Область техники, к которой относится изобретение
Настоящее изобретение относится к гуманизированным антителам к ЕrВ2 и способам лечения рака с помощью антител к ЕrВ2, таких как гуманизированные антитела к ЕrВ2.
Предпосылки создания изобретения
Семейство ErbB2-рецепторов тирозинкиназ включает важные медиаторы роста, дифференцировки и выживания клеток. Семейство рецепторов включает 4 различных представителя, в том числе рецептор эпидермального фактора роста (EGFR или ErbB1), HER2 (ErbB2 или p185neu), HER3 (ErbB3) и HER4 (ErbB4 или tyro2).
Известно, что EGFR, кодируемый геном erbB1, вызывает злокачественные болезни у людей. В частности, повышенный уровень экспрессии EGFR был обнаружен при раке молочной железы, мочевого пузыря, легкого, головы, шеи и желудка, а также при глиобластомах. Повышенный уровень экспрессии рецептора EGF часто связывают с повышенным производством лиганда EGFR, т.е. трансформирующего фактора роста альфа (TGF-α), указанными клетками, что приводит к активации рецептора посредством аутокринного пути стимуляции (Baselga и Mendelsohn, Pharmac Ther. 64: 127-154 (1994)). Моноклональные антитела к EGFR и его лигандам TGF-α, и EGF, были изучены в качестве терапевтических агентов для лечения указанных злокачественных заболеваний (см., например, Baselga и Mendelsohn, выше, Masui и др., Cancer Research 44: 1002-1007 (1984) и Wu и др., J.Clin. Invest. 95: 1897-1905 (1995)).
Второй представитель ErbB-семейства, 185neu, был впервые выявлен как продукт трансформирующего гена из нейробластом, обработанных химическими агентами крыс. Активированная форма проонкогена neu возникает в результате точковой мутации (замена валина на глутаминовую кислоту) в трансмембранной области кодируемого протеина. Амплификация человеческого гомолога neu была обнаружена при раке молочной железы и яичника, и она коррелирует с неблагоприятным прогнозом в отношении развития болезни (Slamon и др., Science, 235: 177-182 (1987); Slamon и др., Science, 244: 707-712 (1989); патент США 4968603). К настоящему времени в человеческих опухолях не обнаружено никаких аналогов точковых мутаций, характерных для проонкогена neu. Сверхэкспрессия ErbB2 (которая часто, но не всегда является результатом амплификации гена) также была обнаружена при других формах рака, включая карциномы желудка, эндометрия, слюнной железы, легкого, почки, ободочной кишки, щитовидной железы, поджелудочной железы и мочевого пузыря (см. среди прочего у King и др., Science, 229: 974 (1985); Yokota и др., Lancet, 1: 765-767 (1986); Fukushigi и др., Mol. Cell Biol, 6: 955-958 (1986); Geurin и др., Oncogene Res., 3: 21-31 (1988); Cohen и др., Oncogene, 4: 81-88 (1989), Yonemura и др., Cancer Res. 51: 1034 (1991); Borst и др., Gynecol. Oncol., 38: 364 (1990); Weiner и др., Cancer Res. 50: 421-425 (1990); Kern и др., Cancer Res. 50: 5184 (1990); Park и др., Cancer Res. 49: 6605 (1989); Zhau и др., Mol. Carcinog., 3: 354-357 (1990); Aasland и др., Br.J.Cancer, 57: 358-363 (1989); Williams и др., Pathiobiology, 59: 46-52 (1991) и McCann и др., Cancer, 65: 88-92 (1990)).
Сверхэкспрессия ErbB2 может наблюдаться при раке предстательной железы (Gu и др., Cancer Lett., 99: 185-189 (1996); Ross и др., Hum. Pathol., 28: 827-833 (1997); Ross и др., Cancer, 79: 2162-2170 (1997) и Sadasivan и др., J.Urol. 150: 126-131 (1993)).
Обнаружены антитела к протеиновым продуктам p185neu крыс и человеческого ErbB2. Drebin с коллегами получили антитела к продукту гена neu крыс, т.е. p185neu (см., например, Drebin и др., Cell, 41: 695-706 (1985); Myers и др., Meth. Enzym., 198: 277-290 (1991) и WO 94/22487. Drebin с коллегами в Oncogene, 2: 273-277 (1988) описали, что для смесей антител, реактивных в отношении двух различных областей p185neu, обнаружено синергетическое противоопухолевое действие в отношении трансформированных neu клеток линии NIH-3T3, которые имплантировали бестимусным мышам (см. также патент 5824311, опубликованный 20 октября 1998 г.).
Hudziak и др., Mol. Cell. Biol., 9(3): 1165-1172 (1985) описали метод получения панели антител к ErbB2, основанный на использовании человеческой линии клеток рака молочной железы SK-BR-3. Относительную пролиферацию клеток SK-BR-3 после обработки антителами оценивали с помощью окрашивания монослоев фиолетовым кристаллическим через 72 ч. В этом анализе максимальное ингибирование было получено при использовании антитела, обозначенного как 4D5, которое ингибирует пролиферацию клеток на 56%. В этом анализе другие антитела панели понижали пролиферацию клеток в меньшей степени. Для антитела 4D5 также установлено, что оно сенсибилизирует сверхэкспрессирущие ErbB2 линии клеток опухоли молочной железы в отношении цитотоксических воздействий TNF-α (см. также патент США 5677171, опубликованный 14 октября 1997 г.). Антитела к ErbB2, описанные Hudziak с коллегами, дополнительно охарактеризованы Fendy и др., Cancer Research, 50: 1550-1558 (1990); Kotts и др., In Vitro, 26(3): 59A (1990); Sarup и др., Growth Regulation, 1: 72-83 (1991); Shepard и др., J.Clin. Immunol., 11(3): 117-127 (1991); Kumar и др., Mol. Cell. Biol., 11(2): 979-986 (1991); Lewis и др., Cancer Immunol. Immunother., 37: 255-263 (1993); Pietras и др., Oncogene, 9: 1829-1838 (1994); Vitetta и др., Cancer Research, 54: 5301-5309 (1994); Sliwkowski и др., J.Biol.Chem., 269(20): 14661-14665 (1994); Scott и др., J.Biol. Chem., 266(20): 14300-14305 (1991); D'souza и др., Proc. Natl. Acad. Sci. USA, 91: 7202-7206 (1994); Lewis и др., Cancer Research, 56: 1457-1465 (1996) и Schaefer и др., Oncogene, 15: 1385-1394 (1997)).
Рекомбинантная гуманизированная версия мышиного антитела к ErbB2 4D5 (huMab4D5-8, rhuMab Her2 или HERCEPTIN®; патент США 5821377) обладает клинической активностью для пациентов с метастатическими опухолями молочной железы, которые характеризуются сверхэкспрессией ErbB2, при ее интенсивном применении перед противоопухолевой терапией (Baselga и др., J.Clin. Oncol., 14: 737-744 (1996)). Управление по контролю над качеством пищевых продуктов, медикаментов и косметических средств 25 сентября 1998 г. разрешило применение антитела, имеющего товарный знак HERCEPTIN®, для лечения пациентов, страдающих метастатическим раком молочной железы, в случае, когда в опухоли происходит сверхэкспрессия протеина ErbB2.
Другие антитела к ErbB2, обладающие различными свойствами, описаны у Tagliabue и др., Int. J.Cancer, 4: 933-937 (1991); McKenzie и др., Oncogene, 4: 543-548 (1990); Stancovski и др., PNAS (USA), 88: 8691-8695 (1991); Bacus и др., Cancer Research, 52: 2580-2589 (1992); Xu и др., Int. J.Cancer, 53: 401-408 (1993); WO 94/00136; Kasprzyk и др., Cancer Research, 52: 2771-2776 (1992); Hancock и др., Cancer Research, 51: 4575-4580 (1991); Shawver и др., Cancer Research, 54: 1367-1373 (1994); Arteaga и др., Cancer Research. 54: 3758-3765 (1994); Harwerth и др., J.Biol. Chem., 267: 15160-15167 (1992); патент США 5783167 и Klapper и др., Oncogene, 14: 2009-2109(1997).
Скрининг в отношении гомологии позволил идентифицировать два представителя семейства ЕrB2-рецепторов: ЕrВ3 (патенты США 5183884 и 5480968, а также Kraus и др., PNAS (USA), 86: 9193-9197 (1989)) и ErbB4 (EP-A 599274; Plowman и др., Proc. Natl. Acad. Sci., USA 90: 1746-1750 (1993) и Plowman и др., Nature, 366: 473-475 (1993)). Повышенный уровень экспрессии обоих этих рецепторов обнаружен по меньшей мере в некоторых линиях клеток рака молочной железы.
ErbB-рецепторы как правило присутствуют в клетках в различных комбинациях, и гетеродимеризация, вероятно, способствует повышению разнообразия клеточных ответов на различные лиганды ErbB (Earp и др., Breast Cancer Research and Treatment, 35: 115-132 (1995)). С EGFR связывается шесть различных лигандов: эпидермальный фактор роста (EGF), трансформирующий фактор роста альфа (TGF-α), амфирегулин, гепаринсвязывающий эпидермальный фактор роста (HB-EGF), бетацеллулин и эпирегулин (Groenen и др., Growth Factors, 11: 235-257 (1994)). Представители семейства протеинов херегулинов, полученных в результате альтернативного сплайсинга одного гена, являются лигандами для ErbB3 и ErbB4. Семейство херегулинов включает альфа-, бета- и гамма-херегулины (Holmes и др., Science, 256: 1205-1210 (1992); патент 5641869 и Schaefer и др., Oncogene, 15: 1385-1394 (1997); neu-факторы дифференцировки (NGF), глиальные факторы роста (GGF); индукторы активности ацетилхолинового рецептора (ARIA) и факторы, происходящие из сенсорных и моторных нейронов (SMDF) (см. обзор Groenen и др., Growth Factors, 11: 235-257 (1994); Lemke G. Molec. & Cell Neurosci, 7: 247-262 (1996) и Lee и др., Pharm. Rev. 47: 51-85 (1995)). В настоящее время обнаружены три дополнительных лиганда ErbB: неурегулин-2 (NRG-2), который связывается либо с ErbB3, либо с ErbB4 (Chang и др., Nature, 387: 509-512 (1997); и Carraway и др., Nature, 387: 512-516 (1997); неурегулин-3, который связывается с ErbB4 (Zhang и др., PNAS (USA), 94(18): 9562-9567 (1997)), и неурегулин-4, который связывается с ErbB4 (Harari и др., Oncogene, 18: 2681-2689 (1999)). HB-EGF, бетацеллулин и эпирегулин также связываются с ErbB4.
Несмотря на то что EGF и TGF-α не связываются с ErbB2, EGF стимулирует EGFR и ErbB2 к образованию гетеродимеров, которые активируют EGFR и в результате приводят к трансфосфорилированию ErbB2 в гетеродимере. Димеризация и/или трансфосфорилирование, вероятно, активирует тирозинкиназу ErbB2 (см. Earp и др., выше). Аналогично этому, когда ErbB3 совместно экспрессируется с ErbB2, образуется активный сигнальный комплекс, и антитела к ErbB2 обладают способностью разрушать этот комплекс (Sliwkowski и др., J.Biol. Chem., 269(20): 14661-14665 (1994)). Кроме того, аффинность ErbB3 к херегулину (HRG) повышается и достигает более высокого уровня аффинности при совместной экспрессии с ErbB2 (см. данные Levi и др., Journal of Neuroscience, 15: 1329-1340 (1995); Morrissey и др., Proc. Natl. Acad. Sci. USA, 92: 1431-1435 (1995) и Lewis и др., Cancer Res., 56: 1457-1465 (1996)), касающиеся комплекса протеинов ErbB2-ErbB3. ErbB4, аналогично ErbB3, образует активный сигнальный комплекс с ErbB2 (Carraway и Cantley, Cell, 78: 5-8(1994)).
Краткое изложение сущности изобретения
Одном из объектов изобретения является способ лечения рака у человека, при котором раковые клетки экспрессируют рецептор эпидермального фактора роста (EGFR), предусматривающий введение человеку терапевтически эффективного количества антитела, которое связывается с ErbB2.
Далее указаны различные преимущества применения антитела, которое связывается с ErbB2, для лечения таких видов рака по сравнению с лекарственными средствами, мишенью которых является EGFR. В частности, для EGFR характерен высокий уровень экспрессии в печени и коже, что приводит к очень значительному снижению концентрации активного лекарственного средства в результате его связывания с EGFR. Кроме того, для других лекарственных средств, мишенью которых является EGFR, таких как химерное антитело к EGFR C225 и низкомолекулярное лекарственное средство ZD1839, которое связывается с EGFR, характерна токсичность для кожи. Предполагается, что антитела, которые связываются с ErbB2, обладают улучшенным профилем безопасности по сравнению с такими лекарственными средствами.
В случае, когда применяемое для терапии согласно изобретению антитело блокирует активацию лигандом рецептора ErbB и/или имеет биологические характеристики моноклонального антитела 2С4, могут достигаться дополнительные преимущества. Например, в то время как лекарственные средства, мишенью которых является EGFR, взаимодействуют только с EGFR, предпочтительные антитела по изобретению (например 2С4, включая его гуманизированные варианты и/или созревшие варианты с выраженной аффинностью) могут взаимодействовать с гетеродимерами EGFR/ErbB2, ErbB3/ErbB4 и ErbB2/ErbB3. Кроме того, антитела по изобретению, которые связываются с ErbB2 и блокируют активацию лигандом рецептора ErbB, могут дополнять лекарственные средства, мишенью которых является EGFR, в то время как лекарственные средства, мишенью которых является EGFR, не могут дополнять друг друга.
Объектом изобретения также является способ лечения рака у человека, ghb при котором для раковых клеток не характерна сверхэкспрессия рецептора ErbB2, предусматривающий введение человеку терапевтически эффективного количества антитела, которое связывается с ErbB2 и блокирует активацию лигандом рецептора ErbB.
Кроме того, объектом изобретения также является способ лечения независящего от гормонов рака у человека, предусматривающий введение человеку терапевтически эффективного количества антитела, которое связывается с ErbB2 и блокирует активацию лигандом рецептора ErbB.
Кроме того, объектом изобретения также является способ лечения рака у человека, предусматривающий введение человеку терапевтически эффективных количеств (а) первого антитела, которое связывает ErbB2 и ингибирует рост раковых клеток, сверхэкспрессирующих ErbB2, и (б) второго антитела, которое связывается с ErbB2 и блокирует активацию лигандом рецептора ErbB.
Объектом изобретения также является способ лечения рака у человека, где рак выбирают из группы, включающей рак ободочной кишки, рак прямой кишки и колоректальный рак, предусматривающий введение человеку терапевтически эффективного количества антитела, которое связывает ErbB2 и блокирует активацию лигандом рецептора ErbB.
Другими объектами изобретения (среди прочего) являются формы для применения согласно вышеуказанным способам. Например, изобретение относится к формам для применения, включающим контейнер и содержащуюся в нем композицию, где композиция включает антитело, которое связывает ErbB2, а также вкладыш, на котором указано, что композиция может применяться для лечения рака, при котором происходит экспрессия эпидермального фактора роста (EGFR).
Изобретение также относится к форме для применения, которая включает контейнер и содержащуюся в нем композицию, где композиция включает антитело, которое связывает ErbB2 и блокирует активацию лигандом рецептора ErbB, а также вкладыш, на котором указано, что композиция может применяться для лечения рака, при котором не происходит сверхэкспрессия рецептора ErbB2.
Изобретение также относится к форме для применения, которая включает контейнер и содержащуюся в нем композицию, где композиция включает антитело, которое связывает ErbB2 и блокирует активацию лигандом рецептора ErbB, а также вкладыш, на котором указано, что композиция может применяться для лечения независящего от гормонов рака.
И еще одним объектом изобретения является форма для применения, которая включает (а) первый контейнер с содержащейся в нем композицией, которая включает первое антитело, которое связывает ErbB2 и ингибирует рост раковых клеток, сверхэкспрессирующих ErbB2, и (б) второй контейнер с содержащейся в нем композицией, которая включает второе антитело, которое связывает ErbB2 и блокирует активацию лигандом рецептора ErbB.
Еще одним объектом изобретения является форма для применения, которая включает контейнер и содержащуюся в нем композицию, где композиция включает антитело, которое связывает ErbB2 и блокирует активацию лигандом рецептора ErbB, а также вкладыш, на котором указано, что композиция может применяться для лечения рака, где рак выбирают из группы, включающей рак ободочной кишки, рак прямой кишки и колоректальный рак.
Объектами изобретения также являются: гуманизированное антитело, которое связывает ErbB2 и блокирует активацию лигандом рецептора ErbB; композиция, включающая гуманизированное антитело и фармацевтически приемлемый носитель; и иммуноконъюгат, представляющий собой гуманизированное антитело, конъюгированное с цитотоксическим агентом.
Кроме того, изобретение относится к выделенной нуклеиновой кислоте, кодирующей гуманизированное антитело; вектору, включающему нуклеиновую кислоту; клетке-хозяину, включающей нуклеиновую кислоту или вектор; а также к способу получения гуманизированного антитела, предусматривающему такое культивирование клетки-хозяина, которая включает нуклеиновую кислоту, при котором происходит экспрессия нуклеиновой кислоты, и необязательно также предусматривающему выделение гуманизированного антитела из культуры клетки-хозяина (например, из культуральной среды клетки-хозяина).
Изобретение также относится к иммуноконъюгату, представляющему собой антитело, которое связывает ErbB2, конъюгированное с одной или несколькими молекулами калихеамицина, и применению таких конъюгатов для лечения рака, при котором происходит экспрессия ErbB2, например рака, при котором происходит сверхэкспрессия ErbB2, у человека. Предпочтительно антитело в конъюгате представляет собой моноклональное антитело 4D5, например гуманизированное антитело 4D5 (и предпочтительно huMab4D5-8 (HERCEPTIN®); или моноклональнок антитело 2С4, например, гуманизированное антитело 2С4). Антитело в конъюгате может представлять собой интактное антитело (например, интактное антитело в виде IgG1) или фрагмент антитела (например, Fab, F(ab)2, двойное антитело и т.д.).
Краткое описание чертежей
На фиг.1А и 1Б представлены результаты картирования эпитопов, представляющих собой остатки 22-645 во внеклеточном домене (ECD) ErbB2 (аминокислотная последовательность, включающая сигнальную последовательность, представлена на фиг.1А; SEQ ID NO: 13), проведенного с помощью анализа укороченных мутантов и сайтнаправленного мутагеназа (Nakamura и др., J. of Virology, 67(10): 6179-6191 (1993) и Renz и др. J.Cell Biol, 125(6): 1395-1406 (1994). Различные укороченные версии ECD ErbB2 или точковые мутации получали из кДНК с помощью метода полимеразной цепной реакции. Мутанты ErbB2 экспрессировали в виде слитых протеинов gD в экспрессионной плазмиде млекопитающих. В этой экспрессионной плазмиде используют промотор цитомегаловируса/энхансер с терминатором SV40 и сигналы полиаденирования, локализованные по ходу транскрипции относительно встроенной кДНК. Плазмидной ДНК трансфектировали клетки линии 293. Через 1 день после трансфекции клетки метаболически метили в течение ночи в несодержащей метионин и цистеин среде DMEM с низким содержанием глюкозы, включающей 1% подвергнутой диализу фетальной телячьей сыворотки и по 25 мкКи 35S-метионина и 35S-цистеина. Собирали супернатанты и к супернатанту добавляли либо моноклональные антитела к ErbB2, либо контрольные антитела и инкубировали в течение 2-4 ч при 4°С. Комплексы осаждали, вносили в градиентный гель, содержащий 10-20% трицина-ДСН, и подвергали электрофорезу при 100 В. Гель подвергали электроблоттингу на мембране и анализировали с помощью авторадиографии. Как видно из фиг.1Б антитела к ErbB2 7С2, 7F3, 2C4, 7D3, 3Е8, 4D5, 2Н11 и 3Н4 обладают способностью связывать различные эпитопы ECD ErbB2.
На фиг.2А и 2Б представлены данные о воздействии моноклональных антител к ErbB2 2C4 и 7F3 на активацию rHRGβ1 клеток линии MCF7. На фиг.2А приведены графики зависимости реакции от дозы для ингибирования с помощью 2C4 и 7F3 стимулированного HRG фосфорилирования тирозина. На фиг.2Б приведены графики зависимости реакции от дозы для ингибирования с помощью 2C4 и 7F3 связывания 125I-rHRGβ1177-244 с клетками линии MCF7.
На фиг.3 представлены данные о ингибировании специфического связывания 125I-rHRGβ1177-244 с панелью линий человеческих клеток опухолей с помощью моноклональных антител к ErbB2 2C4 или 7F3. Моноклональные антитела, применяемые в качестве контроля, представляют собой изотипические меченые мышиные антитела, которые не блокируют связывание rHRG. Неспецифическое связывание 125I-rHRGβ1177-244 определяли из параллельных опытов, в которых инкубации осуществляли в присутствии 100 нМ rHRGβ1. Для всех изученных линий клеток неспецифическое связывание 125I-rHRGβ1177-244 составляло менее 1% от общего связывания.
На фиг.4А и 4Б представлены данные о воздействии моноклональных антител 2C4 и 4D5 на пролиферацию клеток линии MDA-MB-175 (фиг.4А) и SK-BR-3 (фиг.4Б). Клетки MDA-MB-175 и SK-BR-3 высевали в 96-луночные планшеты и давали им прилипнуть в течение 2 ч. Эксперимент осуществляли в среде, содержащей 1% сыворотки. Добавляли антитела к ErbB2 или только среду и клетки инкубировали в течение 2 ч при 37°С. Затем добавляли rHRGβl (1 нМ) или только среду и клетки инкубировали в течение 4 дней. Монослои промывали и окрашивали/фиксировали с помощью 0,5%-ного кристаллического фиолетового. Для оценки пролиферации клеток измеряли абсорбцию при 540 нм.
На фиг.5А и 5Б представлены данные о воздействии моноклонального антитела 2C4, антитела HERCEPTIN® или антитела к EGFR на зависящее от херегулина (HRG) связывание ErbB2 с ErbB3 в клетках линии MCF7, экспрессирующих низкие/нормальные уровни ErbB2 (фиг.5А), и в клетках линии SK-BR-3, экспрессирующих высокие уровни ErbB2 (фиг.5Б); см. пример 2 ниже.
На фиг.6А и 6Б приведено сравнение активностей интактного мышиного моноклонального антитела 2С4 (mu 2С4) и химерного Fab-фрагмента 2С4. На фиг.6А представлено ингибирование связывания 125I-HRG с клетками линии MCF7 с помощью химерного Fab-фрагмента 2С4 или интактного мышиного моноклонального антитела 2С4. Клетки линии MCF7 высевали в 24-луночные планшеты (1·105 клеток/лунку) и выращивали до примерно 85%-ной конфлюэнции в течение 2 дней. Эксперименты по связыванию осуществляли согласно методу, описанному у Lewis и др. Cancer Research, 56: 1457-1465 (1996). На фиг.6Б представлены данные об ингибировании активации с помощью rHRGβ1 фосфорилирования тирозина р180 в клеткх линии MCF7, что осуществляли согласно методу, описанному у Lewis и др., Cancer Research, 56: 1457-1465 (1996).
На фиг.7А и 7Б представлен сравнительный анализ аминокислотных последовательностей вариабельных областей легких (VL) (фиг.7А) и вариабельных областей тяжелых (VH) (фиг.7Б) цепей мышиного моноклонального антитела 2С4 (SEQ ID NO:1 и 2 соответственно); VL- и VH-областей гуманизированной версии 574 2С4 (SEQ ID NO:3 и 4 соответственно); и человеческих консенсусных каркасных участков (hum k1, т.е. подгруппа I легкой каппа-цепи; humIII, т.е. подгруппы III тяжелой цепи) (SEQ ID NO:5 и 6 соответственно). Звездочками обозначены различия между каркасными участками гуманизированной версии 574 2С4 и мышиным моноклональным антителом 2С4 или между гуманизированной версией 574 2С4 и человеческим каркасным участком. Гиперварибельные участки (CDR) заключены в скобки.
На фиг.8А-8В представлены данные о связывании химерного Fab-фрагмента 2С4 (Fab v1) и нескольких гуманизированных вариантов антитела 2С4 к внеклеточному домену (ECD) ErbB2, которое определяли с помощью метода ELISA, описанного в примере 3.
На фиг.9 приведена ленточная диаграмма VL- и VH-областей моноклонального антитела 2С4, где белым цветом обозначены каркасные области CDR (L1, L2, L3, H1, Н2, Н3). Также обозначены боковые цепи VH-области, которые выявляли с помощью мутагенеза в процессе гуманизации (см. пример 3, таблица 2).
На фиг.10 представлены данные о воздействии моноклонального антитела 2С4 или HERCEPTIN® на опосредуемую EGF, TGF-α или HRG активацию активируемой митогеном протеинкиназы (МАРК).
На фиг.11 представлена столбчатая диаграмма, на которой показано воздействие антител к ErbB2 (по отдельности или в комбинации) на ксенотрасплантаты (трехкратный уровень (3+) сверхэкспрессии ErbB2) аденокарциномы легкого линии Саlu3. Примечание: обработку прекращали на 24-й день.
На фиг.12 представлены данные о воздействии рекомбинантного гуманизированного моноклонального антитела 2С4 (rhuMAb 2C4) или HERCEPTIN® на рост клеток линии MDA-175 при оценке анализом с использованием Alamar Blue (аламарового синего).
На фиг.13 представлены данные об эффективности rhuMAb 2C4 в отношении ксенотрансплантатов MCF7.
Подробное описание предпочтительных вариантов осуществления
I. Определения
Понятие "рецептор ErbB" обозначает рецептор протеина тирозинкиназы, который принадлежит к семейству ErbB-рецепторов и включает рецепторы EGFR, ErbB2, ErbB3 и ErbB4, в будущем следует ожидать обнаружение других представителей этого семейства. Рецептор ErbB, как правило, включает внеклеточный домен, который может связывать лиганд ErbB; липофильный трансмембранный домен; консервативный внутриклеточный тирозинкиназный домен; С-концевой сигнальный домен, несущий несколько остатков тирозина, которые могут быть фосфорилированы. Рецептор ErbB может иметь "нативную последовательность" рецептора ErbB или "вариант его аминокислотной последовательности". Предпочтительно рецептор ErbB имеет нативную последовательность человеческого рецептора ErbB.
Понятия "ErbB1", "рецептор эпидермального фактора роста" и "EGFR" в контексте настоящего описания используются взаимозаменяемо и обозначают EGFR, например, у Carpenter и др., Ann. Rev. Biochem., 56: 881-914 (1987), включая его встречающиеся в естественных условиях мутантные формы (например, полученный в результате делеции мутант EGFR, описанный у Humphrey и др., PNAS (USA), 87: 4207-4211 (1990). Понятие "erbB1" обозначает ген, кодирующий протеин EGFR.
Понятия "ErbB2" и "HER2" в контексте настоящего описания используются взаимозаменяемо и обозначают человеческий протеин HER2, описанный, например, у Semba и др., PNAS (USA), 82: 6497-6501 (1985) и у Yamamoto и др., Nature, 319: 230-234 (1986) (регистрационный номер Genebank X03363). Понятие "erbB2" обозначает ген, кодирующий человеческий ErbB2, а "neu" обозначает ген, кодирующий крысиный p185neu. Предпочтительно ErbB2 имеет нативную последовательность человеческого ErbB2.
Понятия "ErbB3" и "HER3" обозначают рецепторный полипептид, который описан, например, в патентах США 5183884 и 5480968, а также у Kraus и др., PNAS (USA), 86: 9193-9197 (1989).
Понятия "ErbB4" и "HER4" обозначают рецепторный полипептид, который описан, например, в ЕР-А 599274; у Plowman и др., Proc. Natl. Acad. Sci. USA, 90: 1746-1750 (1993) и у Plowman и др., Nature, 366: 473-475 (1993), включая его изоформы, описанные, например, в WO 99/19488, которая опубликована 22 апреля 1999 г.
Понятие "лиганд ErbB" обозначает полипептид, который связывается с рецептором ErbB и/или активирует его. Представляющий особенный интерес лиганд ErbB представляет собой лиганд, имеющий нативную последовательность человеческого лиганда ErbB, такого как эпидермальный фактор роста (EGF) (Savage и др., J.Biol. Chem., 247: 7612-7621 (1972)); трансформирующий фактор роста альфа (TGF-α) (Marquardt и др., Science, 223: 1079-1082) (1984); амфирегулин, также известный как шваннома или кератиноцитный аутокринный фактор роста (Shoyab и др., Science, 243: 1074-1076 (1989); Kimura и др., Nature, 348: 257-260 (1990); и Cook и др., Mol. Cell. Biol., 11: 2547-2557 (1991); бетацеллулин (Shing и др., Science, 259: 1064-1607 (1990) и Sasada и др., Biochem. Biophys. Res. Commun., 190: 1173 (1993); гепаринсвязывающий эпидермальный фактор роста (HB-EGF) (Higashiyama и др., Science, 251: 936-939 (1991); эпирегулин (Toyoda и др., J. Biol. Chem., 270: 7495-7500 (1995) и Komurasaki и др., Oncogene, 15: 2841-2848 (1997)); херегулин (см. ниже); неурегулин-2 (NRG-2) (Carraway и др., Nature, 387: 512-516 (1997); неурегулин-3 (NRG-3) (Zhang и др., Proc. Natl. Acad. Sci. USA, 94: 9562-9567 (1997)); неурегулин-4 (NRG-4) (Harari и др., Oncogene, 18: 2681-2689 (1999) или крипто (CR-1) (Kannan и др., J.Biol. Chem., 272(6): 3330-3335 (1997)). Лиганды ErbB, которые связываются с EGFR, включают EGF, TGF-α, амфирегулин, бетацеллулин, HB-EGF и эпирегулин. Лиганды ErbB, которые могут связываться с ErbB3, включают херегулины. Лиганды ErbB, которые могут связываться с ErbB4, включают бетацеллулин, эпирегулин, НВ-EGF, NRG-2, NRG-3, NRG-4 и херегулины.
Понятие "херегулин" (HRG) в контексте настоящего описания обозначает полипетид, кодируемый херегулиновым генным продуктом, как он описан в патенте США 5641869 или у Marchionni., Nature, 362: 312-318 (1993). Примеры херегулинов включают херегулин-α, херегулин-β1, херегулин-β2 и херегулин-β3 (Holmes и др., Science, 256: 1205-1210 (1992) и патент США 5641869); neu-фактор дифференцировки (NGF) (Peles и др., Cell, 69: 205-216 (1992); индукторы активности ацетилхолинового рецептора (ARIA) (Fals и др., Cell, 72: 801-815 (1993)); глиальные факторы роста (GGF) (Marchionni и др. Nature, 362: 312-318 (1993)); факторы, происходящие из сенсорных и моторных нейронов (SMDF) (Но и др., J.Biol. Chem., 270: 14523-14532 (1995)); γ-херегулин (Schaefer и др., Oncogene, 15: 1385-1394 (1997)). Понятие включает биологически активные фрагменты и/или варианты аминокислотной последовательности нативной последовательности полипептида HRG, такие как фрагмент его EGF-подобного домена (например, HRGβ1177-244).
Понятие "гетероолигомер ErbB" в контексте настоящего описания обозначает нековалентно связанный олигомер, включающий, по меньшей мере, два различных рецептора ErbB. Такие комплексы могут образовываться, когда клетку, экспрессирующую два или большее количество рецепторов ErbB, обрабатывают лигандом ErbB, и они могут быть выделены путем иммунопреципитации и проанализированы с помощью ДСН-ПААГ согласно методике, например, описанной у Silwkowski и др., J. Biol. Chem., 269 (20): 14661-14665 (1995). Примеры таких гетероолигомеров ErbB включают комплексы EGFR-ErbB2, ErbB2-ErbB3 и ErbB3-ErbB4. Кроме того, гетероолигомер ErbB может включать два или большее количество рецепторов ErbB2, объединенных с другим рецептором ErbB, таким как ErbB3, ErbB4 или EGFR. В гетероолигомер могут быть включены другие протеины, такие как субъединица рецептора цитокина (например, gp130).
Понятие "активация лигандом рецептора ErbB" обозначает трансдукцию сигнала (например, вызываемую фосфорилированием остатков тирозина внутриклеточного киназного домена рецептора ErbB или субстратного полипептида), которая опосредована связыванием лиганда ErbB с гетероолигомером ErbB, включающим представляющий интерес рецептор ErbB. Как правило, это может включать связывание лиганда ErbB с гетероолигомером ErbB, что активирует киназный домен одного или нескольких рецепторов ErbB в гетероолигомере и тем самым приводит к фосфорилированию остатков тирозина в одном или нескольких рецепторах ErbB и/или к фосфорилированию остатков тирозина в дополнительном(ых) субстратном(ых) полипептиде(ах). Активация рецептора ErbB может быть количественно оценена с помощью различных методов анализа фосфорилирования тирозина.
Понятие полипептид, имеющий "нативную последовательность", бозначает полипептид, который характеризуется такой же аминокислотной последовательностью, что и полипептид (например, рецептор ErbB или лиганд ErbB), имеющий естественное происхождение. Такие полипептиды, имеющие нативные последовательности, могут иметь естественное происхождение или быть получены рекомбинантным или синтетическим путями. Так, полипептид с нативной последовательностью может иметь аминокислотную последовательность встречающегося в естественных условиях человеческого полипептида, мышиного полипептида или полипептида, выделенного из любых других видов млекопитающих.
Понятие "вариант аминокислотной последовательности" относится к полипептидам аминокислотные последовательности, которых отличаются в определенной степени от нативной последовательности полипептида. Как правило, варианты аминокислотной последовательности должны быть, по меньшей мере, примерно на 70% гомологичны, по меньшей мере, с одним связывающимся с рецептором доменом нативного лиганда ErbB или, по меньшей мере, с одним связывающимся с лигандом доменом нативного рецептора ErbB, и предпочтительно они должны быть, по меньшей мере, примерно на 80%, более предпочтительно они должны быть, по меньшей мере, примерно на 90% гомологичны таким связывающимся с рецептором или лигандом доменам. Варианты аминокислотной последовательности несут замены, делеции и/или инсерции в определенных положениях в аминокислотной последовательности нативной аминокислотной последовательности.
