Область техники
Данное изобретение относится к способу производства L-аминокислот посредством ферментации. Аминокислоты с разветвленной цепью, такие как L-лейцин и L-изолейцин, используют в качестве пищевых продуктов, кормовых добавок, материалов для синтеза лекарственных средств и сельскохозяйственных химических препаратов, и т.д.
Область предпосылки
В качестве способов производства L-аминокислот посредством прямой ферментации известно несколько способов производства L-лейцина с использованием микроорганизмов, относящихся к роду Escherichia, Serratia, Corynebacterium или Arthrobacter. Известные способы производства L-лейцина с использованием микроорганизмов, относящихся к роду Escherichia, включают: способ с использованием микроорганизма, который устойчив к β-2-тиенилаланину (не рассмотренная опубликованная заявка на выдачу патента Японии No. 72695/81); способ с использованием микроорганизма, который устойчив к L-этионину (не рассмотренная опубликованная заявка на выдачу патента Японии No. 55194/84); способ с использованием микроорганизма, который устойчив к 2-кетомасляной кислоте (не рассмотренная опубликованная заявка на выдачу патента Японии No. 9982/96) и способ с использованием микроорганизма, который устойчив к 4-азалейцину или 5,5,5-трифторлейцину (не рассмотренная опубликованная заявка на выдачу патента Японии No. 70879/96).
Также известны способы производства L-изолейцина с использованием микроорганизмов, относящихся к роду Escherichia, Serratia, Corynebacterium или Arthrobacter. Известные способы производства L-изолейцина с применением микроорганизмов, относящихся к роду Escherichia, включают: способ с использованием микроорганизма, который устойчив к тиаизолейцину, гидроксамату изолейцина, гидроксамату аргинина, DL-этионину и т.д. (не рассмотренная опубликованная заявка на выдачу патента Японии No. 130882/93); способ с использованием микроорганизма, который устойчив к 2-кетомасляной кислоте (не рассмотренная опубликованная заявка на выдачу патента Японии No. 9982/96) и способ с использованием микроорганизма, который быстро растет в среде, содержащей L-гомосерин в качестве единственного источника азота (не рассмотренная опубликованная заявка на выдачу патента Японии No. 322583/96).
Однако в случае указанных способов проблема состоит в том, что в качестве побочных продуктов в значительных количествах образуются L-аминокислоты, отличные от требуемых L-аминокислот. В частности, образование L-валина в процессе производства L-лейцина или L-изолейцина вызывает повышение стоимости производства или снижение выхода при очистке и чистоты продукта, так как трудно осуществить отделение и удаление L-валина на стадии очистки.
Аланин-валинтрансаминаза (трансаминаза С) является ферментом, который катализирует обратимую сопряженную реакцию трансаминирования между L-аланином и пировиноградной кислотой, и 2-оксоизовалериановой кислотой и L-валином, как показано ниже:
L-аланин+2-оксоизовалериановая кислота 
пировиноградная кислота+L-валин
Также известно, что указанный фермент катализирует обратимую сопряженную реакцию трансаминирования между L-аланином и пировиноградной кислотой, и 2-оксомасляной кислотой и 2-аминомасляной кислотой, как показано ниже:
L-аланин+2-оксомасляная кислота
пировиноградная кислота+2-аминомасляная кислота
Ген, кодирующий аланин-валинтрансаминазу, обнаружен в таких микроорганизмах, как Escherichia coli и Salmonella typhimurium. Что касается аланин-валинтрансаминазы, полученной из Escherichia coli, были сообщения о клонировании гена, кодирующего фермент (avtA), и о нуклеотидной последовательности гена [J.Bacteriol., 169, 4228 (1987); Gene, 65, 195 (1988); Science, 277, 1453 (1997); Genbank, инвентарный No. AE00434 (1988)]. Также сообщалось о том, что аланин-валинтрансаминазная активность усиливается при амплификации гена avtA [J.Bacteriol., 169, 5610 (1987)].
О физиологической роли аланин-валинтрансаминазы сообщалось следующее.
Escherichia coli, которая дефицитна по ilvE, кодирующему трансаминазу аминокислот с разветвленной цепью (трансаминазу В), проявляет абсолютную потребность в L-изолейцине, но не в L-валине (фенотип неполного блокирования биохимического процесса), свидетельствуя о том, что аланин-валинтрансаминаза участвует в превращении 2-оксоизомасляной кислоты в L-валин, в качестве второй трансаминазы, заменяющей трансаминазу аминокислот с разветвленной цепью [Escherichia coli и Salmonella typhimurium, American Society for Microbiology, Washington, D.C. (1987)]. Кроме того, частичная потребность в L-валине (фенотип неполного блокирования биохимического процесса) указанного выше мутанта комплементируется амплификацией гена avtA [Escherichia coli и Salmonella typhimurium, American Society for Microbiology, Washington, D.C. (1987)].
Однако не сообщалось о том, что образование L-валина можно уменьшить посредством усиления активности аланин-валинтрансаминазы.
