Предисловие
В соответствии с настоящей заявкой заявляется преимущество предварительной заявки номер 60/346 402, поданной 1 ноября 2001 г, и испрашивается приоритет по РСТ/US01/14151, поданной в мае 2001 г., которая в настоящее время находится на рассмотрении. В соответствии с РСТ/US01/14151 заявляется преимущество временной заявки номер 60/201 344, поданной 2 мая 2000 г. Заявки, преимущество которых заявляется в соответствии с настоящей заявкой, приведены полностью в качестве ссылок в данном описании.
Настоящее изобретение было сделано при поддержке правительственного гранта NCI P50 CA 89019-01, предоставленного Национальным Институтом по изучению злокачественных опухолей, и при поддержке гранта NIH R03-AR44982, предоставленного Национальным Институтом по изучению артрита и скелетно-мышечных и кожных заболеваний (National Institute of Arthritis and Musculoskeletal and Skin Diseases). Правительство обладает определенными правами на настоящее изобретение.
Область техники, к которой относится изобретение
Настоящее изобретение относится к антителу, которое способно специфически связываться с одним типом ассоциированного с фактором некроза опухоли (далее называемым как TNF) рецептора лиганда, индуцирующего апоптоз (далее называемого как TRAIL), более конкретно, настоящее изобретение относится к моноклональному антителу, которое индуцирует in vivo и in vitro апоптоз клеток, экспрессирующих один тип рецептора, и к способам лечения на его основе.
Предпосылки создания изобретения
TRAIL представляет собой член семейства белков TNF, которое включает в себя TNF-α и Fas-лиганд [1]. Указанные белки являются индукторами апоптоза. К настоящему времени идентифицировано пять рецепторов TRAIL, два из которых, DR4 (TRAIL-R1) и DR5 (TRAIL-R2), способны к передаче апоптозного сигнала [2-7], тогда как другие три, DcR1 (TRAIL-R3), DcR2 (TRAIL-R4) и остеопротегерин (OPG), не способны к передаче апоптозного сигнала [8-12]. Все пять рецепторов для TRAIL характеризуются наличием существенной гомологии во внеклеточных доменах, связывающихся с лигандом. Аналогично Fas и рецептору-I TNF (далее называемому как TNFRI), внутриклеточные сегменты DR4 и DR5 содержат домен гибели и передают апоптозный сигнал по механизму, в котором участвуют ассоциированный с Fas белок, содержащий домен гибели (далее называемого как FADD), и каспазы 8 [6, 7]. В дополнение к передаче апоптозного сигнала рецепторы DR4 и DR5 могут также активировать путь, вовлекающий NFkb [6, 7].
Было показано, что биологические функции TRAIL включают его способность селективно индуцировать апоптоз трансформированных опухолевых клеток, тогда как нормальные остаются относительно устойчивыми к TRAIL-опосредованному апоптозу [13-15]. Указанная селективность позволяет предполагать, что, в отличие от Fas-лиганда, введение TRAIL сопровождается очень низким уровнем токсичности, как было показано при системном введении TRAIL животным моделям без индукции значительной токсичности [13]. Так, было сделано предположение о том, что TRAIL является мощным средством, индуцирующим апоптоз, которое может стать подходящим лекарственным средством для лечения злокачественных опухолей и других заболеваний, ассоциированных с аномальной пролиферацией клеток. Было также высказано предположение о том, что TRAIL является мощным апоптоз-индуцирующим средством, которое может стать подходящим лекарственным средством для лечения аутоиммунных и воспалительных заболеваний. Было показано, что TRAIL-опосредованный апоптоз участвует в индуцируемой активацией гибели Т-клеток, выполняя, таким образом, функцию механизма, альтернативного Fas-лиганду [16, 17]. TRAIL-опосредованный апоптоз может также действовать в плане индукции апоптоза Т-клеток и других воспалительных клеток [18] и играть роль в активности NK-клеток по уничтожению клеток [19-21] и в иммуномодулирующей функции дендритных клеток [22, 23]. Так, TRAIL-опосредованный апоптоз может также функционировать при иммунокомпетентности (immunoprivilege) и иммунном контроле.
Рецепторная система для TRAIL является сложной и включает по меньшей мере два рецептора, вызывающих гибель клеток, DR4 и DR5, и по меньшей мере два неапоптозных рецептора, DcR1 и DcR2. Все указанные рецепторы не только характеризуются гомологией с точки зрения сходства аминокислотной последовательности, но также демонстрируют аналогичную связывающую аффинность в отношении TRAIL [2-12]. Способность рецепторов DcR1 и DcR2 конкурировать за связывание с TRAIL без индукции апоптоза дает основание полагать, что они могут действовать в качестве ложных рецепторов, блокирующих или модулирующих активность TRAIL-лиганда. Кроме того, имеется сообщение о том, что нетрансформированные клетки экспрессируют более высокие уровни ложных рецепторов, чем это делают трансформированные клетки. Таким образом, было высказано предположение о том, что дифференцированная модуляция экспрессии рецепторов, вызывающих гибель клеток, и ложных рецепторов может представлять собой ключевой механизм, который определяет чувствительность клеток к TRAIL-опосредованному апоптозу, в случае отсутствия специфичных к рецептору антител [2]. Несмотря на то что экспрессия и функционирование DR4 и DR5 интенсивно изучались, прогресс сдерживался недостатком моноклональных антител, специфичных к рецептору. Относительно экспрессии DR5 на клеточной поверхности нет опубликованных данных. Сообщалось о том, что была получена совокупность антител к рецептору TRAIL, которые способны индуцировать апоптоз в клетках меланомы in vitro, но только при иммобилизации антител, которая усиливает их сшивку и в ряде случаев требуется культивирование клеток с актиномицином D [24]. Было получено несколько антител против DR5 [24]. Однако ранее полученные моноклональные антитела против DR5 обладают низкой активностью по индукции апоптоза in vitro даже в условиях наличия поперечных сшивок. При этом отсутствуют сообщения об их активности in vivo. Указанные антитела не использовались для изучения экспрессии рецепторов TRAIL на клеточной поверхности [24]. Таким образом, имеется потребность в моноклональном антителе, селективном для данного специфического рецептора TRAIL, которое не только было бы способно связываться с рецептором клеточной поверхности, но также могло в значительной мере индуцировать апоптоз в различных типах аномальных клеток, включая опухолевые клетки, как in vivo так и in vitro, без потребности в сшивании или иммобилизации. Такое антитело могло бы не только представлять собой потенциальное терапевтическое средство, но также стать диагностическим инструментом для проведения функционального анализа рецептора TRAIL. Таким образом, существует определенная потребность в антителе, специфичном к каждому из рецепторов DR4 и DR5, способных индуцировать гибель клеток.
При развитии или прогрессировании многих заболеваний часто имеют место случаи, когда клетки не удаляются. При многих аутоиммунных заболеваниях и воспалительных состояниях выживающие активированные клетки атакуют нормальные ткани или клетки. Далее прогрессирование опухолегенеза и формирование пролиферативного состояния при ревматоидном артрите характеризуется отсутствием контроля пролиферации клеток. Таким образом, недостаточный апоптоз приводит к развитию заболевания, а варианты использования лиганда, индуцирующего апоптоз, или агонистического моноклонального антитела, усиливающего апоптоз, рассматриваются как потенциальные терапевтические подходы для устранения таких нежелательных клеток.
Так, например, ревматоидный артрит (называемый в описании РА) представляет собой распространенное аутоиммунное заболевание человека. В настоящее время представление о патофизиологии РА сводится к тому, что аутоиммунные Т-клетки и В-клетки инициируют воспалительную реакцию в суставах, которая ведет к гиперпролиферации синовиоцитов. Вследствие гиперпролиферации синовиальных клеток металлопротеиназы (называемые в описании как «МП») образуются в избыточных количествах, что ведет к характерной для РА эрозивной деструкции хряща и кости [25]. Таким образом, контроль гиперпролиферации синовиальных клеток при воспалении является ключевой стадией при лечении РА. Молекулярные механизмы, ведущие к гиперпролиферации синовиальных клеток, все еще неизвестны. Хотя гиперпролиферирующие синовиальные клетки не являются злокачественными и не являются трансформированными, в ряде исследований высказывалось предположение о том, что они характеризуются некоторыми общими особенностями с трансформированными клетками [46]. Указанные клетки, так называемые «синовиоциты, похожие на трансформированные», характеризуются наличием плотного шероховатого эндоплазматического ретикулюма, многочисленными ядрами неправильной формы и изменениями нормальной веретенообразной формы клеток. Было высказано предположение, что включение онкогенов и генов вирусного происхождения может быть первичным механизмом запуска появления трансформированной формы синовиальных клеток при РА [46].
По меньшей мере два аспекта, свойственные РА, позволяют предположить, что нерегулируемый апоптоз может вносить определенный вклад в развитие заболевания и что изучение возможностей терапевтического применения апоптоза может стать эффективным способом лечения: недостаточное удаление активированных Т-клеток дает основание предполагать наличие дефекта в пути уничтожения указанных Т-клеток, индуцированных активацией, который представляет собой процесс, вовлекающий Fas-опосредованный апоптоз и TRAIL-опосредованный апоптоз, а гиперпролиферативная природа синовиальных клеток при РА представляет собой фактор, действующий на более поздних стадиях патофизиологии РА. Фактически было показано, что введение антитела против Fas в воспаленный сустав подавляет развитие хронического артрита у трансгенных мышей tax, которые представляют собой модель животных для изучения РА человека [26]. Более того, локализованная трансдукция гена fas-ligand с помощью аденовирусного вектора является эффективной с точки зрения профилактики артрита, вызванного коллагеном [27]. В обоих случаях наблюдается подавление пролиферации воспаленных синовиальных клеток за счет усиления Fas-опосредованного апоптоза. Хотя Fas-лиганд является сильным индуктором апоптоза в синовиальных клетках при РА, применение апоптоза, опосредованного Fas-лигандом, для лечения людей ограничено его летальной гепатотоксичностью. Таким образом, апоптоз, индуцированный рецептором TRAIL, представляет более безопасный и более эффективный терапевтический подход для лечения РА, чем апоптоз, индуцированный Fas-лигандом. Апоптоз, индуцированный рецептором TRAIL, также представляет более безопасный и более эффективный терапевтический подход для лечения злокачественной опухоли, чем апоптоз, индуцированный Fas-лигандом. Известно, что TRAIL-опосредованный апоптоз специфически индуцирует апоптоз трансформированных опухолевых клеток, не затрагивая нормальные клетки. Было показано, что системное введение тримеризованного растворимого TRAIL не вызывает токсичности у экспериментальных животных, когда он уже обладает способностью индуцировать регрессию имплантированных опухолей [13, 28]. Его потенциал в качестве средства для вспомогательной терапии в традиционных курсах лечения был пересмотрен в свете последних результатов, согласно которым экспрессия DR5 и чувствительность к TRAIL-индуцированному апоптозу клеток рака молочной железы усиливается при облучении, что дает основание полагать, что в случае объединения с облучением эффективность TRAIL может быть усилена при терапии злокачественной опухоли [29].
Кроме того, ген, кодирующий TRAIL-рецептор DR5, был картирован на хромосоме в локусе 8p21-22, который отличается высокой частотой мутации в некоторых злокачественных клетках [30]. Имеются сообщения о том, что по меньшей мере два вида опухолевых клеток, мелкоклеточный рак легкого [31] и рак головы и шеи [32], демонстрируют мутации в домене гена DR5, связанном с уничтожением клеток. Таким образом, в рамках исследования злокачественной опухоли существует потребность в антителе против DR5 для определения эффекта вариаций в эпитопе рецептора на развитие и прогрессирование злокачественной опухоли. Далее, функциональная значимость мутаций TRAIL-рецептора могла бы обеспечить полезный инструмент для клинической диагностики, а в сочетании с другими биологическими маркерами использоваться для ранней диагностики злокачественной опухоли и прогнозирования агрессивности опухоли.
Краткое описание сущности изобретения
В одном варианте настоящее изобретение относится к антителу, которое распознает TRAIL-рецептор DR5 и которое индуцирует апоптоз DR5-экпрессирующей клетки in vivo или in vitro. Далее описывается антитело, которое распознает DR5, но не DR4, DcR1 или DcR2. Конкретно, подробно описывается моноклональное антитело DR5, продуцируемое гибридомой.
В другом варианте осуществления настоящее изобретение относится к антителу, которое распознает TRAIL-рецептор DR4 и которое индуцирует апоптоз DR4-экпрессирующей клетки in vivo или in vitro. Далее описывается антитело, которое распознает DR4, но не DR5, DcR1 или DcR2. Конкретно подробно описывается моноклональное антитело DR4, продуцируемое гибридомой.
Предлагаемый способ представляет собой индукцию апоптоза в целевых клетках или ингибирование пролиферации целевой клетки путем контакта клетки с терапевтическим количеством антитела, способного связываться с DR5 или DR4. В различных вариантах реализации указанного способа апоптоз может быть индуцирован или пролиферация клеток может быть подавлена путем контакта целевых клеток с обоими видами антител.
Описывается также фармакологическая композиция, которая включает терапевтическое количество моноклонального антитела, активного против DR5 или DR4, фармацевтически приемлемый носитель и необязательно контейнер, включающий указанное антитело и носитель. Далее в изобретении предлагается использование антитела, распознающего DR5, или антитела, распознающего DR4, для изготовления лекарственного средства, применяемого с целью селективного апоптоза аномальных или нерегулируемых клеток.
Антитело по настоящему изобретению взаимодействует с индуцирующим апоптоз рецептором лиганда, связанным с фактором некроза опухоли, таким как DR4, DR5, DrR1, DrR2 или OPG, индуцирующим апоптоз в клетке, экспрессирующей такой рецептор. Описываемое в настоящем изобретении антитело представляет собой антитело, способное селективно связываться с эпитопом рецептора лиганда агонистического или антагонистического фактора некроза опухоли.
Настоящее изобретение относится к лечению связанного с апоптозом заболевания, такого как злокачественная опухоль, воспалительное заболевание или аутоиммунное заболевание, с использованием способа, который включает контактирование целевой ткани, подверженной заболеванию, с терапевтическим количеством антитела по настоящему изобретению, одним или в сочетании с другими антителами, индуцирующими апоптоз, и/или другими терапевтическими средствами или видами лечения.
Далее описывается белок слияния, который включает аминокислотную последовательность антигенного рецептора TRAIL, содержащего по меньшей мере десять оснований, связанных с иммуноглобулиновым белком или его фрагментом, способным вызывать иммунную реакцию у субъекта.
Настоящее изобретение относится к способу генной терапии, при которой целевая клетка подвергается трансфекции последовательностью нуклеиновой кислоты для рецептора TRAIL, включенной в вектор экспрессии, таким образом, что указанный TRAIL-рецептор экспрессируется на целевой клетке. Затем целевая клетка подвергается воздействию антитела, которое селективно связывает TRAIL-рецептор.
В соответствии с изобретением предлагаются разработанные последовательности нуклеиновой кислоты и аминокислотные последовательности, кодирующие тяжелую и легкую цепь иммуноглобулинов антитела, селективного для DR5. Предлагаются также последовательности для антитела, которые селективно связываются с DR4. Далее подробно описываются также векторы, которые включают последовательность нуклеиновой кислоты по настоящему изобретению и трансформированные клетки-хозяева, содержащие такой вектор по настоящему изобретению.
Настоящее изобретение относится к гуманизированному антителу DR5 (то есть TRA-8) и гуманизированному DR4 (то есть 2E12), а также к трансфицированной клетке, продуцирующей гуманизированное антитело DR5, и трансфицированной клетке, продуцирующей гуманизированное антитело DR4.
Описывается также способ получения гуманизированного антитела DR5 или антитела DR4, при котором клетка-хозяин трансформируется последовательностями нуклеиновой кислоты, кодирующими легкую цепь гуманизированного иммуноглобулина и тяжелую цепь гуманизированного иммуноглобулина, после чего трансформированную клетку-хозяин инкубируют в течение заданного периода времени.
В изобретении также описывается способ индукции апоптоза в целевых клетках или подавления пролиферации клеток, который включает контактирование целевой клетки с фармацевтически эффективным количеством гуманизированного антитела DR5, гуманизированного антитела DR4 или их сочетания, в присутствии или в отсутствие других терапевтических средств и режимов лечения.
Предлагается коммерческий набор для индукции апоптоза, который включает гуманизированное антитело TRA-8, селективное для DR5, или гуманизированное антитело, селективное для DR4 (например, гуманизированное 2Е12), упакованные в подходящий контейнер и, необязательно, при наличии инструкции по использованию.
Краткое описание чертежей
Фиг.1. Характеристика TRA-8. (a) Специфичность по связыванию TRA-8: Результат анализа по методу вестерн-блоттинга (верхний план): Рекомбинантные слитые белки из семейства TNFR с зондом в виде TRA-8 или антитела против человеческого IgG. Линия 1: Слитый белок DR5/hIgG1 (иммуноген): Линия 2: DR4/hIgG1 (TRAIL-R1); Линия 3: DR5/hIgG1; Линия 4: TRAIL-R3 (DcR-1)/hIgG1; Линия 5: TRAIL-R4 (DcR-2)/hIgG1; Линия 6: CD95/hIgG1; Линия 7: растворимый TNFRI. Результат анализа по методу ELISA (нижний план): Номера лунок обозначают варианты вестерн-блоттинга за исключением лунки 8, которая представляет собой слитый белок мышиного DR5/hIgG1. (b) Связывающая активность TRAIL и TRA-8 в отношении DR5 и DR4: на планшеты для ELISA наносят DR5/hIgG1 (левый план) или DR4/hIgG1 (средний план) и затем инкубируют с TRAIL или TRA-8. (c) Анализ по методу проточной цитометрии экспрессии DR5 на клеточной поверхности. Cos-7 клетки трансфицируют вектором экспрессии pcDNA3, содержащим кДНК DR5 полной длины (гистограмма со сплошными линиями), кДНК DR4 (открытая гистограмма, сплошная линия) или пустой вектор (открытая гистограмма, пунктирная линия). Через сорок восемь часов после трансфекции клетки окрашивают TRA-8 и затем подвергают воздействию антитела против мышиного IgG1, конъюгированной с ФЭ. (d) Иммуногистохимическая реактивность in situ для DR5. Цитоспиновые слайды Cos-7 клеток, трансфицированных экспрессируемым DR5 или контрольным вектором, окрашивают TRA-8 через сорок восемь часов после трансфекции. (е) Активность TRA-8 по уничтожению клеток: Клетки Jurkat инкубируют с указанными концентрациями TRA-8. Жизнеспособность клеток определяют методами ATPLite, MTT и по исключению PI после культивирования в течение ночи. Результаты тестов ATPLite и MTT представлены в виде процента от значения контроля, представляющего собой среду, и результаты теста PI представлены в виде процента от значения PI для отрицательных клеток. (f) Результаты вестерн-блоттинг-анализа активации каспазы: Клетки Jurkat инкубируют с 500 нг/мл TRA-8 в течение указанного периода времени. Клеточные лизаты разделяют электрофорезом в 15% ДСН-ПААГ, проводят блоттинг и подвергают воздействию зондов с антителами к каспазе. Стрелки указывают расщепленные субъединицы каждой каспазы. (g) Тест на ингибирование каспазы: Клетки Jurkat инкубируют с 50 нг/мл TRA-8 в течение ночи в присутствии различных концентраций указанных ингибиторов каспазы. Жизнеспособность клеток оценивают по методу ATPLite.
Фиг.2. Экспрессия DR5 на клеточной поверхности и чувствительность к DR5-опосредованному апоптозу. Выделяют из свежей периферической крови нормальные T- и B-клетки, клеточные линии, включающие T-клетки (a и a'), глиомные клетки (b и b'), клетки рака предстательной железы (c) и B клетки (d), и инкубируют с TRA-8 или контрольным антителом IgG1 изотипа мышей, после чего вносят конъюгированное с ФЭ козье антитело против мышиного IgG1. Открытые гистограммы обозначают изотип контрольного антитела, тогда как жирные гистограммы обозначают окрашивание под действием TRA-8. Апоптоз определяют по методу ATPLite после ночной инкубации с растворимым TRAIL (открытые кружки) или TRA-8 (зачерненные кружки), как показано для а, b' и d.
На фиг.3a' показан результат инкубирования Т-клеточной линии U937 с TRA-8 или мышиным контрольным антителом IgG1 изотипа. Апоптоз определяют по методу ATPLite после ночного инкубирования с растворимым TRAIL (открытые кружки) или TRA-8 (зачерненные кружки).
На фиг.3 показаны клеточные линии глиомы (b) и рака предстательной железы (c), инкубируемые с TRA-8 или мышиным контрольным антителом IgG1 изотипа. Апоптоз определяют по методу ATPLite после инкубирования в течение ночи с растворимым TRAIL (открытые кружки) или TRA-8 (зачерненные кружки).
На фиг.4 показана серия графиков, характеризующих жизнеспособность человеческих клеток Jurkat после экспозиции с указанными концентрациями (А) антитела, штаммов TRA-1, -8 и -10 и (В) TRAIL в присутствии фиксированных концентраций штаммов антитела по настоящему изобретению, отраженных на фиг.4А.
Фиг.5. Экспрессия DR5 в нормальных тканях и тканях злокачественных опухолей: гомогенаты нормальной ткани и ткани злокачественной опухоли анализируют с TRA-8 в качестве зонда и анализируют по хемилюминесценции. (а) Анализ по методу вестерн-блоттинга белка DR5 в нормальных тканях: линия 1: печень, линия 2: мозг, линия 3: легкое, линия 4: почка, линия 5: селезенка, линия 6: яички, линия 7: яичник, линия 8: сердце, линия 9: поджелудочная железа. (b) Анализ по методу вестерн-блоттинга белка DR5 в тканях злокачественных опухолей. Анализируют с помощью зонда блот из ткани злокачественной опухоли, содержащей раковые клетки яичника (линия 1), легкого (линия 2), печени (линия 3), прямой кишки (линия 4), шейки матки (линия 5), кожи (линия 6), яичек (линия 7), щитовидной железы (линия 8), матки (линия 9), желудка (линия 11), области глотки и пищевода (линия 12) и поджелудочной железы (линия 13). Показана иммуногистохимия in situ нормальных человеческих тканей (с) и тканей злокачественных опухолей (d). Замороженные среды подвергают иммунному окрашиванию с использованием TRA-8.
Фиг.6 Активность TRA-8 по эпилигнации опухолевых клеток. Мышам SCID инокулируют подкожно 1321N1 клетки. Мышам инъецируют внутривенно однократную дозу 100 мкг TRA-8 на второй день после инокуляции опухоли (a) или три дозы 100 мкг TRA-8, начиная с седьмого дня после инокуляции опухоли (b). Рост опухоли определяют по весу и проводят ее гистологический анализ с использованием H&E окрашивания. На фотографиях показан рост жизнеспособной опухоли у контрольных мышей и его отсутствие у мышей, которым был введен TRA-8 (с, верхний план) и при H&E окрашивании опухоли (с, нижний план). Мышам SCID инъецируют внутривенно 106 клеток Jurkat и на второй день после инъекции вводят однократную дозу TRA-8. Через семь дней отбирают клетки селезенки, окрашивают антителом против человеческого CD3 и анализируют методом проточной цитометрии (d.) или иммуногистохимическим методом (е).
На фиг.7 показана экспрессия на клеточной поверхности DR5 при РА (А) и ОА (В) в синовиальных клетках. 1Х106 первичных культивированных синовиальных клеток окрашивают очищенным аффинной хроматографией TRA-8 и затем обрабатывают ФЭ-конъюгированным козьим антителом против мышиного IgG1. Флуоресценцию 10000 жизнеспособных клеток анализируют с использованием FACSvantage.
На фиг.8 показана серия фотографий, демонстрирующих жизнеспособность клеток как функцию концентрации TRAIL и TRA-8, индуцирующих апоптоз репрезентативных штаммов РА (А) и ОА (В) синовиальных клеток с разными концентрациями рекомбинантного растворимого TRAIL (открытые кружки) или очищенного аффинной хроматографией TRA-8 (зачерненные кружки). Жизнеспособность клеток выражают в виде процента значения имп/мин для обработанных клеток относительно значения ипм/мин для необработанных клеток.
На фиг.9 представлена серия фотографий, демонстрирующих зависимость от каспазы DR5-опосредованного апоптоза синовиальных клеток при РА. Синовиальные клетки при РА (RA512) инкубируют с 50 нг/мл растворимого лиганда Fas (открытые квадраты) и антителом против Fas (CH-11) (зачерненные квадраты), растворимым TRAIL (открытые кружки) или антителом против DR5 (TRA-8) (зачерненные кружки) в присутствии варьирующих концентраций ингибиторов каспазы. После ночного культивирования определяют жизнеспособность клеток по методу ATPLite.
На фиг.10А приведены результаты гель-электрофореза, указывающие на активацию NFκb. RА1016-клетки инкубируют с 20 нг/мл TNF-a, 50 нг/мл растворимого TRAIL или 50 нг/мл TRA-8 в указанные временные точки перед проведением электрофореза.
На фиг.10В и С приведены графики, демонстрирующие уровень образования MMP-1 и MMP-3. 1·106/мл указанных синовиальных клеток при РА инкубируют с указанными концентрациями TNF-a (открытые кружки), TRAIL (открытые треугольники) или TRA-8 (зачерненные кружки). После ночного культивирования собирают супернатанты из культуральной среды. Уровень ММР в культуральных супернатантах определяют по методу ELISA.
На фиг.11 показано, что TRA-8 не индуцирует токсичность для клеток печени. (а) Нормальные ткани печени не экспрессируют DR5. Парафиновые срезы двух нормальных печеночных тканей, одной ткани карциномы клеток печени и цитоспиновый препарат клеток HepG2 подготавливают для окрашивания H&E и соответствующие замороженные среды окрашивают с использованием TRA-8. (b) Методом проточной цитометрии анализируют наличие экспрессии DR5 на клеточной поверхности. Гепатоциты, выделенные из двух нормальных печеночных тканей, из одного варианта ткани карциномы печени и клетки HepG2 окрашивают TRA-8, антителом против Fas (DX2) или изотипом контрольного антитела. Жирная гистограмма указывает окрашивание под действием TRA-8 или DX2, и открытая гистограмма соответствуют контрольным изотипам.
На фиг.12 показано, что TRAIL, но не TRA-8 индуцирует токсичность для клеток печени. Свежие нормальные гепатоциты человека поддерживают в культуральной среде для гепатоцитов. (а) Апоптоз гепатоцитов индуцируют с помощью 1 мкг/мл растворимого TRAIL плюс сшиватель или TRA-8 в указанные временные точки. Жизнеспособность клеток определяют методом ATPLite. Результаты представлены в виде процента уровня жизнеспособных клеток относительно контроля, представляющего собой среду. Затененные прямоугольники показывают TRAIL, а черные прямоугольники показывают TRA-8. (b) Конденсированные ядра гепатоцитов окрашивают Hoechst 33352 и анализируют методом проточной цитометрии. (c) Эффект циклогексимида на апоптоз гепатоцитов. Гепатоциты культивируют в контрольной среде или с добавлением 1 мкг/мл TRAIL или TRA-8 в присутствии (закрытые прямоугольники) или в отсутствие (открытые прямоугольники) 1 мкг/мл циклогексимида в течение 8 часов. Жизнеспособность клеток определяют по методу ATPLite. Результаты представлены в виде среднего значения ±СКО (SEM) для трех повторов культур в двух экспериментах. (d) Сравнение чувствительности нормальных гепатоцитов к DR5 и Fas-опосредованному апоптозу. Свежевыделенные гепатоциты инкубируют с указанными концентрациями растворимого TRAIL, TRA-8, растворимого FasL или анти-Fas мАт (mAb) CH11 в течение 6 часов. Жизнеспособность клеток определяют по методу ATPLite. Результаты представлены в виде процента жизнеспособных клеток относительно контроля, представляющего собой среду. В случае нормальных гепатоцитов представлены средние данные ±СКО для четырех нормальных индивидуумов. Результаты по клеткам карциномы печени от одного пациента и по клеткам HepG2 представлены в виде среднего значения результатов исследования культур в тройном повторе.
Фиг.13. Показано, что TRAIL индуцирует гепатит. Мышам В6 внутривенно инокулируют 109 БОЕ аденовирусного вектора, кодирующего человеческий TRAIL полной длины, под контролем элемента транскрипции «Тет-он». Экспрессия TRAIL индуцируется указанными дозами тетрациклина. (а) Анализ по методу нозерн-блоттинга экспрессии человеческого TRAIL в печени. Через 24 часа после инокуляции вектора и индукции тетрациклином из печени выделяют суммарную РНК и подвергают воздействию зондом кДНК человеческого TRAIL или β-актина. (b) Сывороточный уровень АСТ. Через 24 часа после трансдукции TRAIL определяются сывороточные уровни ACT (AST). (c) TRAIL-опосредованная гибель клеток гепатоцитов, инфицированных аденовирусным вектором: мышам В6 внутривенно инокулируют индуцируемый тетрациклином аденовирусный вектор. Через 48 часов после инокуляции выделяют гепатоциты из инокулированных мышей и контрольных неинокулированных мышей и инкубируют с указанными концентрациями TRAIL в течение 8 часов (левый план). Жизнеспособность гепатоцитов определяют по методу ATPLite. Мышам, которым инокулировали указанный выше аденовирусный вектор, через 48 часов внутривенно вводят 10 мкг растворимого человеческого TRAIL. Через 24 часа после инъекции TRAIL (правый план) измеряют сывороточный уровень ACT. (d и e) Гистологический анализ повреждения печени, индуцированного TRAIL. Печень отбирают через 24 часа (d) или 7 дней (е) после трансдукции TRAIL. Парафиновые срезы окрашивают по методу H&E и фотографируют с увеличением ×100 (верхний план) и ×400 (нижний план).
На фиг.14 приведена серия фотографий, из которых следует, что активированные Т-клетки и В-клетки, выделенные и очищенные из человеческих PBMC, экспрессируют повышенный уровень DR5, по результатам измерения методом проточной цитометрии покоящихся (не заштрихованные) и активированных (затененные) клеток.
Фиг.15 представляет собой график, показывающий жизнеспособность клеток как функцию концентрации TRA-8, для очищенных Т-клеток и В-клеток, изображенных на фиг.14, которые были стимулированы в течение 48 часов под действием анти-CD3 или анти-μ, активированных бластных клеток различной плотности, собранных с использованием Ficoll-Paque. Жизнеспособность клеток определяют по методу ATPLite.
Фиг.16 представляет собой гистограмму и график, построенный по результатам исследования методом проточной цитометрии экспрессии CD3 в отсортированной популяции лимфоцитов мышей NOD/SCID с недостаточностью NK-клеток, которым были инъецированы РВМС и TRA-8 или IgG (контроль).
На фиг.17 показаны микрофотографии клеток, окрашенных CD3 и по методу TUNEL, взятых из ткани селезенки мышей, описанных более подробно в примере 13.
На фиг.18 приведены графики цитотоксичности для случая хронического лимфолейкоза (ХЛЛ, CCL) и для нормальных В-клеток человека в присутствии TRA-8, BISVIII и их сочетания.
На фиг.19А показано специфичное связывание 2Е12 с DR4. На пластинки для проведения ELISA наносят растворимую форму слитых белков рецептора человеческого TRAIL, человеческого IgG1 Fc, как показано, и инкубируют с указанными концентрациями мАт 2Е12 и затем с ПХ-конъюгированным антителом против мышиного IgG1. Реакция проходит при использовании субстратного буфера TMB, и значения ОП измеряют при длинах волн 450/650 нМ.
На фиг.19В показано связывание 2Е12 с DR5 на клеточной поверхности. Cos-7 клетки трансфицируют вектором, содержащим кДНК для DR4 полной длины (жирная гистограмма) или контрольный вектор (открытая гистограмма). Трансфицированные клетки окрашивают 10 мкг/мл 2Е12 и ФЭ-конъюгированным антителом против мышиного IgG1. Клетки анализируют по методу проточной цитометрии.
На фиг.19С показана апоптоз-индуцирующая активность 2Е12. Клетки лимфомы человека Ramos B инкубируют в течение ночи с указанной концентрацией 2Е12 в присутствии 2 мкг/мл антитела против мышиного IgG1. Жизнеспособность клеток определяют по методу ATPLite. (D) Активация каспазы индуцируется добавлением 2Е12. Клетки Ramos обрабатывают 2Е12 и антителом против мышиного IgG в указанные временные точки. Активацию каспазы и расщепление под действием PARP определяют вестерн-блот-анализом с использованием специфических антител к каспазе или PARP.
На фиг.20 показан эффект 2Е12 и адриамицина у бестимусных мышей nude, содержащих ксенотрансплантаты рака молочной железы. Клетки 2LMP (3·107) инъецируют подкожно бестимусным мышам nude на день 0. Двум группам мышей инъецируют внутрибрюшинно 200 мкг 2Е12 в дни 7, 10, 14, 17, 21 и 24. Две группы мышей получают в/в адриамицин (6 мг/кг) в дни 8, 12 и 16. Одной группе мышей вводят антитело. Полученные результаты выражают в виде среднего значения изменений размеров опухоли относительно ее размера на день 7 (n=8 мышей/группу).
На фиг.21 показан эффект TRA-8, 2Е12 и адриамицина у бестимусных мышей nude, содержащих ксенотрансплантаты рака молочной железы. Клетки 2LMP (3·107) инъецируют подкожно (п/к) бестимусным мышам nude на день 0. Двум группам инъецируют внутрибрюшинно (в/б) 200 мкг TRA-8 и 2Е12 в дни 7, 10, 14, 17, 21 и 24. Двум группам мышей вводят внутривенно (в/в) адриамицин (6 мг/кг) в дни 8, 12 и 16. Одной группе мышей не вводят антитело. Полученные результаты выражают в виде среднего значения изменений размеров опухоли относительно ее размера на день 7 (n=8 мышей/группу).
На фиг.22А показаны результаты анализа методом проточной цитометрии экспрессии DR5 на клеточной поверхности на плане клеточных линий рака молочной железы человека. Клетки рака молочной железы собирают с использованием ЭДТА и окрашивают с использованием 10 мкг/мл мАт TRA-8 в течение 1 часа при температуре 4°С и затем обрабатывают ФЭ-конъюгированным козьим антителом против мышиного IgG с последующим анализом с использованием флуоресцентного клеточного сортировщика FACScan и программного обеспечения CellQuest. Жирные линии гистограммы показывают наличие окрашивания под действием TRA-8, а тонкие линии гистограммы указывают на результаты инкубации с контрольным мышиным антителом IgG1 изотипа.
На фиг.22В показана цитотоксичность TRA-8 для клеточных линий рака молочной железы человека. Клетки обрабатывают трипсином и вновь высевают на планшеты с плотностью 1000 клеток/лунку в 96-луночном планшете. После посева клеток добавляют антитело TRA-8 и проводят инкубацию в течение 24 часов при 37°С. Жизнеспособность клеток оценивают через 24 часа после добавления TRA-8 с использованием теста ATPLite. Определяют уровень АТФ относительно необработанных контрольных клеток в виде среднего значения и СКО для 2-3 определений в независимых экспериментах, выполненных в тройном повторе.
На фиг.23А показана цитотоксичность сочетанной обработки с использованием TRA-8 и адриамицина для клеточных линий рака молочной железы человека. Клетки (1000/лунка) подвергают воздействию различной концентрации адриамицина в течение 24 часов при 37°С, начиная с точки 24 часа после высева клеток. Через 24 часа после добавления адриамицина вносят TRA-8 и определяют уровни АТФ еще через 24 часа. Определяют среднее значение и величину СКО для тройных повторных определений, выполненных в ходе 2-4 независимых экспериментов, и выражают их относительно результатов, полученных для необработанных контрольных клеток.
На фиг.23B показана цитотоксичность для клеточных линий рака молочной железы человека сочетанной обработки TRA-8 и паклитакселом. Клетки (1000/лунка) подвергают воздействию различных концентраций паклитаксела в течение 24 часов при 37°С, начиная с точки 24 часа после высева клеток. Через 24 часа после внесения паклитаксела добавляют TRA-8 и еще через 24 часа определяют уровни АТФ. Определяют среднее значение и величину СКО для тройных повторных определений, выполненных в ходе 2-4 независимых экспериментов, и выражают их относительно результатов, полученных для необработанных контрольных клеток.
На фиг.24 показан эффект TRA-8 на рост опухоли у бестимусных мышей nude, содержащих установленные ксенотрансплантаты линии рака молочной железы человека 2LMP. Клетки 2LMP (3·107) инъецируют п/к на день 0. Двум группам мышей инъецируют внутрибрюшинно 200 мкг или 600 мкг TRA-8 в дни 7, 10, 14, 17, 21 и 24. Одной группе мышей не вводят антитело. Представленные данные отражают среднее значение изменений размеров опухоли (продукт двух диаметров) относительно ее размера на день 7 (n=8 мышей/группу).
На фиг.25 показан эффект воздействия TRA-8 и адриамицина на рост опухоли у бестимусных мышей nude, содержащих ксенотрансплантаты клеточных линий рака молочной железы. Клетки 2LMP (3·107) инъецируют п/к бестимусным мышам nude на день 0. Двум группам инъецируют в/б 200 мкг TRA-8 в дни 7, 10, 14, 17, 21 и 24. Две группы мышей получают в/в адриамицин (6 мг/кг) в дни 8, 12 и 16. Одной группе мышей не вводят антитело. Полученные результаты выражают в виде среднего значения изменений размеров опухоли относительно ее размера на день 7 (n=8 мышей/группу).
На фиг.26 показан эффект воздействия TRA-8 и паклитаксела на бестимусных мышей nude, содержащих ксенотрансплантаты клеток рака молочной железы. Клетки 2LMP (3·107) инъецируют п/к бестимусным мышам nude на день 0. Двум группам мышей инъецируют в/б 200 мкг TRA-8 в дни 7, 10, 14, 17, 21 и 24. Двум группам мышей вводят в/в паклитаксел (20 мг/кг) в дни 8, 12, 16, 20 и 24. Одной группе мышей не вводят антитело. Полученные результаты выражают в виде среднего значения изменений размеров опухоли относительно ее размера на день 7 (n=8 мышей/группу).
На фиг.27 показан эффект TRA-8, адриамицина и облучения 60Co на рост опухоли у бестимусных мышей nude, содержащих ксенотрансплантаты клеток рака молочной железы. Клетки 2LMP (3Х107) инъецируют п/к бестимусным мышам nude на день 0. Трем группам мышей инъецируют в/б 200 мкг TRA-8 в дни 7, 10, 14, 17, 21 и 24. Двум группам мышей вводят в/в адриамицин (6 мг/кг) в дни 8, 12 и 16. Четыре группы мышей получают облучение 60Co в дозе 3 Gy в дни 9 и 17. Одной группе мышей не вводят антитело. Полученные результаты выражают в виде среднего значения изменений размеров опухоли (продукт двух диаметров) относительно ее размера на день 7 (n=8 мышей/группу).
Подробное описание изобретения
Неспособность удалять клетки связана с дефектами в системе индукции апоптоза, которые определяются дефектами, включающими экспрессию или функционирование лиганда, рецептора или внутриклеточных регуляторных и эффекторных молекул. Настоящее изобретение предлагает способ коррекции системы индукции недостаточного апоптоза, а также выяснения специфических дефектов, присущих данной дефектной системе индукции апоптоза.
Настоящее изобретение относится к новому классу моноклональных антител, которые являются селективными по апоптоз-индуцирующей активности in vivo и in vitro в отношении специфических TRAIL-рецепторов, включая DR5, DR4, DсR1 и DсR2. Таким образом, антитела по настоящему изобретению специфически связываются с одним из TRAIL-рецепторов. Термины «селективное связывание» или «специфическое распознавание» означают, что антитело связывается только с одним рецептором TRAIL и демонстрирует незначительное или отсутствие связывания с другими типами TRAIL-рецепторов, как следует из данных традиционного анализа по методу вестерн-блоттинга. Антитело DR5 по настоящему изобретению селективно связывает с DR5 и демонстрирует уровень связывания, превышающий фоновое связывание с DR4, DсR1 или DсR2 в 1,5 раза. Аналогично антитело DR4 по настоящему изобретению связывается селективно с DR4 и не демонстрирует связывания, превышающего фоновое связывание DR5, DсR1 или DсR2 более чем в 1,5 раза. Настоящее изобретение может быть применимо для использования в качестве реагента в исследованиях сигнальных механизмов апоптоза, а также в качестве терапевтически эффективного средства против клеток, экспрессирующих TRAIL-рецепторы, включающих в качестве иллюстративных примеров широкий перечень классов злокачественных клеток, клеток с разбалансированной системой апоптоза, активированные лимфоциты или другие активированные иммунные клетки (например, лимфоидные клетки и миелоидные клетки), клетки, инфицированные вирусом и аномально пролиферирующие синовиальные клетки (например, синовиальные клетки при ревматоидном артрите, включая воспалительные синовиальные клетки, активированные лимфоидные и миелоидные клетки синовиальной оболочки, макрофагоподобные синовиоциты и фибробластоподобные синовиоциты) при аутоиммунных заболеваниях. Антитела по настоящему изобретению характеризуются специфичностью по связыванию лишь с определенными типами TRAIL-рецепторов несмотря на существующую между ними гомологию. Антитела по настоящему изобретению вызывают направленный апоптоз только тех клеток, которые экспрессируют целевой TRAIL-рецептор или, альтернативно, блокируют TRAIL-опосредованный апоптоз клеток, экспрессирующих целевой рецептор.
Моноклональное антитело DR5 или моноклональное антитело DR4 по настоящему изобретению служит мощным индуктором апоптоза в клетках, экспрессирующих DR5 или DR4 соответственно in vitro и мощным индуктором апоптоза in vivo. Гуманизированные последовательности фрагментов CDR, имеющиеся на скелете молекулы гуманизированного антитела и слитом белке на основе DR5 или DR4 антител по настоящему изобретению демонстрируют аналогичные апоптозные свойства.
К настоящему времени недоступно моноклональное антитело, которое связывалось бы с DR5 на клеточной поверхности и которое даже в низких концентрациях индуцировало бы апоптоз клеток, экспрессирующих DR5 in vitro и in vivo в отсутствие сшивателя. Настоящее изобретение относится к антителу DR5, функционирующему в качестве терапевтического средства при лечении множества заболеваний. Хотя было показано, что растворимый TRAIL эффективен в плане индукции апоптоза опухолевых клеток in vivo, его активность по уничтожению клеток, по всей видимости, является очень низкой на фоне потребности во введении больших и повторных доз [13]. Настоящее изобретение предлагает очищенное антитело, которое связывает TRAIL-рецептор DR5, причем указанное антитело в своей растворимой форме при низких концентрациях обладает апоптоз-индуцирующей активностью in vivo и in vitro в отношении целевых клеток, экспрессирующих DR5. В предпочтительном варианте осуществления изобретения очищенное антитело связывает TRAIL-рецептор DR5 в отсутствие сшивателя для антитела. Предпочтительно указанное антитело не индуцирует значительного апоптоза нормальных фибробластных клеток. Предпочтительно апоптоз-индуцирующая активность характеризуется достижением менее чем 60%, 50%, 40%, 30%, 20%, 10%, 5% жизнеспособности или любого другого процентного уровня в интервале между указанными значениями целевых клеток в концентрации антитела менее чем 0,1, 1, 5, 10 или 20 мкг/мл или любой концентрации в интервале между ними. Очищенное антитело специфически связывается с TRAIL-рецептором DR5 и не связывается с рецепторами DR4, DсR1, DсR2, как следует из результатов рутинного анализа по методу вестерн-блоттинга. В предпочтительном варианте осуществления изобретения указанное антитело представляет собой моноклональное антитело, предпочтительно характеризующееся той же эпитопной специфичностью, что и мышиная-мышиная гибридома TRA-8, зарегистрированная с номером доступа ATCC PTA-1428.
TRA-8, один из серии антител DR5 по настоящему изобретению, является фармацевтически эффективным средством у животных, содержащих трансген человеческого DR5, и также может быть полезным с точки зрения установления модели для исследования роли DR5 и TRAIL.
В различных вариантах осуществления настоящего изобретения предлагаются антитела, которые индуцируют апоптоз в присутствии или в отсутствие сшивки. Так, например, в предпочтительном варианте осуществления изобретения антитело DR5 (например, TRA-8) индуцирует апоптоз в отсутствие сшивателя. В других вариантах осуществления изобретения предлагаются антитела, которые индуцируют апоптоз в присутствии сшивателей, включая, например, предпочтительный вариант с использованием антитела DR4 (2Е12).
Таким образом, настоящее изобретение предлагает очищенное антитело, которое специфически связывается с TRAIL-рецептором DR4, причем указанное антитело в своей растворимой форме обладает in vivo и in vitro активностью по индукции апоптоза в целевых клетках, экспрессирующих DR4. В качестве одного из вариантов осуществления настоящего изобретения предлагаемое антитело является моноклональным антителом, характеризующимся той же эпитопной специфичностью, что и гибридома 2Е12 с номером доступа ATCC PTA-3798, депонированная 24 октября 2001 года под названием «клон гибридомы 2Е12 против человеческого DR4» («2E12 Hybridoma Clone Against Human DR4») от имени исследовательского фонда UAB. 2Е12, относящийся к серии DR4 по настоящему изобретению, представляет собой фармацевтически активное средство, действующее в направлении снижения размера опухоли в сравнении с необработанными контрольными животными или в сравнении с размером до лечения опухоли у животных in vivo, содержащих раковые клетки, экспрессирующие DR4.
Антитела к DR4 и DR5 эффективны в растворимой форме в малых дозах, причем под малыми дозами понимаются дозы или концентрации менее чем примерно от 0,01 до 1 мкг/мл in vitro, и менее чем примерно 1-10 мг/кг in vivo. Предпочтительной особенностью антител по настоящему изобретению является их способность индуцировать апоптоз селективно в клетках, экспрессирующих рецепторы DR5 или DR4, не индуцируя апоптоз в нормальных неактивированных нетрансформированных гепатоцитах, фиброцитах, синовиоцитах, и т.п. Антитело по настоящему изобретению против TRAIL-рецептора отбирают, согласно настоящему изобретению, от экспериментального животного, но оно может быть получено любыми методами получения или синтеза антител, известными в данной области. Гуманизацию антитела по настоящему изобретению осуществляют с целью поддержания его активности по связыванию с рецептором при проявлении сниженного и терапевтически толерантного иммунного ответа у человека, так что получаемое гуманизированное антитело против TRAIL-рецептора по настоящему изобретению используется в качестве терапевтического агониста или антагониста для данного рецептора. Поскольку данное изобретение описывает терапевтическое средство, действующее in vivo, то вторичное сшивание антитела против TRAIL-рецептора уже становится необязательным.
Настоящее изобретение относится не просто к одному антителу против TRAIL-рецептора, обладающему агонистическим или антагонистическим апоптозным эффектом. Скорее, оно относится к двум или более антителам против TRAIL-рецептора, которые приводятся в контакт с культурой клеток in vitro или с тканью организма in vivo для достижения улучшенного результата лечения. Под термином «улучшенный результат лечения» следует понимать любой аддитивный, синергический или потенциирующий эффект. Так, например, клеточная линия глиомы U87 и гемопоэтические линии U937 и Molt-4 приводят к достижению синергической реакции на агонистические антитела против DR4 и DR5, тогда как воздействие одного агонистического антитела против DR5 демонстрирует лишь ограниченный успех по индуцированию апоптоза.
Дополнительно антагонистические антитела против TRAIL-рецептора могут иметь особый вариант применения согласно настоящему изобретению, при котором антитело обладает специфичностью по связыванию с одним из ложных рецепторов DсR1, DсR2 или OPG. Селективное блокирование ложного рецептора антителом по настоящему изобретению дает эффект в типах клеток, экспрессирующих ложные рецепторы, путем переключения равновесия по связыванию TRAIL к тем рецепторам TRAIL, которые способны к трансдукции апоптозного сигнала. Таким образом, согласно другому варианту комбинированной терапии по настоящему изобретению антитело, связывающееся с ложным рецептором, сенсибилизирует экспрессирующую клетку к агонистическому связыванию TRAIL-рецептора, переносящего апоптозный сигнал.
В другом варианте осуществления изобретения предлагается способ выявления агонистических и антагонистических эпитопов для данного TRAIL-рецептора. Далее выявляется полиморфизм, имеющийся между индивидуумами, связанный с данным TRAIL-рецептором, по способу настоящего изобретения путем использования панели моноклональных антител, каждая из которых содержит отличающийся вариабельный или CDR участок. Характерная панель моноклональных антител обеспечивает возможность определять агонистические и антагонистические эпитопы и наличие полиморфизма. Таким образом, панель моноклональных антител по настоящему изобретению может найти применение при поиске лекарственных средств и/или скрининге субъектов на склонность к заболеванию.
В еще одном варианте настоящее изобретение относится к слитым белкам, включающим антигенный фрагмент TRAIL-рецептора, связанный с иммуноглобулиновым белком, полипептидом или их фрагментом. Фрагмент рецептора TRAIL определяется как такой фрагмент, которые содержит достаточное число оснований для проявления иммуногенной реакции на нативный TRAIL-рецептор, экспрессируемый на клеточной поверхности субъекта. Фрагмент слияния с TRAIL-рецептором включает по меньшей мере десять аминокислот. Иммуноглобулиновый слитый белок или его фрагмент в контексте настоящего описания включает нативный или синтетический белок или полипептидный сегмент, содержащий достаточное количество аминокислотных оснований для активации иммуногенного каскада реакций у субъекта. Иммуноген по настоящему изобретению, включающий слитый белок на основе фрагмента TRAIL-рецептора, сопряженного с иммуноглобулиновым фрагментом, может быть применим в качестве терапевтического средства in vivo для выявления у субъекта антитела против TRAIL-рецептора in situ.
В еще одном варианте своего осуществления настоящее изобретение применимо в качестве способа генной терапии. Таким образом, настоящее изобретение предлагает способ селективной индукции апопотоза в целевых клетках, включающий стадии трансфекции целевых клеток вектором, включающим экспрессируемую последовательность нуклеиновой кислоты для TRAIL-рецептора, экспрессию в указанных клетках TRAIL-рецептора, кодируемого указанной последовательностью нуклеиновой кислоты для TRAIL-рецептора, и контакт указанных клеток с апоптоз-индуцирующим антителом, селективным по связыванию с данным TRAIL-рецептором. В аспекте применимости в генной терапии по настоящему изобретению целевые клетки подвергают трансфекции вектором, несущим экспрессируемую последовательность, соответствующую TRAIL-рецептору, при этом вектором является традиционный вектор, выбираемый на основе чувствительности данной целевой клетки к вектору. Векторы для генной терапии могут включать в качестве примеров аденовирус pAdCMV5. При экспрессии в целевых клетках или тканях трансфицированного TRAIL-рецептора указанные клетки или ткани подвергаются воздействию антитела по настоящему изобретению, специфичному по связыванию с трансфицированным TRAIL-рецептором. Считается, что антитело к TRAIL-рецептору обладает агонистическим или антагонистическим действием, что соответствует желательному терапевтическому результату.
Антитела по настоящему изобретению также функционируют в сочетании с сенсибилизатором. Сенсибилизатор в контексте настоящего описания определяется как любой стимул, который индуцирует апоптоз, включая ультрафиолетовый свет, органические молекулы, конкретно класс бисиндолмалеимидов, тяжелые металлы и свободные радикалы.
В том что касается лечения злокачественной опухоли, существенно, что TRA-8 способен индуцировать апоптоз большей части TRAIL-чувствительных опухолевых клеток зависимым от каспазы образом в отсутствие вторичного сшивания. И TRA-8, и 2Е12 поодиночке или в сочетании проявляют сильную активность in vivo по уничтожению опухолевых клеток. Активность TRA-8 или 2Е12 по индукции апоптоза большей части TRAIL-чувствительных опухолевых клеток подтверждает тот факт, что одного DR5 или DR4 достаточно для запуска апоптоза. Большая часть опухолевых клеток из числа подробно описываемых в настоящем изобретении экспрессирует на клеточной поверхности DR5, и их чувствительность к TRA-8-индуцированной гибели клеток согласуется с их чувствительностью к TRAIL, указывая на то, что DR5 представляет собой первичный рецептор, вызывающий гибель клеток по механизму TRAIL-опосредованного апоптоза, в большей части опухолевых клеток. Аналогичные результаты были получены с антителами, специфичными к DR4 (например, к 2Е12). Таким образом, дифференциальная экспрессия DR5 или DR4 нормальными или злокачественными клетками используется в механизме селективности TRAIL-опосредованного апоптоза. TRA-8 обходит ложные рецепторы, индуцируя TRAIL-опосредованный апоптоз. Лишь незначительное число TRAIL-резистентных опухолевых клеток чувствительны к TRA-8, что, однако, указывает на то, что ложные рецепторы, по всей видимости, не играют важной роли в устойчивости опухолевых клеток к TRAIL-опосредованному апоптозу.
Несмотря на то что предшествующие исследования показали, что системное введение растворимой формы TRAIL животным индуцирует регрессии опухоли без сопутствующей токсичности [3,4,22], связанной с мембраной форма человеческого TRAIL индуцирует поражение печени у мышей, как будет показано далее в настоящем описании. Однако токсичность TRAIL для лечения является значительно менее выраженной, чем Fas-лиганда, на что указывает меньшая чувствительность нормальных гепатоцитов к TRAIL-индуцированному повреждению в сравнении с повреждением Fas-лигандом и отсутствие летальности от TRAIL in vivo. Таким образом, титрование с использованием TRAIL может использоваться в лечении злокачественной опухоли.
Показано, как будет более подробно описано далее, что для нормальных гепатоцитов нехарактерна экспрессия DR5 белка в существенной мере, что связано с устойчивостью гепатоцитов к апоптозу, индуцированному TRA-8. Сшивания DR5 с моноклональным антителом недостаточно для организации гомополимерных форм в рецепторе, вызывающем гибель клеток, которая способна бы была запустить апоптоз.
Эксперименты с мартышками доказали отсутствие признаков токсичности для печени при введении TRA-8. Таким образом, агонистическое моноклональное антитело DR5, скорее всего, представляет собой более селективное и безопасное средство, чем растворимый TRAIL, в качестве терапевтического средства. Аналогично DR4 экспрессируется трансформированными или активированными клетками и не экспрессируется в заметных количествах или экспрессируется лишь в значительно меньших количествах нормальными клетками, например фибробластами. DR4 по настоящему изобретению индуцирует апоптоз определенных целевых клеток, не вызывая при этом в значимых количествах гибели нецелевых клеток, таких как фибробласты и т.п. В контексте настоящего описания отсутствие эффекта или отсутствие заметного или выраженного эффекта включает полное отсутствие эффекта или эффект, который ниже или соответствует фоновому или контрольному уровню, и не превосходит фоновый или контрольный уровень более чем в 1,5 раза.
В качестве инструмента для скрининга или для визуализации настоящее изобретение хорошо подходит для целей обнаружения небольших кластеров DR4 или DR5 клеток, которые еще могут демонстрировать морфологию нормальной клетки. Так, например, окрашивание in situ срезов злокачественных клеток человека, включающих раковые клетки легкого, предстательной железы и печени, меченными антителами по настоящему изобретению, позволяет легко идентифицировать злокачественные клетки. Антитела по настоящему изобретению также полезны при скрининге проявления других заболеваний, включая, например, различные воспалительные или аутоиммунные заболевания, такие как, например, ревматоидный артрит. Такой скрининг может быть полезен даже до проявления других клинических симптомов и может использоваться для скрининга риска развития заболевания у субъекта, так что профилактическое лечение может начаться до проявления других признаков или симптомов. Конкретно было показано, что злокачественные клетки экспрессируют очень высокий уровень DR5 в сравнении с нормальными клетками того же типа. Таким образом, настоящее изобретение может быть применимо как чувствительный способ скрининга ранней стадии злокачественных новообразований в тканях, включая по меньшей мере ткань легкого, предстательной железы, толстого кишечника, крови, шейки матки, молочной железы и печени. Способ лечения, направленный на подавление аномальной пролиферации клеток, связанной с заболеваниями, включающими в качестве примера в числе других злокачественные новообразования и лимфолейкозы, будет подробно описан в настоящем изобретении.
В настоящем изобретении особенно подробно описывается моноклональное антитело против человеческого DR5, обозначенное как TRA-8, зарегистрированное под номером PTA-1428. Признано, что методики и результаты, отработанные для агонистического моноклонального антитела TRA-8 против человеческого DR5, полностью могут быть применимы к антагонистическим антителам против DR5, а также к антителам против DR4, DсR1, DсR2, действующих как агонистическим, так и антагонистическим образом. Таким образом, настоящее изобретение подробно описывается применительно к апоптоз-индуцирующему антителу, которое специфично к человеческому DR4. В одном варианте осуществления изобретения антитело имеет ту же эпитопную специфичность, что и гибридома 2Е12, депонированная 24 октября 2001 года и получившая номер доступа от имени исследовательского фонда UAB в Американской Коллекции типовых культур в Роквилле (American Type Culture Collection, Rockville, Md.). Описание депонированного материала называется «Клон гибридомы 2Е12 против человеческого DR4» («2E12 Hybridoma Clone Against Human DR4»), депонированный штамм получил обозначение 2Е12, и файлу с документом присвоен номер PCT/US01/14151. Уровни экспрессии апоптозного рецептора, такого как Fas, не обязательно коррелируют с чувствительностью клеток к апоптозу. Для случая TRAIL-опосредованного апоптоза было высказано предположение, что экспрессия ложных рецепторов для TRAIL влияет на чувствительность клеток. Кроме того, было высказано предположение о том, что DR5 должен ассоциироваться с DR4 для достижения эффективной трансдукции апоптозного сигнала через пути, включающие FADD и каспазу 8. Доступность агонистического моноклонального антитела против DR5 позволяет оценивать регуляцию сигнальной функции DR5 и ее относительную роль в TRAIL-опосредованном апоптозе. Сравнение чувствительности клеток к TRA-8-опосредованонму апоптозу с их чувствительностью к TRAIL-опосредованному апоптозу позволяет прояснить роль DR5 в TRAIL-опосредованном апоптозе и механизмы, которые могут воздействовать на чувствительность. Аналогичные плюсы дает использование антитела к DR4.
Указанное преимущество в основном распространяется на гуманизированные антитела DR5 и DR4 по настоящему изобретению. Так, например, молекулярный клон антитела против DR5 получают с использованием известных методик, которые подробно описаны в приведенных ниже примерах. В настоящем изобретении использованы методы рекомбинантной ДНК [33] для конструирования последовательностей нуклеиновых кислот, которые кодируют молекулу моноклонального антитела или ее участок, ответственный за связывание с антигеном.
Настоящее изобретение позволяет конструировать гуманизированные антитела к TRAIL-рецептору, которые, скорее всего, не будут индуцировать образование человеческого антитела против мышиного иммуноглобулина (обозначаемого в описании как HAMA) [34], но все еще будут оставаться активными эффекторами функции антитела. Полностью соответствующие человеческим антитела могут быть также получены путем иммунизации мышей, способных к образованию полного человеческого антитела (например, мышей, генетически модифицированных для целей продукции человеческих антител), скрининга клонов, которые связывают DR5 или DR4, индуцируют апоптоз и конкурируют с TRA-8 или 2Е12 эпитопом. См., например, работу Лонберга и Хушара (Lonberg and Huszar (1995) Human antibodies from transgenic mice, Int. Rev. Immonol. 13:65-93), которая включена в настоящее описание полностью в качестве ссылки, содержащая описание способов получения полностью сформированных человеческих антител. В контексте настоящего описания термины «человеческий» и «гуманизированный» в отношении антитела относятся к любому антителу, которое, как ожидается, будет вызывать терапевтически толерантный слабый иммуногенный ответ у человека.
Настоящее изобретение относится к антителу DR5, гуманизированному антителу против DR5, TRA-8-иммуноглобулинам с тяжелой и легкой цепью и к гуманизированным иммуноглобулинам с тяжелой и легкой цепью. Настоящее изобретение также относится к антителу DR4, гуманизированному антителу против DR4, иммуноглобулинам антитела DR4 с тяжелой и легкой цепью и к гуманизированным иммуноглобулинам с тяжелой и легкой цепью, нуклеиновым кислотам, которые кодируют антитела и тяжелую и легкую цепи, к векторам, включающим указанные нуклеиновые кислоты, и к клеткам, включающим указанные векторы. Некоторые усеченные формы таких белков или генов осуществляют регуляторную или энзиматическую функции полной последовательности белка или гена. Так, например, последовательности нуклеиновой кислоты, кодирующие их, могут быть изменены путем замещений, добавлений, делеций или путем мультимерной экспрессии, которые обеспечивают получение функционально эквивалентных белков или генов. В связи с вырожденностью кодирующих последовательностей нуклеиновой кислоты в практике осуществления настоящего изобретения могут использоваться другие последовательности, которые кодируют по существу те же самые аминокислотные последовательности, что и природные белки. Указанные последовательности включают, не ограничиваясь, последовательности нуклеиновой кислоты, включающие полностью или части последовательностей нуклеиновых кислот, кодирующих указанные выше полипептиды, которые были изменены путем замещения различных кодонов, кодирующих функционально эквивалентные аминокислотные остатки в последовательности с достижением таким образом молчащих изменений. Считается, что нуклеотидная последовательность иммуноглобулина по настоящему изобретению толерантна к вариациям с точки зрения гомологии последовательностей до 25%, как следует из расчетов по стандартным методам («Current Methods in Sequence Comparison and Analysis», Macromolecule Sequencing and Synthesis, Selected Methods and Applications, pp. 127-149, 1998, Alan R. Liss, Inc.), т.е. в той мере, в которой такой вариант образует функциональное антитело, способное распознавать TRAIL-рецептор DR5.
Так, например, один или более аминокислотных остатков в полипептидной последовательности могут быть замещены другой аминокислотой со сходной полярностью, которая действует как функциональный эквивалент, приводя к образованию молчащего изменения. Заместители аминокислоты в данной последовательности могут быть выбраны из других представителей того класса, к которому принадлежит данная аминокислота (т.е. при консервативном замещении). Так, например, неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин. Полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глютамин. Положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин. Отрицательно заряженные (кислые) аминокислоты включают аспарагиновую кислоту и глютаминовую кислоту. В область настоящего изобретения включаются также белки или их фрагменты или производные, которые были по-разному модифицированы во время трансляции или после нее, например, путем гликозилирования, протеолитического расщепления, связывания с молекулой антитела или другими клеточными лигандами и т.п. Дополнительно, рекомбинантный вектор, кодирующий последовательности нуклеиновой кислоты антител по настоящему изобретению, может быть сконструирован генно-инженерными методами с целью модификации процессинга или экспрессии такого вектора. Другие модификации могут быть осуществлены в последовательности нуклеиновой кислоты или аминокислотной последовательности без снижения или без существенного снижения апоптозной активности антитела. Такие модификации могут быть осуществлены в CDR или в участках, не содержащих CDR, с использованием известных в данной области методик. См., например, работу Янга с соавт. (Yang et al. (1995), J. Mol. Biol. 254:392-403), которая включена в настоящее описание полностью в отношении методик проведения мутагенеза CDR-участков.
Дополнительно ингибитор, кодирующий последовательность нуклеиновой кислоты, может быть мутирован in vitro или in vivo с созданием и/или разрушением последовательности трансляции, инициации и/или терминации, или с созданием вариаций в кодирующих участках, и/или с созданием новых сайтов для рестрикционных эндонуклеаз, или с разрушением ранее существовавших сайтов для облегчения дальнейших модификаций in vitro. Могут использоваться любые методики мутагенеза, известные в данной области, включающие, без ограничения, направленный мутагенез in vitro (J. Biol. Chem. 253:6551), использование Tab линкеров (Pharmacia) и т.п.
Данные рентгеновской кристаллографии указывают на то, что складчатая структура иммуноглобулина антитела в основном представляет собой длинную цилиндрическую структуру, включающую два слоя антипараллельных b-полос, каждая из которых состоит из трех или четырех b-цепей. В вариабельном участке три петли из каждого из V-доменов кластера H- и L-цепи вместе образуют антигенсвязывающий сайт. Каждая из этих петель носит название комплементарного детерминирующего участка (КДУ (CDR)). КДУ обладают наибольшей вариабельностью по аминокислотной последовательности в антителе. Части вариабельного участка, которые не входят в КДУ, носят название «рамочные регионы» («FR» регионы) и в основном играют роль в поддержании структуры КДУ. Предпочтительно все КДУ в данном антителе переходят на акцепторное антитело для сохранения связывающего участка на эпитопном регионе TRAIL-рецептора. Считается, что в доноре при пересаживании на него части от общего числа КДУ поддерживается соответствующее функционирование. Очевидно, что процесс пересаживания в целом обеспечивается заменой остатка за остатком аминокислоты или соответствующего участка на другой. Однако иногда, особенно при переносе участка один или более остатков могут быть добавлены, или изъяты, или замещены при желании, и такие делеции и вставки, а также соответствующие замены и инверсии также известны в данной области техники.
Антитело по настоящему изобретению получают, например, путем пересаживания каждого КДУ от субъединицы L-цепи или H-цепи моноклонального антитела против TRAIL-рецептора в соответствующий КДУ человеческого антитела, что дает гуманизированное мышиное моноклональное антитело, эффективное против TRAIL-рецептора.
Фрагменты антитела, которые содержат идиотип соответствующей молекулы в функциональном виде, могут быть также получены с использованием известных методик. Так, например, такие фрагменты включают фрагмент анти-TRAIL-рецептора (AB')2, который может быть получен путем расщепления пепсином молекулы антитела, фрагменты антитела к TRAIL-рецептору AB', полученные путем восстановления дисульфидных связей во фрагменте TRAIL-рецептора (AB')2 и фрагмент антитела, который получают при обработке молекулы антитела папаином и восстановителем.
Антитела по настоящему изобретению могут быть получены с использованием многочисленных методик, известных в технике. В качестве примера можно отметить, что моноклональное антитело TRA-8 против DR5 может быть получено при культивировании гибридомы, которая, в свою очередь, может быть получена путем иммунизации мыши человеческим DR5 с последующим слиянием клеток селезенки или клеток лимфатического узла мыши с миеломными клетками мыши.
Получение моноклонального антитела включает в качестве наглядных примеров следующие стадии:
а) очистка биологической макромолекулы с целью использования ее в качестве антигена;
b) получение клеток, продуцирующих антитело, после первой иммунизации животного с использованием инъекций антигена с последующим отбором крови у животного и определением титра антител с целью определения времени, подходящего для удаления селезенки;
c) получение миеломных клеток;
d) слияние клеток, продуцирующих антитело, и миеломных клеток;
е) отбор гибридомы, продуцирующей желательное антитело;
f) получение клона одной клетки (клонирование);
g) необязательное культивирование клеток гибридомы или выращивание животных, которым были трансплантированы клетки гибридомы, для крупномасштабного получения моноклонального антитела;
h) тестирование биологической активности и специфичности или анализ маркерных свойств полученного моноклонального антитела.
Процедура получения моноклонального антитела подробно описана ниже со ссылками на указанные выше стадии. Описываемый способ получения антитела по настоящему изобретению дается лишь с целью иллюстрации способов получения и не ограничивает настоящее изобретение. Могут быть использованы другие известные методики, или указанные методики могут быть модифицированы, например, путем использованием в качестве клеток, продуцирующих антитело, иных клеток, отличных от клеток селезенки и миеломных клеток.
(а) Получение антигена
Рекомбинантный белок (называемый в описании как «рекомбинантный человеческий DR5» или «рекомбинантный человеческий DR4»), эффективный в качестве антигена, получают при трансфекции клеток QBI-293A вектором pAdDR5-IgG с получением слитого белка, включающего внеклеточный домен человеческого DR5 или DR4 и Fc участка человеческого антитела IgG1 (далее обозначаемого как «IgG»), (№ PTA-1428) для экспрессии его с использованием набора ADENO-Quesi (Quantum Biotechnologies Inc., Канада) с последующим отбором и частичной очисткой продукта экспрессии. Плазмиду pAdDR5-IgG конструируют путем вставки ДНК, кодирующей человеческий DR5 или DR4 и человеческий слитый белок IgG, в pAdCMV5, который представляет собой вектор экспрессии для клеток животного. Могут использоваться также другие материалы, такие как ДНК, кодирующая DR5 или DR4, вектор и клетка-хозяин.
Слитый белок человеческих DR5 или DR4 и IgG, получаемый в супернатанте культуры клеток QBI-293A, трансфицированных вектором pAdDR5-IgG, могут быть очищены аффинной хроматографией на колонке, содержащей белокА-сефарозу, или аффинной хроматографией на колонке, содержащей белокG-сефарозу, или путем ионообменной хроматографии на колонке Resourse Q (торговое наименование, Pharmacia).
Альтернативно в качестве антигена может использоваться очищенный DR5 или DR4, полученный из клеточных мембран клеточных линий человека. Далее, поскольку первичная структура DR4 и DR5 известны (№ PTA-1428), пептид, включающий аминокислотную последовательность SEQ ID No. 1, может быть химически синтезирован известным способом, таким как метод Сэнджера, и использоваться дальше в качестве антигена.
(b) Получение клеток, продуцирующих антитело
Мышь иммунизируют иммуногеном, полученным на стадии (а), смешанным с адъювантом, таким как полный или неполный адъювант Фрейда или алюм. Другие примеры подходящих для эксперимента животных включают крыс, морских свинок, кроликов, собак, кур, лошадей, свиней, коров и овец.
Подходящие способы введения с целью иммунизации экспериментального животного включают подкожный, внутрибрюшинный, внутривенный, внутрикожный и внутримышечный способы инъекции, при этом подкожный и внутрибрюшинный способы инъекции являются предпочтительными.
Иммунизацию необязательно проводят путем введения однократной дозы или путем введения нескольких повторных доз с соответствующими интервалами (предпочтительно от 1 до 5 недель). Иммунизированных животных отслеживают на наличие в сыворотке титра антител, и животных с достаточно высоким титром антител отбирают в качестве источника клеток, продуцирующих антитело. Отбор животных с высоким титром делает последующий процесс более эффективным. Клетки для проведения впоследствии слияния в основном собирают от животного через 3-5 дней после последней иммунизации.
Способы определения титра антител включают различные известные методики, такие как радиоиммуноанализ (обозначаемый далее как «РИА»), иммуноферментный твердофазный анализ (обозначаемый далее как «ELISA»), флуоресцентный анализ антител и пассивный тест на гемагглютинацию, при этом методы РИА и ELISA являются предпочтительными в связи с их чувствительностью по обнаружению, быстроте, точности и возможности выполнять в автоматизированном режиме.
Определение титра антител может быть осуществлено, например, в рамках ELISA указанным ниже способом. Вначале очищенный или частично очищенный DR5 или DR4 сорбируют на поверхности твердой фазы, такой как 96-луночный планшет для проведения ELISA, с последующим блокированием оставшейся незаполненной поверхности, с которой DR5 и DR5 не должны связываться, белком, отличным от антигена, таким как бычий сывороточный альбумин (БСА). После промывки поверхности лунок приводят в контакт с серией разбавленных образцов мышиной сыворотки для создания условий связывания DR5 или DR4 антитела в образцах с антигеном. Добавляют меченое антимышиное антитело, такое как вторичное антитело, для связывания с мышиным антителом. Метка может включать ферментативную метку, флуоресцентную метку и другие известные в технике метки. После промывки добавляют ферментный субстрат и оценивают титр антитела при определении изменения поглощения по развитию окраски, вызванной изменением субстрата и т.п.