"Гомологию" выражают в виде процента остатков в варианте аминокислотной последовательности, которые оказались идентичными после сравнительного анализа последовательностей и введения при необходимости брешей для достижения максимального процента гомологии. Методы и компьютерные программы для сравнительного анализа последовательностей хорошо известны в данной области. Одна из таких компьютерных программ представляет собой "Align 2", разработанную на фирме Genetech Inc., которая передана вместе с документацией для пользователя в United States Copyright Office, Washington, DC 20559, 10 декабря 1991 г.
Понятие "антитело" в контексте настоящего описания используется в наиболее широком смысле и, в частности, относится к интактным моноклональным антителам, поликлональным антителам, мультиспецифическим антителам (например, биспецифическим антителам), образованным по меньшей мере из двух интактных антител, и фрагментам антител, в том случае, если они обладают требуемой биологической активностью.
Понятие "моноклональное антитело" в контексте настоящего описания обозначает антитело, полученное из популяции практически гомогенных антител, т.е. входящие в популяцию индивидуальные антитела идентичны за исключением возможных встречающихся в естественных условиях мутаций, которые могут присутствовать в небольших количествах. Моноклональные антитела являются высокоспецифическими и представляют собой антитела к отдельному сайту антигена. Кроме того, в противоположность препаратам поликлональных антител, которые включают различные антитела к различным детерминантам (эпитопам), каждое моноклональное антитело направлено к одной детерминанте антигена. Помимо их специфичности преимуществом моноклональных антител является то, что они могут быть синтезированы без загрязнения другими антителами. Прилагательное "моноклональное" свидетельствует о том, что антитело получено из практически гомогенной популяции антител, при этом не предполагается, что оно должно быть получено с помощью какого-либо конкретного метода. Например, моноклональные антитела, которые могут применяться согласно изобретению, могут быть получены с помощью метода на основе гибридом, впервые описанного Kohler и др., Nature, 256: 495 (1975), или могут быть сконструированы методами рекомбинатной ДНК (см., патент США 4816567). "Моноклональные антитела" могут быть выделены также из фаговых библиотек антител с помощью методик, описанных, например, у Clackson и др., Nature, 352: 624-628 (1991) и у Marks и др., J.Mol. Biol., 222: 581-597 (1991).
Моноклональные антитела в контексте настоящего описания включают, в частности, "химерные" антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям антител, полученным из конкретных видов, или принадлежит к конкретным классам или подклассам антител, а остальной участок цепи(ей) идентичен или гомологичен соответствующим последовательностям антител, полученным из других видов, или принадлежит к другим классам или подклассам антител, включая фрагменты таких антител, в случае, если они обладают требуемой биологической активностью (патент США 4816567; и Morrison и др., Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984)). Представляющие интерес в контексте настоящего описания химерные антитела включают "приматизированные" антитела, которые содержат антигенсвязывающие последовательности вариабельных областей, полученные из приматов, не включая людей (например, низших узконосых обезьян, обезьян и т.д.), и последовательности человеческих константных областей.
"Фрагмент антитела" включает часть интактного антитела, предпочтительно включает его антигенсвязывающую или вариабельную области. Примеры фрагментов антитела включают Fab-, Fab-', F(ab')2- и Fv-фрагменты; двойные антитела, линейные антитела; молекулы одноцепочечных антител; и мультиспецифические антитела, полученные из фрагмента(ов) антител.
"Интактное антитело" представляет собой антитело, которое содержит вариабельную антигенсвязывающую область, а также константную область легкой цепи (СL) и константные области тяжелой цепи Сн 1, Сн 2 и Сн 3. Константные области могут представлять собой константные области, имеющие нативные последовательности (например, человеческие константные области, имеющие нативные последовательности), или варианты их аминокислотных последовательностей. Предпочтительно интактное антитело обладает одной или несколькими эффекторными функциями.
Понятие "эффекторные функции" антитела относится к биологическим активностям, связанным с Fc-областью (нативной последовательностью Fc-области или с вариантом аминокислотной последовательности Fc-области) антитела. Примерами эффекторных функций антитела являются С1q-связывание; зависящая от комплемента цитотоксичность; связывание Fc-рецептора; опосредуемая антителами клеточная цитотоксичность (ADCC); фагоцитоз; понижающая регуляция рецепторов клеточной поверхности (например, В-клеточного рецептора, BCR) и т.д.
В зависимости от аминокислотной последовательности константной области их тяжелых цепей интактные антитела могут быть разделены на различные "классы". Известно 5 основных классов интактных антител: IgA, IgD, IgE, IgG и IgM, при этом некоторые из них могут быть подразделены на "подклассы" (изотипы): например, IgG1, IgG2, IgG3, IgG4, IgA и IgA2. Константные области тяжелой цепи, которые соответствуют различным классам антител, обозначают α, δ, ε, γ и μ соответственно. Строение субъединиц и трехмерная конфигурация различных классов иммуноглобулинов хорошо известны.
Понятия "опсредуемая антителом клеточная цитотоксичность" и "ADCC" обозначают опосредуемую клеткой реакцию, при которой неспецифические цитотоксические клетки, которые экспрессируют Fc-рецепторы (FcR) (например, естественные киллеры (NK-клетки), нейтрофилы и макрофаги) распознают связанное антитело на клетке-мишени и далее вызывают лизис клетки-мишени. Основные клетки, для которых характерна ADCC, т.е. NK-клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Данные о экспрессии FcR на поверхности гематопоэтических клеток обобщены в таблице 3 на стр. 464 статьи Ravetch и Kinet, Ann. Rev. Immunol., 9: 457-492 (1991). Для оценки ADCC-активности представляющей интерес молекулы может применяться ADCC-анализ in vitro, например, описанный в патентах США 5500362 или 5821337. Эффекторные клетки, которые могут применяться для таких анализов, включают мононуклеарные клетки периферической крови (РВМС) и естественные киллеры (NK-клетки). В другом или дополнительном варианте для оценки ADCC-активности представляющей интерес молекулы может применяться анализ in vivo, например, с использованием в качестве моделей животных согласно методу, описанному у Clynes и др., PNAS (USA), 95: 652-656 (1998).
"Человеческие Эффекторные клетки" представляют собой лейкоциты, которые экспрессируют один или несколько FcR и обладают эффекторными функциями. Предпочтительно клетки экспрессируют, по меньше мере, FcγRIII и обладают ADCC-эффекторным функциями. Примеры человеческих лейкоцитов, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (РВМС), естественные киллеры (NK-клетки), моноциты, цитотоксические Т-клетки и нейтрофилы; предпочтительными являются РВМС и NK-клетки. Эффекторные клетки могут быть выделены из их естественного источника, например, из крови или РВМС, согласно описанной ниже методике.
Понятия "Fc-рецептор" или "FcR" в контексте настоящего описания относятся к рецептору, который связывается с Fc-областью антитела. Предпочтительный FcR имеет нативную последовательность человеческого FcR. Кроме того, предпочтительный FcR представляет собой FcR, который связывается с антителом в виде IgG (гамма-рецептор) и включает подклассы FcγRI-, FcγRII- и FcγRIII-рецепторов, в том числе аллельные варианты и полученные в результате альтернативного сплайсинга формы этих рецепторов. FcγRII-рецепторы включают FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), которые имеют одинаковые аминокислотные последовательности, но прежде всего отличаются своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит в своем цитоплазматическом домене иммунорецепторный тирозинсодержащий активирующий мотив (ITAM). Ингибирующий рецептор FcγRIIB содержит в своем цитоплазматическом домене иммунорецепторный тирозинсодержащий ингибирующий мотив (ITIM) (обзор данных см. у М.Daeron, Ann. Rev. Immunol., 15: 203-243 (1997)). Данные о FcR обобщены у Ravetch и Kinet, Ann. Rev, Immunol., 9: 457-492 (1991); Capel и др., Immunomethods, 4: 25-34 (1994) и у de Haas и др., J.Lab. Clin. Med., 126: 330-341 (1995). Другие FcR, включая FcR, которые будут обнаружены в будущем, подпадают под указанное понятие "FcR". Понятие также включает неонатальный рецептор, FcRn, который ответствен за перенос материнских IgG в эмбрион (Guyer и др., J.Immunol., 117: 587 (1976)) и Kim и др., J.Immunol., 24: 249 (1994)).
Понятие "зависящая от комплемента цитотоксичность" или "CDC" обозначает способность молекулы вызывать лизис мишени в присутствии комплемента. Путь активации комплемента инициируется связыванием первого компонента системы комплемента (C1q) с молекулой (например, антителом), входящей в комплекс с родственным антигеном. Для оценки активации комплемента может применяться CDC-анализ, например, описанный у Gazzano-Santoro и др., J.Immunol. Methods, 202: 163(1996).
"Нативные антитела" обычно представляют собой гетеротетрамерные гликопротеины с молекулярной массой примерно 150000 Да, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь связана с тяжелой цепью посредством одной ковалентной дисульфиной связи, в то время как количество дисульфидных мостиков между тяжелыми цепями различных изотипов иммуноглобулинов варьируется. Каждая тяжелая и легкая цепь также включает правильно расположенные в пространстве дисульфидные мостики внутри цепей. Каждая тяжелая цепь имеет на одном конце вариабельную область (VH), за которой следуют несколько константных областей. Каждая легкая цепь имеет вариабельную область на одном конце (VL) и константную область на другом конце. Константная область легкой цепи сопоставима с первой константной областью тяжелой цепи, а вариабельная область легкой цепи сопоставима с вариабельной областью тяжелой цепи. Вероятно, определенные аминокислотные остатки образуют поверхность раздела между вариабельными областями легкой цепи и тяжелой цепи.
Понятие "вариабельный" относится к ситуации, когда определенные части вариабельных областей различных антител в значительной степени отличаются по их последовательности, и они используются для специфического связывания каждого конкретного антитела с конкретным антигеном. Однако вариабельность распределена неравномерно по вариабельным областям антител. Она сконцентрирована в трех сегментах, которые называют гипервариабельными участками в вариабельных областях как легкой цепи, так и тяжелой цепи. Наиболее консервативные участки вариабельных областей называют каркасными участками (FR). Каждая вариабельная область нативных легких и тяжелых цепей включает четыре FR, хорошо адаптированных к β-складчатой конформации, соединенные тремя гипервариабельными участками, которые образуют петлю, связывающую и в некоторых случаях образующую часть β-складчатой структуры. Гипервариабельные участки в каждой цепи поддерживаются в тесной связи друг с другом посредством FR и вместе с гипервариабельными участками другой цепи участвуют в образовании антигенсвязывающего центра антител (см. Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд. Public Health Service, National Institutes of Health, Bethesda, MD (1991)). Константные области не участвуют непосредственно в связывании антитела с антигеном, но они обладают различными эффекторными функциями, такими как участие антитела в опосредуемой антителом клеточной цитотоксичности (ADCC).
Понятие "гипервариабельный участок" в контексте настоящего описания обозначает аминокислотные остатки антитела, которые ответственны за связывание с антигеном. Гипервариабельный участок, как правило, включает аминокислотные остатки антитела из "гипервариабельной области" или "CDR" (например, остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельной области легкой цепи и 31-35 (H1), 50-65 (Н2) и 95-102 (Н3) вариабельной области тяжелой цепи (см. Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд. Public Health Service, National Institutes of Health, Bethesda, MD (1991)) и/или аминокислотные остатки из "гипервариабельной петли" (например, остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельной области легкой цепи и 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) вариабельной области тяжелой цепи (см. Chothia и Lesk, J.Mol. Biol., 196: 901-917 (1987)). Остатки "каркасного участка" или "FR" представляют собой такие остатки вариабельной области, которые отличны от остатков гипервариабельного участка, как он определен в настоящем описании.
При расщеплении антител папаином образуются два идентичных антигенсвязывающих фрагмента, которые называют "Fab"-фрагментами, каждый с антигенсвязывающим центром, и остается "Fc"-фрагмент, название которого отражает его способность легко кристаллизоваться. После обработки пепсином образуется F(ab')2-фрагмент, который несет два антигенсвязывающих центра и еще сохраняет способность к перекрестному связыванию с антигеном.
"Fv" обозначает минимальный фрагмент антитела, который содержит полный антигенраспознающий и антигенсвязывающий центр. Эта область состоит из димера, который включает одну вариабельную область тяжелой цепи и одну вариабельную область легкой цепи в тесной нековалентной связи. Именно в этой конфигурации три гипервариабельных участка каждой вариабельной области взаимодействуют с определенным антигенсвязывающим центром на поверхности VL-VH-димера. Всего шесть гипервариабельных участков обусловливают специфичность антитела в отношении связывания с антигеном. Однако даже одна вариабельная область (или половина Fv, включающая только три гипервариабельных участка, специфических для антигена) обладает способностью распознавать антиген и связываться с ним, хотя и с более низкой аффинностью по сравнению с полным связывающим сайтом.
Fab-фрагмент также содержит константную область легкой цепи и первую константную область (СН1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на С-конец СН1-области тяжелой цепи, включая один или несколько остатков цистеина из шарнирной области антитела. Fab'-SH в контексте настоящего описания обозначен как Fab'-фрагмент, в котором остаток(ки) цистеина константных областей несет(ут) по меньшей мере одну свободную тиольную группу. F(ab')2-фрагменты антитела в исходном состоянии получают в виде пары Fab'-фрагментов, между которыми расположены остатки цистеина шарнирной области. Также известны другие химические связи фрагментов антител.
"Легкие цепи" антител любых видов позвоночных могут быть отнесены к одному из двух хорошо различимых типов, называемых каппа (κ) и лямбда (λ) на основе аминокислотных последовательностей их константных областей.
"Одноцепочечный Fv-" или "scFv"-фрагменты антитела включают" VL- и VH-области антитела, где эти области присутствуют в виде одноцепочечной полипептидной цепи. Предпочтительно Fv-полипептид дополнительно включает полипептидный линкер между VL- и VH-областями, который придает scFv-фрагменту строение, требуемое для связывания с антигеном. Обзор данных, касающихся scFv, см. у Pluckthin в: The Pharmacology of Monoclonal Antibodies, том 113, ред-ры Rosenbrug и Moore, Springer-Verlag, New York, стр.269-315 (1994). scFv-фрагменты антитела к ErbB2 описаны в WO 93/16185; патенте США 5571894 и патенте США 5587458.
Понятие "двойные антитела" относится к небольшим фрагментам антител с двумя антигенсвязывающими центрами, эти фрагменты содержат вариабельную область тяжелой цепи (VH), связанную с вариабельной областью легкой цепи (VL) в одной и той же полипептидной цепи (VH-VL). С помощью линкера, который является слишком коротким, что позволить происходить спариванию между двумя областями на одной цепи, происходит принудительное спаривание областей с комплементарными областями другой цепи, и создаются два антигенсвязывающих центра. Двойные антитела более подробно описаны, например, в ЕР 404097; WO 93/11161 и у Hollinger и др. Proc. Natl. Acad. Sci, USA 90: 6444-6448 (1993).
"Гуманизированные" формы антител животных кроме человека (например, грызунов) представляют собой химерные антитела, которые включают минимальную последовательность, полученную из иммуноглобулина животного кроме человека. Основная часть гуманизированных антител представлена человеческими иммуноглобулинами (антитело-реципиент), в которых остатки из гипервариабельной области реципиента заменены остатками из гипервариабельной области других видов кроме человека (антитело-донор), таких как мышь, крыса, кролик или приматы кроме человека, обладающими требуемой специфичностью, аффинностью и потенциалом. В некоторых случаях остатки каркасного участка (FR) человеческого иммуноглобулина заменяют соответствующими остатками из других видов кроме человека. Кроме того, гуманизированные антитела могут включать остатки, которые не присутствуют в антителе-реципиенте или в антителе-доноре. Эти модификации осуществляют с целью дополнительного усовершенствования характеристик антитела. В целом, гуманизированное антитело должно включать практически полностью, по меньшей мере, одну, а как правило две, вариабельные области, в которых все или практически все гипервариабельные петли соответствуют петлям указанных иммуноглобулинов других животных кроме человека, а все или практически все FR соответствуют последовательности иммуноглобулина человека. Гуманизированное антитело необязательно также может включать, по меньшей мере, часть константной области иммуноглобулина (Fc), как правило, человеческого иммуноглобулина. Дополнительные более подробные данные см. у Jones и др., Nature, 321: 522-525 (1986); Reichmann и др., Nature, 332: 323-329 (1988) и Presta, Curr. Op. Struct. Biol, 2: 593-59 (1992).
Гуманизированные антитела к ErbB2 включают huMab4D5-1, huMab4D5-2, huMab4D5-3, huMab4D5-4, huMab4D5-5, huMab4D5-6, huMab4D5-7 и huMab4D5-8 (HERCEPTIN®), представленные в таблице 3 патента США 5821337, который полностью включен в настоящее описание в качестве ссылки; гуманизированное антитело 520С9 (WO 93/21319) и гуманизированное антитело 2С4, описанное далее.
"Выделенное" антитело представляет собой антитело, которое идентифицировано и отделено от и/или получено из компонента его естественного окружения. Загрязнителями в его естественном окружении являются материалы, которые могут оказывать воздействие при диагностическом или терапевтическом применении антитела, и они могут включать ферменты, гормоны и другие белковые или небелковые растворы. Предпочтительно антитело должно быть очищено (1) до более 95% в пересчете на массу антитела, что определяют методом Лоури, и наиболее предпочтительно до более 99 мас.%, (2) до степени, достаточной для получения, по меньшей мере, 15 N-концевых остатков или внутренней аминокислотной последовательности с помощью секвинатора с вращающейся чашкой, или (3) до гомогенного состояния по данным ДСН-ПААГ в восстанавливающих или невосстанавливающих условиях при окрашивании кумасси бриллиантовым голубым или предпочтительно при окрашивании серебром. Выделенные антитела включают антитела, находящиеся in situ в рекомбинантных клетках, если в них не присутствует ни один компонент естественного окружения антитела. Однако обычно выделенное антитело должно быть получено с использованием, по меньшей мере, одной стадии очистки.
Антитело, "которое связывается" с представляющим интерес антигеном, например, антигеном ErbB2, представляет собой антитело, которое обладает способностью связывать этот антиген с достаточной аффинностью, так чтобы это антитело могло применяться в качестве терапевтического агента, мишенью которого является клетка, экспрессирующая антиген. Если антитело представляет собой антитело, которое связывается с ErbB2, то оно главным образом связывается с ErbB2, а не с другими рецепторами ErbB, и может представлять собой антитело, которое не обладает существенной перекрестной реактивностью на другие протеины, такие как EGFR, ErbB3 или ErbB4. В таких вариантах степень связывания антитела с этими протеинами кроме ErbB2 (например, связывания антитела на поверхности клетки с эндогенным рецептором) должна составлять менее 10% при определении с помощью клеточного сортера с возбуждением флуоресценции (FACS) или с помощью радиоиммунопреципитации (РИА). В некоторых случаях антитело к ErbB2 не должно обладать существенной перекрестной реактивностью на крысиный протеин neu, например, как описано у Schecter и др., Nature, 312: 513 (1984) и Drebin и др., Nature, 312: 545-548(1984).
Антитело, которое "блокирует" активацию лигандом рецептора ErbB, представляет собой антитело, которое снижает или предупреждает такую активацию, как она определена выше, при этом антитело обладает способностью блокировать активацию лигандом рецептора ErbB значительно более эффективно, чем моноклональное антитело 4D5, например, примерно с эффективностью, соответствующей эффективности моноклональных антител 7F3 или 2С4 или их Fab-фрагментов, и предпочтительно примерно с эффективностью, соответствующей эффективности моноклонального антитела 2С4 или его Fab-фрагмента. Например, антитело, которое блокирует активацию лигандом рецептора ErbB, представляет собой антитело, которое примерно на 50-100% более эффективно, чем 4D5 в отношении блокады образования гетероолигомера ErbB. Блокада активации лигандом рецептора ErbB может происходить с помощью любого процесса, например, в результате воздействия на: лиганд, который связывается с рецептором ErbB, образование ErbB-комплекса, тирозинкиназную активность рецептора ErbB в ErbB-комплексе и/или фосфорилирование тирозинкиназного(ых) остатка(ов) в рецепторе или с помощью рецептора ErbB. Примеры антител, которые блокируют активацию лигандом рецептора ErbB, включают моноклональные антитела 2С4 и 7F3 (которые блокируют активацию с помощью HRG гетероолигомеров ErbB2/ErbB3 и ErbB2/ErbB4; и активацию с помощью EGF, TGF-α, амфирегулина, HB-EGF и/или эпирегулина гетероолигомера EGFR/ErbB2); и антитела L26, L96 и L288 (Klapper и др., Oncogene, 14: 2099-2109 (1997)), которые блокируют связывание EGF и NDF с клетками линии T47D, которые экспрессируют EGFR, ErbB2, ErbB3 и ErbB4.
Антитело, которое имеет "биологические характеристики" требуемого антитела, например, такого как моноклональное антитело, обозначенное как 2С4, представляет собой антитело, обладающее одной или несколькими биологическими характеристиками указанного антитела, которое отличается от других антител, обладающих способностью связываться с одним и тем же антигеном (например, ErbB2). Например, антитело, имеющее биологические характеристики 2С4, может блокировать активацию с помощью HRG гетероолигомера ErbB, включающего ErbB2 и ErbB3 или ErbB4; блокировать опосредуемую EGF, TGF-α, HB-EGF, эпирегулином и/или амфирегулином активацию рецептора ЕгЬ, включая EGFR иErbB2; блокировать опосредуемую EGF, TGF-α и/или HRG активацию МАРК; и/или связывается с тем же эпитопом во внеклеточном домене ErbB2, с которым связывается 2С4 (например, которое блокирует связывание моноклонального антитела 2С4 с ErbB2).
Если не указано иное, то выражение "моноклональное антитело 2С4" обозначает антитело, которое имеет атигенсвязывающие остатки мышиного антитела 2С4, описанного ниже в примерах, или которое получено из этого антитела. Например, моноклональное антитело 2С4 может представлять собой мышиное моноклональное антитело 2С4 или его вариант, такой как гуманизированное антитело 2С4, процессирующее антигенсвязывающие аминокислотные остатки мышиного моноклонального антитела 2С4. Примеры гуманизированных антител 2С4 представлены ниже в примере 3. Если не указано иное, то понятие "rhuMAb 2C4" в контексте настоящего описания обозначает антитело, включающее вариабельные области легкой (VL) и тяжелой (VH) цепей, имеющие последовательности SEQ ID NO: 3 и 4 соответственно, слитые с последовательностями константных областей легкой и тяжелой цепи человеческого IgG1 (не А-аллотипа), которое необязательно экспрессируют в клетках яичника китайского хомячка (СНО).
Если не указано иное, то понятие "моноклональное антитело 4D5" обозначает антитело, которое имеет атигенсвязывающие остатки мышиного антитела 4D5 (АТСС CRL 10463), или которое получено из этого антитела. Например, моноклональное антитело 4D5 может представлять собой мышиное моноклональное антитело 4D5 или его вариант, такой как гуманизированное антитело 4D5, процессирующее антигенсвязывающие аминокислотные остатки мышиного моноклонального антитела 4D5. Примеры гуманизированных антител 4D5 включают huMab4D5-1, huMab4D5-2, huMab4D5-3, huMab4D5-4, huMab4D5-5, huMab4D5-6, huMab4D5-7 и huMab4D5-8 (HERCEPTIN®), которые описаны в патенте США 5821337, при этом huMab4D5-8 (HERCEPTIN®) представляет собой предпочтительное гуманизированное антитело 4D5.
Понятие "ингибирующий рост агент" в контексте настоящего описания относится к соединению или композиции, которые ингибируют рост клетки, прежде всего экспрессирующей ErbB раковой клетки, либо in vitro, либо in vivo. Например, ингибирующий рост агент может представлять собой агент, который в значительной степени снижает процент экспрессирующих ErbB, находящихся на S-фазе развития. Примеры ингибирующих рост агентов включают агенты, которые блокируют цикл клеточного развития (на фазе, отличной от S-фазы), такие как агенты, которые индуцируют прекращение G1-фазы или М-фазы. Классические блокаторы М-фазы включают производные винка (винкристин и винбластнин), таксаны, и ингибиторы topo II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Действие тех агентов, которые индуцируют прекращение G1-фазы, также распространяется на прекращение S-фазы, например, таких агентов, как тамоксифен, преднизон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-флуорацил и ара-С. Дополнительную информацию можно найти в Molecular Basis of Cancer, pe-ры Mendelson и Israel, часть 1, озаглавленная "Cell cycle regulation, oncogenes, and antineoplastic drugs", Murakami и др. (WB Saunders: Philadelphia, (1995)), прежде всего на стр.13.
Примерами "ингибирующих рост" антител являются антитела, которые связываются с ErbB2 и ингибируют рост раковых клеток, сверхэкспрессирующих ErbB2. Предпочтительные ингибирующие рост антитела к ErbB2 ингибируют рост клеток опухоли молочной железы линии SK-BR-3 в культуре клеток более чем на 20% и предпочтительно более чем на 50% (например, от примерно 50 до примерно 100%) при концентрации антитела примерно от 0,5 до 30 мкг/мл, когда ингибирование роста определяют чрез 6 дней после обработки клеток линии SK-BR-3 антителом (см. патент США 5677171, выданный 14 октября 1997 г.). Метод анализа ингибирования роста клеток линии SK-BR-3 изложен более подробно ниже в настоящем описании. Предпочтительным ингибирующим рост антителом является моноклональное антитело 4D5, например, гуманизированное 4D5.
Антитело, которое "индуцирует гибель клеток", представляет собой антитело, которое приводит к превращению жизнеспособных клеток в нежизнеспособные. Клетка обычно представляет собой клетку, которая экспрессирует рецептор ErbB2, прежде всего клетку, для которой характерна сверхэкспрессия рецептора ErbB2. Предпочтительно клетка представляет собой раковую клетку, например клетку рака молочной железы, яичника, желудка, эндометрия, слюнной железы, легкого, почки, ободочной кишки, щитовидной железы, поджелудочной железы или мочевого пузыря. В опытах in vitro клетки могут представлять собой клетки линий SK-BR-3, ВТ474, Calu 3, MDA-MB-453, MDA-MB-361 или SKOV3. Гибель клеток in vitro следует определять в отсутствии эффекторных клеток комплемента и эффекторных иммуннокомпетентных клеток для того, чтобы отличать гибель, вызванную опосредуемой антителом клеточной цитотоксичностью (ADCC) и зависящей от комплемента цитотоксичностью (CDC). Так, анализ гибели клеток можно осуществлять с помощью инактивированной нагреванием сыворотки (т.е. в отсутствие комплемента) и в отсутствие эффекторных иммуннокомпетентных клеток. Для определения того, способно ли антитело индуцировать гибель клеток, можно оценивать потерю целостности мембран по поглощению йодида пропидия (PI), трипанового синего (см. Moore и др., Cytotechnology, 17: 1-11 (1995)) или 7AAD по сравнению с необработанными клетками. Предпочтительными индуцирующими гибель клеток антителами являются антитела, которые индуцируют поглощение PI в опыте по поглощению PI клетками линии ВТ474 (см. ниже).
Антитело, которое "индуцируют апоптоз", представляет собой антитело, которое индуцирует запрограммированную гибель клеток, что определяют по связыванию аннексина V, фрагментации ДНК, сжатию клетки, расширению эндоплазматического ретикулума, фрагментации клеток и/или образованию мембранных пузырьков (так называемых апоптозных телец). Клетка обычно представляет собой клетку, в которой происходит сверхэкпрессия рецептора ErbB2. Предпочтительно клетка представляет собой раковую клетку, например клетку рака молочной железы, яичника, желудка, эндометрия, слюной железы, легкого, почки, ободочной кишки, щитовидной железы, поджелудочной железы или мочевого пузыря. В опытах in vitro клетки могут представлять собой клетки линий SK-BR-3, ВТ474, Calu 3, MDA-MB-453, MDA-MB-361 или SKOV3. Различные методы могут применяться для оценки случаев гибели клеток, связанной с апоптозом. Например, по связыванию аннексина может быть оценена транслокация фосфатидилсерина (PS); фрагментация ДНК может быть оценена по образованию ступеней ДНК; конденсация ядер/хроматина наряду с фрагментаций ДНК может быть определена по увеличению количества гиподиплоидных клеток. Предпочтительно антитело, которое индуцирует апоптоз, представляет собой антитело, которое приводит примерно к 2-50-кратной, предпочтительно примерно к 5-50-кратной и наиболее предпочтительно к 10-50-кратной индукции связывания аннексина по сравнению с необработанными клетками, при анализе связывания аннексина с использованием клеток линии ВТ474 (см. ниже). В некоторых случаях проапоптозное антитело может представлять собой антитело, которое дополнительно блокирует активацию лигандом ErbB рецептора ErbB (например, антитело 7F3); т.е. антитело, которое обладает биологическими характеристиками моноклонального антитела 2С4. В других вариантах антитело представляет собой антитело, которое не обладает способностью в значительной степени блокировать активацию лигандом ErbB рецептора ErbB (например, антитело 7С2). Кроме того, антитело может представлять собой подобное 7С2 антитело, которое, обладая способностью индуцировать апоптоз, не приводит к значительному снижению процента клеток, находящихся в S-фазе (например, антитело, которое индуцирует только 0-10%-ное снижение количества таких клеток по сравнению с контролем).
Понятие "эпитоп 2С4" обозначает область во внеклеточном домене ErbB2, с которой связывается антитело 2С4. С целью отбора антител, которые связываются с эпитопом 2С4, можно осуществлять обычный анализ перекрестного связывания, такой как анализ, описанный в Antibodies. A Laboratory Manual, Cold Spring Harbor Laboratory, ред-ры Harlow и David Lane (1988). В другом варианте картирование эпитопа можно осуществлять для анализа того, связывается ли антитело с эпитопом 2С4 ErbB2 (например, с любым одним или несколькими остатками в области примерно от остатка 22 до примерно остатка 584 ErbB2 включительно; см. фиг.1А-Б).
Понятие "эпитоп 4D5" обозначает область во внеклеточном домене ErbB2, с которой связывается антитело 4D5 (АТСС CRL 10463). Этот эпитоп примыкает к трансмембранному домену ErbB2. С целью отбора антител, которые связываются с эпитопом 4D5, можно осуществлять обычный анализ перекрестного связывания, такой как анализ, описанный в Antibodies. A Laboratory Manual, Cold Spring Harbor Laboratory, ред-ры Harlow и David Lane (1988). В другом варианте картирование эпитопа можно осуществлять для анализа того, связывается ли антитело с эпитопом 4D5 ErbB2 (например, с любым одним или несколькими остатками в области примерно от остатка 529 до примерно остатка 625 включительно, см. фиг.1А-Б).
Понятие "эпитоп 3Н4" обозначает область во внеклеточном домене ErbB2, с которой связывается антитело 3Н4. Эпитоп включает остатки примерно от 541 до примерно 599, включительно, аминокислотной последовательности внеклеточного домена ErbB2; см. фиг.1А-Б.
Понятие "эпитоп 7C2/7F3" обозначает область на N-конце внеклеточного домена ErbB2, с которой связываются антитела 7С2 и/или 7F3 (каждое из которых депонировано в АТСС, см. ниже). С целью отбора антител, которые связываются с эпитопом 7C2/7F3, можно осуществлять обычный анализ перекрестного связывания, такой как анализ, описанный в Antibodies. A Laboratory Manual, Cold Spring Harbor Laboratory, ред-ры Harlow и David Lane (1988). В другом варианте картирование эпитопа может быть осуществлено для анализа того, связывается ли антитело с эпитопом 7C2/7F3 ErbB2 (например, с любым одним или несколькими остатками в области, примерно от остатка 22 до примерно остатка 53 ErbB2; см. фиг.1А-Б).
Понятие "лечение" относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам. Пациенты, которые нуждаются в лечении, включают пациентов, которые уже имеют нарушение, и тех, у которых появление нарушения следует предупреждать. Следовательно, у млекопитающего, подлежащего лечению согласно изобретению, нарушение уже может быть диагностировано, или млекопитающее может быть предрасположено или чувствительно к нарушению.
Понятие "млекопитающее", подлежащее лечению, включает любое животное, относящееся к классу млекопитающих, включая людей, домашних и сельскохозяйственных животных, а также живущих в зоопарке животных, предназначенных для спорта или комнатных животных, таких как собаки, лошади, кошки, коровы и т.д. Предпочтительно млекопитающее представляет собой человека.
Понятие "нарушение" включает любое состояние, которое может облегчаться в результате лечения с помощью антитела к ErbB2. Оно включает хронические и острые нарушения или болезни, в том числе патологические состояния, которые приводят к предрасположенности млекопитающего к рассматриваемому нарушению. Примеры нарушений, которые могут лечиться согласно изобретению включают (но не ограничиваясь ими) доброкачественные и злокачественные опухоли; лейкозы и лимфоидные злокачественные заболевания; нарушения относящиеся к нервной клетке, глии, астроциту, гипоталамусу и другим железам, к макрофагам, эпителию, строме и бластоцелю; а также воспалительные, ангиогенные и иммунологические нарушения.
Понятие "терапевтически эффективное количество" относится к количеству лекарственного средства, эффективному для лечения заболевания или нарушения у млекопитающего. При раке терапевтически эффективное количество лекарственного средства может снижать количество раковых клеток; уменьшать размер опухоли; ингибировать (т.е. в некоторой степени замедлять или предпочтительно прекращать) инфильтрацию раковых клеток в периферические органы; ингибировать (т.е. в некоторой степени замедлять или предпочтительно прекращать) метастазы опухоли; ингибировать в определенной степени рост опухоли и/или облегчать в некоторой степени один или несколько симптомов, связанных с раком. По уровню, с которым лекарственное средство может предупреждать рост и/или вызывать гибель существующих раковых клеток, оно может быть цитостатическим /или цитоксическим. В терапии рака эффективность может, например, оцениваться по времени развития болезни (ТТР) и/или путем определения скорости реакции (RR).
