Эта заявка испрашивает приоритет предварительной заявки на патент США № 60/475598, поданной 4 июня 2003 г., приведенной здесь в качестве ссылки в полном объеме.
Область, к которой относится изобретение
Настоящее изобретение относится к антителам, которые связываются с фактором роста соединительной ткани (CTGF). Эти антитела в частности направлены на области CTGF, вовлеченные в биологическую активность, связанную с различными заболеваниями.
Предпосылки изобретения
Фактор роста соединительной ткани (CTGF)
CTGF представляет собой богатый цистеином связывающий гепарин, секретируемый гликопротеид с молекулярной массой 36 кДа, первоначально выделенный из культуральной среды эндотелиальных клеток пупочной вены человека (см. например, Bradham et al. (1991) J Cell Biol 114:1285-1294; Grotendorst and Bradham, патент США № 5408040). CTGF относится к семейству белков CCN (CTGF, Cyr61, Nov) (секретируемых гликопротеидов), к которому относится индуцируемый сывороткой ранний генный продукт Cyr61, предполагаемый онкоген Nov, связанный с ECM белок FISP-12, src-индуцибельный ген CEF-10, Wnt-индуцибельный секретируемый белок WISP-3 и антипролиферативный белок HICP/rCOP (Brigstock (1999) Endocr. Rev. 20: 189-206; O'Brian et al. (1990) Mol. Cell Biol., 10: 3569-3577; Joliot et al. (1992) Mol Cell Biol., 12: 10-21; Ryseck et al. (1990) Cell Growth and Diff., 2: 225-233; Simmons et al. (1989) Proc. Natl. Acad. Sci. USA, 86: 1178-1182; Pennica et al. (1998) Proc. Natl. Acad. Sci. USA, 95: 14717-14722; и Zhang et al. (1998) Mol. Cell Biol., 18: 6131-61416). Белки CCN отличаются тем, что у них консервативны 38 цистеиновых остатков, которые составляют более 10% общего содержания аминокислот и обеспечивают возникновение модулярной структуры с N- и C-концевыми доменами. Модулярная структура CTGF включает консервативные мотивы для белка, связывающего подобный инсулину фактор роста (IGF-BP), и фактора фон Виллебранда (VWC) в N-концевом домене и тромбоспондин (TSP1) и мотив цистеинового узла в С-концевом домене.
Экспрессия CTGF индуцируется членами суперсемейства трансформирующего фактора роста бета (TGFβ), которое включает TGFβ-1, -2 и -3, костный морфогенетический белок (ВМР)-2 и активин, а также различными другими регуляторными модуляторами, включая дексаметазон, тромбин, сосудистый эндотелиальный фактор роста (VEGF) и ангиотензин II; и стимулами окружающей среды, включая гипергликемию и гипертонию (см. например, Franklin (1997) Int. J. Biochem. Cell Biol., 29: 79-89; Wunderlich (2000) Graefes Arch. Clin. Exp. Ophthalmol, 238: 910-915; Denton and Abraham (2001) Curr. Opin. Rheumatol, 13: 505-511 и Riewald (2001) Blood, 97: 3109-3116; Riser et al. (2000) J. Am. Soc. Nephrol, 11: 25-38 и международную публикацию WO 00/13706). Стимуляция экспрессии CTGF под действием TGFβ является быстрой и длительной и не требует продолжительного применения (Igarashi et al. (1993) Mol. Biol. Cell, 4:637-645). Усиленная экспрессия CTGF TGFβ включает транскрипционную активацию посредством регуляторных элементов ДНК, присутствующих в промотере CTGF (Grotendorst et al. (1996) Cell Growth Differ, 7: 469-480; Grotendorst and Bradham, патент США №6069006; Holmes et al. (2001) J. Biol. Chem., 276: 10594-10601).
Было показано, что CTGF увеличивает стационарную транскрипцию коллагена α1 (I), интегрина α5 и мРНК фибронектина, а также промотирует клеточные процессы, включая пролиферацию и хемотаксис различных типов клеток в культуре (см. например, Frazier et al. (1996) J. Invest. Dermatol., 107: 406-411; Shi-wen et al. (2000) Exp. Cell Res., 259: 213-224; Klagsburn (1997) Exp. Cell Res., 105: 99-108; Gupta et al. (2000) J. Kidney Int., 58: 1389-1399; Wahab et al. (2001) Biochem. J., 359 (Pt 1): 77-87; Uzel et al (2001) J. Periodontol., 72: 921-931 и Riser and Cortes (2001) Ren. Fail., 23: 459-470). Подкожная инъекция CTGF новорожденным мышам приводит к местному отложению грануляционной ткани. Аналогичным образом подкожная инъекция TGFβ генерирует формирование грануляционной ткани и вызывает высокие уровни мРНК CTGF в местных фибробластах. Более того, комбинированное или последовательное лечение TGFβ и CTGF приводит к развитию более стойкой гранулемы (Mori et al. (1999) J. Cell Physiol., 181: 153-159). Таким образом, представляется, что CTGF опосредует подгруппу эффектов, вызываемых TGFβ, в частности, продукцию и отложение внеклеточного матрикса (ЕСМ). Кроме того, способность реагировать на CTGF или степень реакции на CTGF может зависеть от стимула примирования, обеспечиваемого обработкой TGFβ, что обеспечивает возможность клеточной "компетентности" (Международная публикация WO 96/08140).
Хотя было охарактеризовано множество взаимодействующих факторов, которые модулируют тканевую организацию, в настоящее время появляется согласие в отношении роли CTGF в регуляции развития скелета, заживлении ран и перестройке внеклеточного матрикса (ЕСМ), фиброзе, развитии опухолей и ангиогенезе. Например, повышенная экспрессия CTGF наблюдалась в печени при циррозе, при пневмосклерозе, воспалительных кишечных заболеваниях, в склеротической коже и келоидах, при десмоплазии и атеросклеротических бляшках (Abraham et al. (2000) J. Biol. Chem., 275: 15220-15225; Dammeier et al. (1998) Ind. J. Biochem. Cell Biol., 30: 909-922; diMola et al. (1999) Ann. Surg. 230(1): 63-71; Igarashi et al. (1996) J. Invest. Dermatol., 106: 729-733; Ito et al. (1998) Kedney Int., 53: 853-861; Williams et al. (2000) J. Hepatol., 32: 754-761; Clarkson et al (1999) Curr. Opin. Nephrol. Hypertens, 8: 543-548; Hinton et al. (2002) Eye, 16: 422-428; Gupta et al. (2000) Kidney Int., 58: 1389-1399; Riser et al. (2000) J. Am. Soc. Nephrol., 11: 25-38).
Стимулирующая регуляция CTGF также происходит при гломерулонефрите, нефропатии, вызванной IgA, очаговом и сегментарном гломерулосклерозе и диабетической нефропатии (см. например, Riser et al. (2000) J. Am. Soc. Nephrol., 11: 25-38). Увеличение количества клеток, экспрессирующих CTGF, также наблюдается в участках хронического канальцево-интерстициального повреждения, и уровни CTGF коррелируют со степенью повреждения (Ito et al. (1998) Kidney Int., 53: 853-861). Кроме того, экспрессия CTGF увеличивается в клубочках и интерстиции канальцев при различных почечных заболеваниях в связи с рубцеванием и склерозом почечной паренхимы. Повышенные уровни CTGF также связаны с фиброзом печени, инфарктом миокарда и пневмосклерозом. Например, у пациентов с идиопатическим пневмосклерозом (IPF) наблюдается сильная стимулирующая регуляция CTGF в биопсиях и клетках жидкости бронхоальвеолярного лаважа (Ujike et al. (2000) Biochem. Biophys. Res. Commun., 277: 448-454; Abou-Shady et al. (2000) Liver, 20: 296-304; Williams et al. (2000) J. Hepatol., 32: 754-761; Ohnishi et al. (1998) J. Mol. Cell Cardiol., 30: 2411-22; Lasky et al. (1998) Am. J. Physiol., 275: L365-371; Pan et al. (2001) Eur. Respir. J., 17: 1220-1227 и Allen et al. (1999) Am. J. Respir. Cell Mol. Biol., 21: 693-700). Таким образом, CTGF является обоснованной мишенью при лечении расстройств, таких как описанные выше.
Была установлена связь CTGF с различными аспектами этих расстройств, и были описаны способы лечения расстройств посредством модуляции CTGF (см. например, Grotendorst and Bradham, патент США № 5783187; Международная публикация № WO 00/13706; и Международная публикация № WO 03/049773). Модуляцию факторов роста, цитокинов и рецепторов клеточной поверхности можно осуществить, используя моноклональные антитела, и несколько терапевтических антител были разрешены к применению или находятся в процессе разработки (см. например, Infliximab (Remicade; Maini et al. (1998) Arthritis Rheum., 41: 1552-1563; Targan et al. (1997) N. Engl. J. Med., 337: 1029-1035); Basiliximab (Simulect) and Daclizumab (Zenapax) (Bumgardner et al. (2001) Transplantation, 72: 839-845; Kovarik et al. (1999) Transplantation, 68: 1288-1294); и Transtuzumab (Herceptin; Baselga (2001) Ann. Oncol., 12 Suppl. 1: S49-55)).
Были генерированы антитела против CTGF, и они оказались эффективными in vivo, например, ингибируя ангиогенез (см. например, Grotendorst and Bradham, патент США № 5408040; международная публикация WO 99/07407 и Shimo et al. (2001) Oncology 61:315-322). Кроме того, оказалось, что модулярная природа CTGF различает домены, участвующие в специфических видах биологической активности. Например, было показано, что N-концевая половина CTGF стимулирует клеточную дифференциацию и продукцию ЕСМ, тогда как С-концевая половина стимулирует клеточную пролиферацию (см. например, Международные публикации WO 00/35936 и WO 00/35939; Brigstock and Harding, патент США № 587670). Это демонстрирует, что антитела, направленные на различные области молекулы CTGF, проявляют различные эффекты в отношении модулирования видов биологической активности CTGF (см. например, Международные публикации WO 00/35936 и WO 00/35939). К настоящему времени не было установлено четкого различия между антителами против CTGF, которые продуцируют желательный эффект, и антителами, которые или продуцируют множественные эффекты, или являются ненейтрализующими (см. например, Международную публикацию WO 99/33878).
Существует явная необходимость в данной области в средствах, которые эффективно нейтрализуют активность CTGF при патологии. Антитела, в частности моноклональные антитела, обеспечивают специфичность и фармакокинетический профиль, подходящий для терапевтического средства, и нейтрализующие антитела, нацеленные на конкретные виды активности CTGF, удовлетворили бы потребность в данной области и могли бы найти применение в терапевтическом лечении связанных с CTGF расстройств, включая легочные расстройства, такие как идиопатический пневмосклероз (IPF) и т.д.; почечные расстройства, такие как диабетическая нефропатия, гломерулосклероз и т.д.; и глазные расстройства, такие как ретинопатия, дегенерация желтого пятна и т.д.
Краткое изложение сущности изобретения
Настоящее изобретение относится к антителам, в частности к моноклональным антителам, и их частям, которые специфически связываются с областью на N-концевом фрагменте полипептида CTGF.
В одном из аспектов антитело по изобретению специфически связывается с областью человеческого CTGF (SEQ ID NO:2), что соответствует аминокислотам от примерно 103 до примерно 164 (SEQ ID NO:21); более конкретно, от аминокислоты примерно 135 до примерно аминокислоты 157 (SEQ ID NO:22) и, еще более конкретно, от примерно аминокислоты 142 до примерно аминокислоты 154 (SEQ ID NO:25); или ортологичной областью CTGF, полученного от другого вида. В определенных вариантах осуществления изобретения антитело имеет такую же специфичность, что и антитело, продуцируемое клеточной линией, обозначенной номером доступа в АТСС (Американской коллекции типовых культур) РТА-6006 (депонирована в АТСС 20 мая 2004 г.). В определенных вариантах осуществления изобретения антитело по существу идентично mAb1, как описано ниже. Предпочтительнее антитело по существу аналогично CLN1, как описано ниже. В еще одном варианте осуществления антитело по изобретению конкурентно связывается с любым из указанных выше антител к полипептиду CTGF.
В одном варианте осуществления настоящее изобретение относится к моноклональному антителу или его части, содержащим по меньшей мере один член группы, состоящей из последовательности тяжелой цепи иммуноглобулина, включающей SEQ ID NO:14, последовательности тяжелой цепи иммуноглобулина, включающей вариабельный домен SEQ ID NO:14, последовательности легкой цепи иммуноглобулина, включающей SEQ ID NO:20, последовательности легкой цепи иммуноглобулина, включающей вариабельный домен SEQ ID NO:20, или их консервативных вариантов. В определенном варианте осуществления антитело содержит вариабельный домен тяжелой цепи иммуноглобулина от аминокислотного остатка 1 до аминокислотного остатка 167 SEQ ID NO:14. В другом определенном варианте осуществления антитело содержит вариабельный домен легкой цепи иммуноглобулина от аминокислотного остатка 1 до аминокислотного остатка 136 SEQ ID NO:20. В конкретном варианте осуществления антитело содержит последовательность тяжелой цепи иммуноглобулина SEQ ID NO:14 и последовательность легкой цепи иммуноглобулина SEQ ID NO:20. В пределах этого варианта осуществления настоящее изобретение, в частности, относится к антителу CLN1 или его части, содержащей по меньшей мере остатки связывающей антиген области CLN1.
В определенных аспектах антитело по изобретению представляет собой поликлональное антитело. В других аспектах антитело представляет собой моноклональное антитело. В определенных вариантах осуществления изобретения антитело представляет собой гуманизированное моноклональное антитело, более предпочтительно человеческое моноклональное антитело. Любое из указанных выше антител может дополнительно иметь различную степень гликозилирования, что осуществляется клеткой, продуцирующей антитело, или применяется и/или модифицируется синтетически; или антитело может быть лишено гликозилирования. Антитело может быть необязательно пегилировано и/или модифицировано аналогичным образом для увеличения периода полувыведения из плазмы и т.д. В различных вариантах осуществления изобретение относится к фрагментам антитела, особенно когда фрагмент представляет собой фрагмент Fab, F(ab)2 или Fv.
В определенных аспектах антитело или его часть продуцируется клонированной клеточной линией. Клеточная линия может быть получена из любой модели на животных, используемой для продукции моноклонального антитела, включая, но не ограничиваясь этим, мышей, коз, кур и т.д. В частности клеточная линия может быть получена у мышей. Мыши могут являться стандартными мышами, используемыми для продукции антител, например, BALB/C, или модифицированной, например, трансгенной мышиной линии, оптимизированной или разработанной для продукции специфического изотипа, идиотипа или видоспецифических моноклональных антител. В одном варианте осуществления изобретения клеточная линия является линией клеток гибридомы, которая продуцирует и секретирует mAb1. В других вариантах осуществления линия клеток продуцирует и секретирует антитело или его часть, которые имеют свойство, по существу эквивалентное mAb1. В еще одних вариантах осуществления линия клеток продуцирует и секретирует антитело или его часть, которые имеют свойство, по существу эквивалентное CLN1. В конкретном варианте осуществления изобретение относится к линии клеток, обозначенной номером доступа в АТСС РТА-6006 (депонирована 20 мая 2004 г.).
В соответствии с другим аспектом изобретения антитело или его часть получены у трансгенного животного, отличного от человека, в частности трансгенного млекопитающего, отличного от человека, способного продуцировать человеческое антитело. Животное может быть любого вида, включая, но не ограничиваясь этим, мышь, курицу, корову, козу и т.д. В частности животное может представлять собой мышь. Такие антитела могут быть получены иммунизацией трансгенного млекопитающего, отличного от человека, фрагментом человеческого CTGF, например, SEQ ID NO:21 или, более конкретно, SEQ ID NO:22, или к ортологичной области CTGF, полученного от видов, отличных от человека. В определенных вариантах осуществления изобретения антитела получают иммунизацией трансгенного млекопитающего, отличного от человека, фрагментом CTGF, выбранным из группы, состоящей из последовательностей с SEQ ID NO:23 по SEQ ID NO:26, или к ортологичной области CTGF, полученного от видов, отличных от человека. В определенных вариантах осуществления антитела получены иммунизацией трансгенной мыши любым из указанных выше фрагментов. В других вариантах осуществления антитела получены иммунизацией трансгенной мыши функциональными эквивалентами любого из указанных выше фрагментов CTGF.
Термин "специфически связывается с областью CTGF" подразумевает, что антитела имеют специфичность связывания к определенной области CTGF, которая может быть определена первичной аминокислотной последовательностью или третичной, т.е. трехмерной, конформацией части полипептида CTGF. Специфичность связывания означает, что аффинность антител к части CTGF по существу больше, чем их аффинность к другим родственным полипептидам. Под термином "по существу большей аффинностью" заявители подразумевают, что имеется измеримое увеличение аффинности к части CTGF по сравнению с аффинностью к другим родственным полипептидам. Предпочтительно аффинность по меньшей мере в 1,5 раз, 2 раза, 5 раз, 10 раз, 100 раз, 103 раз, 104 раз, 105 раз, 106 раз больше к определенной части CTGF, чем к другим белкам. Предпочтительно специфичность связывания определяется с помощью аффинной хроматографии, иммунопреципитации или анализа связывания in vitro, такого как радиоиммуноанализ (RIA) или иммуноферментный анализ (ELISA), или анализ сортировки клеток с активацией флуоресценции (FASC). Более предпочтительно специфичность связывания определяется с помощью RIA или аффинной хроматографии, как описано ниже.
В предпочтительных вариантах осуществления изобретения антитела имеют аффинность, которая равна или больше, чем аффинность mAb1, описанная ниже, как определено, например, анализом Скэтчарда Munson and Pollard (1980, Anal Biochem 107:220). Аффинность антитела определяется как сила всех нековалентных взаимодействий между одним антигенсвязывающим сайтом антитела и одним эпитопом антигена. Аффинность рассчитывается путем измерения константы ассоциации (Ka), так что
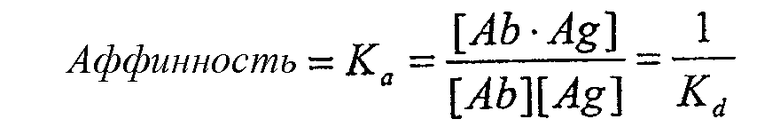
где [Ab] представляет собой концентрацию свободного антигенсвязывающего сайта антитела, [Ag] представляет собой концентрацию свободного антигена, [Ab·Ag] представляет собой концентрацию антигенсвязывающего сайта антитела, занятого антигеном, и Kd представляет собой константу диссоциации комплекса антитело-антиген. Предпочтительно антитела по изобретению имеют аффинность к CTGF, которая больше чем Kd=10-8, предпочтительно больше чем 10-9, предпочтительно больше чем 10-10, особенно для терапевтического применения. Преимущественно антитело в соответствии с изобретением имеет аффинность, аналогичную или большую, чем аффинность mAb1 (то есть Kd-9). Однако антитела, разделяющие эпитоп, связывающийся с mAb1, но имеющие более низкую аффинность (т.е. более высокое значение Kd), чем mAb1, также входят в объем изобретения и могут потенциально использоваться в различных анализах и диагностических видах применения, как описано в настоящем описании. Такие антитела могут дополнительно использоваться в терапевтических видах применения, особенно если они обладают высокой авидностью к антигену, как описано ниже.
Антитела в соответствии с изобретением могут быть одновалентными, двухвалентными или они могут быть многовалентными. В определенных вариантах осуществления изобретения предпочтительно, чтобы антитела по изобретению были двухвалентными или многовалентными. Любыми антителами по изобретению можно манипулировать для улучшения авидности, например, объединением связывающих эпитоп сайтов в один конструкт антитела, например, тритело, и т.д. Антитела в соответствии с изобретением могут представлять собой одноцепочечные антитела.
В некоторых случаях может быть полезно, чтобы антитела по изобретению проявляли подходящую аффинность к CTGF от других видов, например, для лечения и предотвращения расстройств у этих видов. Например, антитело по изобретению, которое проявляет подходящий Kd к собачьему CTGF, можно использовать для лечения связанного с CTGF расстройства у собак. Антитела по изобретению, которые проявляют перекрестно-видовую аффинность, такие как mAb1, также можно использовать в качестве инструментов исследования для изучения связанных с CTGF расстройств на различных моделях у животных. В другом аспекте антитело или его часть кодируется генетическим материалом, первоначально полученным у человека. Антитело может быть генерировано клетками в культуре, например, с использованием техники фагового дисплея, или оно может продуцироваться у животного, например, трансгенного животного, отличного от человека, содержащего гены иммуноглобулина, полученные у человека.
Кроме того, изобретение относится к рекомбинантным конструктам, содержащим части любых антител по изобретению, как описано выше, и белок, полученный из другого источника. Конкретно предусмотрены варианты осуществления, охватывающие химерные антитела, содержащие вариабельную область, полученную из моноклонального антитела, которое специфически связывается с областью на N-концевом фрагменте CTGF, и константную область, полученную из другого источника. Вариабельная область может быть получена из любого антитела, определенного изобретением, и, в частности, охватываются антитела, которые связываются с областью человеческого CTGF от примерно аминокислоты 97 до примерно аминокислоты 180 SEQ ID NO:2, или, более конкретно, от примерно аминокислоты 103 до примерно аминокислоты 164 SEQ ID NO:2, или, более конкретно, от примерно аминокислоты 134 до примерно аминокислоты 158 SEQ ID NO:2, или, еще более конкретно, от примерно аминокислоты 143 до примерно аминокислоты 154 SEQ ID NO:2, или к ортологичной области CTGF, полученного от другого вида. Константная область может быть получена из любого источника. В некоторых вариантах осуществления константную область получают из константной области человеческого иммуноглобулина.
Настоящее изобретение также относится к любому из описанных выше антител, где антитело дополнительно включает метку, способную обеспечить выявляемый сигнал, одно или вместе с другими веществами. Такие средства, осуществляющие мечение, могут быть выбраны из группы, состоящей из фермента, флуоресцентного вещества, хемилюминесцентого вещества, биотина, авидина и радиоактивного изотопа, но не ограничиваются ими. Настоящее изобретение также относится к любому из описанных выше антител, где антитело дополнительно включает цитотоксическое средство или фермент.
В других вариантах осуществления изобретения описанные выше антитела по изобретению дополнительно нейтрализуют по меньшей мере одну активность, связанную с CTGF. Такие виды активности, связанные с CTGF, включают, но не ограничиваются этим, стимуляцию клеточной миграции, продукцию внеклеточного матрикса клеткой in vivo или ex vivo и/или уменьшение фиброза у индивидуума. В конкретных вариантах осуществления биологическая активность выбрана из группы, состоящей из клеточного роста, дифференциации фибробластов и/или эндотелиальных клеток и индукции экспрессии белков, участвующих в формировании и перестройке внеклеточного матрикса, включая, например, коллагены, включающие, но не ограничивающиеся ими, типы I, II, III и IV, и фибронектин.
В определенных вариантах осуществления изобретения антитела специфически ингибируют миграцию клеток в анализах ex vivo. Предпочтительно антитела ингибируют стимулированную CTGF миграцию гладкомышечных клеток в анализе в камере Boyden. Например, в анализе миграции клеток, описанном ниже, антитела по изобретению повторно и воспроизводимо ингибируют вызванную CTGF миграцию. В различных вариантах осуществления антитела специфически уменьшают фиброз в случае моделей на животных. Предпочтительно антитела ингибируют развитие в случае моделей на животных пневмосклероза и фиброза почек. Например, антитела ослабляют вызванный блеомицином пневмосклероз у мышей на 60-70%, как было определено путем ингибирования легочного накопления гидроксипролина (коллагена) и/или гистологического исследования препаратов ткани, описанных ниже. Кроме того, антитела уменьшают накопление коллагена в случае модели остатка почки крысы (т.е. после нефрэктомии на 5/6) и у мышей после односторонней обструкции мочеточника (UUO), как описано ниже.
В других вариантах осуществления антитела по изобретению модулируют взаимодействие между полипептидом CTGF и секретируемым или ассоциированным с мембраной кофактором, нейтрализуя посредством этого биологическую активность CTGF. Кофактор может представлять собой любой белок, углеводород и/или липид; в определенных вариантах осуществления кофактором является член семейства ростовых факторов TGF-β, ВМР-4 и т.д.
В другом аспекте антитело уменьшает фиброз у субстанции. В различных вариантах осуществления субстанция представляет собой ткань или орган. В другом варианте осуществления субстанцией является животное, предпочтительно млекопитающее, наиболее предпочтительно человек. Когда субстанция представляет собой ткань, изобретение, в частности, предусматривает как эндогенные ткани, так и ткани ex vivo, например ткани трансплантата, ткань, выращенную в культуре и т.д. В различных вариантах осуществления ткань выбрана из группы, состоящей из эпителиальной, эндотелиальной и соединительной ткани. Когда субстанция представляет собой орган, изобретение, в частности, предусматривает органы, выбранные из группы, включающей почку, легкое, печень, глаз, сердце и кожу. В предпочтительных вариантах осуществления субстанцией является животное, в частности животное вида млекопитающих, включая вид крыс, кроликов, коров, овец, свиней, мышей, лошадей и приматов. В самом предпочтительном варианте осуществления субстанцией является человек.
В определенных вариантах осуществления изобретения антитело используется для лечения или профилактики связанного с CTGF расстройства у субъекта, у которого есть связанное с CTGF расстройство или риск его развития. Такие расстройства включают, но не ограничиваются этим, различные виды рака, включая острый лимфобластический лейкоз, дерматофибромы, рак молочной железы, глиому и глиобластому, рабдомиосаркому и фибросаркому, десмоплазию, ангиолипому, ангиолейомиому, десмопластические формы рака и рост и метастазы рака предстательной железы, яичников, толстой и прямой кишки, поджелудочной железы, желудочно-кишечного тракта и печени и других опухолей. Связанные с CTGF расстройства также включают различные фибротические расстройства, включая, но не ограничиваясь этим, идиопатический пневмосклероз, фиброз почек, гломерулосклероз, глазной фиброз, остеоартрит, склеродермию, сердечный фиброз и фиброз печени. Фиброз может возникнуть в любом органе или ткани, включая орган, выбранный из почки, легкого, печени, сердца и кожи, но не ограничиваясь ими; или в ткани, выбранной из эпителиальной, эндотелиальной и соединительной ткани, но не ограничиваясь ими. В других вариантах осуществления связанное с CTGF расстройство может быть вызвано любым инициирующим фактором, включая, но не ограничиваясь этим, контакт с химическими веществами или биологическими агентами, воспалительную реакцию, аутоиммунную реакцию, травму, хирургические процедуры и т.д. Связанные с CTGF расстройства также включают, но не ограничиваются этим, расстройства вследствие гипергликемии и гипертонии. Такие расстройства могут возникнуть, например, вследствие сахарного диабета, ожирения и т.д., и включают диабетическую нефропатию, ретинопатию и сердечно-сосудистые заболевания.
