ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к новому способу амплификации (увеличения количества) и получения характеристик нуклеиновых кислот.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Удлинение праймера (олигонуклеотида - затравки синтеза комплементарной нити) нуклеиновой кислоты на матрице целевой нуклеиновой кислоты - это очень важный процесс, имеющий многочисленные применения, в том числе для обнаружения (детектирования), диагностики и количественного определения нуклеиновых кислот. В частности, удлинение праймера на матрице:
(i) дает прямое доказательство того, что произошла гибридизация (взаимодействие комплементарных нитей с образованием двунитевой структуры) праймера на матрице,
(ii) подтверждает наличие на этой матрице нуклеотидной последовательности, комплементарной праймеру, и таким образом
(iii) подтверждает наличие данного праймера или целевой нуклеиновой кислоты.
Используя близкородственные праймеры, отличающиеся всего лишь одним нуклеотидом, обычно удается различить нуклеиновые кислоты, различающиеся в своих нуклеотидных последовательностях единственным нукпеотидом. Критическое ограничение в использовании удлинения праймера на матричной нуклеиновой кислоте как способа обнаружения, диагностики и количественного определения нуклеиновой кислоты состоит в том, что в типичной реакции удлинения праймер отжигается (гибридизируется) с матрицей, но удлиняется всего лишь один раз. Поэтому количество матрицы в образце определяет количество удлиненного праймера. Для большинства применений в областях, связанных с обнаружением, диагностикой и количественным определением нуклеиновых кислот, количество матрицы часто слишком мало, чтобы обеспечить непосредственное обнаружение удлиненного праймера, если только эта реакция удлинения каким-то образом не повторяется или не осуществляется циклически.
Чтобы преодолеть это критическое ограничение, необходимы процессы амплификации. Многие такие процессы были описаны ранее и они по существу включают:
(a) условия циклической диссоциации, когда удлиненный праймер с помощью нагревания отделяется от матричной нуклеиновой кислоты, позволяя осуществить гибридизацию и последующее удлинение нового праймера;
(b) повторяющееся создание праймера в ДНК с помощью фермента рестрикции, чтобы произвести однонитевой разрыв в сайте распознавания в немодифицированной нити наполовину модифицированной ДНК и создать возможность для ДНК-полимеразы, лишенной 5'-3' экзонуклеазной активности, удлинить 3'-конец ДНК в месте однонитевого разрыва и заместить нисходящую нить ДНК;
(c) процесс, где матричная нуклеиновая кислота может быть включена в циклический процесс, так что удлинение праймера осуществляется непрерывно, и/или
(d) процесс, где способом транскрипции создаются копии матрицы, которые служат матрицами в последующих реакциях удлинения праймера.
Амплификация нуклеиновой кислоты (создание многих копий) до такого уровня, когда можно их детектировать (обнаружить) и производить с ними определенные действия, используется очень широко. Такие процессы амплификации, дающие достаточные количества специфической целевой нуклеиновой кислоты, являются, как правило, первым ключевым этапом, необходимым для получения характеристик нуклеиновой кислоты. Амплификация специфической последовательности нуклеиновой кислоты прямо из пробы позволяет определить, присутствует или отсутствует указанная нуклеиновая кислота в этой пробе, и очень широко применяется поэтому в ДНК / генной диагностике. Амплификация специфической последовательности нуклеиновой кислоты прямо из пробы и последующее получение ее характеристик могут осуществляться для многих целей, в том числе для диагностики наличия или отсутствия вариаций ДНК, таких как мутации и полиморфизмы в определенной нуклеиновой кислоте. Во многих случаях процессы амплификации могут быть спланированы таким образом, чтобы обеспечить и амплификацию, и получение полных или частичных характеристик амплифицированной матрицы.
Каждый из известных к настоящему времени способов амплификации и получения характеристик нуклеиновых кислот имеет различные недостатки, особенно в части процесса, необходимого для диссоциации праймера, создания праймера, специфичности, разносторонности, создания однонитевой ДНК и облегчения умножения и высокопроизводительного генотипирования, в особенности генотипирования однонуклеотидных полиморфизмов (single nuclear polymorphisms, SNP), описанных ниже.
Удлинение праймера
Как таковое, удлинение праймера по матрице очень хорошо документировано в литературе. Оно может быть осуществлено отжигом (гибридизацией) праймера с комплементарной ему последовательностью на матрице или целевой нуклеиновой кислоте с последующей инкубацией с ДНК-полимеразой в присутствии предшественников ДНК, обычно dATP, dGTP, dCTP и dTTP (соответственно дезоксирибоаденозин-, дезоксирибогуанозин-, дезоксирибоцитидин- и дезоксириботимидинтрифосфатов), в результате чего происходит удлинение праймера с его 3'-ОН (гидроксильного) конца (окончания) в направлении от 5' к 3' (в праймере) на матричной нити и создается вновь синтезированная нить ДНК, комплементарная исходной матрице. Удлинение праймера может происходить только тогда, когда праймер имеет свободный 3'-ОН конец. Для амплификации этой комплементарной нити на матрице способами удлинения праймера необходимо:
(a) чтобы удлинение праймера происходило более одного раза на одну гибридизированную молекулу праймера,
(b) чтобы новый праймер мог быть повторно гибридизирован с матрицей и удлинен и
(c) чтобы комплементарная нить служила матрицей для последующего удлинения второго праймера.
Полимеразная цепная реакция (ПЦР)
Амплификация матрицы нуклеиновой кислоты может быть осуществлена с помощью полимеразной цепной реакции (ПЦР) (Saiki R.K. и др. // Science. 1988. Т.239. С.487-491).
Амплификацию целевого сегмента матрицы нуклеиновой кислоты методом ПЦР обычно проводят с использованием в ПЦР синтетических олигонуклеотидных праймеров вместе с термостабильной ДНК-полимеразой и предшественниками ДНК. Многократные циклы денатурации, гибридизации и удлинения праймера дают в результате экспоненциальную амплификацию целевого сегмента. Поэтому обнаружение продукта указывает на наличие в специфическом локусе определенной последовательности. Длина амплифицированного продукта определяется суммарной длиной праймеров, расстоянием между их 3'-концами на матрице и так далее.
Для ПЦР абсолютно необходима термостабильная ДНК-полимераза. ПЦР включает также процесс циклического изменения температуры (термоциклизация), поэтому для автоматизации этого метода требуется специальное оборудование. В типичной ПЦР для инициирования реакции обычно необходимы два праймера, поэтому это вносит в реакцию дополнительную сложность, особенно, если хотят произвести в одной пробирке несколько различных реакций амплификации. Это приводит к повышению числа праймеров, присутствующих в реакции, и, следовательно, повышает вероятность ошибочной и неспецифической амплификации. Кроме того, усложняются также планирование и оптимизация ПЦР, поскольку температура гибридизации в реакции должна годиться для обоих праймеров, используемых в реакции. Это представляет собой дополнительное неудобство, если мулитиплицируются различные реакции амплификации, поскольку одна температура гибридизации должна годиться для всех реакций амплификации, подлежащих мультиплицированию.
Кроме того, имеется опасность загрязнения ампликона (амплифицируемого участка), так как в ходе реакции создается множество копий матричной нуклеиновой кислоты, которые должны служить последующими матрицами.
Способы амплификации на основе транскрипции
Амплификация на основе транскрипции (Kwoh D.Y. и др. // Proc. Natl. Acad. Sci. USA. 1989. Т.86. С.1173-1177; Guatelli J.C. и др. // Proc. Natl. Acad. Sci. USA. 1990. Т.87. С.1874-1878; Compton J. // Nature. 1991. Т.350. С.91-92) представляет собой способ амплификации, который основывается на удлинении праймеров на целевой нуклеиновой кислоте, происходящем так, что в восходящем направлении от целевой области, подлежащей амплификации, создается промотор для РНК-полимеразы. По существу с матричной нуклеиновой кислотой инкубируются два праймера, фланкирующих целевую последовательность. Один из праймеров имеет последовательность промотора для РНК-полимеразы в восходящем направлении (5') от последовательности, которая комплементарна матрице и гибридизируется на ней. Другой праймер комплементарен сегменту нити, комплементарной матрице. Праймер с последовательностью промотора для РНК-полимеразы удлиняется на матричной нити с помощью полимеразы нуклеиновой кислоты (такой, как обратная транскриптаза) и предшественников ДНК. После тепловой денатурации гибридной нуклеиновой кислоты (матричная нить и вновь синтезированная комплементарная нить) или ферментативного разрушения матричной нити на вновь синтезированной комплементарной нити гибридизируется и удлиняется второй праймер. В результате получается продукт, который является двунитевым и имеет промотор для РНК-полимеразы, присоединенный к целевой последовательности. Инкубация этого продукта с РНК-полимеразой и предшественниками РНК приводит к продуцированию многочисленных РНК-транскриптов этой целевой последовательности. Каждый РНК-транскрипт служит, в свою очередь, матрицей для продуцирования комплементарной нити ДНК, и этот процесс продолжается как самоподдерживающийся циклический процесс при изотермических условиях, пока не исчерпаются или не инактивируются компоненты реакции. Это приводит к крупномасштабной амплификации целевой последовательности нуклеиновой кислоты.
Главный недостаток состоит в том, что эта техника абсолютно зависит от РНК, которая имманентно более нестабильна, чем ДНК, и более восприимчива к деструкции. Следовательно, эта реакция сверхчувствительна к примесным рибонуклеазам. Обычно для этого способа требуется РНК в качестве матрицы и он не подходит или не оптимален для ДНК-матрицы.
Этот способ также требует для инициирования реакции наличия, по меньшей мере, двух праймеров, причем один из праймеров должен быть специально сконструирован таким образом, чтобы он включал сайт инициации транскрипции для РНК-полимеразы.
Кроме того, имеется опасность загрязнения ампликона, так как в ходе реакции создается множество копий матричной нуклеиновой кислоты, которые должны служить последующими матрицами.
Эти способы не могут быть использованы для обнаружения мутаций или полиморфизмов без введения дополнительных стадий.
Амплификация с замещением нити
Реакция с замещением нити (strand displacement amplification, SDA) основана на способности фермента рестрикции создать в немодифицированной нити, наполовину модифицированной двунитевой ДНК-однонитевой разрыв в специфическом сайте распознавания и способности ДНК-полимеразы, лишенной 5-3' экзонуклеазной активности, удлинить 3'-конец нити в месте полученного однонитевого разрыва и в этом процессе заместить нисходящую нить ДНК (Walker G.T. и др. // PNAS. 1992. Т.89. С.392-396). Экспоненциальная амплификация целевой ДНК достигается сопряжением реакций на матрице, в которых нити, высвобожденные из реакции на матричной нити, служат объектом для реакции на комплементарной нити и, наоборот. Весьма важно, что тепловая денатурация образца ДНК создает два однонитевых фрагмента ДНК (Т1 и Т2). В избытке присутствуют два праймера амплификации ДНК (Р1 и Р2). 3'-конец Р1 связывается с 3'-концом Т1, образуя дуплекс с 5' перекрываниями. Подобным же образом Р2 связывается с Т2. Перекрывания 5' Р1 и Р2 содержат распознаваемую последовательность для фермента рестрикции, такого как Hincll. ДНК-полимераза Е.coli в форме, свободной от 5-3' экзонукпеазной активности, удлиняет 3'-концы дуплексов с участием предшественников ДНК dGTP, dCTP, dTTP и модифицированного предшественника дезоксирибоаденозин-5'-[α-тио]-трифосфата, что приводит к формированию на Р1Т1 и Р2Т2 гемифосфотиоатных сайтов распознавания. Hincll производит однонитевые разрывы в незащищенных праймерных нитях гемифосфотиоатных сайтов распознавания, оставляя модифицированные комплементарные нити интактными. ДНК-полимераза удлиняет 3'-конец в месте разрыва на Р1Т1 и замещает нисходящую нить, которая функционально эквивалентна Т2. Подобным же образом удлинение в месте разрыва в Р2Т2 приводит к замещению нисходящей нити, которая функционально эквивалентна Т1. Стадии образования однонитевого разрыва и полимеризации/замещения повторяются циклически на Р1Т1 и Р2Т2, поскольку удлинение в месте разрыва воссоздает способный к расщеплению сайт распознавания для Hincll. Амплификация целевой последовательности экспоненциальна, так как нити, замещенные с Р1Т1, служат мишенями для (гибридизации) Р2, а нити, замещенные с Р2Т2, служат мишенями для Р1.
Главный недостаток SDA состоит в том, что необходимо использовать специально сконструированные праймеры, включающие сайт для специфического фермента рестрикции. Обычно для проведения реакции на каждый ампликон необходимо иметь два или больше специфичных к определенной последовательности праймера. Кроме того, амплифицированные фрагменты, получаемые в этой реакции с использованием единственного праймера, не склонны иметь определенный 3'-конец. Это также очень важный недостаток SDA, так как именно определенный 3'-конец на замещенном фрагменте позволяет ему инициировать последующую реакцию с тем же самым фрагментом.
В заявке WO 97/03210 раскрыт способ быстрого обнаружения наличия или отсутствия конкретной последовательности нуклеиновой кислоты в локусе-кандидате в образце целевой нуклеиновой кислоты, включающий следующие этапы:
(i) введение в указанный рассматриваемый локус (локус-кандидат) в одном или нескольких выбранных заранее положениях модифицированного гетероциклического основания, являющегося субстратом для ДНК-гликозилазы;
(ii) вырезание модифицированного основания с помощью ДНК-гликозилазы таким образом, чтобы создать лишенное основания звено;
(iii) расщепление фосфатных связей при лишенных основания звеньях, созданных в (ii); и
(iv) анализ продуктов расщепления этапа (iii) таким образом, чтобы идентифицировать в указанной последовательности целевой нуклеиновой кислоты наличие или отсутствие указанной конкретной последовательности нуклеиновой кислоты в указанном локусе-кандидате.
Способ применим, в частности, для обнаружения в образце ДНК специфических мутаций, в том числе обнаружения в ДНК многочисленных известных мутаций.
В заявке WO 99/54501 раскрыт способ получения характеристик молекул нуклеиновой кислоты, включающий следующие этапы:
(i) введение в молекулу ДНК модифицированного основания (например, урацила), которое является субстратом для ДНК-гликозилазы;
(ii) удаление (вырезание) модифицированного основания с помощью указанной ДНК-гликозилазы таким образом, чтобы создать лишенное основания звено;
(iii) расщепление ДНК у лишенного основания звена таким образом, чтобы создать направленный вверх фрагмент ДНК, который можно удлинить; и
(iv) инкубация способного к удлинению направленного вверх фрагмента в присутствии фермента (например, полимеразы или лигазы), обеспечивающего его удлинение, и матричной нуклеиновой кислоты и анализ полученного фрагмента (полученных фрагментов).
Однако для способа, описанного в заявке WO 99/54501, расщепление ДНК в звене, где удалено основание, является критичным.
В случае способа, проиллюстрированного примерами в заявке WO 99/54501, реакционную смесь, содержащую амплифицированную целевую нуклеиновую кислоту, обрабатывали экзонуклеазой I, чтобы расщепить не удлиненные на стадии амплификации праймеры, и щелочной фосфатазой из креветок, чтобы разрушить не использованные в амплификации дезоксирибонукпеозидтрифосфаты (dNTP). Поэтому дальнейшая амплификация матрицы нуклеиновой кислоты не происходит и способ ограничивается одиночным циклом.
