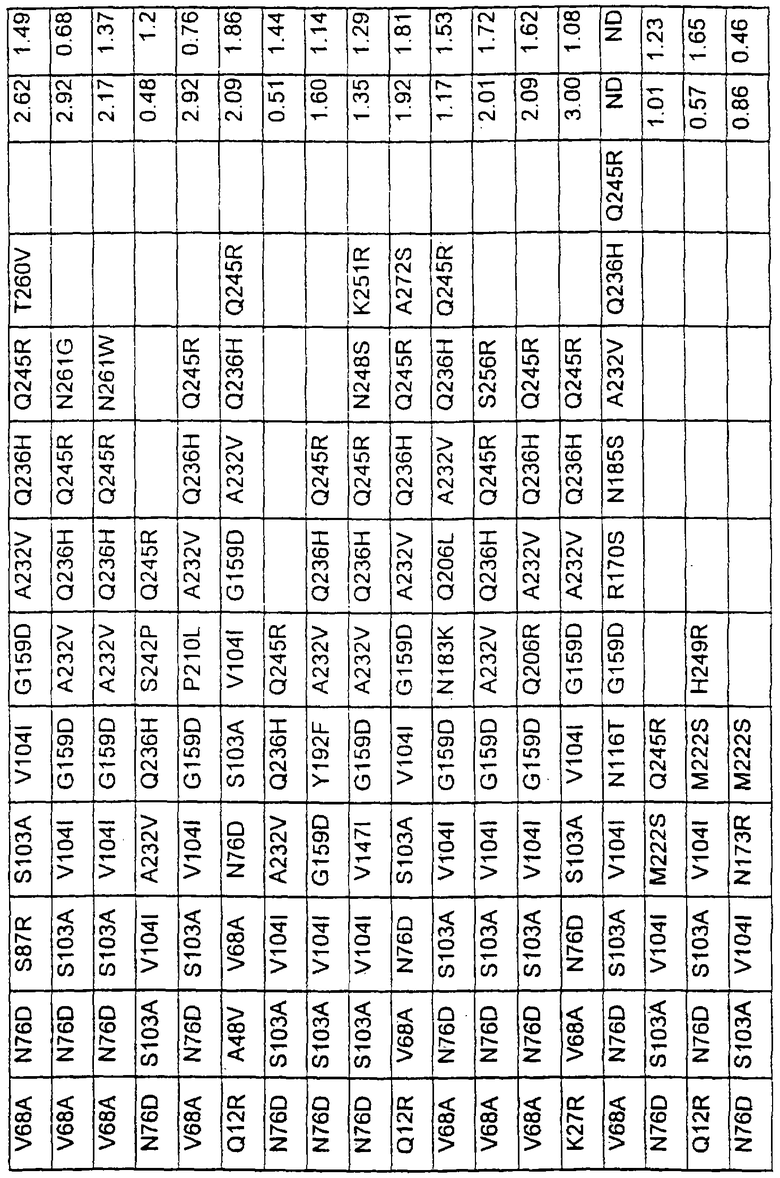
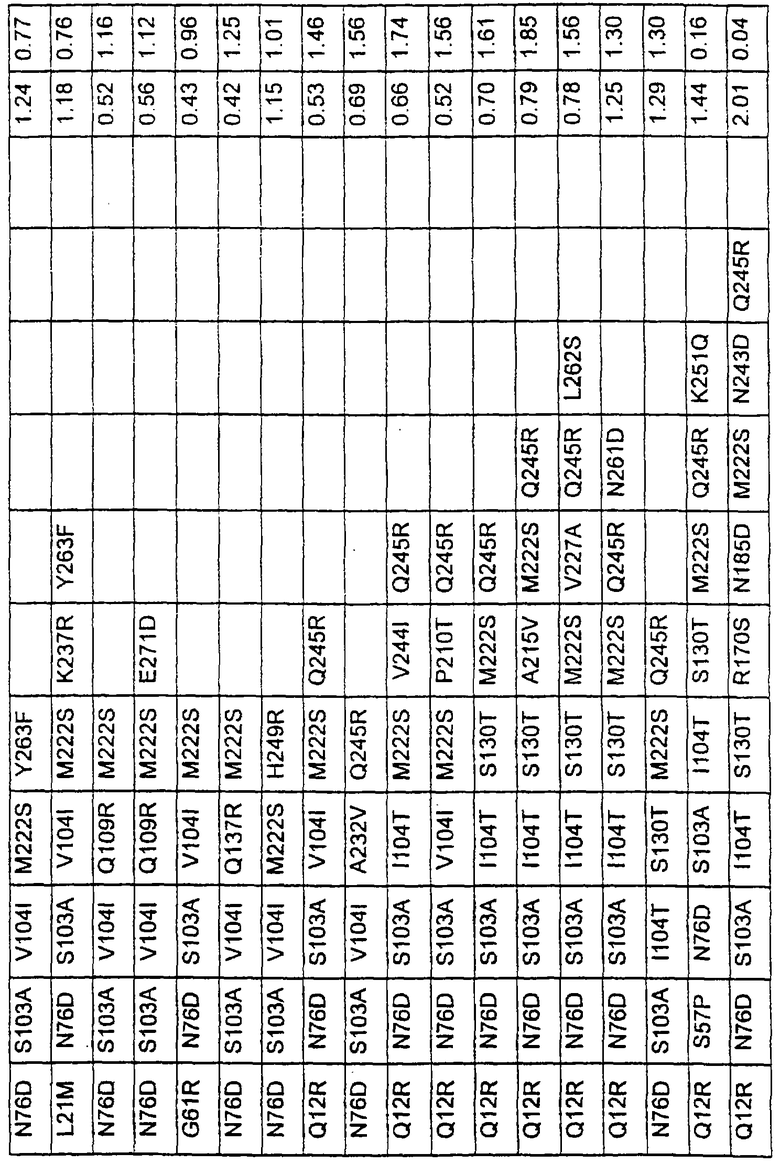
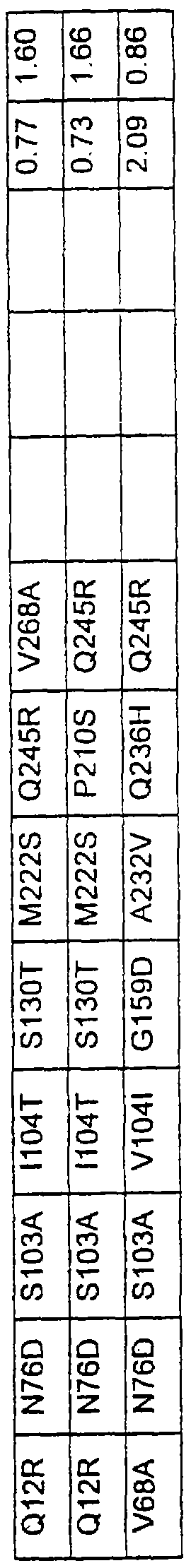
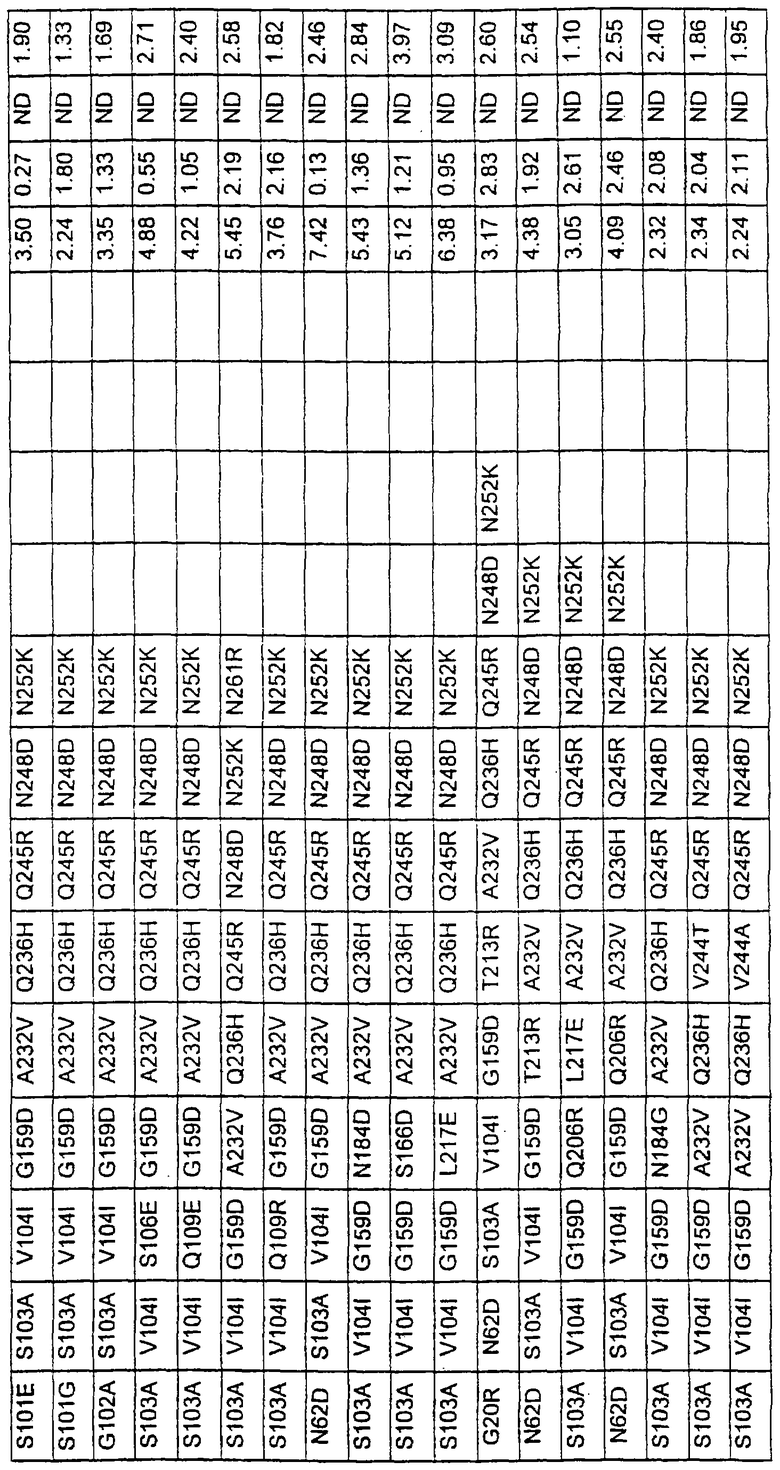
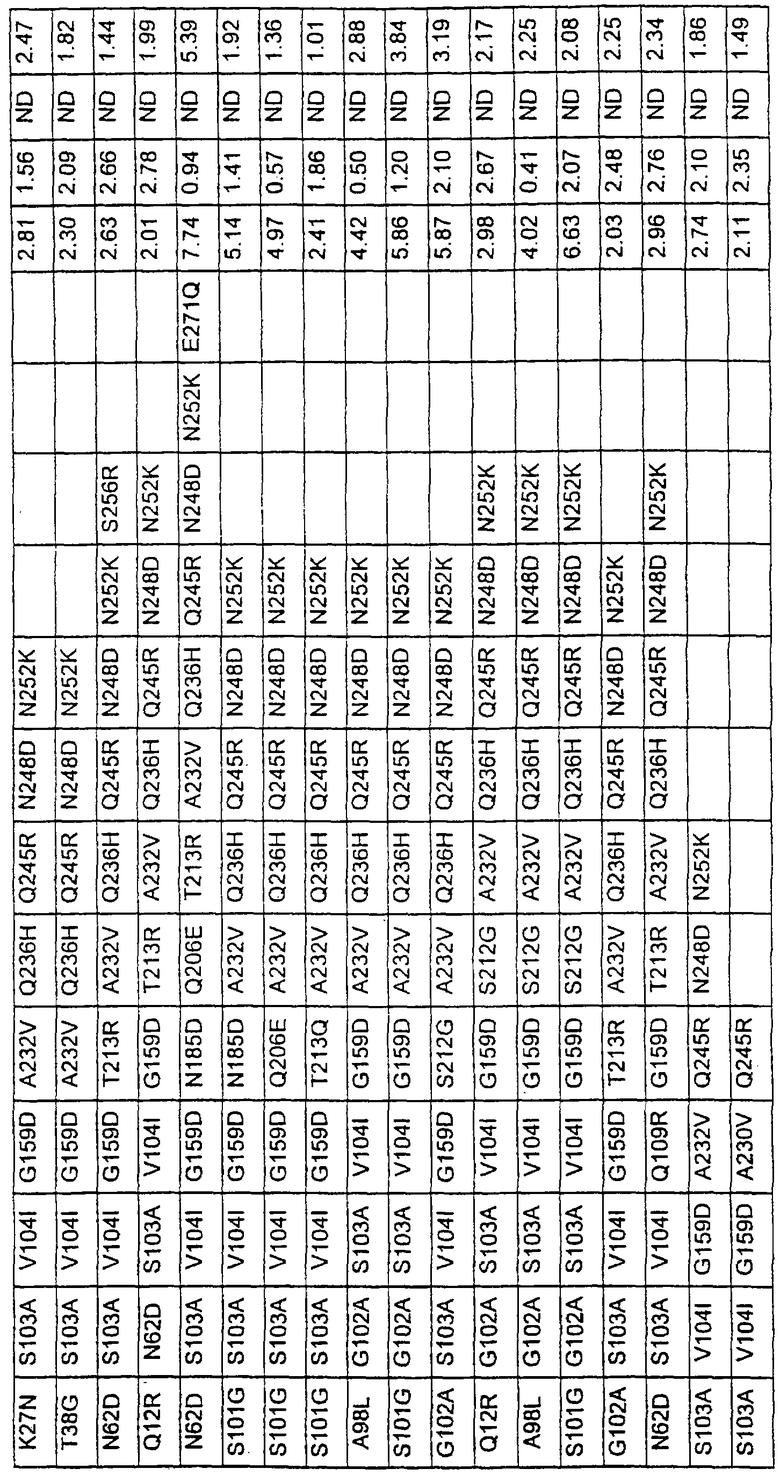
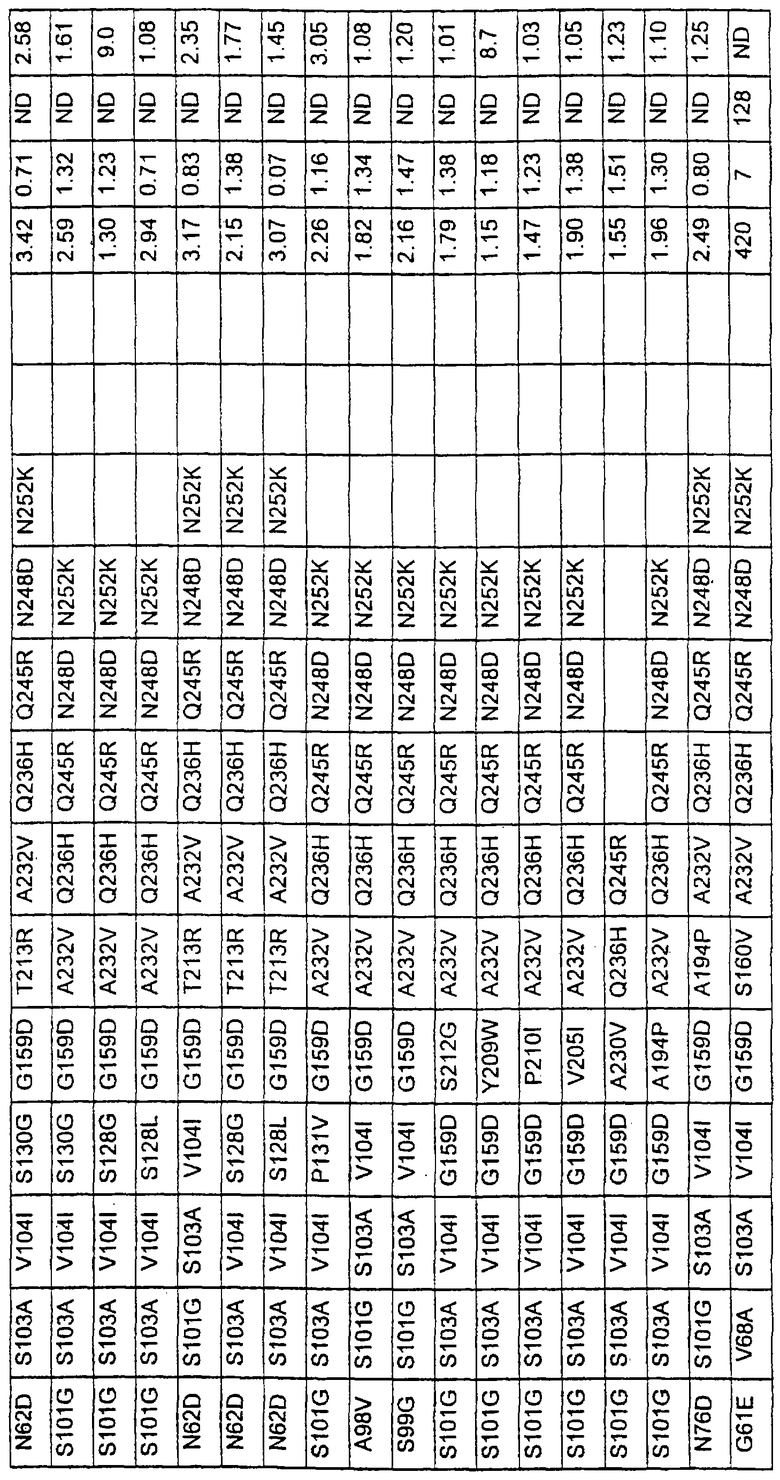
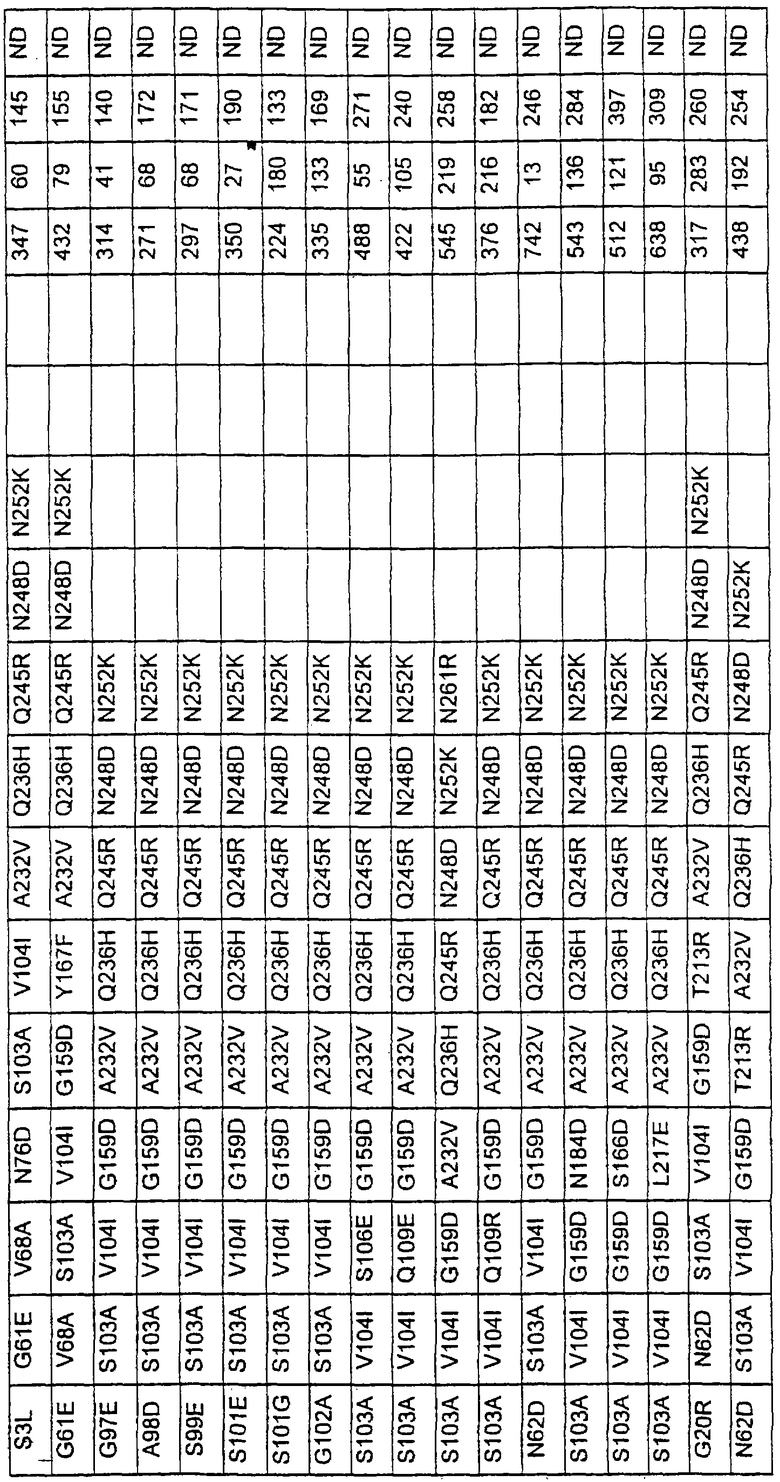
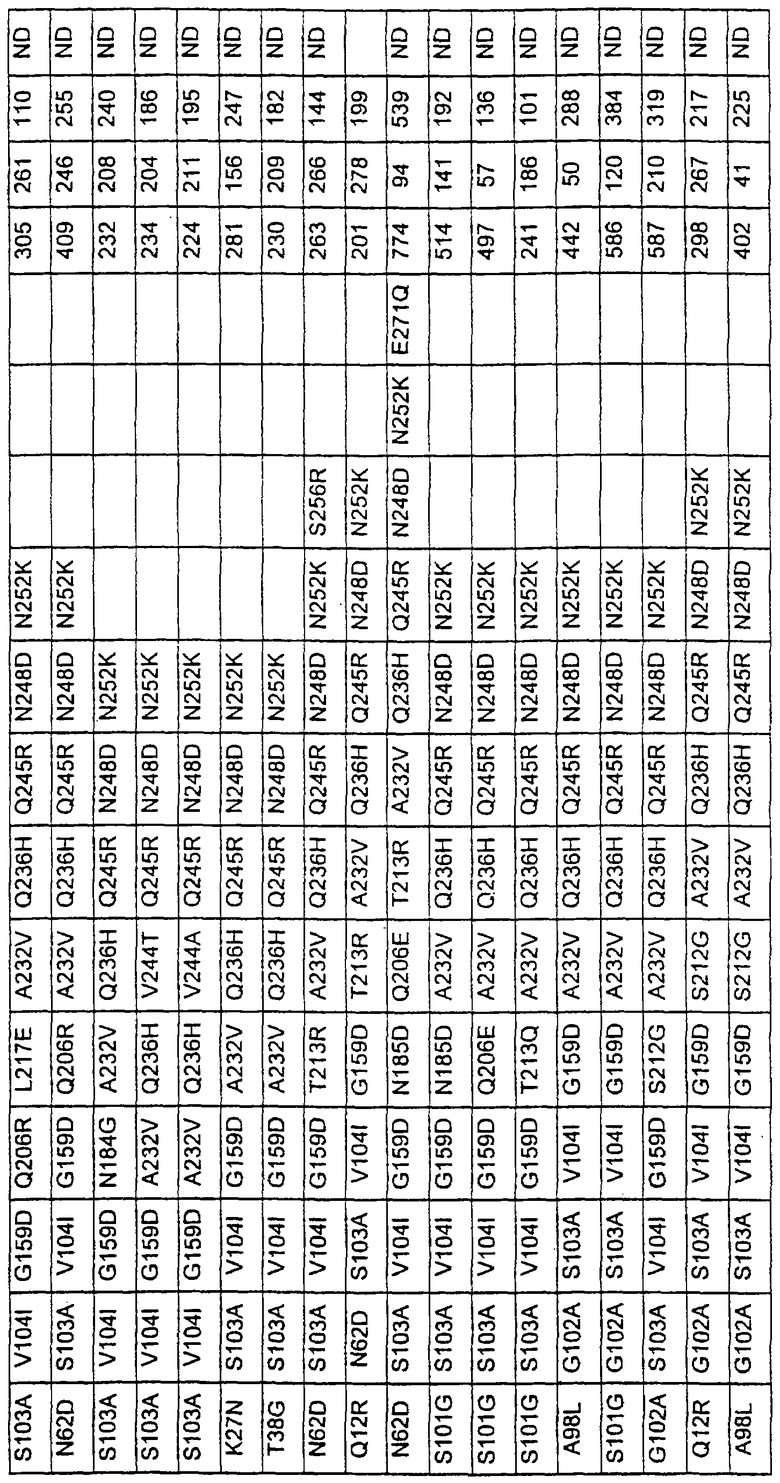
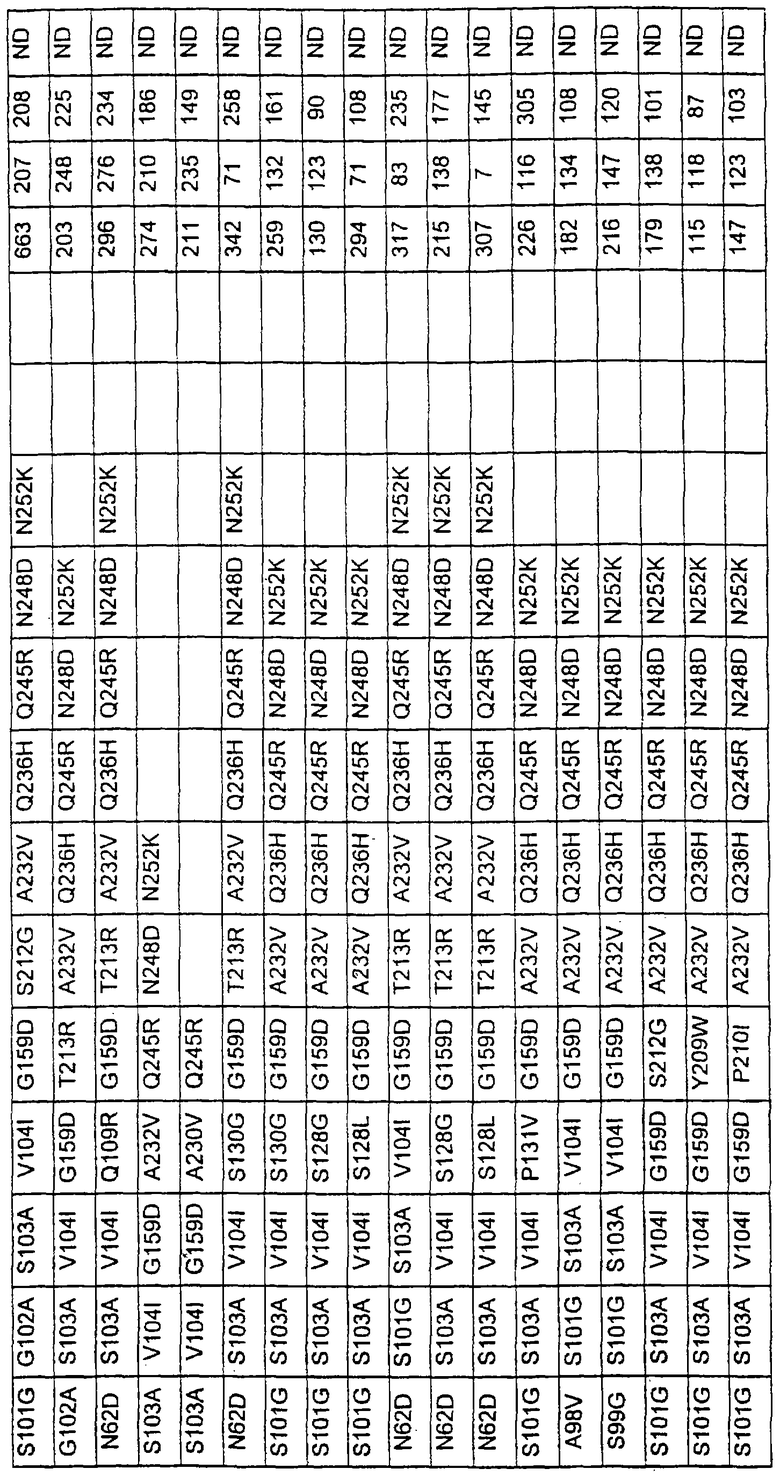
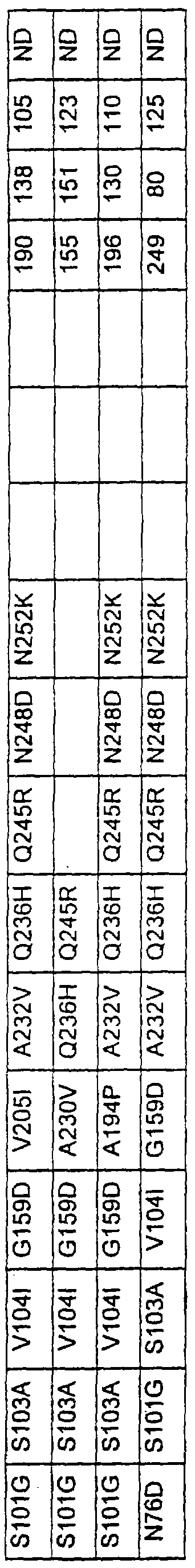
Более предпочтительные варианты протеазы имеют совокупности замещений, выбираемых из