(с) Получение миеломных клеток
Клетки из установленных клеточных линий мыши служат в качестве источника миеломных клеток, включающего, например, мышиную линию, устойчивую к 8-азагуанину, полученную из BALB/с миеломных штаммов P3X63Ag8U.1 (P3-U1) (35), P3/NSI/1-Ag4-1(NS-1) (36), Sp2/0-Ag14 (SP-2) (37), P3X63Ag8.653 (653) (38) и P3X63Ag8 (X63) [39]. Отобранную клеточную линию подвергают серийной трансфекции в соответствующую среду, такую как 8-азагуаниновая среда. 8-азагуаниновая среда включает Дульбекко модифицированную среду Искове (Iscove's Modified Dulbecco's medium) (обозначаемую далее как «IMDM») или Дульбекко модифицированную среду Игла (обозначаемую далее как «DMEM»), среду RPMI-1640 с добавкой глютамина, 2-меркаптоэтанола, гентамицина, фетальной сыворотки теленка (обозначаемой далее как «ФСТ») и 8-азагуанина. Затем за 3-4 дня до проведения слияния клетки переносят в нормальную среду, такую как ASF104 (Ajinomoto, K. K.), содержащую 10% ФСТ, с тем чтобы обеспечить наличие по меньшей мере 2·107 клеток ко дню проведения слияния.
(d) Слияние клеток
Лимфоциты и клетки плазмы, полученные из любой подходящей части животного, представляют собой клетки-предшественники для получения антитела. Источники лимфоцитов или плазматических клеток включают в качестве примера селезенку, лимфатические узлы, периферическую кровь или их любое приемлемое сочетание, при этом клетки селезенки являются наиболее частым источником.
После проведения последней бустерной инъекции ткань, в которой присутствуют антителообразующие клетки, отбирают из мыши, имеющей заданный титр антител. Наиболее часто используемая методика для проведения слияния клеток селезенки с миеломными клетками, полученными на стадии с), включает применение полиэтиленгликоля.
Методика слияния включает промывку клеток селезенки и миеломных клеток бессывороточной средой (такой как RPMI 1640) или фосфатно-буферным раствором (обозначаемым далее как ФБР), так чтобы отношение количества клеток селезенки к клеткам миеломы составляло примерно от 5:1 до 10:1, и затем проводят центрифугирование. После отбрасывания супернатанта осадок клеток разбавляют 1 мл бессывороточной среды, содержащей 50% (вес/объем) полиэтиленгликоля (М.в. 1000-4000) путем добавления его по каплям при перемешивании. Затем медленно добавляют 10 мл бессывороточной среды и центрифугируют. Супернатант снова отбрасывает и остаток клеток суспендируют в соответствующем количестве НАТ среды, содержащей раствор гипоксантина, аминоптерина и тимидина (обозначаемый далее как «HAT») и мышиный интерлейкин-2 (обозначаемый далее как «IL-2»). Полученную суспензию затем распределяют по лункам микротитрационного планшета для культивирования тканей (далее называемого просто как «планшет») и инкубируют в атмосфере 5 об.% CO2 при 37°C в течение примерно 2 недель с добавкой HAT среды по потребности.
(е) Отбор гибридом
В том случае когда используют миеломный штамм, резистентный к 8-азагуанину, то есть который является дефицитным по гипоксантингуанинфосфорибозилтрансферазе (ГГФРТ), любые неслившиеся миеломные клетки и результат любого слияния по типу миелома-миелома не способны выживать в HAT среде. С другой стороны, клетки, полученные при слиянии клеток, продуцирующих антитело, друг с другом, а также гибридом антителопродуцирующих клеток с миеломными клетками могут выживать, при этом первый вариант отличается ограниченной продолжительностью жизни. Соответственно длительное инкубирование в НАТ среде приводит к отбору только желательных гибридом.
Полученные гибридомы выращивают до образования колоний, которые затем переносят в НАТ среду, не содержащую аминопетрин (НТ среда). После этого собирают аликвоты культурального супернатанта для определения титра антител против Fas, например, по методу ELISA. Когда в качестве антигена в методике ELISA используют указанный выше слитый белок, необходимо также устранять клоны, продуцирующие антитело, которые специфически связываются с Fc-участком человеческого IgG1. Наличие или отсутствие такого клона может быть подтверждено, например, методом ELISA с использованием в качестве антигена Fas-IgG1 или IgG1.
(f) Клонирование
Гибридомы, которые, как было показано, продуцируют специфические антитела, при использовании для определения титра антител методики, аналогичной таковой, описанной для стадии (b), затем переносят на другой планшет для клонирования. Подходящие методы клонирования включают: метод ограниченного разбавления, при котором гибридомы разбавляют до содержания одной клетки на лунку планшета и затем культивируют; метод культивирования на мягком агаре, при котором колонии получают после культивирования в мягкой агаровой среде; метод с использованием микроманипулятора для отбора одной клетки с целью последующего культивирования; и метод «сортировки клона», при котором с помощью клеточного сортировщика отбирают единичные клетки.
Методику клонирования с использованием, например, способа ограниченного разбавления повторяют 2-4 раза для каждой лунки с определением титра антител и клоны, имеющие стабильные титры антител, выбирают в качестве гибридом, продуцирующих моноклональное антитело против DR5. Гибридомы, продуцирующие антимышиное антитело к DR5, выбирают по аналогичной методике с получением клеточной линии, продуцирующей моноклональное антитело против DR5.
Мышиная-мышиная гибридома TRA-8, которая является основой для получения антител по настоящему изобретению, была депонирована в Американской Коллекции типовых культур (American Type Culture Collection) 1 марта 2000 года с номером доступа PTA-1428. Гибридома 2Е12 была депонирована в Американской Коллекции типовых культур (American Type Culture Collection) 24 октября 2001 года, как было указано выше, с номером доступа ATCC No. PTA-3798. Соответственно при получении антитела с использованием мышиной-мышиной гибридомы TRA-8 или любой другой установленной гибридомы указанное получение может быть осуществлено по методике, начиная со стадии (g), приведенной ниже, а стадии (a)-(f) могут быть пропущены.
(g) Культивирование гибридомы для получения моноклонального антитела
Гибридому, полученную путем клонирования, культивируют далее в нормальной среде, т.е. не в HT-среде. Крупномасштабную культуру получают при культивировании ее в роллерных бутылях с использованием больших культуральных бутылей или путем культивирования на качалке. Затем супернатант от культуры, полученной при крупномасштабном культивировании, отбирают и очищают соответствующим методом, таким как гель-фильтрация, который хорошо известен специалистам в данной области, с получением моноклонального антитела DR5 или DR4, которое представляют собой основной источник антител по настоящему изобретению. Гибридома также может быть выращена внутрибрюшинно в сингенной мыши, такой как BALB/с мышь или nu/nu мышь, с получением асцитов, содержащих в больших количествах моноклональное антитело DR5 или DR4. Для очистки собранных антител удобно использовать коммерчески доступные наборы для очистки моноклональных антител (например, MAbTrap GII Kit; Pharmacia).
Моноклональное антитело, полученное по указанному выше способу, характеризуется высокой специфичностью в отношении человеческого DR5 или DR4 соответственно.
(h) Анализ моноклонального антитела
Соответствующие методики идентификации изотипа и подкласса моноклонального антитела включают метод Оучтерлони (Ouchterlony method), ELISA и РИА. Предпочтительно используют коммерческий набор для идентификации, такой как набор Mouse Typer Kit (торговое наименование, BioRad).
Количественное определение белка может быть проведено по методу Фолина-Лоури или путем расчета на основании значения поглощения при 280 нм (1,4 (ОП280) = 1 мг/мл иммуноглобулина).
Идентификация эпитопа, который распознает моноклональное антитело, осуществляется следующим образом. Вначале определяют структуры разных частей молекулы, которые распознает моноклональное антитело. Частичные структуры определяют в соответствии с методом, в рамках которого путем синтеза получают различные пептидные части молекулы по известным методикам синтеза олигопептидов или по методу, при котором ДНК, кодирующую желательный частичный полипептид, включают в соответствующую плазмиду экспрессии и экспрессируют в подходящей клетке-хозяине, такой как E. coli, с получением пептидов. В основном часто используют обе методики в сочетании с указанным выше объектом. Так, например, серия полипептидов, имеющих соответствующим образом сниженную длину, работающих на С- или N-конце антигенного белка, может быть получена с помощью отработанных методик генно-инженерной технологии. Путем установления того, какие фрагменты реагируют с антителом, получают примерные данные о сайте эпитопа.
Более точно эпитоп идентифицируют путем синтеза множества более мелких соответствующих ему олигопептидов или путем получения мутантов пептида с использованием отработанных методик синтеза олигопептидов для определения способности пептида к связыванию с моноклональным антителом против DR5, например, что является основой для получения антитела по настоящему изобретению и для достижения конкурентного ингибирования связывания пептида с антигеном моноклональным антителом. Могут использоваться коммерчески доступные наборы, такие как SPOTs Kit (Genosys Biotechnologies, Inc.) и серия мультипиновых наборов для пептидного синтеза на основе мультипинового метода синтеза (Chiron Corp.) с получением множества олигопептидов.
Антитело по настоящему изобретению обладает разнообразными функциональными свойствами от а) до f), описанными ниже, каждое из которых подтверждено, например, с помощью описанной ниже методики.
а) Специфическое связывание TRA-8 с клетками, экспрессирующими человеческий DR5
Уникальная особенность настоящего изобретения заключается в способности связывать DR5 клеточной поверхности. Указанная способность может быть продемонстрирована с помощью метода проточной цитометрии клеток, экспрессирующих DR5. Вначале специфическое связывание DR5 на клеточной поверхности подтверждается на COS-7 клетках, трансфицированных с использованием кДНК полной длины, кодирующей человеческий DR5. Конкретно TRA-8 распознает только Cos-7 клетки, трансфицированные DR5, но не пустой контрольный вектор или вектор, кодирующий DR4. Во-вторых, тестируются клетки из трех разных источников происхождения: гемопоэтического, глиомного и из раковых клеток предстательной железы человека. Большая часть из указанных трансформированных опухолевых клеток экспрессирует значительные количества DR5 на клеточной поверхности, хотя уровень экспрессии может широко варьировать. В-третьих, исследуются две панели человеческих первичных синовиальных фибробластных клеток от пациентов с РА и ОА. Все синовиальные клетки при РА экспрессируют значительно больший уровень DR5 в сравнении с ОА клетками.
b) Индукция апоптоза в человеческих клетках со злокачественной опухолью in vitro в отсутствие сшивания
Усиление способности антитела по настоящему изобретению распознавать TRAIL-рецептор и непосредственно индуцировать апоптоз клеток злокачественной опухоли человека определяется по методу жизнеспособности клеток (ATPLite) при культивировании клеток in vitro c различными концентрациями антитела, конкретно TRA-8. Большинство опухолевых клеток чувствительны к TRA-8-индуцированному апоптозу. В случае некоторых клеток TRA-8 демонстрирует мощную активность по индукции апоптоза, например, TRA-8 способен индуцировать апоптоз человеческих клеток Jurkat, на уровне пг/мл. Важно, что TRA-8 индуцирует апоптоз без потребности в сшивании, и в большинстве клеток TRA-8 демонстрирует более сильную активность по индукции апоптоза, чем рекомбинантный растворимый TRAIL в присутствии энхансера.
с) Активность TRA-8 по уничтожению опухолевых клеток in vivo
Активность TRA-8 по уничтожению опухолевых клеток оценивают на двух моделях опухоли SCID/человека. Вначале мышам SCID внутривенно инокулируют клетки Jurkat лейкоза человека и вводят однократно дозу TRA-8 (100 мкг). Результаты показывают, что большая часть имплантированных клеток Jurkat элиминируется из периферической крови и селезенки при вводе TRA-8, о чем свидетельствуют результаты анализа методом проточной цитометрии и иммунохимического окрашивания клеток Jurkat in situ. Во-вторых, клетки астроцитомы человека 1321N1 подкожно инокулируют мышам SCID и затем мышам, получившим опухоль, вводят однократную дозу TRA-8. Рост имплантированных 1321N1 клеток значительно подавляется у мышей после введения TRA-8, что видно по размерам опухоли и по данным гистологического анализа.
d) Идентификация синовиальных клеток при РА с помощью TRA-8
Первичные синовиальные клетки выделяют у 8 пациентов с РА и 4 пациентов с ОА и тестируют на наличие экспрессии DR5 на клеточной поверхности. TRA-8 способен положительно окрашивать все РА клетки, а все ОА клетки окрашивать отрицательно. Таким образом, РА может быть дифференцирован от ОА посредством выявления с помощью TRA-8 экспрессии DR5 на клеточной поверхности.
е) Индукция апоптоза синовиальных фибробластных клеток при РА под действием TRA-8
Способность TRA-8 индуцировать апоптоз синовиальных клеток при РА определяют в тесте на жизнеспособность клеток в культуре in vitro в присутствии различных концентраций TRA-8. Все РА клетки демонстрируют чувствительность к 100 нг/мл TRA-8 в степени от высокой до умеренной. И наоборот, все ОА клетки по существу устойчивы к апоптозу, индуцированному TRA-8. Важно, что TRA-8 демонстрирует лучшую апоптоз-индуцирующую способность в синовиальных клетках при РА, чем растворимый TRAIL с энхансером. Более того, в сравнении с антителом против Fas (CH-11), TRA-8 демонстрирует лучшую селективность к синовиальным клеткам при РА.
f) TRA-8 не индуцирует образование ММП в синовиальных клетках при РА
Поскольку TRA-8 способен индуцировать активацию NF-κb в синовиальных клетках при РА, таких как TNF-a, определяют эффект TRA-8 на продукцию ММП-1 и ММП-3 в синовиальных клетках. Поскольку TNF-a индуцирует зависимое от дозы повышение уровня ММП, TRA-8 не способен индуцировать образование в каких-либо количествах ММП, а в некоторых концентрациях TRA-8 незначительно снижает образование ММП в синовиальных клетках при РА.
g) TRA-8 индуцирует множественную активацию каспазы
Поскольку каспазы играют решающую роль в индукции апопотоза, определяют способность TRA-8 индуцировать активацию каспазы в человеческих клетках Jurkat. Когда клетки Jurkat инкубируют с низкой дозой (50 нг/мл) TRA-8, активация каспазы 8, каспазы 9 и каспазы 3 наблюдается уже к 15 минутам после начала активации, что видно по данным вестерн-блоттинга и по анализу расщепления каспазой. С точки зрения времени, количества и интенсивности активации каспазы, антитела по настоящему изобретению, включающие репрезентативное антитело TRA-8, демонстрируют гораздо большую активность, чем известные апоптоз-индуцирующие антитела, такие как антитела против человеческого Fas (CH-11).
Антитело 2Е12 специфически связывается с DR4 в его растворимой форме, обладает in vivo и in vitro апоптоз-индуцирующей активностью в отношении целевых клеток, экспрессирующих DR4 (включая, например, злокачественные клетки, синовиальные клетки при ревматоидном артрите, активированные иммунные клетки наподобие активированных лимфоцитов и вирус-инфицированные клетки), обладает активностью по уничтожению опухолевых клеток in vivo (предпочтительно при отсутствии токсичности к неопухолевым клеткам). Предпочтительно DR4 антитело по настоящему изобретению обладает апоптоз-индуцирующей активностью, характеризующейся достижением менее чем примерно 60%, 50%, 40%, 30%, 20% или 10% жизнеспособности целевых клеток при концентрации антитела менее чем 30 мкг/мл, 3 мкг/мл, 0,3 мкг/мл или 0,03 мкг/мл и активностью по уничтожению опухолевых клеток, характеризующейся снижением размера опухоли на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100%. Таким образом, антитело по настоящему изобретению представляет собой вещество, обладающее способностью селективно индуцировать апоптоз в патогенных клетках, как было указано при описании его эффекта в пунктах (a) и (g). Соответственно оно полезно как профилактическое и лечебное средство в случае заболеваний, связанных с несоответствующим выживанием клеток или с несоответствующей пролиферацией клеток, таких как заболевания, определяемые дисбалансом в системах апоптоза, включая систему Fas/Fas-лигандов.
Способность антитела по настоящему изобретению индуцировать апоптоз подтверждается при культивировании клеток, таких как клетки клеточной линии лейкоза человека Jurkat (Американская Коллекция типовых культур No. TIB-152) и клетки клеточной линии астроцитомы 1321N1 в среде, к которой добавляют исследуемый образец, с последующим определением уровня выживания, например в тесте ATPLite.
Антитело по настоящему изобретению, особенно антитела DR5 и DR4, обладающие такой же иммуногенностью для человека, что и человеческие антитела, используют в качестве средства профилактики или лечения заболеваний, связанных с несоответствующим выживанием или несоответствующей пролиферацией клеток, включая те заболевания, которые связаны с дисбалансом в системах апоптоза при воспалительных и аутоиммунных заболеваниях, включающих в качестве примера системную красную волчанку, болезнь Хашимото, ревматоидный артрит, реакцию отторжения трансплантата, синдром Шегрена, злокачественную анемию, болезнь Аддисона, склеродерму, синдром Гудпастура, болезнь Крона, аутоиммунную гемолитическую анемию, стерильность, тяжелую псевдопаралитическую миастению, рассеянный склероз, базедову болезнь, тромбоцитопеническую пурпуру, инсулинзависимый сахарный диабет, аллергию, астму, атопическую болезнь, атеросклероз; миокардит; кардиомиопатию, гломерулонефрит; гипопластическую анемию, реакцию отторжения после пересадки органов и различные виды злокачественных новообразований легкого, предстательной железы, печени, яичника, толстого кишечника, шейки матки, лимфатических тканей и тканей молочной железы. Антитела по настоящему изобретению могут использоваться для нацеливания и селективной индукции апоптоза в активированных иммунных клетках, включающих активированные лимфоциты, лимфоидные клетки, миелоидные клетки и синовиальные клетки при ревматоидном артрите (включая воспалительные синовиоциты, макрофагоподобные синовиоциты, фибробластоподобные синовиоциты) и инфицированные вирусом клетки (включая, например, клетки, инфицированные ВИЧ) в той мере, в которой целевые клетки экспрессируют или могут быть начать экспрессировать специфические TRAIL-рецепторы (например, DR4 и DR5).
Такое профилактическое или лекарственное средство может вводиться в различных формах. Подходящие способы введения включают пероральное введение, такое как в виде таблеток, капсул, гранул, порошков и сиропов, или парентеральное введение, такое как путем инъекции, или с использованием суппозитория.
Антитело или лекарственное средство может вводиться перорально, ректально, внутригрудинно, внутрижелудочковым способом, внутричерепным способом, интратекально, внутрисуставным способом, внутривагинально, парентерально (внутривенно, внутримышечно или подкожно), местным способом (в виде порошков, мазей или капель) или внутрибрюшинной инъекцией, чрескожно, путем ингаляции, трансбуккально или с помощью назального спрея. Точное количество требуемого антитела или лекарственного средства варьирует от субъекта к субъекту в зависимости от возраста, веса и общего состояния здоровья субъекта, тяжести заболевания, которое предстоит лечить, от локализации и размера опухоли, конкретной природы используемых соединений, способа введения и т.п. Соответствующее количество может быть определено специалистом со средним уровнем знаний в данной области с использованием обычных экспериментов, приведенных в настоящем описании. Типичные однократные дозировки антитела варьируют в диапазоне от 0,1 до 10000 микрограмм, предпочтительно от 1 до 100 микрограмм. Типичные концентрации антитела в носителе варьируют в диапазоне от 0,2 до 2000 нанограмм на доставляемый миллилитр. Для инъекции в сустав объемы антитела и носителя будут варьировать в зависимости от сустава, но примерно будут составлять 0,5-10 мл и предпочтительно 1-5 мл, в случае инъекции в колено человека, и примерно от 0,1 до 5 мл и предпочтительно 1-2 мл, в случае инъекции в голеностопный сустав.
В зависимости от целевого способа введения антитело или лекарственное средство может входить в состав фармацевтических композиций, имеющих вид твердых, полутвердых или жидких дозированных форм, таких как таблетки, суппозитории, пилюли, капсулы, порошки, жидкости или суспензии, предпочтительно в виде стандартных дозированных форм, приемлемых для однократного введения точной дозировки. Композиция включает эффективное количество выбранного субстрата в сочетании с фармацевтически приемлемым носителем и вдобавок может включать другие медицинские средства, фармацевтические средства, носители или разбавители. Термин «фармацевтически приемлемый» означает материал, который не является биологически или в любом другом контексте нежелательным, который может вводиться индивидууму вместе с выбранным субстратом, не вызывая значительных нежелательных биологических эффектов или взаимодействия в неблагоприятном режиме с любыми другими компонентами фармацевтической композиции, в которой он содержится.
Композиции, пригодные для парентерального введения, могут включать физиологически приемлемые стерильные водные или неводные растворы, дисперсии, суспензии или эмульсии, а также стерильные порошки для разведения с получением стерильных инъецируемых растворов или дисперсий. Примеры подходящий водных и неводных носителей, разбавителей, растворов или наполнителей включают воду, этанол, полиолы (пропиленгликоль, полиэтиленгликоль, глицерин и т.п.), их подходящие смеси, растительные масла (такие как оливковое масло) и инъецируемые органические сложные эфиры, такие как этилолеат. Соответствующая текучесть может поддерживаться, например, за счет использования покрытия, такого как лецитин, посредством поддержания нужного размера частиц в случае дисперсии или за счет использования поверхностно-активных веществ.
Указанные композиции могут также содержать вспомогательные вещества, такие как консервант, смачивающее вещество, эмульгатор и средство, способствующее диспергированию. Предотвращение действия микроорганизмов может быть обеспечено различными противобактериальными и противогрибковыми средствами, например парабенами, хлорбутанолом, фенолом, сорбиновой кислотой и т.п. Могут также при желании включаться средства, способствующие изотоничности, например сахара, хлорид натрия и т.п. Пролонгируемого всасывания инъецируемой фармацевтической формы можно добиться путем использования средств, задерживающих всасывание, например, моностеарата алюминия и желатина.
Твердые дозированные формы для перорального введения включают капсулы, таблетки, пилюли, порошки и гранулы. В составе таких твердых дозированных форм активное соединение смешивают по меньшей мере с одним обычным инертным наполнителем (или носителем), таким как цитрат натрия или фосфат дикальция, или с (а) наполнителями или средствами, увеличивающими объем, такими как, например, крахмалы, лактоза, сахароза, глюкоза, маннит или кремневая кислота, (b) связующим веществом, например, карбоксиметилцеллюлозой, альгинатами, желатином, поливинилпирролидоном, сахарозой и аравийской камедью, (с) увлажнителями, например, глицерином, (d) средствами, способствующими разложению, такими как, например, агар-агар, карбонат кальция, картофельный крахмал или крахмал тапиоки, альгиновая кислота, некоторые комплексные соединения силикатов и карбонат натрия, (е) средствами, замедляющими растворение, такими как, например, парафин, (f) ускорителями всасывания, такими как, например, соединения четвертичного аммония, (g) смачивающими веществами, такими как, например, цетиловый спирт и моностеарат глицерина, (h) адсорбентами, такими как, например, каолин и бентонит, и (i) замасливателями, такими как, например, тальк, стеарат кальция, стеарат магния, твердые полиэтиленгликоли, лаурилсульфат натрия или их смеси. В случае капсул, таблеток и пилюль дозированные формы могут также включать забуферивающие вещества.
Твердые композиции аналогичного типа могут также использоваться в составе наполнителей в мягких и твердых желатиновых капсулах с использованием таких эксципиентов, как лактоза или молочный сахар, а также высокомолекулярные полиэтиленгликоли и т.п.
Твердые дозированные формы, такие как таблетки, драже, капсулы, пилюли и гранулы, могут быть приготовлены с использованием покрытий и оболочек, таких как энтеросолюбильные покрытия и другие известные в технике виды покрывающих оболочек. Они могут содержать вещества, делающие материал непрозрачным, и могут также иметь такую композицию, которая позволяет высвобождать активное соединение или соединения в определенной части желудочно-кишечного тракта задержанным способом. Примеры таких приемлемых для использования погруженных композиций включают полимерные вещества и воски. Активные соединения могут также представлять собой микроинкапсулированную форму, которая в случае приемлемости включает один или более указанных выше эксципиентов.
Жидкие дозированные формы для перорального введения включают фармацевтически приемлемые эмульсии, растворы, суспензии, сиропы и эликсиры. Кроме активных соединений жидкие дозированные формы могут содержать инертные разбавители, обычно используемые в данной области техники, такие как вода или другие растворители, солюбилизирующие средства и эмульгаторы, например этиловый спирт, изопропиловый спирт, этилкарбонат, этилацетат, бензиловый спирт, бензилбензоат, пропиленгликоль, 1,3-бутиленгликоль, диметилформамид, масла, в частности хлопковое масло, арахисовое масло, кукурузное масло, оливковое масло, касторовое масло и кунжутное масло, глицерин, тетрагидрофурфуриловый спирт, полиэтиленгликоли и сложные эфиры жирных кислот и сорбитана или смеси указанных веществ и т.п.
Кроме указанный инертных разбавителей композиции могут также включать вспомогательные средства, такие как смачивающие вещества, эмульгаторы, средства, способствующие суспендированию, подсластители, вкусовые вещества и отдушки.
Суспензии вдобавок к активным соединениям могут содержать средства, способствующие суспендированию, такие как, например, этоксилированые изостеариловые спирты, сложные эфиры полиоксиэтиленсорбита и сорбитана, микрокристаллическая целлюлоза, метагидроксид алюминия, бентонит, агар-агар и трагакант или смеси указанных веществ и т.п.
Композиции для ректального введения представляют собой предпочтительно суппозитории, которые могут быть приготовлены при смешивании соединений по настоящему изобретению с соответствующими нераздражающими эксципиентами или носителями, такими как, например, масло какао, полиэтиленгликоль или воск для свечей, которые при обычных температурах представляют собой твердые вещества, но становятся жидкими при температуре тела, в связи с чем они размягчаются в прямой кишке или в вагинальной полости и высвобождают активный компонент.
Дозированные формы для местного введения соединения по настоящему изобретению включают мази, порошки, аэрозоли и ингаляторы. Активные компоненты смешивают в стерильных условиях с физиологически приемлемым носителем и любыми консервантами, буферами или пропеллентами в зависимости от потребностей. Глазные композиции мази, порошки и растворы также входят в область настоящего изобретения.
Термин «фармацевтически приемлемые соли, сложные эфиры, амиды и пролекарства» в контексте настоящего изобретения относятся к карбоксилатным солям, солям добавления аминокислот, сложным эфирам, амидам и пролекарствам соединений по настоящему изобретению, которые по результатам медицинского заключения рассматриваются как приемлемые для использования в контакте с тканями пациента, не вызывая чрезмерной токсичности, раздражения, аллергической реакции и т.п., и которые характеризуются разумным соотношением польза-риск, являются эффективными для целевого применения, а также к цвиттерионным формам, где это приемлемо, соединений по настоящему изобретению. Термин «соли» применяется относительно нетоксичных солей добавления неорганических и органических кислот соединений по настоящему изобретению. Указанные соли могут быть приготовлены in situ в ходе выделения и очистки соединений или путем проведения отдельной реакции очищенного соединения в его свободной основной форме с соответствующей органической или неорганической кислотой при последующем выделении полученной таким образом соли. Репрезентативные соли включают гидробромидные, гидрохлоридные, сульфатные, бисульфатные, нитратные, ацетатные, оксалатные, валератные, олеатные, пальмитатные, стеаратные, лауратные, боратные, бензоатные, лактатные, фосфатные, тозилатные, цитратные, малеатные, фумаратные, сукцинатные, тартратные, нафтилатмезилатные, глюкогептонатные, лактобионатные, метансульфонатные и лаурилсульфонатные соли и т.п. Они могут включать катионы щелочных и щелочноземельных металлов, таких как натрий, литий, калий, кальций, магний и т.п., а также нетоксичные аммониевые, четвертичные аммониевые и аминные катионы, которые включают, не ограничиваясь, аммоний, тетраметиламмоний, тетраэтиламмоний, метиламин, диметиламин, триметиламин, триэтиламин, этиламин и т.п. (См. например, работу S.M. Barge et al., «Pharmaceutical Salts», J. Pharm. Sci., 1977, 66:1-19, которая включена в настоящее описание в качестве ссылки).
Термин «пролекарство» относится к соединениям, которые быстро подвергаются трансформации in vivo с образованием исходных соединений указанных выше формул, например, путем гидролиза в крови.
Обширное обсуждение пролекарств приведено в работе Хигучи и Стелла (T. Higuchi and V. Stella, «Pro-drugs as Novel Delivery Systems», Vol. 14 of the A.C.S. Symposium Series, and in Bioreversible Carriers in Drug Design, ed. Edward B. Roche, American Pharmaceutical Association and Pergamon Press, 1987).
Целевая клетки представляет собой клетку животного, примеры которой включают клетку человека, примата, отличного от человека, кошек, собак, крысы, мыши, морской свинки, кролика, козы, овцы, коровы, лошади, курицы, свиньи, мартышки и африканского хорька.
Дополнительно антитело и терапевтическое средство по настоящему изобретению могут иметь вид как несольватированных, так и сольватированных форм с фармацевтически приемлемыми растворителями, такими как вода, этанол и т.п. В целом, для целей настоящего изобретения сольватированные формы рассматриваются как эквивалентные несольватированным формам.
Молекулы антитела очищают известными методиками, примеры которых включают аминоабсорбционную или аминоаффинную хроматографию, хроматографические методики, такие как высокоэффективная жидкостная хроматография, или их сочетания.
В другом аспекте настоящее изобретение включает фармацевтический продукт для использования при доставке биологически активного антитела против TRAIL-рецептора или гуманизированного антитела против TRAIL-рецептора у позвоночных животных. Фармацевтический продукт включает фармацевтически эффективное количество антитела против TRAIL-рецептора или его фрагмент, фармацевтически приемлемый носитель и контейнер, включающий носитель и антитело в стерильном виде.
В предпочтительном варианте осуществления настоящего изобретения фармацевтически эффективное количество антитела по настоящему изобретению подавляет пролиферацию клеток или индуцирует апоптоз при контакте с целевой клеткой или с целевыми клетками. Фармацевтически эффективное количество или количество антитела, распознающего DR5 или DR4, или гуманизированного антитела, распознающего DR5 или DR4, представляет собой вводимое индивидууму количество, которого достаточно для того, чтобы вызвать желательный эффект. В контексте настоящего описания термины «фармацевтически эффективное количество» и «терапевтическое количество» являются синонимами. Желательные эффекты введения фармацевтически эффективного количества антител, распознающих DR5 или DR4, включают гибель одной или более целевых клеток, подавление роста одной или более целевых клеток, стимуляцию DR5 или DR4 соответственно, связывание с DR5 или DR4 соответственно, и повышение уровня NFkB или его активности в целевой клетке. Целевая клетка представляет собой клетку, которая экспрессирует DR5 или DR4 и которая в качестве примера включает аномально растущие клетки и опухолевые клетки, такие как папилломы и бородавки; рак молочной железы, рак толстого кишечника, гепатомы, лейкозы, рак легкого, меланому, миеломы, остеосаркомы, рак яичника, рак поджелудочной железы, рак предстательной железы, рак головы и шеи, рак щитовидной железы, рак матки, опухоли мозга, такие как астроцитомы, а также активированные иммунные клетки (например, активированные лимфоциты, лимфоидные и миелоидные клетки), воспалительные клетки, синовиальные клетки при ревматоидном артрите и клетки, инфицированные вирусом. Целевая клетка in vivo представляет собой клетку индивидуума в патологическом состоянии, включая такие состояния, при которых клеточная пролиферация является аномальной, или разрегулированной, такие как злокачественные или доброкачественные опухоли и ревматоидный артрит.
В другом предпочтительном варианте осуществления изобретения целевая клетка приводится в контакт с терапевтическим средством.
Терапевтическое средство представляет собой соединение или композицию, эффективные в плане ослабления патологического состояния. Иллюстративный пример терапевтического средства включает противораковое соединение.
Противораковое соединение представляет собой соединение или композицию, эффективные по подавлению или остановке роста аномально растущей клетки. Фармацевтически эффективное количество противоракового соединения представляет собой вводимое индивидууму количество, которого достаточно для того, чтобы вызвать подавление или остановку роста аномально растущей клетки. Иллюстративные примеры противораковых соединений включают: блеомицин, карбоплатину, хлорамбуцил, цисплатину, колхицин, циклофосфамид, даунорубицин, дактиномицин, диэтилстилбэстрол-доксорубицин, этопозид, 5-фторурацил, флоксуридин, мелфалан, метотрексат, митомицин, 6-меркаптопурин, тенипозид, 6-тиогуанин, винкристин и винбластин. Другие примеры противораковых соединений и соответствующих терапевтических средств приведены в руководстве Мерка (The Merck Manual of Diagnosis and Therapy, 15th El., Berkow et al., eds., 1987, Rahway, N.J. and Sladek et al. Metabolism and Action of Anti-Cancer Drugs, 1987, Powis et al., eds., Taylor and Francis, New York, N.Y.).
Антитело по настоящему изобретению может быть также объединено с другими терапевтическим стратегиями, такими как химиотерапия и/или радиотерапия, при лечении злокачественных опухолей, и терапевтическая эффективность может быть повышена путем ввода соединений, индуцирующих апоптоз, таких как бисиндолмалеимид VIII (BisVIII), или других сенсибилизирующих средств, таких как SN-50 или LY294002. Таким образом, в одном варианте антитело или антитела по настоящему изобретению могут быть объединены с BisVIII и химиотерапевтическим средством (например, адриамицином). В другом варианте антитело или антитела по настоящему изобретению могут быть объединены с нестероидным противовоспалительным средством (например, сульфидом сулиндака или другими ингибиторами COX-1 или COX-2) и химиотерапевтическим средством (например, адриамицином). При лечении других заболеваний, например аутоиммунных и воспалительных заболеваний, могут также использоваться сочетания различных стратегий. Так, например, антитело может быть введено в сочетании с другими терапевтическими средствами, такими как химиотерапевтические средства, противовоспалительные средства, DMARD, метотрексат, бисиндолмалеимид VIII, или другими сенсибилизирующими средствами и т.п. Радиотерапия может быть также объединена с другими терапевтическими средствами при лечении воспалительных и аутоиммунных заболеваний. Специалист в данной области может адаптировать определенный вид радиотерапии к конкретному воспалительному или аутоиммунному заболеванию (например, лучевую синовэктомию при лечении артрита).
Метод лечения, при котором используется антитело по настоящему изобретению, может быть также объединен с методом лечением, при котором используется другое антитело по настоящему изобретению. Так, например, антитело против DR5 может быть введено субъекту, при необходимости, вместе, до или после введения антитела против DR4. Такая комбинированная терапия антителом может быть далее объединена с введением одного или более лекарственных средств (например, с введением химиотерапевтических средств, доксорубицина и/или метотрексата), с радиотерапией или с обоими указанными методами лечения.
В сравнении с исследованным ранее антителом против DR5 [24], апоптоз-индуцирующая активность иллюстративного антитела TRA-8 по настоящему изобретению высока, указанное антитело способно индуцировать апоптоз клеток Jurkat in vitro на уровнях пг/мл и характеризуется более высокой активностью по уничтожению опухолевых клеток in vivo в сравнении с ранее известным растворимым TRAIL. Внутривенного введения однократной дозы TRA-8 достаточно для подавления роста клеток твердых опухолей и клеток гемопоэтических опухолей, тогда как для индукции in vivo регрессии опухоли с использованием растворимого TRAIL требуется более высокая доза (500 мкг каждый день в течение 10 дней). Антитела против TRAIL-рецептора по настоящему изобретению, по всей видимости, более безопасны, чем растворимый TRAIL, поскольку иллюстративное антитело TRA-8 не индуцирует апоптоз нетрансформированных фибробластных клеток. Аналогичные результаты были получены с антителами против DR4.
Векторы по настоящему изобретению включают последовательность нуклеиновой кислоты, кодирующую тяжелую или легкую иммуноглобулиновую цепь антитела DR5 или DR4, функционирующего в оперативно связанном состоянии с регулирующим элементом, таким как промотор или энхансер. Термин «оперативно связанный» относится к конфигурации нуклеотидных последовательностей, организованных так, что указанная конфигурация позволяет выполнять обычные функции. Так, регуляторный элемент, оперативно связанный с нуклеотидной последовательностью, кодирующей полипептид, способен направлять транскрипцию, репликацию и/или трансляцию полипептида. Специалистам в данной области понятно, что один вектор необязательно включает кодирующие последовательности для тяжелой и легкой цепи иммуноглобулина DR5 или DR4 антитела. В одном варианте осуществления изобретения векторы кодируют гуманизированную тяжелую или легкую цепь иммуноглобулинов.
Приведенные ниже примеры даны для иллюстрации способов и результатов настоящего изобретения. Их не следует рассматривать как исчерпывающие все аспекты настоящего изобретения, они, скорее, являются иллюстрацией его репрезентативных способов и результатов. Приведенные примеры не должны рассматриваться как исключающие эквиваленты и вариации настоящего изобретения, которые являются очевидными для специалистов в данной области.
Пример 1. Приготовление антигена DR5
1.1 Клонирование кДНК для DR5
ДНК, кодирующую белок DR5, клонируют с помощью указанного ниже метода ОП-ПЦР с использованием следующих компонентов:
а) Матрица
Суммарную РНК из клеток HELa экстрагируют с использованием реагента TRIzol (GIBCO BRL). В качестве матрицы для ПЦР реакции используют кДНК, которую получают с использованием набора для синтеза ДНК First-Strand (American Pharmacia Biotech) в соответствии с инструкцией, прилагаемой к набору.
b) Праймеры для ПЦР
Для ПЦР синтезируют указанные ниже олигонуклеотидные праймеры:
5'-gacgatgcccgatctactttaaggg-3' (DR5p1:SEQ ID No. 1);
5'-ccactgggtgatgttggatggg-3' (DR5p2:SEQ ID No. 2);
Если особо не оговорено иное, все олигонуклеотиды, приведенные в указанных примерах, синтезируют по методике Lifetechnologies. Все олигонуклеотиды хранят при температуре -20°С после растворения в дистиллированной воде.
с) Реакция ПЦР
Раствор для проведения реакции ПЦР имеет следующий состав:
матричная кДНК, 5 мкл в 33 мкл реакционной смеси
праймер DR5р1, 10 пмоль;
праймер DR5р2, 10 пмоль;
10 × концентрированный ПЦР буфер (прилагаемый к набору), 10 мкл;
дНТФ (каждый по 2,5 мМ), 4 мкл; и
Taq-полимераза (Promega), 5 единиц.
К раствору добавляют стерильную дистиллированную воду, доводя до объема 100 мкл.
Если особо не оговорено иное, используемые дНТФ включают эквимолярную смесь дАТФ, дЦТФ, дГТФ и дТТФ (по 2,5 мМ каждый).
ПЦР проводят следующим образом. Вначале раствор нагревают до 94°С в течение 2 минут, после чего проводят цикл, включающий нагревание до 94°С в течение 30 секунд, нагревание до 52°С в течение 1 минуты и нагревание до 72°С в течение 3 минут с повтором процедуры 40 раз. После завершения указанной процедуры реакционный раствор нагревают при 72°С в течение 10 минут.
Полученный таким образом фрагмент амплифицированной ДНК, разделяют в 1% геле агарозы, содержащем 0,25 мкг/мл этидиум бромида. Полосы, в которых показано содержание желательных фрагментов ДНК, вырезают с использованием лезвия и из них высвобождают ДНК с использованием набора Gene Clean (BIO101). Фрагмент ДНК клонируют с использование набора для клонирования TA Cloning (Invitrogen, CA). Клонирование проводят следующим образом.
Фрагмент ДНК, выделенный из раствора для ПЦР, вместе с 50 нг вектора рCR2.1, прилагаемого к набору TA Cloning, смешивают с 1 мкл 10× буфера для лигазной реакции (6 мМ Tris-HCl (pH 7,5), 6 мМ хлорид магния, 5 мМ хлорид натрия, 7 мМ β-меркаптоэтанол, 0,1 мМ АТФ, 2 мМ ДТТ, 1 мМ спермидин и 0,1 мг/мл бычий сывороточный альбумин), к которому добавляют 4 единицы ДНК-лигазы Т4 (1 мкл). Общий объем смеси доводят до 10 мкл стерильной деионизированной водой и полученный лигазный раствор инкубируют при 14°С в течение 15 часов. По истечении указанного периода времени добавляют 2 мкл раствора для проведения лигазной реакции к 50 мкл компетентного штамма Е. coli TOP10F', который прилагается к набору для клонирования TA и который подготавливают к работе в соответствии с руководством для пользователя, далее добавляют 2 мкл 0,5 М β-меркаптоэтанола и полученную смесь выдерживают на льду в течение 30 минут, затем при 42°С в течение 30 секунд и снова на льду в течение 5 минут. Затем к культуре добавляют 500 мкл среды, содержащей 2 об.% триптона, 0,5% (вес./об.) дрожжевого экстракта, 0,05% (вес./об.) хлорида натрия, 2,5 мМ хлорида калия, 1 мМ хлорида магния и 20 мМ глюкозы (далее обозначаемой как среда «SOC»), и смесь инкубируют в течение 1 часа при 37°С при качании. По прошествии указанного периода времени культуру распределяют по чашке с L-бульонным агаром (1 об./об.% триптона, 0,5% (вес./об.) дрожжевого экстракта, 0,05% (вес./об.) хлорида натрия, 0,1% (вес./об.) глюкозы и 0,6% (вес/об.) бактоагара (Difco)), содержащим 100 мкг/мл ампициллина. Устойчивые к ампициллину колонии, появившиеся на чашке, отсортировывают, соскребают с помощью платиновой петли и далее культивируют в L-бульонной среде, содержащей 100 мкг/мл ампициллина, при 37°С в течение ночи при качании со скоростью 200 об/мин. После инкубации клетки собирают центрифугированием, после чего получают плазмидную ДНК щелочным методом. EcoRI-EcoRI фрагмент ДНК DR5c из полученной таким образом плазмиды клонируют в плазмиде pcDNA3 (Invitrogen, CA). Ген DR5 полной длины в pcDNA3 секвенируют и сравнивают с опубликованной последовательностью. Полученную таким образом плазмиду обозначают как плазмида pcDNA3-DR5.
1.2 Конструирование вектора экспрессии DR5-IgG
Для того чтобы получить растворимую форму человеческого DR5, не содержащего трансмембранный домен, конструируют плазмидный вектор экспрессии. Указанный вектор разрабатывают с целью кодирования слитого белка, который должен включать внеклеточный домен человеческого DR5, слитый с ДНК Fc IgG1 человека [41]. ДНК, кодирующую человеческий DR5, которая не содержит трансмембранный домен, получают с помощью приведенной ниже реакции ПЦР.
а) Матрица
В качестве матрицы для данной ПЦР используют pcDNA3-DR5.
b) Праймеры для ПЦР
Синтезируют приведенные ниже олигонуклеотидные праймеры для проведения ПЦР:
5'-gacgatgcccgatctactttaaggg-3' (DR5p1:SEQ ID No. 1);
5'-ggatccgtggacacattcgatgtc-3' (DR5p3:SEQ ID No. 3);
Если особо не оговорено иное, все олигонуклеотиды, приведенные в указанных примерах, синтезируют по методике Lifetechnologies. Все олигонуклеотиды хранят при температуре -20°С после растворения в дистиллированной воде.
с) Реакция ПЦР
Проводят ПЦР и выделяют амплифицированную ДНК по методике, описанной в примере 1.1(с).
Полученную таким образом плазмиду обозначают как плазмида pCR-ΔDR5. BamHI-EcoRI фрагмент, кодируемый человеческим Fc-фрагментом, который восстанавливают из pmFas-hIgG1Fc, субклонируют в множественных сайтах клонирования для BamHI и EcoRI в pcDNA3. Полученную таким образом плазмиду обозначают как pcDNAFc. Далее фрагмент BamHI-BamHI, кодирующий участок растворимого человеческого DR5, который восстанавливают из pCR-ΔDR5, субклонируют его в сайте BamHI плазмиды pcDNAFc. Полученную таким образом плазмиду обозначают как плазмида pcDNAΔDR5-Fc. В EcoRI-фрагменте, кодирующем участок, включающий человеческий растворимый DR5-человеческий IgG Fc, который получают из плазмиды pcDNAΔDR5-Fc, делают липкие концы с использованием фрагмента ДНК-полимеразы Кленова (Klenow, GIBCO BRL) и затем подвергают субклонированию в шаттл-векторе pAdCMV5 (Quantum Biotechnologies Inc., Канада), в котором создают липкие концы после расщепления BamHI. Полученную таким образом плазмиду обозначают как плазмида pAdΔDR5-Fc.
1.3 Экспрессия и очистка слитого белка человеческого DR5-IgG1
Клетки QBI-293A (прилагаемые к набору ADENO-Quest), подвергают совместной трансфекции с использованием pAdΔDR5-Fc и QBI-вирусной ДНК (прилагаемых к набору ADENO-Quest) с использованием набора ADENO-Quest (Quantum Biotechnologies Inc., Канада) в соответствии с инструкцией для пользователя. Культивируют рекомбинантные вирусные бляшки и далее проводят скрининг на экспрессию слитого белка DR5-IgG при анализе супернатанта по методу ELISA. Положительные бляшки амплифицируют в клетках QBI-293A и хранят при температуре -80°С в качестве вирусного раствора для хранения. Пятьдесят чашек (150 мм) с клетками QBI-293A трансфицируют рекомбинантным вирусом pAdΔDR5-Fc с множественностью инфекции 10 (м.и.). Культуральные среды собирают после завершения трансфекции через 48 часов.
Трансфицированные клетки, содержащие ген DR5-IgG, выращивают до плотности клеток 1·106 клетки/мл при инкубировании в 500 мл среды DMEM (GIBCO), содержащей 10 об./об.% ФСТ, при 37°С в атмосфере 5 об./об.% CO2 в течение 2 дней. Затем культуру центрифугируют (1000 об/мин, 5 минут) и собирают супернатант. Очистка DR5-IgG из супернатанта достигается с использованием аффинной хроматографии на белкеА-Сефарозе CL-4B (Pharmacia) в следующих условиях:
колонка: БелокА-Сефароза CL-4В (размер колонки 2 мл, Pharmacia);
элюирующий буфер: 0,1 М глицин (pH 2,4) 0,15 М NaCl;
нейтрализующий буфер: 1 М Трис-HCl (pH 8,5).
После нанесения всего супернатанта на колонку ее промывают три раза 20 мл ФБР и затем добавляют 10 раз 1 мл элюирующего буфера. Измеряют оптическую плотность каждой элюированной фракции (1 мл). Фракции от второй до пятой (со значениями ОП280≥0,1 собирают и после добавления 100 мкл нейтрализующего буфера элюаты помещают по отдельности в пробирки для диализа и диализуют элюаты против 1 литра ФБР (pH 7,5) при 4°С. Буфер для диализа меняют дважды.
Элюаты оценивают на наличие экспрессии продукта гена DR5-IgG по методу ELISA. Вначале 100 мкл каждой фракции помещают по отдельности в лунки 96-луночного микротитрационного планшета (Costar) и инкубируют при 37°C в течение 1 часа. По прошествии указанного времени раствор из лунок удаляют и планшет промывают 3 раза по 100 мкл/лунку ФБР, содержащим 0,1 об./об.%. Твин 20 (далее обозначаемым как «ФБР-Твин»). После промывки добавляют ФБР, содержащий 2% (вес/об.) бычьего сывороточного альбумина (далее обозначаемого как БСА) в количестве 100 мкл/лунку и планшет инкубируют при 37°С в течение 1 часа. По прошествии указанного периода времени лунки промывают еще 3 раза по 100 мкл/лунку ФБР-Твином, после чего к каждой лунке добавляют по 100 мкл/лунку раствора моноклонального антитела против человеческого IgG1, разбавленного в 1000 раз ФБР-Твином, и планшет снова инкубируют при 37°С в течение 1 часа. Дале лунки промывают 3 раза по 100 мкл/лунку ФБР-Твином. Затем добавляют 3,3',5,5'-тетраметилбензидиновую (далее обозначаемый как "ТМБ") жидкую субстратную систему (Sigma) в количестве по 100 мкл/лунку и планшет выдерживают при комнатной температуре в течение 5 минут, после чего реакцию останавливают добавлением 0,2N H2SO4 в количестве 100 мкл/лунку. Величину поглощения в каждой лунке определяют при длине волны 450 нм для определения концентрации связанного антитела и с определением поглощения при 650 нм для получения значения контрольного варианта. Поглощение измеряют с использованием счетчика для микротитрационных планшетов (Molecular Devices). Образование IgG1 подтверждается с помощью метода ELISA. Молекулярную массу экспрессированного слитого белка DR5-IgG1 определяют анализом по методу вестерн-блоттинга, в котором используют мАТ против человеческого IgG1 (Sigma) для обнаружения антитела в геле. Экспрессированный слитый белок DR5-IgG1 имеет примерную молекулярную массу 50 кДА. При этом достигается чистота более 90% при оценке электрофорезом в ДСН-ПААГ и путем обнаружения белка окрашиванием кумасси синим.
Пример 2. Образование моноклональных антител против человеческого DR5
2.1 Иммунизация
Самок 6-8-недельных мышей Balb/c (Jackson Laboratory, Bar Harbor, ME) иммунизируют человеческим слитым белком DR5/hIgG1, очищенным аффинной хроматографией. Для сенсибилизации подушечек ног слитый белок (50 мкг) эмульгируют в полном адъюванте Фрейда (Difco, Detroit, MI). Далее проводят бустер-иммунизацию мышей посредством четырех инъекций по 50 мкг слитого белка, вводимого без адъюванта через день. Через три дня после инъекции лимфоциты из периферических лимфатических узлов сливают с миеломными клетками NS-1 и гибридомы культивируют в средах F104 с добавкой 10% фетальной сыворотки теленка. Положительные гибридомы отбирают с помощью метода ELISA, в котором планшеты покрывают либо 1 мкг/мл DR5/hIgG1, либо тем же самым количеством Fas/hIgG1 в качестве контроля. Изотип гибридомы определяют по методу ELISA с использованием панели козьих антител, специфичных к изотипу Ig мыши (Southern Biotechnology, Birmingham, AL). Моноклональные антитела очищают аффинной хроматографией с использованием иммобилизованных антител против мышиного IgG1 или белка G (Sigma).
2.2 Слияние клеток
На третий день после бустерной инъекции у мышей удаляют периферические лимфоузлы и помещают в 10 мл бессывороточной среды RPMI 1640 (GIBCO BRL), содержащей 50 единиц/мл пенициллина, 50 мкг/мл стрептомицина и 300 мкг/мл L-глютаминовой кислоты и разрушают пропусканием органа через сетку (Cell Strainer; Falcon) с использованием шпателя. Полученную суспензию клеток центрифугируют для осаждения клеток периферических лимфоузлов, которые затем дважды промывают бессывороточной средой RPMI. Промытые клетки повторно суспендируют в бессывороточной среде RPMI и подсчитывают.
Тем временем миеломные клетки NS1 (Американская Коллекция типовых культур, American Type Culture Collection TIB-18) растят до плотности клеток, не превышающей 1·108 клеток/мл в среде АSF104 (Ajinomoto, K.K.), содержащей 10 об./об.% ФСТ (Gibco BRL) (среда ASF с сывороткой) при 37°С и в атмосфере 5 об./об.% CO2 и все разрушают аналогичным образом, промывают, ресуспендируют и подсчитывают число клеток.
Количество клеточной суспензии NS1, в которой, по подсчетам, содержится 3·107 клеток, смешивают с количеством суспензии клеток селезенки, в которой, по подсчетам, содержится 3·108 клеток. Полученную смесь центрифугируют и отбрасывают супернатант. Следующие стадии слияния клеток проводят при выдерживании пластиковых пробирок, содержащих осадок, при 37°С с стакане с теплой водой.
Затем к пробирке медленно добавляют один мл 50% (вес/об.) полиэтиленгликоля 1500 (Boehringer Manheim), все время перемешивая осадок концом пипетки. После этого медленно в виде 2 порций добавляют 1 мл бессывороточной среды RPMI, предварительно подогретой до 37°C, и затем добавляют еще 7 мл бессывороточной среды RPMI. Далее полученную смесь центрифугируют, супернатант отбрасывают и добавляют 10 мл среды HAT, содержащей 10 об./об.% ФСТ, при осторожном перемешивании концом пипетки. Добавляют еще 20 мл среды HAT, содержащей 10 об./об.% ФСТ, и суспензию распределяют по лункам 96-луночных микропланшетов для культивирования клеток по 100 мкл/лунку и инкубируют при 37°С в атмосфере 5 об./об.% CO2. Через 7 или 8 дней используют свежую HAT-среду в количестве по 100 мкл/лунку для замены среды во всех лунках, демонстрирующих желтоватый оттенок. Слитые белки из указанных лунок клонируют при ограниченном разбавлении, как описано ниже.
2.3 Клонирование при ограниченном разбавлении
Удаляют тимусы у 4-8-недельных самок мышей Balb/c (от Japan SLC, Inc.), разрушают на луночной сетке (Cell Strainer; Falcon), как указано выше, и разрушенные клетки промывают дважды средой HT, содержащей 10 об./об.% ФСТ. Количество тимусных клеток, соответствующих их количеству, отобранному от одной мыши, суспендируют в 30 мл среды HT, содержащей 10 об./об.% ФСТ, с получением питающей суспензии клеток. Препарат слитых клеток, полученный, как описано выше в примере 2.2, разбавляют указанной питающей суспензией клеток в 10-100 раз и затем делают серийное разбавление питающей суспензией клеток с получением суспензии, характеризующейся плотностью слитых клеток 5, 1 и 0,5 клеток/мл. Полученные таким образом образцы распределяют по 96-луночным микропланшетам для культивирования клеток по 100 мкл/лунку и инкубируют в течение 5 дней при 37°С в атмосфере 5 об./об.% CO2.
2.4 Скрининг
Культуральные супернатанты от растущих гибридом подвергают скринингу по методу ELISA с использованием планшетов, покрытых либо 1 мкг/мл DR5/hIgG1, либо тем же количеством Fas/hIgG1 (41) в качестве контроля. Связанные антитела выявляют с использованием антимышиных иммуноглобулинов, конъюгированных с пероксидазой хрена (ПХ) (Southern Biotechnology, Birmingham, AL) с использованием ТМБ в качестве субстрата (Sigma, St Louis, MI). Очищенный DR5-IgG1 в концентрации 1 мкг/мл или такое же количество Fas-hIgG1 вносят в лунки 96-луночного планшета ELISA/RIA STRIP PLATE (Costar, NY). Планшет выдерживают при 4°С в течение ночи для адсорбции белка на поверхности лунки. По истечении указанного периода времени раствор из лунок отбрасывают и каждую лунку промывают 3 раза ФБР-Твином. Затем к каждой лунке добавляют 100 мкл ФБР, содержащего 1% (вес/об.) бычьего сывороточного альбумина (А3803; Sigma Chemical Co.) и планшет инкубируют при 37°C в течение 1 часа. Затем лунки промывают еще 3 раза ФБР-Твином и к каждой лунке добавляют по 50 мкл каждого из культуральных супернатантов от растущих гибридом. Далее планшет инкубируют при 37°С в течение 1 часа и лунки снова промывают 4 раза ФБР-Твином. После промывания метят 50 мкл пероксидазы хрена козье антитело против мышиного иммуноглобулина (Southern Biotechnology, Birmingham, AL), разбавленное в 1000 раз ФБР, и вносят в лунки, затем планшет снова инкубируют при 37°С в течение одного 1 часа, после чего лунки промывают 4 раза ФБР-Твином. Далее добавляют 3,3',5,5'-тетраметилбензидиновую (ТМБ) жидкую субстратную систему (Sigma) в количестве по 100 мкл/лунку и планшет выдерживают при комнатной температуре в течение 5 минут, после чего реакцию останавливают добавлением 0,2N H2SO4 в количестве 100 мкл/лунку. Величину поглощения в каждой лунке определяют при 450 нм (в случае контроля при 650 нм) с использованием счетчика для микропланшетов (Molecular Devices) и слившиеся клетки отбирают из образца, который имеет поглощение (450 нм-650 нм, значение ОП>0,5) явно выше тех, к которым не добавляли супернатант от слившихся клеток (значение ОП≈0,03). Далее культуральный супернатант из растущих гибридом подвергают функциональному скринингу путем измерения апоптоз-индуцирующей активности в клетках Jurkat. Пятьдесят мкл среды RPMI, содержащей клетки Jurkat (1000 клеток на лунку) и 5 мкМ бисиндолилмалеимида VIII (BisVIII, Alexis, San Diego, CA), добавляют к лункам 96-луночных планшетов в присутствии 50 мкл культуральных супернатантов от растущих гибридом. Клетки культивируют в увлажненном инкубаторе при 37°С в течение ночи. Апоптоз определяют по жизнеспособности клеток с использованием набора для тестирования ATPLite по инструкции от производителя (Packard Instrument) и в образцах проводят подсчет с использованием счетчика TopCounter (Packard Instrument).
2.5 Определение связывания TRAIL и TRA-8 с рецепторами по методу ELISA
На планшеты для ELISA наносят 2 мкг/мл слитого белка DR4-Ig или DR5-Ig на ночь. После блокирования с использованием 3% БСА добавляют TRAIL-FLAG или TRA-8 в указанных концентрациях и инкубируют при 37°С в течение одного часа. Связывание TRAIL или TRA-8 определяют с использованием анти-Flag антитела, конъюгированного с ПХ (Alexis) или антитела против мышиного IgG1, конъюгированного с ПХ (Southern Biotechnology), соответственно. Реакции развиваются с субстратным буфером ТМП и течение их оценивают путем измерения на счетчике для микропланшетов Benchmark Microplate (BioRad). Значения Kd вычисляют на основе односайтовой модели связывания методом нелинейной регрессии с использованием программного обеспечения GraphPad Prism (GraphPad Software, San Diego, CA). Для проведения конкурентного ELISA добавляют 100 нг/мл TRAIL-FLAG и инкубируют в присутствии разных концентраций TRA-8. Связывание TRAIL определяют, как указано выше.
2.6 Клонирование
Стадии, указанные выше в примерах 2.3 и 2.4, повторяют 5 раз для клеток, отобранных в соответствии с пунктом 2.4, позволяющим сделать отбор нескольких клонов гибридомы, каждый из которых продуцирует одно антитело, которое связывает DR5-IgG, но не связывает Fas-IgG. В результате такой процедуры отбора может быть получена мышиная-мышиная гибридома, обозначенная как TRA-8 и продуцирующая антитело, связывающееся с DR5-IgG, но не с Fas-IgG. Указанная гибридома TRA-8 была депонирована в Американской Коллекции типовых культур 1 марта и имеет номер доступа PTA-1428.
Подкласс антитела, продуцируемого мышиной-мышиной гибридомой TRA-8 (далее обозначаемой просто как «TRA-8»), относится к IgG1, как было показано после тестирования с набором для изотипирования моноклонального антитела (Pierce).
С использованием человеческого слитого белка DR5-IgG1 в качестве иммуногена было получено семь гибридомных клонов при первоначальном скрининге по методу ELISA, и все они обладали высоко выраженными позитивными свойствами в отношении DR5-IgG, но не в отношении слитого белка Fas-IgG, указывая на то, что полученные гибридомы продуцируют антитела, которые распознают внеклеточную часть DR5, но не Fc-часть IgG1 (данные не показаны).
2.7 Анализ по методу вестерн-блоттинга
Фильтры для проведения вестерн-блот анализа человеческого гомогената нормальной ткани и ткани злокачественной опухоли получают от компании Geno Technology (St Louis, MO). Для каждой линии вносят равное количество белка по данным определения анти-β-актинового антитела. Блоты зондируют в течение ночи с использованием 1 мкг/мл TRA-8 и затем козьего антитела против мышиного IgG1, конъюгированного с ПХ (Southern Biotechnology), при комнатной температуре в течение одного часа и затем проявляют в условиях хемилюминесценции.
2.8 Иммуногистохимия in situ
Ткани человека получают из Центра заготовки ткани (Tissue Procurement Center UAB). Замороженные секции фиксируют 70%-ным этанолом, блокируют 10% лошадиной сывороткой в ФБР и затем инкубируют с 10 мкг/мл очищенного аффинной хроматографией TRA-8 при комнатной температуре в течение 60 минут. Для визуализации реакционной способности используют в качестве колориметрического субстрата набор АВС с антителом против мышиного IgG и диаминобензидином (Vector, Burlingame), CA).
2.9 Анализ каспазной активации
Клетки Jurkat (1·106/мл) инкубируют с 500 нг/мл TRA-8. Аликвоты (30 мкг белка) клеточного лизата разделяют электрофорезом на 15% ДСН-ПААГ, блоты переносят на нейлоновые мембраны, далее зондируют с использованием антител против каспазы 8, 9 и 3 (BD Pharmingen, San Diego, CA) и затем вторичного антитела, конъюгированного с ПХ, после чего проводят визуализацию расщепленных продуктов хемилюминесцентным методом. Набор ингибиторов каспазы получают от компании R&D System (Minneapolis, MN). Каждый ингибитор каспазы добавляют в культуру в указанных концентрациях.
Пример 3. Очистка моноклонального антитела TRA-8
Мышиную-мышиную гибридому TRA-8 выращивают до плотности клеток 1·106 клеток/мл при инкубации в 500 мл среды ASF, содержащей 10 об./об.% ФСТ, при 37°C в атмосфере 5 об./об.% СО2 в течение 5 дней. Затем культуру центрифугируют (1000 об/мин, 5 минут) и собирают супернатант. Очистку TRA-8 из супернатанта проводят аффинной хроматографией, включающей белокG-сефарозу CL-4B (Pharmacia) в следующих условиях:
колонка: белокG-сефароза CL-4B (размер колонки 2 мл; Pharmacia);
элюирующий буфер: 0,1 М глицин (рН 2,4), 0,15 М NaCl;
нейтрализующий буфер: 1 М Трис-HCl (pH 8,5).
После внесения супернатаната на колонку проводят промывание ее три раза с использованием каждый раз по 20 мл ФБР и затем добавляют 10 раз по 1 мл элюирующего буфера. Промеряют оптическую плотность каждой элюированной фракции. Отдельно собирают фракции с № 2 по № 5(>ОП280=0,1).
После добавления 100 мкг нейтрализующего буфера элюаты помещают по отдельности в диализную пробирку и диализуют против 1 литра ФБР (pH 7,5) при 4°C. Буфер для диализа меняют дважды. Указанный образец анализируют на наличие активности против антитела DR5 методом ELISA с использованием человеческого слитого белка DR5-IgG, приготовленного по описанной выше методике.
Пример 4. Приготовление антигена DR4, вектора экспрессии DR4-IgG и моноклонального антитела к DR4
Процедуры, описанные в примерах 1-3, повторяют с использованием матричной кДНК для DR4 и соответствующих праймеров вместо тех, что были подробно описаны в примере 1, с получением антигена DR4, который используют в примерах 1.2-3 для получения моноклонального антитела, специфичного к DR4.
Пример 5. Моноклональные антитела против DcR1 и DcR2
Моноклональные антитела против ложных рецепторов DcR1 и DcR2 получают путем замены соответствующими вариантами кДНК и праймеров из примера 1 с образованием соответствующих антигенов, аналогично тому, как было показано в примере 1. Векторы экспрессии для слитых белков DcR1 или DcR2 с иммуноглобулином G и образуемые в итоге очищенные антитела конструируют по методикам, описанным в примерах 2 и 3.
Пример 6. Специфичность моноклонального антитела
Как в случае всех рецепторов для TRAIL и других белков семейства TNFR, характеризующихся высокой гомологией, специфичность рассматриваемого в качестве иллюстративного примера антитела TRA-8 к DR5 определяют методом вестерн-блоттинга с использованием двух разных слитых белков DR5-IgG и растворимых рекомбинантных форм других родственных белков. Первый слитый белок DR5-Ig был сконструирован путем слияния кДНК на участке остатков 1-180 во внеклеточной части DR5 и кДНК, кодирующей константный участок человеческого IgG1. Полученную после слияния кДНК клонируют в рекомбинантном векторе аденовируса (Quantum Biotechnologies, Inc., Montreal, Canada). Экспрессируемый слитый белок DR5/hIgG1, имеющий относительную молекулярную массу 50 кДа, очищают с использованием аффинной хроматографии на колонке с антителом против человеческого IgG (Sigma, St.Louis, MO). Для анализа специфичности методом вестерн-блоттинга приобретают второй рекомбинатный человеческий слитый белок DR5/IgG1 (аа 52-212), а также слитые белки TRAIL-R1, R3 и R4 от компании Alexis. Растворимые формы человеческих Fas и TNFR1 были любезно предоставлены Dr. Carl Edwards of Amgen, Inc., Thousands Oaks, CA, USA. Применявшиеся растворимые формы рекомбинантных человеческих молекул DR4, DcR1, DcR2, TNFR1, R4 и Fas представляли собой слитые белки на основе человеческого IgG1. По 0,5 мкг каждого из белков отделяют электрофорезом в 10% ДСН-ПААГ и переносят на нитроцеллюлозную мембрану. Блоты блокируют добавлением 5% сухого молока в ФБР при комнатной температуре на один час и затем добавляют в качестве зонда 1 мкг/мл очищенного моноклонального антитела к DR5 (клон: TRA-8) или 0,1 мкг/мл ПХ-конъюгированного козьего антитела против человеческого IgG при 4°С в течение ночи. Козье антитело против мышиного IgG, конъюгированного с пероксидазой хрена (ПХ), используют в качестве вторичного антитела для обнаружения связанного TRA-8. Блоты выявляют с помощью хемилюминесценции.
Cos-7 клетки после трансфекции вектором pcDNA3 (Clontech, Palo Alto, CA), содержащие DR5 или DR4 полной длины или пустой вектор, используют в анализе методом проточной цитометрии. кДНК полной длины, кодирующую человеческий TRAIL или мышиный Fas-лиганд, клонируют в векторе pTRE по ходу транскрипции от контролируемого тетрациклином промотора (Clontech). Фрагменты XhoI-HindIII в pTRE-hTRAIL или pTRE-mFasL далее клонируют в шаттл-векторе аденовируса pAdBN (Quantum Biotechnologies, Inc.). Клетки-хозяева 293 подвергают совместной трансфекции линеаризированным вектором pAd-TRE-hTRAIL или pAD-TRE-mFasL и крупным фрагментом ДНК аденовируса. Экспрессию функционального человеческого TRAIL или мышиного Fas-лиганда отслеживают с использованием теста на высвобождением 51Cr в клетках Jurkat, взятых в качестве мишени.
TRA-8 вступает в мощное взаимодействие со слитым белком DR5-IgG (≈50 кДа), который используется для иммунизации, как показано на фиг.1а, DR5#1, и слабо взаимодействует со вторым слитым белком, DR5-IgG (≈60 кД), как показано на фиг.1а, DR5, #2. Не отмечается значительного связывания TRA-8 с DR4, DcR1, DcR2, Fas (CD95) или TNFRI. Приведенные результаты указывают на то, что TRA-8 распознает эпитопы, которые специфичны для DR5, но не для других представителей данного семейства.
TRA-8 не вступает во взаимодействие с представителями суперсемейства TNF-рецепторов, такими как Fas (CD95) и TNF-рецептор I, а также TRA-8 не вступает в перекрестную реакцию с мышиным гомологом DR5, что видно по соотношению значений оптического поглощения при 450 нм и 650 нм, приведенных на нижней панели чертежа под номерами 1-7 (фиг.1а, колонка 8 в нижней панели). Растворимый TRAIL и TRA-8 связываются в сравнимой степени с иммобилизованным DR5 (фиг.1b, левая панель). В отличие от указанной ситуации TRAIL связывается с DR4, но TRA-8 не демонстрирует какой-либо связывающей активности в отношении DR4 (фиг.1b, средняя панель). Значения Kd для связывания TRAIL и TRA-8 с DR5 подсчитаны для 59 нМ и 3 нМ соответственно. Важно отметить, что TRA-8 эффективно конкурирует с TRAIL за связывание с DR5, но не за связывание с DR4, как видно по результатам конкурентного ELISA (фиг.1b, правая панель). Приведенные результаты демонстрируют специфичность TRA-8 в отношении человеческого DR5.
TRA-8 способен выявлять экспрессию DR5 на клеточной поверхности, при этом анализ методом проточной цитометрии указывает на наличие специфического связывания с клеточной поверхностью Cos-7 клеток, трансфицированных человеческим DR5 полной длины, но не Cos-7 клеток, трансфицированных DR4 или пустым вектором (фиг.1с). Аналогично иммуногистохимический анализ in situ указывает на реакционную способность TRA-8 в отношении Cos-7 клеток, трансфицированных ДНК DR5 полной длины, но не клеток, трансфицированных контрольным вектором (фиг.1d). TRA-8 не индуцирует апоптоз нетрансфицированных Cos-7 клеток, при этом ОТ-ПЦР на основе РНК из Cos-7 клеток с использованием пары праймеров, кодирующих человеческий DR5, не выявляет специфических продуктов ПЦР. Дополнительный функциональный анализ с использованием человеческих клеток Jurkat в качестве мишеней показывает, что в отсутствие сшивания TRA-8 обладает мощной способностью по индукции гибели клеток, что подтверждается тремя разными анализами по оценке жизнеспособности клеток, включая ATPLite, MTT и PI исключения (фиг.1е). Отмечается гибель более 50% клеток Jurkat под действием нанограммовых количеств TRA-8, на что указывают результаты анализа ATPLite. Активность TRA-8 по уничтожению клеток специфична для DR5, поскольку она может блокироваться DR5-Ig, но не слитым белком DR4-Ig (данные не показаны). Расщепление под действием каспаз 8, 9 и 3 может быть обнаружено по вестерн-блот анализу уже через 30 минут после обработки TRA-8 клеток Jurkat (фиг.1f), тогда как гибель клеток Jurkat полностью подавляется основным ингибитором каспазы (Z-VAD) (фиг.1g). Индивидуальные ингибиторы каспазы 8, 3, 9 и 10 частично подавляют гибель клеток, дополнительно указывая на тот факт, что TRA-8-опосредованная гибель клеток происходит в основном через зависимый от каспазы механизм апоптоза.
Пример 7. Анализ методом проточной цитометрии экспрессии на клеточной поверхности DR5: основного рецептора, опосредующего гибель многих опухолевых клеток, но не нормальных клеток
Способность TRA-8 связываться с DR5, экспрессированным на клеточной поверхности, и специфичность указанной реакции оценивают затем с использованием COS-7 клеток (Американская Коллекция типовых культур (American Type Culture Collection No. CRL-1651)), трансфицированных вектором экспрессии, содержащим кДНК для человеческого DR5 или DR4 полной длины или пустой вектор в качестве контроля. Антитело против мышиного IgG1, конъюгированного с фикоэритрином (ФЭ) (Pharmigen), используют в качестве второго антитела для выявления связанного TRA-8. Измеряют флуоресценцию 1·104 клеток с использованием проточного цитометра (FACSVantage) в следующих условиях:
Длина волны возбуждения: 488 нм;
Длина волны обнаружения: 600 нм.
Анализ с использованием метода проточной циометрии показывает, что TRA-8 окрашивает примерно 30% COS-7 клеток, трансфицированных вектором DR5, как видно на гистограмме, приведенной в виде жирных линий на фиг.1с. Величина приведенного процента коррелирует с эффективностью трансфекции, как следует из результатов анализа трансфекции с использованием зеленого флуоресцентного белка (GFP) (данные на показаны). TRA-8 окрашивает лишь незначительно клетки, трансфицированные DR4 (открытая гистограмма) или контрольным вектором (гистограмма с пунктирными линиями), указывая на то, что TRA-8 специфичен для DR5, имеющегося на клеточной поверхности.
Несмотря на то что экспрессия DR5 в опухолевых клетках исследовалась очень широко на уровне мРНК (см. приведенные в описании ссылки), экспрессия DR5 на клеточной поверхности не описана. Таким образом, доступность моноклонального антитела к DR5 позволяет определить уровень DR5 на клеточной поверхности и сопоставить экспрессию с чувствительностью клеток к TRAIL-опосредованному апоптозу. Указанную ниже панель клеток (1·106) инкубируют с 10 мкг/мл TRA-8 при комнатной температуре в течение 30 минут и затем окрашивают конъюгированным с ФЭ антителом против мышиного IgG1 (Pharmingen) еще в течение 30 минут. В проточном цитометре FACSvantage анализируют 10000 жизнеспособных клеток при соблюдении указанных ниже условий:
Длина волны возбуждения: 488 нм;
Длина волны обнаружения: 600 нм.
Пять исследованных клеточных линий включают клетки Jurkat, CEM-6, Molt-4, H-9 и U937. Экспрессия DR5 обнаруживается на клеточной поверхности Jurkat, CEM-6, H-9 и U937 клеток, но практически не выявляется на Molt-4 клетках, как показано на фиг.2а и 2а'. Несмотря на то что раньше публиковались данные о высоком уровне экспрессии РНК DR5 (43), результаты FAC анализа указывают на то, что данные клетки не экспрессируют высоких уровней поверхностного DR5. Приведенные результаты показывают, что экспрессия DR5 на клеточной поверхности не коррелирует с экспрессией DR5 на уровне транскрипции DR5, что не является неожиданным для такого рецептора. Уровень экспрессии DR5 может быть специфичным для направления дифференцировки клеток, поскольку большая часть клеток гематопоэтического происхождения экспрессирует низкие уровни, тогда как большая часть глиомных клеток и клеток предстательной железы экспрессирует высокие уровни DR5.
Моноклональное антитело TRA-8 используют для определения роли DR5 в индукции TRAIL-опосредованного апоптоза путем изучения экспрессии на клеточной поверхности панели опухолевых клеток человека разного типа, а также чувствительности указанных клеток к TRAIL- и TRA-8-опосредованному апоптозу. Первичные Т-клетки периферической крови не экспрессируют существенных количеств DR5 на поверхности клеток и характеризуются устойчивостью к TRAIL- и TRA-8-опосредованному апоптозу (фиг.2a, 2a' и 3a'). Несмотря на то что пять клеточных линий человеческого Т-клеточного лейкоза экспрессируют относительно низкие уровни DR5 на клеточной поверхности, две из них (Jurkat и CEM-6) отличаются высокой чувствительностью к TRAIL-опосредованному и TRA-8-опосредованному апоптозу, что свидетельствует о том, что одного DR5 достаточно для индукции апоптоза в данных клетках. Клетки Molt-4 и U937 характеризуются частичной чувствительностью к TRAIL-опосредованному апоптозу, но являются относительно устойчивыми к TRA-8-опосредованному апоптозу, и это дает основание полагать, что другие TRAIL-рецепторы также могут участвовать в трансдукции апоптозного сигнала. H-9 клетки устойчивы к TRAIL- и TRA-8-опосредованному апоптозу, представляя блок внутриклеточного антиапоптозного пути.
Исследованная панель клеток включает клеточные линии злокачественных глиомных клеток человека Hs683, U251MG, D37MG, D54MG, U373MG, CH235MG, U87 и нормальные человеческие астроциты, которые были предоставлены Dr. Yancey Gillespie из Neurosurgery Department of the University of Alabama в Birmingham. Клеточные линии рака предстательной железы человека Du154, PC3 и LnCap были предоставлены Dr. William Grizzle из Pathology Department of the University of Alabama в Birmingham, который получил указанные клеточные линии из American Type Culture Collection. Человеческие Т-клеточные линии лейкоза, В-клеточной лимфомы, клетки HepG2 Jurkat (Американская Коллекция типовых культур TIB-152) и клетки CCRF-CEM CEM-6 (Американская Коллекция типовых культур CCL-119); моноцитные клеточные линии, U937 (Американская Коллекция типовых культур CRL-2367) были получены из Американской Коллекции типовых культур. Все указанные выше клеточные линии культивируют в среде RPMI 1640 с добавкой 1-% ФСТ. Клеточная линия астроцитомы человека 1321N1 была любезно предоставлена Dr. Richard Jope of Psychiatry Department из University of Alabama в Birmingham и культивировалась в среде DMEM с добавкой 5% ФСТ.
Растворимый рекомбинантный человеческий TRAIL, полученный от компании Alexis Corporation, San Diego, CA, представляет собой слитый белок, включающий внеклеточный домен человеческого TRAIL (аминокислотные остатки 95-281), слитый на N-конце с FLAF-tag и пептидным линкером из 8 аминокислот. В отличие от описанного ранее His-tag-содержащего TRAIL данный препарат TRAIL сам не индуцирует сильной апоптозной реакции в клетках Jurkat и требует наличия анти-FLAG антитела в качестве сшивателя для усиления апоптоза. Анти-FLAG антитело было также получено от компании Alexis.
Все 10 злокачественных линий глиомных клеток человека экспрессируют выявляемые количества DR5 на клеточной поверхности. Большая часть из них экспрессирует DR5 на уровне от среднего до высокого, как показано на фиг.2b. Три линии D-54MG, U373MG и CH-235MG экспрессируют высокие уровни DR5, тогда как шесть линий Hs-683, U251-MG, D37-MG, U87, SMK1 и 1321N1 экспрессируют DR5 на среднем уровне. Только одна клеточная линия H-465 экспрессирует низкий уровень DR5. Все три клеточных линии предстательной железы человека экспрессируют высокие уровни DR5, как показано на фиг.2с.
Как и нормальные первичные Т-клетки, первичные В-клетки не экспрессируют значительных количеств DR5 и не подвергаются апоптозу после обработки TRAIL или TRA-8 (фиг.2d). Три (SKW 6.4, EB-3 и Raji) из четырех клеточных линий В-лимфомы экспрессируют относительно высокие уровни DR5 и очень чувствительны к TRAIL- и TRA-8-опосредованному апоптозу. Четвертая клеточная линия Daudi экспрессирует очень низкие уровни DR5 и характеризуется намного меньшей чувствительностью к TRAIL- или TRA-8-опосредованному апоптозу. Несмотря на то что первичные астроциты не экспрессируют выявляемых количеств DR5 на клеточной поверхности (фиг.2b'), все четыре глиомных клеточных линии экспрессируют высокие уровни DR5. Более высокий уровень экспрессии DR5 в глиомных клетках, чем в Т- и В-клетках не сопровождается их значительно более высокой чувствительностью к TRAIL-DR5-опосредованному апоптозу, что позволяет предположить, что уровень экспрессии на клеточной поверхности DR5 не обязательно коррелирует с уровнем апоптоза опухолевых клеток. ПЦР-ОТ, проведенная с целью определения уровня матричной РНК для DR4, DR5 и DCR2, выявила матричную РНК во всех исследованных клетках (таблица 1). Однако в целом первичные нормальные клетки экспрессируют относительно низкие уровни DR5 в сравнении с трансформированными опухолевыми клетками.
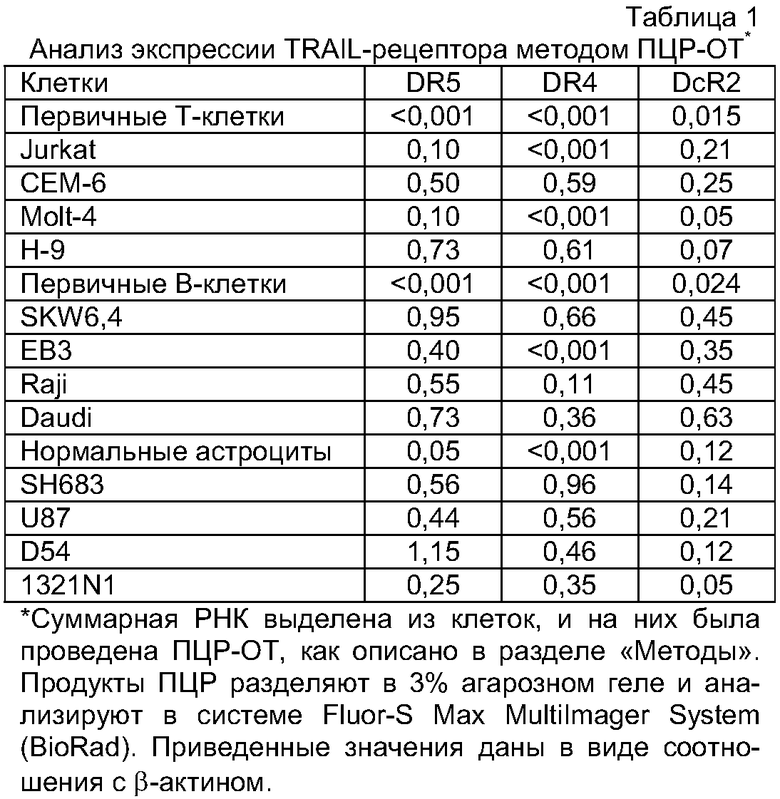
Пример 8. Индукция апоптоза in vitro в злокачественных клетках
Для выяснения того факта, индуцирует ли TRA-8 апоптоз в трансформированных клетках in vitro, все DR5-положительные опухолевые клетки исследуют на чувствительность к апоптозу, индуцированному либо TRA-8, либо TRAIL.
Целевые клетки (1·103 на лунку) культивируют в 96-луночных микротитрационных планшетах в присутствии указанных концентраций растворимого TRAIL плюс сшиватель (Alexis) или TRA-8 при 37°C в течение ночи. Жизнеспособность клеток оценивают с использованием (1) набора ATPLite в соответствии с инструкцией производителя (Packard Instruments, Meriden, CT); (2) набора MTT для оценки пролиферации/жизнеспособности клеток (Sigma); или (3) путем PI окрашивания мертвых клеток с последующим анализом методом проточной цитометрии. В конце культивирования клетки окрашивают PI в концентрации 10 мк/мл и PI-негативные клетки отсортировывают как жизнеспособные клетки. Для анализа конденсированных ядер гепатоцитов клетки окрашивают PI в концентрации 10 нг/мл с использованием Hoechst 33352 (Molecular Probes) и анализируют проточной цитометрией.
Антитело TRA-8 способно индуцировать апоптоз в большей части клеточных линий злокачественных глиомных клеток человека (9/10), в 2 из 3 клеточных линий рака предстательной железы и в 2 из 4 DR5-положительных гематопоэтических клеточных линий. Оно не индуцирует апоптоз в клеточной линии Molt-4, которая экспрессирует практически невыявляемые уровни DR5 на клеточной поверхности. Однако степень чувствительности клеток к TRA-8-опосредованному апоптозу варьирует в значительной мере между клеточными линиями.
Вариабельность в чувствительности клеток к антителу TRA-8, индуцирующему апоптоз, позволяет предположить, что, хотя и требуется минимальный уровень экспрессии DR5 на клеточной поверхности, указанный уровень экспрессии DR5 на клеточной поверхности не обязательно представляет собой основной показатель, определяющий чувствительность, а имеются другие факторы, влияющие на данный процесс. Несмотря на то что все глиомные клетки в основном экспрессируют более высокие уровни DR5 на клеточной поверхности, чем гематопоэтические клетки, чувствительность глиомных клеток к апоптозу, индуцированному TRA-8, непропорционально высока в сравнении с гематопоэтическими клетками. Чувствительность пяти клеточных линий глиомных клеток D-37MG, D54-MG, U373-MG, CH235-MG и 1321N1 к TRA-8-индуцированному апоптозу высока и эквивалента их чувствительности к TRAIL-опосредованному апоптозу, как показано на фиг.3b. Две глиомных клеточных линии H-456 и SMK1 намного менее чувствительны к апоптозу, индуцированному TRA-8. В случае Н-456 клеток экспрессия DR5 на поверхности клеток осуществляется на низком уровне, однако уровень экспрессии DR5 на поверхности SMK1 аналогична таковой, показанной для более чувствительных клеточных линий, что позволяет предположить, что также другие механизмы могут играть роль в определении чувствительности TRAIL-опосредованного апоптоза. Несмотря на то что все три клеточных линии клеток рака предстательной железы экспрессируют высокий уровень DR5, наибольшей чувствительностью к TRA-8-индуцированному апоптозу характеризуются клетки Du145, клетки PC3 характеризуются частичной чувствительностью, а клетки LnCAP являются полностью резистентными, как показано на фиг.3с. В отношении гемопоэтических клеток было показано, что клетки Jurkat и CEM-6 очень чувствительны к апоптозу под действием TRA-8, как видно из фиг.2а, хотя, как было показано, обе указанных клеточных линии экспрессируют очень низкие уровни DR5. Несмотря на то что DR5 выявляется в клетках U937, указанные клетки резистентны к TRA-8-индуцированному апоптозу. Аналогично несмотря на то что H-9-клетки экспрессируют обнаруживаемые количества DR5, H-9-клетки резистентны к апоптозу, индуцированному TRA-8. Приведенные результаты указывают на существование регуляторных механизмов, которые влияют на DR5-опосредованный апоптоз.
Дополнительные антитела, связывающиеся на поверхности клеток, против DR5 получают с использованием процедур, описанных в примерах 1-3. Были изучены два дополнительных антитела против DR5, обозначенные как TRA-1 и TRA-10, в сочетании с TRA-8 для определения их сравнительной способности индуцировать апоптоз, действуя, таким образом, в качестве агониста, или, наоборот, блокировать TRAIL-опосредованный апоптоз, действуя, таким образом, в качестве антагониста. Были использованы человеческие клетки Jurkat в качестве мишени для определения агонистической и/или антагонистической активности трех антител против DR5, обозначенных как TRA-1, TRA-8 и TRA-10. Как видно из фиг.4, жизнеспособность клеток составляет примерно 90%, 70% и 20% для TRA-10, TRA-1 и TRA-8 соответственно при инкубировании в течение ночи с 2,5 мкг на мл. TRA-8 индуцирует мощную апоптозную реакцию зависимым от дозы образом, тогда как TRA-1 индуцирует лишь умеренную апоптозную реакцию и TRA-10 индуцирует слабую реакцию. В этой связи TRA-8 классифицируется как агонист антитела против DR5. На фиг.4 проиллюстрирована жизнеспособность человеческих клеток Jurkat в виде функции TRAIL-индуцированного апоптоза. TRA-10 блокирует в значительной мере апоптоз человеческих клеток Jurkat даже в низкой дозе, как было показано при изучении TRAIL-индуцированного апоптоза. Таким образом, TRA-10 классифицируется как антагонист антитела против DR5.
Чувствительность пяти глиомных клеточных линий: D-37MG, D54-MG, U373-MG, CH235-MG и 1321N1 к TRA-8-индуцированному апоптозу эквивалентна их чувствительности к TRAIL-опосредованному апоптозу, как показано на фиг.3b, указывая на то, что TRAIL-индуцированный апоптоз в данных клетках осуществляется в основном через DR5. Кроме того, две глиомных линии Hs683 и U251-MG демонстрируют устойчивость к TRAIL-индуцированному апоптозу, но частично являются чувствительными к TRA-8-индуцированному апоптозу, указывая на функционирование ложных рецепторов в данных клетках, а также на то, что использование антитела TRA-8 позволяет обойти такой регуляторный механизм. В клеточных линиях рака предстательной железы несмотря на вариации в чувствительности к апоптозу, индуцированному TRA-8, параллельно наблюдается чувствительность клеток к апоптозу, индуцированному TRAIL, что вновь дает основание полагать, что DR5 играет важную роль в механизме TRAIL-опосредованного апоптоза в клетках рака предстательной железы. Среди гемопоэтических клеток было обнаружено, что клетки Jurkat и CEM-6 очень чувствительны как к TRA-8-, так и к TRAIL-опосредованному апоптозу. Уровень апоптоза, индуцированного TRA-8, сравним с таковым в случае индукции TRAIL, как показано на фиг.2а и 3а'. Только одна глиомная клеточная линия U87 и две гемопоэтических клеточных линии U937 и Molt-4 демонстрируют чувствительность к TRAIL-индуцированному апоптозу, но являются менее чувствительными или устойчивыми к TRA-8-индуцированному апоптозу. Одна клеточная линия H-9 экспрессирует выявляемые количества DR5, но является устойчивой к апоптозу, индуцированному TRA-8 или TRAIL. При том что для TRA-8-индуцированного апоптоза требуется минимальный уровень экспрессии DR5, уровень экспрессии DR5 не обязательно свидетельствует о чувствительности клеток к TRA-8-опосредованному апоптозу; ложные рецепторы играют роль в модуляции TRAIL-опосредованного апоптоза в некоторых клетках, но, по всей видимости, не играют заметной роли в большинстве исследованных до настоящего времени клеток; как ожидается, антитело TRA-8 будет обходить механизмы воздействия ложных рецепторов; функциональные мутации DR5 рецептора могут осуществляться в трансформированных клетках; и, наконец, внутриклеточные регуляторные механизмы могут быть так же важны или более важны, чем ложные рецепторы при определении чувствительности клеток к TRAIL- и DR5-опосредованному апоптозу.
В предыдущих исследованиях было показано, что мРНК для DR5 широко представлена в нормальных тканях [7]. Для оценки экспрессии DR5 в уровне белка проводят зондирование панели гомогенатов нормальной человеческой ткани (Geno Technology, St. Louis, MO) TRA-8-антителом в рамках анализа методом вестерн-блоттинга. Из девяти нормальных человеческих тканей мозговая ткань демонстрирует наименее выраженную положительную реакцию (фиг.5а, линия 2). При использовании TRA-8 в качестве диагностирующего инструмента белок DR5 не выявляется в печени (линия 1), легком (линия 3), почке (линия 4), селезенке (линия 5), яичках (линия 6), яичниках (линия 7), сердце (линия 8) или поджелудочной железе (линия 9). И наоборот, все тринадцать тканей злокачественных опухолей человека окрашиваются положительно при использовании TRA-8 (фиг.5b), включая варианты рака яичников (линия 1), легкого (линия 2), печени (линия 3), прямой кишки (линия 4), шейки матки (линия 5), кожи (линия 6), яичек (линия 7), щитовидной железы (линия 8), матки (линия 10), желудка (линия 11), ларингофарингеальной области (линия 12) и поджелудочной железы (линия 13). Более того, при проведении in situ иммуногистохимического анализа нормальных тканей и тканей злокачественных опухолей с использованием TRA-8 было подтверждено, что за исключением нескольких рассеянных положительных клеток в селезенке экспрессия DR5 в нормальных тканях молочной железы, легкого и селезенки не обнаруживается (фиг.5с). Тогда как соответствующие ткани злокачественных опухолей, включая инфильтрующую карциному протоков молочной железы, мелкоклеточный рак легкого и лимфому, реагируют положительно с TRA-8 (фиг.5d). Из 22 исследованных тканей злокачественных опухолей положительную реакцию с TRA-8 проявляют 5 из 6 вариантов рака молочной железы, 2 из 2 вариантов рака шейки матки, 4 из 5 вариантов рака печени, 5 из 8 вариантов лимфомы, 2 из 2 вариантов рака легкого и 2 из 2 вариантов рака предстательной железы. Приведенные результаты согласуются с данными анализа методом проточной цитометрии и показывают, что злокачественные клетки экспрессируют более высокие уровни белка DR5, чем нормальные клетки.
Пример 9. Активность TRA-8 in vivo по уничтожению опухолевых клеток
По разным причинам средства, которые рассматриваются как перспективные в исследованиях in vitro, не демонстрируют эффективности in vivo. Поэтому особенно важно проанализировать эффективность TRA-8 на модели животных in vivo. Для проведения такой оценки антитело TRA-8 против человеческого DR5 вводят мышам, содержащим человеческий ксенотрансплантат, который экспрессирует молекулу человеческого DR5. В исследовании используют 6-8-недельных мышей NOD/SCID (Jackson Laboratory), которым инокулируют подкожно клетки астроцитомы человека 1321N1 (1·107) или внутривенно клетки лейкоза человека Jurkat (1·106). На 2 день после инокуляции мышам инокулируют внутривенно TRA-8 (100 мкг). Через пять дней после введения TRA-8 оценивают рост опухолевых клеток 1321N1 по размеру и весу опухолевой массы. Рост клеток Jurkat оценивают по весу селезенки и по процентному содержанию CD3-положительных клеток Jurkat в селезенке животных после инокуляции. Отбирают образцы опухолевой ткани для биопсии и проводят их гистологическое исследование.
Показано, что раннее введение внутривенной дозы 100 мкг TRA-8, в первый день после инокуляции опухоли, полностью ингибирует образование клетками 1321N1 твердой опухоли (фиг.6а). А введение трех доз по 100 мкг TRA-8 в более поздние сроки, через одну неделю после инокуляции опухоли, снижает вес опухоли в 4 раза или более (фиг.6b). Образование опухоли не отмечается у тех животных, которым вводят TRA-8 в ранние временные точки (фиг.6c, верхняя панель). Гистологической анализ указывает на резкую дегенерацию опухолевой ткани у животных, которым вводили TRA-8 (фиг.6c, нижняя панель). Аналогично введение TRA-8 подавляет размножение клеток Jurkat в селезенке, на что указывает очень малое число CD3-положительных клеток Jurkat в селезенке (фиг.6d, 6e). Гистологический анализ имплантированной опухоли показывает наличие нескольких рассеянных опухолевых клеток в мягкой ткани животных после введения им TRA-8, тогда как в контролях отмечается образование твердой опухоли, как видно из фиг.6c. Как показано на модели клеток Jurkat при анализе методом проточной цитометрии (фиг.6а) и при окрашивании in situ с использованием CD3 (фиг.6с), число клеток Jurkat в селезенках животных после введения им TRA-8 составляет менее 2% против примерно 10% в селезенке контрольных животных.
Приведенные результаты подтверждают тот факт, что системное введение сшитого рекомбинантного TRAIL ингибирует рост опухоли in vivo [13]. Данные результаты указывают также на то, что однократная доза TRA-8 характеризуется высокой эффективностью по уничтожению опухолевых клеток in vivo.
Поскольку в исследовании на мышиной модели было использовано антитело человека, токсичность TRA-8 не оценивают. Однако исследования с введением TRAIL in vivo указывают на то, что указанное введение все-таки сопровождается незначительной токсичностью [13].
Пример 10. Синовиальные клетки при РА чувствительны к TRAIL- и TRA-8-индуцированному апоптозу
В большинстве работ, выполненных к настоящему времени, исследование TRAIL-опосредованного апоптоза проводилось в основном на злокачественных клетках. Согласно настоящему изобретению TRAIL-опосредованный апоптоз является терапевтическом инструментом также в случае аутоиммунных и воспалительных заболеваний, таких как РА.
10.1 Анализ методом проточной цитометрии экспрессии DR5 на клеточной поверхности синовиальных клеток при РА
Экспрессию DR5 в панели восьми культур синовиальных клеток, взятых у пациентов с РА, сравнивают с экспрессией в панели восьми культур синовиальных клеток, взятых у пациентов с остеоартритом (обозначаемым далее как OA). Восемь культур первичных синовиальных клеток при РА: RA-1014, RA-1016, RA-1021, RA-512, RA-707, RA-811, RA-716 и RA-929, были любезно предоставлены Dr. M. Ohtsuki, Sankyo Co., Ltd., Tokyo, Japan), которые культивируют в среде DMEM с добавкой 10% ФСТ, пенициллина, стрептомицина и глютамина. Выделяют семь культур первичных синовиальных клеток из синовиальных тканей пациентов с ОА по стандартной методике выделения с коллагеназой и культивируют в тех же условиях. Число пассажей всех первичных клеток составляет 10. Экспрессию DR5 определяют с использованием флюоресцентной сортировки клеток (FACS-анализ), как описано в примере 5.
Все культуры первичных РА клеток экспрессируют высокие уровни DR5 на своей поверхности и имеются лишь небольшие вариации в уровне экспрессии среди синовиальных клеток, выделенных от разных пациентов, как видно на фиг.7а. И наоборот, экспрессия DR5 на поверхности синовиальных клеток, выделенных от пациентов с OA, очень низка или вообще не выявляется, как показано на фиг.7b. Было показано, что трансформированные под действием SV40 синовиальные клетки экспрессируют высокие уровни DR5, сравнимые с уровнем, показанным для РА клеток. И наоборот, нетрансформированные фибробластные клетки экспрессируют низкие уровни DR5, сравнимые с уровнем, показанным для ОА клеток, как показано на фиг.7b.
10.2 Чувствительность синовиальных клеток при РА к апоптозу, опосредованному TRA-8 или TRAIL
В целом все синовиальные клетки, выделенные от пациентов с РА, чувствительны и к TRAIL, и к антителу против DR5, индуцирующим апоптоз, и все OA клетки устойчивы и к TRAIL, и к антителу против DR5, индуцирующим апоптоз, как показано на фиг.8а, b. Указанные исследования свидетельствуют о том, что антитело TRA-8 нацелено на действие предпочтительно против измененных клеток сравнительно с нормальными клетками. Кроме того, картина чувствительности или устойчивости с апоптозу, индуцированному TRAIL, коррелирует с таковыми, индуцированными антителом против DR5, указывая на то, что синовиальные клетки используют прежде всего DR5 для запуска TRAIL-апоптоза.
Как и в случае злокачественных клеток, чувствительность к апоптозу, индуцированному TRAIL или антителом против DR5, варьирует в синовиальных клетках при РА, хотя они и экспрессируют сходные уровни DR5. RA-512 и RA-707 характеризуются наибольшей чувствительностью, достигающей свыше 80% клеток, которые уничтожаются концентрациями TRAIL или антитела против DR5 ниже 20 нг/мг. RA-1014, RA-811, RA-716 и RA929 относятся к вариантам с промежуточной чувствительностью к TRAIL или TRA-8, при этом примерно 100% гибель клеток достигается в присутствии высоких концентраций (>50 нг/мл) TRAIL или TRA-8. В клетках RA-1016 и RA1021, хотя большая часть клеток (более 60%) и уничтожается низкой дозой TRAIL или TRA-8, наличие части клеток, выживающих при высоких концентрациях TRAIL или TRA-8, указывает на наличие субпопуляции клеток, устойчивых к TRAIL-опосредованному апоптозу. И наоборот, все клетки при OA намного менее чувствительны к TRAIL- и TRA-8-индуцированному апоптозу. Доля клеток не выше, чем 60%, уничтожается в OA52F и OA69F даже в присутствии высокой концентрации TRAIL или TRA-8. Клетки OA72M полностью устойчивы к TRAIL- или TRA-8-индуцированному апоптозу. SV40-трансформированные синовиальные клетки также чувствительны к TRAIL- или TRA-8-индуцированному апоптозу (данные не приведены). И наоборот, нетрансформированные фибробластные клетки, по-видимому, устойчивы к TRAIL и TRA-8.
Ранее было показано, что DR5 использует путь, зависимый от FADD/каспазы 8, для запуска апоптоза [44]. Для определения зависимости от каспазы DR5-опосредованного апоптоза в синовиальных клетках при РА, указанные РА клетки культивируют с TRAIL или антителом против DR5 в присутствии специфических ингибиторов каспазы. Из числа исследованных восьми ингибиторов каспазы было показано, что ингибиторы каспазы 6, 8 и 10 способны ингибировать апоптоз в синовиальных клетках при РА, индуцируемый и TRAIL, и DR5, как показано на фиг.9, что свидетельствует о том, что указанные три каспазы участвуют в DR5-опосредованном апоптозе.
10.3 TRA-8 или TRAIL индуцируют активацию NF-κb в синовиальных клетках при РА без повышения высвобождения МП
Имеется достаточно данных, поддерживающих концепцию о существовании тесных связей между сигнальной функцией апоптоза и сигнальной функцией пролиферации [45]. Было установлено, что DR5 способен активировать путь через NF-κb, дополнительно к участию в функции сигнального переноса через апоптоз и что активация NF-κb может участвовать в переносе антиапоптозного сигнала. Исходя из этого, проводят гель-шифт-анализ. Клетки стимулируют 50 нг/мл рекомбинантного растворимого TRAIL, Fas-лигандом в присутствии 1 мг/мл энхансера или 50 нг/мл TRA-8 в указанные временные точки. Готовят экстракты ядер и инкубируют их с зондом [32P]-меченной олигонуклеотидной цепи ДНК с двойной окраской. Полученные результаты анализируют при визуализации с использованием циклонного сепаратора для фосфора (TopCount NXT, Packard Instrument Company, CT). После инкубации РА клеток с TNF-α или TRAIL проводят активацию NF-κb по способу, зависимому от времени. Антитело TRA-8 способно сильно активировать NF-κb. И наоборот, Fas-лиганд не способен индуцировать активацию NF-κb, как показано на фиг.10а.
Таким образом, хотя TRAIL и антитело TRA-8 индуцируют мощную апоптозную реакцию в синовиальных клетках при РА, они также активируют NF-κb, при этом считается, что активация NF-κb вносит вклад в провоспалительную функцию TNF-α при РА. Так что вполне вероятно, что TRAIL подобно TNF-α может действовать как провоспалительный цитокин. Для выяснения того, имеется ли аналогичное биологическое следствие активации NF-κb, индуцированной TRAIL и TNF-α, определяют по методу ELISA продукцию МП. Синовиальные клетки культивируют в одной только среде или с добавкой 50 нг/мл интерлейкина 1b, 10 нг/мл TNF-α, 50 нг/мл TRAIL или 50 нг/мл TRA-8 в течение ночи. Уровень МП-1 и МП-3 в супернатантах культур определяют по методу ELISA.
Когда РА синовиальные клетки культивируют с провоспалительным цитокином TNF-α или IL-1b, продукция МП-1, 3 и 13 возрастает в сравнении со средой, взятой в качестве контроля, как видно на фиг.10b, c. И наоборот, добавление TRAIL или антитела DR5 не связано с повышенным высвобождением указанных МП.
Пример 11а. Отсутствие индукции токсичности для клеток печени
Для 24-часовых тестов на жизнеспособность клеток свежевыделенные нормальные гепатоциты человека приобретают от компании In Vitro Technology (Baltimore, MD). Гепатоциты культивируют в среде для гепатоцитов, содержащей 1 мкг/мл TRAIL или TRA-8. Для 6-часовых тестов на жизнеспособность клеток нормальные гепатоциты или раковые клетки печени, выделенные из свежих хирургических образцов, получают от компании UAB Tissue Procurement Center. Все реагенты, используемые для выделения человеческих гепатоцитов, включая буфер для перфузии гепатоцитов, среду для расщепления, промывочную среду и среду для прикрепления, получают от компании Gibco. Тканевые срезы расщепляют в среде для расщепления гепатоцитов при температуре 37°C при встряхивании (50 об/мин) в течение одного часа. Выделенные гепатоциты собирают путем центрифугирования с низкой скоростью (50g, 3 мин) и шесть раз промывают промывочной средой для гепатоцитов. Суспензию единичных клеток гепатоцитов культивируют в среде для прикрепления, содержащей 10% FCS, в лунки 96-луночного планшета (Martigel BD) в течение шести часов. Неприкрепленные гепатоциты удаляют при двойной промывке предварительно нагретой средой для прикрепления. Прикрепленные гепатоциты далее инкубируют с разными концентрациями растворимого TRAIL или FasL в присутствии сшивателя или TRA-8, или CH11 в течение 6 часов.
TRAIL содержит по меньшей мере два рецептора (DR4 и DR5), которые способны индуцировать апоптоз. TRA-8 используют для определения того, достаточно ли одного сшивания DR5 для индукции апоптоза в нормальных гепатоцитах. Экспрессию DR5 на уровне белка оценивают вначале в пяти нормальных тканях печени человека и в пяти раковых тканях печени путем иммуногистохимического анализа in situ с использованием TRA-8. Срезы из нормальных тканей печени характеризуются нормальной структурой и морфологией клеток при H&E окрашивании (фиг.11а, левые верхние панели) в отсутствие положительной реакции с TRA-8 для DR5 (фиг.11а, левые нижние панели). И наоборот, ткань карциномы клеток печени человека реагирует положительно с TRA-8, и указанная реакция согласуется с наличием DR5 в клеточной мембране и цитоплазме злокачественных клеток. Клеточная линия карциномы клеток печени человека HepG2 также положительно реагирует с DR5. Приведенные данные согласуются с результатами, полученными по пяти нормальным тканям печени человека, и лишь один вариант (аденома печени) злокачественных опухолей из пяти тканей человека был DR5-отрицательным. Приведенные данные согласуются также с результатами, полученными при проведении анализа по методу вестерн-блоттинга, как показано на фиг.5а, согласно которым, как и другие нормальные ткани, нормальная ткань печени человека не экспрессирует значительных количеств белка DR5. Кроме того, по данным вестерн-блот-анализа, в нормальных человеческих гепатоцитах при зондировании их с использованием TRA-8 не выявляются заметные количества DR5.
Анализ экспрессии DR5 на клеточной поверхности человеческих гепатоцитов методом проточной цитометрии показывает, что свежевыделенные нормальные гепатоциты не экспрессируют обнаруживаемых количеств DR5 на клеточной поверхности (фиг.11b, верхние левые панели). Он не обнаруживается ни в нормальных человеческих гепатоцитах, которые были подвергнуты криоконсервации, ни в случае их помещения в условия краткосрочного культивирования. И наоборот, свежевыделенные нормальные гепатоциты, так же как и клетки HepG2, экспрессируют DR5 на клеточной поверхности. С использованием Fas в качестве варианта сравнения было показано, что нормальные гепатоциты, клетки карциномы печени и клетки HepG2 - все экспрессируют уровни, эквивалентные для случая Fas (фиг.11b, нижние панели). Приведенные данные согласуются с результатами, полученными in situ при проведении иммуногистохимического анализа и анализа по методу вестерн-блоттинга, согласно которым DR5 на клеточной поверхности экспрессируется в высокой степени в раковых клетках печени, но не в нормальных гепатоцитах. Наличие мРНК для DR4, DR5, DcR1 и DcR2 в человеческих гепатоцитах, как показано при анализе в рамках ОТ-ПЦР[23], позволяет предположить, что человеческие гепатоциты могут экспрессировать очень низкие количества белка DR5, уровень которого ниже порога определения при использовании TRA-8.
Для выяснения, индуцирует ли TRA-8 токсичность для клеток печени, определяют чувствительность нормальных гепатоцитов человека к апоптозу, индуцированному TRA-8 и растворимым TRAIL плюс сшиватель. При культивировании нормальных гепатоцитов в присутствии высоких концентраций TRAIL наблюдается зависимое от времени снижение жизнеспособности клеток, по данным тестов ATPLite (фиг.12а) и MTT. TRAIL-опосредованная гибель клеток нормальных гепатоцитов отмечается уже через четыре часа после добавления TRAIL. В конце 24-часового культивирования более 80% гепатоцитов уничтожается под действием TRAIL. И наоборот, в течение того же периода времени TRA-8 не индуцирует значительной гибели нормальных гепатоцитов. Количество конденсированных ядер, окрашиваемых по Hoechst, представляющих собой характерный признак апоптоза, увеличивается в случае TRAIL-обработанных, но не TRA-8-обработанных гепатоцитов (фиг.12b). Число апоптозных гепатоцитов хорошо коррелирует со снижением жизнеспособности клеток при определении по методу ATPLite, что позволяет предположить, что TRAIL-индуцированная гибель гепатоцитов опосредована апоптозом. Указанное предположение подтверждается тем фактом, что Z-VAD способен ингибировать TRAIL-опосредованную токсичность для гепатоцитов. В связи с тем что циклогексимид является мощным энхансером апоптоза, исследуют эффект этого соединения на гепатоциты, обработанные TRAIL и TRA-8 соответственно. В ходе четырехчасового культивирования циклогексимид существенно повышает уровень гибели гепатоцитов, индуцированной TRAIL, при этом более 70% гепатоцитов уничтожаются TRAIL в присутствии циклогексимида (фиг.12с). Однако обработка циклогексимидом не позволяет повысить TRA-8-опосредованную гибель клеток в культуре гепатоцитов. Для сравнения характеристик апоптоза, индуцированного TRA-8, с таковыми в случае индукции TRAIL в отношении гепатоцитов, нормальные гепатоциты, а также злокачественные клетки инкубируют с варьирующими количествами растворимого TRAIL в сочетании со сшивателем или TRA-8. В ходе 6-часового культивирования TRAIL индуцирует умеренную апоптозную реакцию в нормальных гепатоцитах. Свыше 20% гепатоцитов уничтожается в присутствии 500 нг/мл TRAIL (фиг.12d, верхняя левая панель). Обработка TRA-8 нормальных гепатоцитов не приводит к сколько-нибудь значительной гибели клеток за тот же период времени. А в отличие от нормальных гепатоцитов первичные клетки карциномы печени (фиг.12d, верхняя средняя панель) и клетки HepG2 (фиг.12d, верхняя правая панель) демонстрируют высокую чувствительность к апоптозу, опосредованному TRAIL или TRA-8. Свыше 80% клеток карциномы печени и примерно 100% клеток HepG2 уничтожаются в ходе 8-часового периода культивирования. Приведенные результаты показывают, что нормальные гепатоциты полностью устойчивы к TRA-8-опосредованному апоптозу и намного менее чувствительны к TRAIL-опосредованному апоптозу, чем раковые клетки печени. С использованием Fas-лиганда и антитела против Fas (CH-11) показано, что отсутствует существенное различие в чувствительности к Fas-опосредованному апоптозу между нормальными гепатоцитами, клетками карциномы печени и клетками HepG2 (фиг.12d, нижние панели).
Сравнительный пример 11В. Индукция человеческим мембрансвязанным TRAIL гепатита in vivo
8-10-недельным самкам мышей B6 инъецируют внутривенно 109 БОЕ Ad/hTRAIL с равным числом Ad/Tet-on. Сразу после инокуляции аденовирусных векторов мышам скармливают разные концентрации тетрациклина с питьевой водой. Поражение печени выявляют по сывороточным уровням ACT с использованием диагностического набора на ACT (Sigma). Экспрессию TRAIL определяют анализом по методу нозерн-блоттинга.
Для определения того, индуцирует ли мембрансвязанная форма TRAIL повреждение печени in vivo, конструируют рекомбинантный аденовирусный вектор, кодирующий человеческий TRAIL полной длины (Ad/hTRAIL), экспрессия которого осуществляется под контролем индуцируемого тетрациклином промотора. Через двадцать четыре часа после внутривенной инокуляции B6 мышам Ad/hTRAIL оценивают индуцируемую тетрациклином экспрессию человеческого TRAIL зависимым от дозы образом с использованием анализа по методу нозерн-блоттинга (фиг.13а). Уровни экспрессии TRAIL коррелируют с повреждением печени, что следует из тетрациклинзависимого повышения сывороточных уровней трансаминаз, также зависимым от дозы образом (фиг.13b). Поскольку инокуляция аденовирусного вектора per se могла повысить чувствительность гепатоцитов к TRAIL-опосредованному апоптозу, выделяют гепатоциты от мышей, которым был инокулирован Ad/TRAIL, и тестируют TRAIL-опосредованную гибель клеток. Не отмечается существенного повышения уровня гибели клеток в инфицированных Ad/TRAIL гепатоцитах в сравнении с уровнем у контрольных мышей (фиг.13с, левая панель). Кроме того, у мышей, которым был инокулирован Ad/TRAIL, не отмечается усиление повреждения печени после внутривенной инъекции растворимого человеческого TRAIL. Таким образом, следует, что гепатит, индуцированный Ad/TRAIL, опосредован высокими уровнями экспрессии TRAIL в связанной с мембраной форме. Гистологический анализ срезов печени показывает, что повреждение гепатоцитов становится заметным уже через 24 часа после инокуляции вектора (фиг.13d) и длится в течение по меньшей мере 7 дней (фиг.13е). Указанные патологические изменения в печени также являются тетрациклинзависимыми и осуществляются зависимым от дозы образом. На ранней фазе в течение 24 часов после обработки, TRAIL-индуцированное повреждение печени характеризуется наличием участков некроза. На данной стадии не наблюдается инфильтрации воспалительных клеток, но имеется кровотечение. К 7 дню после инокуляции становятся заметными диффузные поражения печени с выраженным нарушением в организации ее долей, с сильной дегенерацией гепатоцитов, включающей разбросанные сгустки цитоплазмы и крупные прозрачные пространства, а также выраженный апоптоз и некроз. Характерной особенностью на данной стадии является обширная инфильтрация мононуклеарных клеток. Данное явление указывает на то, что человеческий TRAIL в его мембраносвязанной форме способен индуцировать повреждение печени in vivo. Несмотря на тенденцию человеческого TRAIL вызывать тяжелый гепатит у мышей, он не вызывает летальной реакции. И наоборот, у мышей, которым были инокулированы аналогичные контролируемые тетрациклином векторы, кодирующие Fas-лиганд, развивается молниеносный гепатит с массивным апоптозом и некрозом гепатоцитов, сопровождаемый тяжелой геморрагией и смертностью, происходящей зависимым от тетрациклина образом в течение 72 часов после инокуляции. Уровень смертности составляет 100% в течение 48 часов в подгруппах животных, которым вводили 3 мг/мл или более тетрациклина. И наоборот, все мыши, которые получали Ad/hTRAIL, независимо от дозы тетрациклина, были все еще живы по прошествии четырех недель после инокуляции. Из этого следует, что in vivo мембраносвязанная форма TRAIL является менее мощным индуктором гепатоцеллюлярного повреждения, чем Fas-лиганд. Предполагается также, что TRAIL может индуцировать повреждение печени через механизм, который отличается от механизма, лежащего в основе токсического действия Fas-лиганда.
Пример 12. Активированные Т- и В-клетки человека экспрессируют повышенные уровни DR5
Для определения, играет ли DR5 определенную роль в TRAIL-опосредованном апоптозе Т-клеток и В-клеток, изучалась экспрессия DR5 на поверхности покоящихся и активированных Т- и В-клеток с использованием TRA-8. Нестимулированные человеческие клетки МНПК не экспрессируют значительных количеств DR5 (фиг.14). К 48 часам после стимуляции с использованием анти-CD3 или Con-A экспрессия DR5 на поверхности клеток значительно повышается. Аналогично нестимулированные В-клетки экспрессируют очень низкие уровни DR5. Стимуляция с использованием анти-μ, но не ЛПС (LPS), приводит к повышению уровня экспрессии DR5 на поверхности клеток. Данный результат указывает на то, что активированные Т- и В-клетки экспрессируют повышенные уровни DR5 на поверхности клеток. Клетки окрашивают с использованием 20 мкг/мл TRA-8 и ФЭ противомышиного IgG1.
Пример 13. Активированные Т- и В-клетки становятся чувствительными к TRA-8-опосредованному апоптозу
Для определения, являются ли чувствительными активированные T- и В-клетки к TRA-8-опосредованному апоптозу, Т-клетки и В-клетки человеческих МНПК стимулируют анти-CD3 или анти-μ in vitro в течение 48 часов соответственно. Жизнеспособные клетки и пролиферирующие бластные клетки собирают центрифугированием в градиенте плотности и инкубируют с разными концентрациями TRA-8. Нестимулированные T- и B-клетки нечувствительны к TRA-8-опосредованному апоптозу (фиг.15). Совокупность стимулированных T-клеток и B-клеток демонстрирует умеренно повышенную чувствительность к TRA-8-опосредованному апоптозу, при этом 20% клеток уничтожается под действием TRA-8 после культивирования в течение ночи. Высокопролиферирующие бластные Т-клетки еще более чувствительны к TRA-8-опосредованному апоптозу. Более 70% бластных Т-клеток могут быть уничтожены под действием TRA-8. Бластные В-клетки также более чувствительны к TRA-8-опосредованному апоптозу в сравнении с другими клетками. Данные результаты указывают на то, что активированные Т- и В-клетки чувствительны к DR5-опосредованному апоптозу.
Пример 14. TRA-8 истощает уровень активированных Т-клеток у человека/мышей SCID
Для определения эффективности против Т-клеток in vivo TRA-8 мышам NOD/SCID инъецируют внутривенно 1·108 человеческих МНПК. В норме человеческие Т-клетки у мышей SCID быстро активируются в ответ на ксеногенную стимуляцию. SCID мышам, содержащим человеческие МНПК, внутрибрюшинно инъецируют 100 мкг TRA-8 или контрольный IgG1 с момента переноса и повторяют ежедневно в течение трех дней. Через пять дней после переноса мононуклеарные клетки выделяют из селезенки и окрашивают антителом против человеческого CD3, а популяцию лимфоцитов просеивают по методу проточной цитометрии и анализируют человеческие Т-клетки, положительные по CD3. Примерно 30% лимфоцитов селезенки представляют собой Т-клетки человека, что следует из результатов окрашивания антитела против человеческого CD3 у контрольных мышей. Однако лишь несколько человеческих Т-клеток (менее 3%) наблюдаются среди лимфоцитов селезенки у мышей, обработанных TRA-8 (фиг.16). Гистологический анализ in situ выявляет, что в селезенке контрольных мышей человеческие Т-клетки вновь распространяются и наблюдается лишь несколько апоптозных клеток, как следует из результатов окрашивания по методике TUNEL. И наоборот, заселение жизнеспособными человеческими Т-клетками не наблюдается в селезенке TRA-8-обработанных мышей, тогда как отмечается множество апоптозных клеток (фиг.17). Приведенные результаты показывают, что TRA-8 обладает in vivo активностью против Т-клеток и свидетельствуют о возможности использования антител по настоящему изобретению для лечения болезни РТПХ.
Пример 15. Противораковая терапевтическая активность TRA-8
15.1 Экспрессия DR5 и его функционирование в тканях и клеточных линиях и тканях злокачественной опухоли человека
i) Экспрессия DR5 в тканях злокачественной опухоли человека при окрашивании in situ TRA-8
Для определения того, характеризуются ли злокачественные клетки и ткани дифференциальной экспрессией более высоких уровней DR5, проводят окрашивание панели тканей злокачественной опухоли человека, включая 20 вариантов рака молочной железы, 6 вариантов рака яичника, 5 вариантов рака толстой кишки и 5 вариантов рака предстательной железы с использованием TRA-8 для иммуногистохимического анализа. Большая часть указанных тканей злокачественных опухолей экспрессирует выявляемые количества DR5. Уровень экспрессии DR5 в указанных тканях злокачественных опухолей варьирует. В целом ткани злокачественных опухолей экспрессируют более высокие уровни DR5, чем другие ткани. Кроме того, экспрессия DR5 явно не коррелирует с мутацией p53.
ii) Экспрессия DR5 и его функционирование в клеточных линиях злокачественной опухоли человека (таблица 2)
Девять клеточных линий рака молочной железы человека, три линии рака яичника, три линии рака толстой кишки и три линии рака предстательной железы исследуют с точки зрения выявления экспрессии DR5 на поверхности клеток и их чувствительности к TRA-8-индуцированному апоптозу in vitro. В 7 из 9 линий рака молочной железы, в 3 из 3 линий рака яичника, в 3 из 3 линий рака толстого кишечника и в 3 из 3 линий рака предстательной железы экспрессируются различные варьирующие уровни DR5 на клеточной поверхности. Из 9 линий рака молочной железы три являются очень чувствительными, три отличаются промежуточной чувствительностью и три - устойчивы к TRA-8-опосредованному апоптозу. Все три линии рака яичников очень чувствительны. Одна из трех линий рака толстого кишечника очень чувствительна, тогда как две другие характеризуются промежуточной чувствительностью. Две из трех линий рака предстательной железы имеют промежуточную чувствительность, и одна является устойчивой.
в злокачественных клетках человека
Примечание:
1Определяют методом проточной цитометрии, клетки окрашивают 20 мкг/мл TRA-8 и сравнивают с контрольным антителом.
2Определяют по тесту ATPLite.
iii) Объединенная цитотоксичность TRA-8 в сочетании с адриамицином
В нескольких клеточных линиях рака молочной железы изучают эффект адриамицина на TRA-8-индуцированный апоптоз. Высокие дозы адриамицина демонстрируют аддитивный эффект. Однако в некоторых TRA-8-резистентных линиях низкие дозы адриамицина синергически повышают TRA-8-индуцированный апоптоз.
iv) In vitro и in vivo связывающая активность TRA-8 в отношении клеток злокачественной опухоли человека
Используют меченный радиоактивной меткой TRA-8. Изучают связывающую активность TRA-8 с раковой линией молочной железы in vitro и in vivo у мышей SCID с имплантированной опухолью. Связывающая активность со злокачественными клетками in vitro была оценена значением Kd, равным 3 нМ, которое не изменилось в сравнении с предыдущими оценками методом ELISA и по меньшей мере в 50 раз превышает уровень, показанный для растворимого TRAIL. In vivo TRA-8 локализуется в имплантированных опухолевых тканях.
15.2 Лечение хронического лимфолейкоза у мышей NOD/SCID с использованием TRA-8
Хронический лимфолейкоз (CLL, ХЛЛ) представляет собой распространенную форму злокачественного заболевания, связанного с В-клетками. Большая часть злокачественных В-клеток при ХЛЛ имеет зрелый фенотип и характеризуется устойчивостью ко многим апоптозным стимулам. Была исследована экспрессия DR5 и его функционирование в В-клетках у пяти пациентов с ХЛЛ. Все пациенты характеризуются высоким числом периферических В-клеток, что составляет более 95% CD19+B клеток в МНПК. В сравнении с нормальными первичными В-клетками В-клетки при ХЛЛ у всех пациентов имеют повышенные уровни DR5 на поверхности клеток и характеризуются большей чувствительностью TRA-8-индуцированному апоптозу in vitro. Интересно, что В-клетки при ХЛЛ также чувствительны к цитотоксичности, индуцированной бисиндолмалеимидом VIII (BisVIII). После комбинированного воздействия с использованием TRA-8 и BisVIII около 50% В-клеток при ХЛЛ уничтожаются, тогда как нормальные В-клетки остаются незатронутыми (фиг.18). Перенос В-клеток от пациентов с ХЛЛ к мышам NOD/SCID приводит к появлению примерно до 25%-30% CD19+B клеток в селезенке реципиентных мышей на пятый день после переноса. Однако три дозы TRA-8 по 100 мкг полностью элюминируют В-клетки с патологией ХЛЛ от четырех из пяти пациентов в селезенке реципиентных мышей SCID. Таким образом, TRA-8 один или в сочетании с другими веществами является активным в качестве терапевтического средства для применения при хроническом лимфолейкозе.
Пример 16. Клонирование кДНК
(1) Определение N-концевой аминокислотной последовательности тяжелой и легкой цепей TRA-8
Для получения кДНК тяжелой и легкой цепей TRA-8, N-концевые аминокислотные последовательности тяжелой и легкой цепи TRA-8 и последовательность клонированных генов TRA-8 определяют с помощью известных методик.
Десять мкг раствора, содержащего антитело TRA-8 против человеческого DR5, подвергают электрофорезу в ДСН-полиакриламидном геле (ДСН-ПААГ) с использованием геля в концентрации 12% (вес/об.) при постоянном напряжении 100 вольт в течение 120 минут. После проведения электрофореза гель погружают на 5 минут в трансферный буфер, содержащий 25 мМ Трис-HCl (pH 9,5), 20% метанола, 0,02 об./об.% ДСН. По истечении указанного периода времени белок, содержащийся в геле, наносят на поливинилидендифторидную мембрану (ПВДФ мембрана, с размером пор 45 мкм, Millipore, Япония), предварительно вымоченную в трансферном буфере, с использованием аппарата для блоттинга (KS-8451; Marysol) в условиях подведения постоянного напряжения 10 вольт при 4°С в течение 14 часов.
По прошествии указанного периода времени ПВДФ мембрану промывают промывочным буфером, содержащим 25 мМ NaCl, 10 мМ натрий-боратного буфера (pH 8,0), затем окрашивают окрашивающим раствором (50 об./об.% метанола, 20 об./об.% уксусной кислоты и 0,05% (вес/об.) Кумасси бриллиантового синего) в течение 5 минут для локализации белковых полос. Затем ПВДФ мембрану обесцвечивают 90 об./об.% водным метанолом и полосы, соответствующие тяжелой цепи, т.е. полосы с меньшей подвижностью, и соответствующие легкой цепи, т.е. полосы с большей подвижностью, предварительно локализованные на ПВДФ мембране, вырезают и промывают деионизированной водой.
N-концевую аминокислотную последовательность тяжелой и легкой цепи определяют по методу Эдмана в автоматизированном режиме (Edman, P., et al. (1967), Eur. J. Biochem., 1, 80) с использованием секвенатора белка в газовой фазе (PPSQ-10; Shimadzu Seisakusyo, K.K.).
Последовательность N-концевой аминокислоты в полосе, соответствующей тяжелой цепи, была определена в указанном ниже виде:
Glu-Val-Met-Leu-Val-Glu-Ser-Gly-Gly-Gly-Leu-Val-Lys-Pro-Gly-Gly-Ser-Leu-Lys-Leu (SEQ ID No. 4 в перечне последовательностей)
и в полосе, соответствующей легкой цепи, определена как:
Asp-Ile-Val-Met-Thr-Gln-Ser-His-Lys-Phe-Met-Ser-Thr-Ser-Val-Gly-Asp-Arg-Val-Ser (SEQ ID No. 5 в перечне последовательностей).
Сравнение указанных аминокислотных последовательностей с аминокислотной последовательностью в базе данных антител, полученных Кабат с соавт. (Kabat E.A., et al. (1991), in "Sequences of Proteins of Immunological Interest Vol. II," U.S. Department of Health and Human Services), показывают, что тяжелая цепь (γ1 цепь) и легкая цепь (k цепь) TRA-8 принадлежат к подтипам 3d и 1 соответственно.
(2) Клонирование кДНК
На основе указанных данных синтезируют олигонуклеотидные праймеры, которые, как ожидается, будут гибридизоваться с частями 5'-нетранслируемых участков и концами 3'-транслируемых участков генов, принадлежащих к указанным мышиным подтипам. Затем кДНК, кодирующие тяжелую и легкую цепь TRA-8, клонируют с использованием приведенного ниже сочетания обратной транскрипции и ПЦР (ОТ-ПЦР).
а) Матрицы
Суммарную РНК из гибридомы TRA-8 (ATCC No. PTA-1428) экстрагируют с использованием реагента TRIzol (GIBCO BRL). В качестве матрицы для ПЦР-реакции используют кДНК, которую получают с использованием набора для синтеза одноцепочечной кДНК (First-Strand kit) (American Pharmacia Biotech) в соответствии с инструкцией, прилагаемой к набору.
b) Праймеры для ПЦР
Для ПЦР синтезируют указанные ниже олигонуклеотидные праймеры:
5′-cagcactgaa cacggacccc-3′ (H5NCS1: SEQ ID No.6 в перечне последовательностей);
5′-aaaggtaatt tattgagaag-3′ (H5NCS2: SEQ ID No.7 в перечне последовательностей);
5′-cctcaccatg aacttcgggc-3′ (H5SS1: SEQ ID No.8 в перечне последовательностей);
5′-ctgttgtatg cacatgagac-3′ (H5SS2: SEQ ID No.9 в перечне последовательностей);
5′-gaagtgatgc tggtggagtc-3′ (H5CS1: SEQ ID No.10 в перечне последовательностей);
5′-agtgtgaagt gatgctggtg-3′ (H5CS2: SEQ ID No.11 в перечне последовательностей);
5′-tttaccagga gagtgggaga g-3′ (H3CR: SEQ ID No.12 в перечне последовательностей);
5′-tgcagagaca gtgaccagag-3′ (H3VR: SEQ ID No.13 в перечне последовательностей);
5′-tgttcaggac cagcatgggc-3′ (L5NCS1: SEQ ID No.14 в перечне последовательностей);
5′-aagacatttt ggattctaac-3′ (L5NCS2: SEQ ID No.15 в перечне последовательностей);
5′-tatcatgaag tctttgtatg-3′ (L5SS1: SEQ ID No.16 в перечне последовательностей;
5′-gatggagaca cattctcagg-3′ (L5SS2: SEQ ID No.17 в перечне последовательностей);
5′-gacattgtga tgacccagtc-3′ (L5CS: SEQ ID No.18 в перечне последовательностей);
5′-ttaacactca ttcctgttga-3′ (L3CR: SEQ ID No.19 в перечне последовательностей) и
5′-gactgggtca tcacaatgtc-3′ (LCSR: SEQ ID No.20 в перечне последовательностей).
Если особо не оговорено иное, все олигонуклеотиды, приведенные в указанных примерах, синтезируются с использованием наборов Pharmacia Biotech. Все олигонуклеотиды хранят при температуре -20°С после растворения в дистиллированной воде.
с) Реакция ПЦР
Состав раствора для проведения реакции ПЦР следующий:
матричная кДНК, 5 мкл в 33 мкл реакционной смеси;
праймер DR5р1, 10 пмоль;
праймер DR5р2, 10 пмоль;
10× концентрированный буфер для ПЦР (прилагаемый к набору), 10 мкл;
смесь дНТФ (каждый по 2,5 мМ), 4 мкл; и
Taq-полимераза (Promega), 5 единиц.
К раствору добавляют стерильную дистиллированную воду, доводя до объема 100 мкл. Если особо не оговорено иное, используемые дНТФ представляют собой эквимолярную смесь дАТФ, дЦТФ, дГТВ и дТТФ (по 2,5 мМ каждый).
Реакцию ПЦР проводят следующим образом. Вначале раствор нагревают до 94°С в течение 2 минут, после чего проводят цикл, включающий нагревание при 94°С в течение 30 секунд, нагревание при 52°С в течение 1 минуты и нагревание при 72°С в течение 3 минут и повторяют указанную процедуру в течение 40 раз. После завершения указанной процедуры реакционный раствор нагревают при 72°С в течение 10 минут.
Полученный таким образом фрагмент амплифицированной ДНК разделяют электрофорезом в 1% геле агарозы, содержащей 0,25 мкг/мл бромида этидия. Полосы, в которых показано содержание желательных фрагментов ДНК, вырезают лезвием и из них выделяют ДНК с использованием набора Gene Clean (BIO101). Фрагмент ДНК клонируют с использованием вектора pGEM-T Easy (Promega). Процедуру проводят следующим образом.
Фрагмент ДНК, выделенный из реакционного раствора для проведения ПЦР вместе с 50 нг вектора pGEM-T Easy (прилагаемого к набору), смешивают с 1 мкл 10× буфера для лигазной реакции (6 мМ Tris-HCl (pH 7,5), 6 мМ хлорид магния, 5 мМ хлорид натрия, 7 мМ β-меркаптоэтанол, 0,1 мМ АТФ, 2 мМ ДТТ, 1 мМ спермидин и 0,1 мг/мл бычий сывороточный альбумин), к которому добавляют 4 единицы ДНК-лигазы Т4 (1 мкл). Общий объем смеси доводят до 10 мкл стерильной деионизированной водой и полученный лигазный раствор инкубируют при 14°С в течение 15 часов. По окончании указанного периода времени добавляют 2 мкл лигазного раствора к 50 мкл компетентного штамма Е. coli JM 109 (прилагаемого к набору и приведенного в компетентное состояние в соответствии с руководством для пользователя), далее добавляют 2 мкл 0,5 М β-меркаптоэтанола и полученную смесь выдерживают на льду в течение 30 минут, затем при 42°С в течение 30 секунд и снова на льду в течение 5 минут. Далее к культуре добавляют 500 мкл среды, содержащей 2 об./об.% триптона, 0,5% (вес/об.) дрожжевого экстракта, 0,05% (вес/об.) хлорида натрия, 2,5 мМ хлорида калия, 1 мМ хлорида магния и 20 мМ глюкозы (далее обозначаемой как среда «SOC»), и смесь инкубируют в течение 1 часа при 37°С при встряхивании. По прошествии указанного периода времени культуру распределяют по чашке с L-бульонным агаром (1 об./об.% триптона, 0,5% (вес/об.) дрожжевого экстракта, 0,5% (вес/об.) хлорида натрия, 0,1% (вес/об.) глюкозы и 0,6% (вес/об.) бактоагара (Difco)), содержащим 100 мкг/мл ампициллина. Появившиеся на чашке колонии, устойчивые к ампициллину, отсортировывают и соскребают с помощью платиновой петли и далее культивируют в L-бульонной среде, содержащей 100 мкг/мл ампициллина при 37°С в течение ночи при встряхивании со скоростью 200 об/мин. После инкубации клетки собирают центрифугированием, выделяя из них плазмидную ДНК щелочным методом. Полученную плазмиду обозначают как pH62 для тяжелой цепи TRA-8 или pL28 для легкой цепи TRA-8. Трансформантные штаммы E. coli, несущие указанные плазмиды, обозначенные как E. coli JM109/pH62 и E. coli JM109/pL28, были депонированы в Международном депозитарии запатентованных организмов при Национальном Институте современной промышленной науки и технологии в Японии (International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology, 1-1, Higashi 1 chome Tsukuba-shi, Ibaraki-ken, 305-5466) 20 апреля 2001 года в соответствии с Будапештским договором по депонированию микроорганизмов с присвоением им номеров FERM-BP-7560 и FERM-BP-7561 соответственно. Нуклеотидные последовательности указанных ДНК, кодирующих тяжелую и легкую цепи TRA-8, подтверждают дидезоксиметодом (Sanger, F.S., et al. (1977), Proc. Natl. Acad. Sci. USA, 74:5463-5467) с использованием анализатора ДНК 3700 (ABI PRISM; Perkin Elmer Applied Biosystems, Japan).
Нуклеотидные последовательности тяжелой и легкой цепей TRA-8 приведены в перечне последовательностей как SEQ ID No. 21 и SEQ ID No. 22 соответственно. Аминокислотные последовательности тяжелой и легкой цепей TRA-8 приведены в перечне последовательностей как SEQ ID No. 23 и SEQ ID No. 24 соответственно. N-концевые аминокислотные последовательности тяжелой и легкой цепей TRA-8 устанавливают в рамках указанных методов. Далее при сравнении полученных аминокислотных последовательностей для тяжелой и легкой цепей с аминокислотными последовательностями антител, приведенных в базе данных, установлено, что в случае тяжелой цепи нуклеотидные последовательности No. 58-414 в SEQ ID No. 21 представляют вариабельный участок, тогда как нуклеотидные последовательности на участке от нуклеотида No. 415-1392 в SEQ ID No. 21 представляет константный участок. В легкой цепи нуклеотидная последовательность на участке от нуклеотида No. 64-387 в SEQ ID No. 22 представляет вариабельный участок, а нуклеотидная последовательность на участке от нуклеотида No. 388-702 в SEQ ID No. 22 представляет константный участок. Расположение и последовательность КДУ также были выявлены при определении уровня гомологичности путем сравнения с вариантами в базе данных. Аминокислотные последовательности КДУ1, КДУ2 и КДУ3 в тяжелой цепи TRA-8 показаны в виде SEQ ID No. 25, No. 26, No. 27, соответственно. Аминокислотные последовательности КДУ1, КДУ2 и КДУ3 в легкой цепи TRA-8 показаны в виде SEQ ID No. 28, No. 29 и No. 30, соответственно.
Пример 17. Конструирование гуманизированного варианта антитела TRA-8
(1) Молекулярное моделирование вариабельного участка TRA-8
Молекулярное моделирование вариабельного участка TRA-8 проводят по методу, известному как моделирование гомологии (Methods in Enzymology, 203, 121-153 (1991)). Первичные последовательности вариабельных участков человеческого иммуноглобулина, зарегистрированные в базе данных по белковым молекулам (Nuc. Acid. Res. 28, 235-242 (2000)), для которых имеются данные по трехмерной структуре, полученные по методу рентгеновской кристаллографии, сравнивают с определенными выше участками рамки для считывания для TRA-8. В результате были выбраны последовательности 1NCD и 1HIL как имеющие наивысшую степень гомологии последовательностей с участками рамки для считывания для легкой и тяжелой цепи TRA-8 соответственно. Трехмерную структуру участков рамки для считывания устанавливают при объединении координат 1NCD и 1HIL, которые соответствуют легкой и тяжелой цепи TRA-8, с получением «модели рамки для считывания». С использованием классификации, введенной Клотиа с соавт. (Chothia et al.), классифицируют следующим образом участки КДУ для TRA-8: CDRL1, CDRL2, CDRH1 и CDRH2, принадлежащие к каноническим классам 2, 1, 1, 3соответственно, тогда как CDRL3 не принадлежит к какому-либо конкретному каноническому классу. Петли КДУ для CDRL1, CDRL2, CDRH1 и CDRH2 приводят в соответствие с конформациями, присущими определенным каноническим классам, и интегрируют с получением модели рамки для считывания. CDRL3 соответствует конформации кластера 8А, по классификации Торнтона с соавт. (Thornton, et al., J. Mol. Biol., 263, 800-815 (1996)) и CDRH3 относят к классу k(8)С по правилу Н3 (FEBS letter 455, 188-197 (1999)). Затем репрезентативные конформации для CDRL3 и CDRH3 интегрируют с получением модели рамки для считывания.
И наконец, проводят расчеты энергетических показателей для удаления неблагоприятных вариантов межатомных контактов, с тем чтобы получить вероятную молекулярную модель вариабельного участка TRA-8 с точки зрения энергетических параметров. Указанную выше процедуру проводят с использованием коммерчески доступной системы для молекулярного моделирования ABM (Oxford Molecular Limited, Inc.). В рамках полученной молекулярной модели точность структуры оценивается далее с использованием программного обеспечения PROCHECK (J. Appl. Cryst. (1993), 26, 283-291).
(2) Создание аминокислотных последовательностей для гуманизированного TRA-8
Конструирование гуманизированных антител TRA-8 проводят по методу, известному как пересадка КДУ (Proc. Natl. Acad. Sci. USA, 86, 10029-10033 (1989)). Акцепторное антитело выбирают на основе аминокислотной гомологии в участке рамки для считывания. Последовательности участка рамки для считывания в TRA-8 сравнивают со всеми последовательностями человеческих рамок для считывания, имеющимися в базе данных Кабата (Kabat) по аминокислотным последовательностям антител (Nuc. Acid. Res. 29, 205-206 (2001)). В результате было выбрано антитело mAB58'CL в качестве акцептора благодаря его наивысшей гомологии по последовательности, составляющей 80% для участка рамки для считывания. Аминокислотные остатки в участке рамки для считывания в mAB58'CL сравнивают с таковыми для TRA-8 и идентифицируют положения, в которых используются разные аминокислоты. Расположение таких остатков анализируют с использованием разработанной выше трехмерной модели TRA-8 и донорные остатки, которые должны быть пересажены на акцептор, выбирают по критериям, данным Квином с соавт. (Queen et al., Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)). Последовательности гуманизированного TRA-8 конструируют, как будет описано в следующем примере, путем переноса нескольких донорных остатков на акцепторное антитело mAB58'CL.
Пример 18. Конструирование вектора экспрессии для тяжелой цепи гуманизированного антитела
(1) Конструирование плазмиды, содержащей ДНК для вариабельного участка гуманизированной тяжелой цепи TRA-8
Для того чтобы определить активность гуманизированного TRA-8, конструируют плазмиду, содержащую гуманизированную тяжелую цепь TRA-8, указанным ниже способом. Однако следует понимать, что гуманизация TRA-8 не ограничивается приведенными примерами.
Как показано в SEQ ID No. 31 в перечне последовательностей, гуманизация аминокислотных последовательностей тяжелой цепи мышиного антитела TRA-8 против человеческого DR5 включает замещение 13-ного аминокислотного остатка (лизина), 19-ного аминокислотного остатка (лизина). 40-ного аминокислотного остатка (треонина), 42-ного аминокислотного остатка (глютаминовой кислоты), 44-ного аминокислотного остатка (аргинина), 84-ного аминокислотного остатка (серина), 88-ного аминокислотного остатка (серина), 93-ного аминокислотного остатка (метионина), 114-ного аминокислотного остатка (треонина), 115-ного аминокислотного остатка (лейцина) глютамином, аргинином, аланином, глицином, глицином, аспарагином, аланином, валином, лейцином и валином соответственно.
Плазмиду, содержащую ДНК, которая кодирует тяжелую цепь вариабельного участка TRA-8 (SEQ ID No. 31 в перечне последовательностей), конструируют следующим образом.
Используют ПЦР для конструирования приведенных ниже последовательностей ДНК, каждая из которых включает указанные выше:
Синтезируют указанные ниже 12 олигонуклеотидов:
5′-ttggataagc ttggcttgac ctcaccatgg gatggagctg tatcatcctc ttcttggtag caacagctac aggtgtccac-3′ (А; SEQ ID No. 32);
5′-tctgaagtaa tgctggtgga gtctggggga ggcttagtac agcctggagg gtccctgaga ctctcctgtg cagcctctgg-3′ (В; SEQ ID No. 33);
5′-attcactttc agtagttatg taatgtcttg ggttcggcag gcaccaggga agggtctgga gtgggttgca accattagta-3′ (С; SEQ ID No. 34);
5′-gtggtggtag ttacacctac tatccagaca gtgtgaaggg ccgattcacc atctccagag acaatgccaa gaacaccctg-3′ (D; SEQ ID No. 35);
5′-tatctgcaaa tgaacagtct gagagcagag gacacggctg tttattactg tgcaagaagg ggtgactcta tgattacgac-3′ (E; SEQ ID No. 36);
5′-ggactactgg ggccaaggga ccctggtcac agtctcctca gcctc cacc aagggcccat cggtc-3′ (F; SEQ ID No. 37);
5′-ctaccaagaa gaggatgata cagctccatc ccatggtgag gtcaagccaa gcttatccaa-3′ (G; SEQ ID No. 38);
5′-tctcagggac cctccaggct gtactaagcc tcccccagac tccaccagca ttacttcaga gtggacacct gtagctgttg-3′ (H; SEQ ID No. 39);
5′-tccagaccct tccctggtgc ctgccgaacc caagacatta cataactact gaaagtgaat ccagaggctg cacaggagag-3′ (I; SEQ ID No. 40);
5′-ctctggagat ggtgaatcgg cccttcacac tgtctggata gtaggtgtaa ctaccaccac tactaatggt tgcaacccac-3′ (J; SEQ ID No. 41);
5′-ccttcttgca cagtaataaa cagccgtgtc ctctgctctc agactgttca tttgcagata cagggtgttc ttggcattgt-3′ (K; SEQ ID No. 42); и
5′-gaccgatggg cccttggtgg aggctgagga gactgtgacc agggtccctt ggccccagta gtccgtcgta atcatagagt cacc-3′ (L; SEQ ID No. 43);
Приведенные ниже праймеры для ПЦР синтезируют по описанной выше методике:
5′-ttggataagc ttggcttgac-3′ (Р1; SEQ ID No. 44); и
5′-gaccgatggg cccttggtgg a-3′ (Р2; SEQ ID No. 45).
