Область техники
Настоящее изобретение относится к фармацевтическим композициям, включающим вещество, обладающее активностью модулирования биологической активности «индуцируемой активацией лимфоцитарной иммуномодуляторной молекулы» (AILIM) (известной также, как «индуцируемый совместный стимулятор» (ICOS)), в частности, трансдукции сигнала, опосредованной AILIM.
Конкретно, настоящее изобретение относится к фармацевтическим композициям, включающим вещество, обладающее активностью регулирования (например, ингибирования) пролиферации клеток, экспрессирующих AILIM, гибели клеток (или апоптоза), или иммунного цитолизиса, или модулирования (например, ингибирования) продукции цитокина (например, интерферона-γ или интерлейкина-4) клетками, экспрессирующими AILIM.
Более конкретно, настоящее изобретение включает вещества, обладающие активностью модулирования трансдукции сигнала посредством AILIM и, особенно предпочтительно, вещества, которые индуцируют гибель клеток, апоптоз или истощение запаса клеток, экспрессирующих AILIM. Настоящее изобретение относится к фармацевтическим композициям для подавления, лечения или профилактики заболеваний, сопровождающих нарушенный иммунитет кишечного тракта (например, воспалительные кишечные заболевания, такие как колит (язвенный колит и подобные заболевания) и болезнь Крона и пищевые аллергии).
Предшествующий уровень техники
Слизистые оболочки желудочно-кишечного тракта постоянно подвержены воздействию не только антигенов, происходящих из пищи и энтеробактериальной флоры, но также различных антигенов, существующих во внешней среде, которые вредны для живого организма, таких как патогенные микроорганизмы. Поэтому слизистые оболочки желудочно-кишечного тракта проявляют цитотоксическую активность для противодействия таким антигенам, вредным для живого организма. Сохраняя способность секретировать антитела для нейтрализации токсинов, указанные слизистые оболочки в то же самое время также имеют уникальный иммунный механизм подавления избыточных иммунных реакций против антигенов, таких как антигены пищевой и энтеробактериальной флор (данный механизм называется иммунитетом слизистых оболочек желудочно-кишечного тракта или кишечным иммунитетом). В частности, нормальный иммунитет слизистых оболочек устанавливается на равновесии между положительными иммунными реакциями против патогенов и отрицательными иммунными реакциями против непатогенных антигенов. Когда указанное равновесие иммунологической гомеостатической поддержки рушится, происходят воспаление, аллергии и инфекции, давая начало кишечным заболеваниям, обычно именуемым воспалительные кишечные заболевания (IBD), и пищевые аллергии.
Наиболее репрезентативными воспалительными кишечными заболеваниями являются болезнь Крона (CD) и колит (в частности, язвенный колит (UC)). Оба указанных заболевания представляют собой заболевания, при которых патоген невозможно определить, и происходят хронические и рецидивирующие приступы боли в животе и диареи, вызывающие значительные и долгосрочные препятствия повседневной жизни больных детей и лиц молодого возраста. Кроме того, поскольку колит (в частности, язвенный колит) может стать причиной рака толстой кишки, имеется неотложная необходимость в выяснении патогенеза колита и разработке эффективных способов лечения.
Хотя различные возможности, такие как генетические факторы и факторы окружающей среды, обсуждались в отношении механизма начала воспалительных кишечных заболеваний, последние исследования указывают на высокую вероятность возможности того, что причиной может быть нарушенный иммунитет кишечного тракта (иммунитет слизистой оболочки желудочно-кишечного тракта). Более конкретно, воспаление или аллергия возникает в слизистых оболочках кишечника вследствие индукции избыточной иммунной реакции, которая происходит по некой причине против антигенов в кишечнике, которые обычно не являются патогенными и имеют низкую иммуногенность, что приводит к началу воспалительного кишечного заболевания.
Кроме того, предполагали, что нарушенный иммунитет против чужеродных патогенов, антигенов, происходящих из пищи, или аутоантигенов глубоко вовлечен в такие воспаления и аллергии кишечника. Кроме того, последние исследования свидетельствовали о возможности того, что нарушенные иммунные реакции в отношении определенных местных бактерий проявляются в виде хронических воспалительных реакций.
Данный механизм начала воспалений и аллергий кишечника вследствие нарушенного иммунитета кишечного тракта подтверждается анализами функции и дифференциации Т-клеток пациентов, а также типа продукции цитокинов в повреждениях или сыворотке. Кроме того, анализ различных недавно разработанных экспериментальных моделей воспалительных кишечных заболеваний на животных также выявляет то, что нарушенный иммунитет слизистых оболочек вызывает хроническое воспаление в кишечнике (Gastroenterology, Vol.109, p.1344-1367, 1995).
Например, ясно, что Т-клетки глубоко вовлечены в начало хронического энтерита, поскольку воспаление кишечника развивается спонтанно у нокаутных мышей с дефицитом α-цепи Т-клеточного рецептора (TCR) (TCRα-/-) (Cell, Vol.75, p.275-282, 1993; J. Exp. Med., Vol.183, p.847-856, 1996). При колите у указанных мышей TCRα-/-продукция интерферона-γ (IFN-γ) в кишечнике возрастает, и на начальной стадии воспаления наблюдается подъем уровней интерлейкинов IL-1α и IL-1β (Laboratory Investigation, Vol.76, p.385-397, 1997). Кроме того, Т-клетки TCRβ (βdim), которые имеют специфический поднабор Vβ и продуцируют IL-4, могут наблюдаться в пищеварительном тракте и лимфоузлах (Gastroenterology, Vol.112, p.1976-1886, 1997). В данной модели, как полагают, дефицит Т-клеток TCRαβ вызывает увеличение фракции патологических Т-клеток, которое затем вызывает патологическую регуляцию продукции цитокинов, становясь медиатором воспаления.
В модели, в которой Т-клетки CD4+/CD45RBhigh вводят мышам с тяжелым комбинированным иммунодефицитом (мышам SCID), индуцируется развитие тяжелого энтерита, сопровождаемого гиперплазией слизистого слоя и инфильтрацией лимфоцитов в кишечнике. Однако данный энтерит не возникает, когда одновременно вводят нефракционированные Т-клетки CD4+ (J. Exp. Med., Vol.178, p.237-244, 1993; Int. Immunol., Vol.5, p.1461-1471, 1993). Т-клетки CD4+ мышей SCID, у которых имеется развившийся энтерит, продуцируют IFN-γ. С другой стороны, поскольку энтерит подавляется введением антител против IFN-γ, считают, что Т-клетки типа Th1 вызывают воспаление (Immunity, Vol.1, p.553-562, 1994).
На основании указанных фактов представляется, что нет сомнений в том, что Т-клетки CD4+ кишечника и их избыточная активация представляют собой важные факторы при воспалительных кишечных заболеваниях.
Кроме того, регрессия энтерита с уменьшением количества Т-клеток CD4+ у пациентов, пораженных и воспалительным заболеванием кишечника, и ВИЧ, также подтверждает глубокое вовлечение патологических Т-клеток CD4+ при воспалительных заболеваниях кишечника (J. Clin. Gastroenterology, Vol.23, p.24-28, 1996). На основании указанных данных предпринимались попытки лечения воспалительных кишечных заболеваний с использованием антитела против CD-4 и сообщалось, что воспалительные поражения подавляются введением антитела против CD-4 (Gut, Vol.40, p.320-327, 1997).
С другой стороны, такая патологическая функциональная регуляция Т-клеток означает, что нарушилось равновесие продукции регуляторных цитокинов.
В действительности, имеются также сообщения о том, что энтерит развивается спонтанно у нокаутных мышей с дефицитом IL-2 и у нокаутных мышей с дефицитом IL-10 (Cell, Vol.75, p.235-261, 1993; Cell, Vol.75, p.263-274, 1993). Кроме того, в указанных моделях наблюдается также избыточная продукция IFN-γ, подтверждая тот факт, что произошла избыточная реакция Т-клеток типа Th1. Избыточная продукция IFN-γ в указанных моделях также согласуется с наблюдением возросшей экспрессии IFN-γ в поражениях, наблюдающаяся при болезни Крона. Энтерит можно лечить у мышей с дефицитом IL-10 введением IL-10. Сообщалось, что энтерит можно подавить указанным способом у мышей SCID, которым вводили Т-клетки CD4+/CD45RBhigh (Immunity, Vol.1, p.553-562, 1994).
Как указано выше, прогресс в анализе механизма начала воспалительных кишечных заболеваний начинался с аспекта патологического иммунитета слизистой оболочки желудочно-кишечного тракта, предполагая возможность лечения воспалительных заболеваний кишечника подавлением возросшей активации Т-клеток CD4+ и избыточно продуцируемых цитокинов. Однако истинный патогенез воспалительных заболеваний кишечника еще не был выявлен, и, кроме того, не был предоставлен эффективный способ лечения.
Активация Т-клеток (приобретение антигенной специфичности) инициируется, когда Т-клетки распознают антигены, представленные антиген-представляющими клетками (APC), такими как макрофаги, В-клетки или дендритные клетки. АРС обрабатывают включенные антигены, и обработанные антигены связываются с главным комплексом гистосовместимости (МНС) и представляются. Т-клетки получают первый сигнал для клеточной активации (приобретения специфичности) в результате распознавания обработанного антигена, представленного АРС посредством комплекса, образованного между рецептором Т-клетки (TCR) на поверхности мембраны Т-клетки и антигеном (комплекс TCR/CD3).
Для достаточной активации Т-клеток в дополнение к первому сигналу необходим второй сигнал, называемый совместным стимулирующим сигналом. Т-клетки активируются специфически в отношении антигена приемом указанного совместного стимулирующего сигнала после приема первого сигнала.
Для трансдукции данного второго сигнала крайне важно взаимодействие (более конкретно, межклеточная адгезия, опосредованная связями, образованными между следующими молекулами) среди CD28 (также известной, как Тр44, Т44 или антиген 9.3), которая представляет собой молекулу клеточной поверхности, экспрессированную, главным образом, на Т-клетках и клетках тимуса, CD80 (также известной, как В7-1, В7, ВВ1 или B7/BB1), которая представляет собой молекулу клеточной поверхности, экспрессированную антиген-представляющими клетками (макрофаги, моноциты, дендритные клетки и т.д.), и CD86 (также известной, как В7-2 или В70), которая также представляет собой молекулу клеточной поверхности на антиген-представляющих клетках.
Кроме того, экспериментально было выявлено взаимодействие (в частности, межклеточная адгезия, опосредованная связями, образуемыми между следующими молекулами) между ассоциированным с цитолитическими Т-лимфоцитами (CTLA-4) антигеном 4, экспрессия которого усиливается в зависимости от второго сигнала, CD80 (B7-1) и CD86 (B7-2) также играет важную роль в регуляции активации Т-клеток указанным вторым сигналом. Более конкретно, было выявлено, что регуляция активации Т-клеток трансдукцией указанного второго сигнала включает, по меньшей мере, взаимодействие между CD28 и CD80/CD86, усиление экспрессии CTLA-4, зависящее, как полагают, от указанного взаимодействия, и взаимодействие между CTLA-4 и CD80/CD86.
Кроме того, недавно, подобно описанным выше CTLA-4 и CD28, была идентифицирована молекула, названная индуцируемой активацией лимфоцитарной иммуномодуляторной молекулой (AILIM; человека, мыши и крысы; Int. Immunol., 12(1), p.51-55, 2000; также называемая индуцируемый совместный стимулятор (ICOS; человека; Nature, 397(6716), p.263-266, 1999); J. Imminol., 166(1), p.1, 2001; J. Immunol., 165(9), p.5035, 2000; Biochem. Biophys. Res. Commun., 276(1), p.335, 2000; Immunity, 13(1), p.95, 2000; J. Exp. Med., 192(1), p.53, 2000; Eur. J. Immunol., 30(4), p.1040, 2000), в качестве третьей, совместно стимулирующей молекулы передачи, которая трансдуцирует второй сигнал (совместно стимулирующий сигнал), необходимый для активации лимфоцитов, таких как Т-клетки, и связана с сигналом, регулирует функцию активированных лимфоцитов, таких как активированные Т-клетки.
Кроме того, была идентифицирована новая молекула, названная B7h, B7RP-1, GL50 или LICOS, которую считают лигандом, взаимодействующим с совместно стимулирующей молекулой передачи AILIM (Nature, Vol.402, №6763, pp.827-832, 1999; Nature Medicine, Vol.5, №12, pp.1365-1369, 1999; J. Immunology, Vol.164, pp.1653-1657, 2000; Curr. Biol., Vol.10, №6, pp.333-336, 2000).
Проводятся трудоемкие исследования биологических функций указанных двух новых молекул, функциональной регуляции молекулами лимфоцитов, таких как Т-клетки, посредством трансдукции третьего совместно стимулирующего сигнала.
С другой стороны, не было даже предположений о взаимосвязи между AILIM (ICOS), которая представляет собой третью совместно стимулирующую молекулу трансдукции, считающуюся существенной для активации Т-клеток, таких как Т-клетки CD4+, и началом указанного выше патологического иммунитета слизистой оболочки кишечника и воспалительных заболеваний кишечника (болезнь Крона и колит (язвенный колит и ему подобный)). Не было также никакого предложения о попытках лечения воспалительных заболеваний кишечника регулированием функции молекулы AILIM.
Описание изобретения
Конкретно, задачей настоящего изобретения является предоставление способов и фармацевтических средств для подавления, лечения или профилактики заболеваний, сопровождающих патологический иммунитет (патологическую активацию Т-клеток, увеличение количества патологических клеток CD4+) кишечного тракта, таких как воспалительные заболевания кишечника (болезнь Крона и колит (язвенный колит и ему подобный)) модулированием посредством лекарственных и фармацевтических способов (например, фармацевтических средств, таких как соединения с низкой молекулярной массой и антибиотики) биологической функции новой молекулы AILIM, которая, как считают, трансдуцирует второй сигнал, существенный для активации лимфоцитов, таких как Т-клетки (совместно стимулирующий сигнал), и регулирует функцию активированных лимфоцитов, таких как активированные Т-клетки.
