Область изобретения
Настоящее изобретение относится к новому микроорганизму, новым ферментам и новой смеси ферментов. Кроме того, настоящее изобретение относится к композиции смеси ферментов, к ее получению и использованию в пищевой промышленности, в производстве кормов и в других отраслях промышленности, включая (но не ограничиваясь ими) бумажную и текстильную промышленность.
Предпосылки изобретения
В течение длительного времени ферменты использовали для широкого круга различных промышленных применений. Известны примеры из хлебопекарной промышленности, виноделия и производства фруктовых соков (где ферменты используют для разрушения пектинов и β-глюканов), текстильной промышленности (где целлюлазы используют для получения мягких и гладких целлюлозных тканей) и также, что является отнюдь не последним применением, для приготовления кормов животным. В этом случае ферменты улучшают переваривание кормов из растительных источников.
Последнее применение обеспечивает более эффективное переваривание животными корма. Ценность корма можно определить по FCR (Feed Conversation Ratio = затраты корма на единицу продукции), питательной ценности, количеству расходуемого корма по отношению к увеличению массы животного. Снижение величины FCR для корма указывает на пропорциональное увеличение массы животного, т.е. на то, что животное способно использовать корм более эффективно.
Плохая усвояемость компонентов корма (крахмал, жир, протеин/аминокислоты) является отличительной особенностью кормов на основе злаковых и, например, особенно тех, в которых высоко содержание ячменя или пшеницы. В этих случаях может оказаться необходимым подготавливать корма так, чтобы они содержали более высокие уровни энергии из других источников и других добавок, таких как аминокислоты. Эти ферменты повышают величину очевидной обменной энергии (Apparent Metabolizabie Energy), включенных в корм злаковых.
Другим подходом к решению этой проблемы было введение ферментных добавок, целлюлаз, эндо-1,3(4)-β-глюканаз (β-глюканазы), эндо-1,4-β-ксиланаз (ксиланазы) и т.д. или смесей ферментных активностей к таким кормам на основе злаковых. Ферментные добавки могут иметь специфическое применение для гидролиза β-глюканов или для гидролиза арабиноксиланов, находящихся в злаковых (обычно в ячмене и пшенице). Добавление ферментов имеет различные цели. Одним из преимуществ, которое, безусловно, обеспечивает эффективность ферментных добавок к кормам, является снижение вязкости материалов в пищеварительном тракте животных, которые получают корм на основе злаковых, содержащий соответствующую ферментную добавку. Более высокая вязкость обусловлена, частично, присутствием β-глюканов и арабиноксиланов в ячмене и пшенице. Более низкая вязкость, являющаяся результатом действия ферментов, обеспечивает более легкую абсорбцию питательных компонентов в пищеварительном тракте животных. Другим преимуществом является выделение питательных веществ, заключенных в стенках клеток злаковых, что снижает необходимость в других дорогостоящих кормовых добавках. Общим результатом является значительное снижение стоимости кормов с аналогичным или более высоким эффектом по измерениям FCR.
Было раскрыто, что ферментные препараты, полученные из широкого круга различных микроорганизмов, повышают усвояемость кормов.
С учетом известного уровня, связанного с использованием ферментов в кормах для животных, авторы указывают Европейскую патентную заявку №0699762, в которой раскрыто использование фитазы, выделенной из Schwanniomyces occidentalis. Эта фитаза представляет собой фитазу, полученную из генетически модифицированного организма, полученного путем встраивания клонированного гена, чего авторы хотели бы избежать в настоящем изобретении.
Что касается патентной заявки WO 95/26398, там также модифицированную целлюлазу получают путем встраивания чужеродной ДНК последовательности в клетку-хозяина, которая модифицирует природу исходного штамма, который выбран из следующего перечня микроорганизмов: Bacillus, Streptomyces, Saccharomyces, Schizosacch aromyces, Aspergillus. Основной целью авторов настоящего изобретения было избежать встраивания чужеродного гена в микроорганизм, который продуцирует фермент.
В патентной заявке WO 96/05739 смесь ферментов (ксиланазу, протеазу и, необязательно β-глюканазу) получают из различных микроорганизмов. Авторы приводят пример (стр.5) смеси ферментов с отношением ксилазной активности к β-глюканазной активности порядка 1:5. Было обнаружено, что когда ксилаза включена в рацион на основе злаковых в оптимальной дозе или около этого, совместное присутствие ферментов, процессирующих β-глюканазную активность, повышает FCR корма, что, естественно, невыгодно. Соответственно, авторы выступают против присутствия β-глюканазы, они рекомендуют максимальное отношение ксиланазной активности к β-глюканазной активности как 1:0-0,25.
В некоторых случаях, для того, чтобы обеспечить присутствие всех ферментных активностей, связанных с применением кормов, приходится получать препараты из препаратов, полученных более чем из одного микроорганизма. В ряде случаев ферментные препараты были получены из микроорганизмов, подвергшихся генетическим модификациям с использованием рекомбинантных ДНК методик.
Авторы обнаружили новый микроорганизм, принадлежащий к классу Penicillum funiculosum, который содержит новые ферменты, и разработали смесь ферментных активностей, которую с успехом можно использовать для повышения, главным образом, усвояемости кормов для животных на основе злаковых.
Краткое содержание изобретения
Соответственно, настоящее изобретение относится к новому микроорганизму, полученному из Penicillum funiculosum, и к способу культивирования этого микроорганизма и выделения ферментов, продуцируемых этим микроорганизмом.
Кроме того, в соответствии с настоящим изобретением предложены новые ферменты, выделенные из этого микроорганизма, их последовательности нуклеиновых кислот и новые композиции, содержащие эти ферменты.
Далее, в соответствии с настоящим изобретением, предложен способ повышения усвояемости аминокислот и кормов для животных на основе аминокислот и злаковых.
Другой целью настоящего изобретения является уменьшение выделения фосфора и выделения аммиака из клеточной батареи, где кормят животных.
Подробное описание изобретения
А. Новый штамм Penicillum funiculosum
Этот новый штамм грибков Penicillum funiculosum депонирован под регистрационным номером IМI378536 в измассатном Международном Депозитарии в соответствии с Будапештским Соглашением (1977), Международным Институтом Микологии (International Mycological Institute (IMI), Bakeham Lane, Englefield Green, Egham, Surrey, TW209TY, UK).
Происхождение
Новый штамм получен из Penicillum funiculosum IMI 134756 после успешного УФ облучения и β-облучения спор, включая скринирование на селективной среде. Никаких генетических модификаций не было получено с помощью рекомбинантных ДНК методик с использованием встраивания чужеродных ДНК или РНК.
Идентификация и типирование
Penicillum funiculosum IMI 134756 был охарактеризован путем выращивания на Czapek Dox агаре при 25°С. Характеристики колонии и микроморфология типичны для Penicillum funiculosum. Идентификация микроорганизма как Penicillum funiculosum была подтверждена в Международном Институте Микологии (International Mycological Institute (IMI), Bakeham Lane, Englefield Green, Egham, Surrey, TW209TY, UK). Рост похож на плотный базальный войлок с аэральным ростом в виде нитей или узелков гифов (funiculose), мицелий белого цвета с подложкой красного цвета в субстрате, поля наоборот бледные, но окрашены красным по направлению к центрам, могут становиться темно-красными. Это типичный пенициллин, он демонстрирует короткий кондиофорез, возникающий, главным образом, из funicles, biverticillate, игольчатых конидиогенных клеток, конидии эллиптичные и гладкие.
Микроорганизм, использованный для получения препарата ферментов настоящего изобретения, выращивают в аэробных условиях в среде, которая содержит целлюлозу, сироп от замачивания кукурузы, карбонат кальция и сульфат аммония.
В. Процесс ферментации
скринирование на селективной среде. Никаких генетических модификаций не было получено с помощью рекомбинантных ДНК методик с использованием встраивания чужеродных ДНК или РНК.
Идентификация и типирование
Penicillum funiculosum IMI 378536 был охарактеризован путем выращивания на Czapek Dox агаре при 25°С. Характеристики колонии и микроморфология типичны для Penicillum funiculosum. Идентификация микроорганизма как Penicillum funiculosum была подтверждена в Международном Институте Микологии (International Mycological Institute (IMI), Bakeham Lane, Engiefield Green, Egham, Surrey, TW209TY, UK). Рост похож на плотный базальный войлок, с аэральным ростом, в виде нитей или узелков гифов (funiculose), мицелий белого цвета с подложкой красного цвета в субстрате, поля наоборот бледные, но окрашены красным по направлению к центрам, могут становиться темно-красными. Это типичный пенициллин, он демонстрирует короткий кондиофорез, возникающий, главным образом, из funicles, biverticillate, игольчатых конидиогенных клеток, конидии эллиптичные и гладкие.
Микроорганизм, использованный для получения препарата ферментов настоящего изобретения, выращивают в аэробных условиях в среде, которая содержит целлюлозу, сироп от замачивания кукурузы, карбонат кальция и сульфат аммония.
В. Процесс ферментации
Эти новые грибки получают путем ферментации депонированного штамма вначале на засеваемой среде, предпочтительно состоящей из (по массе):
Температура инкубирования 27°С-36°С
Ферментационная среда предпочтительно имеет следующий состав (по массе):
Температура инкубирования 27°С-36°С.
Для ферментации загружают ферментер достаточным количеством воды, добавляют ингредиенты к воде в подходящем контейнере с приспособлением для перемешивания, перемешивают до тех пор, пока ингредиенты не растворятся. Стерилизуют, герметизируя ферментер и повышая температуру содержимого обычно до 121°С. Содержимое ферментера инокулируют затравочной ферментационной средой.
Основным источником углерода, который добавляют в процессе ферментации, является целлюлоза; из различных источников целлюлозы авторы предпочитают использовать ARBOCEL, SOLKAFLOC CLAROCEL, ALPHACEL, FIBRACEL различных степеней чистоты.
Величину рН в процессе ферментации предпочтительно контролируют, добавляя серную кислоту или другую кислоту и аммиак в газообразном или жидком виде, или другое основание.
В конце времени ферментации твердые вещества удаляют фильтрованием или центрифугированием, собирают жидкую фазу и концентрируют, например, ультрафильтрацией или на органических или минеральных мембранах.
Эти ферменты можно также получить с помощью рекомбинантных ДНК методик, и таким образом они будут продуцироваться рекомбинантными гомологичными или гетерологичными видами. Хозяина для переноса гена, кодирующего фермент, можно выбрать из видов грибков, бактериальных клеток или растительных клеток. Для встраивания гена, кодирующего представляющий интерес фермент, в такие клетки-хозяева как плазмиды (интегративно или нет), векторы фагов и векторы вирусов, можно использовать любые удобные способы. Penicillum funiculosum, включающий вставку гетерологичных генов или модификацию генома гомологичными генами за счет вставок, делений или модификаций указанного гомологичного гена, также являются частью настоящего изобретения.
В соответствии с настоящим изобретением можно получить фермент в виде выделенного чистого препарата фермента или в виде неочищенного препарата, такого, как культуральная среда, в которой выращивали Penicillum funiculosum.
Можно также включить этот фермент или эти ферменты в композиции, содержащие еще один фермент, тип которого зависит от предполагаемого использования композиции. Добавляемые ферменты можно выбрать, например, из карбогидраз, липаз и протеаз.