Синтез ДНК, кодирующей полипептидную цепь, включающую сигнальную последовательность секреции, гуманизированную тяжелую цепь вариабельного участка TRA-8 и 8 аминокислотных остатков на N-конце IgG-CH1 участка, проводят с использованием сочетания соответствующих ПЦР.
Фрагмент ДНК получают следующим образом:
Состав раствора для проведения ПЦР реакции:
олигонуклеотид А, 10 пмоль;
олигонуклеотид B, 10 пмоль;
олигонуклеотид C, 10 пмоль;
олигонуклеотид D, 10 пмоль;
олигонуклеотид E, 10 пмоль;
олигонуклеотид F, 10 пмоль;
олигонуклеотид G, 10 пмоль;
олигонуклеотид H, 10 пмоль;
олигонуклеотид I, 10 пмоль;
олигонуклеотид J, 10 пмоль;
олигонуклеотид K, 10 пмоль;
олигонуклеотид L, 10 пмоль;
олигонуклеотидный праймер Р1, 2 мкМ;
олигонуклеотидный праймер Р2, 2 мкМ;
10× Pyrobest буфер II, 10 мкл;
смесь дНТФ, 8 мкл;
ДНК-полимераза Pyrobest, 0,5 мкл; и
и повторная дистиллированная вода, вносимая до конечного объема 50 мкл.
Реакцию ПЦР проводят следующим образом. Вначале раствор нагревают при 94°С в течение 5 минут, после чего проводят цикл, включающий нагревание до 98°С в течение 10 секунд, нагревание при 55°С в течение 30 секунд и нагревание при 72°С в течение 1 минуты с повторением указанного цикла 7 раз. После завершения указанной процедуры реакционный раствор нагревают при температуре 72°С в течение 15 минут.
Добавляют равный объем фенол-хлороформной смеси (50 об./об.% фенола, насыщенного водой, 48 об./об.% хлороформа, 2 об./об.% изоамилового спирта) к 200 мкл каждого из продуктов ПЦР и энергично перемешивают в течение 1 минуты. Затем смесь центрифугируют при 10000g и отобранный водный слой смешивают с равным объемом хлороформ-изоамилового спирта (96 об./об.% хлороформа и 4 об./об.% изоамилового спирта), снова энергично центрифугируют при 10000g и отбирают водный слой. Серия стадий, приведенная в данном абзаце, рассматривается как «фенольная экстракция».
Затем проводят осаждение этанолом отобранного водного слоя. В контексте настоящего описания термин «осаждение этанолом» включает добавление при перемешивании одной десятой объема 3М ацетата натрия (pH 5,2) и 2,5 объемов 100% этанола к обрабатываемому раствору и замораживание смеси сухим льдом. Полученную смесь затем центрифугируют при 10000g с получением осадка ДНК.
После проведения фенольной экстракции и осаждения этанолом полученный осадок ДНК сушат в вакууме, растворяют в минимальном количестве повторно дистиллированной воды и разделяют электрофорезом в 3% геле агарозы. После проведения электрофореза гель окрашивают водным раствором бромида этидия в концентрации 1 мкг/мл для обнаружения полос ДНК в УФ свете. Полосу ДНК, соответствующей ДНК гуманизированного TRA-8, вырезают лезвием, элюируя из геля с использованием набора Geneclean Spin (BIO 101, CA, USA). После фенольной экстракции элюированную ДНК концентрируют путем центрифугирования при 7500g и затем проводят осаждение этанолом и, наконец, растворяют в 5 мкл дистиллированной воды.
Полученную экстрагированную ДНК клонируют с использованием вектора pGEM-T Easy (Promega) следующим образом:
Фрагмент ДНК, выделенный из реакционной смеси для ПЦР, 5 мкл;
10× буфер для Taq-полимеразы, 1 мкл;
смесь дНТФ, 1 мкл;
Taq-полимераза (5 единиц/мл), 1 мкл; и
добавляют повторно дистиллированную воду до конечного объема 10 мкл.
После того как указанный выше раствор прореагирует при 70°С в течение 30 минут, лигируют каждый раствор ДНК и вектор pGEM-T Easy с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2,0) (Takara Shuzo Co., Ltd.) по инструкции от производителя.
После 4 часов инкубации при 15°C 2 мкл инкубированного реакционного раствора смешивают со 100 мкл компетентного штамма Е. coli JM109 с клеточной плотностью 1-2·109 клеток/мл (Takara Shuzo Co., Ltd.) и смесь выдерживают на льду в течение 30 минут, затем при 42°С в течение 30 секунд, и снова на льду в течение 1 минуты. К смеси добавляют 500 мкл среды SOC (2 об./об.% триптона, 0,5% (вес/об.) дрожжевого экстракта, 0,05% (вес/об.) хлорида натрия, 2,5 мМ (вес/об.) хлорида калия, 1 мМ хлорида магния и 20 мМ глюкозы) и все инкубируют еще в течение одного часа при встряхивании. Затем выделяют трансформированные штаммы и получают плазмидную ДНК из штаммов, как описано в руководстве («Molecular Cloning A Laboratory Manual»). Нуклеотидные последовательности указанных ДНК, которые кодируют гуманизированную тяжелую цепь TRA-8, подтверждают дидезоксиметодом (Sanger, F.S., et al. (1977), Proc. Natl. Acad. Sci. USA, 74:5463-5467) с использованием анализатора ДНК 3700 (ABI PRISM; Perkin Elmer Applied Biosystems, Japan).
Полученную плазмиду обозначают как pHB14 (плазмида, несущая кДНК, кодирующую гуманизированную тяжелую цепь TRA-8). Трансформированный штамм E. coli, несущий указанную плазмиду, обозначенный как E. coli JM109/pHB14, депонируют в Международном депозитарии запатентованных организмов при Национальном Институте современной промышленной науки и технологии Японии (International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology, 1-1, Higashi 1 chome Tsukuba-shi, Ibaraki-ken, 305-5466) 20 апреля 2001 года в соответствии с Будапештским договором по депонированию микроорганизмов под номером FERM BP-7556.
(2) Конструирование плазмид экспрессии, содержащих вариабельный участок ДНК для гуманизированной тяжелой цепи TRA-8
Рекомбинантный вектор экспрессии для клеток животных конструируют при вставке ДНК, кодирующей гуманизированную тяжелую цепь TRA-8 (клонированной как указано выше) указанным ниже способом.
Один мкг плазмиды pSRHHH3 (заявка на европейский патент EP 0909816 A1), содержащей вариабельный участок для гуманизированной тяжелой цепи против Fas моноклонального антитела HFE7A и константный участок геномной ДНК для IgG1, а также вектор экспрессии для клеток млекопитающих, расщепляют ферментами рестрикции HindIII и ApaI и разделяют электрофорезом в 3% агарозном геле. После электрофореза гель окрашивают водным раствором бромида этидия с концентрацией 1 мкг/мл и выявляют ДНК в УФ свете. Полосы векторной ДНК, содержащей константный участок геномной ДНК для человеческого IgG1 без вариабельного участка для гуманизированной тяжелой цепи HFE7A, вырезают лезвием и элюируют из геля с использованием набора Geneclean Spin (BIO 101, CA, USA).
Затем после фенольной экстракции элюированную ДНК концентрируют центрифугированием при 7500g с последующим осаждением этанолом и в итоге растворяют в 5 мл дистиллированной воды, после чего подвергают дефосфорилированию с использованием CIP. Полученную расщепленную дефосфорилированную плазмиду (100 нг) лигируют с 1 мкг фрагмента ДНК pHB14, содержащим ДНК, кодирующую гуманизированную тяжелую цепь вариабельного участка TRA-8, который уже был расщеплен HindIII и ApaI, с использованием набора для лигирования ДНК (Ligation Kit Version 2,0) (Takara Shuzo Co., Ltd.). Далее указанную лигирующую смесь используют для трансформации E. coli JM109, который затем высевают на чашки с агаром LB, содержащим 50 мкг/мл ампициллина.
Трансформанты, полученные указанным способом, культивируют в 2 мл жидкой среды LB, содержащей 50 мкг/мл ампициллина, при 37°С в течение ночи и затем плазмидную ДНК экстрагируют из полученной культуры по методу, включающему использование щелочи и ДСН.
Экстрагированную плазмидную ДНК расщепляют HindIII и ApaI и подвергают электрофорезу в 3% (вес/об.) агарозном геле, подтверждая наличие или отсутствие вставки ДНК, кодирующей гуманизированную тяжелую цепь вариабельного участка TRA-8. Наличие вставки и ориентация желательного фрагмента ДНК в векторе подтверждается секвенированием ДНК с использованием анализатора генной последовательности (ABI Prism 3700 DNA Analyzer; Applied Biosystems). Полученную плазмиду экспрессии, включающую кДНК, которая кодирует гуманизированную тяжелую цепь TRA-8, обозначают как pHB14-1.
Пример 19. Конструирование вектора экспрессии для гуманизированной легкой цепи антитела
(1) Конструирование векторов для гуманизированных вариантов легких цепей антитела TRA-8
Как показано в SEQ ID No. 46 в перечне последовательностей, в аминокислотной последовательности гуманизированной легкой цепи мышиного антитела TRA-8 против человеческого DR5 8-й аминокислотный остаток (гистидин), 9-й аминокислотный остаток (лизин), 10-й аминокислотный остаток (фенилаланин), 11-й аминокислотный остаток (метионин), 13-й аминокислотный остаток (треонин), 20-й аминокислотный остаток (серин), 42-й аминокислотный остаток (глютамин), 43-й аминокислотный остаток (серин), 60-й аминокислотный остаток (аспарагиновая кислота), 63-й аминокислотный остаток (треонин), 77-й аминокислотный остаток (аспарагин), 78-й аминокислотный остаток (валин), 80-й аминокислотный остаток (серин), 83-й аминокислотный остаток (лейцин), 85-й аминокислотный остаток (аспарагиновая кислота), 87-й аминокислотный остаток (фенилаланин), 99-й аминокислотный остаток (глицин), 103-й аминокислотный остаток (лейцин) и 108-й аминокислотный остаток (аланин) от N-конца аминокислотной последовательности легкой цепи TRA-8 замещены пролином, серином, серином, лейцином, аланином, треонином, лизином, аланином, серином, серином, серином, лейцином, пролином, фенилаланином, треонином, тирозином, глютамином, валином и треонином соответственно. Полученная последовательность была обозначена как LM2.
Плазмиды экспрессии, содержащие указанные аминокислотные последовательности в легкой цепи антитела TRA-8 против человеческого DR5, конструируют приведенным ниже способом.
1) Синтез праймеров для получения вариабельного и константного участков гуманизированной легкой цепи TRA-8
ДНК, кодирующие полипептидную цепь LM2 (SEQ ID No. 46 в перечне последовательностей), каждая из которых представляет собой слияние вариабельного участка гуманизированной легкой цепи анти-DR5 антитела TRA-8 и константного участка легкой цепи человеческого Ig (κ цепи), синтезируют с использованием соответствующих сочетаний ПЦР.
Далее до 7AL1P (SEQ ID No. 47) и 7ALCN (SEQ UD No. 48) синтезируют следующие олигонуклеотидные праймеры по методике ПЦР:
5'-gtcccccaca gatgcagaca aagaacttgg agattgggtc atcacaatgt caccagtgga-3' (HKSPR11; SEQ ID No. 49);
5'-ccaagttctt tgtctgcatc agtaggagac agggtcacca tcacctgc-3' (HKCDF11 SEQ ID No. 50);
5'-agtgtgccgg gtggatgccc agtaaatcag tagtttagga gctttccctg gtttctg-3' (HKCDR12; SEQ ID No. 51);
5'-tgggcatcca cccggcacac tggggtccca agcaggttta gtggcagt-3' (HKCDF22 SEQ ID No. 52);
5'-ataactacta tattgctgac agtaataggt tgcaaaatcc tccggctgca gactagagat ggt-3' (HKCDR22 SEQ ID No. 53) и
5'-cagcaatata gcagctatcg gacgttcggt caaggcacca aggtggaaat caaacggact gtg-3' (HKCF12 SEQ ID No. 54).
2) Конструирование плазмиды pCR3.1/M2-1 (клонирование гуманизированной легкой цепи TRA-8)
Фрагмент LM2-ДНК, определенный в SEQ ID No. 55 в перечне последовательностей, который кодирует аминокислотную последовательность, определенную в SEQ ID No. 46, полученный при проведении 2-стадийной ПЦР, вставляют в плазмидный вектор и клонируют в E. coli.
а) Первая стадия ПЦР
Фрагмент LM2-F1-ДНК, кодирующий сигнальную последовательность секреции и часть участка FRL1 с сайтом расщепления рестрикционным ферментом HindIII, добавленный к 5'-концу, получают в следующих условиях. Матричные плазмиды pHSGHM17 и pSRPDHH получают по способу, описанному в заявке на европейский патент EP 0909816 A1.
Реакционная среда имеет указанный ниже состав:
ДНК плазмиды pHSGHM17 (заявка на европейский патент EP 0909816 А1), 25 нг;
олигонуклеотидный праймер 7AL1P, 50 пмоль;
олигонуклеотидный праймер HKSPR11, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq (Perkin Elmer), 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при температуре 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающий нагревание при температуре 94°С в течение 1 минуты, при температуре 55°С в течение 1 минуты и при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз, после чего проводят нагревание при 72°С в течение 10 минут.
Фрагмент LM2-F2-ДНК, кодирующий часть FRL1, CDRL1, FRL2 и CDRL2, получают в указанных ниже условиях.
Реакционная смесь имеет следующий состав:
ДНК плазмиды pL28, 25 нг;
олигонуклеотидный праймер HKCDF11, 50 пмоль;
олигонуклеотидный праймер HKCDR12, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единиц.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при температуре 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающий нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при температуре 72°С в течение 10 минут.
Фрагмент LM2-F3-ДНК, кодирующий CDRL2, FRL3 и часть CDRL3 получают в указанных ниже условиях.
Реакционный раствор имеет следующий состав:
ДНК плазмиды pSRPDHH (заявка на европейский патент EP 0909816 А1), 25 нг;
олигонуклеотидный праймер HKCDF22, 50 пмоль;
олигонуклеотидный праймер HKCDR22, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют в методике ПЦР.
Температурные условия для проведения ПЦР: нагревают при температуре 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающий нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем повторяют нагревание при температуре 72°С в течение 10 минут.
Фрагмент LM2-F4-ДНК, кодирующий CDRL3, FRL4 и константный участок с сайтом расщепления для рестрикционного фермента EcoRI, добавленным к 3'-концу, получают в указанных ниже условиях:
Реакционная смесь имеет следующий состав:
ДНК плазмиды pSRPDHH, 25 нг;
олигонуклеотидный праймер HKCF12, 50 пмоль;
олигонуклеотидный праймер 7ALCN, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающий нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при температуре 72°С в течение 10 минут.
Фрагменты амплифицированной ДНК после проведения ПЦР разделяют электрофорезом в 5% полиакриламидном геле. Гель после электрофореза окрашивают добавлением 1 мкг/мл бромида этидия для выявления полученной ДНК в УФ свете. Идентифицируют соответствующие полосы ДНК и вырезают лезвием.
b) Вторая стадия ПЦР
ДНК LM2, которая содержит слитые фрагменты LM2-F1-DNA, LM2-F2-DNA, LM2-F3-DNA и LM2-F4-DNA, получают в следующих условиях:
Реакционный раствор имеет следующий состав:
гелевый фрагмент LM2-F1-DNA, полученный на первой стадии ПЦР,
гелевый фрагмент LM2-F2-DNA, полученный на первой стадии ПЦР,
гелевый фрагмент LM2-F3-DNA, полученный на первой стадии ПЦР,
гелевый фрагмент LM2-F4-DNA, полученный на первой стадии ПЦР,
олигонуклеотидный праймер 7AL1P, 50 пмоль,
олигонуклеотидный праймер 7ALCN, 50 пмоль,
смесь дНТФ, 5,0 мкл,
10× буфер для ПЦР, 5,0 мкл,
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при температуре 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающий нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при температуре 72°С в течение 10 минут.
Полученный таким образом фрагмент LM2-ДНК вставляют в ДНК плазмиды pCR3.1 с использование набора для клонирования (Eukaryotic TA cloning Kit (Invitrogen)) по инструкции от производителя и вводят в компетентные клетки E. coli TOP10F', входящие в состав набора. Нуклеотидные последовательности указанных ДНК, кодирующих гуманизированную легкую цепь TRA-8, одтверждают дидезоксиметодом (Sanger, F. S., et al. (1977), Proc. Natl. Acad. Sci. USA, 74:5463-5467) с использованием анализатора ДНК 3700 (ABI PRISM; Perkin Elmer Applied Biosystems, Japan).
Полученные плазмиды обозначают как pCR3.1/M2-1 (плазмида, несущая кДНК, кодирующую вариабельный участок гуманизированной легкой цепи TRA-8 и константный участок легкой цепи человеческого Ig).
Полученную плазмиду pCR3.1/М2-1, содержащую фрагмент LM2-ДНК, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной ДНК плазмиды pHSG399 расщепляют рестрикционными ферментами Hind III и EcoR I и затем подвергают дефосфорилированию с использованием CIP. Полученные дефосфорилированная ДНК pHSG399 и фрагмент LM2-ДНК, которые были расщеплены рестрикционными ферментами Hind III и EcoR I, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем E. coli DH5α трансформируют лигированной ДНК и распределяют по агаровой среде LB, содержащей 0,1 мМ IPTG, 0,1%X-Gal и 50 мкг/мл хлорамфеникола (конечные концентрации). Полученные белые трансформанты культивируют в жидкой среде LB, содержащей 50 мкг/мл хлорамфеникола, и плазмидную ДНК экстрагируют из полученной культуры по методу, включающему обработку щелочью и ДСН. Экстрагированную плазмидную ДНК расщепляют Hind III и EcoR I и затем клон, содержащий фрагмент LM2-ДНК, выделяют электрофорезом в 1% геле агарозы.
В результате проведенной процедуры получают плазмиду pHSG/М2-1-4, содержащую фрагмент слияния вариабельного участка гуманизированной легкой цепи LM2 TRA-8 и константного участка цепи человеческого Igκ. Трансформированный штамм E. coli, несущий плазмиду, обозначенный как E. coli DH5α/pHSG/M2-1-4, депонируют в Международном депозитарии запатентованных организмов при Национальном Институте современной промышленной науки и технологии Японии (International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology, 1-1, Higashi 1 chome Tsukuba-shi, Ibaraki-ken, 305-5466) 20 апреля 2001 года в соответствии с Будапештским договором по депонированию микроорганизмов под номером FERM BP-7563.
3) Конструирование плазмиды pSR/M2-1 (плазмида экспрессии для гуманизированной легкой цепи LM2 TRA-8)
Полученную плазмиду pHSG/M2-1-4, содержащую фрагмент слияния вариабельного участка гуманизированной легкой цепи LM2 TRA-8 и константного участка цепи человеческого Igκ, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной ДНК плазмиды pSRPDHH (заявка на европейский патент EP 0909816 A1) расщепляют рестрикционными ферментами Hind III и EcoR I и затем подвергают дефосфорилированию с использованием CIP. Образовавшиеся дефосфорилированная ДНК pSRPDHH и фрагмент ДНК Hind III-EcoR I, полученный из pHSG/M2-1-4, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем E. coli DH5α трансформируют лигированной ДНК и распределяют по агаровой среде LB. Полученные трансформанты культивируют в жидкой среде LB, содержащей 100 мкг ампициллина, и плазмидную ДНК экстрагируют из полученных культур по методу, включающему обработку щелочью и ДСН. Наличие вставки и ориентацию желательного фрагмента ДНК в векторе pSRPDHH подтверждают секвенированием ДНК с использованием анализатора генной последовательности (ABI PRISM 3700 DNA Analyzer; Applied Biosystems).
Полученную плазмиду экспрессии, несущую кДНК, кодирующую гуманизированную легкую цепь TRA-8, обозначают как pSR/M2-1.
Пример 20. Получение гуманизированного антитела
Трансфекцию COS-7 клеток, то есть клеточной линии, полученной из почки обезьяны, содержащей плазмиды экспрессии для гуманизированной тяжелой цепи TRA-8 и для гуманизированной легкой цепи TRA-8, указанных выше, проводят по методике с использованием реагента для трансфекции FUGENE6 (Boehringer Mannheim Biochemica) в соответствии с инструкцией от производителя, прилагаемой к набору.
Клетки COS-7 (Американская Коллекция типовых культур) (American Type Culture Collection No. CRL-1651) растят до состояния полуслияния (3·106 клеток/чашку) в чашках для культивирования (площадь культивирования: 57 см2; Sumitomo Bakelite), содержащих модифицированную Дульбекко среду Игла (далее обозначаемую как «D-MEM»; Gibco BRL) с добавкой 10% фетальной сыворотки теленка (далее обозначаемой как «ФСТ»; Moregate).
Тем временем смешивают плазмидную ДНК для экспрессии гуманизированной тяжелой цепи DR5 (pHA15-1) в количестве 10 мкг/чашку (всего 5 чашек) и плазмидную ДНК для экспрессии гуманизированной легкой цепи DR5 в количестве 10 мкг/чашку, полученные по методу, включающему обработку щелочью и ДСН и центрифугирование в градиенте плотности хлорида цезия, после чего проводят осаждение этанолом и далее суспендируют в количестве 5 мкл/чашку dH2O.
После смешивания реагента для трансфекции FUGENE6 в количестве 15 мкл/чашку с D-MEM без ФСТ в количестве 180 мкг/чашку указанный раствор FUGENE (185 мкл/чашку) смешивают с 5 мкл/чашку раствором ДНК, содержащим в количестве 10 мкг/чашку плазмидную ДНК для экспрессии гуманизированной тяжелой цепи DR5 и в количестве 10 мкг/чашку плазмидную ДНК для экспрессии гуманизированной легкой цепи DR5. После 15-минутной инкубации при комнатной температуре полученную плазмидную суспензию (200 мкг) добавляют к предварительно приготовленным чашкам с COS-7. После инкубации в атмосфере 5% CO2 при 37°C в течение 24 часов культуральную среду заменяют на D-MEM без ФСТ. После инкубации в атмосфере 5% CO2 при температуре 37°C в течение 72 часов культуральный супернатант регенерируют до очищения продуктов экспрессии в супернатантных жидкостях. По описанному выше методу клетки COS-7 трансфицируют одним из указанных ниже вариантов сочетаний:
(А): без плазмидной ДНК
(В): совместная трансфекция с использованием pHB14-1 и pSR/M2-1
Далее культуру центрифугируют (1000 об/мин, 5 минут) и отбирают супернатант. Супернатант снова центрифугируют (9800 об/мин, 15 минут) и фильтруют через фильтр с размером пор 0,45 мкм (ADVANTEC TOYO DISMIC-25cs, Cat # 25CS045 AS). Очистку IgG из фильтратов проводят с использованием аффинной хроматографии на белке G-POROS (Applied Biosystems) в указанных ниже условиях:
ВЭЖХ: BioCAD 700E (Applied Biosystems);
колонка: Белок-G-ID с сенсорным картриджем (размер колонки: 2,1 мм (внутр. диаметр) × 30 мм длина, объем слоя: 0,1 мл, Cat# 2-1002-00 Applied Biosystems);
элюирующий буфер: 0,1М глицин-HCl (pH 2,5);
нейтрализующий буфер: 1М Трис-HCl (pH 8,5);
выявление: при длине волны 280 нм;
скорость течения: 1 мл/мин;
размер фракции: 0,5 мл/0,5 мин;
пробирка для сбора фракций: полипропиленовая микропробирка на 1,5 мл;
температура: 4°С.
После нанесения всего фильтрата на колонку используют 30 мл ФБР (Sigma, Cat # 1000-3) для промывки колонки. После добавления элюирующего буфера включают коллектор фракций. Каждая микропробирка для сбора фракций содержит внесенные ранее 55 мкл 1М NaCl, 110 мкл нейтрализующего буфера и 74 мкл бычьего сывороточного альбумина в концентрации 2 мг/мл (Sigma, Cat # А-7030) в ФБР. Собирают фракции с No. 8 до No. 10 и проводят диализ против 1 литра ФБР (pH 7,5) при 4°С в течение 1 дня с использованием Slide-A lyzer (Pierce, Cat # 66450). Буфер для диализа меняют дважды.
Подтверждение экспрессии гуманизированных антител и количественный анализ продуктов экспрессии в жидкостях супернатантных проводят по методу ELISA с использованием антитела против человеческого IgG.
К каждой лунке в 96-луночном микропланшете (MaxiSorp, Nunc) добавляют 100 мкл козьего специфичного поликлонального антитела против человеческого IgG Fc (Kappel), растворенного до конечной концентрации 0,5 мкг/мл в буфере для адсорбции (0,05 М гидрокарбонат натрия, 0,02% азид натрия, pH 9,6), и планшет инкубируют при 37°С в течение 2 часов для адсорбции антитела. Затем планшет промывают пять раз 300 мкл ФБР(-), содержащим 0,05% Твин-20 (BioRad) (далее обозначаемого как «ФБР-Т»). К лункам после промывки добавляют культуральный супернатант, разбавленный D-MEM, содержащим 10% ФСТ, и инкубируют при 37°С в течение 2 часов. После новой промывки с использованием ФБР-Т к каждой лунке добавляют 100 мкл козьего специфичного поликлонального антитела против человеческого IgG Fc, меченного щелочной фосфатазой (Jackson Immuno Research Lab.), разбавленного в 10000 раз ФБР-Т, и инкубируют при 37°С в течение 2 часов. После еще одной промывки с использованием ФБР-Т добавляют субстратный раствор, содержащий p-нитрофенилфосфат, который входит в состав набора для субстратов щелочной фосфатазы (BioRad), в соответствии с инструкцией от производителя, прилагаемой к набору. После инкубирования при 37°С в течение периода времени от 0,5 часа до 1 часа измеряют поглощение при длине волны 405 нм. В настоящих экспериментах человеческий иммуноглобулин G подкласса 1 из плазмы человека (IgG1) (Biopure AG), разбавленный D-MEM, содержащим 10% ФСТ, до определенных концентраций, используют в качестве эталонных образцов гуманизированных антител DR5, содержащихся в жидкостях супернатантов культур.
В результате экспрессированные и очищенные продукты из культурального супернатанта специфично выявляются при использовании антитела против человеческого IgG. Количество антитела против человеческого IgG составляет 8,96 мкг (800 мкл).
Пример 21. Апоптоз-индуцирующая активность гуманизированного антитела
Клетки Jurkat (ATCC No. TIB-152) используют для выявления апоптоз-индуцирующей активности очищенного гуманизированного антитела TRA-8.
Клетки Jurkat, культивированные в среде RPMI1640 с добавкой 10% ФСТ (Gibco BRL) при 37°С в течение 3 дней в атмосфере 5% CO2, далее разливают в каждую лунку 96-луночного микропланшета (Sumitomo Bakelite) по 50 мкл на лунку. Гуманизированное TRA-8, полученное по методике примера 20, доводят до концентрации готового рассматриваемого продукта, равной 100 нг/мл, с использованием RPMI1640, содержащей 10% ФСТ, при оценке концентраций в жидкостях по методике, описанной в примере 20. Каждый из растворов экспрессированных продуктов, доведенных до концентрации 100 нг/мл, используют для получения серийных разбавлений с повтором серийных 2-кратных разбавлений средой RPMI1640, содержащей 10% ФСТ. Каждый из разбавленных растворов гуманизированного TRA-8 добавляют в каждую лунку по 50 мкл на лунку. После реакции в течение 12 часов при 37°С добавляют 50 мкл 20 мкМ ФМС (феназинметосульфат; Sigma Chemical Co.), содержащего 1 мг/мл ХТТ (2,3-bis[2-метокси-4-нитро-5-сульфофенил]-2Н-тетразолий-5-карбоксиангидрида внутрикомплексная соль; Sigma, Chemical Co.) (конечная концентрация 250 мкг/мл для ХТТ и 5 мкМ для ФМС). После инкубации в течение 3 часов измеряют поглощение при длине волны 450 нм в каждой лунке и вычисляют жизнеспособность клеток с использованием в качестве показателя восстановительной способности митохондрий.
Жизнеспособность клеток в каждой из лунок вычисляют по следующей формуле:
Жизнеспособность (%) = 100 · (a-b)/(c-b)
где «а» обозначает величину, измеренную в исследуемой лунке, «b» обозначает результат измерения в лунке, не содержащей клеток, и «c» обозначает результат измерения в лунке, в которую не добавлено антитело.
При этом было показано, что продукт экспрессии, полученный в примере 20 (иммунизированный TRA-8), индуцирует апоптоз в клетках клеточной линии Т-лимфомы, экспрессирующих человеческих антиген DR5.
Пример 22. Реакционная способность TRA-8 в отношении различных молекул DR5
Для того чтобы определить реакционную способность TRA-8 к различным молекулам DR5, определяют реакционную способность TRA-8 с использованием активированных лимфоцитов указанным ниже способом.
Вначале отбирают образцы крови у человека (30 мл), мартышки (3 мл) и обезьян циномолгус (20 мл). Образцы крови содержат добавленный к ним 1 мл гепарина (Novoheparin; Novo), затем образцы медленно наслаивают на равный объем раствора Ficoll-Paque PLUS (Amersham Pharmacia Biotech.) с удельной плотностью 1,077 во всех случаях, за исключением варианта для обезьян циномолгус, в случае которых используют раствор с удельной плотностью 1,072) и центрифугируют со скоростью 1700 об/мин в течение 30 минут с получением фракции мононуклеарных клеток периферической крови. Фракцию мононуклеарных клеток промывают дважды сбалансированным солевым раствором Хэнкса и затем суспендируют в среде RPMI1640 c добавкой 10 об./об.% ФСТ до плотности клеток 1·106 клеток/мл. Добавляют фитогемагглютинин-Р (PHA-P, Sigma Chemical Co.) к полученной суспензии до конечной концентрации 5 мкг/мл и образец инкубируют при 37°С в атмосфере 5 об./об.% CO2 в течение 24 часов. По истечении указанного периода времени клетки регенерируют центрифугированием, промывают и суспендируют в среде RPMI1640, содержащей 10 об./об.% ФСТ. Затем для активации регенерированных клеток к суспензии добавляют интерлейкин-2 (Amersham Pharmacia Biotech.) до конечной концентрации 10 единиц/мл и инкубируют при 37°С в атмосфере 5 об./об.% CO2 с в течение 72 часов.
Количество активированного препарата, который по расчету содержит 1·106 активированных лимфоцитарных клеток, вносят в пробирку и либо суспендируют в 50 мкл 0,5, 1, 5, 10 мкг/мл TRA-8 в ФБР, либо только в 50 мкл одного ФБР. Полученную суспензию оставляют на льду на 1 час, после чего клетки промывают 3 раза аликвотами по 500 мкл ФБР и затем суспендируют в 50 мкл FITC-меченного антитела против мышиного IgG (Bioresource) с концентрацией 20 мкг/мл в ФБР. С использованием клеток, суспендированных в 500 мкл ФБР в качестве контроля, измеряют интенсивность флуоресценции на проточном цитометре (FACSCalibur; Becton Dickinson).
Получают характеристики распределения числа клеток по интенсивности флуоресценции и вычисляют пропорцию количества окрашенных клеток к общему количеству клеток. Далее вычисляют значение Kd использованием концентрации TRA-8 и данных о пропорции количества окрашенных клеток к общему количеству клеток. Каждая частота реакционной способности активации лимфоцитов человека, мартышки и обезьян циномологус примерно одна и та же. Соответственно TRA-8 способен связываться с широким перечнем DR5 приматов, включая человека, для которого TRA-8 был исходно приготовлен.
Пример 23. Исследование TRA-8 у мартышек по методу повышения дозы
Предварительное исследование токсичности с повышением дозы TRA-8 проводят на 1 самце и 1 самке мартышек. Проводят три варианта однократного внутривенного дозирования, разделенные 7-дневным периодом без введения дозы. Дозу TRA-8 устанавливают в количестве 50, 250 и 1250 мкг/тело. Через сорок восемь часов после каждого введения отбирают кровь из бедренной вены и получают плазму. На анализаторе (FUJI DRI-CHEM: Fuji Film Medical Co., Ltd.) измеряют плазменную активность аспартатаминотрансферазы и аланинаминотрансферазы. Все образцы крови отбирают без анестезии. В результате по данным биохимического анализа плазмы в каждом из вариантов введения не было получено доказательств наличия повреждения печени.
Пример 24. Фармакологические исследования действия TRA-8 in vitro и in vivo против злокачественных клеток
Для определения наличия у TRA-8 терапевтической эффективности в качестве средства для противораковой терапии оценивают его активность по уничтожению клеток in vitro с использованием различных клеточных линий злокачественной опухоли, как показано ниже.
Различные злокачественные клетки ((2-8)·103 клетка/50 мкл), культивируемые в среде RPMI1640 (для Jurkat), в среде DMEM (для HCT-116), в MEM-R (для WiDR) или DMEM-F12 (для COL2-Jck), полученных от Gibco BRL, с добавлением 10% ФСТ (Gibco BRL) при 37°С в атмосфере 5% CO2, вносят в лунки 96-луночного микротитрационного планшета (Sumitomo Bakelite). Концентрацию TRA-8 доводят до концентрации соответствующего готового продукта (100 нг/мл) средой, содержащей 10% ФСТ. Раствор TRA-8 (100 нг/мл) используют для получения серийных разбавлений с повтором серийных 2-кратных разбавлений средой, содержащей 10% ФСТ. К лункам добавляют каждый из разбавленных растворов TRA-8 в количестве по 50 мкл на лунку и инкубируют при 37°С. После реакции при 37°С в течение 72 часов добавляют 50 мкл 25 мкМ ФМС (феназинметосульфат; Sigma Chemical Co.), содержащей 1 мг/мл ХТТ (конечная концентрация 250 мкг/мл для ХТТ и 5 мкМ для ФМС). После инкубации в течение 3 часов измеряют поглощение при длине волны 450 нм в каждой лунке и вычисляют жизнеспособность клеток с использованием в качестве показателя восстановительной способности митохондрий.
Жизнеспособность клеток в каждой из лунок вычисляют по следующей формуле:
Жизнеспособность (%) = 100 · (a-b)/(c-b),
где «а» обозначает результат измерения в опытной лунке, «b» обозначает результат измерения в лунке без клеток, и «c» обозначает результат измерения в лунке, в которой не добавлено антитело.
Ниже в таблице 3 показаны результаты.
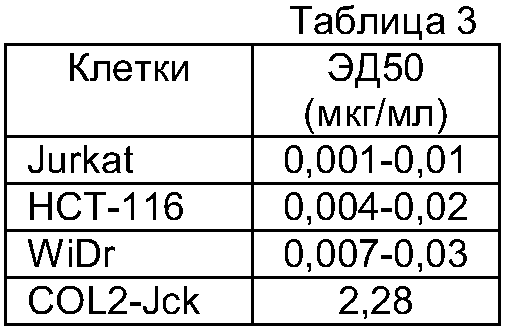
Показано, что в различных злокачественных клетках происходит мощная индукция апоптоза под действием TRA-8 в условиях in vitro.
Далее определяют противоопухолевый эффект TRA-8 in vivo у мышей nude с трансплантированными клетками WiDr, поскольку TRA-8 не вступает в перекрестную реакцию с мышиным DR5.
Антитело TRA-8 против человеческого DR5 вводят мышам nude, несущим человеческие ксенотрансплантаты, которые экспрессируют молекулы человеческого DR5. Используют 6-недельных мышей BALb/c nude/nude (самки получены от Clea Japan Inc.), которым были трансплантированы клеточные линии WiDr (5 мм3) рака толстой кишки человека. На первый день после трансплантации опухоли указанных мышей с трансплантированной опухолью подвергают ежедневной внутрисуставной инъекции TRA-8 (5 мкг/тело) в течение 14 раз. Рост опухоли WiDr определяют ежедневно при определении размера опухолевой массы. Ниже в таблице 4 показаны полученные результаты.
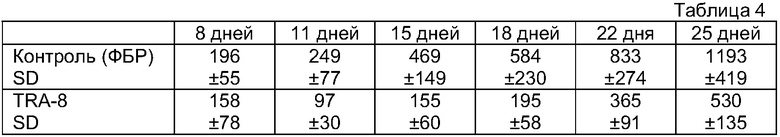
В рамках указанной модели у всех необработанных животных отмечается явный рост опухоли, тогда как у животных, которым вводили TRA-8, наблюдается подавление роста опухоли, что видно по размеру опухоли. Данный результат показывает, что TRA-8 является эффективным средством для элиминации опухолевых клеток in vivo.
Пример 25. Сочетанное исследование TRA-8
Клеточная линия раковых клеток предстательной железы человека PC-3, полученная из Американской Коллекции типовых культур (ATCC), поддерживается на питательной смеси F-12K Nutrient Mixture (21127-022 Gibco BRL), содержащей 10% фетальной сыворотки теленка (ФСТ, Hyclone), 1% L-глютамина, 200 мМ (25030-149; Gibco BRL) и 0,5% раствор пенициллина-стрептомицина (P-7539, Sigma). В описанном ниже эксперименте используют среду RPMI1640 (MED-008, IWAKI) с добавкой 10% ФСТ и 0,5% раствора пенициллина-стрептомицина. Растущие РС-3 клетки в экспоненциальной стадии роста собирают с проведением обработки трипсином и двойной промывкой свежей средой. Затем клетки подсчитывают, ресуспендируют в свежей среде с плотностью 5·104 клеток/мл и распределяют в тройном повторе по лункам 96-луночного микропланшета с плоским дном (3598, Corning-Coster) в общем объеме 100 мкл/лунка за один день до начала эксперимента. Репрезентативное противораковое средство Паклитаксел (169-18611, Wako), растворенное в диметилсульфоксиде (10 мг/мл), разбавляют свежей средой и затем добавляют к лункам 96-луночного планшета, содержащим клетки в количестве 50 мкл/лунку. Конечная концентрация диметилсульфокисида меньше, чем 0,1%. После инкубации в течение 24 часов при 37°С в атмосфере 5% CO2 к лункам добавляют TRA-8, разбавленный свежей средой. После инкубации еще в течение 24 часов к лункам добавляют 50 мкл минимальной эссенциальной среды (11095-098, Gibco BRL), содержащей 1 мг/мл XTT и 25 мМ ФМС и планшет инкубируют в течение 6 часов. Затем измеряют величину поглощения ОП450 на спектрофотометре SPECTRA MAX 250 (Molecilar Devices) и подсчитывают жизнеспособность клеток, как указано ниже.
Жизнеспособность клеток (%) = (ОП450 для лунки, содержащей клетки, обработанные Таксолом и/или TRA-8 агент(ы)) - ОП450 для лунки, не содержащие ни клеток, ни средства) · 100/(ОП450 для лунки, содержащей клетки без средства - ОП450 для лунки, не содержащей ни клеток, ни средства).
Результат указанного выше тестирования варианта, включающего использование TRA-8 в сочетании с репрезентативным противораковым средством паклитаксел, выглядит следующим образом. Паклитаксел снижает жизнеспособность РС-3 клеток, но более 40% сигналов, указывающих на наличие жизнеспособных злокачественных клеток, еще остается при концентрациях до 200 нМ. Следует отметить, что добавление 0,1% нг/мл TRA-8 очень сильно снижает жизнеспособность раковых клеток, вплоть до 10%, даже в том случае, когда не отмечается снижения жизнеспособности клеток после однократного внесения TRA-8 в данной концентрации. Приведенный результат четко указывает на то, что TRA-8 демонстрирует противораковую активность синергично в случае объединения с другими противораковыми средствами.
Пример 26. Анализ других гуманизированных антител TRA-8
(1) Разработка гуманизированных антител
Конструирование гуманизированного варианта TRA-8 проводят по методу, известному как пересадка КДУ. Антитело mAB58'CL, используемое как акцептор в соответствии с описанной в стандартном примере 2 процедурой, и КДУ антитела TRA-8 пересаживают на акцептор. На участке рамки для считывания некоторые аминокислоты пересаживают на акцептор, взятые либо из TRA-8, либо из человеческой консенсусной последовательности, в соответствии с критериями, определенными Квином с соавт. (Queen et al., Ptoc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)) и последовательности гуманизированного TRA-8 конструируют, как будет описано ниже.
(2) Конструирование плазмиды, несущей ДНК для вариабельного участка тяжелой цепи других типов гуманизированных или мышиных TRA-8
Как показано на SEQ ID No. 56 в перечне последовательностей, гуманизация аминокислотных последовательностей типа Н1 тяжелой цепи мышиного антитела TRA-8 против человеческого DR5 включает замену 3-ного аминокислотного остатка (метионина), 13-ного аминокислотного остатка (лизина), 19-ного аминокислотного остатка (лизина), 40-ного аминокислотного остатка (треонина), 42-ного аминокислотного остатка (глютаминовой кислоты), 44-ного аминокислотного остатка (аргинина), 84-ного аминокислотного остатка (серина), 88-ного аминокислотного остатка (серина), 93-ного аминокислотного остатка (метионина), 114-ного аминокислотного остатка (треонина), 115-ного аминокислотного остатка (лейцина) глютамином, глютамином, аргинином, аланином, глицином, глицином, аспарагином, аланином, валином, лейцином и валином соответственно.
Как показано в SEQ ID No. 59 в перечне последовательностей, гуманизация аминокислотных последовательностей Н3 типа тяжелой цепи мышиного антитела TRA-8 против человеческого DR5 включает замену 13-ного аминокислотного остатка (лизина), 19-ного аминокислотного остатка (лизина), 40-ного аминокислотного остатка (треонина), 42-ного аминокислотного остатка (глютаминовой кислоты), 44-ного аминокислотного остатка (аргинина), 88-ного аминокислотного остатка (серина), 93-ного аминокислотного остатка (метионина), 114-ного аминокислотного остатка (треонина), 115-ного аминокислотного остатка (лейцина) глютамином, аргинином, аланином, глицином, глицином, аланином, валином, лейцином и валином соответственно.
Как показано в SEQ ID No. 60 в перечне последовательностей, гуманизация аминокислотных последовательностей Н4 типа тяжелой цепи мышиного антитела TRA-8 против человеческого DR5 включает замену 13-ного аминокислотного остатка (лизина), 19-ного аминокислотного остатка (лизина), 88-ного аминокислотного остатка (серина), 93-ного аминокислотного остатка (метионина), 114-ного аминокислотного остатка (треонина), 115-ного аминокислотного остатка (лейцина) глютамином, аргинином, аланином, валином, лейцином и валином соответственно.
Как показано в SEQ ID No. 61 в перечне последовательностей, плазмида, содержащая ДНК для тяжелой цепи вариабельного участка химерного TRA-8, обозначена как плазмида «М типа». Кроме того, гуманизированный TRA-8, описанный в примерах 17 и 18, обозначен как относящийся к «Н2 типу».
Плазмиды, содержащие ДНК, которая кодирует тяжелую цепь вариабельного участка гуманизированного или химерного TRA-8, конструируют согласно описанной ниже процедуре.
Используют ПЦР для конструирования приведенных ниже последовательностей ДНК, каждая из которых включает описанную выше:
Синтезируют приведенные ниже 24 олигонуклеотила:
5'-ttggataagc ttggcttgac ctcaccatgg gatggagctg tatcatcctc ttcttggtag caacagctac aggtgtccac-3' (A; SEQ ID No. 32);
5'-tctgaagtaa tgctggtgga gtctggggga ggcttagtac agcctggagg gtccctgaga ctctcctgtg cagcctctgg-3' (В; SEQ ID No. 33);
5'-tctgaagtac agctggtgga gtctggggga ggcttagtac agcctggagg gtccctgaga ctctcctgtg cagcctctgg-3' (B2; SEQ ID No. 57);
5'-tctgaagtaa tgctggtgga gtctggggga ggcttagtaa agcctggagg gtccctgaaa ctctcctgtg cagcctctgg-3' (B3; SEQ ID No. 66);
5'-attcactttc agtagttatg taatgtcttg ggttcggcag gcaccaggga agggtctgga gtgggttgca accattagta-3' (С; SEQ ID No. 34);
5'-attcactttc agtagttatg taatgtcttg ggttcggcag actccagaga agaggctgga gtgggttgca accattagta-3' (C2; SEQ ID No. 64);
5'-gtggtggtag ttacacctac tatccagaca gtgtgaaggg ccgattcacc atctccagag acaatgccaa gaacaccctg-3' (D; SEQ ID No. 35);
5'-tatctgcaaa tgaacagtct gagagcagag gacacggctg tttattactg tgcaagaagg ggtgactcta tgattacgac-3' (E; SEQ ID No. 36);
5'-tatctgcaaa tgagcagtct gagagcagag gacacggctg tttattactg tgcaagaagg ggtgactcta tgattacgac-3' (E2; SEQ ID No. 62);
5'-tatctgcaaa tgagcagtct gagatctgag gacacggcta tgtattactg tgcaagaagg ggtgactcta tgattacgac-3' (E3; SEQ ID No. 67);
5'-ggactactgg ggccaaggga ccctggtcac agtctcctca gcctccacc aagggcccat cggtc-3'(F; SEQ ID No.37);
5'-ggactactgg ggccaaggga ccactctcac agtctcctca gcctccacc aagggcccat cggtc-3'(F2; SEQ ID No.68);
5'-ctaccaagaa gaggatgata cagctccatc ccatggtgag gtcaagccaa gcttatccaa-3' (G; SEQ ID No. 38);
5'-tctcagggac cctccaggct gtactaagcc tcccccagac tccaccagca ttacttcaga gtggacacct gtagctgttg-3' (H; SEQ ID No. 39);
5'-tctcagggac cctccaggct gtactaagcc tcccccagac tccaccagct gtacttcaga gtggacacct gtagctgttg-3' (H2; SEQ ID No. 58);
5'-tttcagggac cctccaggct ttactaagcc tcccccagac tccaccagca ttacttcaga gtggacacct gtagctgttg-3' (H3; SEQ ID No. 69);
5'-tccagaccct tccctggtgc ctgccgaacc caagacatta cataactact gaaagtgaat ccagaggctg cacaggagag-3' (I; SEQ ID No. 40);
5'-tccagcctct tctctggagt ctgccgaacc caagacatta cataactact gaaagtgaat ccagaggctg cacaggagag-3' (I2; SEQ ID No. 65);
5'-ctctggagat ggtgaatcgg cccttcacac tgtctggata gtaggtgtaa ctaccaccac tactaatggt tgcaacccac-3' (J; SEQ ID No. 41);
5'-ccttcttgca cagtaataaa cagccgtgtc ctctgctctc agactgttca tttgcagata cagggtgttc ttggcattgt-3' (К; SEQ ID No. 42);
5'-ccttcttgca cagtaataaa cagccgtgtc ctctgctctc agactgttca tttgcagata cagggtgttc ttggcattgt-3' (K2; SEQ ID No. 63);
5'-ccttcttgca cagtaataca tagccgtgtc ctcagatctc agactgctca tttgcagata cagggtgttc ttggcattgt-3' (К3; SEQ ID No. 70);
5'- gaccgatggg cccttggtgg aggctgagga gactgtgacc agggtccctt ggccccagta gtccgtcgta atcatagagt cacc-3' (L; SEQ ID No. 43) и
5'-gaccgatggg cccttggtgg aggctgagga gactgtgaga gtggtccctt ggccccagta gtccgtcgta atcatagagt cacc-3' (L2; SEQ ID No. 71).
Приведенные ниже 2 праймера ПЦР синтезируют по описанной выше методике.
5'-ttggataagc ttggcttgac-3' (PI; SEQ Ю No. 44); и
5'-gaccgatggg cccttggtgg a-3' (P2; SEQ ID No. 45).
Синтез ДНК Н1 типа, кодирующей полипептидную цепь, которая включает сигнальную последовательность секреции, вариабельный участок гуманизированной тяжелой цепи TRA-8 и 8 аминокислотных остатков на N-конце IgG-CH1 участка, проводят с использованием сочетания соответствующих ПЦР.
Фрагмент ДНК Н1 типа получают следующим образом:
Состав раствора для проведения ПЦР реакции:
олигонуклеотид А, 10 пмоль;
олигонуклеотид B2, 10 пмоль;
олигонуклеотид C, 10 пмоль;
олигонуклеотид D, 10 пмоль;
олигонуклеотид E, 10 пмоль;
олигонуклеотид F, 10 пмоль;
олигонуклеотид G, 10 пмоль;
олигонуклеотид H2, 10 пмоль;
олигонуклеотид I, 10 пмоль;
олигонуклеотид J, 10 пмоль;
олигонуклеотид K, 10 пмоль;
олигонуклеотид L, 10 пмоль;
олигонуклеотидный праймер Р1, 2 мкМ;
олигонуклеотидный праймер Р2, 2 мкМ;
10× буфер Pyrobest II, 10 мкл;
смесь дНТФ, 8 мкл;
ДНК-полимераза Pyrobest, 0,5 мкл; и
и повторная дистиллированная вода, вносимая до конечного объема 50 мкл.
Реакцию ПЦР проводят следующим образом. Вначале раствор нагревают при 94°С в течение 5 минут, после чего проводят цикл, включающий нагревание до температуры 98°С в течение 10 секунд, нагревание при температуре 55°С в течение 30 секунд и нагревание при температуре 72°С в течение 1 минуты с повторением указанного цикла 7 раз. После завершения указанной процедуры реакционный раствор нагревают при 72°С в течение 15 минут.
После проведения фенольной экстракции и осаждения этанолом полученный осадок ДНК сушат в вакууме, растворяют в минимальном количестве повторно дистиллированной воды и разделяют электрофорезом в 3% геле агарозы. После проведения электрофореза гель окрашивают водным раствором бромида этидия в концентрации 1 мкг/мл для обнаружения полос ДНК в УФ свете. Полосы ДНК, соответствующие ДНК Н1 типа, вырезают лезвием и элюируют из геля с использованием набора Geneclean Spin (BIO 101, CA, USA). После фенольной экстракции элюированную ДНК концентрируют центрифугированием при 7500g с последующим осаждением этанолом и в конце растворяют в 5 мкл дистиллированной воды.
Синтез ДНК Н3 типа, кодирующей полипептидную цепь, которая включает сигнальную последовательность секреции, вариабельный участок гуманизированной тяжелой цепи TRA-8 и 8 аминокислотных остатков на N-конце IgG-CH1 участка, проводят с использованием сочетания соответствующих ПЦР.
Фрагмент ДНК Н3 типа получают следующим образом:
Состав раствора для проведения ПЦР реакции:
олигонуклеотид А, 10 пмоль;
олигонуклеотид B, 10 пмоль;
олигонуклеотид C, 10 пмоль;
олигонуклеотид D, 10 пмоль;
олигонуклеотид E2, 10 пмоль;
олигонуклеотид F, 10 пмоль;
олигонуклеотид G, 10 пмоль;
олигонуклеотид H, 10 пмоль;
олигонуклеотид I, 10 пмоль;
олигонуклеотид J, 10 пмоль;
олигонуклеотид K2, 10 пмоль;
олигонуклеотид L, 10 пмоль;
олигонуклеотидный праймер Р1, 2 мкМ;
олигонуклеотидный праймер Р2, 2 мкМ;
10× буфер Pyrobest II, 10 мкл;
смесь дНТФ, 8 мкл;
ДНК-полимераза Pyrobest, 0,5 мкл; и
повторная дистиллированная вода, вносимая до конечного объема 50 мкл.
Реакцию ПЦР проводят следующим образом. Вначале раствор нагревают при 94°С в течение 5 минут, после чего проводят цикл, включающий нагревание до 98°С в течение 10 секунд, нагревание при температуре 55°С в течение 30 секунд и нагревание при 72°С в течение 1 минуты с повторением указанного цикла 7 раз. После завершения указанной процедуры реакционный раствор нагревают при температуре 72°С в течение 15 минут.
После проведения фенольной экстракции и осаждения этанолом полученный осадок ДНК сушат в вакууме, растворяют в минимальном количестве повторно дистиллированной воды и разделяют электрофорезом в 3% геле агарозы. После проведения электрофореза гель окрашивают водным раствором бромида этидия в концентрации 1 мкг/мл для обнаружения полос ДНК в УФ свете. Полосы ДНК, соответствующие ДНК Н3 типа, вырезают лезвием и элюируют из геля с использованием набора Geneclean Spin. После фенольной экстракции элюированную ДНК концентрируют центрифугированием при 7500g с последующим осаждением этанолом и в конце растворяют в 5 мкл дистиллированной воды.
Синтез ДНК Н4 типа, кодирующей полипептидную цепь, которая включает сигнальную последовательность секреции, вариабельный участок гуманизированной тяжелой цепи TRA-8 и 8 аминокислотных остатков на N-конце IgG-CH1 участка, проводят с использованием сочетания соответствующих ПЦР.
Фрагмент ДНК Н4 типа получают следующим образом:
Состав раствора для проведения ПЦР реакции:
олигонуклеотид А, 10 пмоль;
олигонуклеотид B, 10 пмоль;
олигонуклеотид C2, 10 пмоль;
олигонуклеотид D, 10 пмоль;
олигонуклеотид E2, 10 пмоль;
олигонуклеотид F, 10 пмоль;
олигонуклеотид G, 10 пмоль;
олигонуклеотид H, 10 пмоль;
олигонуклеотид I2, 10 пмоль;
олигонуклеотид J, 10 пмоль;
олигонуклеотид K2, 10 пмоль;
олигонуклеотид L, 10 пмоль;
олигонуклеотидный праймер Р1, 2 мкМ;
олигонуклеотидный праймер Р2, 2 мкМ;
10× буфер Pyrobest II, 10 мкл;
смесь дНТФ, 8 мкл;
ДНК-полимераза Pyrobest, 0,5 мкл; и
и повторная дистиллированная вода, вносимая до конечного объема 50 мкл.
Реакцию ПЦР проводят следующим образом. Вначале раствор нагревают при 94°С в течение 5 минут, после чего проводят цикл, включающий нагревание до 98°С в течение 10 секунд, нагревание при температуре 55°С в течение 30 секунд и нагревание при температуре 72°С в течение 1 минуты с повторением указанного цикла 7 раз. После завершения указанной процедуры реакционный раствор нагревают при 72°С в течение 15 минут.
После проведения фенольной экстракции и осаждения этанолом полученный осадок ДНК сушат в вакууме, растворяют в минимальном количестве повторно дистиллированной воды и разделяют электрофорезом в 3% геле агарозы. После проведения электрофореза гель окрашивают водным раствором бромида этидия в концентрации 1 мкг/мл для обнаружения полос ДНК в УФ свете. Полосы ДНК, соответствующие ДНК Н4 типа, вырезают лезвием и элюируют из геля с использованием набора Geneclean Spin. После проведения фенольной экстракции элюированную ДНК концентрируют центрифугированием при 7500g с последующим осаждением этанолом и в конце растворяют в 5 мкл дистиллированной воды.
Синтез ДНК М типа, кодирующей полипептидную цепь, которая включает сигнальную последовательность секреции, вариабельный участок тяжелой цепи химерного TRA-8 и 8 аминокислотных остатков на N-конце IgG-CH1 участка, проводят с использованием сочетания соответствующих ПЦР.
Фрагмент ДНК М типа получают следующим образом:
Состав раствора для проведения ПЦР реакции:
олигонуклеотид А, 10 пмоль;
олигонуклеотид B3, 10 пмоль;
олигонуклеотид C2, 10 пмоль;
олигонуклеотид D, 10 пмоль;
олигонуклеотид E3, 10 пмоль;
олигонуклеотид F2, 10 пмоль;
олигонуклеотид G, 10 пмоль;
олигонуклеотид H3, 10 пмоль;
олигонуклеотид I2, 10 пмоль;
олигонуклеотид J, 10 пмоль;
олигонуклеотид K3, 10 пмоль;
олигонуклеотид L2, 10 пмоль;
олигонуклеотидный праймер Р1, 2 мкМ;
олигонуклеотидный праймер Р2, 2 мкМ;
10× буфер Pyrobest II, 10 мкл;
смесь дНТФ, 8 мкл;
ДНК-полимераза Pyrobest, 0,5 мкл; и
и повторно дистиллированная вода, вносимая до конечного объема 50 мкл.
Реакцию ПЦР проводят следующим образом. Вначале раствор нагревают при температуре 94°С в течение 5 минут, после чего проводят цикл, включающий нагревание до 98°С в течение 10 секунд, нагревание при температуре 55°С в течение 30 секунд и нагревание при температуре 72°С в течение 1 минуты с повторением указанного цикла 7 раз. После завершения указанной процедуры реакционный раствор нагревают при температуре 72°С в течение 15 минут.
После проведения фенольной экстракции и осаждения этанолом полученный осадок ДНК сушат в вакууме, растворяют в минимальном количестве повторно дистиллированной воды и разделяют электрофорезом в 3% геле агарозы. После проведения электрофореза гель окрашивают с использованием водного раствора бромида этидия в концентрации 1 мкг/мл для обнаружения полос ДНК в УФ свете. Полосы ДНК, соответствующие ДНК М типа, вырезают лезвием и элюируют из геля с использованием набора Geneclean Spin. После фенольной экстракции элюированную ДНК концентрируют центрифугированием при 7500g с последующим осаждением этанолом и в конце растворяют в 5 мкл дистиллированной воды.
Полученную после каждой экстракции ДНК (H1 типа, H3 типа, H4 типа и M типа) клонируют с использованием вектора pGEM-T Easy (Promega) следующим образом:
Фрагмент ДНК, регенерированный из реакционной среды для ПЦР (Н1, Н3, Н4 или М типа), 5 мкл;
10× Taq-полимеразный буфер, 1 мкл;
смесь дНТФ, 1 мкл;
Taq-полимераза (5 единиц/мл), 1 мкл; и
повторно дистиллированная вода, добавляемая до конечного объема 10 мкл.
После того как каждый раствор прореагирует при 70°С в течение 30 минут, проводят лигирование каждого из растворов ДНК с вектором pGEM-T Easy с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2,0) (Takara Shuzo Co., Ltd.) по инструкции от производителя.
После 4-часовой инкубации при 15°C 2 мкл инкубированного реакционного раствора смешивают со 100 мкл компетентного штамма Е. coli JM109 c плотностью клеток (1-2)·109 клеток/мл (Takara Shuzo Co., Ltd.) и смесь выдерживают на льду в течение 30 минут, затем при 42°С в течение 30 секунд, и снова на льду в течение 1 минуты. К смеси добавляют 500 мкл среды SOC (2 об./об.% триптона, 0,5% (вес/об.) дрожжевого экстракта, 0,05% (вес/об.) хлорида натрия, 2,5 мМ (вес/об.) хлорида калия, 1 мМ хлорида магния и 20 мМ глюкозы) и все инкубируют еще в течение одного часа при встряхивании. Затем выделяют трансформированные штаммы и получают плазмидную ДНК из указанных штаммов, как описано в руководстве по молекулярному клонированию («Molecular Cloning A Laboratory Manual»). Нуклеотидную последовательность указанной ДНК, которая кодирует гуманизированную тяжелую цепь или мышиного TRA-8, подтверждают дидезоксиметодом соответственно (Sanger, F.S., et al. (1977), Proc. Natl. Acad. Sci. USA, 74:5463-5467) с использованием анализатора ДНК 3700 (ABI PRISM; Perkin Elmer Applied Biosystems, Japan).
Полученную плазмиду обозначают как pHА15 (плазмида, содержащая кДНК, кодирующую гуманизированную тяжелую цепь Н1 типа TRA-8), pHС10 (плазмида, содержащая кДНК, кодирующую гуманизированную тяжелую цепь Н3 типа TRA-8), pHD21 (плазмида, содержащая кДНК, кодирующую тяжелую цепь Н4 типа гуманизированного TRA-8) и рМ11 (плазмида, содержащая кДНК, кодирующую тяжелую цепь химерного TRA-8). Трансформированные штаммы E. coli, несущие указанные плазмиды, обозначенные как E. coli JM109/pHА15, E. coli JM109/pHC10, E. coli JM109/pHD21 и E. coli JM109/pМ11 соответственно, были депонированы в Международном депозитарии запатентованных организмов при Национальном Институте современной промышленной науки и технологии Японии (International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology, 1-1, Higashi 1 chome Tsukuba-shi, Ibaraki-ken, 305-5466) 20 апреля 2001 года в соответствии с Будапештским договором по депонированию микроорганизмов под номерами FERM-BP-7555, FERM-BP-7557, FERM-BP-7558 и FERM-BP-7559 соответственно.
(3) Конструирование плазмид экспрессии, содержащих вариабельный участок ДНК тяжелой цепи нескольких гуманизированного или мышиного TRA-8
Рекомбинантный вектор экспрессии для клеток животных конструируют путем вставки ДНК, кодирующей гуманизированную тяжелую цепь Н1 типа, Н3 типа и Н4 типа или М типа химерного TRA-8 (клонированного, как указано выше) указанным ниже способом.
Один мкл плазмиды pSRHHH3 (заявка на европейский патент EP 0909816 A1), содержащей вариабельный участок гуманизированной тяжелой цепи анти-Fas моноклонального антитела HFE7A и константный участок геномной ДНК человеческого IgG1, а также вектор экспрессии для клеток млекопитающих, расщепляют с использованием рестрикционных ферментов HindIII и ApaI и затем разделяют полученные фрагменты электрофорезом в 3% агарозном геле. После электрофореза гель окрашивают водным раствором бромида этидия с концентрацией 1 мкг/мл для выявления ДНК в УФ свете. Полосы векторной ДНК, содержащей константный участок геномной ДНК для человеческого IgG1 без вариабельного участка гуманизированной тяжелой цепи HFE7A вырезают лезвием и элюируют из геля с использованием набора Geneclean Spin. После фенольной экстракции элюированную ДНК концентрируют центрифугированием при 7500g с последующим осаждением этанолом и в конце растворяют в 5 мл дистиллированной воды, после чего подвергают дефосфорилированию с использованием CIP. Полученную расщепленную дефосфорилированную плазмиду (100 нг) лигируют с 1 мкг фрагмента ДНК pHA15, pHC10, pHD21 или pM11, содержащим ДНК, кодирующую вариабельный участок тяжелой цепи гуманизированного или химерного TRA-8, который уже был расщеплен HindIII и ApaI, с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2,0) (Takara Shuzo Co., Ltd.). Затем указанную лигирующую смесь используют для трансформации штамма E. coli JM109, который затем высевают на чашки с LB агаром, содержащим 50 мкг/мл ампициллина.
Трансформанты, полученные указанным способом, культивируют в 2 мл жидкой среды LB, содержащей 50 мкг/мл ампициллина, при 37°С в течение ночи и затем плазмидную ДНК экстрагируют из полученной культуры по методу, включающему использование щелочи и ДСН.
Экстрагированную плазмидную ДНК расщепляют HindIII и ApaI и подвергают электрофорезу в 3% (вес/об.) агарозном геле для подтверждения наличия или отсутствия вставки ДНК, кодирующей вариабельный участок тяжелой цепи гуманизированного или химерного TRA-8. Наличие вставки и ориентация желательного фрагмента ДНК в векторе подтверждается секвенированием ДНК с использованием анализатора генной последовательности (ABI Prism 3700 DNA Analyzer; Applied Biosystems). Полученные плазмиды экспрессии, содержащие кДНК, которая кодирует тяжелую цепь гуманизированного или химерного TRA-8, обозначают как pHA15-1, pHC10-3, pHD21-1 и pM11-1 соответственно.
(4) Конструирование векторов для гуманизированных легких цепей
(4.1) Конструирование вектора экспрессии для гуманизированной легкой цепи антитела (LM1 типа)
Как показано в SEQ ID No. 72 в перечне последовательностей, другие виды гуманизации (LM1 тип) аминокислотных последовательностей легкой цепи мышиного антитела TRA-8 против человеческого DR5 включают замену 3-ного аминокислотного остатка (валина), 8-ного аминокислотного остатка (гистидина), 9-ного аминокислотного остатка (лизина), 10-ного аминокислотного остатка (фенилаланина), 11-ного аминокислотного остатка (метионина), 13-ного аминокислотного остатка (треонина), 20-ного аминокислотного остатка (серина), 42-ного аминокислотного остатка (глютамина), 43-ного аминокислотного остатка (серина), 60-ного аминокислотного остатка (аспарагиновой кислоты), 63-ного аминокислотного остатка (треонина), 77-ного аминокислотного остатка (аспарагина), 78-ного аминокислотного остатка (валина), 80-ного аминокислотного остатка (серина), 83-ного аминокислотного остатка (лейцина), 85-ного аминокислотного остатка (аспарагиновой кислоты), 87-ного аминокислотного остатка (фенилаланина), 99-ного аминокислотного остатка (глицина), 103-ного аминокислотного остатка (лейцина) и 108-ного аминокислотного остатка (аланина) от N-конца аминокислотной последовательности легкой цепи TRA-8 глютамином, пролином, серином, серином, лейцином, аланином, треонином, лизином, аланином, серином, серином, серином, лейцином, пролином, фенилаланином, треонином, тирозином, глютамином, валином и треонином, соответственно. Полученная последовательность обозначена как LM1.
Плазмиду экспрессии, содержащую указанный тип аминокислотных последовательностей гуманизированной легкой цепи антитела TRA-8 против человеческого DR5 (LM1 типа, SEQ ID No. 72 в перечне последовательностей), конструируют приведенным ниже способом.
1) Синтез праймеров для получения вариабельного и константного участков гуманизированной легкой цепи TRA-8 (LM1 типа)
ДНК, кодирующую полипептидную цепь LM1 (SEQ ID No. 72 в перечне последовательностей), каждая из которых представляет собой слияние вариабельного участка гуманизированной легкой цепи антитела TRA-8 против человеческого DR5 (LM1 типа) и константного участка легкой цепи человеческого Ig (κ цепь), синтезируют соответственно с использованием подходящих сочетаний ПЦР.
Далее процедура включает к 7AL1P (SEQ ID No. 47), 7ALCN (SEQ ID No. 48), HKCDF11 (SEQ ID No. 50), HKCDR12 (SEQ ID No. 51), HKCDF22 (SEQ ID No. 52), HKCDR22 (SEQ ID No. 53) и HKCF12 (SEQ ID No. 54).
Для проведения ПЦР синтезируют приведенные ниже олигонуклеотидные праймеры:
5'-gtcccccaca gatgcagaca aagaacttgg agattgggc atctgaatgt caccagtgga-3' (HKSPR12; SEQ ID No. 77).
2) Конструирование плазмиды pCR3.1/LM1-2 (клонирование гуманизированной легкой цепи TRA-8 LM1 типа)
Фрагмент LM1-ДНК, кодирующий аминокислотную последовательность SEQ ID No. 72, получают при проведении 2-стадийной ПЦР, далее вставляют его в плазмидный вектор и клонируют в E. coli.
а) Первая стадия ПЦР
Фрагмент LM1-F1-ДНК, кодирующий сигнальную последовательность секреции и часть участка FRL1 с сайтом расщепления рестрикционным ферментом HindIII, добавленным к 5'-концу, получают в следующих условиях. Матричные плазмиды pHSGHM17 и pSRPDHH получают по способу, описанному в заявке на европейский патент EP 0909816 A1.
Реакционная среда имеет следующий состав:
плазмидная ДНК pHSGHM17, 25 нг;
олигонуклеотидный праймер 7AL1P, 50 пмоль;
олигонуклеотидный праймер HKSPR12, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq (Perkin Elmer), 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагмент LM1-F2-ДНК, кодирующий часть FRL1, CDRL1, FRL2 и CDRL2, получают в следующих условиях:
Реакционная смесь имеет следующий состав:
ДНК плазмиды pL28, 25 нг;
олигонуклеотидный праймер HKCDF11, 50 пмоль;
олигонуклеотидный праймер HKCDR12, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагмент LM1-F3-ДНК, кодирующий CDRL2, FRL3 и часть CDRL3, получают в следующих условиях:
Реакционная смесь имеет следующий состав:
ДНК плазмиды pSRPDHH, 25 нг;
олигонуклеотидный праймер HKCDF22, 50 пмоль;
олигонуклеотидный праймер HKCDR22, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагмент LM1-F4-ДНК, кодирующий CDRL3, FRL4 и константный участок с сайтом расщепления для рестрикционного фермента EcoRI, добавленным к 3'-концу, получают в следующих условиях:
Реакционная смесь имеет следующий состав:
ДНК плазмиды pSRPDHH, 25 нг;
олигонуклеотидный праймер HKCF12, 50 пмоль;
олигонуклеотидный праймер 7ALCN, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагменты амплифицированной ДНК разделяют электрофорезом в 5% полиакриламидном геле. После электрофореза гель окрашивают водным раствором бромида этидия с концентрацией 1 мкг/мл для выявления полученной ДНК в УФ свете. Сответствующие полосы ДНК, выявленные таким образом, вырезают лезвием.
b) Вторая стадия ПЦР
LM1-ДНК, в которой слиты указанные выше фрагменты LM1-F1-ДНК, LM1-F2-ДНК, LM1-F3-ДНК и LM1-F4-ДНК, получают в следующих условиях:
Реакционный раствор имеет следующий состав:
гелевый фрагмент LM1-F1-ДНК, полученный на первой стадии ПЦР,
гелевый фрагмент LM1-F2-ДНК, полученный на первой стадии ПЦР,
гелевый фрагмент LM1-F3-ДНК, полученный на первой стадии ПЦР,
гелевый фрагмент LM1-F4-ДНК, полученный на первой стадии ПЦР,
олигонуклеотидный праймер 7AL1P, 50 пмоль,
олигонуклеотидный праймер 7ALCN, 50 пмоль,
смесь дНТФ, 5,0 мкл,
10× буфер для ПЦР, 5,0 мкл,
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Полученный таким образом фрагмент LM1-ДНК вставляют в плазмидную ДНК pCR3.1 с использованием набора для клонирования эукариотической ТА (InVitrogen) по инструкции от производителя и вводят в компетентные клетки E. coli TOP10F', входящие в состав набора. Нуклеотидные последовательности указанных ДНК, которые кодируют гуманизированную легкую цепь TRA-8 (LM1 типа), подтверждают дидезоксиметодом (Sanger, F. S., et al. (1977), Proc. Natl. Acad. Sci. USA, 74:5463-5467) с использованием анализатора ДНК (3700 DNA Analyzer, ABI PRISM; Perkin Elmer Applied Biosystems, Japan).
Полученную плазмиду обозначают как pCR3.1/LM1-2 (плазмида, несущая кДНК, кодирующую вариабельный участок гуманизированной легкой цепи TRA-8 (LM1 типа) и константный участок легкой цепи человеческого Ig).