группы, включающей положения остатков субтилизина Bacillus amyloliquefaciens, соответствующие положениям, указанным в таблице 1.
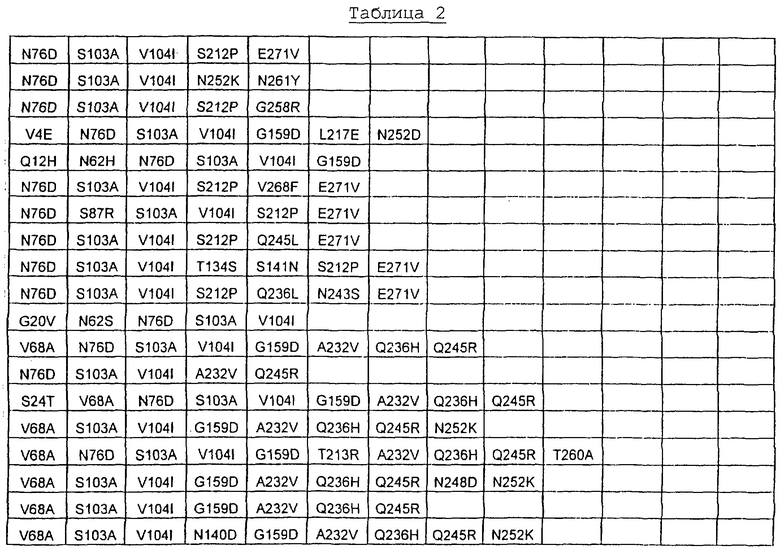
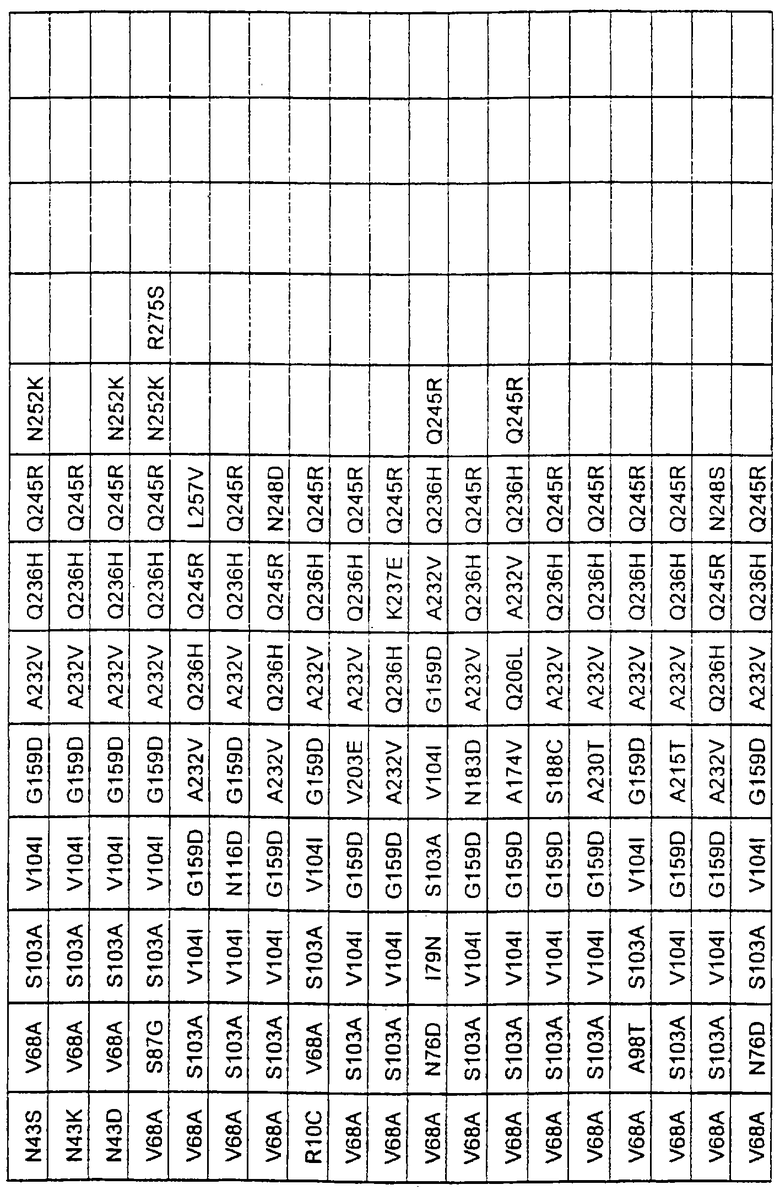
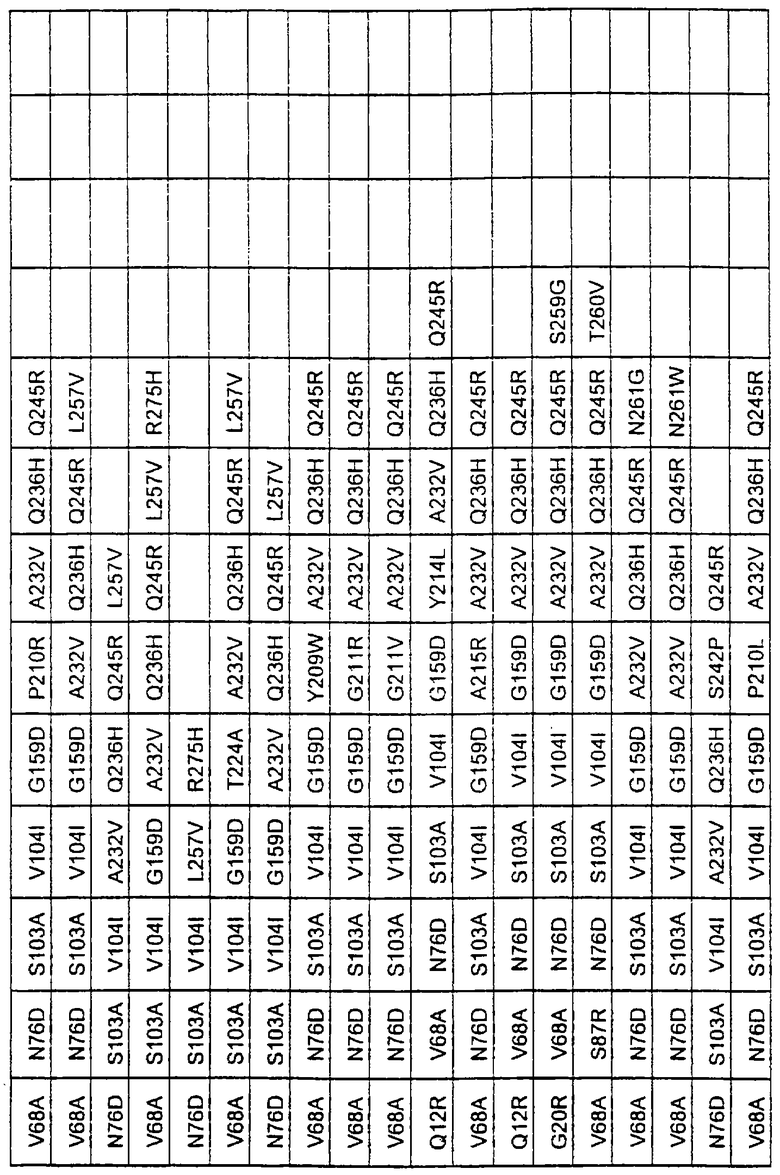
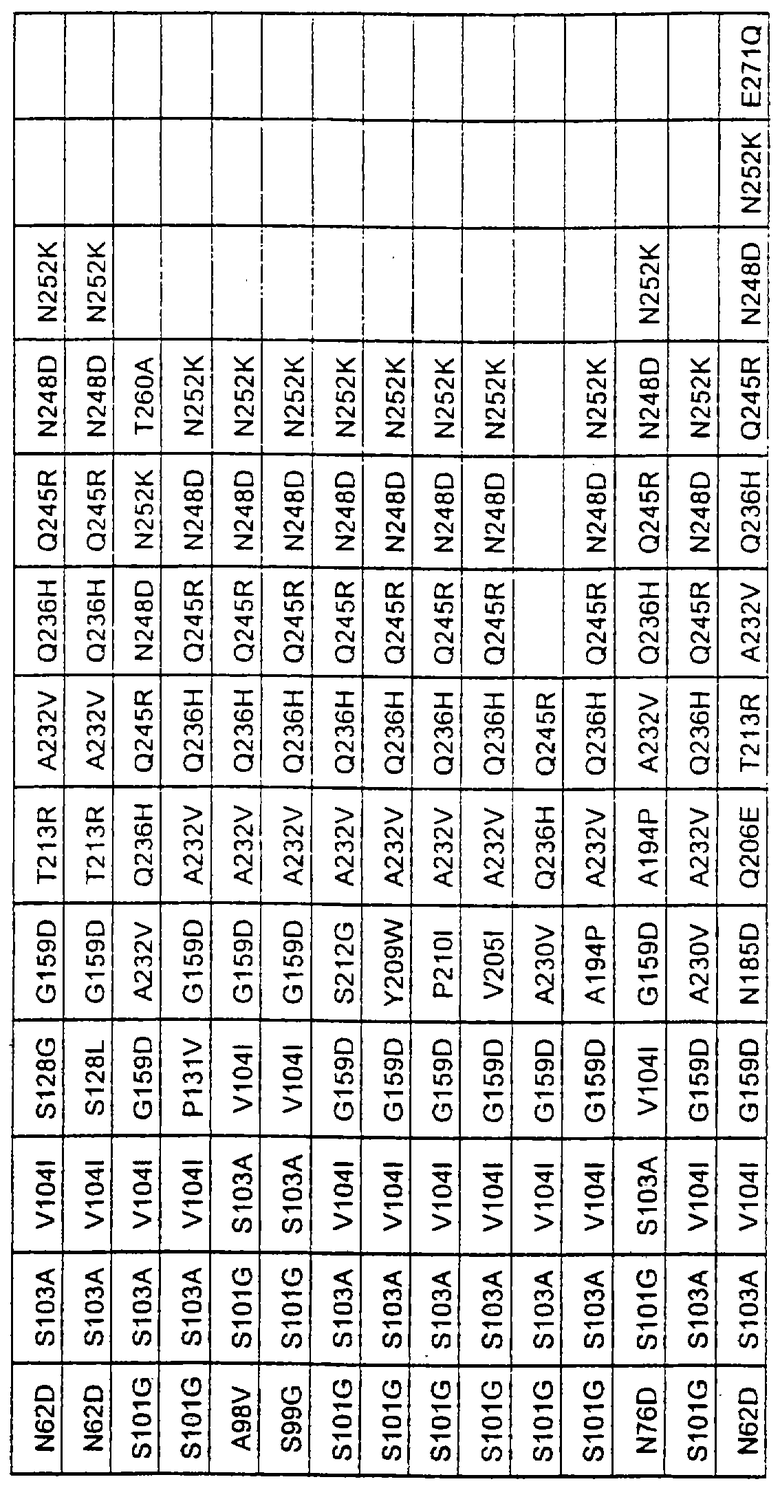
Наиболее предпочтительные варианты протеазы имеют совокупности замещений, выбираемых из группы, включающей положения остатков субтилизина Bacillus amyloliquefaciens, соответствующие положениям, указанным в таблице 2.