Еще одной задачей является применение таких фармацевтических средств, которые модулируют биологическую функцию AILIM (например, фармацевтические средства, такие как соединения с низкой молекулярной массой и антибиотики) для предоставления способов усиления терапевтического эффекта существующих фармацевтических средств, широко применяемых для лечения воспалительных заболеваний кишечника (гормонов коры надпочечников, салазосульфапиридина и т.д.).
Обширные исследования способов подавления биологической функции AILIM (ICOS) млекопитающих, пищевых аллергий и воспалительных заболеваний кишечника, при которых может быть глубоко вовлечен патологический иммунитет кишечного тракта (в частности, болезнь Крона и колит (язвенный колит и ему подобный)), привели авторов настоящего изобретения к открытию того, что фармацевтические средства, которые регулируют функцию AILIM, значительно подавляют воспалительные заболевания кишечника (в частности, болезнь Крона и колит (язвенный колит и ему подобный)). Таким образом, было создано настоящее изобретение.
Фармацевтическую композицию настоящего изобретения можно применять в качестве лекарственного средства для модулирования различных реакций in vivo, в которых участвует трансдукция совместно стимулирующего сигнала к клеткам, экспрессирующим AILIM, опосредованная AILIM (например, пролиферации клеток, экспрессирующих AILIM, продукции цитокина (цитокинов) клетками, экспрессирующими AILIM, иммунного цитолизиса или гибели клеток, апоптоза или истощения запаса клеток, экспрессирующих AILIM, и активности индукции зависимой от антител клеточной цитотоксичности против клеток, экспрессирующих AILIM), и/или в качестве лекарственного средства для профилактики начала и/или прогрессирования различных заболеваний, при которых участвует трансдукция сигнала, опосредованная AILIM, и для лечения или профилактики указанных заболеваний.
Конкретно, фармацевтическая композиция изобретения может модулировать (подавлять и усиливать) пролиферацию клеток, экспрессирующих AILIM, гибель, апоптоз или истощение запаса клеток, или иммунный цитолизис, или может модулировать (ингибировать или усиливать) продукцию цитокинов (например, интерферона-γ, или интерлейкина-4) клетками, экспрессирующими AILIM, и может предотвращать различные патологические состояния, запускаемые различными физиологическими феноменами, в которых участвует трансдукция сигнала, опосредованная AILIM, и обеспечивает возможность лечения или профилактики различных заболеваний.
Таким особенно предпочтительным вариантом реализации фармацевтических композиций данного изобретения являются фармацевтические композиции, включающие вещество, которое вызывает гибель клеток, апоптоз или истощение запаса клеток, экспрессирующих AILIM.
Применяя фармацевтические композиции данного изобретения, можно подавить, предотвратить и/или лечить заболевания, которые могут быть вызваны патологическим иммунитетом кишечного тракта, более конкретно, воспалительные заболевания кишечника (в частности, болезнь Крона и колит (язвенный колит и ему подобный)) и пищевые аллергии.
Кроме того, фармацевтические композиции данного изобретения могут усилить терапевтический эффект при воздействии на воспалительные заболевания кишечника при применении в комбинации с существующим фармацевтическим средством, назначаемым для лечения таких воспалительных заболеваний кишечника.
Более конкретно, настоящее изобретение описано в следующих пунктах (1)-(10).
(1) Фармацевтическая композиция для подавления, лечения или профилактики заболевания, которое сопровождает патологический иммунитет кишечного тракта, причем фармацевтическая композиция включает вещество, обладающее активностью модулирования трансдукции сигнала посредством AILIM, и фармацевтически приемлемый носитель.
(2) Фармацевтическая композиция по п.(1), где указанное вещество обладает активностью вызывать гибель клеток, экспрессирующих AILIM.
(3) Фармацевтическая композиция по п.(1) или (2), где указанное заболевание представляет собой воспалительное заболевание кишечника.
(4) Фармацевтическая композиция по п.(3), где указанное воспалительное заболевание кишечника представляет собой колит.
(5) Фармацевтическая композиция по п.(3), где указанное воспалительное заболевание кишечника представляет собой болезнь Крона.
(6) Фармацевтическая композиция по п.(1) или (2), где указанное заболевание представляет собой пищевую аллергию.
(7) Фармацевтическая композиция по любому одному из пп.(1)-(6), где указанное вещество представляет собой белковое вещество.
(8) Фармацевтическая композиция по п.(7), где указанное белковое вещество выбирают из группы, состоящей из:
а) антитела, которое связывается с AILIM, или части указанного антитела;
b) полипептида, включающего всю внеклеточную область AILIM или ее часть;
c) гибридного полипептида, включающего всю или часть внеклеточной области AILIM и всю или часть константной области тяжелой цепи иммуноглобулина, и
d) полипептида, который связывается с AILIM.
(9) Фармацевтическая композиция по любому одному из пп.(1)-(6), где указанное вещество представляет собой небелковое вещество.
(10) Фармацевтическая композиция по п.(9), где указанное небелковое вещество представляет собой ДНК, РНК или химически синтезированное соединение.
Настоящие изобретения подробно описаны здесь ниже определением терминов и способов продукции веществ, применяемых в данном изобретении.
В настоящем описании термин «млекопитающее» означает человека, корову, козу, кролика, мышь, крысу, хомяка и морскую свинку; предпочтительным является человек, корова, крыса, мышь или хомяк, а особенно предпочтительным является человек.
"AILIM" данного изобретения представляет собой аббревиатуру «индуцируемой активацией лимфоцитарной иммуномодуляторной молекулы» и означает молекулу клеточной поверхности млекопитающего, имеющую структуру и функцию, описанные в предыдущих сообщениях (J. Immunol., 166(1), p.1, 2001; J. Immunol., 165(9), p.5035, 2000; Biochem. Biophys. Res. Commun., 276(1), p.335, 2000; Immunity, 13(1), p.95, 2000; J. Exp. Med., 192(1), p.53, 2000; Eur. J. Immunol., 30(4), p.1040, 2000; Int. Immunol., 12(1), p.51, 2000; Nature, 397(6716), p.263, 1999; GenBank Accession Number: BAA82129 (человек); BAA82128 (крыса); BAA82127 (мутантная крыса); BAA82126 (мышь)).
Особенно предпочтительно, термин обозначает AILIM, полученную у человека (например, International Immunology, Vol.12, №1, p.51-55, 2000; GenBank Accession Number: BAA82129).
Данную AILIM также называют ICOS (Nature, Vol.397, №6716, p.263-266, 1999) или антиген JTT-1/антиген JTT-2 (не рассмотренная опубликованная заявка на патент Японии №(JP-A) Hei 11-29599, международная патентная заявка №WO98/38216), и данные молекулы взаимно относятся к одной и той же молекуле.
Кроме того, "AILIM" в данном изобретении включает аминокислотные последовательности AILIM от каждого млекопитающего, описанного в ранее опубликованных литературных источниках, а особенно предпочтительно, полипептид, имеющий по существу ту же аминокислотную последовательность, что и последовательность AILIM человека. Кроме того, мутанты AILIM человека, аналогичные ранее идентифицированному мутанту AILIM, полученному у крысы (GenBank Accession Number: BAA82127), также включены в "AILIM" данного изобретения.
В настоящем описании выражение «имеющая по существу такую же аминокислотную последовательность» означает, что "AILIM" настоящего изобретения включает полипептиды, имеющие аминокислотные последовательности, в которых множественные аминокислоты, предпочтительно, от 1 до 10 аминокислот, особенно предпочтительно, от 1 до 5 аминокислот, были замещены, подвергнуты делеции и/или модифицированы, и полипептиды, имеющие аминокислотные последовательности, в которых множественные аминокислоты, предпочтительно, от 1 до 10 аминокислот, особенно предпочтительно, от 1 до 5 аминокислот, были добавлены, до тех пор, пока полипептиды имеют по существу такие же биологические свойства, как полипептид, включающий аминокислотную последовательность, показанную в предыдущих сообщениях.
Такие замещения, делеции или инсерции аминокислот можно достичь в соответствии с обычным способом (Experimental Medicine: SUPPLEMENT, "Handbook of Genetic Engineering" (1992), и т.д.).
Примерами являются сайт-направленный мутагенез с использованием синтетического олигонуклеотида (дуплексный способ с гэпами), точечный мутагенез, посредством которого точечную мутацию вводят случайным образом обработкой нитритом или сульфитом, способ, посредством которого получают делеционный мутант с помощью фермента Bal31, и т.д., кассетный мутагенез, способ сканирования линкера, способ ошибки включения, способ с использованием затравки с ошибочным спариванием, способ синтеза сегмента ДНК и т.д.
Сайт-направленный мутагенез с использованием синтетического олигонуклеотида (дуплексный способ с гэпами) можно осуществить, например, следующим образом. Область, которую желательно подвергнуть мутагенезу, клонируют в вектор фага М13, имеющий амбер-мутацию, для получения одноцепочечной ДНК фага. После того как ДНК RF I вектора М13, не имеющего амбер-мутации, линеаризируют обработкой рестрикционным ферментом, ДНК смешивают с указанной выше одноцепочечной ДНК фага, денатурируют и подвергают отжигу, образуя посредством этого «дуплексную ДНК с гэпами». Синтетический олигонуклеотид, в который введены мутации, гибридизируют с дуплексной ДНК с гэпами, и замкнутую в кольцо двухцепочечную ДНК получают путем реакции с ДНК-полимеразой и ДНК-лигазой. Клетки mutS E. coli, дефицитные по активности реапарации ошибочного спаривания, трансфицируют данной ДНК. Клетки E. coli, не имеющие супрессорной активности, инфицируют выращенными фагами и проводят скрининг только фагов, не имеющих амбер-мутации.
В способе, посредством которого вводят точечную мутацию нитритом, используют, например, принцип, указанный ниже. Если ДНК обрабатывают нитритом, нуклеотиды деаминируются с изменением аденина в гипоксантин, цитозина в урацил и гуанина в ксантин. Если в клетки вводят деаминированную ДНК, "A:T" и "G:С" соответственно замещаются "G:C" и "A:T", потому что при репликации ДНК основание гипоксантина, урацила и ксантина конъюгируется соответственно с цитозином, аденином и тимином. На самом деле, фрагменты одноцепочечной ДНК, обработанные нитритом, гибридизируют с «дуплексной ДНК с гэпами», а затем мутантные штаммы отделяют манипулированием таким же образом, как при сайт-направленном мутагенезе с использованием синтетических олигонуклеотидов (дуплексный способ с гэпами).
Термин «цитокин», как в выражении «продукция цитокина клетками, экспрессирующими AILIM», в настоящем изобретении означает произвольный цитокин, продуцируемый клетками, экспрессирующими AILIM (в частности, Т-клетками).
Примерами Т-клеток являются Т-клетки типа Th1 или типа Th2, а цитокин настоящего изобретения, в частности, означает цитокин, продуцируемый Т-клетками типа Th1, и/или произвольный цитокин, продуцируемый Т-клетками типа Th2.
Цитокины, продуцируемые Т-клетками типа Th1, включают IFN-γ, IL-2, фактор опухолевого некроза (TNF), IL-3, а цитокины, продуцируемые Т-клетками типа Th2, включают IL-3, IL-4, IL-5, IL-10 и TNF (Cell, Vol.30, №9, pp.343-346, 1998).
Используемое в настоящем изобретении выражения «вещество», «вещество, обладающее активностью модулирования трансдукции сигнала, опосредованной AILIM», «вещество, обладающее активностью ингибирования пролиферации клеток, экспрессирующих AILIM, или ингибирования продукции цитокина клетками, экспрессирующими AILIM» или «вещество, обладающее активностью индукции гибели клеток, экспрессирующих AILIM» означают встречающееся в природе вещество или искусственно полученное произвольное вещество.
Особенно предпочтительным вариантом реализации «вещества» в соответствии с данным изобретением является вещество, обладающее активностью индукции гибели клеток, апоптоза или истощения запаса клеток, экспрессирующих AILIM.
В настоящем описании выражение «трансдукция сигнала, опосредованная AILIM», означает трансдукцию сигнала посредством AILIM, приводящую к описанному выше изменению любого фенотипа клеток, экспрессирующих AILIM, или в последующих примерах (изменение клеточной пролиферации, активация клеток, инактивация клеток, апоптоз и/или способность продуцировать произвольный цитокин из клеток, экспрессирующих AILIM).
«Вещество» можно, главным образом, классифицировать на «белковое вещество» и «небелковое вещество».
Примерами «белковых веществ» являются следующие полипептиды, антитела (поликлональные антитела, моноклональные антитела или части моноклональных антител).
Когда вещество представляет собой антитело, оно предпочтительно является моноклональным антителом. Когда вещество представляет собой моноклональное антитело, оно включает не только моноклональные антитела, происходящие от млекопитающего, кроме человека, но также следующее: рекомбинантные химерные моноклональные антитела, рекомбинантные гуманизированные моноклональные антитела и человеческие моноклональные антитела.
Когда вещество представляет собой полипептид, оно включает следующее: полипептиды, фрагменты полипептидов (олигопептиды), гибридные полипептиды и химически модифицированные полипептиды. Примерами олигопептидов являются пептиды, включающие от 5 до 30 аминокислот, предпочтительно, от 5 до 20 аминокислот. Химическую модификацию можно проектировать в зависимости от различных назначений, например, для увеличения периода полувыведения из крови в случае введения in vivo, или увеличения устойчивости к разложению, или увеличения всасывания в пищеварительном тракте при пероральном введении.
Примерами полипептидов являются следующие:
(1) Полипептид, содержащий всю или часть внеклеточной области AILIM;
(2) Гибридный полипептид, включающий всю или часть внеклеточной области AILIM и всю или часть константной области тяжелой цепи иммуноглобулина; или
(3) Полипептид, который связывается с AILIM.
Примерами «небелковых веществ» являются ДНК, РНК и химически синтезированные соединения.
В данном описании «ДНК» означает «ДНК, включающую частичную нуклеотидную последовательность антисмысловой ДНК, полученной на основании нуклеотидной последовательности ДНК (включая кДНК и геномную ДНК), кодирующей указанную выше AILIM (предпочтительно, человеческую AILIM), или ее химически модифицированную форму», которую можно применять в качестве лекарственного препарата антисмысловой ДНК. В частности, антисмысловая ДНК может ингибировать транскрипцию ДНК, кодирующей AILIM, в мРНК, или трансляцию мРНК в белок путем гибридизации с ДНК или РНК, кодирующей AILIM.