Поэтому в различных вариантах осуществления изобретение относится к антителам, которые могут использоваться для лечения или профилактики связанных с CTGF расстройств у индивидуума. Настоящее изобретение также относится к применению таких антител при изготовлении лекарственного средства для лечения связанных с CTGF расстройств.
В другом аспекте изобретение относится к способу нейтрализации активности, связанной с CTGF, включающему контактирование антитела по изобретению и полипептида CTGF, нейтрализуя посредством этого биологическую активность CTGF, такую как виды активности, описанные выше. Биологическая активность может представлять собой любую активность CTGF, включая, но не ограничиваясь этим, стимуляцию миграции клеток и продукцию внеклеточного матрикса. В различных вариантах осуществления нейтрализация происходит in vitro. В других вариантах осуществления нейтрализация происходит у индивидуума in vivo.
В еще одном аспекте изобретение относится к способам применения антитела, как описано выше, для лечения связанного с CTGF расстройства у нуждающегося в нем пациента, где способ включает введение антитела или его фармацевтической композиции пациенту, осуществляя посредством этого лечение расстройства. Индивидуум может представлять собой пациента с диагностированным или предполагаемым связанным с CTGF расстройством, включая, например, расстройство, возникающее в результате избыточной продукции внеклеточного матрикса. В определенных аспектах связанное с CTGF расстройство выбрано из рака или фибротического расстройства. Виды рака включают, но не ограничиваются этим, острый лимфобластический лейкоз, дерматофибромы, рак молочной железы, карциному молочной железы, глиому и глиобластому, рабдомиосаркому и фибросаркому, десмоплазию, ангиолипому, ангиолейомиому, десмопластические формы рака и рак предстательной железы, яичников, толстой и прямой кишки, поджелудочной железы, желудочно-кишечного тракта и печени, и фибротические расстройства включают, но не ограничиваются этим, идиопатический пневмосклероз, фиброз почек, гломерулосклероз, глазной фиброз, дегенерацию желтого пятна, остеоартрит, склеродермию, хроническую сердечную недостаточность, сердечный фиброз и фиброз печени. В других вариантах осуществления связанное с CTGF расстройство может быть вызвано любым инициирующим фактором, включая, но не ограничиваясь этим, контакт с химическими веществами или биологическими агентами, воспалительную реакцию, аутоиммунную реакцию, травму, хирургические процедуры и т.д. Связанные с CTGF расстройства также включают, но не ограничиваются этим, расстройства вследствие гипергликемии и гипертонии. Такие расстройства могут возникнуть, например, вследствие сахарного диабета, ожирения и т.д., и включают диабетическую нефропатию, ретинопатию и сердечно-сосудистые заболевания.
В другом аспекте настоящее изобретение относится к композиции, содержащей антитело, как описано выше, и по меньшей мере один другой компонент. Компоненты могут включать любое соединение, молекулу или вещество, включая, например, белки, нуклеиновые кислоты, углеводороды, липиды и т.д. Кроме того, компоненты могут включать различные растворители, соли и другие носители и/или эксципиенты. В некоторых вариантах осуществления композиция представляет собой фармацевтическую композицию, содержащую антитело, как описано выше, и по меньшей мере один дополнительный компонент, выбранный из растворителя, стабилизатора или эксципиента. В определенном варианте осуществления фармацевтическая композиция включает антитело в смеси с фармацевтически приемлемым носителем. Фармацевтическая композиция может дополнительно содержать второе терапевтическое средство, например ингибитор ангиотензин-превращающего фермента (АСЕ), средство для расщепления или ингибирования конечного продукта гликирования и т.д. Изобретение, кроме того, относится к лекарственным средствам, содержащим антитело, как определено выше, для лечения индивидуума, имеющего связанное с CTGF расстройство. Такие расстройства включают, но не ограничиваются этим, различные формы рака и фибротические расстройства; расстройства, возникающие в результате таких состояний, как инфаркт миокарда, артрит и воспаление; и расстройства вследствие сахарного диабета, ожирения и им подобных, которые могут включать диабетическую нефропатию, ретинопатию и сердечно-сосудистые заболевания.
В другом варианте осуществления изобретение относится к полипептидной последовательности, выбранной из группы, состоящей из SEQ ID NO:14, аминокислот 1-167 SEQ ID NO:14, SEQ ID NO:20, и аминокислот 1-136 SEQ ID NO:20. Изобретение также охватывает консервативные варианты полипептидов. В другом варианте осуществления изобретение относится к специфическим фрагментам CTGF человека, выбранным из группы, состоящей из последовательностей от SEQ ID NO:21 до SEQ ID NO:26, и ортологичным фрагментам CTGF, полученным от видов, отличных от человека.
Указанные выше полипептиды могут представлять собой "измененные" полипептиды, как определено ниже.
В другом варианте осуществления изобретение относится к полинуклеотидной последовательности, кодирующей антитело по изобретению, или его часть. В определенных вариантах осуществления изобретения полинуклеотидная последовательность выбрана из группы, состоящей из полинуклеотидной последовательности, кодирующей SEQ ID NO:14, полинуклеотидной последовательности, кодирующей аминокислоты 1-167 SEQ ID NO:14, полинуклеотидной последовательности SEQ ID NO:13, и полинуклеотида, включающего нуклеотиды 1-501 SEQ ID NO:13. В другом варианте осуществления полинуклеотидная последовательность выбрана из группы, состоящей из полинуклеотидной последовательности, кодирующей SEQ ID NO:20, полинуклеотидной последовательности, кодирующей аминокислоты 1-136 SEQ ID NO:20, полинуклеотида SEQ ID NO:19, и полинуклеотида, включающего нуклеотиды 1-408 SEQ ID NO:19.
Указанные выше полинуклеотиды могут представлять собой "измененные" полинуклеотиды, как определено ниже.
Изобретение, кроме того, относится к рекомбинантным полинуклеотидам, содержащим любую из полинуклеотидных последовательностей, описанных выше, операбельно связанных с последовательностью вектора, которая содержит последовательности регулировки репликации и траскрипции. В одном из аспектов рекомбинантный полинуклеотид кодирует аминокислотную последовательность SEQ ID NO:14 или вариабельный домен в ней. В другом аспекте рекомбинантный полинуклеотид включает SEQ ID NO:13. В еще одном аспекте рекомбинантный полинуклеотид кодирует аминокислотную последовательность SEQ ID NO:20 или вариабельный домен в ней. В еще одном аспекте рекомбинантный полинуклеотид включает SEQ ID NO:19.
Изобретение также относится к клеткам-хозяевам, трансфецированным по меньшей мере одним из описанных выше рекомбинантных полинуклеотидов. Клетки-хозяева включают любую прокариотическую и эукариотическую клетку-хозяина, включая, например, клонированные клеточные линии, поддерживаемые способами культивирования, известными специалистам в данной области. Клетки-хозяева также включают трансгенные растения и животных, полученных из трансформированных клеток, например, стволовых клеток. В одном варианте осуществления клетка-хозяин включает клетку, совместно трансфецированную полинуклеотидом, кодирующим SEQ ID NO:14, и полинуклеотидом, кодирующим SEQ ID NO:20, и которая продуцирует функциональное антитело с характеристиками, по существу такими же как у mAb1. В определенных вариантах осуществления изобретения антитело представляет собой CLN1. В другом конкретном варианте осуществления клетка-хозяин обозначена номером доступа в АТСС РТА-6006 (депонирована 20 мая 2004 г.).
Эти и другие варианты осуществления настоящего изобретения будут легко понятны специалистам в данной области в свете представленного описания, и все такие варианты осуществления конкретно предусмотрены настоящим изобретением.
Краткое описание чертежей
На фиг.1А и 1В показано, что структура и последовательности фактора роста соединительной ткани сохраняются. На фиг.1А показана модулярная структура домена CTGF, который включает консервативные мотивы для белка, связывающего подобный инсулину фактор роста (IGF-BP), и фактора фон Виллебранда (VWC) в N-концевом фрагменте и мотив тромбоспондина (TSP1) и узла цистеина (СТ) в С-концевом фрагменте. На фиг.1В показано упорядочение множественных последовательностей между N-концевыми фрагментами ортологов человеческого (hCTGF), коровьего (bCTGF), свиного (pCTGF), крысиного (rCTGF) и мышиного (FIPS12)CTGF. Упорядочение было создано с использованием программы CLUSTAL W (версия 1.74; Thompson et al. (1994) Nucleic Acids Res., 22: 4673-4680) с использованием параметров по умолчанию. На чертеже звездочка (*) показывает полное сохранение аминокислотного остатка среди представленных видов.
На фиг.2А и 2В показаны графики Скэтчарда конкурентного связывания между меченым и немеченым человеческим CTGF соответственно с антителами против CTGF, mAb2 и mAb1. mAb1 представляет собой иллюстративное антитело по настоящему изобретению.
На фиг.3А показан фрагмент антитела Fab (Mr 45 кДа), полученный после расщепления папаином соответствующего IgG антитела mAb1 и последующей аффинной хроматографии на белке А-сефарозе (полоса 2), что было показано с помощью SDS-PAGE (электрофореза в полиакриламидном геле с додецилсульфатом натрия). На фиг.3В показано связывание фрагмента Fab и соответствующего IgG с CTGF в зависимости от увеличивающейся концентрации хаотропного агента (тиоцианата).
На фиг.4А и 4В показаны графики Скэтчарда конкурентного связывания между меченым рекомбинантным человеческим CTGF и немеченым крысиным CTGF соответственно с антителами против CTGF, mAb2 и mAb1.
На фиг.5А, 5В и 5С представлены терапевтические благоприятные эффекты антитела по изобретению в случае модели интерстициального фиброза в легком. На фиг.5А показан эффект лечения антителом в случае вызванного блеомицином увеличения содержания гидроксипролина в легких мышей. Количество животных в каждой группе показано в скобках под каждым столбцом, а группы лечения указаны по оси х. SA: солевой раствор; BL: блеомицин; AbsJ: пул из 3 моноклональных антител по изобретению; mAb1, иллюстративное антитело по изобретению. Величины выражены в виде средней величины ± SE (стандартная ошибка). На фиг.5В и 5С показаны окрашенные гематоксилином и эозином парафиновые срезы легочных проксимальных ацинусов от мышей, подвергнутых воздействию блеомицина при внутритрахеальной инъекции, и последующему лечению соответственно солевым раствором или антителами по изобретению. На фиг.5В тонкие межальвеолярные перегородки ацинуса имеют патологический внешний вид, и присутствуют воспалительные клетки и фиброз. На фиг.5С паренхима в основном нормальная и имеется лишь умеренное утолщение межальвеолярных перегородок.
На фиг.6А, 6В и 6С представлены терапевтические благоприятные эффекты антитела по изобретению в случае модели канальциево-интерстициального фиброза почки. На фиг.6А показано уменьшение фиброза благодаря односторонней обструкции мочеточника (UUO) после лечения антителом по изобретению, mAb1, или антителом mAb3, направленным на С-конец CTGF. Степень фиброза выражена в виде соотношения между гидроксипролином и пролином в почке с обструкцией по сравнению с почкой без обструкции (средняя величина ± SE). На фиг.6В и 6С показаны окрашенные трихромом парафиновые срезы почки с обструкцией, в случае лечения соответственно солевым раствором или антителом.
На фиг.7А и 7В представлен терапевтический благоприятный эффект антитела по изобретению в случае модели клубочкового фиброза в почке. На фиг.7А и 7В показаны микрофотографии окрашенной трихромом ткани остатка почки после получения лечения соответственно солевым раствором или антителом.
На фиг.8А, 8В, 8С, 8D, 8E, 8F и 8G показана индукция локализованных подкожных гранулем у новорожденных мышей. Слева на фиг.8А и 8В показано образование гранулемы в участке подкожной инъекции одного TGFβ или TGFβ и CTGF, соответственно. Справа на фиг.8С-G показана гистологическая панель, представляющая систему балльной оценки (от 0 [нормальная] до 4 [фибротическая]), используемую для оценки терапевтического благоприятного эффекта антитела.
На фиг.9 показана степень фиброза в модели локализованной подкожной гранулемы с лечением антителом против CTGF и без него. mAb1 представляет собой иллюстративное антитело по изобретению, тогда как mAb3 представляет собой антитело против CTGF, которое специфически связывается с С-концевым эпитопом CTGF.
На фиг.10А, 10В, 10С и 10D представлен терапевтический благоприятный эффект антитела по изобретению при фиброзе органа в случае использования модели системного склероза (склеродермии). На каждой панели представлено изменение накопления коллагена в соответствующих органах после лечения солевым раствором (контроль), TGFβ и CTGF или лечения TGFβ и CTGF в сочетании с лечением антителом.
На фиг.11А и 11В показано графическое изображение клонирования тяжелой и легкой цепей иммуноглобулина иллюстративного антитела по изобретению, mAb1. На фиг.11А показано упорядочение фрагментов тяжелой цепи PCR (полимеразной реакции синтеза цепи), используемых для определения кодирующей последовательности тяжелой цепи (CDS) mAb1. На фиг.11В показано упорядочение фрагментов легкой цепи PCR, используемых для определения кодирующей последовательности легкой цепи (CDS) mAb1.
На фиг.12А и 12В представлены результаты исследования связывания между CTGF и TGFβ. На фиг.12А показана степень связывания между TGFβ и либо CTGF, фрагментом CTGF, кодируемым экзоном 3 (Экзоном 3), либо фрагментом CTGF, кодируемым экзоном 5 (Экзоном 5), в присутствии или в отсутствие антитела против CTGF. На фиг.12В показана степень, до которой антитела против CTGF ингибируют взаимодействие TGFβ и CTGF. На фигуре антитела включают иллюстративные антитела по изобретению, mAb1 и mAb4 и антитело, которое специфически связывается с С-концевым эпитопом CTGF, mAb3.
Описание изобретения
Прежде чем описывать настоящие композиций и способы, следует понимать, что изобретение не ограничивается описанными конкретными методологиями, последовательностями операций, линиями клеток, анализами и реагентами, поскольку они могут варьироваться. Следует также понимать, что используемая в настоящем описании терминология предназначена для описания конкретных вариантов осуществления настоящего изобретения и никоим образом не предназначена для ограничения объема настоящего изобретения, как изложено в прилагаемой формуле изобретения.
Пока нет других определений, все используемые в настоящем описании технические и научные термины имеют такие же значения, которые обычно понятны среднему специалисту в данной области, для которого предназначено это изобретение. Хотя в практике или тестировании настоящего изобретения можно использовать любые способы и материалы, аналогичные или эквивалентные тем, которые описаны в настоящем изобретении, здесь описаны предпочтительные способы, устройства и материалы. Все приведенные в настоящем описании публикации приведены здесь в качестве ссылки с целью описания и раскрытия методологий, реагентов и инструментов, о которых сообщалось в публикациях, которые могут использоваться в связи с изобретением. В настоящем описании ничто не следует толковать как допущение того, что изобретение не дает право предвосхищать такое раскрытие на основании предыдущего изобретения.
При отсутствии иных указаний при осуществлении настоящего изобретения используются обычные способы химии, биохимии, молекулярной биологии, клеточной биологии, генетики, иммунологии и фармакологии в пределах навыков в данной области. Полное объяснение таких методик имеется в литературе (см. например, Gennaro A.R., ed. (1990) Remington`s Pharmaceutical Sciences, 18th ed., Mack Publishing Co.; Colowinck S. et al, Eds., Methods In Enzymology, Academic Press, Inc.; Handbook of Experimental Immunology, Vols. I-IV (D.M. Weir and C.C. Blackwell, eds., 1986, Blackwell Scientific Publications); Maniatis T. et al., eds. (1989) Molecular Cloning: A Laboratory Manual, 2nd edition, Vols. I-III, Cold Spring Harbor Laboratory Press; Ausubel F.M. et al., eds. (1999) Short Protocols on Molecular Biology, 4th edition, John Wiley & Sons; Ream et al., eds. (1998) Molecular Biology Techniques: An Intensive Laboratory Course, Academic Press); PCR (Introduction to Biotechniques Series), 2nd ed. (Newton & Graham eds., 1997, Springer Verlag).
Определения
"Фактор роста соединительной ткани" или "CTGF" относится к аминокислотным последовательностям по существу очищенного CTGF, полученного у любого вида, в частности вида млекопитающих, включая крысу, кролика, корову, овцу, свинью, мышь, лошадь и человекообразную обезьяну, предпочтительно у человека, и из любого источника, натурального, синтетического, полусинтетического или рекомбинантного.
Термин "N-концевой фрагмент" CTGF относится к любому полипептиду, содержащему последовательности, полученные из амино-концевой части полипептида CTGF, или к любому его варианту или фрагментам. N-концевые фрагменты могут не включать или включать весь CTGF или его части от начального метионинового остатка до лишенного цистеина шарнира, как показано на фиг.1А и 1В. Далее, N-концевые фрагменты могут не включать или включать весь белковый мотив, связывающий инсулиновый фактор роста, и/или домен фон Виллебранда типа С (SEQ ID NO:21) или их части, как показано на фиг.1В. N-концевые фрагменты CTGF могут также не включать или включать всю свободную от цистеина область или её части. Далее, N-концевые фрагменты CTGF могут представлять собой любые 15 или более соседних аминокислот, содержащихся в пределах любого определенного выше предшествующего N-концевого фрагмента.
В одном аспекте "N-концевой фрагмент" CTGF относится к полипептидным последовательностям, полученным из амино-концевой части человеческого CTGF. Такие фрагменты могут охватывать всю область от аминоксилотного остатка 1 до примерно аминокислотного остатка 198 SEQ ID NO:2 или от примерно аминокислоты 23 до примерно аминокислоты 198 SEQ ID NO:2. Граница N-концевого фрагмента в пределах шарнира может быть необязательно определена одним из нескольких сайтов расщепления протеазы, определенных в SEQ ID NO:2, таких как сайты расщепления химотрипсина между остатками 179 и 180, между остатками 182 и 183 и между остатками 188 и 189; сайты расщепления плазмина между остатками 183 и 184 и между остатками 196 и 197; и сайт расщепления костного морфогенетического белка-1 между остатками 169 и 170. Кроме того, N-концевые фрагменты человеческого CTGF могут не включать или включать всю или части области аминокислот 27-97 SEQ ID NO:2, аминокислот 103-166 SEQ ID NO:2 или аминокислот 167-198 SEQ ID NO:2. Далее, N-концевые фрагменты человеческого CTGF могут представлять собой любые 15 или более соседних аминокислот, содержащихся в пределах любого определенного выше предшествующего N-концевого фрагмента.
В определенных вариантах осуществления изобретения N-концевые фрагменты CTGF настоящего изобретения содержат последовательности, выбранные из следующих областей человеческого CTGF (SEQ ID NO:2) и его ортологичных фрагментов, полученных от различных видов, в частности вида млекопитающих, включая крысу, кролика, корову, овцу, свинью, мышь, лошадь: аминокислотные остатки 23-96 (кодируемые экзоном 2); аминокислотные остатки 27-97 (мотив IGF-BP); аминокислотные остатки 97-180 (кодируемые экзоном 3); аминокислотные остатки 103-166 (домен VWC); аминокислотные остатки 167-198 (лишенный цистеина шарнир); аминокислотные остатки 23-180 (кодируемые экзонами 2 и 3); аминокислотные остатки 27-166 (IGF-BP и VWC); и аминокислотные остатки 23-198 (см. фиг.1В).
Термин "С-концевой фрагмент" CTGF относится к любому полипептиду, содержащему последовательности, полученные из карбокси-концевой части аминокислотной полипептидной последовательности CTGF, или к любым его вариантам или фрагментам. С-концевые фрагменты CTGF могут не включать или включать всю или части лишенной цистеина области полипептида CTGF (аминокислоты 167-198 SEQ ID NO:2).
С-концевые фрагменты могут не включать или включать всю или части CTGF от лишенного цистеина шарнира до конца белка. Далее, С-концевые фрагменты могут не включать или включать всю или части мотива тромбоспондина и/или мотива цистеинового узла. Далее, С-концевые фрагменты CTGF могут представлять собой любые 15 или более соседних аминокислот, содержащихся в пределах любого определенного выше предшествующего С-концевого фрагмента.
В некоторых аспектах изобретения С-концевые фрагменты могут охватывать всю область от аминокислотного остатка 181 по примерно аминокислотный остаток 349 SEQ ID NO:2. Граница С-концевого фрагмента внутри шарнира может быть необязательно определена одним из нескольких сайтов расщепления протеазы, определенных в SEQ ID NO:2, таких как определенные выше сайты расщепления химотрипсина, плазмина и костного морфогенетического белка-1. Кроме того, С-концевые фрагменты включают последовательности, выбранные из следующих областей человеческого CTGF (SEQ ID NO:2) и их ортологичных фрагментов, полученных у различных видов, в частности видов млекопитающих, включая крысу, кролика, корову, овцу, свинью, мышь и лошадь: аминокислоты 201-242 SEQ ID NO:2, аминокислоты 247-349 SEQ ID NO:2, аминокислоты 248-349 SEQ ID NO:2 или аминокислоты 249-346 SEQ ID NO:2. Далее, С-концевые фрагменты человеческого CTGF могут представлять собой любые 15 или более соседних аминокислот, содержащихся внутри любого определенного выше предшествующего С-концевого фрагмента.
Термины "лишенная цистеина область" или "шарнир" CTGF относятся к любому полипептиду, происходящему от примерно аминокислотного остатка 167 до примерно аминокислотного остатка 198 человеческого CTGF (SEQ ID NO:2), и его ортологичным фрагментам, полученным от различных видов, в частности вида млекопитающих, включая крысу, кролика, корову, овцу, свинью, мышь, лошадь.
Используемые в настоящем описании термины "аминокислотная последовательность" или "полипептид" или "полипептиды" относятся к олигопептидным, пептидным, полипептидным или белковым последовательностям и их фрагментам и к природным или синтетическим молекулам. Полипептидный фрагмент или аминокислотный фрагмент представляет собой часть полипептида, которая сохраняет по меньшей мере одну структурную и/или функциональную характеристику полипептида. Фрагменты CTGF включают любую часть полипептидной последовательности CTGF, которая сохраняет по меньшей мере одну структурную и/или функциональную характеристику CTGF. Когда "аминокислотная последовательность" указывается для обозначения полипептидной последовательности природной молекулы белка, термин "аминокислотная последовательность" и подобные термины не предназначены для ограничения аминокислотной последовательности полной нативной последовательностью, связанной с указанной молекулой белка.
Термин "иммуногенность" относится к способности вещества при введении в организм стимулировать иммунный ответ и продукцию антитела. Средство, проявляющее свойство иммуногенности, назывется иммуногенным. Иммуногенные средства могут включать, но не ограничиваются этим, разнообразные макромолекулы, такие как, например, белки, липопротеиды, полисахариды, нуклеиновые кислоты, бактерии и бактериальные компоненты и вирусы и вирусные компоненты. Иммуногенные средства часто имеют молекулярную массу более чем 10 кДа. Антигенными фрагментами именуются фрагменты полипептида CTGF, предпочтительно фрагменты длиной примерно от 5 до 15 аминокислот, которые сохраняют по меньшей мере один биологический или иммунологический аспект активности полипептида CTGF.
Термин "антитело" относится к интактным молекулам, а также к их фрагментам, таким как Fab, F(ab')2 и Fv, которые способны связывать эпитопную детерминанту, и включают поликлональные и моноклональные антитела. Антитела, которые связывают CTGF или фрагменты CTGF, можно получить с использованием интактных полипептидов или с использованием фрагментов, содержащих мелкие интересующие пептиды в качестве иммунизирующего антигена. Полипептид или олигопептид, используемый для иммунизации животного (например, мыши, кролика, курицы, индюшки, козы и т.д.), может быть получен в результате трансляции РНК или синтезирован химически, и при желании он может быть конъюгирован с белком-носителем. Обычно используемые носители, химически соединенные с пептидами, включают, например, бычий сывороточный альбумин, тиреоглобулин и гемоцианин лимфы улитки (KLH).
Используемый в настоящем описании термин "моноклональное антитело" относится по существу к однородной популяции антител, т.е. к отдельным антителам, включающим популяцию, которая является идентичной по специфичности и аффинности, за исключением возможных природных мутаций, которые могут присутствовать в небольших количествах. Следует отметить, что композиция моноклонального антитела может содержать более одного моноклонального антитела.
Моноклональные антитела, включенные в пределы объема изобретения, включают гибридные и рекомбинантные антитела (например, "гуманизированные" антитела), независимо от видов происхождения или обозначения класса или подкласса иммуноглобулина, а также фрагменты антител (например, Fab, F(ab')2 и Fv), имеющие по меньшей мере одну из определенных характеристик антител, описанных в настоящем описании. Предпочтительные варианты осуществления включают антитела, способные связываться по существу с тем же эпитопом, что и эпитоп, распознаваемый моноклональным антителом mAb1, и/или имеющие аффинность к этому эпитопу, которая больше или равна аффинности mAb1.
Термин "моноклональное" указывает на характер антитела как по существу однородной популяции антител и не должен трактоваться как требующий продукцию антитела каким-либо определенным способом. Например, моноклональные антитела по изобретению могут быть изготовлены методом гибридомы, впервые описанным Kohler and Milstein (1975, Nature 256:495-497), или могут быть изготовлены методами рекомбинантной ДНК (например, см., Celltech Therapeutics Ltd., европейский патент EP-0120694; Cabilly et al, патент США № 4816567 или Mage and Lamoyi (1987; In: Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, pp. 79-97).
Используемый в настоящем описании термин "нейтрализующее антитело" относится к антителу, предпочтительно моноклональному антителу, которое способно существенно ингибировать или устранить биологическую активность CTGF. Обычно нейтрализующее антитело ингибирует связывание CTGF с кофактором, таким как TGFβ c CTGF-специфичным рецептором, связанным с клеткой-мишенью или с другой биологической мишенью. В определенном варианте осуществления нейтрализующее антитело ингибирует биологическую активность CTGF в степени, приблизительно равной или большей, чем mAb1, предпочтительно нейтрализующее антитело ингибирует биологическую активность CTGF в степени, приблизительно равной или большей, чем CLN1.