Следовательно, важно разработать усовершенствованный способ амплификации и получения характеристик нуклеиновых кислот, который является более разносторонним, более специфичным, дает более высокую производительность, улучшает мультиплицирование реакции и позволяет получать однонитевую ДНК.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В соответствии со сказанным выше настоящее изобретение предлагает способ амплификации матричной нуклеиновой кислоты, который состоит в одновременном проведении следующих этапов:
(i) проведение реакции праймера нуклеиновой кислоты с указанной матричной нуклеиновой кислотой, нормальными нуклеотидами-предшественниками ДНК, по меньшей мере, одним модифицированным нуклеотидом-предшественником ДНК и ДНК-полимеразой таким образом, чтобы получить удлиненный праймер нуклеиновой кислоты, причем указанный праймер нуклеиновой кислоты остается связанным с указанной матрицей;
(ii) расщепление содержащего модифицированное основание удлиненного праймера нуклеиновой кислоты таким образом, чтобы создать свободный 3'-ОН конец, который может быть удлинен указанной ДНК-полимеразой; и
(iii) повторение этапов (i) и (ii) на созданных таким образом фрагментах ДНК.
В одном из примеров осуществления изобретения содержащий модифицированное основание удлиненный праймер нуклеиновой кислоты расщепляют 3'-эндонуклеазой.
В этом примере осуществления 3'-эндонуклеаза предпочтительно является эндонуклеазой V из E.coli или ее гомологами, которые можно найти в других организмах.
В альтернативном примере осуществления способ включает следующие этапы:
(i) проведение реакции праймера нуклеиновой кислоты с указанной матричной нуклеиновой кислотой, нормальными нуклеотидами-предшественниками ДНК, по меньшей мере, одним модифицированным нуклеотидом-предшественником ДНК, который является субстратом для ДНК-гликозилазы, и ДНК-полимеразой таким образом, чтобы получить удлиненный праймер нуклеиновой кислоты, остающийся связанным с указанной матрицей;
(ii) удаление модифицированного основания в модифицированном нуклеотиде-предшественнике ДНК из удлиненного праймера нуклеиновой кислоты с помощью ДНК-гликозилазы таким образом, чтобы создать лишенное основания звено;
(iii) расщепление удлиненного праймера нуклеиновой кислоты в лишенном основания звене таким образом, чтобы создать свободный 3'-ОН конец, который может быть удлинен указанной ДНК-полимеразой; и
(iv) повторение этапов (i)-(iii) на созданных таким образом фрагментах ДНК.
Способ в соответствии с настоящим изобретением имеет много специфических преимуществ, как указывается далее. Однако в более общем смысле способ настоящего изобретения имеет существенные преимущества перед существующими технологиями в том, что он более разносторонен и более гибок в смысле обеспечения одного процесса с высокой производительностью, который легко может быть приспособлен (адаптирован) ко многим различным форматам в области обнаружения (детектирования), количественного определения и получения характеристик ДНК.
Настоящее изобретение будет далее описано преимущественно со ссылкой на примеры осуществления, включающие применение ДНК-гликозилазы. Для данного способа в соответствии с настоящим изобретением авторами создан термин «амплификация при посредстве гликозилазы» ("glycosilase mediated amplification", GMA). Однако акроним GMA относится здесь в собирательном значении и ко всем примерам осуществления настоящего изобретения.
Таким образом, в способе по настоящему изобретению модифицированный нуклеотид-предшественник ДНК может быть субстратом для ДНК-гликозилазы или может распознаваться 3'-эндонуклеазой, как здесь описано.
Обычно матричной нитью нуклеиновой кислоты может быть любая нить природной или искусственно синтезированной нуклеиновой кислоты.
Предпочтительно матричной нуклеиновой кислотой является ДНК.
В способе настоящего изобретения затравка/инициация производится праймером нуклеиновой кислоты. Для удобства праймер, ответственный за инициацию реакции, назван здесь инициирующим праймером (ИП).
ИП может быть любая нуклеиновая кислота со свободным 3'-ОН концом, которая может быть удлинена ДНК-полимеразой. ИП может быть искусственно синтезированным, например синтетическим, олигонуклеотидом или может быть получен прямо или опосредованно из имеющейся в природе нуклеиновой кислоты.
В одном из примеров осуществления праймером нуклеиновой кислоты является праймер ДНК.
Нормальными нуклеотидами-предшественниками ДНК являются dATP, dCTP, dGTP и dTTP. При определенных лимитирующих условиях могут быть также использованы и включены в реакцию дидезоксинуклеотидтрифосфаты. Таким образом, возможные нормальные предшественники включают также ddATP, ddCTP, ddGTP и ddTTP.
Предпочтительно нуклеотиды-предшественники ДНК выбраны из dATP, dCTP, dGTP и dTTP.
В реакции GMA может быть использована любая из нескольких полимераз нуклеиновой кислоты. Если используют матрицу ДНК, применяют ДНК-полимеразу. Если используют матрицу РНК, применяют ДНК-полимеразу, которая может работать по матрице РНК, обычно таким ферментом является обратная транскриптаза. Обычно используют два класса ДНК-полимераз в зависимости от того, что необходимо: замещение или расщепление (переваривание) нуклеиновой кислоты в нисходящем направлении от удлиненного праймера. Если необходимо замещение нити, создающее таким путем замещенные фрагменты, используют ДНК-полимеразу, не имеющую 5'-3' экзонуклеазной активности. Наоборот, если используют ДНК-полимеразу с 5-3' экзонуклеазной активностью, в каждом цикле реакции GMA нисходящая ДНК разрушается. Это также приводит к формированию обнаруживаемого продукта, как описано далее.
Существует несколько известных модифицированных нуклеотидов-предшественников, которые, будучи включены в ДНК, становятся субстратом для ДНК-гликозилазы и/или распознаются 3'-эндонуклеазой, в зависимости от необходимости. В последнем случае модифицированный нуклеотид-предшественник направляет расщепление ферментом 3'-эндонуклеазой на фосфодиэфирную связь, находящуюся в положении 3' по отношению к сайту его включения.
В каждом случае расщепление зависит от наличия в ДНК модифицированного основания и именно оно диктует расщепление ДНК и диктует локализацию места расщепления, выполняемого по одному из двух путей, а именно:
1) выщепление модифицированного основания гликозилазой и расщепление в полученном лишенном основания звене или
2) расщепление ферментом 3'-эндонуклеазой удлиненного праймера нуклеиновой кислоты во второй фосфодиэфирной связи с 3' стороны звена с включенным модифицированным основанием.
В одном из примеров осуществления модифицированным предшественником нуклеиновой кислоты является dUTP (дезоксирибоуридинтрифосфат).
Модифицированный нуклеотид-предшественник dUTP представляет собой основание с сахаром и фосфатом, содержит основание урацил и сахаро-фосфатный остаток. Удлинение праймера на матрице с использованием нуклеотидов-предшественников dATP, dCTP, dGTP и dUTP вместо dTTP приводит в результате к образованию вновь синтезированной ДНК, комплементарной матрице, в которой тимин полностью замещен на урацил.
Однако специалистам в данной области должно быть понятно, что могут быть использованы другие модифицированные предшественники нуклеиновой кислоты, такие как dITP (дезоксирибоинозинтрифосфат) и 8-OH-dGTP.
Модифицированный нуклеотид-предшественник dITP представляет собой основание с сахаром и фосфатом, содержит основание гипоксантин и сахаро-фосфатный остаток. Модифицированный нуклеотид-предшественник 8-OH-dGTP представляет собой основание с сахаром и фосфатом, содержит основание 8-ОН-гуанин и сахаро-фосфатный остаток.
Предшественники гликозилазного субстрата dUTP, dITP и 8-OH-dGTP, будучи включенными в ДНК, дают субстраты для гликозилазы - соответственно основания урацил, гипоксантин и 8-ОН-гуанин.
В одном из примеров осуществления ДНК-гликозилазой является урацил-ДНК-гликозилаза (УДГ, UDG).
UDG специфически распознает урацил в ДНК и выщепляет его из ДНК. UDG также распознает другие родственные урацилу основания, если они присутствуют в ДНК.
Было описано много ДНК-гликозилаз. Эти ферменты расщепляют N-гликозидную связь, соединяющую основание, являющееся субстратом для гликозилазы, со скелетом ДНК. Это высвобождает основание из ДНК и создает лишенное основания звено.
Другие подходящие ДНК-гликозилазы включают алкилпурин-ДНК-гликозилазы (ADG) или формамидопиримидин-ДНК-гликозилазу (FPG).
Алкилпурин-ДНК-гликозилазы (ADG) специфически распознают и удаляют из ДНК гипоксантин. Этот фермент также распознает и выщепляет N3-метиладенин, N3-метилгуанин, О2-метилцитозин и О2-метилтимин, если они имеются в ДНК. ДНК-гликозилазы FPG специфически распознают и высвобождают из ДНК 8-ОН-гуанин. Этот фермент также распознает и выщепляет другие пурины с разомкнутым кольцом, если они имеются в ДНК.
Известны несколько агентов, которые расщепляют в ДНК фосфодиэфирные связи у лишенных основания звеньев. Расщепление связи может происходить с 5'-стороны или с 3'-стороны лишенного основания звена. Расщепление с 5'-стороны может происходить проксимально или дистально по отношению к фосфатному остатку и создавать направленный вверх фрагмент, который имеет 3'-окончание соответственно со свободной 3'-ОН группой или с 3'-фосфатной группой. Окончания 3'-ОН способны удлиняться ДНК-полимеразами на матрице, тогда как окончания с 3'-фосфатом к удлинению не способны. Такие 3'-фосфатные окончания могут, вообще говоря, быть сделаны способными к удлинению обработкой ферментом фосфатазой, такой как полинуклеотидкиназа Т4, которая имеет 3'-фосфатазную активность. Агентами, которые расщепляют фосфодиэфирную связь в положении 5' по отношению к фосфатному остатку и создают 3'-конец со свободной 3'-ОН группой, являются ферменты с апурин- и апиримидинэндонуклеазной (АР endonuclease) активностью, такие как АР эндонуклеаза IV из E.coli. Агентами, расщепляющими связь с 3' стороны фосфатного остатка и создающими 3'-конец с 3'-фосфатной группой, являются щелочи, тепло и определенные ферменты репарации ДНК, такие как FPG, а также основные белки и пептиды. Агенты, выполняющие расщепление с 3' конца лишенного основания звена, включают тепло и ферменты репарации ДНК с активностью АР лиазы, такие как эндонуклеаза III из E.coli. Такие 3'-дезоксирибофосфатные (dRp) концы могут быть, вообще говоря, превращены в удлиняемые с помощью обработки АР эндонуклеазой. ДНК-гликозилаза FPG расщепляет связь как с 5' стороны, так и с 3' стороны лишенного основания звена.
Предпочтительно, чтобы нуклеиновая кислота расщеплялась у лишенного основания звена ферментом, который ведет расщепление у лишенного основания звена нуклеиновой кислоты.
Далее, предпочтительно, чтобы ферментом была АР эндонуклеаза, особенно АР эндонуклеаза IV, которая ведет расщепление с 5' стороны лишенного основания звена и создает свободный 3'-ОН конец.
В том случае, когда удлиненный праймер расщепляется 3'-эндонуклеазой, расщепление удлиненного праймера таким ферментом зависит от наличия в удлиненном праймере модифицированного основания и расщепляется фосфодиэфирная связь с 3' стороны (то есть в нисходящем направлении) от места включения модифицированного основания. В противоположность действию гликозилазы и расщеплению у апуринового звена для расщепления удлиненного праймера 3'-эндонуклеазой не нужно удаления модифицированного основания и создания лишенного основания звена. Расщепление 3'-эндонуклеазой зависит от наличия в удлиненном праймере модифицированного основания и распознавания указанного модифицированного основания ферментом. Расщепление обычно происходит по второй фосфодиэфирной связи с 3' стороны модифицированного основания/нуклеотида. Результатом этого события расщепления является формирование в нити ДНК однонитевого разрыва с образованием 3'-ОН и 5'-фосфорильной групп. Присутствующая в реакции ДНК-полимераза может затем удлинить нить от свободной 3'-ОН группы. Как указывалось выше, 3'-эндонуклеазой может быть эндонуклеаза V из Escherichia coli. Эндонуклеаза V распознает в ДНК несколько модифицированных оснований, в том числе остатки урацила, гипоксантина (инозина) и мочевины. Кроме расщепления ДНК с модифицированными основаниями, эндонукпеаза V может также расщеплять ДНК, содержащую лишенные оснований звенья. Таким образом, при определенных условиях эндонуклеазу V можно использовать для расщепления удлиненного праймера в комбинации с действием ДНК-гликозилазы, создающей в ДНК лишенное основания звено.
В одном из вариантов осуществления модифицированный нуклеотид-предшественник частично замещает один из нормальных нуклеотидов-предшественников.
Например, удлинение праймера на матрице при использовании нуклеотидов-предшественников dATP, dCTP, dGTP и dTTP в добавление к модифицированному нуклеотиду-предшественнику dUTP приводит к образованию вновь синтезированной ДНК, комплементарной матрице, в которой тимин заменен случайным образом (статистически) урацилом. В ходе процесса синтеза ДНК урацил включается во вновь синтезируемую нить ДНК в положениях, комплементарных остаткам аденина в матричной нити ДНК. Поэтому границы направленных в нисходящую сторону (downstream) замещенных фрагментов определяются положениями случайного включения dUMP (дезоксирибоуридинмонофосфата) во вновь синтезированную ДНК. В результате создаются замещенные фрагменты различных размеров вследствие пермутаций - включения dUTP вместо dTTP в комплементарную нить напротив остатков аденина в матричной нити нуклеиновой кислоты.
Подобный подход с заменой dGTP на dITP или dGTP на 8-OH-dGTP приводит к замещению всех или части гуанинов соответственно гипоксантином или 8-ОН-гуанином во вновь синтезированной ДНК, комплементарной матрице, во всех положениях, комплементарных остатку цитозина в матричной нити ДНК. Полная или частичная замена одного или нескольких нормальных предшественников ДНК одним или несколькими обрывающими дидезоксинуклеотидами (это нуклеотид, который, будучи включенным в цепь, запрещает дальнейшее удлинение праймера на матрице) может быть использована при желании для остановки удлинения праймера ДНК-полимеразой. Частичная замена одного или нескольких нормальных предшественников ДНК одним или несколькими обрывающими дидезоксинуклеотидами обрывает удлинение праймера во многих различных положениях на матричной нити и создает оборванные праймеры со многими различными длинами. Полная замена одного или нескольких нормальных предшественников ДНК одним или несколькими обрывающими дидезоксинуклеотидами обрывает удлинение праймера в специфических позициях на матричной нити и создает оборванные праймеры специфической длины. В GMA можно использовать несколько модифицированных нуклеотидов-предшественников с одной или несколькими ДНК-гликозилазами. Могут быть использованы две ДНК-гликозилазы, одна из которых высвобождает модифицированное основание из праймера, а другая высвобождает модифицированное основание, если оно включено во вновь синтезированную ДНК.
