Настоящее изобретение относится к классу генов точки контроля, которые контролируют прогресс на протяжении клеточного цикла в эукариотических клетках.
Предпосылки создания изобретения
Контроль клеточного цикла является фундаментальным для роста и поддержания эукариотических организмов, от дрожжей до млекопитающих. Эукариотические клетки имеют развитые контрольные пути, названные "точками контроля" ("checkpoints"), которые обеспечивают, чтобы индивидуальные стадии клеточного цикла завершались до того, как происходит следующая стадия. В ответ на повреждение ДНК, клеточное выживание возрастает как за счет непосредственных механизмов восстановления ДНК, так и за счет замедления прогресса на протяжении клеточного цикла в эукариотических клетках. В зависимости от положения клетки в пределах цикла во время облучения, повреждение ДНК в клетках млекопитающего может препятствовать (а) переходу из GI в S фазу, (b) прогрессу на протяжении S фазы или (с) переходу из G2 в митоз. Предполагается, что такие точки контроля должны предотвращать разрушительные события такие, как репликация поврежденной ДНК и сегрегация фрагментированных хромосом во время митоза (Hartwell and Kastan, 1994).
Rad3 ген Schizosaccharomyces pombe требуется для точек контроля, которые отвечают за повреждение ДНК и репликацию блоков. Rad3 является членом липид киназного подкласса киназ, который обладает областями, имеющими последовательность, гомологичную липид киназному доменому р110 субъединицы фосфатидилинозитол-3 киназы (PI-3). Этот подкласс также включает АТМ белок, поврежденный у пациентов с атаксия-телеангиэктазией. Клетки от пациентов с атаксия телеангиэктазией (AT клетки) утрачивают замедление в S фазе после облучения и, как оказывается, проявляют радиоустойчивость при синтезе ДНК (Painter and Young, 1989). AT клетки, облученные в S фазе, аккумулируются в G2 с летальным повреждением, вероятно, как следствие стремления реплицировать поврежденную ДНК. AT клетки, облученные во время G2, проявляют различный фенотип: они не прекращают митоз после повреждения ДНК и развиваются на протяжении митоза с поврежденной ДНК (Beamish and Lavin, 1994). Мутации в локусе, к которому АТМ ген картирован, приводит таким образом к разрушению нескольких точек контроля, которые требуются для соответствующего ответа на ионизирующее облучение. Эти элементы липид киназного подкласса включают:
Tellp (Greenwell et al., 1985), ген, вовлеченный в поддержание собственной теломерной длины в Saccharomyces cerevisiae: Esrip: Meclp и продукт Drozophila melanogaster mei-41 ген точки контроля (Hari et al., 1995).
Раскрытие изобретения
Мы проанализировали S. pombe rad3 ген и нашли, что он имеет аминокислотную последовательность из 2386 аминокислот полной длины, но не из 1070 аминокислот, описанных Seaton et al., 1992. Определено, что он является прямым гомологом S. cerevisiae Esrip, и что он имеет ту же самую общую структуру, что и АТМ ген. С-терминальная область rad3 белка содержит липид киназный домен, который требуется для функции Rad3. Было показано, что Rad3 способен к самоассоциации. Мы также определили протеин киназную активность, связанную с Rad3.
Кроме того, мы обнаружили человеческий гомолог к rad3. Этот ген, который мы назвали ATR (атаксия и rad родственный "ataxia and rad"), проявляет значительно более высокую гомологию к rad3, чем он проявляет ее к АТМ гену.
ATR кДНК последовательность человека представляют в Seq.ID No.1. Аминокислотную последовательность ORF из нуклеотидов 80 и 80II представляют в Seq.ID.No.2.
ДНК последовательность открытой рамки считывания (ORF) rad3 показывают как Seq.ID No.3. 2386 аминокислотная трансляция гена (нуклеотиды 585-7742 Seq.ID Nо.3.) показывают как Seq.ID Nо.4.
Соответственно, в первом аспекте, изобретение представляет собой ATR белок Seq.ID No.2 и его гомологи, его полипептидные фрагменты, а также антитела, способные к связыванию ATR белка или его полипептидных фрагментов. ATR белки, гомологи и его фрагменты рассматриваются ниже как полипептиды изобретения.
В другом аспекте, настоящее изобретение представляет собой полинуклеотид в существенно изолированной форме, способный гибридизироваться селективно с Seq.ID No.1 или с его комплементом (т.е. противоположной цепью). Обеспечиваются также полинуклеотиды, кодирующие полипептиды изобретения. Такие полинуклеотиды будут рассматриваться как полинуклеотид изобретения. Полинуклеотиды изобретения включают ДНК Seq.ID Nos 1 и фрагменты их, способные селективно гибридизироваться с этим геном.
В следующем аспекте, изобретение представляет собой рекомбинантные векторы, несущие полинуклеотид изобретения, включая экспрессионные векторы, и способы роста таких векторов в подходящей клетке хозяине, например, при условиях, при которых происходит экспрессия белка или полипептида, кодируемого последовательностью изобретения.
В дополнительном аспекте, изобретение представляет собой наборы, включающие полинуклеотиды, полипептиды или антитела изобретения, и способы использования таких наборов в диагностике наличия или отсутствия ATR и его гомологов, или его вариантов, включая вредные ATR мутанты.
Изобретение, кроме того, представляет собой способы анализа для скринирования веществ кандидатов для применения в качестве соединений для ингибирования или активирования ATR акивности, или активности мутированных форм ATR, которые являются дефицитными в активности точки контроля. Изобретение также представляет собой способы анализа для скринирования веществ кандидатов для применения в качестве соединений для ингибирования взаимодействий между ATR и другими сединениями, которые взаимодействуют с ATR, включая и сам ATR.
В близком аспекте, изобретение представляет собой полинуклеотидную последовательнось Seq.ID Nо.3 в существенно выделенной форме, и белок Seq.ID No.4 в существенно выделенной форме, и их новые варианты и фрагменты.
Детальное описание изобретения
А. Полинуклеотиды
Полинуклеотиды изобретения могут включать ДНК или РНК. Они могут быть также полинуклеотидами, которые имеют в своем составе синтетические или модифицированные нуклеотиды. В данной области известен ряд различных типов модификаций до олигонуклеотидов. Они включают метилфосфонатную и фосфоротиоатную основные цепи, дополнительно акридиновую или полилизиновые цепи при 3’ и/или 5’ концах молекулы. Для целей настоящего изобретения должно быть понятно, что Полинуклеотиды, описанные здесь, могут быть модифицированы любым способом, который доступен в данной области. Такие модификации могут проводиться для того, чтобы повысить активность in vivo или продолжительность жизни полинуклеотидов изобретения.
Полинуклеотиды изобретения, способные селективно гибридизироваться с ДНК Seq.ID No.1, будут составлять, по крайней мере, 70%, предпочтительно, по крайней мере, 80 или 90%, и более предпочтительно, по крайней мере, 95% гомологичных к соответствующей ДНК Seq.ID No.1 в пределах области, по крайней мере, 20, предпочтительно, по крайней мере, 25 или 30, например, по крайней мере, 40, 60 или 100 или более, соседних нуклеотидов.
Должно быть понятно, что специалисты в этой области могут, используя известные технические приемы, осуществить нуклеотидные замещения, которые не затрагивают полипептидную последовательность, кодированную полинуклеотидами изобретения для отражения использования кодона любого конкретного организма хозяина, в котором полипептиды изобретения должны экспрессироваться.
Может быть использована любая комбинация вышеупомянутых степеней гомологии и минимальных размеров для определения полинуклеотидов изобретения, с более строгими комбинациями (т.е. более высокая гомология в пределах больших длин), являющихся предпочтительными. Так, например, полинуклеотид, который является, по крайней мере, на 80% гомологичным в пределах 25 нуклеотидов, предпочтительно 30 нуклеотидов, представляет один аспект изобретения, так же как и полинуклеотид, который является на 95% гомологичным в пределах 40 нуклеотидов.
Полинуклеотиды изобретения могут применяться для продуцирования праймера, например, PCR праймера, праймера для альтернативной реакции амплификации, пробы, например, меченной меткой, обнаруживаемой стандартными средствами с использованием радиоактивных или нерадиоактивных меток, или полинуклеотиды могут быть клонироваными в векторах. Такие праймеры, пробы и другие фрагменты будут представлять, по крайней мере, 15, предпочтительно, по крайней мере, 20, например, по крайней мере, 25, 30 или 40 нуклеотидов в длину, и охватываются также названными нуклеотидами изобретения, как они использованы здесь.
Полинуклеотиды, такие, как ДНК полинуклеотиды и праймеры согласно изобретению, могут продуцироваться рекомбинантно, синтетически, или любыми другими средствами, доступными специалистам в данной области. Они могут также клонироваться стандартными техническими приемами.
В основном, праймеры могут продуцироваться синтетическим способом, включая ступенчатое производство одного нуклеотида желаемой последовательности нуклеиновой кислоты за один раз. Технические приемы для выполнения этого способа, использующие автоматизированные способы, являются легко доступными в данной области.
Более длинные полинуклеотиды будут, в основном, продуцироваться с использованием рекомбинантных способов, например, использующих PCR (полимеразная цепная реакция) способы клонирования. Способ клонирования будет включать изготовление пары праймеров (например, около 15-30 нуклеотидов) с областью ATR гена, которая является желательной для клонирования, приведение праймеров в контакт с мРНК или кДНК, полученными из клетки человека (например, делением клетки такой, как периферический лейкоцит крови), выполнение реакции полимеразной цепи при условиях, которые возникают при амплификации желаемой области, выделение амплифицированного фрагмента (т.е. очисткой реакционной смеси на агарозном геле) и выделение амплифицированной ДНК. Праймеры могут конструироваться так, чтобы они содержали подходящие сайты узнавания фермента рестрикции, с тем чтобы амплифицированная ДНК могла клонироваться в пригодном клонирующем векторе.
Для получения всей или части ATR последовательности, могут использоваться технические приемы, описанные здесь. Могут быть также получены аналогичным способом геномные клоны, содержащие ATR ген и его интроны и промоторные области, начиная с геномной ДНК клетки человека, например, клетки печени.
Хотя, в основном, технические приемы, упомянутые здесь, хорошо известны в науке, ссылка может быть сделана, в частности, на Sambrook et al. (Molecular Cloning: A Laboratory Manual, 1989).
Полинуклеотиды, которые не являются на 100% гомологичными последовательностям настоящего изобретения, но попадают в объем изобретения, можно получать рядом приемов.
Другие аллельные варианты человека ATR последовательности, описанные здесь, могут быть получены, например, опробованием геномных ДНК библиотек, созданных из ряда представителей, например, представителей из различных популяций.
Кроме того, могут быть получены гомологи ATR других животных, особенно млекопитающих (например, мышей, крыс или кроликов), более конкретно приматов, и такие гомологи и их фрагменты, в основном, могут быть способны селективно гибридизироваться с Seq.ID No.1. Такие последовательности могут быть получены опробированием кДНК библиотек, созданных из разделенных клеток или тканей или геномных ДНК библиотек от других животных видов, и опробированием таких библиотек пробами, включающими всю или часть Seq.ID 1 в условиях среды с высокой строгостью (например, 0.03 М хлорида натрия и 0.03 М цитрата натрия при температуре от около 50°С до около 60°С).
Аллельные варианты и видовые гомологи могут быть также получены с использованием дегенеративной PCR, которая будет использовать праймеры, сконструированные для нацеливания последовательностей внутри вариантов и гомологов, кодирующих сохраненные аминокислотные последовательности. Сохраненные последовательноси могут быть предсказаны сравнением ATR аинокислотной последовательности с последовательностью rad3. Праймеры будут содержать одну или больше дегенеративных позиций и будут использоваться в строгих условиях более низких, чем те, которые применяются для клонирования последовательностей с единичной последовательностью праймеров по отношению к известным последовательностям.
Альтернативно, такие полинуклеотиды могут быть получены посредством сайт направленного мутагенеза ATR последовательностей или их аллельных вариантов. Это может применяться там, где, например, требуются изменения молчащего кодона для последовательностей, чтобы оптимизировать преимущества кодона для конкретной клетки хозяина, в которой экспрессируется полинуклеотидная последовательность. Могут быть желательны изменения других последовательностей для того, чтобы ввести узнаваемые сайты фермента рестрикции, или изменить свойство или функцию полипептидов, кодированных полинуклеотидами, Могут быть желательными дальнейшие изменения для того, чтобы представить особые кодирующие изменения, найденные в ATR, которые дают увеличение мутантных ATR генов, которые утратили функции точки контроля. Пробы, базирующиеся на таких изменениях, могут использоваться как диагностические пробы для обнаружения таких ATR мутантов.
Изобретение далее представляет двухцепочные полинуклеотиды, включающие полинуклеотид изобретения и его комплемент.
Полинуклеотиды или праймеры изобретения могут нести обнаруживаемую метку. Подходящие метки включают радиоизотопы, такие, как 32P или 35S, ферментные метки, или другие белковые метки, такие, как биотин. Такие метки могут добавляться к полинуклеотидам или праймерам изобретения и могут обнаруживаться с использованием технических средств, известных по существу.
Полинуклеотиды или праймеры изобретения или их фрагменты, меченные или немеченные, могут применяться специалистами в данной области в опытах на основе нуклеиновой кислоты для обнаружения или секвенирования ATR в теле человека или животного.
Такие опыты по обнаружению обычно включают приведение образца тела человека или животного, содержащего ДНК или РНК, в контакт с пробой, включающей полинуклеотид или праймер изобретения, при условиях гибридизации, и обнаружение любого дуплекса, образованного между пробой и нуклеиновой кислотой в образце. Такое обнаружение может быть достигнуто применением технических приемов, таких как PCR, или путем иммобилизации пробы на твердую подложку с удалением нуклеиновой кислоты, которая не гибридизируется с пробой, и затем определяя нуклеиновую кислоту, которая гибридизировалась с пробой. Альтернативно, образец нуклеиновой кислоты можно иммобилизовать на твердую подложку, и количество пробы, связанное с такой подложкой, можно определить. Подходящие методы анализа этого и любых других форматов можно найти, например, в WO 89/03891 и WO 90/13667.