Сущность изобретения
Целью данного изобретения является предоставление эффективного и выгодного при промышленном применении ферментативного способа производства L-аминокислот за счет уменьшения образования других аминокислот в качестве побочных продуктов.
Авторы данного изобретения провели всесторонние исследования с целью уменьшения образования нежелательных аминокислот в качестве побочных продуктов при производстве L-аминокислоты посредством ферментации. В результате авторы изобретения обнаружили, что образование нежелательных аминокислот можно уменьшить посредством усиления активности аланин-валинтрансаминазы штамма, продуцирующего требуемую L-аминокислоту, и осуществили изобретение.
Данное изобретение относится к следующим пунктам с (1) по (11).
(1) Способ производства L-аминокислоты, который включает в себя культивирование в среде микроорганизма, обладающего способностью продуцировать L-аминокислоту, аланин-валинтрансаминазная активность которого повышена по сравнению с его родительским штаммом, обеспечение возможности образования и накопления в культуре L-аминокислоты, и извлечение L-аминокислоты из культуры.
(2) Способ по п.(1), где микроорганизм выбран из группы, состоящей из мутанта, линии слитых клеток, трансдуктанта и рекомбинантного штамма, сконструированного с помощью технологии рекомбинантной ДНК.
(3) Способ по п.(1) или (2), где микроорганизм выбран из группы, состоящей из родов Escherichia, Serratia, Corynebacterium и Arthrobacter.
(4) Способ по п.п. с (1) по (3), где микроорганизмом является Escherichia coli H-8719/pAD27 (FERM BP-7063) или Escherichia coli H-9156/pAD27.
(5) Способ по п.(1), где L-аминокислота выбрана из группы, состоящей из L-лейцина и L-изолейцина.
(6) Способ по п.(1), где аланин-валинтрансаминазная активность микроорганизма повышена посредством увеличения в клетках микроорганизма уровня экспрессии гена аланин-валинтрансаминазы.
(7) Способ по п.(1), где аланин-валинтрансаминазная активность микроорганизма повышена посредством увеличения в клетках микроорганизма числа копий гена аланин-валинтрансаминазы.
(8) Микроорганизм, обладающий способностью продуцировать L-аминокислоту, аланин-валинтрансаминазная активность которого повышена по сравнению с его родительским штаммом.
(9) Микроорганизм по п.(8), который выбран из группы, состоящей из мутанта, линии слитых клеток, трансдуктанта и рекомбинантного штамма, сконструированного с помощью технологии рекомбинантной ДНК.
(10) Микроорганизм по п.(8) или (9), который выбран из группы, состоящей из родов Escherichia, Serratia, Corynebacterium и Arthrobacter.
(11) Микроорганизм по п.п. с (8) по (10), который представляет собой Escherichia coli H-8719/pAD27 (FERM BP-7063) или Escherichia coli H-9156/pAD27.
Подробно данное изобретение описано ниже.
В данном изобретении можно использовать любой организм при условии, что его аланин-валинтрансаминазная активность усилена по сравнению с его родительским штаммом, и он обладает способностью продуцировать L-аминокислоту. Термин "родительский штамм" в используемом здесь смысле означает микроорганизм, используемый в качестве исходного микроорганизма для того, чтобы сконструировать мутант, линию слитых клеток, трансдуктант или рекомбинантный штамм. Микроорганизмом, аланин-валинтрансаминазная активность которого усилена по сравнению с его родительским штаммом, может быть любой мутант, трансдуктант, рекомбинантный штамм и любая линия слитых клеток. Примеры микроорганизмов включают микроорганизмы, выбранные из микроорганизмов, относящихся к родам Escherichia, Serratia, Corynebacterium и Arthrobacter. Предпочтительными примерами являются так называемые коринеформные штаммы, продуцирующие глутаминовую кислоту, такие как Corynebacterium glutamicum и Corynebacterium lactofermentum, и Escherichia coli, которые используют в ферментации аминокислот.
Конкретными примерами родительских штаммов являются Escherichia coli Н-8719 (штамм, продуцирующий L-лейцин, который устойчив к 4-азалейцину, полученный из штамма FERM ВР-4704), обладающий способностью продуцировать L-лейцин, и Escherichia coli H-9156 (FERM BP-5056), обладающий способностью продуцировать L-изолейцин.
Аланин-валинтрансаминазную активность указанных микроорганизмов можно, например, усилить следующими способами.
I. Способ, при котором мутант, обладающий повышенной аланин-валинтрансаминазной активностью, выбран из микроорганизмов, полученных посредством обработки мутагеном микроорганизма, несущего ген аланин-валинтрансаминазы.
II. Способ, при котором в ген аланин-валинтрансаминазы вводят мутацию in vitro, и из мутированных генов выбирают ген, кодирующий аланин-валинтрансаминазу, активность которой выше, чем перед введением мутации, и выбранный ген вводят в микроорганизм-хозяин.
III. Способ, при котором в клетках увеличивают число копий гена, кодирующего аланин-валинтрансаминазу.
IV. Способ, при котором in vitro модифицируют район, ответственный за экспрессию гена, кодирующего аланин-валинтрансаминазу, чтобы увеличить уровень экспрессии гена, и модифицированным геном заменяют ген, кодирующий аланин-валинтрансаминазу, в хромосоме хозяина.