С. Композиции, состоящие из "смеси ферментных активностей"
1. Жидкая композиция
Для жидкой композиции после добавления противомикробных агентов осуществляют измерение концентрации ферментов и разбавление до нужной концентрации. Предпочтительным составом жидкого раствора (по массе) является следующий:
Забуференный с доведением рН до 3-5
Противомикробный агент выбирают из таких продуктов, как сорбиновая кислота и ее соли, бензойная кислота и ее соли, метил-4-гидроксибензоат и н-пропил-4-гидроксибензоат, фумаровая кислота, соли и сложные эфиры. Можно также использовать такие соли, как хлорид натрия или хлорид калия.
Наиболее предпочтительными антифризами являются 1,2-пропандиол, этиленгликоль, глицерин.
2. Порошковая композиция
Для получения порошковых препаратов полученный концентрированный раствор сушат необязательно в присутствии носителя. Порошок, полученный в результате сушки концентрированного раствора без присутствия носителя, можно впоследствии смешать с подходящим носителем.
Предпочтительным составом порошковой формы композиции является следующий:
Предпочтительные носители выбирают из пшеничной муки, крахмала, гипса, мальтодекстрина, твердых продуктов кукурузы, побочных продуктов, полученных при обработке злаковых культур, такие, как кукурузная крупа, второсортная пшеница, пшеничные отруби, отходы при обработке риса, смесь минералов.
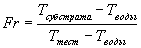
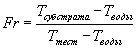
D. Характеристики ферментов
Получена новая смесь ферментов, продуцируемых Penicillum funiculosum. Эта смесь ферментов содержит новые ферменты, такие, как целлюлазы, β-глюканазы, ксиланазы, такие сопутствующие ксиланазные ферменты, как арабинофуранозидаза и ферулоил-эстеразы.
1. Процедура
Ферментный препарат охарактеризован в различных анализах, которые включают анализы активностей целлюлазы, целлобиогидролазы, β-глюкозидазы, эндо-1,3(4)-β-глюканазы, ламинариназо-эндо-1,4-β-ксиланазы (с использованием различных субстратов), β-ксилозидазы, арабинофуранозидазы и ферулоилэстеразы (с использованием различных субстратов).
1.1. DNS CMC способ анализа целлюлазы
Анализ активности целлюлазы основан на ферментном гидролизе гликозидных связей в карбоксиметилцеллюлозе (CMC), β-1,4-глюкане. Продукты реакции, олигосахариды β-1,4-глюкана, определяют по увеличению уровня восстановления (как глюкозу).
Раствор, содержащий 1 мл 1% (масса/объем) раствора CMC в 0,1 М натрийацетатном буфере, рН 5,0 (или другие значения рН); 1 мл соответствующим образом разбавленного ферментного раствора инкубируют при 50°С в течение 10 минут. Ферментную реакцию останавливают, добавляя 2 мл DNS раствора (1% (масса/ объем)) 3,5-динитросалициловой кислоты, 16% (масса/объем) гидроксида натрия, 30% (масса/объем) калий натрий (+)-тартрата в дистиллированной воде). Этот раствор перемешивают и помещают в баню с кипящей водой (минимум 95°С) на 5 минут, затем охлаждают до 25°С. К раствору добавляют 10 мл дистиллированной воды, и поглощение измеряют при 540 нм, используя стеклянную кювету толщиной 2 см.
Полученный результат превращают в мкмоли восстановленного сахара (как глюкоза), сравнивая со стандартной кривой для 2 мл 0,00-0,04% (масса/объем) растворов глюкозы, обработанных аналогичным образом раствором DNS.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой DNS раствор добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности целлюлазы определяют как количество фермента, которое продуцирует 1 мкмоль глюкозного эквивалента в минуту в условиях анализа (50°С и рН 5,0 или при других значениях рН).
1.2 Анализ целлобиогидролазы с использованием р-нитрофенил β-D-целлобиопиранозида
Анализ целлобиогидролазы основан на ферментном гидролизе р-нитрофенил β-D-целлобиопиранозида. Продукт реакции, р-нитрофенол, определяют колориметрически.
Раствор, содержащий 1 мл 0,1% (масса/объем) р-нитрофенил β-D-целлобиопиранозида в дистиллированной воде; 1 мл дистиллированной воды; 1 мл 0,2 М натрийацетатного буфера, рН 5,0; 1 мл соответствующим образом разбавленного ферментного раствора инкубируют при 50°С в течение 30 минут. Ферментную реакцию останавливают, добавляя 4 мл 0,4 М раствора глицина. Этот раствор перемешивают и охлаждают до 20°С. Поглощение измеряют на 400 нм, используя стеклянную кювету толщиной 1 см.
Полученный результат превращают в мкмоли р-нитрофенола, сравнивая с коэффициентом молярной экстинкции р-нитрофенола в этих же условиях.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой раствор глицина добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности целлобиогидролазы определяют как количество фермента, которое продуцирует 1 мкмоль р-нитрофенола из р-нитрофенил β-D-целлобиопиранозида в минуту в условиях анализа (50°С и рН 5,0).
1.3. Анализ β-глюкозидазы с использованием р-нитрофенил β-D-глюкопиранозида
Анализ β-глюкозидазы основан на ферментном гидролизе р-нитрофенил β-D-глюкопиранозида. Продукт реакции, р-нитрофенол, определяют колориметрически.
Раствор, содержащий 1 мл 0,1% (масса/объем) р-нитрофенил β-D-глюкопиранозида в дистиллированной воде; 1 мл дистиллированной воды; 1 мл 0,2 М натрийацетатного буфера, рН 5,0; 1 мл соответствующим образом разбавленного ферментного раствора инкубируют при 50°С в течение 30 минут. Ферментную реакцию останавливают, добавляя 4 мл 0,4 М раствора глицина. Этот раствор перемешивают и охлаждают до 20°С. Поглощение измеряют на 400 нм, используя стеклянную кювету толщиной 1 см.
Полученный результат превращают в мкмоли р-нитрофенола, сравнивая с коэффициентом молярной экстинкции р-нитрофенола в этих же условиях.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой раствор глицина добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности β-глюкозидазы определяют как количество фермента, которое продуцирует 1 мкмоль р-нитрофенола из р-нитрофенил β-D-глюкопиранозида в минуту в условиях анализа (50°С и рН 5,0).
1.4. Анализ эндо-1,3(4)-β-глюканазы с использованием DNS и β-глюкана ячменя
Анализ активности эндо-1,3(4)-β-глюканазы основан на ферментном гидролизе гликозидных связей в β-глюкане ячменя, β-1,3(4)-глюкане. Продукты реакции, олигосахариды β-1,3(4)-глюкана, определяют по конечному увеличению уровня восстановления (как глюкозы).
Раствор, содержащий 1 мл 1% (масса/объем) раствора β-глюкана ячменя в 0,1 М натрийацетатном буфере, рН 5,0 (или при других значениях рН); 1 мл соответствующим образом разбавленного ферментного раствора инкубируют при 50°С в течение 10 минут. Ферментную реакцию останавливают, добавляя 2 мл DNS раствора (1% (масса/объем)) 3,5-динитросалициловой кислоты, 1,6% (масса/объем) гидроксида натрия, 30% (масса/объем) калий натрий (+)-тартрата в дистиллированной воде). Этот раствор перемешивают и помещают в баню с кипящей водой (минимум 95°С) на 5 минут, затем охлаждают до 25°С. К раствору добавляют 10 мл дистиллированной воды, и поглощение измеряют при 540 нм, используя стеклянную кювету толщиной 2 см.
Полученный результат превращают в мкмоли восстановленного сахара (как глюкозы), сравнивая со стандартной кривой для 2 мл 0,00-0,04% (масса/объем) растворов глюкозы, обработанных аналогичным образом раствором DNS.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой DNS раствор добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности эндо-1,3(4)-β-глюканазы определяют как количество фермента, которое продуцирует 1 мкмоль глюкозного эквивалента в минуту в условиях анализа (50°С и рН 5,0 или при других значениях рН).
1.5. Анализ эндо-1,3(4)-β-глюканазы с использованием β-глюкана азо-ячменя
Анализ активности эндо-1,3(4)-β-глюканазы основан на ферментном гидролизе β-глюкана ячменя, у которого имеется связанный хромофор (β-глюкан азо-ячменя). Продукты реакции, олигомеры, которые растворимы после осаждения этанолом, определяют по наблюдаемому увеличению поглощения при 590 нм.
Раствор, содержащий 0,5 мл субстрата β-глюкана азо-ячменя (в форме, готовой для использования) и 0,2 мл разбавления фермента (содержащего от 0,15 до 0,60 ед/мл в CMC в 0,01 М натрийацетатном буфере, рН 4,6), инкубируют при 30°С точно в течение 20 минут. Ферментную реакцию останавливают, добавляя 2,5 мл осаждающего раствора (содержащего 18,1 г ацетата натрия и 3,0 г цинковой смеси, перемешанных в 300 мл стакане с дистиллированной водой, рН доводят до 5,0 соляной кислотой, переносят содержимое в 1 л мерную колбу и доводят до нужного объема 96% (объем/объем) этанолом). Раствор перемешивают и оставляют при комнатной температуре на 10 минут. Раствор переносят в центрифужную ампулу и центрифугируют при 1000 д в течение 10 минут в лабораторной центрифуге. Поглощение надосадочной жидкости измеряют при 590 нм, используя стеклянную кювету толщиной 1 см.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой осаждающий раствор добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности эндо-1,3(4)-β-глюканазы определяют как количество фермента, которое гидролизует субстрат, обеспечивая величину поглощения 0,820 ед при 590 нм, используя стандартный субстрат в условиях анализа (30°С и рН 4,6).
1.6. Анализ ламинариназы (эндо-1,3-β-глюканазы) с использованием DNS и ламинарина
Анализ активности ламинариназы (эндо-1,3-β-глюканазы) основан на ферментном гидролизе гликозидных связей в ламинарине, β-1,3-глюкане. Продукты реакции, олигосахариды β-1,3-глюкана, определяют по конечному увеличению уровня восстановления (как глюкозы).
Раствор, содержащий 1 мл 1% (масса/объем) раствора ламинарина в 0,1 М натрийацетатном буфере, рН 5,0; 1 мл соответствующим образом разбавленного ферментного раствора инкубируют при 50°С в течение 10 минут. Ферментную реакцию останавливают, добавляя 2 мл DNS раствора (1% (масса/объем)) 3,5-динитро-салициловой кислоты, 1,6% (масса/объем) гидроксида натрия, 30% (масса/объем) калий натрий (+)-тартрата в дистиллированной воде). Этот раствор перемешивают и помещают в баню с кипящей водой (минимум 95°С) на 5 минут, затем охлаждают до 25°С. К раствору добавляют 10 мл дистиллированной воды, и поглощение измеряют при 540 нм, используя стеклянную кювету толщиной 2 см.
Полученный результат превращают в мкмоли восстановленного сахара (как глюкозы), сравнивая со стандартной кривой для 2 мл 0,00-0,04% (масса/объем) растворов глюкозы, обработанных аналогичным образом раствором DNS.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой DNS раствор добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности ламинариназы определяют как количество фермента, которое продуцирует 1 мкмоль глюкозного эквивалента в минуту в условиях анализа (50°С и рН 5,0).
1.7. Анализ эндо-1,4-β-ксиланазы с использованием DNA и ксилана древесины березы
Анализ активности эндо-1,4-β-ксиланазы основан на ферментном гидролизе ксилозидных связей в ксилане древесины березы, β-1,4 ксилане. Продукты реакции, олигосахариды β-1,4-ксилана, определяют по конечному увеличению уровня восстановления (как ксилозы).