Понятия "рак" и "раковый (злокачественный)" относится к физиологическому состоянию или описывает физиологическое состояние у млекопитающего, которое обычно характеризуется нерегулируемым ростом клеток. Примеры рака включают (но не ограничиваясь ими) карциному, лимфому, бластому, саркому и лейкоз или лимфоидные злокачественные заболевания. Более конкретные примеры таких видов рака включают плоскоклеточный рак (например, эпителиальный плоскоклеточный рак), рак легкого, включая мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого и плоскоклеточный рак легкого, перитонеальный рак, рак клеток печени, рак пищевода или желудка, включая желудочно-кишечный рак, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак ободочной кишки, рак прямой кишки, колоректальный рак, карциному эндометрия или матки, карциному слюнных желез, рак почки, рак предстательной железы, рак вульвы, рак щитовидной железы, печеночную карциному, анальную карциному, карциному полового члена, а также рак головы и шеи.
Понятие "ErbB-экспрессирующий рак" относится к раковой опухоли, включающей клетки, на клеточной поверхности которых присутствует протеин ErbB. Понятие "ErbB2-экспрессирующий рак" относится к раковой опухоли, на поверхности клеток которой продуцируются уровни ErbB2, достаточные для того, чтобы антитело к ErbB2 могло связывать их и оказывать терапевтическое воздействие на раковую опухоль.
Понятие "рак, характеризующейся чрезмерной активацией" рецептора ErbB, обозначает рак, при котором уровень активации рецептора ErbB в значительной степени превышает уровень активации этого рецептора в незлокачественных клетках ткани такого же типа. Такая чрезмерная активация может быть результатом сверхэкспрессии рецептора ErbB и/или более высоким по сравнению с нормальным уровнем лиганда ErbB, доступного для активирующегося рецептора ErbB в раковых клеток. Такая чрезмерная активация может быть причиной и/или вызываться злокачественным состоянием раковой клетки. В некоторых вариантах осуществления рак должен подвергаться диагностическому или прогностическому анализу для определения того, имеет ли место амплификация и/или сверхэкспрессия рецептора ErbB, которая может привести к такой чрезмерной активации рецептора ErbB. В альтернативном или дополнительном варианте рак может подвергаться диагностическому или прогностическому анализу для определения того, имеет ли место амплификация и/или сверхэкспрессия лиганда ErbB, которая может быть связана с чрезмерной активацией рецептора. В подгруппе таких раковых заболеваний чрезмерная активация рецептора может быть результатом аукринного пути стимуляции.
При "аукринном" пути стимуляции происходит самостимуляция с помощью раковой клетки, продуцирующей как лиганд ErbB, так и его родственный рецептор ErbB. Например, раковая клетка может экспрессировать или сверхэкпрессировать EGFR и также экспрессировать или сверхэкпрессировать лиганд EGFR (например, EGF, TGF-α или HB-EGF). В другом варианте раковая клетка может экспрессировать или сверхэкпрессировать ErbB2 и также экспрессировать или сверхэкпрессировать херегулин (например, γ-HRG).
Рак, при котором происходит "сверхэкпрессия" рецептора ErbB, представляет собой рак, при котором на клеточной поверхности значительно повышены уровни рецептора ErbB, такого как ErbB2, по сравнению с незлокачественной клеткой этого же типа ткани. Такая сверхэкпрессия может вызываться амплификацией гена или повышенным уровнем транскрипции или трансляции. Сверхэкпрессию рецептора ErbB можно определять с помощью диагностического или прогностического анализа, который позволяет оценить повышенные уровни протеина ErbB на поверхности клетки (например, с помощью иммуногистохимического анализа; ИГХ). В альтернативном или дополнительном варианте можно измерять уровни кодирующей ErbB нуклеиновой кислоты в клетке, например, с помощью флуоресцентной гибризации in situ (FISH; см. WO 98/45479, опубликованную в октябре 1998 г.), Саузерн-блоттинга или методов на основе полимеразной цепной реакции (ПЦР), таких как количественная экспресс-ПЦР (ОТ-ПЦР). Сверхэкспрессию рецептора ErbB также можно изучать путем оценки выделения антигена (например, внеклеточного домена ErbB) в биологическую жидкость, такую как сыворотка (см., например, патент США 4933294, выданный 12 июня 1990 г.; WO 91/05264, опубликованную 18 апреля 1991 г.; патент США 5401638, выданный 28 марта 1995 г. и у Sias и др., J.Immunol. Methods, 132: 73-80 (1990)). Помимо указанных выше методов, специалисту в данной области известны различные анализы in vivo. Например, можно обрабатывать клетки внутри организма пациента антителом, необязательно помеченным маркером, который можно обнаружить, например, радиоактивным изотопом, а связывание антитела с клеткой в организме пациента можно определять, например, с помощью внешнего сканирования радиоактивности или путем анализа биопсии, взятой из организма пациента, до обработки антителом.
И наоборот, рак, для которого "не характерна сверхэкспрессия ErbB" представляет собой рак, при диагностическом анализе которого не выявлены уровни экспрессии рецептора ErbB2, превышающие нормальные уровни по сравнению с незлокачественной клеткой этого же типа ткани.
Рак, при котором происходит "сверхэкспрессия" лиганда ErbB, представляет собой рак, при котором продуцируются существенно более высокие уровни лиганда ErbB по сравнению с незлокачественной клеткой этого же типа ткани. Такая сверхэкспрессия может вызываться амплификацией гена или повышенным уровнем транскрипции или трансляции. Сверхэкпрессию лиганда ErbB можно определять с помощью диагностического анализа, который позволяет оценить уровни лиганда (или кодирующей его нуклеиновой кислоты) у пациента, например, с помощью биопсии опухоли или различных диагностических анализов, таких как ИГХ, FISH, Саузерн-блоттинг, ПЦР или описанных выше анализов in vivo.
"Независящий от гормонов" рак представляет собой рак, при котором пролиферация не зависит от присутствия гормона, обладающего способностью связывать рецептор, который экспрессируется клетками при раке. Такие типы рака не поддаются клинической регрессии при введении фармацевтических агентов или с помощью хирургического вмешательства, посредством которого снижается концентрация гормона в опухоли или близи нее. Примеры независящих от гормонов видов рака включают независящий от андрогена рак предстательной железы, независящий от эстрогена рак молочной железы, рак эндометрия и рак яичника. Такие виды рака могут начинаться как зависящие от гормонов опухоли и развиваться, переходя после гормональной терапии от чувствительной к гормону стадии к устойчивой к гормону опухоли.
Понятие "цитотоксичный агент" в контексте настоящего описания относится к субстанции, которая ингибирует или препятствует функции клеток и/или вызывает разрушение клеток. Подразумевается, что понятие включает радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивные изотопы Lu), химиотерапевтические агенты и токсины, такие как токсины, имеющие небольшой размер молекул, или обладающие ферментативной активностью токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты.
"Химиотерапевтический агент" представляет собой химическое соединение, которое применяют для лечения рака. Примеры химиотерапевтических агентов включают алкилирующие агенты, такие как тиотепа и циклофосфамид (CYTOXANФ); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбохуон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфаорамид, триметилоломеламин; азотные аналоги горчичного газа, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлоретамин, гидрохлорид оксида мехлорметамина, мелфалан, новембихин, фенэстерин, преднимустин, трофосфамид, урациловый аналог горчичного газа; нитрозмочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин, ранимустин; антибиотики, такие как аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, калихеаминцин, карабицин, карминомицин, карзинофелин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диаза-5-оксо-L-норлейцин, доксорубицин, эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, микофенольная кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, хеломицин, родарубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-ФУ); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; пуриновые аналоги, такие как флударабин, 6-меркаптопурин, тиамитрин, тиогуанин; пиримидиновые аналоги, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуредин, доксифлуридин, эноцитабин, флоксуридин, 5-ФУ; андрогены, такие как калустерон, дромостанолонпропионат, эпитиостанол, мепитиостан, тестолактон; антиадренергетики, такие как аминоглутетимид, митотан, трилостан; заменитель фолиевой кислоты, такой как пролиновая кислота; ацеглатон; альдофосфамидгликозид; аминолевулиновую кислоту; амсакрин; бестрабуцил; бисантрен; эдатраксат; дефофамин; демеколцин; диазихон; элформитин; ацетат эллиптиния; этоглюцид; нитрат галлия; гидроксимочевину; лентинан; лонидамин; митогуазон; митоксантрон; мопидамол; нитракрин; пентостатин; фенамед; пирарубицин; подафиллиновую кислоту; 2-этилгидразид; прокарбазин; PSK®; разоксан; сизофиран; спирогерманий; тенуазоновую кислоту; триазихон; 2,2',2''-трихлортриэтиламин; уретан; виндесин; дакарбазин; манномустрин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид; ("Ara-С"); циклофосфамид; тиотепа; таксаны, например патклитаксел (TAXOL®, Bristol-Myers Squibb Oncology, Принстон, штат Нью-Джерси) и доцетаксел (TAXOTERE®, Rhone-Poulenc Rorer, Энтони, Франция); хлорамбуцил; гемцитабин; 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин; платина; этопозид (VP-16); ифосфамид; митомицин С; митоксантрон; винкристин; винорелбин; навелбин; новантрон; тенипозид; дауномицин; аминоптерин; кселода; ибандронат; СРТ-11; ингитор топоизомеразы RFS-2000; дифторметилорнитин (ДМПО); ретиноевую кислоту; эсперамицины; капецитабин и их фармацевтически приемлемые соли, кислоты или производные любого из указанных выше производных. Также под объем этого определения подпадают антигормональные агенты, которые регулируют или ингибируют действие гормона на опухоли, такие как антиэстрогены, включая, например, тамоксифен, раллоксифен, ингибирующие ароматазу 4(5)-имидазолы, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон, иторемифен (фарестон) и антиандрогены, такие как флутамид, нилутамид, бикалутамид, леупролид и госерелин, а также фармацевтически приемлемые соли, кислоты и производные любого из указанных выше соединений.
В контексте настоящего описания понятие "лекарственное средство, мишенью которого является EGFR" обозначает терапевтический агент, который связывается с EGFR и необязательно ингибирует активацию EGFR. Примеры таких агентов включают антитела и небольшие молекулы, которые связываются с EGFR. Примеры антител, которые связываются с EGFR, включают Mab 579 (АТСС CRL НВ 8506), Mab 455 (АТСС CRL НВ 8507), Mab 225 (АТСС CRL НВ 8506), Mab 528 (АТСС CRL НВ 8509) (см. патент США 4943533, на имя Mendelsohn и др.) и их варианты, такие как химерное антитело 225 (С225) и реконструированное человеческое антитело 225 (Н225) (см. WO 96/40210, на имя Imclone Systems Inc.); антитела, которые связываются с мутантом типа II EGFR (см. патент США 5212290); гуманизированные и химерные антитела, которые связываются с EGFR, описанные в патенте США 5891996; и гуманизированные антитела, которые связываются с EGFR (см. WO 98/50433 на имя Abgenix). Антитело к EGFR может быть конъюгировано с токсическим агентом с получением иммуноконъюгата (см., например, ЕР 659439А на имя Merk Patent GmbH). Примеры небольших молекул, которые связываются с EGFR, включают ZD1839 (Astra Zeneca), CP-358774 (OSI/Pfizer) и AG1478.
Понятие "антиангиогенный агент" обозначает соединение, которое блокирует или оказывает определенное воздействие на развитие кровеносных сосудов. Антиангиогенный фактор, например, может представлять собой небольшую молекулу или антитело, которое связывается с фактором роста или рецептором фактора роста, принимающими участие в усилении ангиогенеза. В контексте настоящего описания предпочтительным антиангиогенным фактором является антитело, которое связывается с сосудистым эндотелиальным фактором роста (VEGF).
Понятие "цитокин" является общим обозначением для протеинов, которые высвобождаются одной популяцией клеток и оказывают воздействие на другую клетку в качестве межклеточных медиаторов. Примерами таких цитокинов являются лимфокины, монокины и традиционные полипептидные гормоны. Под понятие цитокины подпадают гормон роста, такой как человеческий гормон роста, N-метионилированный человеческий гормон роста и бычий гормон роста; паратиреодный гормон; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (FSH), тиреостимулирующий гормон (TSH) и лютеинизирующий гормон (LH); печеночный фактор роста; фибробластный фактор роста; пролактин; плацентарный лактоген; фактор некроза опухоли-α и -β; муллерианингибирующую субстанцию; мышиный связанный с гонадотропином пептид; ингибин; активин; сосудистый эндотелиальный фактор роста; интегрин; тромбопоэтин (ТРО); факторы роста нерва, такие как NGF-β; тромбоцитарный фактор роста; трансформирующие факторы роста (TGF), такие как TGF-α и TGF-β; инсулинподобные факторы типа I и II; эритропоэтин (ЕРО); остеоиндуктивные факторы; интерфероны, такие как интерферон-α, -β и -γ; колониестимулирующие факторы (CSF), такие как макрофаг-CSF (M-CSF); гранулоцит-макрофаг-CSF (GM-CSF); и гранулоцит-CSF (G-CSF); интерлейкины (IL), такие как IL-1, IL-1a, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10 IL-11, IL-12; фактор некроза опухоли, такой как TNF-α или TNF-β; и другие полипептидные факторы, включая LIF и набор лигандов (KL). В контексте настоящего описания понятие цитокин включает протеины, имеющие естественное происхождение или полученные из культуры рекомбинантных клеток и биологически активные эквиваленты природных последовательностей цитокинов.
Понятие "пролекарство" в контексте настоящего описания обозначает предшественник или производное фармацевтически активной субстанции, которые менее цитотоксичны для опухолевых клеток по сравнению с исходным лекарственным средством и обладают способностью активироваться или превращаться в более активную исходную форму с помощью ферментов (см., например, Wilman в: "Prodrugs in Cancer Chemotherapy", Biochemicals Society Transactions, 14, стр.375-382, 615-й симпозиум в Белфасте (1986); и Stella и др. в: "Prodrugs: A Chemical Approach to Targeted Drug Delivery", Directed Drug Delivery, Borchardt и др. (ред-ры), стр.247-267, Humana Press (1985)). Пролекарства по изобретению включают (но не ограничиваясь ими) фосфатсодержащие пролекарства, тиофосфатсодержащие пролекарства, сульфатсодержащие пролекарства, пептидсодержащие пролекарства, модифицированные D-аминокислотой пролекарства, гликозилированные пролекарства, β-лактамсодержащие пролекарства, пролекарства, содержащие необязательно замещенный феноксиацетамид, или пролекарства, содержащие необязательно замещенный фенилацетамид, 5-флуороцитозин- и другие 5-флуороуридинсодержащие пролекарства, которые могут превращаться в более активную цитотоксическую свободную лекарственную форму. Примеры цитотоксических лекарственных средств, которые могут образовывать производные в виде пролекарства и применяются согласно изобретению, включают (но не ограничиваясь ими) описанные выше химиотерапевтические агенты.
"Липосома" обозначает небольшой пузырек, включающий различные типы липидов, фосфолипидов и/или детергент, который может применяться для введения млекопитающему лекарственного средства (такого как описанные выше антитела к ErbB2, и необязательно химиотерапевтический агент). Компоненты липосом обычно имеют двуслойное строение, аналогичное строению биологических мембран.
Понятие "вкладыш для упаковки" в контексте настоящего описания обозначает инструкции, которыми обычно снабжают поступающие в продажу упаковки терапевтических продуктов, содержащие информацию о показаниях, применении, дозах, пути введения, противопоказания и/или предостережения, касающиеся применения таких терапевтических продуктов.
"Кардиозащитное средство" обозначает соединение или композицию, которые предупреждают или снижают дисфункцию миокарда (т.е. кардиомиопатию или застойную сердечную недостаточность), связанную с введением пациенту лекарственного средства, такого как антрациклиновый антибиотик и/или антитело к ErbB2. Кардиозащитное средство может, например, блокировать или уменьшать опосредуемое свободными радикалами кардиотоксическое действие и/или предупреждать или снижать повреждение, вызванное окислительным стрессом. Примеры кардиозащитных средств, подпадающих под объем изобретения, включают образующий хелатные комплексы с железом агент, такой как дексразоксан (ICRF-187) (Seifert и др. в: The Annals of Pharmacotherapy, 28: 1063-1072 (1994)); снижающий уровень липидов агент и/или антиоксидант, такой как пробукол (Signal и др., J.Mol. Cell Cardiol., 27: 1055-1063 (1995)); амифостин (сложный аминотиол-2-[(3-аминопропил)амино]этантиоловый эфир кислого первичного фосфата, также называемый WR-2721, и его дефосфорилированная проникающая в клетку форма, называемая WR-1065) и S-3-(3-метиламинопропиламино)пропилфосфоротиоевая кислота WR-151327) (см. также Green и др., Cancer Research, 54: 738-741 (1994)); дигоксин (Bristow M.R. в: Bristow M.R. (ред), Drug-Induced Heart Disease, New York: Elsevier, 191-215 (1980)); бета-блокаторы, такие как метопролол (Hjamarson и др., Drugs, 47: Suppl. 4: 31-9(1994); и Shaddy и др., Am. Heart J., 129: 197-199 (1995)); витамин Е; аскорбиновую кислоту (витамин С); поглотители свободных радикалов, такие как олеаноловая кислота, урсоловая кислота и N-ацетилцистеин (NAC); соединения, представляющие собой спиновые ловушки, такие как альфа-фенил-трет-бутилнитрон (PBN) (Paracchini и др., Anticancer Res., 13: 1607-1612 (1993)); селенорганические соединения, такие как Р251 (элбесен) и т.п.
"Выделенная" молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая идентифицирована и отделена от, по меньшей мере, одной молекулы нуклеиновой кислоты-примеси, с которой она обычно связана в естественном источнике нуклеиновой кислоты антитела. Выделенная молекула нуклеиновой кислоты отличается от той формы или набора, в которых она находится в естественных условиях. Таким образом, выделенная молекула нуклеиновой кислоты отличается от молекулы нуклеиновой кислоты, существующей в природных клетках. Однако выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, находящуюся в клетках, в которых в норме происходит экспрессия антитела, например, в случае, если молекула нуклеиновой кислоты имеет локализацию в хромосоме, отличную от ее локализации в природных клетках.
Выражение "контролирующие последовательности" относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в определенном организме-хозяине. Пригодные для прокариот контролирующие последовательности включают, например, промотор, необязательно оператор и сайт связывания рибосомы. Как известно, в эукариотических клетках присутствуют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота "функционально связана", если она находится в функциональной связи с другой нуклеотидной последовательностью. Например, ДНК предпоследовательности или секреторного лидера функционально связана с ДНК полипептида, если она экспрессируется в виде предпротеина, который принимает участие в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он оказывает воздействие на транскрипцию последовательности; сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, что может облегчать трансляцию. Как правило, "функционально связан" обозначает, что связанные последовательности ДНК являются смежными, а в случае секреторного лидера, являются смежными и находятся в фазе считывания. Однако энхансеры не обязательно должны быть смежными. Связывание осуществляется путем встраивания лигированием в существующий сайт рестрикции. Если такие сайты не существуют, то согласно известной практике применяют синтетические олигонуклеотидные адаптеры или линкеры.
В контексте настоящего описания понятия "клетка", "линия клеток" и "культура клеток" используются взаимозаменяемо, и все такие понятия включают потомство. Так, понятия "трансформация" и "трансформированные клетки" включают первичную представляющую интерес клетку и полученные из нее культуры, вне зависимости от количества пересевов. Следует также понимать, что все потомство может не быть точно идентичным по содержанию ДНК вследствие преднамеренных или случайных мутаций. Под понятие подпадает мутантное потомство, обладающее такой же функцией или биологической активностью, которые выявлены у исходной трансформированной клетки. Если подразумеваются другие понятия, они должны быть очевидны из контекста.
II. Получение антитела к ErbB2
Ниже описаны примеры методик получения антител, которые применяются согласно изобретению. Антиген ErbB2, который должен применяться для получения антител, может, например, представлять собой растворимую форму внеклеточного домена ErbB2 или его часть, содержащую требуемый эпитоп. В другом варианте для получения антител можно применять клетки, экспрессирующие ErbB2 на клеточной поверхности (например, клетки линии NIH-3T3, трансформированные с целью сверхэкспрессии ErbB2; или линия клеток карциномы, такая как линия клеток SK-BR-3, см. Stancovski и др., PNAS (USA), 88: 8691-8695 (1991)). Специалистам в данной области должны быть очевидны другие формы ErbB2, которые можно применять для получения антител.
(I) Поликлональные антитела
Поликлональные антитела предпочтительно получают в организме животных с помощью нескольких подкожных (sc) или внутирибрюшинных (ip) инъекций соответствующего антигена и адъюванта. Может оказаться целесообразным использовать конъюгат соответствующего антигена и протеина, который является иммуногенным для видов, подлежащих иммунизации, например, гемоцианин лимфы улитки, сывороточный альбумин, бычий тироглобулин или соевый ингибитор трипсина, применяя бифункциональный или дериватизирующий агент, например, сложный малеимидобензоилсульфосукцимидный эфир (конъюгация через остатки цистеина), N-гидроксисукцинимид (конъюгация через остатки лизина), глутаровый альдегид, янтарный ангидрид, SOCl2 или R1N=C=NR, где R и R1 обозначают различные алкильные группы.
Животных иммунизируют антигеном, иммуногеными конъюгатами или их производными путем объединения, например, 100 мкг или 5 мкг протеина или конъюгата (для кроликов или мышей соответственно) с 3 объемами полного адъюванта Фрейнда и внутрикожной инъекции раствора в несколько мест. Через 1 месяц животных подвергают бустерной инъекции с использованием 1/5-1/10 исходного количества пептида или конъюгата в полном адъюванте Фрейнда путем подкожной инъекции в несколько мест. Через 7-14 дней у животных берут кровь и анализируют в сыворотке титр антител. Животных подвергают бустерным инъекциям до тех пор пока титр антител не достигнет постоянного уровня. Предпочтительно животных подвергают бустерным инъекциям с использованием конъюгата, который включает этот же антиген, но другой протеин, и/или сшитого с помощью другого реагента. Конъюгаты также можно получать в культуре рекомбинантных клеток в виде слитых протеинов. Для усиления иммунного ответа также можно использовать агрегирующие агенты, такие как квасцы.
(II) Моноклональные антитела
Моноклональные антитела получают из популяции практически гомогенных антител, т.е. из популяции, в которой индивидуальные антитела идентичны, за исключением наличия возможных встречающихся в естественных условиях мутаций, которые могут присутствовать в небольших количествах. Таким образом, определение "моноклональный" относится к варианту, когда антитело не является смесью дискретных антител.
Моноклональные антитела можно создавать, например, с помощью метода на основе гибридом, который впервые описан Kohler и др., Nature, 256: 495 (1975), или с помощью методов рекомбинантной ДНК (патент США 4816567).
При использовании метода на основе гибридом мышь или другое пригодное животное-хозяин, такое как хомячок, иммунизируют согласно описанной выше методике, с тем чтобы вызывать образование лимфоцитов, которые продуцируют или могут продуцировать антитела, обладающие способностью специфично связывать протеин, применяемый для иммунизации. Согласно другому варианту лимфоциты можно подвергать иммунизации in vitro. Затем лимфоциты можно сливать с клетками миеломы с помощью приемлемого связывающего агента, такого как полиэтиленгликоль, с получением клетки гибридомы (Goding, Monoclonal Antibodies: Principles and Practice, стр.59-103 (Academic Press, 1986)).
Полученные таким образом клетки гибридомы высевают и выращивают в пригодной культуральной среде, которая предпочтительно содержит одно или несколько веществ, ингибирующих рост или выживание неслитых, родительских клеток миеломы. Например, если родительские клетки миеломы не содержат фермента гипоксантингуанинфосфорибозилтрансферазы (HGPRT или HPRT), культуральная среда для гибридом, как правило, должна включать гипоксантин, аминоптерин и тимидин (НАТ-среда), т.е. вещества, которые препятствуют росту клеток с дефицитом HGPRT.
Предпочтительными клетками миеломы являются клетки, которые легко поддаются слиянию, поддерживают стабильный высокий уровень производства антител выбранными продуцирующими антитела клетками и чувствительны к среде, такой как НАТ-среда. Из них предпочтительными линиями клеток миелом являются линии мышиной миеломы, такие как созданные на основе мышиных линий опухолевых клеток МОРС-21 и МРС-11, которые могут быть получены из Salk Institite Cell Disrtibution Center, Сан-Диего, штат Калифорния, США, и SP-2 или Х63-AgS-653, которые могут быть получены из Американской коллекции типовых культур, Роквилл, штат Мэриленд, США. Также описано применение линий клеток человеческой миеломы и гетеромиеломы типа "мышь-человек" для производства моноклональных аетител (Kozbor, J.Immunol., 133: 3001 (1984); и Brodeur и др., Monoclonal Antibody Production Techniques and Applications, стр.51-63 (Marcel Dekker Inc., New York, 1987)).
Культуральную среду, в которой выращивают клетки гибридомы, анализируют в отношении производства моноклональных антител к антигену. Предпочтительно специфичность связывания моноклональных антител, полученных с использованием клеток гибридомы, определяют методом иммунопреципитации или с помощью анализа связывания in vitro, такого радиоиммунный анализ (РИА) или твердофазный имммуноферментный анализ (ELISA).
Аффинность связывания моноклонального антитела можно, например, определять с помощью Scatchard-анализа, описанного Munson и др., Anal. Biochem., 107: 220 (1980).
После того как выявлены клетки гибридомы, которые продуцируют антитела требуемой специфичности, аффинности и/или активности, клоны можно субклонировать с помощью процессов ограничивающего разведения и выращивать стандартными методами (Goding, Monoclonal Antibodies: Principles and Practice, стр. 59-103 (Academic Press, 1986)). Пригодные для этой цели среды включают, например, среду D-MEM или RPMI-1640. Кроме того, клетки гидрибомы можно выращивать in vivo в виде асцитных опухолей в организме животных.
Моноклональные антитела, секретируемые субклонами, можно отделять от культуральной среды, асцитной жидкости или сыворотки с помощью обычных методов очистки антител, например, с использованием протеин А-сефарозы, хроматографии с использованием гидроксилапатитов, гель-электрофореза, диализа или аффинной хроматографии.
ДНК, кодирующую моноклональные антитела, можно легко выделить и секвенировать с помощью общепринятых процедур (например, с использованием олигонуклеотидных зондов, которые обладают способностью специфично связывать гены, кодирующие тяжелые и легкие цепи мышиных антител). В качестве предпочтительного источника такой ДНК служат клетки гибридомы. После выделения ДНК можно включать в экспрессионные векторы, которыми затем трансфектируют клетки-хозяева, такие как клетки Е.coli, обезьяньи COS-клетки, клетки яичника китайского хомячка (СНО) или клетки миеломы, которые без трансфекции не продуцируют протеин антитела, что приводит к синтезу моноклональных антител в рекомбинантных клетках-хозяевах. Обзор статей о рекомбинантной экспрессии в бактериях ДНК, кодирующей антитела, см. у Skerra и др., Curr. Opinion in Imunol., 5: 256-262 (1993) и Plückthun, Immunol. Revs. 130: 151-188 (1992).
Согласно еще одному варианту моноклональные антитела или фрагменты антител можно выделять из фаговых библиотек антител, созданных с помощью методов, описанных у McCafferty и др., Nature, 348: 552-554 (1990). Clackson и др., Nature, 352: 624-628 (1991) и Marks и др., J.Mol. Biol, 222: 581-597 (1991) описали выделение мышиных и человеческих антител соответственно с помощью библиотек фагов. В последующих публикациях было описано выделение высокоаффинных (нМ-диапазон) человеческих антител с помощью перестановки цепи (Marks и др., Bio/Technology, 10: 779-783 (1992)), а также комбинаторная инфекция и рекомбинация in vivo в качестве стратегии конструирования очень больших библиотек фагов (Waterhouse и др., Nucl. Acids. Res., 21: 2265-2266 (1993)). Таким образом, эти методы представляют собой реальную альтернативу традиционным методам выделения моноклональных антител на основе гибридом моноклональных антител.
ДНК также можно модифицировать, например, заменой кодирующими последовательностями константных областей легкой человеческой и тяжелой цепи гомологичных мышиных последовательностей (патент США 4816567 и Morrison и др., Proc. Natl. Acad. Sci. USA: 81: 6851 (1984)) или с помощью ковалентного связывания с кодирующей последовательностью иммуноглобулина всей или части кодирующей последовательности полипептида, не относящегося к иммуноглобулинам.
Как правило, такие не относящиеся к иммуноглобулинам полипептиды заменяют на константные области антитела или их заменяют на вариабельные области антигенсвязывающего центра антитела, создавая химерное бивалентное антитело, которое содержит один антигенсвязывающий центр, обладающий специфичностью в отношении антигена, и другой антигенсвязывающий центр, обладающий специфичностью в отношении другого антигена.
(III) Гуманизированные антитела
Методы создания "гуманизированных" антител животных кроме человека описаны в данной области. Предпочтительно гуманизированное антитело имеет один или несколько встроенных в него аминокислотных остатков, полученных из источника, отличного от человека. Эти полученные из источника, отличного от человека, аминокислотные остатки часто называют "импортными" остатками, поскольку их обычно получают из "импортной" вариабельной области. Гуманизацию в целом можно осуществлять согласно методу Winter с соавторами (Jones и др., Nature, 321: 522-525 (1986); Riechmann и др., Nature, 332: 323-327 (1988); Verhoeyen и др., Science, 239: 1534-1536 (1988)) путем замены последовательностей гипервариабельного участка на соответствующие последовательности человеческого антитела. Таким образом, "гуманизированные" антитела представляют собой химерные антитела (патент США 4816567), в которых участок, существенно меньший, чем вариабельная интактная человеческая область, заменен соответствующей последовательностью, полученной из видов кроме человека. На практике гуманизированные антитела представляют собой обычные человеческие антитела, в которых часть остатков гипервариабельного участка и возможно часть остатков FR заменены остатками из аналогичных участков антител грызунов.
Выбор вариабельных человеческих областей как легкой, так и тяжелой цепи, предназначенных для получения гуманизированных антител, очень важен для снижения антигенности. Согласно так называемому методу "наилучшего подбора" последовательность вариабельной области антитела грызунов подвергают скринингу в отношении полной библиотеки известных последовательностей вариабельных человеческих областей. Человеческую последовательность, которая наиболее близка к последовательности грызунов, затем применяют в качестве человеческого каркасного участка (FR) для гуманизированного антитела (Sims и др., J.Immunol., 151: 2296 (1993); Chothia и др., J.Mol.Biol., 196: 901 (1987)). В другом методе используется определенный каркасный участок, полученный из консенсусной последовательности всех человеческих антител определенной подгруппы легких или тяжелых цепей. Один и тот же каркасный участок может применяться для нескольких различных гуманизированных антител (Carter и др., Proc. Natl. Acad. Sci. USA: 89: 4285 (1992); Prestan др., J.Immunol, 151: 2623 (1993)).
Также важно, чтобы антитела были гуманизированы с сохранением высокой аффинности к антигену и других важных биологических свойств. Для решения этой задачи согласно предпочтительному способу гуманизированные антитела получают с помощью анализа родительских последовательностей и умозрительных различных гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов общедоступны и хорошо известны специалистам в данной области. Известны компьютерные программы, которые иллюстрируют и отображают возможные трехмерные конформационные структуры выбранных перспективных последовательностей иммуноглобулинов. Изучение этих изображений позволяет осуществлять анализ возможной роли остатков в функции перспективной последовательности иммуноглобулина, т.е. анализ остатков, которые влияют на способность перспективного иммуноглобулина связывать антиген. Таким путем могут быть выбраны FR-остатки и объединены с реципиентными и импортными последовательностями для достижения требуемых характеристик антитела, таких как повышенная аффинность к антигену(ам)-мишени(ням). Как правило, остатки гипервариабельного участка оказывают непосредственное и наиболее существенное влияние на связывание антигена.
Ниже в примере 3 описано производство приведенных в качестве примера гуманизированных антител к ErbB2, которые связывают ErbB2 и блокируют активацию лигандом рецептора ErbB. Представляющее наибольший интерес гуманизированное антитело блокирует опосредуемую EGF, TGF-α и/или HRG активацию МАРК практически столь же эффективно, что и мышиное моноклональное антитело 2С4 (или его Fab-фрагмент). Гуманизированное антитело по изобретению может, например, включать остатки гипервариабельного участка из отличного от человека источника, встроенные в вариабельную человеческую область тяжелой цепи, и может дополнительно включать замену в каркасном участке (FR) в положении, выбранном из группы, включающей 69Н, 71Н и 73Н (используемая система нумерации вариабельной области предложена Kabat с соавторами. Sequences of Proteins of Immunological Interest, 5-ое изд. Public Health Service, National Institutes of Health, Bethesda, MD (1991)). В одном из вариантов осуществления гуманизированное антитело включает замены в FR в двух или во всех положениях 69Н, 71Н и 73Н.
Представляющее интерес приведенное в качестве примера гуманизированное антитело по изобретению включает остатки, определяющие комплементарность вариабельной области тяжелой цепи, такие как GFTFTDYTMX, где Х предпочтительно обозначает D или S (SEQ ID NO:7); DVNPNSGGSIYNQRFKG (SEQ ID NO:8); и/или NLGPSFYFDY (SEQ ID NO:9), небязательно включающие аминокислотные модификации этих CDR-остатков, например, если модификации в значительной степени сохраняют или повышают аффинность антитела. Например, вариант представляющего интерес антитела может иметь от примерно 1 до примерно 7 или примерно 5 аминокислотных замен в указанных выше последовательностях вариабельных CDR тяжелых цепей. Такие варианты можно получать путем созревания антител с выраженной аффинностью, например, согласно описанному ниже методу. Наиболее предпочтительное гуманизированное антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи, представленную в SEQ ID NO:4.
