Область техники
Данное изобретение относится к применению кардиотрофина (СТ-1) для стимуляции регенерации печени и защиты гепатоцитов от апоптоза и некроза. В частности, данное изобретение относится к применению кардиотрофина для лечения острого, подострого, быстро развивающегося и хронического гепатита и циррозов печени, для стимуляции регенерации печени после резекции и трансплантации печени, а также для стимуляции пролиферации и трофики гепатоцитов или предшественников гепатоцитов в культуре.
Предпосылки изобретения
Печень человека и животных обладает уникальной способностью регулировать свой рост и массу. При разрушении вредным веществом части паренхимы печени выжившие гепатоциты способны реплицировать и заменять поврежденную паренхиму. Если при резекции печени или поражении гепатоцитов вирусного, токсического, иммунологического или метаболического характера затрагивается очень большая часть паренхимы, превышающая регенеративную способность оставшейся печеночной ткани, возникает печеночная недостаточность, которая может привести к летальному исходу. В настоящее время отсутствуют лекарственные средства, оказывающие на печень защитное и регенеративно-стимулирующее действие, которые можно использовать при острой или хронической печеночной недостаточности. Поэтому совершенно необходимо расширить спектр лекарственных средств, используемых в гепатологии, лечебными препаратами, предназначенными для указанных целей. Гепатозащитным средством является продукт или активное вещество, способное защитить гепатоциты от разных факторов, оказывающих токсическое действие и/или вызывающих повреждение гепатоцитов и в конечном счете некроз или апоптоз. Таким образом, при любых поражениях печени введение гепатозащитных средств в правильных дозах должно повысить выживаемость гепатоцитов и как следствие этого облегчить регенерацию печени, способствовать нормализации функции печени и в экстремальных случаях сохранить жизнь субъекту. Поражение печени может быть вызвано токсическими веществами (включая алкоголь), вирусами, аутоиммунными нарушениями, ишемией, ишемией/реперфузией (как это имеет место в случае поражения, возникающего в имплантированной печени в процессе трансплантации) и любыми воспалительными процессами. Хорошее гепатозащитное средство должно предотвращать или ослаблять развитие поражения печени и гибель клеток печени в указанных ситуациях.
Под регенерацией печени авторы настоящего изобретения понимают реакцию печени, компенсирующую уменьшение ее функциональной массы (уменьшение ткани или утрату клеток) путем пролиферации нормальных гепатоцитов до восстановления массы печени. Существует несколько клинических случаев, когда регенерация печени играет важную роль, включая резекцию печени при хирургическом вмешательстве (резекция печени или трансплантация печени от живых доноров) или вышеуказанные поражения печени (токсическими веществами, вирусами, ишемией, ишемией/реперфузией и т.д.). Средством, стимулирующим регенерацию печени, является такое средство, которое способно индуцировать пролиферацию гепатоцитов, помогающую сократить смертность, связанную с уменьшением функциональной массы печени.
Настоящее изобретение относится к применению кардиотрофина при заболеваниях печени.
Кардиотрофин (именуемый также CHF или фактором гипертрофии сердца) ранее использовали для лечения сердечных нарушений, нейродегенеративных и неврологических заболеваний (WO 95/29237) в качестве модулятора локальных воспалительных процессов, обусловленных LIFRβ-рецептором (WO 97/30146), для диагностики и лечения опухолей (WO 00/43790) и для лечения латерального амиотрофического склероза и болезни Паркинсона (WO 97/39629).
Настоящее изобретение не имеет отношения ни к одному из вышеуказанных применений, так как его объектом является применение СТ-1 в лечебных композициях, предназначенных для воздействия на гепатоциты, в частности, в качестве средства, защищающего указанные клетки от апоптоза и некроза, и в качестве средства, стимулирующего общую регенерацию печени.
СТ-1 является одним из так называемых нейропоэтических цитокинов, относящихся к семейству IL-6 [1]. Рецепторы цитокинов указанного семейства состоят из отдельных субъединиц, но все они имеют субъединицу gp130 [2]. Некоторые члены данного семейства (IL-6 и IL-11) вызывают гомодимеризацию gp130 [3], в то время как другие члены, такие как фактор ингибирования лейкоза (LIF), онкостатин и цилиарный нейротрофический фактор (CNTF), вызывают гетеродимеризацию субъединицы gp130 с рецептором LIF размером 190 кДа [4]. Рецептор СТ-1 содержит цепь gp130, β-субъединицу рецептора LIF (LIFRβ) и третий элемент, известный как α-субъединица рецептора СТ-1 [5, 6]. Последний элемент участвует в образовании трехкомпонентного комплекса, который сообщает высокую чувствительность и специфичность СТ-1. Активация рецептора СТ-1 индуцирует ряд внутриклеточных сигналов, которые включают раннюю активацию тирозинкиназ семейства JAK (JAK-1, JAK-2 и Tyk2). Главными эффекторами JAK является группа цитозольных факторов транскрипции STAT (STAT-1 и STAT-3; трансдуцирующие сигнал активаторы транскрипции). Активация JAK также вызывает передачу сигнала по пути, опосредуемому Ras-MAP-киназой, и активирует путь передачи сигналов, опосредуемый PI3-K (фосфатидилинозитол-3-киназа) [2].
СТ-1 первоначально был идентифицирован как гипертрофический фактор кардиомиоцитов [7, 8], так как было установлено, что указанный фактор стимулирует развитие кардиомиоцитов в эмбрионе и защищает кардиомиоциты от апоптоза, вызываемого гипоксией, ишемией, поражением вследствие ишемии-реперфузии и другими поражениями [8, 9, 10, 11, 12]. В научной литературе описано защитное действие, оказываемое СТ-1 на миокард в случае сердечной недостаточности [10]. Кроме того, СТ-1 стимулирует выживание двигательных и допаминэргических нейронов [13, 14].
Библиографические ссылки















Описание изобретения
В данном описании изобретения использована нижеследующая терминология, соответствующая целям настоящего изобретения:
i) активная фракция СТ-1 означает любую неполную полипептидную последовательность СТ-1, которая сохраняет физиологическое действие нативного белка, являющегося объектом настоящего изобретения;
ii) полипептидное производное, обладающее активностью СТ-1, означает любую полипептидную последовательность, характеризующуюся гомологией с нативным СТ-1 более чем на 80% и сохраняющую физиологическое действие нативного белка, являющегося объектом настоящего изобретения;
iii) полинуклеотидные последовательности, кодирующие указанные активные неполные последовательности СТ-1 или полипептидные производные СТ-1, описанные в пунктах i) и ii), также входят в объем настоящего изобретения;
iv) кардиотрофин-1 или СТ-1 означает нативную форму белка, любую форму рекомбинантного белка (используемого отдельно или в препаратах пролонгированного действия), любую полинуклеотидную форму, кодирующую или экспрессирующую нативный белок СТ-1, или удлиняющий сегмент любой из форм, описанных в пунктах i), ii) и iii).