Опыты по секвенированию ATR включали приведение образца тела человека или животного, содержащего мишень ДНК или РНК, в контакт с пробой, включающей полинуклеотид или праймер изобретения, в условиях гибридизации и определение последовательности посредством, например, способа терминации Ganger дидеокси цепи (см. Sambrook et al.).
Такой способ обычно включает удлинение в присутствии пригодных реагентов, праймера путем синтеза цепи комплементарной к ДНК мишени или РНК мишени и селективно терминирующий реакцию удлинения при одном или большем количестве А, С, Q или T/U остатка; обеспечение возможности для осуществления удлинения цепи и реакции терминации; разделение согласно размеру удлиненных продуктов для определения последовательности нуклеотидов, при которых произошла селективная терминация. Подходящие реагенты включают фермент ДНК полимеразы, деоксинуклеотиды dATP, dCTP, dGTP и dTTP, буфер и АТР. Дидеоксинуклеотиды используются для селективной терминации.
Опыты для обнаружения или секвенирования ATR в теле человека или животного могут использоваться для определения ATR последовательностей внутри клеток у отдельных представителей, которые имеют, или предполагают, что имеют, измененную последовательность ATR гена, например, в раковых клетках, включая клетки лейкемии и твердые опухоли такие, как опухоли молочной железы, яичника, легкого, толстой кишки, поджелудочной железы, яичка, печени, мозга, мышечных и костных опухолей.
Кроме того, открытие ATR будет выявлять роль этого гена в наследственных заболеваниях, которые будут исследоваться, способом, аналогичным способу АТМ гена. В основном это будет включать установление статуса ATR (например, использованием анализа PCR последовательности) в клетках, выделенных из пациентов с заболеваниями, которые могут быть связаны с повреждением до реплицирования клеток, например, наследственная предрасположенность к раку, хромосомный разрыв или нестабильный фенотип или восстановимо-поврежденный чувствительный фенотип.
Пробы изобретения могут традиционно упаковываться в форме опытного набора в соответствующем контейнере. В таких наборах проба может связываться с твердой подложкой, где формат анализа, для которого конструируется набор, требует такого связывания. Набор также может содержать пригодный реагент для обработки образца, который будет опробоваться, гибридизируя пробу с нуклеиновой кислотой в образце, контрольные реагенты, инструкции, и тому подобное.
Настоящее изобретение также представляет полинуклеотиды, кодирующие полипептиды изобретения, описанные ниже. Из-за того, что такие полинуклеотиды будут пригодны как последовательности для рекомбинантного образования полипептидов изобретения, для них не является необходимым быть селективно гибридизируемыми с последовательностью Seq.ID No.1, хотя это будет обычно желательным. Иначе говоря, такие полипетиды могут метиться, использоваться и готовиться, как описано выше, если это желательно. Полипептиды изобретения описываются ниже.
Особенно предпочтительными полинуклеотидами изобретения являются те, которые получаются из домена липид киназы ATR, его аллельных вариантов и видовых гомологов. Домен липид киназы представляется нуклеотидами от 7054 до 8011 Seq.ID No.1. Полинуклеотиды изобретения, которые включают этот домен, являются особенно предпочтительными. Термин "липид киназный домен" относится к домену, который является гомологичным с другими известными липид киназами, в частности р110 субъединицей PI-3 киназы, как определено рядами последовательности.
Другими предпочтительными полинуклеотидами изобретения являются те, которые включают нуклеотиды, кодирующие аминокислоты от 181 до 302 Seq.ID No 2 (нуклеотиды от 620 до 985 Seq.ID No.1), которые, как предполагается, являются областью лейциновой "молнии", предполагаемым сайтом белок-белкового взаимодействия, и аминокислот от 1358 до 1366 (нуклеотиды от 4151 до 4177), которые также сохраняются.
В дополнительном аспекте, полинуклеотиды изобретения включают те Seq.ID No 3 и их фрагменты, которые способны селективно гибридизироваться с этой последовательностью другой, чем фрагмент, состоящий из нуклеотидов от 2482 до 6599, в котором проведены следующие изменения: делеция остатков 2499, 2501, 2507 и 2509; инсерция С между 5918/5919.
Особенно предпочтительные фрагменты включают те, которые содержат остаток от 6826 до 7334 (домен липид киназы) и области лейциновой "молнии" от 1476 до 1625 и от 2310 до 2357. Кроме того, является предпочтительным фрагмент, включающий сохраненную область от 3891 до 3917. Такие полипептиды и фрагменты могут быть приготовлены и использованы, как описано выше.
В. Полипептиды
Полипептиды изобретения включают полипептиды по существу в выделенной форме, которые включают последовательность, представленную в виде Seq ID No.2.
Полипептиды, кроме того, включают варианты таких последовательностей, включая естественно возникшие алельные варианты и синтетические варианты, которые являются существенно гомологичными указанным полипептидам. В этом контексте, существенная гомология рассматривается как последовательность, которая имеет, по крайней мере, 70%, например, 80% или 90% аминокислотной гомологии (идентичности) в пределах 30 аминокислот с последовательностью Seq ID No.2, за исключением липид киназного домена и С-терминальной части (остатки от 2326 до 2644), где существенная гомология рассматривается как, по крайней мере, 80% гомология, предпочтительно 90% гомология (идентичность) в пределах 50 аминокислот.
Полипептиды также включают другие полипептиды, которые кодируют ATR гомологи от других видов, включая животных таких, как млекопитающие (например, мыши, крысы или кролики), особенно приматов, и варианты их, как указано выше.
Полипептиды изобретения также включают фрагменты вышеупомянутых полипептидов полной длины и их варианты, включая фрагменты последовательности, представленной в виде Seq ID No.2.
Предпочтительные фрагменты включают те, которые включают эпитоп, особенно эпитоп. Пригодные фрагменты будут составлять, по крайней мере, около 5, например, 10, 12, 15 или 20 аминокислот по размеру. Полипептидные фрагменты ATR белка и его аллельные и видовые варианты могут содержать одно или большее количество (например, 2, 3, 5, или 10) замещений, делеций, или инсертов, включая сохраненные замещения.
Сохраненные замещения могут готовиться в соответствии со следующей таблицей, которая показывает сохраняемые замещения, где аминокислоты одного и того же блока во второй колонке и предпочтительно на одной и той же строке в третьей колонке могут замещаться друг на друга:
Варианты полипептидов изобретения могут также включать полипептиды, где одна или больше специфических (т.е. естественно кодированнных) аминокислот делецируется или замещается, или где добавляются одна или больше неспецифических аминокислот: (1) без потери активности киназы, специфичной к полипептидам изобретения; или (2) при отсутствии акивности киназы, специфичной к полипептидам изобретения; (3) при отсутствии способности к взаимодействию с участниками или регуляторами пути точки контроля клеточного цикла.
Эпитопы могут определяться любым техническим средством, таким, как средство пептидного сканирования, как описано Geysen et al. Mol. lmmunol., 23; 709-715 (1986).
Полипептиды изобретения могут находиться по существу в выделенной форме. Должно быть понятно, что полипептиды могут смешиваться с носителями или разбавителями, которые не будут влиять на предполагаемое назначение полипептида, и все же могут рассматриваться как существенно выделенные. Полипептид изобретения может также быть в существенно очищенной форме, в том случае, когда полипептид будет содержаться в препарате более чем на 90%, например, 95%, 98% или 99% полипептида в препарате, он является полипептидом изобретения. Полипептиды изобретения могут быть модифицированы, например, добавлением остатков гистидина, для облегчения их очистки, или добавлением сигнальной последовательности, чтобы способствовать их выделению из клетки.
Полипептиды изобретения могут метиться обнаруживаемой меткой. Обнаруживаемая метка может быть любой пригодной меткой, которая позволяет обнаруживать полипептид. Пригодные метки включают радиоизотопы, например 125I, ферменты, антитела, полинуклеотиды и линкеры, такие, как биотин. Меченные полипептиды изобретения могут использоваться в диагностических процедурах, таких, как иммуноанализы, для того, чтобы определять количество полипептида в образце. Полипептиды или меченные полипептиды изобретения могут также использоваться в серологических или связанных с клеткой иммуных анализах для определения иммунной реактивности к указанным полипептидам в организмах животных и людей при использовании стандартных протоколов.
Полипептиды или меченные полипептиды изобретения или фрагменты их могут также фиксироваться на твердой фазе, например, на поверхности чашек для иммуного анализа или измерительного стержня.
Такие меченные или иммобилизованные полипептиды могут упаковываться в наборы в подходящем контейнере, вместе с подходящими реагентами, контрольными составами, инструкциями и тому подобным.
Такие полипептиды и наборы могут использоваться в методах обнаружения антител к ATR белку или его аллельных или видовых вариантов с помощью иммуных анализов.
Способы иммуного анализа хорошо известны в данной области и будут обычно включать:
(a) обеспечение полипептида, содержащего эпитоп, способный к связыванию с помощью антитела по отношению к указанному белку;
(b) инкубирование биологического образца с указанным полипептидом при условиях, которые позволяют образование антитело-антигенного комплекса;
и
(c) определение, образовался ли антитело-антигенный комплекс, содержащий указанный полипептид.
Полипептиды изобретения могут быть образованы либо синтетическими средствами (например, как описано Geysen et al.), либо рекомбинантно, как описано ниже.
Особено предпочтительные полипептиды изобретения включают те, которые протягиваются или внутри домена липид киназы, а именно от аминокислот 2326 до 2644 Seq.ID.2, или последовательностей, существенно гомологичных ему. Фрагменты, как определено выше, из этой области являются особенно предпочтительными. Полипептиды и фрагменты их могут содержать аминокислотные сдвиги, как описано выше, включая замещения при одной или больше позиций 2475, 2480 и 2494, которые соответствуют позициям rad3 заместителей, описанных в примерах ниже. Предпочтительные замещения включают D2475A, N2480K и D2494E.
Полипептиды изобретения могут использоваться in vitro или in vivo в системах клеточной культуры для изучения роли ATR в качестве гена точки контроля. Например, усеченные или модифицированные (например, модифицированные в домене липид киназы) ATR-ы могут быть введены в клетку для разрушения нормальных функций точки контроля, которые происходят в клетке.
Полипепиды изобретения могут вводиться в клетку посредством in situ эспрессии полипептида из рекомбинантного экспрессионного вектора (см. ниже). Экспрессионный вектор необязательно несет индуцибельный промотор для контроля экспресии полипептида.
Применение клеток хозяев млекопитающего, как ожидается, обеспечит такие пост-трансляционные модификации (например, миристолирование, гликозилирование, усечение, лапидацию и тирозин, серин или треонин фосфорелирование), которые могут оказаться нужными, чтобы придать оптимальную биологическую активность продуктам рекомбинантной экспрессии изобретения.
Системы такой клеточной культуры, в которых экспрессируется полипептид изобретения, могут использоваться в системах анализа для идентификации веществ кандидатов, которые мешают или усиливают функции точки контроля в клетке (см. ниже).
В дополнительном аспекте, полипептиды изобретения включают белок Seq.ID No.4 и фрагменты его из области другой, чем фрагмент, содержащий аминокислоты от 713 до 1778. Особенно предпочтительные фрагменты включают те, которые содержат остатки от 2082 до 2386 (домен липид киназы) и области лейциновой "молнии" от 298 до 347 и от 576 до 591. Кроме того, фрагмент, включающий сохраненную область от 1103 до 1111, является предпочтительным. Такие полипептиды и фрагменты могут готовиться и использоваться, как описано выше.
Изобретение также представляет полипептиды, существенно гомологичные белку Seq.ID No.4 и фрагментам его. В этом контексте, существенная гомология рассматривается как последовательность, которая имеет, по крайней мере, 70%, например, 80% или 90% аминокислотной гомологии (идентичности) в пределах 30 аминокислот с последовательностью Seq.ID No.4, за исключением липид киназного домена и С-терминальной части (остатки от 2082 до 2386), где существенная гомология рассматривается как, по крайней мере, 80%, предпочтительно, по крайней мере, 90% гомологии (идентичности) в пределах 50 аминокислот.
С. Векторы
Полинуклеотиды изобретения можно вводить в рекомбинантный способный к репликации вектор. Вектор можно использовать для репликации нуклеиновой кислоты в совместимой клетке хозяине. Таким образом, в следующем варианте, изобретение представляет собой способ приготовления полинуклеотидов изобретения введением полинуклеотида изобретения в способный к репликации вектор, введением вектора в совместимую клетку хозяин и выращиванием клетки хозяина при условиях, которые приводят к репликации вектора. Вектор может выделяться из клетки хозяина. Пригодные клетки хозяева описаны ниже в связи с экспрессионными векторами.
Р. Экспрессионные векторы
Предпочтительно, полинуклеотиды изобретения в векторе являются способными к связыванию с контрольной последовательностью, которая является способной обеспечить экспрессию кодирующей последовательности клеткой хозяином, т.е. вектор является экспрессионным вектором.
Термин "способный к связыванию" ("operably linked") относится к наложению прямой мутации и супрессора (juxtaposition), где описанные компоненты находятся во взаимосвязи, позволяющей им функционировать предназначенным им способом. Контрольная последовательность, "способная к связыванию" с кодирующей последовательностью, лигируется таким образом, что экспрессия кодирующей последовательности достигается при условиях, совместимых с контрольными последовательностями.