Гуманизированное антитело может включать остатки, определяющие комплементарность вариабельной области легкой цепи, такие как KASQDVSIGVA (SEQ ID NO:10); SASYX1X2X3, где X1 предпочтительно обозначает R или L, X2 предпочтительно обозначает Y или Е и X3 предпочтительно обозначает Т или S (SEQ ID NO:11); и/или QQYYIYPYT (SEQ ID NO:12), например, в дополнение к тем CDR-остаткам вариабельной области тяжелой цепи, которые описаны в предыдущем разделе. Такие гуманизированные антитела необязательно включают аминокислотные модификации указанных выше CDR-остатков, например, в которых модификации в значительной степени сохраняют или повышают аффинность антитела. Например, вариант представляющего интерес антитела может иметь от примерно 1 до примерно 7 или примерно 5 аминокислотных замен в указанных выше последовательностях вариабельных CDR легких цепей. Такие варианты можно получать путем созревания антител с выраженной аффинностью, например, согласно описанному ниже методу. Наиболее предпочтительное гуманизированное антитело содержит аминокислотную последовательность вариабельной области легкой цепи, представленную в SEQ ID NO:3.
Настоящее изобретение также включает созревшие антитела с выраженной аффинностью, которые связываются с ErbB2 и блокируют активацию лигандом рецептора ErbB. Родительское антитело может представлять собой человечеакое антитело или гуманизированное антитело, например, антитело, включающее последовательности вариабельной области легкой и/или тяжелой цепи, которые представлены в SEQ ID NO:3 и SEQ ID NO:4 соответственно (т.е. вариант 574). Созревшие антитела с выраженной аффинностью предпочтительно связываются с рецептром ErbB2 с аффинностью, превышающей аффинность мышиного 2С4 или варианта 574 (например, от примерно 2-кратно или примерно 4-кратно до примерно от 100-кратно до примерно 1000-кратно повышенной аффинностью, например, установленной с помощью ELISA, проведенного с использованием внеклеточного домена ErbB2 (ECD)). Примеры CDR-остатков вариабельной области тяжелой цепи, в которых могут быть произведены замены, включают Н28, H30, Н34, H35, Н64, Н96, Н99 или комбинации из двух или большего количества (например, 2, 3, 4, 5, 6 или 7 этих остатков). Примеры CDR-остатков вариабельной области легкой цепи, в которых могут быть произведены замены, включают L28, L50, L53, L56, L91, L92, L93, L94, L96, L97 или комбинации из двух или большего количества (например, 2, 3, 4, 5, 6 или вплоть до примерно 10 этих остатков).
Различные формы гуманизированного антитела или созревшего антитела с выраженной аффинностью также подпадают под объем изобретения. Например, гуманизированное антитело или созревшее антитело с выраженной аффинностью может представлять собой фрагмент антитела, такой как Fab, который необязательно конъюгируют с одним или несколькими цитотоксическим(и) агентом(ами) с целью получения иммуноконъюгата. В другом варианте гуманизированное антитело или созревшее антитело с выраженной аффинностью может представлять собой интактное антитело, такое как интиктное антитело в виде IgG1.
(IV) Человеческие антитела
Альтернативой гуманизации является получение человеческих антител. Например, в настоящее время можно получать трансгенных животных (например, мышей), которые после иммунизации могут продуцировать полный спектр человеческих антител в отсутствие производства эндогенного иммуноглобулина. Например, было описано, что гомозиготная делеция областей стыка гена тяжелой цепи антитела (JH-сегмента) в химерных мышах и в мышах с мутацией в зародышевой линии приводит к полному ингибированию производства эндогенного антитела. Перенос набора зародышевой линии гена человеческого иммуноглобулина в такую мутантную зародышевую линию мышей приводит к производству человеческих антител после контрольного заражения антигеном (см., например, Jakobovis и др., Proc. Natl. Acad. Sci. USA: 90: 2551 (1993); Jakobovis и др., Nature, 362: 25-258 (1993); Bruggermann и др., Year in Immuno., 7: 33 (1993); и патенты США 5591669, 5589369 и 5545807)).
В альтернативном варианте для получения человеческих антител и фрагментов антител in vitro из спектра генов вариабельной области (V) иммуноглобулина иммунизированных доноров можно использовать технологию проявления с помощью фага (McCafferty и др., Nature, 348: 552-553 (1990)). Согласно этой методике гены V-области антитела клонируют в рамке считывания либо с основным, либо с минорным геном протеина оболочки нитчатого бактериофага, такого как М13 или fd, и проявляют в виде функциональных фрагментов антитела на поверхности фаговой частицы. Поскольку нитчатая частица содержит копию одноцепочечной ДНК фагового генома, отборы, основанные на функциональных свойствах антитела, также приводят к отбору гена, кодирующего антитело, которое обладает этими свойствами. Таким образом, фаг имитирует некоторые свойства В-клетки. Проявление с помощью фага можно осуществлять в различных форматах (обзор которых см., например, у Johnson, Kevin S. и Chiswell David J., Current Opinion in Structural Biology, 3: 564-571 (1993)). Для проявления с помощью фага можно использовать различные источники сегментов V-генов. Clackson и др., Nature, 352: 624-628 (1991) выделили различные наборы антител к оксазолону из небольшой произвольной комбинаторной библиотеки V-генов, полученной из селезенки иммунизированных мышей. Можно сконструирован спектр V-генов из иммунизированных людей-доноров, и антитела к различному набору антигенов (включая аутоантигены) можно выделять в целом согласно методам, описанным у Marks и др., J.Mol. Biol, 222: 581-597 (1991) или у Griffith и др., ЕМВО J., 12: 725-734 (1993) (см. также патенты США 5565323 и 5537905).
Как описано выше, человеческие антитела также могут вырабатываться in vitro активированными В-клетками (см. патенты США 5567610 и 5229275).
Человеческие антитела к ErbB2 описаны в патенте США 5772997, выданном 30 июня 1998 г. и в WO 97/00271, опубликованной 3 января 1997 г.
(V) Фрагменты антител
Для получения фрагментов антител разработаны различные методы. Традиционно эти фрагменты получали путем протеолитического расщепления интактных антител (см., например, Morimoto и др., Journal of Biochemical and Biophysical Methods, 24: 107-117 (1992) и Brennan и др., Science, 229: 81 (1985)). Однако в настоящее время эти фрагменты можно получать непосредственно с помощью рекомбинантных клеток-хозяев. Например, фрагменты антител можно выделять из описанных выше фаговых библиотек антител. Согласно другому варианту Fab'-SH-фрагменты можно непосредственно выделять из Е.coli и химически сшивать с образованием F(ab')2-фрагментов (Carter и др., Bio/Technology, 10: 163-167 (1992)). Согласно другому подходу F(ab')2-фрагменты можно выделять непосредственно из культуры рекомбинантных клеток-хозяев. Специалистам в данной области должны быть очевидны другие методики получения фрагментов антител. В других вариантах осуществления выбранное антитело представляет собой одноцепочечный Fv-фрагмент (scFv) (см. WO 93/16185; патент США 5571894 и патент США 5587458). Фрагмент антитела может также представлять собой "линейное антитело", например, описанное в патенте США 5641870. Такие фрагменты линейного антитела могут быть моноспецифическими или биспецифическими.
(VI) Биспецифические антитела
Биспецифические антитела представляют собой антитела, которые специфично связываются, по меньшей мере, с двумя различными эпитопами. Например, биспецифические антитела могут связывать два различных эпитопа протеина ErbB2. Другие такие антитела могут наряду с сайтом связывания с ErbB2 включать сайт(ы) связывания с EGFR, ErbB3 и/или ErbB4. В альтернативном варианте плечо антитела к ErbB2 можно объединять с плечом, которое связывается со стимулирующей молекулой на поверхности лейкоцита, такой как молекула Т-клеточного рецептора (например, CD2 или CD3), или с Fc-рецептором для IgG (FcγR), таким как FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD 16), в результате чего происходит фокусирование защитных механизмов клетки на экспрессирующей ErbB2 клетке. Биспецифические антитела также могут применяться для локализации цитотоксических агентов в клетках, которые экспрессируют ErbB2. Эти антитела несут ErbB2-связывающее плечо и плечо, которое связывается с цитотоксическим агентом (например, сапорином, антиинтерфероном-α, винка-алкалоидом, цепью рицина А, метотрексатом или с меченым гаптеном). Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител (например, F(ab')2-фрагментов биспецифических антител).
В WO 96/16673 опасано биспецифическое антитело к ErbB2/FcγRIII, а патенте США 5837234 описано биспецифическое антитело к ErbB2/FcγRII. Биспецифическое антитело к ErbB2/Fcα описано в WO 96/16673, а в патенте США 5821 описано биспецифическое антитело к ErbB2/CD3.
Методы создания биспецифических антител известны в данной области. Общепринятое получение полноразмерных биспецифических антител основано на совместной экспрессии двух пар тяжелых и легких цепей иммуноглобулина, где обе цепи имеют различную специфичность (Millstein и др., Nature, 305: 537-539 (1983)). Из-за случайного набора тяжелых и легких цепей иммуноглобулина, эти гибридомы (квадромы) потенциально могут продуцировать смесь из 10 различных молекул антител, из которых только одно имеет правильную биспецифическую структуру. Очистка правильной молекулы, которую обычно осуществляют в несколько стадий с помощью аффинной хроматографии, является довольно трудоемкой, а выход продукта - низким. Аналогичные процессы описаны в WO 93/08829 и у Traunecker и др., ЕМВО J., 10: 3655-3659 (1991).
Согласно другому подходу вариабельные области антитела с требуемой специфичностью связывания (антигенсвязывающие центры антитела) сливают с последовательностями константной области иммуноглобулина. Слияние предпочтительно осуществляют с константной областью тяжелой цепи иммуноглобулина, которая включает, по меньшей мере, часть шарнирной, СН2- и СН3-областей. Предпочтительно, чтобы, по меньшей мере, в одном из слияний присутствовала первая константная область тяжелой цепи (CH1), содержащая сайт, необходимый для связывания легкой цепи. ДНК, кодирующие слияния тяжелой цепи иммуноглобулина и при необходимости легкой цепи иммуноглобулина, встраивают в различные экспрессионные векторы и ими совместно трансфектируют приемлемый организм-хозяин. Это обеспечивает большую гибкость в подборе общих пропорций трех полипептидных фрагментов в вариантах, когда в конструкции используют неравные соотношения трех полипептидных цепей с целью оптимизации выходов. Однако также можно встраивать кодирующие последовательности в две или во все три полипептидные цепи в одном экспрессионном векторе, когда экспрессия, по меньшей мере, двух полипептидных цепей в равных пропорциях обеспечивает высокие выходы или когда соотношения не имеют решающего значения.
В предпочтительном варианте осуществления указанного подхода биспецифические антитела включают гибрид тяжелой цепи иммуноглобулина, обеспечивающий первую специфичность связывания в первом плече, и гибрид пары тяжелая цепь - легкая цепь иммуноглобулина (что обеспечивает вторую специфичность связывания) во втором плече. Было обнаружено, что эта асимметричная структура облегчает отделение требуемого биспецифического соединения от нежелательных комбинаций цепей иммуноглобулинов, поскольку в присутствии легкой цепи иммуноглобулина только в одной половине биспецифической молекулы имеется возможность облегчения разделения. Этот подход описан в WO 94/04690. Дополнительные детали получения биспецифических антител можно обнаружить, например, у Suresh и др., Methods in Enzymology, 121: 210 (1986).
Согласно следующему подходу, описанному в патенте США 5731168, можно сконструировать область контакта между парой молекул антитела с целью повышения до максимального уровня процента гетеродимеров, которые получают из культуры рекомбинантных клеток. Предпочтительная область контакта включает, по меньшей мере, часть СH3-домена константной области антитела. Согласно этому методу одну или несколько небольших аминокислот с боковыми цепями из области контакта первой молекулы антитела заменяют более крупными молекулами с боковыми цепями (например, тирозин или триптофан). Уравновешивающие "полости", идентичные или близкие по размеру большой(им) боковой(ым) цепи(ям) создают в области контакта второй молекулы антитела путем замены аминокислот с крупными боковыми цепями на более мелкие (например, аланин или треонин). Это обеспечивает механизм, способствующий повышению выхода гетеродимера относительно других нежелательных конечных продуктов, таких как гомодимеры.
Биспецифические антитела включают поперечно сшитые антитела или "гетероконъюгаты". Например, одно из антител в гетероконъюгате может быть сшито с авидином, а другое с биотином. Такие антитела можно использовать, например, для направления клеток иммунной системы к нежелательным клеткам (патент США 467690) и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/200373 и ЕР 03089). Антитела-гетероконъюгаты можно создавать с помощью любого из обычных методов введения перекрестных сшивок. Приемлемые кросс-линкеры хорошо известны в данной области и описаны в патенте США 4676980 наряду с различными методами введения перекрестных сшивок.
Методы получения биспецифических антител из фрагментов антител также описаны в литературе. Например, биспецифические антитела можно получать с помощью химического связывания. Brennan и др., Science, 229: 81 (1985) описали методику, согласно которой интактные антитела подвергают протеолитическому расщеплению, получая F(ab')2-фрагменты. Эти фрагменты восстанавливают в присутствии агента, образующего комплексы с дитиолом, такого как арсенит натрия, для стабилизации соседних дитиолов и предупреждения образования дисульфидных связей. Полученные Fab'-фрагменты затем превращают в тионитробензоатное (TNB) производное. Одно из Fab'-TNB-производных затем повторно превращают в Fab'-тиол путем восстановления меркаптоэтиламином и смешивают с эквимолярным количеством другого Fab'-'TNB-производного, получая биспецифическое антитело. Полученные биспецифические антитела могут применяться в качестве агентов для избирательной иммобилизации ферментов.
Достигнутый в настоящее прогресс позволяет облегчить непосредственное выделение Fab'-SH-фрагментов из Е.coli, которые можно химически сшивать с образованием биспецифических антител. Shalaby и др., J.Exp. Med., 175: 217-225 (1992) описали получение F(ab')2-фрагмента полностью гуманизированного биспецифического антитела. Каждый Fab'-фрагмент по отдельности секретировался из Е.coli и его подвергали непосредственному химическому связыванию in vitro с получением биспецифического антитела. Полученное таким образом биспецифическое антитело обладало способностью связывать клетки, для которых характерна сверхэкспрессия рецептора ErbB2, и с нормальными человеческими Т-клетками, а также стимулировать литическую активность человеческих цитотоксических лимфоцитов, мишенью которых является опухоль молочной железы человека.
Также описаны различные методы получения и выделения фрагментов биспецифических антител непосредственно из культуры рекомбинантных клеток. Например, биспецифические антитела получали с помощью лейциновых "застежек-молний" (Kostelny и др., J.Immunol., 148(5): 1547-1553 (1992)). Пептиды лейциновых "застежек-молний" из протеинов Fos и Jun связывали с Fab'-фрагментами двух различных антител путем генного слияния. Гомодимеры антител восстанавливали в шарнирной области с получением мономеров, а затем путем повторного окисления получали гетеродимеры антител. Этот метод может применяться для получения гомодимеров антител. Технология "двойных антител", описанная Hollinger и др., Proc. Natl. Acad. Sci. USA: 90: 6444-6448 (1993) представляет собой другой механизм получения фрагментов биспецифических антител. Фрагменты включают вариабельную область тяжелой цепи (VH), связанную с вариабельной областью легкой цепи (VL) линкером, который является слишком коротким, для того чтобы позволить произойти спариванию между двумя областями одной и той же цепи. Таким образом, VH- и VL-области одного фрагмента должны спариваться с комплементарными VL- и VH-областями другого фрагмента, тем самым образуя два антигенсвязывающих центра. Также описана другая стратегия получения фрагментов биспецифических антител, основанная на применении одноцепочечных (Fv) (sFv) димеров (см. Gruber и др., J.Immunol., 152: 5368 (1994)).
Под объем изобретения также подпадают антитела с более чем двумя валентностями. Например, могут быть получены триспецифические антитела (Tutt и др., J.Immunol., 147: 60 (1991)).
(VII) Другие модификации аминокислотных последовательностей
Приведенная(ые) в описании модификация(и) аминокислотных последовательностей антител к ErbB2 подпадает(ют) под объем изобретения. Например, может требоваться улучшение аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотных последовательностей антитела к ErbB2 получали путем введения соответствующих нуклеотидных замен в нуклеиновую кислоту антитела к ErbB2 или путем пептидного синтеза. Такие модификации включают, например, делеции и/или инсерции и/или замены остатков в аминокислотных последовательностях антитела к ErbB2. В конечной конструкции можно осуществлять любую комбинацию делеции, инсерции и замен, при условии, что конечная конструкция будет обладать требуемыми характеристиками. Амнокислотные замены также могут изменить посттрансляционные процессы антитела к ErbB2, такие как изменение количества или положения сайтов гликозилирования.
Пригодный метод для идентификации определенных остатков или областей антитела к ErbB2, которые имеют предпочтительные местоположения для мутагенеза, называемый "мутагенез на основе сканирования аланина" описан Cunningham и Wells, Sciente, 244: 1081-1085 (1989). Согласно этому методу идентифицируют остаток или группу остатков-мишеней (например, заряженных остатков, таких как arg, asp, his, lys и glu) и заменяют нейтральной или отрицательно заряженной аминокислотой (наиболее предпочтительно аланин или полиаланин), оказывая тем самым воздействие на взаимосвязь аминокислот с антигеном ErbB2. Те локализации аминокислот, для которых установлена функциональная чувствительность к заменам, затем подвергают усовершенствованию путем введения дополнительных или других вариантов сайтов, в которых производят замену. Таким образом, в то время как сайт, в который вводят вариант аминокислотной последовательности, является предопределенным, не требуется, чтобы природа мутации per se была предопределена. Например, для анализа характеристики мутации данного сайта осуществляют мутагенез на основе сканирования аланина или неспецифический мутагенез кодона- или области-мишени и проводят скрининг экспрессируемого варианта антитела к ErbB2 в отношении требуемой активности.
Вставки аминокислотных последовательностей включают N- и/или С-концевые слияния, размер которых варьируется по длине от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутрь последовательности одного или нескольких аминокислотных остатков. Примеры концевых вставок включают антитело к ErbB2 с N-концевым метионильным остатком или антитело, слитое с цитотоксическим полипептидом. Другие варианты вставок молекулы антитела к ErbB2 включают слияние N- или С-конца антитела к ErbB2 с ферментом (например, ADEPT) или полипептидом, который повышает продолжительность полужизни антитела в сыворотке.
Другим типом варианта является вариант аминокислотной замены. Эти варианты включают замену, по меньшей мере, одного аминокислотного остатка в молекуле антитела к ErbB2 на другой остаток. Сайты, представляющие собой наибольший интерес для мутагенеза методом замены, включают гипервариабельные участки, но также могут быть созданы изменения в FR. Консервативные замены приведены в таблице 1, под заголовком "Предпочтительные замены". Если такие замены приводят к изменению биологической активности, то могут быть введены более существенные замены, обозначенные как "Примеры замен" в таблице 1, или другие замены, описанные ниже при ссылке на классе аминокислот, и затем продукты могут быть подвергнуты скринингу.
Существенных модификаций биологических свойств антитела достигают путем отбора замен, которые в значительной степени различаются по их воздействию на поддержание (а) структуры каркаса полипептида в области замены, например, β-складчатой или спиральной конформации, (б) заряда или гидрофобности молекулы в сайте-мишени или (в) размера боковой цепи. Встречающиеся в естественных условиях остатки подразделяют на группы на основе общих свойств боковых цепей:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: cys, ser, thr;
(3) кислые: asp, glu;
(4) основные: asn, gln, his, lys, arg;
(5) остатки, которые влияют на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены предусматривают замену представителя одного из этих классов на представителя другого класса.
Любой из остатков цистеина, не участвующий в поддержании соответствующей конформации антитела к ErbB2, также может быть заменен, как правило, серином, с целью повышения устойчивости молекулы к окислению и предупреждения аномального поперечного сшивания. И наоборот, цистеиновая(ые) связь(и) может(гут) быть введена(ы) в антитела для повышения его стабильности (особенно, если антитело представляет собой фрагмент антитела, такой как Fv-фрагмент).
Особенно предпочтительный тип варианта замены включает замену одного или нескольких остатков гипервариабельного участка родительского антитела (например, гуманизированного или человеческого антитела). Как правило, полученный(ые) в результате вариант(ы), отобранные для дальнейшего усовершенствования, должны обладать улучшенными биологическими свойствами относительно родительского антитела, из которого оно(они) пролучено(ы). Приемлемый путь получения таких созревших вариантов с выраженной аффинностить на основе замены включает использование проявления с помощью фага. В целом метод состоит в следующем: несколько сайтов гипервариабельного участка (например, 6-7 сайтов) подвергают мутации, получая все возможные аминокислотные замены в каждом сайте. Полученные таким образом варианты антитела проявляют в одновалентой форме на поверхности частиц нитчатого фага в виде слияний с продуктом гена III фага М13, который упакован в каждой частице. Варианты, которые проявляются фагом, затем подвергают скринингу в отношении их биологической активности (например, аффинности к связыванию) согласно методикам, приведенным в настоящем описании. Для того чтобы выявить перспективные с точки зрения модификации сайты гипервариабельного участка, можно осуществлять мутагенез на основе сканирования аланина с целью идентификации остатков гипервариабельного участка, которые наиболее важны для связывания антигена. В альтернативном или дополнительном варианте может оказаться целесообразным анализировать кристаллическую структуру комплекса антиген-антитело с целью выявления точек соприкосновения между антителом и человеческим ErbB2. Такие остатки в точках соприкосновения и соседние остатки является кандидатами для замены с помощью разработанных методов. После получения таких вариантов панель вариантов подвергают скринингу согласно представленному в данном описании методу, и антитела с улучшенными свойствами по данным одного или нескольких соответствующих анализов отбирают для дальнейшей разработки.
Другой тип аминокислотных вариантов антитела изменяет исходную схему гликозилирования антитела. Под изменением понимают делецию одного или нескольких углеводных фрагментов, присутствующих в антителе, и/или добавление одного или нескольких сайтов гликозилирования, которые в норме не присутствуют в антителе.
Гликозилирование антител обычно происходит посредством либо N-связывания, либо O-связывания. N-связывание предусматривает присоединение углеводного фрагмента к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, где Х обозначает любую аминокислоту кроме пролина, представляют собой распознаваемые последовательности, предназначенные для ферментативного присоединения углеводного фрагмента к боковой цепи остатка аспарагина. Так, присутствие любой их этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. O-связанное гликозилирование предусматривает присоединение одного из сахаров, таких как N-ацетилгалактозамин, галактоза или ксилоза, к гидроксиаминокислоте, наиболее часто к серину или треонину, хотя также могут применяться 5-гидроксипролин или 5-гидроксилизин.
Введение дополнительных сайтов гликозилирования в антитело можно осуществлять путем такого изменения аминокислотной последовательности, чтобы она содержала одну или несколько описанных выше трипептидных последовательностей (для сайта N-гликозилирования). Изменение также может включать добавление или замену одного или нескольких остатков серина или треонина в последовательности исходного антитела (для сайтов O-гликозилирования).
Молекулы нуклеиновых кислот, кодирующие варианты аминокислотных последовательностей антитела к ErbB2, получают различными известными в данной области методами. Эти методы включают (но не ограничиваясь ими) выделение из природного источника (в случае встречающихся в естественных условиях вариантов аминокислотных последовательностей) или получение с помощью сайтнаправленного мутагенеза с использованием олигонуклеотидов (или сайтнаправленного мутагенеза), ПЦР-мутагенез и мутагенез с использованием кассеты ранее полученного варианта или не включенной в указанные варианты версии антитела к ErbB2.
Может оказаться желательным модифицировать антитело по изобретению в отношении его эффекторной функции, например, с тем, чтобы повысить опосредванную антителом клеточную цитотоксичность (ADCC) и/или зависящую от комплемента цитотоксичность (CCD) антитела. Это может достигаться путем введения одной или нескольких аминокислотных замен в Fc-область антитела. В альтернативном или дополнительном варианте остаток(тки) цистеина может(гут) быть интродуцированы в Fc-область, что приводит к образованию в этой области дисульфидной связи между цепями. Полученное таким образом гомодимерное антитело может обладать улучшенной способностью к интернализации и/или повышенной зависящей от комплемента цитотоксичностью и опосредванной антителом клеточной цитотоксичностью (ADCC) (см. Caron и др., J.Exp. Med., 176: 1191-1195 (1992) и Shopes, J.Immunol., 148: 2918-2922 (1992)). Гомодимерные антитела с повышенной противоопухолевой активностью также могут быть получены с использованием гетеробифункциональных кросс-линкеров, согласно методике, описанной у Wolff и др., Cancer Research, 53: 2560-2565 (1993). В другом варианте может быть сконструировано антитело, имеющее удвоенное количество Fc-областей, которое вследствие этого может иметь повышенные способности к зависящему от комплемента лизису и ADCC (см. Stevenson и др., Ant-Cancer Drug Design, 3:219-230 (1989)).
Для увеличения времени полужизни антитела в сыворотке в антитело может быть включен связывающий рецептор эпитоп-"спасатель" (особенно в фрагмент антитела), например, согласно методу, описанному в патенте США 5739277. В контексте настоящего описания понятие "связывающий рецептор эпитоп-"спасатель" относится к эпитопу Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), который ответствен за увеличение времени полужизни в сыворотке in vivo молекулы IgG.
(VIII) Скрининг антител с требуемыми свойствами
Выше описаны методы получения антител. Далее при необходимости можно осуществлять дополнительный отбор антител с определенными биологическими характеристиками.
Для идентификации антитела, которое блокирует активацию лигандом рецептора ErbB, можно определять способность антитела блокировать связывание лиганда с клетками, экспрессирующими рецептор ErbB (например, в конъюгатах ErbB с другим рецептором ErbB, с которым представляющий интерес рецептор ErbB образует гетероолигомеры ErbB). Например, клетки, которые в естественных условиях экспрессируют или которые трансфектированы, для того чтобы экспрессировать рецепторы ErbB гетероолигомера ErbB, можно инкубировать с антителом и затем обрабатывать меченым лигандом ErbB. После этого можно оценивать способность антитела к ErbB2 блокировать связывание лиганда с рецептором ErbB в гетероолигомере ErbB.
Например, ингибирование связывания HRG с линиями клеток опухоли молочной железы MCF7 антителами к ErbB2 можно осуществлять с использованием монослойных культур MCF7 на льду в формате 24-луночных планшетов в целом согласно методу, описанному ниже в примере 1. Моноклональные антитела к ErbB2 можно добавлять в каждую лунку и инкубировать в течение 30 мин. Затем можно добавлять 125I-rHRGβ1177-244 (25 пм) и инкубацию продолжать в течение 4-16 ч. На основании этих данных могут быть построены графики зависимости реакции от дозы и рассчитаны значения IC50 для представляющего интерес антитела. Согласно одному из вариантов антитело, которое блокирует активацию лигандом рецептора ErbB, в этом опыте имеет значение IC50 для ингибирования связывания HRG с клетками линии MCF7 примерно 50 нМ или ниже, более предпочтительно 10 нМ или ниже. Если антитело представляет собой фрагмент антитела, такой как Fab-фрагмент, то значение IC50 для ингибирования связывания HRG с клетками линии MCF7 в этом опыте составляет примерно 100 нМ или ниже, более предпочтительно 50 нМ или ниже.
С использованием альтернативного или дополнительного варианта способа можно оценить способность антитела к ErbB2 блокировать стимулируемое лигандом ErbB фосфорилирование тирозина рецептора ErbB, присутствующего в гетероолигомере ErbB. Например, клетки эндогенно экспрессирующие рецепторы ErbB или трансфектированные с целью их экспрессии, можно инкубировать с антителом и затем оценивать зависящую от лиганда ErbB активность в отношении фосфорилирования тирозина с помощью моноклонального антитела к фосфотирозину (необязательно конъюгированного с обнаруживаемой меткой). Анализ активации киназного рецептора, описанный в патенте США 5766863, также можно применять для определения активации рецептора ErbB и блокады антителом его активности.
Согласно одному из вариантов способа можно осуществлять скрининг в отношении антитела, которое ингибирует стимуляцию HRG фосфорилирования тирозина р180 в клетках линии MCF7, в целом согласно методу, описанному ниже в примере 1. Например, клетки линии MCF7 можно высевать в 24-луночные планшеты и добавлять в каждую лунку моноклональные антитела к ErbB2 и инкубировать с течение 30 мин при комнатной температуре; затем можно в каждую лунку добавлять rHRGβ1177-244 до конечной концентрации 0,2 нМ и инкубацию продолжать в течение 8 мин. Среды могут быть удалены аспирацией из каждой лунки и реакции прекращены добавлением 100 мкл образца ДСН-буфера (5% ДСН, 25 мМ ДТТ и 25 мМ Трис-HCl, рН 6,8). Каждый образец (25 мкл) после электрофореза в 4-12%-ном градиентном геле (фирма Novex) может быть электрофоретически пересен на поливинилиденфторидную мембрану. Можно получать иммуноблоты антитела к фосфотирозину (1 мкг/мл) и интенсивность активной основной полосы, соответствующей Mr примерно 180000 тысяч количество оценивать с помощью денситометра для измерения оптической плотности в отраженном свете. {PRIVATE } В этом анализе выбранное антитело предпочтительно должно в значительной степени ингибировать стимуляцию HRG фосфорилирования тирозина р180, по меньшей мере, примерно до 0-35% по сравнению с контролем. На основании этих данных, полученных с помощью денситометрии в отраженном свете, могут быть построены графики зависимости ингибирования стимуляции HRG фосфорилирования тирозина р180 от дозы и рассчитаны значения IC50 для представляющего интерес антитела. Согласно одному из вариантов осуществления в этом анализе для антитела, которое блокирует активацию лигандом рецептора ErbB, значение IC50 для ингибирования стимуляции HRG фосфорилирования тирозина р180 составляло примерно 50 нМ или менее, более предпочтительно 10 нМ или менее. Если антитело представляло собой фрагмент антитела, такой как Fab-фрагмент, то в этом анализе значение IC50 для ингибирования стимуляции HRG фосфорилирования тирозина р180 составляло, например, примерно 100 нМ или менее, более предпочтительно 50 нМ или менее.
Также можно оценить ингибирующие воздействия антитела на рост клеток линии MDA-MB-175, например, в основном согласно методу, описанному у Schaefer и др., Oncogene, 15: 1385-1394 (1997). Согласно этому анализу клетки линии MDA-MB-175 можно обрабатывать моноклональным антителом к ErbB2 (10 мкг/мл) в течение 4 дней и окрашивать кристаллическим фиолетовым. После инкубации с антителом к ErbB2 было выявлено ингибирующее действие на рост этой линии клеток, аналогичное действию моноклонального антитела 2С4. Согласно еще одному варианту экзогенный HRG не должен оказывать существенного обратного воздействия на ингибирование. Предпочтительно антитело обладает способностью ингибировать пролиферацию клеток линии MDA-MB-175 в большей степени, чем моноклональное антитело 4D5 (и необязательно в большей степени, чем моноклональное антитело 7F3), как в присутствии экзогенного HRG, так и без него.
Согласно одному из вариантов представляющее интерес антитело к ErbB2 может блокировать зависящее от херегулина связывание ErbB2 с ErbB3 как в клетках линии MCF7, так и SK-BR-3, что установлено в эксперименте по коиммунопреципитации, описанном в примере 2, более эффективно по сравнению с моноклональным антителом 4D5 и предпочтительно более эффективно по сравнению с моноклональным антителом 7F3.
Для выявления ингибирующих рост антител к ErbB2 можно осуществлять скрининг в отношении антител, которые ингибируют рост раковых клеток, обладающих способностью сверхэкспрессировать ErbB2. Согласно одному из вариантов выбранное ингибирующее рост антитело обладает способностью ингибировать рост клеток линии SK-BR-3 в культуре клеток примерно на 20-100% и предпочтительно примерно на 50-100% при использовании концентрации примерно от 0,5 до 30 мкг/мл. Для выявления таких антител можно осуществлять анализ с использованием клеток линии SK-BR-3, описанный в патенте США 5677171. Согласно этому анализу клетки линии SK-BR-3 выращивают в смеси 1:1 среды F12 и среды DMEM, дополненной 10% фетальной бычьей сыворотки, глутамином и пенициллином и стрептомицином. Клетки линии SK-BR-3 высевают из расчета 20000 клеток на 35-миллиметровую чашку для культивирования (чашка: объем 2 мл/диаметр 35 мм). В чашку добавляют от 0,5 до 30 мкг/мл антитела к ErbB2. Через 6 дней с помощью электронного счетчика клеток типа COULTER™ подсчитывают количество клеток по сравнению с необработанным контролем. В качестве антител, обладающих способностью ингибировать рост, выбирают антитела, которые ингибируют рост клеток линии SK-BR-3 примерно на 20-100% или примерно на 50-100%.
Для отбора антител, которые индуцируют гибель клеток, можно осуществлять оценку потери целостности мембраны, например, по поглощению PI, трипанового синего или 7AAD в сравнении с контролем. Предпочтительный анализ основан на оценке поглощения PI с использованием клеток линии ВТ474. Согласно этому анализу клетки линии ВТ474 (которые могут быть получены из Американской коллекции типовых культур (Роквилл, штат Мэриленд)), культивируют в смеси (50:50) модифицированная по способу Дульбекко среда Игла (D-MEM):среда Хама F-12, дополненной 10% термически активированной ФБС (фирма Hyclone) и 2 мМ глутамином. (Таким образом, этот анализ осуществляют в отсутствие комплемента и иммунокомпетентных эффекторных клеток). Клетки линии ВТ474 высевают с плотностью 3х106/на чашку в чашки для культивирования размером 100х20 мм и дают прикрепиться в течение ночи. Затем среду удаляют и заменяют только свежей средой или средой, содержащей 10 мкг/мл соответствующего моноклонального антитела. Клетки инкубируют в течение 3 дней. После каждой обработки монослои промывают ЗФР и отщепляют путем трипсинизации. Затем клетки центрифугируют при 12000 об/мин в течение 5 мин при 4°С, дебрис ресуспендируют в 3 мл охлажденного на льду связывающего Са++ буфера (10 мМ Hepes, pH 7,4, 140 мМ NaCl, 2,5 мМ CaCl2) и аликвоты вносят в 33-миллиметровые пробирки размером 12×75, закрытые фильтром (1 мл на пробирку, по 3 пробирки на каждую обрабатываемую группу), для удаления скоплений клеток. Затем в пробирки вносят PI (10 мкг/мл). Образцы можно анализировать с помощью проточного цитометратипа FACSCAN™ и программного обеспечения FACSCONVERT™ CellQuest (фирма Becton Dickinson). В качестве антител, которые индуцируют гибель клеток, отбирают антитела, которые индуцируют статистически значимые уровни гибели клеток при оценке по степени поглощения PI.