Указанный выше способ, при котором мутант, обладающий повышенной активностью аланин-валинтрансаминазы, выбирают из микроорганизмов, полученных путем обработки мутагеном микроорганизма, несущего ген аланин-валинтрансаминазы, выполняют, например, следующим образом. Микроорганизм, несущий ген аланин-валинтрансаминазы, подвергают мутагенезу известным способом, используя такой мутаген, как N-метил-N'-нитро-N-нитрозогуанидин, и из обработанных мутагеном микроорганизмов отбирают микроорганизм, обладающий повышенной аланин-валинтрансаминазной активностью по сравнению с его родительским штаммом, подвергаемым мутагенезу. Аланин-валинтранс-аминазную активность микроорганизма можно, например, измерить, культивируя микроорганизм в соответствующей среде, подвергая культивируемые клетки центрифугированию, разрушая полученные клетки известным способом, чтобы получить неочищенный раствор ферментов, выполняя ферментативную реакцию с использованием неочищенного раствора ферментов и L-аланина или пировиноградной кислоты и L-валина в качестве субстрата, и измеряя количество L-валина или L-аланина, образованного в результате ферментативной реакции.
Примеры способов, при которых вводят мутацию в ген аланин-валинтрансаминазы in vitro, и из мутированных генов отбирают ген, кодирующий аланин-валинтрансаминазу, активность которой выше, чем активность до введения мутации, включают следующие способы;
1) способ, при котором в ген аланин-валинтрансаминазы вводят делению, замену или присоединение основания посредством сайт-специфичного мутагенеза [Nucleic Acids Research, 10, 6487 (1982); Proc. Natl. Acad. Sci., USA, 79, 6409 (1982); Proc. Natl. Acad. Sci., USA, 81, 5662 (1984); Science, 224, 1431 (1984); заявка РСТ WO 85/00817 (1985); Nature, 316, 601 (1985); Gene, 34, 315 (1985); Nucleic Acids Research, 13, 4431 (1985); Current Protocols in Molecular Biology, John Wiley and Sons (1987-1997)], чтобы получить ген, кодирующий аланин-валинтрансаминазу, активность которой выше, чем перед мутагенезом; и
2) способ, при котором мутацию, такую как замена основания, случайным образом вводят в ген аланин-валинтрансаминазы посредством полимеразной цепной реакции, подверженной ошибкам [Bio/Technol., 9, 1073 (1991) (в дальнейшем полимеразную цепную реакцию называют ПЦР)], и из мутированных генов отбирают ген, кодирующий аланин-валинтрансаминазу, активность которой выше, чем активность перед мутагенезом.
Число копий гена, кодирующего аланин-валинтрансаминазу, можно, например, увеличить посредством клонирования гена, кодирующего аланин-валинтрансаминазу, и затем
1) лигирования фрагмента ДНК, содержащего ген, кодирующий аланин-валинтрансаминазу, в плазмидный вектор, способный к автономной репликации в клетках требуемого микроорганизма, и введения полученной в результате плазмиды в микроорганизм; или
2) интеграции рекомбинантной ДНК, содержащей ген, кодирующий аланин-валинтрансаминазу, в хромосому штамма, используемого в качестве хозяина, путем гомологичной рекомбинации, или с использованием фага или транспозона.
Примеры способов, при которых район, ответственный за экспрессию гена, кодирующего аланин-валинтрансаминазу, модифицируют in vitro, чтобы увеличить уровень экспрессии гена, и модифицированным геном заменяют ген, кодирующий аланин-валинтрансаминазу, в хромосоме хозяина, включают следующие способы:
1) способ, при котором район, ответственный за экспрессию гена в микроорганизме, используемом в качестве хозяина для введения и экспрессии гена, заменяют известным промотором, обладающим сильной промоторной активностью; и
2) способ, при котором делецию, замену или присоединение основания вводят в ДНК, имеющую нуклеотидную последовательность, ответственную за экспрессию гена, путем описанного выше сайт-специфичного мутагенеза или склонной к ошибкам ПЦР, и из мутантных ДНК отбирают ДНК, увеличивающую уровень экспрессии гена, кодирующего аланин-валинтрансаминазу, по сравнению с уровнем перед мутагенезом.
Ген, кодирующий аланин-валинтрансаминазу, подходящий для данного изобретения, можно получить из клеток, предпочтительно микроорганизма, более предпочтительно микроорганизма, относящегося к роду Escherichia или Salmonella.
Ген, кодирующий аланин-валинтрансаминазу, можно, например, получить следующими способами.
I. В том случае, когда известна нуклеотидная последовательность гена, кодирующего аланин-валинтрансаминазу микроорганизма, как в случае с Escherichia coli или Salmonella typhimurium, ген можно получить в ПЦР, основанной на нуклеотидной последовательности, с использованием хромосомной ДНК микроорганизма в качестве матрицы [PCR Protocols, Academic Press (1990)].