Полученную плазмиду pCR3.1/LМ1-2, содержащую фрагмент LM1-ДНК, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pHSG399 расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Полученные дефосфорилированная ДНК pHSG399 и фрагмент LM1-ДНК, которые были расщеплены рестрикционными ферментами Hind III и EcoR I, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем клетки E. coli DH5α трансформируют лигированной ДНК и распределяют по агаровой среде LB, содержащей 0,1 мМ IPTG, 0,1% X-Gal и 50 мкг/мл хлорамфеникола (конечные концентрации). Полученные белые трансформанты культивируют в жидкой среде LB, содержащей 50 мкг/мл хлорамфеникола, и плазмидную ДНК экстрагируют из полученных культур по методу, включающему обработку щелочью и ДСН. Экстрагированную плазмидную ДНК расщепляют с использованием Hind III и EcoR I и затем клон, содержащий фрагмент LM1-ДНК, выделяют электрофорезом в 1% геле агарозы.
В результате проведенной процедуры получают плазмиду pHSG/M1-2-2, содержащую фрагмент слияния вариабельного участка гуманизированной легкой цепи TRA-8 типа LM1 и константного участка человеческой цепи человеческого Igκ. Трансформированный штамм E. coli, несущий указанную плазмиду, обозначенный как E. coli DH5α/pHSG/M1-2-2, был депонирован в Международном депозитарии запатентованных организмов при Национальном Институте современной промышленной науки и технологии Японии (International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology, 1-1, Higashi 1 chome Tsukuba-shi, Ibaraki-ken, 305-5466) 20 апреля 2001 года в соответствии с Будапештским договором по депонированию микроорганизмов под номером FERM-BP-7352.
3) Конструирование плазмиды pSR/LM1-2 (плазмида экспрессии гуманизированной легкой цепи TRA-8 типа LM1)
Полученную плазмиду pHSG/M1-2, содержащую фрагмент слияния вариабельного участка гуманизированной легкой цепи TRA-8 типа LM1 и константного участка цепи человеческого Igκ, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pSRPDHH (заявка на европейский патент EP 0909816 A1) расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Образованные дефосфорилированная ДНК pSRPDHH и фрагмент ДНК Hind III-EcoR I, полученный из pHSG/M1-2-2, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем проводят трансформацию клеток E. coli DH5α лигированной ДНК и распределяют по поверхности LB агара. Полученные трансформанты культивируют в жидкой среде LB, содержащей 100 мкг/мл ампициллина, и из клеток полученной культуры экстрагируют плазмидную ДНК по методу, включающему обработку щелочью и ДСН. Наличие вставки и ориентацию желательного фрагмента ДНК в векторе pSRPDHH подтверждают секвенированием с использованием анализатора генной последовательности.
Полученную плазмиду экспрессии, несущую кДНК, которая кодирует гуманизированную легкую цепь TRA-8 типа LM1, обозначают как pSR/LM1-2.
(4.2) Конструирование вектора экспрессии гуманизированной легкой цепи антитела (LM3 типа)
Как показано в SEQ ID No. 73 в перечне последовательностей, гуманизация (LM3 типа) аминокислотных последовательностей легкой цепи мышиного антитела TRA-8 против человеческого DR5, включает замену 8-ного аминокислотного остатка (гистидина), 9-ного аминокислотного остатка (лизина), 10-ного аминокислотного остатка (фенилаланина), 11-ного аминокислотного остатка (метионина), 13-ного аминокислотного остатка (треонина), 20-ного аминокислотного остатка (серина), 42-ного аминокислотного остатка (глютамина), 43-ного аминокислотного остатка (серина), 77-ного аминокислотного остатка (аспарагина), 78-ного аминокислотного остатка (валина), 80-ного аминокислотного остатка (серина), 83-ного аминокислотного остатка (лейцина), 85-ного аминокислотного остатка (аспарагиновой кислоты), 87-ного аминокислотного остатка (фенилаланина), 99-ного аминокислотного остатка (глицина), 103-ного аминокислотного остатка (лейцина) и 108-ного аминокислотного остатка (аланина) на N-конце аминокислотной последовательности легкой цепи TRA-8 пролином, серином, серином, лейцином, аланином, треонином, лизином, аланином, серином, лейцином, пролином, фенилаланином, треонином, тирозином, глютамином, валином и треонином соответственно. Полученная последовательность была обозначена как LM3.
Плазмиды экспрессии, несущие указанный тип аминокислотных последовательностей гуманизированной легкой цепи антитела TRA-8 (LM3 типа, SEQ ID No. 73 в перечне последовательностей) против человеческого DR5 конструируют указанным ниже способом.
(1) Синтез праймеров для получения вариабельного и константного участков гуманизированной легкой цепи TRA-8 LM3 типа
ДНК, кодирующую полипептидную цепь LM3 (SEQ ID No. 73 в перечне последовательностей), каждая из которых представляет собой слияние вариабельного участка гуманизированной легкой цепи антитела TRA-8 против DR5 и константного участка легкой цепи человеческого Ig (цепь κ), синтезируют с использованием соответствующих комбинаций ПЦР.
Далее к 7AL1P (SEQ ID No. 47) и 7ALCN (SEQ ID No. 48) процедура включает синтез следующих нуклеотидных праймеров для проведения ПЦР:
5'-atctagttct cagagatgga gacagacaca atcctgctat gggtgctgct gctctgggtt ccagg-3' (MOD1F1; SEQ ID No. 78);
5'-cagcacccat agcaggattg tgtctgtctc catctctgag aactagatga gaggatgctt cttaagctt-3' (MOD1R1; SEQ ID No. 79);
5'-ctccactggt gacattgtga tgacccaatc tccaagttct ttgtctgcat ctgtggggga cagggtc-3' (MOD1F22; SEQ ID No. 80);
5'-acttggagat tgggtcatca caatgtcacc agtggagcct ggaacccaga gcag-3' (MOD1R22; SEQ ID No. 81);
5'-accatcacct gcaaggccag tcaggatgtg ggtactgctg tagcctggta ccaacagaaa ccaggaa-3' (MOD1F3; SEQ ID No.82);
5'-tacagcagta cccacatcct gactggcctt gcaggtgatg gtgaccctgt cccccacaga tgcagacaaa ga-3' (MOD1R3; SEQ ID No. 83);
5'-aagcacccaa actcctcatc tattgggcat ccacccggca cactggggtc ccagataggt ttacaggcag t-3' (MOD1F42; SEQ ID No. 84);
5'-cccagtgtgc cgggtggatg cccaatagat gaggagtttg ggtgcttttc ctggtttctg ttggtaccag gc-3' (MOD1R4; SEQ ID No. 85);
5'-gggtctggga cagacttcac cctcaccatc tctagtctgc agccggagga ttttgcaacc tat-3'(MOD1F5; SEQ ID No.86);
5'-actagagatg gtgagggtga agtctgtccc agacccactg cctgtaaacc tatctgggac-3' (MOD1R52; SEQ ID No. 87);
5'-tactgtcagc aatatagcag ctatcggacg ttcggtcaag gcaccaaggt ggaaatc-3' (MOD1F6; SEQ ID No. 88);
5'-cgtccgatag ctgctatatt gctgacagta ataggttgca aaatcctccg gctgcac-3' (MOD1R6; SEQ ID No. 89)
5'-aaacggactg tggctgcacc atctgtcttc atcttcccgc catctgatga g-3' (MOD1F7; SEQ ID No. 90);
5'-gaagatgaag acagatggtg cagccacagt ccgtttgatt tccaccttgg tgccttgacc gaa-3' (MOD1R7; SEQ ID No. 91); и
5'-agatttcaac tgctcatcag atggcgggaa (LR17; SEQ ID No. 101).
2) Конструирование плазмиды pCR3.1/LM3-3-44 (клонирование гуманизированной легкой цепи TRA-8 LM3 типа)
Фрагмент LM3-ДНК, кодирующий аминокислотную последовательность SEQ ID No. 73, получают при проведении 2-стадийной ПЦР, затем вставляют в плазмидный вектор и клонируют в E. coli.
а) Первая стадия ПЦР
Фрагмент LM3-F31В-ДНК, кодирующий участок сигнальной последовательности с сайтом расщепления рестрикционным ферментом Hind III, добавленным к 5'-концу, FRL1, CDRL1, FRL2 и CDRL2, FRL3, CDRL3 FRL4 и часть константного участка, получают в следующих условиях.
Реакционная среда имеет следующий состав:
олигонуклеотидный праймер MOD1F1, 5 пмоль;
олигонуклеотидный праймер MOD1R1, 5 пмоль;
олигонуклеотидный праймер MOD1F22, 5 пмоль;
олигонуклеотидный праймер MOD1R22, 5 пмоль;
олигонуклеотидный праймер MOD1F3, 5 пмоль;
олигонуклеотидный праймер MOD1R3, 5 пмоль;
олигонуклеотидный праймер MOD1F42, 5 пмоль;
олигонуклеотидный праймер MOD1R4, 5 пмоль;
олигонуклеотидный праймер MOD1F5, 5 пмоль;
олигонуклеотидный праймер MOD1R52, 5 пмоль;
олигонуклеотидный праймер MOD1F6, 5 пмоль;
олигонуклеотидный праймер MOD1R6, 5 пмоль;
олигонуклеотидный праймер MOD1F7, 50 пмоль;
олигонуклеотидный праймер MOD1R7, 5 пмоль;
олигонуклеотидный праймер 7AL1P, 50 пмоль;
олигонуклеотидный праймер LR17, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды с последующим использованием в ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагмент LM3-F31C-ДНК, кодирующий части константного участка с сайтом расщепления рестрикционным ферментом EcoRI, добавленным на 3'-конце, получают в следующих условиях:
Матричные плазмиды pSRPDHH получают в соответствии с описанием, приведенным в заявке на европейский патент ЕР 0909816А1.
Реакционная смесь имеет следующий состав:
ДНК плазмиды pSRPDHH, 25 нг;
олигонуклеотидный праймер MOD1F7, 50 пмоль;
олигонуклеотидный праймер 7ALCN, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагменты амплифицированной ДНК, полученные после проведения ПЦР, разделяют электрофорезом в 5% полиакриламидном геле. Гель после электрофореза окрашивают при добавлении 1 мкг/мл бромида этидия для выявления полученной ДНК в УФ свете. Выявленные при этом соответствующие полосы ДНК вырезают лезвием.
b) Вторая стадия ПЦР
LM3-ДНК, в которой слиты указанные выше фрагменты LM3-F31B-ДНК и LM3-F31C-ДНК, получают в следующих условиях:
Реакционный раствор имеет следующий состав:
гелевый фрагмент LM3-F31В-ДНК, полученный на первой стадии ПЦР,
гелевый фрагмент LM3-F31С-ДНК, полученный на первой стадии ПЦР,
олигонуклеотидный праймер 7AL1P, 50 пмоль,
олигонуклеотидный праймер 7ALCN, 50 пмоль,
смесь дНТФ, 5,0 мкл,
10× буфер для ПЦР, 5,0 мкл,
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Полученный таким образом фрагмент LM3-ДНК вставляют в плазмидную ДНК pCR3.1 с использованием набора для клонирования эукариотической ТА (InVitrogen) по инструкции от производителя и вводят в компетентные клетки E. coli TOP10F', входящие в состав набора. Нуклеотидные последовательности указанных ДНК, кодирующих гуманизированную легкую цепь TRA-8 LM3 типа, подтверждают дидезоксиметодом (Sanger, F. S., et al. (1977), Proc. Natl. Acad. Sci. USA, 74:5463-5467) с использованием анализатора ДНК (3700 DNA Analyzer; ABI PRISM; Perkin Elmer Applied Biosystems, Japan).
Полученную плазмиду обозначают как pCR3.1/LM1-3-3-44 (плазмида, несущая кДНК, кодирующую вариабельный участок гуманизированной легкой цепи TRA-8 LM3 типа и константный участок легкой цепи человеческого Ig).
Полученную плазмиду pCR3.1/LМ3-3-44, содержащую фрагмент LM3-ДНК, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pHSG399 расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Полученные дефосфорилированная ДНК pHSG399 и фрагмент LM3-ДНК, которые были подвергнуты расщеплению рестрикционными ферментами Hind III и EcoR I, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем клетки E. coli DH5α трансформируют лигированной ДНК и распределяют клетки по агаровой среде LB, содержащей 0,1 мМ IPTG, 0,1% X-Gal и 50 мкг/мл хлорамфеникола (конечные концентрации). Полученные белые трансформанты культивируют в жидкой среде LB, содержащей 50 мкг/мл хлорамфеникола, и плазмидную ДНК экстрагируют из полученных культур по методу, включающему обработку щелочью и ДСН. Экстрагированную плазмидную ДНК расщепляют с использованием Hind III и EcoR I и затем клон, содержащий фрагмент LM3-ДН, выделяют электрофорезом в 1% геле агарозы.
В результате проведенной выше процедуры получают плазмиду pHSG/M3-3-22, содержащую фрагмент слияния вариабельного участка гуманизированной легкой цепи TRA-8 типа LM3 и константного участка цепи человеческого Igκ. Трансформированный штамм E. coli, несущий указанную плазмиду, обозначенный как E. coli DH5α/pHSG/M3-3-22, депонируют в Международном депозитарии запатентованных организмов при Национальном Институте современной промышленной науки и технологии Японии (International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology, 1-1, Higashi 1 chome Tsukuba-shi, Ibaraki-ken, 305-5466) 20 апреля 2001 года в соответствии с Будапештским договором по депонированию микроорганизмов под номером FERM-BP-7564.
3) Конструирование плазмиды pSR/LM3-3-44-10 (плазмида экспрессии гуманизированной легкой цепи TRA-8 LM3 типа)
Полученную плазмиду pHSG/M3-3-22, содержащую фрагмент слияния вариабельного участка гуманизированной легкой цепи TRA-8 LM3 типа и константного участка цепи человеческого Igκ, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pSRPDHH (заявка на Европейский патент EP 0909816 A1) расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Образованные дефосфорилированная ДНК pSRPDHH и фрагмент ДНК Hind III-EcoR I, полученный из pHSG/M3-3-22, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем проводят трансформацию клеток E. coli DH5α лигированной ДНК и распределяют их по поверхности LB агара. Полученные трансформанты культивируют в жидкой среде LB, содержащей 100 мкг/мл ампициллина, и из полученной культуры экстрагируют плазмидную ДНК по методу, включающему обработку щелочью и ДСН. Наличие вставки и ориентацию желательного фрагмента ДНК в векторе pSRPDHH определяют секвенированием ДНК с использованием анализатора генной последовательности (ABI, Prism 3700 DNA Analyzar; Applied Biosystems).
Полученную плазмиду экспрессии, несущую кДНК, кодирующую гуманизированную легкую цепь TRA-8 LM3 типа, обозначают как pSR/LM3-3-44-10.
(4.3) Конструирование вектора экспрессии легкой цепи гуманизированного антитела (LM4 типа)
Как показано в SEQ ID No. 74 в перечне последовательностей, другой вид гуманизации (LM4 типа) аминокислотных последовательностей легкой цепи мышиного антитела TRA-8 против человеческого DR5, включает замену 8-ного аминокислотного остатка (гистидина), 9-ного аминокислотного остатка (лизина), 10-ного аминокислотного остатка (фенилаланина), 11-ного аминокислотного остатка (метионина), 13-ного аминокислотного остатка (треонина), 20-ного аминокислотного остатка (серина), 42-ного аминокислотного остатка (глютамина), 43-ного аминокислотного остатка (серина), 77-ного аминокислотного остатка (аспарагина), 78-ного аминокислотного остатка (валина), 80-ного аминокислотного остатка (серина), 83-ного аминокислотного остатка (лейцина), 85-ного аминокислотного остатка (аспарагиновой кислоты), 99-ного аминокислотного остатка (глицина), 103-ного аминокислотного остатка (лейцина) и 108-ного аминокислотного остатка (аланина) на N-конце аминокислотной последовательности легкой цепи TRA-8 пролином, серином, серином, лейцином, аланином, треонином, лизином, аланином, серином, лейцином, пролином, фенилаланином, треонином, глютамином, валином и треонином соответственно. Полученная последовательность была обозначена как LM4.
Плазмиды экспрессии, несущие указанный тип аминокислотных последовательностей гуманизированной легкой цепи антитела TRA-8 против человеческого DR5 (LM4 типа) (SEQ ID No. 74 в перечне последовательностей), конструируют указанным ниже способом.
(1) Синтез праймеров для получения вариабельного и константного участков гуманизированной легкой цепи TRA-8 типа LM4
ДНК, кодирующую полипептидную цепь LM4 (SEQ ID No. 74) в перечне последовательностей), каждая из которых представляет собой слияние вариабельного участка гуманизированной легкой цепи антитела TRA-8 против DR5 и константного участка легкой цепи человеческого Ig (цепь κ), синтезируют с использованием соответствующих комбинаций ПЦР.
Далее к 7AL1P (SEQ ID No. 47), 7ALCN (SEQ ID No. 48), MOD1F1 (SEQ ID No. 78), MOD1R1 (SEQ ID No. 79), MOD1F22 (SEQ ID No. 80), MOD1R22 (SEQ ID No. 81), MOD1F3 (SEQ ID No. 82), MOD1R3 (SEQ ID No. 83), MOD1F42 (SEQ ID No. 84), MOD1R4 (SEQ ID No. 85), MOD1F5 (SEQ ID No. 86), MOD1R52 (SEQ ID No. 87), MOD1F7 (SEQ ID No. 90) и MOD1R7 (SEQ ID No. 91), LR17 (SEQ ID No. 101) синтезируют следующие олигонуклеотидные праймеры для проведения ПЦР:
5′-ttctgtcagc aatatagcag ctatcggacg ttcggtcaag gcaccaaggt ggaaatc-3′ (MOD1F62; SEQ ID No.92);
5′-cgtccgatag ctgctatatt gctgacagaa ataggttgca aaatcctccg gctgcag-3′ (MOD1R62; SEQ ID No.93).
(2) Конструирование плазмиды pCR3.1/LM4-5-3 (клонирование гуманизированной легкой цепи TRA-8 типа LM4)
Фрагмент LM4-ДНК, кодирующей аминокислотную последовательность SEQ ID No. 74, получают при проведении 2-стадийной ПЦР, с последующей вставкой в плазмидный вектор и клонированием в E. coli.
а) Первая стадия ПЦР
Фрагмент LM4-F41В-ДНК, кодирующий участок сигнальной последовательности с сайтом расщепления рестрикционным ферментом Hind III, добавленным к 5'-концу, FRL1, CDRL1, FRL2, CDRL2, FRL3, CDRL3, FRL4 и часть константного участка получают в следующих условиях.
Реакционная среда имеет следующий состав:
олигонуклеотидный праймер MOD1F1, 5 пмоль;
олигонуклеотидный праймер MOD1R1, 5 пмоль;
олигонуклеотидный праймер MOD1F22, 5 пмоль;
олигонуклеотидный праймер MOD1R22, 5 пмоль;
олигонуклеотидный праймер MOD1F3, 5 пмоль;
олигонуклеотидный праймер MOD1R3, 5 пмоль;
олигонуклеотидный праймер MOD1F42, 5 пмоль;
олигонуклеотидный праймер MOD1R4, 5 пмоль;
олигонуклеотидный праймер MOD1F5, 5 пмоль;
олигонуклеотидный праймер MOD1R52, 5 пмоль;
олигонуклеотидный праймер MOD1F62, 5 пмоль;
олигонуклеотидный праймер MOD1R62, 5 пмоль;
олигонуклеотидный праймер MOD1F7, 50 пмоль;
олигонуклеотидный праймер MOD1R7, 5 пмоль;
олигонуклеотидный праймер 7AL1P, 50 пмоль;
олигонуклеотидный праймер LR17, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторной дистиллированной воды с последующим использованием в ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагмент LM4-F41C-ДНК, кодирующий часть константного участка с сайтом расщепления рестрикционным ферментом EcoRI, добавленным на 3'-конце, получают в следующих условиях:
Матричные плазмиды pSRPDHH получают в соответствии с описанием, приведенным в заявке на европейский патент ЕР 0909816 А1.
Реакционная смесь имеет следующий состав:
ДНК плазмиды pSRPDHH, 25 нг;
олигонуклеотидный праймер MOD1F7, 50 пмоль;
олигонуклеотидный праймер 7ALCN, 50 пмоль;
смесь дНТФ, 5 мкл;
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единиц.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагменты амплифицированной ДНК, полученные после проведения ПЦР, разделяют электрофорезом в 5% полиакриламидном геле. Гель после электрофореза окрашивают при добавлении 1 мкг/мл бромида этидия для выявления полученной ДНК в УФ свете. Выявленные при этом соответствующие полосы ДНК вырезают лезвием.
b) Вторая стадия ПЦР
LM4-ДНК, в которой слиты указанные выше фрагменты LM4-F41B-ДНК и LM4-F31C-ДНК, получают в следующих условиях:
Реакционный раствор имеет следующий состав:
гелевый фрагмент LM4-F41В-ДНК, полученный на первой стадии ПЦР,
гелевый фрагмент LM4-F41С-ДНК, полученный на первой стадии ПЦР,
олигонуклеотидный праймер 7AL1P, 50 пмоль,
олигонуклеотидный праймер 7ALCN, 50 пмоль,
смесь дНТФ, 5,0 мкл,
10× буфер для ПЦР, 5,0 мкл,
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Полученный таким образом фрагмент LM4-ДНК вставляют в ДНК плазмиды pCR3.1 с использованием набора для клонирования эукариотической ТА (InVitrogen) по инструкции от производителя и вводят в компетентные клетки E. coli TOP10F', входящие в состав набора. Нуклеотидные последовательности указанных ДНК, которые кодируют гуманизированную легкую цепь TRA-8 типа LM4, подтверждают дидезоксиметодом (Sanger, F. S., et al. (1977), Proc. Natl. Acad. Sci. USA, 74:5463-5467) с использованием анализатора ДНК (3700 DNA Analyzer; ABI PRISM; Perkin Elmer Applied Biosystems, Japan).
Полученную плазмиду обозначают как pCR3.1/LM4-5-3 (плазмида, несущая кДНК, кодирующую вариабельный участок гуманизированной легкой цепи TRA-8 типа LM4 и константный участок легкой цепи человеческого Ig).
Полученную плазмиду pCR3.1/LМ4-5-3, содержащую фрагмент LM4-ДНК, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pHSG399 расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Полученные дефосфорилированная ДНК pHSG399 и фрагмент LM4-ДНК, которые были подвергнуты расщеплению рестрикционными ферментами Hind III и EcoR I, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем клетки E. coli DH5α трансформируют лигированной ДНК и распределяют их по агаровой среде LB, содержащей 0,1 мМ IPTG, 0,1% X-Gal и 50 мкг/мл хлорамфеникола (конечные концентрации). Полученные белые трансформанты культивируют в жидкой среде LB, содержащей 50 мкг/мл хлорамфеникола, и плазмидную ДНК экстрагируют из полученных культур по методу, включающему обработку щелочью и ДСН. Экстрагированную плазмидную ДНК расщепляют с использованием Hind III и EcoR I и затем клон, содержащий фрагмент LM4-ДНК, выделяют электрофорезом в 1% геле агарозы.
В результате проведенной процедуры получают плазмиду pHSG/M4-5-3-1, содержащую фрагмент слияния вариабельного участка гуманизированной легкой цепи TRA-8 типа LM4 и константного участка цепи человеческого Igκ. Трансформированный штамм E. coli, несущий указанную плазмиду, обозначенный как E. coli DH5α/pHSG/M4-5-3-1, был депонирован в Международном депозитарии запатентованных организмов при Национальном Институте современной промышленной науки и технологии Японии (International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology, 1-1, Higashi 1 chome Tsukuba-shi, Ibaraki-ken, 305-5466) 20 апреля 2001 года в соответствии с Будапештским договором по депонированию микроорганизмов под номером FERM-BP-7565.
3) Конструирование плазмиды pSR/LM4-5-3-3 (плазмида экспрессии гуманизированной легкой цепи TRA-8 типа LM4)
Полученную плазмиду pHSG/M4-5-3-1, содержащую фрагмент слияния вариабельного участка гуманизированной легкой цепи TRA-8 LM4 типа и константного участка цепи человеческого Igκ, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pSRPDHH (заявка на европейский патент EP 0909816 A1) расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Образованные дефосфорилированная ДНК pSRPDHH и фрагмент ДНК Hind III-EcoR I, полученный из pHSG/M4-5-3-1, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем проводят трансформацию клеток E. coli DH5α лигированной ДНК и распределяют по поверхности LB агара. Полученные трансформанты культивируют в жидкой среде LB, содержащей 100 мкг/мл ампициллина, и из клеток полученной культуры экстрагируют плазмидную ДНК по методу, включающему обработку щелочью и ДСН. Наличие вставки и ориентацию желательного фрагмента ДНК в векторе pSRPDHH определяют секвенированием с использованием анализатора генной последовательности (ABI Prism 3700 DNA Analyzer; Applied Biosystems).
Полученная плазмида экспрессии, несущая кДНК, которая кодирует гуманизированную легкую цепь TRA-8 LM4 типа, была обозначена как pSR/LM4-5-3-3.
(4.4) Конструирование вектора экспрессии легкой цепи гуманизированного антитела (LM5 типа)
Как показано в SEQ ID No. 75 в перечне последовательностей, другой вид гуманизации (LM5 типа) аминокислотных последовательностей легкой цепи мышиного антитела TRA-8 против человеческого DR5 включают замещение 8-ного аминокислотного остатка (гистидина), 9-ного аминокислотного остатка (лизина), 10-ного аминокислотного остатка (фенилаланина), 11-ного аминокислотного остатка (метионина), 13-ного аминокислотного остатка (треонина), 20-ного аминокислотного остатка (серина), 42-ного аминокислотного остатка (глютамина), 43-ного аминокислотного остатка (серина), 77-ного аминокислотного остатка (аспарагина), 78-ного аминокислотного остатка (валина), 80-ного аминокислотного остатка (серина), 83-ного аминокислотного остатка (лейцина), 103-ного аминокислотного остатка (лейцина) и 108-ного аминокислотного остатка (аланина) на N-конце аминокислотной последовательности легкой цепи TRA-8 пролином, серином, серином, лейцином, аланином, треонином, лизином, аланином, серином, лейцином, пролином, фенилаланином, валином и треонином соответственно. Полученная последовательность была обозначена как LM5.
Плазмиды экспрессии, несущие указанный тип аминокислотных последовательностей гуманизированной легкой цепи антитела TRA-8 против человеческого DR5 (LM5 типа) (SEQ ID No. 75 в перечне последовательностей), конструируют следующим образом.
(1) Синтез праймеров для получения вариабельного и константного участков гуманизированной легкой цепи TRA-8 LM5 типа
ДНК, кодирующую полипептидную цепь LM5 (SEQ ID No. 75 в перечне последовательностей), каждая из которых представляет собой слияние вариабельного участка гуманизированной легкой цепи антитела TRA-8 против DR5 и константного участка легкой цепи человеческого Ig (цепь κ), синтезируют с использованием соответствующих комбинаций ПЦР.
Далее к 7AL1P (SEQ ID No. 47), 7ALCN (SEQ ID No. 48), MOD1F1 (SEQ ID No. 78), MOD1R1 (SEQ ID No. 79), MOD1F22 (SEQ ID No. 80), MOD1R22 (SEQ ID No. 81), MOD1F3 (SEQ ID No. 82), MOD1R3 (SEQ ID No. 83), MOD1F42 (SEQ ID No. 84), MOD1R4 (SEQ ID No. 85), MOD1R52 (SEQ ID No. 87), MOD1F7 (SEQ ID No. 90) и LR17 (SEQ ID No. 101) синтезируют следующие олигонуклеотидные праймеры для проведения ПЦР:

(2) Конструирование плазмиды pCR3.1/LM5-3-42 (клонирование гуманизированной легкой цепи TRA-8 LM5 типа)
Фрагмент LM5-ДНК, кодирующей аминокислотную последовательность SEQ ID No. 75, получают при проведении 2-стадийной ПЦР, с последующей вставкой в плазмидный вектор и клонированием в E. coli.
а) Первая стадия ПЦР
Фрагмент LM5-F51В-ДНК, кодирующий участок сигнальной последовательности с сайтом расщепления рестрикционным ферментом Hind III, добавленным к 5'-концу, FRL1, CDRL1, FRL2, CDRL2, FRL3, CDRL3, FRL4 и часть константного участка, получают в следующих условиях.
Реакционная среда имеет следующий состав:
олигонуклеотидный праймер MOD1F1, 5 пмоль
олигонуклеотидный праймер MOD1R1, 5 пмоль
олигонуклеотидный праймер MOD1F22, 5 пмоль
олигонуклеотидный праймер MOD1R22, 5 пмоль
олигонуклеотидный праймер MOD1F3, 5 пмоль
олигонуклеотидный праймер MOD1R3, 5 пмоль
олигонуклеотидный праймер MOD1F42, 5 пмоль
олигонуклеотидный праймер MOD1R4, 5 пмоль
олигонуклеотидный праймер MOD1F52, 5 пмоль
олигонуклеотидный праймер MOD1R52, 5 пмоль
олигонуклеотидный праймер MOD1F63, 5 пмоль
олигонуклеотидный праймер MOD1R63, 5 пмоль
олигонуклеотидный праймер MOD1F7, 50 пмоль
олигонуклеотидный праймер MOD1R72, 5 пмоль
олигонуклеотидный праймер 7AL1P, 50 пмоль
олигонуклеотидный праймер LR17, 50 пмоль
смесь дНТФ, 5 мкл
10× буфер для ПЦР, 5 мкл
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторной дистиллированной воды с последующим использованием в ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагмент LM5-F51C-ДНК, кодирующий часть константного участка с сайтом расщепления рестрикционным ферментом EcoRI, добавленным на 3'-конце, получают в следующих условиях. Матричные плазмиды pSRPDHH получают в соответствии с описанием, приведенным в заявке на европейский патент ЕР 0909816 А1.
Реакционная смесь имеет следующий состав:
ДНК плазмиды pSRPDHH, 25 нг
олигонуклеотидный праймер MOD1F7, 50 пмоль
олигонуклеотидный праймер 7ALCN, 50 пмоль
смесь дНТФ, 5 мкл
10× буфер для ПЦР, 5 мкл
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагменты амплифицированной ДНК, полученные после проведения ПЦР, разделяют электрофорезом в 5% полиакриламидном геле. Гель после электрофореза окрашивают при добавлении 1 мкг/мл бромида этидия для выявления полученной ДНК в УФ свете. Выявленные при этом соответствующие полосы ДНК вырезают лезвием.
b) Вторая стадия ПЦР
LM5-ДНК, в которой слиты указанные выше фрагменты LM5-F51B-ДНК и LM5-F51C-ДНК, получают в следующих условиях:
Реакционный раствор имеет следующий состав:
гелевый фрагмент LM5-F51В-ДНК, полученный на первой стадии ПЦР,
гелевый фрагмент LM5-F51С-ДНК, полученный на первой стадии ПЦР,
олигонуклеотидный праймер 7AL1P, 50 пмоль
олигонуклеотидный праймер 7ALCN, 50 пмоль
смесь дНТФ, 5,0 мкл
10× буфер для ПЦР, 5,0 мкл
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Полученный таким образом фрагмент LM5-ДНК вставляют в плазмидную ДНК pCR3.1 с использованием набора для клонирования эукариотической ТА (InVitrogen) по инструкции от производителя и вводят в компетентные клетки E. coli TOP10F', входящие в состав набора. Нуклеотидные последовательности указанных ДНК, которые кодируют гуманизированную легкую цепь TRA-8 LM5 типа, подтверждают дидезоксиметодом (Sanger, F. S., et al. (1977), Proc. Natl. Acad. Sci. USA, 74:5463-5467) с использованием анализатора ДНК.
Полученную плазмиду обозначают как pCR3.1/LM5-3-42 (плазмида, несущая кДНК, кодирующую вариабельный участок гуманизированной легкой цепи TRA-8 LM5 типа и константный участок легкой цепи человеческого Ig).
Полученную плазмиду pCR3.1/LМ5-3-42, содержащую фрагмент LM3-ДНК, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pHSG399 расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Полученные дефосфорилированная ДНК pHSG399 и фрагмент LM5-ДНК, которые были подвергнуты расщеплению рестрикционными ферментами Hind III и EcoR I, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем клетки E. coli DH5α трансформируют лигированной ДНК и распределяют по агаровой среде LB, содержащей 0,1 мМ IPTG, 0,1% X-Gal и 50 мкг/мл хлорамфеникола (конечные концентрации). Полученные белые трансформанты культивируют в жидкой среде LB, содержащей 50 мкг/мл хлорамфеникола, и плазмидную ДНК экстрагируют из клеток полученных культур по методу, включающему обработку щелочью и ДСН. Экстрагированную плазмидную ДНК расщепляют с использованием Hind III и EcoR I и затем клон, содержащий фрагмент LM5-ДНК, отбирают с помощью электрофореза в 1% геле агарозы.
В результате проведенной процедуры получают плазмиду pHSG/M5-3-27, содержащую фрагмент слияния вариабельного участка гуманизированной легкой цепи TRA-8 LM5 типа и константного участка цепи человеческого Igκ.
(3) Конструирование плазмиды pSR/LM5-3-27-1 (плазмида экспрессии гуманизированной легкой цепи TRA-8 LM5 типа)
Полученную плазмиду pHSG/M5-3-27, несущую фрагмент слияния вариабельного участка гуманизированной легкой цепи TRA-8 LM5 типа и константного участка цепи человеческого Igκ, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pSRPDHH (заявка на Европейский патент EP 0909816 A1) расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Образованные дефосфорилированная ДНК pSRPDHH и фрагмент ДНК Hind III-EcoR I, полученный из pHSG/M5-3-27, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем проводят трансформацию клеток E. coli DH5α лигированной ДНК и распределяют их по поверхности LB агара. Полученные трансформанты культивируют в жидкой среде LB, содержащей 100 мкг/мл ампициллина, и из клеток полученной культуры экстрагируют плазмидную ДНК по методу, включающему обработку щелочью и ДСН. Наличие вставки и ориентацию желательного фрагмента ДНК в векторе pSRPDHH определяют секвенированием с использованием анализатора генной последовательности (ABI Prism 3700 DNA Analyzer; Applied Biosystems).
Полученная плазмида экспрессии, несущая кДНК, кодирующую гуманизированную легкую цепь TRA-8 LM5 типа, была обозначена как pSR/LM5-3-27-1.
(4.5) Конструирование вектора экспрессии гуманизированной легкой цепи антитела (химерного типа)
Последовательность, приведенная в SEQ ID No. 76 в перечне последовательностей, представляющая собой аминокислотную последовательность легкой цепи TRA-8 химерного типа, была обозначена как LM6.
Плазмиды экспрессии, несущие указанный тип аминокислотных последовательностей гуманизированной легкой цепи антитела TRA-8 против человеческого DR5 (LM6 типа) (SEQ ID No. 75 в перечне последовательностей), конструируют следующим образом.
1) Синтез праймеров для получения вариабельного и константного участков гуманизированной легкой цепи TRA-8 LM6 типа
ДНК, кодирующая полипептидную цепь LM6 (SEQ ID No. 75 в перечне последовательностей), каждая из которых представляет собой слияние вариабельного участка легкой цепи мышиного антитела TRA-8 против DR5 (LM6 типа) и константного участка легкой цепи человеческого Ig (цепь κ), синтезируют с использованием соответствующих комбинаций ПЦР.
Далее к 7AL1P (SEQ ID No. 47) и 7ALCN (SEQ ID No. 48) синтезируют следующие нуклеотидные праймеры для проведения ПЦР:

(2) Конструирование плазмиды pCR3.1/LM6-1-16 (клонирование гуманизированной легкой цепи TRA-8 LM6 типа)
Фрагмент LM6-ДНК, кодирующий аминокислотную последовательность, определенную в SEQ ID No. 75, получают при проведении 2-стадийной ПЦР с последующей вставкой в плазмидный вектор и клонированием в E. coli.
а) Первая стадия ПЦР
Фрагмент LM6-F1-ДНК, кодирующий сигнальную последовательность секреции, и участок FRL1 с добавленным к 5'-концу сайтом расщепления рестрикционным ферментом Hind III получают в следующих условиях. Матричные плазмиды pHSGHM17 и pSRPDHH получают в соответствии с описанием, приведенным в заявке на европейский патент ЕР 0909816 А1.
Состав реакционного раствора:
ДНК плазмиды pHSGHM17, 25 нг
олигонуклеотидный праймер 7AL1P, 50 пмоль
олигонуклеотидный праймер HKSPR13, 50 пмоль
смесь дНТФ, 5 мкл
10× буфер для ПЦР, 5 мкл
ДНК-полимераза ampliTaq (Perkin Elmer), 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагмент LM6-F2-ДНК, кодирующий часть FRL1, CDRL1, FRL2, CDRL2, FRL3, CDRL3,, FRL4 и часть константного участка получают в следующих условиях:
Реакционная смесь имеет следующий состав:
ДНК плазмиды pL28, 25 нг
олигонуклеотидный праймер MVF11, 50 пмоль
олигонуклеотидный праймер MVR12, 50 пмоль
смесь дНТФ, 5 мкл
10× буфер для ПЦР, 5 мкл
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагмент LM6-F3-ДНК, кодирующий часть FRL4 и константный участок с добавленным к 3'-концу сайтом расщепления рестрикционным ферментом EcoRI, получают в следующих условиях:
Состав реакционного раствора:
ДНК плазмиды pSRPDHH, 25 нг
олигонуклеотидный праймер MCF11, 50 пмоль
олигонуклеотидный праймер 7ALCN, 50 пмоль
смесь дНТФ, 5 мкл
10× буфер для ПЦР, 5 мкл;
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Фрагменты амплифицированной ДНК, полученной после проведения ПЦР, разделяют электрофорезом в 5% полиакриламидном геле. Гель после электрофореза окрашивают при добавлении 1 мкг/мл бромида этидия для выявления полученной ДНК в УФ свете. Выявленные при этом соответствующие полосы ДНК вырезают лезвием.
b) Вторая стадия ПЦР
LM6-ДНК, в которой слиты указанные выше фрагменты LM6-F1-ДНК, LM6-F2-ДНК и LM6-F3-ДНК, получают в следующих условиях:
Реакционный раствор имеет следующий состав:
гелевый фрагмент LM6-F1-ДНК, полученный на первой стадии ПЦР,
гелевый фрагмент LM6-F2-ДНК, полученный на первой стадии ПЦР,
гелевый фрагмент LM6-F3-ДНК, полученный на первой стадии ПЦР,
олигонуклеотидный праймер 7AL1P, 50 пмоль
олигонуклеотидный праймер 7ALCN, 50 пмоль
смесь дНТФ, 5,0 мкл
10× буфер для ПЦР, 5,0 мкл
ДНК-полимераза ampliTaq, 2,5 единицы.
Реакционный раствор, имеющий указанный выше состав, доводят до конечного объема 50 мкл при добавлении повторно дистиллированной воды и используют для проведения ПЦР.
Температурные условия для проведения ПЦР: нагревают при 94°С в течение 2 минут, после чего проводят цикл термической обработки, включающей нагревание при температуре 94°С в течение 1 минуты, нагревание при температуре 55°С в течение 1 минуты и нагревание при температуре 72°С в течение 2 минут с повторением указанного цикла 30 раз и затем проводят нагревание при 72°С в течение 10 минут.
Полученный таким образом фрагмент LM6-ДНК вставляют в плазмидную ДНК pCR3.1 с использованием набора для клонирования эукариотической ТА (InVitrogen) по инструкции от производителя и вводят в компетентные клетки E. coli TOP10F', входящие в состав набора. Нуклеотидные последовательности указанных ДНК, кодирующих гуманизированную легкую цепь TRA-8, подтверждают дидезоксиметодом с использованием анализатора ДНК.
Полученная плазмида была обозначена как pCR3.1/LM6-1-16 (плазмида, несущая кДНК, кодирующую вариабельный участок легкой цепи мышиного TRA-8 LM3 и константный участок легкой цепи человеческого Ig).
Полученную плазмиду pCR3.1/LМ6-1-16, содержащую фрагмент LM6-ДНК, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pHSG399 расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Полученные дефосфорилированная ДНК pHSG399 и фрагмент LM6-ДНК, которые были подвергнуты расщеплению рестрикционными ферментами Hind III и EcoR I, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем клетки E. coli DH5α трансформируют лигированной ДНК и распределяют их по поверхности агаровой среды LB, содержащей 0,1 мМ IPTG, 0,1% X-Gal и 50 мкг/мл хлорамфеникола (конечные концентрации). Полученные белые трансформанты культивируют в жидкой среде LB, содержащей 50 мкг/мл хлорамфеникола, и плазмидную ДНК экстрагируют из клеток полученных культур по методу, включающему обработку щелочью и ДСН. Экстрагированную плазмидную ДНК расщепляют с использованием Hind III и EcoR I и затем клон, содержащий фрагмент LM6-ДНК, выделяют электрофорезом в 1% геле агарозы.
В результате проведенной процедуры получают плазмиду pHSG/M6-1-4-1, содержащую фрагмент слияния вариабельного участка легкой цепи мышиного TRA-8 и константного участка цепи человеческого Igκ. Трансформированный штамм E. coli, несущий указанную плазмиду, обозначенный как E. coli DH5α/pHSG/M6-1-4-1, был депонирован в Международном депозитарии запатентованных организмов при Национальном Институте современной промышленной науки и технологии Японии (International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology, 1-1, Higashi 1 chome Tsukuba-shi, Ibaraki-ken, 305-5466) 20 апреля 2001 года в соответствии с Будапештским договором по депонированию микроорганизмов под номером FERM BP-7566.
3) Конструирование плазмиды pSR/LM6-1-4-6 (плазмида экспрессии легкой цепи TRA-8 LM6 химерного типа)
Полученную плазмиду pHSG/M6-1-4-1, несущую фрагмент слияния вариабельного участка легкой цепи мышиного TRA-8 и константного участка цепи человеческого Igκ, расщепляют рестрикционными ферментами Hind III и EcoR I.
Один мкг клонированной плазмидной ДНК pSRPDHH расщепляют рестрикционными ферментами Hind III и EcoR I и затем дефосфорилируют с использованием CIP. Образованная дефосфорилированная ДНК pSRPDHH и фрагмент ДНК Hind III-EcoR I, полученный из pHSG/LM6-1-4-1, лигируют с использованием набора для лигирования ДНК (DNA Ligation Kit Version 2.0) (Takara Syuzo, Co. Ltd.). Затем проводят трансформацию клеток E. coli DH5α лигированной ДНК и распределяют их по поверхности LB агара. Полученные трансформанты культивируют в жидкой среде LB, содержащей 100 мкг/мл ампициллина, и экстрагируют плазмидную ДНК из клеток полученной культуры по методу, включающему использование щелочи и ДСН. Наличие вставки и ориентацию желательного фрагмента ДНК в векторе подтверждают секвенированием с использованием анализатора генной последовательности.
Полученная плазмида экспрессии, несущая кДНК, кодирующую легкую цепь TRA-8 (химерного типа), была обозначена как pSR/LM6-1-4-6.
(5) Получение нескольких типов гуманизированного или химерного антитела TRA-8
Проводят трансфекцию COS-7 клеток по методике, включающей использование реагента для трансфекции FUGENE6 (Boehringer Mannheim Biochemica) в соответствии с инструкцией от производителя, прилагаемой к набору.
Клетки COS-7 (American Type Culture Collection No. CRL-1651) выращивают до состояния полуслияния (3·106 клеток/чашку) в чашках для культивирования (площадь культивирования: 57 см2; Sumitomo Bakelite), содержащих модифицированную Дульбекко среду Игла (далее обозначаемую как «D-MEM»; Gibco BRL) с добавкой 10% фетальной сыворотки теленка (далее обозначаемой как «ФСТ»; Moregate).
Тем временем получают в количестве 10 мкг/чашку (всего 5 чашек) плазмидную ДНК для экспрессии гуманизированной тяжелой цепи DR5 (pHA15-1) и в количестве 10 мкг/чашку плазмидную ДНК для экспрессии гуманизированной легкой цепи DR5 по методу, включающему обработку щелочью и ДСН, центрифугируют в градиенте плотности хлорида цезия и смешивают и затем проводят осаждение этанолом и далее суспендируют в количестве 5 мкл/чашку dH2O.
После смешивания в количестве 15 мкл/чашку реагента для трансфекции FUGENE6 со 180 мкл/чашку D-MEM без ФСТ указанный раствор FUGENE (185 мкл/чашку) смешивают с раствором ДНК в количестве 5 мкл/чашку, содержащим 10 мкг/чашку плазмидной ДНК для экспрессии гуманизированной тяжелой цепи DR5 и 10 мкг/чашку плазмидной ДНК для экспрессии гуманизированной легкой цепи DR5. После 15 минут инкубации про комнатной температуре полученную плазмидную суспензию (200 мкл) добавляют к приготовленным заранее чашкам с клетками COS-7. После инкубации в атмосфере 5% CO2 при 37°C в течение 24 часов культуральную среду заменяют средой D-MEM без ФСТ. После инкубации в атмосфере 5% CO2 при температуре 37°C в течение 72 часов культуральный супернатант регенерируют для очищения продуктов экспрессии в супернатантных жидкостях. По описанному выше методу клетки COS-7 трансфицируют каждым из указанных ниже видов сочетаний:
(А): совместная трансфекция с использованием pHA15-1 и pSR/LM1-2 (H1L1)
(В): совместная трансфекция с использованием pHB14-1 и pSR/M2-1 (H2L2)
(C): совместная трансфекция с использованием pHB14-1 и pSR/LM3-3-44-10 (H2L3)
(D): совместная трансфекция с использованием pHB14-1 и pSR/LM4-5-3-3 (H2L4)
(E): совместная трансфекция с использованием pHC10-3 и pSR/M2-1 (H3L2)
(F): совместная трансфекция с использованием pHC10-3 и pSR/LM3-3-44-10 (H3L3)
(G): совместная трансфекция с использованием pHC10-3 и pSR/LM4-5-3-3 (H3L4)
(H): совместная трансфекция с использованием pHD21-1 и pSR/LM5-3-27-1 (H4L5)
(I): совместная трансфекция с использованием pM11-1 и pSR/LM6-1-4-6 (химерный тип)
Далее культуру центрифугируют (35000 об/мин, 15 минут) и отбирают супернатант. Супернатант фильтруют через фильтр с размером пор 0,45 мкм (ADVANTEC TOYO DISMIC-25cs, Cat # 25CS045 AS). Очистку IgG из фильтратов проводят с использованием аффинной хроматографии на белке G-POROS (Applied Biosystems) в следующих условиях:
Система для проведения ВЭЖХ: BioCAD 700E (Applied Biosystems)
колонка: БелокG-ID с сенсорным картриджем
(размер колонки: 2,1 мм (внутр. диаметр) Ч 30 мм (длина), объем слоя: 0,1 мл, Applied Biosystems)
элюирующий буфер: 0,1М глицин-HCl (pH 2,5)
нейтрализующий буфер: 1М Трис-HCl (pH 8,5)
обнаружение: при длине волны 280 нм
скорость течения: 1 мл/мин
размер фракции: 0,5 мл/0,5 мин
пробирка для сбора фракций: 1,5 мл полипропиленовая микропробирка
температура: 4°С.
После нанесения всего фильтрата на колонку используют 50 мл ФБР (Sigma, Cat # 1000-3) для промывки колонки. С момента нанесения элюирующего буфера начинается работа коллектора фракции. Каждая микропробирка для сбора фракций содержит предварительно внесенные 55 мкл 1М NaCl, 110 мкл нейтрализующего буфера и 74 мкл бычьего сывороточного альбумина в концентрации 2 мг/мл (Sigma, Cat # А-7030) в ФБР. Собирают фракции с No. 7 до No. 8.
Подтверждение экспрессии гуманизированных антител и количественный анализ продуктов экспрессии в полученных жидкостях из культурального супернатанта проводят методом ELISA с использованием антитела против человеческого IgG.
К каждой лунке в 96-луночном микропланшете (MaxiSorp, Nunc) добавляют 100 мкл козьего поликлонального антитела, специфичного к человеческому IgG Fc (Kappel), растворенному до конечной концентрации 0,5 мкг/мл в адсорбционном буфере (0,05 М гидрокарбонат натрия, 0,02% азид натрия, pH 9,6) и планшет инкубируют при 37°С в течение 2 часов для адсорбции антитела. Затем планшет промывают 350 мкл ФБР-Т 5 раз. К лункам после промывки добавляют культуральный супернатант, разбавленный D-MEM, содержащим 10% ФСТ, и инкубируют при 37°С в течение 2 часов. После промывки снова с использованием ФБР-Т к каждой лунке добавляют 100 мкл козьего специфичного поликлонального антитела против человеческого IgG Fc, меченного щелочной фосфатазой (Jackson Immuno Research Lab.), разбавленного в 10000 раз ФБР-Т, и инкубируют при 37°С в течение 2 часов. После новой промывки с использованием ФБР-Т добавляют субстратный буфер, содержащий п-нитрофенилфосфат, полученный из набора для щелочной фосфатазы (BioRad) в соответствии с инструкцией от производителя, прилагаемой к набору. После инкубирования при 37°С в течение от 0,5 до 1 часа измеряют поглощение при длине волны 405 нм. В настоящих экспериментах человеческий иммуноглобулин G подкласса 1 из плазмы человека (IgG1) (Biopure AG), разбавленный D-MEM, содержащий 10% ФСТ до определенных концентраций, используют в качестве эталонных образцов со стандартной концентрацией гуманизированных антител DR5, содержащихся в жидкостях культурального супернатанта.
В результате с помощью антитела против человеческого IgG осуществляется специфическое выявление наличия экспрессии и очищенных продуктов в культуральном супернатанте. Конечная концентрация человеческого IgG антитела составляет 44,03 мкг/мл (H1L1), 39,8 мкг/мл (H2L2), 26,7 мкг/мл (H2L3), 41,0 мкг/мл (H2L4), 39,3 мкг/мл (H3L2), 24,7 мкг/мл (H3L3), 21,5 мкг/мл (H3L4), 16,7 мкг/мл (H4L5) и 18,3 мкг/мл (химерный тип) соответственно.
(6) Апоптоз-индуцирующая активность нескольких типов гуманизированного антитела или химерного антитела
Клетки Jurkat (ATCC No. TIB-152) используют для изучения индуцирующей апоптоз активности очищенного гуманизированного антитела TRA-8.
Клетки Jurkat культивируют в среде RPMI1640 с добавкой 10% ФСТ (Gibco BRL) при 37°С в течение 3 дней в атмосфере 5% CO2 и затем распределяют по лункам 96-луночного микропланшета (Sumitomo Bakelite) по 50 мкл на лунку. Гуманизированные TRA-8, приготовленные по методике примера 26, доводят до концентрации готового интересующего продукта, равной 100 нг/мл, с использованием среды RPMI1640, содержащей 10% ФСТ, при оценке их концентрации в жидкостях по методике, описанной в примере 26. Каждый из растворов экспрессированных продуктов, доведенных до концентрации 100 нг/мл, используют для получения серийных разбавлений путем повторных серийных 2-кратных разбавлений средой RPMI1640, содержащей 10% ФСТ. Каждый из разбавленных растворов гуманизированного TRA-8 (H1L1, H2L2, H2L3, H2L4, H3L3, H3L4 или H4L5) добавляют к лункам по 50 мкл на лунку. После реакции в течение 12 часов при 37°С добавляют 50 мкл 25 мкМ ФМС, содержащий 1 мг/мл ХТТ (конечная концентрация составляет 250 мкг/мл для ХТТ и 5 мкМ для ФМС). После инкубации в течение 3 часов измеряют поглощение при длине волны 450 нм в каждой лунке и вычисляют жизнеспособность клеток с использованием в качестве параметра восстановительной функции митохондрий.
Жизнеспособность клеток в каждой из лунок вычисляют по следующей формуле:
Жизнеспособность (%) = 100 · (a-b)/(c-b)
где «а» обозначает результат измерения в опытной лунке, «b» обозначает результат измерения в лунке без добавленных клеток и «c» обозначает результат измерения в лунке без добавления антитела.
В результате было показано, что исследуемые гуманизированные антитела индуцируют апоптоз в клетках клеточной линии Т-лимфомы, экспрессирующих человеческий антиген DR5.
Далее была изучена апоптоз-индуцирующая активность гуманизированного антитела TRA-8 к PC3 при добавлении таксола с использованием метода, описанного в примере 25.
Клеточную линию раковых клеток предстательной железы человека PC-3 (ATCC No. CRL-1435), полученную из Американской Коллекции типовых культур (ATCC), поддерживают на питательной смеси F-12K (21127-022, Gibco BRL), содержащей 10% фетальной сыворотки теленка (ФСТ, Hyclone), 1% L-глютамина - 200 мМ (25030-149, Gibco BRL) и 0,5% раствор пенициллина-стрептомицина (P-7539, Sigma). В проведенном ниже эксперименте используют среду RPMI1640 (MED-008, IWAKI) с добавкой 10% ФСТ и 0,5% раствора пенициллина-стрептомицина. РС-3 клетки в экспоненциальной фазе роста собирают с проведением обработки трипсином и двойной промывкой свежей средой. Затем клетки подсчитывают, ресуспендируют в свежей среде с плотностью 5·104 клеток/мл и распределяют в тройном повторе по лункам 96-луночного микропланшета с плоским дном (3598, Corning-Coster) до общего объема 100 мкл/лунку за один день до начала эксперимента. Репрезентативное противораковое средство Поклитаксел (169-18611, Wako), растворенное в диметилсульфоксиде (10 мг/мл), разбавляют свежей средой и затем добавляют к лункам 96-луночного планшета, содержащим клетки в количестве 50 мкл/лунку. Конечные концентрации диметилсульфоксида ниже, чем 0,1%. После инкубации в течение 24 часов при 37°С в атмосфере 5% CO2 к лункам добавляют гуманизированное антитело TRA-8 (H1L1, H2L2, H2L3, H2L4, H3L2, H3L3, H3L4 и H4L5), разбавленное свежей средой. После инкубации еще в течение 24 часов к лункам добавляют 50 мкл минимальной эссенциальной среды (11095-098, Gibco BRL), содержащей 1 мг/мл XTT и 25 мМ ФМС и планшеты инкубируют в течение 6 часов. Величину поглощения ОП450 измеряют с использованием счетчика (ARVO HTS 1420 Multilabel Counter; Wallac Berthold) и подсчитывают, как указано ниже, жизнеспособность клеток.
Жизнеспособность клеток (%) = (ОП450 для лунки, содержащей клетки, обработанные Таксолом и гуманизированным TRA-8 (агент(ы)) -ОП450 для лунки, не содержащие ни клеток, ни агента) · 100/(ОП450 для лунки, содержащей клетки и не содержащей агенты - ОП450 для лунки, не содержащей ни клеток, ни агента)
В результате было показано, что исследуемые гуманизированные антитела индуцируют апоптоз в клеточных линиях рака предстательной железы, экспрессирующих человеческий антиген DR5.
Пример 27. Продукция антитела DR4
Белок слияния, содержащий внеклеточный домен человеческого DR4 (аминокислотные остатки 1-236) и Fc часть человеческого IgG1 экспрессируется в клетках Cos-7, трансфицированных рекомбинантным аденовирусным вектором. Белок слияния очищают аффинной хроматографией с белком А. Мышей линии Balb/c иммунизируют очищенным белком слияния, как было описано выше. Один клон гибридомы 2E12 (IgG1, κ), характеризующийся специфическим связыванием с DR4 и способностью индуцировать апоптоз в клетках лимфомы В человека Ramos, подвергают субклонированию три раза. Специфичность по связыванию с 2E12 определяют методом ELISA и вестерн-блоттинга с использованием DR5, DcR1 DcR2 и белка слияния IgG1 в качестве контрольных антигенов. Связывание 2E12 с DR4 на клеточной поверхности определяют методом проточной цитометрии Cos-7 клеток, трансфицированных кДНК полной длины, кодирующей человеческий DR4. Апоптоз-индуцирующую активность определяют при инкубации Ramos клеток с 1 мкг/мл 2E12 в присутствии козьего противомышиного IgG1. Жизнеспособность клеток определяют по методу ATPLite, как было описано выше.
Пример 28. Характеристика антитела DR4
Моноклональное антитело DR4 (2Е12) является специфическим для человеческого DR4, но не связывается с другими рецепторами TRAIL, такими как DR5, DcR1 и DcR2, по результатам оценки методом ELISA (фиг. 19А). 2Е12 распознает DR4 на клеточной поверхности, как было показано при проведении проточной цитометрии Cos-7 клеток, трансфицированных DR4 (фиг. 19В). 2Е12 способно индуцировать апоптоз лимфомных клеток Ramos в присутствии второго сшивающего антитела зависимым от дозы образом (фиг. 19С). Обработка in vitro клеток Ramos 2Е12 приводит к зависимой от времени активации каспазы 8, 9 и 3 и к расщеплению PARP (фиг. 19D). Приведенные результаты показывают, что 2Е12 является антагонистическим антителом против DR4, которое индуцирует апоптоз зависимым от каспазы способом.
С использованием анти-DR4 (2Е12) и далее ФЭ-конъюгированного козьего антитела против мышиного IgG1 с проведением проточной цитометрии было показано, что антитело DR4 связывается с клетками клеточной линии фибросаркомы (Hs913T) и несколькими клеточными линиями рака молочной железы (2LMP, MDA-MB231 и MDA-MB-453), но незначительно связывается с нормальной клеточной линией фибробластов кожи человека (Malme-3).
Пример 29. Активность антител DR4 по уничтожению опухолевых клеток
Активность 2Е12 по уничтожению опухолевых клеток исследуют на моделях раковой опухоли молочной железы. Мышам nude инокулируют п/к клеточную линию рака молочной железы человека 2LMP.
Обработку вводимыми в/б дозами 2Е12 по 200 мкг проводят в дни 7, 10, 14, 17, 21 и 24 после инъекции опухолевых клеток. Животным вводят в/в адриамицин (доксорубицин) (6 мг/кг) в дни 8, 12 и 16. Лечение с использованием 2Е12 и адриамицина (фиг. 20) приводит к большему подавлению роста опухоли, чем при использовании 2Е12 или адриамицина поодиночке.
Активность TRA-8 по уничтожению опухолевых клеток в сочетании с 2Е12 исследуют с использованием тех же самых моделей раковой опухоли молочной железы. Введение в/б 200 мкг TRA-8 и 2Е12 проводят в дни 7, 10, 14, 17, 21 и 24 после инъекции опухолевых клеток. Животным вводят в/в адриамицин (6 мг/кг) в дни 8, 12 и 16. Лечение с использованием TRA-8 плюс 2Е12 или TRA-8 плюс 2Е12 и адриамицин приводит к 88% и 100% полной регрессии опухоли, соответственно (фиг. 21).
Пример 30. Активность антител DR4/DR5 по уничтожению опухолевых клеток в сочетании с другими видами лечения
(1) Клеточные линии и реагенты
Субклон 2LMP клеточной линии рака молочной железы человека MDA-MB-231, субклон LCC6 MDA-MB-435 и субклон DY36T2 MDA-MB-361, полученные от Dr. Marc Lipmann из Georgetown University, Washington, D.C., поддерживают на улучшенной среде MEM с добавкой 10% ФБР (Hyclone, Logan, UT). Клеточные линии рака молочной железы человека MDA-MB-231, MDA-MB-453, MDA-MB-468, BT-474, SK-BR-3 и ZR-75-1 получают из Американской Коллекции типовых культур (Manassas, VA). Клетки MDA-MB-231, MDA-MB-453 и MDA-MB-468 растят в среде DMEM с добавкой MEM витаминов, MEM заменимых аминокислот, 1 мМ пирувата натрия и 10% ФБР. Клетки BT-474 растят в среде RPMI1640 с добавкой 10 мкг/мл инсулина, 4,5 г/л глюкозы, 10 мМ HEPES, 1 мМ пирувата натрия и 10% ФБР. Клетки SK-BR-3 растят в среде McCoy's с добавкой 15% ФБР. Клетки ZR-75-1 растят в среде Ham's F12K с добавкой 20% ФБР. Все клетки поддерживают на среде без антибиотиков при 37°С в атмосфере 5% CO2 и проводят традиционный анализ на заражение микоплазмой.
Очищенное моноклональное антитело TRA-8 (IgG1), произведенное в UAB, было также получено от компании Sankyo Co., Ltd. (Токио. Япония). Конъюгированный с фикоэритрином козий противомышиный IgG1 и специфичное к изотипу IgG1 контрольное антитело получают от компании Southern Biotechnology Associates (Birmingham, AL). Адриамицин и паклитаксел получают от компании Sigma Chemical CO. (St. Louis, MO) и готовят для использования в виде 10 мМ раствора для хранения в дистиллированной воде или ДМСО соответственно. При исследовании на животных клинические композиции паклитаксела (Bristol-Myers Squibb Co., Princeton, NJ) получают из университета Алабамы, фармакологического отделения при Бирмингемской больнице. Указанный препарат разбавляют 1:5 ФБР непосредственно перед использованием.
(2) Непрямая иммунофлуоресценция и анализ проточной цитометрией экспрессии DR5
Клетки в экспоненциальной фазе роста промывают один раз ФБР в модификации Дульбеко (дефицитной по Ca2+ и Mg2+) и собирают с использованием 4 мМ ЭДТА/0,5% KCl при 37°С. Клетки собирают центрифугированием при 4°С в течение 5 минут со скоростью 1000 об/мин, промывают один раз и ресуспендируют в ФБР, содержащем 1% БСА и 0,01% азида натрия (FACS буфер) при 4°С. Клетки инкубируют с 10 мкг/мл очищенного TRA-8 или с контрольным антителом, специфичным к изотипу IgG1, в течение 60 минут при 4°С, промывают один раз буфером и затем инкубируют с 10 мкг/мл ФЭ-конъюгированного козьего противомышиного IgG1 в течение 20 минут при 4°С. После окрашивания антителом клетки промывают один раз FACS буфером и фиксируют 1% параформальдегидом в течение 15 минут на льду. Образцы измеряют с использованием флуоресцентного анализатора Becton Dickinson Facscan (San Jose, CA) и полученные данные анализируют с помощью программного обеспечения CellQuest.
(3) Анализ жизнеспособности клеток по методу ATPLite
Клетки обрабатывают трипсином и ресуспендируют в полной культуральной среде. Одну тысячу клеток на лунку помещают в оптически чистые 96-луночные черные планшеты (Costar #3904, Corning, NY) и инкубируют в течение ночи при 37°С до начала обработки. Лекарственные средства и антитело разбавляют культуральной средой непосредственно перед использованием, и конечная концентрация ДМСО всегда ≤0,001%. Жизнеспособность клеток оценивают через 24 часа после воздействия одного TRA-8. В случае сочетанных обработок цитотоксическими средствами клетки предварительно обрабатывают лекарственным средством за 24 часа перед добавлением антитела и инкубируют еще в течение 24 часов перед оценкой жизнеспособности клеток путем измерения клеточного уровня АТФ в тесте ATPLite, основанном на люминесценции (Packard Instruments, Meriden, CT). Следуют инструкции от производителя с тем исключением, что все реакционные объемы (культуральная среда, реагенты) снижают наполовину. Все тесты анализируют в тройном повторе и результаты выражают в виде среднего значения ± СКО результатов минимум трех независимых экспериментов.
(4) Исследование результатов лечения с использованием одного TRA-8 или в сочетании с химиотерапией или радиационным облучением на атимических мышах Nude, содержащих ксенотрансплантаты рака молочной железы
Атимичным мышам nude инъецируют п/к 3·107 2LPM клеток. На 7 день после инъекции опухолевых клеток вводят в/б 200 или 600 мкг (10 или 30 мг/кг) TRA-8, после чего проводят 5 дополнительных инъекций в дни 10, 14, 17, 21 и 24. В течение указанного времени отслеживают рост опухолей. В последующих исследованиях животным, содержащим введенные п/к клетки опухоли 2LMP, инъецируют в/б 200 мкг TRA-8 в дни 7, 10, 14, 17, 21 и 24, один или в сочетании с адриамицином (6 мг/кг, в/в, в дни 8, 12 и 16) или с паклитакселом (20 мг/кг, в/б, в дни 8, 12, 16, 20 и 24). Определяют размер опухоли и уровень регрессии. Кроме того, проводят исследование с использованием TRA-8 и адриамицина в описанном выше режиме в сочетании с облучением ксенотрансплантатов 2LMP дозой 3 Gy60Co в дни 9 и 17.
(5) Анализ апоптоза в ксенотрансплантатах
Атимичным мышам nude инъецируют п/к 3·107 2LPM клеток (в день 0) и затем вводят в/б 100 мкг TRA-8 в дни 7 и 10. Мышам в группах по 2 мыши в каждой вводят адриамицин (3 мг/кг) в дни 8 и 11, паклитаксел (10 мг/кг) в дни 8 и 11 или сочетание TRA-8 и адриамицина или паклитаксела в той же дозе и в том же режиме. Одну группу мышей оставляют необработанной. Ксенотрансплантаты рассекают для исследования апоптоза на 14 день после инъекции опухолевых клеток. Причина значительного снижения интенсивности лечения в сравнении со стандартной процедурой заключалась в том, чтобы создать условия для адекватного сравнения опухолевых тканей при проведении анализа на 14 день. Тест Tunel на апоптоз в ксенотрансплантатах опухоли проводят следующим образом. Парафиновые срезы ткани по пять микрон кладут на слайды SuperFrost/Plus и нагревают при 58°С в течение 1 часа. Далее из срезов тканей удаляют парафин обработкой ксилолом с тремя заменами и регидратируют с одной заменой абсолютного этанола, 95% этанола и 70% этанола, каждую из обработок проводят с 5-минутными интервалами. Затем срезы помещают в Трис-забуференный солевой раствор (0,5M Трис-основание, 0,15 M NaCl, 0,0002% Тритон Х-100, pH 7,6). Апоптозные ядра выявляют с использованием набора на пероксидазу (Intergen, Purchase, NY). К образцам ткани добавляют протеиназу К (20 мкг/мл в дистиллированной деионизированной воде) и инкубируют при комнатной температуре в течение 15 минут. Эндогенные пероксидазы гасят добавлением водного раствора 3% пероксида водорода в течение 5 минут. Срезы обрабатывают уравновешивающим буфером в течение 30 минут и затем инкубируют с TdT/ферментом (разбавленным реакционной смесью для мечения) в течение 1 часа при 37°С с использованием парафильмовых покровных стекол. В ходе указанной инкубации TdT фрагмент связывает 3'-OH концы фрагментов ДНК и катализирует добавление дигоксигенин-меченых и немеченых дезоксинуклеотидов. Отрицательные контроли инкубируют с дистиллированной Н2О (разбавленный реакционной смесью для мечения), добавляемой вместо ТдТ фермента. Останавливающий буфер добавляют на 10 минут при комнатной температуре для прекращения реакции мечения. К каждому слайду на 30 минут добавляют анти-дигоксигениновый конъюгат. Используют хромаген 3,3' ДАБ для визуализации меченого 3'-OH конца фрагментов ДНК. Затем слайды промывают деионизированной водой и подвергают легкому контрастному окрашиванию гематоксилином, дегидратируют с использованием очищенных спиртов и ксилола и закрывают с использованием Permount. Оценивают примерно 10 выбранных случайным образом полей для определения процента окрашивания по методу Tunel и процента интенсивно окрашенных апоптозных тел в ткани.
(6) Статистический анализ
(А) Анализ взаимодействия RTA-8 с цитотоксическим лекарственным средством in vitro
Данные по цитотоксичности анализируют для оценки того, какими являются цитотоксические эффекты вводимого сочетания средств: аддитивными, менее чем аддитивными (антагонистическими), или более чем аддитивными (синергическими). Взаимосвязь дозы и реакции для средств поодиночке и в сочетании моделируют с использованием модели поверхностной реакции второго порядка в линейном, квадратичном и интерактивном режимах для каждой из 9 клеточных линий (Montgomery, D,C, Design and Analysis of Experiments, New York: Wiley, 2001) в соответствии с рекомендациями Gennings (On Testing for Drug/Chtmical Interaction: Definitions and Inference, pp. 457-468, 2000). Значимый параметр взаимодействия классифицируется как синергический или антагонистический в зависимости от того, был ли указанный параметр взаимодействия отрицательным, вызывая более чем аддитивную, цитотоксичность, или положительным, вызывая менее чем аддитивную цитотоксичность. Если параметр взаимодействия был незначимым, то взаимосвязь между TRA-8 и адриамицином или TRA-8 и паклитакселем рассматривается аддитивной при условии, что параметр аддитивности был значительным.
(B) Анализ эффекта TRA-8, химиотерапии, радиационного облучения и сочетанной терапии в индивидуальных экспериментах с животными
Данные, полученные в шести независимых исследованиях, анализируют в независимом эксперименте. Лечение с использованием сочетанной обработки сравнивают с результатами по противоопухолевой эффективности in vitro, то есть по подавлению роста опухоли, которую измеряют в трех точках: по увеличению времени удвоения размера опухоли, по проценту регрессии опухоли и по относительной скорости роста с течением времени. Фактическое число дней, в течение которых опухоль удваивает площадь поверхности (выражаемую в виде значений двух диаметров) относительно базовой линии на 7 день после инъекции опухоли, используют в анализе времени удвоения. Непараметрический тест Крускала-Валлиса (Krusral-Wallis) используют для сравнения средних значений времени удвоения опухоли между обработками. Для сравнения соотношения уровня регрессий опухоли и уровня регрессии без рецидивов в группах лечения используют тест на точность Фишера. Для определения того, дает ли какое-либо сочетание видов лечения значимое синергическое подавление роста опухоли, то есть более чем аддитивный эффект, проводят сравнение кривых роста, полученных при измерениях серий площадей, с использованием линейного смешанного подхода, в течение первых 3 недель после начала лечения (Lindsey, J.K. Models for Repeated Measurements, pp. 100-142, Oxford, 1993). Для определение наличия синергических эффектов сочетанного лечения в модель включают параметр, оценивающий взаимодействие. Если параметр взаимодействия является значимым и наблюдаемым эффектом является подавление роста со скоростью большей, чем аддитивный эффект, то такое взаимодействие рассматривается как синергическое.
(С) Агрегатный анализ эффекта лечения
В агрегатный анализ включают всего 166 животных, 10 групп лечения и проводят 6 независимых экспериментов. Результаты сочетанного лечения сравнивают по противоопухолевой эффективности in vivo. Среднее значение времени удвоения опухоли анализируют с использованием теста Крускала-Валлиса, и тест на точность Фишера используют для сравнения соотношения уровня регрессии опухоли и уровня регрессии без рецидивов в группах лечения.
Все статистические анализы проводят с использованием SAS® (Sas/Star User's Guide, SAS OnlineDoc, Version 8, Cary NC: SAS Institute Inc., 1999).