Для отбора антител, которые индуцируют апоптоз, можно применять анализ, основанный на связывании аннексина с клетками линии ВТ474. Клетки линии ВТ474 культивируют и высевают в чашки, описанные в предыдущем разделе. Затем среду удаляют и заменяют только свежей средой или средой, содержащей 10 мкг/мл моноклонального антитела. После трехдневной инкубации монослои промывают ЗФР и отщепляют путем трипсинизации. Затем клетки центрифугируют, ресуспендируют в связывающем Са++ буфере и аликвоты вносят в пробирки, как описано выше для анализа гибели клеток. Затем в пробирки вносят меченый аннексин (например, аннексин V-FTIC) (1 мкг/мл). Образцы можно анализировать с помощью проточного цитометра типа FACSCAN™ и программного обеспечения FACSCONVERT™ CellQuest (фирма Becton Dickinson). В качестве антител, которые индуцируют апоптоз, отбирают антитела, которые индуцируют статистически значимые уровни связывания аннексина по сравнению с контролем.
Помимо анализа, основанного на оценке связывания аннексина, можно применять метод, основанный на окрашивании ДНК с использованием клеток линии ВТ474. Для осуществления этого анализа клетки линии ВТ474, которые были обработаны представляющим интерес антителом согласно методике, описанной в двух приведенных выше разделах, инкубируют с 9 мкг/мл HOECHST 33342™ в течение 2 ч при 37°С, затем анализируют с помощью проточного цитометра типа EPICS ELITE™ (фирма Coulter Corporation), используя программное обеспечение MODFITLT™ (фирма Verity Software House). С помощью этого анализа в качестве проапоптозных антител отбирают антитела, индуцирующие изменение процента апоптозных клеток, которое в 2 раза или более (и предпочтительно в 3 раза или более) превышает соответствующую величину для необработанных клеток (вплоть до 100% апоптозных клеток).
Для отбора антител, которые связываются с эпитопом на ErbB2, связанном с представляющим интерес антителом, можно осуществлять общепринятый анализ поперечного связывания, например, описанный в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, ред-ры Harlow и David Lane (1988). В альтернативном или дополнительном варианте можно осуществлять картирование эпитопа с использованием методов, хорошо известных в данной области (см. фиг.1А и 1Б настоящего описания).
(IX) Иммуноконъюгаты
Изобретение также относится к иммуноконъюгатам, которые включают антитело, конъюгированное с цитотоксическим агентом, таким как химиотерапевтический агент, токсин (например, токсин, имеющий небольшой размер молекул, или обладающий ферментативной активностью токсин бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты) или радиоактивный изотоп (т.е. радиоконъюгат).
Химиотерапевтические агенты, которые могут применяться для получения таких иммуноконъюгатов, описаны выше. Под объем изобретения также подпадают конъюгаты антитела и одного или нескольких токсинов, имеющих небольшой размер молекул, таких как калихеамицин, маитансин (патент США 5208020), трихотен и СС1065.
Согласно одному из предпочтительных вариантов осуществления изобретения антитело конъюгируют с одной или несколькими молекулами маитансина (например, от примерно 1 до примерно 10 молекул маитансина на молекулу антитела). Маитансин, например, можно превратить в May-SS-Me, который можно восстановить до May-SH3 и подвергнуть взаимодействию с модифицированным антителом (Chari и др., Cancer Research, 52: 127-131 (1992)) с получением иммуноконъюгата маитансиноид-антитело.
Другой представляющий интерес иммуноконъюгат включает антитело к ErbB2, конъюгированное с одной или несколькими молекулами калихеамицина. Представители семейства калихеамициновых антибиотиков в субпикомолярных концентрациях обладают способностью вызывать разрывы двухцепочечных ДНК. Структурные аналоги калихеамицина, которые могут применяться, включают (но не ограничиваясь ими)γ1 1', α2 1, α3 1, N-ацетил-γ1 1, PSAG и θ1 1 (Hinman и др., Cancer Research, 58: 2925-2928 (1998)) (см. также патенты США 5714586; 5712374; 5264586 и 5773001, полностью включенные в настоящее описание в качестве ссылки).
Обладающие ферментативной активностью токсины и их фрагменты, которые могут применяться, включают цепь А дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модеццина, альфа-сарцин, протеины Aleurites fordii, протеины диантина, протеины Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, цурцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены (см., например, WO 93/21232, опубликованную 28 октября 1993 г.).
Настоящее изобретение также относится к иммуноконъюгату, включающему антитело и соединение, обладающее нуклеолитической активностью (например, рибонуклеазу или ДНК-эндонуклеазу, такую как дезоксирибонуклеаза; ДНКаза).
Радиоактивные различные изотопы могут применяться для получения радиоконъюгатов антител к ErbB2. Их примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и радиоактивные изотопы Lu.
Конъюгаты, включающие антитело и цитоксический агент, могут быть созданы с использованием целого ряда бифункциональных протеинсвязывающих агентов, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат, иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат•HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бисазидосоединения (такие как бис(пара-азидобензоил)гександиамин), производные бисдиазония (такие как бис(пара-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицина может быть получен согласно методу, описанному у Vitetta и др., Science, 238: 1098 (1987). Меченная с помощью С14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является примером хелатирующего агента для конъюгации радионуклеотида с антителом (см. WO 94/11026). Линкер может представлять собой "расщепляемый линкер", который облегчает высвобождение цитотоксического лекарственного средства в клетке. Например, может применяться неустойчивый в кислой среде линкер, чувствительный к действию пептидаз линкер, диметильный линкер или содержащий дисульфидную связь линкер (Chari и др., Cancer Research, 52: 127-131 (1992)).
В другом варианте слитый протеин, содержащий антитело к ErbB2 и цитотоксический агент, может быть создан, например, с помощью методов рекомбинантной ДНК или пептидным синтезом.
Согласно еще одному варианту осуществления может быть создан конъюгат антитела с "рецептором" (таким как стрептавидин) с целью применения для предварительной обработки опухоли, когда конъюгат антитело-рецептор вводят пациенту с последующим удалением несвязанного конъюгата из кровотока с помощью реагента для осветления и затем вводят "лиганд" (например, авидин) в виде конъюгата с цитотоксическим агентом (например, радионуклеотидом).
(X) Терапия с использованием пролекарства, активируемого ферментом, зависящим от антитела (ADEPT)
Антитела по настоящему изобретению также могут применяться для ADEPT путем конъюгации антитела с пролекарством, активируемым ферментом, который превращает пролекарство (такое как пептидильный химиотерапевтический агент, см. WO 81/01145) в активное противораковое лекарство (см., например, WO 88/07378 и патент США 4975278).
Компонент конъюгата, представляющий собой фермент, который может применяться для ADEPT, включает любой фермент, обладающий способностью оказывать такое воздействие на пролекарство, в результате которого оно превращается в более активную цитотоксическую форму.
Ферменты, которые могут применяться в способе по изобретению, включают (но не ограничиваясь ими) щелочную фосфатазу, которая может превращать фосфатсодержащие пролекарства в свободные лекарства; арилсульфатазу, которая может превращать сульфатсодержащие пролекарства в свободные лекарства; цитозиндеаминазу, которая может превращать нетоксичный 5-флуорцитозин в противораковое лекарство, 5-флуорацил; протеазы, такие как протеаза Serratia, термолизин, субтилизин, карбоксипептидазы и катепсины (такие как катепсины В и L), которые могут применяться для превращения пептидсодержащих пролекарств в свободные лекарства; D-аланилкарбоксипептидазы, которые могут применяться для превращения пролекарств, которые содержат D-аминокислотные заместители; расщепляющие углеводы ферменты, такие как β-галактозидаза и нейраминидаза, которые могут применяться для превращения гликозилированных пролекарств в свободные лекарства; β-лактамаза, которая может применяться для превращения лекарств, являющихся производными β-лактамов, в свободные лекарства; и амидазы пенициллинов, такие как амидаза пенициллина V или амидаза пенициллина G, которые могут превращать лекарства, дериватизированные по азотам амина с помощью феноксиацетильных или фенилацетильных групп соответственно, в свободные лекарства. В другом варианте антитела, обладающие ферментативной активностью, также известные в данной области под названием "каталитические антитела (абзимы)", могут применяться для превращения пролекарств по изобретению в свободные активные лекарства (см., например, Massey, Nature, 328: 457-458 (1987)). Конъюгаты антитела-абзимы могут быть получены согласно представленному в данном описании способу введения абзимов в популяцию опухолевых клеток.
Ферменты по изобретению можно ковалентно связывать антитела к ErbB2 с помощью методов, хорошо известных в данной области, например, основанных на применении описанных выше гетеробифункциональных перекрестосшивающих реагентов. В другом варианте с помощью хорошо известных в данной области методов рекомбинантной ДНК можно конструировать слитые протеины, содержащие, по меньшей мере, один антигенсвязывающий центр антитела по изобретению, соединенный, по меньшей мере, с одной функционально активной частью фермента по изобретению (см., например, Neuberger и др., Nature, 312: 604-608 (1984)).
(XI) Другие модификации антител
Ниже описаны другие модификации антител. Например, антитело может связывать однин из множества небелковых полимеров, например полиэтиленгликолем, полипропиленгликолем, полиоксиалкиленами или сополимерами полиэтиленгликоля и полипропиленгликоля. Антитело также может представлять собой инкапсулированное в микрокапсулы антитело, которые получают, например, с помощью методов коацервации или межфазной полимеризации (например, микрокапсулы из гидроксиметилцеллюлозы или желатина и поли(метилметацилата) соответственно), в коллоидных системах для введения лекарственных средств (например, в липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие методы описаны в Remington's Pharmaceutical Science, 16-oe изд., ред. Oslo A.
Антитела к ErbB2, представленные в настоящем описании, также можно включать в формы типа иммунолипосом. Содержащие антитела липосомы получают с помощью хорошо известных в данной области методов, описанных у Epstein и др., Proc. Natl. Acad. Sci. USA: 82: 3688 (1985); Hwang и др., Proc. Natl. Acad. Sci. USA: 77: 4030 (1980); патентах США 4485045 и 4544545; и в WO 97/38731, опубликованной 23 октября 1997 г. Липосомы с удлиненным временем жизни в кровотоке описаны в патенте США 5013556.
Особенно предпочтительные липосомы могут быть получены методом упаривания с обратной фазой с использованием липидной композиции, включающей фосфатидилхолин, холестерин и ПЭГ-дериватизированный фосфатидилэтаноламин (ПЭГ-ФЭ). Липосомы экструдируют через фильтры с определенным размером пор, получая липосомы требуемого диаметра. Fab'-фрагменты антитела по настоящему изобретению могут быть включены в липосомы согласно методу, описанному у Martin и др., J.Biol. Che., 257: 286-288 (1982), посредством реакции образования дисульфидной связи. В липосомы необязательно может быть включен химиотерапевтический агент (см. Gabizon и др., J.National Cancer Inst., 81(19): 1484 (1989)).
III. Векторы, клетки-хозяева и методы рекомбинации
Изобретение также относится к выделенной нуклеиновой кислоте, кодирующей гуманизированное антитело к ErbB2, векторам и клеткам-хозяевам, содержащим нуклеиновую кислоту, и к методам рекомбинации для получения антитела.
Для рекомбинантного получения антитела кодирующую его нуклеиновую кислоту выделяют и встраивают в способный к репликации вектор для дальнейшего клонирования (амплификация ДНК) или для экспрессии. ДНК, кодирующая моноклональное антитело, может быть легко выделена и секвенирована с использованием общепринятых методик (например, с помощью олигонуклеотидных зондов, которые обладают способностью специфично связывать гены, кодирующие тяжелые и легкие цепи антитела). Могут применяться многочисленные векторы. Векторы обычно включают один или несколько из следующих компонентов (но не ограничиваясь ими): сигнальную последовательность, сайт инициации репликации, один или несколько маркерных генов, энхансер, промотор и терминатор транскрипции.
(I) Компонент, представляющий собой сигнальную последовательность
Антитело к ErbB2 по изобретению может быть получено с помощью рекомбинации не только индивидуально, но также в виде слитого полипептида с гетерологичным полипептидом, который предпочтительно представляет собой сигнальную последовательность или другой полипептид, несущий специфичный сайт расщепления на N-конце зрелого протеина или полипептида. Выбранная гетерологичная сигнальная последовательность предпочтительно представляет собой последовательность, которая распознается клеткой-хозяином и процессируется в ней (т.е. расщепляется сигнальной пептидазой). Для прокариотических клеток-хозяев, которые не распознают и в которых не процессируется сигнальная последовательность нативного антитела к ErbB2, сигнальную последовательность заменяют на прокариотическую сигнальную последовательность, выбранную, например, из группы, включающей щелочную фосфатазу, пенициллиназу, lpp, или лидерные последовательности термостабильного энтеротоксина II. Для секреции из дрожжей нативная сигнальная последовательность может быть заменена, например, лидерной последовательностью инвертазы дрожжей, лидерной последовательностью α-фактора (включая лидерные последовательности α-фактора Saccharomyces и Kluyveromyces) или лидерной последовательностью кислой фосфатазы, лидерной последовательностью глюкоамилазы С.albicans или сигнальной последовательностью, описанной в WO 90/13646. Для экспрессии в клетках млекопитающих применяют сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например, сигнальную последовательность вируса герпеса простого gD.
ДНК такой области-предшественника встраивают путем лигирования в рамку считывания ДНК, которая кодирует антитело к ErbB2.
(II) Компонент, представляющий собой сайт инициации репликации
Как экспрессионные, так и клонирующие векторы содержат нуклеотидную последовательность, которая инициирует репликацию вектора в одной или нескольких выбранных клетках-хозяевах. Как правило, в клонирующих векторах эта последовательность представляет собой последовательность, которая инициирует репликацию вектора независимо от хромосомной ДНК хозяина и включает сайты инициации репликации или автономно реплицирующиеся последовательности. Такие последовательности хорошо известны для различных бактерий, дрожжей и вирусов. Сайт инициации репликации из плазмиды pBR322 пригоден для большинства грамотрицательных бактерий, сайт инициации плазмиды 2μ пригоден для дрожжей, а различные вирусные сайты инициации (SV40 (обезьяний вирус), вирус полиомы, аденовирус, VSV (вирус везикулярного стоматита) или BPV (вирус папилломы крупного рогатого скота)) пригодны для клонирующих векторов в клетках млекопитающих. Как правило, компонент, представляющий собой сайт инициации репликации, не нужен для экспрессионных векторов млекопитающих (сайт инициации SV40, как правило, применяют только из-за того, что он содержит ранний промотор).
(III) Компонент, представляющий собой селектирующий ген
Экспрессионные и клонирующие векторы могут содержать селектирующий ген, также обозначаемый как селектируемый маркер. Как правило, селектирующие гены кодируют протеины, которые (а) обусловливают устойчивость к антибиотикам или другим токсинам, таким как ампициллин, неомицин, метотрексат или тетрациклин, (б) дополняют дефицит ауксотрофности или (в) пополняют имеющие решающее значение питательные вещества, которые не поступают из комплексных сред, например, ген, кодирующий D-аланинрацемазу для Bacilli.
Один из примеров схемы селекции основан на использовании лекарства для прекращения роста клеток-хозяев. Те клетки, которые успешно трансформированы гетерологичным геном, продуцируют протеин, придающий устойчивость к лекарству, и в результате этого выживают в указанном режиме селекции. Примеры таких агентов для доминантной селекции, основанной на применении лекарств, включают неомицин, микофенольную кислоту и гигромицин.
Примерами других приемлемых селектируемых маркеров для клеток млекопитающих являются маркеры, которые позволяют идентифицировать клетки, обладающие способностью поглощать нуклеиновую кислоту антитела к ErbB2, такие как DHFR, тимидинкиназа, металлотионеин-I и -II, предпочтительно гены металлотионеина приматов, аденозиндеаминаза, орнитиндекарбоксилаза и т.д.
Например, клетки, трансформированные селектирующим геном DHFR, сначала идентифицируют путем культивирования всех трансформантов в культуральной среде, которая содержит метотрексат (Mtx), конкурентный антагонист DHFR. Пригодной клеткой-хозяином при применении DHFR дикого типа является линия клеток яичника китайского хомячка (СНО) с дефицитом DHFR-активности.
В другом варианте клетки-хозяева (прежде всего хозяева дикого типа, которые содержат эндогенный DHFR), трансформированные или совместно трансформированные последовательностями ДНК, кодирующими антитело к ErbB2, протеин DHFR дикого типа и другой селектируемый маркер, такой как аминогликозид-3'-фосфотрансфераза (АРН), могут быть отобраны путем выращивания клеток в среде, содержащей селектирующий агент для селектируемого маркера, такой как аминогликозидовый антибиотик, например канамицин, неомицин или G418 (см. патент США 4965199).
Приемлемый селектирующий ген для применения в дрожжах представляет собой ген trp1, который присутствует в полученной из дрожжей плазмиде YRp7 (Stinchcomb и др., Nature, 282:39 (1979)). Ген trp1 является селектирующим маркером для мутантного штамма дрожжей, лишенного способности расти в среде с добавлением триптофана, например, штамма АТСС №44076 или РЕР4-1 (Jones, Genetics, 85: 12 (1977)). Наличие повреждения trp1 в геноме дрожжевой клетки-хозяина создает благоприятные условия для выявления трансформации путем выращивания в отсутствие триптофана. Аналогично этому штаммы дрожжей с дефицитом Leu-2 (АТСС 20622 или 38626) дополняются с помощью известных плазмид, несущих ген Leu-2.
Кроме того, векторы, полученные из кольцевой плазмиды pKDI размером 1,6 мкм, могут применяться для трансформации дрожжей Kluyveromyces. Другая экспрессионная система для крупномасштабного производства рекомбинантного телячьего химосина описана для К. lactis (Van den Berg, Bio/Technology, 8: 135 (1990)). Также описаны стабильные экспрессионные векторы с множеством копий, предназначенные для обеспечения секреции зрелого рекомбинантного человеческого сывороточного альбумина применяемыми в промышленном масштабе штаммами Kluyveromyces (Fleer и др., Bio/Technology, 9: 968-975 (1991)).
(IV) Компонент, представляющий собой промотор
Экспрессионные и клонирующие векторы обычно содержат промотор, который распознается организмом-хозяином и функционально связан с нуклеиновой кислотой антитела к ErbB2. Промоторы, которые пригодны для применения в прокариотических клетках-хозяевах, включают промотор phoA, промоторные системы β-лактамазы и лактозы, промоторные системы щелочной фосфатазы, триптофана (trp) и гибридные промоторы, такие как промотор tac. Однако могут применяться и другие известные бактериальные промоторы. Промоторы, предназначенные для применения в бактериальных системах, также содержат последовательность Шайна-Дальгарно (S.D.), функционально связанную с ДНК, кодирующей антитело к ErbB2.
Известны также промоторные последовательности, пригодные для эукариот. Практически все гены эукариот имеют область с высоким содержанием AT, расположенную на расстоянии примерно от 25 до 30 пар оснований против хода транскрипции от сайта, в котором инициируется транскрипция. Другая последовательность, которая расположена на расстоянии 70-80 оснований от начала транскрипции против хода транскрипции многих генов, представляет собой CNCAAT-область, в которой N обозначает любой нуклеотид. На 3'-конце большинства эукариотических генов находится последовательность ААТААА, которая может являться сигналом для добавления поли-А-хвоста на 3'-конец кодирующей последовательности. Все эти последовательности можно встраивать в эукариотические экспрессионные векторы.
Примеры промоторных последовательностей, которые могут применяться в дрожжевых клетках-хозяевах, включают промоторы 3-фосфоглицераткиназы или других гликолитических ферментов, таких как энолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа.
Другие промоторы дрожжей, которые представляют собой индуцибельные промоторы, имеющие дополнительное преимущество, связанное с транскрипцией, которая контролируется условиями роста, являются промоторные области алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, отщепляющих ферментов, связанных с метаболизмом азота, металлтионеина, глицеральдегид-3-фосфатдегидрогеназы и ферментов, ответственных за утилизацию мальтозы и галактозы. Приемлемые векторы и промоторы, которые могут применяться при экспрессии в дрожжах, дополнительно описаны в ЕР 73657. С промоторами дрожжей также целесообразно применять энхансеры дрожжей.
Транскрипцию антитела к ErbB2 из векторов в клетках млекопитающих - хозяевах контролирует, например, с помощью промоторов, полученных из геномов вирусов, таких как вирус полиомы, fowlpox-вирус, аденовирус (такой как Аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и наиболее предпочтительно вирус обезьян 40 (SV40), из гетерологичных промоторов млекопитающих, например, промотора актина или промотора иммуноглобулина, из промоторов теплового шока при условии, что такие промоторы совместимы с системами клетки-хозяина.
Ранний и поздний промоторы вируса SV40 удобно получать в виде рестрикционного SV40-фрагмента, который также содержит сайт инициации репликации SV40. Прямой ранний промотор человеческого цитомегаловируса может быть получен в виде HindIII Е-рестрикционного фрагмента. Система для экспрессии ДНК в хозяевах-млекопитающих с помощью вируса папилломы крупного рогатого скота описан в патенте США 4419446. Модификация этой системы описана в патенте США 4601978 (данные, касающиеся экспрессии кДНК человеческого β-интерферона в мышиных клетках под контролем промотора тимидинкиназы вируса герпеса простого см. также у Reyes и др., Nature, 297: 598-601 (1982)). В другом варианте в качестве промотора может применяться длинный концевой повтор вируса саркомы Рауса.
(V) Компонент, представляющий собой энхансер
Транскрипцию ДНК, кодирующей антитело к ErbB2 по изобретению, высшими эукариотами часто повышают путем встраивания в вектор энхансерной последовательности. Известны многие энхансерные последовательности генов млекопитающих (глобин, эластаза, альбумин, α-фетопротеин и инсулин). Однако, как правило, применяют энхансер вируса эукариотической клетки. Примеры включают энхансер SV40, расположенный на удаленной стороне сайта инициации репликации (пары оснований 100-270), энхансер раннего промотора цитомегаловируса, энхансер полиомы, расположенный на удаленной стороне сайта инициации репликации, и энхансеры аденовирусов. Элементы, усиливающие активацию эукариотических промоторов, описаны также у Yaniv, Nature, 297: 17-18 (1982). Энхансер можно встраивать путем сплайсингав вектор в положении 5' или 3' последовательности, кодирующей антитело к ErbB2, но предпочтительно его помещают в 5'-сайт по отношению к промотору.
(VI) Компонент, представляющий собой терминатор транскрипции
Экспрессионные векторы, используемые в эукариотических клетках-хозяевах (клетки дрожжей, грибов, насекомых, растения, животных, людей или ядросодержащие клетки других многоклеточных организмов), должны содержать также последовательности, необходимые для терминации транскрипции и стабилизации мРНК. Такие последовательности обычно получают из 5'- и иногда из 3'-нетранслируемых областей эукариотических или вирусных ДНК или кДНК. Эти области содержат нуклеотидные сегменты, транскрибированные в виде полиаденилированных фрагментов в нетранслируемую область мРНК, кодирующей антитело к ErbB2. Одним из пригодных компонентов, обеспечивающих терминацию транскрипции, является область полиаденилирования бычьего гормона роста (см. WO 94/11026 и приведенное там описание экспрессионного вектора).
(VII) Отбор и трансформация клеток-хозяев
Клетки-хозяева, которые могут использоваться согласно изобретению для клонирования или экспрессии ДНК в векторах, представляют собой описанные выше прокариотические клетки, клетки дрожжей или клетки высших эукариотических организмов. Прокариотические организмы, которые могут применяться для этой цели, включают эубактерии, такие как грамотрицательные или грамположительные организмы, например, представителей Enterobacteriaceae, таких как Esherichia, например Е.coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans, и Shigella, а также Bacilli, таких как В. subtilis и В.lichenformis (например, В.lichenformis 41P, описанный в DD 266710, опубликованной 12 апреля 1989), Pseudomonas, таких как Р.aeruginosa, и Streptomyces. Одним из предпочтительных хозяев для клонирования с использованием E.coli является штамм E.coli W3110 (АТСС 31446), хотя могут применяться также и другие штаммы, такие как E.coli В, E.coli X1776 (АТСС 31573) и E.coli W3110 (АТСС 27325). Эти примеры приведены с целью иллюстрации, но не направлены на ограничение объема изобретения.
Помимо прокариотических организмов в качестве хозяев для клонирования или экспрессии векторов, кодирующих антитело к ErbB2, могут использоваться эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи. Наиболее часто применяемым низшим эукариотическим микроорганизмом-хозяином является Saccharomyces cerevisiae или обычные пекарские дрожжи. Однако широко известно большое количество других родов, видов и штаммов, которые могут применяться в качестве хозяев согласно изобретению, такие как Schizosaccharomyces pombe; Kluyveromyces, например, К.lactis, К.fragilis (ATCC 12424), К.bulgaricus (ATCC 16045), К.wickeramii (ATCC 24178), К.waltii (ATCC 56500), К.drosophilarum (ATCC 36906), К.thermotolerans и К.marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, например Schwanniomyces occidentalis; и нитчатые грибы, такие как, например, Neurospora, Penicillium, Tolypocladium и Aspergillus, такие как A.nidulans и A.niger.
Клетки-хозяева, которые могут использоваться для экспрессии гликозилированного антитела к ErbB2, получают из многоклеточных организмов. Примерами клеток беспозвоночных являются клетки растений и насекомых. Были выявлены многочисленные бакуловирусные штаммы и варианты и соответствующие пригодные для них в качестве хозяев клетки насекомых, таких как Spodoptera frugiperda (гусеница), Aedes aegipti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая муха) и Bombyx mori. Широко известны разнообразные штаммы вирусов, которые могут использоваться для трансфекции, например вариант Autographa NPV L-1 и Bombix mori NPV, и такие вирусы могут применяться согласно изобретению в качестве вирусов, прежде всего для трансфекции клеток Spodoptera frugiperda.
В качестве хозяев можно также использовать культуры клеток растений, таких как хлопчатник, кукуруза, картофель, соя, петунья, томат и табак.
Однако наибольший интерес представляли клетки позвоночных и в настоящее время размножение клеток позвоночных в культуре (культура ткани) стало стандартной процедурой. Примерами пригодных в качестве хозяев линий клеток млекопитающих являются линия клеток почки обезьяны СV1, трансформированная с помощью SV40 (COS-7, ATCC CRL 1651); линия клеток почки эмбриона человека, 293 или клетки линии 293, субклонированные с целью выращивания в суспензионной культуре, Graham и др., J.Gen. ViroL, 36: 59 (1977)); клетки почки детеныша хомячка (ВНК, ATCC CCL 10); клетки яичника китайского хомячка/-DHFR (СНО, Urlaub и др., Ргос. Natl. Acad. Sci. USA, 77: 4216 (1980)); клетки Сертоли мыши (ТМ4, Mather, Biol. Reprod., 23: 243-251 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени бычьей крысы (BRL 3А, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Нер G2, НВ 8065); клетки опухоли молочной железы мыши (ММТ 060562, ATCC CCL51); клетки TRI (Mather и др., Annals N.Y. Acad. Sci., 383: 44-68 (1982)); клетки MRC 5; клетки FS4 и линия клеток гепатомы человека (Нер G2).
Клетки-хозяева трансформируют с помощью описанных выше экспрессионных или клонирующих векторов с целью производства антитела к ErbB2 и культивируют в пригодных питательных средах, соответствующим образом модифицированных для индукции промоторов, отбора трансформантов или амплификации генов, кодирующих требуемые последовательности.
(VIII) Культивирование клеток-хозяев
Клетки-хозяева, используемые для получения антител к ErbB2 по настоящему изобретению, можно культивировать в различных средах. Для культивирования клеток-хозяев можно использовать имеющиеся в продаже среды, такие как среда Хама F10 (фирма Sigma), минимальная поддерживающая среда (MEM, фирма Sigma), RPMI-1640 (фирма Sigma) и модифицированная по способу Дульбекко среда Игла (DMEM, фирма Sigma). Кроме того, в качестве сред для культивирования клеток-хозяев могут использоваться любые из сред, описанных у Ham и др., Meth. Enz., 58: 44 (1979), Barnes и др., Anal. Biochem., 102: 255 (1980), в патентах США 4767704, 4657866, 4927762, 4560655 или 5122468; в WO 90/03430, WO 87/00195; или в US Patent Re. 30985. Любую из этих сред при необходимости можно дополнять гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид и фосфат натрия, кальция, магния), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство GENTAMYCIN®), микроэлементами (т.е. неорганическими соединениями, обычно присутствующими в конечных концентрациях, находящихся в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. Специалисту в данной области должно быть очевидно, что можно также включать любые другие необходимые добавки в соответствующих концентрациях. Условия культивирования, такие как температура, значение рН и т.п., представляют собой условия, которые использовались ранее для клеток-хозяев, отобранных для экспрессии, и они должны быть очевидны специалисту в данной области.
(IX) Очистка антитела к ErbB2
При использовании методов рекомбинации антитело может продуцироваться внутри клетки, в периплазматическом пространстве или непосредственно выделяться в среду. Если антитело продуцируется внутри клетки, то на первой стадии удаляют, например, с помощью центрифугирования или ультрафильтрации, дебрис, состоящий из отдельных частиц, либо из клеток-хозяев, либо из лизированных фрагментов. У Carter и др., Bio/Technology, 10: 163-167 (1992) описан метод выделения антител, секретируемых в периплазматическое пространство Е.coli. В целом метод состоит в следующем. Клеточную пасту подвергают оттаиванию приблизительно в течение 30 мин в присутствии ацетата натрия (рН 3,5), ЭДТК и фенилметилсульфонилфторида (ФМСФ). Клеточный дебрис можно удалять центрифугированием. Если антитело секретируется в среду, супернатанты таких экспрессионных систем, как правило, сначала концентрируют с помощью имеющегося в продаже фильтра для концентрирования протеинов, например, устройства для ультрафильтрации типа Amicon или Millipore Pellicon. На любой из предыдущих стадий для ингибирования протеолиза может быть добавлен ингибитор протеазы, такой как ФМСФ, и для предупреждения увеличения нежелательных примесей могут быть добавлены антибиотики.
Композицию антитела, полученную из клеток, можно очищать, например, хроматографией с использованием гидроксилапатита, гель-электрофорезом, диализом и аффинной хроматографией, при этом предпочтительным методом очистки является аффинная хроматография. Применимость протеина А в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, присутствующего в антителе. Протеин А можно использовать для очистки антител, основанных на человеческих γ1-, γ2- или γ4-тяжелых цепях (Lindmark и др., J.Immunol. Meth., 62: 1-13 (1983)). Протеин G рекомендуется для применения для всех мышиных изотипов и для человеческой γ3-цепи (Guss и др., ЕМВО J., 5: 1567-1575 (1986)). В качестве матрицы, с которой связывается аффинный лиганд, наиболее часто применяют агарозу, однако могут использоваться и другие матрицы. Устойчивые к механическим воздействиям матрицы, такие как стекло или поли(стиролдивинил)бензол с определенным размером пор, позволяют достичь более высоких скоростей потока и более коротких времен процессинга по сравнению с характеристиками, достигаемыми при использовании агарозы. Если антитело содержит СH3-домен, то для очистки можно использовать смолу типа Bakerbond АВХ® (J.T.Baker, Филлипсбург, штат Нью-Джерси). В зависимости от антитела, подлежащего выделению, могут применяться также другие методы очистки протеина, такие как фракционирование на ионообменной колонке, осаждение этанолом, ЖХВР с обращенной фазой, хроматография на силикагеле, хроматография на гепарин-SEPHAROSE™, хроматография на анионо- или катионообменной смоле (например, на колонке с полиаспартамовой кислотой), хроматофокусирование, ДСН-ПААГ и осаждение сульфатом аммония.
После любой из предыдущих стадий очистки смесь, содержащая представляющее интерес антитело, и примеси могут быть подвергнуты хроматографии на основе гидрофобного взаимодействия при низком значении рН с использованием буфера для элюирования с рН приблизительно от 2,5 до 4,5, которую предпочтительно осуществляют при низких концентрациях солей (например, с использованием приблизительно 0-0.25 М солей).
IV. Фармацевтические композиции
Терапевтические композиции антител, применяемых согласно настоящему изобретению, приготавливают с целью их дальнейшего хранения путем смешения антитела, имеющего необходимую степень чистоты, с необязательно используемыми фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences, A.Osol (ред.) (1980)) в форме лиофилизированных композиций или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы должны быть нетоксичными для реципиентов в используемых дозах и концентрациях, они включают буферы, такие как фосфатный, цитратный буферы, а также буферы на основе других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и мета-крезол); полипептиды с низкой молекулярной массой (содержащие менее приблизительно 10 остатков); протеины, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТК; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; содержащие металл комплексы (например, комплексы Zn-протеин); и/или неионогенные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (ПЭГ). Предпочтительные лиофилизированные композиции антител к ErbB2 описаны в WO 97/04801, которая специально включена в настоящее описание в качестве ссылки.
Композиция по изобретению может также включать более одного действующего вещества, если это необходимо для лечения конкретного состояния, предпочтительно она включает действующие вещества, дополнительные активности которых не оказывают неблагоприятного влияния друг на друга. Например, может оказаться необходимым дополнительно включить в одну композицию антитела, которые обладают способностью связывать EGFR, ErbB2 (например, антитело, которое связывается с другим эпитопом ErbB2), ErbB3, ErbB4 или сосудистый эндотелиальный фактор (VEGF). В альтернативном или дополнительном варианте композиция может дополнительно включать химиотерапевтический агент, цитотоксический агент, цитокин, ингибитор роста, антигормональный агент, лекарственное средство, мишенью которого является EGFR, антиангиогенное лекарственное средство и/или кардиозащитное средство. Такие молекулы должны присутствовать в композиции в количествах, которые являются эффективными для конкретной цели.