Такие векторы можно трансформировать в пригодной клетке хозяине, как описано выше, для обеспечения экспрессии полипептида изобретения. Таким образом, в следующем аспекте изобретение представляет собой процесс приготовления полипептидов согласно изобретению, который включает культивирование клетки хозяина, трансформированной или трансфекцированной экспрессионным вектором, как описанно выше, при условиях, которые обеспечивают экспрессию вектором кодирующей последовательности, которая кодирует полипептиды, и выделение экспрессированных полипептидов.
Векторы могут быть, например, плазмидой, вирусом или фаговым вектором, снабженным изначально репликацией, необязательно промотором для экспрессии указанного полинуклеотида и необязательно регулятором промотора. Векторы могут содержать один или больше селектируемых генов маркеров, например, ген, устойчивый к ампицилину в случае бактериальной плазмиды, или ген, устойчивый к неомицину для вектора млекопитающего. Векторы могут применяться in vitro, например, для приготовления РНК или применяться для трансфекции или трансформации клетки хозяина. Векторы можно приспосабливать также для применения in vivo, например, в способе генной терапии.
Следующий вариант изобретения представляет клетки хозяев, трансформированные или трансфекцированные векторами для репликации и экспрессии полинуклеотидов изобретения. Клетки будут отбираться так, чтобы они были совместимыми с указанным вектором, и могут быть бактериальными, дрожжевыми, клетками насекомого или млекопитающего.
Полинуклеотиды, согласно изобретению, можно также инсерцировать в векторы, описанные выше, в антисмысловой ориентации, для того, чтобы обеспечить приготовление антисмысловой РНК. Антисмысловая РНК или другие антисмысловые полинуклеотиды могут также продуцироваться синтетическими способами. Такие антисмысловые полинуклеотиды могут использоваться в способе контролирования уровней ATR или его вариантов или видовых гомологов.
Промоторы и другие экспрессионные регулирующие сигналы могут выбираться так, чтобы они были совместимыми с клеткой хозяином, для которой конструируется экспрессионный вектор. Например, дрожжевые промоторы включают S.cerevisiae GAL4 и ADH промоторы, S.pombe nmtl и adh промотор. Промоторы млекопитающего включают металлотионеиновый промотор, который может быть введен в ответ на тяжелые металлы, такие, как кадмий. Могут также использоваться вирусные промоторы, такие, как SV40 большой Т антигеновый промотор или аденовирусные промоторы. Все эти промоторы являются легко доступными в этой области.
Е. Антитела
Изобретение также представляет моноклональные или поликлональные антитела к полипептидам изобретения или их фрагментам. Изобретение далее представляет собой процесс получения моноклональных или поликлональных антител к полипептидам изобретения. Моноклональные антитела могут быть приготовлены с помощью традиционной гибридомной технологии, использующей полипептиды изобретения или их пептидные фрагменты в качестве иммуногенов. Поликлональные антитела могут также готовиться традиционными средствами, которые включают инокулирование животного хозяина, например, крысы или кролика, полипептидом изобретения или его пептидным фрагментом и выделение иммуной сыворотки.
Для того чтобы готовить такие антитела, изобретение должно обеспечить полипептиды изобретения или их фрагменты, гаптенизированные с другим полипептидом для использования в качестве иммуногенов в организмах животных или людей.
Предпочтительные антитела изобретения будут способны к селективному связыванию ATR белка человека, который обладает сродством, по крайней мере, 10-кратным, предпочтительно, по крайней мере, 100-кратным сродству rad3 белка. Такие антитела могут получаться известными приемами, например, отбором областей ATR белка с последовательностями, отличными от соответствующих областей rad3, приготовлением пептидов, содержащих такие последовательности, и использование таких пептидов в качестве иммуногенов. Вслед за получением антител, может быть определено связывание указанных антител. Предпочтительные антитела изобретения включают те, которые являются способными селективно связывать домена липид киназы (как определено выше) ATR белка человека. Кроме того, антитела, которые являются способными к связыванию липид киназных доменов человека и дрожжей (S.pombe) аналогичным сродством, но не к доменам АТМ семейства белков, образуют следующий аспект изобретения. Такие антитела могут выращиваться в противоположность пептидам из липид киназных доменов, которые соответствуют областям, найденным как идентичные, или существенно идентичные, в генах дрожжей и человека.
Для целей этого изобретения, термин "антитело", если не оговорено особо, включает фрагменты всего антитела, которое сохраняет свою связывающую активность для антигена мишени опухоли. Такие фрагменты включают Fv, F(ab) и F(ab’)2 фрагменты, так же как одноцепочные антитела. Кроме того, антитела и их фрагменты могут быть очеловеченными антителами, например, как описано в ЕР-А-239400.
Антитела могут применяться в способе обнаружения полипептидов изобретения, представленных в биологических образцах способом, который включает:
(a) обеспечение антитела изобретения;
(b) инкубирование биологического образца указанным антителом при условиях, которые позволяют образование антитело-антигенового комплекса; и
(c) определение, образовался ли антитело-антигеновый комплекс, включающий указанное антитело.
Пригодные образцы включают экстракты из разделенных клеток, например, лейкоцитов или раковых клеток, включающих клетки лейкемии, и твердых опухолей таких, как опухоли молочной железы, яичника, легкого, толстой кишки, поджелудочной железы, яичка, печени, мозга, мышечных и костных опухолей.
Антитела изобретения могут быть связаны с твердой подложкой и/или упакованы в наборы в соответствующем контейнере вместе с подходящими реагентами, контрольными составами, инструкциями и им подобными.
F. Анализы
Ликвидация точек контроля клеточного цикла является потенциальной стратегией для развития или конструирования лекарств для противораковой терапии, как нового лечения, так частично и объединенной терапии для усиления специфической токсичности существующих химиотерапевтических агентов. Например, алкилирующие агенты, такие, как азотсодержащий горчичный газ, используются как терапевтические агенты, которые повреждают ДНК в отношении быстрого деления клеток, приводя к смерти клетки. Токсичность таких агентов может быть уменьшена за счет восстановления ДНК и механизмов точки контроля. Ликвидация таких механизмов будет таким образом усиливать эффективность терапевтических соединений, сконструированных для повреждения ДНК. Ликвидация ATR точки контроля будет особенно полезна там, где опухолевые клетки утратили другую точку контроля или гены, ответственные за повреждение, так как эти другие гены могут быть способны комплементировать потерю функции ATR в неопухолевых клетках, приводя к еще большему усилению эффективности химиотерапевтического агента.
Липид киназная активность АТР является целью при разработке противораковых соединений, так как результаты, представленные в следующих примерах, показывают, что для функции ATR требуется домен киназы. Таким образом, настоящее изобретение представляет собой способ анализа для скринирования веществ кандидатов при противораковой терапии, который включает:
(a) обеспечение полипептида изобретения, который сохраняет активность липид киназы и субстрат для указанной киназы, при условиях и с реагентами такими, что активность киназы будет влиять на субстрат;
(b) приведение указанного полипептида и субстрата в контакт с веществом кандидатом;
(c) измерение степени уменьшения активности киназы полипептида; и
(d) выбор вещества кандидата, которое обеспечивает уменьшение активности.
Анализ можно проводить in vitro, например, в лунках микротитрического планшета. Такой формат можно легко приспособить для автоматического применения, позволяя использовать большое количество веществ кандидатов для скринирования.
Субстрат, на который будет воздействовать полипептид изобретения, может быть белковым или липидным субстратом естественного или синтетического происхождения. Обычно, полипептид изобретения будет фосфорилировать субстрат.
Любой пригодный формат может применяться специалистом в этой области из категории производительных анализов. Обычно, полипептид изобретения, который сохраняет активность липид киназы, будет связываться с твердой подложкой в присутствии субстрата и клеточных или других компонентов, которые обычно требуются для активности. Меченый фосфат и вещество кандидат будут добавляться в смесь одновременно или последовательно один за другим. После прохождения соответствующего времени реакции (обычно несколько минут, но в любом случае достаточного для того, чтобы произошло фосфорелирование субстрата в отсутствие вещества кандидата), определяется количество свободного фосфата, например, осаждением фосфата. Вещества кандидаты, которые ингибируют активность киназы, будут ингибировать вхождение свободного фосфата в субстрат и, таким образом, место, где находится свободный фосфат, является показателем ингибирования.
Другие форматы анализа могут быть использованы специалистом в данной области.
Вещества кандидаты могут использоваться в начальном скрининге в группах из, например, 10 соединений за реакцию, и соединениях тех групп, которые показывают игибирование при индивидуальном тестировании.
Пригодные вещества кандидаты включают пептиды, особенно от около 5 до 20 аминокислот по размеру, основанные на последовательности домена киназы, или варианты таких пептидов, в которых один или больше остатков замещены, как описано выше. Могут использоваться пептиды из панелей пептидов, включающих статистические последовательности или последовательности, которые изменены по составу для обеспечения максимального разнообразия панели пептидов. Кроме того, вещества кандидаты включают киназные ингибиторы, которые являются маленькими молекулами, такими, как циклоспорин подобные и стауроспорин подобные соединения, или другими соединениями, промышленно доступными, в панелях малых молекулярных ингибиторов.
Вещества кандидаты, которые проявили активность в in vitro скринингах, таких, как описанные выше, могут затем испытываться в in vivo системах, таких, как дрожжевые клетки или клетки млекопитающего, которые будут подвергаться воздействию ингибиторов и испытываться на активность точки контроля.
Было также показано, что Rad3 обладает активностью протеин киназы. Субстраты мишени активности Rad3 протеин киназы могут идентифицироваться введением испытываемых соединений в анализы на активность киназы. Rad3 белок повторно суспендируется в буфере киназы и инкубируется при наличии или отсутствии испытываемого соединения (например, казеина, гистона H1, или соответствующего субстратного пептида). Количество молей фосфата, перенесенное киназой к испытываемому соединению, измеряется авторадиографированием или сцинтилляционным подсчетом. Переход фосфата к испытываемому соединению является показателем, что испытываемое соединение является субстратом киназы.
Агенты, которые модулируют активность Rad3/ATR липид киназы или Rad3 протеин киназы, могут идентифицироваться инкубированием испытываемого соединения и Rad3/ATR иммуноочищенного из клеток естественно экспрессирующих Rad3/ATR, с Rad3/ATR, полученным из рекомбинантных прокариотических или эукариотических клеток экспрессирующих фермент, или с очищенным Rad3/ATR, и затем определением влияния испытываемого соединения на Rad3/ATR активность. Активность Rad3/ATR липид киназы или доменов Rad3 протеин киназы может измеряться определением молей 32Р-фосфата, перенесенного киназой из гамма-32Р-АТР или к себе (автофосфорелирование) или к экзогенному субстрату, такому, как липид или белок. Количество фосфата, введенного в субстрат, измеряется сцинтилляционным подсчетом или авторадиографированием. Увеличение молей фосфата, перенесенного в субстрат в присутствие испытываемого соединения, по сравнению с молями фосфата, перенесенного в субстрат в отсутствии испытываемого соединения, показывает, что испытываемое соединение является активатором указанной активности киназы. Наоборот, уменьшение молей фосфата, перенесенного в субстрат в присутствии испытываемого соединения, по сравнению с молями фосфата, перенесенного в субстрат в отсутствие испытываемого соединения, показывает, что модулятор является ингибитором указанной активности киназы.
В предпочтительном теперь анализе, Rad3/ATR антитело, связанное с гранулами агарозы, инкубируется клеточным лизатом, приготовленным из клеток хозяев экспрессирующих Rad3/ATR. Гранулы промывали для удаления белков, связывающихся неспецифично с гранулами, и гранулы затем ресуспендировали в буфере киназы (таком как 25 мМ K-HEPES рН 7.7, 50 мМ хлорида калия, 10 мМ хлорида магния, 0.1% Nonidet-P-40, 20% глицерина, 1 мМ DTT). Реакция инициируется добавлением 100 мкМ гамма-32Р-АТР (4 Ci/мМ) и экзогенного субстрата, такого, как липид или пептид, и реакция проводится при 30°С в течение 10 минут. Активность киназы измеряется определением молей 32Р-фосфата, перенесенного киназой или к себе или к добавленному субстрату. В предпочтительном варианте, клетки хозяева испытывают недостаток в эндогенной активности Rad3/ATR киназы. Селективность соединения, которое модулирует активность липид киназы Rad3/ATR, может оцениваться сравнением его активности на Rad3/ATR с его активностью, например, на других известных фосфатидилинозитол-3(РI-3)-родственных киназах. Комбинация рекомбинантных Rad3/ATR продуктов изобретения с другими рекомбинантными РI-3 родственными киназными продуктами в серии независимых анализов обеспечивает систему для создания селективных модуляторов Rad3/ATR киназной активности. Аналогично, селективность соединения, которое модулирует активность протеин киназы Rad3, может определяться ссылкой на другие протеин киназы, например, ДНК зависимую протеин киназу или АТМ.
Кроме того, демонстрация того, что rad мутант rad.D2249E (смотри примеры) может действовать как доминантный негативный мутант, указывает на включение в один или больше белковых комплексов, и такие комплексы сами могут нацеливаться для терапевтических внедрений. Было показано, например, что Rad3 может одинаково самоассоциироваться и ассоциироваться с ATR. Следовательно, это похоже на то, что Rad/ATR функционирует как мультимерные молекулы. Мутантный дрожжевой rad или ATR гены человека, или их производные, которые также испытывают недостаток Rad/ATR активности, могут вводиться в клетки, чтобы действовать в качестве доминантных негативных мутантов. Таким образом, например, если экспрессия доминантного негативного мутанта (например, ATR D2475A, N2480K или D2494E) в опухолевых клетках ведет к повышенной чувствительности к радиации, это указывает на то, что природная ATR все еще функционирует и таким образом выступает в качестве мишени для терапевтических агентов.