Другим объектом этого изобретения являются последовательности ДНК, кодирующие такие варианты протеазы, а также экспрессирующие векторы, содержащие последовательности ДНК таких вариантов.
Еще одним объектом этого изобретения являются клетки-хозяева, трансформированные такими векторами, а также клетки-хозяева, способные экспрессировать такую ДНК для продуцирования вариантов протеазы внутриклеточно или внеклеточно.
Данное изобретение относится далее к очищающей композиции, содержащей вариант протеазы по настоящему изобретению.
Кроме того, это изобретение относится к корму для животных, содержащему вариант протеазы по настоящему изобретению.
Данное изобретение относится также к композиции для обработки ткани, содержащей вариант протеазы по настоящему изобретению.
Краткое описание чертежей
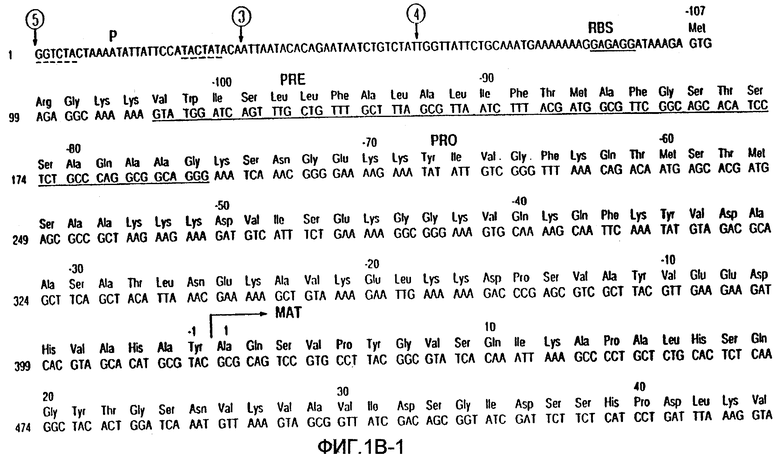
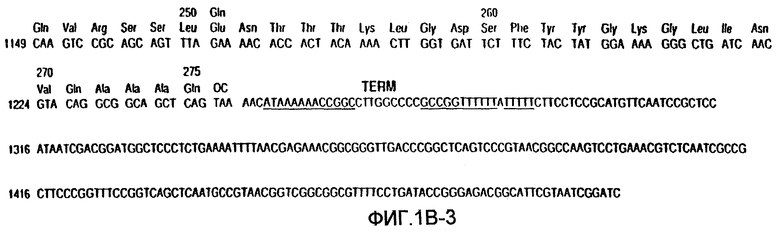
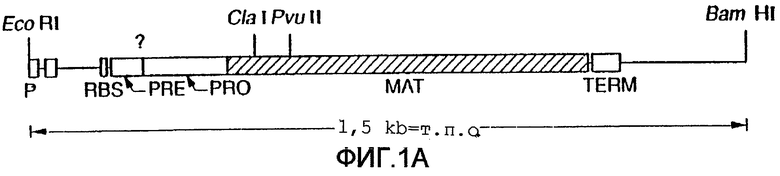
На фиг.1А-В показаны последовательность ДНК и аминокислотная последовательность для субтилизина Bacillus amyloliquefaciens и частичная рестрикционная карта этого гена.
На фиг.2 показаны консервативные аминокислотные остатки субтилизинов из Bacillus amyloliquefaciens (bpn)’ и Bacillus lentus (дикого типа).
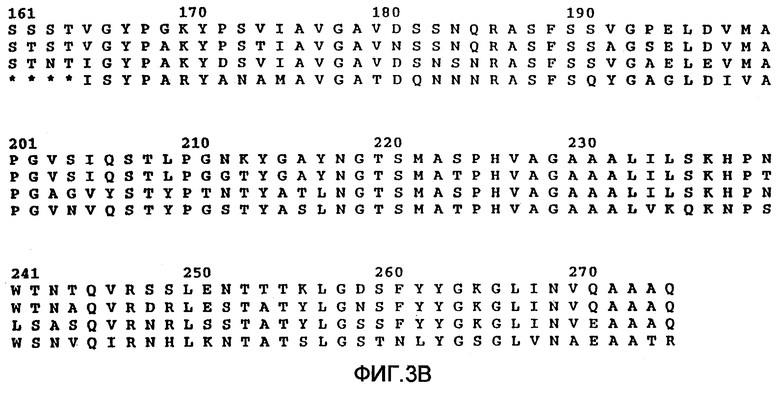
На фиг.3А и 3В показаны аминокислотные последовательности четырех субтилизинов. Верхний ряд относится к аминокислотной последовательности субтилизина из Bacillus amyloliquefaciens (который иногда определяется как субтилизин BPN’). Второй ряд относится к аминокислотной последовательности субтилизина из Bacillus subtilis. Третий ряд относится к аминокислотной последовательности субтилизина из B.licheniformis. Четвертый ряд относится к аминокислотной последовательности субтилизина из Bacillus lentus (который именуется также субтилизином 309 в РСТ WO 89/06276). Символ * означает отсутствие специфических аминокислотных остатков по сравнению с субтилизином BPN’.
Подробное описание изобретения
Протеазы являются карбонильными гидролазами, которые обычно расщепляют пептидные связи белков или пептидов. В используемом здесь значении термин "протеаза" означает природную или рекомбинантную протеазу. К природным протеазам относятся гидролаза α-аминоацилпептида, гидролаза пептидиламинокислоты, ациламиногидролаза, сериновая карбоксипептидаза, металлокарбоксипептидаза, тиолпротеиназа, карбоксилпротеиназа и металлопротеиназа. В объем этого изобретения входят сериновые протеазы, металлопротеазы, тиолпротеазы и кислотные протеазы, а также эндо- и экзопротеазы.
Настоящее изобретение относится к ферментам протеазы, которые не являются природными вариантами карбонильной гидролазы (вариантами протеазы), так как они обладают другой протеолитической активностью, устойчивостью, специфичностью к субстрату, профилем рН и/или эффективностью по сравнению с карбонильной гидролазой-предшественником, из которой получена аминокислотная последовательность этого варианта. В частности, такие варианты протеазы содержат аминокислотную последовательность, не встречающуюся в природе, которую получают путем замещения нескольких аминокислотных остатков протеазы-предшественника другими аминокислотами. Протеаза-предшественник может быть природной или рекомбинантной протеазой.
В вариантах протеазы по настоящему изобретению могут быть замещены любые из девятнадцати природных L-аминокислот в указанных положениях аминокислотных остатков. Такие замещения могут быть произведены в субтилизине-предшественнике (прокариотический субтилизин, эукариотический субтилизин, субтилизин млекопитающих и т.д.). В этом описании изобретения разные аминокислоты обозначены общепринятыми одно- и трехбуквенными кодами. Такие коды приведены в справочнике Dale M.W. (1989), Molecular Genetics of Bacteria, John Wiley & Sons, Ltd., Appendix B.
Варианты протеазы по этому изобретению предпочтительно получают из субтилизина Bacillus. Более предпочтительно варианты протеазы получают из субтилизина Bacillus lentus и/или субтилизина 309.
Субтилизины являются протеазами бактерий или грибов, которые обычно расщепляют пептидные связи белков или пептидов. В используемом здесь значении термин "субтилизин" означает природный или рекомбинантный субтилизин. Известно, что разные виды микроорганизмов продуцируют и часто секретируют целый ряд природных субтилизинов. Аминокислотные последовательности этих субтилизинов не являются полностью гомологичными. Однако субтилизины, входящие в эту группу, обладают одинаковой или подобной протеолитической активностью. Этот класс сериновых протеаз имеет общую аминокислотную последовательность, определяющую каталитическую триаду, которая отличает их от класса сериновых протеаз, родственных химотрипсину. Как субтилизины, так и родственные химотрипсину сериновые протеазы имеют каталитическую триаду, состоящую из аспартата, гистидина и серина. В протеазах, родственных субтилизину, эти аминокислоты при считывании от аминоконца к карбоксильному концу расположены в следующем относительном порядке: аспартат-гистидин-серин. Однако родственные химотрипсину протеазы имеют следующий относительный порядок расположения этих аминокислот: гистидин-аспартат-серин. Таким образом, субтилизин относится к сериновой протеазе, имеющей каталитическую триаду родственных субтилизину протеаз. Примерами таких субтилизинов являются, но не ограничиваются ими, субтилизины, показанные на фиг.3. Нумерация этих аминокислот в протеазах обычно и в соответствии с целями настоящего изобретения соответствует номерам, присвоенным последовательности зрелого субтилизина Bacillus amyloliquefaciens, изображенной на фиг.1.
"Рекомбинантный субтилизин" или "рекомбинантная протеаза" означают субтилизин или протеазу, в которых последовательность ДНК, кодирующая субтилизин или протеазу, модифицирована с возможностью продуцирования варианта (или мутанта) последовательности ДНК, кодирующей замещение, делецию или вставку одной или нескольких аминокислот в природной аминокислотной последовательности. Приемлемые способы выполнения такой модификации, которые можно использовать в сочетании со способами, описанными в этом изобретении, включают способы, описанные в патентах США №№ RE 34606, 5204015, 5185258, 5700676, 5801038 и 5763257.
"Субтилизины, не принадлежащие человеку" и кодирующие их ДНК можно получить из многих прокариотических и эукариотических микроорганизмов. Приемлемыми примерами прокариотических микроорганизмов являются грамотрицательные микроорганизмы, такие как E.coli или Pseudomonas, и грамположительные бактерии, такие как Micrococcus или Bacillus. Примерами эукариотических микроорганизмов, из которых может быть получен субтилизин и его гены, являются дрожжи, такие как Saccharomyces cerevisiae, и грибы, такие как Aspergillus sp.
"Вариант протеазы" имеет аминокислотную последовательность, полученную из аминокислотной последовательности "протеазы-предшественника". Протеазы-предшественники являются природными и рекомбинантными протеазами. Аминокислотную последовательность варианта протеазы получают из аминокислотной последовательности протеазы-предшественника путем замещения, делении или вставки одной или нескольких аминокислот в аминокислотной последовательности предшественника. Это достигается путем модификации "последовательности ДНК-предшественника", кодирующей аминокислотную последовательность протеазы-предшественника, а не манипуляцией с ферментом протеазы-предшественника per se. Приемлемые способы такой модификации последовательности ДНК-предшественника включают описанные здесь способы, а также способы, известные специалистам в этой области (см., например, европейский патент №0328299, WO 89/06279 и приведенные выше патенты и заявки на патент США).
В этом описании изобретения идентифицированы специфические замещения аминокислот в положениях одного или нескольких остатков, соответствующих положениям остатков, выбираемым из группы, включающей положения 62, 212, 230, 232, 252 и 257 субтилизина Bacillus amyloliquefaciens.
Предпочтительными вариантами являются варианты с комбинацией замещений в положениях остатков, соответствующих положениям субтилизина Bacillus amyloliquefaciens в таблице 1.
Более предпочтительными вариантами являются варианты с комбинациями замещений в положениях остатков, соответствующих положениям субтилизина Bacillus amyloliquefaciens в таблице 2.