Раствор, содержащий 1 мл 1% (масса/объем) раствора ксилана древесины березы в 0,1 М натрийацетатном буфере, рН 5,0 (или при других значениях рН); 1 мл соответствующим образом разбавленного ферментного раствора инкубируют при 50°С в течение 10 минут. Ферментную реакцию останавливают, добавляя 2 мл DNS раствора (1% (масса/объем)) 3,5-динитросалициловой кислоты, 1,6% (масса/объем) гидроксида натрия, 30% (масса/объем) калий натрий (+)-тартрата в дистиллированной воде). Этот раствор перемешивают и помещают в баню с кипящей водой (минимум 95°С) на 5 минут, затем охлаждают до 25°С. К раствору добавляют 10 мл дистиллированной воды, и поглощение измеряют при 540 нм, используя стеклянную кювету толщиной 2 см.
Полученный результат превращают в мкмоли восстановленного сахара (как ксилозы), сравнивая со стандартной кривой для 2 мл 0,00-0,03% (масса/объем) растворов ксилозы, обработанных аналогичным образом раствором DNS.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой DNS раствор добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности эндо-1,4-β-ксиланазы определяют как количество фермента, которое продуцирует 1 мкмоль ксилозного эквивалента в минуту в условиях анализа (50°С и рН 5,0 или при других значениях рН).
1.8. Анализ эндо-1,4-β-ксиланазы с использованием DNS и арабиноксилана пшеницы
Анализ активности эндо-1,4-β-ксиланазы основан на ферментном гидролизе ксилозидных связей в арабиноксилане пшеницы, арабинозой замещенном β-1,4-ксилане. Продукты реакции, олигосахариды арабино-β-1,4-ксилана, определяют по конечному увеличению уровня восстановления (как ксилозы).
Раствор, содержащий 1 мл 1% (масса/объем) раствора арабиноксилана в 0,1 М натрийацетатном буфере, рН 5,0 (или при других значениях рН); 1 мл соответствующим образом разбавленного ферментного раствора инкубируют при 50°С в течение 10 минут. Ферментную реакцию останавливают, добавляя 2 мл DNS раствора (1% (масса/объем)) 3,5-динитросалициловой кислоты, 1,6% (масса/объем) гидроксида натрия, 30% (масса/объем) калий натрий (+)-тартрата в дистиллированной воде). Этот раствор перемешивают и помещают в баню с кипящей водой (минимум 95°С) на 5 минут, затем охлаждают до 25°С. К раствору добавляют 10 мл дистиллированной воды, и поглощение измеряют при 540 нм, используя стеклянную кювету толщиной 2 см.
Полученный результат превращают в мкмоли восстановленного сахара (как ксилоза), сравнивая со стандартной кривой для 2 мл 0,00-0,03% (масса/объем) растворов ксилозы, обработанных аналогичным образом раствором DNS.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой DNS раствор добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности эндо-1,4-β-ксиланазы определяют как количество фермента, которое продуцирует 1 мкмоль ксилозного эквивалента в минуту в условиях анализа (50°С и рН 5,0 или при других значениях рН).
1.9. Анализ эндо-1,4-β-ксиланазы вискозиметрическим способом с использованием арабиноксилана пшеницы
Анализ активности эндо-1,4-β-ксиланазы основан на ферментном гидролизе стандартного раствора арабиноксилана пшеницы, причем активность определяют по уменьшению относительной вязкости в зависимости от времени.
Раствор, содержащий 1 мл 1% (масса/объем) раствора арабиноксилана пшеницы в 0,1 М натрийацетатном буфере, рН 5,5 (или при других значениях рН); 3 мл дистиллированной воды и 1 мл соответствующим образом разбавленного ферментного раствора вводят в микровискозиметр Haake (используя золотой шарик, калиброванный до 0,1-2,0 мПа·сек), и измеряют время падения шарика (Ттест) (в мсек относительно определенной высоты падения) каждые 30 сек на протяжении 15-20 минут при 30°С. Средние времена падения шарика измеряют для воды (5 мл дистиллированной воды) и субстратного (1 мл 1% (масса/объем) раствора арабиноксилана пшеницы в 0,1 М натрийацетатном буфере, рН 5,5 и 4 мл дистиллированной воды) как Твода и Тсубстрат, соответственно. Проводят контрольные измерения аналогичным образом. Относительную текучесть (Fr) рассчитывают для каждого значения Ттест следующим образом:
Наклон кривой Fr в зависимости от времени (прошедшее время, при котором проведено каждое измерение Ттест), рассчитывают как изменение относительной вязкости в минуту (ΔFr/мин), и оно пропорционально концентрации фермента.
Одну единицу активности эндо-1,4-β-ксиланазы определяют как количество фермента, которое гидролизует субстрат, уменьшая вязкость раствора, обеспечивая изменение относительной текучести в 1 (безразмерная единица) в минуту в условиях анализа (30°С и рН 5,5 или при других значениях рН).
1.10. Анализ β-ксилозидазы с использованием р-нитрофенил-β-ксилопиранозида
Анализ β-ксилозидазы основан на ферментном гидролизе р-нитрофенил β-D-ксилопиранозида. Продукт реакции, р-нитрофенол, определяют колориметрически.
Раствор, содержащий 1 мл 0,1% (масса/объем) р-нитрофенил-β-D-ксилопиранозида в дистиллированной воде; 1 мл дистиллированной воды; 1 мл 0,2 М натрийацетатного буфера, рН 5,0; 1 мл соответствующим образом разбавленного ферментного раствора инкубируют при 50°С в течение 30 минут. Ферментную реакцию останавливают, добавляя 4 мл 0,4 М раствора глицина. Этот раствор перемешивают и охлаждают до 20°С. Поглощение измеряют при 400 нм, используя стеклянную кювету толщиной 1 см.
Полученный результат превращают в мкмоли р-нитрофенола, сравнивая с коэффициентом молярной экстинкции р-нитрофенола в этих же условиях.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой раствор глицина добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности ксилозидазы определяют как количество фермента, которое продуцирует 1 мкмоль р-нитрофенола из р-нитрофенил-β-D-ксилопиранозида в минуту в условиях анализа (50°С и рН 5,0).
1.11. Анализ α-N-арабинофуранозидазы с использованием р-нитрофенил α-L-арабинофуранозида
Анализ α-N-арабинофуранозида (арабинофуранозида) основан на ферментном гидролизе р-нитрофенил α-L-арабинофуранозида. Продукт реакции, р-нитрофенол, определяют колориметрически.
Раствор, содержащий 1 мл 0,1% (масса/объем) р-нитрофенил α-L-арабинофуранозида в дистиллированной воде; 1 мл дистиллированной воды; 1 мл 0,2 М натрийацетатного буфера, рН 5,0; 1 мл соответствующим образом разбавленного ферментного раствора инкубируют при 50°С в течение 30 минут. Ферментную реакцию останавливают, добавляя 4 мл 0,4 М раствора глицина. Этот раствор перемешивают и охлаждают до 20°С. Поглощение измеряют на 400 нм, используя стеклянную кювету толщиной 1 см.
Полученный результат превращают в мкмоли р-нитрофенола, сравнивая с коэффициентом молярной экстинкции р-нитрофенола в этих же условиях.
В наблюдаемое поглощение для растворов ферментных реакций вносят поправку на неспецифическое поглощение, осуществляя реакцию, в которой раствор глицина добавляют к смеси перед добавлением ферментного раствора. Одну единицу активности арабинофуранозидазы определяют как количество фермента, которое продуцирует 1 мкмоль р-нитрофенола из р-нитрофенил α-L-арабинофуранозида в минуту в условиях анализа (50°С и рН 5,0).
1.12. Аналиа ферулоил-эстеразы FAXX методом
Анализ ферулоил-эстеразы (эстеразы феруловой кислоты) основан на ферментном гидролизе O-[5-O-(транс-ферулоил)-α-L-арабинофуранозил]-(1→3)-O-β-D-ксилопиранозил-(1→4)-D-ксилопиранозы (FAXX). FAXX получают из ферментативно гидролизованных пшеничных отрубей, очищают и характеризуют с помощью ЯМР. FAXX гидролиз определяют спектрофотометрически.
Ферментативную реакцию соблюдают при длине волны 325 нм, используя кювету толщиной 1 см, в растворе, содержащем 0,050 мМ FAXX в 0,1 М MOPS буфере, рН 6,0 при 37°С.
Одну единицу активности ферулоил-эстеразы на FAXX определяют как количество фермента, которое превращает 1 мкмоль субстрата в продукт в минуту в условиях анализа (37°С и рН 6,0).
1.13. Анализ ферулоил-эстеразы с помощью Ara2F
Анализ ферулоил-эстеразы (эстеразы феруловой кислоты) основан на ферментативном гидролизе Ara2F (феруловая кислота, связанная по 1,2 с арабинозой). Ara2F получают из ферментативно гидролизованной пульпы сахарной свеклы, очищают и характеризуют с помощью ЯМР. Ara2F гидролиз определяют спектрофотометрически.
Ферментативную реакцию наблюдают при длине волны 325 нм, используя кювету толщиной 1 см, в растворе, содержащем 0,050 мМ Ara2F в 0,1 М MOPS буфере, рН 6,0 при 37°С.
Одну единицу активности ферулоил-эстеразы на Ara2F определяют как количество фермента, которое превращает 1 мкмоль субстрата в продукт в минуту в условиях анализа (37°С и рН 6,0).
1.14. Анализ ферулоил-эстеразы с использованием гидролиза метиловых сложных эфиров: метилферуловой кислоты (MFA); метилкофеиновой кислоты (МСА); метилсинаповой кислоты (MSA); метил-р-кумаровой кислоты (MpCA)
Анализ ферулоил-эстеразы (эстеразы феруловой кислоты) основан на ферментативном гидролизе метиловых сложных эфиров феруловой кислоты (MFA); кофеиновой кислоты (МСА); синаповой кислоты (MSA); и р-кумаровой кислоты (МрСА). Гидролиз метиловых сложных эфиров определяют в 0,1 М MOPS буфере, рН 6,0 при 37°С. Анализ основан на двух различных методиках.
В спектрофотометрическом способе концентрация субстрата метиловых сложных эфиров составляет 0,10 мМ, и за гидролизом сложных эфиров наблюдают при длине волны 325 нм, используя кювету толщиной 1 см. В этом способе начальная концентрация субстрата ограничена.
В способе с использованием ВЭЖХ концентрация субстрата метиловых сложных эфиров составляет 1,0 мМ, и за гидролизом сложных эфиров следят, измеряя выделение свободной кислоты с помощью ВЭЖХ с 10-30 минутными интервалами. В этом способе нет ограничения по концентрации субстрата, и измеряемые активности значительно выше, нежели концентрации, определяемые спектрофотометрически.
Одну единицу активности ферулоил-эстеразы определяют как количество фермента, которое превращает 1 мкмоль субстрата в продукт в минуту в условиях анализа (37°С и рН 6,0).
1.15. Определение концентрации протеина с помощью модифицированного анализа Bradford связывания протеина с Coomassie blue
Анализ концентрации протеина основан на модифицированном анализе Bradford связывания протеина с Coomassie blue с использованием Brilliant Blue G (кумасси брильянтовый голубой), измеряемом спектрофотометрически при длине волны 595 нм, используя кювету толщиной 1 см. Этот способ (Sigma В 6916) стандартизован с использованием альбумина бычьей сыворотки (Sigma P094).