Используемое в настоящем описании выражение «частичная нуклеотидная последовательность» относится к частичной нуклеотидной последовательности, включающей произвольное количество нуклеотидов в произвольной области. Частичная нуклеотидная последовательность включает от 5 до 100 последовательных нуклеотидов, предпочтительно, от 5 до 70 последовательных нуклеотидов, более предпочтительно, от 5 до 50 последовательных нуклеотидов, а еще предпочтительнее, от 5 до 30 последовательных нуклеотидов.
Когда ДНК применяют в качестве лекарственного препарата антисмысловой ДНК, последовательность ДНК можно химически частично модифицировать для удлинения периода полувыведения (устойчивости) из крови, когда ДНК вводят пациентам, для увеличения проницаемости ДНК через цитоплазматическую мембрану, или для увеличения устойчивости к разрушению или всасывания перорально введенной ДНК в пищеварительных органах. Химические модификации включают, например, модификацию фосфатной связи, рибозы, нуклеотида, сахарной части и 3'-конца и/или 5'-конца в структуре ДНК олигонуклеотида.
Модификации фосфатных связей включают, например, превращение одной или более связей в сложный эфир двухосновной фосфорной кислоты (D-олиго), фосфортиоатные связи, фосфордитиоатные связи (S-олиго), метилфосфонатные связи (МР-олиго), фосфорамидатные связи, нефосфатные связи или метилфосфонотиоатные связи, или их комбинации. Модификация рибозы включает, например, превращение в 2'-фторрибозу или 2'-О-метилрибозу. Модификация нуклеотида включает, например, превращение в 5-пропинилурацил или 2-аминоаденин.
В данном описании термин «РНК» означает «РНК, включающую частичную нуклеотидную последовательность антисмысловой РНК, полученной на основании нуклеотидной последовательности РНК, кодирующей указанную выше AILIM (предпочтительно, человеческую AILIM), или ее химически модифицированную форму», которую можно применять в качестве лекарственного препарата антисмысловой РНК. Антисмысловая РНК может ингибировать транскрипцию ДНК, кодирующей AILIM, в мРНК или трансляцию мРНК в белок путем гибридизации с ДНК или РНК, кодирующей AILIM.
Используемое здесь выражение «частичная нуклеотидная последовательность» относится к частичной нуклеотидной последовательности, включающей произвольное количество нуклеотидов в произвольной области. Частичная нуклеотидная последовательность включает от 5 до 100 последовательных нуклеотидов, предпочтительно от 5 до 70 последовательных нуклеотидов, более предпочтительно от 5 до 50 последовательных нуклеотидов, а еще предпочтительнее от 5 до 30 последовательных нуклеотидов.
Последовательность антисмысловой РНК можно химически частично модифицировать для удлинения периода полувыведения (устойчивости) из крови, когда РНК вводят пациентам, для увеличения проницаемости РНК через цитоплазматическую мембрану, или для увеличения устойчивости к разрушению или всасывания перорально введенной РНК в пищеварительных органах. Химические модификации включают такие, как модификации, которые относятся к указанной выше антисмысловой ДНК.
Примерами «химически синтезированного соединения» являются произвольное соединение, исключая указанные выше ДНК, РНК и белковые вещества, имеющее молекулярную массу от около 100 до около 1000 или менее, предпочтительно, вещество, имеющее молекулярную массу от около 100 до около 800, а предпочтительнее, молекулярную массу от около 100 до около 600.
Термин «полипептид», включенный в определение указанного выше «вещества», означает часть (фрагмент) полипептидной цепи, составляющей AILIM (предпочтительно, человеческой AILIM), предпочтительно, всю или часть внеклеточной области полипептида, составляющего AILIM (от 1 до 5 аминокислот можно необязательно добавить в N-конец и/или С-конец области).
AILIM в соответствии с настоящим изобретением представляет собой трансмембранную молекулу, пронизывающую клеточную мембрану, включающую 1 или 2 полипептидных цепи.
В настоящем описании «трансмембранный белок» означает белок, который соединен с клеточной мембраной посредством гидрофобной области пептида, который пронизывает липидный бислой мембраны один или несколько раз, и структура которого в целом составлена из трех основных областей, то есть внеклеточной области, трансмембранной области и цитоплазматической области, как наблюдается во многих рецепторах или молекулах клеточной поверхности. Такой трансмембранный белок составляет каждый рецептор или молекулу клеточной поверхности в виде мономера или в виде гомодимера, гетеродимера или олигомера, связанного с одной или несколькими цепями, имеющими одну и ту же или различную аминокислотную последовательность (последовательности).
В настоящем описании «внеклеточная область» означает всю или часть частичной структуры (частичной области) всей структуры упомянутого выше трансмембранного белка, где частичная структура существует снаружи от мембраны. Другими словами, она означает всю или часть области трансмембранного белка, исключая область, включенную в мембрану (трансмембранная область), и область, существующую в цитоплазме, следом за трансмембранной областью (цитоплазматическая область).
«Гибридный полипептид», включенный в указанное выше «белковое вещество», означает гибридный полипептид, включающий всю или часть внеклеточной области полипептида, составляющего AILIM (предпочтительно, человеческую AILIM), и «всю или часть константной области тяжелой цепи иммуноглобулина (Ig, предпочтительно, человеческого Ig)». Предпочтительно, гибридный полипептид представляет собой гибридный полипептид, имеющий внеклеточную область AILIM и часть константной области тяжелой цепи человеческого IgG, и особенно предпочтительно, гибридный полипептид включает внеклеточную область AILIM и область (Fc) тяжелой цепи человеческого IgG, включающую шарнирную область, домен СН2 и домен СН3. В качестве IgG предпочтителен IgG1, а в качестве AILIM предпочтительна человеческая, мышиная или крысиная AILIM (предпочтительно, человеческая).
Используемое здесь выражение «вся или часть константной области тяжелой цепи иммуноглобулина (Ig)» означает константную область области Fc тяжелой цепи иммуноглобулина (цепи Н), полученного у человека, или ее часть. Иммуноглобулин может представлять собой любой иммуноглобулин, относящийся к любому классу и любому подклассу. В частности, иммуноглобулин включает IgG (IgG1, IgG2, IgG3 и IgG4), IgM, IgA (IgA1 и IgA2), IgD и IgE. Предпочтительно, иммуноглобулин представляет собой IgG (IgG1, IgG2, IgG3 или IgG4) или IgM. Примерами особенно предпочтительных иммуноглобулинов настоящего изобретения являются иммуноглобулины, относящиеся к IgG (IgG1, IgG2, IgG3 или IgG4), полученными у человека.
Иммуноглобулин имеет Y-образную структурную единицу, в которой 4 цепи, составленные из 2 гомологичных легких цепей (L-цепей) и двух гомологичных тяжелых цепей (Н-цепей) соединены посредством дисульфидных связей (S-S связей). Легкая цепь составлена из вариабельной области легкой цепи (VL) и константной области легкой цепи (CL). Тяжелая цепь составлена из вариабельной области тяжелой цепи (VН) и константной области тяжелой цепи (CН).
Константная область тяжелой цепи составлена из нескольких доменов, имеющих аминокислотные последовательности, уникальные для каждого класса (IgG, IgM, IgA, IgD и IgE) и каждого подкласса (IgG1, IgG2, IgG3 и IgG4, IgA1 и IgA2).
Тяжелая цепь IgG (IgG1, IgG2, IgG3 и IgG4) составлена из VH, домена СН1, шарнирной области, домена СН2 и домена СН3 в указанном порядке от N-конца.
Аналогичным образом, тяжелая цепь IgG1 составлена из VH, домена Сγ11, шарнирной области, домена Сγ12 и домена Сγ13 в указанном порядке от N-конца. Тяжелая цепь IgG2 составлена из VH, домена Сγ21, шарнирной области, домена Сγ22 и домена Сγ23 в указанном порядке от N-конца. Тяжелая цепь IgG3 составлена из VH, домена Сγ31, шарнирной области, домена Сγ32 и домена Сγ33 в указанном порядке от N-конца. Тяжелая цепь IgG4 составлена из VH, домена Сγ41, шарнирной области, домена Сγ42 и домена Сγ43 в указанном порядке от N-конца.
Тяжелая цепь IgA составлена из VH, домена Сα1, шарнирной области, домена Сα2 и домена Сα3 в указанном порядке от N-конца.
Аналогичным образом, тяжелая цепь IgA1 составлена из VH, домена Сα11, шарнирной области, домена Сα12 и домена Сα13 в указанном порядке от N-конца. Тяжелая цепь IgA2 составлена из VH, домена Сα21, шарнирной области, домена Сα22 и домена Сα23 в указанном порядке от N-конца.
Тяжелая цепь IgD составлена из VH, домена Сδ1, шарнирной области, домена Сδ2 и домена Сδ3 в указанном порядке от N-конца.
Тяжелая цепь IgM составлена из VH, домена Сμ1, домена Сμ2, домена Сμ3 и домена Сμ4 в указанном порядке от N-конца и не имеет шарнирной области, которая наблюдается у IgG, IgA и IgD.
Тяжелая цепь IgE составлена из VH, домена Сε1, домена Сε2, домена Сε3 и домена Сε4 в указанном порядке от N-конца и не имеет шарнирной области, которая наблюдается у IgG, IgA и IgD.
Если, например, IgG обрабатывают папаином, он расщепляется у слегка N-концевой стороны за пределами дисульфидных связей, существующих в шарнирной области, где дисульфидные связи соединяют две тяжелых цепи, с образованием двух гомологичных Fab, в которых фрагмент тяжелой цепи, составленный из VH и CH1, соединен с одной легкой цепью посредством дисульфидной связи; и одного Fc, в котором два гомологичных фрагмента тяжелой цепи, составленные из шарнирной области, домена СН2 и домена СН3, соединены посредством дисульфидных связей (см. "Immunology Illustrated", original 2nd ed., Nankodo, pp.65-75 (1992); и "Focus of Newest Medical Science 'Recognition Mechanism of Immune System'", Nankodo, pp.4-7 (1991); и т.д.).
А именно, указанная выше «часть константной области тяжелой цепи иммуноглобулина» означает часть константной области тяжелой цепи иммуноглобулина, имеющую структурные характеристики, как указано выше, и, предпочтительно, представляет собой константную область без домена С1 или область Fc. В частности, ее примером является область, составленная из шарнирной области, домена С2 и домена С3 из каждого из IgG, IgA и IgD, или представляет собой область, составленную из домена С2, домена С3 и домена С4 из каждого из IgM и IgE. Особенно предпочтительным ее примером является область Fc IgG1, полученного у человека.
Указанный выше гибридный полипептид имеет преимущество крайне легкой очистки путем использования аффинной колоночной хроматографии с использованием свойства белка А, который специфично связывается с фрагментом иммуноглобулина, потому что гибридный полипептид настоящего изобретения имеет часть константной области (например, Fc) иммуноглобулина, такого как IgG, как указано выше, в качестве партнера по слиянию. Кроме того, поскольку имеются различные антитела против Fc различных иммуноглобулинов, можно легко выполнить иммунологический анализ для выявления гибридных полипептидов с помощью антител против Fc.
«Полипептид, который связывается с AILIM», охватывается «полипептидом», включенным в определение указанного выше «вещества».
Конкретным примером «полипептида, который связывается с AILIM», является весь или часть полипептида, составляющего известную молекулу, называемую B7h, B7RP-1, GL50 или LICOS, которая представляет собой лиганд, взаимодействующий с AILIM (Nature, Vol.402, №6763, pp.827-832, 1999; Nature Medicine, Vol.5, №12, pp.1365-1369, 1999; J. Immunology, Vol.164, pp.1653-1657, 2000; Curr. Biol., Vol.10, №6, pp.333-336, 2000).
Предпочтительно, полипептид представляет собой полипептид, включающий всю или часть внеклеточной области указанного выше лиганда (B7h, B7RP-1, GL50, LICOS), или гибридный полипептид, включающий полипептид, и всю или часть константной области тяжелой цепи иммуноглобулина (предпочтительно, человеческого иммуноглобулина). В настоящем описании выражения «внеклеточная область» и «константная область тяжелой цепи иммуноглобулина» имеют такие же значения, как указано выше.
Указанные выше полипептиды, части полипептида (фрагменты) и гибридные полипептиды можно получить не только технологией рекомбинантной ДНК, как указано ниже, но также способом, хорошо известным в данной области, таким как способ химического синтеза или способ культуры клеток или модификации указанных способов.
«Антитело» настоящего изобретения может представлять собой поликлональное антитело (антисыворотку) или моноклональное антитело против AILIM млекопитающих (особенно предпочтительно, человеческой AILIM), определенный выше, и предпочтительно, моноклональное антитело.
В частности, антитело представляет собой антитело, обладающее активностью ингибирования пролиферации клеток, экспрессирующих AILIM, путем связывания с AILIM, или ингибирования продукции интерферона-γ или интерлейкина-4 клетками, экспрессирующими AILIM, посредством связывания с AILIM.
Антитела настоящего изобретения могут быть природными антителами, полученными иммунизацией млекопитающих, таких как мыши, крысы, хомяки, морские свинки и кролики, антигеном, таким как природные клетки, клеточные линии, опухолевые клетки и т.д., экспрессирующие AILIM настоящего изобретения, трансформанты, полученные с использованием технологии рекомбинантной ДНК с тем, чтобы избыточно экспрессировать AILIM на их поверхности, полипептиды, составляющие AILIM, или указанные выше гибридные полипептиды, включающие полипептид AILIM или внеклеточную область AILIM. Антитела настоящего изобретения также включают химерные антитела и гуманизированные антитела (CDR-привитые антитела), которые можно получить технологией рекомбинантной ДНК, и антитела человека, которые можно получить с использованием трансгенных животных, продуцирующих человеческие антитела.
Моноклональные антитела включают антитела, имеющие любой изотип IgG, IgM, IgA, IgD или IgE. Предпочтителен IgG или IgM.
Поликлональное антитело (антисыворотки) или моноклональное антитело можно получить известными способами. А именно, млекопитающее, предпочтительно мышь, крысу, хомяка, морскую свинку, кролика, кошку, собаку, свинью, козу, лошадь или корову, или предпочтительнее мышь, крысу, хомяка, морскую свинку или кролика иммунизируют, например, указанным выше антигеном с адъювантом Фрейнда, если необходимо.