В основе настоящего изобретения лежит открытие того, что ген СТ-1 сверхэкспрессируется в процессе регенерации печени после хирургической резекции части паренхимы печени, достигая максимальной экспрессии через 48 часов после резекции, что совпадает с моментом максимальной пролиферации гепатоцитов. На основании данного открытия было исследовано влияние СТ-1 на процесс регенерации печени, при этом было обнаружено, что трансдукция паренхимы печени генными последовательностями, кодирующими СТ-1, существенно стимулирует регенерацию печени после частичной гепатэктомии и предотвращает гибель животного после почти полной гепатэктомии. Аналогичным образом, было продемонстрировано, что трансдукция печени последовательностями, кодирующими СТ-1, обеспечивает эффективную защиту гепатоцитов от разных гепатотоксических веществ, существенно уменьшая апоптоз/некроз гепатоцитов. И наконец, полученные результаты показывают, что СТ-1 является сильнодействующим защитным средством для гепатоцитов от воздействия веществ, вызывающих гибель клеток и, кроме того, обладает свойством стимуляции процессов регенерации печени.
Таким образом, настоящее изобретение относится к применению СТ-1, активной фракции СТ-1, полипептидного производного, обладающего активностью СТ-1, полинуклеотидной последовательности, кодирующей и экспрессирующей СТ-1, активную фракцию СТ-1 или полипептидное производное, обладающее активностью СТ-1, для получения композиций, которые можно использовать для стимуляции регенерации печени после хирургической резекции печени или после поражения печени, вызванного химическими веществами, биологическими веществами, воспалительными или иммунологическими факторами, а также в качестве гепатозащитного средства при разных формах острого, подострого, быстро развивающегося и хронического гепатита, имеющего токсическую, вирусную, иммунологическую или метаболическую этиологию, и для стимуляции регенерации печени, защиты гепатоцитов и улучшения функции печени в случае циррозов печени, имеющих алкогольную, вирусную, метаболическую или иммунологическую этиологию, и при трансплантации печени.
Примеры
1. Аденовирусный вектор, содержащий генную последовательность, кодирующую СТ-1 (AdCT-1)
Дефектный аденовирус (с делецией в положениях Е1 и Е3), содержащий ген кардиотрофина-1 (AdCT-1), был создан способом, подробно описанным ниже. кДНК СТ-1 мыши была выбрана из библиотеки мышечной кДНК мыши при помощи зонда PCR, соответствующего нуклеотидам 20-639 последовательности кДНК мыши (№ доступа U18366 в банке генов GenBank). Полученную кДНК клонировали в векторе pGEM-T/CT-1 и правильность клонирования проверяли секвенированием. Затем кДНК СТ-1 клонировали в векторе pKS с образованием плазмиды pKS-CT-1, содержащей кассету экспрессии, состоящую из промотора вируса саркомы Рауса (нуклеотиды RSV 4526-5108 под № доступа М83237 в банке генов GenBank), пептидного сигнала фактора роста нервов (нуклеотиды NGF 298-378 под № доступа V00836 в банке генов GenBank), кДНК СТ-1 мыши (нуклеотиды 20-639 под № доступа U18366 в банке генов GenBank) и сигнала полиаденилирования SV40 (нуклеотиды 2546-2775 под № доступа NC0016691 в банке генов GenBank). Кассету экспрессии удаляли из плазмиды pKS-CT-1 при помощи BamHI/SalI и лигировали с шаттл-плазмидой аденовируса pGY63 на сайте HinfI с образованием плазмиды pGY63-CT-1. Полученная плазмида pGY63-CT-1 содержит левосторонний инвертированный концевой повтор аденовируса (ITR), сигнал упаковки (ps), часть гена pIX и кассету экспрессии СТ-1, расположенную между двумя последними элементами. Плазмиду pGY63-CT-1 котрансформировали в электрокомпетентных клетках SF800 E.coli вместе с плазмидой pXL2689, содержащей геном аденовируса для гомологичной рекомбинации. Правильные рекомбинанты расщепляли с помощью PacI и трансфицировали в клетки 293 (почечные клетки человеческого эмбриона, трансформированные с помощью ДНК аденовируса 5, ссылочный номер АТСС CRL-1573) для продуцирования аденовирусов. Структура AdCT-1 представлена на фиг.1. Трансформированные штаммы E.coli были включены в качестве депозита в Испанскую коллекцию типовых культур (СЕСТ) 12 сентября 2001 г. (E.Coli PKSCT1, CECT № 5980) в Университете Валенсии (Burjasot, Valencia, Spain).
Для продуцирования аденовирусов использовали клетки 293, инфицированные супернатантом, содержащим рекомбинантный аденовирус. Клетки 293 сначала высевали на 6-луночные планшеты при слиянии, равном примерно 80%, используя 2% среду DMEM. Через несколько часов культуральную среду удаляли и клетки инфицировали 0,5 мкл супернатанта, содержащего рекомбинантный аденовирус, разведенный в 3 мл DEMEM. Клетки инкубировали в течение 1 часа при 37°С, после чего инокулят удаляли и добавляли 4 мл агара. Клетки культивировали в течение 5-7 дней при 37°С. При помощи пастеровской пипетки образец вируса собирали из вирусной бляшки, образовавшейся в монослое клеток; агаровый цилиндр вновь суспендировали в 500 мкл DMEM с 2% фетальной телячьей сыворотки и хранили при -80°С. Для идентификации рекомбинантного аденовируса клетки 293 высевали на 12-луночные планшеты и инфицировали ранее выделенным вирусом в объеме 250 мкл. После обнаружения цитопатического действия клетки собирали отдельно из каждой лунки. Затем клетки трижды замораживали и оттаивали с целью их разрушения и высвобождения максимального количества вирусных частиц. Клеточный лизат из каждой серии центрифугировали со скоростью 1500 оборотов/мин в течение 10 минут. Супернатант, содержащий вирус, использовали для еще одного инфицирования клеток 293, культивируемых на 6-луночных планшетах. Как только клетки начинали приобретать округлую форму, супернатант собирали и проверяли наличие вируса путем обнаружения вирусных ДНК и РНК в указанном супернатанте. Супернатант с высокими уровнями экспрессии вируса отбирали для амплификации с целью создания рекомбинантных аденовирусов.
Клетки 293 культивировали в 150 мм чашках (от 50 до 100 чашек) и инфицировали аденовирусом из полученного запаса при множественности заражения, равной 10 (10 бляшкообразующих единиц, БОЕ/клетку). Клетки, проявляющие цитопатическое действие, собирали и центрифугировали со скоростью 1500 оборотов в минуту в течение 10 минут, вновь суспендировали в 0,1 М Трис-буфере (рН 8) и замораживали при -80°С до последующей очистки.
Рекомбинантный аденовирус очищали градиентами хлорида цезия. Для этой цели клетки, хранившиеся при -80°С, вновь суспендировали в 0,01 М Трис-буфере и обрабатывали 5% дезоксихолатом натрия в отношении 1:10 (об/об) в течение 30 минут. Затем клетки разрушали в предварительно охлажденном ручном стеклянном гомогенизаторе до получения полужидкого раствора. После этого клеточный экстракт добавляли к насыщенному раствору хлорида цезия в отношении 5,8 мл раствора хлорида цезия на 10 мл клеточного экстракта. Данную смесь получали в специальных запаянных полигаломерных пробирках (Quick-seal, Beckman Instruments, CA, USA). Центрифугирование производили в роторе с фиксированным углом наклона Beckman 50 Ti со скоростью 35000 оборотов/мин в течение 16-20 часов при 4°С. Полосу, соответствующую вирусу, собирали шприцем со стерильной иглой и еще раз центрифугировали в аналогичных условиях. Полосу экстрагировали и подвергали диализу против 0,01 М Трис-буфера, рН 8, при 4°С, в виде двух независимых процессов продолжительностью по 1,5 часа каждый. Аликвоты вирусного препарата помещали в пробирки со стерильным глицерином (ICN, USA) в количестве 10% (об/об), замораживали и хранили в жидком азоте до последующего использования.