1.16. Определение изоэлектрической точки с помощью изоэлектрического фокусирования
Изоэлектрические точки протеинов определяют стандартными способами, используя предварительно отлитые вертикальные 5% полиакриламидные гели, такие, как гели от NOVEX® для интервала рН 3-10 (р1 интервал 3,5-8,5) или рН 3-7 (р1 интервал 3,0-6,5) в кювете NOVEX® XCell II™ Mini-Cell. Используют NOVEX® катод, анод и IFE образцовые буферы для рН 3-10 или рН 3-7. Используют NOVEX® стандартный протокол для изоэлектрического фокусирования, фиксирования, окрашивания красителем Coomassie R-250 Blue и обессоливания.
1.17. SDS-PAGE (гель-электрофорез в натрийдодецилсульфат полиакриламидном геле)
Аналитическое разделение и определение молекулярной массы протеинов осуществляют стандартными SDS-PAGE методами. Предварительно отлитые NOVEX® NuPAGE гели (NuPAGE™ Bis-Tris гели или NuPAGE™ Tris-ацетатные гели с NOVEX® рекомендованными рабочими буферами используют в NOVEX® XCell II™ Mini-Cell. Используют NOVEX® образцовые препаративные и рабочие буферы и стандарты молекулярных масс. Используют NOVEX® стандартные протоколы для SDS-PAGE, фиксирования, окрашивания красителем Coomassie R-250 Blue и обессоливания.
2. Результаты анализа смеси ферментов
2.1. Оптимальные значения pH
2.1.1. Активность эндо-1,3(4)-β-глюканазы
Анализ эндо-1,3(4)-β-глюканазы из Penicillum funiculosum осуществляют в стандартных условиях при 50°С, с помощью способа с использованием DNS и β-глюкана ячменя. Ферментативную активность измеряют при значениях рН между 3,0 и 7,0. Оптимальные значения рН для активности фермента составляют 4,0-5,0.
2.1.2. Активность эндо-1,4-β-ксиланазы
Анализ эндо-1,4-β-ксиланазы из Penicillum funiculosum осуществляют в стандартных условиях при 50°С, с помощью способа с использованием DNS и ксилана древесины березы.
2.2. Оптимальная температура
2.2.2. Активность эндо-1,3(4)-β-глюканазы
Анализ эндо-1,3(4)-β-глюканазы из Penicillum funiculosum осуществляют в стандартных условиях при рН 5,0 (оптимальное значение рН для этого фермента) с помощью способа с использованием DNS и β-глюкана ячменя. Ферментативную активность измеряют при значениях температуры между 30 и 70°С. Значения оптимальных температур находятся между 50 и 60°С, причем наивысшая активность наблюдается при 60°С. Подробные результаты в форме таблицы в зависимости от температур приведены в таблице
2.2.2. Активность эндо-1,4-β-ксиланазы
Анализ эндо-1,4-β-ксиланазы из Penicillum funiculosum осуществляют в стандартных условиях при рН 5,5 и 3,5, с помощью способа с использованием DNS и ксилана древесины березы.
Ферментативную активность измеряют при значениях температуры между 30 и 70°С. Значения оптимальных температур находятся между 50 и 60°С, причем наивысшая активность наблюдается при 50°С для рН 5,5 и при 60°С для рН 3,5. Подробные результаты в форме таблицы в зависимости от температур приведены в таблице
Ферменты, продуцируемые Penicillum funiculosum, отличаются высокими уровнями целлюлазы, эндо-1,3(4)-β-глюканазы и других глюканолитических активностей. Кроме того, они также характеризуются высокими уровнями эндо-1,4-β-ксиланазы и активностями сопутствующих ферментов ксиланазы. Широкий спектр гемицеллюлолитических ферментов является характеристикой ферментных препаратов, получаемых из этого микроорганизма.
Каждую из измеряемых активностей можно представить как отношение к основной активности для этого препарата. Примеры полученных результатов представлены в таблице А. Эти отношения могут меняться в препаратах, полученных из различных партий ферментации.
3. Свойства компонентов в смеси ферментов
3.1 Способы очистки
Хроматография гидрофобных взаимодействий
Препараты, полученные после фильтрации и концентрирования ферментационной среды до концентрации протеина 112,6 мг/мл, разбавляют 1/1 буфером для хроматографии гидрофобных взаимодействий (HIC) (50 мМ фосфатный буфер, рН 7,0/1,7 М (NH4)2SO4/0,04% азид натрия), заменяют на HIC буфер (PD-10 колонки; Pharmacia). Порции (по 10 мл) высокоэффективного HIC геля (Pharmacia) вводят в колонку (диаметр 10×5 см, 200 мл) PhenylSepharoes™, и разделяют, используя градиент сульфата аммония (NH4)2SO4) с понижающейся концентрацией (1,7-0,0 М) за 10 минут. Фракции (по 10 мл) собирают и анализируют на активность ксиланазы.
HIC дает два основных пика активности ксиланазы. Первый, названный А, элюируется из колонки, когда концентрация (NH4)2SO4 снижается до около 0,6 М, тогда как второй, названный В, элюируется при концентрации (NH4)2SO4 около 0,25 М. Фракции, содержащие пики А и В, из каждого ввода собирают отдельно. Полная фракция А соответствует 2,8% от полной активности ксиланазы, тогда как фракция В соответствует 97,2% от полной активности ксиланазы. Выход составляет 77%.
Ионообменная хроматография
Объединенные фракции пиков А и В из HIC осаждают, повышая концентрацию (NH4)2SO4 до 100% насыщения с последующим центрифугированием (10000×g в течение 30 минут). Осадки снова растворяют в смеси 20 мл Tris-HCl буфера, рН 8,0/0,04% азид натрия и обессоливают до того же самого буфера, используя PD-10 колонку. Образцы (5 мл) вводят в MonoQ™ HR 10/10 анионообменную колонку (Pharmacia), предварительно уравновешенную смесью 20 мМ Tris-HCl буфера рН 8,0/0,04% азид натрия, и элюируют со скоростью 2 мл/мин с повышающейся концентрацией хлорида натрия (NaCl(0-1 М) в том же самом буфере. Фракции (2 мл) собирают и анализируют на активность ксиланазы.
Пик А:
Выделение пика А с помощью анионообменной хроматографии приводит к получению одного пика активности ксиланазы, который элюируется при около 0,3 М NaCl. Наиболее активные фракции собирают и анализируют с помощью SDS-PAGE (гель-электрофорез в натрийдодецилсульфат полиакриламиде). Получают одну основную полосу с молекулярной массой 48 kDa. Выделение активности ксиланазы после IEF (изоэлектрическое фокусирование) подтверждает, что эта основная Coomassie-окрашенная полоса является ксиланазой.
Пик В:
Выделение пика В с помощью анионообменной хроматографии дает два основных пика активности ксиланазы, один из которых элюируется в пике несорбирующегося материала (несвязанный материал; пик В-I), а другой - при 0,1 М NaCl (пик В-II). Присутствуют также два небольших пика, которые элюируются при 0,13 и 0,19 М NaCl. Активные фракции, соответствующие каждому пику, собирают и анализируют с помощью SDS-PAGE, но ни один из образцов не является чистым.
Гель-фильтрационная хроматография
Объединенные фракции, содержащие В-I и В-II, сушат вымораживанием, снова растворяют в воде и обессоливают (используя PD-10 колонки). Образцы (0,2 мл) вводят в Superdex™ 75 HR колонку (Pharmacia) и элюируют при скорости 0,4 мл/мин буфером 20 мМ Bis-Tris, рН 6,0/0,2 М NaCl/0,04% азид натрия. Фракции (0,4 мл) собирают и анализируют на ксиланазную активность.
3.2. Характеристики ксиланаз
3.2.1. Определение изоэлектрической точки с помощью изоэлектрического фокусирования
Изоэлектрические точки протеинов определяют стандартными способами, используя предварительно отлитые вертикальные 5% полиакриламидные гели от NOVEX® для рН 3-10 и рН 3-7. Используют NOVEX® катод, анод и IEF образцовые буферы, и стандартные протоколы для изоэлектрического фокусирования, фиксации, окрашивания Coomassie R-250 Blue красителем, и обесцвечивания.
Для ксиланазы А используют образец после MonoQ. Для ксиланаз В-I и В-II используют образец после HlC, ксиланазу В. Для каждого из А и В небольшой образец (10 мкл) помещают в отдельную ячейку, и большой образец (50 мкл) помещают в тройную ячейку. После фокусирования образцов гель разрезают на две части так, что одна часть содержит два небольших образца (А+В) и маркеры молекулярного веса (эта половина окрашена Coomassie), тогда как другая часть содержит два крупных образца. Часть геля, которая содержит большие образцы, разрезают так, чтобы разделить полосы двух образцов, а затем каждую из полос разделяют на участки по 2 мм. Каждый из 2 мм участков вымачивают отдельно в течение ночи в 100 мМ MOPS буфере, рН 6,0/0,04% азид натрия. Фракции анализируют на активность ксиланазы.
Для образца А ксиланазы окрашенный IEF гель демонстрирует одну основную полосу pI 3,55 маркера и несколько небольших примесных полос. Ксиланазная активность обнаружена только во фракции, соответствующей этой полосе, подтверждая, что основной пик соответствует ксиланазе.
Для образца В окрашенный IEF гель демонстрирует несколько полос в интервале значений pI. Ксиланазная активность имела место в двух разделенных фракциях неокрашенного геля и соответствует протеинам с pI 4,2 и 4,8.
3.2.2 Определение молекулярной массы с помощью SDS-PAGE
Для подтверждения молекулярных масс ксиланаз в пике В из HIC фракции с ксиланазной активностью, элюированной из IEF геля, обессоливают, сушат вымораживанием и разделяют с помощью SDS-PAGE. Денатурирование PAGE осуществляют, используя 10% Tris-глициновый гель (NOVEX®) с дитиотреитолом (DTT 50 мМ), включенным в образцовый буфер в качестве восстанавливающего агента.
Окрашенный гель показывает, что обе ксиланазы оказались чистыми, с молекулярными массами 36 kDa и 15 kDa для ксиланазы В-I и В-II, соответственно.
Все три очищенные ксиланазы анализируют с помощью SDS-PAGE: фракцию ксиланазы А после анионообменной хроматографической обработки, фракции ксиланазы В-I и В-II после гель-фильтрационной хроматографической обработки. Ксиланаза А дает одну полосу с молекулярной массой 48 kDa. Ксиланаза В-I дает одну основную и четыре небольшие полосы после окрашивания Coomassie. Подтверждается, что основная полоса соответствует ксиланазе, так как ее молекулярная масса равна 36 kDa. Степень чистоты оценивают как 90%. Ксиланаза В-II дает основную полосу с молекулярной массой 15 kDa и 2-3 небольшие полосы. Степень чистоты этой ксиланазы примерно 95%.
3.2.3. Активность ферментов
Тесты по измерению активности ферментов были описаны выше.
3.2.3.1. Анализ ксиланазы А
[Протеин] 0,4 (мг/мл)
Но - не определяли
Н/п - неприменимо
Активность ксиланазы в отношении ксилана древесины березы в зависимости от рН
3.2.3.2. Анализ ксиланазы В-I
[Протеин] 0,096 (мг/мл)
Но - не определяли
Н/п - неприменимо
Активность ксиланазы в отношении ксилана древесины березы в зависимости от рН
3.2.3.3. Анализ ксиланазы В-II
[Протеин] 0,165 (мг/мл)
Но - не определяли
Н/п - неприменимо
Активность ксиланазы в отношении ксилана древесины березы в зависимости от рН
3.2.4 Последовательности
Один из вариантов настоящего изобретения относится к последовательностям аминокислот и нуклеиновых кислот для описанных выше протеинов или их вариантов.