Этапы (i) и (ii) или (i)-(iii), соответствующие способу настоящего изобретения, могут продолжаться циклически, пока один из реагентов не станет лимитирующим.
Способ настоящего изобретения может осуществляться при изотермических условиях. Поэтому, если способ изотермический, циклического изменения температуры не требуется.
Способ настоящего изобретения является единственной изотермической реакцией амплификации, способной амплифицировать несколько вновь синтезированных и дискретных сегментов ДНК в реакции удлинения праймера с использованием единственного праймера.
Необходимо понимать, что способ настоящего изобретения может приводить к накоплению замещенных однонитевых направленных вниз фрагментов нуклеиновой кислоты, определяемых положениями модифицированных оснований в комплементарной нити нуклеиновой кислоты.
Поэтому способ настоящего изобретения предусматривает путь создания многих копий дискретных однонитевых праймеров в нисходящем направлении от инициирующего праймера. Это привносит исключительную специфичность для целей обнаружения, поскольку дискретные направленные вниз праймеры могут создаваться только в том случае, когда присутствует целевая матричная нуклеиновая кислота. Поэтому в том, что касается специфичности, GMA для диагностики ДНК является существенным усовершенствованием по сравнению со всеми ранее описанными способами амплификации.
В высшей степени желательна амплификация многих сегментов ДНК с единственного матричного образца и это постоянно является ограничением для амплификационных технологий. Это ограничение в основном основано на том факте, что в существующих технологиях используется экспоненциальная амплификация и/или они являются более громоздкими и создают двунитевой продукт или же однонитевой продукт большого размера.
Способ по настоящему изобретению обладает дополнительными преимуществами перед существующими способами множественной амплификации сегментов ДНК.
Способ настоящего изобретения может быть использован для создания многочисленных копий дискретных однонитевых праймеров в нисходящем направлении от инициирующего праймера нуклеиновой кислоты.
Замещенные направленные вниз фрагменты могут быть удлинены во вторичной реакции.
Кроме того, замещенные направленные вниз фрагменты могут быть удлинены на вторичной матричной нуклеиновой кислоте.
Множественные вторичные матрицы могут также быть иммобилизованы на ДНК-чипе (микроячеечный планшет для микроколичественного обнаружения комплементарности тестируемых ДНК к стандарту).
Эти аспекты настоящего изобретения будут дополнительно описаны далее.
Способ по настоящему изобретению может быть применен в детекционной диагностике. Так, например, способ может быть применен для обнаружения патогенов.
Способ настоящего изобретения может быть также применен для обнаружения наличия или отсутствия мутаций и для обнаружения полиморфизмов.
Следует отметить, что способ настоящего изобретения может быть применен для количественного определения уровня содержания нуклеиновой кислоты в образце.
В способе GMA по настоящему изобретению измерения могут быть произведены качественно и количественно различными способами. Следовательно, характеристики и количество ИП (инициирующего праймера) и/или место его комплементарной гибридизации на матричной нуклеиновой кислоте могут быть установлены по способности ИП служить затравкой реакции GMA на этой матрице. При необходимости разрешение, достигаемое согласно настоящему изобретению, может быть таким высоким, чтобы определить различия в одном нуклеотиде между инициирующими праймерами и/или матрицей или целевой нуклеиновой кислотой, и это разрешение определяется успешностью затравки реакции GMA. Поскольку ИП могут быть получены прямо или опосредованно из существующей в природе нуклеиновой кислоты, a GMA обеспечивает получение качественных и количественных характеристик ИП, поэтому GMA может быть использована для получения качественных и количественных характеристик нуклеиновых кислот. Это в высокой степени востребовано в областях обнаружения, диагностики и количественного определения нуклеиновых кислот. Это включает, например, обнаружение и количественное определение патогенных микроорганизмов, как некоторые бактерии и вирусы, обнаружение их вариантов, обнаружение мутаций, вызывающих заболевания человека, обнаружение однонуклеотидных полиморфизмов и количественное определение или титрование специфических типов мРНК в образцах тканей.
В области количественного определения способ по настоящему изобретению имеет существенные преимущества перед существующими технологиями. Так как кинетика GMA является линейной, реакцию GMA легче обнаруживать и измерять (количественно характеризовать) по сравнению с существующими технологиями амплификации с экспоненциальной кинетикой.
Способ по настоящему изобретению имеет также существенные преимущества перед существующими технологиями в области контроля загрязнений. Причина этого состоит в том, что в отличие от существующих способов, GMA в своем базовом формате не синтезирует новых матриц для последующего использования и кинетика процесса линейна.
Способ по настоящему изобретению может быть также применен для амплификации сигнала от любой нуклеиновой кислоты, которая может функционировать как праймер или как матрица.
Способ по настоящему изобретению имеет существенные преимущества перед известными технологиями в том, что он обеспечивает амплификацию сигнала от инициирующего праймера с использованием одиночной линейной матрицы. GMA не вставляет инициирующий праймер в амплифицированные замещенные направленные вниз фрагменты. Это придает то преимущество, что замещенные направленные вниз фрагменты не имеют 5'-«хвоста», который всегда является инициирующим праймером. Более того, это означает, что замещенные направленные вниз фрагменты могут быть удлинены во вторичной реакции с получением комплементарных замещенных направленных вниз фрагментов, не имеющих последовательностей, комплементарных инициирующему праймеру.
Способ по настоящему изобретению уникален в том, что он может быть осуществлен в одной реакционной емкости, причем удлинение ИП на матрице создает и амплифицирует новые праймеры, которые отличаются от ИП и которые могут затем служить как ИП на той же матрице, с которой они были получены, или на иной матрице.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1 представляет собой последовательную диаграмму одного из вариантов осуществления способа настоящего изобретения, как оно описано среди прочего в Примере 1.
Фиг.2 представляет собой последовательную диаграмму другого варианта осуществления способа настоящего изобретения, как оно описано в Примере 4.
Один из вариантов осуществления настоящего изобретения иллюстрирован на Фиг.1 следующим образом:
Событие 1. Праймер связывается с комплементарной последовательностью на матрице;
Событие 2. После связывания 3'-ОН конец праймера удлиняется ДНК-полимеразой. Это осуществляется полимеризацией нуклеотидов-предшественников на 3'-ОН конце праймера. Нуклеотиды-предшественники (dATP, dCTP, dGTP и/или dTTP) включаются в удлиняющийся праймер вместе с модифицированным нукпеотидом-предшественником в соответствии с нукпеотидной последовательностью матрицы. Модифицированный нуклеотид-предшественник обычно замещает (либо полностью, либо частично) один из нормальных нуклеотидов-предшественников. Вновь синтезированная нить ДНК комплементарна исходной матрице и называется здесь комплементарной матричной нитью;
Событие 3. После того как модифицированный нуклеотид-предшественник включился во вновь синтезированную ДНК, эта ДНК теперь содержит модифицированное основание, являющееся субстратом для специфической ДНК-гликозилазы. Вследствие этого каждый раз, как модифицированное основание появляется во вновь синтезированной ДНК, оно высвобождается из ДНК благодаря расщеплению N-гликозидной связи, которая соединяет это основание с остатком дезоксирибозы в ДНК. В результате это приводит к созданию лишенного основания звена, которое по существу представляет собой остаток дезоксирибозы, соединенный с фланкирующими участками ДНК фосфодиэфирными связями на проксимальной и дистальной сторонах (то есть с 5' и 3' стороны остатка дезоксирибозы, причем 5'-связь ближе всего к исходному праймеру);
Событие 4. Лишенное основания звено представляет собой, например, субстрат для фермента АР эндонуклеазы (АРЕ). Следовательно, каждый раз, как возникает лишенное основания звено, в этом месте АРЕ производит расщепление. Этот фермент расщепляет фосфодиэфирную связь с 5' стороны остатка дезоксирибозы, создавая свободный 3'-ОН конец направленного вверх сегмента ДНК, и остаток дезоксирибозы, присоединенный к 5'-окончанию направленного вниз сегмента.
Событие 5. Присутствующая в реакционной смеси ДНК-полимераза синтезирует новую ДНК на этом вновь созданном 3'-ОН конце направленного вверх фрагмента каждый раз, как он возникает, и, действуя таким образом, замещает ДНК как одну нить в нисходящем направлении полимеризации. Следствием этого является включение во вновь синтезируемую комплементарную матричную нить новых нуклеотидов-предшественников, в том числе модифицированных нуклеотидов-предшественников, и появление нового модифицированного основания в каждой позиции вновь синтезированной нити, расположенной напротив комплементарного ему основания в матричной нуклеиновой кислоте.
Таким образом, этапы реакции с (i) no (iii) по настоящему изобретению осуществляются циклически до тех пор, пока один из реагентов не становится лимитирующим.
Каждый свободный 3'-ОН конец, возникший в цикле реакции, удлиняется один раз в каждом последующем цикле реакции, что сопровождается замещением направленных вниз сегментов ДНК. Поскольку реакция непрерывна, полезный результат состоит в повторяющемся синтезе новой ДНК на каждом возникшем 3'-ОН конце и накоплении замещенной направленной вниз ДНК в виде дискретных однонитевых фрагментов с дискретными размерами, которые здесь названы замещенными направленными вниз фрагментами, или замещенных фрагментов, границы которых определены местоположениями модифицированных оснований в комплементарной нити и/или 3'-концом, возникшим вследствие обрыва реакции синтеза ДНК-полимеразой.
Праймеры, которые были удлинены и расщеплены, могут быть немедленно заново удлинены полимеразой путем включения нуклеотидов-предшественников, в том числе модифицированных нуклеотидов-предшественников. Поскольку нормальные нуклеотиды-предшественники, модифицированные нуклеотиды-предшественники, полимераза, гликозилаза и расщепляющие агенты все одновременно присутствуют в одной и той же реакционной смеси, непрерывные циклы удлинения и расщепления дают в результате амплификацию множественных копий замещенных направленных вниз фрагментов.
Если модифицированным нукпеотидом-предшественником является dUTP, тогда модифицированным основанием является урацил и специфической ДНК-гликозилазой, если таковая используется, является урацил-ДНК-гликозилаза. Следовательно, границы замещенных направленных вниз фрагментов обозначены или определены положениями урацила в комплементарной нити и, следовательно, положениями адениновых оснований в матричной нуклеиновой кислоте, так как урацил с аденином образуют нормальную пару оснований Уотсона-Крика.
Таким образом, вкратце, праймер связывается с матрицей и удлиняется ДНК-полимеразой. Дезоксирибонукпеозидтрифосфаты dATP, dCTP, dGTP и dUTP включаются в удлиняемый праймер. Затем урацил-ДНК-гликозилаза выщепляет урацилы во вновь синтезированной нити, и фермент АР эндонуклеаза производит расщепление у возникших лишенных основания звеньев.
Альтернативно, 3'-эндонуклеаза распознает урацил во вновь синтезированной нити и расщепляет нить в месте второй фосфодиэфирной связи в положении 3' относительно остатка урацила.
После этого ДНК-полимераза начинает синтезировать новую ДНК на новых возникших 3'-ОН окончаниях каждый раз, как они создаются, и замещает 3' или нисходящую ДНК в ходе полимеризации. Результатом этого вновь является включение дополнительных урацилов во вновь синтезируемую ДНК, которые затем выщепляются или распознаются, ДНК расщепляется и на новых 3'-ОН окончаниях инициируется полимеризация.
Амплификацию GMA можно проводить либо в мезофильных, либо в термофильных условиях. При мезофильных условиях (температура обычно между 25 и 42°С, типичная температура 37°С) можно использовать ДНК-полимеразу, такую как фрагмент Кленова ехо- ДНК-полимеразы Е.coli, тогда как в термофильных условиях (типичная температура от 50 до 80°С, хотя она может быть и выше) можно использовать термостабильные ДНК-полимеразы, как, например, заменяющую нить ДНК-полимеразу из Thermus aquaticus (фрагмент Stoffel). Полимеразы обоих типов можно добавлять в реакционную смесь вместе или поочередно. Если требуется высокая процессивность, так что праймер удлиняется до значительной длины, прежде чем полимераза отделится от ДНК, можно использовать высокопроцессивную полимеразу. Наоборот, если требуется низкая процессивность, так что праймер удлиняется до малой длины, прежде чем полимераза отделится от ДНК, можно использовать полимеразу с низкой процессивностью. Если в качестве матрицы используется РНК, для реакции GMA можно использовать обратную транскриптазу с замещающей нить активностью.
В самом простом случае для инициирования реакции GMA на заданной матрице или целевой нуклеиновой кислоте в качестве ИП вносят искусственный или синтетический праймер. ИП выбирают так, чтобы он гибридизировался со специфической целевой последовательностью в матрице. После гибридизации ИП происходит инициация GMA и удлиненный ИП повторяющимся образом удлиняется в циклической реакции, в результате чего амплифицируются замещенные направленные вниз фрагменты ДНК. Эти замещенные фрагменты можно охарактеризовать качественно и количественно с помощью многих различных приемов в соответствии с опубликованными методиками.
Прямое обнаружение замещенных фрагментов можно осуществить различными способами, например они могут быть подходящим образом помечены.
Введение метки в замещенные фрагменты может быть осуществлено многими способами, включая добавление к фрагментам в ходе или после синтеза радиоактивного, флуоресцирующего или различаемого каким-либо способом лиганда. Использование в любой из реакций удлинения меченого нукпеотида-предшественника способствует обнаружению этих фрагментов. Методы прямого окрашивания ДНК, такие как окрашивание серебром или бромистым этидием, облегчает их обнаружение после разделения по размерам на основе электрофоретической подвижности. Для идентификации фрагментов можно использовать гибридизацию с ними комплементарных или зондовых нуклеиновых кислот, и такие комплементарные или зондовые нуклеиновые кислоты могут быть иммобилизованы и непосредственно гибридизированы с замещенными фрагментами. В этой связи очень полезны устройства для макроанализа ДНК (DNA macroarrays), устройства для микроанализа ДНК (DNA microarrays) и ДНК-чипы (DNA chips). В качестве альтернативы замещенные фрагменты могут служить несущими молекулами для гибридизации, так что одна зондовая нуклеиновая кислота, которая может быть иммобилизована, гибридизируется с частью замещенного фрагмента, а вторая зондовая или репортерная нуклеиновая кислота гибридизируется с оставшейся частью замещенного фрагмента. В этой связи также очень полезны устройства для макроанализа ДНК и устройства для микроанализа ДНК.
Комплементарные или зондовые нуклеиновые кислоты могут быть подходящим образом помечены любым из многих прямых или непрямых методов мечения, например методами мечения флуоресцентным красителем с сопряжением репортера и гасителя. Поскольку замещенные фрагменты являются однонитевыми, после гибридизации с комплементарными молекулами будут образовываться двунитевые нуклеиновые кислоты, которые можно обнаруживать с помощью специфических зондов для двунитевых нуклеиновых кислот, таких как реагент SYBR зеленый. Этого можно достигнуть в реакции GMA по настоящему изобретению, вводя ДНК, комплементарную замещенным фрагментам, одновременно с реагентом SYBR зеленым, который специфически связывается с двунитевой ДНК.