Действующие вещества могут быть также включены в микрокапсулы, например, с помощью методов коацервации или межфазной полимеризации, например, в гидроксипропилметилцеллюлозные или желатиновые микрокапсулы и поли(метилметакрилатные) микрокапсулы соответственно, в коллоидные системы доставки лекарственного средства (например, в липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методы описаны в Remington's Pharmaceutical Sciences, A.Osol (ред.) (1980).
Могут быть приготовлены композиции с пролонгированным высвобождением. Примеры композиций с пролонгированным высвобождением, которые могут применяться согласно изобретению, включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, при этом матрицы могут представлять собой продукт, имеющий определенную форму, например, пленки или микрокапсулы. Примеры матриц, которые могут использоваться для пролонгированного высвобождения, включают полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт), полилактиды (патент США 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразложимые сополимеры этилена и винилацетата, разложимые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (предназначенные для инъекции микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата леупролида), и поли-D-(-)-3-гидроксимасляную кислоту.
Композиции, предназначенные для применения in vivo, должны быть стерильными. Это легко может быть достигнуто путем фильтрации через стерильные фильтрующие мембраны.
V. Лечение с использованием антител к ErbB2
Подразумевается, что согласно настоящему изобретению антитела к ErbB2 могут применяться для лечения различных болезней или нарушений. Примеры таких состояний или нарушений включают доброкачественные или злокачественные опухоли; лейкозы и злокачественные лимфоидные заболевания; другие нарушения, такие как нейронные, глиальные, астроцитальные, гипоталамические, гландулярные, макрофагиальные, эпителиальные, стромальные, бластоцельные, воспалительные, ангиогенные и иммунологические нарушения.
Как правило, болезнь или нарушение, подлежащее лечению, представляет собой рак. Примеры рака, которые могут лечиться согласно настоящему изобретению, включают (но не ограничиваясь ими) карциному, лимфому, бластому, саркому и лейкоз или злокачественные лимфоидные заболевания. Более конкретными примерами таких видов рака являются плоскоклеточный рак (например, эпителиальный плоскоклеточный рак), рак легкого, включая мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого и плоскоклеточную карциному легкого, рак брюшины, гепатоцеллюлярный рак, рак желудка или пищевода, включая желудочно-кишечный рак, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак ободочной кишки, рак прямой кишки, колоректальный рак, карциному эндометрия или матки, карциному слюнных желез, рак почки, рак предстательной железы, рак вульвы, рак щитовидной железы, карциному печени, анальную карциному, карциному полового члена, а также рак головы и шеи.
Рак обычно характеризуется наличием клеток, экспрессирующих ErbB2, поэтому антитело к ErbB2 по настоящему изобретению обладает способностью связывать раковые клетки. Хотя подлежащие лечению виды рака могут отличаться наличием сверхэкспрессии рецептора ErbB2, но в настоящем изобретении также предложен способ лечения видов рака, которые не относятся к раку, при котором происходит сверхэкспрессия ErbB2. Для выявления экспрессии ErbB2 в раковых клетках могут применяться различные методы диагноза/прогноза. Согласно одному из вариантов осуществления сверхэкспрессию ErbB2 можно анализировать с помощью ИГХ, например, с использованием HERCEPTEST® (фирма DACO). Срезы ткани, полученные при биопсии опухоли и погруженные в парафин, можно подвергать ИГХ-анализу и результаты можно оценивать в баллах с использованием следующих критериев интенсивности окрашивания протеина ErbB2:
Балл 0:
Не выявлено окрашивания или окрашивание мембран наблюдается у менее чем 10% клеток опухоли.
Балл 1+
Выявлено слабое/едва заметное окрашивание мембран у более чем 10% клеток опухоли. Окрашена только часть мембран клеток.
Балл 2+
Выявлено полное окрашивание мембран от слабого до среднего у более чем 10% клеток опухоли.
Балл 3+
Выявлено полное окрашивание мембран от среднего до сильного у более чем 10% клеток опухоли.
Опухоли, для которых получены баллы 0 или 1+, могут быть охарактеризованы как опухоли, в которых не происходит сверхэкспрессия ErbB2, а опухоли, для которых получены баллы 2+ или 3+, могут быть охарактеризованы как опухоли, в которых происходит сверхэкспрессия ErbB2.
В альтернативном или дополнительном варианте для определения уровня сверхэкспрессии (если она присутствует) ErbB2 в опухоли может быть проведен FISH-анализ, например, с помощью INFORM™ (поступает в продажу от фирмы Ventana, штат Аризона) или PATHVISION™ (фирма Vysis, штат Иллинойс), с использованием образцов ткани опухоли, зафиксированных формалином и погруженных в парафин.
В одном из вариантов рак может представлять собой любой из видов рака, при котором раковые клетки экспрессируют (и могут сверхэкспрессировать) EGFR. Примеры видов рака, при которых может происходить экспрессия/сверхэкспрессия EGFR, включают плоскоклеточный рак (например, эпителиальный плоскоклеточный рак), рак легкого, включая мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого и плоскоклеточную карциному легкого, рак брюшины, гепатоцеллюлярный рак, рак желудка или пищевода, включая желудочно-кишечный рак, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак ободочной кишки, рак прямой кишки, колоректальный рак, карциному эндометрия или матки, карциному слюнных желез, рак почки, рак предстательной железы, рак вульвы, рак щитовидной железы, карциному печени, анальную карциному, карциному полового члена, а также рак головы и шеи.
Рак, подлежащий лечению, может представлять собой один из видов рака, отличающегося повышенной активацией рецептора ErbB, например, EGRF. Такая повышенная активация может обусловливаться сверхэкспрессией или повышенным производством рецептора ErbB или лиганда EGFR. Согласно одному из вариантов осуществления изобретения для определения того, характеризуется ли рак у пациента наличием повышенной активации рецептора ErbB, следует осуществлять диагностический или прогностический анализ. Например, можно выявлять амплификацию гена ErbB и/или сверхэкспрессию рецептора ErbB в раковых клетках. Для выявления такой амплификации/сверхэкспрессии могут применяться различные методы анализа, известные в данной области, которые включают описанные выше ИГХ-, FISH-анализы и анализ с использованием антигена. В альтернативном или дополнительном варианте можно определять с помощью известных методов уровни лиганда ErbB, такого как TGF-α, в опухоли или связанные с опухолью. С помощью таких анализов можно обнаружить в тестируемом образце протеин и/или кодирующую его нуклеиновую кислоту. Согласно одному из вариантов осуществления уровни лиганда ErbB в опухоли можно определять с помощью иммуногистохимического анализа (ИГХ) (см., например, Scher и др., Clin, Cancer Research, 1: 545-550 (1995)). В альтернативном или дополнительном варианте можно оценивать в тестируемом образце уровни нуклеиновой кислоты, кодирующей лиганд ErbB; например, с помощью FISH-анализа, Саузерн-блоттинга или метода ПЦР.
Кроме того, сверхэкспрессию или амплификацию рецептора ErbB или лиганда ErbB можно выявлять с помощью диагностического анализа in vivo, например, путем введения молекулы (такой как антитело), которая обладает способностью связываться с подлежащей выявлению молекулой и несет обнаруживаемую метку (например, радиоактивный изотоп), и последующего сканирования пациента с целью определения местонахождения метки.
Если подлежащий лечению рак представляет собой независящий от гормонов рак, то экспрессию гормона (например, андрогена) и/или родственного ему рецептора в опухоли можно оценить с помощью любого пригодного метода, например, из числа описанных выше методов. В альтернативном или дополнительном варианте пациент может быть диагностирован как пациент, имеющий независящий от гормонов рак, на основе того, что он более не поддается терапии с использованием антиандрогенов.
Согласно одому из вариантов пациенту вводят иммуноконъюгат, представляющий собой антитело к ErbB2, конъюгированное с цитотоксическим агентом. Предпочтительно иммуноконъюгат и/или протеин ErbB2, с которым он связывается, интернализируется(ются) клеткой, что приводит к повышенной терапевтической эффективности иммуноконъюгата в отношении уничтожения раковой клетки, с которой он связывается. В предпочтительном варианте цитотоксический агент направленно действует на нуклеиновую кислоту или взаимодействует с ней в раковой клетке. Примеры таких цитотоксических агентов включают майтанзиноиды, калихеамины, рибонуклеазы и ДНК-эндонуклеазы.
Антитела к ErbB2 или иммуноконъюгаты вводят больному человеку с помощью известных методов, таких как внутривенное введение, например, в виде болюса, или путем непрерывной инфузии в течение определенного промежутка времени, путем внутримышечного, внутрибрюшинного, интрацереброспинального, подкожного, интраартикулярного, внутрисуставного, внутриоболочечного, перорального, местного введения или путем ингаляции. Предпочтительным является внутривенное или подкожное введение антитела.
В сочетании с введением антитела к ErbB2 могут применяться другие режимы терапии. Совместное введение включает одновременное введение с использованием раздельных композиций или единой фармацевтической композиции и последовательное введение в произвольном порядке, при этом предпочтительно должен существовать промежуток времени, в течение которого оба (или все) действующие вещества одновременно проявляют свои биологические активности.
Согласно одному из предпочтительных вариантов осуществления пациента лечат с помощью двух различных антител к ErbB2. Например, пациента можно лечить с помощью первого антитела к ErbB2, которое блокирует активацию лигандом рецептора ErbB или антителом, обладающим биологическими характеристиками моноклонального антитела 2С4, а также с помощью второго антитела к ErbB2, представляющего собой ингибитор роста (например, HERCEPTIN®) или антитело к ErbB2, которое индуцирует апоптоз клеток, сверхэкспрессирующих ErbB2 (например, 7С2, 7F3 или их гуманизированные варианты). Предпочтительно такая совместная терапия приводит к синергетическому терапевтическому действию. Можно, например, лечить пациента с помощью HERCEPTIN® и после этого лечить с помощью rhuMab 2C4, например, если пациент не реагирует на терапию с использованием HERCEPTIN®. Согласно другому варианту пациента можно сначала лечить с помощью rhuMab 2C4, а затем с помощью HERCEPTIN®. Согласно еще одному варианту осуществления пациента можно лечить одновременно с помощью rhuMab 2C4 и HERCEPTIN®.
Может оказаться целесообразным объединять введение антитела или антител к ErbB2 с введением антитела к другому связанному с опухолью антигену. В этом случае другое антитело может, например, связывать EGFR, ErbB3, ErbB4 или сосудистым эндотелиальным фактором роста (VEGF).
В одном из вариантов осуществления лечение согласно настоящему изобретению предусматривает совместное введение антитела (или антител) к ErbB2 и одного или нескольких химиотерапевтических агентов или ингибиторов роста, включая одновременное введение смеси различных химиотерапевтических агентов. Предпочтительные химиотерапевтические агенты включают таксаны (такие как паклитаксел и доцетаксел) и/или антрациклиновые антибиотики. Для таких химиотерапевтических агентов могут применяться композиции и режим доз, указанные в инструкциях производителя, или они могут быть определены эмпирически специалистом в данной оласти. Композиции и режимы доз для такой химиотерапии описаны также в Chemotherapy Service, M.C. Perry (ред.), Williams & Williams, Baltimore, MD (1992).
Антитело можно объединять с антигормональным соединением; например, антиэстрогенным соединением, таким как тамоксифен; антипрогестероном, таким как онапристон (см. ЕР 616812); или антиандрогеном, таким как флутамид, в дозах, известных для таких веществ. Если рак, подлежащий лечению, представляет собой независящий от гормонов рак, то пациент может сначала быть подвергнут антигормональной терапии, а затем, после того как рак станет независимым от гормонов, пациенту может вводиться антитело к ErbB2 (и необязательно другие описанные выше агенты).
Иногда может оказаться целесообразным также одновременно вводить пациенту кардиозащитное средство (для предупреждения или уменьшения дисфункции миокарда, связанной с терапией) или один или несколько цитокинов. Можно также одновременно вводить лекарственное средство, мишенью которого является EGFR, или антиангиогенное лекарственное средство. В дополнение к указанным выше терапевтическим режимам пациент может быть подвергнут хирургической операции по поводу удаления раковых клеток и/или лучевой терапии.
Антитела к ErbB2 согласно настоящему изобретению можно также объединять с лекарством, мишенью которого является EGFR, таким как одно из лекарств, перечисленных в разделе "Определения", что приводит к дополнительному и возможно синергетическому терапевтическому действию.
Приемлемые дозы любого из перечисленных выше совместно вводимых агентов представляют собой дозы, в которых они применяются в настоящее время, и эти дозы могут быть уменьшены благодаря объединенному действию (синергизм) агента и антитела к ErbB2.
Необходимая для предупреждения или лечения болезни доза антитела должна зависеть от типа указанной выше болезни, подлежащей лечению, серьезности и течения болезни, от того, для какой цели, профилактической или терапевтической, вводится антитело, от предыдущей терапии, анамнеза пацента и его реакции на антитело, а также от решения штатного врача больницы. Антитело можно вводить пациенту в виде однократной дозы или с помощью серии обработок. В зависимости от типа и серьезности болезни в качестве начальной пробной дозы пациенту вводят от приблизительно 1 мкг/кг до 15 мг/кг (например, 0,1-20 мг/кг) антитела, например, в виде однократной дозы или в виде нескольких раздельных доз, или путем непрерывной инфузии. Типичная суточная доза может варьироваться от приблизительно 1 мкг/кг до 100 мг/кг в зависимости от указанных выше факторов. В случае повторных введений в течение нескольких дней или более в зависимости от состояния лечение продолжают до достижения необходимого уровня супрессии симптомов болезни. Предпочтительные дозы антитела составляют от приблизительно 0,05 до приблизительно 10 мг/кг. Так, пациенту можно вводить одну или несколько доз, каждая из которых составляет приблизительно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любую их комбинацию). Такие дозы можно вводить периодически, например, каждую неделю или каждые три недели (например, таким образом, чтобы пациент получил от приблизительно двух до приблизительно двадцати, например приблизительно шесть доз антитела к ErbB2). Сначала можно вводить начальную более высокую ударную дозу, а затем одну или несколько более низких доз. Примером режима доз является введение начальной ударной дозы, составляющей приблизительно 4 мг/кг, а затем еженедельно поддерживающей дозы, составляющей приблизительно 2 мг/кг антитела к ErbB2. Однако может применяться и другой режим доз. Успех применения такой терапии легко может быть оценен с помощью обычных методов и анализов.
Помимо введения пациенту протеина антитела в настоящем описании предложено введение антитела с помощью генной терапии. Такое введение нуклеиновой кислоты, кодирующей антитело, подпадает под объем понятия "введение терапевтически эффективного количества антитела" (см., например, WO 96/07321, опубликованную 14 марта 1996, касающуюся применения генной терапии для получения внутриклеточных антител).
Существует два основных подхода для введения нуклеиновой кислоты (необязательно содержащейся в векторе) в клетки пациента, а именно in vivo и ex vivo. Для доставки in vivo нуклеиновую кислоту инъецируют непосредственно пациенту, как правило, в область, где необходимо присутствие антитела. При лечении ех vivo выделяют клетки из организма пациента, встраивают в эти выделенные клетки нуклеиновую кислоту и модифицированные клетки вводят пациенту либо непосредственно, либо, например, в форме, когда они инкапсулированы в пористые мембраны, которые имплантируют пациенту (см, например, патенты США 4892538 и 5283187). Существуют различные методы, которые могут использоваться для введения нуклеиновых кислот в живые клетки. Методы могут варьироваться в зависимости от того, переносят нуклеиновую кислоту in vitro в культивируемые клетки или in vivo в клетки хозяина, подлежащего лечению. Методы, которые можно применять для переноса нуклеиновой кислоты в клетки млекопитающего in vitro, включают использование липосом, электропорацию, микроинъекцию, слияние клеток, использование ДЭАЭ ((диэтиламино)этилцеллюлоза)-декстрана, метод преципитации с помощью фосфата кальция и т.д. Как правило, вектор, используемый для доставки гена ех vivo, представляет собой ретровирус.
Предпочтительные в настоящее время методы переноса нуклеиновой кислоты in vivo включают трансфекцию вирусными векторами (такими как аденовирус, вирус герпеса простого I или аденоассоциированный вирус) и системы на основе липидов (примерами липидов, которые могут применяться для опосредуемого липидами переноса гена, являются DOTMA, DOPE DC-Chol). В некоторых ситуациях целесообразно источник нуклеиновой кислоты связывать агентом, обеспечивающим направленный перенос в клетки-мишени, таким как антитело, специфическое в отношении протеина поверхности клеточной мембраны или клетки-мишени, лиганд рецептора на клетке-мишени и т.д. Если применяются липосомы, то для направленного переноса и/или облегчения поглощения можно использовать протеины, которые связываются с протеином поверхности клеточной мембраны, опосредующим эндоцитоз, например, капсидные протеины или их фрагменты, которые являются тропическими для конкретного типа клеток, антитела к протеинам, которые подвергаются интернализации при циклизации, и протеины, которые обеспечивают направленный перенос внутрь клетки и увеличивают время полужизни внутри клетки. Метод опосредуемого рецептором эндоцитоза описан, например, у Wu и др., J.Biol. Chem., 262: 4429-4432 (1987); и у Wagner и др., Proc. Natl. Acad. Sci. USA, 87: 3410-3414 (1990). Обзор известных в настоящее время методов маркирования генов и протоколов генной терапии приведен у Anderson и др., Science, 256: 808-813 (1992) (см. также WO 93/2567 и процитированные там ссылки).
VI. Формы для применения
Еще одним объектом изобретения является форма для применения, содержащая продукты, которые можно использовать для лечения описанных выше нарушений. Форма для применения включает контейнер и этикетку или вкладыш для упаковки, которые находятся на контейнере или прилагаются к нему. Контейнеры, которые можно использовать для этой цели, включают, например, бутылки, пузырьки, шприцы и т.д. Контейнеры могут изготавливаться из различных материалов, таких как стекло или пластик. Контейнер содержит композицию, обладающую эффективностью в отношении лечения определенного состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой баллон или пузырек для раствора, предназначенного для внутривенного введения, снабженный пробкой, которая может быть проколота иглой для подкожной инъекции). По крайней мере, одно действующее вещество в композиции представляет собой антитело к ErbB2. На этикетке или вкладыше для упаковки указывается, что композиция может применяться для лечения определенного состояния, такого как рак. В одном из вариантов осуществления на этикетке или вкладыше указывается, что композиция, содержащая антитело, которое обладает способностью связывать ErbB2, может применяться для лечения рака, при котором происходит экспрессия рецептора ErbB, выбранного из группы, включающей рецептор эпидермального фактора роста (EGFR), ErbB3 и ErbB4, предпочтительно EGFR. Кроме того, на этикетке или вкладыше для упаковки может быть указано, что подлежащий лечению пациент является пациентом, страдающим раком, который характеризуется повышенной активацией рецептора ErbB, выбранного из группы, включающей рецептор эпидермального фактора роста EGFR, ErbB3 или ErbB4. Например, рак может представлять собой один из видов рака, при котором происходит сверхэкспрессия одного из этих рецепторов и/или сверхэкспрессия лиганда ErbB (такого как TGF-α). На этикетке или вкладыше для упаковки может быть указано, что композицию можно применять для лечения рака, который не характеризуется сверхэкспрессией рецептора ErbB2. Например, в то время как на существующем в настоящее время вкладыше для упаковки для HERCEPTIN® указано, что антитело используется, для лечения пациентов, страдающих метастатическим раком молочной железы, при котором опухолевые клетки сверхэкспрессируют протеин ErbB2, на вкладыше для упаковки по настоящему изобретению может быть указано, что антитело или композиция используется для лечения рака независимо от уровня сверхэкспрессии ErbB2. В других вариантах осуществления на вкладыше для упаковки может быть указано, что антитело или композиция может применяться для лечения рака молочной железы (например, метастатического рака молочной железы); независящего от гормонов рака; рака предстательной железы (например, независящего от андрогена рака предстательной железы); рака легкого (например, немелкоклеточного рака легкого); рака ободочной кишки, прямой кишки или колоректального рака; или любой из других болезней или нарушений, перечисленных в настоящем описании. Кроме того, форма для применения может представлять собой (а) первый контейнер с содержащейся в нем композицией, которая включает первое антитело, которое обладает способностью связывать ErbB2 и ингибировать рост раковых клеток, сверхэкспрессирующих ErbB2, и (б) второй контейнер с содержащейся в нем композицией, которая включает второе антитело, которое обладает способностью связывать ErbB2 и блокировать активацию лигандом рецептора ErbB. Согласно этому варианту осуществления форма для применения может дополнительно включать вкладыш для упаковки, на котором указано, что композиции первого и второго антител могут применяться для лечения рака. Кроме того, вкладыш для упаковки может содержать инструкцию для потребителя композиции (которая включает антитело, обладающее способностью связывать ErbB2 и блокировать активацию лигандом рецептора ErbB) по применению совместной терапии с использованием антитела и любой из описанных в предыдущем разделе вспомогательных терапий (например, с помощью химиотерапевтического агента, лекарственного средства, мишенью которого является EGFR, антиангиогенного агента, антигормонального соединения, кардиозащитного средства и/или цитокина). В альтернативном или дополнительном варианте форма для применения может дополнительно включать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (БСВИ), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Кроме того, она может включать другие продукты, необходимые с коммерческой точки зрения и с точки зрения потребителя, в частности другие буферы, разбавители, фильтры, иглы и шприцы.
VII. Нетерапевтические применения антитела к ErbB2
Антитела (например, гуманизированные антитела к ErbB2) по изобретению могут применяться в других нетерапевтических целях.
Например, антитела могут использоваться в качестве агентов для аффинной очистки. Согласно этому методу антитела иммобилизируют на твердой фазе, такой как смола Sephadex или фильтровальная бумага, с помощью методов, хорошо известных в данной области. Иммобилизированное антитело приводят в контакт с образцом, содержащим протеин ErbB2 (или его фрагмент), который требуется очистить, после чего подложку промывают пригодным растворителем, позволяющим удалить практически весь продукт из образца за исключением протеина ErbB2, связанного с иммобилизованным антителом. Наконец, подложку промывают другим пригодным растворителем, таким как глициновый буфер, рН 5,0, который позволяет отделить протеин ErbB2 от антитела.
Антитела к ErbB2 можно также применять в диагностических анализах протеина ErbB2, например, для обнаружения его экспрессии в определенных клетках, тканях или сыворотке.
Для применений в области диагностики антитело как правило метят с помощью обнаруживаемого фрагмента. Можно применять многочисленные метки, которые в целом могут быть подразделены на следующие категории:
(а) Радиоизотопы, такие как 35S, 14С, 125I, 3H и 131I. Мечение антитела с использованием радиоизотопа можно осуществлять с помощью методов, описанных, например, в Current Protocols in Immunology, тома 1 и 2, Coligen и др. (ред-ры) Wiley-Interscience, New York, изд. (1991), а радиоактивность можно измерять с помощью сцинтилляционного счетчика.
(б) Флуоресцентные метки, такие как хелаты редкоземельных металлов (хелаты европия) или флуоресцеин и его производные, родамин и его производные, дансил, лиссамин, фикоэритрин и техасский красный. Флуорецентные метки можно конъюгировать с антителом, например, с помощью методов, описанных в Current Protocols in Immunology (см. выше). Флуоресценцию можно количественно оценивать с помощью флуориметра.
(в) Могут применяться различные метки на основе комбинаций фермент-субстрат, в патенте США 4275149 приведен обзор некоторых из них. Фермент обычно катализирует химическое изменение хромогенного субстрата, которое можно оценить с помощью различных методов. Например, фермент может катализировать изменение окраски субстрата, которое можно измерить спектрофотометрическим путем. В альтернативном варианте фермент может изменять флуоресценцию или хемилюминисценцию субстрата. Методы количественной оценки изменения флуоресценции описаны выше. Хемилюминисцентный субстрат подвергается возбуждению электронов в результате химической реакции и затем может испускать свет, который можно измерять (например, с использованием хемиолюминометра), или становится донором энергии для флуоресцентного акцептора. Примерами ферментативных меток являются люциферазы (например, люцифераза светляков и бактериальная люцифераза; патент США 4737456), люциферин, 2,3-дигидрофталазиндионы, малатдегидрогеназа, уреаза, пероксидаза, такая как пероксидаза из хрена (HRPO), щелочная фосфатаза, (3-галактозидаза, глюкоамилаза, лизозим, сахаридооксидазы (например, глюкозооксидаза, галактозооксидаза и глюкозо-6-фосфат-дегидрогеназа), гетероциклические оксидазы (такие как уриказа и ксантиноксидаза), лактопероксидаза, микропероксидаза и т.п. Методы конъюгации ферментов с антителами описаны у O'Sullivan, Methods for the Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay, в Methods in Enzym. (ред-ры J. Langone и Н. Van Vunakis), Academic Press, New York, 75: 147-166 (1982). Примерами комбинаций фермент-субстрат являются, например:
(I) пероксидаза из хрена (HRPO) и кислая пероксидаза в качестве субстрата, при этом кислая пероксидаза обладает способностью окислять предшественник красителя (например, ортофенилендиамин (ОФД) или гидрохлорид 3,3'5,5'-тетраметилбензидина (ТМБ));
(II) щелочная фосфатаза (ЩФ) и пара-нитрофенилфосфат в качестве хромогенного субстрата; и
(III) β-D-гапактозидаза (β-D-Gal) и хромогенный субстрат (например, пара-нитрофенил-β-D-галактозидаза) или флуорогенный субстрат 4-метилумбеллиферил-β-D-галактозидаза.
Специалист в данной области может использовать многочисленные другие комбинации фермент-субстрат. Общий обзор таких комбинаций приведен в патентах США 4275149 и 4318980.
Иногда метку конъюгируют с антителом косвенным образом. Специалисту в данной области должны быть очевидны различные методы осуществления такой коъюгации. Например, антитело можно конъюгировать с биотином и любую из относящихся к указанным выше трем широким категориям меток можно конъюгировать с авидином или наоборот. Биотин обладает способностью избирательно связывать авидин и поэтому можно осуществлять конъюгацию метки с антителом таким косвенным путем. В альтернативном варианте для осуществления конъюгации метки с антителом косвенным путем антитело конъюгируют с малым гаптеном (например, дигоксином) и одну из относящихся к перечисленным выше трем различным типам меток конъюгируют с антителом к гаптену (например, с антителом к дигоксину). Таким путем может быть осуществлена косвенная конъюгация метки с антителом.
В другом варианте осуществления не требуется мечения антитела к ErbB2, а его присутствие можно обнаруживать с помощью меченого антитела, которое обладает способностью связывать антитело к ErbB2.
Антитела по настоящему изобретению могут применяться в любом из известных методов анализа, таких как конкурентный анализ связывания, прямой и косвенный сэндвич-анализ и анализ с использованием иммунопреципитации (см. Zola, Monoclonal Antibodies: A manual of Techniques, стр.147-158 (CRC Press, Inc., 1987).
Для иммуногистохимического анализа можно использовать образец опухоли, который является свежим или замороженным или который погружен в парафин и зафиксирован с помощью консерванта, например, такого как формалин.
Антитела также можно использовать для диагностических анализов in vivo. В целом метод состоит в следующем. Антитело метят с помощью радионуклеида (такого как 111In, 99Tc, 14С, 131I, 125I, 3H, 32Р или 35S), так что месторасположение опухоли может быть установлено с помощью иммуносцинтиографии. Для удобства антитела по настоящему изобретению могут поставляться в наборе, т.е. в упаковке, содержащей реагенты в заранее определенном количестве и инструкцию для проведения диагностического анализа. Если антитело метят с помощью фермента, то набор должен включать субстраты и кофакторы, наобходимые для фермента (например, предшественник субстрата, который превращается в обнаруживаемый хромофор или флуорофор). Кроме того, могут быть включены другие добавки, такие как стабилизаторы, буферы (например, блокирующий буфер или буфер для лизиса) и т.п. Относительные количества различных реагентов могут варьироваться в широких пределах для обеспечения таких концентраций реагентов в растворе, которые позволяют добиться оптимальной чувствительности анализа. В частности, реагенты могут поставляться в виде сухих порошков, как правило, лиофилизированных, и эксципиентов, которые после растворения должны обеспечить получение раствора реагентов, имеющего соответствующую концентрацию.
VIII. Депонирование
В Американской коллекции типовых культур, 10801 University Boulevard, Манассас, штат Вирджиния, VA 20110-2209, США (АТСС), были депонированы следующие линии клеток гибридомы:
Ниже изобретение проиллюстрировано с помощью примеров, не ограничивающих его объем. Содержание всех процитированных в описании документов, во всей полноте включено в настоящее описание в качестве ссылки.
Пример 1
Получение и определение характеристик моноклонального антитела 2С4
Мышиные моноклональные антитела 2С4, 7F3 и 4D5, которые обладают способностью специфически связывать внеклеточный домен ErbB2, получали согласно методу, описанному у Fendly и др., Cancer Research, 50: 1550-1558 (1990). В целом метод состоит в следующем. Клетки линии NIH 3T3/HER2-3400 (экспрессирующие приблизительно 1×105 молекул ErbB2/клетку), которые получали согласно методу, описанному у Hudziak и др., Proc. Natl. Acad. Sci. (США), собирали с использованием забуференного фосфатом физиологического раствора (ЗФР), содержащего 25 мМ ЭДТК, и использовали для иммунизации мышей линии BALB/c. Мышам вводили путем i.p. инъекции 107 клеток в 0,5 мл ЗФР через 0, 2, 5 и 7 недель. Мышам с антисывороткой, которая обладала способностью иммуноосаждать меченный с помощью Р32 ErbB2, вводили через 9 и 13 недель путем i.p. инъекции экстракт мембран ErbB2, очищенный с помощью агглютинина из проростков пшеницы-сефарозы (WGA). После этого путем i.v. инъекции вводили 0,1 мл препарата ErbB2 и спленоциты сливали с линией клеток мышиной миеломы Х63-Ag8.653.
Супернатанты гибридомы подвергали скринингу в отношении способности связываться с ErbB2 с помощью ELISA и радиоиммунопреципитации.
Эпитопы ErbB2, связанные с моноклональными антителами 4D5, 7F3 и 2С4, выявляли с помощью анализа конкурентного связывания (Fendy и др., Cancer Research, 50: 1550-1558 (1990)). Исследования перекрестной блокады проводили с использованием антител путем непосредственного измерения флуоресценции от интактных клеток с помощью устройства PANDEX™ Screen Machine, позволяющего проводить количественную оценку флуоресценции. Каждое моноклональное антитело конъюгировали с изотиоцианатом флуоресцеина (ФИТЦ) с помощью известных методов (Wofsy и др., Selected Methods in Cellular Immunology, стр.287, Mishel и Schiigi (ред-ры), San Francisco; W.J. Freeman Co. (1980)). Конфлуэнтные монослои клеток линии NIH 3T3/HER2-3400 обрабатывали трипсином, однократно промывали и ресуспендировали в концентрации 1,75×106 клеток/мл в холодном ЗФР, содержащем 0,5% бычьего сывороточного альбумина (БСА) и 0,1% NaN3. Для уменьшения закупоривания планшетных мембран PANDEX™ добавляли частицы латекса (фирма IDC, Портланд, штат Орегон) в конечной концентрации 1%. В лунки планшета PANDEX™ добавляли 20 мкл суспензии клеток и 20 мкл очищенных моноклональных антител (от 100 до 0,1 мкг/мл) и инкубировали в течение 30 мин на льду. В каждую лунку добавляли предварительно определенные разведения моноклональных антител, меченных с помощью ФИТЦ, инкубировали в течение 30 мин, промывали и количественно оценивали флуоресценцию с помощью PANDEX™. Предполагалось, что моноклональные тела имеют эпитоп, если каждое из них блокировало связывание другого на 50% или более по сравнению с контролем без использования моноклонального антитела. В этом эксперименте эпитопы для моноклональных антител 4D5, 7F3 и 2С4 были обозначены как I, G/F и F соответственно.
Характеристики моноклональных антител 2С4, 7F3 и 4D5, касающиеся ингибирования роста, оценивали с использованием линии клеток опухоли молочной железы SK-BR-3 (см. Hudziak и др., Molec. Cell. Biol., 9(3): 1165-1172 (1989)). В целом метод состоит в следующем. Клетки линии SK-BR-3 обрабатывали 0,25 об.% трипсином и суспендировали в полной среде с плотностью 4×105 клеток/мл. Аликвоты по 100 мкл (4×104 клеток) вносили в лунки 96-луночных планшетов для микроразведений, клеткам давали возможность прилипнуть и затем добавляли 100 мкл либо только среды, либо среды, содержащей моноклональное антитело (конечная концентрация 5 мкг/мл). Через 72 ч планшеты дважды промывали ЗФР (рН 7,5), окрашивали кристаллическим фиолетовым красителем (0,5%-ным в метаноле) и анализировали в отношении относительной пролиферации клеток согласно методу, описанному у Sugarman и др., Science, 230: 943-945 (1985). Моноклональные антитела 2С4 и 7F3 ингибировали относительную пролиферацию клеток линии SK-BR-3 приблизительно на 20% и приблизительно на 38% соответственно, в то время как ингибирование с использованием моноклонального антитела 4D4 составляло приблизительно 56%.
Моноклональные антитела 2С4, 4D5 и 7F3 оценивали в отношении их способности ингибировать стимулируемое HRG фосфорилирование тирозина протеинов с молекулярными массами Mr приблизительно 180000 из лизатов целых клеток линии MCF7 (Lewis и др., Cancer Research, 56: 1457-1465 (1996)). Известно, что клетки линии MCF7 экспрессируют все известные рецепторы ErbB, но с относительно низкими уровнями. Поскольку ErbB2, ErbB3 и ErbB4 имеют практически одинаковые размеры молекул, то при оценке лизатов целых клеток с помощью Вестерн-блоттинга невозможно определить, в каком протеине происходит фосфорилирование тирозина.
Однако эти клетки идеально подходят для анализов стимулируемого HRG фосфорилирования тирозина, поскольку в отсутствие выделяемого экзогенным путем HRG они характеризуются малыми или необнаруживаемыми уровнями фосфорилирования тирозина протеинов с Мr приблизительно 180000.