Взаимодействующие белки, которые включают компоненты комплексов мультимерного белка, включающего Rad3 или ATR, могут идентифицироваться следующими анализами.
Первый анализ, рассмотренный в изобретении, представляет ди-гибридное скринирование. Ди-гибридная система была выращена в дрожжах [Chien et.al., (1991)] и основана на функциональной in vivo реконструкции фактора транскрипции, который активирует ген репортер. Конкретно, полинуклеотид кодирующий белок, который взаимодействует с Rad3/ATR, выделяется с помощью: трансформирования или трансфекцирования соответствующих клеток хозяев ДНК конструкцией, включающей ген репортер под контролем промотора, регулированного фактором транскрипции, имеющим ДНК связывающий домен и активирующий домен; экспрессирования в клетках хозяевах первой гибридной ДНК последовательности, кодирующей первый гибрид части или всего и либо ДНК связывающий домен, либо активирующий домен фактора транскрипции; экспрессирования в клетках хозяевах библиотеки вторых гибридных ДНК последовательностей, кодирующих вторые гибриды части или всех предполагаемых Rad3/ATR связывающих белков и ДНК связывающего домена, или активирующего домена фактора транскрипции, который не вводится в первый гибрид; обнаружения связывания Rad3/ATR взаимодействующего белка с Rad3/ATR в конкретной клетке хозяине за счет обнаружения продуцирования генного продукта репортера в клетке хозяине; и выделения вторых гибридных ДНК последовательностей, кодирующих взаимодействующий белок из конкретной клетки хозяина. Теперь предпочтительными для использования в анализе являются lехА промотор для проведения экспресии гена репортера, lacZ ген репортер, фактор транскрипции, включающий lехА ДНК связывающий домен и GAL4 трансактивационный домен и дрожжевые клетки хозяева.
Другие анализы для идентификации белков, которые взаимодействуют с Rad3 или ATR, могут включать иммобилизацию Rad3/ATR или испытываемого белка, обнаруживание метящего неиммобилизованного связывающего партнера, инкубирование связывающих партнеров вместе и определение величины меченной связи. Связанная метка указывает на то, что испытываемый белок взаимодействует с Rad3/ATR.
Другой тип анализа для идентификации Rad3 или ATR взаимодействующих белков включает иммобилизацию Rad3/ATR или его фрагмента на твердую подложку, покрытую (или импрегнированную флуоресцирующим агентом) флуоресцирующим агентом, мечение испытываемого белка соединением, способным возбуждать флуоресцирующий агент, контактирование иммобилизованного Rad3/ATR с меченным испытываемым белком, обнаружение излучения света флуоресцирующим агентом и идентификацию взаимодействующих белков в качестве испытываемых белков, которая приводит к излучению света флуоресцирующим агентом. Или же, предполагаемый взаимодействующий белок может быть иммобилизован и Rad3/ATR может быть меченным в анализе.
Соединения, которые модулируют взаимодействие между Rad3/ATR и другими клеточными компонентами, могут быть использованы в методах лечения рака. Например, если конкретная форма рака возникает из мутации гена другого чем ATR, такого, как р53 гена, агент, который ингибирует транскрипцию или ферментативную активность ATR и таким образом G2 контрольную точку клеточного цикла, может использоваться для превращения раковых клеток, более восприимчивых к химиотерапии или радиационной терапии. Терапевтический эффект такого агента подчеркивает фактически, что текущая радиационная терапия или химиотерапия в большинстве случаев не перекрывает способности р53 мутанта раковой клетки к ощущению и корректировке ДНК повреждения, возникающего в результате лечения. В результате, раковая клетка может просто исправить ДНК повреждение. Модулирующие агенты изобретения могут поэтому быть адъювантами химиотерапии и радиационной терапии или могут быть сами непосредственно активными в качестве химиотерапевтических лекарств.
Анализы для идентификации соединений, которые модулируют взаимодействие Rad3/ATR с другими белками, могут включать: трансформирование или трансфекцирование соответствующих клеток хозяев ДНК конструкцией, включающей ген репортер под контролем промотора, регулированного фактором транскрипции, имеющим ДНК связывающий домен и активирующий домен; экспрессирование в клетках хозяевах первой гибридной ДНК последовательности, кодирующей первый гибрид части или всего Rad/ATR и ДНК связывающий домен, или активирующий домен фактора транскрипции; экспрессирование в клетках хозяевах второй гибридной ДНК последовательности, кодирующей часть или весь белок, который взаимодействует с Rad3/ATR и ДНК связывающим доменом, или активирующим доменом фактора транскрипции, который не вводится в первый гибрид; оценку влияния испытываемого соединения на взаимодействие между Rad3/ATR и взаимодействующим белком за счет обнаружения связывания взаимодействующего белка с Rad3/ATR в конкретной клетке хозяине; за счет измерения образования продукта гена репортера в клетке хозяине в присутствии или отсутствие испытываемого соединения: и идентификацию модулирующих соединений в качестве таких испытываемых соединений, которые меняют продуцирование продукта гена репортера, по сравнению с продуцированием продукта гена репортера в отсутствие модулирующего соединения. Теперь предпочтительными для использования в анализе являются lехА промотор для проведения экспресии гена репортера, lacZ ген репортер, фактор транскрипции, включающий lехА ДНК домен и GAL4 трансактивационный домен и дрожжевые клетки хозяева.
Другой тип анализа для идентификации соединений, которые модулируют взаимодействие между Rad3/ATR и взаимодействующим белком, включает иммобилизацию Rad3/ATR или натурального Rad3/ATR взаимодействующего белка, обнаруживание метящего неиммобилизованного связывающего партнера, инкубирование связывающих партнеров вместе и определение влияния испытываемого соединения на величину меченой связи, где снижение в меченой связи в присутствии испытываемого соединения по сравнению с величиной меченой связи в отсутствие испытываемого соединения указывает на то, что испытываемый агент является ингибитором Rad3/ATR взаимодействия с белком. Напротив, увеличение в связывании в присутствии испытываемого соединения к величине меченой связи в отсутствие сравниваемого соединения указывает на то, что предполагаемый модулятор является активатором Rad3/ATR взаимодействия с белком.
Еще один способ, рассматриваемый изобретением для идентификации соединений, которые модулируют связывание между Rad3/ATR и взаимодействующим белком, включает иммобилизацию Rad3/ATR или его фрагмента на твердую подложку, покрытую (или импрегнированную) флуоресцирующим агентом, мечение взаимодействующего белка соединением, способным возбуждать флуоресцирующий агент, контактирование иммобилизованного Rad3/ATR с меченым взаимодействующим белком, в присутствии или отсутствие испытываемого соединения, обнаружение излучения света флуоресцирующим агентом и идентификацию модулирующих соединений в качестве тех испытываемых соединений, которые влияют на излучение света флуоресцирующим агентом, по сравнению с излучением света флуоресцирующим агентом в отсутствие испытываемого соединения. Или же Rad3/ATR взаимодействующий белок может быть иммобилизованным и Rad3/ATR может быть меченным в анализе.
Было показано, что Rad3 взаимодействует с ATR. Поэтому вышеупомянутые анализы могут быть также использованы для идентификации соединений, которые модулируют взаимодействие между Rad3 и ATR где взаимодействие белка, описанное в методах анализа, представляет либо Rad3 либо ATR.
Было также показано, что Rad3 связывается сам с собой, предполагая в значительной степени, что ATR также может связываться сам с собой. Поэтому, вышеупомянутые анализы могут быть также использованы для идентификации соединений, которые модулируют Rad3-Rad3 взаимодействия и ATR-ATR взаимодействия.
Такие соединения могут быть использованы терапевтически для разрушения взаимодействий ATR-ATR и увеличения чувствительности к опухолевым клеткам в химиотерапии и/или радиотерапии. Таким образом, изобретение представляет собой метод анализа для скринирования веществ кандидатов для противораковой терапии, который включает:
(а) (i) инкубирование полипептида изобретения с другим полипептидом изобретения, который может быть таким же или отличающимся от первого полипептида, в условиях, которые позволяют первому полипептиду связываться со вторым полипептидом с образованием комплекса;
(ii) приведение полученного таким образом комплекса в контакт с веществом кандидатом;
или
(а) инкубирование полипептида изобретения с другим полипептидом изобретения, который может быть таким же или отличающимся от первого полипептида, в условиях, которые позволяют первому полипептиду связываться со вторым полипептидом с образованием комплекса и в присутствии вещества кандидата;
и
(б) определение того, ингибирует ли вещество кандидат связывание первого полипетида со вторым полипетидом и
(с) выбор вещества кандидата, которое ингибирует связывание первого полипетида со вторым полипетидом.
Предпочтительно, первый и второй полипептиды могут быть отличными друг от друга, Например, первый полипептид и второй полипетид могут оба быть ATR, или могут оба быть Rad3, или один может быть ATR и один может быть Rad3 или производными либо ATR либо Rad3, которые сохраняют связывающую активность. Когда оба полипептида являются ATR или Rad3, в этих анализах, предпочтительно, будут использоваться две различаемые формы ATR/Rad3. Они могут быть различимыми, например, мечением любого из полипептидов. Примеры меток включают радиоактивные метки, эпитопные мишени или другие полипептидные мишени, такие, как глутатионин-S-трансфераза. Например, одна форма Rad3 может иметь одну форму эпитопной мишени и другая форма будет иметь другую эпитопную мишень, позволяя им быть различимыми иммунологически, так что связывание одной формы с другой может быть установлено качественно или количественно. В предпочтительном способе, первый полипептид может быть, например, иммобилизованным в агарозные гранулы или на твердую подложку, а второй полипетид может находиться в свободном растворе. Связывание определяется затем с использованием способов, описанных выше и хорошо известных специалистам в этой области.
Настоящим изобретением рассматриваются также продукты антител (например, моноклональные и поликлональные антитела, антитела единичной цепи, химирные антитела, CDR-привитые антитела и им подобные) и другие связывающие белки (такие, как те, которые идентифицированы в анализах, приведенных выше), которые являются специфическими для Rad3 протеин киназного домена или Rad3/ATR липид киназных доменов. Связывающие белки могут быть сконструированы с использованием выделенных натуральных или рекомбинантных ферментов. Связывающие белки могут быть разработаны с использованием выделенных естественных или рекомбинантных ферментов. Связывающие белки являются полезными, в свою очередь, для очистки рекомбинантных ферментов и ферментов естественного происхождения и идентификации клеток, продуцирующих такие ферменты. Анализы для обнаружения и количественного определения белков в клетках и жидкостях могут включать определение вещества единичного антитела или веществ множества антител в формате “сэндвичевого” анализа. Связывающие белки являются также, очевидно, полезными в модулировании (т.е. блокировании, ингибировании или стимулировании) взаимодействий фермент/субстрат или фермент/регулятор.
Модуляторы Rad3/ATR могут оказывать влияние на его киназную активность, его локализацию в клетке, и/или его взаимодействие с элементами пути точки контроля клеточного цикла. Селективные модуляторы могут включать, например, полипептиды или пептиды, которые специфически связываются с Rad3/ATR или Rad3/ATR нуклеиновой кислотой, и/или другими непептидными соединениями (например, выделенными или синтетическими органическими молекулами), которые специфически реагируют с Rad3/ATR или Rad3/ATR нуклеиновой кислотой. Мутантные формы Rad3/ATR, которые влияют на ферментативную активнность или клеточную локализацию Rad3/ATR дикого типа, также рассматриваются изобретением.
Кроме того, комбинаторные библиотеки, пептиды и пептидные миметики, определенные химические вещества, олигонуклеотиды и библиотеки натуральных продуктов могут быть скринированы на активность в качестве модуляторов Rad3/ATR киназной актиности и Rad3/ATR взаимодействий в анализах, таких, как те, которые описаны выше.
F. Терапевтические применения.
Модуляторы Rad3/ATR активности, включающие ингибиторы их липид киназной и протеин киназной активностей, могут быть использованы в противораковой терапии. В частности, они могут быть использованы для увеличения восприимчивости раковых клеток к химиотерапии и/или радиотерапии посредством их способности к разрыву регуляторных функций клеточного цикла Rad3/ATR.
Таким образом, изобретение представляет применение соединений, которые модулируют Rad3/ATR активность, идентифицированных с помощью скринирующих анализов, описанных выше, в способе лечения рака, В одном варианте, указанные соединения являются способными к ингибированию rad3/ATR липид киназной и/или Rad3 протеин киназной активности. В другом варианте, указанные соединения являются способными к ингибированию взаимодействий между ATR и им самим и/или между ATR и другими взаимодействующими белками, которые могут, например, нормально образовывать часть мультимерного белкового комплекса.
Должно быть понятно, что термин “соединение” в этом контексте относится также к веществам кандидатам, выбранным в вышеописанных анализах.
Обычно, соединения готовятся для клинического введения путем смешения их с фармацевтически приемлемым носителем или разбавителем. Например, они могут быть составлены для точечного, парентерального, внутривенного, внутримышечного, подкожного, внутриглазного или трансдермального введения. Предпочтительно, соединение используется в инъецируемой форме. Прямая инъекция в опухоль пациента является целесообразной, потому что делает возможным концентрировать терапевтический эффект на уровне поврежденных тканей. Поэтому соединение может быть смешано с любым наполнителем, который является фармацевтически приемлемым для инъецируемой композиции, предпочтительно, для прямой инъекции в участок, который подвергается лечению. Фармацевтический носитель или разбавитель могут быть, например, стерильными или изотоническими растворами.
Доза используемого соединения может регулироваться в соответствии с различными параметрами, особенно, в соответствии с используемым соединением, возрастом, весом и состоянием пациента, который подвергается лечению, формой используемого введения, патологией опухоли и требуемым клиническим режимом. В качестве руководства, количество соединения, введенного с помощью инъекции, является пригодным от 0.01 мг/кг до 30 мг/кг, предпочтительно, от 0.1 мг/кг до 10 мг/кг.