Нуклеотидную последовательность замещенных фрагментов и, в частности, их 3'-конца можно определить по их способности функционировать в роли инициирующих праймеров в последующей или той же самой реакции GMA. Весьма важно, что такое определение основано на способности этих фрагментов и в особенности их 3'-концов, гибридизироваться с выбранной комплементарной последовательностью матрицы при выбранных условиях и выполнять роль ИП во вторичной реакции GMA. Следует отметить, что существует много возможностей выбора комплементарных последовательностей на матрице и самих матричных молекул. Тем не менее, способность замещенного фрагмента по праву функционировать в качестве ИП в реакции GMA является мерой его гибридизации или отсутствия его гибридизации с выбранной целевой последовательностью и поэтому это позволяет определить природу последовательности части или всего замещенного фрагмента.
Очень выгодно обнаруживать замещенный фрагмент на основании перспективы специфичности, так как его создание зависит от (а) успешной гибридизации ИП с целевой матрицей и (b) инициирования реакции GMA на правильной матрице. Таким образом, обнаружение ожидаемого замещенного фрагмента представляет собой доказательство того, что ИП гибридизировался с правильным местом на правильной матрице.
Идентичность нуклеотидной последовательности замещенного фрагмента можно определить с помощью различных приемов, включая гибридизацию, измерение массы, а также определение его способности быть пришитым непосредственно к нуклеиновой кислоте или служить комплементарной нитью, необходимой для пришивки (лигирования) одной или нескольких молекул нуклеиновой кислоты. Например, он может быть обнаружен и охарактеризован путем оценки его способности служить несущей молекулой для гибридизации для сшивания 5'- и 3'-концов линейной зондовой ДНК с образованием кольца. Гибридизированный замещенный фрагмент, или дополнительный праймер, может затем служить ИП для реакции GMA на новой кольцевой матрице, что приведет к амплификации ДНК по механизму репликации вращающегося кольца (rolling circle replication).
Необходимо также отметить, что, кроме действия в роли ИП, замещенный направленный вниз фрагмент может также служить матрицей в последующей реакции GMA.
Существует много методов, которыми может быть создан ИП. Во всех случаях внесенный или созданный ИП должен иметь свободный 3'-ОН конец, чтобы он мог служить затравкой для последующего этапа полимеризации ДНК.
Искусственный синтез ИП обеспечивает много возможностей в синтезе, конструировании и модификации ИП. Ранее были описаны многие различные модификации искусственно синтезированных праймеров. Они включают модификации основания, сахара и фосфодиэфирной связи, в том числе такие модификации, когда в праймер включались такие основания-субстраты для гликозилазы, как урацил, гипоксантин и 8-ОН-гуанин.
Обычно стандартный или модифицированный ИП синтезируют таким образом, чтобы он специфически соответствовал комплементарной ему последовательности на матричной нуклеиновой кислоте. Хорошо известно, что комплементарность между основаниями 3'-конца потенциально способного к удлинению праймера и матрицы является одним из ключевых параметров, который определяет, будет ли ИП удлинен на данной матрице. ИП, полностью комплементарный участку матрицы, может быть удлинен ДНК-полимеразой, тогда как ИП, который комплементарен всему участку матрицы, кроме основания на его 3'-конце, не удлиняется при строгих условиях. Таким образом, удлинение ИП на матрице может быть использовано, чтобы различить близкородственные ИП, отличающиеся всего лишь одним основанием. Подобным же образом следует отметить, что удлинение ИП на матрице может быть использовано, чтобы различить близкородственные матрицы, отличающиеся всего лишь одним основанием. Этот подход особенно важен в области генетики человека, где он позволяет обнаруживать мутации и полиморфизмы, такие как однонуклеотидные полиморфизмы (single nucleotide polymorfisms, SNPs).
Хорошо известно, что строгость условий гибридизации ИП с матрицей может сильно меняться. Условия низкой строгости позволяют осуществить гибридизацию с низкой специфичностью между молекулами ДНК. Так, в условиях низкой строгости могут гибридизироваться друг с другом частично комплементарные молекулы ДНК. Поэтому при таких условиях с матрицей может гибридизироваться частично комплементарный ИП. При таких условиях одиночный ИП может гибридизироваться и может быть удлинен на одном или нескольких из частично комплементарных мест на матричной нуклеиновой кислоте. Когда строгость условий так низка, что инициирование осуществляется во многих местах, такой процесс называется случайным инициированием (хотя инициирование не вполне случайно, так как обычно все же требуется существенное соответствие между пятью наиболее близкими к 3'-концу праймера основаниями и матрицей). При увеличении строгости условий гибридизация становится все более специфической и могут быть легко найдены такие условия гибридизации, при которых происходит гибридизация только полностью комплементарного праймера, но исключается гибридизация частично комплементарного праймера, даже если он отличается только на один нуклеотид. В ходе гибридизации имеется несколько параметров, которыми может быть изменена строгость условий гибридизации, например температура. Если температура повышается, строгость гибридизации возрастает. Вследствие этого в ферментативных процессах, которые зависят от гибридизации между молекулами ДНК, при более высоких температурах достигается более высокая специфичность. Однако для ферментативного процесса при повышенной температуре обычно необходимы термостабильные ферменты.
ИП может быть создан после расщепления искусственно синтезированного праймера или природной нуклеиновой кислоты таким образом, что образуется новый свободный 3'-ОН конец. Расщепление может быть однонитевым или двунитевым в зависимости от присутствия модифицированного основания в одной нити или в обеих нитях, может зависеть от последовательности нуклеотидов, от наличия области неполного спаривания в двойной спирали или от наличия специфической структуры. Так, праймер может быть создан в ходе направляемого гликозилазой расщепления зонда, содержащего в одной нити или в обеих нитях модифицированное основание, распознаваемое специфической гликозилазой. ИП может быть также создан расщеплением зонда, имеющего неспаренное основание, которое распознается на уровне двойной спирали эндонуклеазой или гликозилазой, специфичными к области неполного спаривания, например, в том случае, когда зонд гибридизирован с матричной нуклеиновой кислотой. Это может быть достигнуто таким конструированием праймера, что в одной или нескольких позициях в гибридизированном сегменте возникает несоответствие оснований у праймера и матрицы, и инкубированием с одной или несколькими из многообразия эндонуклеаз или ДНК-гликозилаз, для которых уже известно, что они в двунитевых нуклеиновых кислотах ведут расщепление у неспаренных оснований. Эти ферменты включают Т7 эндонуклеазу I, ДНК-гликозилазу MutY, специфичную к неспариванию тимина ДНК-гликозилазу и эндонуклеазу V.
ИП может быть также создан путем расщепления комплекса, образующегося при гибридизации перекрывающихся олигонуклеотидных зондов, с помощью такого специфичного к структуре фермента, как Cleavase.
Праймер может быть синтезирован с блокированным 3'-концом, так что удлинение праймера невозможно, пока в ходе расщепления праймера не будет удалена блокирующая группа. Такой праймер здесь назван 3'-блокированным праймером. Блокированный 3'-конец праймера можно получить несколькими методами, в том числе 3'-фосфорилированием, включением на 3'-конце дидезоксинуклеотида, синтезом праймера с 3'-аминогруппой или 3'-тиольной группой, синтезом праймера с одним или более инвертированными нуклеотидами на 3'-конце или расщеплением у лишенного основания звена ДНК-лиазой.
Таким образом, в одном из вариантов осуществления может быть синтезирован праймер с некомплементарным 3'-концом (то есть некомплементарным к матрице на его 3'-конце), так что удлинение праймера невозможно, пока 3'-конец не освобожден путем расщепления праймера.
Может быть синтезирован праймер с некомплементарным 5'-концом, так что расщепление приводит к диссоциации праймера от матричной нуклеиновой кислоты. Диссоциированный праймер может затем служить ИП в реакции GMA на другой матрице.
Расщепление праймера таким образом, что 3'-блокированный праймер разблокируется, а некомплементарный 3'-конец или некомплементарный 5'-конец высвобождается, можно сделать зависящим от гибридизации праймера со всей или с частью матричной нуклеиновой кислоты, так что после расщепления создается ИП с новым свободным 3'-ОН концом, что позволяет удлинить ИП на той же или другой матрице. В этом случае для такого расщепления праймера, что он может быть удлинен на матрице, необходимо, чтобы праймер был расщеплен в одной или нескольких позициях в гибридизированном сегменте этого праймера. Это можно осуществить конструированием праймера, содержащего модифицированное основание гипоксантин, которое является субстратом для 3-алкилпурин-ДНК-гликозилазы (например, AlkA).
В другом варианте осуществления настоящего изобретения при использовании термостабильных расщепляющих агентов можно расщепить гибридизированный праймер у модифицированного или неспаренного основания таким образом, что, как только проба расщеплена, два или несколько фрагментов становятся термически нестабильными (в смысле устойчивости их гибридизации на матрице) и отделяются от целевой нуклеиновой кислоты, что позволяет гибридизироваться другому праймеру полной длины. Этот осциллирующий процесс амплифицирует сигнал (усиливается создание расщепленного праймера). Расщепленный продукт с 3'-ОН концом может затем функционировать как ИП в последующей или сопряженной реакции GMA.
ИП может быть также создан расщеплением праймера у модифицированного основания, когда такое расщепление зависит от удлинения праймера на матрице. Это позволяет создавать ИП, который меньше по размеру, чем исходный праймер, и может быть охарактеризован различными способами. Например, урацил-ДНК-гликозилаза Е.coli не высвобождает урацил в последней или предпоследней 3'-позиции праймера. Однако, если праймер удлинен на матрице, урацил, который был до этого в последней или предпоследней 3'-позиции праймера, теперь находится далеко от 3'-конца вновь удлиненной нуклеиновой кислоты и поэтому будет теперь высвобожден. Таким образом, создается праймер со свободным 3'-ОН концом, который короче исходного праймера на один или два нуклеотида и поэтому может являться ИП для последующей реакции GMA.
ИП может быть получен из существующей в природе или амплифицированной нуклеиновой кислоты путем полного или частичного ферментативного или химического расщепления агентами, расщепляющими ДНК или РНК, такими как ДНКазы, РНКазы, рестрикционные эндонуклеазы, ДНК-гликозилазы (после превращения нормального основания ДНК в основание-субстрат для гликозилазы или внедрения такого основания в ходе амплификации), АР эндонуклеазы после частичной или полной депуринизации или депиримидинизации ДНК, ферменты, расщепляющие РНК или ДНК в гибридах РНК : ДНК (такие, как РНКаза Н), и ферменты, расщепляющие ДНК в положениях неполного спаривания, возникающих после денатурации и обратной гибридизации гибридных молекул нуклеиновой кислоты (таких, как гибриды РНК : ДНК и гибриды ДНК : ДНК). Ферментативное или химическое расщепление ДНК или РНК может создать 3'-концы, не удлиняемые ДНК-полимеразами. Такие 3'-концы могут быть, вообще говоря, сделаны способными к удлинению путем обработки одним или несколькими ферментами, такими как АР эндонуклеаза IV или Т4 полинуклеотидкиназа, имеющая 3'-фосфатазную активность. Хорошо известно, что расщепляющие агенты могут быть специфичными к двунитевой структуре, однонитевой структуре или к последовательности нуклеотидов. Двунитевую или однонитевую нуклеиновую кислоту можно расщепить соответственно до малых двунитевых или однонитевых фрагментов, используя перед реакцией GMA один или несколько расщепляющих агентов. Это дает способ ограничения размера замещенных фрагментов.
Расщепление или создание однонитевого разрыва в одной нити дуплексной молекулы дает участок нуклеиновой кислоты со свободным 3'-ОН концом, который может функционировать в реакции GMA в качестве ИП на матрице, с которой он гибридизирован, или на иной матрице. Расщепление или создание однонитевого разрыва в одной нити дуплексной молекулы можно осуществить с помощью определенных расщепляющих агентов. В нуклеиновой кислоте можно создать неспецифический однонитевой разрыв с помощью малого количества фермента нуклеазы (такого, как ДНКаза), что приведет к созданию многих различных замещенных направленных вниз фрагментов во многих позициях матрицы нуклеиновой кислоты.
Однонитевые разрывы в ДНК могут быть произведены более специфическими агентами, такими как фермент рестрикции N.BstNB I, который делает разрыв в ДНК с 3'-стороны распознаваемой последовательности GAGTC на 4 нуклеотида ниже. В качестве альтернативы, разрыв в нуклеиновой кислоте может быть произведен ферментом рестрикции (например, Hincll или BsoBI), который разрывает немодифицированную нить наполовину модифицированной двунитевой ДНК в специфическом сайте распознавания, создавая таким образом свободный 3'-ОН конец. РНК можно расщепить неспецифически некоторыми РНКазами, а более специфически - специфическими к нуклеотидной последовательности или структуре РНКазами (такими, как рибозимы). РНКаза Н расщепляет РНК в гибриде РНК : ДНК. Поэтому РНК можно расщепить РНКазой Н после синтеза кДНК (комплементарной ДНК) на матрице РНК с помощью обратной транскриптазы. РНК можно расщепить специфически РНКазой Н после гибридизации молекул олигонуклеотидной ДНК с одной или несколькими последовательностями в РНК.
В частности, добавление олиго / поли (дТ) дает способ сделать двунитевым хвост поли(А) на 3'-конце мРНК (матричной РНК). Добавление РНКазы Н приводит к ферментативному гидролизу хвоста поли(А), что придает каждому виду мРНК уникальный 3'-ОН конец. Этот способ дает путь, в котором каждый вид мРНК в образце может потенциально функционировать как ИП в реакции GMA.
Двунитевую нуклеиновую кислоту можно расщепить таким образом, чтобы высвободить ее свободный 3'-конец, что позволит расщепленной ДНК выполнять роль ИП в реакции GMA. Например, двунитевую ДНК можно денатурировать нагреванием. В качестве альтернативы, двунитевую ДНК можно обработать Т7 (ген 6) экзонуклеазой, которая гидролизует дуплекс ДНК с 5'-конца в ступенчатой непроцессивной реакции. Фермент не действует на однонитевую ДНК и поэтому останавливается, когда больше нет дуплексных участков.
Важно, что специфическое расщепление одной нити дуплексной молекулы в участке неспаренности создает фрагмент нуклеиновой кислоты с 3'-ОН концом, который может функционировать как ИП прямо на той матрице, с которой он гибридизирован, или на другой матрице. Это дает способ идентификации в нуклеиновых кислотах мутаций и полиморфизмов.
В связи с этим настоящее изобретение позволяет исследовать наличие или отсутствие мутации или полиморфизма в специфической позиции в нуклеиновой кислоте (локус-кандидат) по следующему пути: гибридизация 3'-блокированного праймера с целевой нуклеиновой кислотой таким образом, что в локусе-кандидате создается нарушение спаривания. Нарушение спаривания обычно располагается внутри праймера, то есть основание, неспаренное с основанием локуса-кандидата, не является основанием 5'-конечного или 3'-конечного нуклеотида праймера. При расщеплении праймера в положении нарушения спаривания специфической к нарушению спаривания гликозилазой и агентом, выполняющим расщепление у лишенного основания звена, расщепленная с 5'-стороны часть праймера имеет 3'-ОН конец и, следовательно, представляет собой ИП, который может инициировать реакцию GMA на той же самой или альтернативной матрице. Поэтому результирующая GMA является индикатором присутствия или отсутствия мутации в зависимости от того, образуется или нет нарушение спаривания при связывании праймера с матрицей, или наоборот.