Для этой цели последовательности ксиланаз идентифицируют по аминокислотным последовательностям очищенных протеинов (ксиланаза А, ксиланаза В-I и ксиланаза В-II) и при сравнении последовательностей аминокислот и нуклеотидов с последовательностями известных грибковых ксиланаз.
Следует понимать, что в рамках настоящего изобретения термин варианты относится к любому полипептиду или любому аналогу протеина, фрагменту протеина, производному или протеину-мутанту из нативного протеина или полипептида и обладающему теми же самыми биологическими функциями, что и указанный нативный протеин или полипептид. В природном состоянии могут существовать различные варианты. Такими вариантами могут быть, например, аллельные варианты, характеризующиеся отличиями в последовательности генов, кодирующих указанный протеин, или могут быть результатом дифференциального сплайсинга или пост-трансдукционных модификаций. Варианты можно получить путем замещения, делеции, добавления и/или модификации одной или более из аминокислот. Все модификации хорошо известны и их можно осуществить любыми известными специалистам способами.
Вариантами являются молекулы, обладающие, например, большим сродством к их субстрату, или обладающие новыми биологическими свойствами.
Другой целью настоящего изобретения является также применение последовательностей для экспрессии описанных протеинов или полипептидов в клетках-хозяевах одноклеточных или многоклеточных организмов. Для этой цели указанные последовательности могут быть введены в геном вектора. Указанным вектором может быть плазмида, фаг или вирус. Поэтому другим вариантом настоящего изобретения является клетка-хозяин, выделенная из одноклеточного или многоклеточного организма, трансфецированная или инфицированная вектором, как описано выше. В предпочтительном варианте клеткой-хозяином является бактерия.
Другим вариантом настоящего изобретения является применение указанных векторов, включающих последовательность нуклеиновых кислот описанных протеинов для экспрессии указанного протеина в любую клетку-хозяина.
3.2.4.1 Последовательности ксиланазы С
Создание зондов основано на сравнениях аминокислотных и нуклеотидных последовательностей известных грибковых ксиланаз. Определяют консервативные участки и используют для создания ПЦР праймеров, продукты которых можно было бы использовать для скринирования геномной библиотеки Penicillum funiculosum.
Создают две пары дегенеративных праймеров. Первую пару создают для амплификации 200 bp (примерно) продукта из гена ксилазы типа В (или типа 2). Вторую пару создают для амплификации 250 bp продукта из гена ксиланазы типа А (тип 1).
Полосу 258 bp получают с праймерами 3 и 4. После клонирования в pGEMT и секвенирования было обнаружено, что имеется значительное сходство последовательности с последовательностью грибковой ксиланазы типа А/1. Плазмиду, содержащую клонированный продукт, называют pPFXYLA.
Полная последовательность ксиланазы С представлена на фиг. 1 и в последовательности ID №1 (ПОСЛЕД. ID №1).
3.2.4.2 Последовательности ксиланазы BI
Для конструирования дегенеративных ПЦР праймеров ((ПОСЛЕД. ID №2 и №3) используют внутреннюю аминокислотную последовательность наряду со сравнениями последовательностей других грибковых целлобиогидролаз. Продукт 290 bp (ПОСЛЕД. ID №4) амплифицируют и клонируют в pGEMT (Promega) для создания pGEMTCB2 и секвенируют. На фиг. 2 видно, что праймерные последовательности подчеркнуты. Этот продукт ПЦР в настоящее время применяют в качестве зонда для скринирования геномной библиотеки Penicillum funiculosum IMI134756.
3.2.4.3. Последовательности ксиланазы BII
Вся последовательность гена ксиланазы BII включает 1,3 kb 5'нетранслируемого и расположенного в обратном направлении участка и 0,85 kb 3'нетранслируемого участка, 54 bp интрон и 669 bp, кодирующих протеин из 223 аминокислот.
Для доказательства существования 54 bp интрона используют ПЦР с обратной транскриптазой (RT-PCR). Полную РНК выделяют из мицелия культур Penicillum funiculosum IMI134756, собранных после 4 дней выращивания на 1% (масса/объем) ксилане овса. Конструируют праймеры для амплификации вплоть до 195 bp фрагмента из информационной РНК (249 bp из геномной ДНК) и до 433 bp (487 bp с геномной ДНК).
Секвенирование 3 kb с 3'конца плазмиды (pPFXYNC2) выявило ген (обозначенный per А), который содержит два предполагаемых интрона, и кодирует полипептид, состоящий примерно из 570 аминокислот. Этот полипептид демонстрирует значительное сходство последовательности с аминокислотными пермеазами грибков.
3.2.4.4. Последовательность ксиланазы А
Была получена внутренняя последовательность ксиланазы А, и она представлена следующей последовательностью аминокислот: AEAINYNQDY
3.3. Свойства ферулоил-эстераз
3.3.1. Очистка
Осуществляют таким же способом, что и для ксиланаз.
Смесь ферментов содержит, по крайней мере, две различные ферулоил-эстеразы. У одной из них (FaeB) молекулярная масса по данным масс-спектрометрии составляет 38,945-41,051 Da (35,450 Da из первичной аминокислотной последовательности и 37 kDa по данным SDS-PAGE). FaeB имеет значение pI 4,2, что соответствует ферулоил-эстеразе типа В и является специфическим для МрСА и Ara2F субстратов (активность в отношении МрСА, МСА, MFA и Ara2F; но не в отношении MSA и FAXX).
У другой ферулоил-эстеразы (FaeA) молекулярная масса составляет 29 kDa (по данным SDS-PAGE). FaeA имеет значение pI 4,65, что соответствует ферулоил-эстеразе типа А, и является специфическим для FAXX и MSA субстратов (активности в отношении MSA, МСА, MFA и FAXX, но не в отношении МрСА Ara2F).
3.3.2. Определение изоэлектрической точки с помощью изоэлектрического фокусирования.
Изоэлектрические точки протеинов определяют стандартными способами. Смесь ферментов наносят в виде широкой полоски (около 20 мм) в IEF гель и осуществляют электрофорез при пониженной температуре (5°С). После фокусировки и сужения полосы гель разрезают по середине полосы образца. Одну часть полосы образца и IEF стандарты фиксируют, окрашивают и обессоливают, используя стандартный протокол. Другую часть полосы нарезают на участки шириной 2 мм, и каждый участок вымачивают в течение ночи в 1 мл 100 мМ MOPSбуфера, рН 6,0. Активность ферулоил-эстеразы определяют для каждого участка геля, используя MFA, МрСА и MSA в качестве субстратов.
Окрашенный IEF гель демонстрирует наличие очень многих протеинов в целлюлазе, pI значения для которых варьируются в интервале от очень кислотных (pI 2,4) до около pI 7. Большинство протеинов являются кислотными (интервал pI 2,4-5). Два пика ферулоил-эстеразной активности обнаруживают во фракциях, вырезанных из геля. Один, соответствующий FaeB, имеет значение pI 4,2 и активность только в отношении MFA и МрСА (но не MSA). Другой, соответствующий FaeA, имеет значение pI 4,65 и активность в отношении всех трех тестированных субстратов.
3.3.3 Молекулярная масса, определенная с помощью SDS-PAGE
Молекулярные массы определяют с помощью SDS-PAGE, используя 10% Tris-глициновые гели. Получают SDS-PAGE гели, фиксируют, окрашивают Coomassie Blue красителем и обессоливают, используя стандартный протокол.
Смесь ферментов содержит, по крайней мере, две различные ферулоил-эстеразы. У одной из них, соответствующей FaeB (pI 4,2), молекулярная масса составляет 37 kDa. У другой, соответствующей FaeA (pI 4,65), молекулярная масса составляет 29 kDa.
Молекулярную массу FaeB оценивают как 34,450 Da из первичной аминокислотной последовательности, и как 38,945-41,051 Da по данным масс-спектрометрии.
3.3.4. Активность ферулоил-эстеразы
Анализ на активность ферулоил-эстераз осуществляют на смеси ферментов, используя спектрофотометрический метод.
Смесь ферментов содержит активности против всех тестированных субстратов. Для метилэстераз наивысшей активностью является активность в отношении МрСА, а самой низкой - в отношении MSA.
Активности в отношении Ara2F и FAXX выше, нежели в отношении метилэстераз, что является показателем того, что активности эстераз связаны с истинными ферулоил-эстеразами, а не с общими эстеразами или побочными активностями других разрушающих стенки клеток эстераз (например, ацетил-ксилан-эстераза, пектин-эстераза).
3.3.5. Последовательности
3.3.5.1 Последовательность FEA-А
В соответствии с расщеплением очищенных протеинов получают внутренние аминокислотные последовательности как:
Последовательность 1
QYTLTLPSNYNPNK
Последовательность 2
AVAVMSGANL
Последовательность 3
TEYSG (С/А) DSEHPVWWIAFDGP
Последовательность 4
DTFVKDDHCTPTNPPAPAAGSGTHIKYV
Конструируют несколько дегенеративных ПЦР праймеров из аминокислотных последовательностей, полученных из очищенного протеина. Многие продукты были клонированы в pGEMT (Promega) и секвенированы.
С помощью ПЦР было обнаружено, что плазмида, названная pGEMTD19 (180 bp) (фиг.3), содержит последовательность, которая распознается как пептидная последовательность 4, представленная выше. Как показано на фиг.3, праймерные последовательности подчеркнуты дважды, а ранее известная последовательность подчеркнута один раз.
Последовательности нуклеиновой кислоты и аминокислот FAE-A представлены в ПОСЛЕД. ID №5.
3.3.5.2. Последовательность FEA-B
Праймеры, сконструированные из пептидной последовательности FAE-B, используют для усиления зонда, который затем используют для скринирования геномной библиотеки Penicillum funiculosum. Клон 2291 bp выделяют и секвенируют (ПОСЛЕД. ID №6). Ген, кодирующий полипептид из 304 аминокислот, имеет один предполагаемый интрон. Предсказанная аминокислотная последовательность представлена на фиг.4, где зрелый протеин (длина зрелого протеина =338) изображен жирным шрифтом. Этот протеин включает два различных домена, разделенные в сильной степени гликозилированным линкером. Как представлено на фиг.4, каталитический домен изображен жирным шрифтом, тогда как связующий домен изображен жирным шрифтом и подчеркнут дважды, а линкер представлен точечной жирной линией.
Протеин также отличается предполагаемым фрагментом активного сайта (серин=нуклеофил), что представлено подчеркиванием на фиг.4, со следующими предположительными каталитическими триадами:
(1) S136/D174/H216
(2) S136/D220/H276.
FAE-B протеин включает также последовательность секреции (353) и 10 цистеинов.
3.4 Свойства глюканаз
Смесь ферментов обрабатывают с помощью 2D гель-электрофореза. IEF осуществляют, используя предварительно отлитые вертикальные 5% полиакриламидные гели от NOVEX® для рН 3-7 (интервал pI 3,0-6,5) в мини-ячейке NOVEX® XCell II™ Mini-Cell. Используют NOVEX® катод, анод и IEF образцовые буферы для рН 3-7, а также NOVEX® стандартный протокол для изоэлектрического фокусирования. Одну полосу отрезают, и обрабатывают электрофоретически во втором направлении, используя 10% Laemmli SDS-PAGE гель. Из геля выделяют вторую полосу, разрезают на 35 фракций, полоски геля вымачивают в буфере, и фракции анализируют на активность фермента. Третью полосу оставляют на геле, фиксируют, окрашивают Coomassie R-250 Blue красителем и обессоливают, используя NOVEX® стандартный протокол.
Значительные активности эндо-1,3(4)-β-глюканазы (способ DNS и β-глюкана ячменя) и целлюлазы (способ DNS и CMC) обнаружены во фракциях, соответствующих протеинам с pI 4,2, М.В. 36 kDa и pI 5,4, М.В. 27 kDa. Для удаления ксиланазы В-1, находящейся в одной из фракций, фракции тестируют на активность, используя способ DNS и ксилана древесины березы. Во фракциях, соответствующих активностям β-глюканазы или целлюлазы, активность ксиланазы не обнаружена.
Список чертежей
Фиг.1: Аминокислотная последовательность протеина ксиланазы С Penicillium funiculosum.
Фиг.2: Нуклеотидная и аминокислотная последовательности продукта ПЦР ксиланазы B1 (XynBI).
Фиг.3: Нуклеотидная и аминокислотная последовательности ПЦР продукта ферулоил-эстеразы A (faeA).
Фиг.4: Аминокислотная последовательность протеина (FAE-V или FAE-I) ферулоил-эстеразы В (faeB) Penicillium funiculosum
Е. Применение смеси ферментов в кормах для животных
Пример 1: Оценка эффективности препарата ферментов, продуцируемых Penicillium funiculosum в отношении энергетической ценности (AMEN) корма из смеси пшеницы и ячменя для бройлеров.
Целью было продемонстрировать эффективность ферментов (активность β-глюканазы: 100 ед/кг и активность ксиланазы: 1100 ед/кг) на кажущуюся обменную энергию, скорректированную по азотному балансу (AMEN) корма, содержащего 50% пшеницы и 22% ячменя.
Эксперименты проводили с помощью контрольных и ферментных препаратов (активность β-глюканазы: 100 ед/кг и активность ксиланазы: 1100 ед/кг), используя Европейский метод сравнения (Bourdillon et al., 1990) при кормлении ad libitum и полном сборе экскрета для возраста между 18 и 21 днем.
При необходимости, полученные экспериментальные ферментные препараты дополнялись отдельными выделенными ферментами для получения указанной ферментативной активности испытуемых ферментных препаратов. В случае, когда активность партии ферментного препарата по β-глюканазе составила 70 ед/кг, то препарат для включения в эксперимент был дополнен выделенным из Penicillium funiculosum и очищенным ферментом до требуемой активности.
а. Материал и методы
Птицы: Порода и условия выращивания
Однодневных петушков Ross бройлеров содержат в клетках коллективной батареи до возраста 12 дней. Им дают стандартный начальный корм. На 12 день птиц взвешивают и равномерно распределяют на 10 отдельных клеток для каждой обработки, а затем им дают экспериментальный корм в течение периода адаптации (минимум 5 дней).
Поддерживают стандартную температуру и влажность. Постоянный световой режим составляет: 23 часа света и 1 час темноты до конца испытаний.
Корма: Птицы получают начальный корм с одного дня жизни до 12 дней, а затем получают экспериментальный корм.
Экспериментальные корма
Корма содержат 50% пшеницы и 22% ячменя (таблица 1.1). Ферментный препарат распыляют на 20 кг зерна.
Восстановление ферментов в кормах измеряют вискозиметрически (Sabatier and Fish, 1996).
Измерение обменной энергии
Баланс начинают определять с 18 дня (Д18) следующим образом:
Д 17, птицам не дают корма в течение ночи;
Д 18, птиц взвешивают, очищают сборочные лотки;
Д 19, фекалии собирают и замораживают;
Д 20, фекалии собирают и замораживают, ночью не дают корма;
Д 21, фекалии собирают и замораживают, птиц взвешивают и снова кормят.
Фекалии сушат вымораживанием и измельчают как корм (1 мм, измельчитель Retsch). Общую энергию корма и экскрета определяют с помощью адиабатического колориметра IKA C5000. Также определяют содержание протеинов (N·6,25, Kjeldahl метод Z130) и липидов (метод Z160). Корректировку по азотному балансу вводят, используя 18% протеин в увеличении массы.
b. Результаты и обсуждение
Очевидная обменная энергия, скорректированная по азотному балансу (AMEN)
Зоотехнические характеристики и величины обменной энергии представлены в таблице 1.2. Между обработками нет различий в зоотехнических характеристиках.
При выращивании бройлеров ферментный препарат повышает (AMEN) корма на основе 50% пшеницы и 22% ячменя на 6,2% (+204 ккал/кг ДМ (Сухого Вещества)).
Более того, вариабельность энергетической усвояемости была снижена с 80 до 62 ккал/кг ДМ.
Столь значительное увеличение демонстрирует пользу обоих активностей (ксиланазы и β-глюканазы), продуцируемых Penicillum funiculosum в отношении гидролиза растворимых не крахмалистых полисахаридов пшеницы и ячменя.
Пример 2: Влияние препарата ферментов, продуцируемых Penicillum funiculosum, на усвояемость корма бройлерами, получающими в качестве корма пшеницу
Проводят испытания с целью определить влияние препарата ферментов, продуцируемых Penicillum funiculosum (активность β-глюканазы: 100 ед/кг и активность ксиланазы: 1100 ед/кг) на очевидную обменную энергию (АМЕ), на усвояемость протеинов и липидов бройлерами, получающими корм, содержащий 54% пшеницы. Исследуют также зависимость от степени измельчения корма
(1) Контроль 1 (54% измельченной пшеницы)
(2) Контроль 1 + Препарат ферментов (активность β- глюканазы 100 ед/кг и активность ксиланазы 1100 ед/кг)
(3) Контроль 2 (30% целых зерен пшеницы, 24% измельченных зерен пшеницы)
(4) Контроль 2 + Препарат ферментов (активность β-глюканазы 100 ед/кг и активность ксиланазы 1100 ед/кг)
В соответствии с Европейским методом сравнения (корм ad libitum и полный сбор экскрета для возраста с 18 до 21 дня) (Bourdillion et al., 1990).
а. Материалы и методы
Птицы: Порода и условия выращивания
Однодневных петушков Ross бройлеров содержат в клетках коллективной батареи до возраста 12 дней. Затем их переводят в индивидуальные клетки для определения баланса усвояемости.
Поддерживают стандартную температуру и влажность. Постоянный световой режим составляет: 23 часа света и 1 час темноты до 8-дневного возраста. Затем световой режим меняют на 15 час 30 мин света и 8 час 30 мин темноты из-за испытаний, проводящихся в том же помещении.
Корма: Птицы получают стандартный начальный корм до 12-дневного возраста, а затем получают экспериментальные корма.
Экспериментальные корма
Корма содержат 54% пшеницы, характеристики представлены в таблице 2.1. Состав корма представлен в таблице 2.2.
Измерение очевидной обменной энергии
Баланс начинают определять с 17 дня по следующей схеме:
Д 17, птицам не дают корма в течение ночи;
Д 18, птиц взвешивают, очищают сборочные лотки;
Д 19, фекалии собирают и замораживают;
Д 20, фекалии собирают и замораживают, ночью не дают корма;
Д 21, фекалии собирают и замораживают, птиц взвешивают и снова кормят.
Фекалии сушат вымораживанием и измельчают как корм (1 мм, измельчитель Retsch). Общую энергию корма и экскрета определяют с помощью адиабатического колориметра IKA C5000. Определяют также содержание протеинов (N·6,25, Kjeidahl метод Z130 для кормов и Z135 для фекалий и липидов (метод Z160).
Состав аминокислот также определяют, используя ВЭЖХ (метод Z100 для кормов и Z080 для фекалий). Содержание фосфора в кормах и экскретах определяют, используя AFNOR метод (NFV18-106).
b. Результаты и обсуждение
Очевидная обменная энергия (АМЕ)
Характеристики роста и данные об обменной энергии представлены в таблице 2.3. Характеристики (увеличение массы, потребление корма), измеряемые в течение трехдневного периода, не отличаются для обработок. АМЕ контрольного корма, содержащего 54% измельченной пшеницы, составляет 3173 ккал/кг. Обменная энергия корма, содержащего то же самое количество пшеницы, но 30% которого представляют целые зерна, повышается на 100 ккал/кг по сравнению с теоретическим значением. Более того, вариабельность, оцененная по стандартным отклонениям измеренных различных критериев, также уменьшается при использовании целых зерен пшеницы.
Ферменты, продуцируемые Penicillum funiculosum (активность β-глюканазы 100 ед/кг и активность ксиланазы 1100 ед/кг), повышают величину обменной энергии для корма на основе 54% пшеницы на +3,4% (122 ккал/кг DM), если пшеница измельчена, и на +2,7% (101 ккал/кг DM), если 30% пшеницы включены в виде целых зерен.
Очевидная усвояемость питательных веществ (липиды, протеины и аминокислоты)
Если измельчена вся пшеница, очевидная усвояемость липидов и протеинов повышается на 7 и 2,7% соответственно, при добавлении Препарата ферментов Penicillum funiculosum. Если часть пшеницы представлена в виде целых зерен, такое увеличение меньше: +3 и +0,6%, соответственно, за счет общей повышенной усвояемости питательных веществ. Действительно, усвояемость питательных веществ для контрольного корма, содержащего целые зерна пшеницы, сходна с усвояемостью экспериментального корма, содержащего только измельченную пшеницу, но с добавкой Препарата ферментов.
Влияние Препарата ферментов на очевидную усвояемость аминокислот представлено в таблице 2.4. Улучшение при использовании Препарата ферментов достигает в среднем +2,9% для измельченной пшеницы и +1,1% для пшеницы в виде целых зерен, что подтверждает влияние на очевидную усвояемость протеинов.
Очевидное удержание фосфора и экскреция фосфора
Влияние Препарата ферментов на очевидное удержание фосфора представлено в таблице 2.5. Очевидное удержание фосфора значительно повышается при добавлении Препарата ферментов: +8,0%. Такое повышение больше, нежели то, которое наблюдается для других питательных веществ (+2,9 до +3,5% в зависимости от критериев: АМЕ, протеины, липиды, аминокислоты). Такое повышение может быть результатом повышенной усвояемости питательных веществ (непосредственный эффект ксиланазы и β-глюканазы), но также и результатом усиленного действия фитазы пшеницы. В результате гидролиза не крахмалистых полисахаридов ксиланаза и β-глюканаза обеспечивают повышенную доступность к фитовой кислоте для эндогенной фитазы пшеницы.
Такая улучшенная усвояемость фосфора приводит к снижению экскреции фосфора: -8%, если выразить в г фосфора на кг увеличения массы.
1: одинарный способ анализа отклонений, эффект корма, н=47; а, b: значения с одинаковыми верхними индексами не отличаются при р<0,05.
2: двойной способ анализа отклонений, н=47 (пшеница: 54% измельченной или 24% измельченных +30% целых зерен пшеницы; фермент; с 0,21/t ксилана или без него;
3: FCR: Показатель превращения корма (г корма: г увеличения массы)
Пример 3. Оценка Препарата ферментов в отношении АМЕN кормов на основе пшеницы для растущих индюшек
Целью этого эксперимента является демонстрация эффективности Препарата ферментов из Penicillum funiculosum (активность β-глюканазы 100 ед/кг и активность ксиланазы 1100 ед/кг) в отношении очевидной обменной энергии (АМЕ) кормов на основе пшеницы в соответствии со следующей схемой эксперимента
(1) Контроль
(2) ЕР 1: Препарат ферментов (активность β-глюканазы 100 ед/кг и активность ксиланазы 1100 ед/кг)
(3) ЕР 2: Препарат ферментов (активность β-глюканазы 150 ед/кг и активность ксиланазы 1650 ед/кг)
В соответствии с Европейским методом сравнения (Bourdillion et al., 1990): корм ad libitum и полный сбор экскрета для возраста с 33 до 37 дня.
а. Материалы и методы
Птицы: Порода и условия выращивания
Однодневных индюков BUT9 содержат в клетках общей батареи до возраста 20 дней. Затем их переводят в индивидуальные клетки для определения баланса усвояемости после адаптационного периода, по крайней мере, в 7 дней.
Поддерживают стандартную температуру и влажность. Постоянный световой режим составляет: 23 часа света и 1 час темноты в течение первых двух недель, а затем световой режим уменьшают до 15 часов света и 9 часов темноты до конца испытаний.
Корма: Птицы получают стандартный полный начальный корм с первого до 21 дня жизни, а затем получают экспериментальные корма.
Экспериментальные корма
Корма содержат 47% пшеницы и 33% соевой муки (таблица 3.1). Распыление ферментов осуществляют на 20 кг частиц контрольных кормов.
Измерение обменной энергии
На 21 день (Д 21) птиц взвешивают, и равномерно распределяют в 10 отдельных клеток для каждой обработки, и дают экспериментальные корма.
Баланс начинают определять с 33 дня по следующей схеме:
Д 32, птицам не дают корма в течение ночи;
Д 33, птиц взвешивают, очищают сборочные лотки;
Д 34 и Д 35, фекалии собирают и замораживают;
Д 36, фекалии собирают и замораживают, ночью не дают корма;
Д 37, фекалии собирают и замораживают, птиц взвешивают и снова кормят.
Фекалии сушат вымораживанием и измельчают как корм (1 мм, измельчитель Retsch). Общую энергию корма и экскрета определяют с помощью адиабатического колориметра IKA C5000.
Величину АМЕ корректируют по азотному балансу, учитывая увеличение массы тела (г) и содержание азота (21% сырого протеина).
Фекалии сушат вымораживанием и измельчают как корм (1 мм, измельчитель Retsch). Общую энергию корма и экскрета определяют с помощью адиабатического колориметра IKA C5000. Определяют также содержание протеинов (N·6,25, Kjeidahl метод Z130 для кормов и Z135 для фекалий). Состав аминокислот также определяют, используя ВЭЖХ (метод Z100 для кормов и Z080 для фекалий).
b. Результаты и обсуждение Очевидная обменная энергия (АМЕ)
Зоотехнические характеристики и величины обменной энергии представлены в таблице 3.2. В характеристиках роста нет значительных различий при балансе между обработками.
При выращивании индюков ферментный препарат повышает АМЕN корма на основе пшеницы на 2,2 и 5,4% для ЕР 1 и ЕР 2, соответственно.
Столь значительное наблюдаемое увеличение демонстрирует пользу обоих активностей (ксиланазы и β-глюканазы), содержащихся в Препарате ферментов, в отношении гидролиза не крахмалистых полисахаридов пшеницы для увеличения энергетической ценности этого злака при выращивании индюков.
1 Одинарный способ дисперсионного анализа отклонений:
Эффект ферментов: н=60; а, b: означает, что то, что не обозначено одинаковыми буквами, значительно отличается при р<0,05; Эффект дозы: 0, 0,2, 0,3 I/t.
2 Среднее ± SEM
Пример 4. Оценка влияния Препарата ферментов, продуцируемых Penicillum funiculosum, на эффективность полного корма на основе пшеницы для растущих свиней.
Цель состоит в оценке влияния добавок ферментов к кормам на основе пшеницы на усвояемость энергии в тонком кишечнике растущих свиней. Нормальные уровни активности Препарата ферментов составляют 1100 ед/кг для ксиланазы и 100 ед/кг для β-глюканазы.
а. Материал и методы
Животные
Контрольные обработки проводят в соответствии со схемой Latin square для трех кормов и трех периодов и двух свиней для каждого из режимов кормления. Корма дают в фиксированных количествах в соответствии с массой свиней на протяжении периода тестирования.
Экспериментальные корма
Корма на основе пшеницы плохого качества и сбалансированные за счет других типичных кормовых ингредиентов дают шести растущим свиньям (см. таблицу 4.1). Дают один из кормов:
1. Корм без добавок (основной);
2. Корм с добавками (1): с Препаратом ферментов 1 × количество (активность β-глюканазы 100 ед/кг и активность ксиланазы 1100 ед/кг).
3. Корм с добавками (2): с Препаратом ферментов 2 × количество (активность β-глюканазы 200 ед/кг и активность ксиланазы 2200 ед/кг).
Точные дозы кормов получают, разбавляя Препарат ферментов кукурузным крахмалом для создания предварительной смеси, которую затем добавляют к корму в соответствии с необходимостью.
Сбор образцов
Соки подвздошной кишки собирают в течение 48 часов каждую неделю в соответствии со стандартными процедурами в RPNA лабораториях. Энергию образцов сока подвздошной кишки и тестируемого корма анализируют с помощью калориметрической бомбы по Сандерсу (Sanders) для определения усвоенной энергии. Аликвоты образцов хранят для дальнейших анализов при необходимости.
Статистический анализ
Усвояемость сырой энергии рассчитывают из результатов для соков подвздошной кишки, полученных с помощью калориметрической бомбы, для кормов и потребления кормов. Анализ отклонений предпринимают в отношении расчетов усвояемости.
b. Результаты и обсуждение
Добавки ксиланазы к корму свиней повышают усвояемость энергии, по крайней мере, на 6%. Это указывает на то, что фермент усиливает разрушение стенок клеток сырого материала(в частности, пшеницы) и высвобождает дополнительную энергию в тонком кишечнике
Пример 5. Влияние Препарата ферментов, продуцируемых Penicillum funiculosum на характеристики кормов из соломы, кукурузного силоса, сена и травяного силоса у жвачных животных.
HFT тест (Hohenheimer Futterwertesten, Menke et al., 1979, 1988) представляет собой in vitro инкубационный тест, позволяющий определять разложение сырого материала путем измерения объема газа, выделяемого при ферментации этих кормов в забуференном соке рубца.
а. Материал и методы
200 мг высушенного и измельченного субстрата инкубируют с 10 мл сока рубца плюс 20 мл буфера в шприцах, содержимое которых осторожно перемешивают на роторе в инкубаторе с контролируемой температурой (39°С). Объем выделившегося газа регистрируют через 24 часа. Для внесения поправок в результаты проводят холостой (без субстрата) и стандартный контроль с сеном и стандартный контроль с концентратом (с известным количеством вырабатываемого объема газа), и рассчитывают полный объем газа, вырабатываемого за 24 часа. Энергетическую ценность (OMD) (усвояемость органического вещества) субстратов рассчитывают, используя объем газа, выделившийся за 24 часа, и используя прогнозирующие уравнения, предложенные Menke с сотр. (1988).
Сок рубца собирают у 2 яловых коров, в рубец вводят канюлю и кормят в 8 утра и 7 вечера кормом, состоящим из 6 кг сена и 2 кг концентрата (отношение 75/25). Сок рубца собирают непосредственно перед утренним кормлением. Сок рубца фильтруют, чтобы избежать попадания пищевых частиц, и содержат в строгих анаэробных условиях.
Целью этого испытания является проверка влияния добавки Препарата фермента на фураж за 15 часов перед HFT инкубированием.
Предварительная обработка Препаратом фермента: раствор фермента распыляют на фураж, разложенный на полу, на солому, на кукурузный силос, на сено и травяной силос. Распыление осуществляют, используя 1 мл Препарата ферментов на 2 кг сухого вещества фуража. Фураж с краев (около 10 см) удаляют, чтобы повысить однородность образца. После обработки фураж смешивают вручную и оставляют при комнатной температуре на 15 часов после опрыскивания. HFT инкубирование осуществляют после 15 часов контакта с Препаратом ферментов для одной серии и 6 повторов для каждой обработки.
b. Результаты и обсуждение
Полный объем газа, выделившегося за 24 часа, представлен в таблице 5.1 для соломы, кукурузного силоса, сена и травяного силоса.
Применение целлюлазы для соломы в результате предварительной обработки дает 18% увеличения полного объема газа по сравнению с контролем. Для кукурузного силоса такое увеличение составляет 8%, для сена 9,5% и для травяного силоса 9%. OMD приведено в таблице 5.2 для различных типов фуража до и после предварительной обработки.
ОМ усвояемость соответственно повышается для соломы, кукурузного силоса, сена и травяного силоса по сравнению с контролем на 8,5% для соломы, 5% для кукурузного силоса, 5,4% для сена и 5% для травяного силоса.
15 часов предварительной обработки фуража (соломы, кукурузного силоса, сена и травяного силоса) Препаратом ферментов повышают интенсивность инкубирования субстрата рубца и ОМ усвояемость субстрата.
Пример 6: Влияние Препарата ферментов, продуцируемых Penicillum funiculosum на производительность несушек, которых кормят пшеницей или ячменем
Этот эксперимент осуществляют с целью определить влияние добавления Препарата ферментов на параметры продуктивности несущихся кур, которых кормят пшеницей или ячменем.
а. Материал и методы
Схема эксперимента: 4 обработки × 8 повторов × 5 клеток × 3 курицы
Обработки: 1. Контроль 1: 60% пшеницы
2. Контроль 1 + Препарат ферментов
3. Контроль 2: 60% ячменя
4. Контроль 2 + Препарат ферментов
Животные, содержание и эксперимент
Эксперименты проводят на 480 коричневых курах штамма Ну-Line. Каждый повтор осуществляют для 5 кур с обычными кормушками, т.е. всего 32 повтора для 15 кур каждый.
Распределенные в двух идентичных помещениях, эти повторы получают программируемое освещение и вентиляцию. Программа освещенности составляет 14 часов света в день по прибытии кур 17-недельного возраста и повышается каждые 2 недели на 30 минут вплоть до максимального значения 17 часов света в день.
Возраст кур в начале эксперимента 22 недели, что продолжается на протяжении 5 месяцев периода яйценоскости.
Корма и кормление
Имеются два вида экспериментального корма на основе 60% пшеницы (корм 1) и 60% ячменя (корм 2), и 10% муки подсолнечника. Их состав приведен в таблице 6.1. Характеристики злаков представлены в таблице 6.2.
Контроль
Химический анализ:
Образцы корма
Контроль за качеством экспериментальных кормов осуществляют, анализируя сухое вещество, сырой протеин, сырой жир и золу.
Активность ксиланазы (Т-1, Т-2) и активность β-глюканазы (Т-3 и Т-4) определяют в смешанных кормах.
Измерения
Расход корма и эффективность корма регистрируют каждые 4 недели. Кур взвешивают в начале и в конце эксперимента. Количество снесенных яиц, массу яиц и % сухого вещества и жира в яйцах записывают ежедневно в течение 5 периодов по 4 недели каждый. Смертность контролируют и регистрируют ежедневно, включая причину смерти.
b. Результаты и обсуждение
Проведение эксперимента
Полученные параметры продуктивности в течение эксперимента представлены в таблицах 6.3-6.5. В первые два периода (недели 22-30) во всем эксперименте % грязных яиц статистически обрабатывается (Р>0,005). Птицы, которым дают корм без ферментов, несут больше грязных яиц. Статистическая значимая разница между обработками была обнаружена для % снесенных яиц (Р>0,05) и для массы яиц (Р>0,005), начиная со второго периода до конца эксперимента. Куры, которых кормили кормом на основе ячменя, демонстрируют больший % снесенных яиц, и они несли более тяжелые яйца, нежели куры, которых кормили пшеницей. Препарат ферментов, по-видимому, повышает эти параметры, но не значительно, с величиной 0,05 уровня вероятности.