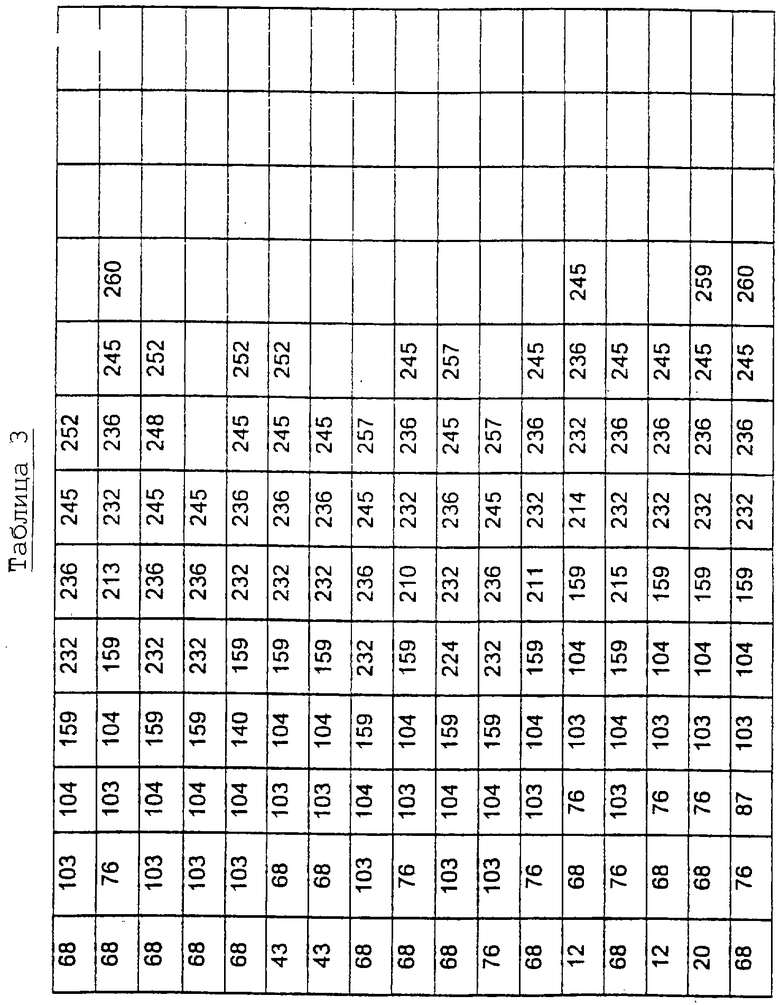
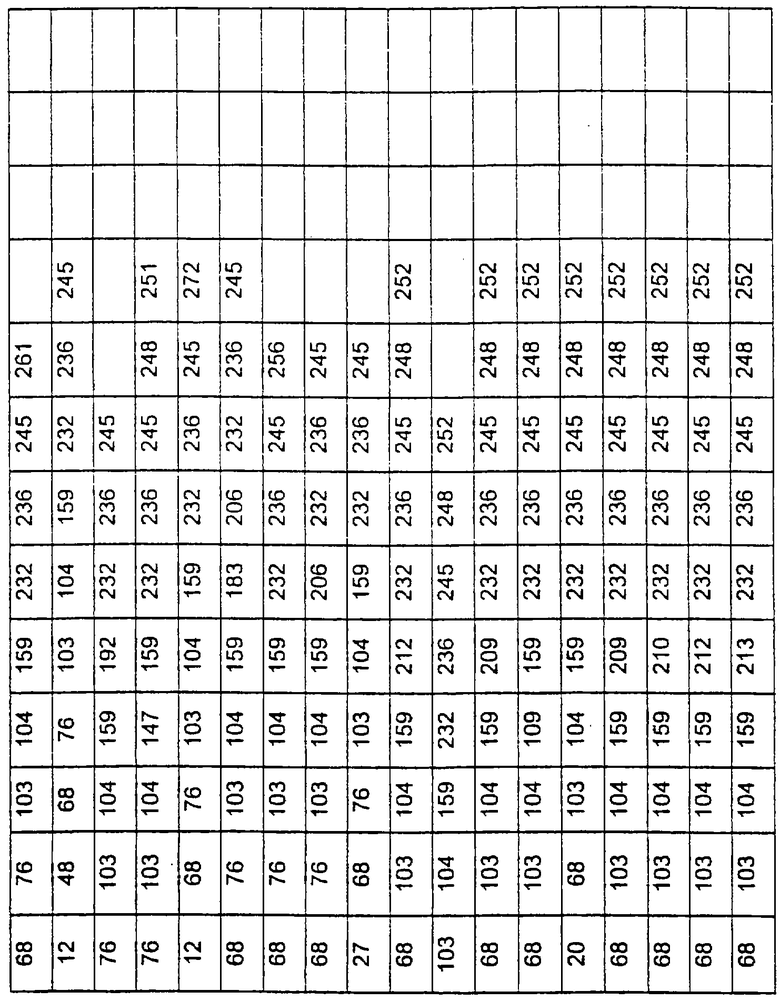
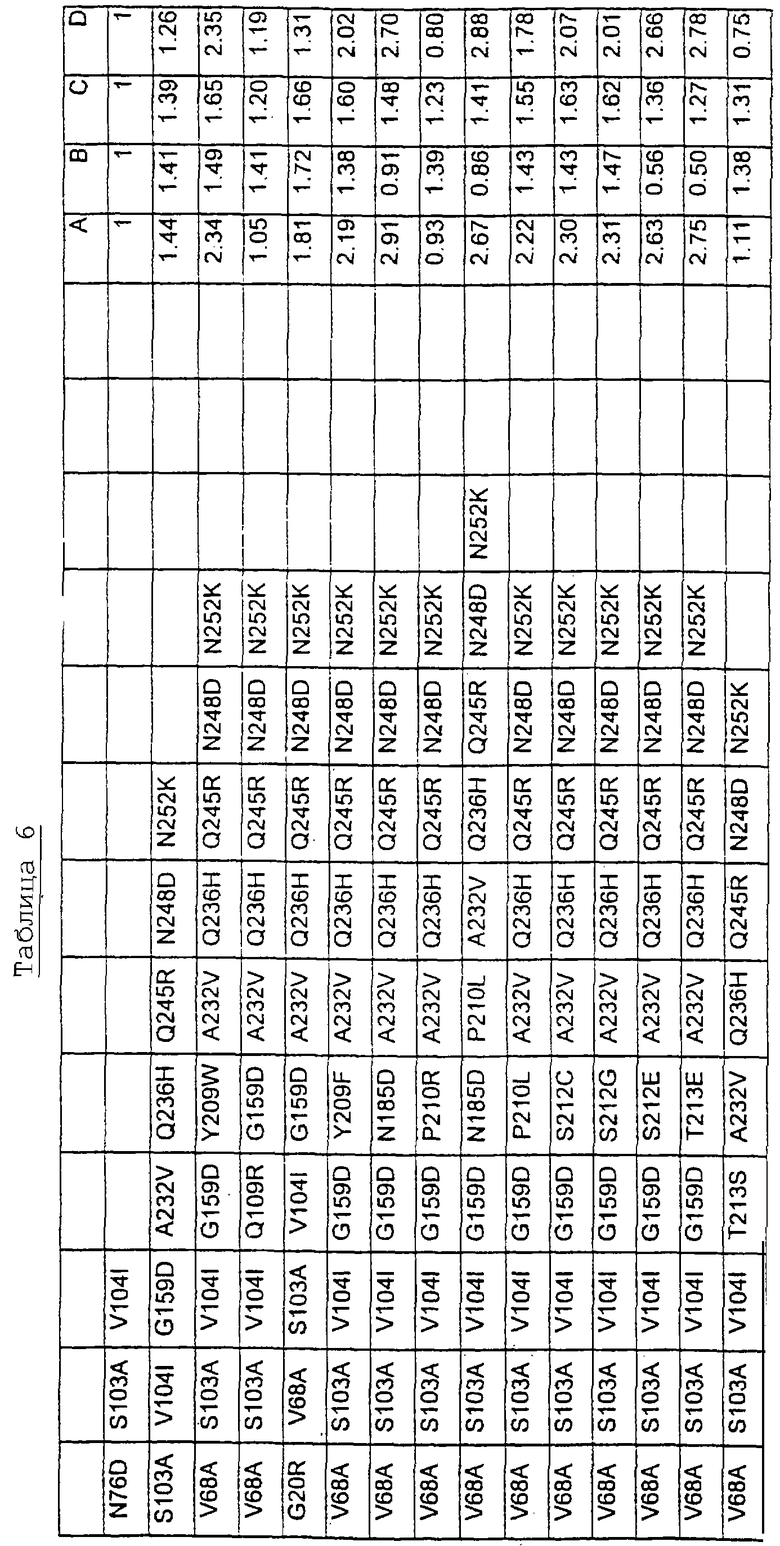
Другими предпочтительными вариантами являются варианты с комбинациями замещений в положениях остатков, соответствующих положениям субтилизина Bacillus amyloliquefaciens в таблице 3.
Номера положений этих аминокислот относятся к номерам, присвоенным последовательности зрелого субтилизина Bacillus amyloliquefaciens, показанной на фиг.1. Однако это изобретение не ограничивается зрелой формой этого субтилизина и включает протеазы-предшественники, содержащие аминокислотные остатки в положениях, которые "эквивалентны" конкретным идентифицированным остаткам в субтилизине Bacillus amyloliquefaciens. В предпочтительном варианте осуществления настоящего изобретения протеаза-предшественник является субтилизином Bacillus lentus, и замещения произведены в положениях, эквивалентных аминокислотным остаткам в B.lentus, которые соответствуют указанным выше.
Положение остатка (аминокислоты) протеазы-предшественника эквивалентно положению остатка субтилизина Bacillus amyloliquefaciens, если он гомологичен (то есть соответствует положению в первичной или третичной структуре) или аналогичен конкретному остатку или части этого остатка в субтилизине Bacillus amyloliquefaciens (то есть обладает такой же или подобной функциональной способностью соединяться, реагировать или химически взаимодействовать).
Для установления гомологии с первичной структурой аминокислотную последовательность протеазы-предшественника сравнивают непосредственно с первичной последовательностью субтилизина Bacillus amyloliquefaciens и, в частности, с набором остатков, которые, как известно, являются неизменяемыми в субтилизинах и определяют характер последовательности. Например, на фиг.2 показаны консервативные остатки, которые являются общими для субтилизинов Bacillus amyloliquefaciens и B.lentus. После выполнения сравнительного анализа консервативных остатков, позволяющего определить вставки и делеции, необходимые для сохранения первичной структуры (то есть во избежание удаления консервативных остатков в результате случайных делеции и инсерции), определяют остатки, эквивалентные определенным аминокислотам в первичной последовательности субтилизина Bacillus amyloliquefaciens. Сравнение консервативных остатков предпочтительно должно выявить наличие 100% таких остатков. Однако наличие более 75% или не менее 50% консервативных остатков также является достаточным для определения эквивалентных остатков. Каталитическая триада, Asp32/His64/Ser221, должна быть полностью сохранена. Siezen et al. (1991) Protein Eng. 4(7):719-737 выполнили сравнительный анализ большого количества сериновых протеаз. Siezen et al. определяют эту группу как субтилазы или субтилизинподобные сериновые протеазы.
Например, на фиг.3 показан сравнительный анализ аминокислотных последовательностей субтилизина из Bacillus amyloliquefaciens, Bacillus subtilis, Bacillus licheniformis (carlsbergensis) и Bacillus lentus, целью которого является выявление максимальной гомологии между этими аминокислотными последовательностями. Сравнение этих последовательностей показывает, что в каждой последовательности имеется несколько консервативных остатков. Эти консервативные остатки (для BPN' и B.lentus) представлены на фиг.2.
Таким образом, эти консервативные остатки можно использовать для определения соответствующих эквивалентных аминокислотных остатков субтилизина Bacillus amyloliquefaciens в других субтилизинах, таких как субтилизин из Bacillus lentus (публикация РСТ № WO 89/06279 от 13 июля 1989 г.), предпочтительный фермент протеазы-предшественника, или субтилизин, определяемый как РВ92 (европейский патент №0328299), который гомологичен предпочтительному субтилизину Bacillus lentus. На фиг.3А и 3В показан сравнительный анализ аминокислотных последовательностей некоторых из этих субтилизинов с последовательностью субтилизина Bacillus amyloliquefaciens, позволяющий выявить максимальную гомологию консервативных остатков. Как видно, в последовательности Bacillus lentus имеется ряд делеций по сравнению с субтилизином Bacillus amyloliquefaciens. Так, например, аминокислота для Vall65 в субтилизине Bacillus amyloliquefaciens эквивалентна изолейцину в других субтилизинах В.lentus и B.licheniformis.
"Эквивалентные остатки" можно также установить, определяя гомологию на уровне третичной структуры для протеазы-предшественника, третичная структура которой исследована рентгеновской кристаллографией. Эквивалентными остатками являются такие остатки, для которых атомные координаты двух или более атомов в главной цепи определенного аминокислотного остатка протеазы-предшественника и субтилизина Bacillus amyloliquefaciens (N перекрывает N, СА перекрывает СА, С перекрывает С и О перекрывает О) после упорядочения находятся в пределах 0,13 нм и предпочтительно в пределах 0,1 нм. Упорядочение первичной структуры достигается тогда, когда ориентация и расположение лучшей модели обеспечивает максимальное перекрывание атомных координат атомов белка, не являющихся водородом, у рассматриваемой протеазы и субтилизина Bacillus amyloliquefaciens. Лучшая модель является кристаллографической моделью, которая позволяет получить наименьший R-фактор для экспериментальных данных дифракции при самом высоком разрешении.
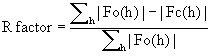
Эквивалентные остатки, которые функционально аналогичны специфическому остатку Bacillus amyloliquefaciens, определяют в качестве аминокислот протеазы-предшественника, которые могут принять такую конформацию, при которой они могут изменять, модифицировать или стимулировать образование белковой структуры, связывание с субстратом или катализ аналогично определенному и свойственному специфическому остатку субтилизина Bacillus amyloliquefaciens. Кроме того, такие остатки являются остатками протеазы-предшественника (третичная структура которой установлена рентгеновской кристаллографией), занимающими аналогичное положение, при котором атомные координаты, по крайней мере, двух атомов в боковой цепи остатка находятся на расстоянии 0,13 нм от соответствующих атомов в боковой цепи субтилизина Bacillus amyloliquefaciens, хотя атомы в главной цепи данного остатка могут не удовлетворять критерию эквивалентности с точки зрения гомологии положения. Координаты трехмерной структуры субтилизина Bacillus amyloliquefaciens приведены в публикации ЕРО №0251446 (соответствует патенту США №5182204, который включен в это описание изобретения в качестве ссылки), и, как указано выше, могут быть использованы для определения эквивалентных остатков на уровне третичной структуры.
Некоторые замещаемые остатки являются консервативными остатками, в то время как другие остатки такими не являются. В случае остатков, которые не являются консервативными, замещение одной или нескольких аминокислот ограничено замещениями, позволяющими получить вариант, имеющий аминокислотную последовательность, которая не соответствует природной последовательности. В случае консервативных остатков такие замещения не должны привести к образованию природной последовательности. Варианты протеазы по настоящему изобретению включают зрелые формы вариантов протеазы, а также про- и препроформы таких вариантов протеазы. Препроформы являются предпочтительной конструкцией, так как они облегчают экспрессию, секрецию и созревание вариантов протеазы.
"Пропоследовательность" означает последовательность аминокислот, связанных с N-концевой частью зрелой формы протеазы, удаление которой вызывает появление "зрелой" формы протеазы. Многие протеолитические ферменты встречаются в природе в виде продуктов трансляции профермента, и при отсутствии посттрансляционного процессинга они экспрессируются в таком виде. Предпочтительной пропоследовательностью для получения вариантов протеазы является предполагаемая пропоследовательность субтилизина Bacillus amyloliquefaciens, хотя можно использовать и другие пропоследовательности протеазы.
"Сигнальная последовательность" или "препоследовательность" означает любую последовательность аминокислот, связанных с N-концевой частью протеазы или с N-концевой частью протеазы, которая может участвовать в секреции зрелой формы или проформы протеазы. Это определение сигнальной последовательности является функциональным и включает все аминокислотные последовательности, кодированные N-концевой частью гена протеазы, которые участвуют в секреции протеазы в естественных условиях. Такие последовательности использованы в настоящем изобретении для осуществления секреции описанных здесь вариантов протеазы. Одна приемлемая сигнальная последовательность содержит семь первых аминокислотных остатков сигнальной последовательности, выделенной из субтилизина Bacillus subtilis, слитых с остальной частью сигнальной последовательности субтилизина Bacillus lentus (ATCC 21536).
"Препро" форма варианта протеазы содержит зрелую форму протеазы, имеющей пропоследовательность, операбельно связанную с аминоконцом протеазы, и "пре" или "сигнальную" последовательность, операбельно связанную с аминоконцом пропоследовательности.
"Экспрессирующий вектор" означает конструкцию на основе ДНК, содержащую последовательность ДНК, которая операбельно связана с приемлемой регуляторной последовательностью, способной экспрессировать указанную ДНК в приемлемом хозяине. Такие регуляторные последовательности включают промотор для осуществления транскрипции, необязательную операторную последовательность для контроля транскрипции, последовательность, кодирующую сайты связывания рибосомы с приемлемой мРНК, и последовательности, контролирующие окончание транскрипции и трансляции. Вектор может быть плазмидой, фаговой частицей или просто потенциальной геномной вставкой. После введения приемлемому хозяину вектор может реплицироваться и функционировать независимо от генома хозяина, и в некоторых случаях он может интегрироваться в сам геном. В этом описании изобретения термины "плазмида" и "вектор" иногда использованы во взаимозаменяемом значении, так как плазмида является наиболее часто используемой формой вектора в настоящее время. Однако в объем этого изобретения входят и другие формы экспрессирующих векторов, которые выполняют эквивалентные функции и известны в этой области.
"Клетки-хозяева", используемые в настоящем изобретении, обычно являются прокариотическими или эукариотическими хозяевами, которые предпочтительно подвергают обработке в соответствии со способами, описанными в патенте США № RE 34606, в результате чего они утрачивают способность секретировать ферментативно активную эндопротеазу. Предпочтительной клеткой-хозяином для экспрессии протеазы является штамм BG2036 Bacillus, который не имеет ферментативно активной нейтральной протеазы и щелочной протеазы (субтилизина). Конструирование штамма BG2036 подробно описано в патенте США №5264366. Другими клетками-хозяевами для экспрессии протеазы являются Bacillus subtilis I168 (которые описаны также в патентах США №№34606 и 5264366, включенных в это описание изобретения в качестве ссылки), а также любой приемлемый штамм Bacillus, такой как B.licheniformis, B.lentus и т.д.
Клетки-хозяева трансформируют или трансфецируют векторами, сконструированными методами рекомбинантных ДНК. Такие трансформированные клетки-хозяева способны реплицировать векторы, кодирующие варианты протеазы, или экспрессировать требуемый вариант протеазы. В случае векторов, кодирующих пре- или препроформу варианта протеазы, такие варианты, будучи экспрессированными, обычно секретируются из клетки-хозяина в среду, содержащую клетки-хозяева.