Используемая в настоящем описании фраза "связанные с CTGF расстройства" относится к состояниям и заболеваниям, связанным с аномальной или измененной экспрессией или активностью CTGF. Аномальная экспрессия CTGF связана с клеточными пролиферативными расстройствами, такими как расстройства, вызванные пролиферацией эндотелиальных клеток; клеточная миграция, виды роста, подобные опухолевому росту; общее рубцевание ткани и различные заболевания, характеризуемые несоответствующим отложением внеклеточного матрикса.
Расстройства, связанные с CTGF, включают, но не ограничиваются этим, расстройства, вовлекающие ангиогенез и другие процессы, которые играют центральную роль при таких состояниях как пролиферативная витреоретинопатия, рак, включая острый лимфобластический лейкоз, дерматофибромы, рак молочной железы, карцинома молочной железы, глиома и глиобластома, рабдомиосаркома и фибросаркома, десмоплазия, ангиолипома, ангиолейомиома, дермопластические раковые опухоли, и рак предстательной железы, яичников, толстой и прямой кишки, поджелудочной железы, желудочно-кишечного тракта и печени; другой опухолевой рост и метастазы и т.д.
Расстройства, связанные с CTGF, также включают фибротические расстройства и связанные с ними состояния, такие как избыточное рубцевание, возникающее в результате локализованного или системного фиброза, хронический или острый фиброз органов, таких как почка, легкие, печень, глаза, сердце, кожа и т.д.; или ткани, выбранной, но не ограничивающейся этим, из эпителиальной, эндотелиальной и соединительной ткани. Фиброз может также возникнуть в глазах и суставах. Такие расстройства, связанные с CTGF, включают, например, сердечный фиброз, включая сердечный реактивный фиброз или сердечная перестройка после инфаркта миокарда или застойной сердечной недостаточности, легочные расстройства, включая интерстициальный легочный фиброз и т.д.; фиброз, связанный с диализом, включая перитонеальный диализ, например постоянный амбулаторный перитонеальный диализ (CAPD); перидуральный фиброз, почечный фиброз, легочный фиброз, интерстициальный фиброз, кожный фиброз и фиброз, возникающий в результате острых или повторных травм, включая хирургические операции, химиотерапию, лучевое лечение, отторжение аллотрансплантата, хроническое и острое отторжение трансплантата (например, почки, печени или другого органа), облитерирующий бронхиолит, например, после трансплантации легких, и воспаление и инфекция, например, вследствие болезни или повреждения.
Кроме того, расстройства, связанные с CTGF, включают, но не ограничиваются этим, склеротические состояния, включая системный склероз, склеродермию, келоиды, гипертрофическое рубцевание и другие дерматологические заболевания и состояния; атеросклероз, такой как состояния, включающие атеросклеротические бляшки. И атеросклероз, связанный с сахарным диабетом, перитонеальным диализом и т.д.; артрит, включая ревматоидный артрит, остеоартрит и другие суставные воспалительные состояния и т.д.; интерстициальные заболевания, включая интерстициальный фиброз; болезнь Крона; воспалительное заболевание кишечника; ретинопатии, включая, например, пролиферативную витреоретинопатию, непролиферативную диабетическую ретинопатию, пролиферативную диабетическую ретинопатию и дегенерацию желтого пятна, включая связанную с возрастом и ювенильную болезнь (Stargardt), и отслоение пигментного эпителия; нефропатии, включая диабетическую нефропатию, нефропатию, связанную с IgA, нефропатию, связанную с токсичностью, волчаночное почечное заболевание и т.д.; и состояния, связанные с химической токсической деструкцией канальцев.
Расстройства, связанные с CTGF, также включают, но не ограничиваются этим, расстройства вследствие гипергликемии, гипертонии, образование конечного продукта гликирования (AGE) и т.д. Такие расстройства могут возникнуть, например, вследствие сахарного диабета, ожирения и т.д. и включают диабетическую нефропатию, ретинопатию и сердечно-сосудистые заболевания. Кроме того, расстройства, связанные с CTGF, могут быть вызваны любым инициирующим фактором, включая, но не ограничиваясь этим, контакт с химическими или биологическими веществами, воспалительную реакцию, аутоиммунную реакцию, травму, хирургические процедуры и т.д. В некоторых вариантах осуществления используются способы для лечения пациента, предрасположенного к расстройству, связанному с CTGF, вследствие состояния, включающего, но не ограничивающегося этим, инфаркт миокарда, артрит и локальное или системное воспаление.
Указанные в настоящем описании "пролиферативные" процессы и расстройства включают патологические состояния, характеризуемые постоянным размножением клеток, приводящим к избыточному росту клеточной популяции в пределах ткани. Клеточные популяции необязательно представляют собой трансформируемые вызывающие рост опухолей или злокачественные клетки, но они могут также включать нормальные клетки. Например, CTGF может участвовать патологически при пролиферативном поражении слоя интимы артериальной стенки, приводя к атеросклерозу или стимуляции неоваскуляризации.
Термин "рак" относится к любому автономному росту ткани, включая неконтролируемый, аномальный рост клеток, или к любой злокачественной опухоли с потенциально неограниченным ростом, которая распространяется локально путем внедрения и системно - метастазированием. Рак также относится к любому аномальному состоянию, обозначаемому раком.
Термин "фиброз" относится к аномальному процессингу фиброзной ткани или к фиброидной или фиброзной дегенерации. Фиброз может возникнуть в результате различных травм или заболеваний, и он может часто возникнуть в результате хронического отторжения трансплантата, связанного с трансплантацией различных органов. Фиброз обычно вовлекает аномальную продукцию, накопление или отложение компонентов внеклеточного матрикса, включая избыточную продукцию и увеличенное отложение, например, коллагена и фибронектина. Термин "фиброз" используется в настоящем описании в его самом широком смысле, и он относится к любой избыточной продукции или отложению белков внеклеточного матрикса. Имеются многочисленные примеры фиброза, включая образование рубцовой ткани после сердечного приступа, что нарушает способность сердца нагнетать кровь. Сахарный диабет часто вызывает повреждение/рубцевание в почках, что ведет к прогрессирующей потере почечной функции; и в глазах, что вызывает потерю зрения. После операции рубцовая ткань может образовываться между внутренними органами, вызывая контрактуру, боль и, в некоторых случаях бесплодие. Основные органы, такие как сердце, почка, печень, глаз и кожа склонны к хроническому рубцеванию, обычно связанному с другими заболеваниями. Гипертрофические рубцы (конгломерат незлокачественной ткани) представляют собой обычную форму фиброза, вызванного ожогами и другой травмой. Кроме того, существует ряд других фибропролиферативных расстройств, включая склеродермию, келоиды и атеросклероз, которые связаны соответственно с общим рубцеванием ткани, видами роста, подобными опухолевому росту, в коже, или стойкое рубцевание кровеносных сосудов, которое нарушает способность переносить кровь.
Термины "нуклеиновая кислота" или "полинуклеотид" или "полинуклеотиды" относятся к олигонуклеотидам, нуклеотидным последовательностям или полинуклеотидам или к любым их фрагментам и к ДНК и РНК природного или синтетического происхождения, которые могут быть одно- или двухнитевыми и могут представлять смысловую или антисмысловую нить, к пептидной нуклеиновой кислоте (PNA), или к любому материалу, подобному ДНК или РНК, природному или синтетическому по происхождению. Полинуклеотидные фрагменты представляют собой любую часть полинуклеотидной последовательности, которая сохраняет по меньшей мере одну структурную или функциональную характеристику полинуклеотида. Фрагменты полинуклеотидов могут быть вариабельной длины, например длиной более чем 60 нуклеотидов, длиной по меньшей мере 100 нуклеотидов, длиной по меньшей мере 1000 нуклеотидов, или длиной по меньшей мере 10000 нуклеотидов.
"Измененные" полинуклеотиды включают полинуклеотиды с делециями, инсерциями или заменами различных нуклеотидов, приводящими к получению полинуклеотида, который кодирует такой же или функционально эквивалентный полипептид. В это определение включены последовательности, проявляющие полиморфизм, который может быть или не быть легко выявляемым с использованием определенных олигонуклеотидных зондов, или посредством делеции дефектной или неожиданной гибридизации в аллели при локусе, отличном от нормального хромосомного локуса для данной полинуклеотидной последовательности.
"Измененные" полипептиды могут содержать делеции, инсерции или замены аминокислотных остатков, которые вызывают молчащее изменение и приводят к получению функционально эквивалентного полипептида. Умышленные замены аминокислот можно осуществить на основании аналогии полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков, пока сохраняется биологическая или иммунологическая активность кодируемого полипептида. Например, отрицательно заряженные аминокислоты могут включать аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты могут включать лизин и аргинин; а аминокислоты с незаряженными полярными головными группами, имеющие одинаковые величины гидрофильности, могут включать лейцин, изолейцин и валин, глицин и аланин, аспарагин и глутамин, серин и треонин и фенилаланин и тирозин.
"Вариант" полипептида или аминокислотный "вариант" представляет собой аминокислотную последовательность, которая изменена одной или несколькими аминокислотами из определенной аминокислотной последовательности. Вариант полипептида может иметь консервативные изменения, при которых замещающая аминокислота имеет структурные или химические свойства, подобные замещенной аминокислоте, например, замещение лейцина изолейцином. Вариант может также иметь неконсервативные изменения, при которых замещающая аминокислота имеет физические свойства, отличные от свойств замещенной аминокислоты, например, замещение глицина триптофаном. Аналогичные небольшие изменения могут также включать делеции или инсерции аминокислот или оба эти изменения. Предпочтительно аминокислотные варианты сохраняют определенные структурные или функциональные характеристики определенного полипептида. Руководство в определении, какие аминокислотные остатки могут быть заменены, инсертированы или подвергнуты делеции, можно получить, например, с использованием компьютерных программ, хорошо известных в данной области, таких как программное обеспечение LASERGENE (DNASTAR Inc., MADISON, WI).
Вариант полинуклеотида представляет собой вариант определенной полинуклеотидной последовательности, полинуклеотидная последовательностиь которого предпочтительно на по меньшей мере примерно 80%, более предпочтительно на по меньшей мере примерно 90%, и, наиболее предпочтительно на по меньшей мере примерно 95% аналогична определенной полинуклеотидной последовательности. Специалистам в данной области понятно, что в результате вырожденности генетического кода может быть продуцировано множество вариантных полинуклеотидных последовательностей, кодирующих определенный белок, где некоторые несут минимальную гомологию с полинуклеотидными последовательностями любого известного и природного гена. Таким образом, изобретение предусматривает каждое и любое возможное изменение полинуклеотидной последовательности, которое может быть произведено выбором комбинаций на основе возможных выборов кодона. Эти комбинации производятся в соответствии со стандартным триплетным генетическим кодом кодона, и все такие изменения также следует рассматривать как являющиеся определенно раскрытыми.
"Делеция" представляет собой изменение аминокислотной или нуклеотидной последовательности, которое приводит к отсутствию одного или нескольких аминокислотных остатков или нуклеотидов.
Термины "инсерция" или "добавление" относятся к изменению полипептидной или полинуклеотидной последовательности, приводящему к добавлению соответственно одного или нескольких аминокислотных остатков или нуклеотидов по сравнению с природной молекулой.
Используемый в настоящем описании термин "функциональный эквивалент" относится к полипептиду или полинуклеотиду, который обладает по меньшей мере одной функциональной и/или структурной характеристикой определенного полипептида или полинуклеотида. Функциональный эквивалент может содержать модификации, которые обеспечивают возможность выполнения определенной функции. Термин "функциональный эквивалент" предназначен для включения фрагментов, мутантов, гибридов, вариантов, аналогов или химических производных молекулы.
Термин "микронабор" относится к любому расположению нуклеиновых кислот, аминокислот, антител и т.д. на субстрате. Субстрат может представлять собой любую подходящую подложку, например шарики, стекло, бумагу, микроцеллюлозу, нейлон или любую соответствующую мембрану и т.д. Субстрат может представлять собой любую жесткую или полужесткую подложку, включая, но не ограничиваясь этим, мембраны, фильтры, тонкие диски, стружки, покровные стекла, волокна, шарики, включая магнитные или немагнитные шарики, гели, трубки, пластины, полимеры, микрочастицы, капилляры и т.д. Субстрат может обеспечить поверхность для покрытия и/или может иметь множество форм поверхности, такие как лунки, штифты, бороздки, каналы и поры, с которыми могут связываться нуклеиновые кислоты, аминокислоты и т.д.
Термин "образец" используется в настоящем описании в его самом широком смысле. Образцы могут быть получены из любого источника, например из биологических жидкостей, выделений, тканей, клеток или клеток в культуре, включая, но не ограничиваясь этим, слюну, кровь, мочу, сыворотку, плазму, стекловидное тело, синовиальную жидкость, спинномозговую жидкость, амниотическую жидкость, и ткань органа (например, ткань, взятую при биопсии); из хромосом, органелл или других мембран, выделенных из клетки; из геномной ДНК, кДНК, РНК, мРНК и т.д.; и из очищенных клеток или тканей или из блотов и импринтингов таких клеток или тканей. Образцы могут быть получены из любого источника, такого как, например, человек или млекопитающее, отличное от человека, и т.д. Предусмотрены также образцы, полученные из любой модели на животных заболевания. Образец может быть в растворе или может быть, например, фиксирован или связан с субстратом. Образец может относиться к любому материалу, подходящему для тестирования для выявления присутствия CTGF или фрагментов CTGF, или подходящему для скрининга молекул, которые связываются с CTGF или с его фрагментами. Способы получения таких образцов находятся в пределах навыков в данной области.
Термин "гибридизация" относится к способу, посредством которого последовательность нуклеиновой кислоты связывается с комплементарной последовательностью посредством спаривания оснований. Условия гибридизации можно определить, например, по концентрации соли или формамида в прегибридизационном и гибридизационном растворах или по температуре гибридизации, и они хорошо известны в данной области. Гибридизация может происходить в условиях различной строгости.
В частности, строгость может увеличиваться при уменьшении концентрации соли, увеличении концентрации формамида или повышении температуры гибридизации. Например, в целях настоящего изобретения гибридизация в очень жестких условиях может происходить в примерно 50% формамиде при температуре примерно от 37°С до 42°С и в менее жестких условиях - в формамиде в концентрации примерно от 35% до 25% при температуре примерно от 30°С до 35°С. В частности, гибридизация в целом происходит в самых жестких условиях при 42°С в 50% формамиде, 5Х SSPE, 0,3% SDS и 200 мкг/мл фрагментированной и денатурированной ДНК спермы лосося.
Температурный диапазон, соответствующий определенному уровню строгости условий, можно дополнительно уменьшить способами, известными в данной области, например, расчетом соотношения между пурином и пиримидином интересующей нуклеиновой кислоты и соответствующей регулировкой температуры. Для удаления неспецифических сигналов блоты можно последовательно промывать, например, при комнатной температуре или выше вплоть до 60°С, во все более жестких условиях вплоть до 0,1Х SSC и 0,5% SDS. Изменения указанного выше диапазона и условий хорошо известны в данной области.
Изобретение
Настоящее изобретение относится к антителам, которые специфически связываются с фактором роста соединительной ткани (CTGF). Антитела представляют собой поликлональные или моноклональные антитела, предпочтительно человеческие моноклональные антитела. Антитела направлены к N-концевому фрагменту CTGF, показанному на фиг.1. Более конкретно антитела направлены к фрагменту CTGF, простирающемуся от примерно остатка 97 до примерно остатка 180 SEQ ID NO:2. В определенных вариантах осуществления изобретения антитела направлены к фрагменту CTGF, простирающемуся от примерно остатка 103 до примерно остатка 164, а конкретнее к фрагменту от примерно остатка 134 до примерно остатка 158 SEQ ID NO:2. Конкретнее антитела направлены к фрагменту CTGF, простирающемуся от примерно остатка 143 до примерно остатка 154 SEQ ID NO:2.
В определенных вариантах осуществления изобретения антитела нейтрализуют биологическую активность CTGF. Виды биологической активности CTGF включают клеточную пролиферацию, дифференциацию, генную экспрессию и т.д. В определенных вариантах осуществления биологическая активность выбрана из группы, состоящей из клеточной дифференциации, например дифференциации или трансдифференциации фибробластов, миофибробластов, эндотелиальных клеток и т.д., из различных клеток-предшественников; индукции экспрессии белка, участвующего в формировании и перестройке внеклеточного матрикса, например коллагена I типа, фибронектина и т.д.; совместной индукции каскадов передачи сигналов, связанных с различными факторами, включая, но не ограничиваясь этим, TGF-β, IGF, VEGF, ангиотензин II, эндотелин и т.д.; и клеточной реакции на различные стимулы окружающей среды, включая, но не ограничиваясь этим, увеличенное содержание глюкозы (гипергликемия), увеличенное механическое напряжение (гипертония) и т.д.
Хотя изобретение не должно ограничиваться механизмом, которым антитела нейтрализуют активность CTGF, антитела могут связываться с CTGF и предотвращать его взаимодействие со специфическими клеточными рецепторами. Рецепторы могут иметь высокую аффинность связывания с CTGF и путем связывания с CTGF стимулируют внутриклеточный сигнал, который ведет к пролиферации, дифференциации, индукции экспрессии генов и/или изменению клеточной морфологии или функции. Определенная биологическая реакция клетки на CTGF зависит от клетки и текущего состояния окружающей микросреды. Альтернативно рецепторы могут иметь низкую аффинность связывания с CTGF и, путем связывания с CTGF, могут, например, располагать CTGF относительно рецепторов с высокой аффинностью для содействия распознавания CTGF и реакции на него. Альтернативно антитела могут связывать CTGF внутри тканей или органов и содействовать титрованию или удалению CTGF из организма.
Альтернативно или в сочетании с описанными выше механизмами антитела могут связываться с CTGF и предотвращать его взаимодействие с секретируемыми или связанными с мембранами кофакторами. Такие кофакторы, в частности, включают члены суперсемейства TGFβ, включая, например, TGFβ-1, -2 и -3; активин-А, -В, -С и -Е; BMP-2, -3, -4, -5, -6, -7, -8a, -8b, -9, -10, -11 и -15; и GDF-3, -5, -6, -7, -9 и -10. Например, было показано, что CTGF связывается с TGFβ-1 и BMP-4 и модулирует их активность (Abreu et al. (2002) Nat Cell Biol 4:599-604). Настоящее изобретение предоставляет доказательство того, что область CTGF, которая связывается с TGFβ, кодируется экзоном 3 (фиг.1В; нуклеотиды 418-669 SEQ ID NO:1), и антитела, которые связываются в пределах этой области, предотвращают взаимодействие между CTGF и TGFβ (см. пример 12 ниже). Кроме того, было показано, что антитела, которые связываются в пределах этой области CTGF, нейтрализуют специфические связанные с CTGF процессы в случае моделей на животных. Например, было показано, что антитела, которые связываются в пределах этой области CTGF, специфически ингибируют клеточную миграцию в анализах ex vivo и уменьшают фиброз в случае моделей на животных. Иллюстративными антителами по изобретению являются mAb1 и CLN1; CLN1 продуцируется линией клеток, обозначенной номером доступа РТА-6006 в АТСС, депонированном в Американской коллекции типовых культур (Manassus VA) 20 мая 2004 г.
Независимо от механизма действия настоящее изобретение относится к способам применения антител для лечения различных заболеваний и расстройств, связанных с CTGF, включая, но не ограничиваются этим, нейропатии, пневмосклерозы, ретинопатии, склеродермию, печеночные фиброзы, сердечную недостаточность, артрит и атеросклероз. Кроме того, расстройства, связанные с CTGF, возникают вследствие различных факторов, включая, но не ограничиваясь этим, гипергликемию, гипертонию, сахарный диабет, ожирение и т.д.; и включают диабетическую нефропатию, ретинопатию, сердечно-сосудистые заболевания и им подобные. Поскольку избыточная экспрессия CTGF имеется при широком разнообразии заболеваний, включая перечисленные выше, изобретение предусматривает лечение пациентов, имеющих расстройство, связанное с CTGF, антителом против CTGF для улучшения или стабилизации патологии, сохранения или восстановления функции органов, улучшения качества жизни и удлинения выживаемости.
Например, антитела, в частности, направлены на области CTGF, участвующие в видах биологической активности, связанных как с фибротическим, так и с нефибротическим аспектами различных расстройств, включая, например, интерстициальный пневмосклероз, диабетическую нефропатию и ретинопатию, дегенерацию желтого пятна и т.д. Изобретение также относится к способам применения антител для лечения расстройств, связанных с CTGF, включающих локализованные и системные фибротические расстройства, такие как расстройства легких, печени, сердца, кожи и почек и т.д.; и локализованное формирование рубца вследствие, например, травмы хирургических процедур и т.д.
Антитела по изобретению могут также использоваться в любом способе, который включает связывание с CTGF. Такие способы включают очистку CTGF или фрагментов CTGF, например, аффинной хроматографией; выявление CTGF или фрагментов CTGF в образце, например, с использованием ELISA или иммуногистохимических методик; диагностику расстройства, связанного с CTGF, путем использования способа выявления CTGF для измерения уровней CTGF в образце от пациента и сравнения уровня CTGF в образце со стандартом.
Антитела, направленные против CTGF
Была продемонстрирована модуляция количества и/или активности секретируемых клеточных факторов с использованием, например, моноклональных антител, и были разрешены к применению или проходят разработку несколько терапевтических антител (см. например, Abciximab (Reopro; Centocor, Inc., Malvern PA), Infliximab (Remicade; Maini et al. (1998) Arthritis Rheum., 41: 1552-1563; Targan et al. (1997) N. Engl. J. Med., 337: 1029-1035); Basiliximab (Simulect) and Daclizumab (Zenapax) (Bumgardner et al. (2001) Transplantation, 72: 839-845; Kovarik et al. (1999) Transplantation, 68: 1288-1294); и Transtuzumab (Herceptin; Baselga (2001) Ann. Oncol., 12 Suppl., 1: S49-55)). Специалистам в данной области известны и доступны многочисленные способы продукции антител, включая продукцию у животных, растений, грибов и бактерий; синтетическую конструкцию и культуру ex vivo.
Антитела по изобретению можно получать, используя любую методику, которая обеспечивает продукцию молекул антител. Методики для продукцию in vivo и in vitro или моноклональных, или поликлональных антител хорошо известны в данной области (см. например, Pound (1998) Immunochemical Protocols, Humana Press, Totowa NJ; Harlow and Lane (1988) Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York; Goding (1986) Monoclonal Antibodies: Principles and Practice, 2nd Edition, Academic Press; Schook (1987) Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc). Продукция химерных антител также хорошо известна в данной области, как и продукция одноцепочечных антител (см. например, Morrison et al. (1984) Proc. Natl. Acad. USA, 81: 6851-6855; Neuberger et al. (1984) Nature, 312: 604-608; Takeda et al. (1985) Nature, 314: 452-454). Антитела с аналогичной специфичностью, но разного идиотипического состава можно генерировать разнообразными имеющимися средствами, например, "перетасовкой" цепи из случайных комбинационных библиотек иммуноглобулина (см. например, Burton (1991) Proc. Natl. Acad. Sci. USA, 88:11120-11123).
Антитела могут быть также получены индукцией продуцирования in vivo в популяции лимфоцитов или скринингом библиотек или панелей иммуноглобулинов реагентов высокоспецифичного связывания (см. например, Orlandi et al. (1989) Proc. Natl. Acad. USA, 86: 3833-3837; Winter and Milstein (1991) Nature, 349: 293-299). Можно также генерировать фрагменты антител, которые содержат специфические сайты связывания полипептида-мишени. Такие фрагменты антител включают, но не ограничиваются этим, фрагменты F(ab')2, которые могут продуцироваться расщеплением пепсином молекулы антитела, и фрагменты Fab, которые могут генерироваться восстановлением дисульфидных мостиков фрагментов F(ab')2. Альтернативно могут быть сконструированы библиотеки экспрессии Fab для обеспечения возможности быстрой и легкой идентификации фрагментов моноклонального Fab с желательной специфичностью (см. например, Huse et al. (1989) Science, 254:1275-1281).
Моноклональные антитела по изобретению можно также получить, используя способ гибридомы (см. например, Kohler and Milstein (1975, Nature 256:495-497) или методы рекомбинантной ДНК (например, см., Celltech Therapeutics Ltd., европейский патент EP-0120694; Cabilly et al, патент США №4816567 или Mage Lamoyi (1987; In: Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, pp. 79-97).
В способе гибридомы мышь или другое соответствующее животное-хозяин иммунизируется CTGF или его фрагментом подкожным, внутрибрюшинным или внутримышечным путем для того, чтобы выявить лимфоциты, которые продуцируют или способны продуцировать антитела, которые специфически связываются с полипептидом, используемым для иммунизации. Альтернативно животное-хозяин может представлять собой трансгенное млекопитающее, имеющее трансгены, кодирующие гены человеческого иммуноглобулина, и имеющие инактивированные локусы эндогенного иммуноглобулина. Трансгенное млекопитающее реагирует на иммуногены продукцией человеческих антител (см. например, Lonberg et al., WO 93/12227 (1993), патент США №5877397 и Nature, 148: 1547-1553 (1994) и Kucherlapati et al. (1991) WO 91/10791). Альтернативно лимфоциты могут быть иммунизированы in vitro и затем слиты с клетками миеломы с использованием подходящего средства гибридизации, такого как полиэтиленгликоль, для формирования клетки гибридомы (см. например, Goding (1986) Monoclonal Antibodies: Principles and Practice, 2nd Edition, Academic Press, pp.59-103). Альтернативно человеческие соматические клетки, способные продуцировать антитело, в частности В лимфоциты, подходят для слияния с линиями клеток миеломы. Хотя можно использовать В-лимфоциты из биопсий селезенок, небных миндалин или лимфоузлов индивидуума, более легко доступные В-лимфоциты периферической крови являются предпочтительными. Кроме того, В-клетки человека могут быть непосредственно иммортализованы вирусом Эпштейна-Барра (см. например, Cole et al. (1995) Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp.77-96).
Предпочтительными линиями клеток миеломы для использования в продуцирующих гибридому процедурах слияния являются те, которые эффективно сливаются, поддерживают устойчивый высокий уровень экспрессии антитела отобранной продуцирующей антитело клеткой, имеют дефицит ферментов, что делает их неспособными к росту в определенных селективных средах, которые поддерживают рост желательных гибридом и которые сами не продуцируют антитело. Примеры линий клеток миеломы, которые можно использовать для продукции гибридом в настоящем изобретении, включают (P3X63Ag8, P3X63Ag8-653, NS1/1.Ag 4.1, Sp210-Ag14, FO, NSO/U, MPC-11, MPC11-X45-GTG 1.7, S194/5XX0 Bul, все получены у мышей; R210.RCY3, Y3-Ag 1.2.3, IR983F и 4B210, все получены у крыс; и U-266, GM1500-GRG2, LICR-LON-HMY2, UC729-6, все получены улюдей. (см. например, Goding (1986) Monoclonal Antibodies: Principles and Practice, 2nd Edition, Academic Press, pp.65-66 и Campbell (1984) In: Monoclonal Antibody technology: Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 13 (Burden and Von Knippenberg, eds.) Amsterdam, Elseview, pp. 75-83).