II. В том случае, когда нуклеотидная последовательность гена, кодирующего аланин-валинтрансаминазу клеток, обладающих аланин-валинтрансаминазной активностью, неизвестна, библиотеку кДНК или библиотеку хромосомной ДНК, полученной из клеток, готовят обычным способом [Molecular Cloning, A Laboratory Manual, Second Edition (1989) (в дальнейшем сокращенно обозначают Molecular Cloning, 2nd ed.)], и затем
1) измеряют аланин-валинтрансаминазную активность каждой из клеток, составляющих библиотеку, и из клеток отбирают клетку, содержащую ген, кодирующий аланин-валинтрансаминазу, или
2)клетку, содержащую ген, кодирующий аланин-валинтрансаминазу, отбирают из клеток, составляющих библиотеку с помощью гибридизации колоний или гибридизации бляшек (Molecular Cloning, 2nd ed.), используя в качестве зонда ген аланин-валинтрансаминазы Escherichia coli или Salmonella typhimurium.
III. Когда известна полная нуклеотидная последовательность хромосомной ДНК, но точно не определен ген, кодирующий аланин-валинтрансаминазу, ген, имеющий нуклеотидную последовательность, в высокой степени гомологичную нуклеотидной последовательности гена аланин-валинтрансаминазы, полученного из Escherichia coli или Salmonella typhimurium, определяют в указанной выше полной нуклеотидной последовательности с использованием компьютерной программы анализа, такой как BLAST [J. Mol. Biol., 215, 403 (1990)] или FASTA [Methods in Enzymology, 183, 63 (1990)] и требуемый ген получают посредством ПЦР.
Клонирование гена avtA, который является геном, кодирующим аланин-валинтрансаминазу Escherichia coli, можно, например, выполнить следующим образом.
Сначала район (приблизительно 1,9 т.п.н.), содержащий ген avtA и его промоторную последовательность, амплифицируют в ПЦР, используя в качестве набора праймеров два вида ДНК-праймеров, сконструированных и синтезированных на основе последовательности гена avtA, полученного из микроорганизма, относящегося к роду Escherichia, и прилегающей к нему последовательности [GenBank, инвентарный No. AE00434 (1998)], например, ДНК, соответственно, имеющие нуклеотидные последовательности, показанные в SEQ ID NO: 1 и 2, и используя в качестве матрицы хромосомную ДНК микроорганизма.
Полученный ген avtA лигируют в плазмидный вектор, способный к автономной репликации в клетках микроорганизма, в который вводят ген, и рекомбинантный вектор вводят в клетки микроорганизма обычным способом, посредством чего можно клонировать ген avtA.
В качестве плазмидного вектора в данном изобретении можно использовать любую плазмиду, способную к автономной репликации в клетках микроорганизма, в который вводят ген, кодирующий аланин-валинтрансаминазу. В том случае, когда микроорганизмом, в который вводят ген, является Escherichia coli, можно использовать любую плазмиду, способную к автономной репликации в клетках Escherichia coli. Подходящие плазмиды включают ZAP Express [Stratagene; Strategies, 5, 58 (1992)], pBluescript II SK(+) [Nucleic Acids Research, 17, 9494 (1989)], λzap II (Stratagene), λgt10, λgt11 [DNA Cloning, A Practical Approach, 1, 49 (1985)], λTriplEx (Clontech Laboratories, Inc.), λBlueMid (Clontech Laboratories, Inc.), λExCell (Pharmacia), pT7T318U (Pharmacia), pcD2 [Mol. Cell. Biol., 3, 280 (1983)], pUC18 [Gene, 33, 103 (1985)], pUC19 [Gene, 33, 103 (1985)] и их производные.
Можно увеличивать уровень экспрессии гена, кодирующего аланин-валинтрансаминазу, с использованием ДНК, полученной путем лигирования гена avtA в положении ниже промотора, который функционирует в клетках микроорганизма, в который вводят ген. Когда микроорганизмом, в который вводят ген, является Escherichia coli, можно использовать любой промотор, способный функционировать в клетках-хозяевах. Подходящие промоторы включают промоторы, полученные из Escherichia coli, фага, и т.д., такие как промотор trp (Ptrp), промотор lac, промотор PL, промотор PR и промотор Т7. Также можно использовать искусственно сконструированные и модифицированные промоторы, такие как промотор, в котором объединены в тандем два PtrpS (Ptrp×2), промотор tac, промотор lасТ7 и промотор letI.
Предпочтительно использовать плазмиду, в которой расстояние между последовательностью Шайн-Далгарно (последовательность, связывающая рибосомы) и кодоном инициации доводят до соответствующей длины (например, от 6 до 18 оснований).
Примеры способов введения плазмиды в клетки микроорганизма, в который вводят ген, включают электропорацию, которая вызывает включение ДНК в клетки в результате приложения электрических импульсов высокого напряжения [Nucleic Acids Res., 16, 6127 (1988)] и способ протопластов (не рассмотренная опубликованная заявка на патент Японии No. 248394/88). В том случае, когда плазмиду вводят в Escherichia coli, также можно использовать способ, при котором проницаемость ДНК увеличивают, используя хлорид кальция (Molecular Cloning, 2nd ed.).