Пути введения и описанные дозы предполагаются только в виде руководства, так как специалист в этой области будет способен легко определить путь введения и дозу для любого конкретного пациента и состояния.
Соединения, которые будут вводиться, могут включать полипептиды или нуклеиновые кислоты. Нуклеиновые кислоты могут кодировать полипептиды или они могут кодировать антисмысловые конструкции, которые ингибируют экспрессию клеточного гена. Нуклеиновые кислоты могут быть введены с помощью, например, липофекции (lipofection) или с помощью вирусных векторов. Например, нуклеиновая кислота может образовывать часть вирусного вектора, такого, как аденовирус. Когда используются вирусные векторы, в общем, введенная доза находится между 104 и 1014 pfu/мл, предпочтительно, 106-1010 рfu/мл. Термин pfu (“plaque forming unit”) (бляшкообразующая единица) соответствует неэффективности вирусного раствора и определяется с помощью инфицирования соответствующей клеточной культуры и измерения, обычно после 48 часов, количества бляшек инфицированных клеток. Методы определения pfu титра вирусного раствора являются хорошо документированными в литературе.
Любые типы рака могут быть подвергнуты лечению этими методами, например, лейкемии, и твердые опухоли, такие, как опухоли молочной железы, яичника, легкого, толстой кишки, поджелудочной железы, яичка, печени, мозга, мышц и кости. Предпочтительно, опухоль имеет нормальную ATR функцию.
На чертеже показано отношение между ATR, rad3, mei-4I, MECI, TELI и АТМ/
А. Общие структуры ATR, Rad3, Mei-4I, Meclp, Tellp и АТМ
Обозначение: открытые квадраты - Rad3 домен; заштрихованные квадраты - киназный домен
В. Дендрограмма на основании рядов последовательностей, генерированных способом Clustal (РАМ 250) с использованием DNAstar software, rad3/ESRI/mei4I/ATR являются более родственными друг другу, чем АТМ и TeL1.
Последовательности rad3 и АТМ являются доступными в базе данных EMBL.
Следующие примеры иллюстрируют изобретение.
Пример 1
rad3 ген S.pombe является одним из шести генов, безусловно требуемых для ДНК структуры точек контроля в S.pombe (AI-Khodairy and Carr; 1992; Al-Khodairy et.al., 1994). Последовательность, представляющая часть rad3 гена, сообщалась Seaton et.al., (1992). При попытке выяснить интрон/эксонную структуру этого гена мы идентифицировали последующие аномалии с обоих концов 5’ и 3’. Мы секвенировали полный ген (см. Экспериментальную часть) и нашли, что rad3 является способным кодировать продукт 2386 аминокислот. С-терминальная область содержит типичные согласованные последовательности подкласса киназ, известных как липид киназы, основной член которого представляет р110 каталитическую субъединицу Р13 киназы (Hiles et.al., 1992).
Усеченный rad3 клон с недостающим аминотерминусом и киназной областью, как, сообщалось, комплементирует rad3::pR3H1.0 разрывающий мутант гена rad3 (Jimenez et.al., 1992). Этот разрывающий мутант не удаляет потенциальный киназный домен. Для выяснения роли этого домена, мы создали нуль мутант с помощью генного замещения. Этот мутант содержит аминокислоты 1477-2271 rad3, включающие киназный согласованный домен замещенный ura4+. Этот штамм rad3.d имеет идентичные дефекты точки контроля и обладает радиационной/гидроксимочевино чувствительностью к rad3.136 мутанту (Nasim and Smith, 1975) и исходному rad3::pR3H1.0 разрывающему мутанту (Jimenez et.al., 1992; Seaton et.al., 1992) (данные не показаны). Мы создали три отдельных точечных мутанта в предполагаемом киназном домене rad3 и использовали их в экспериментах по генному замещению в конструкициях штаммов с определенными киназными нуль мутациями. Все три штамма rad3.D2230A, rad3.N2235K и rad3.D2249E имели фенотипы, идентичные rad3.d нуль мутанту (данные не показаны), предполагая, что киназная активность требуется для Rad3 функции. В свете наших находок, интерпретация результатов Seaton et al. (1992) и Jimenez et al. (1992) состоит в том, что частичный клон может показывать внутригенную комплементацию между плазмидно рожденным усеченным геном и геномной частичной делецией, которая сохраняет киназную функцию. Такая интерпретация будет согласовываться с Rad3, действующим в виде димера или мультимера.
Когда киназный нуль аллель rad3.D2249E умеренно сверх-экспрессировался в клетках дикого типа под контролем модифицированного nmtl промотора (Maundrell, 1990), он вызывал чрезвычайную радиационную чувствительность, проанализированную с помощью испытаний с УФ полосами, и выступал в качестве доминантного негативного мутанта (данные не показны). Когда та же киназная нуль конструкция была экспрессирована при более высоком уровне, она ингибировала рост (данные не показаны). Исследование клеток указывает на то, что деление продолжается очень медленно, и при меньшем клеточном размере клеток дикого типа и клеток, содержащих пустой вектор, делятся приблизительно на 15 микрон, в то время как rad3 и rad3.D2249E сверх-экспрессирующие клетки делятся приблизительно на 11.2 микрона (данные не показаны). В S pombe это обычно указывает на повышение митоза.
Человеческий rad3 гомолог ATR.
Для идентификации человеческой формы rad3, была применена комбинация способов. Через эти подходы мы клонировали полностью кодирующую область человеческого гена (см. материалы и способы), которую мы назвали ATR (ataxia and rad related). ATR является способным кодировать 2644 аминокислотный белок, который является гораздо более родственным продуктам S.pombe rad3, S cerevisiae ESRI (Kato and Ogawa, 1994) и D melanogaster mei-4I генам (Hari et al., 1995), чем человеческому АТМ и S cerevisiae Теl1 белкам (Savitsky et al., 1995; Greenwell et al., 1995), и, вероятно, является истинным гомологом rad3. ESR1 является аллельным по отношению к mec1/sad3 мутантам точки контроля (Allen et al., 1994; Weinert et al., 1994), которые имеют эквивалентный фенотип по отношению к rad3. ATR является менее близко родственным человеческому АТМ гену точки контроля, содержащему С-терминальный предполагаемый липид киназный домен и имеющему аналогичную общую структуру. Ряды последовательностей четко демонстрируют, что rad3/ESR1 (MEC1/SAD3) /mei-4I/ATR гены являются более родственными друг другу, чем любые гены, которые родственны АТМ или TEL1, и что АТМ является более гомологичным TEL1 (см. чертеж).
АТМ ген экспрессируется в широкое разнообразие тканей ((Savitsky et al,, 1995). В S.cerevisiae, ESR1 показывает низкий уровень экспрессии в митотических клетках, но быстро индуцируется в процессе мейоза (Kato and Ogawa, 1994). Используя анализ Нозерн блоттинга, мы продемонстрировали, что ATR также слабо экспрессируется во многие ткани, но что он является более высоко экспрессированным в яичко (данные не показаны). Показано, что ATR, Rad3 и Esrlp белки являются более родственными друг другу, чем АТМ, самая высокая ATR экспрессия в яичко согласуется с наблюдением о том, что Esrlp играет роль в мейотической рекомбинации (Kato and Ogawa, 1994). Используя анализы FISH и PCR, мы картировали ATR в хромосому 3q22-3q25 (данные не показаны). Эта область не ассоциируется с известными пронированными (prone) синдромами рака.
Для того чтобы далее исследовать возможности того, что Rad3 выступает в качестве мультимера, мы создали две раздельные меченые конструкции полной длины rad3 в pREP на основании индуцибельных векторов. В одном случае, Rad3 транслировался с двумя mуc эпитопными метками в N терминусе, в то время, как в другом эти конструкции замещались тройной НА эпитопной меткой. Когда обе конструкции экспрессировали вместе в клетки дикого типа, стало возможным со-осаждение НА меченого Rad3 с mуc специфическим антителом, и mуc меченого Rad 3 с НА специфическим антителом (данные не показаны). Это демонстрирует, что in vivo, Rad3 белок является способным к самоассоциации и является полностью совпадающим с данными комплементации Jimenez et al. (1992).
Хотя ATR ген может не комплементировать фенотип rad3 мутантов, мы исследовали способность ATR к образованию белкового комплекса с S.pombe Rad3 путем экспрессирования и ATR и mуc-меченого S.pombe Rad3 в тех же самых дрожжевых клетках. Используя анти-ATR антитело (которое не осаждает S.pombe Rad3, см. материалы и способы), у нас появилась возможность со-осадить дрожжевой белок. Мы были способны также осадить человеческий ATR белок с mуc-специфическими антителами, которые узнавали S.pombe Rad3 (данные не показаны). Эти данные предполагают, что человеческие и дрожжевые белки могут образовывать гетеродимерный комплекс, который подтверждает спор, основанный на подобии последовательности, близко функциоанального отношения между этими гомологами. Rad3 белки обладают ассоциированной киназной активностью.
Так как эксперименты по мутагенезу предполагают, что киназная активность Rad3 белков in vivo является, по-видимому, существенной для их функции, мы исследовали эту активность далее. Используя S.pombe Rad3::ura4-клетки, экспрессирующие НА нацеленный S.pombe Rad3, мы были способны обнаружить значительную протеин киназную активность, которая ускорялась НА-специфическими антителами только тогда, когда индуцировался Rad3, который не изменялся после облучения (данные не показаны). Эта активность, которая является специфической для Rad3 или со-ускоряющей киназы, проявляется для отражении фосфорелирования самого Rad3, так как большая полоса выше 200RD, которая является фосфорелированной частью, может обнаруживаться с помощью анализа Вестерн блоттинга с анти-НА антителом (данные не показаны). Попытки убедительной идентификации in vitro субстратов, таких, как миелиновый основной белок, RP-A и некоторых очищенных белков с S.pombe точкой контроля, как было доказано, оказались безуспешными. Если IP киназный анализ in vitro проводили с клетками, сверх-экспрессирующими “киназную-нуль” D2249E версию Rad3, ассоциированная киназная активность, ускоряемая НА-специфическим антителом, значительно понижается (данные не показаны). Этому имеется несколько возможных объяснений. Измеренная киназная активность может непосредственно отражать Rad3 активность. В этом случае, наблюдаемая остаточная активность с киназным мертвым Rad3 может отражать тот факт, что он является неизвестным для эквивалентных D-Е мутаций в других протеин киназах для продуцирования биологически инертного белка с остаточной in vitro биохимической активностью. Или же, киназная активность, которая позволяет фосфорелировать Rad3, может быть обусловлена ассоциированными белками, и эти белки могут взаимодействовать менее эффективно с D2249E мутантным белком.
Обсуждение
Пути точек контроля, контролирующие развитие клеточного цикла после повреждения ДНК или интерференцию в индивидуальных событиях, которые включает цикл, являются в значительной степени важными в поддержании генетической стабильности и могут считаться путями, которые подавляют онкогенез. Некоторые опухолевые супрессорные гены тесно включаются в субнаборы путей точки контроля (обзоры, приведенные в Hartwell and Kastan, 1994), особенно те, которые оказывают влияние на переход от G1 в S фазу и коммитируют в клеточный цикл. Конвергенция двух дрожжевых модельных систем для точек контроля четко указывает на то, что гены, включенные в эти пути, сохраняются. Настоящая работа расширяет это сохранение до метазоановых (metazoan) клеток, и проясняет отношение между rad3, ESR1/(MEC1/SAD3), mei-41 и АТМ геном.
В этой работе демонстрируется, что правильная последовательность rad3 гена помещает его продукт в семейство белок/липидных киназ родственных АТМ. Сверх-экспрессия киназно-поврежденного rad3 мутанта в S.pombe вызывает появление доминантного негативного фенотипа, которая предполагает, что Rad3 выступает в качестве компонента белкового комплекса, чья однородность является необходимой для функции точки контроля. Это согласуется с наблюдением о том, что rad1, rad9, rad17, rad26 и hus1 мутанты делеции все обладают фенотипами, неотличимыми от rad3.d (Sheldrick and Carr, 1993). Неожиданно, в отличие от остающихся rad генов точки контроля, высокий уровень сверх-экспрессии аллелей либо дикого типа, либо мутантных rad3 ингибирует клеточный рост и вызывает появление митоза при уменьшенном размере клетки, указывая на преждевременное вхождение в митоз. Этот “semi-wee” (“полу-крошечный”) фенотип не наблюдается в нуль мутанте и может указывать на вмешательство во второй путь, чья функция перекрывается с функцией Rad3 и выступает в ингибировании митоза. Кандидатом для такого пути является ATM/TEL1 путь, который, как показано, имеет некоторые функции, перекрывающиеся с ESR1(MEC1/SAD1) путем (Morrow et al., 1995).
Структура АТМ является наиболее близко родственной TelIp, который включается в поддержание длины теломера (Greenwell et al., 1995). Однако, АТМ функция оказывается также родственной функции Rad3/Esrlp/mei-41 продуктам. После первоначального открытия АТМ гена и его связи последовательности с rad3/ESR1 генами и с TEL1, не ясно, является ли ген, как во многих случаях дрожжей, дуплицированным или дивергированным в дрожжи, или же два дрожжевых белка определенно сохраняют субсемейство близко родственных генов. Значительной находкой этой работы является идентификация человеческого гена ATR, который является более родственным rad3/ESR1 /mei-41. Эта находка определяет два структурно четких родственных субсемейства с точкой контроля, субсемейства белок/липидных киназ, которые сохраняются на протяжении эукариотической эволюции. Хотя белки в этих двух субсемействах могут иметь некоторые перекрывающиеся функции, они, вероятно, контролируют различные процессы. Например, rad3 суб-семейство в дрожжах контролирует все G1 и G2 точки контроля повреждения ДНК в ответ на УФ и ионизирующее излучение, и точке контроля S фазы, которая предотвращает митоз после ингибирования репликации (AI-Khodairy and Carr, 1992; Allen et al., 1994; Weinert et al., 1994). Напротив, А-Т клетки имеют аномальный ответ в узкой области ДНК повреждающих агентов, включая ионизационное излучение, повреждение блеомицином и неокарциностатином, которые продуцируют разрушение цепи в ДНК как следствие радикальной атаки. Ответ на УФ излучение и большинство химических карциногенов является обычным, как и ответ на ингибирование синтеза ДНК. Возможно, что некоторые или все остающиеся точки контроля повреждения ДНК и точка контроля S фазы контролируются ATR.