Клетки гибридомы высевают и выращивают в подходящей культуральной среде, которая предпочтительно содержит одно или несколько веществ, которые ингибируют рост или выживание неслитых, материнских клеток миеломы. Например, если в материнских клетках миеломы отсутствует фермент гипоксантин-гуанин-фосфорибозил-трансфераза (HGPRT или HPRT), культуральная среда для гибридом обычно будет содержать вещество, такое как гипоксантин, аминоптерин и тимидин (среда НАТ), что предотвращает рост клеток с дефицитом HGPRT.
Культуральную среду, в которой растут клетки гибридомы, анализируют для выявления продукции моноклональных антител, направленных против CTGF или фрагментов CTGF. Предпочтительно специфичность связывания определяется аффинной хроматографией, иммунопреципитацией или анализом связывания in vitro, таким как радиоиммуноанализ (RIA) или ферментный иммуносорбентный анализ (ELISA), или анализом сортировки активируемой флуоресценции клеток (FASC). Моноклональные антитела по изобретению представляют собой антитела, которые связываются с CTGF, и, кроме того, антитела, которые нейтрализуют биологическую активность CTGF, как проиллюстрировано ниже.
Необязательно проводится скрининг антител, продуцированных, например, как описано выше, для выявления антител, которые связываются по существу в N-концевым фрагментом CTGF. В одном варианте осуществления антитела направлены против фрагмента CTGF, простирающегося от примерно остатка 24 до примерно остатка 180 SEQ ID NO:1. В другом варианте осуществления антитела направлены против фрагмента CTGF, простирающегося от примерно остатка 96 до примерно остатка 180 SEQ ID NO:1. В определенном варианте осуществления скрининг выявляет антитела, которые связываются по существу с тем же эпитопом, который распознается антителом mAb1, что определяется, например, с помощью конкурентных анализов описанного выше типа. В другом определенном варианте осуществления скрининг выявляет антитела, которые связываются по существу с тем же эпитопом, который распознается антителом CLN1, что определяется, например, с помощью конкурентных анализов описанного выше типа. Следует учитывать, что "тот же эпитоп" не означает точную аминокислоту или углеводород, с которым связывается эталонное антитело, что можно определить, например, картированием эпитопа с использованием сканированных аланином вариантов CTGF. "Тот же эпитоп" означает домен CTGF, который блокирован связыванием с CTGF нативного эталонного антитела в интактной форме. Конечно, "тот же эпитоп" включает остатки домена CTGF или углеводород, который структурно взаимодействует или связывается с эталонными определяющими комплементарность областями (CDR) mAb1 или CLN1.
В предпочтительном варианте осуществления изобретения моноклональное антитело должно иметь аффинность, которая равна или больше, чем аффинность mAb1, что определяется, например, анализом Скэтчарда Munsin and Pollard (1980, Anal. Biochem., 107:220).
После идентификации клеток гибридомы, которые продуцируют нейтрализующие антитела желательной специфичности и аффинности, клоны обычно субклонируют с помощью процедур органичивающего разведения и выращивают стандартными способами (Goding (1986) Monoclonal Antibodies: Principles and Practice, 2nd Edition, Academic Press, pp.59-104). Подходящие для этой цели культуральные среды включают, например, Модифицированную Дульбекко среду Игла или среду RPMI-1640. Кроме того, клетки гибридомы могут выращиваться in vivo в виде асцитических опухолей у животного.
Моноклональные антитела, секретируемые субклонами, можно выделить из культуральной среды, асцитической жидкости или сыворотки обычными методами очистки иммуноглобулина, такими как, например, белок А-сефароза, хроматография на гидроксиапатите, гель-электрофорез или аффинная хроматография.
ДНК, кодирующая моноклональные антитела по изобретению, легко выделяется и секвенируется с использованием обычных процедур, например использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи антител. После выделения ДНК может быть лигирована в векторы экспрессии или клонирования, которые затем трансфецируются в клетки-хозяева, такие как обезьяньи клетки COS, клетки яичников китайского хомячка (CHO), или клетки миеломы, которые иначе не продуцируют белок иммуноглобулина. Клетки, трансформированные таким образом, культивируются в условиях, подходящих для синтеза моноклональных антител в культуре рекомбинантных клеток-хозяев. Иллюстративная линия клеток представлена в АТСС под номером доступа PTA-6006, депонированная в АТСС 20 мая 2004 г.
ДНК необязательно модифицируется для изменения характера кодированного иммуноглобулина. Варианты иммуноглобулинов хорошо известны. Например, химерные антитела получают замещением кодирующей последовательности для константных доменов тяжелой и легкой цепи от одного вида, например мыши, гомологичными последовательностями от другого вида, например человека (см. например, Boss et al., международная публикация WO 84/03712; Cabilly et al., патент США № 4816567 или Morrison et al. (1984) Proc. Nat. Acad. Sci, 81: 6851). В определенном варианте осуществления гуманизированные формы мышиных антител могут быть получены замещением определяющих комплементарность областей (CDR), т.е. вариабельных доменов, мышиного антитела, каркасным доменом, т.е. константной областью, человеческого антитела (см. например, международную публикацию WO 92/22653). В некоторых вариантах осуществления выбранные мышиные каркасные участки замещаются человеческим реципиентным иммуноглобулином. Кроме того, выбранный домен Fc может представлять собой IgA, IgD, IgE, IgG-1, IgG-2, IgG-3, IgG-4 или IgM. Домен Fc необязательно способен к эффекторным взаимодействиям, таким как связывание комплемента.
Антитела против CTGF настоящего изобретения могут также слиты с частями, которые обеспечивают дополнительные возможности, такие как выявление или цитотоксические эффекты. Слияния иммуноглобулинов настоящего изобретения и цитотоксических частей получают, например, путем лигирования в кодирующую последовательность иммуноглобулина всей или части кодирующей последовательности цитотоксического неиммуноглобулинового полипептида. Такие неиммуноглобулиновые полипептиды включают полипептидные токсины, такие как рицин, дифтерийный токсин или экзотоксин Pseudomonas. Конъюгаты могут также быть получены способами in vitro. Например, иммунотоксины могут быть сконструированы с использованием реакции дисульфидного обмена или путем формирования тиоэфирной связи между иммуноглобулином и полипептидом токсина. Примеры подходящих реагентов для этой цели включают иминотиолат и метил-4-меркаптобутиримидат. Обычно такие неиммуноглобулиновые слитые полипептиды заменяют константными доменами антитела по изобретению. Альтернативно они заменяются вариабельными доменами одного антигенсвязывающего сайта антитела по изобретению.
Замена Fv или CDR антитела, имеющего специфичность к антигену, не являющемуся CTGF, создаст химерное антитело, содержащее один антигенсвязывающий сайт, имеющий специфичность к CTGF, и другой антигенсвязывающий сайт, имеющий специфичность к другому антигену. В таких вариантах осуществления происходит делеция легкой цепи и Fv тяжелой цепи заменяется желательным полипептидом. Эти антитела называются бивалентными или поливалентными в зависимости от количества "плеч" иммуноглобулина, которыми обладает используемый домен Fc; например, IgG будут бивалентными, а IgM будут поливалентными. Помимо указанных выше иммуноглобулинов, антителу также придается мультивалентность путем рекомбинации антител, которые имеют более одной специфичности. Например, антитело в некоторых вариантах осуществления способно связывать CTGF, как уже описано в настоящем описании, но оно также способно связывать второй фактор роста, например TGFβ, VEGF, FGF, другие члены семейства CCN, например CYR61 и им подобные, или цитокин. Иллюстративные антитела, направленные против этих факторов, хорошо известны. Мультиспецифические, мультивалентные антитела получают путем совместной трансформации клетки ДНК, кодирующей тяжелую и легкую цепи обоих антител и часть экспрессированных антител, имеющих желательную структуру, получают иммуноаффинной хроматографией или подобным способом. Альтернативно такие антитела получают из моновалентных антител, которые распознаются in vitro обычным образом.
Моновалентные антитела также получают способами, которые сами по себе являются обычными. Подходящей является рекомбинантная экспрессия легкой цепи и модифицированной тяжелой цепи. Тяжелая цепь является усеченной обычно в любой точке в области Fc с тем, чтобы предотвратить поперечную сшивку тяжелой цепи. Способы in vitro также используются для получения моновалентных антител, например, фрагменты Fab получают ферментным расщеплением интактного антитела.
Диагностика
Антитела настоящего изобретения можно использовать для количественного и качественного выявления CTGF в образце. Образцы могут быть из любого источника, включая кондиционированные среды из клеток, выращенных в культуре; образцы ткани, например биопсии ткани и трансплантатов органов; биологические жидкости, включая кровь, мочу, жидкость пузырьковых высыпаний, спинномозговую жидкость, стекловидное тело и синовиальную жидкость, и т.д. В одном варианте осуществления выявление CTGF используется для диагностики состояния клеток, выращенных в культуре, например, в отношении дифференциации, продукции матрикса и т.д. CTGF оказывает различные аутокринные и паракринные эффекты на культивируемые клетки, и уровень CTGF, связанного с клеточным слоем, или присутствующим в кондиционированных средах, может быть показателем текущего состояния клетки или прогнозировать будущее состояние клетки (см. например, международную публикацию WO 96/38168). В других вариантах осуществления выявление CTGF используется для определения состояния ткани или органа. Например, орган, предназначенный для трансплантата, можно оценить путем измерения уровней CTGF, причем уровень CTGF, экспрессируемый клетками в органе, указывает на относительное здоровье органа и пригодность для трансплантата. Уровни CTGF можно также определять в биопсии ткани для определения состояния органа или стадии и возможного метастатического потенциала рака.
В предпочтительных вариантах осуществления антитела используются для диагностики заболевания или расстройства, связанного с CTGF (см. например, международную публикацию WO 03/024308). В одном из аспектов изобретение относится к антителам для диагностики связанного с CTGF расстройства путем получения образца, выявления и количественной характеристики уровня CTGF в образце, и сравнения уровня CTGF в образце с уровнем стандартного количества CTGF, где увеличенное или уменьшенное количество CTGF в образце указывает на присутствие связанного с CTGF расстройства. Расстройства, связанные с аберрантными (например, повышенными или сниженными) уровнями CTGF, включают, но не ограничиваются этим, расстройства, связанные с измененной экспрессией белков, связанных с внеклеточного матрикса. Такие расстройства включают, например, различные виды рака, такие как рак молочной железы, поджелудочной железы и желудочно-кишечного тракта; атеросклероз, артрит, ретинопатии, такие как диабетическая ретинопатия; нефропатии, такие как диабетическая нефропатия; фиброз сердца, легкого, печени и почки, и заболевания, связанные с хроническим воспалением и/или инфекцией. Расстройства, связанные с CTGF, также связаны с такими состояниями, как инфаркт миокарда, сахарный диабет, перитонеальный диализ, хроническое и острое отторжение трансплантата, химиотерапия, лучевая терапия и хирургическая операция.
В другом аспекте изобретение относится к антителам для идентификации того, имеет или нет индивидуум предрасположенность к развитию расстройства, связанного с CTGF. На предрасположенность может первоначально указывать гипергликемия, гипертония или ожирение у субъекта. Кроме того, предрасположенность может быть заподозрена вследствие эпизода, например, инфаркта миокарда, хирургической операции, ортопедической или паралитической иммобилизации, застойной сердечной недостаточности, беременности или варикозного расширения вен у субъекта.
В другом аспекте изобретение относится к антителам для мониторинга прогрессирования связанного с CTGF расстройства или мониторинга терапевтической эффективности лечения связанного с CTGF расстройства. Например, способ использования антител может включать получение образцов у субъекта в течение времени; выявление и количественное определение уровня CTGF в таком образце; и сравнение уровня CTGF в последующих образцах с уровнями CTGF в более ранних или предыдущих образцах. Изменение уровня CTGF между образцами в течение времени указывает на прогрессирование расстройства, связанного с CTGF, или терапевтическую эффективность лечения расстройства, связанного с CTGF.
В случае применения для диагностики видов применения антитела по изобретению обычно метят выявляемой составной частью. Выявляемая составная часть может представлять собой любую часть, способную продуцировать, прямо или косвенно, выявляемый сигнал. Например, выявляемая часть может представлять собой радиоизотоп, такой как 3Н, 14С, 32Р, 35S или 125I, флуоресцентное или хемилюминесцентное соединение, такое как изоцианат флюоресцеина, родамин или люцифирин; или фермент, такой как щелочная фосфатаза, бета-галактозидаза или пероксидаза хрена.
Можно использовать любой способ, известный в данной области, для отдельного конъюгирования антитела с выявляемой составной частью (см. например, Hunter et al. (1962) Nature, 144: 945; David et al. (1974) Biochemistry, 13: 1014; Pain et al. (1981) J. Immunol. Meth., 40: 219 и Nygren (1982) J. Histochem. Cytochem., 30: 407). Антитела по настоящему изобретению можно использовать в любом известном способе анализа, таком как анализы конкурентного связывания, прямой и непрямой сэндвич-анализы и анализы иммунопреципитации (Zola (1987) In: Monoclonal Antibodies: A Manual of Techniques, CRC Press, Inc., pp.147-158).
Анализы конкурентного связывания основаны на способности меченого стандарта (который может представлять собой CTGF или его иммунологически реактивную часть) конкурировать с анализируемым веществом тестируемого образца (CTGF) за связывания с ограниченным количеством антитела. Количество CTGF в тестируемом образце обратно пропорционально количеству стандарта, которое оказывается связанным с антителами. Чтобы облегчить определение количества стандарта, который связывается, антитела в целом становятся нерастворимыми перед или после конкуренции, так что стандарт и анализируемое вещество, которые связаны с антителами, могут быть удобно отделены от стандарта и анализируемого вещества, которые остаются несвязанными.
Сэндвич-анализы включают использование двух антител, каждое из которых способно связываться с другой иммуногенной частью или эпитопом подлежащего выявлению белка. В сэндвич-анализе анализируемое вещество в тестируемом образце связывается с первым антителом, которое иммобилизовано на твердой подложке, и затем второе антитело связывается с анализируемым веществом, формируя, таким образом, нерастворимый комплекс, состоящий из трех частей (David and Greene, патент США № 4376110). Второе антитело само может быть меченным выявляемой составной частью (прямые сэндвич-анализы) или может быть измерено с использованием антитела против иммуноглобулина, которое мечено выявляемой частью (непрямой сэндвич-анализ). Например, один тип сэндвич-анализа представляет собой анализ ELISA, и в этом случае выявляемая часть представляет собой фермент. Иллюстративный анализ, в котором можно использовать антитела по изобретению, например, описан в международной публикации WO 03/024308.
Антитела по изобретению можно также использовать для визуализации in vivo, при которой антитело, меченное выявляемой частью, такой как рентгеноконтрастное вещество, радиоизотоп или флуоресцентная часть, такая как зеленый флуоресцентный белок (GFP), вводится хозяину предпочтительно в поток крови и анализируется присутствие и локализация меченого антитела у хозяина. Эту методику визуализации можно использовать при определении стадии и лечении связанных с CTGF расстройств, таких как фибротические расстройства. Антитело может быть меченным любой частью, которую можно выявить у хозяина, или ядерным магнитным резонансом, рентгенологическим или другими средствами выявления, известными в данной области.
Терапевтические средства
Настоящее изобретение относится к антителам для лечения различных болезней и расстройств, связанных с CTGF. Было обнаружено, что антитела по изобретению уменьшают вредные эффекты продукции или активности CTGF при нескольких расстройствах, как проиллюстрировано ниже. Кроме того, антитела проявляют благоприятную фармакокинетику, что делает их превосходными терапевтическими средствами для лечения расстройств, связанных с CTGF.
Антитела против CTGF настоящего изобретения ингибируют развитие фиброза в случае моделей на животных, например, пневмосклероза и фиброза почек. В частности, антитела ослабляют вызванный блеомицином пневмосклероз у мышей на 60-70%, что определяется по ингибированию легочного накопления гидроксипролина (коллагена) и путем гистологического исследования препаратов ткани. Кроме того, антитела снижают накопление коллагена в модели остатка почки крыс (т.е. после нефрэктомии 5/6) и у мышей после односторонней обструкции мочеточника (UUO). Антитела также уменьшают фиброз, вызванный комбинированным подкожным или внутрибрюшинным вливанием CTGF и TGFβ у новорожденных мышей. Кроме того, антитела уменьшают осложнения, связанные с недостаточностью органа, например улучшают почечную функцию в случае различных моделей хронической и острой почечной недостаточности. При использовании этих антител у животных не наблюдали токсичности. Поскольку CTGF избыточно экспрессируется при широком разнообразии фибротических заболеваний, включая диффузную и ограниченную склеродермию, остеоартрит, диабетическую нефропатию и ретинопатию и т.д., изобретение предусматривает лечение пациентов, имеющих расстройство, связанное с CTGF, антителом против CTGF для улучшения или стабилизации течения патологии, восстановления функции органов, улучшения качества жизни и продления выживаемости.
Поэтому антитела по изобретению особенно полезны в случае терапевтического применения для предотвращения или лечения расстройств, связанных с CTGF, у субъекта. Такие расстройства включают, но не ограничиваются этим, ангиогенез и другие процессы, которые играют центральную роль при таких состояниях, как атеросклероз, глаукома и т.д.; и при раке, включая острый лимфобластический лейкоз, дерматофибромы, рак молочной железы, карциному молочной железы, глиому и глиобластому, рабдомиосаркому и фибросаркому, десмоплазию, ангиолипому, ангиолейомиому, десмопластические формы рака и рак предстательной железы, яичников, толстой и прямой кишки, поджелудочной железы, желудочно-кишечного тракта и печени и другой опухолевой рост и метастазы.
Кроме того, антитела по изобретению можно использовать для терапевтического применения для предотвращения или лечения связанных с CTGF расстройств, включающих фиброз. В одном из аспектов антитела по изобретению вводятся субъекту для предотвращения или лечения связанного с CTGF расстройства, включая, но не ограничиваясь этим, расстройства, проявляющие измененную экспрессию и отложение белков, связанных с внеклеточного матрикса, например фибротические расстройства. В различных аспектах фиброз может быть локализован в определенной ткани, такой как эпителиальная, эндотелиальная или соединительная ткань; или в органе, таком как почка, легкое или печень. Фиброз может также возникнуть в глазу и суставах. В других аспектах фиброз может быть системным и вовлекать множественные органные и тканевые системы. Связанные с CTGF расстройства включают, например, атеросклероз, артрит, ретинопатии, такие как диабетическая ретинопатия, нефропатии, такие как диабетическая нефропатия; сердечный, легочный, печеночный и почечный фиброз, и заболевания, связанные с хроническим воспалением и/или инфекцией.
В другом аспекте изобретение относится к антителам для предотвращения связанного с CTGF расстройства у субъекта, имеющего предрасположенность к развитию такого расстройства. Предрасположенность может включать, например, гипергликемию, гипертонию или ожирение у субъекта. Такие расстройства могут возникать, например, вследствие сахарного диабета, ожирения и т.д. и включают диабетическую нефропатию, ретинопатию и сердечно-сосудистые заболевания. Кроме того, предрасположенность может быть заподозрена вследствие явления, например, инфаркта миокарда, хирургической операции, перитонеального диализа, хронического или острого отторжения трансплантата, химиотерапии, лучевой терапии, травмы, ортопедической или паралитической иммобилизации, застойной сердечной недостаточности, беременности или варикозного расширения вен у субъекта.
В конкретных вариантах осуществления, как проиллюстрировано в настоящем описании, антитела по настоящему изобретению вводятся субъекту для лечения фиброза органа, например легкого или почки. В настоящем описании показано, что антитела приводят к благоприятному действию в случае различных моделей пневмосклероза и почечного фиброза (см. например, примеры 7-9). В другом конкретном варианте осуществления антитела по настоящему изобретению вводятся субъекту для уменьшения локального или системного склероза (см. например, примеры 11 и 12). В дополнительных вариантах осуществления антитела вводятся субъекту для лечения или профилактики глазных расстройств, таких как пролиферативная витреоретинопатия, диабетическая ретинопатия, дегенерация желтого пятна и т.д. Поскольку CTGF вовлечен в широкий спектр расстройств, изобретение, кроме того, предусматривает лечение пациентов, имеющих связанное с CTGF расстройство, с использованием антитела по изобретению для улучшения или стабилизации патологического процесса и функции органа, улучшения качества жизни и продления выживаемости.
При терапевтическом применении антитела по изобретению вводятся млекопитающему, предпочтительно человеку, в фармацевтически приемлемой лекарственной форме. Антитела можно вводить внутривенно в виде болюса или путем непрерывного вливания в течение периода времени и/или внутримышечным, подкожным, внутрисуставным, внутрисиновиальным, подоболочечным, интравитреальным (в стекловидное тело), внутричерепным, пероральным, местным или ингаляционным путями. Когда антитело обладает подходящей активностью, можно также использовать пути введения в опухоль, вокруг опухоли, внутрь поражения или около поражения для оказания местного, а также системного терапевтических эффектов.
Такие лекарственные формы включают фармацевтически приемлемые носители, которые являются по существу нетоксичными и нетерапевтическими. Примеры таких носителей включают ионообменные вещества, окись алюминия, стеарат алюминия, лецитин; сывороточные белки, такие как альбумин сыворотки человека; буферы, такие как фосфат или глицин; сорбиновую кислоту, сорбат калия, частичные глицеридовые смеси насыщенных растительных жирных кислот, воду, соли; или электролиты, такие как протаминсульфат, хлорид натрия, соли металлов, коллоидный оксид кремния, трисиликат магния, поливинилпирролидон, целлюлозные полимеры и полиэтиленгликоль. Носители для местных или основанных на геле форм антитела включают полисахариды, такие как карбоксиметилцеллюлоза или метилцеллюлоза натрия, поливинилпирролидон, полиакрилаты, блок-сополимеры полиоксиэтилена-полиоксипропилена, полиэтиленгликоль и спирты воска дерева. Обычные депо-формы включают, например, микрокапсулы, нано-капсулы, липосомы, пластыри, сублингвальные таблетки и полимерные матрицы, такие как сополимеры полилактида:полигликолида. В случае присутствия в водной лекарственной форме, а не в лиофилизированном виде, антитело обычно содержится в композиции в концентрации примерно от 0,1 мг/мл до 100 мг/мл, хотя допускается колебание далеко за пределы этих границ.
Для предотвращения или лечения заболевания соответствующая дозировка антитела будет зависеть от типа подлежащего лечению заболевания, как определено выше, тяжести и течения заболевания, от того, вводится ли антитело в профилактических или лечебных целях, от курса предыдущей терапии, клинического анамнеза пациента и реакции на антитело и от суждения лечащего врача. Антитело можно вводить пациенту один раз или в ходе серии лечебных циклов.
В зависимости от типа и тяжести заболевания примерно от 0,015-15 мг антитела/кг массы пациента является начальной возможной дозировкой для введения пациенту, например, путем одного или нескольких отдельных введений или непрерывного вливания. При повторных введениях в течение нескольких дней или более в зависимости от состояния лечение повторяют до тех пор, пока не произойдет желательное подавление симптомов заболевания. Однако можно использовать другие схемы дозировки, и они не исключены из настоящего изобретения.
В соответствии с другим вариантом осуществления эффективность антитела при предотвращении или лечении заболевания может быть повышена введением антитела серийно или в комбинации с другим средством, которое эффективно для той же клинической цели, таким как другое антитело, направленное против эпитопа, отличного от эпитопа, против которого направлено основное антитело, или с одним или несколькими обычными терапевтическими средствами, которые, как известно, применяются по предполагаемому показанию, например для предотвращения или лечения состояний, связанных с избыточной продукцией внеклеточного матрикса, таких как фиброз или склероз, ингибирования роста или метастазирования опухолевых клеток, ингибирования неоваскуляризации или уменьшения воспаления. Такие средства могут облегчить симптомы или улучшить исход посредством аналогичного механизма действия, например антитела против TGFβ, или другого механизма, например интерферон-γ. Такие средства могут дополнительно облегчить симптомы, прямо или косвенно связанные с расстройством, связанным с CTGF, или предрасположенностью к развитию расстройства, связанного с CTGF, например, ингибиторы ангиотензин-превращающего фермента (АСЕ) и блокаторы ангиотензиновых рецепторов (Arbs).
Например, больные склеродермией, получающие вливания устойчивого агониста простациклина илопроста, часто отмечают уменьшение уплотнения кожи, что согласуется с ингибирующим эффектом на формирование рубцовой ткани фибробластами кожи. Было показано, что простаноиды оказывают ингибирующий эффект на синтез коллагена, и ряд доказательств демонстрирует, что илопрост блокирует индукцию CTGF при склеродермии (Korn et al (1980) J. Clin. Invest., 65: 543-554; Goldstein and Polger (1982) J. Biol. Chem., 257: 8630-8633 и Stratton et al. (2001) J. Clin. Invest., 108: 241-250). Уровень CTGF возрастает в 7 раз в жидкости пузырьковых высыпаний у пациентов со склеродермией по сравнению со здоровыми контрольными лицами, однако у пациентов, получающих внутривенное введение илопроста, проявляется выраженное снижение уровня CTGF в жидкости пузырьковых высыпаний (Stratton et al. (2001) J. Clin. Invest., 108:241-250). Взятые вместе, эти результаты свидетельствуют о том, что некоторые из положительных эффектов терапии илопростом при склеродермии могут быть получены в результате антифибротических эффектов, опосредованных через снижение уровней CTGF. Поскольку имеются опасения относительно применения активного сосудорасширяющего и антитромбоцитарного аналога простациклина при хроническом системном введении пациентам со склеродермией, лечение с использованием одного антитела против CTGF или в сочетании со сниженными уровнями илопроста может обеспечить безопасное и эффективное лечение в случае склеродермии.
Дополнительные способы применения
Антитела по изобретению можно также использовать в качестве средств аффинной очистки. При этом способе антитела против CTGF иммобилизуются на подходящей подложке, такой как смола Sephadex или фильтровальная бумага, с использованием способов, хорошо известных в данной области. Затем иммобилизованное антитело контактирует с образцом, содержащим подлежащий очистке CTGF, а затем подложка промывается подходящим растворителем, который удаляет по существу весь материал в образце, кроме CTGF, который связан с иммобилизованным антителом. Наконец, подложка промывается другим подходящим растворителем, таким как глициновый буфер (рН 5,0), который освобождает CTGF от антитела.