Поликлональное антитело можно получить из сыворотки, полученной у животного, иммунизированного таким образом. Кроме того, моноклональные антитела получают следующим образом. Получают гибридомы из продуцирующих антитела клеток, полученных от иммунизированного таким образом животного, и клеток миеломы, которые не способны продуцировать аутоантитела. Гибридомы клонируют и проводят скрининг клонов, продуцирующих моноклональные антитела, проявляющие специфическое сродство к антигену, использованному для иммунизации млекопитающего.
В частности, моноклональное антитело можно получить следующим образом. Иммунизации проводят инъекцией или имплантацией один или несколько раз указанного выше антигена в качестве иммуногена, если необходимо, с адъювантом Фрейнда, подкожно, внутримышечно, внутривенно, через подушечку лапы или внутрибрюшинно млекопитающему, кроме человека, в частности мыши, крысе, хомяку, морской свинке или кролику, предпочтительно мыши, крысе или хомяку (включая трансгенное животное, созданное с тем, чтобы продуцировать антитела, полученные от другого животного, такого как указанная ниже трансгенная мышь, продуцирующая человеческое антитело). Обычно иммунизацию проводят от одного до четырех раз, каждый раз на 14 день после первой иммунизации. Клетки, продуцирующие антитела, получают от млекопитающего, иммунизированного таким образом, приблизительно через 1-5 дней после последней иммунизации. Частоту и интервал между иммунизациями можно соответствующим образом подобрать в зависимости от, например, свойства используемого иммуногена.
Гибридомы, которые секретируют моноклональное антитело, можно получить способом Kohler и Milstein (Nature, Vol.256, pp.495-497 (1975)) или его модификацией. А именно, гибридомы получают слиянием продуцирующих антитела клеток, содержащихся в селезенке, лимфоузлах, костном мозге или небных миндалинах, полученных у млекопитающего, кроме человека, иммунизированного, как указано выше, предпочтительно, из селезенки, с миеломами без способности продуцировать аутоантитела, которые получены, предпочтительно, у млекопитающего, такого как мышь, крыса, морская свинка, хомяк, кролик или человек, или, предпочтительнее, мышь, крыса или человек.
Например, в качестве миеломы для слияния клеток можно использовать миелому мыши P3/X63-AG8.653 (653), P3/NSI/1-Ag4-1 (NS-1), P3/X63-Ag8.U1 (P3U1), SP2/0-Ag14 (Sp2/0, Sp2), PAI, F0, NSO или BW5147, миелому крысы 210RCY3-Ag.2.3, или миелому человека U-266AR1, GM1500-6TG-A1-2, UC729-6, CEM-AGR, D1R11 или СЕМ-Т15.
Скрининг гибридом, продуцирующих моноклональные антитела, можно проводить культивированием гибридом, например, в микротитрационных планшетах и измерением реактивности надосадочной жидкости культуры в лунках, в которых наблюдается рост гибридомы, на иммуноген, используемый для указанной выше иммунизации, например, ферментным иммунологическим анализом, таким как радиоиммуноанализ (RIA) и твердофазный иммуноферментный анализ (ELISA).
Моноклональные антитела можно получить из гибридом культивированием гибридом in vitro или in vivo, таких как асцитическая жидкость мыши, крысы, морской свинки, хомяка или кролика, предпочтительно, мыши или крысы, предпочтительнее, мыши, и выделением антител из полученной надосадочной жидкости культуры или асцитической жидкости млекопитающего.
Культивирование гибридом in vitro можно проводить в зависимости от, например, свойства клеток, которые предстоит культивировать, цели исследования и различных условий способа культивирования, используя известные питательные среды или любые питательные среды, полученные из известных основных сред для роста, поддержания и хранения гибридом для получения моноклональных антител в надосадочной жидкости культуры.
Примерами основных сред являются среды с низкой концентрацией кальция, такие как среда Ham'F12, среда MCDB153 или среда МЕМ с низкой концентрацией кальция, и среды с высокой концентрацией кальция, такие как среда MCDB104, среда МЕМ, среда D-MEM, среда RРMI1640, среда ASF104 или среда RD. Основные среды могут содержать, например, сыворотки, гормоны, цитокины и/или различные неорганические и органические вещества, в зависимости от цели.
Моноклональные антитела можно выделить и очистить из указанной выше надосадочной жидкости культуры или асцитической жидкости осаждением насыщенным сульфатом аммония, способом осаждения эуглобулином, способом капроновой кислоты, способом каприловой кислоты, ионообменной хроматографией (DEAE или DE52) и аффинной хроматографией с использованием антииммуноглобулиновой колонки или колонки с белком А.
«Рекомбинантное химерное моноклональное антитело» представляет собой моноклональное антитело, полученное приемами генетической инженерии и, в частности, означает химерное антитело, такое как мышиное/человеческое химерное моноклональное антитело, вариабельные области которого происходят из иммуноглобулина млекопитающего, кроме человека (мыши, крысы, хомяка и т.д.), а константная область которого происходит из иммуноглобулина человека.
Константная область, происходящая из иммуноглобулина человека, имеет аминокислотную последовательность, уникальную для каждого изотипа, такого как IgG (IgG1, IgG2, IgG3, IgG4), IgM, IgA, IgD и IgE. Константная область рекомбинантного химерного моноклонального антитела может представлять собой константную область иммуноглобулина человека, принадлежащего к любому изотипу. Предпочтительно, она представляет собой константную область человеческого IgG.
Химерное моноклональное антитело можно получить, например, следующим образом. Нет необходимости говорить, что способ получения не ограничивается им.
Мышиное/человеческое химерное моноклональное антитело можно получить в соответствии со способом, описанным в публикации Experimental Medicine: SUPPLEMENT, Vol.1.6, №10 (1988) и рассмотренной опубликованной заявке на патент Японии №(JP-B) Hei 3-73280. А именно, его можно получить, оперативно вставляя ген СН (ген С, кодирующий константную область Н-цепи), полученный из ДНК, кодирующей иммуноглобулин человека, «правее» активных генов VH (перестроенного гена VDJ, кодирующего вариабельную область Н-цепи), полученных из ДНК, кодирующей мышиное моноклональное антитело, выделенной из гибридомы, продуцирующей мышиное моноклональное антитело, и гена CL (гена С, кодирующего константную область L-цепи), полученного из ДНК, кодирующей иммуноглобулин человека, «правее» активных генов VL (перестроенного гена VJ, кодирующего вариабельную область L-цепи), полученных из ДНК, кодирующей мышиное моноклональное антитело, выделенной из гибридомы, в один и тот же вектор или разные векторы экспрессируемым образом с последующей трансформацией клеток-хозяев вектором экспрессии, а затем культивированием трансформантов.
Конкретно, ДНК сначала экстрагируют из гибридом, продуцирующих мышиные монокланоальные антитела, обычным способом, расщепляют соответствующими рестрикционными ферментами (например, EcoRI и HindIII), подвергают электрофорезу (с использованием, например, 0,7% агарозного геля) и анализируют Саузерн-блоттингом. После окрашивания подвергнутого электрофорезу геля, например, этидием бромидом, и его фотографирования в геле устанавливают положения маркеров, гель дважды промывают водой и вымачивают в 0,25 М HCl в течение 15 минут. Затем гель вымачивают в 0,4 н. растворе NaOH в течение 10 минут при осторожном перемешивании. ДНК переносят на фильтр в течение 4 часов обычным способом. Фильтр извлекают и промывают дважды раствором 2×SSC. После достаточной сушки фильтра его прогревают при 75°С в течение 3 часов. После прогрева фильтр обрабатывают 0,1×SSC/0,1% раствором додецилсульфата натрия (SDS) при 65°С в течение 30 минут. Затем его вымачивают в 3×SSC/0,1% SDS. Полученный фильтр обрабатывают предгибридизационным раствором в пластиковом мешке при 65°С в течение 3-4 часов.
Затем меченый 32Р ДНК-зонд и раствор для гибридизации добавляют в мешок и проводят реакцию при 65°С в течение 12 часов. После гибридизации фильтр промывают при соответствующей концентрации соли, температуре и времени реакции (например, 2×SSC/0,1% SDS, комнатной температуре, 10 минут). Фильтр помещают в пластиковый мешок с небольшим объемом 2×SSC и подвергают авторадиаографии после герметичного укупоривания мешка.
Перестроенный ген VDJ и ген VJ, кодирующие Н-цепь и L-цепь мышиного моноклонального антитела, идентифицируют указанным выше саузерн-блоттингом. Область, включающую идентифицированный фрагмент ДНК, фракционируют центрифугированием в градиенте плотности сахарозы и производят инсерцию в фаговый вектор (например, Charon 4A, Charon 28, λEMBL3 и λEMBL4). E. coli (например, LE392 и NM539) трансформируют фаговым вектором с получением геномной библиотеки. Скрининг геномной библиотеки проводят методикой гибридизации бляшек, такой как способ Benton-Davis (Science, Vol.196, pp.180-182 (1977)), с использованием соответствующих зондов (ген J Н-цепи, ген J L-цепи (κ) и т.д.) для получения положительных клонов, включающих перестроенный ген VDJ или ген VJ. Получением рестрикционной карты и определением нуклеотидной последовательности полученных клонов подтверждают, были ли получены гены, включающие желаемый, перестроенный ген VH (VDJ) или VL (VJ).
Отдельно выделяют человеческий ген СН и человеческий ген CL, используемые для химеризации. Например, когда получают химерное антитело с человеческим IgG1, ген Сγ1 выделяют в качестве гена СН, а ген Сκ - в качестве гена CL.Указанные гены можно выделить из человеческой геномной библиотеки с помощью мышиного гена Cγ1 и мышиного гена Сκ, соответствующих человеческому гену Cγ1 и человеческому гену Сκ, соответственно, в качестве зондов, используя преимущество высокой гомологии между нуклеотидными последовательностями гена мышиного иммуноглобулина и гена человеческого иммуноглобулина.
Конкретно, фрагменты ДНК, включающие человеческий ген Сκ и область энхансера выделяют из человеческой геномной библиотеки λ Charon 4А HaeIII-AluI (Cell, Vol.15, pp.1157-1174 (1978)), например, с использованием HindIII-BamHI-фрагмента размером 3 т.п.н. клона Ig146 (Proc. Natl. Acad. Sci. USA, Vol.75, pp.4709-4713 (1978)) и EcoRI-фрагмента размером 6,8 т.п.н. клона МЕР10 (Proc. Natl. Acad. Sci. USA, Vol.78, pp.474-478 (1981)) в качестве зондов. Кроме того, например, после расщепления ДНК человеческих плодных гепатоцитов с помощью HindIII и фракционирования электрофорезом в агарозном геле, производят инсерцию фрагмента размером 5,9 т.п.н. в λ788, а затем человеческий ген Сγ1 выделяют с помощью указанных выше зондов.
Используя полученный таким образом мышиный ген VH, мышиный ген VL, человеческий ген СН и человеческий ген CL, и учитывая область промотора и область энхансера, производят инсерцию человеческого гена СН «правее» мышиного гена VH, а инсерцию человеческого гена СL производят «правее» мышиного гена VL в вектор экспрессии, такой как pSV2gpt или pSV2neo, с помощью соответствующих рестрикционных ферментов и ДНК-лигазы обычным способом. В данном случае можно произвести инсерцию химерных генов мышиного гена VH/человеческого гена СН и мышиного гена VL/человеческого гена СL, соответственно, в один и тот же вектор экспрессии или в различные векторы экспрессии.
Полученный таким образом вектор (векторы) экспрессии с инсерцией химерных генов вводят в миеломы, которые не продуцируют антитела, например, клетки P3X63·Ag8·653 или клетки SP210, способом слияния протопласта, способом с DEAE-декстраном, способом с фосфатом кальция или способом электропорации. Проводят скрининг трансформантов культивированием в средах, содержащих лекарственное средство, соответствующее гену устойчивости к лекарственному средству, введенного в вектор экспрессии, а затем получают клетки, продуцирующие желательные химерные моноклональные антитела.
Желательные химерные моноклональные антитела получают из надосадочной жидкости культуры продуцирующих антитела клеток, выявленных таким скринингом.
«Гуманизированное моноклональное антитело (CDR-привитое антитело)» настоящего изобретения представляет собой моноклональное антитело, полученное методами генетической инженерии, и, в частности, означает гуманизированное моноклональное антитело, в котором области, определяющие комплементарность, гипервариабельной области или их части происходят из областей, определяющих комплементарность, гипервариабельной области моноклонального антитела млекопитающего, кроме человека (мыши, крысы, хомяка и т.д.), каркасные участки происходят из каркасных участков вариабельной области из человеческого иммуноглобулина, а константная область происходит из константной области иммуноглобулина человека.
Определяющие комплементарность области гипервариабельной области существуют в гипервариабельной области в вариабельной области антитела и означают три области, которые непосредственно и комплементарно связываются с антигеном (остатки, определяющие комплементарность, CDR1, CDR2 и CDR3). Каркасные участки вариабельной области означают четыре сравнительно консервативных области, лежащие «левее», «правее» или между тремя областями, определяющими комплементарность, (каркасный участок, FR1, FR2, FR3 и FR4).
Другими словами, гуманизированное моноклональное антитело означает такое антитело, в котором все области, за исключением определяющих комплементарность областей гипервариабельной области моноклонального антитела, полученного у млекопитающего, кроме человека или их части, были замещены их соответствующими областями, полученными из иммуноглобулина человека.
Константная область, происходящая из иммуноглобулина человека, имеет аминокислотную последовательность, уникальную для каждого изотипа, такого как IgG (IgG1, IgG2, IgG3, IgG4), IgM, IgA, IgD и IgE. Константная область гуманизированного моноклонального антитела в настоящем изобретении может быть областью из иммуноглобулина человека, относящегося к любому изотипу. Предпочтительно, она представляет собой константную область человеческого IgG. Каркасные участки константной области, происходящие из иммуноглобулина человека, конкретно не ограничены.
Гуманизированное моноклональное антитело можно получить, например, следующим образом. Нет необходимости говорить, что способ получения не ограничивается им.