Экспериментальная часть
Штаммы, плазмиды и среда
Стандартные генетические процедуры, условия роста и среда для S.pombe описываются в Gutz et al. (1974). S.pombe штамм sp011 (ura4.D18, leu1.32 ade6.704 h) были описаны ранее (Murray et al., 1992). Плазмида pSUB41 была подарена S.Subramani (Seaton et al., 1992).
Клонированив S.pombe rad3
4.0 kb Kpn1 фрагмент был вырезан из pSUB41 и секвенирован в обоих направлениях для получения 5’ rad3 последовательности. 3’ клон был идентифицирован из геномной библиотеки (Barbet et al., 1992) путем гибридизации колонии с использованием 1 kb 3’ пробы, полученной из опубликованной rad3 последовательности и секвенированной в обоих направлениях. Таким путем была изготовлена последовательность полного rad3 гена.
Нуль и “мертвые киназные” rad3 мутанты
Конструкция rad3, в которой 794 аминокислот между аа1477 и аа2271 (включая киназный домен) заменяли ura4+ геном, создавалась с использованием методологии, описанной в Barbet et al. (1992). Линейный фрагмент этой конструкции использовали для трансформирования sp011 для урацил прототропии и единичная копия интеграции при rad3 локусе проверяли с помощью анализа Саузерн блоттинга. Для создания сайт специфических киназных нуль мутаций, С-терминальный 3.01 kb BamHI-SaI1 фрагмент rad3 был подвергнут мутации либо с (A: GTTTTGGCCAGGCGCGCTCCCAAACCCAA, B: TTCATCAAACAATATCTTTTCGCCATGGCG, или C: CAAAAAGACAGTTGAATTCGACATGGATAG) для того, чтобы ввести либо D2230A, N2235K или D2249E мутации в киназный домен. Аналогичные изменения были ранее использованы в анализе Р13 киназы VPS34 S.cerevisiae (Schu et al., 1993). Эти фрагменты затем были использованы для трансформирования rad3 нуль мутанта и замещения гена, выбранного с помощью их способности к росту на FOA содержащей среде (Grimm et al., 1988). Все штаммы были проанализированы с помощью анализа Саузерн блоттинга. Экспрессионные конструкции полной длины rad3.D2230A были созданы в pREP1 и pREP41 (Maundrell, 1990) с помощью стандартного субклонирования после введения Ndel сайта при ATG и делеции трех внутренних Ndel сайтов.
Стрип тесты на чувствительность к УФ излучению
Экспрессия из REP1 (высокого) и REP41 (промежуточного) была индуцирована при отсутствии тиамина в течение 18 часов до посева. Чашки были подвергнуты облучению с градиентом дозы УФ облучения вдоль чашки от 0 до 300 Jm-2 в соответствии с наборами на Stratagene Stratalinker.
Клонирование и экспрессия ATR
Для выделения соответствующей пробы для идентификации кДНК-т, соответствующих человеческому rad3 гомологу, вырожденные олигонуклеотиды были сконструированы по отношению к аминокислотам LGLGDRH (5’ олиго; oDH18) и HVDF[D/N]C (3’ олиго; oDH-16) Rad3/Esrlp. Инозин вводили в положения четырехкратной дегенерации, и праймеры были наращены BamHI (oDH18) и EcoRI (oDH16) для облегчения клонирования. Анализ ДНК последовательности ~ 100 bp PCR продукта, полученного при амплификации кДНК периферийных лейкоцитов крови, продемонстрировал значительное подобие MECl/rad3. Эта последовательность была использована для синтеза ненарушенного праймера (oDH-23; GACGGAGAATTCACCAGTCAAAGAATCAAAGAG) для PCR с дополнительным нарушенным праймером (oDH17), сконструированным по отношению к аминокислотной последовательности KFPP/[I/V][L/F]Y[Q/E]WF Rad3/Esrlp. Продукт 174 bp этой реакции был использован непосредственно для скринирования микрофаговой библиотеки кДНК. Были выделены четыре положительных клона (самый большой приблизительно 3kb).
Параллельно, с помощью базы данных исследовали S.pombe rad3 полной длины, полученный из EMBL основных данных человеческого кДНК клона, HSAAADPDG, в качестве потенциального гомолога rad3, если позволял единичный сдвиг рамки для последовательности в 233 bp. Эта 233 bp последовательность содержится внутри 1.6 kb клона, полученного от Dr.N.Affara, Human Molecular Research Group, Cambridge University, UK. Полный клон (1.6 kb) был секвенирован и лежал в пределах кДНК клонов, идентифицированных за счет нарушения PCR и скринирований бибилиотеки. Для идентификации всего гена, были проведены RACE PCR эксперименты на кДНК, полученной из плацентальной и тимусной мРНК с использованием инструкций, обеспеченных Clontech Marathon Kit. Были получены генные специфические праймеры из кДНК клонов. Из этих экспериментов была составлена 8239bр кДНК последовательность с внутренними ORF 2644 аминокислотами, 79 bp 5’ некодирующей областью, 194 bp 3’ некодирующей областью и поли А- хвостом. Части последовательности были определены исключительно с помощью PCR. Для того чтобы избежать ошибок, клоны из минимум трех независимых PCR реакций были секвенированы в обоих направлениях. 233 bp последовательность соответствовала последовательности нуклеотидов 6809-7042 (234-тому в общем) Seq.ID No.1, за исключением единичной основной делеции в положении 6942. Эта последовательность кодировала аминокислоты 2244-2320 Seq.ID No.2.
Последовательность “1.6 kb” инсерта соответствовала нуклеотидам 5725-7104 (1353 - ему) Seq.ID No.1 и кодировала аминокислоты 1892-2340 Seq.ID No.2.
Гибридизация Нозерн блоттинг: 1.3kb PCR продукт был амплифицирован в присутствии 32P-dCTP с использованием праймеров 279-3 (TGGATGGATGATGACAGCTGTGTC) и 279-6 (TGTAGTCGCTGCTGCTCAATGTC). Нейлоновая мембрана, содержащая 2 мкг фракционированной по размеру полиА+РНК из различных источников человеческой ткани (Clontech Laboratories) была апробирована как предложено производителем, за исключением того, что конечную промывку проводили скорее при 55°С, чем при 50°С для снижения возможности перекрестной гибридизации с родственными последовательностями.
Картирование ATR
Мы картировали ATR ген в хромосому 3 за счет комбинации флуоресцентной in situ гибридизации и реакции полимеразной цепи (PCR), основанной на анализах. FISH анализ, использующий кДНК клон, идентифицировал ATR ген на хромосоме 3, приблизительно в положении q22-23. PCR также идентифицировал ATR на хромосоме 3. Два праймера (oATR23: GACGCAGAATTCACCAGTCAAAGAATCAAAGAG и oATR26:
TGGTTTCTGAGAACATTCCCTGA), которые амплифицировали 257 bp фрагмент ATR гена, были использованы на ДНК, полученной из гибридов человеческой/грызуновой соматической клетки, содержащей различные человеческие хромосомные панели, доступные из NIGMS Human Genetic Mutant Cell Repository (Drwinga et al., 1993). PCR с теми же самыми праймерами была использована для суб-локализации ATR в специфическую область на хромосоме 3. Матрицы для этих амплификаций состояли из ДНК образцов от пациентов с усечениями вдоль хромосомы 3 (Leach et al., 1994).
Иммунопреципитация (ШЗ) и анализы киназы RAD3
S.pombe rad3 и человеческие ATR гены были клонированы в pREP41 векторе экспрессии для исследований комплементации. Для того чтобы пометить белки, версии этих векторов, содержащих в рамке N-терминальные последовательности-метки, были использованы либо двойная myc, либо тройная НА метка (Griffiths et al., 1995). Меченые белки были экпрессированы за счет роста в среде без тиамина (Maundrell, 1990). Клетки дрожжей лизировали в лизисном буфере (25 мМ ТрисСl рН 7.5. 60 мМ В-глицерофосфата, 0.1 мМ Na3VO4, 1% Тритон Х-100, 50 мМ NaCl, 2 мМ EDTA, 50 мМ NaF, 1 мМ фенилметилсульфонилхлорида [PMSF], 5 мкг/мл лейпептина, 5 мкг/мл апротина, 1 мМ DTT) путем добавления стеклянных гранул с последующей обработкой в устройстве для измельчения в течение 2 минут. Для IP 300 мкг полный белковый экстракт был проинкубирован во льду соответствующим антителом в течение 30 минут и иммунные комплексы осаждались путем смешения с гранулами Protein G в течение дальнейших 30 минут при 4°С. Для киназных анализов, иммунные комплексы промывали 4 раза лизисным буфером, один раз киназным буфером (25 мМ Hepes, рН 7,7; 50 мМ KCl, 10 мМ MgCl2, 0.1% NP-40 2% глицерина, 1 мМ DTT), и инкубировали в киназном буфере с 10 мкМ АТР [50 Сi/ммоль] в течение 15 минут при 30°С. Реакции останавливали добавлением 20 унл 2Х SDS буфера образца до разделения на 6% полиакриламидных гелях. Rad3 IP’s содержали несколько фосфорелированных продуктов, включая один, который совместно мигрировал с самим Rad3 белком в анализах Вестерн блоттинга.
Источники информации
Al-Khodairy, F., and. Carr, A.M. (1992). DNA repair mutants defining G2 checkpoint pathways in Schizosaccharomyces pombe. EMBO J.11, 1343-1350.
Al-Khodairy, F., Fotou, E., Sheldrick. K.S., Griffiths, DJ.F., Lehmann, A.R. and Саrr, А.М. (1994). Identification and characterization of new elements involved in checkpoints and feedback controls in fission yeast Mol. Biol. Cell 5, 147-160.
Allen, J.B., Zhou. Z., Siede. W., Friedberg, B.C. and Elledge. SJ. (1994) The SAD1/RAD53 protein kinase controls multiple checkpoints and DNA damage-induced transcription in yeast. Genes Dev.8, 2416-2428.
Barbet, N.C., Muriel, W.J., and Can, A.M. (1992) Versatile shuttle vectors and genomic libraries for use with Schizosaccharomyces pombe. Gene 114, 59-66.
Beamish, H. and Lavin, M.F. (1994) Radiosensitivity in ataxia-telangiectasia: anomalies in radiation-induced cell cycle delay. Int. J. Radiat Biol. 65, 175-184.
Carr, A.M. and Hoekstra, M.F. (1995) The Cellular Responses to DNA Damage. Trends in Cell Biology 5, 32-40.
Chien et al., (1991) Proc. Natl. Acad Sci USA 88, 9578-9582.
Deng, С., Zhang, P., Harper, J.W., EIledge. S.J. and Lcder. PJ. (1995) Mice lacking p21CIPl/WAFl undergo nonmal development, but are defective in Gl checkpoint control. Cell (in press).
Drwinga. H.L., Tojia, L.H., Kim, C.H, Grcene, A.E., and Mulovor, R.A. (1993). NIGMS Human/Rodent Somatic Cell Hybrid Mapping Panels 1 and 2. Genomics 16:311-314.
El-Deiry. W.S., Tokino, Т., Velculescu, V.E., Levy, D.B., Parson, R., Trent. J.M., Lin. D., Mercer, W.E, Kinder, K.W. and Vogelstein, B. (1993) WAF1. a potential mediator of p53 tumour suppression. Cell 75, 817-825.
Enoch, Т., Carr, A.M. and Nurse, P. (1992). Fission yeast genes involved in coupling mitosis to completion of DNA-replication. Genes Dev.6, 2035-2046.
Greenwell. P.W., Kronmal, S.L., Porter. S.E., Gassenhuber. J., Obermaier. B. and Petes, T.D. (1995) TEL I, a gene involved in controlling telomere length in Saccharomyces cerevisiae, is homologous to the human ataxia telangiectasia (ATM) gene. Cell submitted.
Grimm, C., Kholi, J. Murray, J.M. and Maundrell, K. (1988) Genetic engineering of Schizosaccharomyces pombe: a system for gene disruption and replacement using the ura4 gene as a selectable marker. Mol. Gen. Genet 275, 81-86.
Gutz, H., Heslot, H. Leupold, U. and Loprieno, N. (1974). In "Handbook of Genetics", King R.C., Ed., Plenum Press, New York, Vol.7, 395-446.
Hari, K.L, Santerre, A., Sekelsky, J.J., McKim, K.S., Boyd, J.B, and Hawiey, R.S. (1995) The mei-41 gene of Drosophila melanogaster is functionally homologous to the human ataxia teiangiec Harper, J.W., Adami. G., Wei, N., Kcyomarsi, K. and EIledge, SJ. (1993) The 21 kD Cdk interacting protein Cipl is a potent inhibitor of G1 cyclin dependent kinases. Cell 75, 805.816.
Harwell, L.H., and Kastan. KB. (1994). Cell cycle control and Cancer. Science 266, 1821-1828.
Hiles, LD., Otsu, M., Volinia, S., Fry, M.J., Gout, I., Dhand, R., Panayotou, G., Ruiz-Larrea, F., Thompson, A., Totty, N.F., Hsuan, J.J., Courtneidge, SA., Parker. P.J. and Waterfield M.D. (1992) Phospbatidylinositol 3-kinase: structure and expression of the 110kd catalytic subunit. Celt 70, 419-429.