ПРИМЕРЫ
Изобретение будет далее разъяснено при ссылке на следующие примеры, которые предназначены лишь для иллюстрации изобретения. Эти примеры представлены только для иллюстрации заявленного изобретения. Объем настоящего изобретения не ограничивается проиллюстрированными вариантами его осуществления, которые предназначены только для иллюстраций одиночных аспектов. Любые способы, которые являются функционально эквивалентными, находятся в пределах объема изобретения. Различные модификации изобретения в дополнение к модификациям, описанным в настоящем описании, станут очевидными для специалистов в данной области из предшествующего описания и сопровождающих чертежей. Такие модификации предназначены для включения в объем прилагаемой формулы изобретения.
Пример 1. Продукция рекомбинантного человеческого CTGF
Бакуловирусный конструкт рекомбинантного человеческого CTGF продуцировали, как описано в публикации Segarini et al. (2001) J. Biol. Chem., 276:40659-40667). Вкратце, кДНК CTGF, содержащую только открытую рамку считывания, генерировали PCR с использованием DB60R32 (Bradham et al. (1991) J. Cell Biol., 114:1285-94) в качестве матрицы и затравок 5'-gctccgcccgcagtgggatccATGaccgccgcc-3' и 5'-ggatccggatccTCAtgccatgtctccgta-3', которые добавляли сайты рестрикционного фермента BamHI к концам амплифицированного продукта. Нативные инициирующие и терминирующие кодоны указаны заглавными буквами.
Полученный амплифицированный фрагмент ДНК расщепляли BamHI, выделяли электрофорезом на агарозном геле и субклонировали непосредственно в сайт BamHI экспрессионной плазмиды PFASTBAC1 бакуловируса (Invitrogen Corp., Carlsbad CA). Последовательность и ориентацию экспрессионной кассеты подтверждали секвенированием ДНК. Затем полученную экспрессионную кассету CTGF переносили в бакмидную ДНК путем сайт-специфической рекомбинацией у бактерий. Затем эту бакмиду использовали для генерирования полностью рекомбинантного бакуловируса CTGF в клетках SF9 насекомого Spodoptera frugiperda в соответствии с протоколами, поставляемыми изготовителем (руководство по Экспрессионной системе BAC-TO-BAC, Invitrogen). Экспансию титров рекомбинантного бакуловируса в клетках Sf9 насекомых выполняли с использованием стандартной процедуры, известной в данной области.
Клетки Hi5 насекомых адаптировали для роста в суспензии путем нескольких пассажей клеток в культуре во стряхиваемой колбе, сопровождаемых обогащением при каждом пассаже для отделенных клеток. Суспензию клеток Hi5 культивировали в 1л среды SF900II SFM (Invitrogen) с добавлением 20 мкг/мл гентамицина (Mediatech, Inc., Herndon VA) и 1х липида (Invitrogen) в одноразовых культуральных колбах Fernbach емкостью 2,8 л (Corning Inc., Acton MA) на платформе шейкера при 110 об/мин при 27°С. Когда клетки достигали плотности 1,0-1,5×106 клеток/мл при жизнеспособности >95%, их инфицировали рекомбинантным бакуловирусом при множественности заражения (MOI) 10. Затем культуры инкубировали при 27°С в течение еще 40-44 ч. Кондиционированную среду, которая содержит rhCTGF, собирали, охлаждали на льду и центрифугировали при 5000 g. Затем супернатант пропускали через фильтр с размером пор 0,45 мм.
Альтернативно рекомбинантный крысиный CTGF продуцировали инсерцией клона 2-4-7, который кодирует крысиный CTGF (Schmidt et al., патент США № 6348329), в экспрессионный вектор pMK33 (сконструированный Michael Koelle, Stanford University Ph.D. dissertation, 1992). Экспрессионный конструкт крысиного CTGF трансфецировали в клетки Schneider 2 (Американская коллекция типовых культур, Manassas VA; Schneider (1972) J Embryol Exp Morphol 27:353-365) с использованием реагента CELLFECTIN (Invitrogen Corp., Carlsbad CA). Клетки выращивали в среде, содержащей 300 мкг/мл гидромицина В, в течение 6 недель, и затем выращивали без отбора в течение 3-х дней. Экспрессию CTGF индуцировали добавлением 500 мкМ CuSO4 и 100 мкМ ZnSO4 и через 4 дня среду собирали и просветляли центрифугированием и фильтрацией, как указано выше.
CTGF, продуцированный любым способом, описанным выше, очищали следующим образом. 4 л кондиционированной среды загружали на гепариновую колонку емкостью 5 мл HI-TRAP (Amersham Biosciences Corp., Piscataway NJ), предварительно уравновешенную 50 мМ Tris (рН 7,5), 150 мМ NaCl. Колонку промывали 10 объемам колонки 350 мМ NaCl, 50 мМ Tris (рН 7,5). CTGF элюировали из колонки с увеличением градиента соли NaCl. Проводили скрининг элюированных фракций с помощью SDS-PAGE и объединяли фракции, содержащие CTGF.
Очищенный гепарином CTGF разбавляли до конечной проводимости 5,7 мС апирогенной дважды дистиллированной водой и рН доводили до 8,0. Сильную анионообменную колонку Q-SEPHAROSE (Amersham Biosciences), содержащую приблизительно 23 мл смолы, соединенную тандемом с карбометиловой полистироловой колонкой (CM) POROS (Applied Biosystems), содержащей приблизительно 7 мл смолы, использовали для удаления эндотоксина и захвата и элюирования очищенного rhCTGF. Перед загрузкой образца тандемную колонку промывали 0,5 М NaOH с последующим промыванием 0,1 М NaOH и, наконец, уравновешивающим буфером. Загруженный образец пропускали через тандемную колонку, колонку Q-Sepharose удаляли, и CTGF элюировали из колонки CM POROS (Applied Biosystems) с увеличением градиента от 350 мМ до 1200 мМ NaCl. Чистоту элюированных фракций, содержащих CTGF, оценивали с помощью анализа SDS-PAGE перед формированием конечного пула образца.
Пример 2. Продукция N-концевого и С-концевого фрагмента CTGF
N-концевые и С-концевые фрагменты CTGF получали следующим образом. Рекомбинантный человеческий CTGF, полученный и очищенный, как описано выше, расщепляли при комнатной температуре в течение 6 ч путем обработки шариками химотрипсина (Sigma Chemical Co., St.Louis, MO) при 1,5 мг CTGF на единицу химотрипсина. Смесь центрифугировали, шарики химотрипсина удаляли и супернатант, содержащий ферментативно расщепленный CTGF, разводили 1:5 50 мМ Tris, pH 7,5. Разбавленный супернатант наносили на гепариновую колонку Hi-Trap. Собирали элюирующиеся содержащие N-концевые фрагменты CTGF. Гепариновую колонку промывали 350 мМ NaCl и связанные С-концевые фрагменты CTGF элюировали с линейным градиентом от 350 мМ до 1200 мМ NaCl, как описано выше. Фракции анализировали SDS-Page, и объединяли фракции, содержащие С-концевые фрагменты CTGF.
Элюент с гепариновой колонки, который содержал N-концевые фрагменты CTGF, доводили до 0,5 М сульфата аммония/50 мМ Tris при рН 7,5 и затем загружали на колонку фенилсефарозы НР на 15 мл (Amersham-Pharmacia), которая была предварительно уравновешена 0,5 М сульфата аммония/50 мМ Tris при рН 7,5. Колонку промывали 15 колоночными объемами 0,5 М сульфата аммония/50 мМ Tris при рН 7,5, и связанные N-концевые фрагменты CTGF элюировали с линейным градиентом от 0,5 до 0 М сульфата аммония/50 мМ Tris при рН 7,5 приблизительно 15 объемами колонки. Фракции анализировали SDS-PAGE и объединяли фракции, содержащие N-концевые фрагменты CTGF. Объединенный раствор концентрировали и буфер обменивали с 50 мМ Tris, 400 мМ NaCl (рН 7,2), используя ультрафильтрационную мембрану ULTRACEL AMICON YM10 (Millipore Corp., Bedford MA).
Пример 3. Продукция человеческих моноклональных антител против CTGF
Полностью человеческие моноклональные антитела к человеческому CTGF получали, используя линии HCo7, HCo12 и HCo7+HCo12 мышей HUMAB (Medarex, Inc., Princeton NJ). Мышей иммунизировали вплоть до 10 внутрибрюшинными (IP) или подкожными (Sc) инъекциями 25-50 мг рекомбинантного человеческого CTGF в полном адъюванте Фрейнда в течение периода 2-4 недель. Иммунный ответ контролировали позадиглазничными кровоизлияниями. Скрининг плазмы проводили с помощью ELISA (как описано ниже), и мышей с достаточными титрами иммуноглобулина против CTGF использовали для гибридизаций. Мышам внутривенно повторно проводили иммунизацию антигеном за 3 и 2 дня до умерщвления и удаления селезенки.
Суспензии одиночных клеток селезеночных лимфоцитов от иммунизированных мышей подвергали гибридизации с одной четвертой количества не секретирующих клеток миеломы мышей P3X63-Ag8.653 (Американская коллекция типовых культур (АТСС), Manassas VA) в 50% PEG (Sigma, St.Louis MO). Клетки высевали в концентрации приблизительно 1 × 105 клеток/лунку в титровочный микропланшет с плоским дном и инкубировали в течение примерно 2 недель в среде DMEM (модифицированной Дульбекко среде Игла) с высоким содержанием глюкозы (Mediatech, Hemdon VA), содержащей L-глутамин и пируват натрия, 10% фетальную телячью сыворотку, 10% кондиционированную среду P388D1 (ATCC), 3-5% ориген (Igen International, Gaithersburg MD), 5 мМ HEPES, 0,055 мМ 2-меркаптоэтанола, 50 мг/мл гентамицина и 1х НАТ (Sigma). Через 1-2 недель клетки культивировали в среде, в которой НАТ был замещен НТ. Затем проводили скрининг отдельных ячеек ELISA (описанный ниже). Секретирующие антитела гибридомы повторно высевали на планшеты, снова проводили скрининг и, если они еще были положительными по антитела против CTGF, субклонировали по меньшей мере дважды при ограничивающем разведении. Затем устойчивые субклоны культивировали in vitro для генерирования небольших количеств антитела в среде для культуры ткани, чтобы охарактеризовать. Один клон от каждой гибридомы, которая сохраняла реактивность материнских клеток, использовали для генерирования банков клеток в 5-10 флакончиках, хранившихся в жидком азоте.
Анализы ELISA выполняли, как описано Fishwild et al. (1996, Nature Biotech., 14:845-851). Вкратце, титровочные микропланшеты покрывали 1-2 мкг/мл очищенного рекомбинантного CTGF в PBS (сыворотке с фосфатным буфером) в концентрации 50 мкл/лунку, инкубировали при 4°С в течение ночи, затем блокировали 200 мкл/лунку 5% куриной сывороткой в PBS/Tween (0,05%). Разведения плазмы от мышей, иммунизированных CTGF, или супернатанты культуры гибридомы добавляли в каждую лунку и инкубировали 1-2 ч при окружающей температуре. Планшеты промывали PBS/Tween и затем инкубировали с козьим поликлональным антителом Fc против человеческого IgG, конъюгированным с пероксидазой хрена (HRP) в течение 1 ч при комнатной температуре. После промывания планшеты проявляли 0,22 мг/мл субстрата ABTS (Sigma) и анализировали с помощью спектрофотометра при 415-495 нм.
Пример 4: Характеристика антитела
Гибридомы, которые продуцировали антитела к человеческому CTGF, получали, как описано в примере 3. Клонированные клетки гибридомы выращивали в модифицированной Дульбекко среде Игла с высоким содержанием глюкозы/RPMI 1640 (50:50) с 8 мМ L-глутамина, 1/2x не незаменимых аминокислот и 10% фетальной телячьей сывороткой. Клетки, размноженные для получения антитела, выращивали в той же среде с 1,5% фетальной телячьей сывороткой с низким содержанием IgG в течение 4-9 дней при 37°С и 6% СО2. Полученную кондиционированную среду очищали от клеток и концентрировали, используя устройство фильтрации в тангенциальном потоке/концентрирования. Концентрат пропускали через колонку с белком А и связанные моноклональные антитела элюировали 100 мМ глицина, рН 3. Элюат нейтрализовали 1 М Tris, pH 8,0 и проводили диализ против PBS.
4.1 Картирование эпитопов
Картирование эпитопов антител с помощью экспериментов конкурентного связывания хорошо известно специалистам в области иммунологии (см. например, Van Der Geld et al. (1999) Clinical and Experimantal Immunology, 118:487-96). Каждую популяцию антител, выделенную из клеток, размножившихся из уникальной клетки клонированной гибридомы, картировали и относили к специфическому домену связывания человеческого CTGF, используя стандартные эксперименты связывания и блокирования (см. например, Antibodies: A Laboratory Manual (1988) Harlow and Lane (eds), Cold Spring Harbor Laboratory Press; Tietz Textbook of Clinical Chemistry, 2nd ed., (1994) Chapter 10 (Immunochemical Techniques), Saunders и Clinical Chemistry: Theory, Analysis, Correlation (1984) Chapter 10 (Immunochemical Techniques) и Chapter 11 (Competitive Binding Assays), C.V. Mosby, St. Louis). Независимые домены связывания первоначально определяли с помощью экспериментов конкуренции антител, в которых 2 различных антитела инкубировали в последовательном порядке на планшетах, покрытых CTGF. Если стерическое препятствие от первого антитела мешало связыванию второго антитела с CTGF, то эти 2 антитела относили к одному и тому же домену связывания. Однако следует понимать, что эти 2 антитела могут иметь различные эпитопы, но все же настолько близкие по отношению друг к другу, чтобы обозначаться как члены одного и того же домена связывания.
Были идентифицированы домены связывания, охватывающие все 4 экзона человеческого CTGF. Все домены связывания конформационно определены, так что антитела связываются с CTGF в невосстанавливающих условиях в анализах вестерн-блоттинга. Некоторые антитела также связываются с CTGF в восстанавливающих условиях в анализах вестерн-блоттинга, это предполагает, что каждое из этих антител связывается с линейным эпитопом белка CTGF. Также антитела, представляющие поднабор доменов связывания, проявляют перекрестную реактивность на мышиный CTGF в анализах вестерн-блоттинга. Антитело из каждой группы, имеющее самую высокую аффинность ко всему CTGF, использовали для дальнейшей характеристики и анализа.
Более точное картирование эпитопов выполняли с помощью анализа ELISA, используя специфические рекомбинантно экспрессированные фрагменты CTGF. Например, антитела, которые распознают эпитопы N-концевого домена CTGF, идентифицировали анализом ELISA против иммобилизованных фрагментов, полученных в результате рекомбинантной экспрессии экзона 2 и/или экзона 3 гена CTGF. Таким образом, отбирали и проводили дальнейшую характеристику антител, которые специфически распознают N-концевые домены или N-концевые фрагменты CTGF. Также отбирали и проводили дальнейшую характеристику антител, которые специфически распознают С-концевые домены или С-концевые фрагменты CTGF.
Группа эпитопов, определенная mAb1, связывается с линейным эпитопом N-концевого фрагмента CTGF, кодируемого экзоном 3. Была генерирована серия усеченных синтетических пептидов, перекрывающих области, кодируемые полинуклеотидом экзона 3, и были проведены тесты ELISA с использованием этих пептидов для дальнейшего определения эпитопа mAb1. Результаты суммированы в табл.1; "+" указывает на связывание между пептидом и mAb1, тогда как "-" указывает на то, что mAb1 не связывается с пептидом. Представленная жирным шрифтом курсивом "С" указывает на остаток цистеина в пептиде, который является существенным для связывания mAb1. Подчеркнутая "С" указывает на остаток цистеина, добавленный на конце и не являющийся частью последовательности нативного CTGF.
Связывание mAb1 с серией усеченных пептидов, кодированных экзоном 3
Поэтому mAb1 является членом класса антител, которые связываются с N-концевой областью CTGF. Линейный эпитоп CTGF, необходимый и достаточный для связывания mAb1, определяется аминокислотными остатками L143-V154 человеческого CTGF (SEQ ID NO:2). Дополнительное подтверждение специфичности связывания mAb1 с этим пептидом было получено с помощью RIA и аффинной хроматографии. Антитела, которые разделяют этот эпитоп частично или полностью, особо включены в настоящее изобретение. Кроме того, антитела, которые конкурируют с mAb1 за связывание с CTGF или его фрагментом, также особо включены в настоящее изобретение.
4.2 Аффинность антител к CTGF
Аффинность антител определяется как сила всех нековалентных взаимодействий между одним антигенсвязывающим сайтом антитела, и одним эпитопом антигена. Аффинность рассчитывается с помощью измерения константы ассоциации (Ка), так что
где [Ab] представляет собой концентрацию свободного антигенсвязывающего сайта антитела, [Ag] представляет собой концентрацию свободного антигена, [Ab·Ag] представляет собой концентрацию антигенсвязывающего сайта антитела, занятого антигеном, и Kd представляет собой константу диссоциации комплекса антитело-антиген.
Аффинность каждой популяции антител, идентифицированной картированием эпитопов, измеряли, используя RIA, где весь rhCTGF связывали с радиоактивным йодом и добавляли в лунки, содержащие иммобилизованное моноклональное антитело, следующим образом. Рекомбинантный человеческий CTGF метили радиоактивным изотопом 125I, используя способ хлорамин-Т (см. Greenwood et al. (1963) Biochem. J., 89:114-123). Обычно включалось по меньшей мере 60% 125I и удельная активность меченого CTGF составляла по меньшей мере 1×105 импульсов/мин/нг, хотя можно использовать при радиоиммуноанализе меченый CTGF более низкой удельной активности. Козье антитело против человеческого IgG, γFc-специфическое антитело захвата (Jackson ImmunoResearch) в DPBS, лишенной Ca2+ и Mg2+ (Mediatech, Herndon VA), добавляли в лунки титровочного микропланшета MAXISORP BREAKAPART (Nalge Nunc International, Rochester NY) и давали возможность связываться в течение ночи при 4°С. Затем лунки блокировали 1% BSA (бычьим сывороточным альбумином) в DPBS, лишенной Ca2+ и Mg2+, в течение по меньшей мере 4 ч при 4°С. Блокирующий раствор удаляли, добавляли 100 мкл тестируемого антитела при 2-50 нг/мл в DPBS, лишенной Ca2+ И Mg2+, и давали возможность связываться в течение ночи при 4°С. Смеси серийных разведений немеченого CTGF при постоянном количестве [125I]rhCTGF добавляли в лунки и инкубировали при комнатной температуре в течение 4-8 ч. Затем лунки промывали 4 раза 0,1% Tween 20 в PBS, лишенной Ca2+ И Mg2+ (Mediatech), и лунки титровочного микропланшета отделяли и проводили подсчет в гамма-счетчике.
Аффинность оценивали графически с помощью способа Скэтчарда (1948, Ann. NY Acad. Sci., 51:660-72). Общую концентрацию меченого CTGF, нанесенного на планшет, рассчитывали как
где "cpm_applied" представляют собой импульсы, полученные из контрольных флакончиков, которые заполнены смесями CTGF, параллельно с лунками титровочного микропланшета; "cpm/fmol" представляет собой удельную активность [125I]CTGF, "[CTGF]охл. исх. раствор" представляет собой концентрацию немеченого CTGF, добавленного в каждую лунку, и "разведение" представляет собой фактор разведения для немеченого CTGF.
Концентрацию CTGF, связанного с антителом, рассчитывали по пропорции импульсов, связанных с лунками, и общей концентрации CTGF, внесенного в лунки.
Концентрация свободного (несвязанного) CTGF представляет собой разность между общей концентрацией нанесенного CTGF и концентрацией связанного CTGF.
Графики Скэтчарда определения аффинности для антител по изобретению показаны на фиг.2. На фиг.2А показан график связывания антитела по изобретению, mAb2, с [125I]rhCTGF в присутствии увеличивающихся концентраций немеченого rhCTGF. На фиг.2В показан график связывания иллюстративного антитела по изобретению, mAb1, с [125I]rhCTGF в присутствии увеличивающихся концентраций немеченого rhCTGF. Большей массе соответствуют точки с одинаковым соотношением связанного и несвязанного CTGF, потому что эти точки имеют количество импульсов связанного CTGF, превышающее контроли (следовательно, определенные в лунке импульсы связанного фактора), но все же существенно меньше, чем общее количество импульсов нанесенного CTGF (следовательно, определенные в лунке импульсы свободного фактора). Максимальное связывание (Bmax) и Kd представлены в виде отрезков, отсекаемых на оси х и оси у соответственно.
Аффинность (Kd) mAb1 к CTGF составляет менее чем 10-9 М, обычно обнаруживаемая аффинность у коммерчески успешных терапевтических средств антител (см. например, Maini et al. (1998) Arthritis Rheum., 41: 1552-1563; Targan et al (1997) N. Engl. J. Med., 337: 1029-1035; Bumgardner et al. (2001) Transplantation, 72: 839-45 и Kovarik et al. (1999) Transplantation, 68: 1288-94). Таким образом, mAb1 является подходящим кандидатом для терапевтического применения, и антитела, которые разделяют связывание эпитопов с mAb1, как описано выше, и имеют аффинность к CTGF, которая аналогична или больше аффинности mAb1 (то есть Kd=10-9), аналогичным образом являются подходящими кандидатами для терапевтического применения. Антитела, разделяющие связывание эпитопов с mAb1, но имеющие более низкую аффинность (т.е. более высокое значение Kd), чем mAb1, также осуществлены в рамках настоящего изобретения и потенциально могут использоваться в различных анализах и диагностическе, как описано в настоящем описании. Такие антитела, кроме того, подходят для терапевтического применения, особенно если они имеют высокую авидность к антигену, как описано ниже.
4.3 Авидность антител
Для антител с более чем одним антигенсвязывающим сайтом (мультивалентность) аффинность одного связывающего сайта не всегда отражает истинную силу взаимодействия антитело-антиген. Когда мультивалентное антитело связывается с антигеном, имеющим множественные повторяющиеся эпитопы, взаимодействие одного антигена, взаимодействующего у одного связывающего сайта антителе, увеличивает возможность взаимодействия антигена с дополнительными связывающими сайтами. Авидность отражает силу функционального объединения антитела с антигеном, которая связана как с аффинностью взаимодействия между эпитопами и паратопами, так и валентностью антитела и антигена. Таким образом, авидность обеспечивает более точную меру тенденции антитела к диссоциации.
Высокая авидность может компенсировать низкую аффинность. Например, сайты связывания антигена IgM обычно имеют более низкую аффинность, чем IgG, но мультивалентность IgM дает ему высокую авидность, позволяя, таким образом, эффективно связывать антиген.
Для определения авидности антител по изобретению фрагмента сначала Fab были получены обычным расщеплением папаином соответствующего иммуноглобулина. Затем иммобилизованный белок А использовали для отделения фрагментов Fab от Fc и непереваренного антитела.
Приблизительно 1 мл суспензии иммобилизованного папаина, содержащей 0,5 мл затвердевшего геля, 250 мкг папаина и 3,5 единиц ВАЕЕ, промывали 3 × 1 мл и 1 × 10 мл буфера расщепления (DB; 20 мМ фосфата натрия, 10 мМ EDTA (этилендиамин-тетрауксусной кислоты), 20 мМ цистеина, рН 7,0). Затем суспензию ресуспендировали в 0,3 мл DB, смешивали с 1,1 мл антитела (приблизительно 5 мг, рН 7) и перемешивали в течение ночи при 37°С. Затем продукт расщепления антитела отделяли от смолы и фрагменты Fab отделяли от фрагментов Fc и непереваренного антитела с помощью аффинной хроматографии с использованием белка А. Чистоту фрагмента Fab контролировали с помощью SDS-PAGE (фиг.3А).
Моновалентное связывание отличали от бивалентного связывания путем элюирования связанных с антигеном антител тиоцианатом с меняющейся концентрацией. При увеличении концентрации хаотропного иона (тиоцианата) в растворе сначала разрушаются ассоциации с более низкой аффинностью (например, моновалентное связывание Fab с антигеном), тогда как ассоциации с более высокой аффинностью (например, бивалентное связывание IgG с лигандом) остаются нетронутыми. Таким образом, с помощью увеличения концентрации тиоцианина можно различить 2 различных связывания.
Планшеты покрывали 10 мкг/мл CTGF или пептидов CTGF в 50 мМ бикарбонатном буфере (рН 8,5) при 4°С в течение ночи, блокировали блокатором казеином/TBS при 4°С в течение ночи и затем инкубировали со 100 мкг/мл антитела или соответствующего Fab в блокаторе казеине/TBS при комнатной температуре в течение ночи при перемешивании. Затем планшеты инкубировали с разведениями (1:1) тиоцианата (0-7,6 М) в 100 мМ фосфатном буфере (рН 6,0) в течение 15 мин при комнатной температуре и перемешивании с последующей инкубацией с конъюгатом щелочной фосфатазы-мышиного антитела против человеческого антигена (Fab')2 (разведение 1:1000) при комнатной температуре в течение 45 мин. Добавляли субстрат щелочной фосфатазы (1 мг/мл; Sigma) в 1 М диэтаноламине, 0,5 мМ MgCl2 (рН 9,8), планшеты инкубировали при комнатной температуре и определяли поглощение при 405 нм через 2, 10, 20 и 60 мин.
Показатель аффинности является концентрацией хаотропного вещества (тиоцианата), которая приводит к снижению первоначального поглощения на 50%. Для иллюстративного антитела по изобретению mAb1, показатель аффинности для диссоциации Fab от CTGF, составил 0,46 М, тогда как показатель аффинности для диссоциации интактного IgG от CTGF составил 1,8 М (фиг.3В). Таким образом, mAb1 связывается с антигеном преимущественно бивалентно (авидность) и диссоциируется от антигена гораздо медленнее, чем антитело, которое связывается моновалентно. Дополнительные антитела по изобретению, которые разделяют показатели связывания эпитопов с mAb1, могут быть аналогичным образом бивалентными, или они могут быть моно- или мультивалентными. Любым из антител по изобретению можно манипулировать для улучшения авидности, например комбинированием сайтов связывания эпитопов в один конструкт антитела, например тритело и т.д. (см. например, Schoonjans et al. (2000) J. Immunol., 165:7050-7057).