Микроорганизм, несущий введенную плазмиду, содержащую ген, кодирующий аланин-валинтрансаминазу, можно отобрать, используя в качестве индикатора устойчивость микроорганизма к лекарственному средству, приобретенную благодаря гену устойчивости к лекарственному средству в плазмиде. То, что полученный таким образом трансформант является требуемым рекомбинантным микроорганизмом, можно установить посредством определения его аланин-валинтрансаминазной активности, рестрикционным ферментативным картированием фрагмента ДНК, встроенного в плазмиду, или определением его нуклеотидной последовательности.
Получение L-аминокислоты с использованием полученного таким образом штамма, продуцирующего L-аминокислоту, аланин-валинтрансаминазная активность которого повышена по сравнению со штаммом-хозяином, можно выполнить обычным способом культивирования, используемым для производства L-аминокислот посредством ферментации. А именно, культивирование проводят в среде, содержащей источники углерода, источники азота, неорганические соли, витамины и другие компоненты, необходимые для роста штамма, в аэробных условиях при установленных на соответствующих уровнях температуре и рН.
Примеры источников углерода включают различные углеводы, такие как глюкоза, фруктоза, сахароза и лактоза, содержащую их мелассу, гидролизат целлюлозы, неочищенный гидролизат сахара и гидролизат крахмала.
Примеры источников азота включают аммиак, аммонийные соли различных неорганических или органических кислот, такие как хлорид аммония, сульфат аммония, ацетат аммония и фосфат аммония, амины и другие азотсодержащие соединения, пептон, мясной экстракт, дрожжевой экстракт, сироп на основе кукурузного экстракта, гидролизат казеина, гидролизат соевого белка, гидролизат соевого жмыха и различные ферментированные клетки микроорганизмов и их расщепленные продукты.
Примеры неорганических солей включают однозамещенный фосфат калия, двузамещенный фосфат калия, фосфат магния, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, сульфат меди и карбонат кальция.
Культивирование проводят в аэробных условиях, например, во встряхиваемой культуре или вращающейся культуре при аэрировании. Температура культивирования предпочтительно составляет от 20 до 40°С. рН среды находится в пределах от 5 до 9, и предпочтительно поддерживается около нейтрального значения. Доведение рН осуществляют, используя карбонат кальция, неорганическую или органическую кислоту, раствор щелочи, аммиак, рН-буфер и т.д. L-аминокислота образуется и накапливается в культуре обычно при культивировании от одного до 7 дней.
L-аминокислоту можно извлечь из культуры известными способами, такими как способ с применением ионообменной смолы, концентрирование, высаливание и осаждение [The Society of Chemical Engineers, Japan (ed.), Handbook of Bioseparation Process, Kyoritsu Shuppan (1996)].
Примеры данного изобретения показаны ниже. Приведенные примеры не следует рассматривать как ограничивающие рамки изобретения.
Краткое описание чертежей
На чертеже показана структура плазмиды pAD27. Символы, используемые на чертеже, представляют собой следующее.
Ampr: ген устойчивости к ампициллину
ori: начало репликации, функционирующее в Escherichia coli
avtA: ген, кодирующий аланин-валинтрансаминазу, полученный из Escherichia coli
Наилучший способ выполнения изобретения.
Пример 1. Клонирование гена avtA Escherichia coli.
Штамм W3110 (АТСС 27325), полученный из Escherichia coli К-12, высевали на агаризованную среду LB на чашках [10 г/л триптона-пептона (Difco Laboratories Inc.), 5 г/л дрожжевого экстракта, 10 г/л хлорида натрия и 15 г/л агара, рН 7,5], с последующим культивированием при 37°С в течение 24 часов. Одну платиновую петлю культивируемых клеток инокулировали в 8 мл среды LB [10 г/л триптона-пептона (Difco Laboratories Inc.), 5 г/л дрожжевого экстракта и 10 г/л хлорида натрия, рН 7,5], после чего культивировали при встряхивании (колебательное встряхивание: 300 об/мин) в большой пробирке (диаметр: 25 мм, длина: 200 мм) при 37°С в течение 24 час. Полученную в результате культуру (1,25 мл) переносили в 250 мл среды LB и культивировали при встряхивании (ротационное встряхивание: 200 об/мин) в колбе Эрленмейера объемом 2 л при 37°С в течение 24 час.
Полученную культуру подвергали центрифугированию при 5000 об/мин при 4°С в течение 10 минут и собирали клетки. Полученные клетки промывали буфером ТЕ [10 ммоль/л трис(гидроксиметил)аминометана, 1 ммоль/л динатрийэтилендиаминтетраацетата, рН 7,5] и затем собирали. Из собранных клеток выделяли хромосомную ДНК согласно способу Saito-Miura [Biochem. Biophys. Acta, 72, 619 (1963)].