Кроме того, заново гибридизированные гибридные молекулы нуклеиновой кислоты можно обработать специфическими к нарушению спаривания ферментами, такими как Т7 эндонуклеаза I, ДНК-гликозилаза MutY, специфичная к неспаренным тиминам ДНК-гликозилаза или эндонуклеаза V. Комбинация генной амплификации методами ПЦР с последующей повторной гибридизацией и расщеплением ДНК в месте неспаренности оснований специфическим к неспаренностям репарирующим ферментом создает свободные 3'-ОН концы, которые после диссоциации с комплементарной нити могут выполнять роль ИП в последующей реакции GMA на другой матрице.
Как альтернатива, удлинение 3'-ОН конца, выполняемое со стороны неспаренности с помощью замещающей нить ДНК-полимеразы в стандартной реакции замещения непосредственно после или в реакции GMA, создает замещенный фрагмент, который может служить ИП в последующей реакции GMA. Поскольку создание ИП зависит от наличия неспаренности, процесс инициирования реакции GMA является индикатором наличия неспаренности. Если при таком подходе использовать метод амплификации полного генома со многими зондами и/или многими матрицами для обнаружения замещенных фрагментов, можно сразу анализировать много продуктов амплификации на наличие неспаренностей. Можно локализовать неспаренности, используя выбранные зонды и/или выбранные матрицы. Если использовать планшеты для параллельного анализа многих проб, то можно применить этот подход для одновременного широкого анализа генома или анализа множества амплифицированных нуклеиновых кислот.
Применение вырожденного ИП в реакции GMA дает путь для осуществления инициирования GMA во многих точках матрицы. Вырожденность ИП может быть очень высокой и, следовательно, можно осуществить случайное инициирование на матрице. В этом случае обычно используют такой ИП, как статистический гексамер или более длинный праймер со статистическим гексамером в качестве его 3'-концевой последовательности, с такой матрицей, как геномная ДНК. С помощью ИП с более низким уровнем вырожденности можно достигнуть намного более специфичного множественного инициирования.
Способ настоящего изобретения может быть применен как новый способ амплификации сигнала, обеспечиваемый реакцией GMA. Удлиненные ИП и замещенные фрагменты, созданные в реакции GMA, имеют свободный 3'-ОН конец и могут функционировать как праймеры, которые могут быть удлинены. При использовании замещающей нить ДНК-полимеразы замещенные фрагменты, создаваемые в реакции GMA на матрице, свободны и могут функционировать как ИП во вторичной реакции GMA, если имеется подходящая матрица. Матрица может быть предоставлена в той же самой реакции или в несопряженной реакции как средство амплификации сигнала. Матрица, предлагаемая для применения в амплификации сигнала, названа здесь матрицей амплификации сигнала или бустерной матрицей. Она особенно важна, когда количество исходной матрицы мало, что часто бывает в том случае, когда геномная ДНК используется в качестве матрицы или целевой нуклеиновой кислоты в исходной реакции GMA. Бустерную матрицу синтезируют так, чтобы она имела внутреннюю нуклеотидную последовательность, комплементарную любому представляющему интерес инициирующему праймеру. Эта последовательность названа здесь «сайтом связывания ИП». Обычно эта последовательность комплементарна удлиненному инициирующему праймеру или замещенному фрагменту, созданному в ходе реакции GMA. Обычно сайт связывания ИП находится на 3'-конце бустерной матрицы (бустерная матрица прочитывается в направлении от 3' к 5' полимеразой, которая ведет полимеризацию и удлиняет ИП в направлении от 5' к 3'. В направлении вверх (5') от сайта связывания ИП на бустерной матрице имеются одно или несколько оснований, которые комплементарны модифицированному основанию, используемому в реакции GMA. Обычно этим основанием является аденин. Это приводит в ходе реакции GMA к включению в удлиняемый праймер в этой позиции модифицированного основания, обычно урацила (U). Следующая за этой позицией в направлении вверх (upstream) последовательность, называемая здесь бустерной последовательностью, не содержит оснований, комплементарных модифицированному основанию, которое было использовано в исходной реакции GMA. Бустерная последовательность может варьироваться по размеру и обычно она длиннее 18 нуклеотидов. Обычно бустерная последовательность, чтобы избежать наличия ложного «самоинициирования», содержит 3'-модификацию, такую как инвертированный нуклеотид. Для предотвращения ложного «самоинициирования» в бустерную последовательность в области, комплементарной ИП, может быть также включено блокирование синтеза ДНК. Когда инициирующий праймер выполняет инициирование GMA по бустерной матрице, он создает комплементарную бустерной последовательности нить, которая здесь названа комплементарной бустерной последовательностью. Полезный эффект этого процесса состоит в том, что комплементарная бустерная последовательность в высокой степени копируется / амплифицируется, поскольку бустерная матрица обычно не является лимитирующей, и каждый созданный ИП может потенциально служить затравкой в реакции GMA для любой неинициированной бустерной матрицы. Комплементарная бустерная последовательность может также служить универсальным репортером для целей обнаружения.
Чтобы достигнуть высокого уровня сигнальной амплификации, за бустерной последовательностью (например, БП №1) могут следовать один или несколько нуклеотидов, комплементарных модифицированному основанию, включенному в реакции GMA. В сою очередь, за этой бустерной последовательностью следует вторая бустерная последовательность (БП №2). Обычно БП №2 идентична БП №1. Когда ИП производит инициирование реакции GMA на бустерной матрице, он создает последовательности, комплементарные БП №1 и БП №2, которые здесь обозначены как комплементарная бустерная последовательность №1 (кБП №1) и комплементарная бустерная последовательность №2 (кБП №2). Последовательности кБП №1 и кБП №2 идентичны и функционируют как ИП, выполняющие инициирование реакции на бустерной матрице по БП №1. Они связываются также с БП №2, но замещаются в каждом цикле реакции GMA. Полезный эффект состоит в том, что кБП №2 может быть амплифицирована в высокой степени, поскольку бустерная матрица обычно не является лимитирующей и каждая последовательность кБП №1 и кБП №2 могут потенциально быть ИП в реакции GMA для любой неинициированной бустерной матрицы.
Существует много возможностей конструирования бустерной матрицы. Если в первичной реакции GMA создается кБП №1, тогда необходима бустерная матрица только с последовательностями БП №1 и БП №2, разделенными нуклеотидом, комплементарным модифицированному нуклеотиду, поскольку инициирующим праймером служит кБП №1. Можно сконструировать бустерную матрицу с несколькими различными сайтами связывания ИП (т.е. позволяя многим ИП с различными последовательностями связываться и выполнять инициирование реакции), за которыми следуют идентичные или различные блоки бустерной последовательности.
Дополнительная возможность обнаружения или мониторинга развития инициированной реакции GMA состоит в мониторинге активности ДНК-полимеразы. Поскольку результатом GMA является постоянное повторное удлинение (то есть полимеризация) фрагментов ДНК на матрице, происходит непрерывное включение дезоксирибонуклеозидмонофосфатов (dNMP) во вновь синтезируемую ДНК и высвобождение пирофосфатной части (PPi) на каждый включенный нуклеозид-монофосфат. Поэтому для косвенного обнаружения или мониторинга активности GMA можно использовать реакцию определения PPi (Nyren P. // Analytical Biochemistry. 1987. Т.167. С.235-238).
В реакции GMA, использующей наряду с модифицированным нукпеотидом-предшественником, таким как dUTP, также нуклеотиды-предшественники dATP, dCTP, dGTP, dTTP, образующиеся замещенные фрагменты имеют различные размеры в соответствии с включением во вновь синтезируемую комплементарную нить dUTP вместо dTTP напротив остатков А в матричной нити. В таком случае подходящим сайтом связывания ИП в бустерной последовательности является последовательность, идентичная участку исходной матрицы, которая расположена с 5'-стороны (по матрице) от того места, где первоначально произвел инициирование ИП. Ожидается, что в последовательности ДНК dUTP и dTTP включается в среднем каждый четвертый нуклеотид, поскольку вероятность ожидания остатка А в каждой данной позиции на матрице составляет 0,25. Поэтому средний ожидаемый размер замещенного фрагмента в реакции GMA, использующей dUTP вместо dTTP, будет составлять три нуклеотида. В то же время, если использовать такое соотношение dUTP к dTTP, когда ДНК-полимераза включает каждый из двух dNTP с вероятностью 0,5, размер создаваемых замещенных фрагментов будет варьировать от минимального размера в 3 нуклеотида до более значительных размеров, где вероятность создания данного замещенного фрагмента уменьшается с увеличением размера этого замещенного фрагмента. Однако, чем больше размер замещенного фрагмента, тем более строгой может быть гибридизация между замещенным фрагментом, действующим как ИП, и сайтом связывания ИП. Поэтому желательно, чтобы сайт связывания ИП на бустерной матрице был идентичен последовательности, расположенной с 5'-стороны от сайта связывания ИП на исходной матричной нуклеиновой кислоте. Например, если замещенный фрагмент выбран таким образом, что у него в 40-нуклеотидном сегменте имеется 10 позиций, в которых может быть включен dUTP или dTTP, нужен сайт связывания ИП на бустерной матрице, комплементарный 40 нуклеотидам замещенной ДНК.
Для обнаружения однонукпеотидных полиморфизмов (SNP) или мутаций ИП, производящий инициирование на матричной нуклеиновой кислоте, может быть помещен с 3'-стороны выбранного положения SNP на матричной нити, так что одна замена созданного замещенного фрагмента имеет 3'-окончание, определяемое сайтом SNP. В таком случае наличие SNP на матричной нуклеиновой кислоте ведет к созданию замещенного фрагмента с уникальным 3'-ОН концом, который не создается, если сайт SNP отсутствует. ИП может быть выбран таким образом, что в реакции GMA с использованием смеси нормального и модифицированного предшественников, таких как dTTP и dUTP, создается замещенный фрагмент такого подходящего размера, чтобы обеспечить его гибридизацию с сайтом связывания ИП на бустерной матрице при строгих условиях. Далее, сайт связывания ИП на бустерной матрице может быть сконструирован таким образом, что инициирование на ней может осуществляться только таким замещенным фрагментом с уникальным 3'-ОН концом, который определен сайтом SNP и статусом этого сайта. Этого можно добиться, конструируя бустерную матрицу таким образом, что ключевой остаток аденина, обеспечивающий осуществление GMA, достаточно близко расположен с 5'-стороны на бустерной матрице, так что 3'-окончание замещенного фрагмента, созданного с 3'-ОН концом, определяемым следующей за сайтом SNP на исходной матрице местом включения dUTP, находится напротив ключевого остатка аденина на бустерной матрице или с 5'-стороны от него. При строгих условиях гибридизации замещенные фрагменты, у которых 3'-ОН конец создается на основании сайта, находящегося в направлении вверх (upstream) от сайта SNP (на нити, комплементарной к той, которая служит матрицей), не будут производить инициирование на бустерной матрице, поскольку они не будут гибридизироваться с сайтом связывания ИП.
Согласно следующему варианту осуществления настоящего изобретения сайт связывания ИП может быть вырожденным, так что он может служить сайтом связывания для многих различных инициирующих праймеров. В реакцию GMA могут быть вовлечены две или более бустерные матрицы, так что кБП, созданная на первой бустерной матрице, может служить инициирующим праймером для второй бустерной матрицы. Это создает дополнительное преимущество, так как вторая бустерная матрица может быть одинаковой для многих различных реакций GMA и может поэтому служить универсальной бустерной последовательностью. Это способствует получению единого рационализованного процесса для обнаружения и амплификации сигнала с помощью одной окончательной бустерной матрицы. Обычно окончательную бустерную матрицу можно сконструировать и синтезировать таким образом, что она служит прямо или опосредованно в качестве репортера для GMA.
ИП в реакции GMA может замещать направленный вниз удлиненный праймер, который служит ИП в последующей реакции GMA. Это имеет важные применения для обнаружения SNP и мутаций. Праймер можно поместить достаточно близко к сайту SNP, так что первый модифицированный предшественник включается в сайт SNP или на расстоянии от него. Поэтому праймер удлиняется на различную длину в зависимости от того, имеется или нет в этом сайте SNP. Желательно создать много копий по-разному удлиненного праймера для последующего получения их характеристик любым из нескольких путей, включая определение их способности функционировать в качестве ИП в последующей реакции GMA на бустерной матрице. Много копий по-разному удлиненного праймера можно получить в процессе циклического изменения температуры, как описано для полимеразной цепной реакции. В качестве альтернативы, по-разному удлиненный праймер может повторно замещаться инициированием реакции GMA в 5'-сторону от этого праймера (то есть продолжая синтез в 3'-сторону по матричной нити). Это достигается с помощью ИП, который гибридизируется с 5'-стороны праймера, локализованного возле (проксимально) сайта SNP, и инициирует реакцию GMA. В каждом цикле направленный вниз праймер будет удлиняться в той же самой реакции, но он будет также в каждом цикле замещаться. Как только свежий праймер замещен, он может гибридизироваться и удлиняться.
Замещенные фрагменты, созданные по матричным нуклеиновым кислотам, и комплементарные бустерные последовательности, созданные по бустерной матрице, могут быть обнаружены (детектированы) анализом с помощью 5'-нуклеазы. Обычно 5'-нуклеазный анализ представляет собой анализ, обнаруживающий конкретную нуклеиновую кислоту по ее способности служить инициирующим праймером для синтеза ДНК по матрице с помощью ДНК-полимеразы с 5-3' экзонуклеазной активностью, что приводит к деструкции зонда, который гибридизирован на матрице в нисходящем направлении. Эта деструкция основана на присутствии 5'-3' экзонуклеазной активности в полимеразе, использованной в реакции. Типичным зондом является олигонуклеотид, содержащий в тесном соседстве и репортерный флуоресцентный краситель, и краситель - гаситель флуоресценции. Когда репортер и гаситель разделены/разъединены друг от друга вследствие деструкции зонда, происходит увеличение интенсивности флуоресценции. Поэтому увеличение флуоресценции указывает на то, что зонд был гибридизирован с матрицей и был деструктирован 5'-3' экзонуклеазной активностью ДНК-полимеразы, как только он удлинил инициирующий праймер на матричной нуклеиновой кислоте.
В варианте этого теста использован подход, в котором зонд является частью матрицы, но комплементарен участку матрицы. Это приводит к тому, что в зонде образуется шпилька за счет спаривания основаниями части матрицы с комплементарным ей участком матричной нуклеиновой кислоты, в результате чего двунитевая шпилька имеет один свободный 5'-конец, а на другом конце имеется петля.
В качестве альтернативы можно использовать самокомплементарный зонд с репортером на одном конце и гасителем на другом конце. В этом случае зонд образует шпильку с петлей вследствие спаривания комплементарных оснований, это приводит репортер в тесный контакт с гасителем, в результате чего флуоресценция гасится. В результате денатурации петельной структуры шпильки в присутствии полностью или частично комплементарного замещенного фрагмента происходит гибридизация между зондом и замещенным фрагментом или кБП. Это расправляет зонд в двунитевую структуру и увеличивает расстояние между репортером и гасителем, что приводит к увеличению интенсивности флуоресценции.