Термин "операбельно связанный", используемый для описания взаимосвязи между двумя областями ДНК, означает, что они функционально связаны друг с другом. Например, препоследовательность операбельно связана с пептидом, если она действует в качестве сигнальной последовательности, участвующей в секреции зрелой формы белка, вызывая расщепление указанной сигнальной последовательности. Промотор операбельно связан с кодирующей последовательностью, если он контролирует транскрипцию последовательности; сайт связывания рибосомы операбельно связан с кодирующей последовательностью, если его положение делает возможной трансляцию.
Гены, кодирующие природную протеазу-предшественника, можно получить обычными методами, известными в этой области. Эти методы обычно включают синтез меченых зондов, имеющих мнимые последовательности, кодирующие области представляющей интерес протеазы, создание геномных библиотек из микроорганизмов, экспрессирующих протеазу, и скрининг этих библиотек в отношении требуемого гена путем гибридизации с зондами. Затем положительно гибридизирующие клоны картируют и секвенируют.
Клонированную протеазу используют для трансформации клетки-хозяина с целью экспрессии протеазы. Ген протеазы затем лигируют в плазмиду с большим количеством копий. Эта плазмида реплицируется в хозяевах, так как она содержит хорошо известные элементы, необходимые для репликации плазмиды: промотор, оперативно связанный с рассматриваемым геном (который может быть введен в виде гомологического промотора гена, если его распознает, то есть транскрибирует, хозяин), область терминации транскрипции и область полиаденилирования (которая необходима для обеспечения устойчивости мРНК, транскрибируемой хозяином из гена протеазы в определенных эукариотических клетках-хозяевах), которая является экзогенной или поддерживается эндогенной областью терминатора гена протеазы, и желательно селектируемый ген, в частности, ген устойчивости к антибиотикам, что делает возможным постоянное культивирование инфицированных плазмидой клеток-хозяев путем выращивания в средах, содержащих антибиотики. Плазмиды с большим количеством копий имеют также источник репликации для хозяина, что позволяет получать большие количества плазмид в цитоплазме без хромосомных ограничений. Однако настоящее изобретение относится также к интеграции нескольких копий гена протеазы в геном хозяина. Это облегчается прокариотическими и эукариотическими микроорганизмами, которые особенно подвержены гомологичной рекомбинации.
Ген может быть природным геном B.lentus. Альтернативно можно получить синтетический ген, кодирующий природную или мутантную протеазу-предшественника. В этом случае определяют последовательность ДНК и/или аминокислотную последовательность протеазы-предшественника. Затем синтезируют несколько перекрывающихся фрагментов синтетической одноцепочечной ДНК, которые после гибридизации и лигирования образуют синтетическую ДНК, кодирующую протеазу-предшественника. Пример конструирования синтетического гена приведен в примере 3 патента США №5204015, который включен в это описание изобретения в качестве ссылки.
После клонирования природного или синтетического гена протеазы-предшественника выполняют ряд модификаций, направленных на усиление синтеза гена по сравнению с природной протеазой-предшественником. Такие модификации включают продуцирование рекомбинантных протеаз, как это описано в патенте США № RE 34606 и в публикации ЕРО №0251446, а также продуцирование описанных здесь вариантов протеазы.
Можно использовать нижеследующий способ кассетного мутагенеза, который облегчает конструирование вариантов протеазы по настоящему изобретению, хотя возможно применение и других способов. Сначала получают природный ген, кодирующий протеазу, который секвенируют полностью или частично. Затем последовательность сканируют для нахождения точки, в которой желательно произвести мутацию (делецию, вставку или замещение) одной или нескольких аминокислот в кодированном ферменте. Последовательности, фланкирующие эту точку, исследуют на наличие сайтов рестрикции для замены короткого сегмента гена пулом олигонуклеотидов, которые в экспрессированном состоянии должны кодировать разные мутанты. Такие сайты рестрикции предпочтительно являются уникальными сайтами в гене протеазы, облегчающими замену сегмента гена. Однако можно использовать любой удобный сайт рестрикции, который не является избыточным в гене протеазы, при условии, что фрагменты гена, полученные в результате рестриктирования, можно вновь собрать в требуемую последовательность. Если сайты рестрикции отсутствуют в местах, расположенных на удобном расстоянии от выбранной точки (от 10 до 15 нуклеотидов), такие сайты можно создать, замещая нуклеотиды в гене так, чтобы в конечной конструкции не были изменены ни рамка считывания, ни кодированные аминокислоты. Мутацию гена с целью изменения его последовательности так, чтобы она соответствовала требуемой последовательности, осуществляют при помощи праймера М13 в соответствии с известными методами. Задача локализации приемлемых фланкирующих областей и определения необходимых изменений для получения двух удобных последовательностей в сайте рестрикции значительно облегчается благодаря наличию избыточного генетического кода, рестрикционной карты фермента гена и большого количества разных рестрикционных ферментов. Следует отметить, что при наличии удобного фланкирующего сайта рестрикции рассмотренный выше метод необходимо использовать только в случае фланкирующей области, не имеющей нужного сайта.
После клонирования природной или синтетической ДНК сайты рестрикции, фланкирующие предназначенные для мутации положения, расщепляют родственными рестрикционными ферментами и лигируют в ген несколько кассет олигонуклеотидов, комплементарных концевым областям. Этот метод позволяет упростить мутагенез, так как все олигонуклеотиды можно синтезировать так, чтобы они имели одинаковые сайты рестрикции, причем для создания сайтов рестрикции не требуются синтетические линкеры.
В используемом здесь значении термин "протеолитическая активность" означает скорость гидролиза пептидных связей на миллиграмм активного фермента. Известно много методов измерения протеолитической активности (K.M.Kalisz, "Microbial Proteinases", Advances in Biochemical Engineering/Biotechnology, A.Fiechter ed., 1988). Помимо или в качестве альтернативы модифицированной протеолитической активности у вариантов ферментов по настоящему изобретению могут быть изменены другие свойства, такие как Кm, kcat, отношение kcat/Km, специфичность к субстрату и/или профиль активности в зависимости от рН. Эти ферменты могут быть предназначены для определенного субстрата, который, как предполагается, будет иметь место, например, при получении пептидов или в гидролитических процессах, применяемых при стирке.
В одном из аспектов этого изобретения целью является получение варианта протеазы с измененными, предпочтительно улучшенными, моющими характеристиками по сравнению с протеазой-предшественником, по крайней мере, в одном моющем составе и/или, по крайней мере, в одном режиме стирки.
Условия стирки, воздействию которых может подвергаться вариант протеазы, являются весьма разнообразными, включая разные моющие средства, разный объем моечной воды, разную температуру моечной воды и продолжительность стирки. Например, моющие составы, используемые в разных регионах, характеризуются разными концентрациями моющих компонентов в моечной воде. Например, детергенты, применяемые в Европе, обычно содержат около 4500-5000 частей на миллион моющих компонентов в моечной воде, в то время как детергенты, применяемые в Японии, обычно содержат примерно 667 частей на миллион моющих компонентов в моечной воде. В Северной Америке, в частности в США, детергенты обычно содержат около 975 частей на миллион моющих компонентов в моечной воде.
Система с низкой концентрацией детергента включает детергенты, характеризующиеся наличием менее 800 частей на миллион моющих компонентов в моечной воде. Японские детергенты обычно считаются системами с низкой концентрацией детергента, так как они содержат примерно 667 частей на миллион моющих компонентов в моечной воде.
Система со средней концентрацией детергента включает детергенты, характеризующиеся наличием от около 800 до около 2000 частей на миллион моющих компонентов в моечной воде. Детергенты, используемые в Северной Америке, обычно считаются системами со средней концентрацией детергента, так как они содержат примерно 975 частей на миллион моющих компонентов в моечной воде. В Бразилии обычно используются детергенты, содержащие примерно 1500 частей на миллион моющих компонентов в моечной воде.
Система с высокой концентрацией детергента включает детергенты, характеризующиеся наличием более 2000 частей на миллион моющих компонентов в моечной воде. Европейские детергенты обычно считаются системами с высокой концентрацией детергента, так как они содержат примерно 4500-5000 частей на миллион моющих компонентов в моечной воде.
Детергенты, используемые в Латинской Америке, обычно являются высокопенными, модифицированными фосфатом детергентами, поэтому их можно классифицировать как системы со средней и высокой концентрацией детергента, так как они характеризуются наличием от 1500 до 6000 частей на миллион моющих компонентов в моечной воде. Как указывалось выше, бразильские детергенты обычно содержат примерно 1500 частей на миллион моющих компонентов в моечной воде. Однако в других регионах, для которых характерно применение высокопенных, модифицированных фосфатом детергентов и которые не ограничиваются странами Латинской Америки, могут использоваться системы с высокой концентрацией детергента, содержащие до 6000 частей на миллион моющих компонентов в моечной воде.
В свете вышеизложенного становится очевидным тот факт, что концентрации детергентов в типичных моечных растворах, используемых в различных регионах мира, изменяются от менее 800 частей на миллион ("регионы применения систем с низкой концентрацией детергента"), например около 667 частей на миллион в Японии, до около 800-2000 частей на миллион ("регионы применения систем со средней концентрацией детергента"), например около 975 частей на миллион в США и около 1500 частей на миллион в Бразилии, и более 2000 частей на миллион ("регионы применения систем с высокой концентрацией детергента"), например около 4500-5000 частей на миллион в Европе и около 6000 частей на миллион в регионах применения высокопенных, модифицированных фосфатом детергентов.
Концентрации обычных моечных растворов определяются эмпирически. Например, в США обычная стиральная машина вмещает примерно 64,4 л моечного раствора. Таким образом, чтобы получить в моечном растворе концентрацию, равную примерно 975 частям на миллион детергента, в 64,4 л моечного раствора нужно добавить примерно 62,79 г моющей композиции. Именно это количество отмеривает потребитель при помощи мерного стаканчика, прилагаемого к детергенту.
В качестве еще одного примера можно привести использование в разных регионах разной температуры стирки. Температура моечной воды в Японии обычно ниже, чем в Европе.
Согласно одному из аспектов настоящее изобретение включает вариант протеазы, который обладает более высоким моющим действием, по крайней мере, в одном режиме стирки.
Другой целью этого изобретения является замещение аминокислоты в положениях одного или нескольких остатков, соответствующих положениям остатков, выбираемым из группы, включающей положения 62, 212, 230, 232, 252 и 257 субтилизина Bacillus amyloliquefaciens, что имеет важное значение для улучшения моющего действия данного фермента.
Эти замещения предпочтительно производят в субтилизине Bacillus lentus (рекомбинантного или нативного типа), хотя подобные замещения могут быть выполнены в любой протеазе Bacillus.
На основании результатов, полученных при исследовании вариантов протеаз, можно отметить, что указанные мутации в субтилизине Bacillus amyloliquefaciens имеют важное значение для протеолитической активности, эффективности и/или устойчивости этих ферментов, а также для очищающего или моющего действия таких вариантов ферментов.
Многие варианты протеазы по этому изобретению полезны для получения разных моющих композиций или средств личной гигиены, таких как шампуни или лосьоны. В композициях, содержащих мутанты протеазы по этому изобретению, можно использовать целый ряд известных соединений, являющихся приемлемыми поверхностно-активными веществами. Эти вещества являются неионогенными, анионными, катионными или цвиттерионными детергентами, описанными в патенте США №4404128, выданном Barry J.Anderson, и в патенте США №4261868, выданном Jiri Flora et al. Приемлемый моющий состав описан в примере 7 патента США №5204015 (ранее включенного в это описание изобретения в качестве ссылки). В этой области известны разные составы, которые можно использовать в качестве очищающих композиций. Понятно, что помимо обычных очищающих композиций варианты протеазы по этому изобретению могут быть использованы в любой области, где применяются нативные протеазы или протеазы дикого типа. Так, эти варианты можно использовать, например, при применениях твердого или жидкого мыла, составов для мытья посуды, растворов или продуктов для очистки контактных линз, составов для гидролиза пептидов, утилизации отходов, обработки тканей, а также в качестве расщепляющих ферментов при получении белков и т.д. Варианты по настоящему изобретению могут сообщать моющему составу более высокую эффективность (по сравнению с предшественником). Более высокая эффективность детергента определяется как лучшее выведение пятен, восприимчивых к конкретному ферменту, таких как пятна растительного происхождения или пятна крови, по методу обычной оценки после стандартного цикла стирки.
Протеазы по этому изобретению могут входить в состав известных порошкообразных и жидких детергентов с рН от 6,5 до 12,0 и концентрацией от около 0,01 вес.% до около 5 вес.% (предпочтительно от 0,1% до 0,5%). Эти очищающие композиции с детергентами могут содержать другие ферменты, в частности известные протеазы, амилазы, целлюлазы, липазы или эндогликозидазы, а также добавки и стабилизаторы.
Введение протеаз по этому изобретению в обычные очищающие композиции не связано с какими-либо особыми ограничениями. Другими словами, любые значения температуры и рН, приемлемые для данного детергента, приемлемы также для композиций по настоящему изобретению, если показатель рН находится в пределах вышеуказанного диапазона и данная температура ниже температуры денатурации описанной протеазы. Кроме того, протеазы по этому изобретению можно использовать в очищающей композиции без детергентов, отдельно или в сочетании с добавками и стабилизаторами.
Настоящее изобретение относится также к очищающим композициям, содержащим варианты протеазы по этому изобретению. Очищающие композиции могут дополнительно содержать добавки, которые обычно используются в таких композициях. Эти добавки включают, но не ограничиваются ими, отбеливатели, поверхностно-активные вещества, модифицирующие компоненты, ферменты и катализаторы отбеливания. Специалисту в этой области должно быть очевидно, что добавки выбирают с учетом их пригодности для данной композиции. Приведенный здесь список не является исчерпывающим и должен рассматриваться только в качестве примеров приемлемых добавок. Специалисту в этой области должно быть также понятно, что необходимо использовать только те добавки, которые совместимы с ферментами и другими компонентами композиции, например с поверхностно-активным веществом.
Количество добавки в очищающей композиции, если она используется, составляет от около 0,01% до около 99,9%, предпочтительно от около 1% до около 95%, более предпочтительно от около 1% до около 80%.
Варианты протеаз по настоящему изобретению могут быть введены в корм для животных в качестве части добавок, как это описано, например, в патентах США №№5612055, 5314692 и 5147642.
Одним из аспектов этого изобретения является композиция для обработки ткани, включающая варианты протеаз по настоящему изобретению. Эту композицию можно использовать для обработки, например, шелка или шерсти, как это описано в таких публикациях, как RD 216034, европейский патент №134267, патент США №4533359 и европейский патент №344259.
Далее настоящее изобретение иллюстрируется примерами, которые не ограничивают объем нижеследующей формулы изобретения.
Все приведенные здесь публикации и патенты полностью включены в это описание изобретения в качестве ссылки.