(6) Экспрессия DR5 и индуцированная TRA-8 цитотоксичность клеточных линий рака молочной железы
Как видно из фиг. 22А, все девять клеточных линий рака молочной железы являются DR5-положительными с варьирующим уровнем экспрессии от очень положительного (LCC6 MDA-MB-453) до слабо положительного (MDA-MB-468 и SK-BR-3). На фиг. 22B показано наличие TRA-8-индуцированной цитотоксичности в девяти клеточных линиях. Четыре клеточных линии демонстрируют чувствительность к TRA-8-индуцированной цитотоксичности со значениями ИК50 17-299 нг/мл (LCC6, 2LMP, MDA-MB-231, MDA-MB-468), тогда как другие являются вполне устойчивыми (DY36T2, BT-474, MDA-MB-453). Не отмечается хорошей корреляции экспрессии DR5 и уровня TRA-8-индуцированной цититоксичности, как показано на клеточных линиях MDA-MB-453 и MDA-MB-468.
Эффект TRA-8 на цитотоксичность, индуцированную химиотерапией, изучают с использованием адриамицина (фиг. 23А) и паклитаксела (фиг. 23В). Результаты анализа, проведенного с целью исследования взаимодействия между антителом и эффектом вводимого лекарственного препарата, представлены в таблице 5. Не отмечается значимого синергического взаимодействия между TRA-8 и паклитакселом, при этом большая часть эффектов является аддитивной. Четыре из девяти клеточных линий соответствуют критерию синергического взаимодействия между TRA-8 и адриамицином. Клеточная линия 2LMP демонстрирует хорошую чувствительность к TRA-8, а также чувствительность к адриамицину или паклитакселу. Указанная клеточная линия была выбрана для исследования in vivo эффективности антитела и/или лекарственных средств.
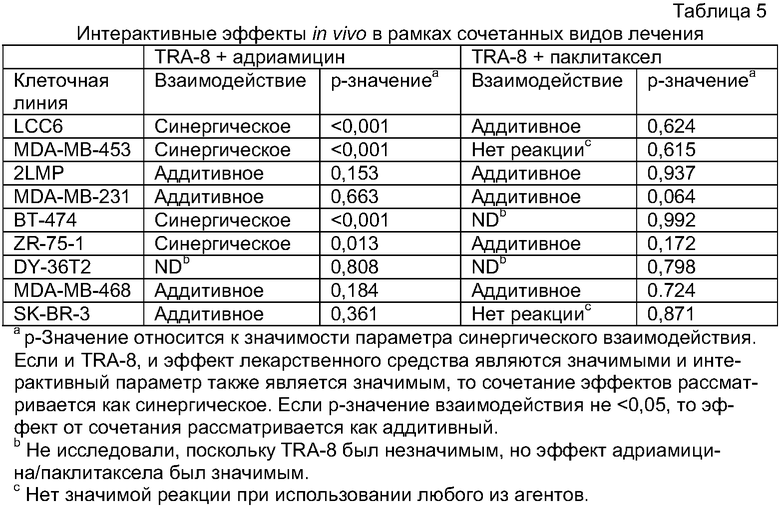
(7) Противоопухолевые эффекты no vivo TRA-8 одного или в сочетании с химиотерапией и/или облучением
TRA-8 в дозах 200 мкг и 600 мкг, вводимый дважды в неделю в виде 6 доз, приводит к аналогичному подавлению роста опухоли в случае хорошо развившихся после п/к введения 2LMP опухолей (фиг. 24). В трех дополнительных независимых экспериментах доза в 200 мкг/режим давала статистически значимое подавление роста опухоли (p<0,004, тест Крускала-Валлиса по времени удвоения опухоли) в сравнении с необработанными контролями, и указанные доза и режим были выбраны для дальнейших исследований. Фиг. 25 иллюстрирует эффект TRA-8, адриамицина или сочетания TRA-8 и адриамицина на противоопухолевую эффективность. В сравнении с необработанными контролями лечение TRA-8 одним или TRA-8 + адриамицин приводит к значимому подавления роста опухоли (p=0,002, тест Крускала-Валлиса), тогда как адриамицин не отличается от контролей. Сочетание TRA-8 плюс адриамицин давало большее подавление роста, чем любое средство поодиночке (р=0,002), а также значительно чаще приводило к полной регрессии опухолевых клеток (в четыре раза), чем любое средство поодиночке, где полная регрессия не отмечалась (р<0,001, тест на точность Фишера). Синергизм действия in vivo TRA-8 и адриамицина оценивается с использованием анализа кривой раннего роста. Параметр взаимодействия является значимым (p<0,001) и указывает на синергическое взаимодействие. Синергическое взаимодействие исследовали во втором независимом эксперименте.
Эффекты TRA-8 и паклитаксела исследуют на той же модели с теми же параметрами исследования (фиг. 26). В сравнении с необработанными контролями TRA-8 и TRA-8 плюс паклитаксел демонстрировали значительное подавление роста опухоли (p<0,001 тест Крускала-Валлиса). Рост опухоли у животных, обработанных TRA-8 плюс паклитаксел, существенно отличался от варианта применения одного паклитаксела (p=0,008) и давал 3/8 от полной регрессии в сравнении с отсутствием такого результата в варианте введения любого из средств поодиночке. Анализ кривых раннего роста опухоли показывает, что синергический эффект был почти значимым (p=0,063), тогда как аддитивный эффект был значимым (p<0,001).
И наконец, анализируют эффект TRA-8, адриамицина и облучения дозой 60Со в качестве единственных средств и в различных сочетаниях, как показано на фиг. 27. Отмечается значимое различие в отношении времени удвоения опухоли (p<0,001), и множественные сравнения показывают, что тройная терапия с использованием TRA-8, адриамицина и облучения дозой 60Со приводит к подавлению роста опухоли, которое значительно отличается от всех других групп лечения, тогда как обе группы с двойной терапией (адриамицин + TRA-8 или 60Со + TRA-8) отличались также от групп лечения одним лишь агентом (p<0,001). Животные, которых лечили путем облучения дозой 60Со, не отличались от необработанных контролей (p=0,926). Все двухпараметровые варианты сочетаний характеризовались значимым синергическим эффектом (p<0,001). Полная регрессия наблюдалась у 6 из 8 животных, проходивших тройную терапию, и у 4 животных не отмечено рецидива опухоли в течение 180 дней наблюдения.
(8) Агрегатный анализ эффекта терапии
В исследования противоопухолевого эффекта in vivo включены 166 животных, для каждого из которых анализируют время удвоения опухоли и частоту полной регрессии опухоли (таблица 6). Анализ вариантов по методу ANOVA средних показателей времени удвоения опухоли указывает на наличие значимого различия между группами с разными видами лечения (p<0,001), и результаты множественного сравнения показывают, что применение TRA-8+паклитаксел, TRA-8+адриамицин и TRA-8+адриамицин+облучение 60Со приводит к достижению значительно большего показателя времени удвоения опухоли, чем в любой группе лечения без TRA-8. Добавление TRA-8 в любом варианте лечения приводит к большему увеличению показателя времени удвоения опухоли, чем сам по себе режим лечения. Аналогично в тесте Крускала-Валлиса по анализу среднего времени удвоения опухоли показано, что средние значения характеризуются существенными различиями (p<0,001). Попарное сравнение вариантов с использованием ранжированного теста Вилкоксина дает сходную картину среднего времени удвоения опухоли, соответствующую результатам, полученным при проведении множественного сравнения по методу ANOVA. Однако указанный анализ недооценивает подавления роста, вызванного наиболее эффективной обработкой в тех группах, которые не достигли удвоения размеров опухоли к концу эксперимента, который был установлен в день его планового окончания. В таблице 6 приведены данные по частоте полной регрессии опухоли и по частоте персистенции регрессии к концу эксперимента. Не отмечалось полной регрессии опухоли у животных, которых лечили либо химиотерапией, либо облучением по таким параметрам, как рост опухоли и агрессивность опухоли. На основании теста на точность Фишера показано наличие значимой разницы по частоте полной регрессии опухоли между группами с разными видами лечения (p<0,001). У тридцати из 166 животных достигнута полная регрессия и 28 из них получали только TRA-8 или его в сочетании с другими режимами лечения. Полная регрессия наблюдается у 1/42 контрольных животных: 1/54 животных получали химиотерапию, облучение или их сочетание, и у 28/68 животных, принимавших TRA-8 или TRA-8 в сочетании с другими вариантами лечения. Группы, для лечения которых использовали TRA-8, характеризовались значительно большей (p<0,001) частотой полной регрессии. Аналогично у 14/68 животных, получавших TRA-8 или сочетанное лечение с TRA-8, не выявлено повторного роста опухоли в сравнении с 1/42 контролями или 0/52 животными, которых лечили химиотерапией и/или облучением. Регрессию без рецидивов наблюдали в течение периода времени от 99 до 171 дня (146±24 дня).
(9) Апоптоз обработанных опухолей
Индукция апоптоза ксенотрансплантатов 2LMP после лечения TRA-8, адриамицином, паклитакселом, TRA-8+адриамицин и TRA-8+паклитаксел оценивают с использованием методики TUNEL. У необработанных животных в опухолях показано наличие 4% окрашенных клеток (1% интенсивно), тогда как в случае лечения адриамицином или паклитакселом указанная величина составляет 8% (6% интенсивно) и 7% (2% интенсивно) окрашенных клеток соответственно. Животные, которым вводят один TRA-8, характеризуются выраженным апоптозом с наличием 25% (15% интенсивно) окрашенных клеток. Вариант лечения TRA-8+ адриамицин дает величину 28% (22% интенсивно) и вариант лечения TRA-8+паклитаксел дает величину 26% (12% интенсивно) окрашенных клеток.
Любые патенты и публикации, приведенные в настоящем описании, являются показателями достигнутого уровня в той области, к которой относится настоящее изобретение. Указанные патенты и публикации включены в настоящее описание в той мере, в которой каждая индивидуальная публикация конкретно и индивидуально относится к теме настоящей заявки.
Настоящее изобретение не ограничивается по тематике указанными выше материалами или описанными в примерах вариантами изобретения, которые следует рассматривать как иллюстрацию некоторых аспектов настоящего изобретения, и любой функционально эквивалентный вариант также входит в область настоящего изобретения. Для специалистов в данной области техники могут быть очевидны различные модификации изобретения в дополнение к настоящему описанию, которые также охватываются рамками прилагаемой формулы изобретения.




| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНАЦИИ АНТИТЕЛ, ОБЛАДАЮЩИХ СЕЛЕКТИВНОСТЬЮ ПО ОТНОШЕНИЮ К РЕЦЕПТОРУ ЛИГАНДА, ИНДУЦИРУЮЩЕМУ АПОПТОЗ, АССОЦИИРОВАННЫЙ С ФАКТОРОМ НЕКРОЗА ОПУХОЛИ, И ДРУГИХ ТЕРАПЕВТИЧЕСКИХ СРЕДСТВ | 2002 |
|
RU2313368C2 |
| АНТИТЕЛО, ОБЛАДАЮЩЕЕ СЕЛЕКТИВНОСТЬЮ ПО ОТНОШЕНИЮ К РЕЦЕПТОРУ ЛИГАНДА, ИНДУЦИРУЮЩЕМУ АПОПТОЗ, АССОЦИИРОВАННЫЙ С ФАКТОРОМ НЕКРОЗА ОПУХОЛИ, И ЕГО ИСПОЛЬЗОВАНИЕ | 2001 |
|
RU2298013C2 |
| АПОПТОТИЧЕСКАЯ ЧУВСТВИТЕЛЬНОСТЬ К Apo2L/TRAIL ПУТЕМ ТЕСТИРОВАНИЯ ЭКСПРЕССИИ GalNac-T14 В КЛЕТКАХ/ТКАНЯХ | 2006 |
|
RU2416097C2 |
| АНАЛИЗЫ И СПОСОБЫ ПРИМЕНЕНИЯ БИОМАРКЕРОВ | 2006 |
|
RU2409817C2 |
| АНАЛИЗЫ И СПОСОБЫ, ИСПОЛЬЗУЮЩИЕ БИОМАРКЕРЫ | 2005 |
|
RU2410438C2 |
| СПОСОБЫ ПРИМЕНЕНИЯ АГОНИСТОВ АРО-2L-РЕЦЕПТОРОВ И АКТИВАТОРОВ NK-КЛЕТОК | 2005 |
|
RU2395294C2 |
| СПОСОБ ПОЛУЧЕНИЯ МУТАНТНОГО БЕЛКА TRAIL ЧЕЛОВЕКА | 2009 |
|
RU2405038C1 |
| АНАЛИЗЫ И СПОСОБЫ С ПРИМЕНЕНИЕМ БИОМАРКЕРОВ | 2005 |
|
RU2431676C2 |
| АНТИ-DR5-АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2735956C1 |
| НОВОЕ АНТИТЕЛО ПРОТИВ DR5 | 2011 |
|
RU2644678C1 |
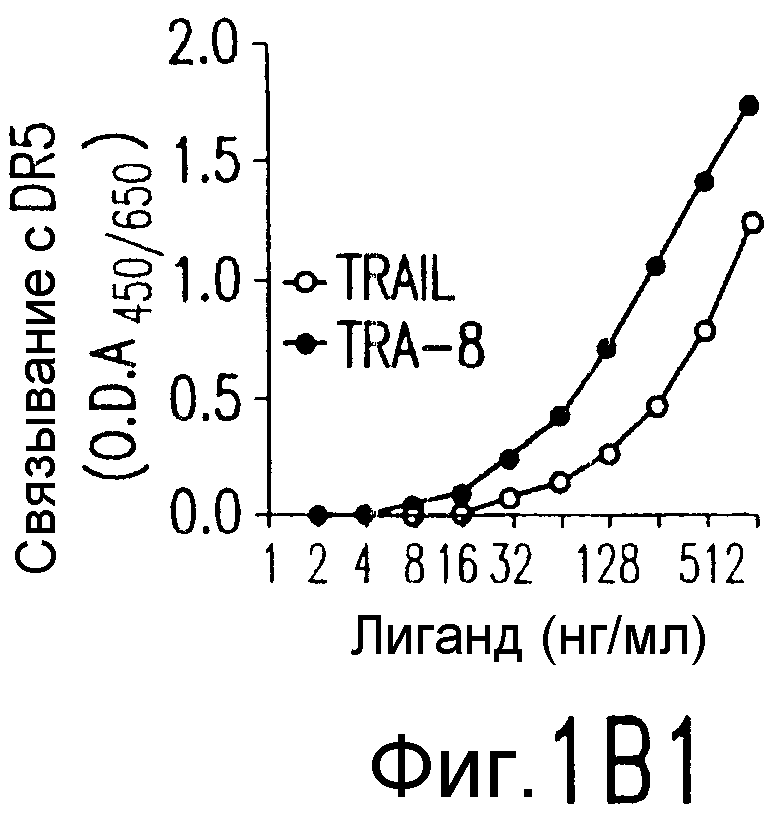
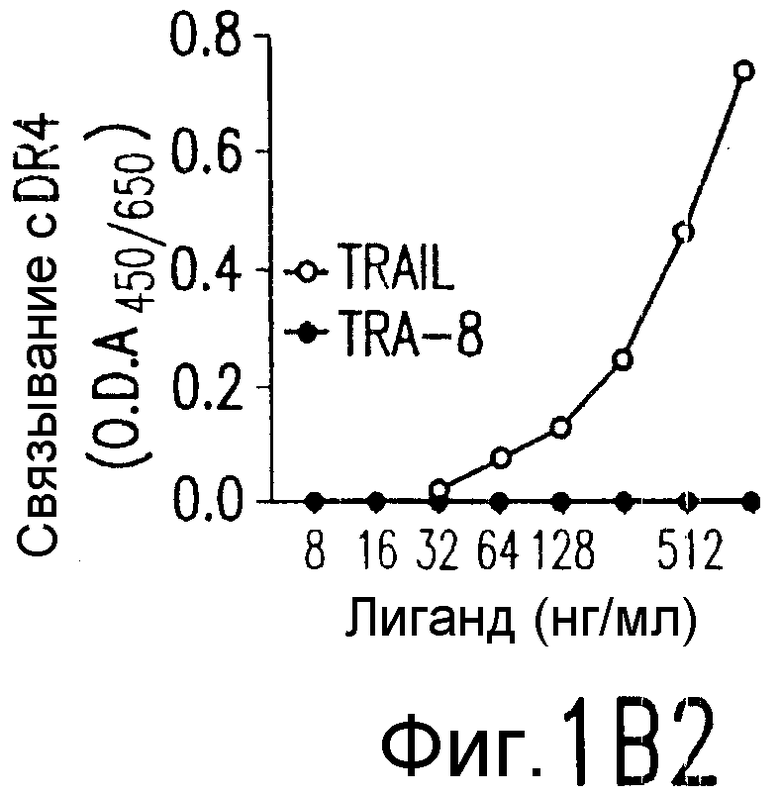
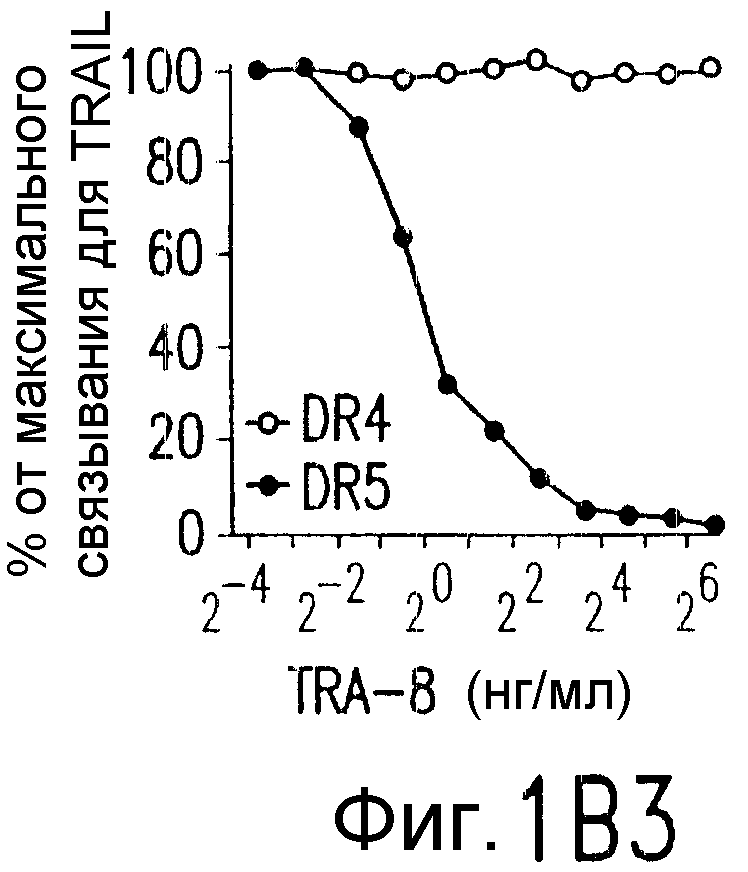
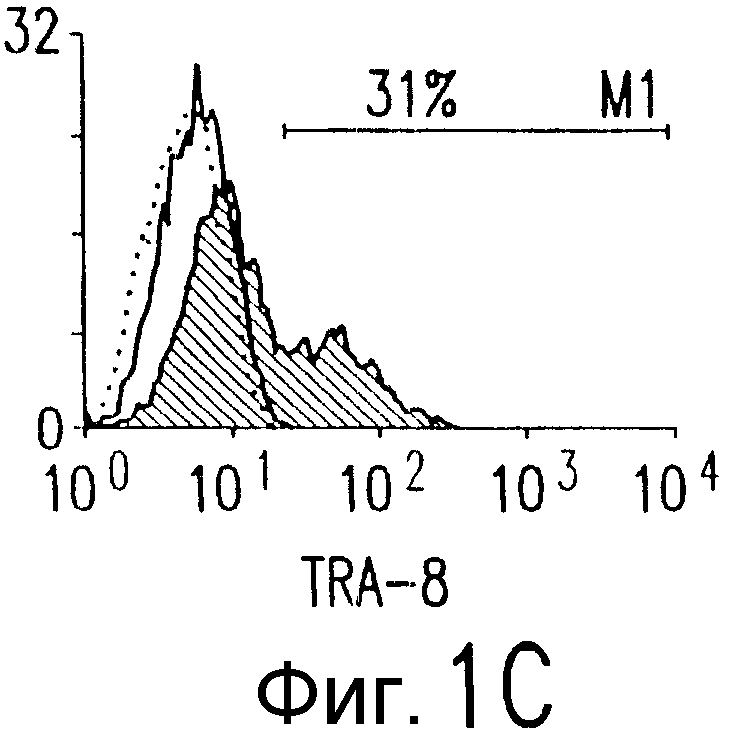

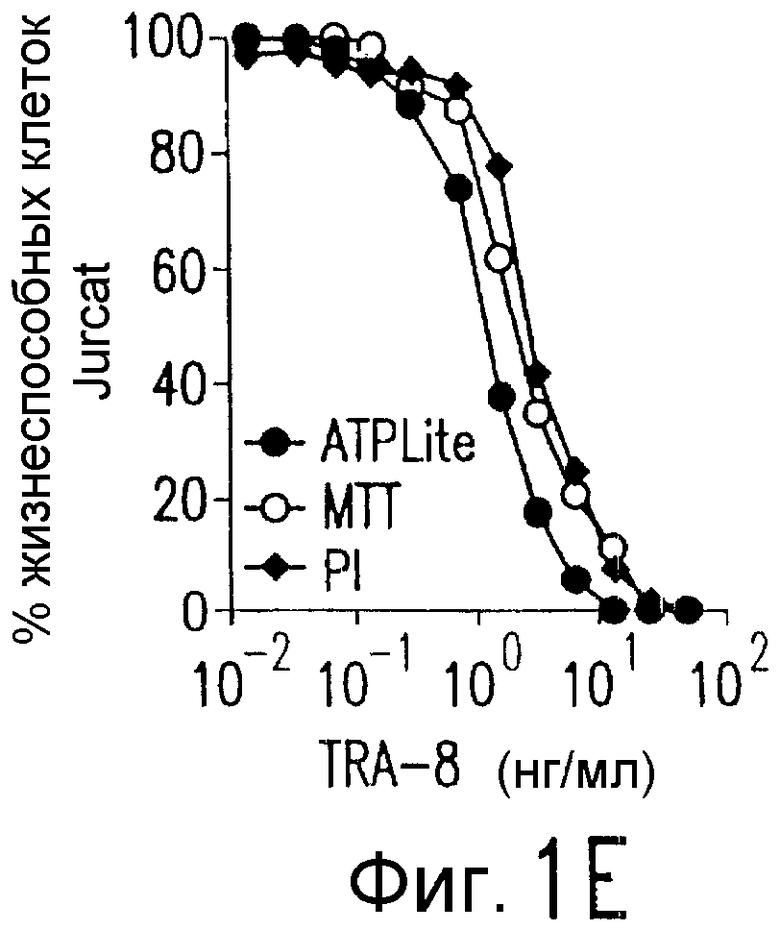
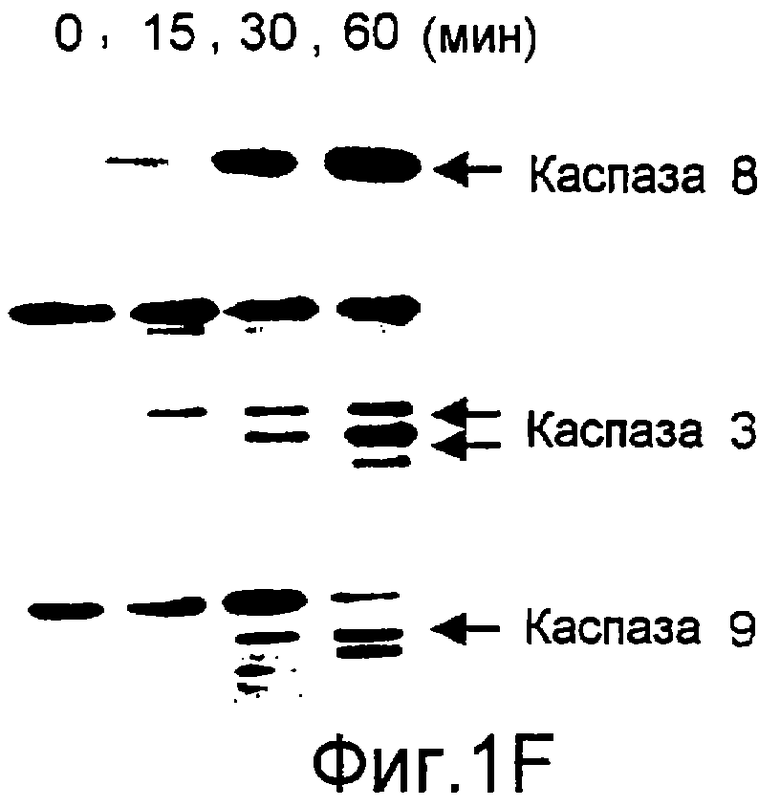