4.4 Перекрестная реактивность
Описанный выше радиоиммуноанализ (пример 4.2) использовали для определения перекрестной реактивности антител за исключением того, что немеченый rhCTGF был заменен другим немеченым конкурентом, крысиным CTGF, полученным из клеток почки нормальной крысы (NRK). Клетки NRK выращивали до их слияния и затем культуральную среду меняли на бессывороточную среду, содержащую 2 нг/мл TGF-β2, 50 мкг/мл гепарина и 250 мкг/мл BSA. Кондиционированную среду собирали через 2 дня культивирования, центрифугировали для удаления клеточных фрагментов и инкубировали с гранулами гепарина-сефарозы (1/100 об/об суспензии гранул:среды) в течение 2 ч при 4°С при перемешивании. Затем смесь центрифугировали; гранулы собирали и промывали PBS и затем лизировали в буфере SDS (додецилсульфата натрия).
График Скэтчарда связывания mAb2 с [125I]rhCTGF в присутствии увеличивающихся концентраций немеченого крысиного CTGF показан на фиг.4А; а график Скэтчарда связывания mAb1 с [125I]rhCTGF в присутствии увеличивающихся концентраций немеченого крысиного CTGF показан на фиг.4В. Как видно из фигуры, mAb1 связывается как с человеческим, так и с крысиным CTGF, тогда как mAb2 связывается с человеческим, но не связывается с крысиным CTGF.
Для mAb1 графики Скэтчарда в случае конкуренции с крысиным CTGF (фиг.4В) имеют меньший наклон, более низкую видимую аффинность и более высокую видимую величину Bmax, чем графики в случае конкуренции с rhCTGF (фиг.2В). Таким образом, хотя крысиный CTGF способен конкурировать с человеческим CTGF за связывание с mAb1, антитело имеет более высокую аффинность к рекомбинантному человеческому CTGF, чем к рекомбинантному крысиному CTGF. mAb1 также перекрестно взаимодействует с мышиным и обезьяним CTGF (данные не показаны). Антитела, проявляющие подходящую аффинность к CTGF от других видов, можно использовать при лечении и предотвращении расстройств у этих видов. Например, антитело по изобретению, которое проявляет подходящий Kd к собачьему CTGF, можно использовать для лечения расстройства, связанного с CTGF, у собак. Антитела по изобретению, которые проявляют перекрестновидовую аффинность, такие как mAb1, можно также использовать в качестве инструментов исследования для изучения расстройств, связанных с CTGF, на различных моделях на животных.
4.5 Гликозилирование
Описанный выше радиоиммуноанализ (пример 4.2) использовали для определения эффекта гликозилирования антител на аффинность связывания антигенов. Антитело mAb1 обрабатывали в течение 8 дней при 37°С в PBS, 0,5 M EDTA, pH 8,0, пептидом N-гликозидазой F (PNGаза F), который отщепляет олигосахариды от N-связанных гликопротеидов. После инкубации реакционный раствор или использовали непосредственно, или фракционировали на колонке с белком А-SEPHAROSE FASTFLOW (Amersham Bioscience, Piscataway NJ) и элюировали 0,1 М глицином-HCl, рН 2,5. Извлечение антитела после фракционирования составило приблизительно 87%, и уровень эндотоксина составил 0,30 единиц энтропии/мг. Дегликозилирование подтверждали SDS-PAGE. Активность связывания дегликозилированного антитела с человеческим рекомбинантным CTGF была идентична в пределах экспериментальной ошибки активности связывания гликозилированной формы антитела.
Поскольку различные клетки продуцируют различные типы гликозилирования, продукция рекомбинантных белков, например антител, в культивированных клетках или негомологичных видах, может генерировать ненативное гликозилирование.
Для активности некоторых белков требуется специфическое гликозилирование, и измененное гликозилирование снижает активность; например в случае антител снижается аффинность к антигену. Продукция белка в определенных системах, например растениях и куриных яйцах, может также продуцировать типы гликозилирования, которые являются иммуногенными, таким образом, снижая возможность использования белков при определенных видах применения. Способность настоящих антител проявлять одинаковую активность в гликозилированной и негликозилированной форме демонстрирует то, что изобретение не ограничивается присутствием гликозилирования, в частности, видоспецифического гликозилирования.
Пример 5. Анализ клеточной миграции
Миграция клеток представляет собой нормальное и важное клеточное явление, например, во время развития и заживления ран. Миграция клеток также является фактором в патологии расстройств, таких как формирование фибротических поражений, и клетки, выделенные из фибротических поражений, больше реагируют на хемотаксические раздражители, чем клетки из соответствующей нормальной ткани.
Антитела по настоящему изобретению анализировали на их способность ингибировать стимулированную CTGF хемотаксическую миграцию гладкомышечных клеток с использованием анализа в камере Бойдена следующим образом. Артериальные гладкомышечные клетки крыс (ASMC) в среде, содержащей 0,1% фетальную телячью сыворотку (FCS), добавляли в верхний отсек камеры Бойдена, а среду, содержащую или 300 нг/мл rhCTGF, 10% FCS, или только 0,1% FCS, добавляли в нижний отсек. Покрытый коллагеном фильтр, имеющий поры диаметром 8 мкм, отделял верхнюю камеру от нижней камеры. Клеткам давали возможность в течение 2-3 ч сцепиться с фильтром и мигрировать через него. Затем фильтр удаляли, клетки на фильтре фиксировали и окрашивали и подсчитывали клетки, которые мигрировали через фильтр. Инкубация с 300 нг/мл rhCTGF увеличивала количество клеток, мигрирующих через фильтр, приблизительно в 5 раз относительно контролей 0,1% FCS. Увеличение миграции, стимулированной CTGF, составило приблизительно 27% хемотаксического эффекта, наблюдавшегося в случае 10% FCS, которая содержит множественные хемотаксические факторы.
Антитела по изобретению испытывали на их способность ингибировать опосредованную CTGF клеточную миграцию, используя описанный выше анализ, за исключением того, что или антитела против CTGF (в концентрации 30 и 300 мг/мл), или объединенный человеческий IgG также добавляли в нижнюю камеру. Подсчитывали 4 поля клеток каждого из 3 отдельных фильтров для каждого образца в каждом анализе. Результаты показаны в табл.2.
Ингибирование опосредованной CTGF клеточной миграции
Как видно в табл.2, антитела, которые связываются с CTGF, в пределах эпитопа, определенного mAb1, ингибируют опосредованную CTGF клеточную миграцию зависимым от дозы образом. Антитела группы этого эпитопа представляли собой единственные испытанные антитела против CTGF, которые повторно и воспроизводимо ингибировали вызванную CTGF миграцию.
Различные процессы, такие как ангиогенез, хондрогенез и онкогенез, требуют изменения адгезии и миграции клеток. CTGF связан как с адгезией, так и с миграцией клеток, и способность антител, направленных против CTGF, дифференциально воздействовать на одну активность в сравнении с другой предоставляет разнообразный репертуар лекарственных средств для лечения состояний, связанных с CTGF. Антитела, полученные в настоящем изобретении, ясно демонстрируют дифференциальную активность, относящуюся к нейтрализации активностей CTGF. Как проиллюстрировано ниже, эти способности предоставляют уникальный терапевтический потенциал в этом классе антитела против CTGF.
Пример 6. Легочные расстройства
Внутритрахеальная (IT) инстилляция блеомицина у мышей представляет собой модельную систему, широко используемую для изучения пневмосклероза и для скрининга потенциально желательных антифибротических средств. Антитела по изобретению испытывали на их способность уменьшать вызванный блеомицином пневмосклероз in vivo с использованием процедуры, описанной Wang et al. (2000) Biochem. Pharmacol., 60:1949-1958, следующим образом.
Самцов мышей C57BL/6 методом случайной выборки делили на 2 группы. Мышей наркотизировали изофлюраном и затем интратрахеально инъецировали или одну дозу блеомицина в 0,9% солевом растворе в дозе 0,1 единицы/50 мкл/мышь, или один 0,9% солевой раствор. Каждую группу делили и лечили сразу и затем 1 раз через день, чтобы всего количество вводимых внутрибрюшинно (в/б) доз или солевого раствора, или антитела было равно 7. Через 14 дней после внутритрахеальной инстилляции мышей подвергали эвтаназии кровопусканием из нисходящей брюшной аорты под наркозом и брали легочную ткань.
Содержание коллагена в легких анализировали, измеряя уровень гидроксипролина и пролина, используя способ Palmerini et al. (1985; J. Chromatogr., 339:285-292), за исключением того, что L-азетидин-2-карбоновая кислота (Aldrich) была заменена на 3,4-дигидропролин в качестве внутреннего стандарта. Вкратце, образцы ткани гидролизировали в 6н. HCl в течение 22 ч при 105°С. Образцы подвергали предколоночной дериватизации о-фталальдегидом и затем 4-хлор-7-нитробензофураном (Aldrich) для образования флуоресцентных аддуктов пролина и гидроксипролина. Флюоресцентные аддукты отделяли ВЭЖХ на обращенной фазе с последующим флуорометрическим выявлением.
На фиг.5 показано сравнение результата терапевтического введения солевого раствора (SA), иллюстративного антитела по изобретению, mAb1, и пула антител, специфичных для CTGF (AbsJ) в случае их способности подавлять пневмосклероз после обработки блеомицином. Как видно на фиг.5А, обработка блеомицином (BL+SA) значительно увеличивала содержание гидроксипролина на 168% по сравнению с контрольной группой (SA+SA; 220 ± 15 мкг/легкое). Однако последующее лечение объединенными антителами по изобретению (BL+ AbsJ) показало снижение на 60% содержания гидроксипролина в легких по сравнению с обработкой одним блеомицином. Аналогичным образом последующее лечение mAb1 (BL+mAb1) показало снижение содержания гидроксипролина в легких на 70% по сравнению с одним блеомицином.
Гистологическое исследование легких мышей выявило нормальную легочную паренхиматозную ткань в контрольной группе (не показана). Однако в легких, обработанных блеомицином, отчетливо наблюдалось увеличение областей фиброза (фиг.5В; стрелки). Терапевтическое введение антитела по изобретению после обработки блеомицином показало отчетливое уменьшение фиброза (фиг.5С), хотя в некоторых долях еще проявлялась легкая степень интерстициального фиброза. Таким образом, антитела по изобретению являются терапевтически выгодными при введении пациентам с риском развития легочного расстройства, такого как идиопатический пневмосклероз (IPF).
Пример 7. Почечные расстройства
7.1. Почечная недостаточность
Канальцево-интерстициальный фиброз представляет собой основной компонент нескольких почечных заболеваний, связанных с прогрессированием конечной стадии почечной недостаточности (Sharma et al. (1993) Kidney Int., 44:774-788). Односторонняя обструкция мочеточника (UUO), характеризуемая сниженной почечной функцией и увеличенным интерстициальным фиброзом, использовалась в качестве экспериментальной модели для индукции канальцево-интерстициального повреждения и фиброза (Fern et al.(1999) J. Clin. Invest., 103:39-46).
Мышей наркотизировали изофлюраном и затем выполняли перевязку левого мочеточника в соответствии со способом, описанным Moriyama et al.(1998; Kidney Int., 54:110-119). Мышей лечили сразу после операции и затем 1 раз через день для обеспечения всего семи доз или солевого раствора, или антитела, вводимого внутрибрюшинно (в/б). Через 14 дней после UUO животных наркотизировали и умерщвляли кровопусканием из нисходящей брюшной аорты. И правую, и левую почки отдельно декапсулировали и взвешивали. Половину каждой почки фиксировали в 10% формалине для гистологического исследования (окрашивание трихромом), а другую половину взвешивали и хранили при -70°С для определения содержания гидроксипролина. Содержание гидроксипролина и пролина определяли, как описано выше.
Как видно на фиг.6А, у каждой мыши в результате UUO увеличивалось содержание коллагена в почке приблизительно в 4 раза по данным измерения соотношения между гидроксипролином и пролином в левой почке, подвергнутой обструкции, относительно не подвергнутой обструкции правой почки. Лечение антителом по изобретению, mAb1, привело к статистически значимому зависимому от дозы снижению фиброза подвергнутой обструкции почки (фиг.6А). Однако антитело, которое связывается с С-концевым эпитопом на CTGF, mAb3, не проявило значимого эффекта. Окрашивание трихромом почки после UUO выявляет области увеличенного накопления коллагена (фиг.6В, стрелки), тогда как лечение антителом по изобретению показывает значительное уменьшение окрашивания коллагена в почке, подвергнутой обструкции (фиг.6С).
Альтернативно фиброз почек можно изучать на модели прогрессирующей почечной недостаточности - остаточной почки крысы. Модель, которая включает одностороннюю нефрэктомию на 2/3 в сочетании с субтотальной резекцией второй почки (нефрэктомия на 5/6), вызывает дегенеративные паренхиматозные изменения с хронической почечной недостаточностью в остатке почки, и у животных развивается уремия и проявляется выраженная альбуминурия, гломерулосклероз, интерстициальный фиброз и атрофия канальцев (см. например, Frazer et al. (2000) Vet. Pathol., 37:328-335; и Gandhi et al. (1998) Kidney Int., 54:1157-1165).
Нефрэктомию на 5/6 выполняли по методу Frazier et al. (2000, Vet Pathol 37:328-335). Пятинедельных самцов крыс Sprague-Dawley (Harlan, Indianapolis IN) со средней массой тела 120 г наркотизировали кетамином и ксилазином и проводили разрез краниальной 1/3 и каудальной 1/3 левой почки. Накладывали марлевый тампон кратковременно для обеспечения гемостаза, брюшную полость промывали солевым раствором, 0,2 мл буторфенола и операционную рану зашивали. Через 1 неделю после первоначальной операции вторую почку полностью удаляли.
Крыс делили на группы лечения солевым раствором и антителом, причем лечение начинали через 2 недели после нефрэктомии на 5/6. Солевой раствор или антитело в дозировке 5 мг/кг вводили в/б инъекцией (0,5 мл каждая) 1 раз/3 дня в течение 15 дней (всего 5 инъекций). Образцы крови и мочи брали еженедельно у рандомизированных крыс, подвергнутых нефрэктомии, для слежения за развитием почечного заболевания и для корреляции функциональных почечных нарушений с гистологическими изменениями. Результаты анализа почечного фиброза, анализа мочи и анализов химического состава сыворотки сравнивали у групп через 18 и 28 дней после начала лечения.
Почечный фиброз оценивался независимо двумя патологоанатомами слепым методом; 3 гистологических среза из каждой почки исследовали, используя 3 различных морфологических красителя: гематоксилин/эозин, трихром Masson и пикриновая кислота-сириус красный. Кроме того, выполняли иммуногистохимическое исследование на замороженных срезах для оценки типа отложения коллагена в каждом участке в почке. Выполняли количественную оценку коллагена (соотношение гидроксипролин/пролин), и почечную функцию оценивали, используя и анализ мочи, и химический анализ сыворотки образцов, взятых во время эвтаназии.
Гистологически были отмечены умеренные различия фиброза в остатках почек без лечения и после лечения антителом (фиг.7). Через 3 дня после лечения осуществляли слепую субъективную оценку, имеющую своим результатом величину средней балльной оценки фиброза 12,6 в группе лечения солевым раствором, в сравнении с 10,7 в группе лечения антителом (p<0,05). Статистически значимые различия гистологической степени фиброза у крыс, получавших лечение солевым раствором и антителом, сохранялись через 14 дней после лечения при средней балльной оценке фиброза, равной 16,9 в группе лечения солевым раствором, в сравнении с 14,4 в группе лечения антителом (p<0,05). Количественный анализ содержания гидроксипролина коллагена также продемонстрировал тенденцию к сниженному фиброзу в группе лечения антителом относительно группы лечения солевым раствором, но различие не было статистически значимым.
Между группами лечения были также отмечены качественные различия. Хотя большая часть отложения коллагена в группах лечения антителом ограничивалась кортикомедуллярным и медуллярным интерстицием, фиброз у крыс, получавших лечение солевым раствором, был многоочаговым с диффузным распределением по коре и мозговому слою. Наиболее выраженное патогистологическое различие наблюдалось в случае в количества клубочкового фиброза. У многих крыс из группы лечения солевым раствором имелся гломерулосклероз от умеренной до тяжелой степени с перикапсулярным фиброзом, утолщением мембраны Боумэна, спайками и запустеванием клубочков. В других группах эти изменения были от минимальных до легких, включая получавшие лечение антителом почки крыс. Накопление коллагена визуализировали окрашиванием трихромом Masson и пикриновой кислотой-сириусом красным.
В обеих моделях прогрессирующей почечной недостаточности антитело по изобретению уменьшало разрушение ткани и улучшало почечную функцию. Таким образом, антитела по изобретению обеспечивают терапевтическую выгоду при введении пациентам с риском развития почечного расстройства, такого как гломерулонефрит, нефропатия, вызванная IgA, гломерулосклероз; почечной недостаточности и разрушения канальцев, вызванного токсинами, и т.д.
7.2. Диабетическая нефропатия
Сахарный диабет ведет к недостаточности множества органов, включая, но не ограничиваясь этим, почки, сердце и глаза. Основным компонентом патоморфологического прогрессирования диабетической недостаточности органа является фиброз. Признанной моделью диабетической нефропатии является мышь, несущая мутацию потери функции рецептора лептина (Ob-R, кодируемого геном db). Ключевые признаки, общие между мышью db/db и диабетической нефропатией человека, включают почечную гипертрофию, увеличение клубочков, альбуминурию и расширение мезангиального матрикса.
Антитела по изобретению испытывали, используя следующую модель на мышах db/db диабетической нефропатии. Восьминедельных мышей db/db (Harlan, Indianapolis IN) и их гетерозиготных особей db/+ того же помета лечили внутрибрюшинной инъекцией либо антитела по изобретению (CLN1; см. ниже), либо контрольного человеческого IgG (cIgG). У всех животных за первоначальной инъекцией 300 мкг антитела следовали дозы 100 мкг, вводимые 3 раза в неделю в течение 60 дней. Брали образцы крови и измеряли массу тела в начале и периодически в течение всего периода лечения. Регистрировали также потребление пищи.
К 11-ой неделе существовало четкое различие между животными с диабетом (db/db) и животными без диабета (db/+) в отношении массы тела, уровней глюкозы в крови и потребления пищи. Лечение или антителом по изобретению, или контрольным антителом не оказывало значимого воздействия ни на один из этих показателей. Однако различные способы определения почечной функции показали отчетливое различие между мышами, пораженными и не пораженными сахарным диабетом. Как видно из табл.3, у мышей с диабетом было выявлено увеличение массы почек, клиренса креатинина и скорости выделения альбумина (AER) относительно мышей без диабета. Однако у животных с диабетом, получавших лечение антителом по изобретению, выявлены нормализованные величины всех показателей. Все данные выражены в виде средней величины ± стандартная ошибка средней. Количество мышей в группе (n) находилось в диапазоне от 9 до 15.
Почечная функция у мышей db/db и db/+
* P<0,01, в сравнении с мышами db/+ и P<0,05 с получавшими clgG мышами db/db.
Поскольку CTGF индуцируется при высоком уровне глюкозы и опосредует различные виды активности, включая продукцию ЕСМ в тканях в результате повреждения, например вследствие образования и накопления конечных продуктов гликирования (AGE), и т.д, патологические состояния, связанные с сахарным диабетом, такие как диабетическая нефропатия, можно предотвратить, используя антитела по изобретению.
Пример 8. Глазные расстройства
Увеличенная экспрессия CTGF связана с различными глазными расстройствами, включая пролиферативную витреоретинопатию (PVR), дегенерацию желтого пятна и диабетическую ретинопатию (см. например, (Hinton et al (2002) Eye, 16: 422-428; He et al. (2003) Arch Ophthalmol., 121: 1283-1288 и Tikellis et al. (2004) Endocrinology, 145: 860-866). Была предположена роль CTGF и применение лекарственных средств против CTGF (см. международную публикацию WO 03/049773). Антитела по настоящему изобретению являются представителями класса уникальных терапевтически эффективных лекарственных средств против CTGF для применения в случае глазных расстройствах. Способность антител по настоящему изобретению облегчать осложнения при глазных расстройствах испытывается на моделях глазных заболеваний следующим образом.
8.1. Диабетическая ретинопатия
Модели на животных сахарного диабета, например мыши db/db, описаны в примере 7.2 выше. Любую из этих моделей можно использовать для демонстрации эффективности лечения диабетической ретинопатии с использованием антител по изобретению. Конкретная модель диабетической ретинопатии представлена ниже, при которой животным инъецируют стрептозотоцин (STZ), известный токсин секретирующих инсулин панкреатических β-островковых клеток.
Сахарный диабет вызывается у крыс (например, Long-Evans, Sprague-Dawley и т.д.) инъекцией, например, внутрибрюшинно, стрептозотоцина (STZ), например, в дозе примерно 60-85 мг/кг массы тела. Для улучшения выживаемости крысам можно давать 10% сахарную воду в течение 24 ч и/или 2-4 единицы инсулина в день после инъекции STZ. Различные факторы, включая, например, массу тела, скорость выделения альбумина с мочой, уровень глюкозы в крови, гликированный гемоглобин, артериальное давление и т.д., измеряют, например, через 4, 8 и 12 недель. Одновременно наблюдаются контрольные животные, которым инъецировали один буфер. Одну половину получавших STZ и контрольных крыс дополнительно лечат антителом по изобретению, инъецируемым, например, внутривенно, внутрибрюшинно или интраокулярно. В течение исследования животные имеют неограниченный доступ к пище и воде. Животных умерщвляют через 12 недель и глаза извлекают и исследуют для выявления гистологических изменений.
Уменьшение патоморфологических изменений у животных, получавших лечение антителом, относительно не получавших лечение контролей указывает на терапевтическую эффективность при диабетической ретинопатии. Поскольку CTGF индуцируется высоким уровнем глюкозы и опосредует различные виды активности, включая продукцию ЕСМ в тканях в результате повреждения, например, вследствие образования и накопления конечных продуктов гликирования (AGE), и т.д, то патологические состояния, связанные с сахарным диабетом, такие как диабетическая ретинопатия, можно предотвратить, используя лекарственные препараты против CTGF (см. например, международную публикацию № WO 03/049773). Антитела по настоящему изобретению являются представителями уникального класса терапевтически эффективных лекарственных средств против CTGF для применения при глазных расстройствах, например диабетической ретинопатии.
8.2. PVR (пролиферативная витреоретинопатия)
Клетки пигментного эпителия сетчатки (RPE) кроликов выделяют из глаз взрослых кроликов и культивируют в среде DMEM с добавкой 10% фетальной телячьей сыворотки. Почти слившиеся культуры (обычно при 2-3 пассаже) используют для всех последующих инъекций. Во время инъекции культивированные клетки RPE собирают и суспендируют в PBS приблизительно до концентрации 2,5 × 106 клеток/мл. Приблизительно 0,2 мл водянистой влаги удаляют из каждого глаза кролика-реципиента, используя иглу 25G, и затем клетки RPE инъецируют через склеру в точку на 3 мм позади от лимба непосредственно над зрительным диском и выше него, используя иглу 27G. После инъекции клеток RPE или 0,1 мл PDGF BB (50-150 нг), CTGF (200-400 нг), или PDGF и CTGF в PBS инъецируется через тот же участок входа. Глаз каждого животного без инъекции служит в качестве контроля. Необязательно, CTGF можно дополнительно инъецировать на 7-ой день и/или 14-й день после первой инъекции. Одну половину животных дополнительно лечат антителом по изобретению, инъецируемым, например, внутривенно, внутрибрюшинно или интраокулярно. В зависимости от участка введения антитело может вводиться ежедневно или водиться менее часто, например на 7-й, 10-й, 14-й день и т.д.
Животных исследуют, используя непрямые офтальмоскопические процедуры, для контроля развития и степени PVR, которая классифицируется в соответствии с параметрами, описанными Fastenberg (Fastenberg et al. (1982) Am. J. Ophthalmol., 93:565-572). Затем животных умерщвляют и глаза анализируют с помощью гистологического исследования для выявления и степени формирования мембраны и фиброза. Кроме того, сетчатку и фибротическую мембрану можно взять для измерения содержания коллагена.
Альтернативно PVR вызывают в глазах кролика, используя подсетчаточную инъекцию диспазы, используя адаптированные модель и процедуру, из публикации Frenzel et al. (1998, Invest. Ophthalmol. Vis. Sci., 39: 2157-2164). Подсетчаточный пузырь формируется с использованием 50 мл (0,05 Ед) диспазы (Sigma Chemical Co.) в PBS. Одну половину животных дополнительно лечат антителом по изобретению, инъецированным, например, внутривенно, внутрибрюшинно или интраокулярно. Отслоение сетчатки вызывают приблизительно у 75% кроликов, которым проводили инъекцию, не получающих антитело по изобретению через 1 неделю после операции и приблизительно у 100% этих животных через 2 недели после операции. Эпиретинальные мембраны исследуют для определения степени фиброза.
Уменьшение патоморфологических изменений у животных, получавших лечение антителом, относительно не получавших лечение контролей указывает на терапевтическую эффективность при PVR. Поскольку CTGF связан с повреждением ткани в случае моделей PVR, средства против CTGF предложены в качестве лекарственных средств для использования при таких расстройствах (см. например, международную публикацию WO 03/049773). Антитела по настоящему изобретению являются представителями уникального класса терапевтически эффективных лекарственных средств против CTGF для применения при глазных расстройствах, например PVR.
Пример 9. Склероз
Склероз в целом характеризуется диффузным фиброзом, дегенеративными изменениями и сосудистыми аномалиями в коже (склеродермия), суставах и внутренних органах, в частности пищеводе, желудочно-кишечном тракте, легких, сердце и почках.
9.1. Индукция локализованной гранулемы
У новорожденных мышей развивается стойкий локализованный фиброз при введении комбинации полученного у человека TGF-β2 и CTGF подкожной инъекцией в течение 7 последовательных дней (Mori et al. (1999) J. Cell Physiol., 181: 153-159; Shinozaki et al. (1997) Biochem. Biophys. Res. Commun., 237: 292-297).
Через 1 день после рождения мышей делили на 3 группы лечения и вводили 40 мкл 1% мышиного сывороточного альбумина (MSA), PBS, содержащий или 800 нг TGF-β2, 400 нг CTGF, или оба TGF-β2 и CTGF, подкожной инъекцией в подлопаточную область в течение 7 последовательных дней. Группу комбинации TGF-β2 и CTGF дополнительно делили на 2 группы, где одна группа дополнительно получала 40 мкг антитела по изобретению, mAb1. На 11-й день животных умерщвляли и срезы участков инъекции обрабатывали и окрашивали трихромом Mason для гистологической оценки. Покровные стекла рандомизировались и качественно оценивались в слепом методе тремя учеными; балльная оценка изменялась в диапазоне от 0 (отсутствие изменений) до 4 (фибротическая ткань) на основании степени фиброза или распространения соединительной ткани (см. фиг.8). Затем рассчитывали кумулятивные балльные оценки по каждому покровному стеклу от всех отдельных исследователей и средние величины сравнивали между группами с использованием теста ANOVA (вариационного анализа).