Адаптация GMA для использования с блоком репортер-гаситель предусматривает уникальную бустерную матрицу, которая обеспечивает одновременно обнаружение ИП и сигнальную амплификацию. Необходимо, чтобы бустерная матрица была сконструирована с сайтом связывания ИП, за которым следует самокомплементарная последовательность, которая приводит соединенные репортер и гаситель в тесную близость при образовании двунитевой шпилечной структуры. Сайт связывания ИП и последовательность, образующая шпилечную структуру типа стержня с петлей, не имеют ни одного основания, комплементарного модифицированному основанию в реакции GMA. Сайт связывания ИП находится с 3'-стороны последовательности шпильки и может содержать последовательность, комплементарную инициирующему праймеру и/или последовательности, которая комплементарна кБП. Одно или несколько оснований, комплементарных модифицированному основанию в реакции GMA, располагаются с 5'-стороны шпильки, а за ними следует другая последовательность, комплементарная кБП. Когда инициирующий праймер удлиняется на бустерной матрице с репортером-гасителем, полезный эффект заключается в том, что сайт связывания ИП и секция последовательности шпильки бустерной матрицы линеаризуется, становится двунитевой в ходе реакции GMA с замещением нити и остается двунитевой. Сигнал амплифицируется путем синтеза дальнейших кБП на 5'-конце бустерной матрицы, которая в свою очередь служит инициирующим праймером для любой неинициированной бустерной матрицы в реакции GMA. Когда шпилька линеаризуется и становится частью двунитевой структуры, интенсивность флуоресценции увеличивается и измерение значений интенсивности флуоресценции дает меру количества ИП в реакции.
Альтернативный способ обнаружения сигнала в соответствии с настоящим изобретением включает конструирование бустерной матрицы таким образом, что часть последовательности или вся последовательность замещенного фрагмента самокомплементарна либо внутри самой себя, либо между другими копиями самой себя и образует двунитевую ДНК. Двунитевую ДНК можно затем легко обнаруживать по связыванию специфических для двунитевой структуры зондов, таких как краситель SYBR зеленый.
Бустерная матрица может быть кольцевой. Такая кольцевая бустерная матрица обеспечивает способ побудить ДНК-полимеразу работать непрерывно в формате репликации по механизму вращающегося кольца. Существует много вариантов, которые могут быть использованы с кольцевой бустерной матрицей. В своей простейшей форме кольцевая бустерная матрица может служить матрицей для амплификации по типу вращающегося кольца, где ИП может служить инициатором репликации ДНК по кольцу. В этом случае ДНК непрерывно синтезируется до тех пор, пока полимераза или предшественники ДНК не начинают лимитировать реакцию. Включение такой бустерной матрицы в реакцию GMA требует, чтобы бустерная матрица была сконструирована таким образом, чтобы она не содержала ни одного звена, комплементарного модифицированному нуклеотиду-предшественнику в реакции GMA.
В качестве альтернативы можно сконструировать бустерную матрицу с одним или несколькими сайтами связывания ИП, за которым(и) следует звено, поддерживающее включение модифицированного предшественника в удлиняющийся ИП в ходе реакции GMA. За ним далее следует одна или несколько бустерных последовательностей. Перед бустерными последовательностями и после них может быть одно или более оснований, комплементарных модифицированному основанию в реакции GMA. Комплемент бустерной последовательности, будучи создан, служит ИП для любых неинициированных бустерных матриц. Инициирование последующих бустерных матриц дает дополнительные ИП для дополнительных неинициированных бустерных матриц и реакция продолжается до тех пор, пока все бустерные матрицы не окажутся инициированными, и репликация продолжается, пока какой-нибудь из реагентов не станет лимитирующим. Полезный эффект такого способа состоит в том, что в каждом цикле создается линейная копия всей бустерной матрицы или ее части. Введение второй комплементарной бустерной матрицы с самоинициируемой бустерной последовательностью дает способ создания многих линейных копий бустерной матрицы. Затем линейные копии двух бустерных матриц гибридизируются друг с другом, образуя двунитевую ДНК, которую можно затем обнаруживать несколькими методами, в том числе с помощью связывающихся с ДНК специфических для двунитевой структуры агентов, таких как краситель SYBR зеленый.
Можно также сконструировать единичную бустерную матрицу таким образом, что она продуцирует кБП, которые спариваются основаниями друг с другом, образуя таким образом двунитевую ДНК. Бустерная матрица особенно пригодна для такого применения, поскольку участки самокомплементарности с кольцом ДНК могут быть инициированы без денатурации с помощью ИП, гибридизирующихся с сайтами связывания ИП, которые отсутствуют в области самокомплементарности. Напротив, для полностью двунитевой линейной комплементарной ДНК необходима денатурация, чтобы обеспечить ИП доступ к сайту связывания ИП.
В другом варианте осуществления настоящего изобретения бустерную или вторичную матрицу можно иммобилизовать на твердой подложке, например на микро- или макропланшете или на ДНК-чипе. Много различных бустерных или вторичных матриц можно иммобилизовать на ДНК-чипе, который затем можно использовать, чтобы охарактеризовать много различных нуклеиновых кислот и многие различные реакции GMA в высокорационализованном режиме. В предпочтительном примере осуществления вторичные матрицы прикрепляют их 3'-концами. Это делает недоступной 3'-ОН группу, которая иначе была бы реакционноспособной. Поэтому вторичная матрица не может быть удлинена полимеразой и функционирует только как матрица, что снижает любое фоновое неспецифическое удлинение.
Иной вариант осуществления настоящего изобретения направлен на применение GMA в молекулярных компьютерах на основе ДНК. Сейчас все более признается, что ДНК благодаря присущим ей физическим и химическим свойствам может дать путь компьютерных решений сложных математических задач. Следует признать, что, сконструировав инициирующие праймеры, матрицы и/или бустерные матрицы так, чтобы они действовали как индивидуальные или комбинированные блоки информации, проблемы это и/или решения, можно выполнить расчеты решений математических задач с помощью реакции GMA. Предпочтительно, чтобы ИП и матрицы были синтезированы искусственно. Будет также понятно, что использование GMA в ДНК-компьютерах обеспечивает одновременные операции и параллельные поиски и может обеспечить полный набор потенциальных решений. В соответствии с этим настоящее изобретение предлагает применение способа, как он здесь описан, в вычислениях на основе ДНК и/или как «инструмент» в вычислениях с помощью ДНК, что здесь собирательно названо как компьютерный ДНК-анализ.
Изобретение будет далее иллюстрировано нижеследующими примерами.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
В следующих примерах были использованы различные ферменты. Некоторые из этих ферментов являются коммерческими продуктами, тогда как другие были очищены после сверхэкспрессии в штаммах Е.coli, как описано далее.
Урацил-ДНК-гликозилаза из Thermotoga maritime (TmaUDG)
Открытую рамку считывания для белка TmaUDG амплифицировали из геномной ДНК Т. maritima с помощью ПЦР. Продукт ПЦР вставляли в сверхэкспрессирующий вектор pBAD-TOPO в соответствии с инструкциями поставщика (InVitrogen). Затем компетентные клетки Е.coli, штамм TOP-10 (InVitrogen) трансформировали полученной конструкцией в режиме теплового шока согласно инструкциям поставщика. Клетки, содержавшие конструкцию pBAD-TOPO/TmaUDG, выращивали до оптической плотности 0,6 при 600 нм (1000 мл) и индуцировали сверхэкспрессию арабинозой, добавляемой до конечной концентрации 0,2%. После инкубации при 37°С в течение 4 ч клетки лизировали и проводили очистку слитого белка аффинной хроматографией на иммобилизованных ионах металла, используя смолу ProBond (InVitrogen) в соответствии с инструкциями поставщика. Собирали фракцию объемом 15 мл, содержавшую элюированный белок (по данным электрофореза в полиакриламидном геле в присутствии SDS). Эту фракцию затем дополнительно очищали ионообменной хроматографией на смоле Mono-S, отбирали фракцию с наибольшей концентрацией элюированного белка объемом 0,5 мл и хранили ее с добавкой глицерина до 50% при -20°С до использования.
Эндонуклеаза IV из Thermotoga maritime (TmaEndolV)
Открытую рамку считывания для белка TmaEndolV амплифицировали из геномной ДНК Т. maritime с помощью ПЦР.Продукт ПЦР вставляли в сверхэкспрессирующий вектор pBAD-TOPO в соответствии с инструкциями поставщика (InVitrogen). Затем компетентные клетки E.coli, штамм TOP-10 (InVitrogen) трансформировали полученной конструкцией в режиме теплового шока согласно инструкциям поставщика. Содержавшие конструкцию pBAD-TOPO/TmaEndolV клетки выращивали до оптической плотности 0,6 при 600 нм (1000 мл) и индуцировали сверхэкспрессию арабинозой, добавляемой до конечной концентрации 0,2%. После инкубации при 37°С в течение 4 ч клетки лизировали и проводили очистку слитого белка аффинной хроматографией на иммобилизованных ионах металла, используя смолу ProBond (InVitrogen) в соответствии с инструкциями поставщика. Собирали фракцию объемом 15 мл, содержавшую элюированный белок (по данным электрофореза в полиакриламидном геле в присутствии SDS). Эту фракцию затем дополнительно очищали ионообменной хроматографией на смоле Mono-S, отбирали фракцию с наибольшей концентрацией элюированного белка объемом 0,5 мл и хранили ее с добавкой глицерина до 50% при -20°С до использования.
Эндонуклеаза V из Thermotoga maritima (TmaEndoV)
Открытую рамку считывания для белка TmaEndoV амплифицировали из геномной ДНК Т. maritima с помощью ПЦР. Продукт ПЦР вставляли в сверхэкспрессирующий вектор pBAD-TOPO в соответствии с инструкциями поставщика (InVitrogen). Затем компетентные клетки Е.coli, штамм TOP-10 (InVitrogen) трансформировали полученной конструкцией в режиме теплового шока согласно инструкциям поставщика. Содержавшие конструкцию pBAD-TOPO/TmaEndoV клетки выращивали до оптической плотности 0,5 при 600 нм (1000 мл) и индуцировали сверхэкспрессию арабинозой, добавляемой до конечной концентрации 0,2%. После инкубации при 37°С в течение 8 ч клетки лизировали и проводили очистку слитого белка аффинной хроматографией на иммобилизованных ионах металла, используя смолу ProBond (InVitrogen) в соответствии с инструкциями поставщика. Собирали и объединяли 15 фракций по 1 мл, содержавших элюированный белок (по данным электрофореза в полиакриламидном геле в присутствии SDS). Объединенные фракции затем дополнительно очищали ионообменной хроматографией на смоле Mono-S, отбирали фракцию с наибольшей концентрацией элюированного белка объемом 0,5 мл и хранили ее с добавкой глицерина до 50% при -20°С до использования.
Эндонуклеаза IV из Е.coli (EcoEndolV)
Открытую рамку считывания для белка EcoEndolV амплифицировали из геномной ДНК Е.coli с помощью ПЦР. Продукт ПЦР вставляли в сверхэкспрессирующий вектор pBAD-TOPO в соответствии с инструкциями поставщика (InVitrogen). Затем компетентные клетки Е.coli, штамм TOP-10 (InVitrogen) трансформировали полученной конструкцией в режиме теплового шока согласно инструкциям поставщика. Содержавшие конструкцию pBAD-TOPO/EcoEndolV клетки выращивали до оптической плотности 0,6 при 600 нм (1000 мл) и индуцировали сверхэкспрессию арабинозой, добавляемой до конечной концентрации 0,2%. После инкубации при 37°С в течение 4 ч клетки лизировали и проводили очистку слитого белка аффинной хроматографией на иммобилизованных ионах металла, используя смолу ProBond (InVitrogen) в соответствии с инструкциями поставщика. Собирали фракцию объемом 15 мл, содержавшую элюированный белок (по данным электрофореза в полиакриламидном геле в присутствии SDS). Эту фракцию затем дополнительно очищали ионообменной хроматографией на смоле Mono-S, отбирали фракцию с наибольшей концентрацией элюированного белка объемом 0,5 мл и хранили ее с добавкой глицерина до 50% при -20°С до использования.
UDG-тиоредоксин из Archaeoglobus fulgidus (AfUDG-Thio)
Открытую рамку считывания для белка AfUDG амплифицировали из геномной ДНК A.fulgidus с помощью ПЦР. Продукт ПЦР вставляли в сверхэкспрессирующий вектор pBAD-TOPO-ThioFusion в соответствии с инструкциями поставщика (InVitrogen). Затем компетентные клетки Е.coli, штамм ТОР-10 (InVitrogen) трансформировали полученной конструкцией в режиме теплового шока согласно инструкциям поставщика. Клетки, содержавшие конструкцию pBAD-TOPO-ThioFusion/AfUDG, выращивали до оптической плотности 0,6 при 600 нм (2000 мл) и индуцировали сверхэкспрессию арабинозой, добавляемой до конечной концентрации 0,2%. После инкубации при 37°С в течение 4 ч клетки лизировали и проводили очистку слитого белка аффинной хроматографией на иммобилизованных ионах металла, используя смолу ProBond (InVitrogen) в соответствии с инструкциями поставщика. Отбирали фракцию объемом 15 мл, содержавшую элюированный белок (по данным электрофореза в полиакриламидном геле в присутствии SDS), и хранили ее при 4°С до использования.
Пример 1
Способ по настоящему изобретению был применен для циклического удлинения 25-звенного олигонуклеотида (инициирующий праймер - ИП), который был комплементарен участку 80-звенного олигонуклеотида (матричная нуклеиновая кислота), который служил матрицей в реакции полимеризации. Комплементарная область простиралась от точки, отстоящей на 10 оснований от 3'-окончания 80-звенника, до точки, отстоящей на 35 оснований от 3'-окончания 80-звенника. Участок 80-звенного олигонуклеотида, примыкающий с 5'-стороны к области комплементарности ИП (сайту связывания ИП), был сконструирован таким образом, чтобы он содержал несколько аденинов, что при циклическом удлинении 25-звенного олигонуклеотида приведет вследствие реакции GMA к продуцированию фрагментов ДНК дискретных размеров. Олигонуклеотид с 80 звеньями был сконструирован также таким образом, что между аденинами (А) расположено определенное число гуанинов (G). Это позволяет пометить получающиеся в ходе реакции замещенные направленные вниз фрагменты ДНК включением [α32P]-dCTP. Диаграмма последовательности этапов способа приведена на Фиг.1. Оба олигонуклеотида были синтезированы искусственно и очищены вырезанием зон из полиакриламидного геля после электрофореза. Задачей было определить, гибридизируется ли 25-звенник с 80-звенником и может ли он быть циклически/повторно удлинен способом настоящего изобретения, давая множественные копии меченых однонитевых фрагментов ДНК (замещенных фрагментов).