Например, рекомбинантное гуманизированное моноклональное антитело, происходящее из мышиного моноклонального антитела, можно получить методами генетической инженерии со ссылкой на опубликованный перевод на японский язык Международной публикации (JP-WA) №Hei 4-506458 и JP-A Sho 62-296890. А именно, по меньшей мере, один ген CDR мышиной Н-цепи и, по меньшей мере, один ген CDR мышиной L-цепи, соответствующий гену CDR мышиной Н-цепи, выделяют из гибридом, продуцирующих мышиное моноклональное антитело, а ген Н-цепи человека, кодирующий все области, за исключением CDR человеческой Н-цепи, соответствующей указанному выше CDR мышиной Н-цепи, и ген человеческой L-цепи, кодирующий всю область, за исключением указанного выше CDR человеческой L-цепи, соответствующий указанному выше CDR мышиной L-цепи, выделяют из генов иммуноглобулина человека.
Оперативно встраивают выделенные таким образом ген (гены) CDR мышиной Н-цепи и ген (гены) человеческой Н-цепи в соответствующий вектор таким образом, чтобы они могли экспрессироваться. Аналогичным образом, оперативно встраивают ген (гены) CDR мышиной L-цепи и ген (гены) человеческой L-цепи в соответствующий другой вектор таким образом, чтобы они могли экспрессироваться. Альтернативно, можно оперативно встроить ген (гены) CDR мышиной Н-цепи/ген (гены) человеческой Н-цепи и ген (гены) CDR мышиной L-цепи/ген (гены) человеческой L-цепи в один и тот же вектор экспрессии экспрессируемым образом. Клетки-хозяева трансформируют полученными таким образом вектором экспрессии с получением трансформантов, продуцирующих гуманизированное моноклональное антитело. Культивированием трансформантов из надосадочной жидкости культуры получают желательное гуманизированное моноклональное антитело.
«Человеческое моноклональное антитело» представляет собой иммуноглобулин, в котором все области, включающие вариабельную и константную область Н-цепи, и вариабельную и константную область L-цепи, составляющих иммуноглобулин, происходят из генов, кодирующих иммуноглобулин человека.
Человеческое антитело (предпочтительно, человеческое моноклональное антитело) можно получить хорошо известными способами, например, таким же образом, как способ получения указанных выше поликлональных или моноклональных антител иммунизацией антигеном трансгенного животного, полученного интегрированием, по меньшей мере, гена человеческого иммуноглобулина в локус гена млекопитающего, кроме человека, такого как мышь.
Например, трансгенную мышь, продуцирующую человеческие антитела, получают способами, описанными в Nature Genetics, Vol.7, pp.13-21 (1994); Nature Genetics, Vol.15, pp.146-156 (1997); JP-WA Hei 4-504365; JP-WA Hei 4-509137; Nikkei Sciences, №6, pp.40-50 (1995); WO 94/25585; Nature, Vol.368, pp.856-859 (1994) и JP-WA №Hei 6-500233.
Кроме того, можно также применять недавно разработанную методику получения белка, полученного у человека, из молока трансгенной коровы или свиньи (Nikkei Science, pp.78-84 (April, 1997)).
Используемое в настоящем изобретении выражение «часть антитела» означает частичную область моноклонального антитела, как указано выше. Оно, в частности, означает F(ab')2, Fab', Fab, Fv (вариабельный фрагмент антитела), sFv, dsFv (Fv, стабилизированный дисульфидом) или dAb (антитело с одним доменом) (Exp. Opin. Ther. Patents, Vol.6, №5, pp.441-456 (1996)).
F(ab')2 и Fab' можно получить обработкой иммуноглобулина (моноклонального антитела) протеазой, такой как пепсин и папаин, и означает фрагмент антитела, образованный расщеплением иммуноглобулина около дисульфидных связей в шарнирных областях, существующих между каждыми из двух Н-цепей. Например, папаин расщепляет IgG выше дисульфидных связей в шарнирных областях, существующих между каждыми из двух Н-цепей, с образованием двух гомологичных фрагментов антитела, в которых L-цепь, составленная из VL (вариабельной области L-цепи) и CL (константной области L-цепи), и фрагмент Н-цепи, составленный из VH (вариабельной области Н-цепи) и области CHγ1 (участок γ1 в константной области Н-цепи), соединены у их С-концевых областей посредством дисульфидной связи. Каждый из таких двух гомологичных фрагментов антитела называется Fab'. Пепсин также расщепляет IgG ниже дисульфидных связей в шарнирных областях, существующих между каждыми из двух Н-цепей, с образованием фрагмента антитела, немного большего, чем фрагмент, в котором два указанных выше Fab' соединены в шарнирной области. Данный фрагмент антитела называется F(ab')2.
Выражение «иммунитет кишечного тракта», «желудочно-кишечный иммунитет» и «иммунитет слизистой оболочки» данного изобретения используют для выражения почти одинакового значения.
Предпочтительным примером «заболевания, сопровождающего патологический иммунитет кишечного тракта» данного изобретения может быть воспалительное заболевание кишечника или пищевая аллергия.
Репрезентативным примером «воспалительного заболевания кишечника» данного изобретения является колит (в частности, язвенный колит (UC)) или болезнь Крона (CD), каждое из которых имеет, например, следующие характеристики.
Воспалительные заболевания кишечника (IBD) можно классифицировать на колит (в частности, язвенный колит) и болезнь Крона. Указанные заболевания часто развиваются у лиц молодого возраста и считаются трудноизлечимыми хроническими воспалительными заболеваниями, которые характеризуются повторными ремиссиями и рецидивами, и их причины неизвестны.
Болезнь Крона представляет собой заболевание, при котором хронические грануломатозные воспаления и язвы возникают во всем пищеварительном тракте от пищевода до анального отверстия, главным образом, в тонкой кишке и толстой кишке, характеризующееся такими симптомами, как боль в животе, диарея, лихорадка, патология области анального отверстия, включая геморрой, и/или уменьшение веса тела. Гистологически наблюдается выраженная лимфоцитарная инфильтрация и нетворожистая эпителиоидная гранулома, свидетельствуя о патологической реакции Т-клеток и антиген-представляющих клеток.
Колит (в частности, язвенный колит) представляет собой хроническое воспаление, которое развивается местно в толстой кишке. Оно, главным образом, поражает слизистую оболочку и образует ссадины и изъязвления. Гистологически наблюдаются значительная инфильтрация лимфоцитами, плазматическими клетками, макрофагами и тучными клетками в слизистой оболочке и в основном слое слизистой и язвы с криптами, сопровождающие инфильтрацию нейтрофилами, и происходит исчезновение бокаловидных клеток.
Существующие фармацевтические средства, применяемые для лечения воспалительных заболеваний кишечника данного изобретения, относятся к одному или более произвольным фармацевтическим средствам, назначаемым в клинических условиях для лечения колита (язвенного колита и ему подобного) и болезни Крона, и примерами являются гормоны коры надпочечников и салазосульфапиридин.
Выражение «фармацевтически приемлемый носитель» настоящего изобретения включает основу, разбавитель, наполнитель, разрыхлитель, стабилизатор, консервант, буфер, эмульгатор, отдушку, краситель, подслащивающее вещество, средство увеличения вязкости, корригент, средство, увеличивающее растворимость, или несколько других добавок. С использованием одного или более таких носителей фармацевтическую композицию можно составить в таблетки, пилюли, порошки, гранулы, растворы для инъекций, растворы, капсулы, пастилки, эликсиры, суспензии, эмульсии, сиропы и т.д.
Фармацевтическую композицию можно вводить перорально или парентерально. Другие виды парентерального введения включают раствор для наружного нанесения, суппозиторий для ректального введения и пессарий, назначаемый обычным способом, включающие один или более активных ингредиентов.
Дозировка может варьировать в зависимости от возраста, пола, веса тела и симптомов у пациента, эффекта лечения, пути введения, периода лечения, вида активного ингредиента (указанного выше «вещества» в соответствии с настоящим изобретением), содержащегося в фармацевтической композиции, и т.д. Обычно фармацевтическую композицию можно вводить взрослому человеку в дозе от 10 мкг до 1000 мг (или от 10 мкг до 500 мг) на одно введение. В зависимости от различных условий в некоторых случаях может быть достаточна дозировка меньшая, чем указанная выше, а у других может быть необходима дозировка большая, чем указанная выше.
В случае раствора для инъекций его можно изготовить растворением или суспендированием антитела в нетоксичном, фармацевтически приемлемом носителе, таком как физиологический солевой раствор или имеющаяся в продаже дистиллированная вода для инъекций, доводя концентрацию до диапазона от 0,1 мкг антитела/мл носителя до 10 мг антитела/мл носителя. Изготовленный таким образом раствор для инъекций можно вводить пациенту-человеку, нуждающемуся в лечении, в диапазоне доз от 1 мкг до 100 мг/кг веса тела, предпочтительно, в диапазоне от 50 мкг до 50 мг/кг веса тела, один или более раз/день. Примерами путей введения являются целесообразные с медицинской точки зрения пути введения, такие как внутривенная инъекция, подкожная инъекция, внутрикожная инъекция, внутримышечная инъекция, внутрибрюшинная инъекция или им подобные, предпочтительно, внутривенная инъекция.
Раствор для инъекций можно также изготовить в неводном растворителе (например, пропиленгликоле, полиэтиленгликоле, растительном масле, таком как оливковое масло, и спирте, таком как этанол), в виде суспензии или эмульсии.
Раствор для инъекций можно стерилизовать фильтрацией через фильтр, фильтрующий бактерии, смешиванием с бактериоцидным средством или облучением. Раствор для инъекций можно получать таким образом, что его готовят во время применения. А именно, его лиофилизируют для того, чтобы стерильная композиция была представлена в виде твердого вещества, которое можно растворить в стерильной дистиллированной воде для инъекций или другом растворителе перед применением.
С использованием фармацевтических композиций настоящего изобретения можно подавлять, предотвращать и/или лечить заболевания, которые могут быть вызваны патологическим иммунитетом кишечного тракта, более конкретно, воспалительных заболеваний кишечника (в частности, болезни Крона и колита (в частности, язвенного колита)) и пищевых аллергий.
Кроме того, с использованием фармацевтической композиции настоящего изобретения можно усилить терапевтический эффект существующих фармацевтических средств, которые назначают для лечения таких воспалительных заболеваний.
Краткое описание чертежей
На фиг.1 показан подавляющий эффект антитела против AILIM на начало воспалительных заболеваний кишечника, когда антитело постоянно вводят (перед началом или после прогрессирования заболевания), и терапевтический эффект на воспалительное заболевание кишечника введения антитела против AILIM, определяемый с использованием в качестве показателя потери веса тела, который характерен для колита.
Фиг.2 представляет собой фотографию, показывающую соответственно состояние толстой кишки здоровых мышей, у которых не развилось воспалительное заболевание кишечника, состояние толстой кишки мышей, пораженных воспалительным заболеванием кишечника, и состояние толстой кишки мышей, у которых было обнаружено, что начало воспалительного заболевания кишечника подавлено введением антитела против AILIM.
На фиг.3 показан подавляющий эффект антитела против AILIM на начало воспалительного заболевания кишечника, когда антитело вводят постоянно, по данным потери веса тела в качестве показателя, характерного для колита.
 : антитело отрицательного контроля (n=20)
: антитело отрицательного контроля (n=20)
 : антитело против AILIM/ICOS (n=20)
: антитело против AILIM/ICOS (n=20)
 : антитело против B7RP-1 (n=7)
: антитело против B7RP-1 (n=7)
 : введение антитела отрицательного контроля мыши BALB/c scid/scid, которой вводили Т-клетки CD4+CD45RBlow вместо Т-клеток CD4+CD45RBhigh (n=7)
: введение антитела отрицательного контроля мыши BALB/c scid/scid, которой вводили Т-клетки CD4+CD45RBlow вместо Т-клеток CD4+CD45RBhigh (n=7)
На фиг.4 показана степень тяжести колита, выраженная в виде гистологических балльных оценок.
На фиг.5 показано количество клеток CD4+, которые инфильтрировали слизистую оболочку толстой кишки (основной слой).
На фиг.6 показана степень апоптоза клеток в тканях толстой кишки.
На фиг.7 показан терапевтический эффект введения антитела против AILIM (введение после прогрессирования заболевания) на воспалительное заболевание кишечника, по данным потери веса тела в качестве показателя, характерного для колита
: антитело отрицательного контроля
: антитело против AILIM/ICOS
На фиг.8 показана степень тяжести колита, выраженная в виде гистологических балльных оценок.
На фиг.9 показано количество клеток CD4+, которые инфильтрировали слизистую оболочку толстой кишки (основной слой).
На фиг.10 показан подавляющий эффект постоянно вводимых антител против AILIM на начало воспалительного заболевания кишечника, по данным потери веса тела в качестве показателя, характерного для колита.
: антитело отрицательного контроля (n=7)
: антитело против AILIM/ICOS (n=7)
На фиг.11 показана степень тяжести колита, выраженная в виде гистологических балльных оценок.
На фиг.12 показано количество клеток CD4+, которые инфильтрировали слизистую оболочку толстой кишки (основной слой).
Лучший способ осуществления изобретения
Далее настоящее изобретение конкретно проиллюстрировано со ссылкой на примеры, но его не следует рассматривать как ограниченное ими.
Пример 1
Терапевтический эффект при колите на модели колита у мыши <Испытание 1>
<1-1> Животные
Использовали мышей BALB/c scid/scid, мышей с тяжелым иммунодефицитом (самки в возрасте 6-8 недель; CLEA Japan) и здоровых мышей BALB/c (самцы в возрасте 6-8 недель; CLEA Japan).
<1-2> Получение моноклонального антитела против AILIM мыши
Получение проводили следующим образом.
С использованием кДНК, кодирующей полноразмерную аминокислотную последовательность мышиной AILIM, о которой сообщали ранее (Int. Immunol., Vol.12, №1, pp.51-55, 2000), получали трансформированную клетку, экспрессирующую мышиную AILIM, в соответствии со стандартными способами с использованием генетической рекомбинантной технологии.