ПРИМЕР 1
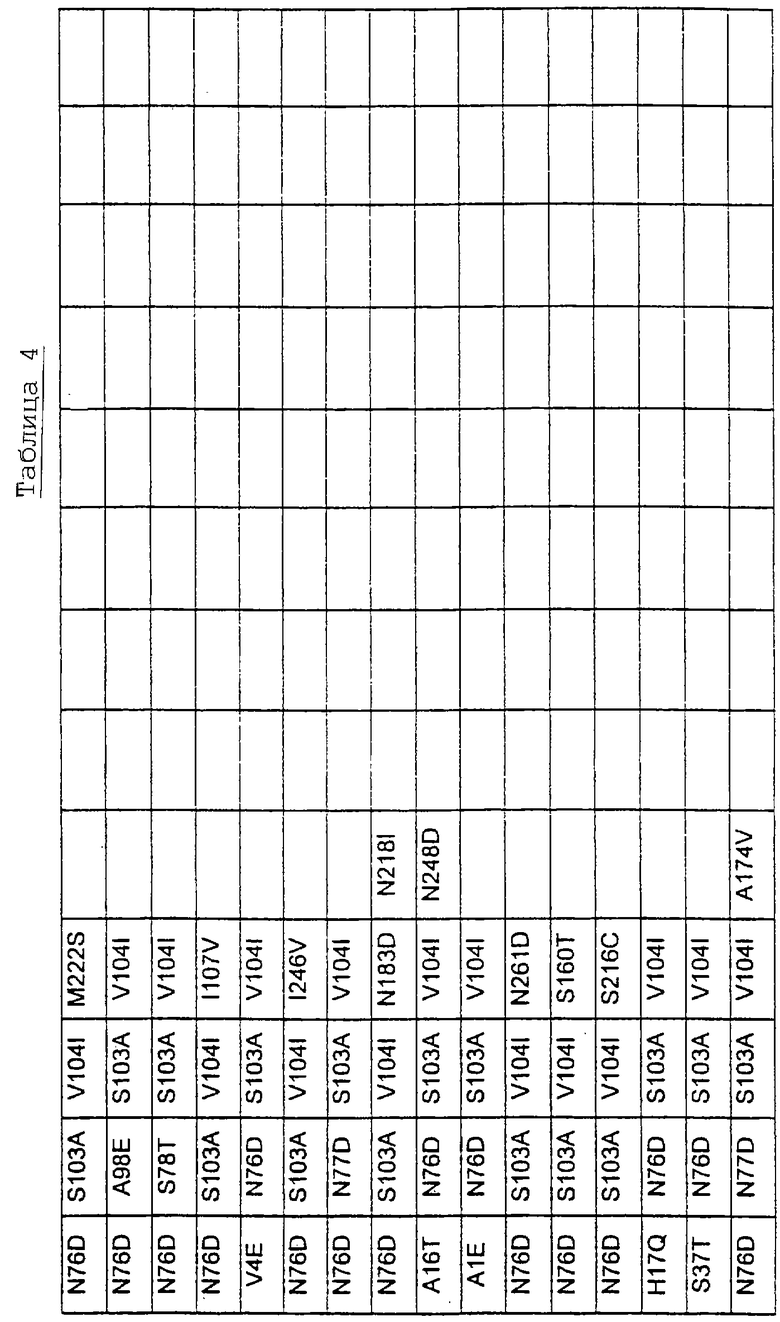
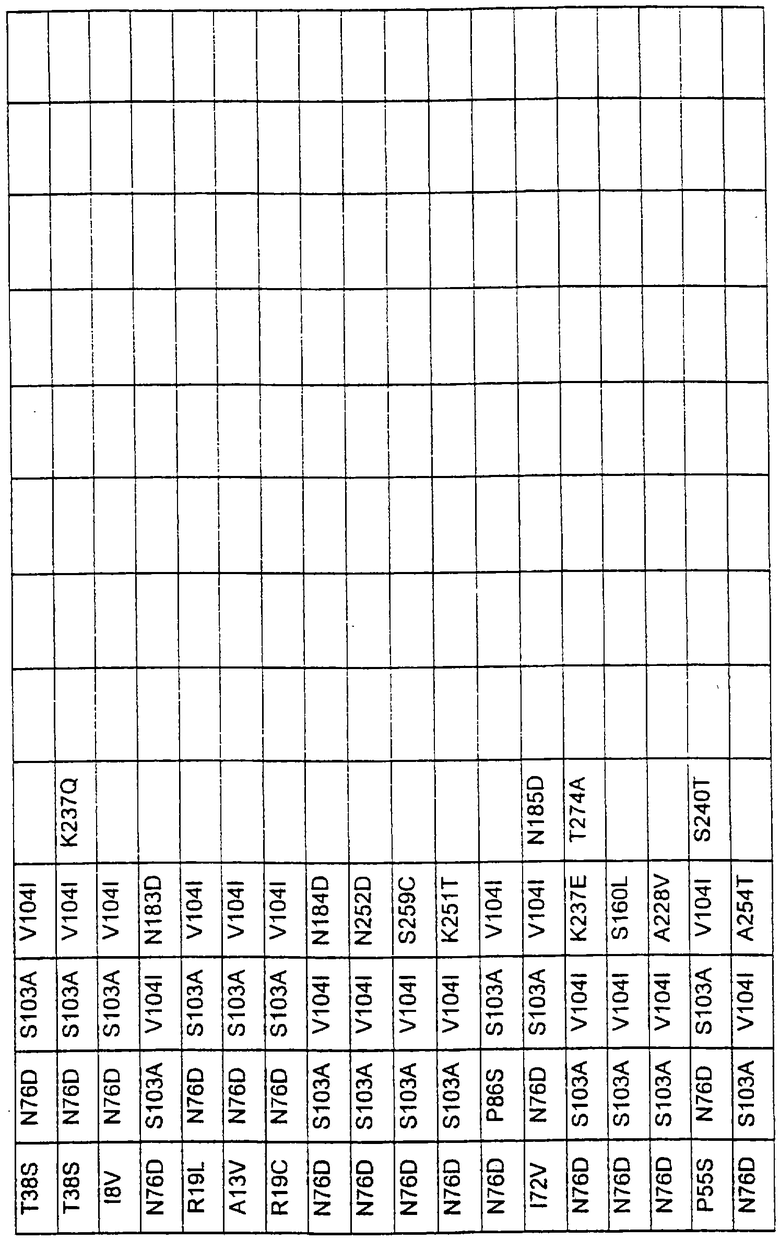
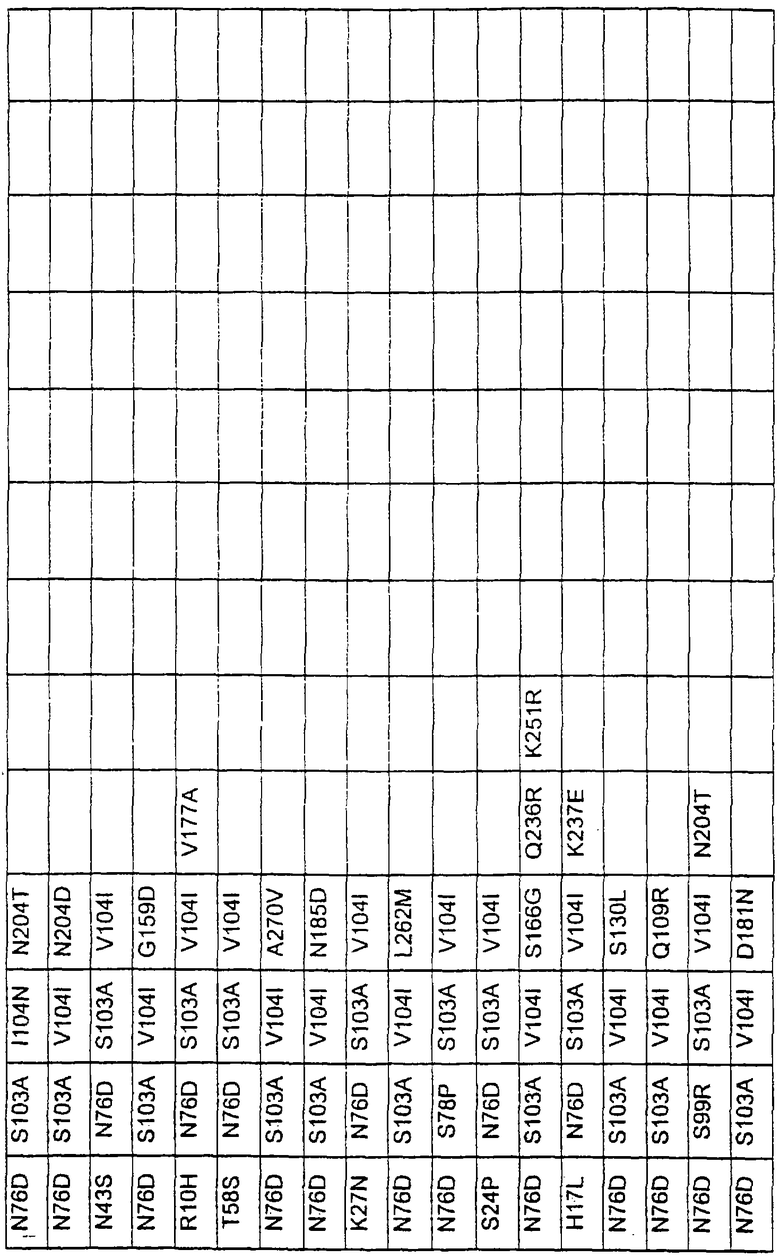
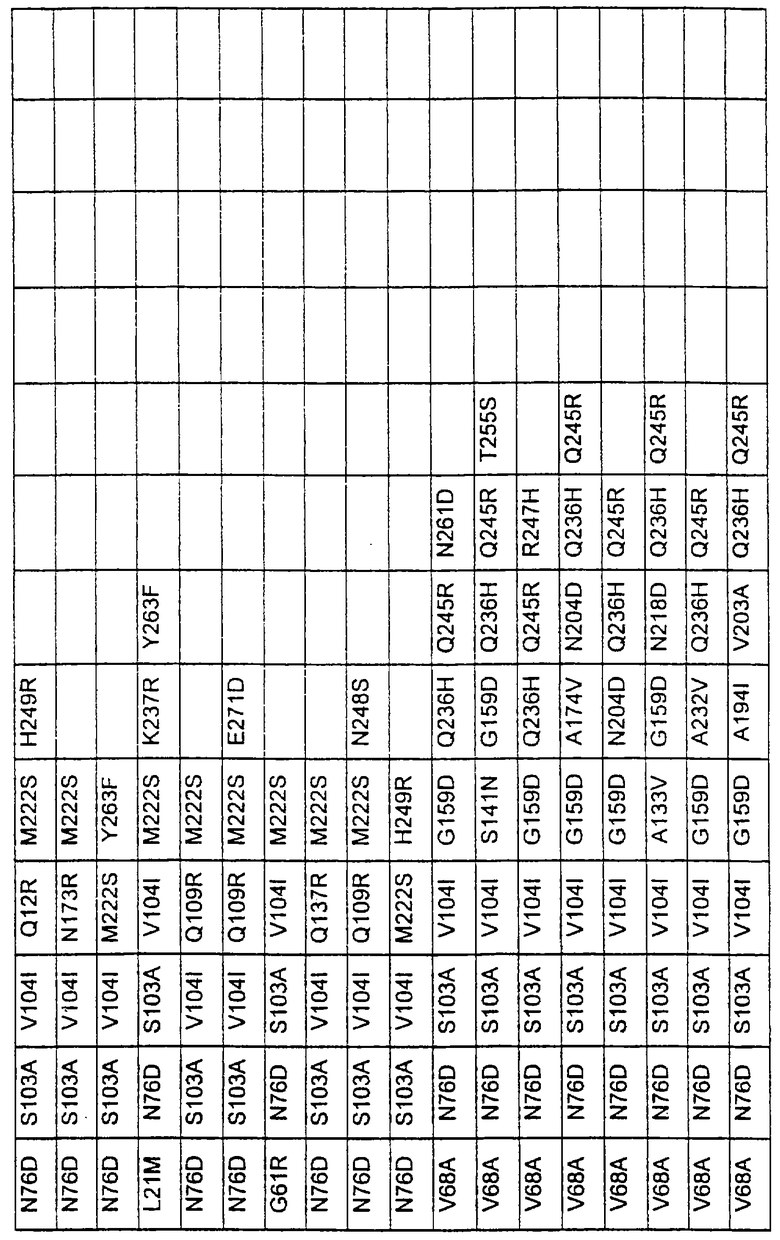
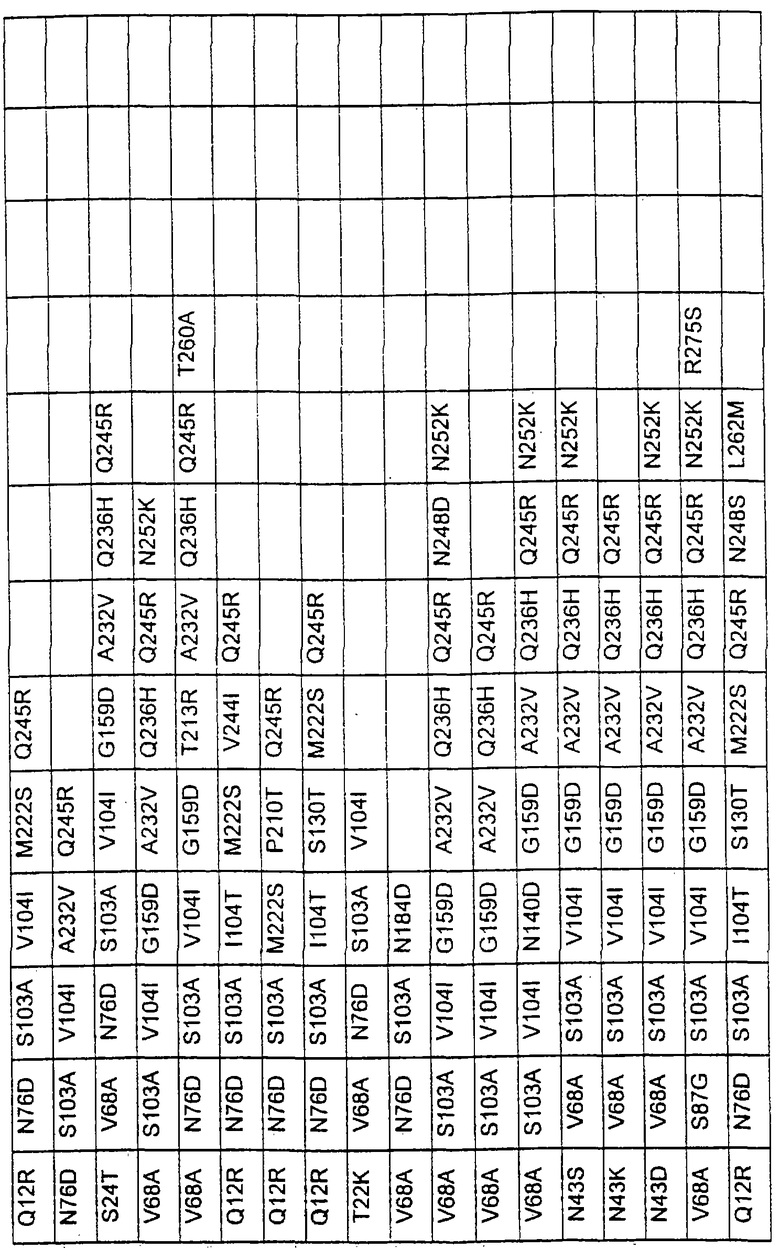
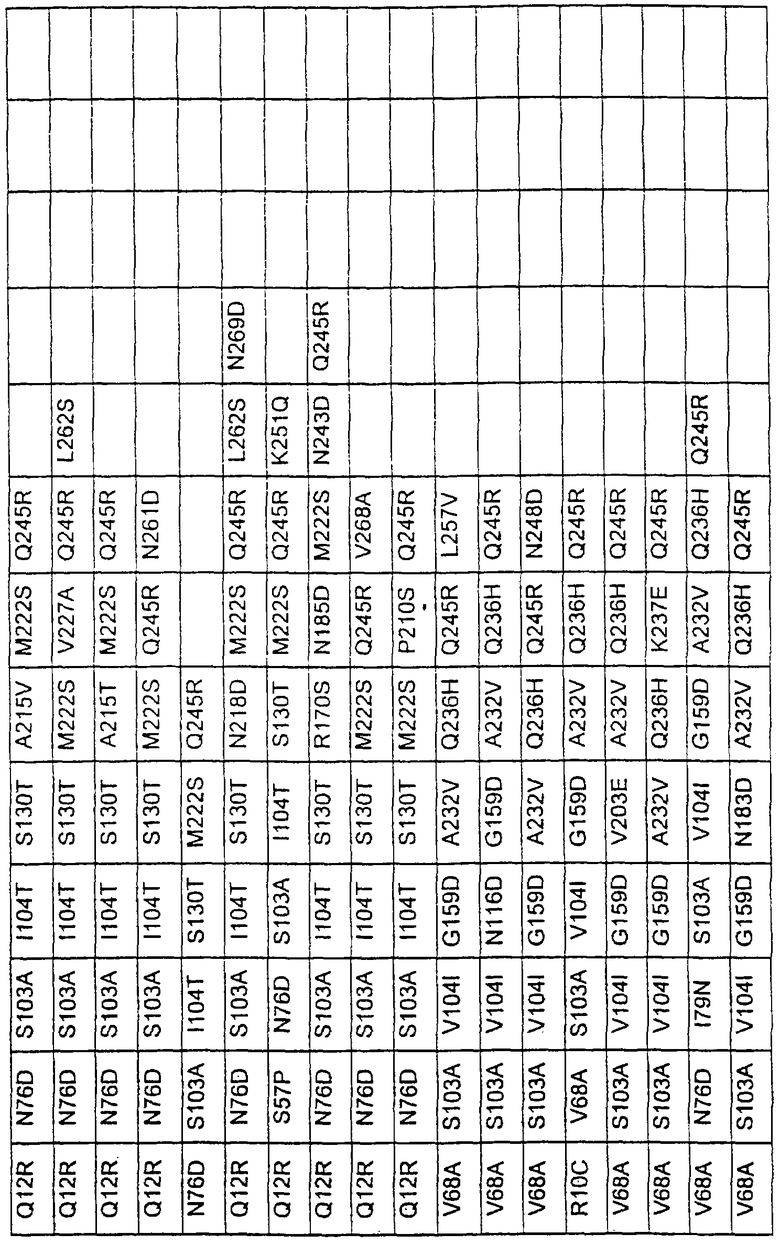
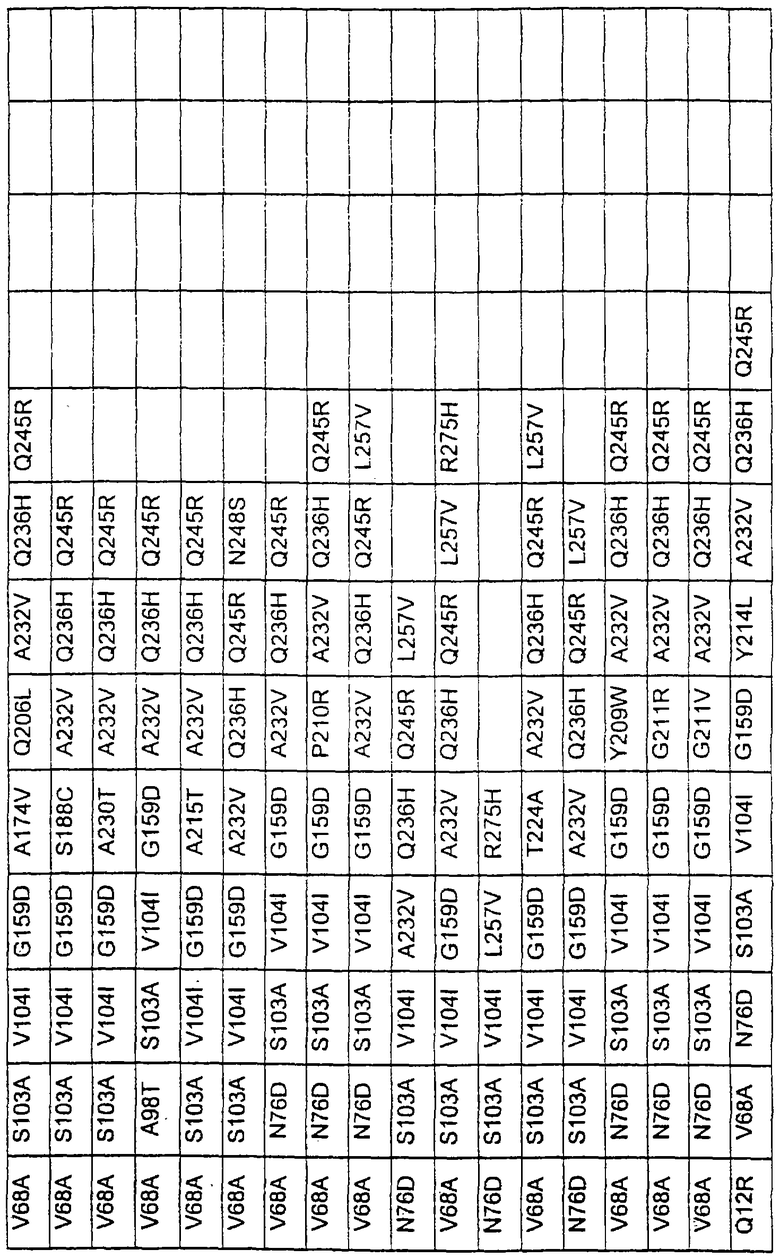
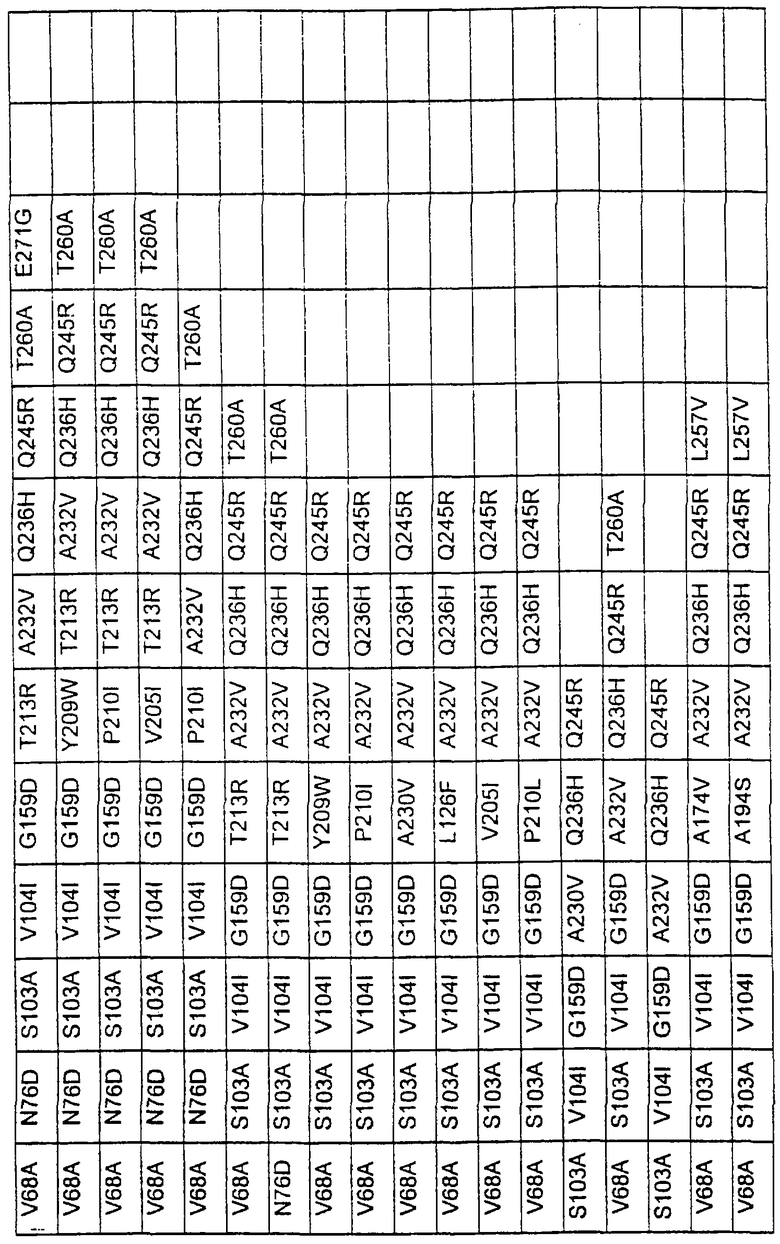