Перед добавлением фермента реакционные смеси содержали в общем объеме 17 мкл: 100 фемтомолей (фмолей) каждого олигонуклеотида, 10 мМ трис-HCl (рН 7,5), 5 мМ MgCl2, 7 мМ дитиотреитол, 0,2 мМ каждого из dATP, dGTP и dUTP, 0,02 мМ dCTP и 0,2 мкл [α32P]-dCTP (10 мКи/мл, ˜800 Ки/моль). Эту смесь покрывали слоем минерального масла и прогревали при 95°С в течение 2 мин. Затем реакционную смесь охлаждали до 37°С и выдерживали при этой температуре. Затем добавляли последовательно 5 единиц активности фрагмента Кленова (3'→5' ехо-), 1 единицу урацил-ДНК-гликозилазы Е.coli и 2 единицы либо эндонуклеазы IV E.coli, либо (в 2 мкл) эндонуклеазы IV Thermotoga maritima. Конечный объем реакционной смеси становился равным 20 мкл. В контрольные реакционные смеси, не содержавшие эндонуклеазы IV, и в контрольные реакционные смеси, не содержавшие ни эндонуклеазы IV, ни урацил-ДНК-гликозилазы, добавляли воду до конечного объема 20 мкл. Из реакционных смесей с 30-минутными интервалами после добавления экзонуклеазы IV отбирали пробы объемом 5 мкл. К пробам добавляли NaOH до конечной концентрации 50 мМ и прогревали их при 95°С в течение 15 мин. Затем добавляли равный объем раствора с формамидом и красителем для нанесения (98% формамид, 0,025% бромфенол синий, 0,025% ксиленцианол). После этого пробы наносили на 20% денатурирующий (7 М мочевина) полиакриламидный гель и проводили электрофорез для определения размеров фрагментов ДНК. После электрофореза гель экспонировали с фосфоресцирующим экраном и изображения (зон) последовательно сканировали прибором Storm 860 (товарный знак).
Анализ полученных изображений показал, что реакционные смеси с эндонуклеазой IV содержали значительное количество меченых фрагментов ДНК различного размера короче 70 нуклеотидов и их количество увеличивалось со временем. Эти фрагменты не появлялись в контрольных реакционных смесях, не содержавших эндонуклеазы IV, или в контрольных реакционных смесях, не содержавших ни эндонуклеазы IV, ни урацил-ДНК-гликозилазы.
Было отмечено, что эндонуклеаза IV Thermatoga maritima содержит присущую ей или примесную 3'→5' экзонуклеазную активность. Это приводит к неспецифической фоновой амплификации в ходе реакции GMA. Чтобы избавиться от этой активности и от неспецифической амплификации, фермент эндонуклеазу IV прогревали перед использованием в реакции GMA. Рабочий раствор фермента эндонуклеазы IV нагревали до 90°С в течение 10 мин, затем помещали в лед на ≥5 мин и после этого центрифугировали в течение ≥5 мин. Было обнаружено, что препараты эндонуклеазы IV E.coli также имели такую активность и для этих препаратов также была необходима инактивация экзонуклеазной активности перед их использованием.
Пример 2
Способ настоящего изобретения был применен для циклического удлинения 41-звенного олигонуклеотида (инициирующий праймер - ИП), который был самокомплементарен на своем 3'-конце, т.е. был палиндромным, и поэтому служил матрицей самому себе для реакции GMA. Комплементарный участок простирался на 20 нуклеотидов от 3'-конца 41-звенника и этот участок олигонуклеотида по существу функционировал как ИП. Участок 41-звенника с 5'-стороны от этой комплементарной области ИП (сайта связывания ИП) конструировали таким образом, чтобы он содержал следом за сайтом связывания ИП один аденин, что приводит после реакции GMA к продукции дискретного фрагмента ДНК длиной 20 нуклеотидов. 41-Звенный олигонуклеотид конструировали также таким образом, чтобы он содержал несколько гуанинов (G). Это позволило пометить получающиеся в ходе реакции замещенные направленные вниз фрагменты ДНК (20-звенные) включением [α32P]-dCTP. Задачей было определить, гибридизируется ли 41-звенник сам с собой и может ли он быть циклически/повторно удлинен способом настоящего изобретения, давая множественные копии меченого однонитевого 20-звенного фрагмента ДНК (замещенного фрагмента).
Перед добавлением фермента реакционные смеси содержали в общем объеме 17 мкл: 100 фмоль 41-звенного олигонуклеотида, 10 мМ трис-HCl (рН 7,5), 5 мМ MgCl2, 7 мМ дитиотреитол, 0,2 мМ каждого из dATP, dGTP и dUTP, 0,02 мМ dCTP и 0,2 мкл [α32P]-dCTP (10 мКи/мл, ˜800 Ки/моль). Эту смесь покрывали слоем минерального масла и прогревали при 95°С в течение 2 мин. Затем реакционную смесь охлаждали до 37°С и выдерживали при этой температуре. После этого добавляли последовательно 5 единиц активности фрагмента Кленова (3'→5' ехо-), 1 единицу урацил-ДНК-гликозилазы E.coli и 2 единицы либо эндонуклеазы IV E.coli, либо (в 2 мкл) эндонуклеазы IV Thermotoga maritima. Конечный объем реакционной смеси становился равным 20 мкл. В контрольные реакционные смеси, не содержавшие эндонуклеазы IV, и в контрольные реакционные смеси, не содержавшие ни эндонуклеазы IV, ни урацил-ДНК-гликозилазы, добавляли воду до конечного объема 20 мкл. Реакцию проводили в течение 30 мин или дольше. Для остановки реакции добавляли EDTA (этилендиаминтетраацетат) до конечной концентрации 10 мМ. Затем добавляли равный объем раствора с формамидом и красителем для нанесения (98% формамид, 0,025% бромфенол синий, 0,025% ксиленцианол). После этого пробы наносили на 20% денатурирующий (7 М мочевина) полиакриламидный гель и проводили электрофорез для определения размеров фрагментов ДНК. После электрофореза гель экспонировали с фосфоресцирующим экраном и изображения (зон) последовательно сканировали прибором Storm 860 (товарный знак).
Анализ полученных изображений показал, что в каждой опытной реакционной смеси, но не в контрольных реакционных смесях, продуцировалось значительное количество меченого и амплифицированного 20-звенного олигонуклеотида.
Пример 3
Способ настоящего изобретения применяли, как описано в Примере 2, со следующими модификациями:
Перед добавлением фермента реакционные смеси содержали в общем объеме 20 мкл: 100 фмоль 41-звенного палиндромного олигонуклеотида, 10 мМ трис-HCl (рН 8,3), 10 мМ KCl, 3 мМ MgCl2, 0,2 мМ каждого из dATP, dUTP и dGTP, 0,02 мМ dCTP и [α32P]-dCTP. Эту смесь покрывали слоем минерального масла и прогревали при 95°С в течение 2 мин. Затем реакционную смесь охлаждали до 60°С и выдерживали при этой температуре. После этого добавляли последовательно 10 единиц фрагмента Stoffel ДНК-полимеразы AmpliTaq (товарный знак), 2 единицы (в 2 мкл) UDG Thermotoga maritima и 4 единицы эндонуклеазы IV Е.coli (или воду в случае контрольной реакционной смеси без эндонуклеазы IV). Конечный объем реакционной смеси был равен 25 мкл. Из реакционных смесей с 30-минутными интервалами после добавления эндонукпеазы IV отбирали пробы объемом 5 мкл. К пробам добавляли EDTA до конечной концентрации 10 мМ. Затем добавляли равный объем раствора с формамидом и красителем для нанесения (98% формамид, 0,025% бромфенол синий, 0,025% ксиленцианол). Анализ размеров ДНК в пробах проводили, как в Примере 1.
Анализ полученных изображений показал, что в реакционных смесях с эндонуклеазой IV содержались большие количества меченой ДНК, которая отсутствовала в контрольных реакционных смесях без эндонуклеазы IV. Количество ДНК в опытных реакционных смесях возрастало со временем, достигая пика через 90 мин после добавления эндонуклеазы IV.
Пример 4
Способ настоящего изобретения применяли, как описано в Примере 2, со следующими модификациями:
Перед добавлением фермента реакционные смеси содержали в общем объеме 20 мкл: 100 фмоль 41-звенного олигонуклеотида, 10 мМ трис-HCl (рН 8,3), 10 мМ KCl, 3 мМ MgCl2, 0,2 мМ каждого из dATP, dUTP и dGTP, 0,02 мМ dCTP и 0,2 мкл [α32P]-dCTP (10 мКи/мл, ˜800 Ки/моль). Эту смесь покрывали слоем минерального масла и прогревали при 95°С в течение 2 мин. Затем реакционную смесь охлаждали до 70°С и выдерживали при этой температуре. После этого добавляли последовательно 10 единиц фрагмента Stoffel ДНК-полимеразы AmpliTaq, 2 мкл AfUDG-Thio и 2 мкл EcoEndolV (или воды в случае контрольной реакционной смеси без эндонуклеазы IV). Конечный объем реакционной смеси был равен 25 мкл. Реакцию останавливали через 60 мин добавлением EDTA до конечной концентрации 10 мМ. Затем добавляли равный объем раствора для нанесения с формамидом и красителем (98% формамид, 0,025% бромфенол синий, 0,025% ксиленцианол). Анализ размеров ДНК в пробах проводили, как в Примере 1. После электрофореза гель экспонировали с фосфоресцирующим экраном и изображения зон обнаруживали на визуализующем приборе Storm 860 (товарный знак).
Анализ полученных изображений показал, что в реакционных смесях с эндонуклеазой IV содержались большие количества ожидаемого меченого 20-звенного олигонуклеотида и меньшие количества несколько более коротких фрагментов, которые отсутствовали в контрольных реакционных смесях без эндонуклеазы IV.
Пример 5
Способ настоящего изобретения был применен для обнаружения удлинения праймера на аттомольных количествах 41-звенного палиндромного олигонуклеотида. Расщепление урацил-ДНК-гликозилазой с последующим расщеплением эндонуклеазой IV продукта, образованного при катализируемом ДНК-полимеразой удлинении 41-звенного олигонукпеотида, когда он сам был матрицей, в качестве одного из продуктов давало 20-звенный однонитевой олигонуклеотид (то есть замещенный фрагмент, который далее может действовать как ИП). Этот олигонуклеотид действует как вторичный праймер. В этом примере способ настоящего изобретения позволяет обнаруживать циклическое удлинение 20-звенного олигонуклеотида вследствие повторной реакции образования петли у продукта. 42-звенный олигонуклеотид (бустерная матрица) был сконструирован, как схематически изображено на Фиг.2. Комплементарная последовательность 20-звенного вторичного праймера/замещенного фрагмента повторяется дважды в этом 42-звеннике. Повторяющиеся последовательности разделены (читая в направлении от 5' к 3') последовательностью AT. Этот олигонуклеотид синтезировали и очищали так же, как и 41-звенный палиндромный олигонуклеотид. Этот 42-звенник вводили в реакцию в большом избытке. Как только образовывался 20-звенный вторичный праймер, как описано выше, он мог после этого гибридизироваться на 42-звенном олигонуклеотиде и быть сам по способу настоящего изобретения циклически удлинен, давая в результате большое число копий самого себя. Эти вновь созданные 20-звенные вторичные праймеры могут затем гибридизироваться с другими копиями 42-звенного олигонуклеотида, инициируя новые этапы циклического удлинения, и так далее. Задача состояла в обнаружении количеств 41-мерного палиндромного олигонуклеотида, количество которого в норме слишком мало, чтобы быть обнаруженным, как описано в Примере 2, по продукции обнаружимых количеств 20-звенного вторичного праймера при применении описанного выше способа настоящего изобретения в модификации образования петли продуктом.
Описанные в Примере 2 реакционные смеси выполнялись в 3 повторах со следующими модификациями. Описанный выше 42-звенник вносили в каждую реакционную смесь в количестве 40 нг. В разных наборах реакционных смесей (контрольные - без эндонуклеазы IV и опытные - с эндонуклеазой IV) концентрация 41-звенного палиндромного олигонуклеотида была 1,0 фмоль, 10-2 фмоль или 10-4 фмоль в объеме реакционной смеси. Реакции проводили в течение 2 ч, после чего добавляли NaOH до конечной концентрации 50 мМ. Содержание меченого фрагмента ДНК в реакционных смесях устанавливали, определяя эквивалент в пробах 5 мкл первоначальной реакционной смеси (до добавления NaOH и раствора с формамидом и красителем для нанесения), как описано в Примере 1.
Анализ изображений зон показал, что в каждой опытной реакционной смеси образовывалось обнаружимое количество меченого 20-звенного олигонуклеотида, который не образовывался в контрольных реакционных смесях.
Подобный же результат наблюдался, когда описанный выше опыт повторяли с использованием термостабильных ферментов - фрагмента Stoffel ДНК-полимеразы, UDG Thermatoga maritima и эндонуклеазы IV Thermatoga maritima. Чтобы избежать какой-либо неспецифической амплификации из-за самоинициирования бустерной матрицы, 3'-конец бустерной матрицы обычно содержит блокирующую группу, предотвращающую ее удлинение ДНК-полимеразой. Блокирующей группой, использованной на описанной выше 42-мерной бустерной матрице, был дидезоксицитидиновый нуклеотид.
Пример 6
Способ настоящего изобретения был применен для циклического удлинения самогибридизирующегося 41-звенного олигонуклеотида. Эта реакция дает 20-звенный олигонуклеотид, который может вслед за этим действовать как инициирующий праймер на 42-звенном олигонуклеотиде (бустер) с блокированным 3'-окончанием (блоком является инвертированный цитидиновый нуклеотид). Бустер сконструирован таким образом, что эта вторичная реакция также дает тот же самый 20-звенный олигонуклеотид путем циклического удлинения (см. Фиг.2). Эти 20-звенники могут затем инициировать новое удлинение на других бустерных молекулах, амплифицируя сигнал из исходной реакции на 41-звеннике; 41-звенник и 42-звенный бустер были сконструированы таким образом, чтобы обеспечить включение радиоактивно меченного [α32P]-dCTP. Реакционные смеси содержали альтернативно: (1) 100 фмоль 41-звенника, (2) 1 фмоль 41-звенника, (3) 100 фмоль бустера или (4) 1 фмоль 41-звенника и 100 фмолей бустера в буфере для фрагмента Stoffel полимеразы Taq (10 мМ трис-HCl, 10 мМ KCl, рН 8,3) с добавлением 3 мМ MgCl2, 0,2 мМ dATP, 0,2 мМ dGTP, 0,2 мМ dUTP, 0,02 мМ dCTP и 0,2 мкл [α32P]-dCTP (10 мКи/мл, ˜800 Ки/моль). Эту реакционную смесь покрывали слоем масла и инкубировали при 95°С в течение 2 мин, после чего охлаждали и выдерживали при 70°С. После этого добавляли последовательно 10 ед. фрагмента Stoffel ДНК-полимеразы Taq, 2 мкл AfUDG-Thio и 2 мкл EcoEndolV. Конечный объем реакционной смеси был равен 25 мкл. Реакцию проводили при 70°С в течение 60 мин и затем останавливали добавлением EDTA до конечной концентрации 10 мМ. Добавляли равный объем раствора для нанесения с 98% формамидом (содержащим 0,025% бромфенол синий, 0,025% ксиленцианол) и пробы анализировали электрофорезом в 20% полиакриламидом геле в денатурирующих условиях. После электрофореза гель совмещали с фосфоресцирующим экраном и изображения зон обнаруживали на визуализующем приборе Storm 860 (товарный знак).