Трансформированную клетку гомогенизировали и подвергали ультрацентрифугированию (100000·g), и осадок после центрифугирования, содержащий фракцию клеточной мембраны, собирали и суспендировали в солевом растворе с фосфатным буфером (PBS). Полученную фракцию клеточной мембраны инъецировали вместе с полным адъювантом Фрейнда в подушечку лапы крысы Wistar для начальной иммунизации (день 0). Кроме того, фракцию клеточной мембраны вводили в качестве антигена в подушечку лапы через интервалы на 7-й день, 14-й день и 28-й день. Через 2 дня после конечной иммунизации собирали клетки лимфоузлов.
Клетки лимфоузлов и клетки мышиной миеломы PAI (JCR №B0113; Res. Disclosure, Vol.217, p.155, 1982) смешивали в соотношении 5:1, и гибридому, продуцирующую моноклональные антитела, получали слиянием клеток с использованием полиэтиленгликоля 4000 (Boehringer Mannheim) в качестве агента слияния. Отбор гибридомы проводили культивированием в среде ASF104, содержащей НАТ (Ajinomoto), содержащей 10% фетальной бычьей сыворотки и аминоптерин.
Измеряли величины интенсивности флюоресценции клеток, окрашенных обеспечением реакции надосадочных жидкостей культур каждой гибридомы с указанными выше трансфицированными клетками, экспрессирующими рекомбинантную мышиную AILIM, а затем обеспечением их реакции с меченым FITC анти-крысиным IgG (Cappe1), с использованием проточного цитометра EPICS-ELITE для подтверждения реактивности моноклональных антител, продуцированных в надосадочной жидкости культуры каждой гибридомы, против мышиной AILIM. В результате получили несколько гибридом, которые продуцировали моноклональные антитела, обладающие реактивностью в отношении мышиной AILIM.
Одну из указанных гибридом назвали «В10.5». Данную гибридому (каждую от 106 до 107 клеток/0,5 мл/мышь) инъецировали внутрибрюшинно мыши ICR nu/nu (самка в возрасте от 7 до 8 недель). Через 10-20 дней у мыши под анестезией проводили лапаротомию, и большое количество моноклонального антитела против мышиной AILIM (IgG2a) получали из асцитической жидкости в соответствии со стандартными процедурами. Далее указанное антитело просто называют «антителом против AILIM».
<1-3> Индукция воспалительного заболевания кишечника
Как описано ниже, воспалительное заболевание кишечника (колит) вызывали введением CD45RBhigh мышам BALB/c scid/scid в соответствии с ранее описанным способом. Известно, что сопровождая начало и прогрессирование воспалительного заболевания кишечника, через 3-5 недель после введения Т-клеток CD45RBhigh на данной модели воспалительного заболевания кишечника происходит значимая потеря веса тела.
Т-клетки CD4+ выделяли и получали из мононуклеарных клеток, полученных из селезенок здоровых мышей BALB/c, с использованием устройства магнитного разделения MACS (Miltenyi Biotec) c антителом против CD4 (L3T4). Более конкретно, клетки селезенки, выделенные у мышей, культивировали при 4°С в течение 30 минут с магнитными бусинами, с которыми связано антитело против CD4, а затем указанные клетки промывали и обогащали пропусканием через магнитную проточную колонку.
После метки полученных Т-клеток CD4+ (чистоту подтверждали на уровне 96-97% с использованием проточного цитометра) антителом против мышиного CD4 (RM4-5; PharMingen), меченым фикоэритрином (РЕ), и антителом против CD4 (16А; PharMingen), меченым флюоресцеином изотиоцианатом (FITC), их сортировали с использованием FASC Vantage (Becton Dickinson) и фракционировали на Т-клетки, имеющие высокую экспрессию CD45RB (CD45RBhigh), и Т-клетки, имеющие низкую экспрессию CD45RB (CD45RBlow).
Затем для того, чтобы вызвать колит у мышей BALB/c scid/scid, полученные Т-клетки CD45RBhigh (5·105 клеток/200 мкл PBS) вводили внутрибрюшинно (в/б) мышам.
<1-4> Введение антитела против AILIM
Каждую группу указанных выше мышей SCID, которым вводили Т-клетки CD45RBhigh, обрабатывали следующим образом.
Группа 1
Антитело против AILIM (250 мкг/250 мкл PBS) вводили внутрибрюшинно непосредственно после введения Т-клеток CD45RBhigh (первое введение), а затем постоянно каждую неделю с частотой 3 раза/неделю.
Группа 2
Антитело отрицательного контроля (крысиный IgG, Sigma, 250 мкг/250 мкл PBS) вводили внутрибрюшинно непосредственно после введения Т-клеток CD45RBhigh (первое введение), а затем постоянно каждую неделю с частотой 3 раза/неделю. Одновременно, после 8-й недели непосредственно после введения Т-клеток CD45RBhigh (первое введение) и в последующем, в дополнение к антителу отрицательного контроля внутрибрюшинно вводили антитело против AILIM (250 мкг/250 мкл PBS) постоянно каждую неделю c аналогичной частотой.
Группа 3
Антитело отрицательного контроля (крысиный IgG, Sigma, 250 мкг/250 мкл PBS) вводили внутрибрюшинно непосредственно после введения Т-клеток CD45RBhigh (первое введение), а затем постоянно каждую неделю с частотой 3 раза/неделю.
Степень прогрессирования воспалительного заболевания кишечника и степень подавления и лечения начавшегося и прогрессирующего заболевания благодаря антителу против AILIM анализировали измерение веса тела в каждой группе с течением времени с момента непосредственно перед введением Т-клеток.
Результаты показаны на фиг.1.
Поэтому, как ожидали, значимая потеря веса тела, сопровождающая прогрессирование воспалительного заболевания кишечника, произошла в группе, которой вводили только антитело отрицательного контроля (группа 3). Однако в группе, которой постоянно вводили антитело против AILIM с момента непосредственно после введения Т-клеток CD45RBhigh (группа 1) абсолютно никакой потери веса тела не наблюдали, и в этой группе начало воспалительного заболевания кишечника было полностью подавлено.
Кроме того, в группе, которой только антитело отрицательного контроля вводили с момента непосредственно после введения Т-клеток CD45RBhigh до 7-й недели и с 8-й недели вводили антитело против AILIM в дополнение к антителу отрицательного контроля (группа 2), наблюдали значимое увеличение (восстановление) веса тела с момента непосредственно после начала введения антитела против AILIM, в сравнении с группой, которой только антитело отрицательного контроля вводили даже после 8-й недели и далее (группа 3). Поэтому было обнаружено, что антитело против AILIM излечивает воспалительное заболевание кишечника.
Более того, степень прогрессирования воспалительного заболевания кишечника, а также степень подавления и лечения начавшегося и прогрессирующего заболевания антителом против AILIM анализировали взятием толстых кишок через 6 недель после введения Т-клеток у нескольких мышей из группы 1 и группы 3 и исследованием их состояния невооруженным глазом. В качестве нормального контроля проводили аналогичные наблюдения на толстой кишке, взятой у мышей BALB/c scid/scid, которым вводили Т-клетки CD45RBhigh и не вводили антитела.
Результаты показаны на фиг.2.
В результате инволюцию толстой кишки (утолщение и укорочение кишечного тракта), сопровождающую прогрессирование воспалительного заболевания кишечника, и не переработанные фекалии наблюдали в группе, которой вводили антитело отрицательного контроля (группа 3). Однако состояние толстой кишки у группы, которой вводили антитело против AILIM (группа 1), было аналогичным состоянию толстой кишки нормального контроля, и было выявлено, что антитело против AILIM значимо подавляет начало и прогрессирование воспалительного заболевания кишечника.
Пример 2
Терапевтический эффект при воспалительном заболевании кишечника на модели колита у мышей <Испытание 2>
<2-1> Животные
Использовали мышей BALB/c scid/scid с иммунодефицитом, мышей C57BL/6 scid/scid и здоровых мышей BALB/c (все самцы в возрасте 6-8 недель; CLEA Japan).
<2-2> Моноклональные антитела
Использовали моноклональное антитело против мышиной AILIM (ICOS), моноклональное антитело против мышиной B7RP-1, которая является лигандом мышиной AILIM (ICOS), и антитело отрицательного контроля.
В10.5 получали, как описано выше, и использовали для моноклонального антитела против мышиной AILIM/ICOS.
Моноклональное антитело против мышиной B7RP-1 получали следующим образом. Крысу SD иммунизировали рекомбинантными L-клетками, которые экспрессируют мышиную B7RP-1, полученным в соответствии со стандартными процедурами. Получали клетки селезенки иммунизированной крысы, и путем слияния клеток с клетками миеломы в соответствии со стандартными процедурами получали гибридомы. Используя надосадочную жидкость культуры каждой из гибридом, с помощью EIA измеряли степень связывания моноклонального антитела против мышиной B7RP-1, содержащегося в надосадочной жидкости культуры, с рекомбинантными клетками (клетками NRK), которые экспрессируют мышиную B7RP-1, и отбирали гибридомы, продуцирующие антитело, которое связывается с мышиной B7RP-1. Моноклональное антитело против мышиной B7RP-1, использованное для исследования, получали из надосадочной жидкости культуры отобранных гибридом. Кроме того, было подтверждено, что моноклональное антитело против мышиной B7RP-1 обладает активностью ингибирования связывания мышиной AILIM-Ig (гибридного полипептида, включающего растворимую область мышиной AILIM/ICOS и Fc иммуноглобулина) с рекомбинантными клетками, которые экспрессируют мышиную B7RP-1.
Крысиный IgG (Sigma), который не вступает в реакцию с мышиной AILIM/ICOS или B7RP-1, использовали в качестве антитела отрицательного контроля.
<2-3> Индукция колита (воспалительного заболевания кишечника)
Воспалительное заболевание кишечника (колит) вызывали введением Т-клеток CD4+CD45RBhigh мышам BALB/c scid/scid в соответствии с имеющимся сообщением (J. Immunol., Vol.164, pp.4878-4882, 2000).
Т-клетки CD4+ выделяли из клеток селезенки здоровых мышей BALB/c с использованием устройства магнитного разделения MACS (Miltenyi Biotec) c антителом против CD4 (L3T4) в соответствии с прилагаемыми инструкциями. Т-клетки CD4+ (чистота 96-97%, анализировавшаяся FASC) метили антителом против мышиного CD4, меченым РЕ (RM4-5; PharMingen), и антителом против CD45RB, меченым FITC (16А; PharMingen), и их сортировали на Т-клетки CD45RBhigh и Т-клетки CD45RBlow с использованием FASC Vantage (Becton Dickinson).
Воспалительное заболевание кишечника (колит) вызывали внутрибрюшинным введением Т-клеток CD45RBhigh (5Ч105 клеток/200 мкл PBS) мышам BALB/c scid/scid.
<2-4> Лечение воспалительного колита антителом против AILIM
Моноклональное антитело против мышиной AILIM/ICOS (250 мкг/250 мкл PBS), моноклональное антитело против мышиной B7RP-1 (250 мкг/250 мкл PBS) или контрольное антитело (250 мкг/250 мкл PBS) вводили внутрибрюшинно с частотой 3 раза/неделю в течение 7 недель от момента введения (неделя 0) Т-клеток CD45RBhigh мышам, у которых было вызвано развитие воспалительного колита, как описано выше.
С другой стороны, для исследования терапевтического эффекта антитела против AILIM/ICOS в состоянии прогрессирующего воспалительного колита моноклональное антитело против AILIM/ICOS вводили (в концентрации 250 мкг/кг веса тела; 3 раза/неделю в/б) в условиях, аналогичных указанным выше, начиная с 3 недели после введения Т-клеток CD45RBhigh,мышам, у которых было вызвано развитие воспалительного колита, как описано выше. Через 7 недель после введения Т-клеток CD45RBhigh тестируемых животных умерщвляли и анализировали состояние воспаления в толстой кишке.
<2-5> Введение мышам патогенных Т-клеток CD4+ от мышей с колитом
Из основных слоев (LP) толстой кишки мышей BALB/c scid/scid, которым вводили Т-клетки CD4+ CD45RBhigh (адаптационный перенос), выделяли клетки CD4+ (Т-клетки LP CD4+) с использованием указанных выше магнитных бусин MACS через 7 недель после введения Т-клеток. Подтвержденная FASC чистота выделенных Т-клеток LP CD4+ составила 95%.
Т-клетки LP CD4+ (1·106 клеток/200 мкл PBS, в/б) вводили мышам BALB/c scid/scid. Затем тестируемым мышам вводили антитело против AILIM/ICOS (в концентрации 250 мкг/250 мкл PBS) или антитело отрицательного контроля (крысиный IgG, 250 мкг/250 мкл PBS) с частотой 3 раза/неделю. Через 4 недели после введения Т-клеток LP CD4+ производили эвтаназию мышей, удаляли толстую кишку и анализировали гистологически.
<2-6> Гистологическое исследование и иммуногистохимическое окрашивание
Образцы ткани толстой кишки, взятые у каждого из указанных выше тестируемых животных, фиксировали в PBS, содержащей 6% формалин. Залитые в парафин среды ткани толстой кишки (5 мкм) окрашивали гематоксилином и эозином (окрашивание НЕ). Анализировали каждый из полученных срезов ткани. Степень воспаления толстой кишки выражали в балльных оценках в соответствии с существующим сообщением (Gastroenterology, Vol.119, pp.715-723, 2000).
С другой стороны, образцы ткани толстой кишки для анализа иммуногистохимическим окрашиванием фиксировали в соединении орто-хлортолуола (ОСТ), замораживали в жидком азоте и хранили при -80°С. Окрашивание срезов ткани проводили способом с использованием авидин-биотинового комплекса.
Срезы ткани (6 мкм) инкубировали с биотинилированным моноклональным антителом против мышиной AILIM/ICOS, биотинилированным моноклональным антителом против B7RP-1 мыши, биотинилированным моноклональным антителом против мышиного CD4 (RM4-5; крысиный IgG1; PharMingen), биотинилированным моноклональным антителом против F4/80 мыши (крысиный IgG2, PharMingen) или биотинилированным, подобранным по изотипу контрольным антителом (PharMingen). Биотинилированное антитело выявляли комплексом стрептавидина-биотинилированной пероксидазы хрена (набор Vectastain ABC Kit; Вектор) и визуализировали с помощью диаминобензидина. Затем проводили контр-окрашивание каждого среза гематоксилином.