Jimenez, G., Yucel. J., Rowley, R. and Subramani S. (1992) The rad3+ gene of Schizosaccharomyces pombe is involved in multiple checkpoint functions and in DNA repair. Proc Natl. Acad. Sci. USA 87, 4952-4956.
Kato, R. and Ogawa, H. (1994) An essential gene, ESR1, is required for mitotic cell growth, DNA repair and Meiotic recombination in Saccharomyces cerevisiae. Nucleic Acids Res. 22, 3104-3112.
Lamb, J.R., Petit-Frere, C., Broughton, B.C., Lehmann, A.R. and Green, M.H.L. (1989) Inhibition of DNA replication by ionizing radiation is mediated by a trans acting factor. Int. J. Radiat. Biol. 56, 125-130.
Leach, R.J., Chinn, R., Reus, B.E., Hayes, S., Sehantz. L., Dubois, В., Overhauser, J., Ballabio, A., Drabkin, H., Lewis. B.T., Mendgen, G., and Naylor, S.L. (1994) Regional Localisation of 188 Sequence Tagged Sites on a Somatic Cell Hybrid Mapping Panel for Human Chromosome 3 Genomics 24, 549-556.
Maundrell, К. (1990). nmtl of fission yeast. A highly transcribed gene completely repressed by thiamine. J. Biol. Chem. 265. 10857-10864.
Morrow. D.M., Tagle, D.A., Shiloh. Y., Collins F.S. and Hieter, P. (1995) HATlITELl, a Saccharomyces cerevisiae homologue of the human gene mutated in ataxia-telangiectasia, is functionally related to the yeast checkpoint gene MECIIESR1. Cell submitted.
Murray, J.M., Doe, C. Schenk, P. Carr, A.M. Lehmann, A.R. and Watts, F.Z. (1992) Cloning and characterization of the S. pombe rad15 gene, a homologue to the S. cerevisiae RAD3 and human ERCC2 genes Nucleic Acids Res. 20, 2673.2678.
Nasim, A. and Smith, B.P. (1975) Genetic control of radiation sensitivity in Schizosaccharomyces pombe. Genetics 79, 573-582.
Painter, R.B. and Young, B.R. (1980) Radiosensitivity in ataxia - telangiectasia: A new explanation. Proc. Natl. Acad. Sci. USA. 77, 7315-7317.
Rowley, R., Subramani. S. and Young, P.G. (1992). Checkpoint controls in Schizosaccharomyces pombe, radl. EMBO J. 11, 1335-1342.
Savitsky. К., Bar-Shira, A., Gilad, S., Rotman. G., Ziv, Y., Vanagaite L., Tagle, D.A., Smith, S., Uziel, Т., Sfez, S., Ashkenazi, M., Pecker, I., Frydman, M., Harnik, R., Patanjali, S.R., Simmons, A., Clines, G.A., Sartiel, A., Gatti, R.A., Chessa, L., Sanal, O., Lavine, M.F., Jaspers, N.G.J., Taylor, M.R., Arlett, C.F., Miki, Т., Weissman. S.M., Lovett, M., Collins, F.S. and Shiloh, Y. (1995). A single ataxia telangiectasia gene with a product similar to PI-3 kinase Science 286, 1749-1753.
Seaton, B.L., Yucel, J., Sunnerhagen P. and Subramani, S. (1992). Isolation and characterisation of the Schizosaccharomyces pombe rad3 gene which is involved in the DNA damage and DNA synthesis checkpoints. Gene 119, 83-89.
Schu, P.V., Takegawa, K., Fry, M.J., Stack, J.H., Waterfield. M.D. and Emr, S.D. (1993) Phosphatidylinositol 3-kinase encoded by yeast VPS34 gene essential for protein sorting. Science 260, 88-91.
Sheldrick, K.S. and Carr. A.M. (1993). Feedback controls and G2 checkpoints, fission yeast as a model system. BioEssays 15, 775-782.
Walworth. N., Davey, S. and Beach, D. (1993). Fission yeast chkl protein kinase links the rad checkpoint pathway to cdc2. Nature 363, 368-371.
Weinert, T.A., and Hartwell, L.H. (1988). The RAD9 gene controls the cell cycle response to DNA damage in Saccharomyces cerevisiae. Science 241, 317-322.
Weinert, T.A., Kiser, G.L. Hartwell, L.H. (1994). Mitotic checkpoint genes in budding yeast and the dependence of mitosis on DNA replication and repair. Genes Dev.8, 652-665.










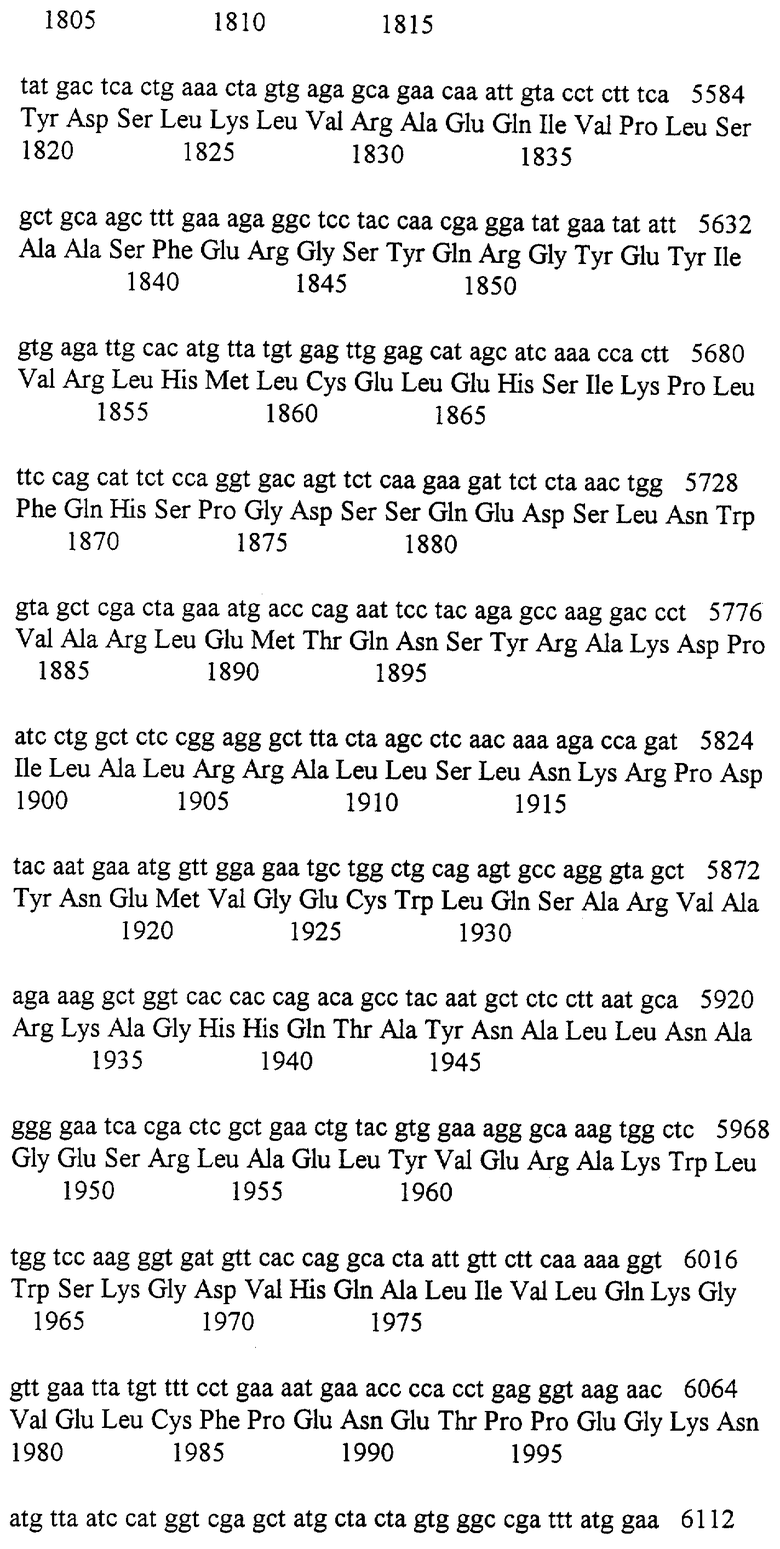

















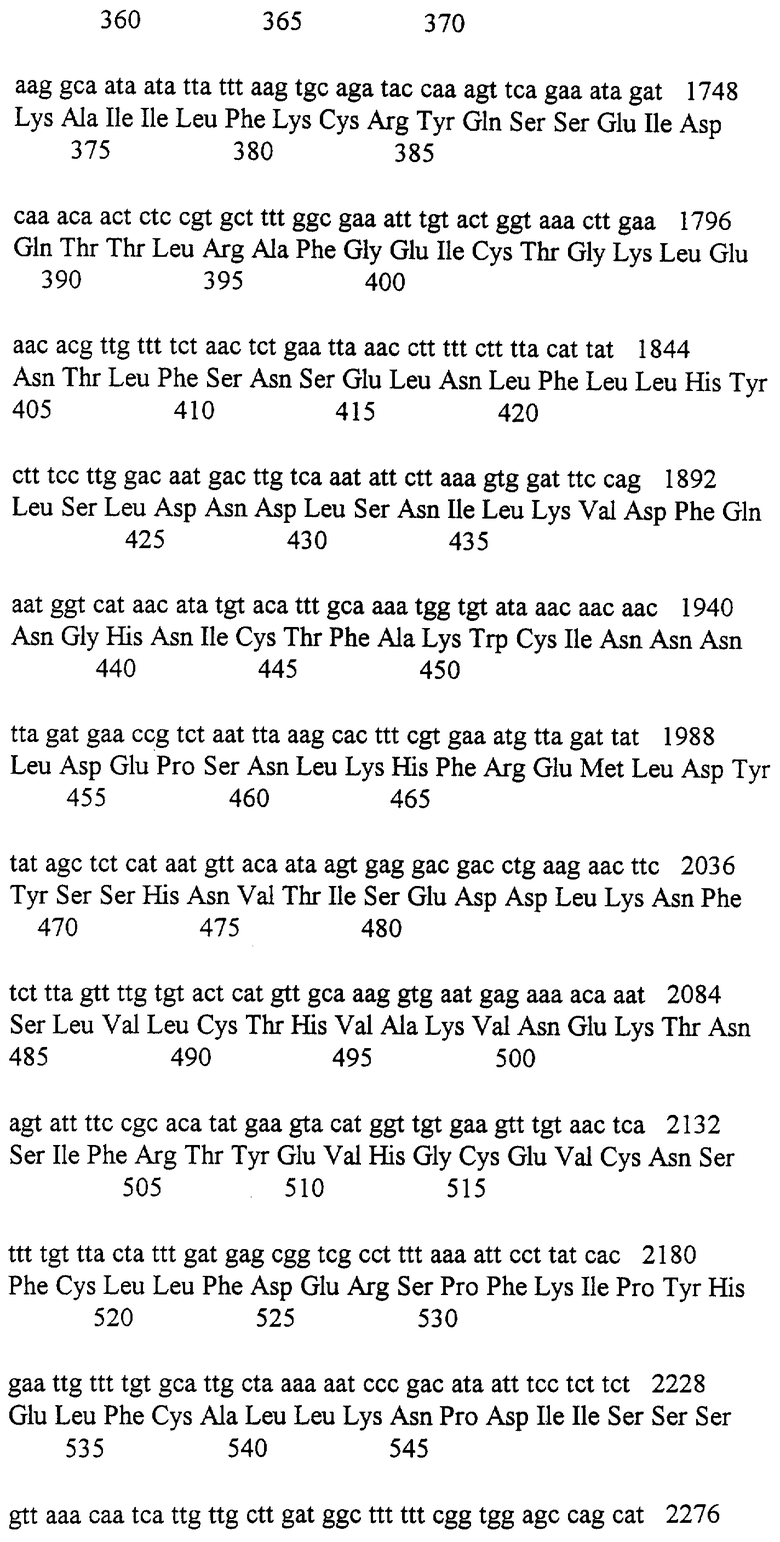

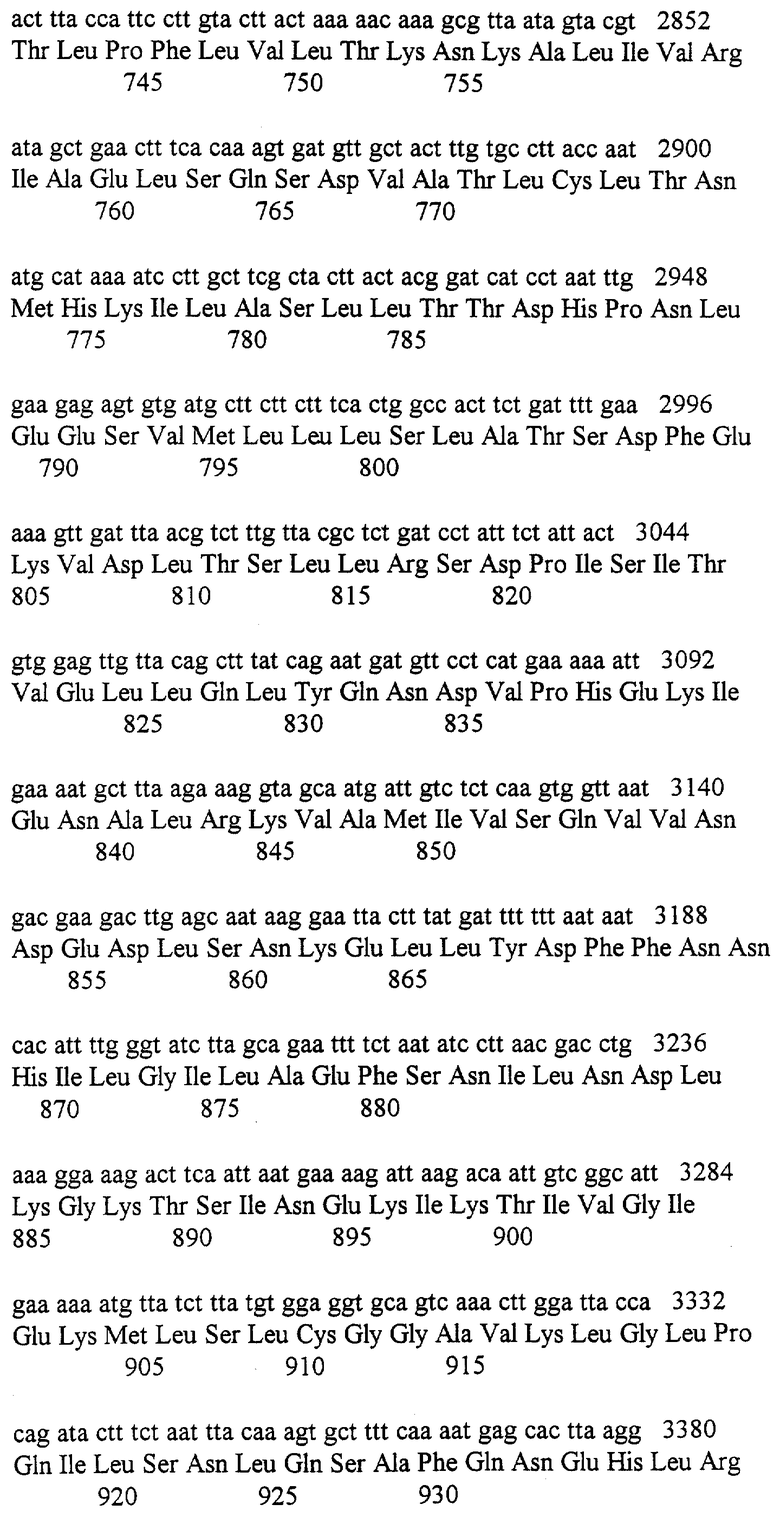





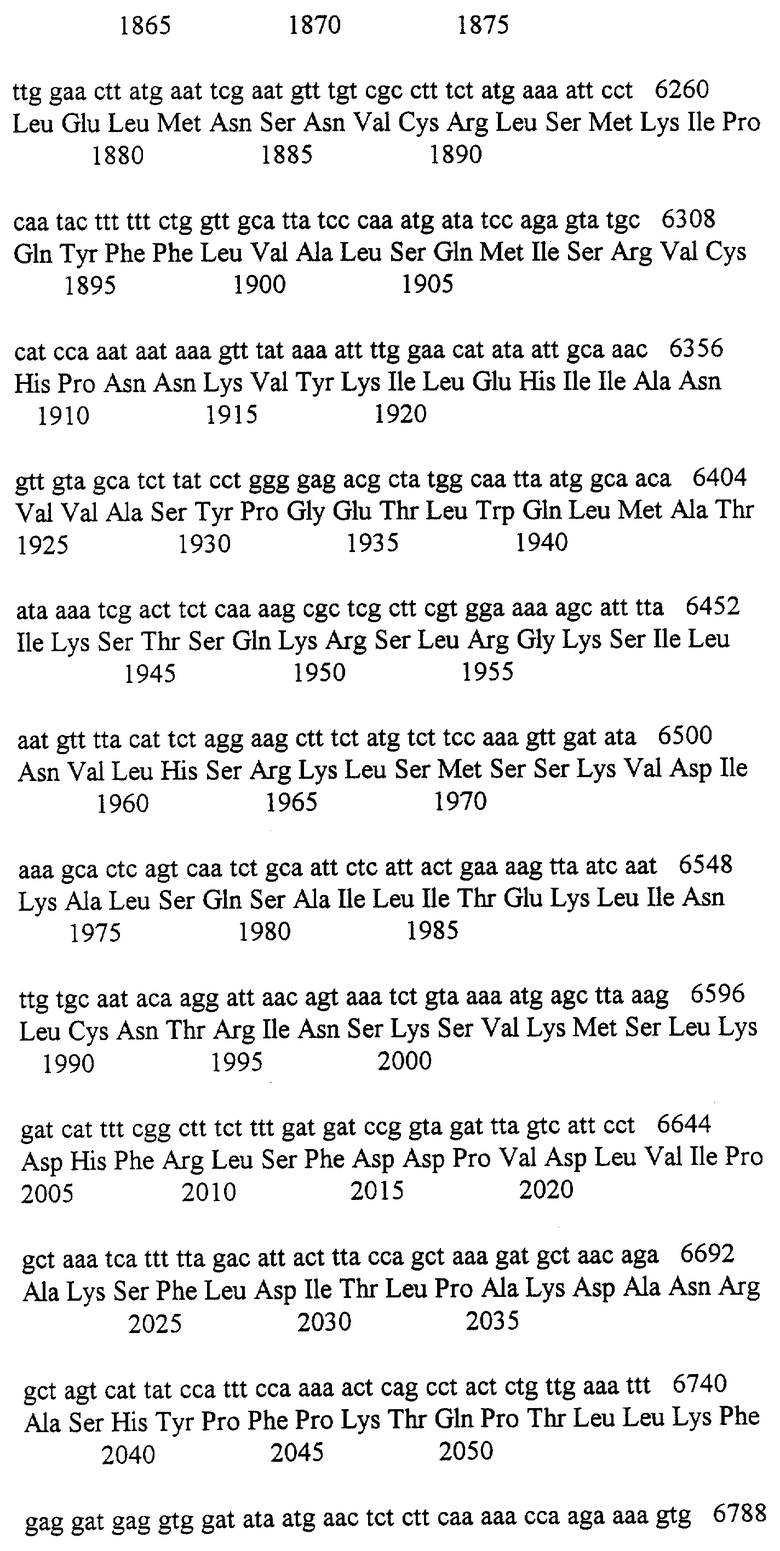












Изобретение относится к генной инженерии, конкретно, к генам точки контроля клеточного цикла и может быть использовано in vitro или in vivo в системах клеточной культуры для изучения роли ATR в качестве гена точки контроля. Полинуклеотид, кодирующий гомолог ATR полипептида rad3, клонируют в экспрессирующий вектор, функциональный в эукариотических клетках. Полипептид гомолога ATR полипептида rad3 получают путем культивирования эукариотической клеточной культуры, трансформированной вектором. Моноклональное антитело к гомологу ATR полипептида rad3 получают с помощью гибридомных технологий. Путем инокуляции полипептида гомолога ATR полипептида rad3 животному-хозяину получают поликлональные антитела. Присутствие полинуклеотида в образце животной ткани обнаруживают путем контактирования данного образца, содержащего ДНК или РНК, с полинуклеотидом, кодирующим гомолог ATR полипептида rad3, в условиях гибридизации. Полипептид в биологическом образце обнаруживают путем контактирования образца с моноклональными или поликлональными антителами. Проводят отбор веществ - кандидатов как антираковых агентов, судя по уменьшению активности полипептида ATR на субстрате или уменьшению комплексообразования гомолога ATR в присутствии веществ-кандидатов. Изобретение позволяет получить гомологи ATR полипептида rad3 человека или S.pombe. 22 н. и 2 з.п. ф-лы, 1 ил.