Анализ изображений показал, что в реакции (1) шло продуцирование 20-звенного ДНК-продукта, в меньшей степени появлялись олигомеры длиной около 60 звеньев и какое-то количество продуктов длиной менее 20 звеньев. Такой же продукт появлялся в реакции (2), но в гораздо меньшем количестве. В реакции (3) не обнаруживалось никакого меченого продукта реакции, однако в реакции (4) продуцировался меченый 20-звенный продукт, причем зона была намного интенсивнее, чем в реакции (2), а количество меченых продуктов длиной менее 20 звеньев было меньше. Было обнаружено, что использование инвертированных нуклеотидов в качестве блоков 3'-концов было гораздо эффективнее, чем использование в качестве оконечных блоков дидезоксинуклеотидов.
Пример 7
Способ настоящего изобретения был применен для циклического удлинения 41-звенного олигонуклеотида (инициирующий праймер - ИП), который был самокомплементарен на своем 3'-окончании, т.е. был палиндромным, и поэтому служил матрицей самому себе для реакции GMA. Комплементарный участок простирался на 20 нуклеотидов от 3'-окончания 41-звенника и этот участок олигонуклеотида по существу функционировал как ИП. Участок 41-звенника с 5'-стороны от этой комплементарной области ИП (сайта связывания ИП) конструировали таким образом, чтобы он содержал следом за сайтом связывания ИП один аденин, что приводит после реакции GMA к продукции путем циклического удлинения 41-звенника дискретного фрагмента ДНК длиной 20 нуклеотидов. 41-звенный олигонуклеотид конструировали также таким образом, чтобы он содержал несколько гуанинов (G). Это позволило пометить получающиеся в ходе реакции замещенные направленные вниз фрагменты ДНК (20-звенные) включением [α32P]-dCTP. Олигонуклеотид синтезировали искусственно и очищали, вырезая зону из полиакриламидного геля после электрофореза. Задачей было определить, гибридизируется ли 41-звенник сам с собой и может ли он быть циклически/повторно удлинен способом настоящего изобретения с использованием экзонуклеазы III Е.coli вместо эндонуклеазы IV E.coli, давая множественные копии меченого однонитевого 20-звенного фрагмента ДНК (замещенного фрагмента). Перед добавлением ферментов реакционные смеси содержали в общем объеме 22 мкл: 100 фмоль 41-звенного олигонуклеотида, 10 мМ трис-HCl (рН 7,5), 5 мМ MgCl2, 7 мМ дитиотреитол, 0,2 мМ каждого из dATP, dGTP и dUTP, 0,02 мМ dCTP и 0,2 мкл [α32P]-dCTP (10 мКи/мл, ˜800 Ки/моль). Эту смесь покрывали слоем минерального масла и прогревали при 95°С в течение 2 мин. Затем реакционную смесь охлаждали до 37°С и выдерживали при этой температуре. После этого добавляли последовательно 5 ед. активности фрагмента Кленова (3'→5' ехо-), 10 ед. урацил-ДНК-гликозилазы E.coli и 0,1 ед. экзонуклеазы III E.coli. Конечный объем реакционной смеси становился равным 25 мкл. В контрольные реакционные смеси, не содержавшие экзонуклеазы III, добавляли воду до конечного объема 25 мкл. Реакцию проводили в течение 30 мин или дольше. Для остановки реакции добавляли EDTA до конечной концентрации 10 мМ. Затем добавляли равный объем раствора с формамидом и красителем для нанесения (98% формамид, 0,025% бромфенол синий, 0,025% ксиленцианол). После этого пробы наносили на 20% денатурирующий (7 М мочевина) полиакриламидный гель и проводили электрофорез для определения размеров фрагментов ДНК. После электрофореза гель экспонировали с фосфоресцирующим экраном и изображения зон обнаруживали с помощью визуализатора Storm 860 (товарный знак).
Анализ изображений показал, что в каждой опытной реакционной смеси, но не в контрольных реакционных смесях, продуцировалось значительное количество меченого 20-звенного олигонуклеотида
Пример 8
Способ по настоящему изобретению был применен для циклического удлинения 21-звенного олигонуклеотида (инициирующий праймер - ИП), который был комплементарен участку 40-звенного олигонуклеотида (матричная нуклеиновая кислота), который служил матрицей в реакции полимеризации. Комплементарная область простиралась от 3'-окончания 40-звенника. Участок 40-звенного олигонуклеотида, примыкающий с 5'-стороны к области комплементарности ИП (сайту связывания ИП), был сконструирован таким образом, чтобы он содержал несколько цитозинов (С), что при циклическом удлинении удлиненного 21-звенного олигонуклеотида приведет вследствие реакции GMA к продуцированию фрагментов ДНК дискретных размеров. Олигонуклеотид с 40 звеньями был сконструирован также таким образом, что между аденинами (А) расположено определенное число гуанинов (G). Это позволяет пометить получающиеся в ходе реакции замещенные направленные вниз фрагменты ДНК включением [α32P]-dCTP. Оба олигонуклеотида были синтезированы искусственно и очищены вырезанием зон из полиакриламидного геля после электрофореза. Задачей было определить, гибридизируется ли 21-звенник с 40-звенником и может ли он быть циклически/повторно удлинен способом настоящего изобретения, давая множественные копии меченых однонитевых фрагментов ДНК (замещенных фрагментов). Перед добавлением фермента реакционные смеси содержали в общем объеме 22 мкл: 100 фмоль каждого олигонуклеотида, 10 мМ трис-HCl (рН 7,5), 6 мМ MgCl2, 10 мМ KCl, 0,2 мМ каждого из dATP, dITP и dTTP, 0,02 мМ dCTP и 0,2 мкл [α32P]-dCTP (10 мКи/мл, ˜800 Ки/моль). Эту смесь покрывали слоем минерального масла и прогревали при 95°С в течение 2 мин. После этого реакционную смесь охлаждали до 70°С и выдерживали при этой температуре. Затем добавляли последовательно 10 ед. фрагмента Stoffel ДНК-полимеразы Taq и 800 пг (пикограмм) TmaEndoV. Конечный объем реакционной смеси становился равным 25 мкл. В контрольные реакционные смеси, не содержавшие эндонуклеазы V, добавляли воду до конечного объема 25 мкл. Реакции останавливали добавлением EDTA до конечной концентрации 10 мМ. Затем добавляли половинный объем раствора с формамидом и красителем для нанесения (98% формамид, 0,025% бромфенол синий, 0,025% ксиленцианол), и продукты реакции денатурировали прогревом при 95°С в течение 5 мин. Затем пробы наносили на 20% денатурирующий (7 М мочевина) полиакриламидный гель и проводили электрофорез для определения размеров фрагментов ДНК. После электрофореза гель экспонировали с фосфоресцирующим экраном и изображения зон обнаруживали прибором для визуализации Storm 860 (товарный знак).
Анализ полученных изображений показал, что реакционные смеси с эндонуклеазой V содержали значительное количество меченых фрагментов ДНК различного размера, устанавливаемых положением включения dITP в ходе удлинения. Количество этих фрагментов увеличивалось с продолжительностью реакции, и эти фрагменты не появлялись в контрольных реакционных смесях, не содержавших эндонуклеазы V.
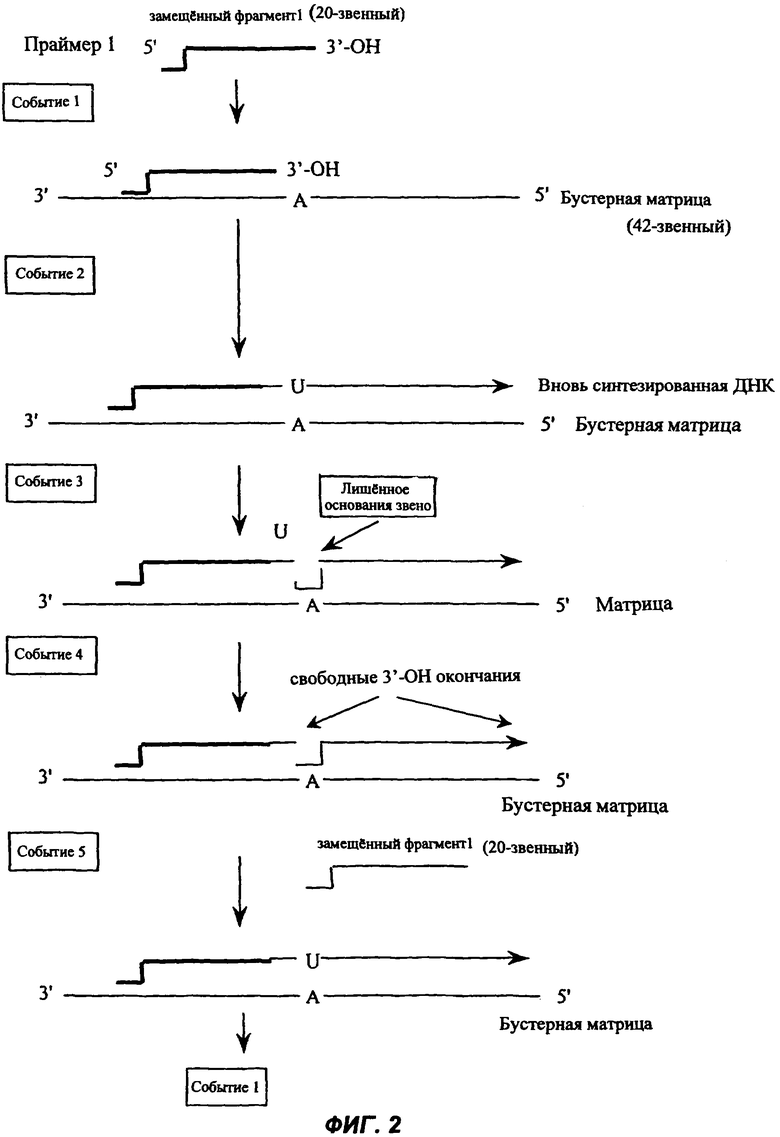
Изобретение относится к области молекулярной биологии и может быть использовано в аналитических методах различного назначения, связанных с определением количественных и качественных характеристик нуклеиновой кислоты (НК). Предложен метод амплификации НК, предусматривающий осуществление реакций удлинения праймера на матрице в присутствии, по крайней мере, одного нуклеотида - предшественника ДНК, распознаваемого специфическим расщепляющим агентом; расщепления удлиненного праймера, сохраняющего связь с матричной НК, указанным агентом с образованием фрагментов, содержащих на 3'-конце свободную ОН-группу, и удлинения полученных фрагментов на матрице. Отличительной особенностью способа по изобретению является объединение всех реакций в одном реакционном сосуде, для чего используется реакционная смесь, содержащая все необходимые для этого компоненты, а ДНК - полимераза, расщепляющий агент и условия реакции подбираются с учетом возможности их одновременного функционирования. 27 з.п., 2 ил.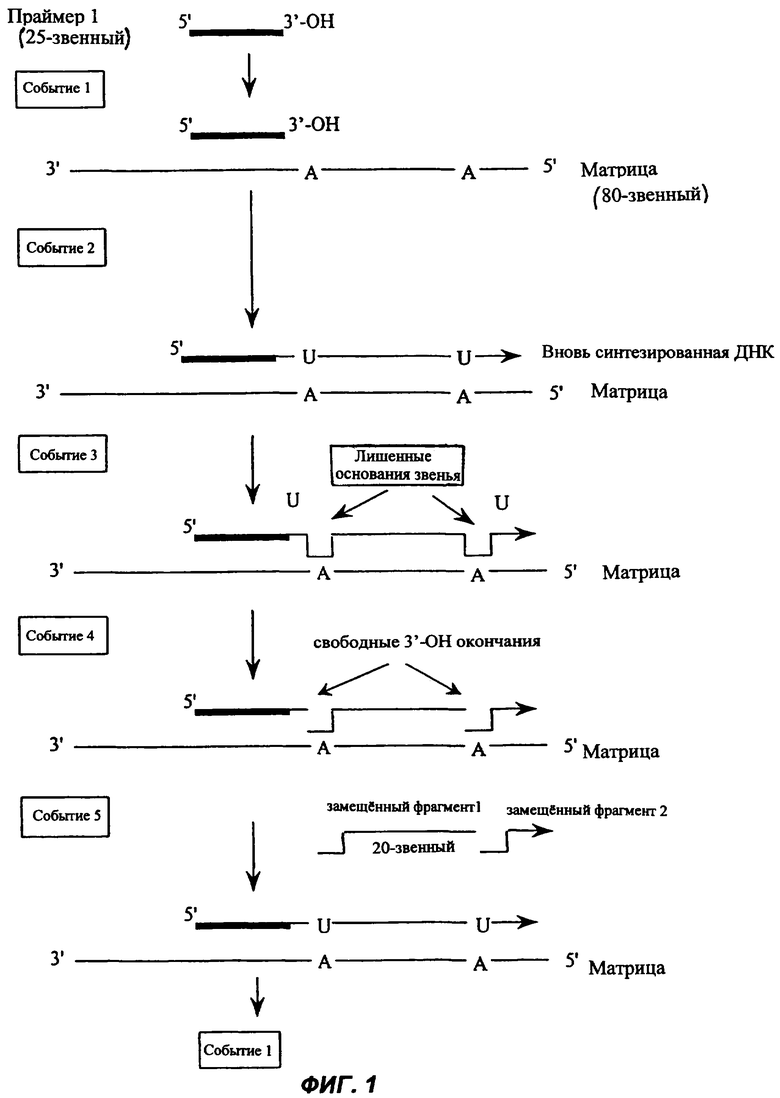
а) удлинения инициирующего праймера на указанной НК в присутствии ДНК-полимеразы, нормальных нуклеотидов-предшественников ДНК и, по меньшей мере, одного модифицированного нуклеотида-предшественника ДНК, обеспечивающего возможность расщепления удлиненного праймера, сохраняющего связь с матричной НК, по сайту встраивания модифицированного основания;
б) расщепления удлиненного праймера агентом, специфически распознающим сайт встраивания модифицированного основания, с образованием фрагментов, имеющих на 3'-конце свободную ОН-группу, и
в) реакцию удлинения ДНК-полимеразой образовавшихся фрагментов, имеющих на 3'-конце свободную ОН-группу, на матричной ДНК, отличающийся тем, что все указанные реакции проводят в одном реакционном сосуде, используя реакционную смесь, содержащую все компоненты, необходимые для их осуществления, а при выборе ДНК-полимеразы, расщепляющего агента и условий учитывают возможность одновременного функционирования названных компонентов.
i) проведение реакции праймера нуклеиновой кислоты с указанной матричной нуклеиновой кислотой, нормальными нуклеотидами-предшественниками ДНК, по меньшей мере, одним модифицированным нуклеотидом-предшественником ДНК, который является субстратом для ДНК-гликозилазы, и ДНК-полимеразой таким образом, чтобы получить удлиненный праймер нуклеиновой кислоты, причем указанный праймер нуклеиновой кислоты остается связанным с указанной матрицей;
ii) выщепление модифицированного основания модифицированного нуклеотида-предшественника ДНК из удлиненного праймера нуклеиновой кислоты с помощью ДНК-гликозилазы таким образом, чтобы создать лишенное основания звено;
iii) расщепление удлиненного праймера нуклеиновой кислоты у лишенного основания звена таким образом, чтобы создать свободный 3'-ОН конец, способный к удлинению указанной ДНК-полимеразой; и
iv) повторение этапов (i)-(iii) на созданных таким образом фрагментах ДНК.
| WO 9954501, 28.10.1999 | |||
| WO 9712061, 03.04.1997. |