Для определения инфекционного титра очищенных рекомбинантных аденовирусов выполняли испытание на предельное разведение на 96-луночных планшетах. В основе данного испытания лежит исследование цитопатического действия, оказываемого вирусом на клетки 293, для определения максимального десятичного разведения суспензии вируса, при котором вирус способен инфицировать клетки 293 и размножаться в указанных клетках. Клетки 293 предварительно высевали на 96-луночные планшеты в количестве 104 клеток/лунку. Затем из лунок удаляли среду и клетки инфицировали аденовирусом в объеме 50 мкл/лунку в виде последовательных дублируемых разведений. Через 6 часов добавляли 150 мкл свежей среды DMEM и инкубировали клетки при 37°С в течение максимального периода, равного 7 дням. По истечении указанного периода оценивали цитопатическое действие вируса на клетки. Для определения титра число клеток, испытывающих цитопатическое действие, умножали на максимальное разведение, вызывающее подобное действие, и полученный результат делили на общий измеренный объем (0,05 мл), что позволило установить число бляшкообразующих единиц (БОЕ) на мл. Указанное определение повторяли для каждого образца по крайней мере три раза.
2. СТ-1, рекомбинантный белок
кДНК, кодирующую СТ-1, получали из плазмиды pGEM-T/CT-1 расщеплением с помощью EcoR1 и клонировали в векторе pET28b (Novagen) (pET28b/CT-1). Данный вектор имеет последовательность, которая кодирует ряд остатков гистидина (1 кДа) и транслируется в соответствии с клонированной кДНК, продуцируя слитый белок, содержащий у своего аминоконца концевой гистидиновый участок длиной 1 кДа и СТ-1, между которыми находится сайт разрезания тромбином.
Для получения указанного белка авторы настоящего изобретения использовали компетентные бактерии штамма BL21 (DE3) (Novagen, Germany, № по каталогу 70235), так как данный штамм содержит ген, индуцируемый РНК-полимеразой Т7, что является необходимым требованием для последующего продуцирования указанного белка. Компетентные бактерии трансформировали ранее полученным вектором: pET14b (вектор pET-14b фирмы Novagen, № по каталогу 69660-3) с кДНК клонированного СТ-1. Трансформированные бактерии отбирали для выращивания в среде LB с ампициллином, так как данный вектор содержит ген устойчивости к указанному антибиотику.
Для получения рекомбинантного СТ-1 трансформированные бактерии выращивали в среде LB с ампициллином при 37°С до достижения оптической плотности, равной 0,4, при 600 нм. Экспрессию рекомбинантного белка индуцировали IPTG, добавляемым до конечной концентрации, равной 0,5 мМ. В результате такой обработки происходит индукция lac-промотора и затем промотора РНК-полимеразы Т7, которая содержит вектор и регулирует экспрессию клонированной кДНК. Культуру выращивали еще 4 часа в аналогичных условиях.
Для получения экстрактов сразу же после выращивания бактерии их подвергали центрифугированию при 4°С. Осажденные бактерии снова суспендировали в буфере, содержащем 10 мМ Трис/HCl, 10% сахарозы, 2 мМ 2-меркаптоэтанола и ингибиторы протеазы. Бактерии инкубировали с лизоцимом при 4°С в течение 30 минут и гомогенизировали ультразвуком. В результате подобной обработки происходило разрушение оболочки бактерии, что улучшало выход экстрагированного продукта. Цитозольный экстракт получали центрифугированием гомогената при 100000 g в течение 90 минут. Полученный белок проверяли, анализируя цитозольную фракцию методом SDS-PAGE.
Слитый белок His-CT-1 очищали хроматографией цитозольного экстракта в 2 мл никелевой колонке. Затем колонку промывали и белок элюировали 1 М имидазолом. Чистый белок обрабатывали тромбином и выделяли СТ-1.
3. Анализы методом нозерн-блоттинга для измерения экспрессии СТ-1 in vivo
Экспрессию генов разных цитокинов (фактор роста гепатоцитов, HGF; LIF; онкостатин; CNTF; CT-1) анализировали в процессе регенерации печени методом нозерн-блоттинга после экстракции мРНК из печени крыс. РНК экстрагировали методом с использованием тиоционат гуанидиния - фенол - хлороформ. Анализ методом нозерн-блоттинга выполняли в соответствии с приведенным ранее описанием [15], используя экспрессию 28S в качестве эталона и зонды, специфические для каждого анализируемого гена.
4. Культуры линий клеток, выделенных из гепатоцитов
Для исследований in vitro были использованы клетки Н35 линии гепатоцитов, выделенных из злокачественной гепатомы крыс. Клетки культивировали в модифицированной по Дульбекко среде Игла (DMEM), содержащей 10% телячьей сыворотки, 2 мМ глутамина, 100 ЕД/мл стрептомицина, 100 мг/мл пенициллина. Культуры клеток инкубировали при 37°С в атмосфере с 5% СО2.
5. Методы анализа апоптоза на основании клеточного цикла и экспрессии аннексина
Клеточный цикл анализировали методом окрашивания ДНК иодидом пропидия. Клетки (0,5·106) делали проницаемыми при помощи 50 мкл 0,1% раствора NP40 до окрашивания 0,5 мкл 50 мкг/мл раствора иодида пропидия и 4 КЕД/мл ДНКазы (набор реагентов DNA-Prep Coulter, Coulter). Клетки инкубировали при 37°С в течение 20 минут и исследовали в цитофлуорометре FACScalibur. Клетки, положительно окрашиваемые иодидом пропидия, анализировали в "дублет-распознающем модуле" (DDM) проточного цитометра (FACScalibur, Becton-Dickinson, USA), исключающем дублеты и использующем параметр FL3. Частота субдиплоидных клеток определяет процентное число клеток, подвергающихся апоптозу.
Присутствие фосфатидилсеринов, ориентированных в направлении наружной оболочки клетки, является одним из параметров, позволяющих определить апоптозную клетку. Аннексин V выявляет апоптозные клетки благодаря своей способности связываться с молекулами фосфатидилсерина, обращенными к наружной стороне клеточной мембраны в тот момент, когда клетка подвергается апоптозу. Клетки (0,5·106) один раз промывали в буфере для инкубации, содержащем: 140 мМ NaCl, 5 мМ KCl, 1,2 мМ MgCl2, CaCl2 и HEPES 10 мМ. Клетки инкубировали в 100 мкл буфера для инкубации и 5 мкл флуоресцеин-изотиоцианатного конъюгата, связанного с аннексином V (Annexin-FITC) в течение 15 минут при комнатной температуре. Затем клетки исследовали в проточном цитометре FACScalibur, используя параметр FL1. Индекс апоптоза определяли на основании процентного содержания клеток с положительной реакцией на аннексин-FITC.
6. Методы анализа белков
Электрофорез. Для анализа белков клетки лизировали в буфере для лизиса (20 мМ Трис, рН 7,5; 150 мМ NaCl, 1 мМ EGTA, 1 мМ EDTA, 1% Triton Х-100, 2,5 мМ пирофосфата натрия, 1 мМ Na3VO4, 1 мкг/мл лейпептида, пепстатина, 10 мкг/мл ингибитора трипсина, 1 мМ PMSF. Лизат 0,5·106 клеток снова суспендировали в объемном отношении в буфере для миграции (125 мМ Трис-HCl (рН 6,8), 10% додецилсульфата натрия, 20% глицерина, 100 мМ дитиотреитола, 0,2% бромфенолового синего). Образцы белкового экстракта нагревали при 100°С в течение 5 минут и подвергали электрофорезу в 10% полиакриламидном геле.