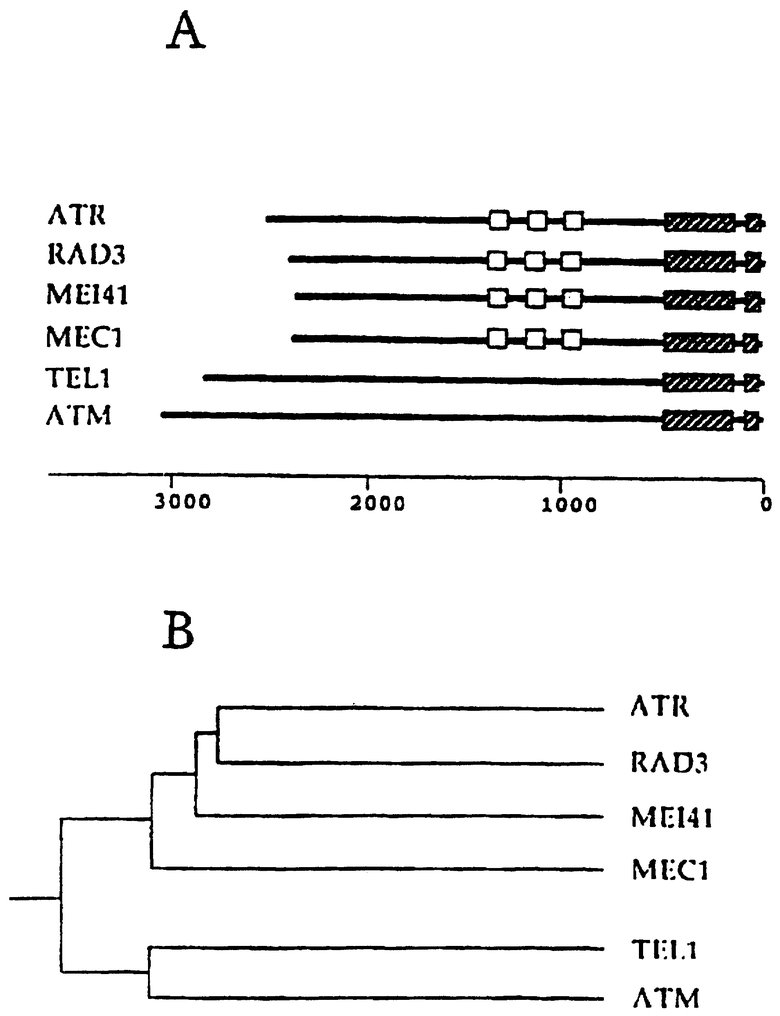
a) полинуклеотида, содержащего последовательность нуклеотидов, представленную в SEQ ID NO: 1; и
b) полинуклеотида, который, по меньшей мере, на 70% является гомологичным полинуклеотиду в а) и который селективно гибридизуется с комплементом полинуклеотида а) в условиях, включающих в себя конечную промывку при 55°С, и который кодирует гомолог ATR, имеющий липидкиназную активность.
a) контактирования образца тела человека или животного, содержащего ДНК или РНК, с пробой, содержащей полинуклеотид по п.3 или его фрагмент, в условиях гибридизации;
b) обнаружения в образце любого дуплекса, образованного между пробой и нуклеиновой кислотой.
a) контактирования биологического образца с антителом по п.6 или 7 в условиях, которые позволяют образовывать комплекс антитело-антиген;
b) определения того, образован ли комплекс антитело-антиген, содержащий это антитело.
a) контактирования полипептида ATR по п.4 и субстрата для указанного полипептида в присутствии или отсутствии вещества-кандидата;
b) идентификации указанного вещества-кандидата в качестве антиракового агента при обнаружении уменьшенной активности полипептида ATR на субстрате в присутствии вещества-кандидата в сравнении с активностью ATR в отсутствии вещества-кандидата.
a) контактирования первого полипептида по п.4 со вторым полипептидом, способным взаимодействовать с гомологом ATR, в условиях, позволяющих образование комплекса между первым и вторым полипептидами и в присутствии и отсутствии вещества-кандидата; причем указанный второй полипептид тождественен первому полипептиду или отличен от него;
b) идентификации вещества-кандидата в качестве антиракового агента, причем уменьшенное образование комплексов между первым полипептидом и вторым полипептидом обнаруживается в присутствии соединения-кандидата в сравнении с образованием комплексов в отсутствии вещества-кандидата.
a) полинуклеотида, содержащего последовательность нуклеотидов, представленную в SEQ ID NO: 3;
b) полинуклеотида, который, по меньшей мере, на 70% является гомологичным полинуклеотиду в а) и который селективно гибридизуется с комплементом полинуклеотида а) в условиях, включающих в себя конечную промывку при 55°С, и который кодирует гомолог ATR, имеющий липидкиназную активность.
a) контактирования образца тела человека или животного, содержащего ДНК или РНК, с пробой, содержащей полинуклеотид по п.15 или его фрагмент в условиях гибридизации;
b) обнаружения в образце любого дуплекса, образованного между пробой и нуклеиновой кислотой.
a) контактирования биологического образца с антителом по п.18 или 19 в условиях, позволяющих образование комплекса антитело-антиген;
b) определения, образовался ли комплекс антитело-антиген, содержащий указанное антитело.
а) контактирования полипептида ATR по п.16 с субстратом для указанного полипептида в присутствии и в отсутствии вещества-кандидата;
б) идентификации указанного вещества-кандидата в качестве антиракового агента при обнаружении уменьшенной активности полипептида ATR на субстрате в присутствии вещества-кандидата в сравнении с активностью ATR в отсутствии вещества-кандидата.
a) контактирования первого полипептида по п.16 со вторым полипептидом, способным взаимодействовать с гомологом ATR, в условиях, позволяющих образование комплекса между первым и вторым полипептидами, и в присутствии и в отсутствии вещества-кандидата, причем указанный второй полипептид тождественен первому полипептиду или отличен от него;
b) идентификации вещества-кандидата в качестве антиракового агента, причем уменьшенное образование комплекса между первым полипептидом и вторым полипептидом обнаруживается в присутствии вещества-кандидата в сравнении с образованием комплекса в отсутствии вещества-кандидата.
| SEATON B.L | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Gene, 1992, v.119, n.1, p.83-89 | |||
| VLAHOS C.J | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| J | |||
| Biol | |||
| Chem | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |