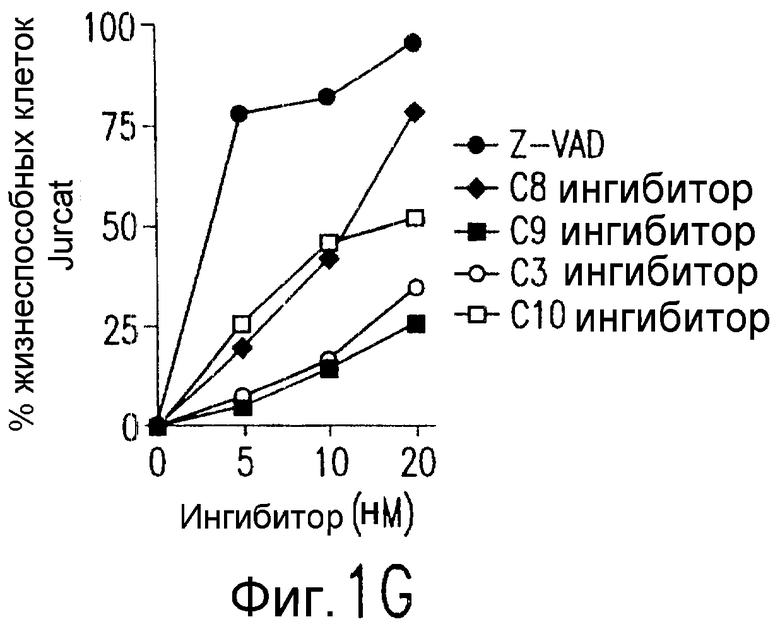
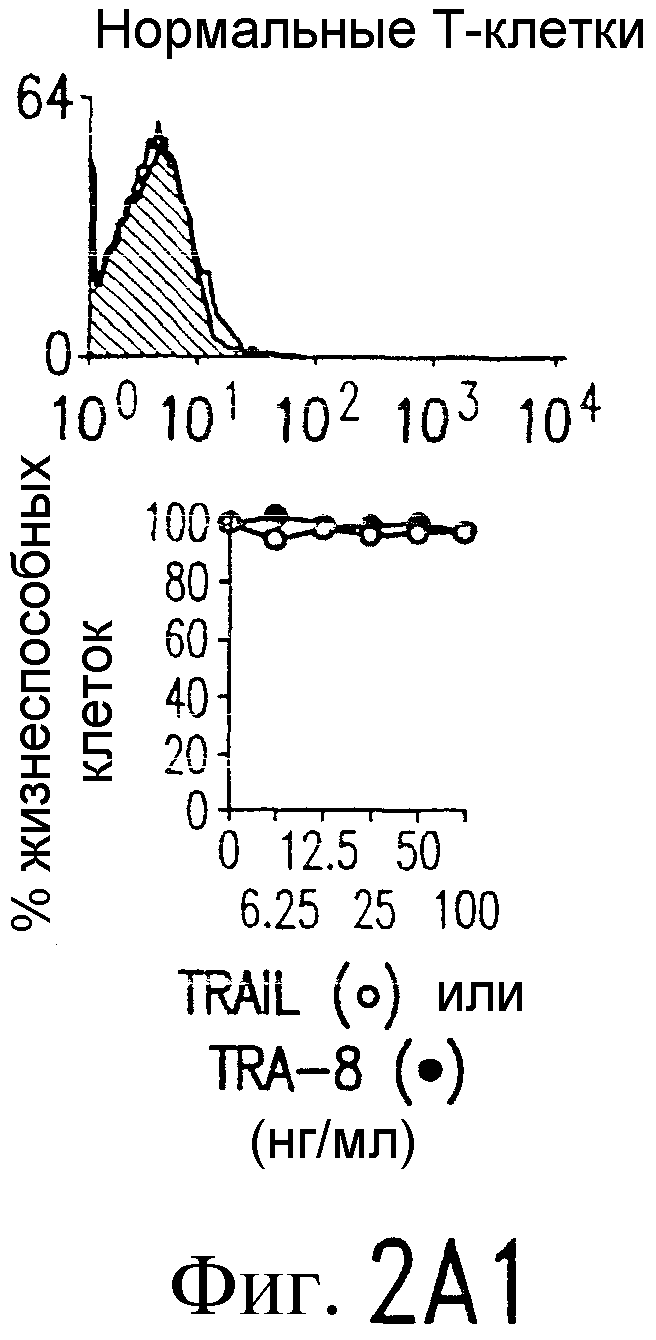
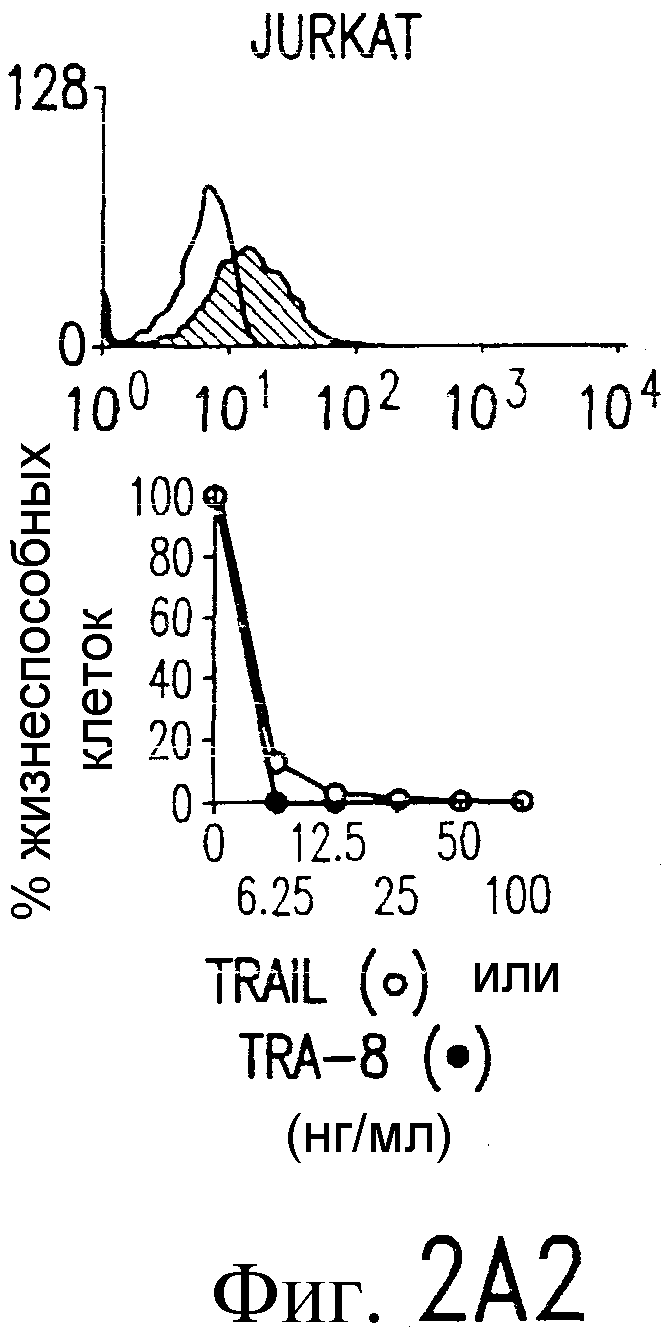
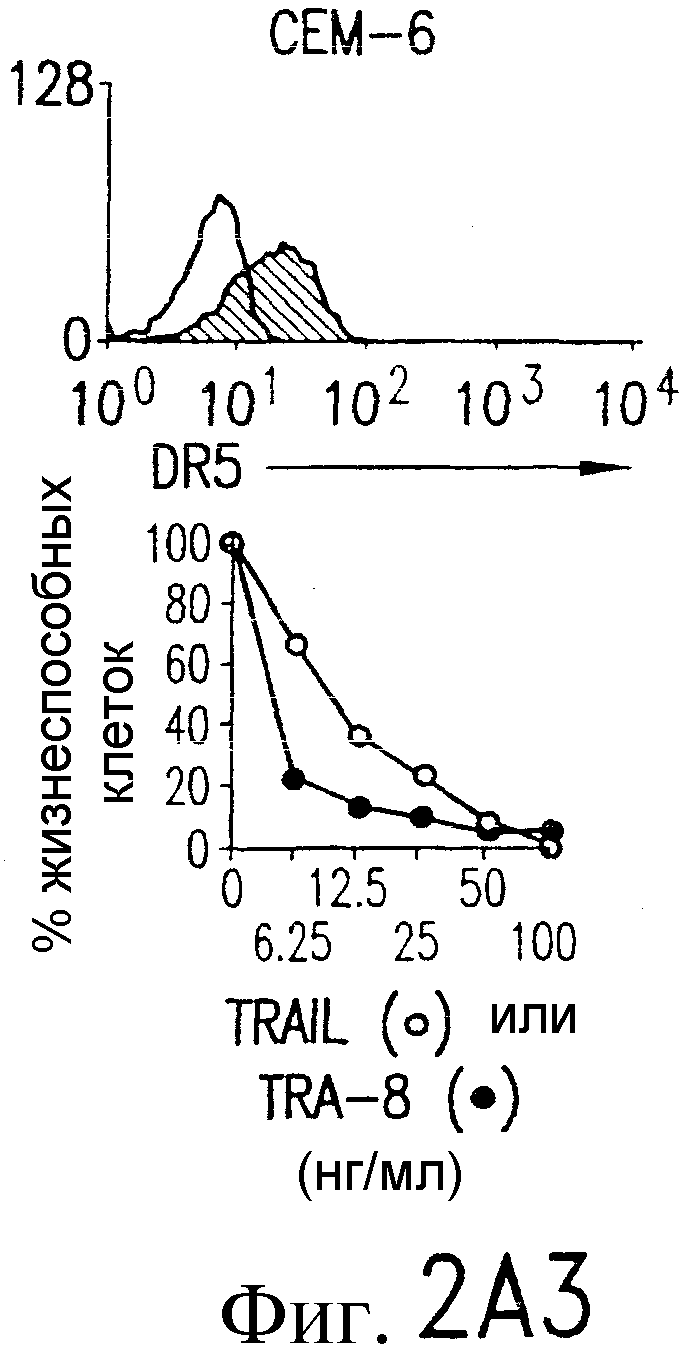
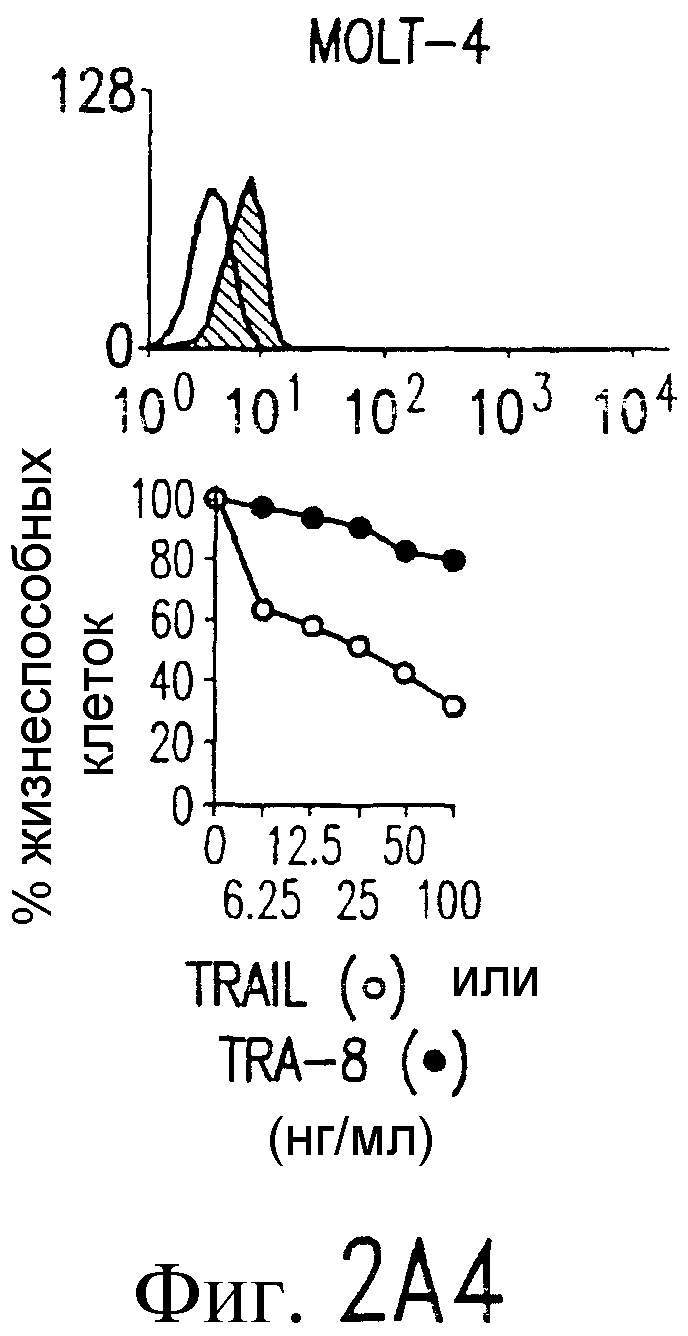
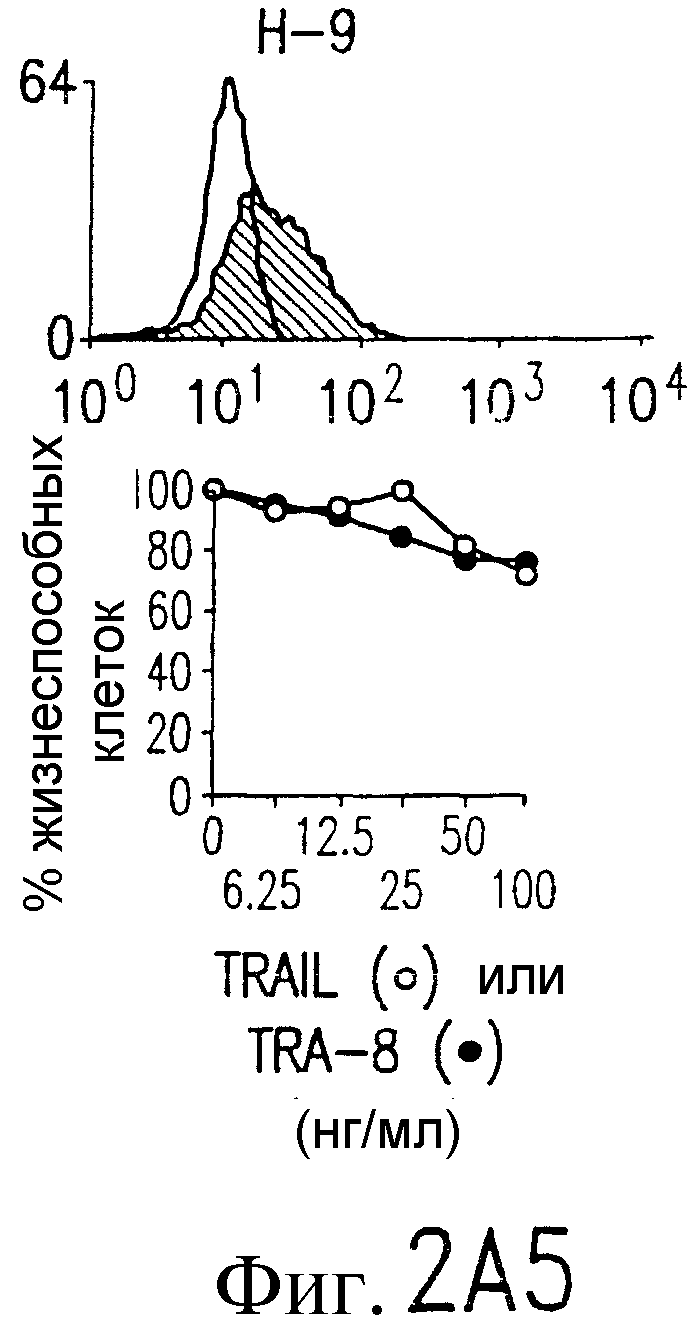
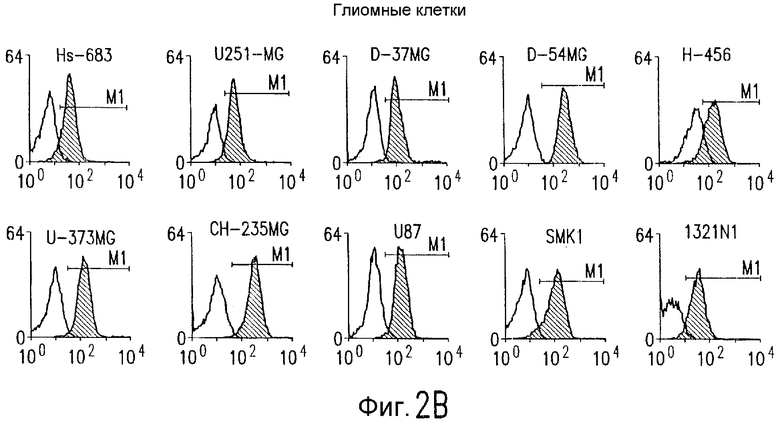
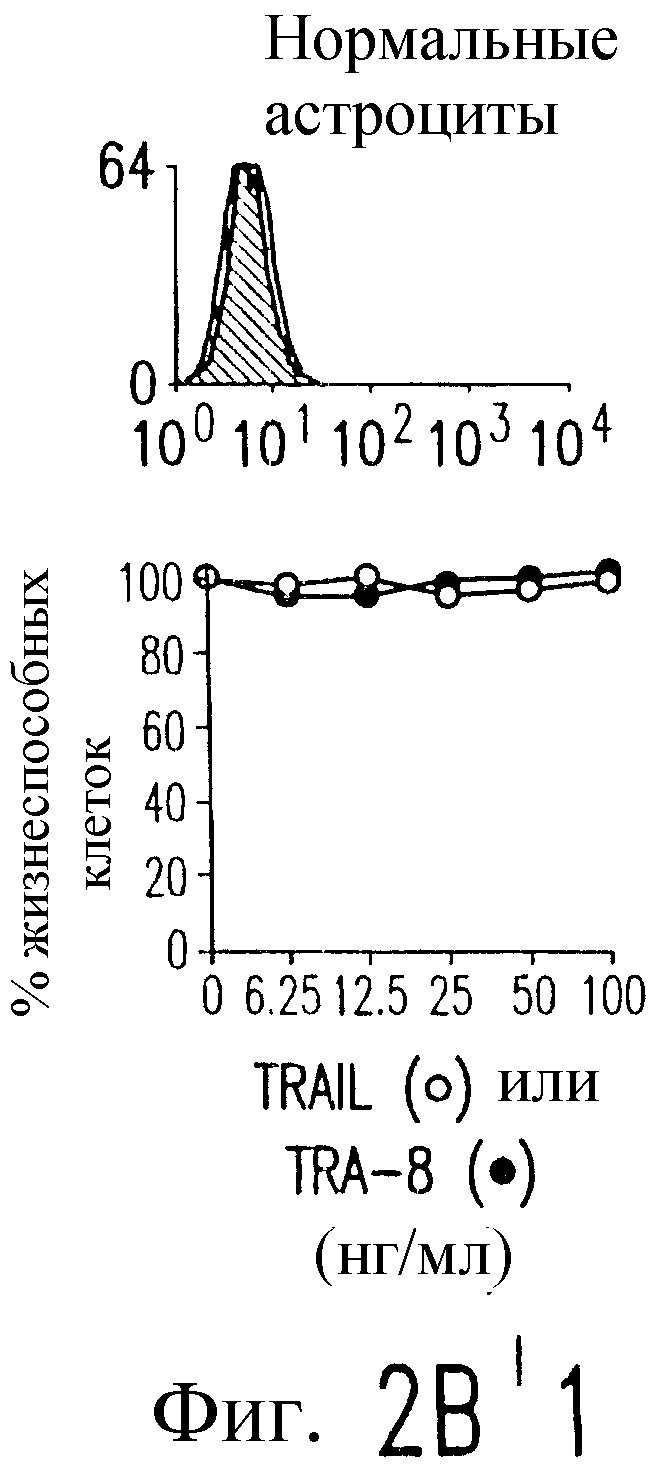
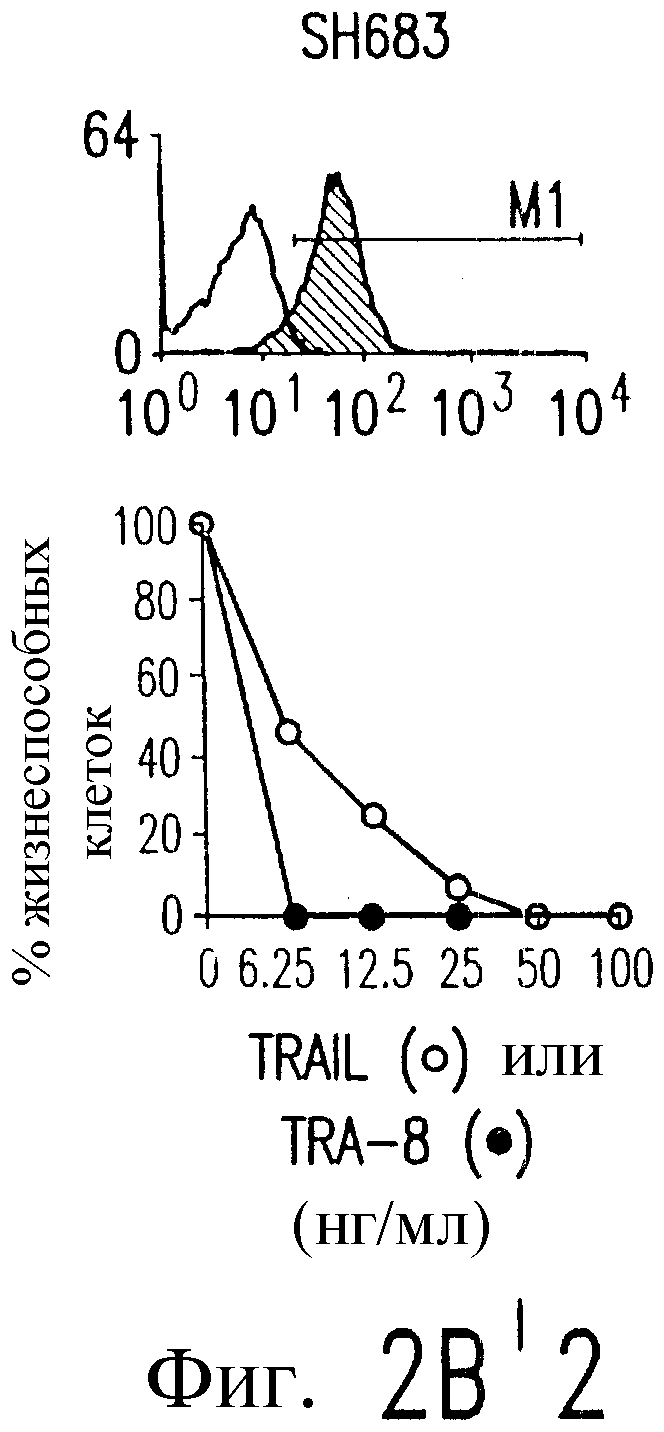
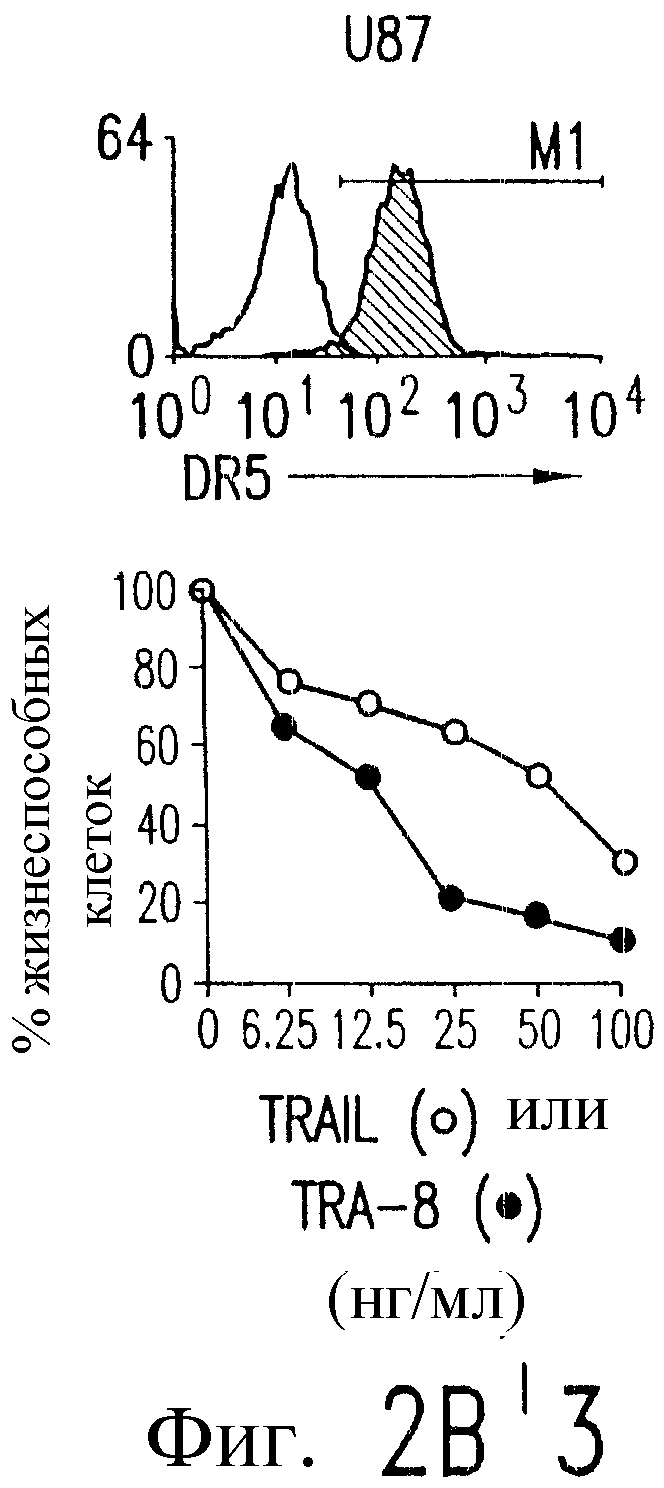
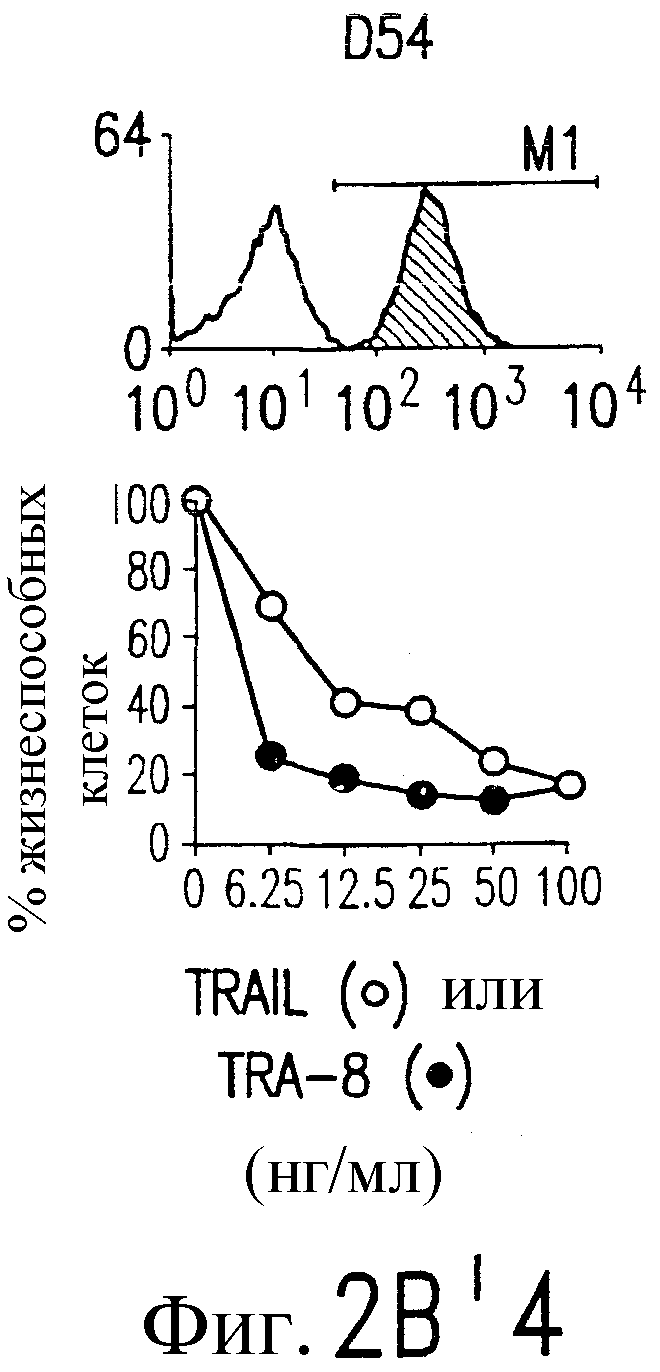
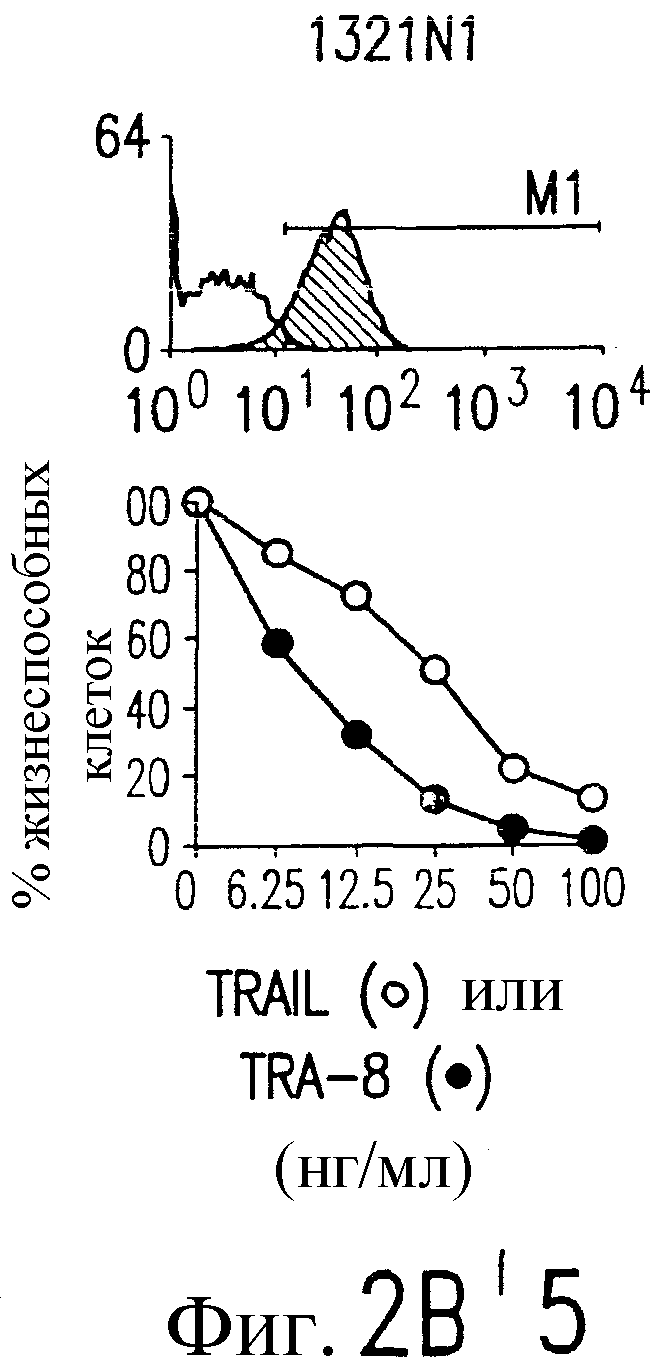
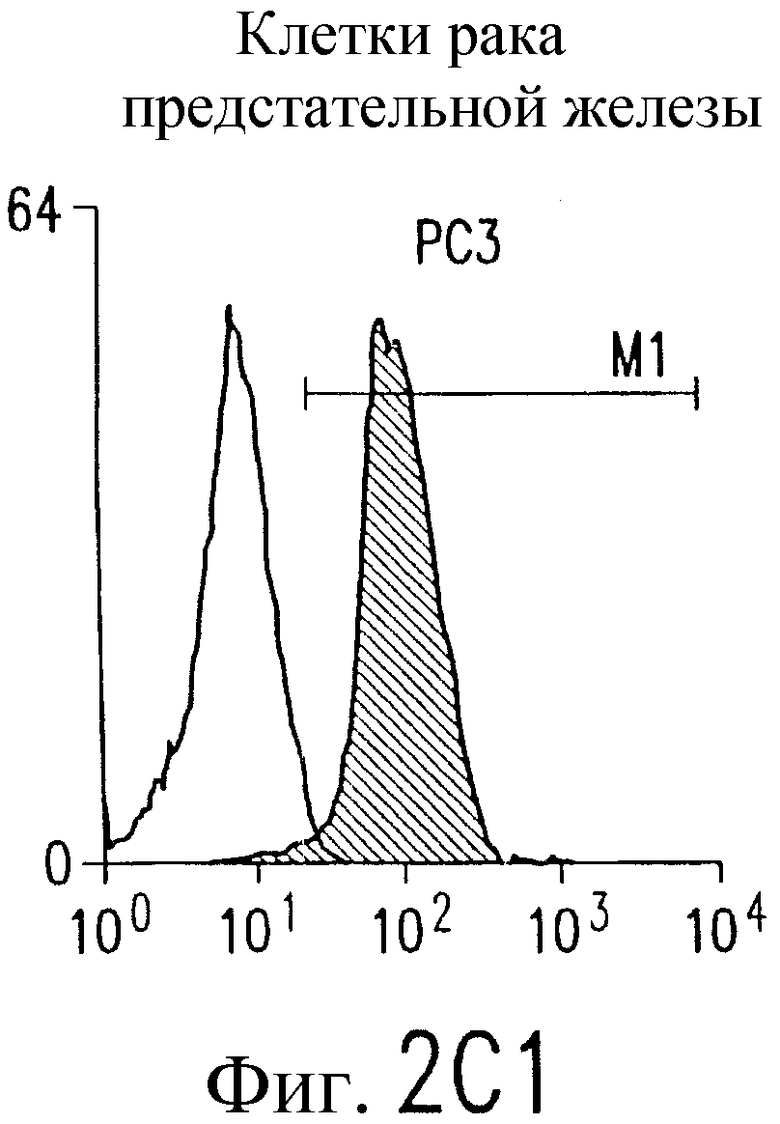
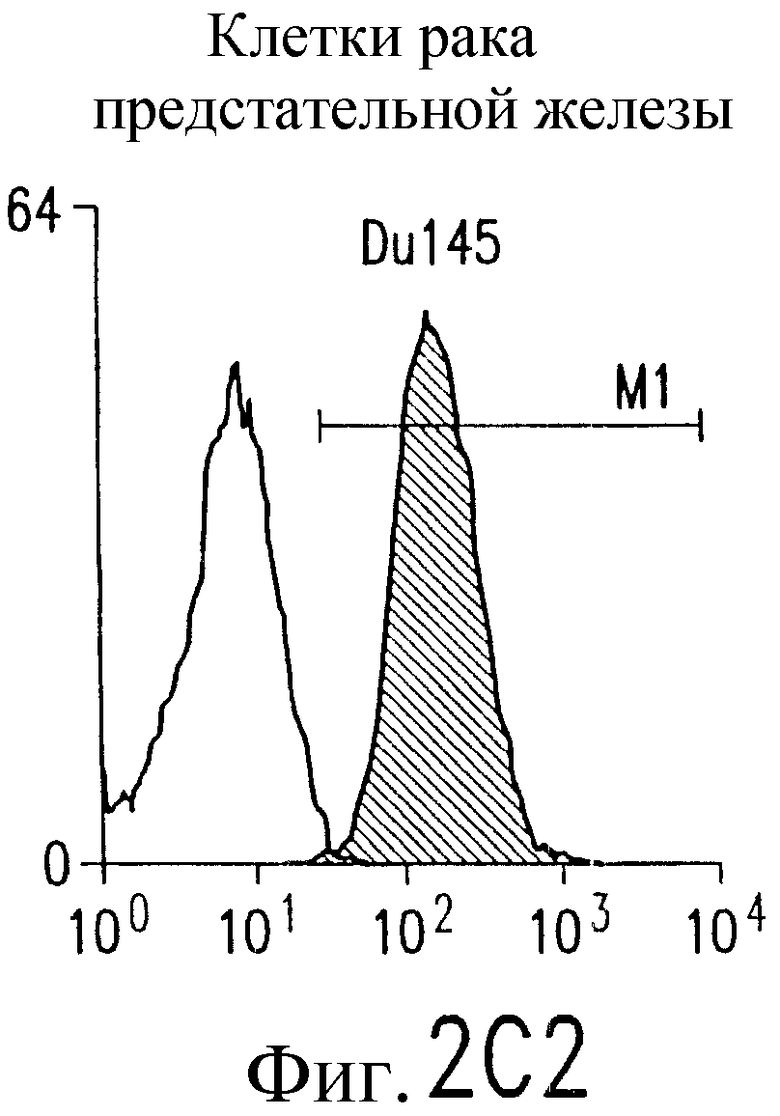
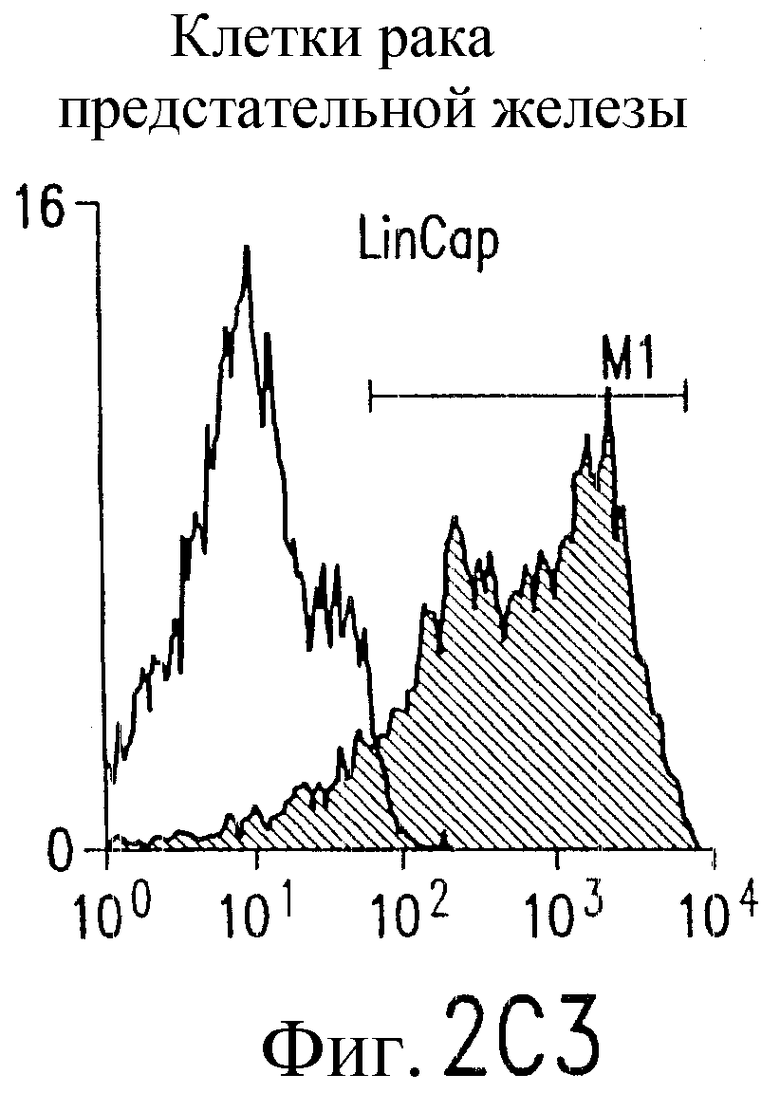
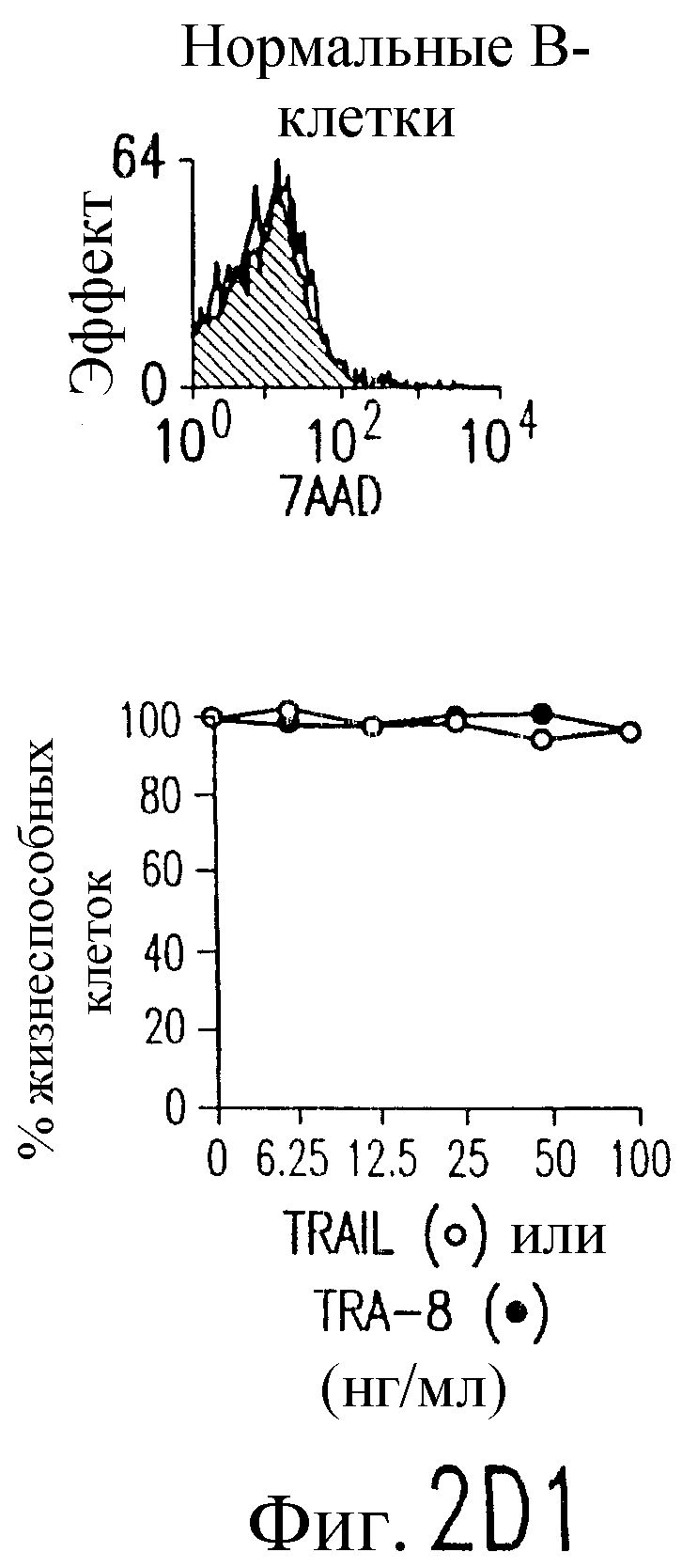
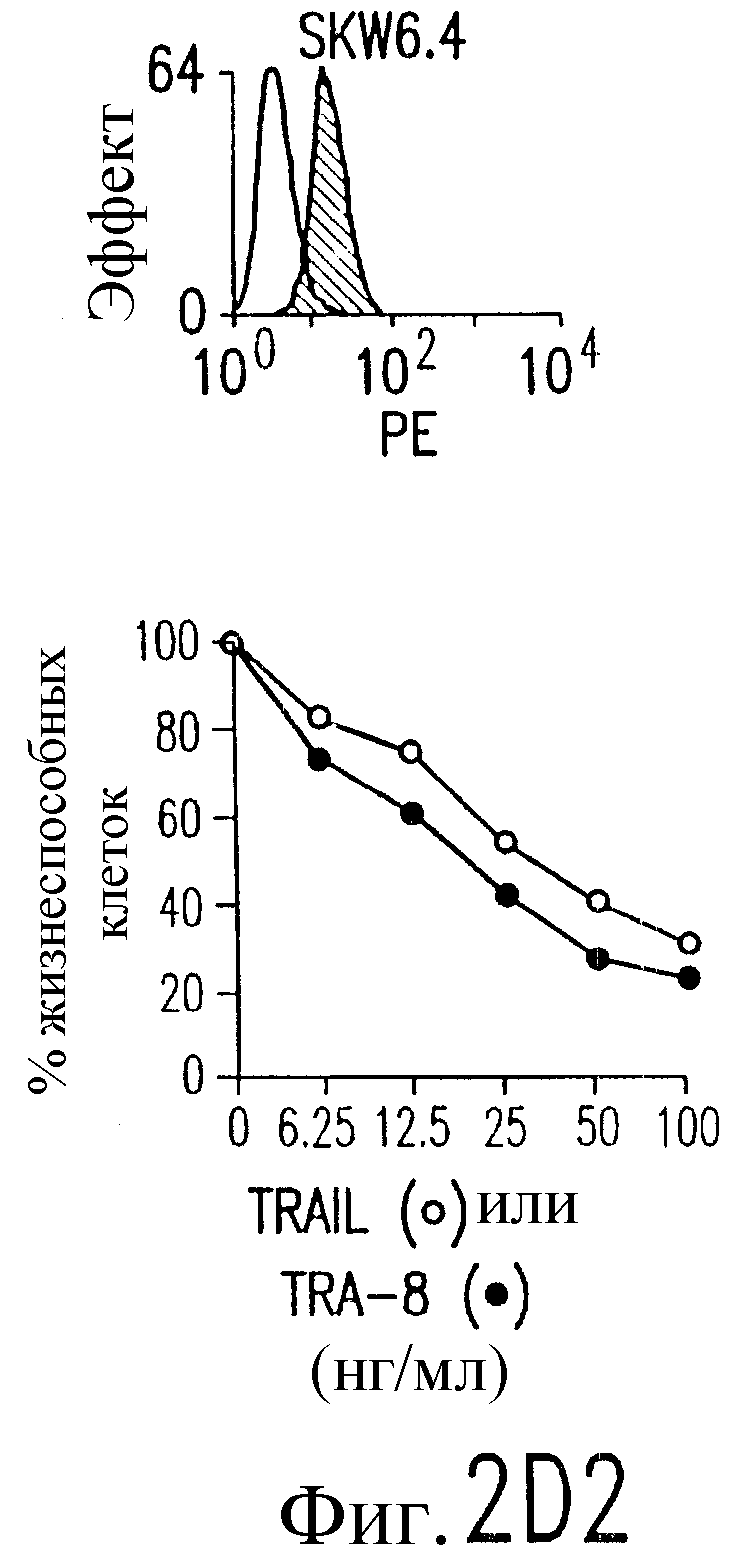
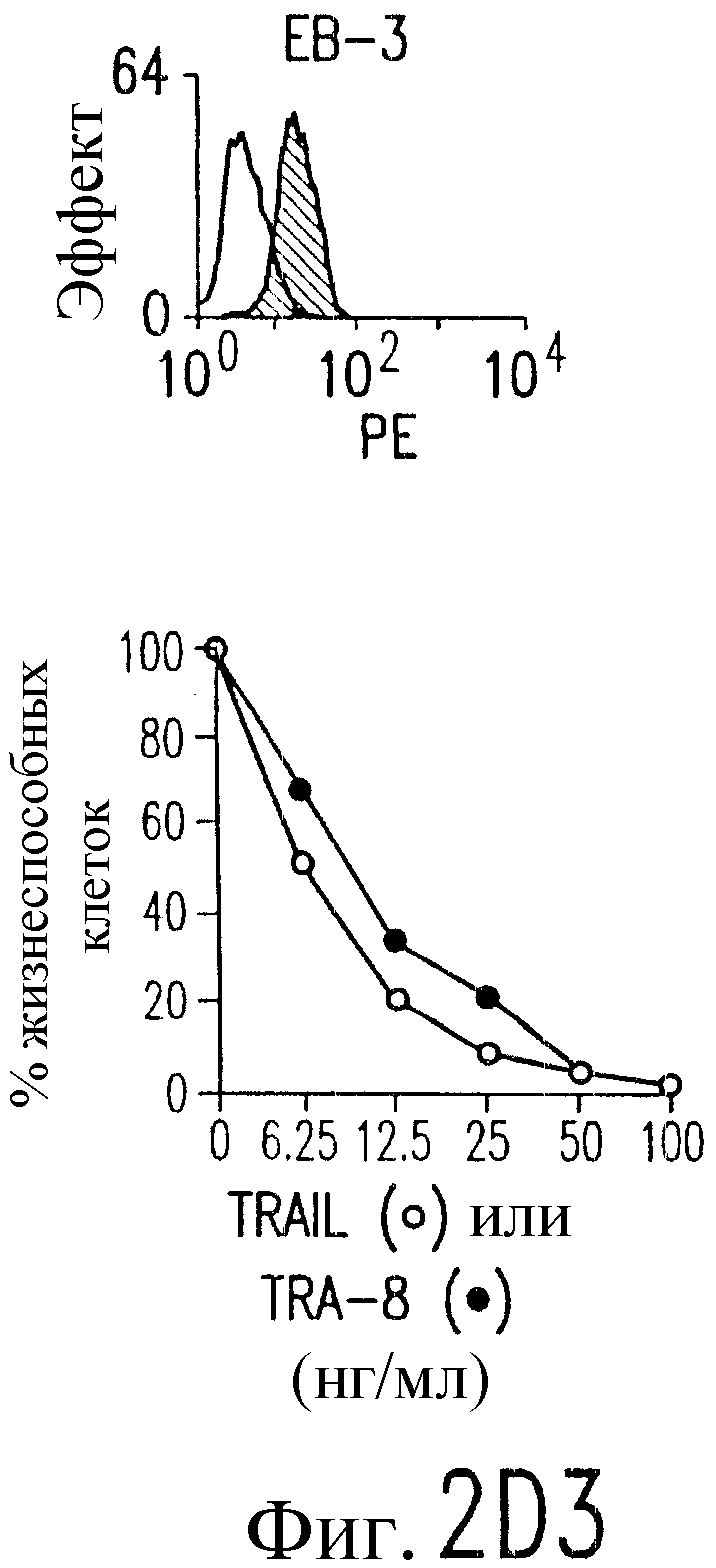
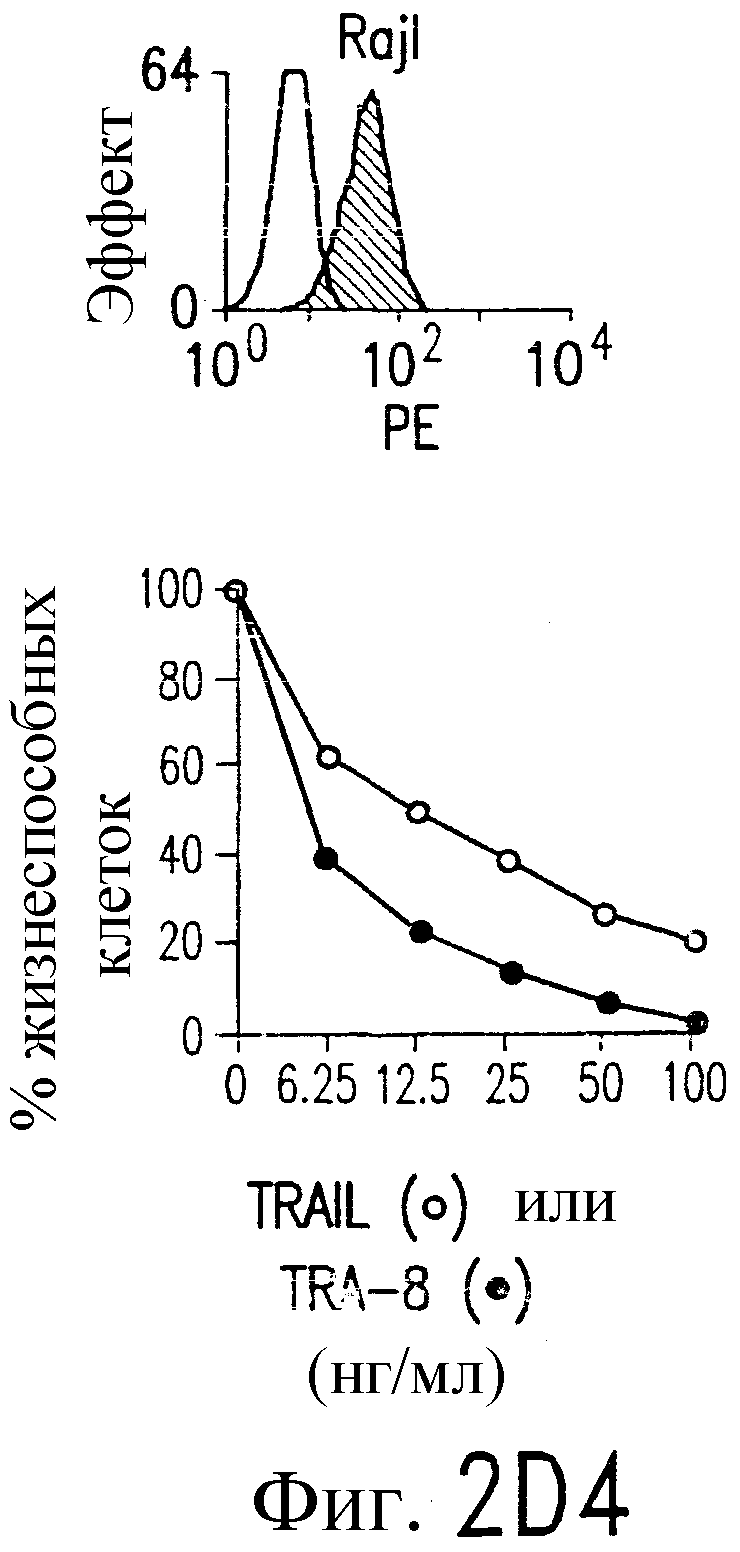
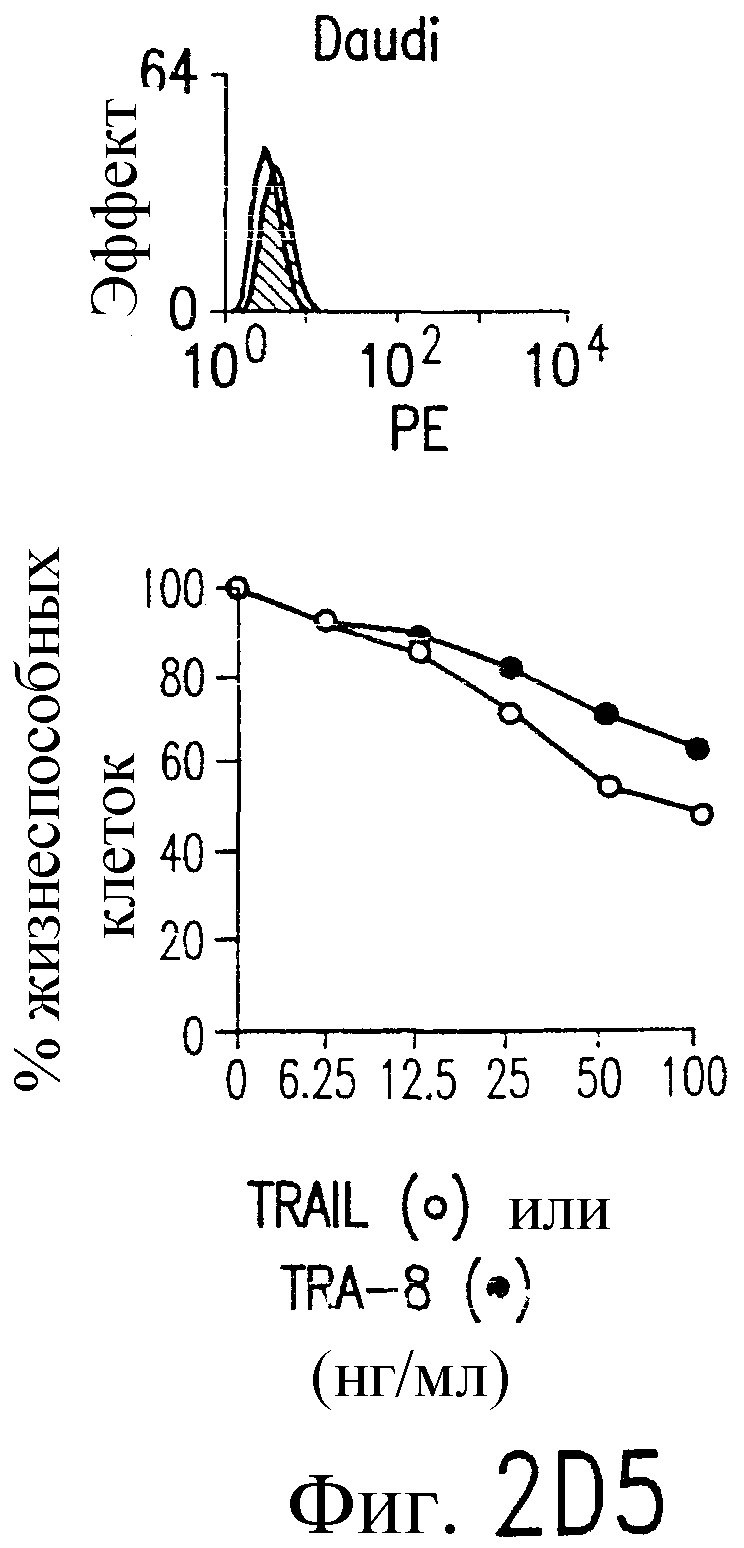
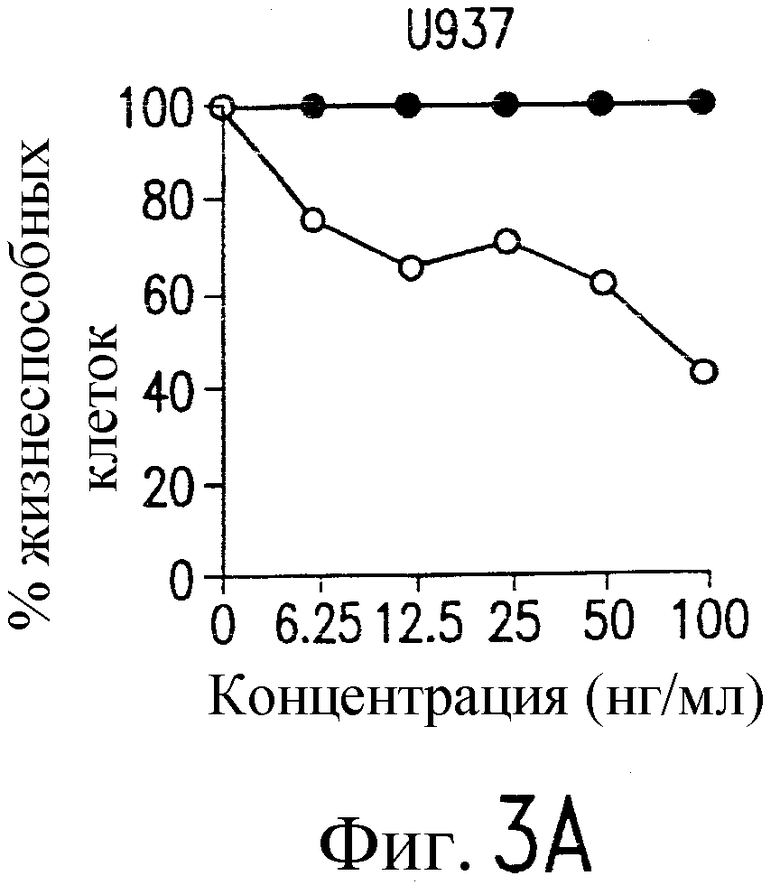
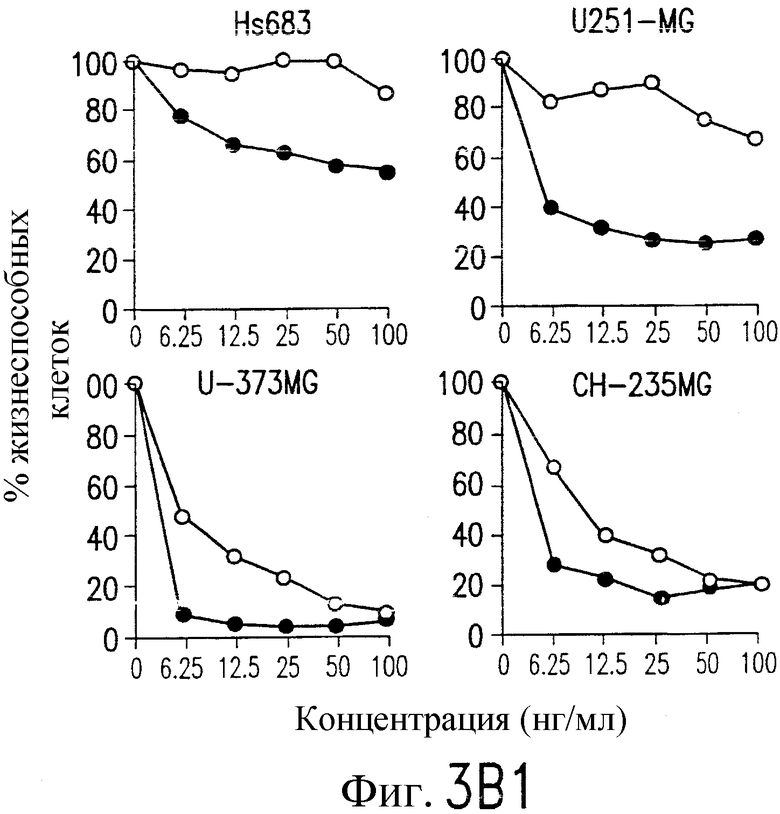
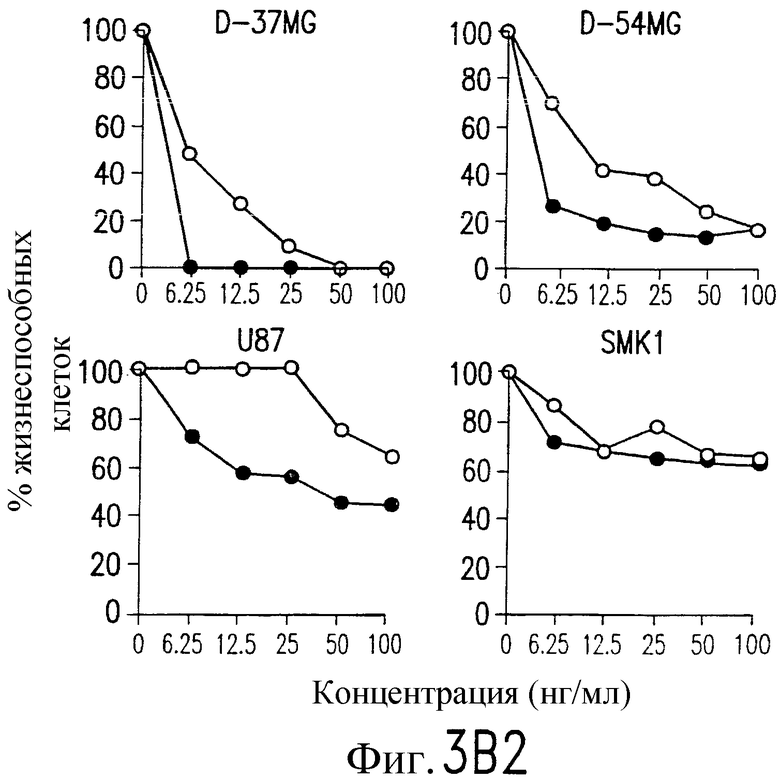
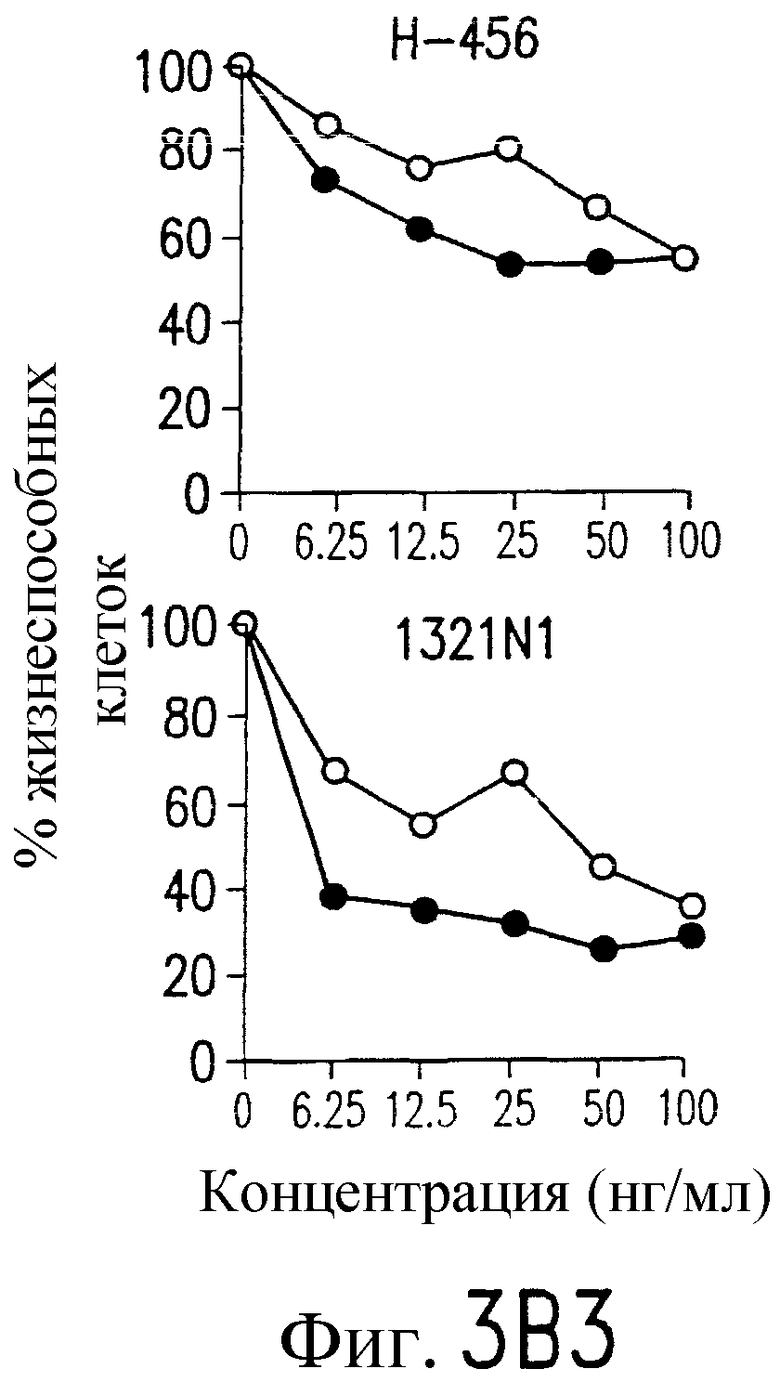
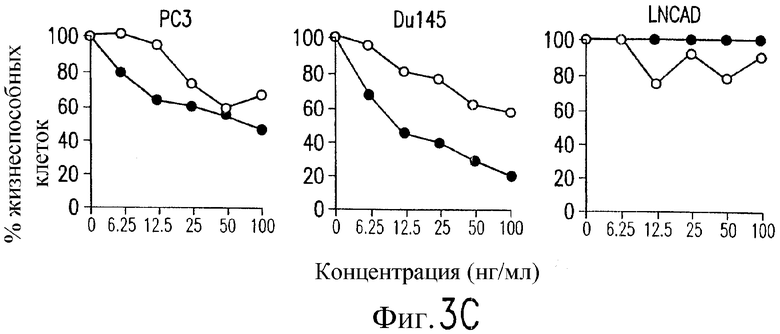
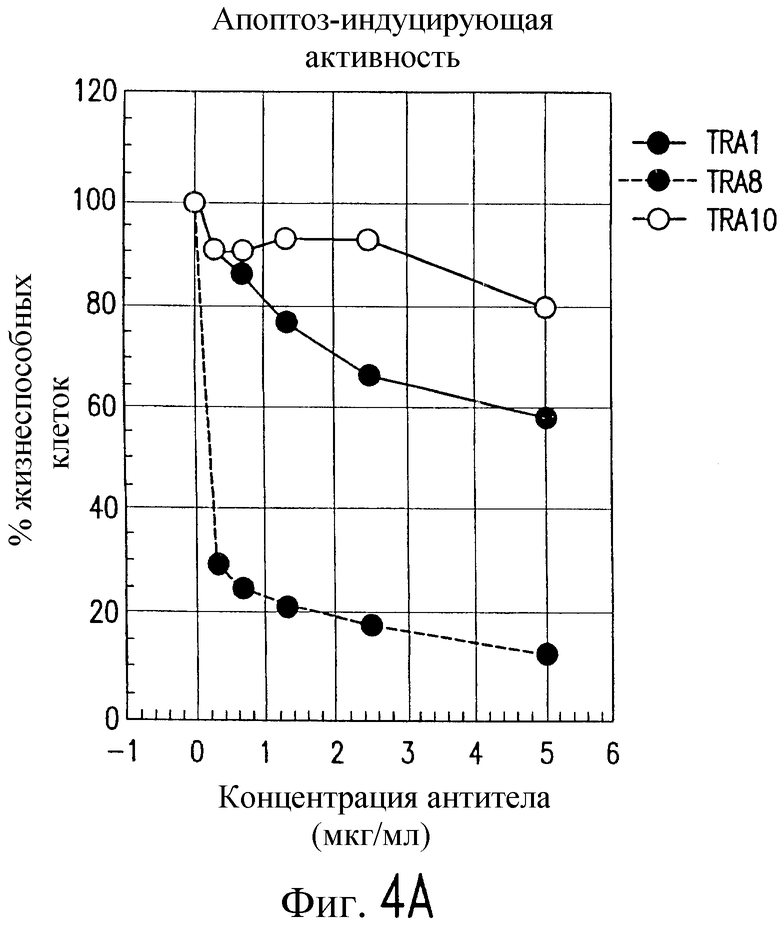
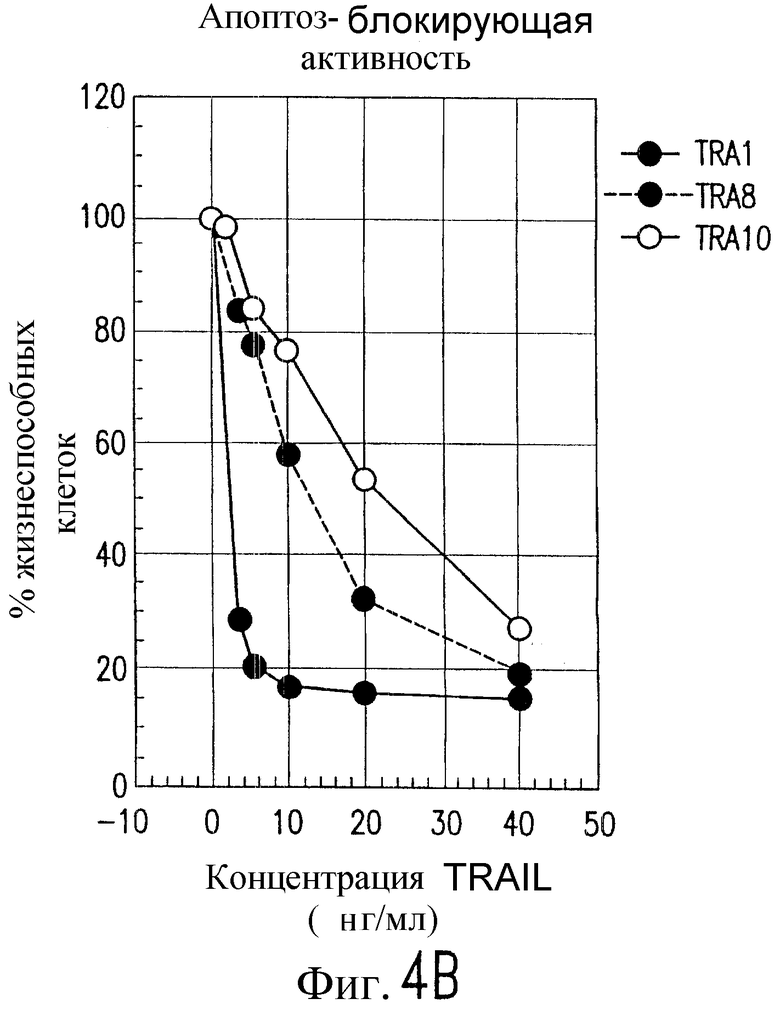


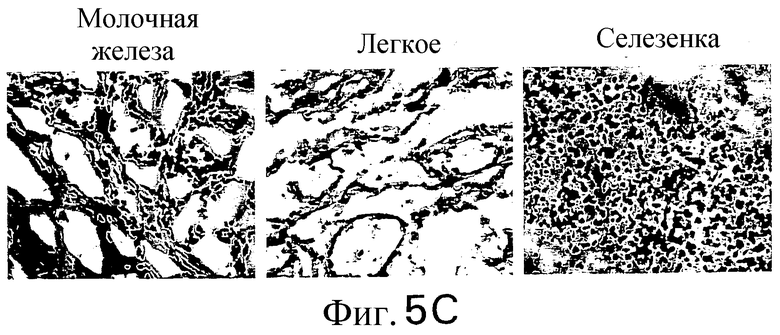
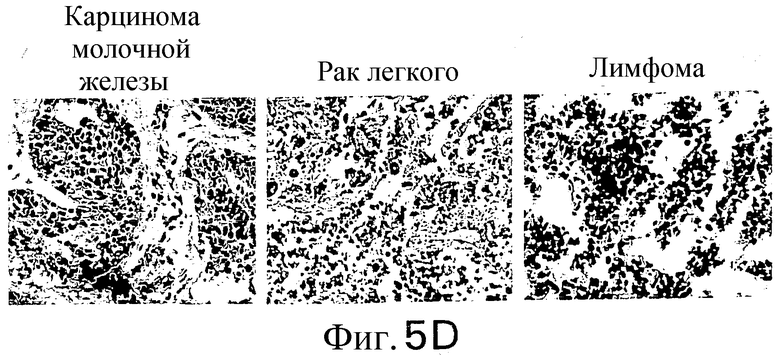
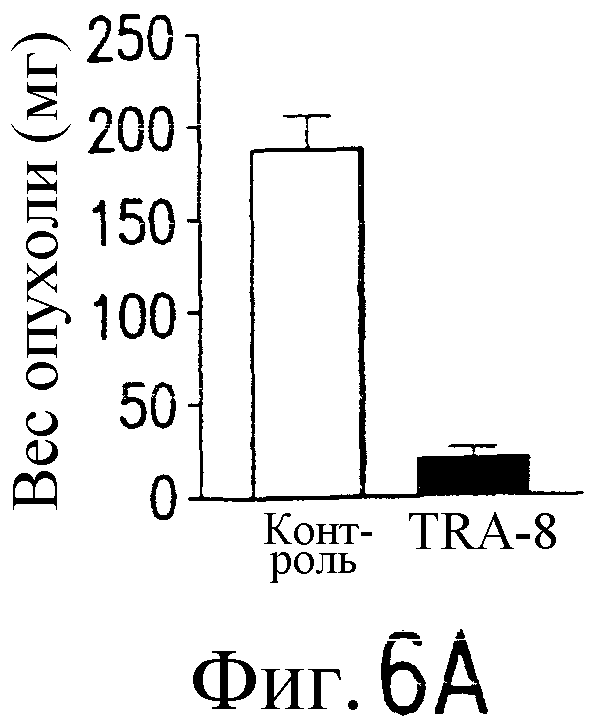
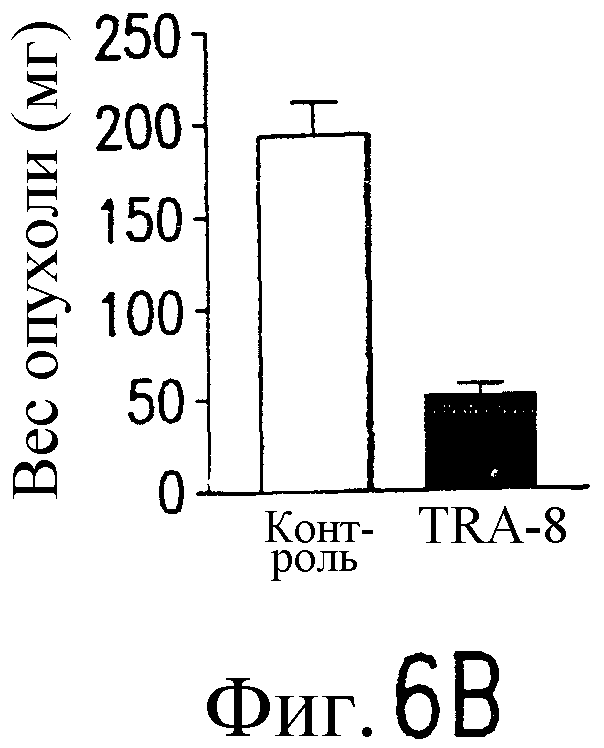
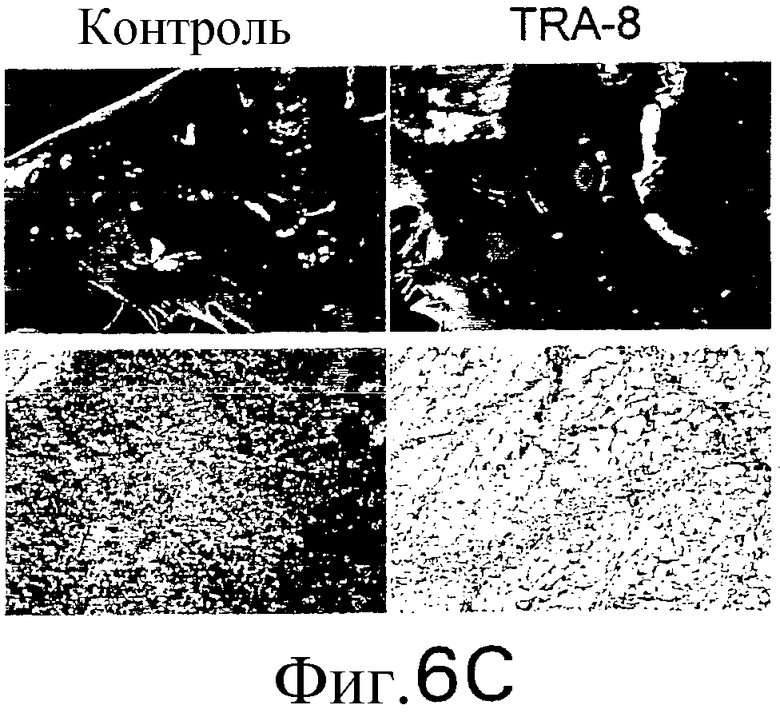
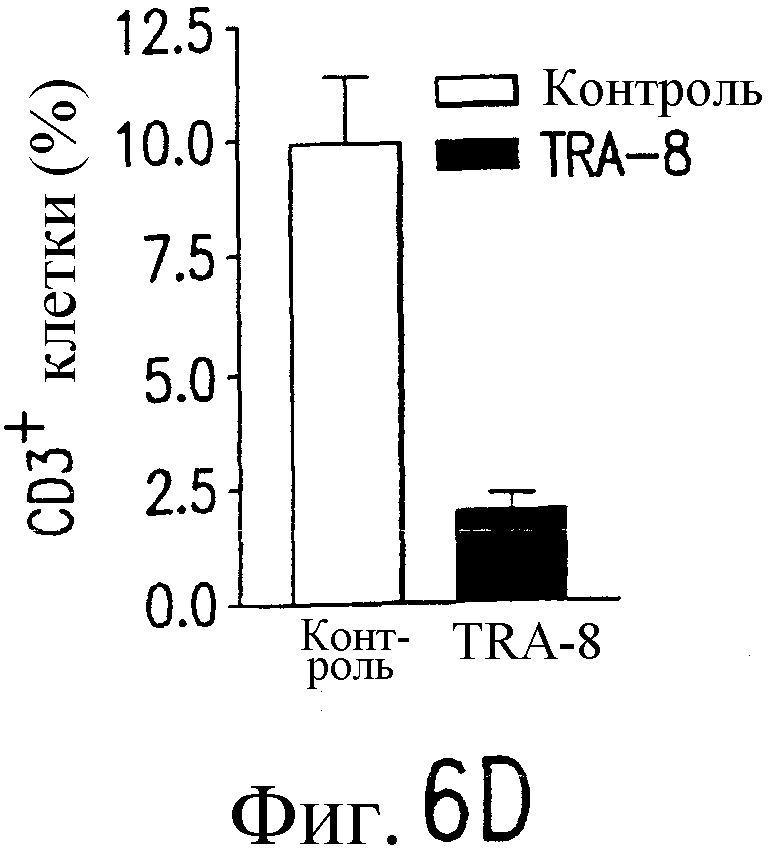
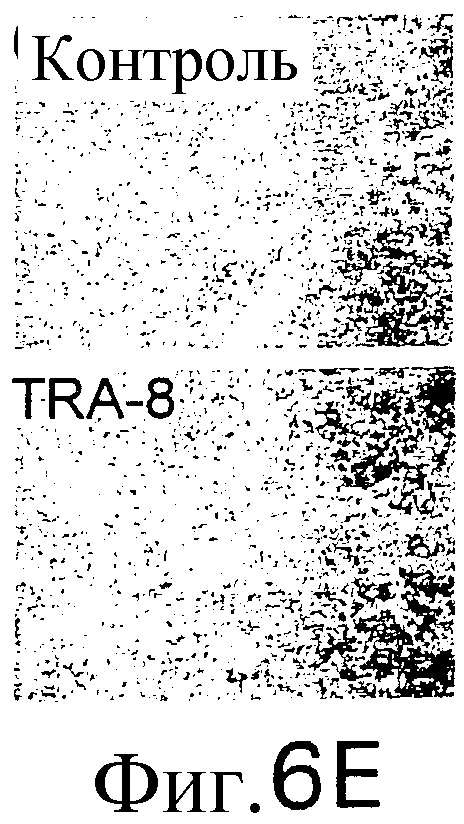
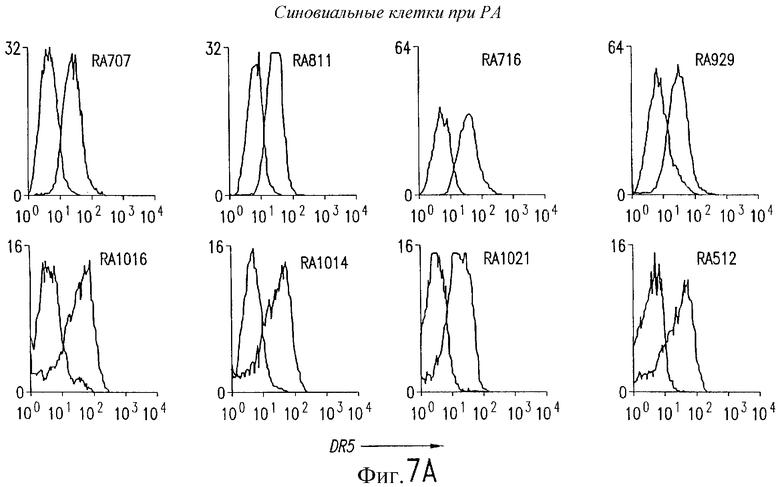
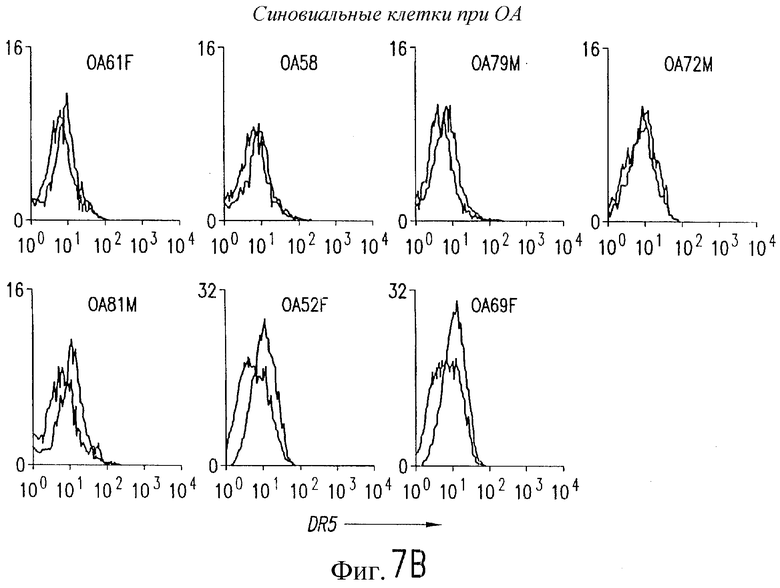
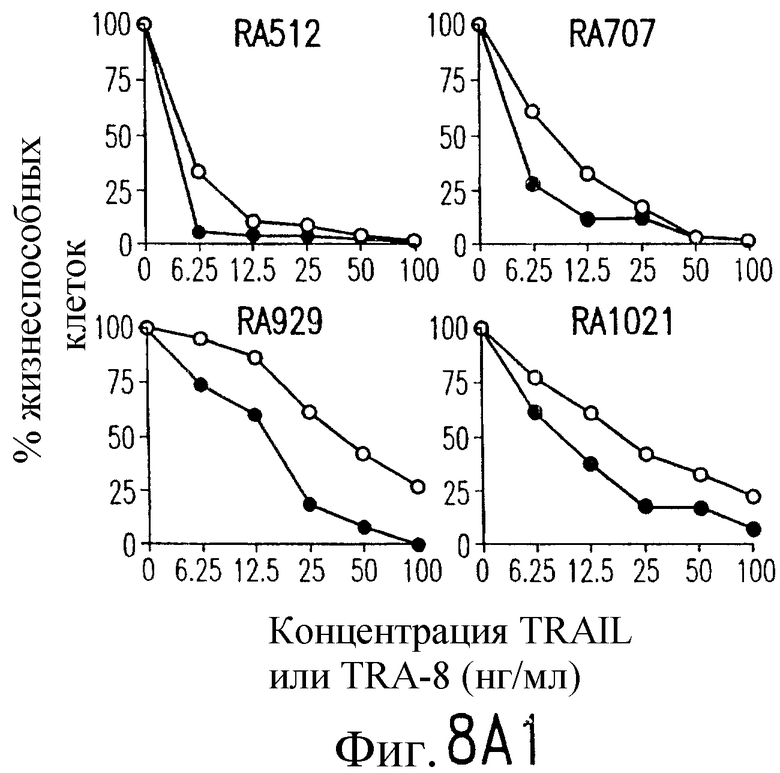
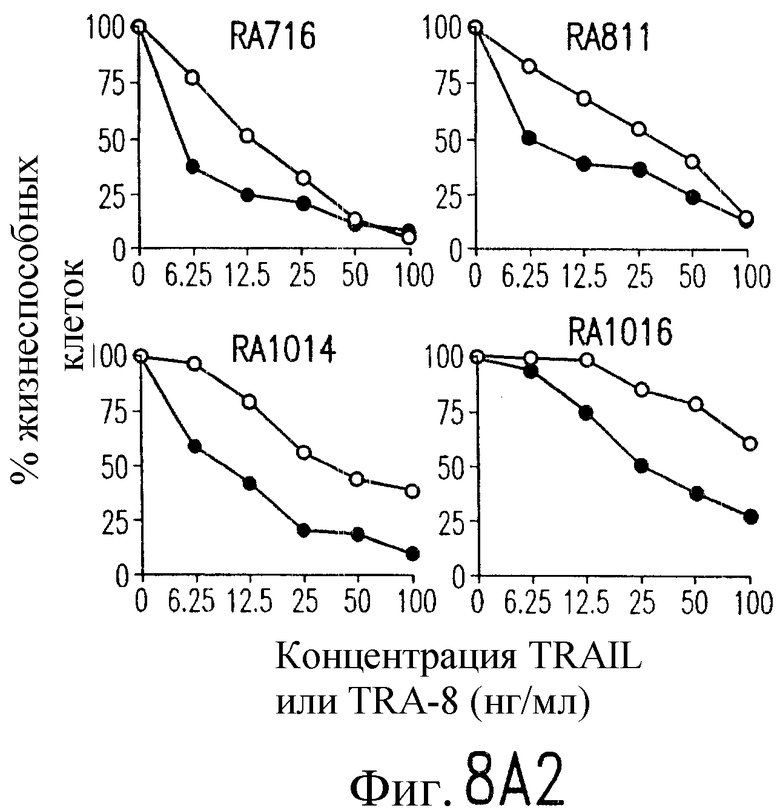
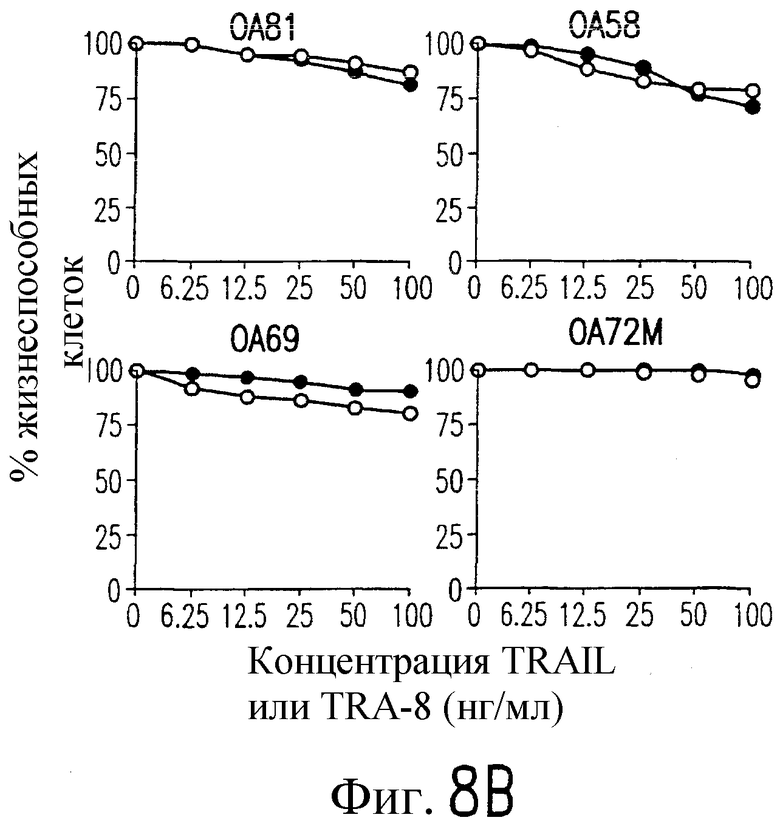
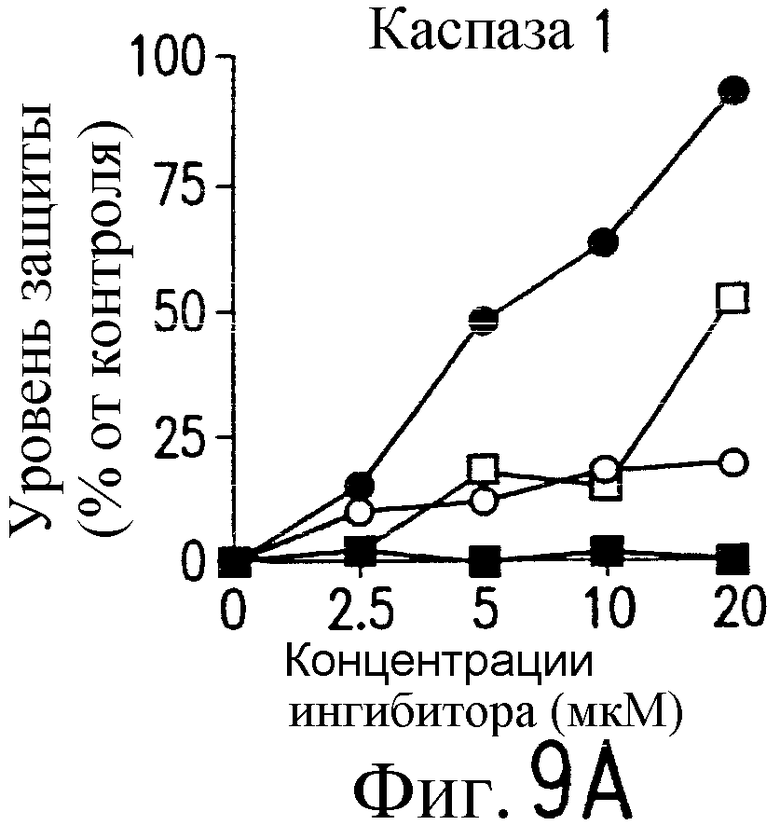
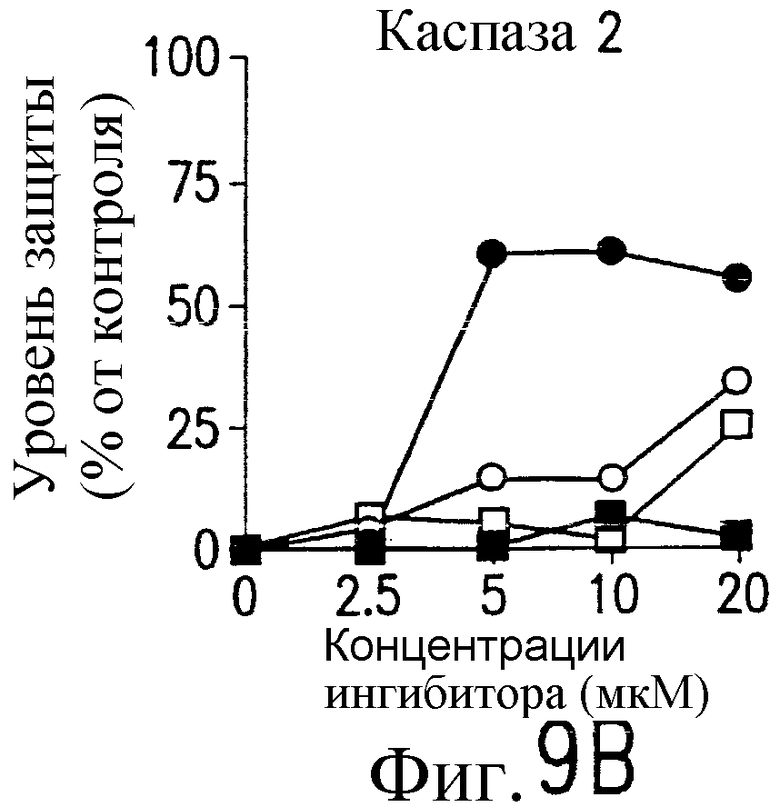
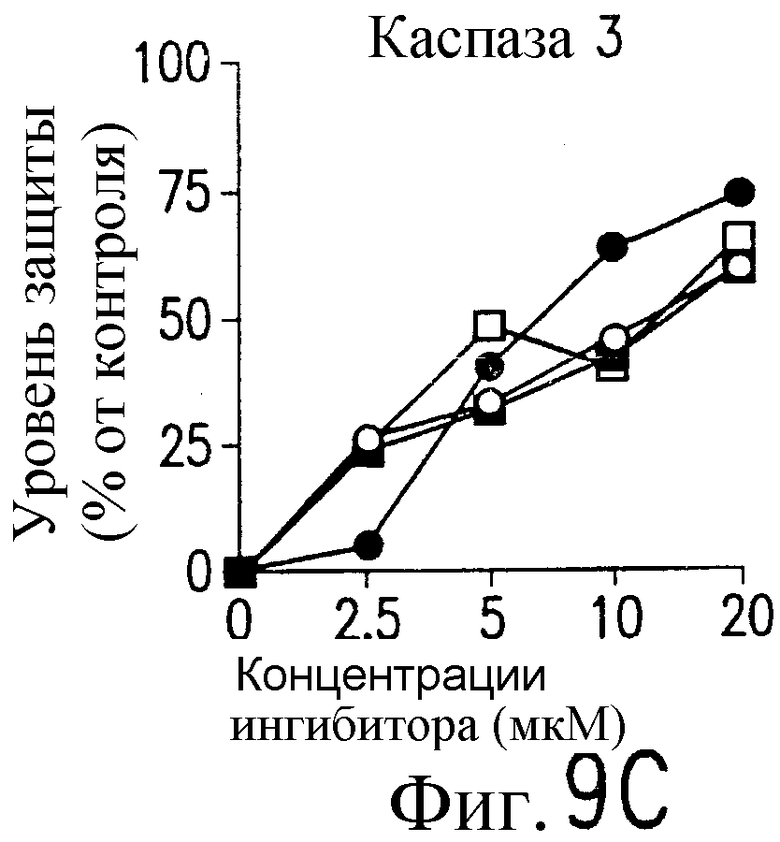
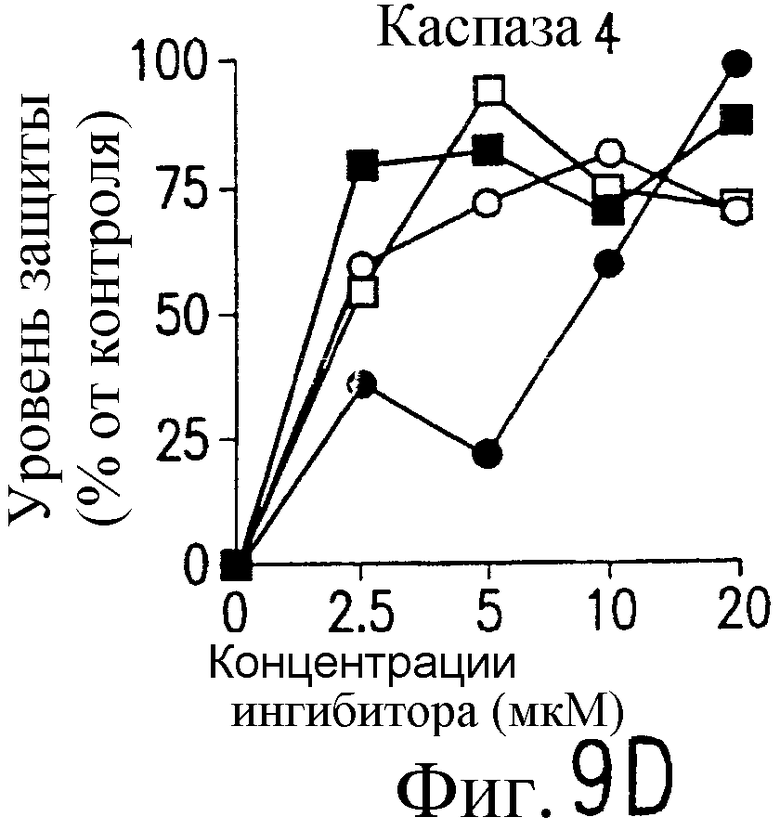
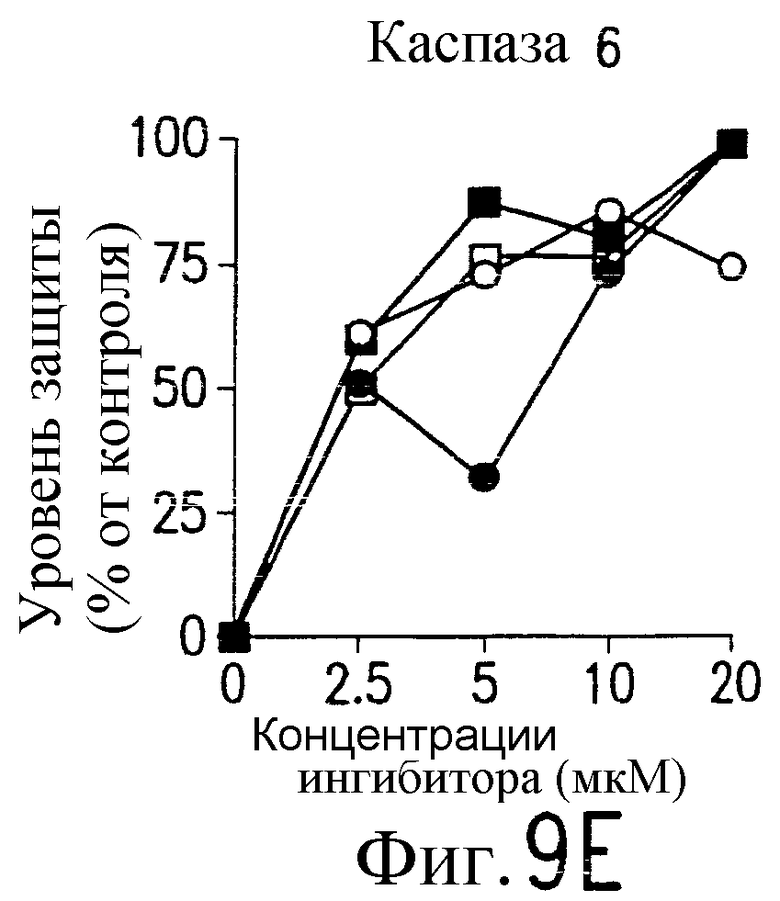
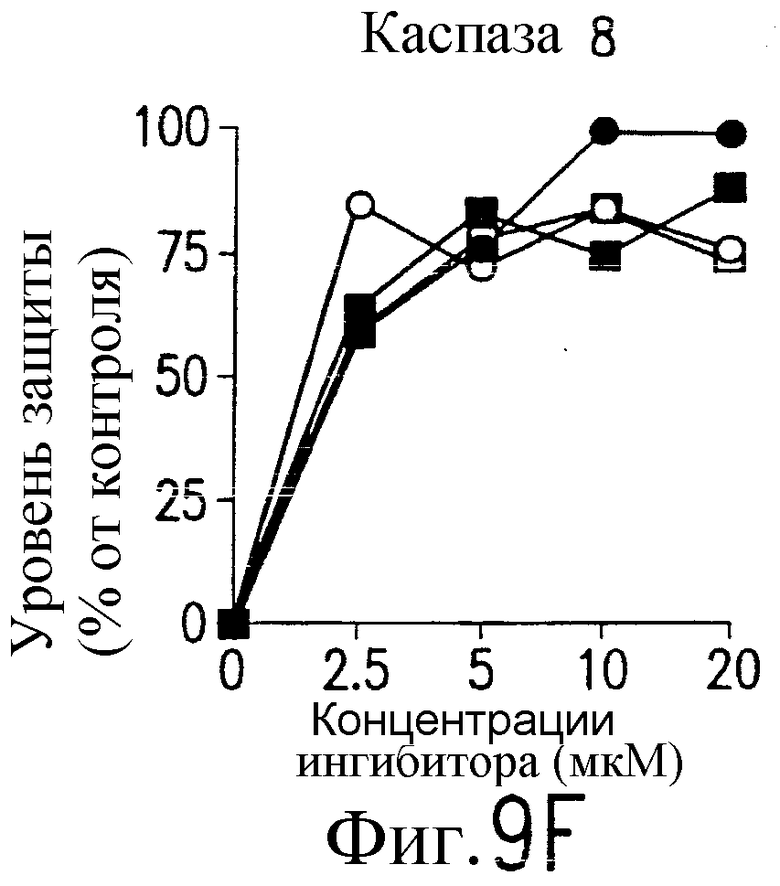
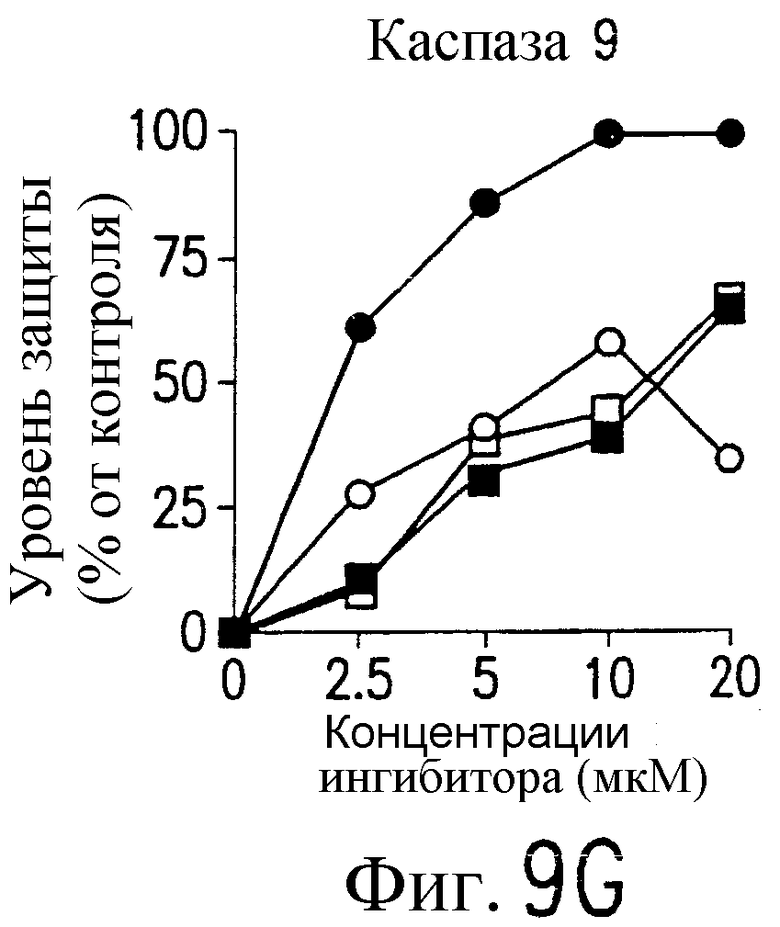
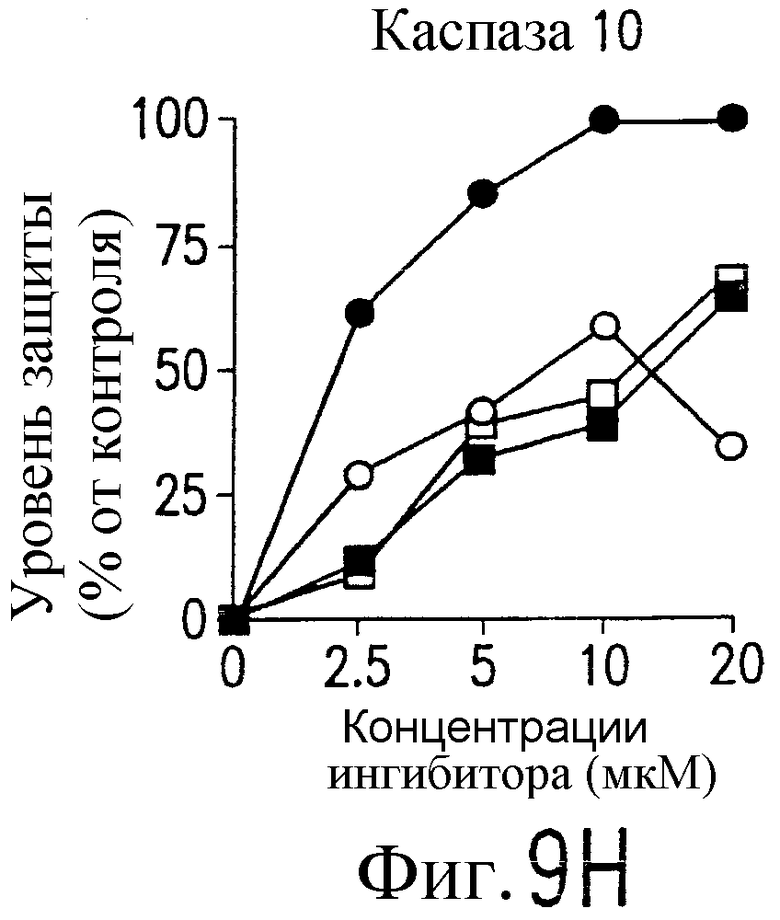
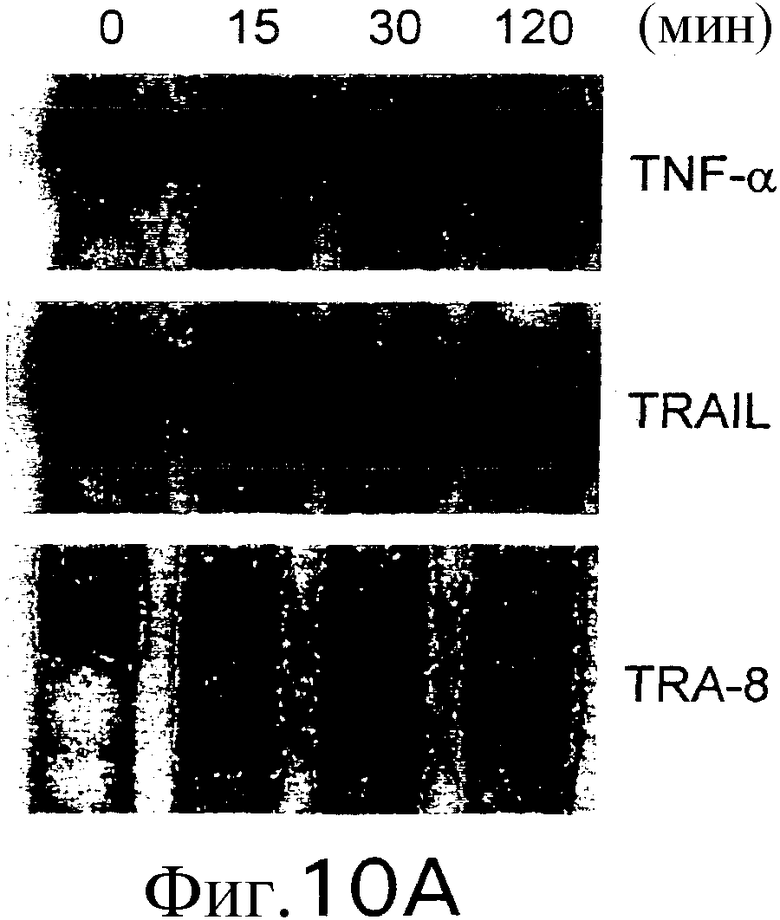
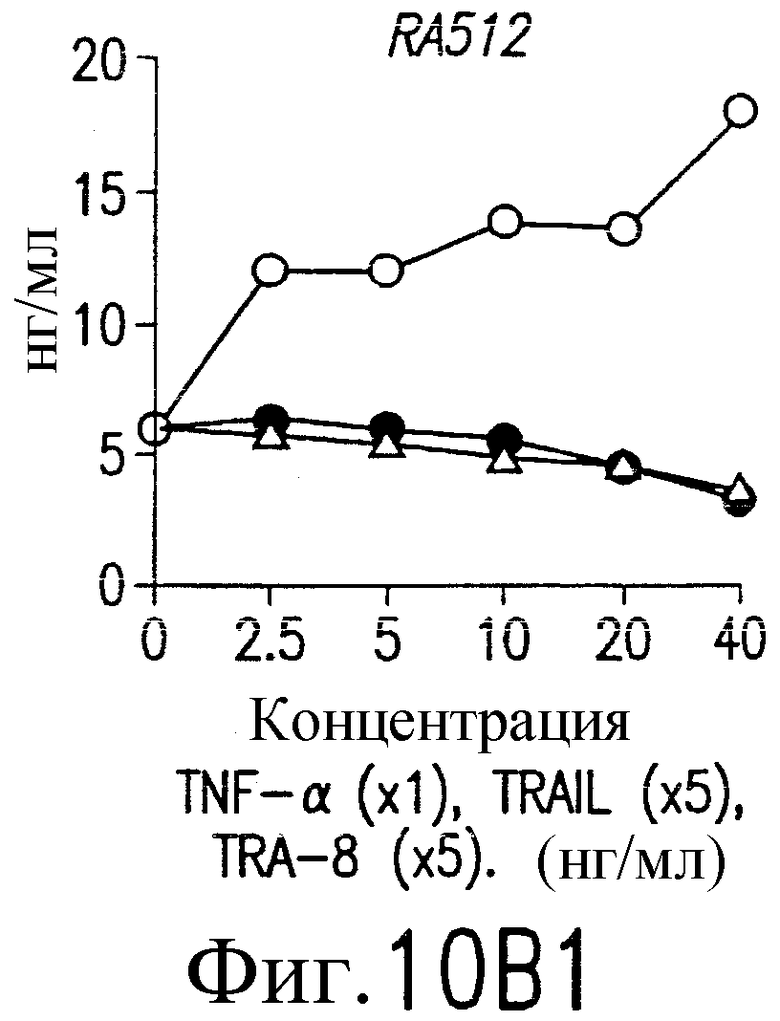
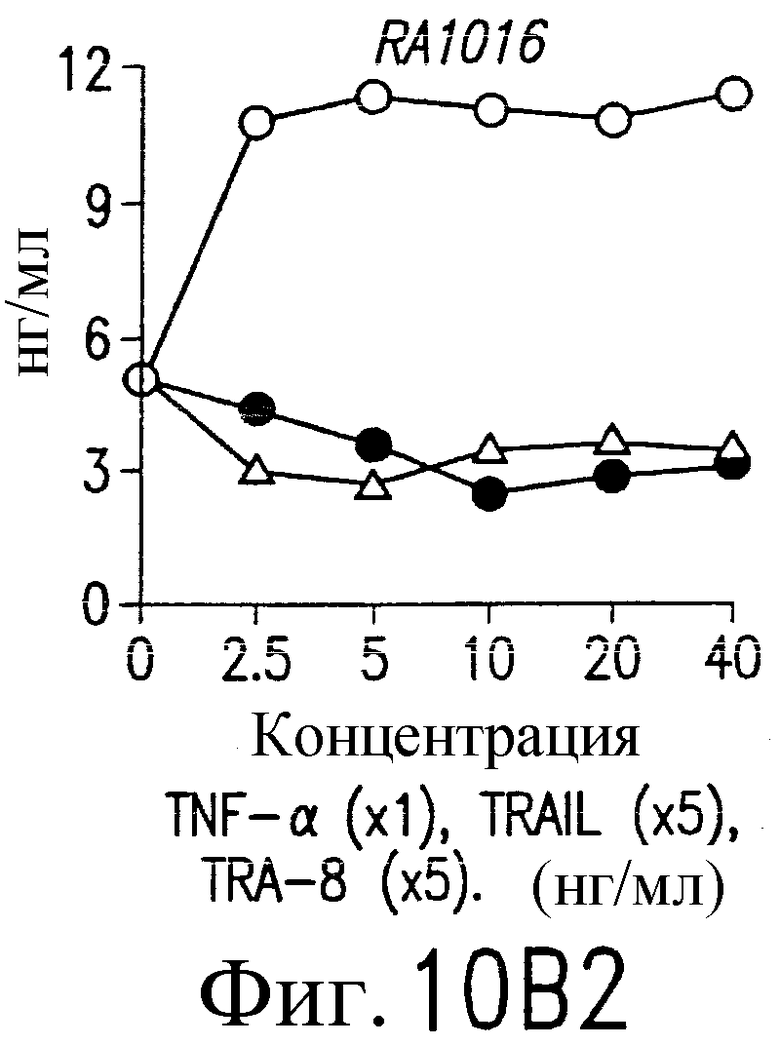
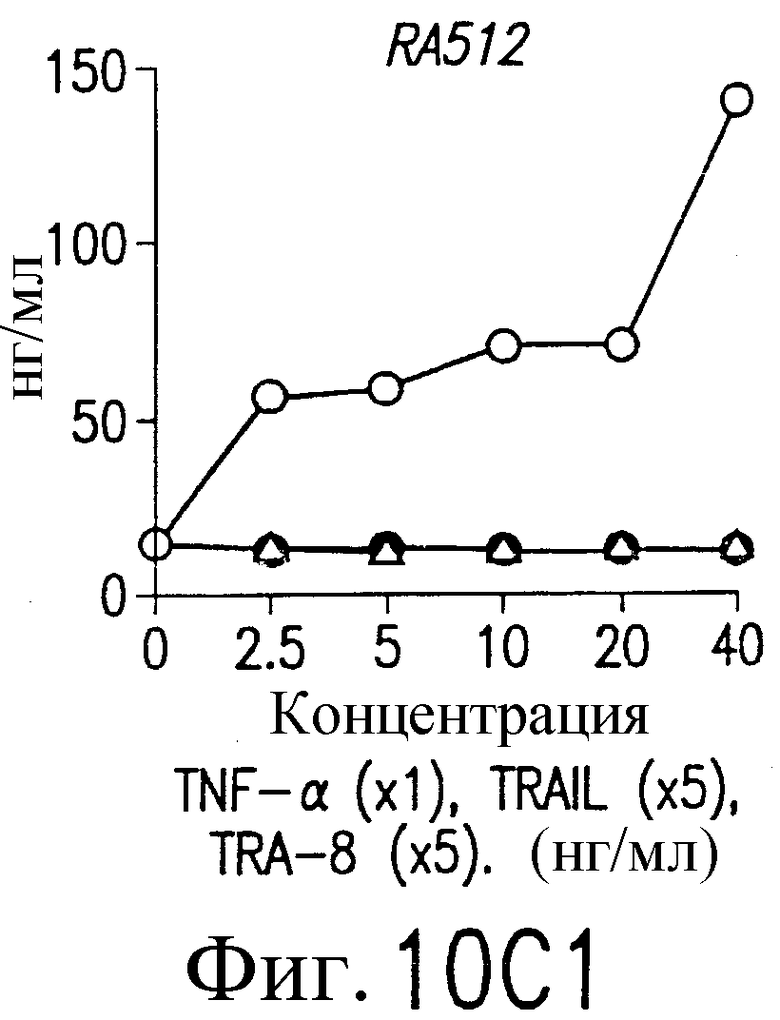
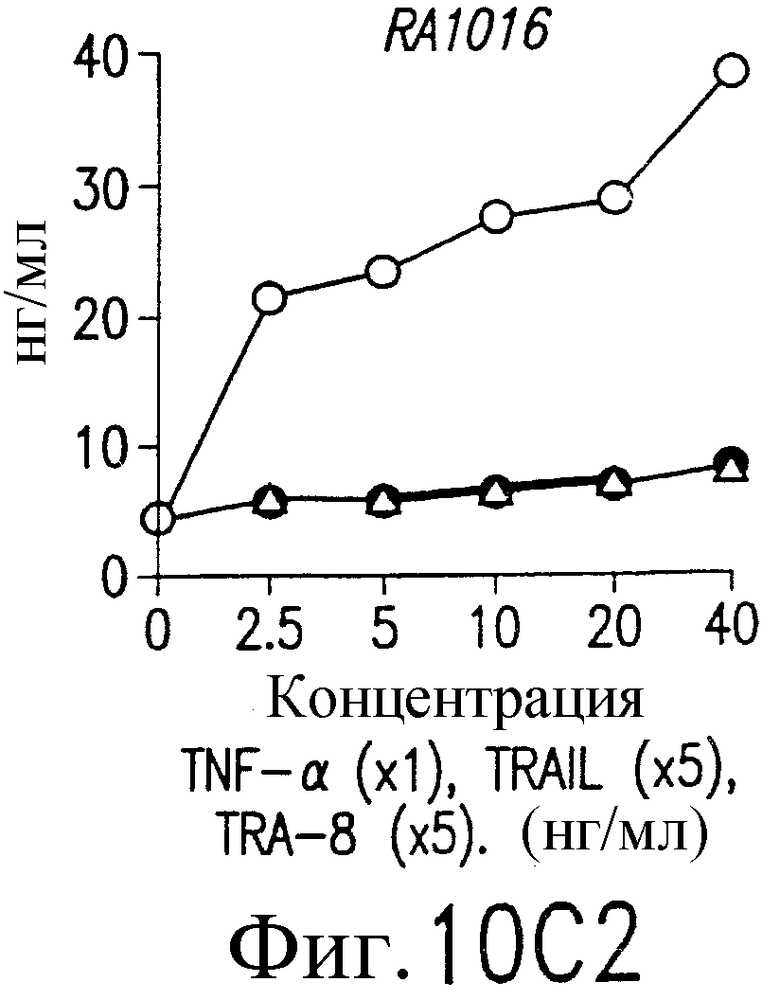
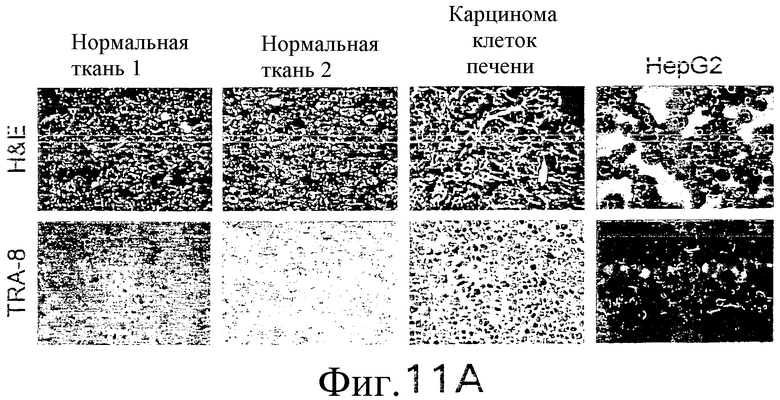
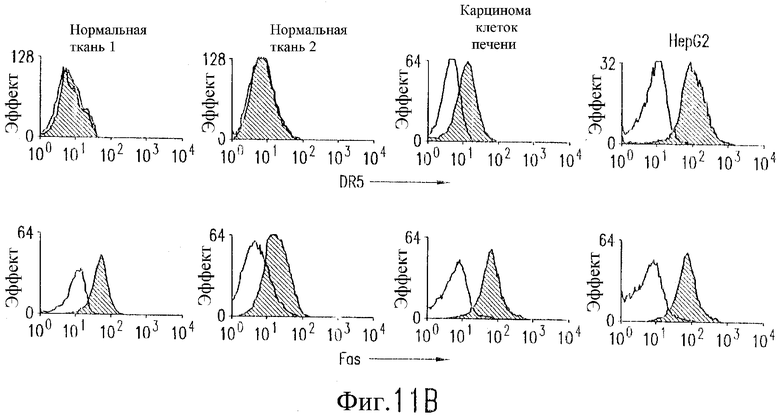
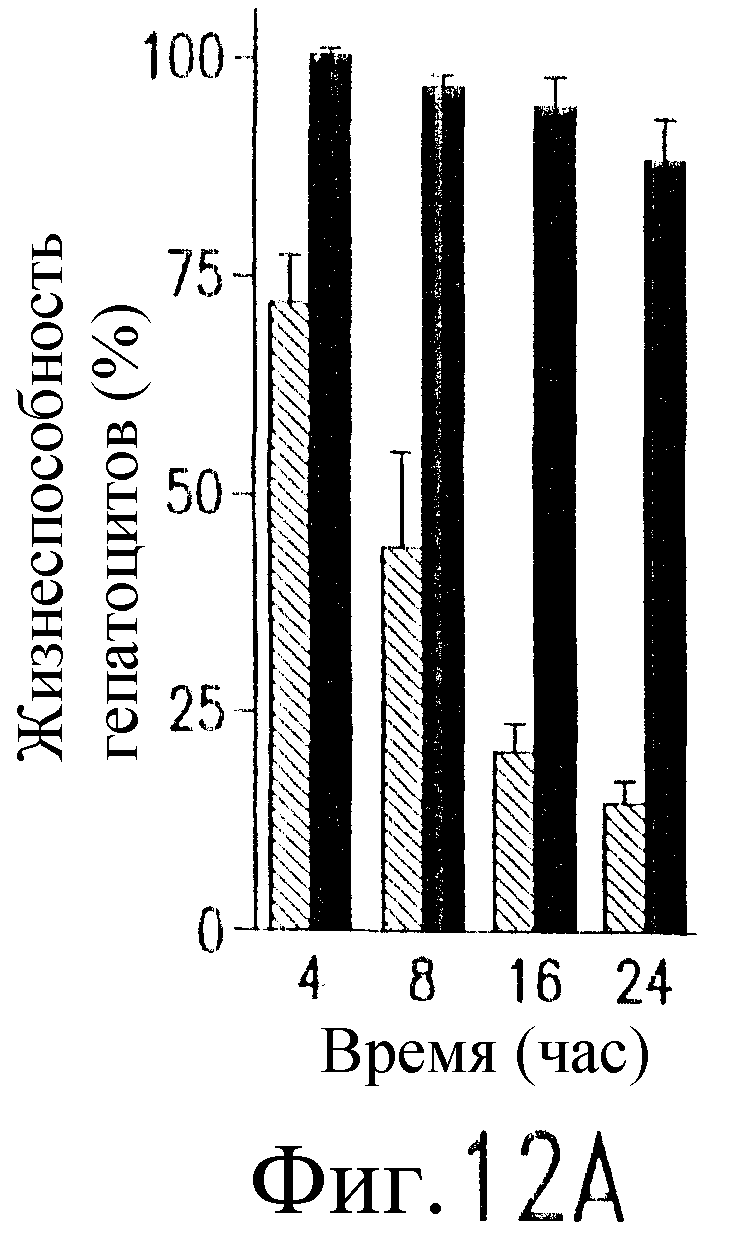
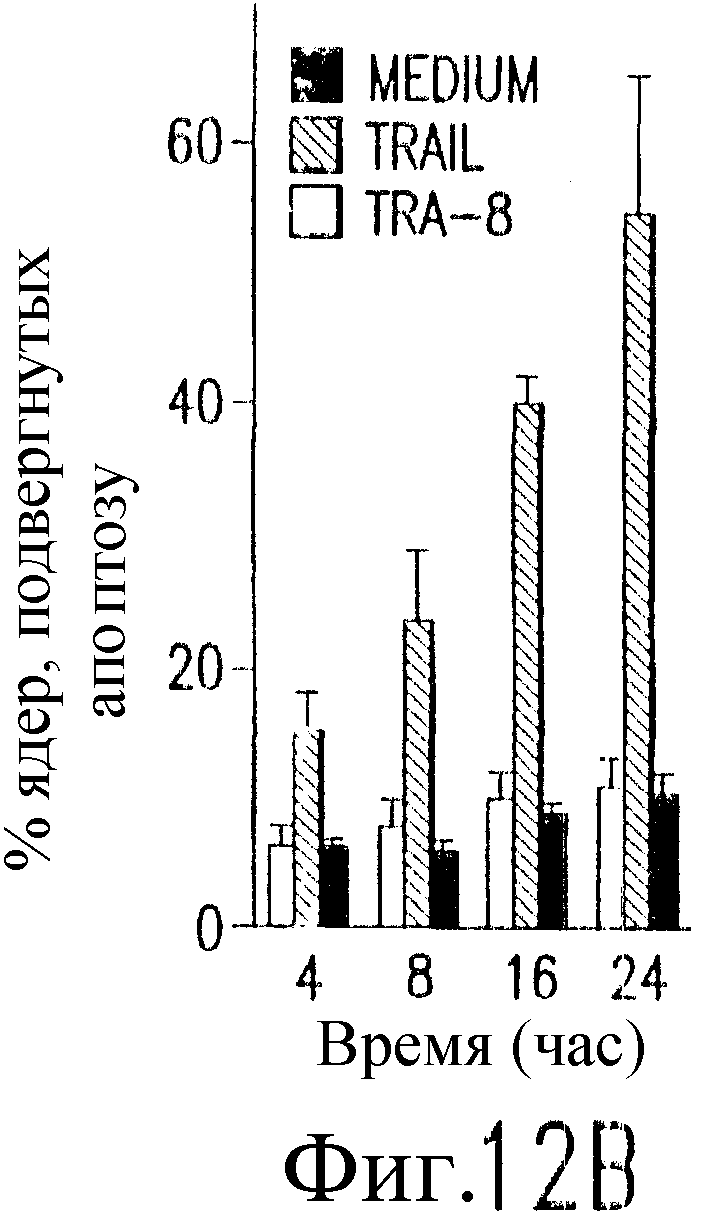
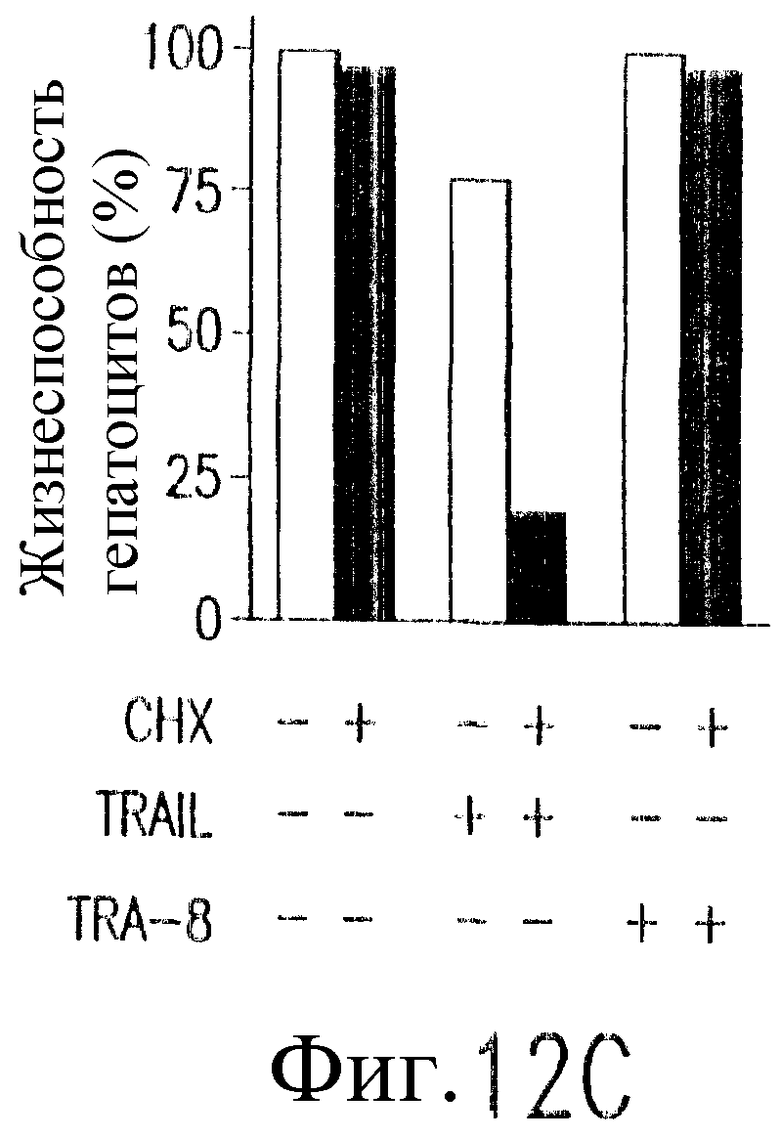
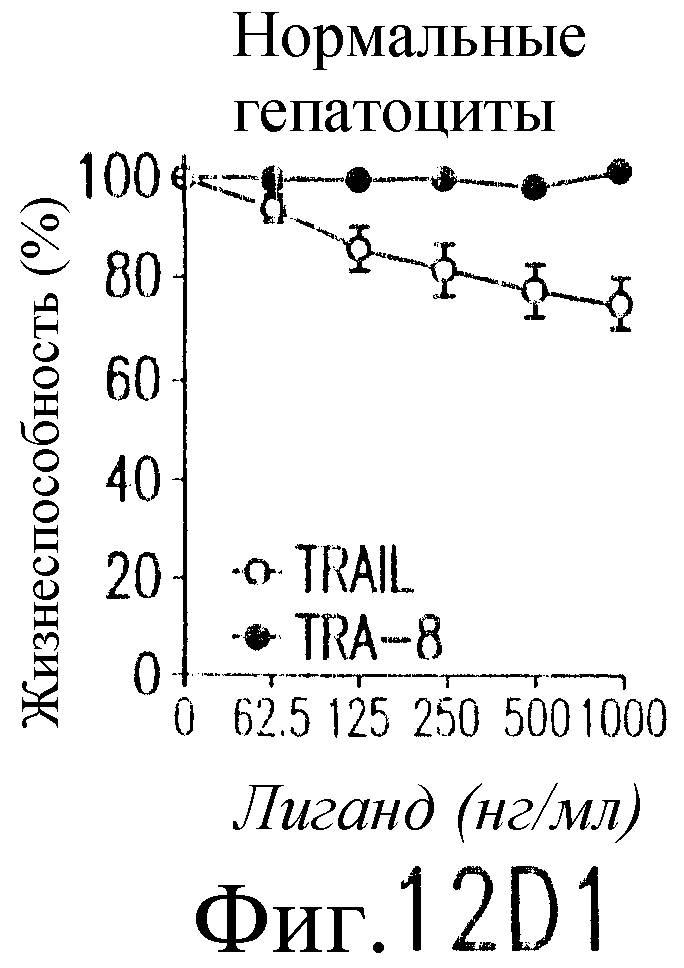
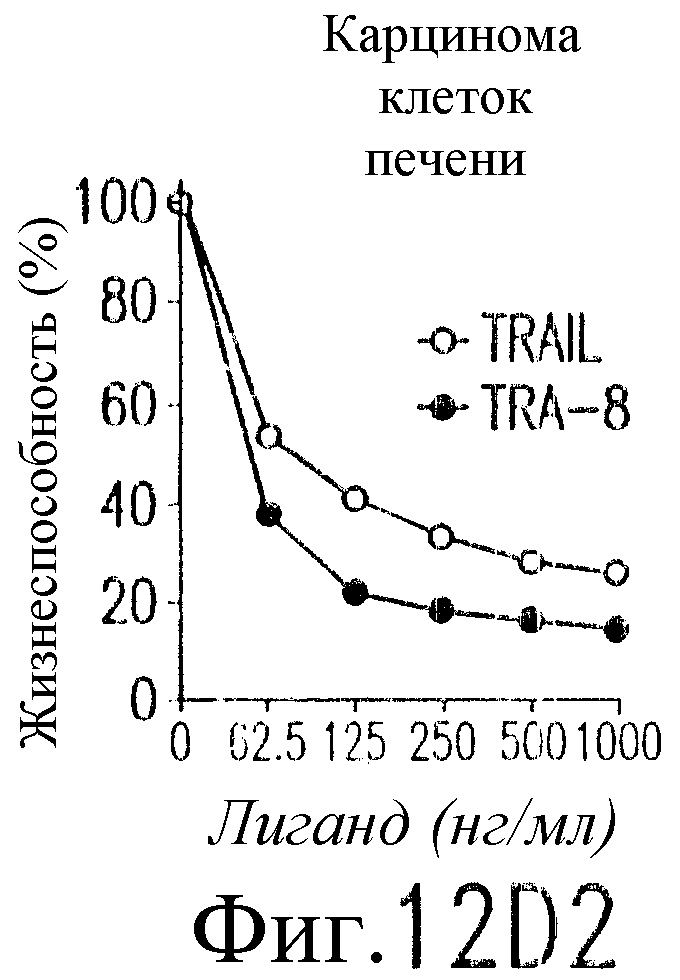
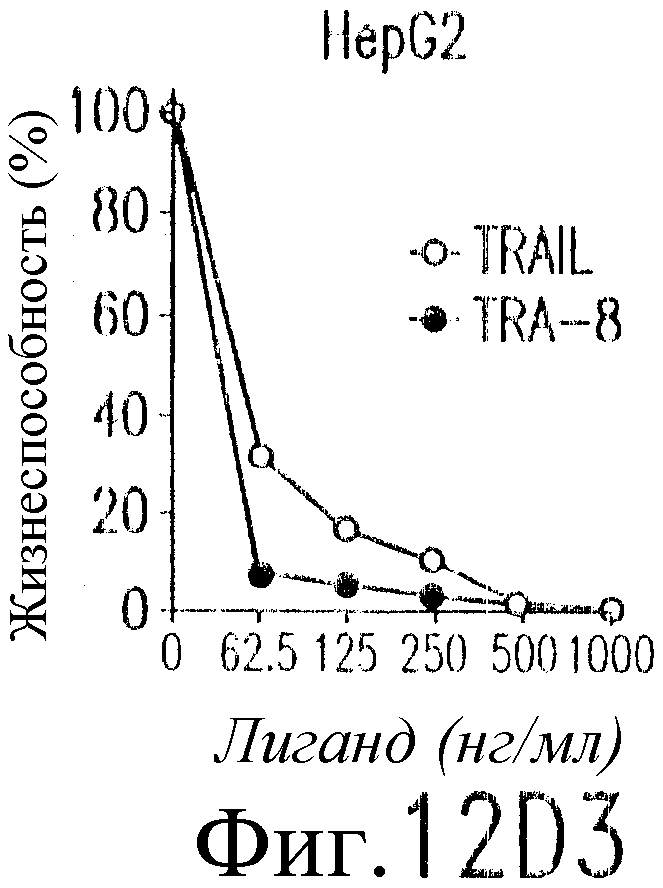
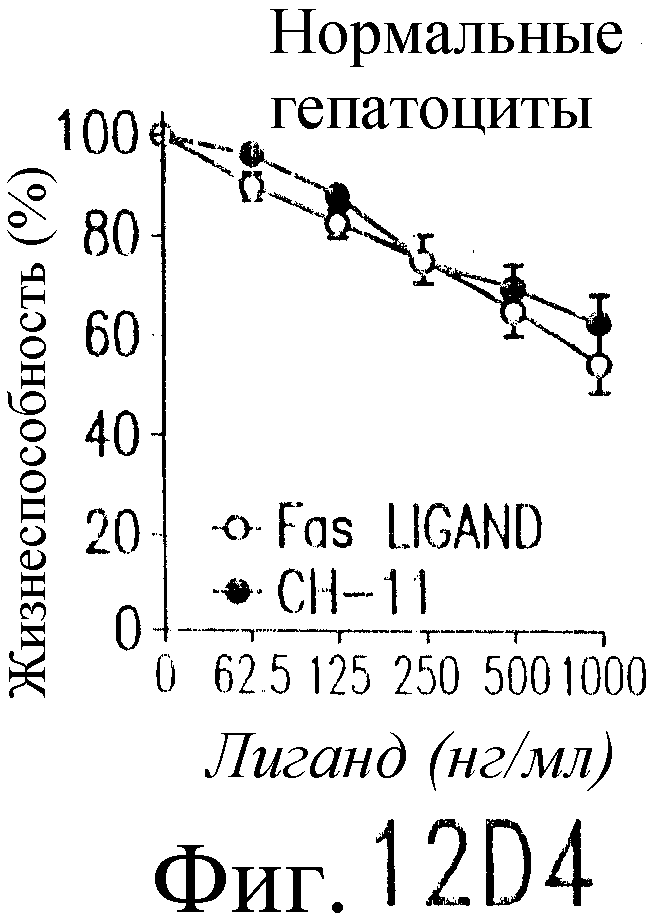
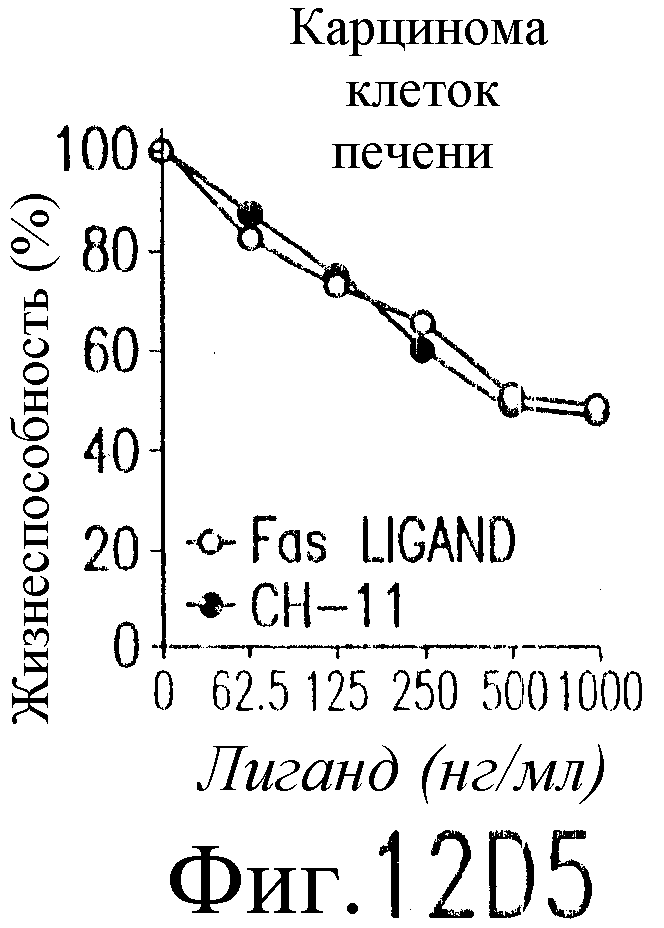
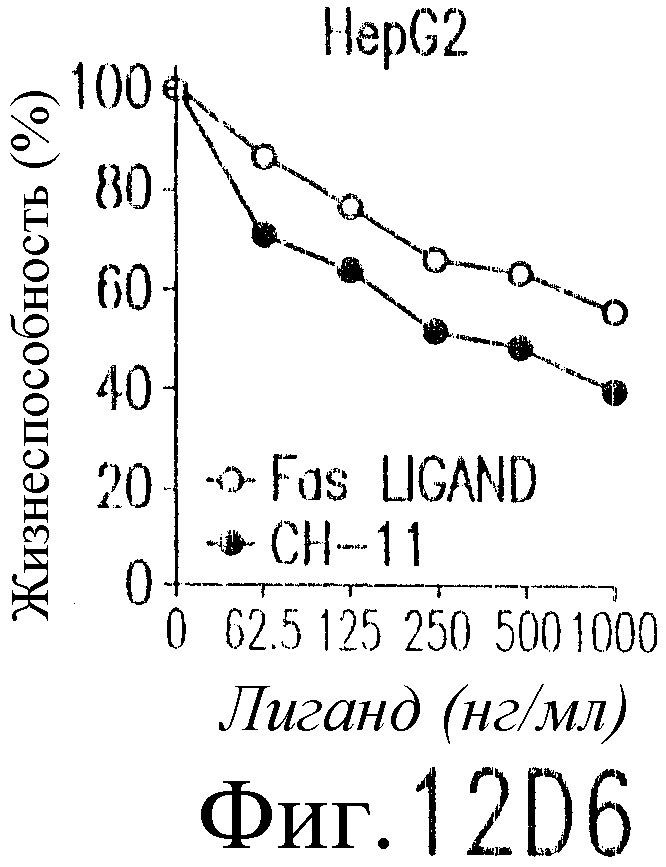
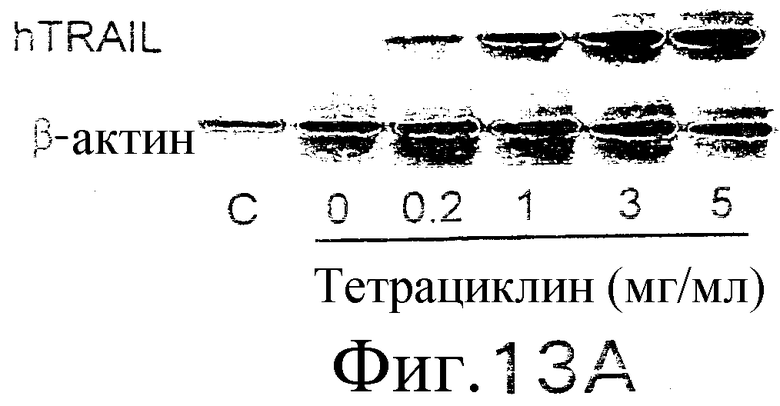
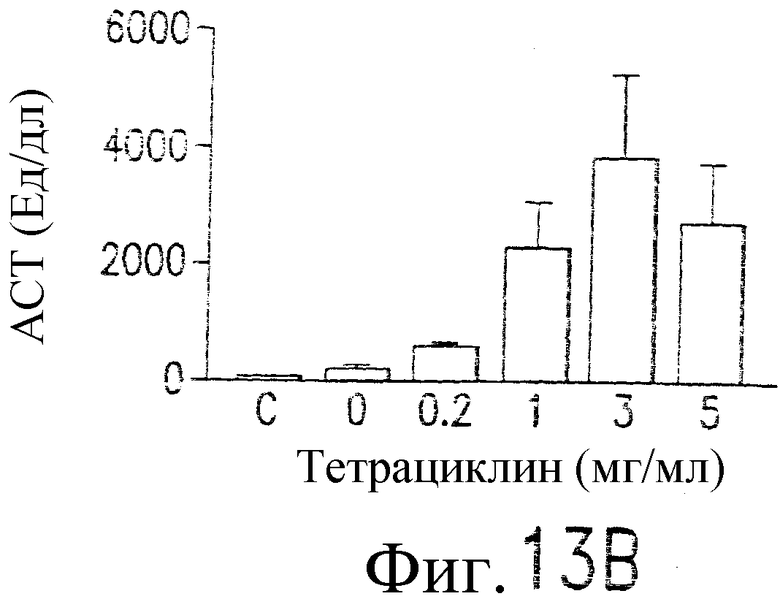
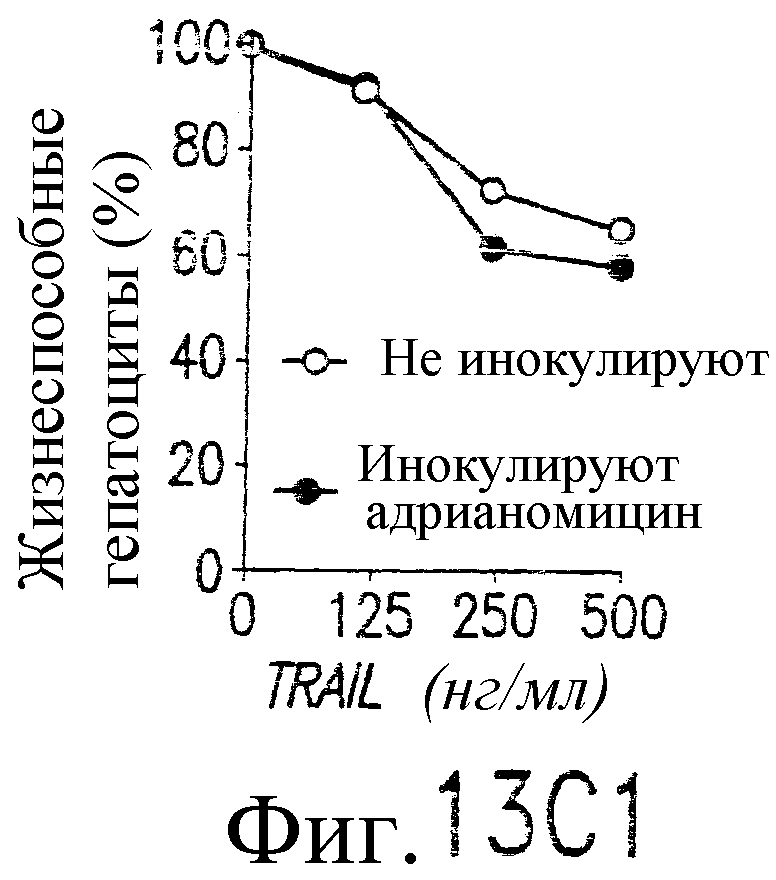
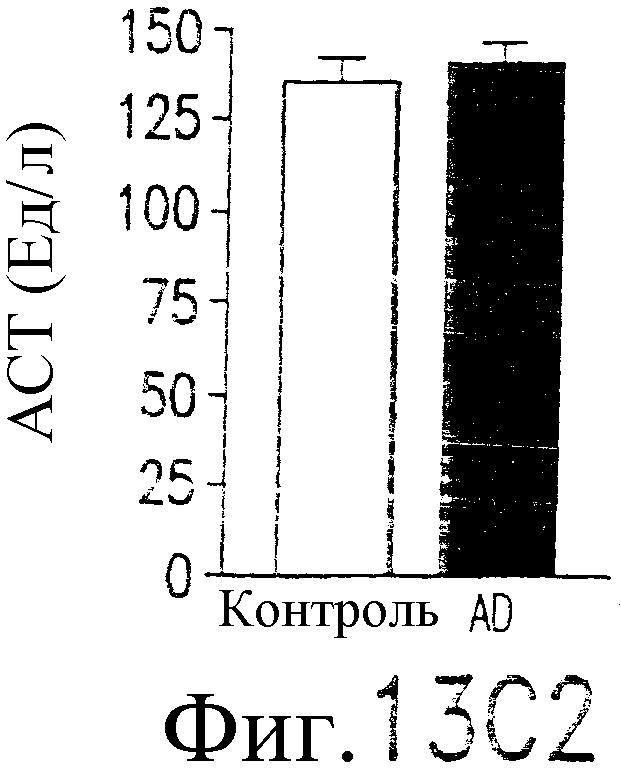


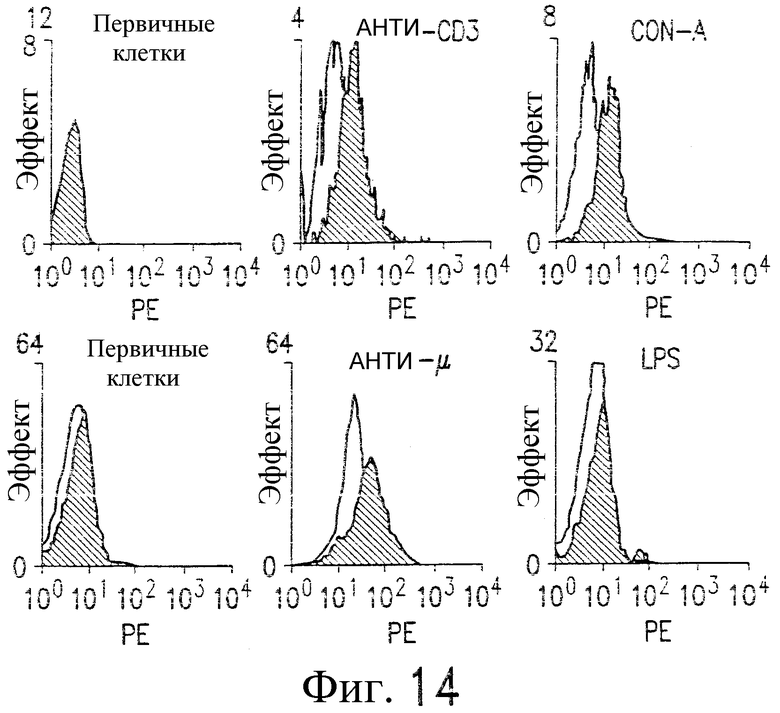
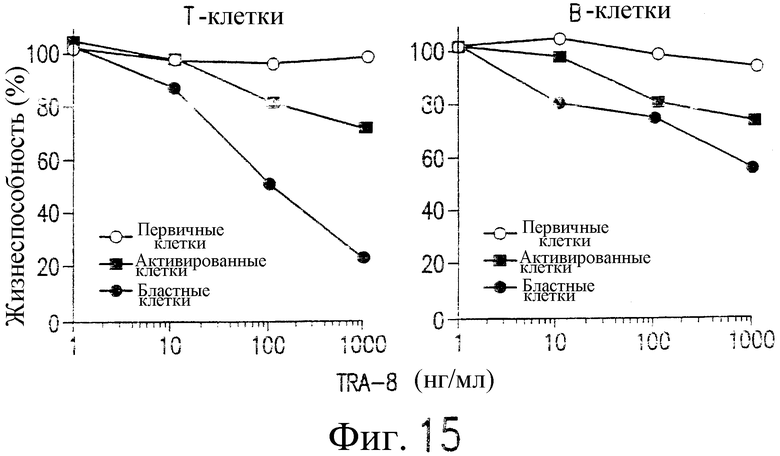
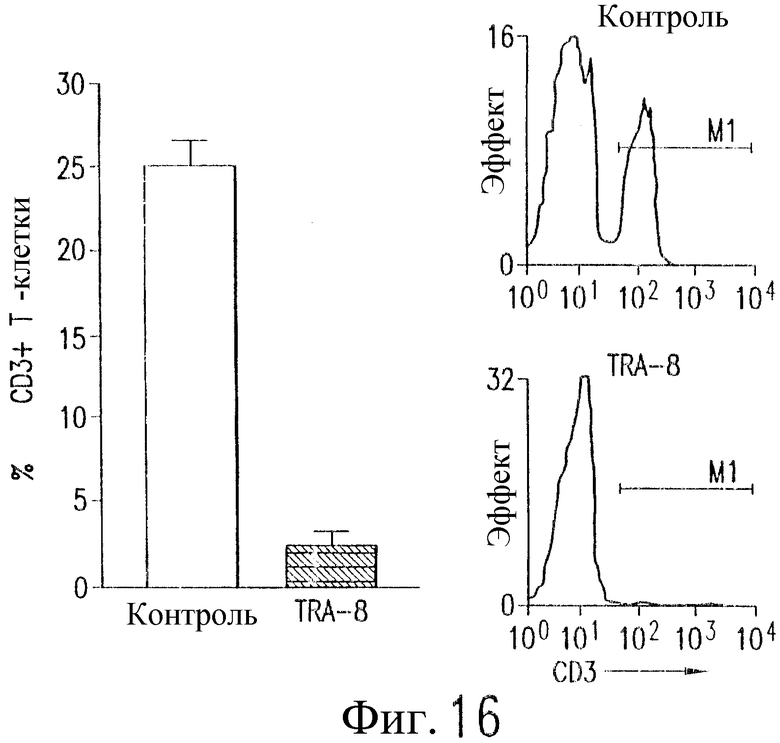
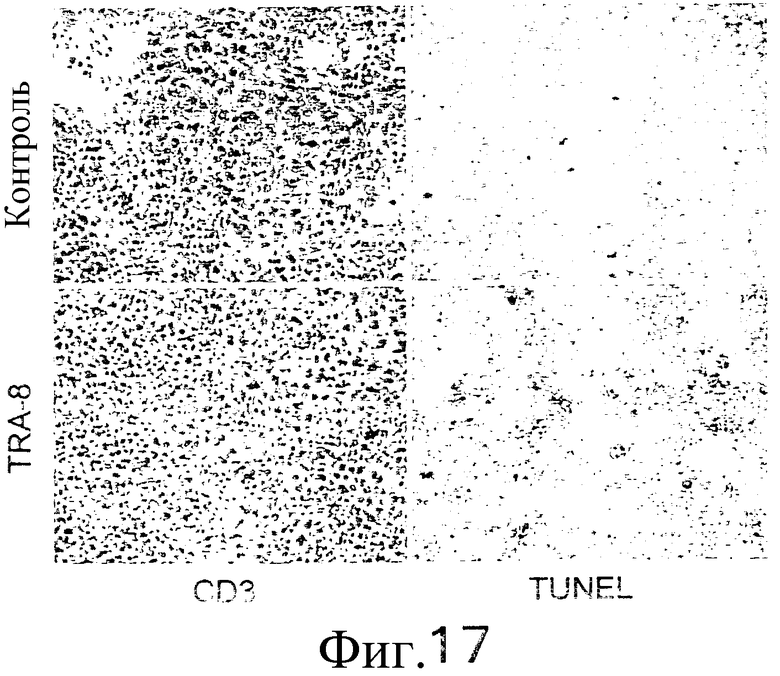
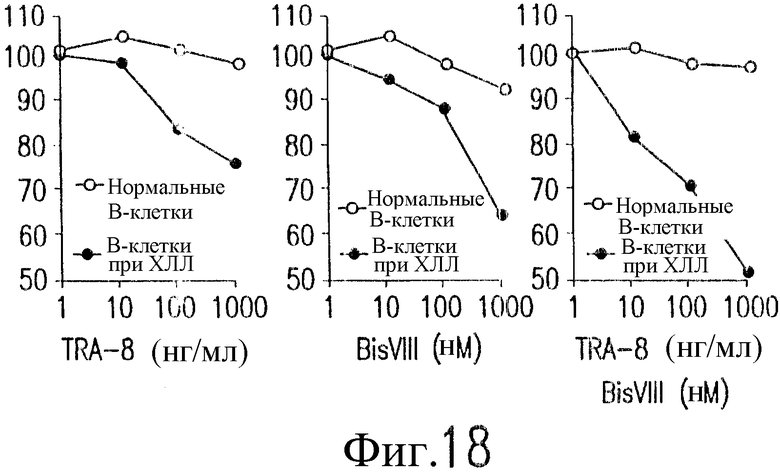
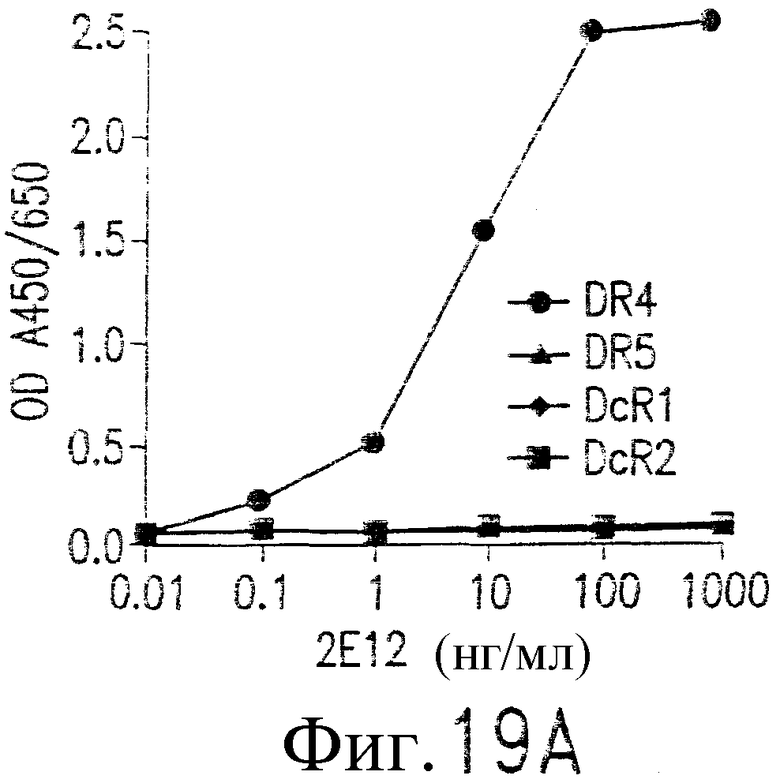
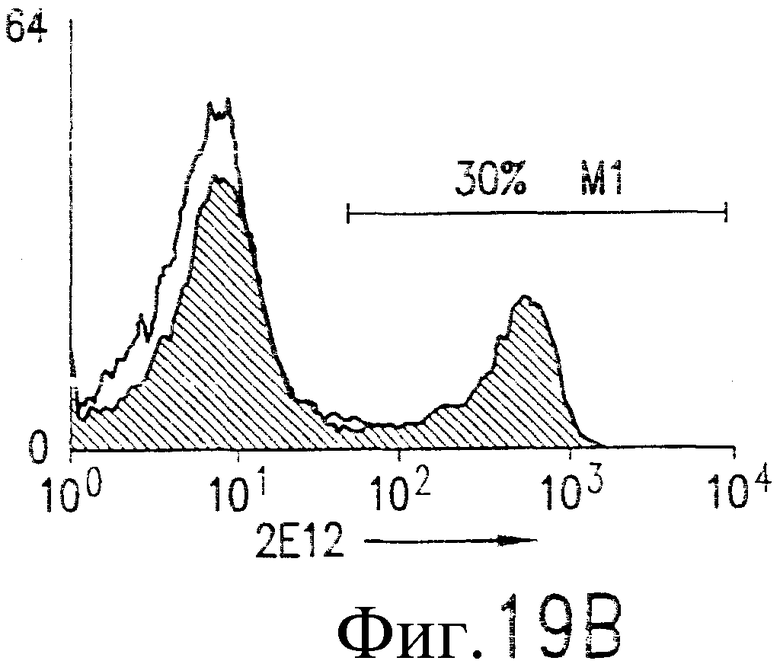
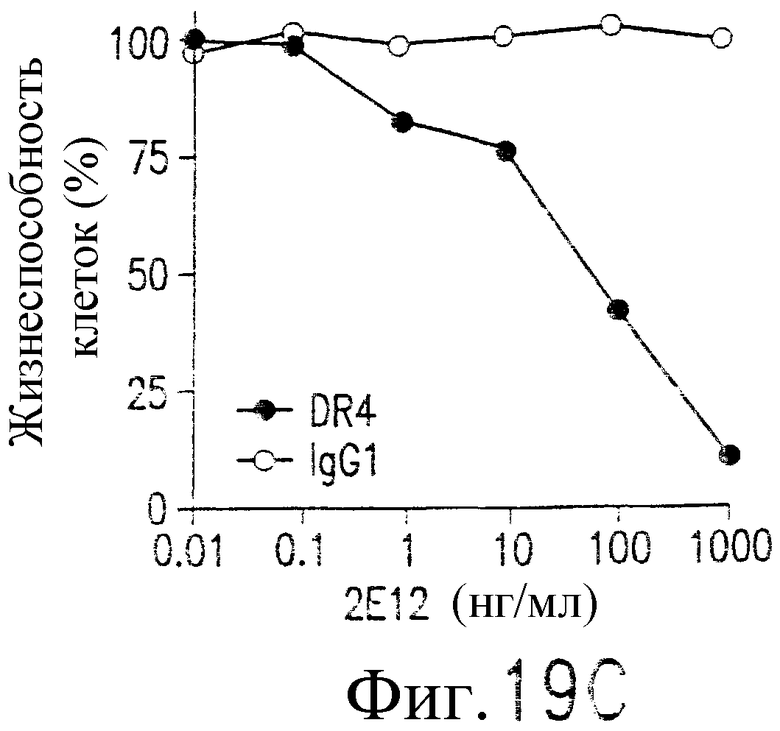
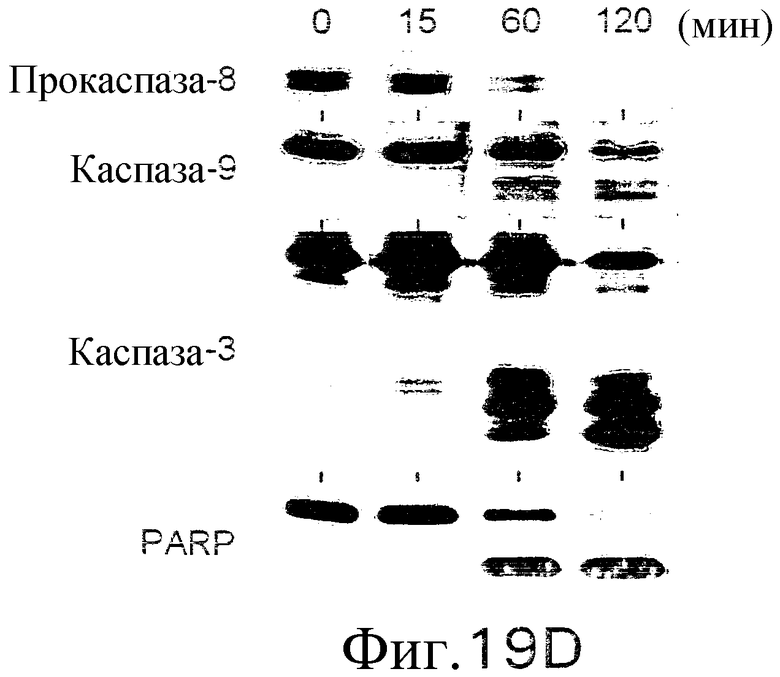
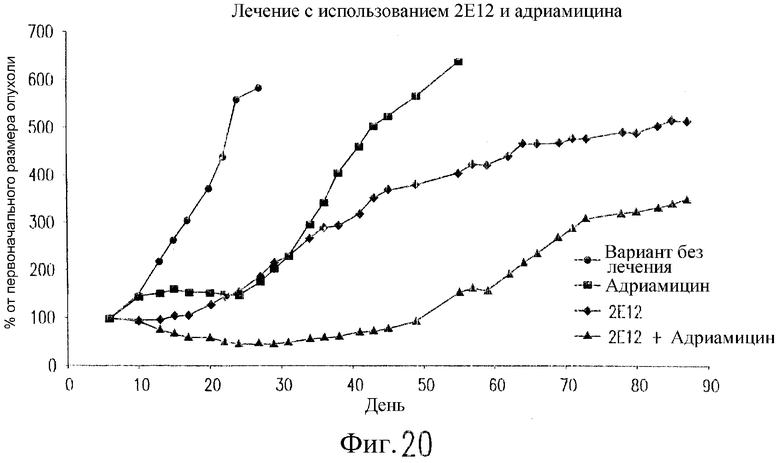
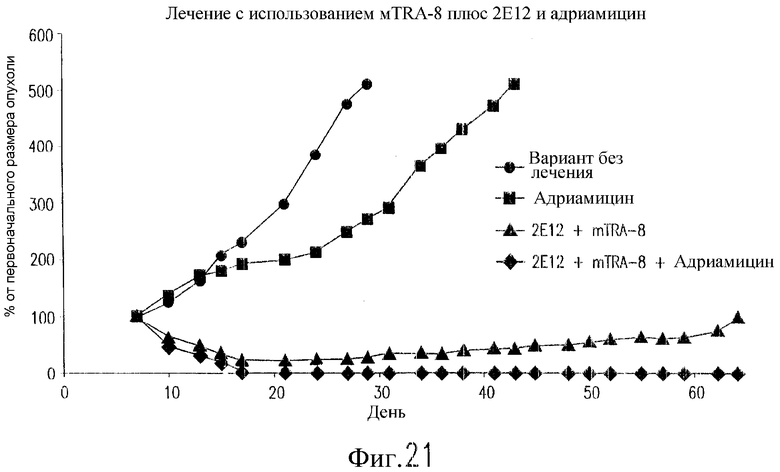
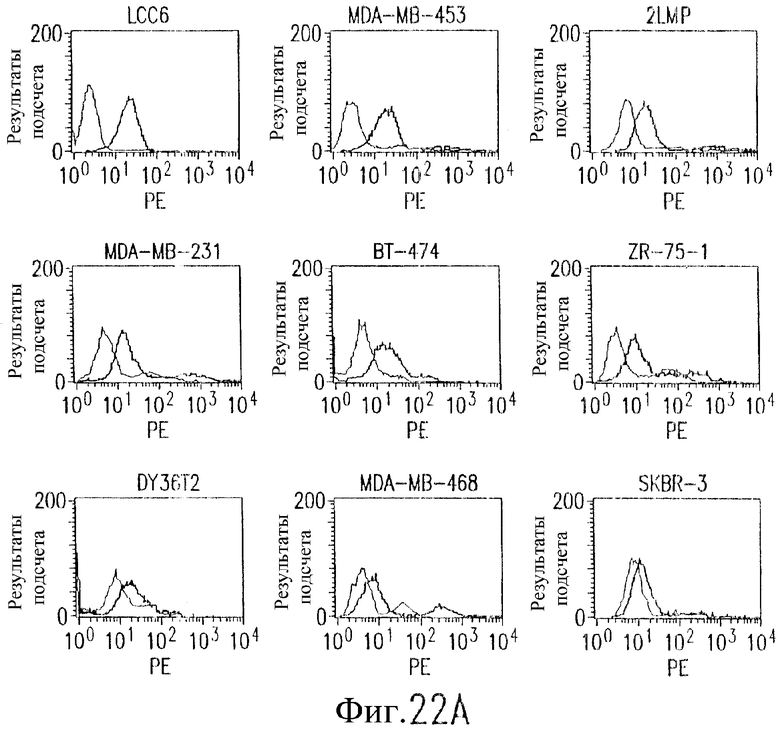
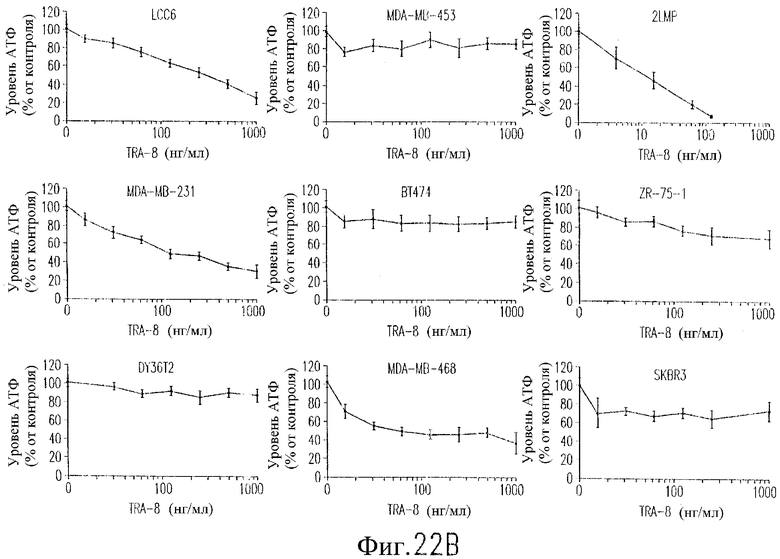
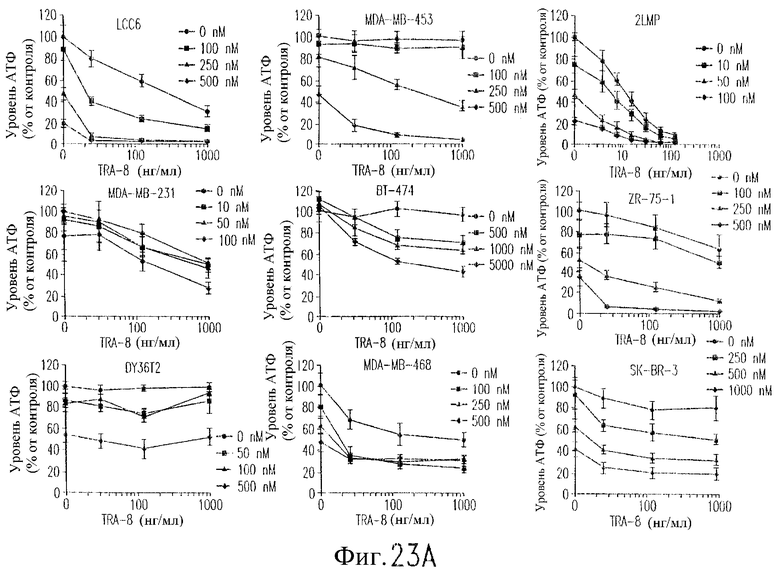
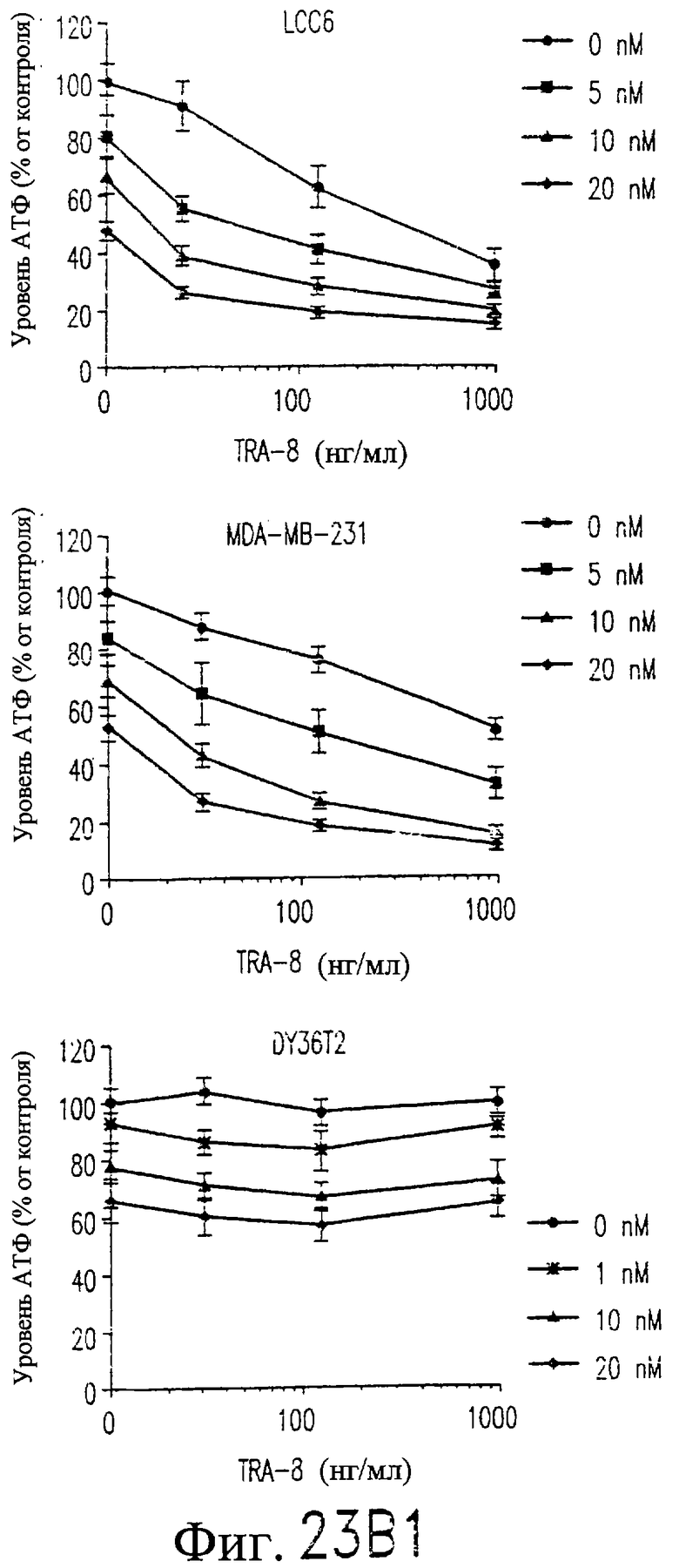
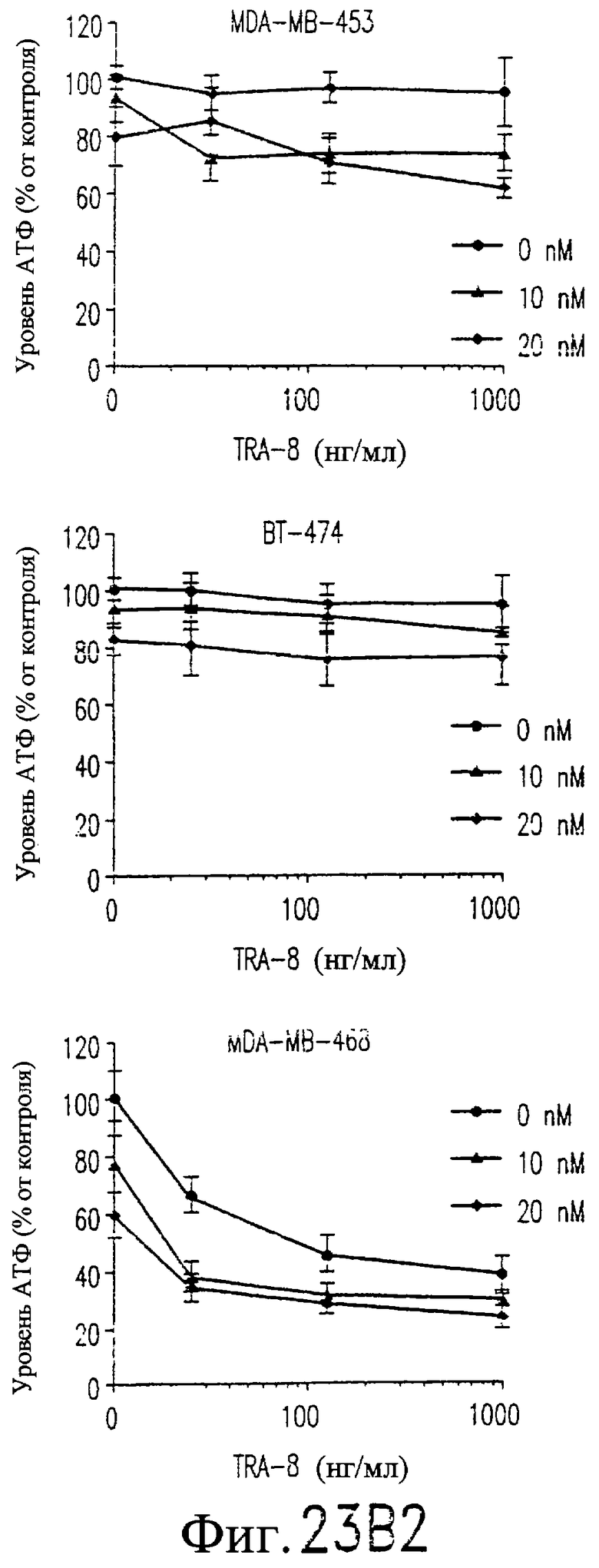
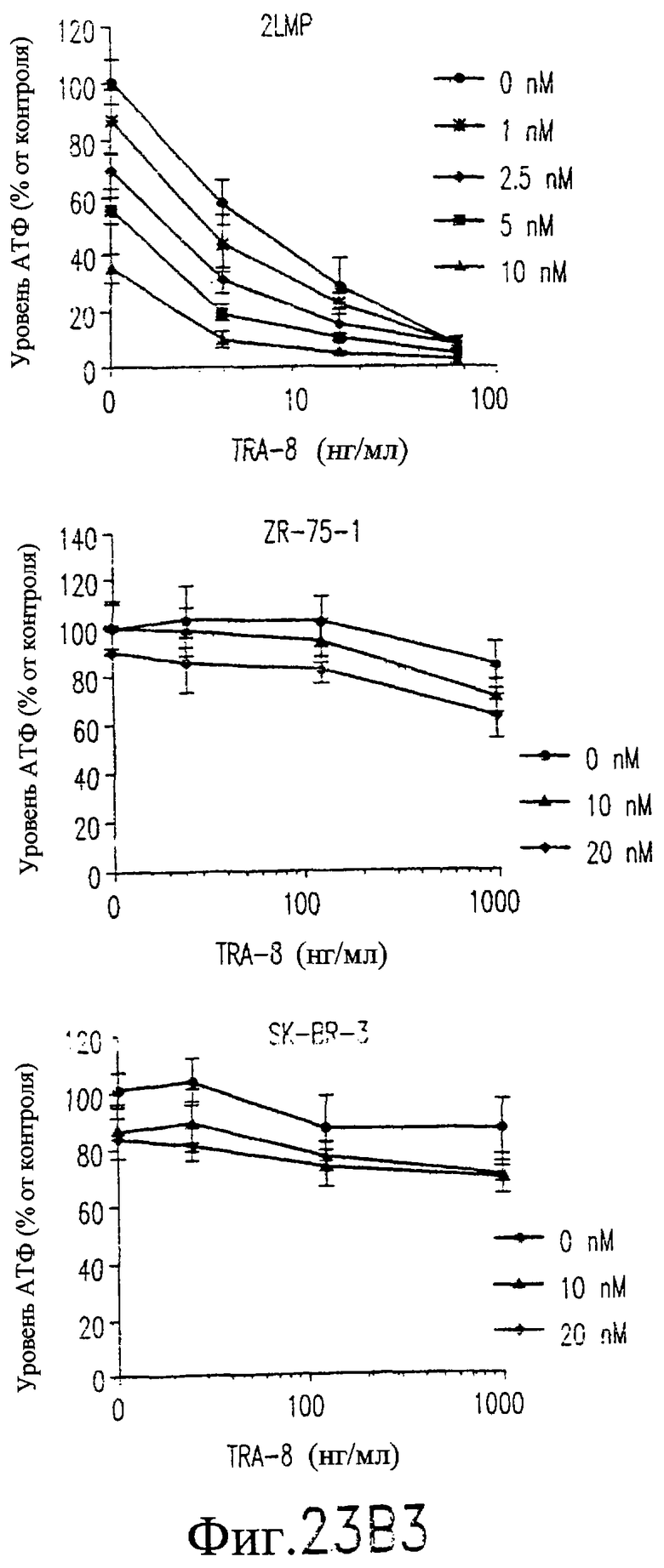
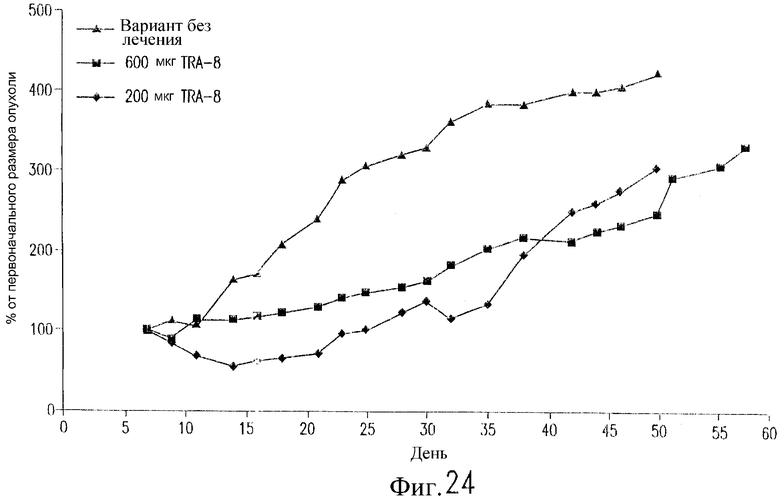
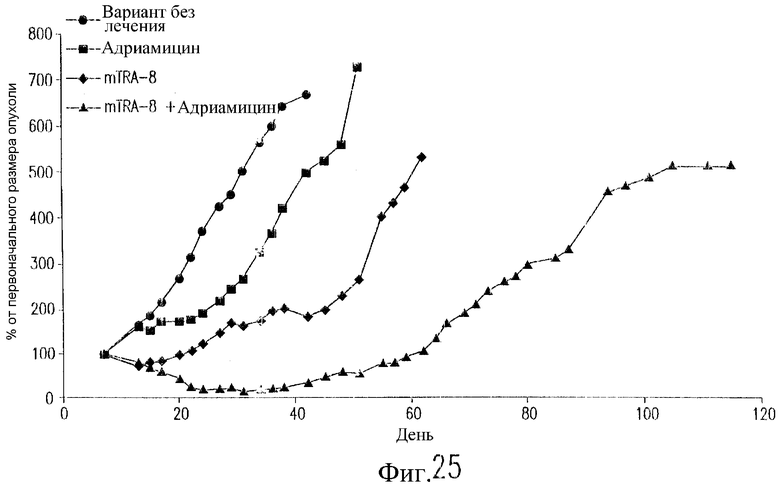
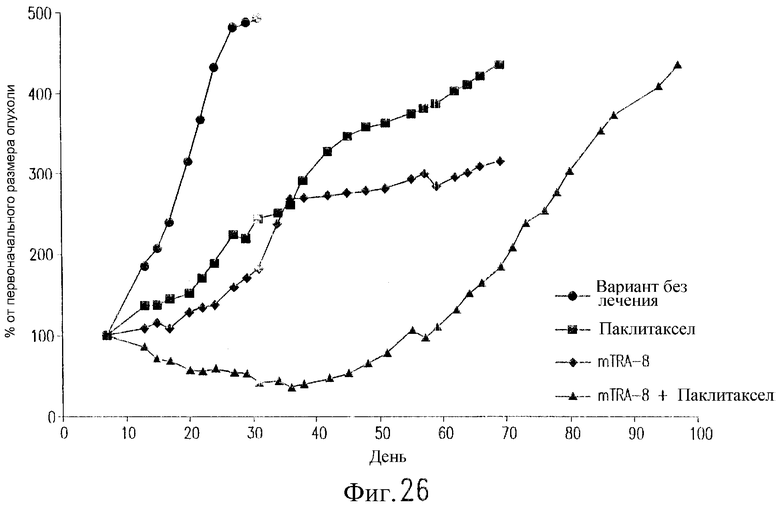
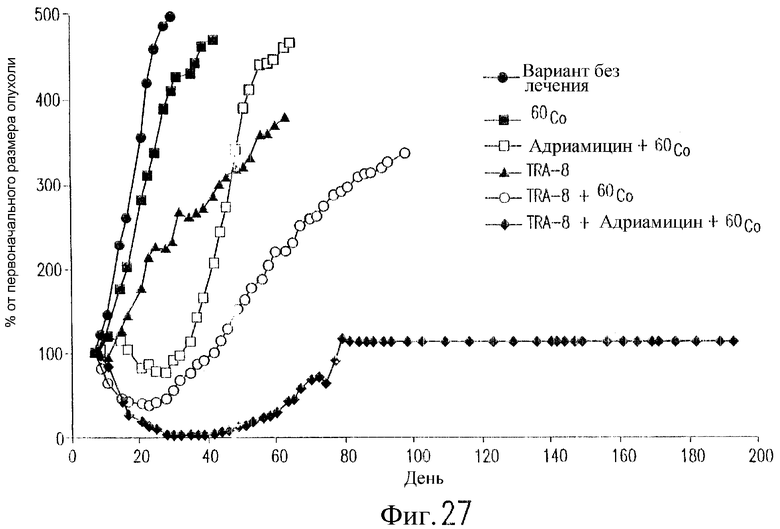
Изобретение относится к биотехнологии и иммунологии. Описаны варианты моноклональных антител специфичных к TRAIL-рецептору DR4. Один из вариантов антитела продуцируется гибридомой 2Е12, зарегистрированной в АТСС под номером РТА-3798. Каждый из вариантов антител обладает апоптоз-индуцирующей активностью как in vivo - при концентрациях менее, чем 10 мг/кг в целевых клетках, экспрессирующих DR4, так и in vitro - в присутствии сшивателя при концентрациях менее 1 мкг/мл в целевых клетках. Раскрыты варианты способов селективной индукции апоптоза в клетках, экспрессирующих DR4, и варианты способов ингибирования пролиферации клеток, экспрессирующих DR4, основанные на использовании антител. Описаны варианты композиций, способов лечения больного воспалительным или аутоиммунным заболеванием и способов лечения больного злокачественной опухолью на основе антитела к DR4 для индуцирования апоптоза в клетках, экспрессирующих DR4. Раскрыты варианты нуклеиновых кислот, очищенных полипептидов, векторов экспрессии и клеток-хозяина для получения антитела. Использование изобретения позволяет замедлить рост опухоли и сократить количество случаев ее регрессии, что может найти применение в терапии опухоли. 21 н. и 82 з.п. ф-лы, 27 ил., 6 табл.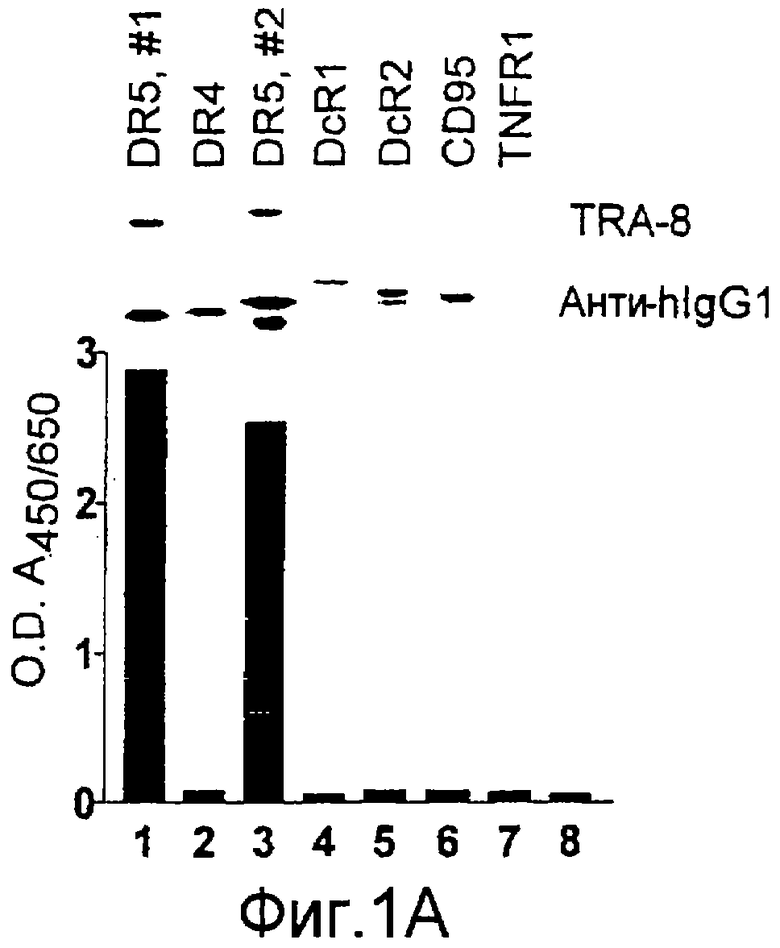
| Chuntharapai et al., Isotype-dependent inhibition of tumor growth in vivo by monoclonal antibodies to death receptor 4, Journal of immunology, 2001, 166 (8), стр.4891-4898 | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| RU 99120779 A, 20.09.2001. | |||