Иммунологический анализ методом вестерн-блоттинга. После электрофореза белки переносили на нитроцеллюлозные мембраны в буфере для переноса (25 мМ Трис, 0,2 М глицина, 20% метанола, рН 8,5) под воздействием тока, равного 300 мА, в течение 1 часа. Перенесенные белки окрашивали раствором пунцового красного для проверки успешного переноса. Затем мембраны использовали для иммунологического анализа специфических белков. Для этой цели мембраны блокировали в буфере для инкубации TBS-T (20 мМ Трис, 137 мМ NaCl при рН 7,6 и 0,5% Твин-20) с 2% BSA (фракция альбумина V) в течение 1 часа. Мембраны инкубировали со специфическими антителами против исследуемого белка в течение 2 часов. Затем мембраны промывали буфером TBS-T в течение 1 часа и снова инкубировали с белком G-HRPO (BIORAD) в течение 1 часа. Мембраны несколько раз промывали в буфере TBS-T, проявляли хемолюминесцентными реагентами (NEN Life Science Products) и сразу же экспонировали на сверхчувствительную пленку (Amersham) в заранее определенные периоды времени.
Иммунопреципитация. Для иммунопреципитации специфических белков лизаты 106 клеток инкубировали в течение 18 часов в присутствии специфического антитела и 20 мкл белка G-Sepharose при 4°С. Иммунокомплексы выделяли центрифугированием, дважды промывали буфером для лизиса и растворяли в буфере для миграции. Затем образцы нагревали до 100°С и вызывали миграцию электрофорезом в 10% гелях. Иммунологический анализ специфических белков выполняли методом вестерн-блоттинга.
7. Определение синтеза ДНК. Анализы пролиферации
Клетки Н-35 высевали на 96-луночные планшеты. Клетки культивировали без сыворотки в течение 24 часов и затем стимулировали СТ-1 (50 нг/мл), разведенным в бессывороточной среде DMEM. Клетки инкубировали с СТ-1 в течение 24 часов и метили 10 мкКи/мл [метил-3Н]тимидина (ICN, Amersham) в течение 12 часов. Радиоактивную среду удаляли, клетки отделяли при помощи 100 мкл трипсина при 37°С и собирали в 25 мкл смеси для сцинтилляционного счета (Ecolite; ICN). Содержание [3H]тимидина анализировали в сцинтилляционном счетчике tri Carb 2900TR (Packard, Meriden, CT).
8. Испытания регенерации печени in vivo после частичной гепатэктомии (хирургическая резекция 75% печени)
Регенерацию печени исследовали с использованием крыс Фишера (самцы с массой тела 180 г). В результате хирургической резекции удаляли 75% печени и умерщвляли крыс в разные периоды времени (1 час, 3 часа, 6 часов, 10 часов, 24 часа, 48 часов, 3 дня, 6 дней и 9 дней). Затем отбирали образцы печени и делили их на три части для гистологического исследования (фиксация в формалине), иммуногистохимического исследования (фиксация в ОСТ) и анализа РНК (замораживание в жидком азоте). Для каждого анализируемого периода времени использовали как минимум 4 крыс. Параметры анализируемой регенерации печени выражали в процентном значении массы печени и экспрессии ядерного антигена пролиферации клеток (PCNA), определяемой иммуногистохимическим методом.
8.1. Экспрессия гена СТ-1 в процессе регенерации печени
Экспрессию генов разных цитокинов (HGF, LIF, онкостатин, CNTF, CT-1) анализировали при помощи модели частичной гепатэктомии с целью исследования их участия в регенерации печени. В данном исследовании авторы настоящего изобретения анализировали образцы печени крыс, полученные в разное время после частичной гепатэктомии (1 час, 3 часа, 6 часов, 10 часов, 24 часа, 48 часов, 3 дня, 6 дней и 9 дней). Каждая группа включала как минимум 4 животных. Помимо этого, анализировали печень здоровых крыс, не подвергавшихся гепатэктомии (контрольные животные). Соответствующие уровни мРНК каждого цитокина определяли методом нозерн-блоттинга. В результате выполнения указанных экспериментов было сделано совершенно новое наблюдение о существенном увеличении уровней мРНК СТ-1 через 24 и 48 часов после гепатэктомии (фиг.2 и 3), соответствующем максимальной пролиферации гепатоцитов, о чем свидетельствует экспрессия PCNA и поглощение бромдезоксиуридина (BrdU) гепатоцитами при иммуногистохимическом исследовании образцов печеночной ткани у подопытных животных. Кроме того, авторы настоящего изобретения обнаружили, что увеличению транскрипционной экспрессии СТ-1 предшествовала максимальная экспрессия HGF, произошедшая через 10 часов после гепатэктомии.
8.2. Влияние СТ-1 на регенерацию печени после частичной гепатэктомии
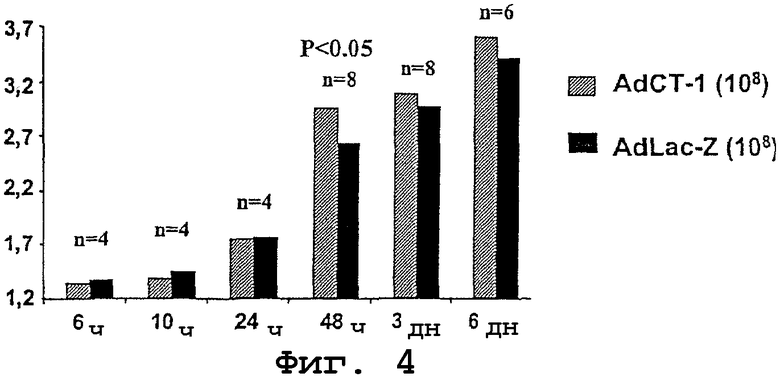
Для исследования роли СТ-1 в регенерации печени животным внутривенно инъецировали аденовирус с СТ-1 (AdCT-1) в дозе 108 БОЕ или аденовирус с репортерным геном LacZ (AdLac-Z) в аналогичной дозе в качестве контрольного образца. Через 48 часов производили хирургическую резекцию 75% печени. Затем крыс умерщвляли в вышеуказанные периоды времени. Для каждого периода времени анализировали как минимум 4 крысы и максимум 8 крыс.
Введение AdCT-1 вызывало увеличение массы печени у крыс, получавших AdCT-1, по сравнению с крысами, которым вводили AdLac-Z, причем между двумя указанными группами наблюдалось существенное различие через 48 часов, то есть во время максимальной пролиферации гепатоцитов (о чем свидетельствуют результаты иммунного окрашивания на PCNA в образцах печени, полученных у крыс после гепатэктомии). Через 3 и 6 дней после гепатэктомии масса печени у крыс, получавших AdCT-1, была больше, чем у контрольных крыс, хотя в указанные периоды времени разница между группами не была статистически значимой (фиг.4). Полученные результаты показывают, что печени, на которые воздействовали СТ-1, характеризовались более быстрой регенерацией и имели большую массу по сравнению с контрольными животными в начальный период после гепатэктомии, но в конечном счете достигали таких же показателей, что и печени контрольных животных под действием гомеостатических механизмов, регулирующих конечный размер печени.