<2-7> Выявление апоптозных клеток
Апоптозные клетки в замороженных срезах ткани выявляли с использованием набора ApoTag Kit (Intergen) в соответствии с ранее опубликованным способом TUNEL. При осмотре под микроскопом определяли количество TUNEL-положительных клеток подсчетом TUNEL-положительных клеток среди 500 мононуклеарных клеток основного слоя (LPMC), которые инфильтрировались в 5 частей одного среза. Процентную долю TUNEL-положительных клеток среди 500 LPMC принимали за показатель апоптоза.
<2-8> Результаты
Результаты показаны на фиг.3-12.
<2-8-1> Подавление колита введением антитела против AILIM/ICOS
У тестируемых мышей (контрольных мышей), которым вводили антитело отрицательного контроля (крысиный IgG), тяжелый колит развился через 4-7 недель после введения Т-клеток CD45RBhigh, и наблюдалась не только значимая потеря веса тела (фиг.3), но также и значимое утолщение кишечной стенки толстой кишки, сопровождающееся диареей и воспалением.
У контрольных мышей средняя величина (на 7-ой неделе) гистологических балльных оценок (указывающих на тяжесть колита), характеризуемого трансмуральным воспалением, при котором большое количество лимфоцитов наблюдается в основном слое и подслизистом слое, и выраженной гиперплазией эпителия, сопровождающей уменьшение количества бокаловидных клеток, составила 5,9±1,2 (фиг.4).
С другой стороны, мыши, получавшие лечение антителом против AILIM/ICOS, были абсолютно здоровы, и признаки колита и утолщение стенки толстой кишки не наблюдали, и с течением времени наблюдали прибавку в весе тела (фиг.3). Кроме того, гистологическая балльная оценка ткани стенки толстой кишки у данных мышей, получавших лечение антителом против AILIM/ICOS (n=7), составила 0,8±0,7, и выраженных патологических изменений не наблюдали (фиг.4).
Среднее количество Т-клеток CD4+, наблюдаемое в основном слое толстой кишки мышей с колитом (контрольные мыши), у которых развилось воспаление, составило 44±9·105 клеток/толстую кишку, в то время как у мышей, получавших лечение антителом против AILIM/ICOS, оно составило 21±3·105 клеток/толстую кишку (p<0,01) (фиг.5).
<2-8-2> Индукция антителом против AILIM/ICOS апоптоза мононуклеарных клеток, инфильтрирующих ткань
В толстой кишке мышей, которым вводили антитело против AILIM/ICOS, наблюдали значимое увеличение количества апоптозных клеток (большинство из них представляли собой мононуклеарные клетки, инфильтрирующие ткань), по сравнению с количеством у мышей, получавших лечение антителом отрицательного контроля (контрольных мышей).
Показатель апоптоза, по данным количественного анализа апоптозных клеток в толстой кишке, был значимо выше у мышей, которым вводили антитело против AILIM/ICOS (n=5), по сравнению с показателем у мышей, которым вводили контрольное антитело (контрольные мыши) (фиг.6).
Полученные результаты показали, что подавляющее воздействие на колит вследствие лечения антителом против AILIM/ICOS связано с истощением запаса патогенных Т-клеток, которые экспрессируют AILIM (ICOS).
<2-8-3> Подавляющее воздействие на колит вследствие введения антитела против AILIM/ICOS после прогрессирования симптомов
Потеря веса тела и инфильтрация ткани толстой кишки лимфоцитами, т.е. болезнь, вызывающая истощение, которая является одним из симптомов в указанной выше модели колита у мышей, начинаются соответственно приблизительно через 3 недели и приблизительно через 2 недели после введения Т-клеток CD45RBhigh. Поэтому, как указано выше, введение антитела против AILIM/ICOS начинали через 3 недели после введения Т-клеток.
У группы, которой вводили антитело против AILIM/ICOS, наблюдали значимое уменьшение потери веса тела (фиг.7) и не наблюдали диарею.
Гистологический анализ срезов ткани толстой кишки мышей из каждой группы показал, что грануломатозное воспаление, лимфоцитарная инфильтрация и утолщение эпителия значимо уменьшались в тканях толстой кишки у группы, которой вводили антитело против AILIM/ICOS, по сравнению с данными у мышей, которым вводили антитело отрицательного контроля (контрольных мышей). Изменения воспаления наблюдали в основном слое, а в определенных случаях - в поражениях, сопровождающих незначительную инфильтрацию лимфоцитами подслизистого слоя, но не мышечного слоя. Кроме того, указанная выше гистологическая балльная оценка, указывающая на тяжесть колита (толстую кишку брали на 7-й неделе), была значимо снижена в группе мышей, которым вводили антитело против AILIM/ICOS (1,62±0,81), по сравнению с оценкой в группе мышей, которым вводили антитело отрицательного контроля (контрольных мышей; 5,93±1,23) (p<0,05; фиг.8). Кроме того, количество инфильтрирующих Т-клеток CD4+ (7-я неделя) значимо уменьшилось у мышей, которым вводили антитело против AILIM/ICOS (5,50±0,7×106 клеток), по сравнению с количеством у контрольных мышей (31,3±7,7×106 клеток) (p<0,05; фиг.9).
<2-8-4> Подавляющее воздействие антитела против AILIM/ICOS на колит, вызванный введением Т-клеток CD4+ основного слоя, полученных у донора