Групповые средние балльные оценки для контроля - носителя, TGF-β2 и комбинации TGF-β2 и CTGF - составили соответственно 0,75, 6,83 и 9,00 (табл.4).
Гистологическая балльная оценка гранулемы у новорожденных мышей
Групповая средняя балльная оценка в случае лечения антителом составила 6,17, что является статистически значимым уменьшением по сравнению с соответствующей комбинацией TGF-β2 и CTGF (p<0,05), тогда как лечение направленным на С-конец ненейтрализующим антителом против CTGF, mAb3, не уменьшало фиброз. Таким образом, антитела по настоящему изобретению, в частности, эффективны при уменьшении локального склеротического поражения тканей.
9.2. Системный фиброз новорожденных
Новорожденных мышей делили на группы и ежедневно в течение 21 дня вводили внутрибрюшинные инъекции 300 мкг/кг/день TGF-β, 300 мкг/кг/день CTGF, комбинацию TGF-β и CTGF по 300 мкг/кг/день каждого, или комбинацию TGF-β и CTGF с предшествующим в/б введением 5 мг/кг антитела по изобретению, mAb1, за 30 мин перед лечением фактором роста. Мышата оставались со своими матерями во время курса лечения. На 21-й день животных умерщвляли, основные органы удаляли и определяли общее содержание пролина и гидроксипролина, как описано выше.
Ежедневные инъекции TGFβ вызывали небольшой системный фиброз, в то время как один CTGF не вызывал реакции. Комбинация TGFβ и CTGF вызывала системный фиброз с обширным отложением коллагена в нескольких органах (фиг.10), включая печень, легкие, сердце, желудочно-кишечный тракт, диафрагму и почки; обширные кишечные спайки и увеличение смертности на 25%. Введение антитела по изобретению в сочетании с лечением фактором роста предотвращало фиброз органов (фиг.10) и кишечные спайки и предотвращало смертность. Таким образом, антитела по настоящему изобретению дополнительно эффективны при системном введении в уменьшении склеротического повреждения различных тканей и органов. Результаты, представленные в примерах 10.1 и 10.2, ясно демонстрируют, что антитела по изобретению при местном или системном введении терапевтически эффективны для лечения склеротических состояний.
9.3. Склеродермия
Антитела согласно изобретению можно использовать для уменьшения фиброза, связанного со склеродермией. Способы, которые измеряют степень и тяжесть поражения кожи при склеродермии, известны в данной области (см. например, Rodhan et al. (1979) Arthritis Rheum, 22:130-40; Aghassi et al. (1995) Arch Dermatol., 131: 1160-1166; Brennan et al. (1982) Br. J. Rheumatol., 31: 457-460; Kahaleh et al. (1986) Clin. Exp. Rheumatol., 4: 367-369; Falanga and Bucalo (1993) J. Am. Acad. Derm., 29: 47-51; Seyger et al. (1997) J. Am. Acad. Derm., 37: 793-796; Seyger et al. (1998) J. Am. Acad. Dermatol., 39: 220-225; Black (1969) Br. J. Dermatol., 81: 661-666; Ballou et al. (1990) J. Reumatol., 17: 790-794 и Enomoto et al. (1996) J. Am. Acad. Dermatol., 35: 381-387).
Например, с помощью модифицированной балльной оценки Rodnan кожи измеряется твердость кожи с использованием цифрового твердомера типа OO Rex DD-3 (Rex Gauge Company, Buffalo Grove IL) в стандартизированных блоках твердомера с 0,1 единицы разрешением. Измерения твердомером выполняются во всех одинаковых участках кожи по данным измерения балльной оценкой Rodnan кожи. Выполнение балльной оценки кожи и получение показаний твердомера осуществляют при исходном скрининге, перед введением антитела по изобретению и 1 раз в 3 месяца в течение периодов введения и наблюдения. Каждое измерение повторяется 4 раза, и структурированный анализ дисперсии и расчет коэффициентов корреляции между классами используется для определения вариабельности между повторениями в связи с вариабельностью участков и пациентов (Fleiss (1971) Physiol. Bull., 76:378-382). Методики корреляции также используются для оценки согласованности между балльными оценками кожи и балльными оценками твердомера. И для общих балльных оценок, и для подгрупповых балльных оценок в данную точку времени. Выполняются также корреляционные анализы со сдвигом фаз (например, отношение балльных оценок твердомера в начале исследования к балльным оценкам кожи во время t+3 месяца или t+6 месяца лечения антителом). Может также быть собрана информация об активности заболевания и функциональном состоянии, включая данные о синтезе коллагена (измерения PIIINP). Уменьшение симптомов и/или осложнений склеродермии по данным измерения с использованием любых описанных выше способов демонстрирует терапевтическую эффективность антител настоящего изобретения.
Пример 10. Остеоартрит
Антитела по настоящему изобретению испытывают на одной из следующих моделей для демонстрации терапевтической эффективности при остеоартрите. Для следующих примеров концентрация используемого антитела находится в диапазоне примерно от 0,015 до 15 мг антитела на 1 кг массы тела индивидуума; например считается целесообразной дозировка примерно 5 мг антитела на 1 кг массы тела.
Животных, например, 12-недельных самцов мышей C57BL/6, содержат в стандартных клетках и без ограничения кормят стандартным рационом с водопроводной водой.
10.1 Внутрисуставная инъекция AdCTGF в коленные суставы мышей
Конструкт вектора экспрессии аденовируса, содержащий CTGF (AdCTGF), получают, используя систему ADEASY (Qbiogene, Carlsbad CA), в соответствии с процедурами, поставляемыми изготовителем. Вкратце, инсерцию в плазмиду PSHUTTLE-CMV (Obiogene) полинуклеотида, кодирующего человеческий CTGF полной длины, производят, используя стандартные методики молекулярного клонирования. Затем конструкт pShuttle-CMV-CTGF линеаризуют и совместно трансфецируют плазмидой PADEASY-1 (Qbiogene) в компетентные клетки E.coli BJ-5183 электропорацией. AdCTGF амплифицируется и очищается с использованием процедур, описанных Kim et al. (2001, J. Biol. Chem., 276:38781-38786); пустой аденовирусный вектор используется в качестве контроля. Бляшкообразующие единицы (диапазон 1,0-2,1 × 1010/мл) и вирусные частицы (диапазон 0,9-1,5 × 1012/мл) одинаковы для AdCTGF и контрольного вируса.
Производится внутрисуставная инъекция AdCTGF или контрольного вируса (1 × 107 бляшкообразующих единиц) и вводятся антитела согласно изобретению путем внутрисуставной, внутривенной, внутрибрюшинной или подкожной инъекции. Антитело может инъецироваться в то же самое время, что и введение аденовируса, или, альтернативно, терапия может начинаться перед или после инъекции AdCTGF. Животным, получающим контрольный аденовирус, аналогичным образом инъецируют или антитело против CTGF, или контрольное антитело. Коленные суставы, в которые не производилась инъекция, служат в качестве контролей для воздействий антитела.
Коленные суставы выделяют в различные дни, например через 1, 3, 7, 14 и/или 28 дней после инъекции AdCTGF, декальцифицируют в течение 14 дней в EDTA/поливинилпирролидоне и хранят при -20°С, используя процедуру, описанную в публикации Stoop et al. (2001, Osteoarthritis Cartilage, 9:308-315). Исследуют гистологию суставов, чтобы измерить синовиальную толщину и истощение запасов протеогликана; гибридизацию in situ и иммуногистохимическое исследование выполняют для идентификации экспрессии CTGF, а также экспрессии дополнительных факторов, включая коллаген (типа I и/или III) и т.д. Синовиальную жидкость собирают для определения уровней CTGF, металлопротеиназ и т.д. Эффективность терапии с использованием антител против CTGF подтверждается уменьшением параметров, связанных с остеоартритом, при сравнении с животными, которым инъецировали AdCTGF, получавшими лечение контрольным антителом.
10.2 Внутрисуставная инъекция AdTGFβ в мышиные коленные суставы
Альтернативно антитела можно испытать на модели на животных остеоартрита, описанной Bakker et al. (2001, Osteoarthritis Cartilage, 9:128-136). Например, антитело по изобретению или контрольное антитело можно инъецировать в то же время, после или перед внутрисуставной инъекцией 1 × 107 бляшкообразующих единиц экспрессирующего TGFβ аденовирусного конструкта (AdTGFβ). Коленные суставы, в которые не производилась инъекция, служат в качестве контроля эффектов антитела. В различные дни, например 3, 7, 14 день и т.д., животных из каждой группы умерщвляют и ткани выделяют и обрабатывают. Гистологию суставов анализируют для измерения синовиальной толщины, истощения запасов протеогликана и формирование остеофитов; гибридизацию in situ и иммуногистохимическое исследование выполняют для идентификации экспрессии CTGF, а также экспрессии дополнительных факторов, включая коллаген (типа I и/или III) и т.д. Синовиальную жидкость собирают для определения уровней TGFβ, CTGF, металлопротеиназ и т.д. Эффективность терапии с использованием антител по изобретению подтверждается уменьшением параметров, связанных с остеоартритом, при сравнении с животными, которым инъецировали ACTGF, получавшими лечение контрольным антителом.
10.3 Внутрисуставная инъекция папаина в мышиные коленные суставы
Альтернативно антитела можно испытывать, используя процедуру, описанную van der Kraan et al. (1989, Am. J. Pathol., 135:1001-1014). Внутрисуставная инъекция папаина вызывает формирование остеофитов, фиброз и истощение запасов протеогликана в суставном хряще. Папаиновая модель инициируется инъекцией 1 единицы раствора папаина (Sigma, St.Louis, MO) в правый коленный сустав мышей. Левый коленный сустав каждого животного служит в качестве внутреннего контроля. Антитела по изобретению вводят внутрисуставной, внутривенной, внутрибрюшинной или подкожной инъекцией в то же самое время, после или перед внутрисуставной инъекции папаина (0,5%/коленный сустав). В различные дни, например 3, 7, 14 день и т.д., животных из каждой группы умерщвляют и ткани выделяют и обрабатывают. Гистологию суставов анализируют для измерения синовиальной толщины, истощения запасов протеогликана и формирование остеофитов; гибридизацию in situ и иммуногистохимическое исследование выполняют для идентификации экспрессии CTGF, а также экспрессии дополнительных факторов, включая коллаген (типа I и/или III) и т.д. Синовиальную жидкость собирают для определения уровней TGFβ, CTGF, металлопротеиназ и т.д. Эффективность терапии с использованием антител против CTGF подтверждается уменьшением параметров, связанных с остеоартритом, при сравнении с животнымих, которым инъецировали папаин и получавшими лечение контрольным антителом.
Пример 11. Клонирование и экспрессия
Хотя следующие примеры иллюстрируют клонирование и экспрессию одного конкретного антитела по изобретению, способы в целом применимы для всех антител, описанных и заявленных в настоящем описании.
Иллюстративное антитело по изобретению, mAb1, было сначала идентифицировано как часть сложного человеческого антитела, секретируемого линией клеток гибридомы (8C12-F10; полученной, как описано в примере 3).
11.1 Клонирование и секвенирование тяжелой цепи mAb1
Матричную РНК изолировали из культуры клеток 8C12-F10, используя набор MICRO-FAST TRACK (Invitrogen), следуя протоколу, предоставленному изготовителем. Затем получали 2 пула кДНК синтезом второй нити, используя набор клеточного цикла кДНК (Invitrogen), следуя протоколу, предоставленному изготовителем, и одну из следующих антисмысловых затравок тяжелой цепи:
Последовательности вариабельной области тяжелой цепи клонировали PCR амплификацией примированного АВ90 пула кДНК, используя затравку АВ90 и одну из серии затравок области V, включая затравки, соответствующие сохраненным секреторным сигнальным последовательностям, которые кодируют 5'-конец соответствующих кодирующих областей, и последовательности каркасной области 1, которые кодируют начало зрелых иммуноглобулинов. Бляшкообразующую единицу ДНК-полимеразы (Stratagene) использовали в соответствии с рекомендуемыми протоколами изготовителя со следующими изменениями: Реакции обычно проводили в общем объеме 50 мкл, содержащем 1 мкл кДНК, по 0,75 мкМ каждой прямой и обратной затравки, по 200 мкМ каждой dNTP и 1 мкл бляшкообразующей единицы ДНК-полимеразы (2,5 единиц на 1 мкл). Использовали программу обратного отсчета термического датчика циклов при первоначальной инкубации при 94°С в течение 2 мин перед добавлением фермента. Затем использовали следующие параметры циклов: 10 циклов при 94°С в течение 45 сек, при 65°С в течение 45 сек и при 72°С в течение 1 мин; 30 циклов при 94°С в течение 45 сек, при 55°С в течение 45 сек и при 72°С в течение 1 мин; и затем 1 цикл при 72°С в течение 10 мин.
Только одна затравка сигнальной последовательности тяжелой цепи, AB87 (ATGGAGTTTGGRCTGAGCTG; SEQ ID NO:5), которая связывается с областями V тяжелой цепи семейства VH3, давала значительный продукт. Продукт PCR нуклеотида 453 клонировали в вектор PCR BLUNT II-TOPO (Invitrogen), проводили скрининг клонов для определения правильного размера вставки и проводили секвенирование трех клонов, соответствующих продуктам PCR. Идентичные последовательности получали для всех трех клонов.
Последовательности константной области тяжелой цепи и области UTR клонировали PCR амплификацией пула кДНК, примированного m19 H1504R. Нуклеотид 601 фрагмента PCR, соответствующий 5'-концу сегмента тяжелой цепи, амплифицировали, используя смысловую затравку VH3-33 29-51F (CGGCGGTGTTTCCATTCGGTGAT; SEQ ID NO:6) и антисмысловую затравку константной области тяжелой цепи m19 H553R (GGGCGCCTGAGTTCCACGACAC; SEQ ID NO:7). Опосредованное топоизомеразой клонирование использовали для клонирования продуктов PCR в вектор PCR-BLUNT II (Invitrogen) в соответствии с инструкциями изготовителя, и затем проводили секвенирование вставки. Аналогичным образом амплифицировали фрагмент PCR нуклеотида 505, используя смысловую затравку m19 H439F (GTCTTCCCCCTGGCACCCTCCTC; SEQ ID NO:8) и антисмысловую затравку m19 H943R (CCCGCGGCTTTGTCTTGGCATTAT; SEQ ID NO:9), и фрагмент PCR нуклеотида 503, используя смысловую затравку m19 H1002F (CTGGCTGAATGGCAAGGAGTA; SEQ ID NO:10) и антисмысловую затравку m19 H1504R. Оба фрагмента отдельно клонировали в вектор PCR-BLUNT II (Invitrogen) и секвенировали, как описано выше. Четвертый фрагмент PCR тяжелой цепи нуклеотидов 586 амплифицировали, используя смысловую затравку m19 H645F (GGGCACCCAGACCTACATC; SEQ ID NO:11) и антисмысловую затравку m19 H1230R (CTCCGGCTGCCCATTGCTCTCC; SEQ ID NO:12) и проводили прямое секвенирование.
На фиг.11А показана схема упорядочения клонированных фрагментов, которые обеспечили нуклеотидную последовательность полной длины (SEQ ID NO:13), которая кодирует тяжелую цепь mAb1 (SEQ ID NO:14). Аминокислотная последовательность вариабельной области тяжелой цепи ближе всего напоминает ген DR-44 зародышевой линии VH3. Хотя было невозможно сказать, какой сегмент D использовался, последовательность mAb1 ближе всего напоминает семейство DH4. Область JH ближе всего соответствует зародышевой линии JH4 и JH5. Константная область тяжелой цепи mAb1 соответствует обозначенной номером доступа ВС016381 в Генном банке, указывая аллотип G1m(3).
11.2 Клонирование и секвенирование легкой цепи mAb1
Матричную РНК выделяли из культуры клеток 8C12-F10, используя набор MICRO-FAST TRACK (Invitrogen), следуя последовательности операций, предоставленной изготовителем. Затем пулы кДНК получали при синтезе второй нити, используя набор клеточного цикла кДНК (Invitrogen), следуя протоколам, предоставленным изготовителем, и одну из следующих затравок антисмысловой кДНК легкой цепи:
Последовательности вариабельной области легкой цепи клонировали амплификацией PCR примированного АВ16 пула кДНК, используя затравку АВ16 и одну из серии затравок области V, включая затравки, соответствующие сохраненным секреторным сигнальным последовательностям, которые кодируют 5'-конец соответствующих колирующих областей, и последовательности каркасной области 1, которые кодируют начало рекомендуемого иммуноглобулина. Бляшкообразующую единицу ДНК-полимеразы (Stratagene) использовали в соответствии с последовательностями операций, рекомендуемыми изготовителем, с описанными выше изменениями и параметрами циклов.
В случае только одной затравки сигнальной последовательности легкой цепи, АВ123 (CCCGCTCAGCTCCTGGGGCTCCTG; SEQ ID NO:17), которая связывается с областями V тяжелой цепи семейства VK1, продуцировалось значительное количество продукта. Продукт PCR нуклеотида 408 клонировали в вектор PCR BLUNT II-TOPO (Invitrogen), проводили скрининг клонов для выявления правильного размера вставки и проводили секвенирование трех клонов, соответствующих продуктам PCR. Идентичные последовательности были получены для всех трех клонов.
Последовательности константной области легкой цепи клонировали амплификацией PCR пула кДНК, примированного Ck-760R. Всю кодирующую последовательность и 5'UTR-область легкой цепи амплифицировали, используя смысловую затравку легкой цепи L15 22m (TCAGWCYCAGTCAGGACACAGC; SEQ ID NO:18) и Ck-760R. Нуклеотидный фрагмент 788 клонировали в вектор PCR BLUNT II (Invitrogen) и секвенировали. Полученную плазмиду обозначили 41m6.
На фиг.11В представлена схема упорядочения клонированных фрагментов PCR, которое обеспечило нуклеотидную последовательность полной длины (SEQ ID NO:19), которая кодирует легкую цепь mAb1 (SEQ ID NO:20). Аминокислотная последовательность вариабельной области легкой цепи ближе всего соответствует областям, кодированным нуклеотидными последовательностями L15 и Jk2 зародышевой линией Vk. Константная область легкой цепи mAb1 идентична известной генной последовательности иммуноглобулина каппа легкой цепи человеческой зародышевой линии (Whitehurst et al. (1992) Nucleic Acids Res 20:4929-4930).
11.3 Продукция конструктов экспрессии тяжелой и легкой цепи
кДНК тяжелой цепи mAb1 полной длины генерировали перекрывающим распространением PCR в два этапа из описанных выше и показанных на фиг.11А продуктов PCR тяжелой цепи. Два 5'-продукта PCR комбинировали с дистальными затравками VH3-33 29-51F и m19 H943R в реакции перекрывающего распространения PCR для получения одного фрагмента из 991 нуклеотида. Аналогичным образом два 3'-продукта PCR комбинировали с дистальными затравками VH3-33 29-51F и m19 H943R в реакции перекрывающего распространения PCR для получения фрагмента из 860 нуклеотидов. Эти продукты реакции распространения PCR были очищены на геле и амплифицированы вместе с использованием дистальных затравок VH3-33 29-51F и m19 H1504R для генерирования кодирующей последовательности кДНК нуклеотида 1407 (остатки 441-1847 SEQ ID NO:13) тяжелой цепи mAb1 полной длины.
кДНК тяжелой цепи затем клонировали в вектор PCR-BLUNT II TOPO (Invitrogen) для получения плазмиды 43а4. Кодирующую область тяжелой цепи mAb1 затем субклонировали расщеплением плазмиды 43а4 рестрикционными эндонуклеазами BamH1 и Xbal с последующим лигированием иссеченной вставки в вектор экспрессии PCDNA5-FRT (Invitrogen), который был предварительно переварен рестрикционными эндонуклеазами BamH1 и Nhe. Последовательность вставки полученной плазмиды экспрессии, 44а1, верифицировали перед осуществлением аналогичного субклонирования в обратной ориентации в вектор PBK-CMV (Clontech) для получения плазмиды 47а4, и в вектор pCEP-Pu (E.Kohfeldt, Max-Planck-Institut fur Biochemie), вектор, полученный из вектора рСЕР4 (Invitrogen), для получения плазмиды 49а1.
кДНК нуклеотида 708 (остатки 415-1122 SEQ ID NO:19), кодирующего легкую цепь mAb1 полной длины, иссекали из плазмиды 41m6, описанной выше, используя рестрикционные эндонуклеазы HindIII и Xho I, и лигировали в вектор PCDNA5-FRT (Invitrogen), который был предварительно расщеплен рестрикционными эндонуклеазами HindIII и Xho I, для получения экспрессионной плазмиды млекопитающих 42b2. Перед осуществлением аналогичного субклонирования в обратной ориентации в вектор PBK-CMV (Clontech) для получения плазмиды 47b3 и в вектор pCEP-Pu (E.Kohfeldt, Max-Planck-Institut fur Biochemie) для получения плазмиды 49b1 верифицировали последовательность вставки плазмиды 42b2.
11.4 Трансфекция и экспрессия конструктов цепи антитела
Клетки COS7 трансфецировали плазмидами 44а1 (тяжелая цепь mAb1) и 42b2 (легкая цепь mAb1) как путем отдельной, так и совместной трансфекции, используя стандартные процедуры. Кондиционированные культуральные среды анализировали для выявления присутствия антитела, как описано в примере 4 (выше). Только в среде клеток, совместно трансфецированных как 44а1, так и 42b2, экспрессировалось человеческое антитело, обладающее активностью связывания CTGF, что было измерено ELISA с использованием описанных выше процедур. Антитело, идентифицируемое в настоящем описании как CLN1, продуцируемое совместно трансфецированными клетками COS7, связывается с N-концевой половиной CTGF с аффинностью 0,8 нМ.
CLN1 также экспрессировалось у генетически модифицированных клеток яичников китайских хомячков (СНО). Линия клеток СНО, экспрессирующих антитело CLN1, депонирована в Американскую коллекцию типовых культур (Manassas VA) 20 мая 2004 г. и обозначена номером доступа PTA-6006. Линии клеток можно оптимизировать и экспрессию антитела можно усилить, используя различные методики, известные в данной области, например, путем генной амплификацией, как описано Wigler et al. (1980; Proc. Natl. Acad. Sci. USA, 77:3567-3570), с модификациями, как описано у Ringold et al. (1981; J. Mol. Appl. Genet., 1: 165-175), Gasser et al (1982; Proc. Natl. Acad. Sci. USA, 79: 6522-6526) и Kaufman et al. (1985; Mol. Cell Biol., 5:1750-1759).
Пример 12: Взаимодействие CTGF с TGFβ
Антитела по изобретению специфически связываются с областями CTGF, определяемыми остатками, кодируемыми экзоном 3 (фиг.1В; нуклеотиды 418-669 SEQ ID NO:1). Эта область охватывает аминокислоты 97-180 SEQ ID NO:2 и включает домен фон Виллебранда типа С (аминокислоты 103-164 SEQ ID NO:2) и эпитоп mAb1 (аминокислоты 134-158 SEQ ID NO:2). Abreu et al. (2002, Nat. Cell Biol., 4:599-604) сообщают о том, что домен, соответствующий домену VWC CTGF, важен для взаимодействия между CTGF и TGFβ или BMP-4 и что указанное взаимодействие модулирует активность TGFβ или BMP-4. Следующие эксперименты демонстрируют, что области, кодируемые экзоном 3, необходимы и достаточны для связывания CTGF с TGFβ, и что антитела по изобретению могут блокировать взаимодействие между CTGF и TGFβ.
Взаимодействие между CTGF и TGFβ анализировали с использованием следующей процедуры. Ячейки 96-луночного планшета MAXISORP ELISA (Nalge Nunc) покрывали на ночь при 4°С 10 мкг/мл либо CTGF, фрагментом CTGF, кодируемым экзоном 3, либо фрагментом CTGF, кодируемым экзоном 5, в PBS; или только PBS. Все лунки затем блокировали 1% BSA в PBS с последующей инкубацией в течение 1 ч при комнатной температуре в 50 мкл раствора, содержащего TGFβ в концентрации 0, 1, 3,3, 10, 33, 100, 333 или 1000 нг/мл, и мышиное моноклональное антитело MAB612 или MAB1835 (R&D Systems, Minneapolis MN) в концентрации 100, 300 или 1000 нг/мл в PBS, 0,05% Tween-20. MAB1835 распознает бычий, мышиный и человеческий TGF-β1 и -β2 и блокирует связывание TGFβ с мышиными тимоцитами. МАВ612 распознает TGF-β2, но не ингибирует активность TGFβ. Лунки промывали PBS, 0,05% Tween-20 и затем инкубировали в течение 1 ч при комнатной температуре в растворе, содержащем конъюгированное щелочной фосфатазой козье антитело против мышиного IgG, разведенное в PBS, 0,05% Tween-20. Планшеты снова промывали и добавляли п-нитрофенилфосфат (PNPP) в 1 М этаноламине, 1 мМ MgSO4, pH 9,8, лунки инкубировали в течение подходящего времени для проявления, и реакцию затем прекращали добавлением NaOH. Измеряли поглощение при λ, равной 405 нм, используя спектрофотометр.
На фиг.12 показано, что CTGF и фрагмент CTGF, кодируемый экзоном 3, способны в эквивалентной степени взаимодействовать с TGFβ, в то время как фрагмент CTGF, кодируемый экзоном 5, не проявлял какой-либо связывающей активности в отношении TGFβ. Интересно, что антитело против TGFβ МАВ612 было также способно выявлять связанный с CTGF TGFβ зависимым от дозы образом, но нейтрализующее антитело, МАВ1835, не позволяло выявлять связанный с CTGF TGFβ в любой испытанной концентрации (данные не показаны). Это свидетельствует о том, что CTGF конкурирует с МАВ1835 за связывание с TGFβ.
Антитела против CTGF были испытаны для выявления их способности блокировать связывание между CTGF и TGFβ. Как показано на фиг.12, антитела согласно изобретению, проиллюстрированные mAb4 и mAb1, блокировали связывание TGFβ как с CTGF, так и с фрагментом CTGF, кодируемым экзоном 3, в то время как антитело против CTGF, направленное на С-концевой фрагмент CTGF, не блокировало связывания. Эти результаты подтверждают механизм действия, при котором антитела согласно изобретению специфически блокируют взаимодействие между CTGF и TGFβ и потенциально между CTGF и другими членами суперсемейства TGFβ.