Для того чтобы амплифицировать ген avtA посредством ПЦР, используя в качестве матрицы хромосомную ДНК, синтезировали олигонуклеотидные праймеры, соответственно имеющие нуклеотидные последовательности, показанные в SEQ ID NO: 1 и 2, на основе известных нуклеотидных последовательностей гена avtA и окружающей его области [GenBank, инвентарный No. АЕ00434 (1998)]. ДНК, имеющие нуклеотидные последовательности, показанные в SEQ ID NO: 1 и 2, являются праймерами, содержащими последовательности, гомологичные или комплементарные последовательности выше гена avtA, содержащей промотор, и последовательности ниже гена avtA, соответственно. Ген avtA амплифицировали посредством ПЦР, используя описанную выше хромосомную ДНК и набор праймеров. ПЦР выполняли, 30 раз повторяя цикл реакций, при котором реакцию проводили при 94°С в течение одной минуты, при 55°С в течение двух минут и при 72°С в течение двух минут.
Фрагмент ДНК (1,9 т.п.н.), амплифицированный посредством ПЦР, затупляли ДНК-полимеразой Т4 и затем лигировали в плазмидный вектор pUC19, расщепленный ферментами рестрикции EcoRI и PstI и затупленный ДНК-полимеразой Т 4, используя ДНК-лигазу Т4. Полученный продукт реакции использовали для трансформации Escherichia coli Н-8719 путем электропорации. Полученную в результате суспензию клеток высевали на агаризованную среду LB на чашках, содержащую 100 мг/л ампициллина, после чего культивировали при 37°С в течение 24 часов. Отбирали колонии, которые вырастали на агаризованной среде на чашках, и с помощью разных ферментов рестрикции анализировали структуру фрагментов ДНК, встроенных в плазмиды, которые несли полученные трансформанты, тем самым подтвердили, что ген avtA существовал во встроенных фрагментах ДНК.
Аланин-валинтрансаминазную активность полученных трансформантов измеряли следующим образом. После культивирования со встряхиванием (колебательное встряхивание: 300 об/мин) в среде LB при 30°С в течение 24 часов культивируемые клетки промывали посредством суспендирования и центрифугирования дважды водным раствором хлорида натрия в концентрации 8,5 г/л и один раз буфером А (25 ммоль/л калий-фосфатного буфера, рН 7,0, 50 мл/л глицерина, 0,1 ммоль/л тринатрийэтилендиаминтетраацетата, 0,2 ммоль/л дитиотреитола и 0,2 ммоль/л пиридоксальфосфата). Затем клетки ресуспендировали в таком же буфере при плотности 100 г влажной массы клеток в литре. Суспендированные клетки разрушали ультразвуком, после чего центрифугировали, чтобы получить неочищенный раствор ферментов. Полученный неочищенный раствор ферментов (20 мкл) добавляли к 980 мкл реакционного раствора, полученного добавлением 10 ммоль/л пировиноградной кислоты и 10 ммоль/л L-валина к буферу А, с последующим протеканием реакции при 37°С в течение 30 минут. Образованный L-аланин определяли высокоэффективной жидкостной хроматографией (ВЭЖХ), чтобы измерить аланин-валинтрансаминазную активность. Условия для анализа ВЭЖХ были следующими.
Колонка: YMC ODS-AQ312
Подвижная фаза: 2,94 г/л цитрата тринатрия, 1,42 г/л сульфата натрия, 63 мл/л n-пропанола, 3 г/л додецилсульфата натрия, рН 3,75 (доводили 2 моль/л серной кислоты)
Скорость потока подвижной фазы: 2 мл/минуту
Реакционный раствор: 18,5 г/л борной кислоты, 11 г/л гидроксида натрия, 3 мл/л Brig-35, 0,6 г/л о-фтальальдегида, 2 мл/л меркаптоэтанола
Скорость потока реакционного раствора: 1 мл/минуту
В результате обнаружено, что аланин-валинтрансаминазная активность трансформанта была более чем в 10 раз выше, чем эта же активность Escherichia coli Н-8719, которую считали равной 1. Полученную таким образом плазмиду назвали pAD27 (фиг.1).
Рекомбинант Escherichia coli H-8719/pAD27, полученный как указано выше, депонировали в National Institute of Bioscience and Human-Technology, National Institute of Advanced Industrial Science and Technology, Ministry of Economy, Trade and Industry, 1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki, 305-0046 Japan, 2 марта 2000 г. с инвентарным номером FERM BP-7063 по условиям Будапештского договора.
Плазмиду pAD27 получали из культивируемых клеток Escherichia coli H-8719/pAD27, и Escherichia coli H-9156, обладающую продуктивностью по L-изолейцину, трансформировали, используя полученную плазмиду, посредством электропорации.
Рекомбинант Escherichia coli H-9156/pAD27, несущий pAD27, получали отбором устойчивого к ампициллину трансформанта таким же способом, который описан выше. Аланин-валинтрансаминазную активность полученного таким образом трансформанта измеряли таким же способом, который описан выше. В результате подтвердили, что аланин-валинтрансаминазная активность трансформант была более чем в 10 раз выше, чем эта же активность Escherichia coli H-9156, принятая за 1.