Клетки линии MCF7 высевали на 24-луночные планшеты, в каждую лунку добавляли моноклональные антитела к ErbB2 и проводили инкубацию в течение 30 мин при комнатной температуре; после чего в каждую лунку добавляли rHRGβ1177-244 в конечной концентрации 0,2 нМ и инкубацию продолжали в течение 8 мин. Среду из каждой лунки осторожно удаляли путем аспирации и реакции прекращали путем добавления 100 мкл ДСН-буфера для образцов (5%-ный ДСН, 25 мМ ДТТ и 25 мМ Трис-HCl, рН 6,8). Каждый образец (25 мкл) подвергали электрофорезу в 4-12%-ном градиентном геле (фирма Novex) и затем электрофоретическим путем переносили на мембрану из поливинилидендифторида. Получали иммунноблотты антитела к фосфотирозину (4G10, фирма UBI, 1 мкг/мл) и интенсивность основной активной полосы, соответствующей Mr ˜ 180000, количественно оценивали с помощью денситометрии в отраженном свете согласно описанному в литературе методу (Holmes и др., Science, 256: 1205-1210 (1992); Sliwkowski и др., J.Biol. Chem. 269: 14661-14665 (1994)).
Моноклональные антитела 2С4, 7F3 и 4D5 существенно ингибировали образование индуцируемого HRG сигнала фосфорилирования тирозина для протеинов с Mr=180000. В отсутствие HRG ни одно из этих антител не обладало способностью стимулировать фосфорилирование тирозина протеинов с молекулярной массой Mr ˜ 180000. Эти антитела также не вступали в перекрестную реакцию с EGFR (Fendy и др., Cancer Research, 50: 1550-1558 (1990)), ErbB3 и ErbB4. Антитела 2С4 и 7F3 существенно (до уровня, составляющего <25% по сравнению с контролем) ингибировали стимуляцию HRG фосфорилирования тирозина р180. Моноклональное антитело 4D5 обладало способностью блокировать стимуляцию HRG фосфорилирования тирозина на ˜50%. На фиг.2А приведены графики зависимости реакции от дозы, полученные в опытах по исследованию способности 2С4 и 7F3 ингибировать стимуляцию HRG фосфорилирования тирозина р180, проведенных с помощью денситометрии в отраженном свете. На основе анализа этих зависимостей ингибирования с использованием аппроксимации экспериментальных данных 4-параметрической кривой, получали значения IC50, составляющие 2,8±0,7 нМ и 29,0±4,1 нМ для 2С4 и 7F3 соответственно.
Опыты по изучению ингибирования связывания HRG с клетками линии опухоли молочной железы MCF7 с помощью антител к ErbB2 осуществляли с использованием монослойных культур на льду в формате 24-луночного планшета (Lewis и др., Cancer Research 56: 1457-1465 (1996)). Моноклональные антитела к ErbB2 добавляли в каждую лунку и инкубировали в течение 30 мин. Добавляли I125-rHRGβ1177-224 (25 пмолей) и инкубацию продолжали в течение 4-16 ч. На фиг.2Б приведены графики зависимости реакции от дозы, полученные в опытах по изучению ингибирования с помощью 2С4 или 7F3 связывания HRG с клетками линии MCF7. Различные концентрации 2С4 или 7F3 инкубировали с клетками линии MCF7 в присутствии I125-rHRGβ1, полученные при этом кривые ингибирования приведены на фиг.2Б. На основе анализа этих данных получали значения IC50, составляющие 2,4±0,3 нМ и 19,0±7,3 нМ для 2С4 и 7F3 соответственно. Величины максимального ингибирования, составляющие для 2С4 и 7F3 ˜74%, находились в соответствии с данными опытов по оценке фосфорилирования тирозина.
Для установления того, носит ли наблюдаемое в отношении клеток линии MCF7 влияние антител к ErbB2 общий характер, различные линии опухолевых клеток человека инкубировали с 2С4 или 7F3 и определяли степень специфического связывания I125-rHRGβ1 (Lewis и др., Cancer Research 56: 1457-1465 (1996)). Результаты этих опытов представлены на фиг.3. Как 2С4, так и 7F3 существенно ингибировали связывание I125-rHRGβ1 для всех линий клеток за исключением линии клеток карциномы молочной железы MDA-MB-468, которая, как известно из опубликованных данных, экспрессирует небольшие количества ErbB2 или совсем не экспрессирует его. Из опубликованных данных известно, что остальные линии клеток экспрессируют ErbB2, при этом уровень экспрессии ErbB2 варьируется в широких пределах для указанных линий клеток. Действительно, уровень экспрессии ErbB2 в изученных линиях клеток различается по величине более, чем на 2 порядка. Например, линии клеток ВТ-20, MCF7 и Caov3 экспрессируют ˜104 рецепторов ErbB2/клетку, в то время как линии клеток ВТ-474 и SK-BR-3 экспрессируют ˜106 ErbB2-рецепторов/клетку. С учетом широкого диапазона уровней экспрессии ErbB2 в этих клетках и приведенных выше данных был сделан вывод о том, что само по себе взаимодействие между ErbB2 и ErbB3 или ErbB4 представляет собой высокоаффинное взаимодействие, которое происходит на поверхности плазматической мембраны.
Оценивали ингибирующие воздействия моноклональных антител 2С4 и 4D5 на рост клеток линий MDA-MB-175 и SK-BR-3 в присутствии экзогенного rHRGβ1 (Schaefer и др., Oncogene, 15: 1385-1394 (1997)) или без него. Установлено, что уровни ErbB2 в клетках линии MDA-MB-175 в 4-6 раз выше, чем уровень, обнаруженный в нормальных эпителиальных клетках молочной железы, и что в клетках линии MDA-MB-175 происходит конститутивное фосфорилирование тирозина ErbB2-ErbB4-рецептора. Клетки линии MDA-MB-175 в течение 4 дней обрабатывали моноклональными антителами 2С4 и 4D5 (10 мкг/мл) к ErbB2. В опытах с использованием окрашивания кристаллическим фиолетовым установлено, что инкубация с 2С4 оказывает сильное ингибирующее действие на рост этой линии клеток (фиг.4А). Присутствие экзогенного HRG не приводит к существенной реверсии этого ингибирования. С другой стороны, не выявлено ингибирующего действия 2С4 на линию клеток SK-BR-3, которые сверхэкспрессируют ErbB2 (фиг.4Б). Моноклональное антитело 2С4, как в присутствии экзогенного HRG, так и без него, обладало способностью ингибировать пролиферацию клеток линии MDA-MB-175 в большей степени, чем моноклональное антитело 4D5. Известно, что ингибирование пролиферации клеток с помощью 4D5 зависит от уровня экспрессии ErbB2 (Lewis и др., Cancer Immunol. Immunother., 37: 255-263 (1993)). Для клеток линии SK-BR-3 оказалось, что максимальное ингибирование составляет 66% (фиг.4Б). Однако это ингибирующее действие может быть превышено при использовании экзогенного HRG.
Пример 2
Зависящее от HRG связывание ErbB2 с ErbB3 блокируется моноклональным антителом 2С4
Способность ErbB3 связывать ErbB2 изучали в опыте с использованием коиммунопреципитации. 1,0×106 клеток линии MCF7 или SK-BR-3 высевали в 6-луночные планшеты для культур тканей в смесь (50:50) DMEM/среда Хама F12, дополненную 10% фетальной телячьей сыворотки и 10 мМ HEPES, рН 7,2 (питательная среда) и давали прикрепиться в течение ночи. Перед началом эксперимента клетки выращивали в течение 2 ч в минимальной питательной среде, не содержащей сыворотку.
Клетки быстро промывали забуференным фосфатом физиологическим раствором (ЗФР) и затем инкубировали либо с 100 нМ указанным антителом, разбавленным 0,2% (мас./об.) бычьего сывороточного альбумина (БСА), либо со средой RPMI, дополненной 10 мМ HEPES, рН 7,2 (буфер для связывания), либо с одним буфером для связывания (контроль). После выдерживания в течение 1 ч при комнатной температуре в половину лунок (+) добавляли HRG до получения конечной концентрации 5 нМ. В остальные лунки (-) добавляли такой же объем буфера для связывания. Инкубацию продолжали в течение приблизительно 10 мин.
Супернатанты удаляли аспирацией и клетки лизировали в среде RPMI, содержащей 10 мМ HEPES, рН 7,2, 1 об% TRITON Х-100™. 1,0% мас./об. CHAPS (буфер для лизиса), включающей 0,2 мМ ФМСФ (фенилметилсульфонилфторид), 10 мкг/мл леупептина и 10 токсических единиц (ТЕ)/мл апротинина. Лизаты очищали от нерастворившегося продукта путем центрифугирования.
ErbB2 подвергали иммунопреципитации с использованием моноклонального антитела, ковалентно связанного с гелем для аффинной хроматографии (типа Affi-Prep 10, фирма Bio-Rad). Указанное антитело (Ab-3, фирма Oncogene Sciences) распознает эпитоп цитоплазматического домена. Иммунопреципитацию осуществляли путем добавления в каждый лизат 10 мкл суспензии геля, содержащей приблизительно 8,5 мкг иммобилизированного антитела, после чего образцам давали смешиваться в течение 2 ч при комнатной температуре. Затем гели собирали центрифугированием. Гели промывали три раза порциями буфера для лизиса для удаления несвязанного продукта. Затем добавляли ДСН-буфер для образца, и образцы в течение короткого промежутка времени нагревали в бане с кипящей водой.
Супернатанты разгоняли на 4-12%-ных полиакриламидных гелях и подвергали электроблоттингу на нитроцеллюлозных мембранах. Присутствие ErbB3 определяли путем зондирования блотов поликлональным антителом к эпитопу его цитоплазматического домена (с-17, фирма Santa Cruz. Biotech). Блоты визуализировали с использованием хемилюминисцентного субстрата (ECL, фирма Amersham).
Как видно по контрольным полосам на фиг.5А и 5Б, полученным для клеток линий MCF7 и SK-BR-3 соответственно, ErbB3 присутствовал в иммунопреципитате ErbB2 только тогда, когда клетки подвергались стимуляции с помощью HRG. Если клетки сначала инкубировали с моноклональным антителом 2С4, то ErbB3-сигнал исчезал в клетках линии MCF7 (фиг.5А, полоса 2С4+) или существенно уменьшался в клетках линии SK-BR-3 (фиг.5Б, полоса 2С4+). Как видно на фиг.5А-Б, моноклональное антитело 2С4 блокировало зависящее от херегулина связывание ErbB3 с ErbB2 как в клетках линии MCF7, так и линии SK-BR-3, существенно более эффективным образом, чем HERCEPTIN®. Предварительная инкубация с HERCEPTIN® приводила к уменьшению ErbB3-сигнала в лизатах MCF7, но оказывала незначительное влияние или не оказывала совсем никакого влияния на количество ErbB3, совместно осажденного из лизатов клеток линии SK-BR-3. Предварительная инкубация с антителом к рецептору EGF (Ab-1, фирма Oncogene Sciences) не оказывала влияния на способность ErbB3 к коиммунопреципитации с ErbB2 во всех линиях клеток.
Пример 3
Гуманизированные антитела 2С4
Вариабельные области мышиного моноклонального антитела 2С4 сначала клонировали в векторе, который позволяет осуществлять получение химерного мышиного/человеческого Fab-фрагмента. Полную РНК выделяли из клеток гибридомы с помощью набора для выделения РНК фирмы Stratagene согласно инструкциям производителя. Вариабельные области амплифицировали с помощью ОТ-ПЦР, очищали с помощью геля и встраивали в плазмиду, являющуюся производной плазмиды с каркасом pUC119, содержащую человеческую константную область каппа-цепи и человеческую СН1-область, согласно ранее опубликованному методу (Carter и др., PNAS (США), 89:4285 (1992); и патент США 5821337). Полученной плазмидой трансформировали штамм E.coli 16C0 с целью экспрессии Fab-фрагмента. Выращивание культур, индукцию экспрессии протеина и очистку Fab-фрагмента осуществляли согласно ранее опубликованному методу (Werther и др., J.Immunol., 157: 4986-4995; Presta и др., Cancer Research, 57: 4593-4599 (1997)).
Очищенный химерный Fab-фрагмент 2С4 сравнивали с мышиным родительским антителом 2С4 в отношении его способности ингибировать связывание 125I-HRG с клетками линии MCF7 и ингибировать опосредуемую rHRG активацию фосфорилирования тирозина р180 в клетках линии MCF7. Как показано на фиг.6А, химерный Fab-фрагмент 2С4 оказался очень эффективным в отношении разрушения образования высокоаффинного сайта связывания ErbB2-ErbB3 на человеческих клетах рака молочной железы линии MCF7. Относительная величина IC50, рассчитанная для интактного мышиного 2С4, составляет 4,0±0,4 нМ, в то время как для Fab-фрагмента она составляет 7,7±1,1 нМ. Как проиллюстрировано на фиг.6Б, одновалентный химерный Fab-фрагмент 2С4 оказался очень эффективным в отношении нарушения опосредуемой HRG активации ErbB2- ErbB3. Величина IC50, рассчитанная для интактного мышиного 2С4, составляет 6,0±2 нМ, в то время как для Fab-фрагмента она составляет 15,0±2 нМ.
Секвенирование ДНК химерного клона позволило осуществить идентификацию остатков CDR (Kabat и др., Sequences of Proteins of Immunological Interest, 5-е изд., Public Health Service, National Institutes of Health, Bethesda, MD (1991)) (фиг.7A и 7Б). С помощью сайтнаправленного мутагенеза с использованием олигонуклеотидов все шесть этих CDR-участков встраивали в полный человеческий каркасный участок (VL каппа-цепиL подгруппа I и VH подгруппа III), содержащийся в плазмиде VX4, согласно ранее опубликованному методу (Presta и др., Cancer Research, 57: 4593-4599 (1997)). Протеин из продукта, полученного путем "CDR-обмена", экспрессировали и очищали согласно описанному выше методу. Для сравнения двух вариантов проводили анализы связывания. В целом метод состоял в следующем. Планшет типа NUNC MAXISORP™ сенсибилизировали в течение ночи при 4°С с помощью 1 мкг/мл внеклеточного домена ErbB2 (ECD; полученный с помощью метода, описанного в WO 90/14357) в 50 мМ карбонатном буфере, рН 9,6, и затем блокировали с помощью разбавителя для ELISA (0,5% БСА, 0,05% полисорбата 20, ЗФР) в течение 1 ч при комнатной температуре. Серийные разведения образцов в разбавителе для ELISA инкубировали на планшетах в течение 2 ч. После промывки связанный Fab-фрагмент выявляли с применением несущего биотиновую метку мышиного антитела к человеческой каппа-цепи (ICN 634771) и затем конъюгированной со стрептавидином пероксидазы из хрена (фирма Sigma), используя в качестве субстрата 3,3',5,5'-тетраметилбензидин (фирма Kirkegaard & Perry Laboratories, Гейтерсбург, штат Мэриленд). Абсорбцию измеряли при длине волны 450 нм. Как проиллюстрировано на фиг.8А, для конструкции человеческого "CPR-обмена", связывание полностью отсутствовало. Для восстановления связывания гуманизированного Fab-фрагмента, полученного путем "CDR-обмена", связывание полностью отсутствовало. Для восстановления связывания гуманизированного Fab-фрагмента конструировали мутанты, используя в качестве матрицы полученный путем "CDR-обмена" продукт. Используя полученную с помощью компьютера модель (фиг.9), создавали мутации с целью замены остатков человеческого каркасного участка на их мышиные копии в положениях, где изменение могло влиять на конформацию CDR или на взаимодействие антитело - антиген. Мутанты перечислены в таблице 2.
Кривые связывания для различных мутантов приведены на фиг.8А-В. Fab-фрагмент гуманизированной версии, имеющий замены ArgH71Val, AspH73Arg и IlaH69Leu, по-видимому, обладает способностью к связыванию, восстановленной до уровня, присущего Fab-фрагменту исходного химерного 2С4. Для того чтобы еще более улучшить или усилить связывание гуманизированного антитела, могут быть сделаны дополнительные модификации остатков FR и/или CDR, таких как L2, L54, L55, L56, Н35, Н48 (например, могут быть сделаны следующие замены: IleL2Thr; ArgL54Leu; TyrL56Ser; AspH35Ser и ValH48Ile). В альтернативном или дополнительном варианте для дальнейшего увеличения или улучшения аффинности и/или других видов биологической активности гуманизированное антитело может быть подвергнуто "созреванию для проявления выраженной аффинности (см. выше).
Гуманизированную версию 574 2С4 подвергали "созреванию для проявления выраженной аффинности" с использованием метода проявления с помощью фага. В целом метод состоит в следующем. Гуманизированный Fab-фрагмент 2С4.574 клонировали в векторе для проявления с помощью фага в виде слияния с геном III. Когда частицы фага индуцируют путем инфицирования фагом-хелпером М13КO7, это слияние дает возможность Fab проявляться на N-конце конструкции отросток фага-файбер-протеин, ген III (Васа и др., J.Biol. Chem., 272: 10678 (1997)).
Для каждого из шести указанных выше CDR создавали индивидуальные библиотеки. В этих библиотеках аминокислоты в CDR, которые были выявлены с использованием созданной с помощью компьютера модели (фиг.9) в качестве потенциально важных для связывания с ErbB2, рандомизировали с помощью олигонуклеотидов, содержащих кодоны "NNS". Затем библиотеки подвергали пэннингу в отношении ECD ErbB2, которым сенсибилизировали планшеты NUNC MAXISORP™, используя вместо всех блокирующих растворов 3%-ное сухое молоко в ЗФР с добавлением 0,2% TWEEN 20® (MPBST). Для отбора фагов с аффинностями выше, чем у 2С4.574, на 3, 4 и 5 циклах пэннинга на стадиях промывки добавляли в качестве конкурента растворимый ECD ErbB2 или растворимый Fab-фрагмент 2С4.574. Времена промывки при комнатной температуре увеличивали на 1 ч.
После 5 циклов пэннинга индивидуальные клоны снова анализировали с помощью ELISA с использованием фага. Индивидуальные клоны выращивали в 96-луночных планшетах для культур тканей типа Costar с U-образным дном и фаги индуцировали, добавляя фаг-хелпер. После выращивания в течение ночи клетки Е.coli пеллетировали и содержащие фаг супернатанты переносили в 96-луночные планшеты, где фаг блокировали с помощью MPBST в течение 1 ч при комнатной температуре. Планшеты NUNC MAXISORP™, сенсибилизированные с помощью ECD ErbB2, также блокировали с помощью MPBST согласно описанному выше методу. Блокированный фаг инкубировали на планшетах в течение 2 ч. После промывки связанные фаги обнаруживали с использованием конъюгированного с пероксидазой из хрена моноклонального антитела к М13 (фирма Amersham Pharmacia Biotech, Inc., 27-9421-01), разбавленного MPBST в соотношении 1:5000, а затем с использованием 3,3',5,5'-тетраметилбензидина в качестве субстрата. Абсорбцию измеряли при длине волны 450 нм.
Из каждой библиотеки производили секвенирование ДНК 48 клонов, которые давали наибольшие сигналы. Клоны, последовательности которых встречались наиболее часто, субклонировали в описанном выше векторе, который позволял осуществлять экспрессию растворимых Fab-фрагментов. Эти Fab-фрагменты индуцировали, протеины очищали, и очищенные Fab-фрагменты анализировали в отношении связывания с помощью ELISA согласно описанному выше методу, и связывание сравнивали со связыванием исходной гуманизированной версии 2С4.574.
После осуществления идентификации представляющих интерес мутаций в отдельных CDR конструировали дополнительные мутанты, имеющие различные комбинации этих мутаций, и проводили их тестирование согласно описанному выше методу. Мутанты, обладающие более высокой способностью к связыванию по сравнению с версией 574, перечислены в таблице 3.
Также были сконструированы перечисленные ниже мутанты, которые в настоящее время проходят тестирование:
В настоящее время конструируются перечисленные ниже мутанты, выбранные на основе выявления гомологии:
Предпочтительной аминокислотой в положении Н34 должен быть метионин. Может быть сделана замена на лейцин, если обнаружено, что в этом положении происходит окисление.
Установлено, что asnH52 и asnH53 являются наиболее предпочтительными для связывания. Замена этих остатков на аланин или аспарагиновую кислоту приводит к очень сильному уменьшению связывания.
Создавали интактное антитело, содержащее вариабельные области легкой и тяжелой цепи гуманизированной версии 574 и константную область тяжелой цепи человеческого IgG1 (см. патент США 5821337). Интактное антитело получали с использованием клеток яичника китайского хомячка (СНО). В настоящем описании молекула обозначена как rhuMAb 2C4.
Пример 4
Моноклональное антитело 2C4 блокирует активацию МАРК, опосредуемую EGF, TGF-α или HRG
Многие рецепторы фактора роста передают сигнал с помощью пути, опосредуемого активируемой митогеном протеинкиназой (МАРК). Такие обладающие двойной специфичностью киназы являются одним из имеющих решающее значение факторов пути трансдукции сигнала, который в конце концов стимулирует деление раковых клеток. Способность моноклонального антитела 2C4 или HERCEPTIN® ингибировать активацию МАРК, опосредуемую EGF, TGF-α или HRG, оценивали следующим образом.
Клетки линии MCF7 (105 клеток/лунку) высевали в среду, содержащую сыворотку, в 12-луночные планшеты для клеточных культур. На следующий день среду для клеток удаляли и в каждую лунку добавляли свежую среду, содержащую 0,1% сыворотки. Затем на следующий день процедуру повторяли и перед анализом среду заменяли бессывороточным буфером для связывания (Jones и др., J.Biol. Chem., 273: 11667-11674 (1998); Schaefer и др., J.Biol. Chem., 274: 859-866 (1999)). Клеткам давали уравновеситься при комнатной температуре и затем их инкубировали в течение 30 мин с 0,5 мл 200 нМ HERCEPTIN® или моноклонального антитела 2C4. Затем клетки обрабатывали в течение 15 мин 1 нМ EGF, 1нМ TGF-α или 0,2 нМ HRG. Реакцию прекращали с помощью аспирации клеточной среды и последующего добавления 0,2 мл буфера для образцов для ДСН-ПААГ, содержащего 1% ДТТ. Активацию МАРК оценивали с помощью Вестерн-блотинга с использованием антитела к МАРК (фирма Promega) согласно ранее опубликованному методу (Jones и др., J.Biol. Chem., 273: 11667-11674 (1998)).
Как следует из данных, представленных на фиг.10, моноклональное антитело 2С4 блокирует активацию МАРК, опосредуемую EGF, TGF-α или HRG, в существенно большей степени, чем HERCEPTIN®. Эти данные позволяют предположить, что моноклональное антитело 2С4 связывается с поверхностью ErbB2, которая нужна для его связывания либо с EGFR, либо с ErbB3, и таким образом препятствует образованию сигнального рецепторного комплекса.
Также было установлено, что моноклональное антитело 2С4 ингибирует опосредуемую херегулином (HRG) активацию АКТ. Активация пути трансдукции сигнала киназы PI3 имеет важное значения для выживания клеток (Carraway и др., J.Biol. Chem., 270: 7111-7116 (1995)). В клетках опухоли активация киназы PI3 может иметь значение для развития инвазивного фенотипа (Tan и др., Cancer Research, 59: 1620-1625 (1999)). Путь выживания в основном опосредуется серин/треонинкиназой АКТ (Bos и др., Trends Biochem. Sci., 20: 441-442 (1995)). Комплексы, образованные ErbB2 либо с ErbB3, либо с EGFR, могут инициировать эти пути в ответ на воздействие херегулина или EGF соответственно (Olayioye и др., Mol. & Cell. Biol., 18: 5042-5051 (1998); Karunagaran и др., EMBO Journal, 15: 254-264 (1996); Krymskaya и др., Am. J.Phisiol., 276: L246-255 (1999)). Инкубация клеток рака молочной железы линии MCF7 с 2С4 ингибирует опосредуемую херегулином активацию АКТ. Более того, исходный уровень активации АКТ, имеющийся в отсутствие херегулина, еще более понижается при добавлении 2С4. Эти результаты позволяют предположить, что 2С4 может ингибировать опосредуемую лигандом ErbB активацию киназы PI3 и что это ингибирование может приводить к апоптозу. Повышенная чувствительность к апоптозу может проявляться в более высокой чувствительности клеток опухоли к токсическим воздействиям химиотерапии.
Таким образом, моноклональное антитело 2С4 ингибирует инициируемую лигандом посылку ErbB-сигнала по двум основным путям трансдукции сигнала - по опосредуемому МАРкиназой (основной путь пролиферации) и Р13-киназой (основной путь выживания/антиапоптоза).
Пример 5
Исследование in vivo эффективности комбинации моноклонального антитела 2С4 и HERCEPTIN®
Для оценки эффективности моноклональных антител к HER2, применяемых индивидуально или в сочетании, в отношении подавления роста опухоли применяли модель на основе ксенотрансплантата с использованием линии аденокарциномы легкого Calu-3. Самкам бестимусных мышей линии NCR подкожно инокулировали 20×106 клеток в объеме 0,1 мл. Изменения опухоли проводили дважды в неделю и когда узлы опухоли достигали объема 100 мм3, животных распределяли случайным образом на 7 групп обработки. Этим группам вводили:
(а) моноклональное антитело Mab 1766 в качестве контроля;
(б) HERCEPTIN®, 10 мг/кг;
(в) моноклональное антитело 7С2, 10 мг/кг;
(г) моноклональное антитело 2С4, 10 мг/кг;
(д) HERCEPTIN® и 7С2, каждое в дозе 10 мг/кг;
(е) HERCEPTIN® и 2С4, каждое в дозе 10 мг/кг; и
(ж) моноклональные антитела 2С4 и 7С2, каждое в дозе 10 мг/кг.
Животных обрабатывали дважды в неделю в течение 24 дней. Объемы опухолей измеряли дважды в неделю в течение 38 дней.
Как следует из столбчатых диаграмм, представленных на фиг.11, обработка мышей, имеющих опухоли Calu-3, с использованием 2С4 или HERCEPTIN® приводила к существенному ингибированию роста опухолей. Комбинация HERCEPTIN® и 2С4 или HERCEPTIN® и 7С2 оказались более эффективными, чем любое моноклональное антитело, вводимое индивидуально.
Пример 6
Лечение рака ободочной кишки с помощью моноклонального антитела 2С4
Человеческие клетки рака ободочной кишки таких линий, как НСА-7, LS 174Т или СаСо-2, подкожно имплантировали бестимусным (лишенным волосяного покрова) мышам согласно методу, описанному у Sheng и др., J.Clin. Invest., 99: 2254-2259 (1997)). Когда опухоли достигали объема приблизительно 100 мм3, группы животных обрабатывали моноклональным антителом 2С4 в дозе 10-50 мг/кг, которую вводили дважды в неделю путем инъекции во внутрибрюшинную полость. Было установлено, что моноклональное антитело подавляло in vivo рост ксенотрансплантатов рака ободочной кишки.
Пример 7
Лечение рака молочной железы с помощью гуманизированного 2С4
Воздействие rhuMab 2C4 или HERCEPTIN® на клетки рака молочной железы, которые не обладают способностью сверхэкспрессировать ErbB2, оценивали в течение 3 дней с помощью анализа с использованием аламарового синего (S.A.Ahmed, J.Immunol. Methods, 170: 211-224 (1994); и Page и др., Int. J. Oncol., 3: 473-476 (1994)). Клетки, которые использовали в этом анализе, представляли собой человеческие клетки рака молочной железы линии MDA-175, которые экспрессируют ErbB2 с уровнем 1+. Как следует из данных, представленных на фиг.12, при добавлении rhuMab 2C4 происходило более существенное ингибирование в зависимости от дозы роста клеток рака молочной железы линии MDA-175, чем при использовании HERCEPTIN®.
Оценивали эффективность rhuMab 2C4 в отношении ксенотрансплантатов клеток линии MCF7, которые являются позитивными в отношении рецептора эстрогена (ER+) и экспрессируют низкие уровни ErbB2. Использовали самок мышей, которым вводили эстроген. rhuMab 2C4 вводили один раз в неделю в дозе 30 мг/кг. Как следует из данных, представленных на фиг.13, rhuMab 2C4 оказался эффективным в отношении ингибирования in vivo роста опухоли рака молочной железы, клетки которой не обладают способностью сверхэкспрессировать ErbB2.
Пример 8
Фармакокинетика, метаболизм и токсикология 2C4
Было установлено, что rhuMab 2C4 обладало стабильностью в человеческой сыворотке. Не было обнаружено наличия или доказательства образования комплексов в биологических матрицах. Из организма мышей rhuMab 2C4 выводилось быстрее, чем HERCEPTIN®. Фармакокинетические исследования позволили установить, что еженедельное введение приблизительно 2-6 мг/кг rhuMab 2C4 должно приводить к возникновению концентраций в сыворотке, близких к концентрациям, возникающим при введении HERCEPTIN® в применяемых в настоящее время дозах. Уровни в сыворотке, возникающие после обработки 2C4, должны существенно превышать величину IC50, определенную в опытах in vitro.
Токсикологические исследования проводили на обезьянах Cynomolgus (по 2 самца и 2 самки в группе). rhuMab 2C4 вводили внутривенно в дозах 0, 10, 50 или 100 мкг/кг дважды в неделю в течение 4 недель. В ходе токсикологического исследования проводили измерения веса тела (за 2 и 1 неделю до опыта, далее еженедельно); оценку поглощения пищи (качественная оценка, ежедневно); физические исследования, включающие измерение кровяного давления, снятие электрокардиограммы (ЭКГ) и измерение температуры тела (за 2 и 1 неделю до опыта, далее через 2 и 4 недели через 4 ч после введения второй дозы, введенной в эту неделю); ультразвуковое обследование сердца (после введения первой дозы в первую неделю и после окончания исследования, т.е. через 4 недели); оценку клинической патологии (исходный уровень и через 2 и 4 недели); анализ мочи (исходный уровень и через 2 и 4 недели); взятие проб для анализа антител (исходный уровень и через 2 и 4 недели); а также аутопсию и гистопатологический анализ.
Все животные во всех группах выжили до конца эксперимента. Не было зафиксировано никаких существенных клинических симптомов или различий между группами. Результаты аутопсии не выявили существенных аномалий в величине органов у всех животных. При микроскопическом исследовании не выявлено существенных аномалий в тканях у всех животных. Не было обнаружено существенных изменений в ЭКГ на протяжении всего исследования. Кроме того, не было отмечено различия между группами.
Пример 9
Увеличение дозы
Пациентам, больным раком, вводили первую дозу rhuMab 2C4 соответствующую одному из пяти уровней доз (0,05, 0,5, 2,0, 4,0 или 10 мг/кг; по 6 пациентов для каждого уровня доз), после чего наступал 4-недельный период выведения. Через 5 недель пациентам вводили такую же дозу 4 раза еженедельно, после чего вновь наступал 4-недельный период выведения. Пациентов с полной реакцией, частичной реакцией или стабильной болезнью отбирали для дальнейших исследований.
Пример 10
Терапия повторного или устойчивого метастатического рака предстательной железы rhuMab 2C4 представляет собой полноразмерное гуманизированное моноклональное антитело к ErbB2 (продуцируемое в клетках СНО). rhuMab 2C4 блокирует связывание ErbB2 с другими представителями к ErbB-семейства, тем самым ингибируя посылку внутриклеточных сигналов ErbB-путем. В отличие от HERCEPTIN® rhuMab 2C4 не только ингибирует рост опухолей, сверхэкспрессирующих ErbB2, но также блокирует рост опухолей, которые требуют наличия опосредуемого лигандом ErbB сигнала.
rhuMab 2C4 может применяться в качестве отдельного агента для лечения пациентов, страдающих устойчивым к действию гормонов (независимым от андрогена) раком предстательной железы. Основные показатели эффективности включают общую выживаемость, достигаемую при применении в качестве отдельного лекарственного средства, взятую в сравнении с выживаемостью при использовании наиболее эффективного лечения (митоксантрон/преднизон), и безопасность. Вспомогательные показатели эффективности включают: время до развития болезни, скорость реакции, качество жизни, боль и/или продолжительность реакции. rhuMab 2C4 вводят внутривенно (i.v.) еженедельно или каждые три недели в дозах 2 или 4 мг/кг соответственно до тех пор, пока продолжается развитие болезни. Антитело поступает в продажу в виде жидкой композиции, содержащей несколько доз (объемом 20 мл с концентрацией 20 мг/мл или более высокой концентрацией).
rhuMab 2C4 может также применяться в сочетании с химиотерапией для лечения пациентов, страдающих устойчивым к действию гормонов (независимым от андрогена) раком предстательной железы. Основные показатели эффективности включают общую выживаемость, взятую в сравнении с выживаемостью при использовании химиотерапии, и безопасность. Вспомогательные показатели эффективности включают: время до развития болезни, скорость реакции, качество жизни, боль и/или продолжительность реакции. rhuMab 2C4 вводят внутривенно (i.v.) еженедельно или каждые три недели в дозах 2 или 4 мг/кг соответственно до тех пор, пока продолжается развитие болезни. Антитело поступает в продажу в виде жидкой композиции, содержащей несколько доз (объемом 20 мл с концентрацией 20 мг/мл или более высокой концентрацией).
Примеры лекарственных средств, которые могут применяться в сочетании с антителом к ErbB2 (которое блокирует активацию лигандом рецептора ErbB2) для лечения рака предстательной железы (например, независимого от андрогена рака предстательной железы) включают ингибитор фарнезилтрансферазы; антиангиогенный агент (например, антитело к VEGF); лекарственное средство, мишенью которого является EGFR (например, С225 или ZD1839); другое антитело к ErbB2 (например, ингибирующее рост антитело к ErbB2, такое как HERCEPTIN®, или антитело к ErbB2, которое индуцирует апоптоз, такое как 7С2 или 7F3, включая их гуманизированные варианты и/или созревшие варианты с выраженной аффинностью); цитокин (например, IL-2, IL-12, G-CSF или GM-CSF); антиандроген (такой как флутамид или ацетат ципротерона); леупролид; сурамин; химиотерапевтический агент, такой как винбластин, эстрамустин, митоксантрон, лиарозол (агент, блокирующий метаболизм ретиноевой кислоты), циклофосфамид, антрациклиновые антибиотики, такие как доксорубицин, таксан (например, паклитаксел или доцетаксел) или метотрексат, или любую комбинацию перечисленных выше лекарственных средств, например винбластин/эстрамустин или циклофосфамид/доксорубицин/метотрексат; преднизон; гидрокортизон; или их комбинацию. Могут вводиться стандартные дозы этих различных лекарственных средств, например 40 мг/м2/еженедельно доксетаксела (TAXOTERE®); 6 (AUC) карбоплатина и 200 мг/м2 паклитаксела (TAXOL®).