9. Анализы регенерации печени in vivo после обширной гепатэктомии (хирургическая резекция >85% печени)
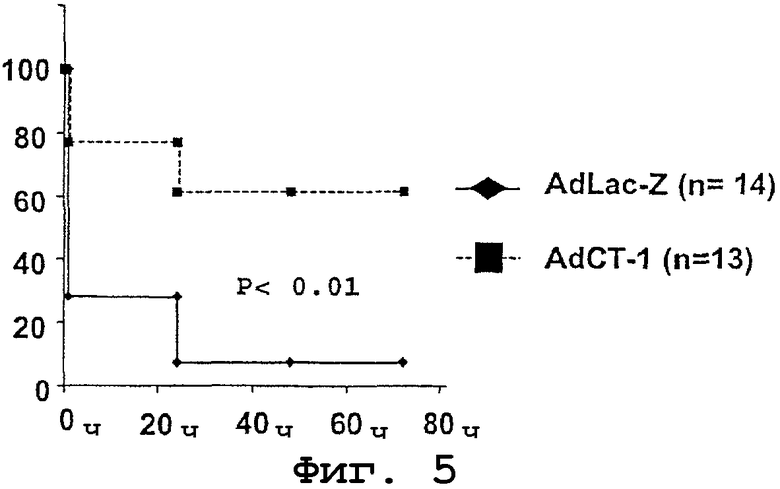
Чтобы определить, способен ли СТ-1 предотвратить гибель животных, подвергнутых почти полной гепатэктомии, выполняли эксперименты с использованием крыс Фишера, у которых в результате хирургической резекции удаляли более 85% печени. В данных экспериментах использовали две группы по 30 крыс в каждой. Одной группе внутривенно вводили AdLac-Z и другой группе вводили AdCT-1 в указанных выше дозах. Хирургическую резекцию подобного типа производили через 48 часов после введения аденовируса. Число крыс, выживших после произведенной хирургической резекции, составило 14 крыс в группе, получавшей AdLac-Z, и 13 крыс в группе, которой инъецировали AdCT-1. У крыс контролировали продолжительность выживания после обширной хирургической резекции.
Было установлено, что в первый час после гепакэктомии смерность составила 77% в группе, получавшей AdLac-Z, и не достигла 20% в группе, получавшей AdCT-1. Через двадцать четыре часа после гепатэктомии были живы только 7% крыс, получавших AdLac-Z, в то время как выживаемость среди крыс, получавших AdCT-1, составила 61%; указанная разница находится на статистически значимом уровне. Вышеуказанные процентные значения сохранялись на тех же уровнях через 4 дня после хирургической резекции (фиг.5). Данные, полученные авторами настоящего изобретения, показывают, что СТ-1 предотвращает смертность, обусловленную обширной резекцией печени.
10. Защитное действие СТ-1 против апоптоза/некроза гепатоцитов in vivo. Испытания для быстро развивающегося поражения печени
Для оценки роли СТ-1 в модуляции поражения печени, вызванного разными вредными веществами, использовали мышей Balb/c (самцы с массой тела 30 г), при этом поражение печени оценивали в трех моделях поражения гепатоцитов: i) поражение, вызванное внутривенным введением 100 мг/кг конканавалина А, Con-A (Sigma, St. Louis, MO., USA); ii) поражение, вызванное комбинацией внутривенного введения TNFα (Peprotech) (0,5 мкг/мышь) и внутрибрюшинного введения 25 мг D-галактозамина, TNFα/D-Gal (Sigma); iii) поражение, вызванное внутривенным введением 1,5 мкг/мышь антитела против Fas (Jo2. Pharmingen). Через шесть часов после введения Con-A, TNFα/D-Gal или антитела против Fas у мышей брали пробы крови, после чего их умерщвляли.
Чтобы определить влияние СТ-1 на поражение печени, группе мышей "А" вводили физиологический раствор, группе "B" вводили AdLac-Z (107 БОЕ) и группе "C" вводили AdCT-1 (107 БОЕ). Через 48 часов в каждой группе трех вышеописанных моделей индуцировалось поражение печени. Группа мышей, которым вводили сыворотку в физиологическом растворе вместо индуктора гепатита, была также включена в данный эксперимент в качестве отрицательной контрольной группы (NC). Каждая группа животных включала 5 мышей. Через 6 часов исследовали степень поражения печени по 2 параметрам: измерение уровня трансаминаз (GPT) в сыворотке при помощи автоматического колориметрического анализа (Technicon RA-1000, Bayer) и измерение апоптоза методом TUNEL в образцах печени, фиксированных в ОСТ, с использованием "Набора для обнаружения гибели клеток in situ" (Roche Diagnostics GmbH, Indianapolis, IN, USA).
У всех мышей брали пробы крови для определения трансаминаз, после чего животных сразу же умерщвляли и производили обработку печени для гистологического исследования (фиксация в формалине) и исследования апоптоза методом TUNEL (замораживание в ОСТ).
В первой модели острого поражения печени, вызванного введением Con-A, было обнаружено, что в то время как у мышей в контрольных группах (животные, получавшие физиологический раствор или AdLac-Z) наблюдались очень высокие показатели GPT, у животных, получавших AdCT-1, уровни трансаминаз были изменены незначительно, и различия между двумя указанными группами были весьма существенными (фиг.6А). При исследовании печеночной ткани методом TUNEL наблюдалось отсутствие апоптоза в образцах печени мышей, получавших AdCT-1, по сравнению с обширными областями некроза и апоптоза у животных, которые получали сыворотку в физиологическом растворе или AdLac-Z до введения Con-A (фиг.7).
Во второй модели острого поражения печени, вызванного введением моноклонального антитела против Fas, было установлено, что введение AdCT-1 предотвращает гибель гепатоцитов (фиг.6В). Через шесть часов после введения антитела против Fas содержание трансаминазы у животных, получавших AdCT-1, было значительно ниже (при статистически значимой разнице), чем у животных, получавших сыворотку в физиологическом растворе или AdLac-Z. Кроме того, при помощи метода TUNEL и гистологического исследования было обнаружено существенное уменьшение апоптозных тел в образцах печеночной ткани мышей, получавших AdCT-1, по сравнению с животными в контрольных группах.
Гепатозащитную роль СТ-1 оценивали также в третьей модели поражения печени, вызванного комбинированным введением TNFальфа и D-галактозамина (TNF-α/DGal). По результатам гистологического анализа и метода TUNEL через шесть часов после поражения печени наблюдалось значительное уменьшение уровней трансаминаз и числа апоптозных гепатоцитов у мышей, получавших AdCT-1, по сравнению с мышами в контрольных группах (фиг.6С).
Все полученные данные показывают, что СТ-1 способен защищать клетки печени от разных факторов, вызывающих апоптоз или некроз клеток печени.
11. Анализ влияния СТ-1 на клеточный цикл и выживание гепатоцитов, выделенных из линий клеток
Используя линию клеток Н35 печени крыс, авторы настоящего изобретения исследовали биологические действия, оказываемые рекомбинантным СТ-1 в качестве цитокинного регулятора апоптоза гепатоцитов. Для испытания стимуляции кардиотрофином СТ-1 клетки предварительно культивировали без сыворотки в течение 18 часов. Испытания стимуляции СТ-1 выполняли без сыворотки.