Во всех экспериментальных периодах потребление корма птицами для Т-3 и Т-4 обработок (ячменный корм) было выше, нежели потребление птицами, которым давали пшеницу, из-за энергетических уровней обоих кормов (ячменный корм соответствует 2600 ккал/кг энергии, тогда как пшеничный корм соответствует 2800 ккал/кг). Учитывая различные величины энергии кормов обоих типов, и потребление птицами корма, во весь период для всех птиц ежедневное потребление энергии было одинаковым.
Эффективность корма (выраженная как г корма/г яиц) для экспериментальных кормов на протяжении первого периода была очень высока из-за малого количества яиц для каждой курицы на протяжении этого отрезка времени. В первые два периода эффективность корма из пшеницы была выше эффективности корма из ячменя; но в третьем периоде, когда был зарегистрирован более высокий % снесенных яиц, оба типа кормов представили сходные эффективности. Начиная с 34 недели до конца эксперимента, ячменные корма демонстрировали более высокую кормовую эффективность, нежели пшеничные корма. Ферменты проявили тенденцию улучшить кормовую эффективность (Р>0,05). Во все периоды β-глюканаза повышала кормовую эффективность ячменного корма (с Р=0,066).
В таблице 6.4 показано, что несушки, которых кормили пшеницей с добавкой Препарата ферментов, имели тенденцию к демонстрации более высоких значений яйценоскости (+1,5 абс. точек), в среднем более тяжелых яиц (+0,37 г) и более низкий показатель превращения корма (-2,7%) по сравнению с кормами без добавок.
Таблица 6.5 демонстрирует, что добавление препарата ферментов к ячменному корму для несушек повышает яйценоскость (+4%) среднюю массу яиц (+0,7%) и показатель превращения корма (-5,7%) по сравнению с контрольными ячменными кормами.
(*) 1 кг корма содержит: витамин А8000 ед ; витамин D3 1600 ед; витамин Е 5 мг; витамин К3 2 мг; витамин В1 1,5 мг; витамин В2 4 мг; витамин В6 3 мг; витамин В12 11,8 мкг; фолиевая кислота 0,35 мг; биотин 150 мкг; пантотенат кальция 10 мг; никотиновая кислота 20 мг; Mn 30 мг; Zn 50 мг; I 0,3 мг; Fe 50 мг; Cu 6 мг; Se 0,1 мг; этоксиквин 125 мг.
1 Сравнение с генетически совершенными курами (данные предоставлены поставщиком кур). Величины являются средними из 8 повторов для 15 кур. Внутри колонок значения с последующими различными индексами значительно отличаются (Р<0,05)
3. Для периода 22-42 недели
Для периода 22-42 недели
Библиография
1. Bourdillon А., Carre В., Conan L, Duperray J., Franscesch М., Fuentes M., Huyghebaert G., Jansen W.M.M.A., Leclercq В., Lessire М., McNab J., Rigoni М., Wiseman J., 1990. European reference method for the in vivo determination of metabolizable energy with adult cockerels: reproductibility, effect of age, comparison with predicted values. British Poultry Science 31, 567-576.
2. Sabatier A.M., Fish N.M. 1996. Method of analysis for feed enzymes: methodological problems? Journal of Applied Poultry Research 5, 408-413.
3. Barrier-Guillot В., Metayer J.P., Bouvarel I., Castaing J., Picard M.. Zwick J.L. 1997.
4. Proceedings of the Xlth European Symposium on Poultry Nutrition, WPSA, Aug 24-28 th Faaborg, Denmark, 237-239.
5. Svihus В., Herstad O., New man C.W., Newman R.K. 1997. British Poultry Science 38, 524-529.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ ГРИБА Penicillium verruculosum B10 EGII ПРОДУЦЕНТ ЭНДО-1.3/1.4-β-ГЛЮКАНАЗЫ, ЦЕЛЛЮЛАЗЫ, β-ГЛЮКОЗИДАЗЫ И КСИЛАНАЗЫ И СПОСОБ ПОЛУЧЕНИЯ КОРМОВОГО КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА | 2012 |
|
RU2532840C2 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ПРОДУЦЕНТ КОМПЛЕКСА ФИТАЗЫ А И ЭНДО-1,4-β-ГЛЮКАНАЗЫ II И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ ДОБАВКИ В КОРМАХ | 2022 |
|
RU2810538C2 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ КОМПЛЕКСА ФЕРМЕНТОВ ЭНДОГЛЮКАНАЗ И КСИЛАНАЗ В КЛЕТКАХ ГРИБА PENICILLIUM VERRUCULOSUM И СПОСОБ ПОЛУЧЕНИЯ КОМПЛЕКСНЫХ ФЕРМЕНТНЫХ ПРЕПАРАТОВ НА ЕГО ОСНОВЕ, ПРЕДНАЗНАЧЕННЫХ ДЛЯ КОРМОПРОИЗВОДСТВА | 2017 |
|
RU2653429C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ЕЕ-105 ПРОДУЦЕНТ КОМПЛЕКСА ВЫСОКОЭФФЕКТИВНЫХ ЭНДОГЛЮКАНАЗ И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ КОРМОВОЙ ДОБАВКИ В ЗЕРНОВЫХ КОРМАХ | 2018 |
|
RU2696036C1 |
| СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА ДЛЯ РАСЩЕПЛЕНИЯ ГЕМИЦЕЛЛЮЛОЗНЫХ ГЕТЕРОПОЛИСАХАРИДОВ КЛЕТОЧНОЙ СТЕНКИ РАСТЕНИЙ И ФЕРМЕНТНЫЙ ПРЕПАРАТ (ВАРИАНТЫ) | 2007 |
|
RU2358756C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ПРОДУЦЕНТ КОМПЛЕКСА ФИТАЗЫ А И ЭНДО-1,4-β-КСИЛАНАЗЫ Е И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ ДОБАВКИ В КОРМАХ | 2023 |
|
RU2819918C1 |
| ШТАММЫ-ПРОДУЦЕНТЫ ФЕРМЕНТОВ И СПОСОБ ПОЛУЧЕНИЯ МУЛЬТИЭНЗИМНОЙ КОМПОЗИЦИИ ПРОТЕОЛИТИЧЕСКОГО И ГЕМИЦЕЛЛЮЛОЛИТИЧЕСКОГО ДЕЙСТВИЯ, ПРЕДНАЗНАЧЕННОЙ ДЛЯ ПРИМЕНЕНИЯ В КОРМОПРОИЗВОДСТВЕ | 2016 |
|
RU2636040C1 |
| ТЕРМОСТАБИЛЬНАЯ КОМПОЗИЦИЯ ДЛЯ ЖИВОТНЫХ, СОДЕРЖАЩАЯ СМЕСЬ ФЕРМЕНТОВ | 2008 |
|
RU2481000C2 |
| Штамм мицелиального гриба TRICHODERMA LONGIBRACHIATUM TW-14-220 - продуцент целлюлаз, бета - глюканаз и ксиланаз для кормопроизводства и способ получения кормового комплексного ферментного препарата | 2017 |
|
RU2654564C1 |
| СПОСОБ ПОЛУЧЕНИЯ КОРМОВОГО КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА (ВАРИАНТЫ) И ШТАММ PENICILLIUM CANESCENS (ВАРИАНТЫ) | 2004 |
|
RU2288267C2 |

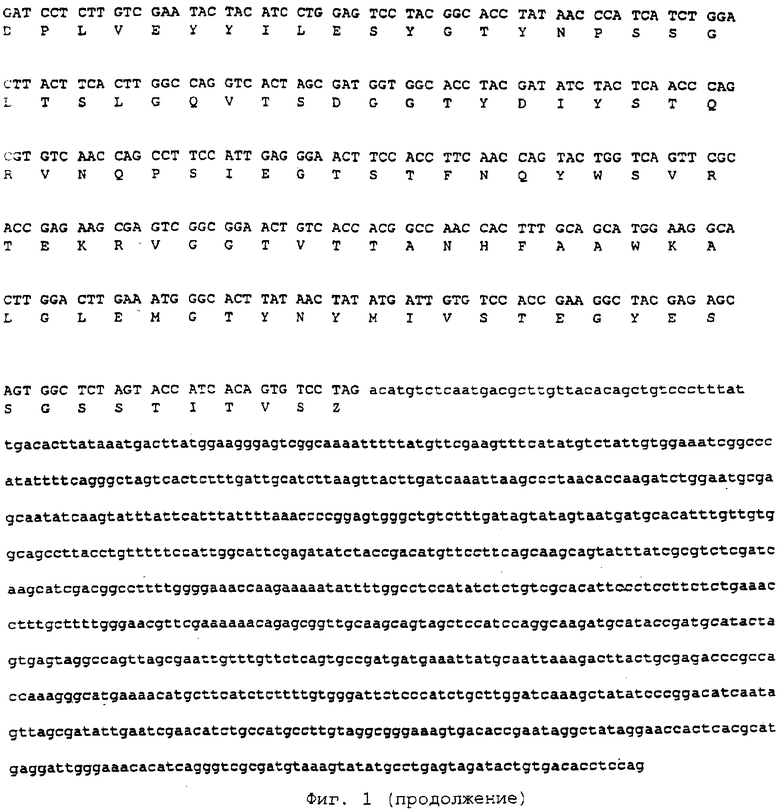



Изобретение относится к биотехнологии и может быть использовано в кормопроизводстве для приготовления кормовой добавки и корма для сельскохозяйственных животных. Штамм Penicillium funiculosum IMI378536 продуцирует комплекс ферментов - целлюлозу, эндо-1,4-β-ксиланазу, целлобиогидролазу, β-глюкозидазу, эндо-1,3(4)-β-глюканазу, ферулоил-эстеразу. Жидкая кормовая добавка для сельскохозяйственных животных, таких как домашняя птица, свиньи и жвачные, содержит микробный продукт, полученный из штамма Penicillium funiculosum IМI378536 по п.1, дополненный, в случае необходимости, ферментом (ферментами) определенным (определенными) в п.1, противомикробный агент, сорбит, антифриз, концентрированный отфильтрованный ферментационный бульон при заданном содержании компонентов. Сухой корм для сельскохозяйственных животных, таких как домашняя птица, свиньи и жвачные, содержит микробный продукт, полученный из штамма Penicillium funiculosum IМI378536 по п.1, дополненный, в случае необходимости, ферментом (ферментами) определенным (определенными) в п.1, носитель, компоненты сухого ферментационного бульона при заданном содержании компонентов. Изобретение обеспечивает повышение усвояемости аминокислот и кормов для животных, уменьшение выделения фосфора и аммиака из помещений, где кормят животных. 3 н. и 22 з.п. ф-лы, 4 ил., 32 табл.
| DE 1945147 А, 14.05.1970 | |||
| WO 9605739 A3, 29.02.1996 | |||
| LACAKE et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Enzyme and Microbial Technology | |||
| Пневматический водоподъемный аппарат-двигатель | 1917 |
|
SU1986A1 |
| СПОСОБ ПРИГОТОВЛЕНИЯ КОМБИКОРМА ДЛЯ ЦЫПЛЯТ-БРОЙЛЕРОВ | 1992 |
|
RU2017433C1 |