Пример 11
Терапия метастатического рака молочной железы
rhuMab 2C4 может применяться в качестве индивидуального агента для лечения пациентов, страдающих метастатическим раком молочной железы, при котором опухолевые клетки не обладают способностью сверхэкспрессировать ErbB2. Основные показатели эффективности включают скорость реакции и безопасность. Вспомогательные показатели эффективности включают: общую выживаемость, время до развития болезни, качество жизни и/или продолжительность реакции. rhuMab 2C4 вводят внутривенно (i.v.) еженедельно или каждые три недели в дозах 2 или 4 мг/кг соответственно до тех пор, пока продолжается развитие болезни. Антитело поступает в продажу в виде жидкой композиции, содержащей несколько доз (объемом 20 мл с концентрацией 20 мг/мл или более высокой концентрацией).
rhuMab 2C4 может также применяться в сочетании с химиотерапией для лечения пациентов, страдающих метастатическим раком молочной железы, при котором опухолевые клетки не обладают способностью сверхэкспрессировать ErbB2. Основные показатели эффективности включают общую выживаемость, взятую в сравнении с выживаемостью при использовании только химиотерапии, и безопасность. Вспомогательные показатели эффективности включают: время до развития болезни, скорость реакции, качество жизни и/или продолжительность реакции. rhuMab 2C4 вводят внутривенно (i.v.) еженедельно или каждые три недели в дозах 2 или 4 мг/кг соответственно до тех пор, пока продолжается развитие болезни. Антитело поступает в продажу в виде жидкой композиции, содержащей несколько доз (объемом 20 мл с концентрацией 20 мг/мл или более высокой концентрацией). Примеры лекарственных средств, которые могут применяться в сочетании с антителом к ErbB2 (которое блокирует активацию лигандом рецептора ErbB2) для лечения рака молочной железы (например, метастатического рака молочной железы, при котором опухолевые клетки не обладают способностью сверхэкспрессировать ErbB2), включают химиотерапевтические агенты, такие как антрациклиновые антибиотики (например, доксорубицин), циклофосфомид, таксан (например, паклитаксел или доцетаксел), навелбин, кселода, митомицин С, соединения на основе платины, оксалиплатин, гемцитабин или комбинацию, включающую два или более таких лекарственных средств, например доксорубицин/циклофосфомид; другое антитело к ErbB2 (например, ингибирующее рост антитело к ErbB2, такое как HERCEPTIN®, или антитело к ErbB2, которое индуцирует апоптоз, такое как 7С2 или 7F3, включая их гуманизированные варианты и/или созревшие варианты с выраженной аффинностью); антиэстроген (например, тамоксифен); ингибитор фарнезилтрансферазы; антиангиогенный агент (например, антитело к VEGF); лекарственное средство, мишенью которого является EGFR (например, С225 или ZD1839); цитокин (например, IL-2, IL-12, G-CSF или GM-CSF) или комбинацию перечисленных выше лекарственных средств. Для таких дополнительных лекарственных средств могут использоваться стандартные дозы.
Кроме того, rhuMab 2C4 может применяться в сочетании с HERCEPTIN® для лечения пациентов, страдающих метастатическим раком молочной железы, при котором опухолевые клетки обладают способностью сверхэкспрессировать ErbB2. Основные показатели эффективности включают скорость реакции и безопасность. Вспомогательные показатели эффективности включают: время до развития болезни, общую выживаемость, взятую в сравнении с выживаемостью при использовании только HERCEPTIN®, качество жизни и/или продолжительность реакции. rhuMab 2C4 вводят внутривенно (i.v.) еженедельно или каждые три недели в дозах 2 или 4 мг/кг соответственно до тех пор, пока продолжается развитие болезни. Антитело поступает в продажу в виде жидкой композиции, содержащей несколько доз (объемом 20 мл с концентрацией 20 мг/мл или более высокой концентрацией). HERCEPTIN® вводят i.v. в начальной ударной дозе 4 мг/кг, а затем еженедельно в поддерживающей дозе 2 мг/кг. HERCEPTIN® поступает в продажу в виде лиофилизированного порошка. Каждая ампула с HERCEPTIN® содержит 440 мг HERCEPTIN®, 9,9 мг гидрохлорида L-гистидина, 6,4 мг L-гистидина, 400 мг дигидрата α-α-трегалозы и 1,8 мг полисорбата 20. Восстановление с помощью 20 мл бактериостатической воды для инъекций (БВИ), содержащей 1,1% бензилового спирта в качестве консерванта, позволяет получить 21 мл раствора, что соответствует нескольким дозам, с концентрацией HERCEPTIN®, равной 21 мг/мл, рН приблизительно 6,0.
Пример 12
Терапия рака легкого
rhuMab 2C4 может применяться в качестве индивидуального агента для лечения стадии IIIb или IV немелкоклеточного рака легкого (НМКРЛ). Основные показатели эффективности включают скорость реакции и безопасность. Вспомогательные показатели эффективности включают: общую выживаемость, время до развития болезни, качество жизни и/или продолжительность реакции. rhuMab 2C4 вводят внутривенно (i.v.) еженедельно или каждые три недели в дозах 2 или 4 мг/кг соответственно до тех пор, пока продолжается развитие болезни. Антитело поступает в продажу в виде жидкой композиции, содержащей несколько доз (объемом 20 мл с концентрацией 20 мг/мл или более высокой концентрацией).
rhuMab 2C4 может также применяться в сочетании с химиотерапией для лечения пациентов, страдающих метастатическим немелкоклеточным раком легкого. Основные показатели эффективности включают общую выживаемость, взятую в сравнении с выживаемостью при использовании стандартной терапии, и безопасность. Вспомогательные показатели эффективности включают: время до развития болезни, скорость реакции, качество жизни и/или продолжительность реакции. rhuMab 2C4 вводят внутривенно (i.v.) еженедельно или каждые три недели в дозах 2 или 4 мг/кг соответственно до тех пор, пока продолжается развитие болезни. Антитело поступает в продажу в виде жидкой композиции, содержащей несколько доз (объемом 20 мл с концентрацией 20 мг/мл или более высокой концентрацией). Примеры дополнительных лекарственных средств, которые могут применяться в сочетании с антителом (которое связывает ErbB2 и блокирует активацию лигандом рецептора ErbB) для лечения рака легкого включают химиотерапевтические агенты, такие как карбоплатин, таксан (например, паклитаксел или доцетаксел), гемцитабин, навелбин, цисплатин, оксалиплатин или комбинацию любых перечисленных лекарственных средств, такую как карбоплатин/доцетаксел; другое антитело к ErbB2 (например, ингибирующее рост антитело к ErbB2, такое как HERCEPTIN®, или антитело к ErbB2, которое индуцирует апоптоз, такое как 7С2 или 7F3, включая их гуманизированные варианты и/или созревшие варианты с выраженной аффинностью); ингибитор фарнезилтрансферазы; антиангиогенный агент (например, антитело к VEGF); лекарственное средство, мишенью которого является EGFR (например, С225 или ZD1839); цитокин (например, IL-2, IL-12, G-CSF или GM-CSF) или комбинацию перечисленных выше лекарственных средств.
Пример 13
Терапия колоректального рака
rhuMab 2C4 может применяться в качестве отдельного агента для лечения метастатического колоректального рака. Основные показатели эффективности включают скорость реакции и безопасность. Вспомогательные показатели эффективности включают: время до развития болезни, качество жизни и/или продолжительность реакции. rhuMab 2C4 вводят внутривенно (i.v.) еженедельно или каждые три недели в дозах 2 или 4 мг/кг соответственно до тех пор, пока продолжается развитие болезни. Антитело поступает в продажу в виде жидкой композиции, содержащей несколько доз (объемом 20 мл с концентрацией 20 мг/мл или более высокой концентрацией).
rhuMab 2C4 может также применяться в сочетании с химиотерапией для лечения пациентов, страдающих метастатическим колоректальным раком. Основные показатели эффективности включают общую выживаемость, взятую в сравнении с выживаемостью при использовании стандартной терапии, и безопасность. Вспомогательные показатели эффективности включают: время до развития болезни, скорость реакции, качество жизни и/или продолжительность реакции. rhuMab 2C4 вводят внутривенно (i.v.) еженедельно или каждые три недели в дозах 2 или 4 мг/кг соответственно до тех пор, пока продолжается развитие болезни. Антитело поступает в продажу в виде жидкой композиции, содержащей несколько доз (объемом 20 мл с концентрацией 20 мг/мл или более высокой концентрацией).
Примеры химиотерапевтических агентов, используемых для лечения колоректального рака, которые могут применяться в сочетании с антителом, которое связывает ErbB2 и блокирует активацию лигандом рецептора ErbB, включают 5-флуорацил (5-FU), леуковорин (LV), СРТ-11, левамизол или комбинацию любых двух или большего количества указанных лекарственных средств, например 5-FU/LV/CPT-11. Могут вводиться стандартные дозы таких химиотерапевтических агентов. Другие лекарственные средства, которые могут применяться в сочетании с антителом к ErbB2 для лечения колоректального рака, включают ингибитор фарнезилтрансферазы; антиангиогенный агент (например, антитело к VEGF); лекарственное средство, мишенью которого является EGFR (например, С225 или ZD1839); цитокин (например, IL-2, IL-12, G-CSF или GM-CSF); другое антитело к ErbB2 (например, ингибирующее рост антитело к ErbB2, такое как HERCEPTIN®, или антитело к ErbB2, которое индуцирует апоптоз, такое как 7С2 или 7F3, включая их гуманизированные варианты и/или созревшие варианты с выраженной аффинностью) или комбинацию перечисленных выше лекарственных средств.







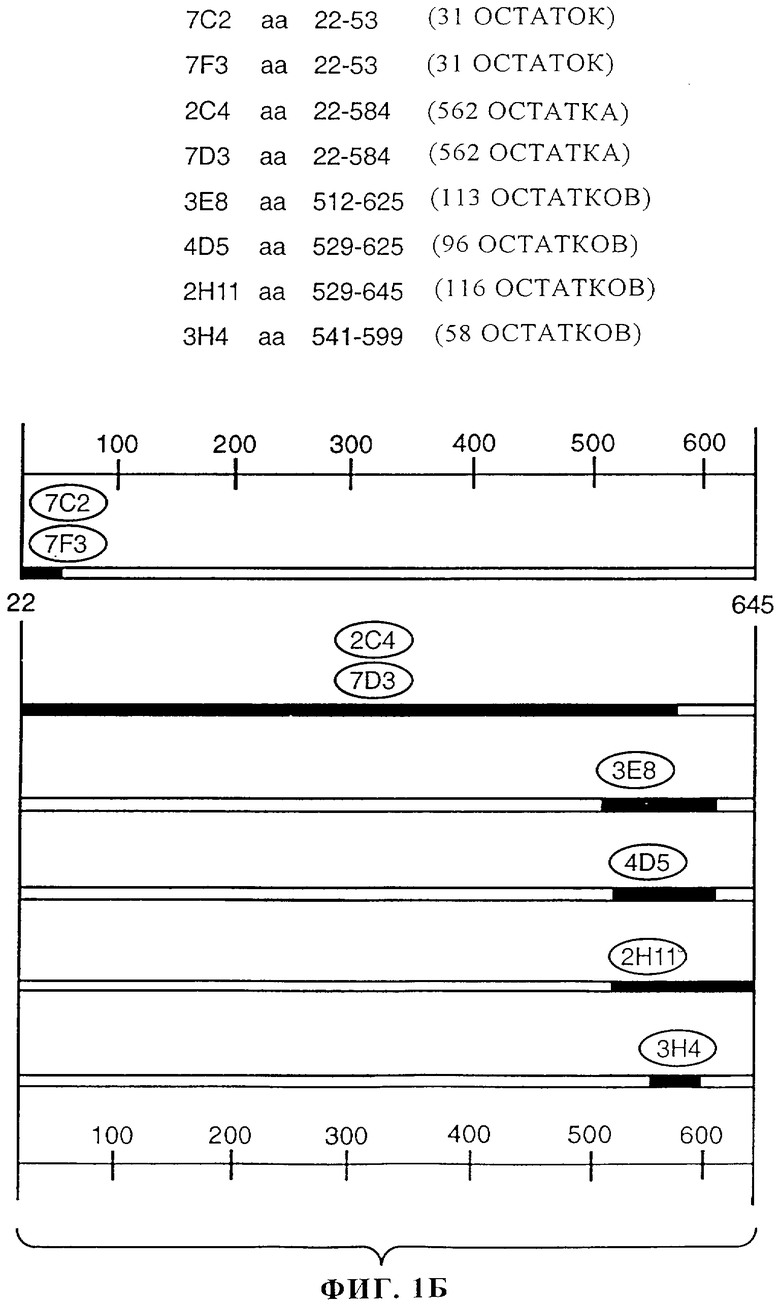
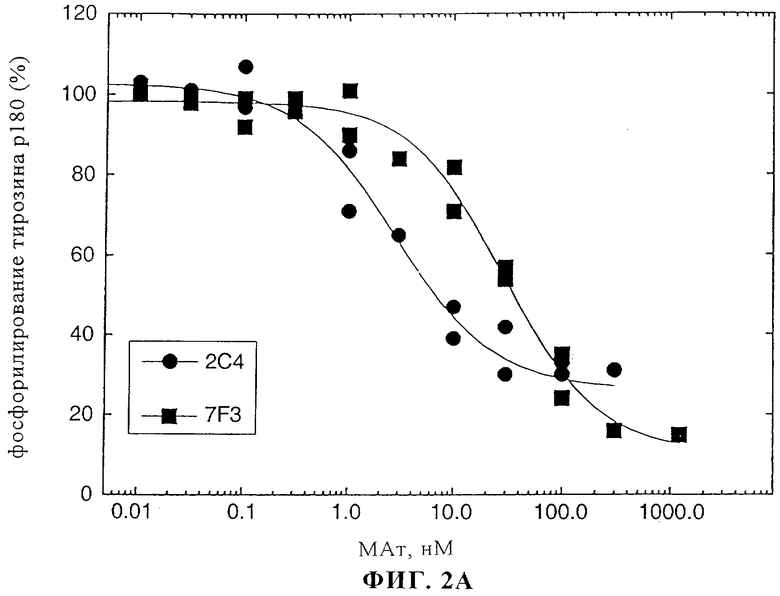

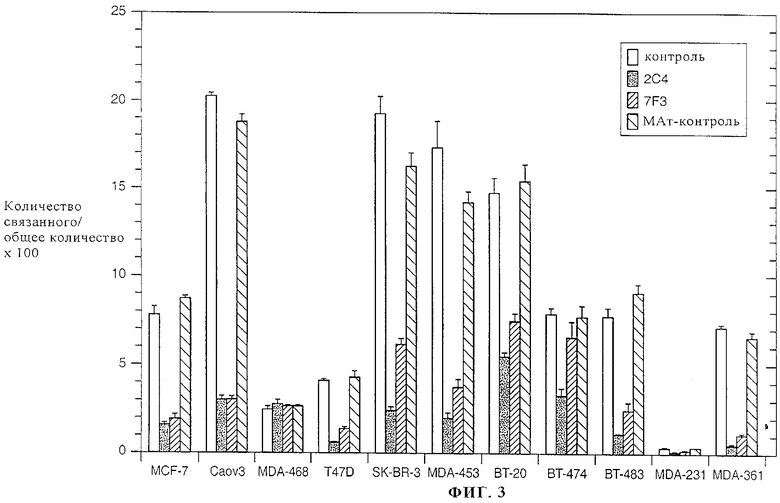
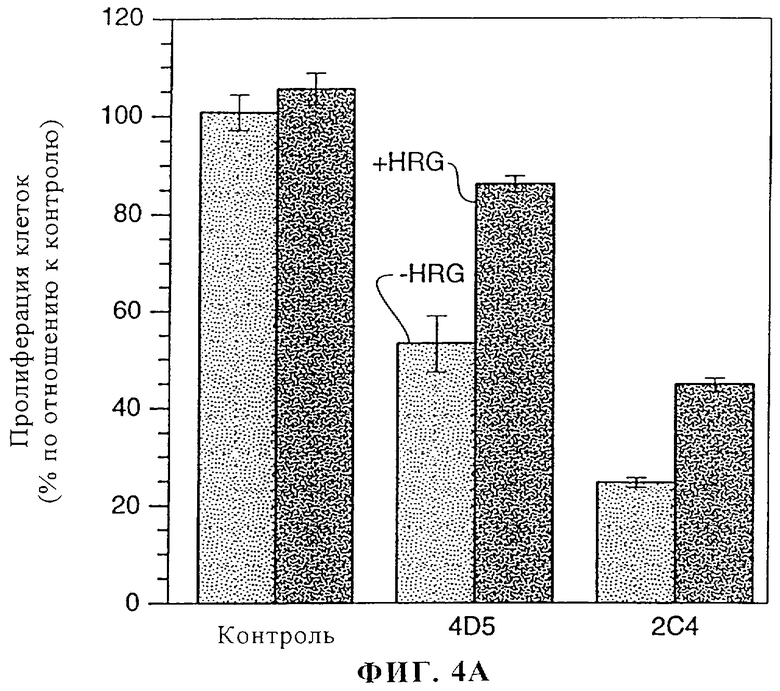
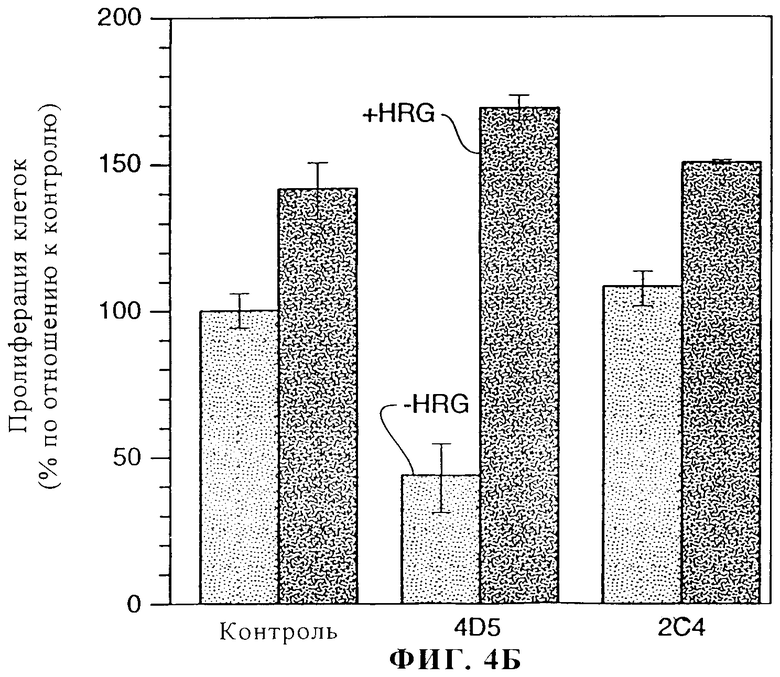

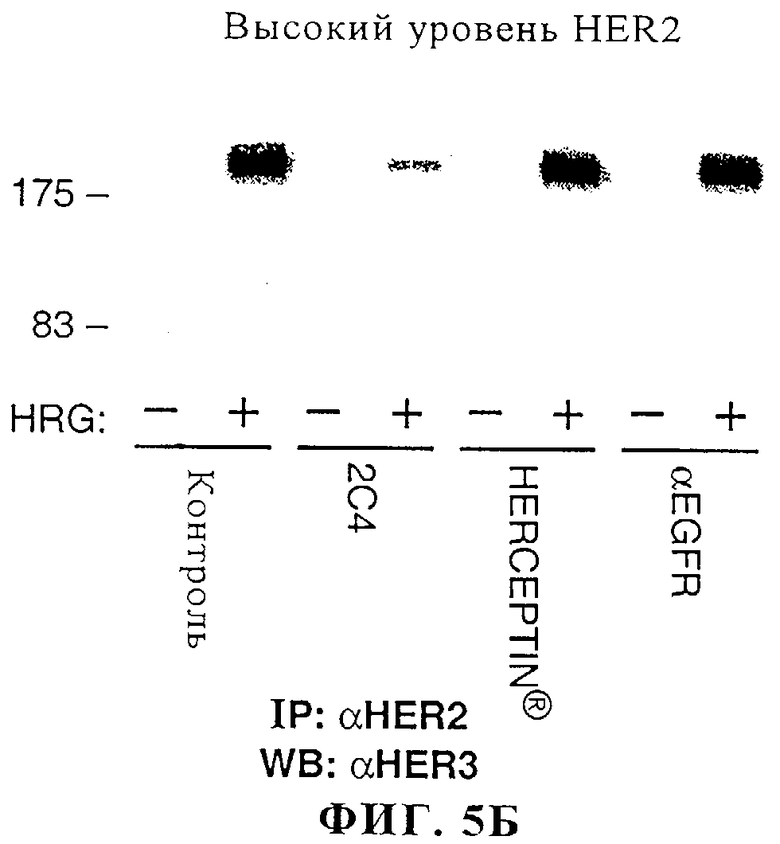
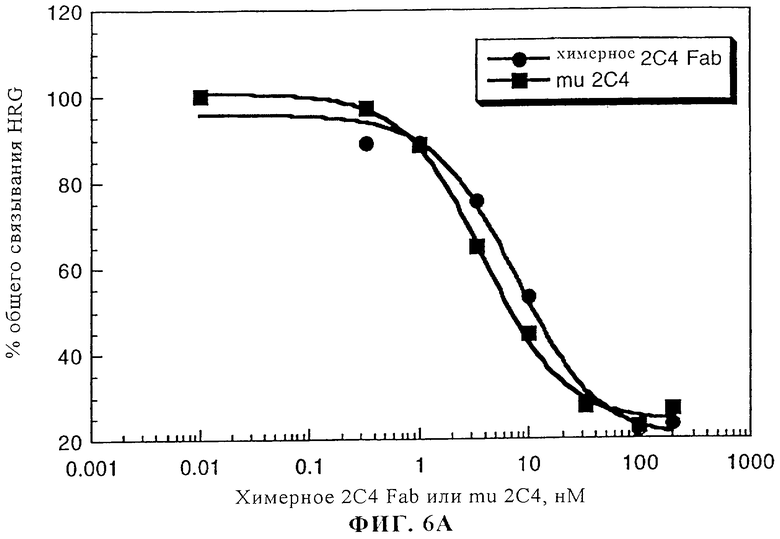
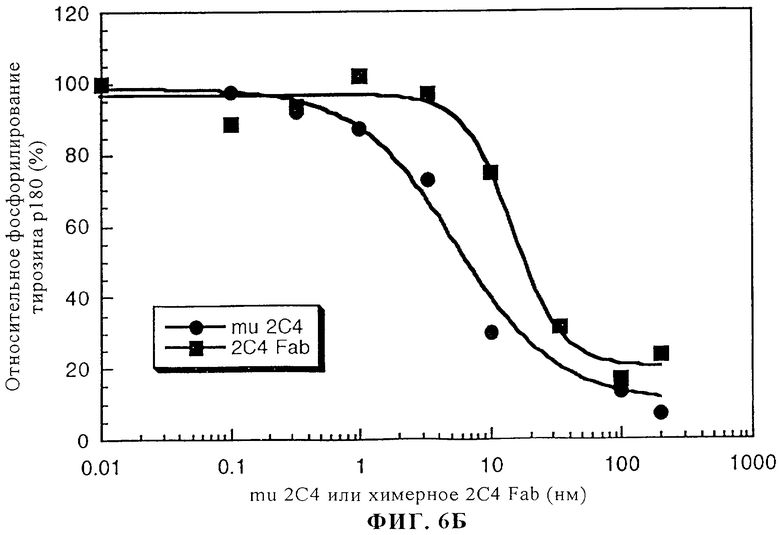


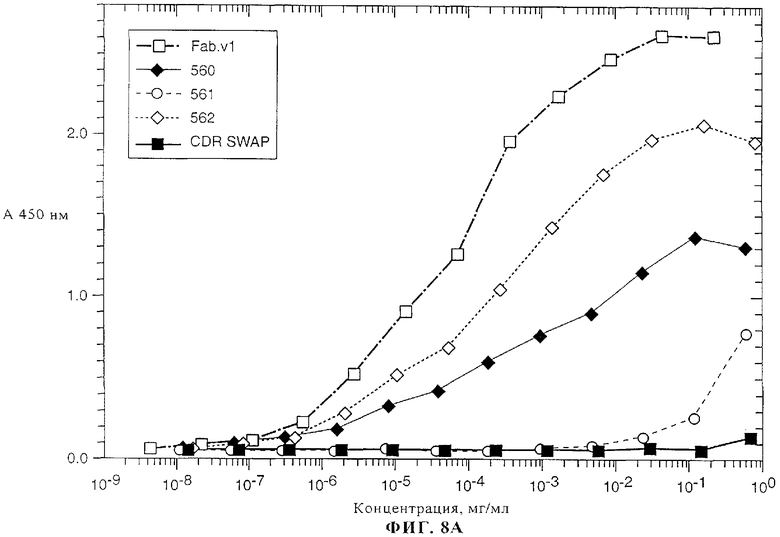
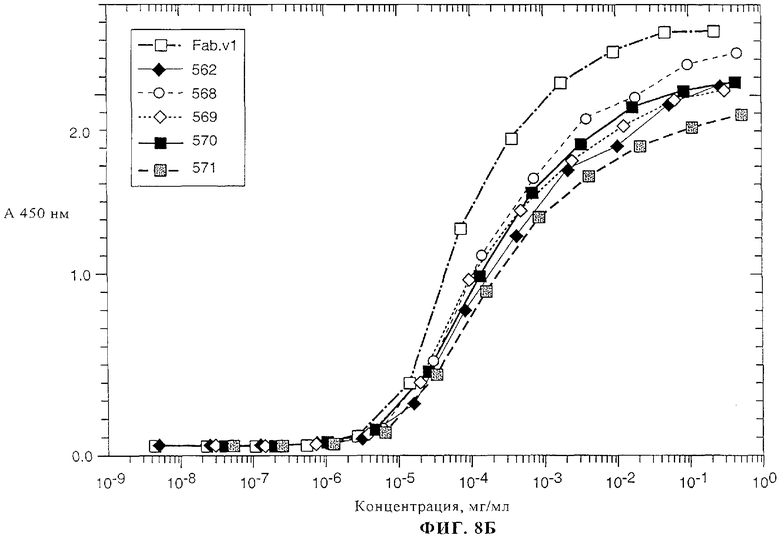
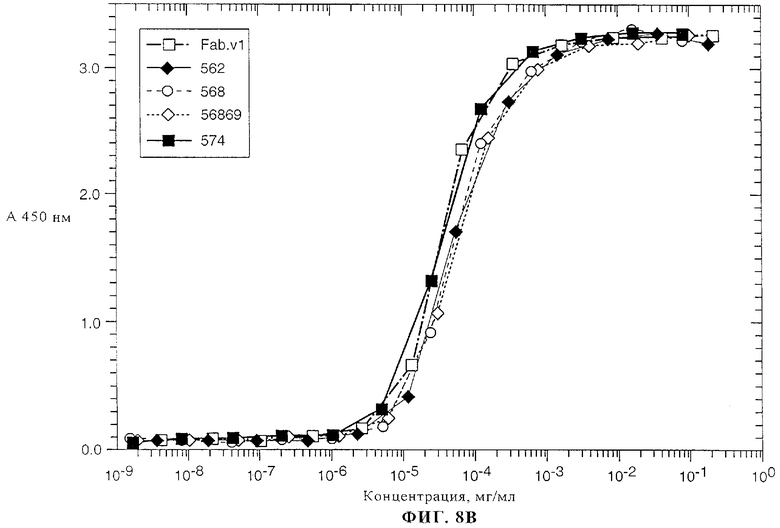
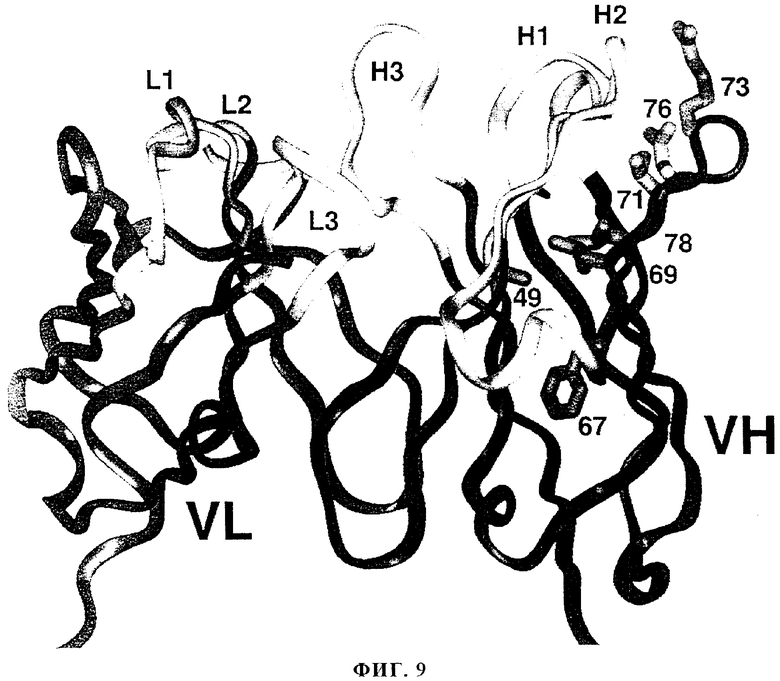
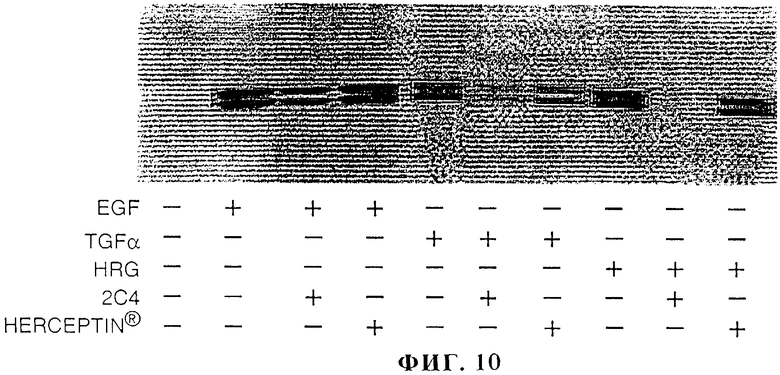
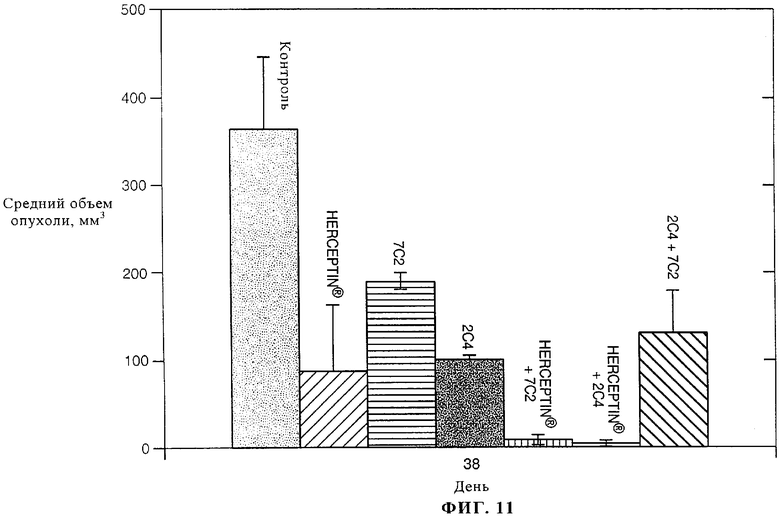

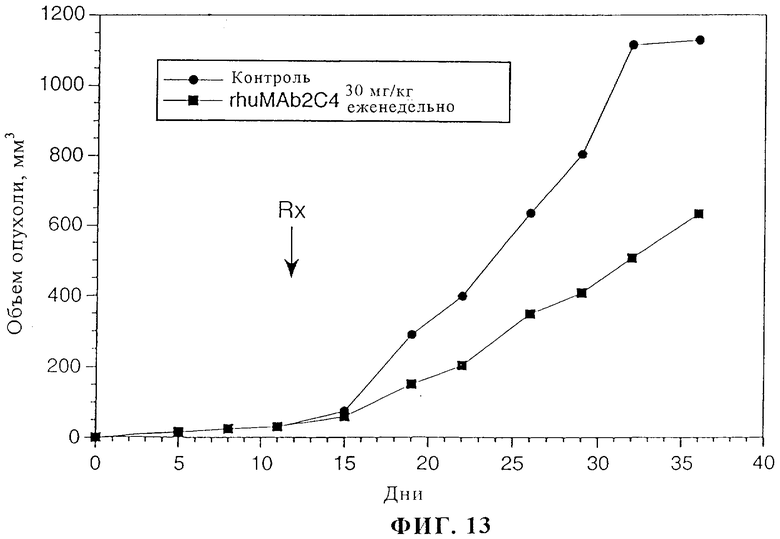
Изобретение относится к области медицины и касается гуманизированных антител с ЕrbВ2. Сущность изобретения заключается в разработке новых гуманизированных антител к рецепторам тирозиназ семейства ЕrbВ2, а также композиции, включающей эти антитела. Преимущество изобретения заключается в расширении области применения указанных антител для лечения форм рака, мишенью которых является рецептор эпидермального фактора роста, EGFR. 3 н. и 11 з.п. ф-лы, 3 табл., 21 ил.
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| US 5677171, 14.10.1997 | |||
| Способ размножения копий рисунков, текста и т.п. | 1921 |
|
SU89A1 |