Авторы настоящего изобретения сначала анализировали влияние СТ-1 на клеточный цикл данной линии клеток печени. Клеточный цикл определяли путем окрашивания ДНК иодидом пропидия с последующим выполнением анализа методом проточной цитофлуорометрии. Апоптоз индуцировали удалением сыворотки из культуральной среды клеток в течение 4 дней. Полученные результаты показали, что в результате четырехдневного культивировании в указанных условиях 86% клеток Н35 подверглись апоптозу. Следует отметить, что при использовании СТ-1 в дозе 50 нг/мл и при отсутствии любых других дополнительных стимулов СТ-1 значительно задерживал начало апоптоза клеток Н35, который наблюдался примерно в 52% клеток (фиг.7).
Аналогичные эксперименты выполняли, культивируя без сыворотки культуры клеток Н35 в течение 3 дней и измеряя способность клеток связывать на своей поверхности очищенный аннексин, связанный с FITC (изотиоцианат флуоресцеина). Связывание аннексин-FITC с поверхностью клеток Н35 исследовали, выполняя анализ методом проточной цитофлуорометрии. Полученные результаты подтверждают, что клетки, культивируемые без СТ-1, характеризуются примерно 21% положительной реакцией на аннексин и клетки, обработанные 50 нг/мл СТ-1, характеризуются примерно 12% положительной реакцией (фиг.8). Таким образом, выполненные эксперименты показали, что СТ-1 в используемой дозе способен оказывать антиапоптозное действие.
12. Анализ влияния СТ-1 на пролиферацию клеток
Используя линию клеток Н-35, авторы настоящего изобретения исследовали способность СТ-1 влиять на синтез ДНК в гепатоцитах. Для этой цели 20000 клеток высевали в одну лунку 96-луночных планшетов. Чтобы обнаружить возможную стимуляцию, клетки предварительно культивировали без сыворотки в течение 24 часов. Испытания стимуляции СТ-1 выполняли без сыворотки, используя СТ-1 в дозе 50 нг/мл, в течение 24 часов. Полученные результаты показали, что клетки, культивируемые в присутствии СТ-1, характеризуются более высоким процентным значением синтеза ДНК по сравнению с контрольными клетками, которые не подвергали воздействию СТ-1 (фиг.9). Таким образом, данные эксперименты подтверждают, что СТ-1 в используемой дозе способен индуцировать синтез ДНК.
13. Исследование и анализ путей передачи сигналов, индуцируемых СТ-1, в линиях клеток, полученных из гепатоцитов
Открытие того, что СТ-1 оказывает антиапоптозное действие на гепатоциты как in vivo, так и in vitro, позволило авторам настоящего изобретения исследовать пути передачи сигналов, участвующие в стимуляции рецептора СТ-1 в гепатоцитах. Стимуляция рецепторов семейства цитокинов IL-6/LIF вызывает немедленное фосфорилирование трансмиттеров сигналов, относящихся к семейству JAK-1. После стимуляции клеток Н35 кардиотрофином СТ-1 в разные периоды времени авторы настоящего изобретения выполняли иммунопреципитацию JAK-1 специфическим антителом (Cell Signaling Technology), выделенным из полных лизатов указанных клеток. При использовании специфического антитела к фосфорилированным тирозинам (4G10, Upstate Biotechnology) и выполнения вестерн-блоттинга было установлено, что СТ-1 индуцирует фосфорилирование тирозином молекулы JAK-1 через 5 минут, при этом сигнал исчезает через 60 минут (фиг.10А).
Фосфорилирование STAT-3 является одним из описанных путей активации, участвующих в передаче сигнала цитокинов семейства IL-6 при помощи JAK. Активация указанной молекулы фосфорилированием в одних случаях обусловлена индукцией клеточной дифференцировки, а в других случаях гипертрофией (миокардиоциты). При помощи вестерн-блоттинга авторы настоящего изобретения анализировали лизаты Н35, обработанные in vitro 50 нг/мл СТ-1 в разные периоды времени. Использование антитела, специфического к фосфорилированному STAT-3 (Santa Cruz Biotechnology), позволило установить, что СТ-1 способен индуцировать фосфорилирование STAT-3, начиная с 5-й минуты после стимуляции и вызывая максимальную индукцию через 30 минут (фиг.10В).
Одним из путей, очевидно, определяющим ингибирование апоптозного сигнала, является путь PI-3/AKT (фосфатидинозитол-3-киназа/АКТ-киназа). Активация PI-3K индуцирует активацию фосфорилированием АКТ в серине 475 и треонине 308. Активация АКТ в свою очередь вызывает фосфорилирование BAD в серине в положениях 112 и 136. BAD является членом семейства Bcl-2 и представляет важный регулятор сигнала выживания. Неактивный BAD образует димеры с белками Bcl-x или Bcl-2, нейтрализуя их антиапоптозную активность. Фосфорилирование BAD вызывает высвобождение Bcl-2 или Bcl-x, которые подавляют путь активации апоптоза. Поэтому предполагается, что фосфорилирование BAD является способом подавления пути активации апоптоза. Авторы настоящего изобретения исследовали, может ли СТ-1 активировать данный путь выживания в клетках Н35. Клетки обрабатывали 50 нг/мл СТ-1 в разные периоды времени, получали цитозольную фракцию и выполняли иммунопреципитацию АКТ при помощи поликлонального антитела против АКТ (Cell Signaling Technology). Затем методом вестерн-блоттинга анализировали наличие фосфорилированного АКТ, используя поликлональное антитело, которое является специфическим к указанной форме АКТ, фосфорилированного в серине 475 (Cell Signaling Technology). Было установлено, что СТ-1 индуцирует фосфорилирование устойчивого АКТ в серине 475 через 15 и 30 минут и исчезает через 60 минут. Таким образом, СТ-1 индуцирует сигнал выживания в линии клеток гепатоцитов (фиг.10С).
Суммируя вышеизложенное, можно сделать вывод, что СТ-1 способен индуцировать путь передачи сигналов JAK/STAT, а также путь выживания PI-3K/AKT. Каскад сигналов, индуцируемых СТ-1 в гепатоцитах, объясняет, каким образом СТ-1 действует в качестве цитокина, оказывая антиапоптозное действие при помощи пути PI-3К/AKT, и, возможно, в качестве индуктора пролиферации и дифференцировки гепатитов при помощи пути JAK/STAT-3.
14. Исследование и анализ путей передачи сигналов, индуцируемых AdCT-1 в моделях острой печеночной недостаточности in vivo
Для анализа защитного действия AdCT-1, наблюдаемого в моделях in vivo острого поражения печени у крыс и мышей, были исследованы пути передачи сигналов, которые в соответствии с данными, полученными при исследовании in vitro, по-видимому, участвуют в стимуляции рецептора СТ-1 гепатитов, с использованием указанных моделей in vivo.
Как было описано выше при исследовании in vitro, СТ-1 может индуцировать активацию трех основных путей, определяющих выживание или подавление апоптоза: STAT-3 (трансдуктор сигналов и активатор транскрипции), PI-3K (фосфатидилинозитол-3-ОН-киназа)/АКТ и Erk1/2 (внеклеточные регулируемые киназы).
а. Модель обширной гепатэктомии у крыс
Как было показано в экспериментах, описанных в примере 9 (резекция печени >85%), самая высокая смертность наблюдалась через 1 час после хирургической резекции (см. фиг.5). По этой причине указанные эксперименты повторяли в 3-х группах животных, которым вводили разные препараты (AdCT-1, AdLac-Z и физиологический раствор), но при этом крыс умерщвляли через 1 час после хирургической резекции для получения образцов печени.