Пример 2. Анализ продукции L-лейцина.
Анализ продукции L-лейцина Escherichia coli H-8719, Н-8719/pAD27, несущей плазмиду pAD27, содержащую ген avtA, и H-8719/pUC19, несущей плазмидный вектор pUC19, выполняли следующим образом.
H-8719/pAD27 и H-8719/pUC19 отдельно высевали на агаризованную среду LB на чашках, содержащую 100 мг/л ампициллина, Н-8719 высевали на агаризованную среду LB на чашках, не содержащую ампициллина. Каждый штамм культивировали при 30°С в течение 24 часов. Одну платиновую петлю культивируемых клеток инокулировали в 6 мл среды для посева [20 г/л глюкозы, 10 г/л пептона, 10 г/л дрожжевого экстракта, 2,5 г/л хлорида натрия и 10 г/л карбоната кальция, рН 7,4], с последующим культивированием при встряхивании (колебательное встряхивание: 300 об/мин) в большой пробирке (диаметр: 25 мм, длина: 200 мм) при 30°С в течение 17 часов. Полученную культуру (0,1 мл) переносили в 6 мл среды для продуцирования [65 г/л глюкозы, 2 г/л сиропа кукурузного экстракта, 16 г/л сульфата аммония, 2 г/л однозамещенного фосфата калия, 40 г/л фосфата магния и 10 г/л карбоната кальция, рН 7,0], с последующим культивированием при встряхивании (колебательное встряхивание: 300 об/мин) в большой пробирке (диаметр: 25 мм, длина: 200 мм) при 30°С в течение 48 часов.
После завершения культивирования количества L-лейцина, образованного и накопленного в культуре, и L-валина, образованного как побочный продукт, определяли посредством ВЭЖХ (способ такой же, как способ определения L-аланина, описанный в примере 1).
Описанный выше анализ культивирования выполняли независимо 20 раз, и средние значения полученных результатов показаны в таблице 1.
Средние значения (n=20)
Три штамма показали почти одинаковую продуктивность по L-лейцину. Отношение L-валина, как побочного продукта, к L-лейцину в случае H-8719/pAD27, у которого уровень экспрессии гена avtA повышен, было всегда уменьшено по сравнению с тем же отношением у хозяина Н-8719, степень уменьшения составляла примерно 54%.
Приведенные выше результаты свидетельствуют о том, что образование L-валина как побочного продукта, который мешает очистке при производстве L-лейцина ферментацией, может быть значительно снижено при усилении активности аланин-валинтрансаминазы, кодируемой геном avtA штамма, продуцирующего L-лейцин.
Пример 3. Анализ продукции L-изолейцина.
Анализ продукции L-изолейцина Escherichia coli H-9156, H-9156/pAD27, несущей плазмиду pAD27, содержащую ген avtA, и H-9156/pUC19, несущей плазмидный вектор pUC19, выполняли следующим образом.
H-9156/pAD27 и H-9156/pUC19 отдельно высевали на агаризованную среду LB на чашках, содержащую 100 мг/л ампициллина, а H-9156 высевали на агаризованную среду LB на чашках, не содержащую ампициллина. Каждый штамм культивировали при 30°С в течение 24 часов. Одну платиновую петлю культивируемых клеток инокулировали в 6 мл среды для посева [20 г/л глюкозы, 10 г/л пептона, 10 г/л дрожжевого экстракта, 2,5 г/л хлорида натрия и 10 г/л карбоната кальция, рН 7,4], с последующим культивированием при встряхивании (колебательное встряхивание: 300 об/мин) в большой пробирке (диаметр: 25 мм, длина: 200 мм) при 30°С в течение 17 часов. Полученную культуру (0,1 мл) переносили в 6 мл среды для продуцирования [65 г/л глюкозы, 2 г/л сиропа кукурузного экстракта, 16 г/л сульфата аммония, 2 г/л однозамещенного фосфата калия, 0,1 г/л DL-метионина, 40 г/л фосфата магния и 10 г/л карбоната кальция, рН 7,0], с последующим культивированием при встряхивании (колебательное встряхивание: 300 об/мин) в большой пробирке (диаметр: 25 мм, длина: 200 мм) при 30°С в течение 48 часов.
После завершения культивирования количества L-изолейцина, образованного и накопленного в культуре, и L-валина, образованного как побочный продукт, определяли посредством высокоэффективной жидкостной хроматографии. Условия измерения были такими же, как условия, описанные в примере 2.
Описанный выше анализ культивирования выполняли независимо 20 раз, и средние значения полученных результатов показаны в таблице 2.
Средние значения (n=20)
Три штамма показали почти одинаковую продуктивность по L-изолейцину. Отношение L-валина, как побочного продукта, к L-изолейцину в случае H-9156/pAD27, у которого уровень экспрессии гена avtA повышен, было всегда уменьшено по сравнению с отношением у хозяина Н-9156, степень уменьшения составляла примерно 48%.