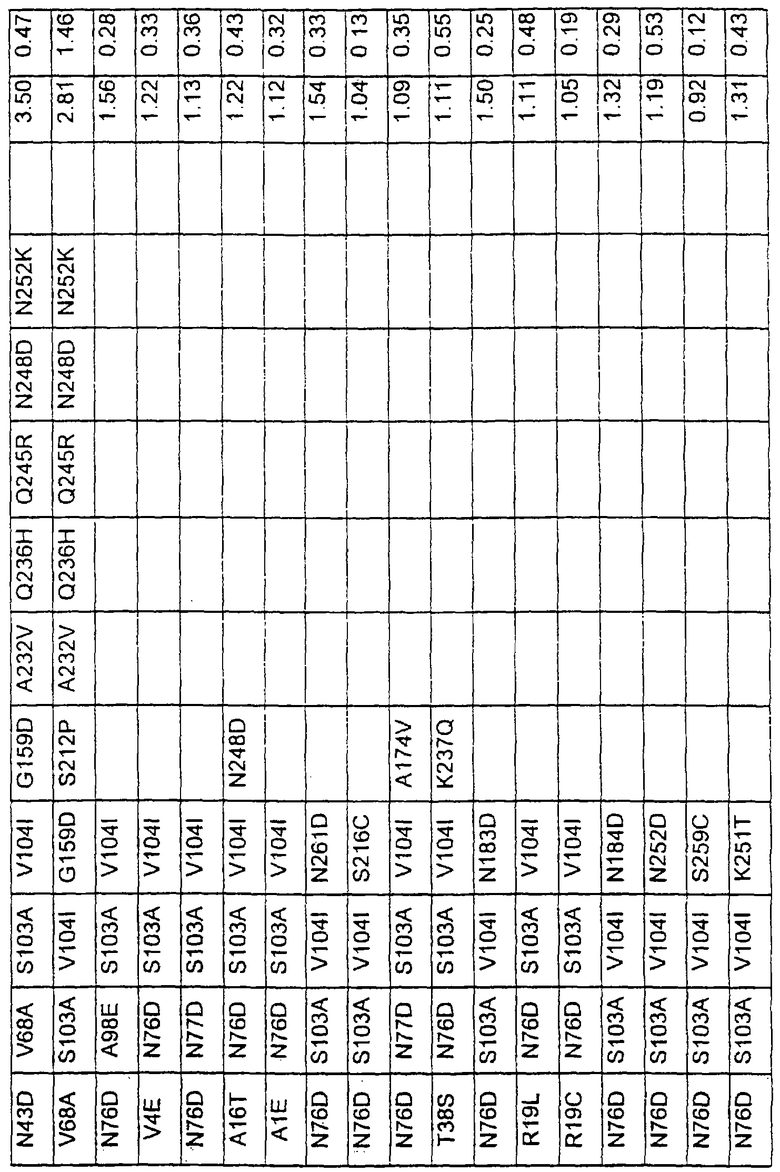
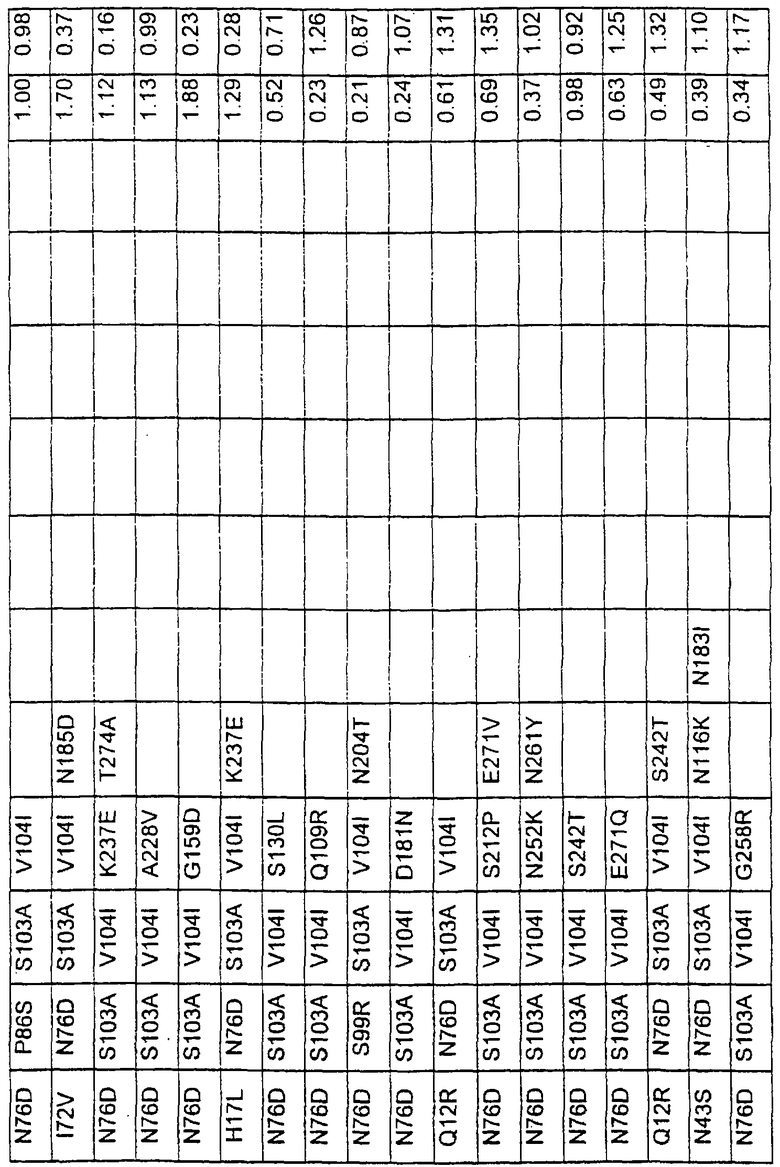
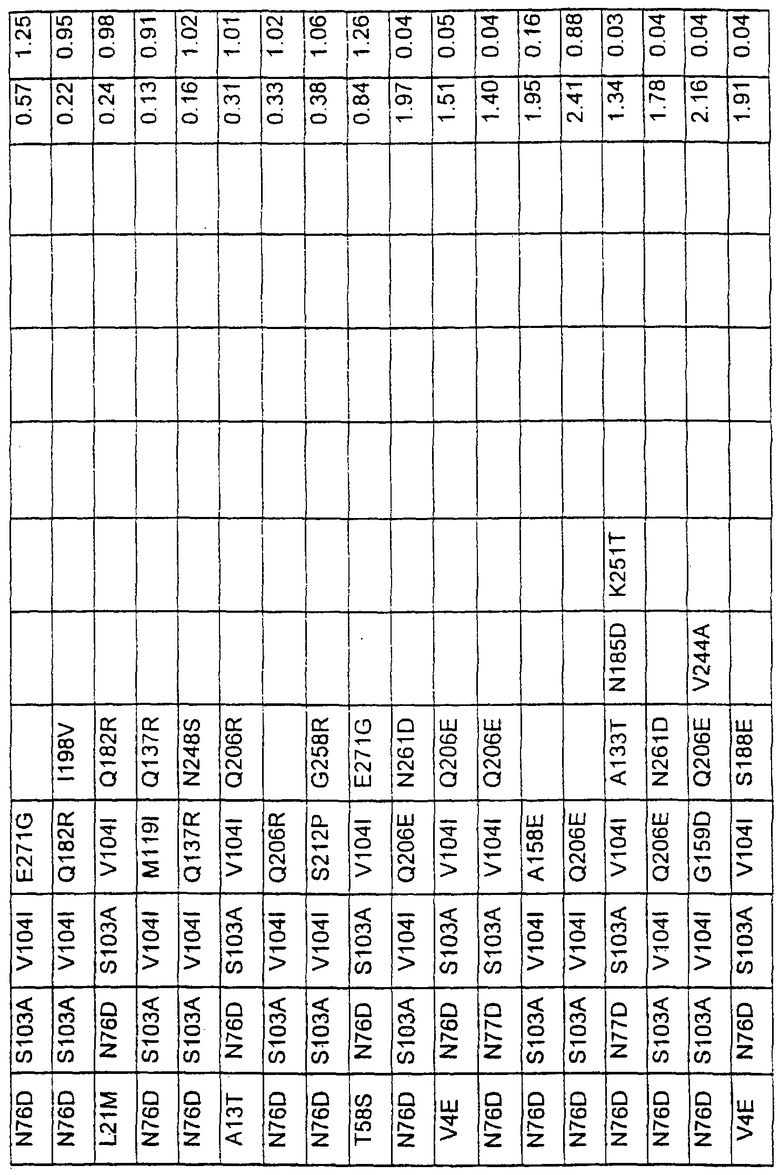
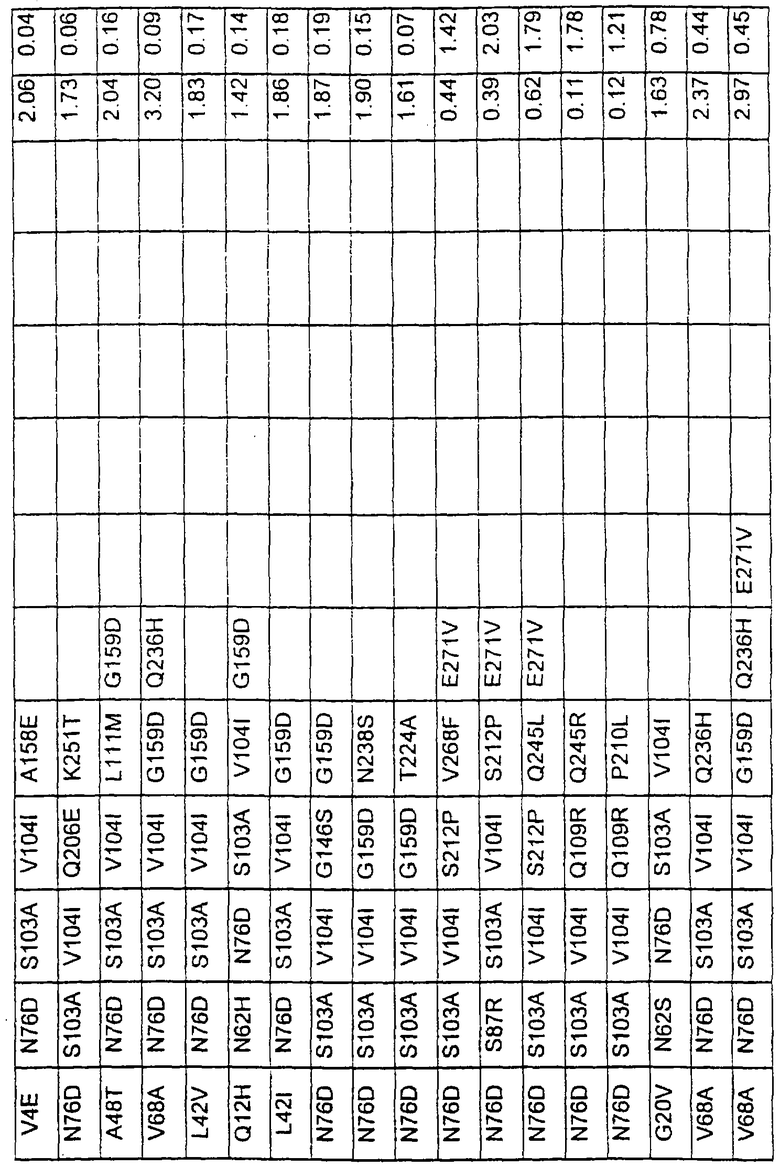
Большое количество вариантов протеазы получено и очищено при помощи методов, хорошо известных в этой области. Все мутации произведены в субтилизине GG36 Bacillus lentus. Эти варианты приведены в таблице 4.