Для исследования терапевтического эффекта антитела против AILIM/ICOS на патогенные Т-клетки, как описано выше, колит вызывали введением Т-клеток LP CD4+, полученных у мышей с колитом, мышам BALB/c scid/scid с последующим введением антитела против AILIM/ICOS.
У мышей, которым вводили антитело против AILIM/ICOS, потеря в весе тела (фиг.10) значимо уменьшалась. Кроме того, гистологическая балльная оценка, показывающая тяжесть колита (толстую кишку брали на 4-й неделе; фиг.11) и инфильтрация клеток CD4+ в основной слой толстой кишки (4-я неделя) (фиг.12) также значимо уменьшилась по сравнению с данными показателями у мышей, которым вводили антитело отрицательного контроля (контрольных мышей).
Промышленная применимость
Фармацевтические композиции настоящего изобретения (особенно предпочтительно содержащие вещество, обладающее активностью индукции гибели клеток, апоптоза или истощения запаса клеток, которые экспрессируют AILIM/ICOS) крайне полезны для подавления, предотвращения и/или лечения заболеваний, которые могут быть вызваны патологическим иммунитетом кишечного тракта, более конкретно, воспалительных заболеваний кишечника (в частности, болезни Крона и колита (язвенного колита и ему подобных)) и пищевых аллергий.
Фармацевтические композиции данного изобретения могут усилить терапевтический эффект при воздействии на воспалительные заболевания кишечника и пищевые аллергии при применении в комбинации с существующими фармацевтическими средствами, назначаемыми для лечения таких воспалительных заболеваний и пищевых аллергий.
Кроме того, фармацевтическая композиция, включающая человеческое антитело против AILIM, которое включено в качестве части фармацевтической композиции данного изобретения, крайне полезна в качестве лекарственного препарата, поскольку она не вызывает никаких побочных эффектов, таких как аллергические реакции, которые наблюдаются при введении людям антител, полученных у мышей.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИММУННЫХ ЗАБОЛЕВАНИЙ | 2000 |
|
RU2203682C2 |
| СРЕДСТВА, ПОДАВЛЯЮЩИЕ ОТТОРЖЕНИЕ ТРАНСПЛАНТАТА | 2002 |
|
RU2263512C2 |
| ЧЕЛОВЕЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ AILIM, КОСТИМУЛИРУЮЩЕЙ МОЛЕКУЛЫ ПЕРЕДАЧИ СИГНАЛА, И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2001 |
|
RU2262511C2 |
| ПРОФИЛАКТИЧЕСКИЙ ИЛИ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ ДЛЯ ЛЕЧЕНИЯ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ КИШЕЧНИКА, СОДЕРЖАЩИЙ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА АНТАГОНИСТ IL-6 | 1999 |
|
RU2195960C2 |
| ТЕРАПЕВТИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2002 |
|
RU2328506C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ IL-21 ЧЕЛОВЕКА | 2008 |
|
RU2504552C2 |
| АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО VISTA И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2746994C2 |
| РЕКОМБИНАНТНЫЕ АНТИТЕЛА ПРОТИВ CD4 ДЛЯ ТЕРАПИИ ЧЕЛОВЕКА | 1996 |
|
RU2232773C2 |
| НОВЫЕ ПОЛНОСТЬЮ ЧЕЛОВЕЧЕСКИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ VAP-1 | 2008 |
|
RU2459832C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ВОСПАЛЕНИЯ В НЕЙРОННОЙ ТКАНИ С ПРИМЕНЕНИЕМ МОНОКЛОНАЛЬНОГО АНТИТЕЛА ИЛИ ЕГО СВЯЗЫВАЮЩЕГО ФРАГМЕНТА | 2007 |
|
RU2565391C2 |
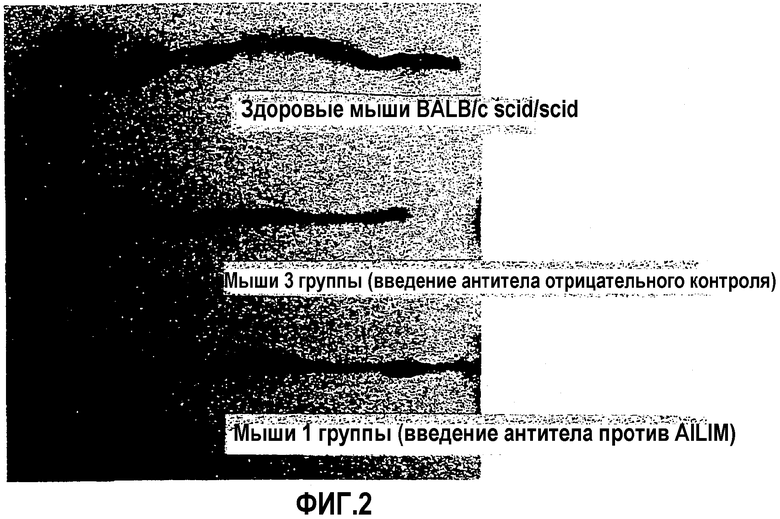
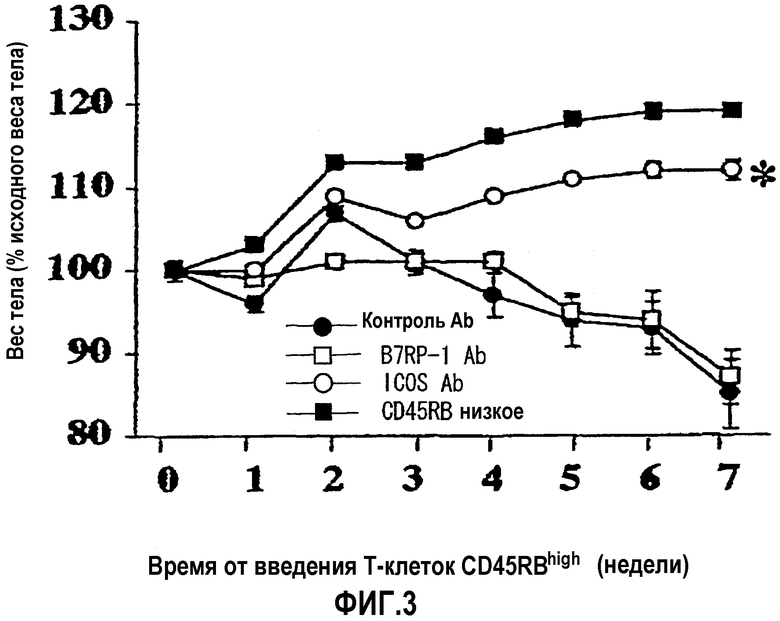
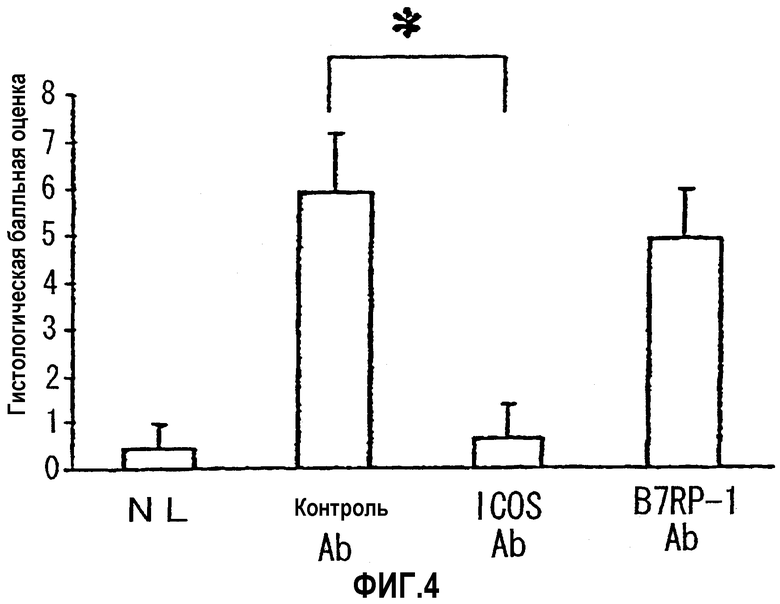
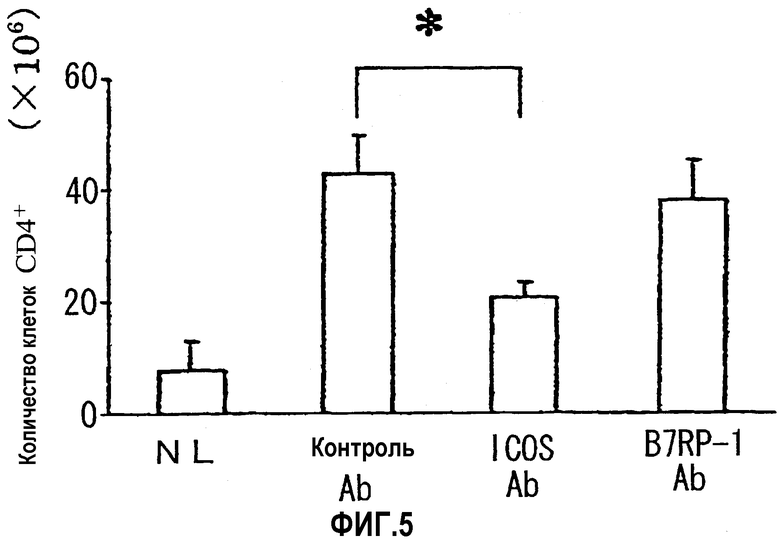
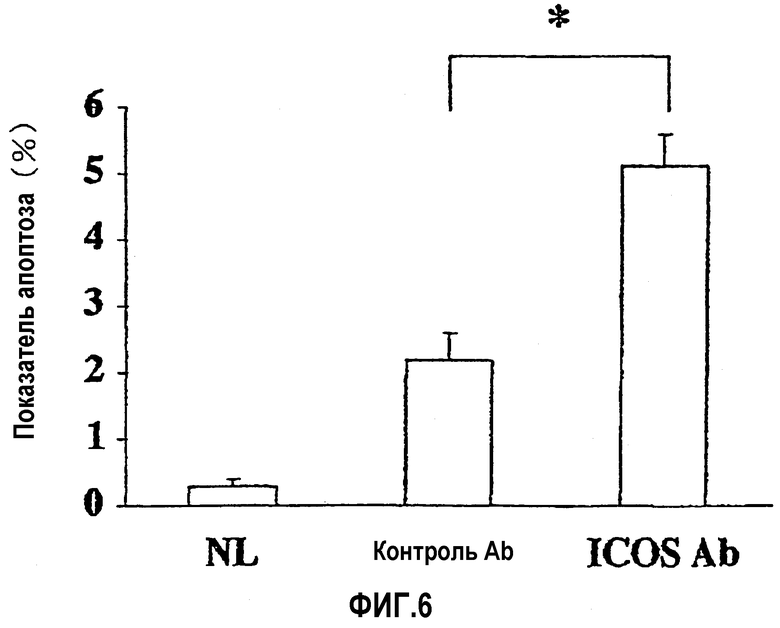
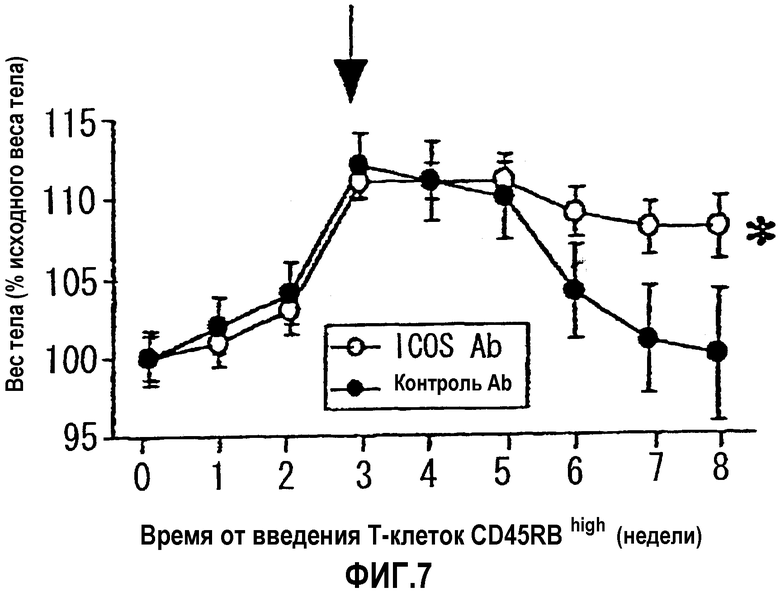
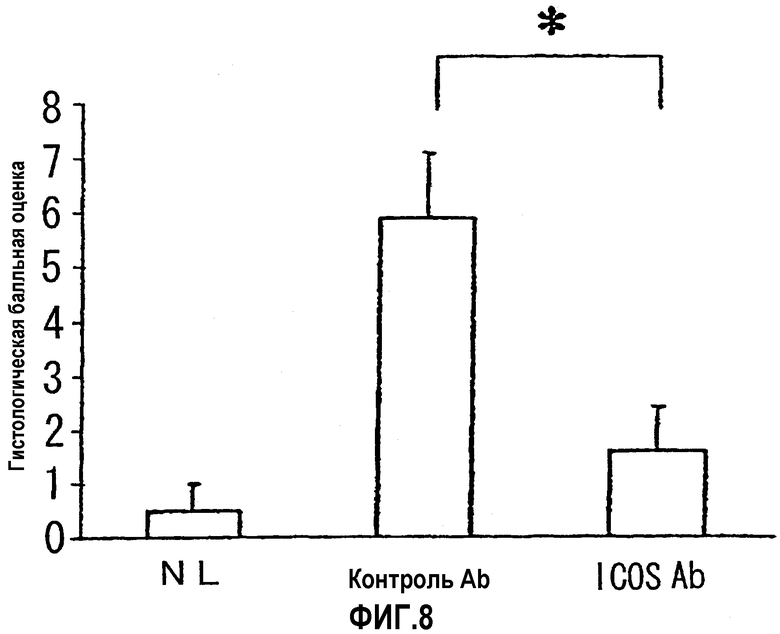
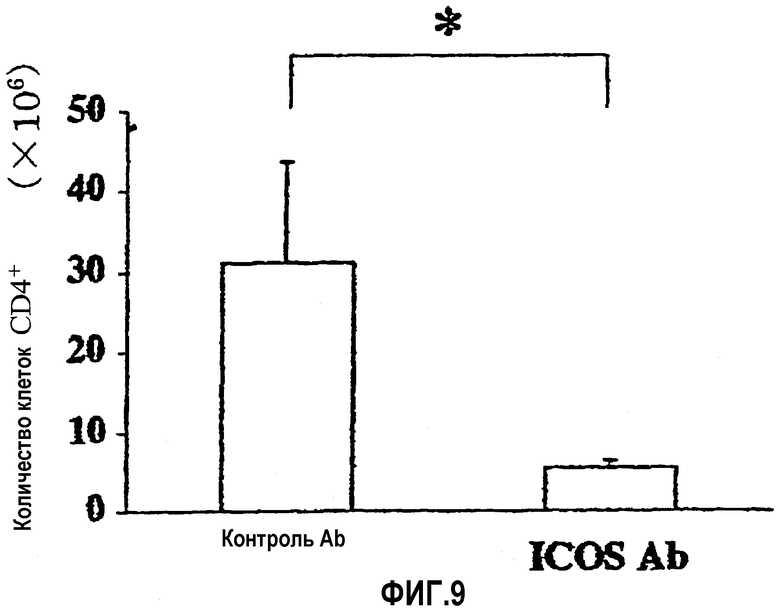
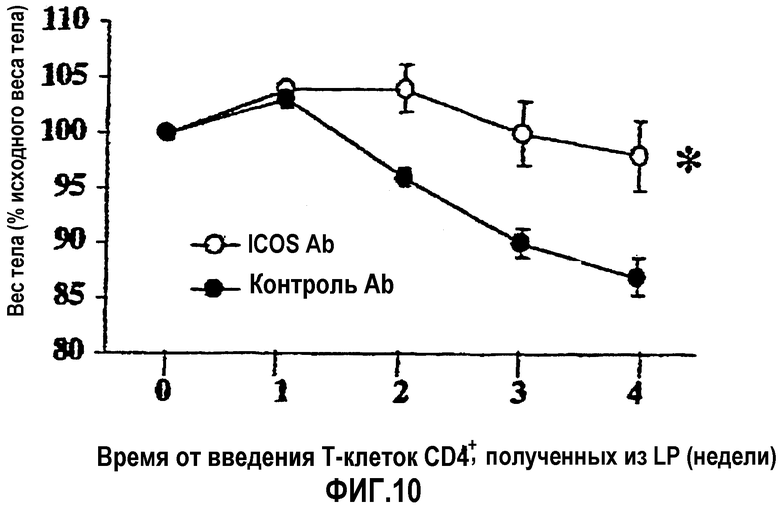
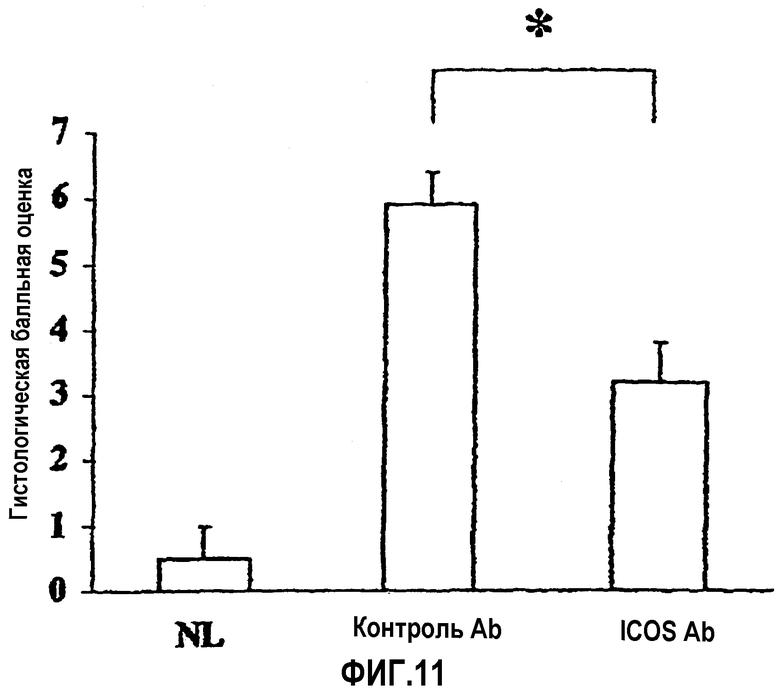
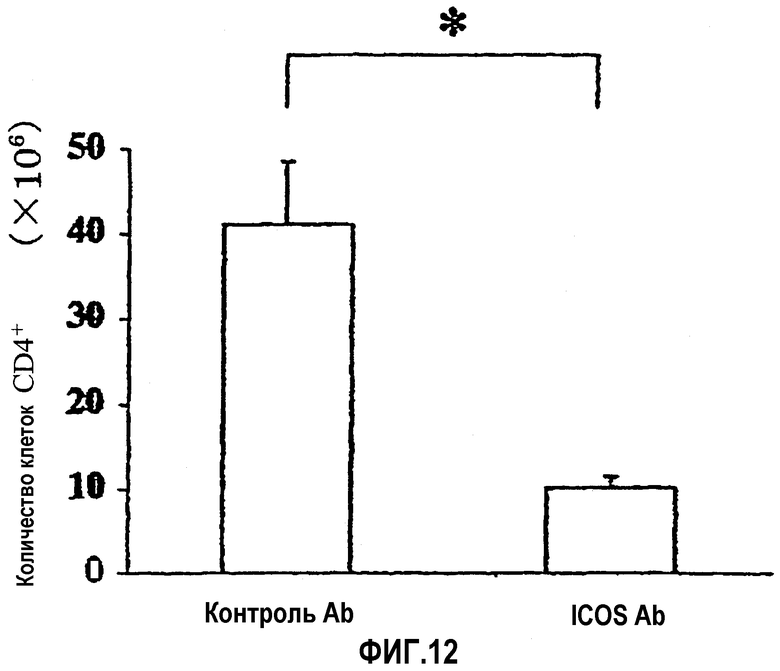
Изобретение относится к фармакологии, медицине и конкретно к композициям, включающим вещество, обладающее активностью регулирования, например ингибирования, пролиферации клеток, экспрессирующих AILIM, для подавления начала воспалительных заболеваний кишечника, в частности болезни Крона и колита. Изобретение обеспечивает усиление терапевтического эффекта. 9 з.п. ф-лы, 12 ил.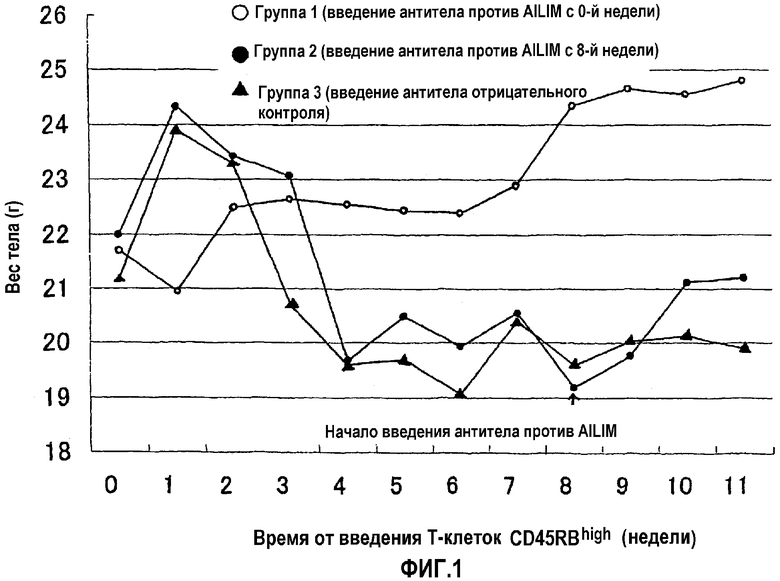
а) антитело, которое связывается с AILIM, или часть указанного антитела;
b) полипептид, содержащий всю внеклеточную область AILIM или ее часть;
c) гибридный полипептид, включающий всю или часть внеклеточной области AILIM, и всю или часть константной области тяжелой цепи иммуноглобулина; и
а) полипептид, который связывается с AILIM.
Приоритет по пунктам:
| Cell, 1993, v.75, p.263-274 | |||
| WO 00/45240 A2, 10.08.2000 | |||
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| Устройство для непрерывной дегазации стали | 1958 |
|
SU115732A1 |
| СТИМУЛЯТОР ЭНДОГЕННОЙ ПРОДУКЦИИ ЦИТОКИНОВ И ГЕПОПОЭТИЧЕСКИХ ФАКТОРОВ И СПОСОБ ЕГО ИСПОЛЬЗОВАНИЯ | 1995 |
|
RU2089179C1 |
| СПОСОБ ДОСТАВКИ БИОАКТИВНОГО АГЕНТА ЖИВОТНОМУ ДЛЯ ИНИЦИАЦИИ ИММУННОГО ОТВЕТА (ВАРИАНТЫ) | 1989 |
|
RU2127118C1 |