Приведенные выше результаты свидетельствуют о том, что образование L-валина как побочного продукта, который мешает очистке при производстве L-изолейцина ферментацией, может быть значительно снижено при усилении активности аланин-валинтрансаминазы, кодируемой геном avtA штамма, продуцирующего L-изолейцин.
Промышленная применимость
Согласно данному изобретению при использовании микроорганизма, обладающего способностью продуцировать L-аминокислоту, аланин-валинтрансаминазная (С-трансаминазная) активность которого повышена, можно снизить образование аминокислот-побочных продуктов, которые мешают очистке при производстве L-аминокислоты ферментацией, и можно получить способ производства L-аминокислоты, обладающий преимуществами при промышленном применении.
Текст без указания последовательностей
SEQ ID NO: 1 - Описание искусственной последовательности: синтетическая ДНК.
SEQ ID NO: 2 - Описание искусственной последовательности: синтетическая ДНК.

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ Escherichia coli - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2001 |
|
RU2215784C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2010 |
|
RU2460793C2 |
| МУТАНТНАЯ АЦЕТОЛАКТАТСИНТАЗА И СПОСОБ ПРОДУКЦИИ РАЗВЕТВЛЕННЫХ L-АМИНОКИСЛОТ | 2006 |
|
RU2355763C2 |
| Микроорганизм, продуцирующий O-ацетилгомосерин, и способ получения O-ацетилгомосерина с использованием этого микроорганизма | 2016 |
|
RU2706535C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae | 2010 |
|
RU2497943C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2368659C2 |
| МУТАНТНАЯ АДЕНИЛАТЦИКЛАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ СЕМЕЙСТВА ENTEROBACTERIACEAE, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2010 |
|
RU2471868C2 |
| ФРАГМЕНТ ДНК ИЗ ESCHERICHIA COLI, ОПРЕДЕЛЯЮЩИЙ ПОВЫШЕННУЮ ПРОДУКЦИЮ L-АМИНОКИСЛОТ (ВАРИАНТЫ), И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 1999 |
|
RU2175351C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2351653C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae | 2010 |
|
RU2501858C2 |
Изобретение относится к биотехнологии L-аминокислот, таких как L-изолейцин или L-лейцин, которые получают культивированием в среде микроорганизма, способного продуцировать L-аминокислоту и который несет ген avtA, кодирующий аланин-валинтрансаминазу (трансаминазу С), и обладает повышенной активностью указанного фермента по сравнению с его родительским штаммом. Накопленные L-изолейцин или L-лейцин выделяют из культуры. Заявленное изобретение позволяет получить L-изолейцин или L-лейцин с высокой степенью эффективности, уменьшая количество аминокислоты, являющейся побочным продуктом, который приводит к осложнениям при очистке указанных L-аминокислот. 3 н. и 5 з.п. ф-лы, 1 ил., 2 табл.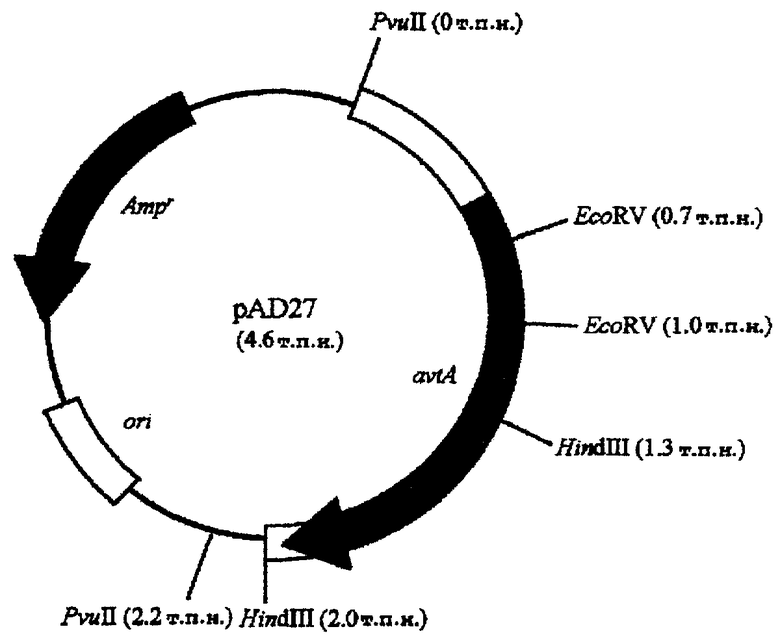
| MING-DER WANG et al., Cloning and characterization of the Escherichia coli K-12 alaninevaline transaminase (AVTA) gene, Journal of Bacteriology, vol 169, no 9, 1987, p.4228-4634 | |||
| Устройство для протяжки рукавного материала в упаковочных машинах | 1978 |
|
SU685555A1 |
| ФРАГМЕНТ ДНК rhtB, КОДИРУЮЩИЙ СИНТЕЗ БЕЛКА RhtB, ПРИДАЮЩЕГО УСТОЙЧИВОСТЬ К L-ГОМОСЕРИНУ БАКТЕРИЯМ ESCHERICHIA COLI, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 1998 |
|
RU2144564C1 |