ПРИМЕР 2
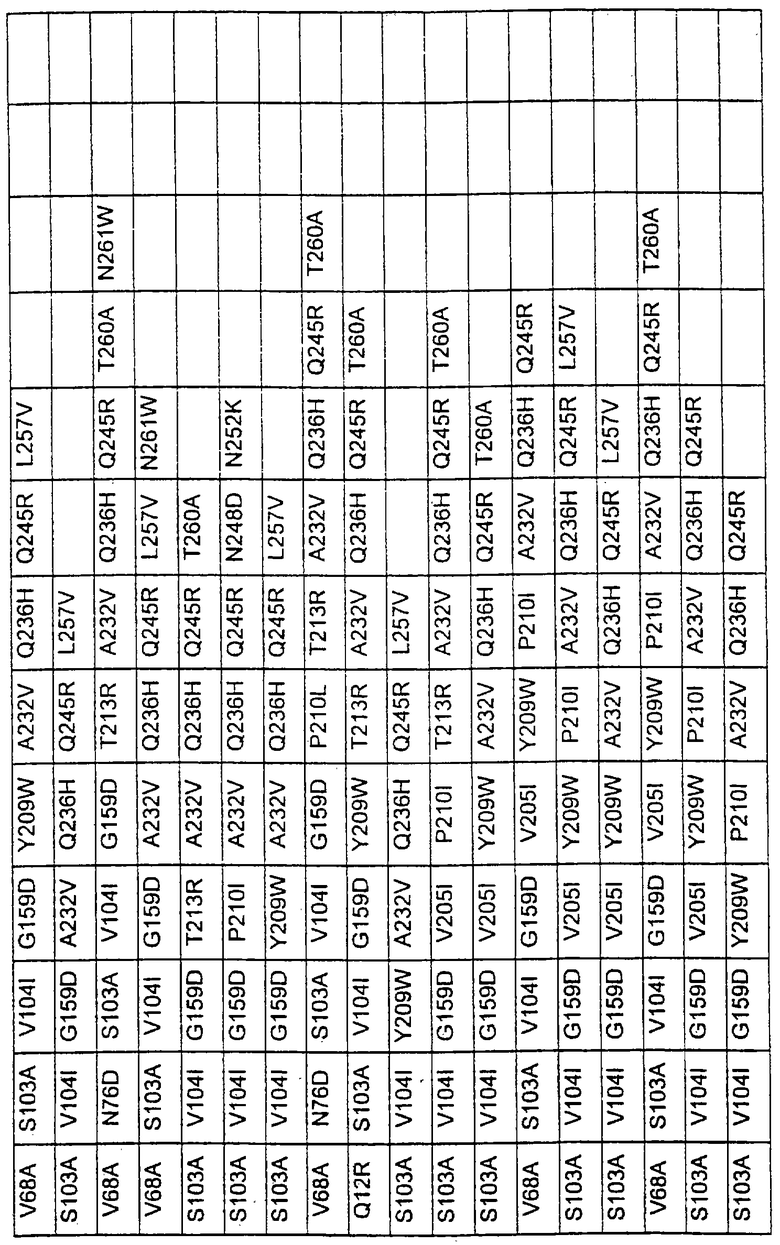
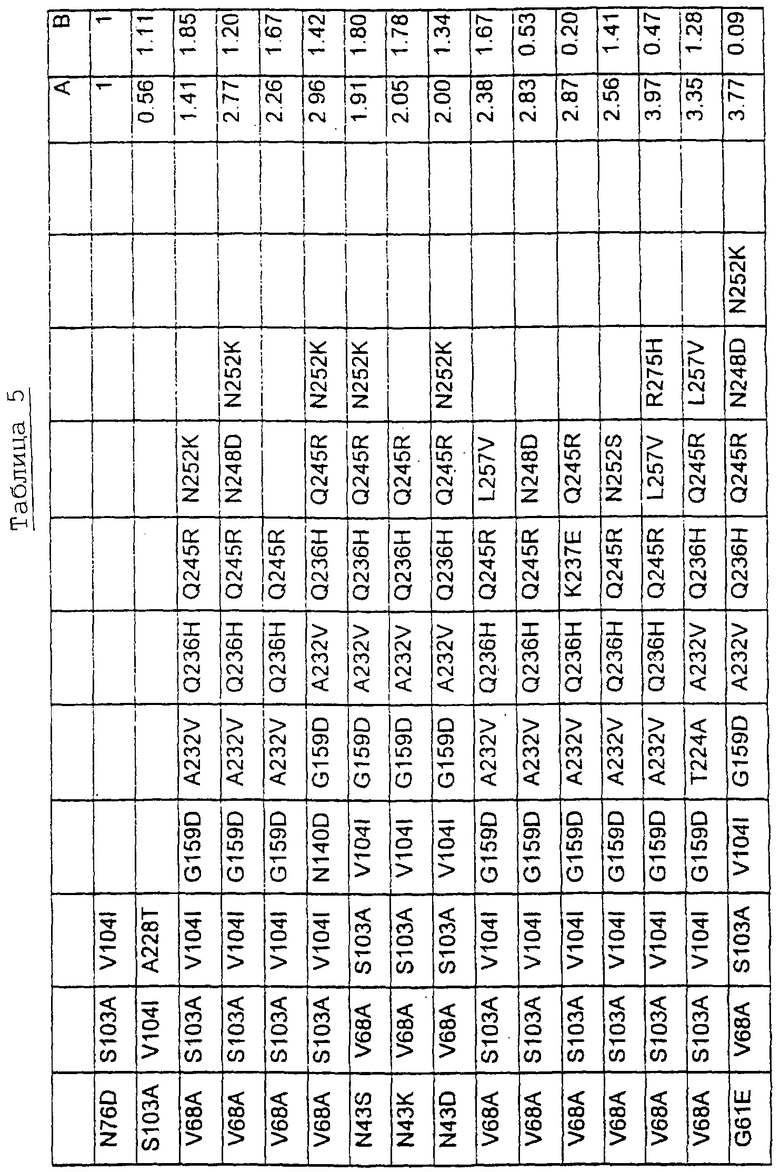
Большое количество вариантов протеазы, полученных в примере 1, испытывали на эффективность в двух типах детергентов и условий стирки при помощи микроскопического анализа, описанного в патенте США №60/068796 "An improved method of assaying for a preferred enzyme and/or preferred detergent composition".
В таблице 5 представлены подвергнутые анализу варианты протеаз и результаты испытания в двух разных детергентах. В столбце А приведены данные для фильтрованного детергента Ariel Ultra (Procter & Gamble, Цинциннати, шт.Огайо, США) с концентрацией 0,67 г/л в растворе, содержащем 42,78 г/м3 (3 грана/галлон) смешанной жесткости Са2+/Мg2+ и 0,3 части на миллион фермента в каждой емкости при 20°С. В столбце В приведены данные для фильтрованного детергента Ariel Futur (Procter & Gamble, Цинциннати, шт.Огайо, США) с концентрацией 3,38 г/л в растворе, содержащем 213,9 г/м3 (15 гран/галлон) смешанной жесткости Са2+/Мg2+ и 0,3 части на миллион фермента в каждой емкости при 40°С.
ПРИМЕР 3
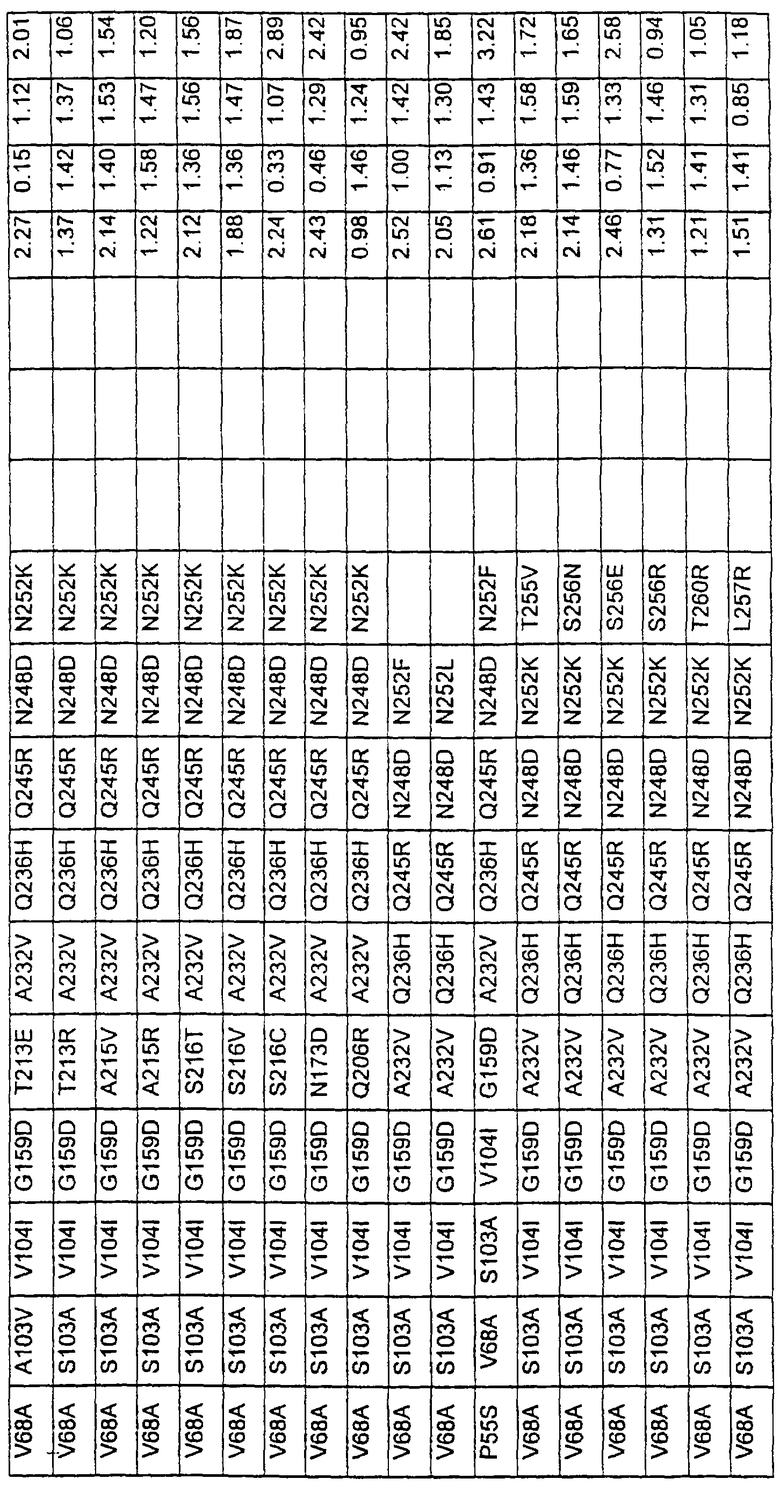
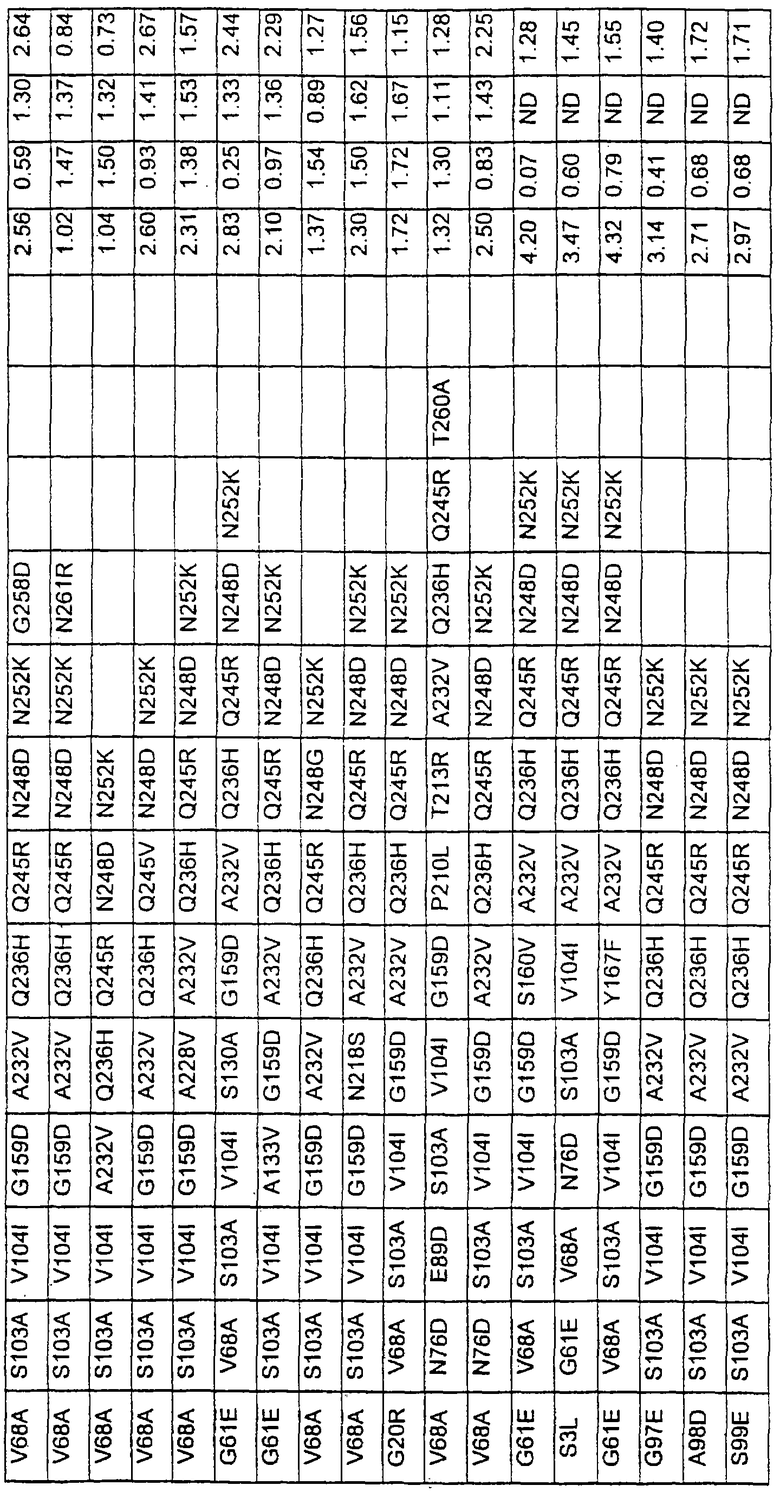
В таблице 6 представлены подвергнутые анализу варианты протеаз по примеру 1 и результаты испытаний в четырех разных детергентах. Указанные варианты протеаз были подвергнуты тем же испытаниям на эффективность, что и в примере 2, с использованием нижеследующих детергентов. В столбце А приведены данные для фильтрованного детергента Ariel Ultra (Procter & Gamble, Цинциннати, шт.Огайо, США) с концентрацией 0,67 г/л в растворе, содержащем 42,78 г/м3 (3 грана/галлон) смешанной жесткости Са2+/Мg2+ и 0,3 части на миллион фермента в каждой емкости при 20°С. В столбце В приведены данные для фильтрованного детергента Ariel Futur (Procter & Gamble, Цинциннати, шт.Огайо, США) с концентрацией 3,38 г/л в растворе, содержащем 213,9 г/м3 (15 гран/галлон) смешанной жесткости Са2+/Мg2+ и 0,3 части на миллион фермента в каждой емкости при 40°С. В столбце С приведены данные для детергента с концентрацией 3,5 г/л HSP1 (Procter & Gamble, Цинциннати, шт.Огайо, США) в растворе, содержащем 114,08 г/м3 (8 гран/галлон) смешанной жесткости Са2+/Мg2+ и 0,3 части на миллион фермента в каждой емкости при 20°С. В столбце D приведены данные для детергента с концентрацией 1,5 г/л Tide КТ (Procter & Gamble, Цинциннати, шт.Огайо, США) в растворе, содержащем 42,78 г/м3 (3 грана/галлон) смешанной жесткости Са2+/Мg2+ и 0,3 части на миллион фермента в каждой емкости при 20°С.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТЫ ПРОТЕАЗЫ, ЗАМЕЩЕННЫЕ В НЕСКОЛЬКИХ ПОЛОЖЕНИЯХ | 1998 |
|
RU2269572C2 |
| ВАРИАНТ СУБТИЛИЗИНА (ВАРИАНТЫ), КОДИРУЮЩАЯ ЕГО ДНК, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, ОЧИЩАЮЩАЯ КОМПОЗИЦИЯ, КОРМ ДЛЯ ЖИВОТНЫХ, КОМПОЗИЦИЯ ДЛЯ ОБРАБОТКИ ТКАНИ | 1998 |
|
RU2252958C2 |
| МОДИФИЦИРОВАННЫЙ СУБТИЛИЗИН, ДНК, КОДИРУЮЩАЯ МОДИФИЦИРОВАННЫЙ СУБТИЛИЗИН, ВЕКТОР ЭКСПРЕССИИ, КОДИРУЮЩИЙ ДНК, И ШТАММ КУЛЬТУРЫ КЛЕТКИ ХОЗЯИНА | 1994 |
|
RU2136756C1 |
| Способы и композиции, содержащие варианты сериновой протеазы | 2012 |
|
RU2663114C2 |
| ВАРИАНТЫ ПРОТЕАЗ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2790443C2 |
| ПРОТЕАЗЫ, СОДЕРЖАЩИЕ ОДНУ ИЛИ НЕСКОЛЬКО КОМБИНИРУЕМЫХ МУТАЦИЙ | 2009 |
|
RU2560978C2 |
| ВАРИАНТЫ СУБТИЛАЗ | 2010 |
|
RU2651525C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ВКЛЮЧАЮЩИЕ В СЕБЯ ПРИМЕНЕНИЕ ВАРИАНТНЫХ МИКРОБНЫХ ПРОТЕАЗ | 2009 |
|
RU2575073C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ВКЛЮЧАЮЩИЕ ВАРИАНТЫ МИКРОБНЫХ ПРОТЕАЗ | 2009 |
|
RU2541786C2 |
| БЕЛКИ, ВЫЗЫВАЮЩИЕ ИЗМЕНЕННУЮ ИММУНОГЕННУЮ РЕАКЦИЮ, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ИСПОЛЬЗОВАНИЯ | 2002 |
|
RU2311458C2 |




Изобретение относится к области генной и белковой инженерии и может быть использовано при создании новых моющих средств и очищающих составов. Предложены мутантные формы субтилизина Bacillus, являющиеся результатом определенной комбинации замен в аминокислотной последовательности нативного фермента, от которой они происходят, и отличающиеся от субтилизина-предшественника повышенной эффективностью очищающего действия. Все варианты субтилизина по изобретению характеризуются либо заменой валином остатка в положении, соответствующем положению 232 аминокислотной последовательности субтилизина Bacillus amyloliquifaciens, либо заменой пролином остатка в положении 212 указанной природной формы субтилизина. Для получения предложенных новых форм фермента соответствующие мутантные последовательности ДНК экспрессированы в клетках-хозяевах, предпочтительно в клетках штаммов Bacillus с пониженным уровнем активности протеаз. Приобретаемые вариантами субтилизина в результате модификации свойства обеспечивают возможность их успешного применения в составе любых очищающих композиций, предусматривающих включение протеаз. 5 с. и 1 з.п. ф-лы, 5 ил., 6 табл.
N76D/S103A/V104I/S212P/E271V;
N76D/S103A/V104I/S212P/G258R;
N76D/S103A/V104I/S212P/V268F/E271 V, и
N76D/S87R/S103A/V104I/S212P/E271V.
Конвенционный приоритет по пп.1-6 установлен от 23.10.1997 согласно дате подачи заявок 08/956,323, 08/956,324 и 08/956,564 в Патентное ведомство США.
| Огнетушитель | 0 |
|
SU91A1 |
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |