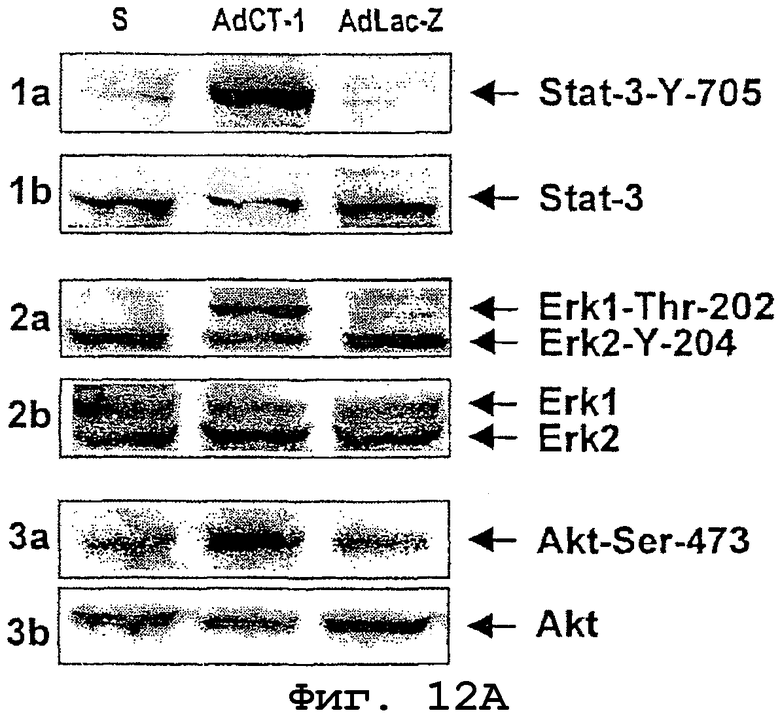
Собранные образцы печени делили на три части для гистологического исследования (фиксация в формалине), иммуногистохимического исследования (фиксация в ОСТ) и анализа белков (замораживание в жидком азоте). Из образцов, замороженных в жидком азоте, получали гомогенаты печени в буфере для лизиса (20 мМ Трис, рН 7,5; 150 мМ NaCl; 1 мМ EGTA; 1 мМ EDTA; 1% Triton Х-100; 2,5 мМ пирофосфата натрия; 1 мМ Na3VO4 и смесь антипротеаз). Методом вестерн-блоттинга с использованием специфических антител фирмы Cell Signaling Technology (Beverly, Mssachusetts) исследовали передачу сигналов в трех группах крыс: получавших AdCT-1, AdLac-Z и физиологический раствор (S). Использовали нижеследующие антитела: против Stat-3, против фосфорилированного Stat-3 (Stat-3-Y-705), против АКТ, против фосфорилированного АКТ (Akt-Ser-473), против Erk1/2 и против фосфорилированного Erk1/2, которые одновременно обнаруживают фосфорилированные формы Erk1 (Erk1-Thr-202) и Erk2 (Erk2-Y-204). Было установлено, что печени крыс, получавших AdCT-1, характеризуются фосфорилированием STAT-3, ERK1/2 и АКТ в отличие от печени крыс, которым вводили AdLac-Z и физиологический раствор, где данный эффект отсутствовал (фиг.12А).
С другой стороны, в возникновении апоптоза в ответ на многие стимулы, включая обширную гепатэктомию (>85%), участвует каспаза-3. По этой причине была исследована активность каспазы-3 с использованием части образцов, находящихся в жидком азоте, в соответствии с методикой испытания, предложенной компанией (CaspACE, Promega, Madison, Wisconsin). Было установлено, что печени крыс, которым ранее вводили AdCT-1, характеризуются меньшей активностью каспазы-3 по сравнению с печенью крыс, которым вводили AdLac-Z и физиологический раствор (фиг. 12В), что свидетельствует о более низком индексе апоптоза в первом случае.
Вышеизложенное позволяет предположить, что защитное действие, оказываемое AdCT-1 на поражение печени у крыс, обусловлено инициацией каскадов антиапоптозных сигналов, индуцируемых СТ-1, с последующим сокращением апоптоза, о чем свидетельствует обнаруженная низкая активность каспазы-3.
b. Модель острого поражения печени, вызванного конканавалином А, у мышей
Для исследования передачи сигналов в данной модели были повторно выполнены эксперименты по индукции, вызываемой Con-A, (см. пример 10) в 3-х экспериментальных группах (AdCT-1, AdLac-Z и физиологический раствор), но при этом мышей умерщвляли через 1 час после введения Con-A. Образцы печени, полученные в момент умерщвления животных, обрабатывали в соответствии с описанием, приведенным выше в примере 14а. Вестерн-блоттинг выполняли аналогичным образом и использовали такие же антитела.
Как показано на фиг.13, введение AdCT-1 вызывало фосфорилирование АКТ и ERK 1/2, главных путей подавления апоптоза и активации выживания, индуцируемых СТ-1. Поэтому полученные результаты позволяют предположить, что AdCT-1 защищает мышей от поражения печени, вызванного Con-A, активируя указанные главные пути подавления апоптоза.
Описание чертежей
Фиг.1. Схематическое изображение структуры аденовирусного вектора AdCT-1, содержащего последовательность, кодирующую СТ-1. RSV: промотор вируса саркомы Рауса; NGF: пептидный сигнал фактора роста нервов; СТ-1: кДНК СТ-1 мыши; SV40: сигнал полиаденилирования вируса SV40. Подавленные области Е1 и Е3 окрашены в черный цвет.
Фиг.2. Обнаружение методом нозерн-блоттинга мРНК кодирующей СТ-1 в образцах печени крыс, полученных в разное время (h = часы; d = дни) после частичной гепатэктомии. 28S: рРНК, используемая в качестве контрольного образца.
Фиг.3. Графическое изображение экспрессии СТ-1 в зависимости от времени (h = часы; d = дни) при выполнении нозерн-блоттинга, показанного на фиг.2. Ордината: произвольные единицы измерения оптической плотности (СТ-1/28S).
Фиг.4. Процентные значения массы печени крыс (на ординате) в разные периоды времени (абсцисса: h = часы; d = дни) после частичной гепатэктомии до введения AdCT-1 или AdLac-Z и после выполнения частичной гепатэктомии.
Фиг.5. Процентные значения выживания (на ординате) крыс, получавших AdCT-1 или AdLac-Z и подвергнутых гепатэктомии (>85%) через 48 часов после введения указанных препаратов. Абсцисса: время (часы) после гепатэктомии.
Фиг.6. Графическое изображение уровней трансаминаз GPT в сыворотке (на ординате, СОЕ/мл) и гистологические изображения печеночной ткани (метод TUNEL для визуализации апоптоза) из 3-х моделей индукции быстро развивающегося гепатипа у мышей: в результате введения конканавалина А, Con-A (фиг.6А); введения антитела против Fas (фиг.6В) и совместного введения TNFα и D-галактозамина, TNFα/D-Gal (фиг.6С). За 48 часов до индукции гепатита животным вводили аденовирусный вектор (AdCT-1 или AdLac-Z) или физиологический раствор (S). Отрицательный контрольный результат (NC) получен в группе мышей, которым вводили сыворотку в физиологическом растворе вместо индуцирующего гепатит вещества.
Фиг.7. Анализ клеточного цикла клеток Н-35 через 1 день (сверху) и 4 дня (снизу) культивирования без сыворотки при отсутствии СТ-1 (С = контрольный образец) и в присутствии СТ-1 (СТ-1). Выбранные зоны слева направо: клетки с ДНК менее, чем 2n (апоптозные клетки, Apo); клетки в фазе G0-G1 (покоящиеся клетки) и клетки в фазе S и М (пролиферирующие клетки). Ордината: число клеток. Абсцисса: содержание ДНК.
Фиг.8. Анализ экспрессии аннексина V в клетках Н-35 методом проточной цитометрии при культивировании без сыворотки в течение 3 дней при отсутствии СТ-1 (С = контрольный образец) и в присутствии СТ-1 (СТ-1). Клетки, культивируемые с СТ-1, характеризовались наличием примерно 12% апоптозных клеток по сравнению с 21% апоптозных клеток, обнаруженных при отсутствии СТ-1.
Фиг.9. Анализ воздействия СТ-1 на пролиферацию клеток, измеренного путем введения [3H]тимидина. Результаты свидетельствуют о процентном увеличении (ордината) пролиферации клеток, обработанных СТ-1 (СТ-1), по сравнению с контрольными клетками, не подвергнутыми обработке (С = контрольный образец).
Фиг.10. Иммунологический анализ фосфорилированных сигнальных белков (Jak-1-Y, Stat-3-Y-705 и АКТ-Ser-475) в лизатах клеток Н35, полученные в разные периоды времени (минуты) после инкубации клеток с СТ-1.
А) Иммунопреципитация клеточных лизатов антителами, специфическими к Jak-1. При выполнении вестерн-блоттинга с использованием антител, специфических к фосфорилированным тирозинам, фосфорилирование молекулы Jak-1 наблюдалось через 5 минут.
В) Вестерн-блоттинг с использованием антител, специфических к фосфорилированному Stat-3 (Stat-3-Y-705), при выполнении которого положительный результат наблюдается через 5 минут после обработки.
С) Иммунопреципитация цитозольной фракции антителами против АКТ с последующим выполнением вестерн-блоттинга с использованием антител, специфических к фосфорилированному АКТ по серину 475 (AKT-Ser-475), свидетельствует о обнаружении индукции через 15 и 30 минут.
Фиг.11. Структура вектора pET-14b.
Фиг.12А. Вестерн-блоттинг сигнальных белков в печени крыс, получавших AdCT-1, AdLac-Z или физиологический раствор (S), которых затем подвергали резекции более 85% печени и умерщвляли через 1 час после резекции.
1а) Вестерн-блоттинг с использованием антител, специфических к фосфорилированному Stat-3 (Stat-3-Y-705).
1b) Вестерн-блоттинг с использованием антител, специфических к Stat-3, для количественного определения общего содержания Stat-3.
Фосфорилирование Stat-3 обнаружено у крыс, получавших AdCT-1.
2а) Вестерн-блоттинг с использованием антител, специфических к фосфорилированному Erk1 и Erk2 (Erk1-Thr-202 и Erk2-Tyr-204).
2b) Вестерн-блоттинг с использованием антител, специфических к Erk1 и Erk2, для количественного определения общего содержания Erk1 и Erk2.
Фосфорилирование ERK1/2 обнаружено у крыс, получавших AdCT-1.
3а) Вестерн-блоттинг с использованием антител, специфических к фосфорилированному Akt (Akt-Ser-473).
3b) Вестерн-блоттинг с использованием антител, специфических к Akt, для количественного определения общего содержания Akt.
Фосфорилирование Akt обнаружено у крыс, получавших AdCT-1.
Фиг.12В. Активность каспазы-3 в печени крыс, подвергнутых обширной резекции печени (>85%). Образцы получены из тех же групп подвергнутых гепатэктомии крыс, которые были использованы в экспериментах, показанных на фиг.12А. На диаграмме показано увеличение активности каспазы-3 (число раз) по сравнению с животными, имеющими здоровую печень.
фиг.13. Вестерн-блоттинг сигнальных белков в печени мышей, получавших соответственно AdCT-1, AdLac-Z или физиологический раствор (S), до введения Con-A для индукции поражения печени. Образцы были получены в момент умерщвления животных, через 1 час после индукции поражения печени при помощи Con-A.
1а) Вестерн-блоттинг с использованием антител, специфических к фосфорилированному Akt (Akt-Ser-473).
1b) Вестерн-блоттинг с использованием антител, специфических к Akt, для количественного определения общего содержания Akt.
Фосфорилирование Akt обнаружено у крыс, получавших AdCT-1.
2а) Вестерн-блоттинг с использованием антител, специфических к фосфорилированному Erk1 и Erk2 (Erk1-Thr-202 и Erk2-Tyr-204).
2b) Вестерн-блоттинг с использованием антител, специфических к Erk1 и Erk2, для количественного определения общего содержания Erk1 и Erk2.
Фосфорилирование ERK1/2 обнаружено у крыс, получавших AdCT-1.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ МОДИФИЦИРОВАННЫХ ЦИКЛОСПОРИНОВ | 2007 |
|
RU2463071C2 |
| ТЕРАПЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ СНИЖЕНИЯ УСТОЙЧИВОСТИ К ИНГИБИТОРАМ ТИРОЗИНКИНАЗЫ | 2016 |
|
RU2735493C2 |
| АНТИФИБРОГЕННЫЕ СОЕДИНЕНИЯ, СПОСОБЫ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2726416C2 |
| ПРИМЕНЕНИЕ ВИРУСА МИКСОМЫ ДЛЯ ТЕРАПЕВТИЧЕСКОГО ЛЕЧЕНИЯ РАКА И ХРОНИЧЕСКОЙ ВИРУСНОЙ ИНФЕКЦИИ | 2004 |
|
RU2362584C2 |
| ТЕРАПЕВТИЧЕСКОЕ СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ НЕРВНОЙ СИСТЕМЫ | 2018 |
|
RU2825666C2 |
| СРЕДСТВА, СТИМУЛИРУЮЩИЕ РЕГЕНЕРАЦИЮ ТКАНЕЙ | 2013 |
|
RU2599289C2 |
| КОМПОЗИЦИЯ ДЛЯ ПОДАВЛЕНИЯ РАКОВЫХ ЗАБОЛЕВАНИЙ КРОВИ ИЛИ МЕТАСТАЗИРОВАНИЯ РАКА, СОДЕРЖАЩАЯ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА МОНОАЦЕТИЛДИГЛИЦЕРИДНОЕ СОЕДИНЕНИЕ | 2014 |
|
RU2632098C2 |
| НОВЫЙ ОПУХОЛЕВЫЙ БИОМАРКЕР | 2010 |
|
RU2567005C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ДИАГНОСТИКИ И ЛЕЧЕНИЯ РАКА | 1996 |
|
RU2174409C2 |
| СПОСОБ СТИМУЛЯЦИИ РЕГЕНЕРАЦИИ ТКАНЕЙ | 2007 |
|
RU2480213C2 |











Изобретение относится к биологии, медицине и раскрывает повышение экспрессии кардиотрофина (СТ-1) в процессе регенерации печени, которая совпадает с максимальной пролиферацией гепатоцитов. Изобретение касается также применения СТ-1 и композиции на его основе в качестве стимулятора регенерации печени, а также гепатозащитной роли СТ-1 в разных моделях острого поражения печени. Изобретение обеспечивает использование СТ-1 для улучшения функции печени при различных ее патологиях. 2 н. и 8 з.п. ф-лы, 13 ил.
| Jin H | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| Peters M et all, "Neu hepatocyte stimulating factor..", FEBS LETTERS, vol.372, n/2-3, 1995, p.177-180 | |||
| US 6046035, 04.04.2000 | |||
| US 6011197, 04.01.2000. | |||