Различные модификации изобретения в дополнение к тем, которые представлены и описаны в настоящем описании, станут очевидными для специалистов в данной области из предшествующего описания. Такие модификации входят в объем прилагаемой формулы изобретения.
Все ссылки, приведенные здесь, включены в полном объеме в качестве ссылки.












| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТ ИНТЕГРИН АЛЬФА-V БЕТА-8 | 2012 |
|
RU2614252C2 |
| АНТИТЕЛА, НЕЙТРАЛИЗУЮЩИЕ ИНТЕГРИН ανβ8 | 2011 |
|
RU2565539C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ ИНТЕРЛЕЙКИНА-33 (IL-33) | 2015 |
|
RU2693084C2 |
| АНТИТЕЛА ПРОТИВ CCR2 | 2009 |
|
RU2547595C2 |
| СКОНСТРУИРОВАННЫЕ АНТИ-TGF-БЕТА АНТИТЕЛА И АНТИГЕН-СВЯЗЫВАЮЩИЕ ФРАГМЕНТЫ | 2014 |
|
RU2681502C2 |
| СВЯЗЫВАЮЩИЕ KIR3DL2 АГЕНТЫ | 2013 |
|
RU2682449C2 |
| Антитела против эотаксина 2, которые распознают дополнительные связывающие CCR3 хемокины | 2015 |
|
RU2705255C2 |
| АНТИТЕЛА ЧЕЛОВЕКА ПРОТИВ СЕМАФОРИНА 4D | 2018 |
|
RU2776443C2 |
| ДИАГНОСТИЧЕСКИЕ АНАЛИЗЫ И НАБОРЫ ДЛЯ ДЕТЕКЦИИ ФОЛАТНОГО РЕЦЕПТОРА 1 | 2013 |
|
RU2668824C2 |
| АНТИТЕЛА ПРОТИВ GDF8 ЧЕЛОВЕКА | 2011 |
|
RU2710156C2 |
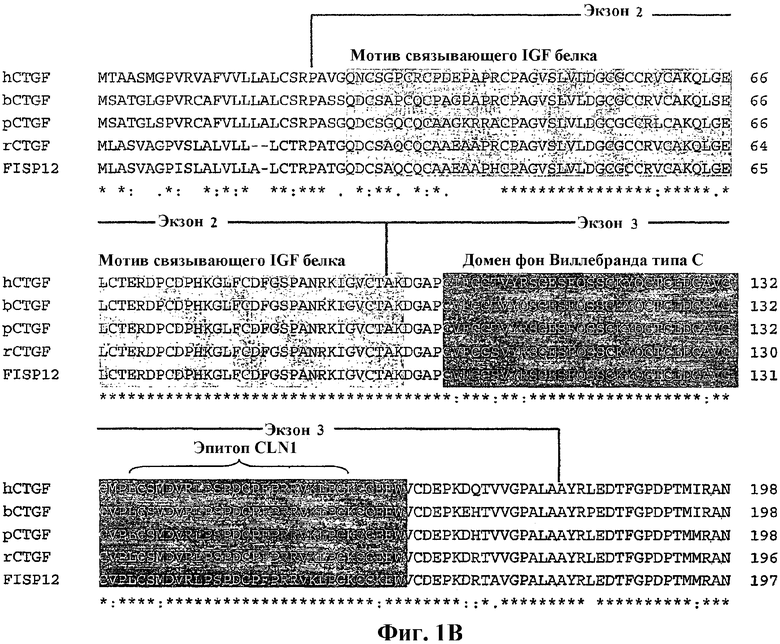
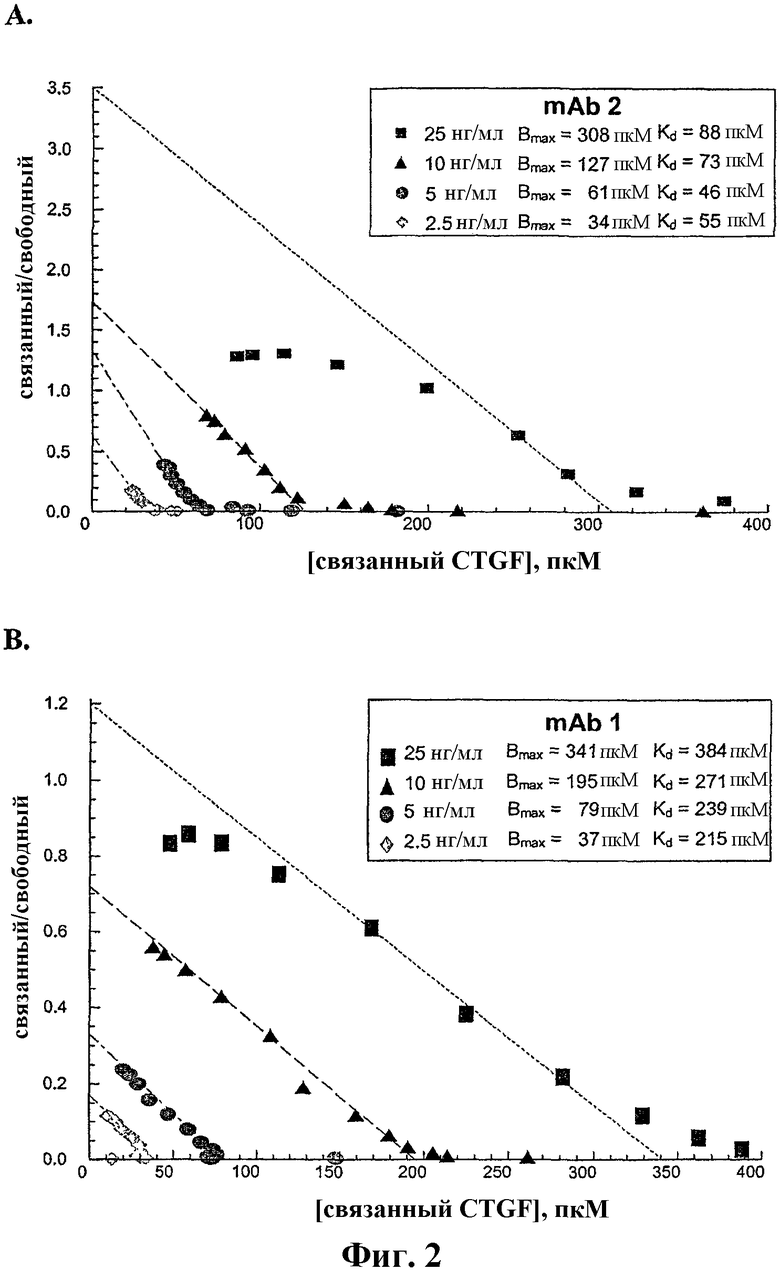
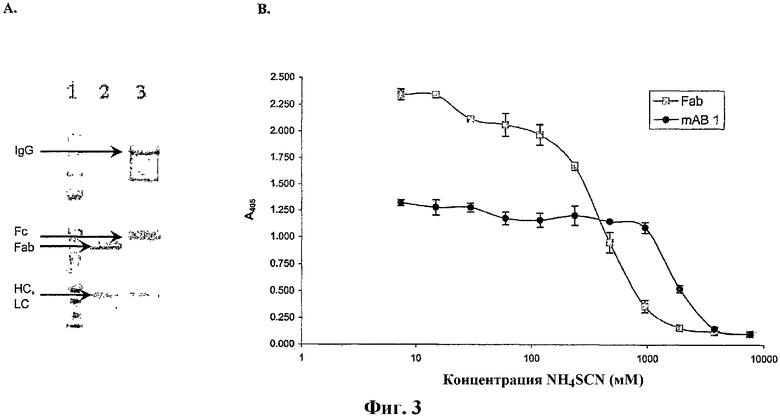
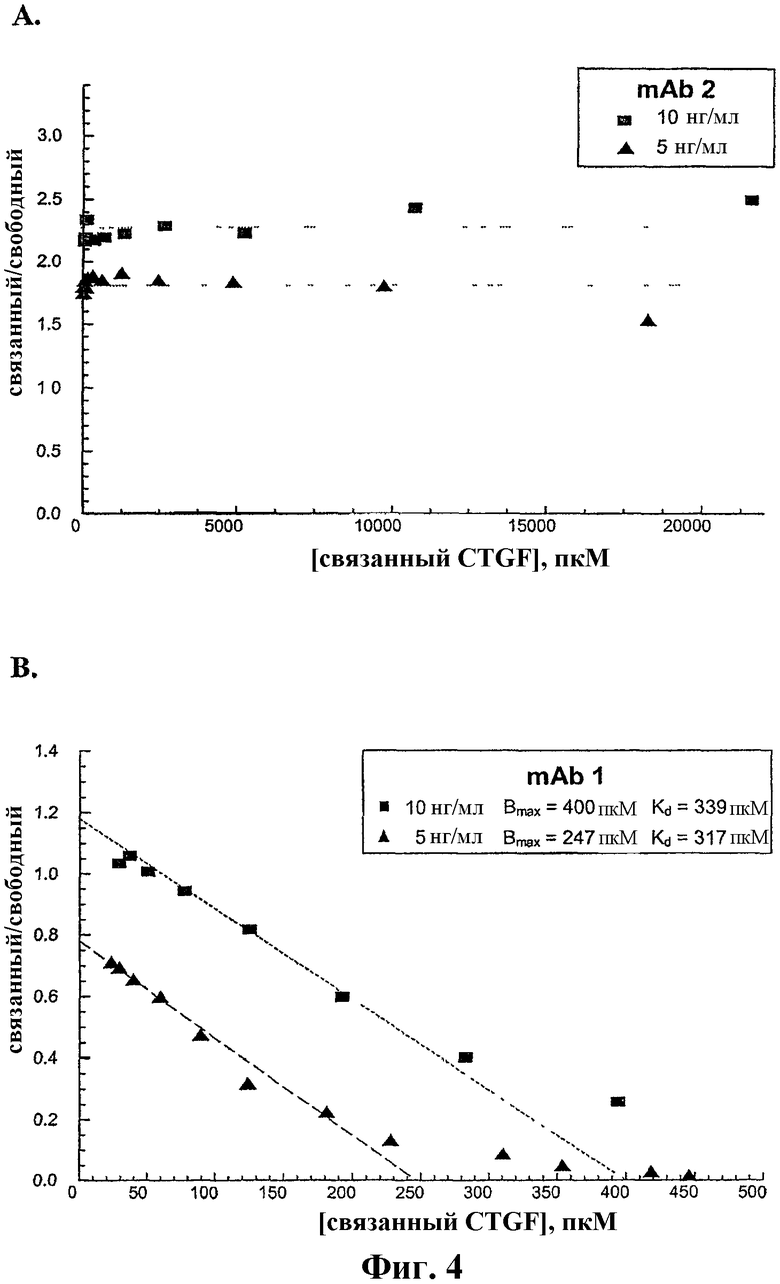
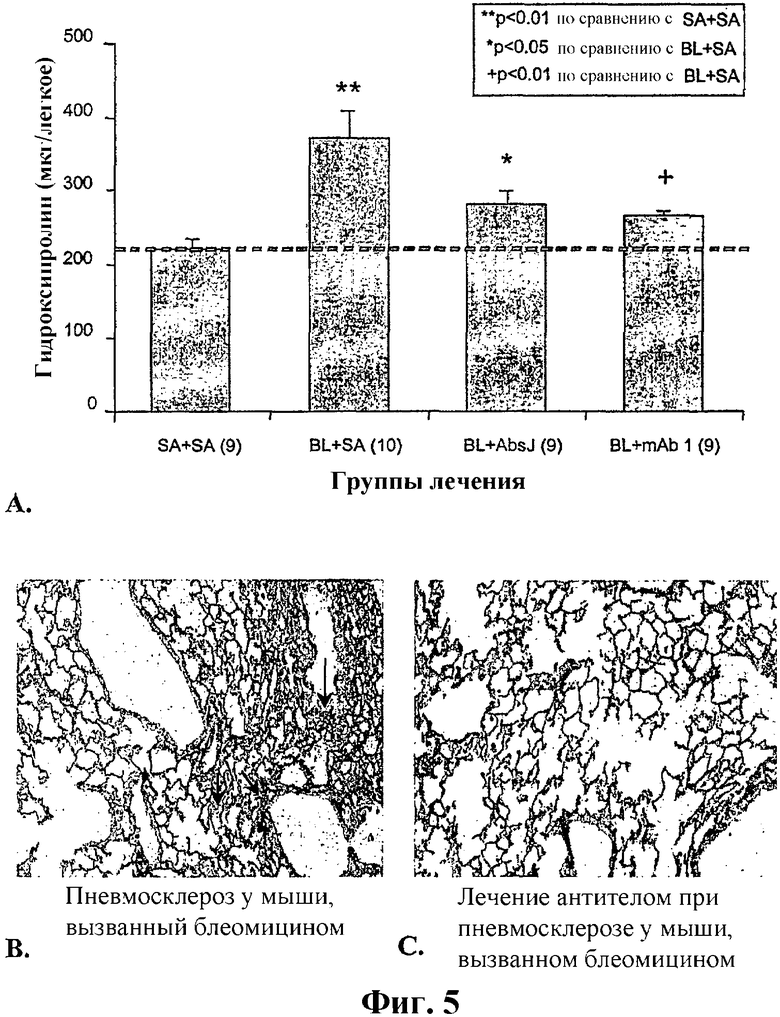
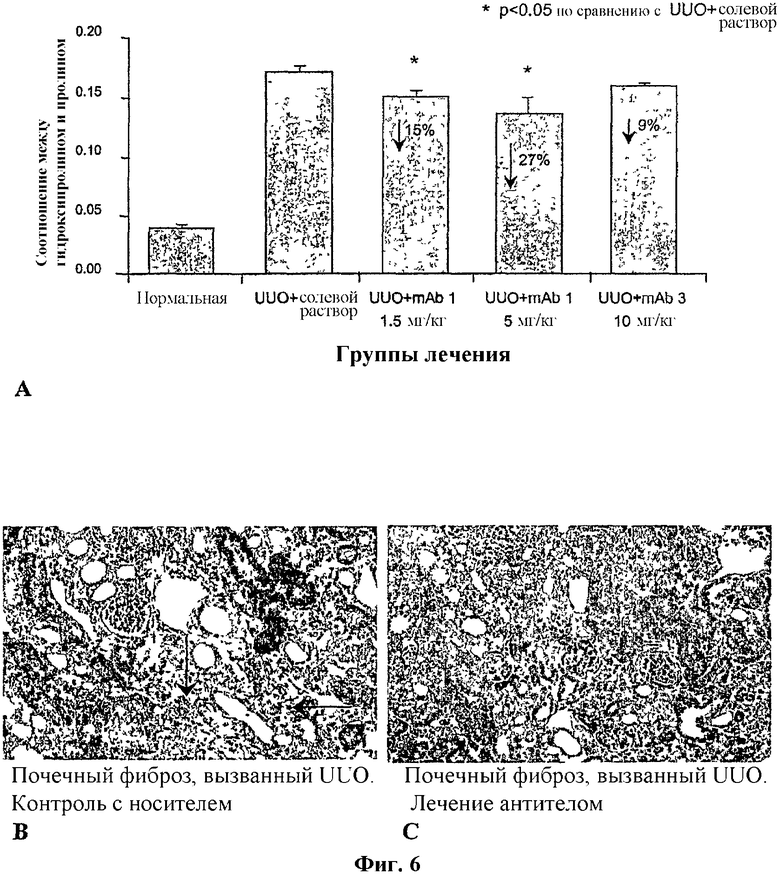

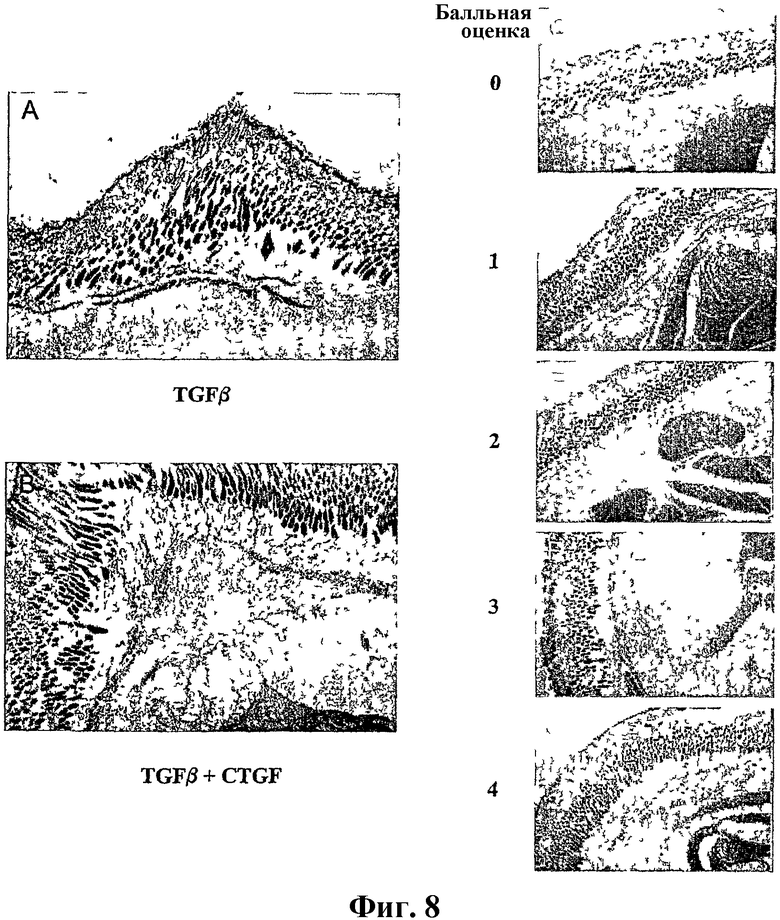
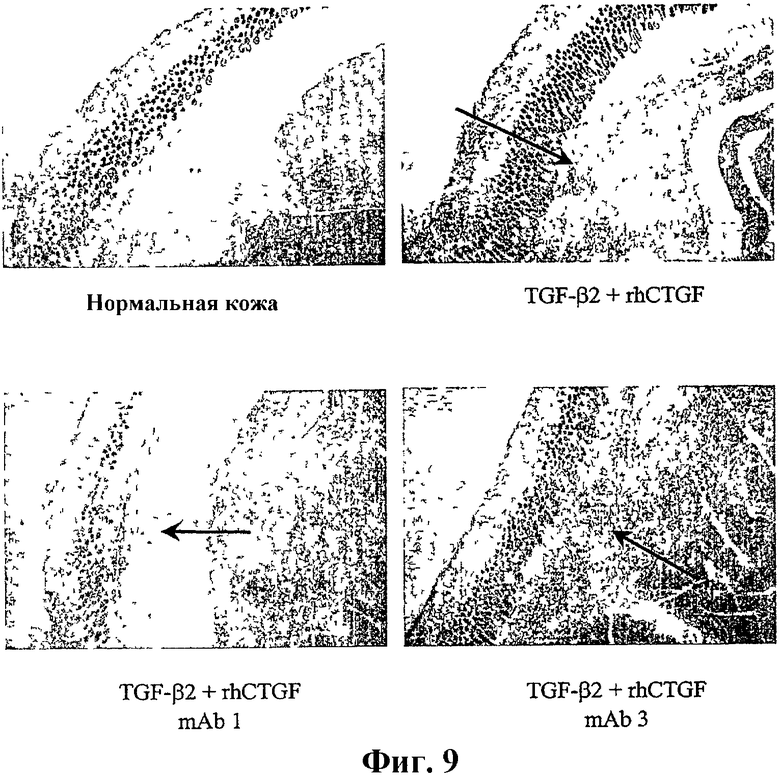
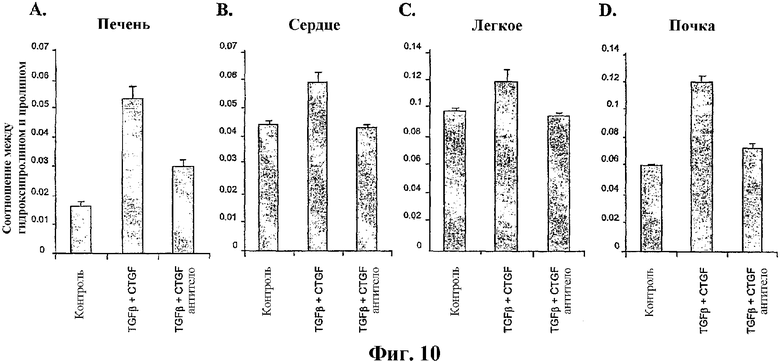
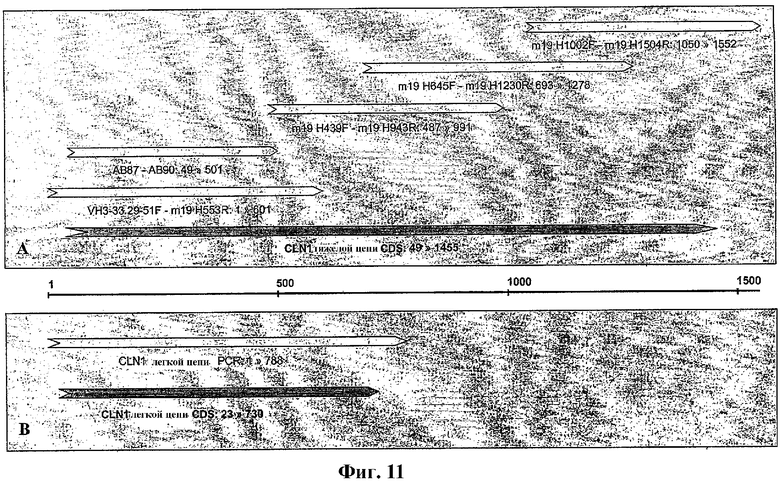
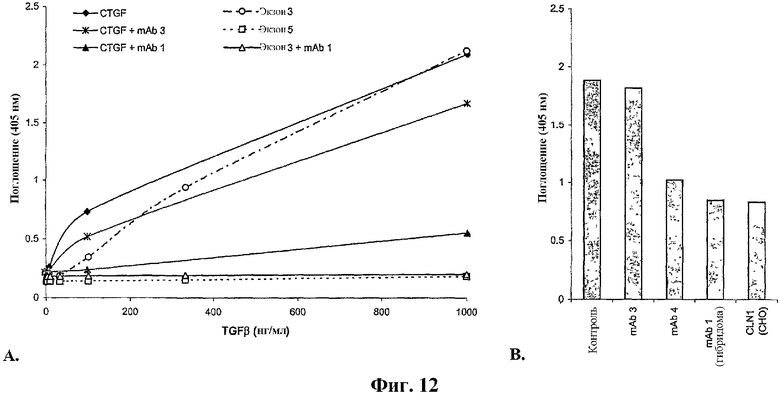
Изобретение относится к антителам, которые связываются с CTGF. Антитела, в частности, направлены на области CTGF, участвующие в различных видах биологической активности, связанной с фиброзом. Изобретение также относится к способам применения антител в составе фармацевтических композиций для лечения расстройств, связанных с CTGF, включая локализованные и системные фибротические расстройства, в том числе расстройства легких, печени, сердца, кожи и почек, для осуществления способа нейтрализации биологической активности CTGF и способа лечения или профилактики заболеваний, связанных с CTGF. Изобретение охватывает полинуклеотидную последовательность и ее варианты, кодирующую указанное антитело, а также клетку-хозяин и клеточную линию №РТА-6006 (АТСС), продуцирующие указанное антитело. Использование изобретения обеспечит новыми специфическими средствами - антителами, которые эффективно нейтрализуют конкретные виды активности CTGF при патологии и обеспечивают специфичность и фармакинетический профиль, подходящий для терапевтического средства. 12 н. и 69 з.п. ф-лы, 12 ил., 4 табл.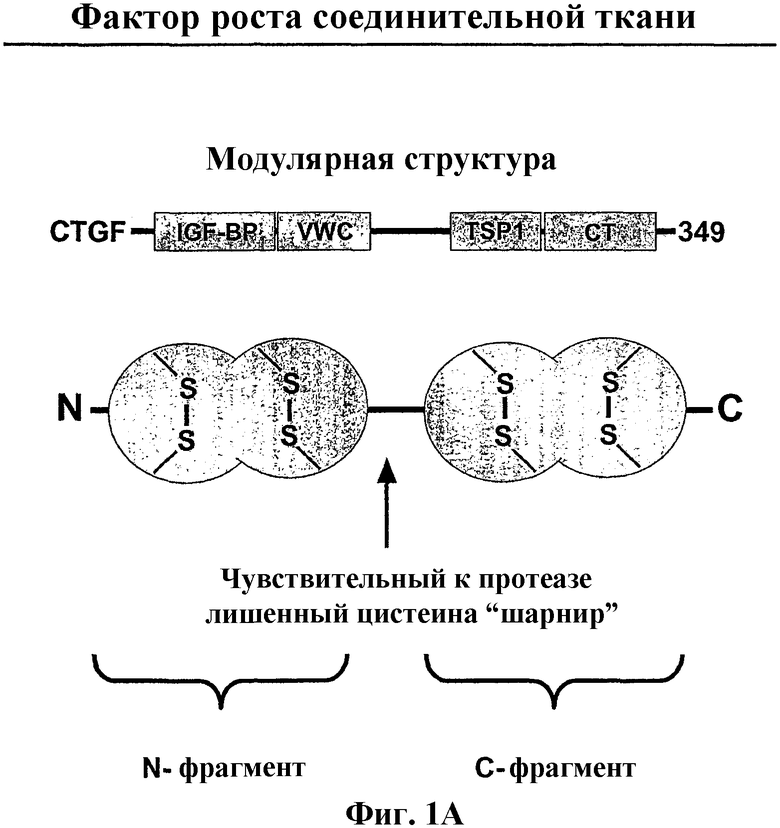
(a) иммуноглобулина с аминокислотной последовательностью SEQ ID NO: 14;
(b) иммуноглобулина с аминокислотной последовательностью SEQ ID NO: 20;
(c) фрагмента иммуноглобулина, содержащего вариабельный домен (а) или (b); и
(d) консервативных вариантов (а), (b) или (с).
(a) полинуклеотидной последовательности, кодирующей последовательность тяжелой цепи иммуноглобулина SEQ ID NO: 14;
(b) полинуклеотидной последовательности, кодирующей вариабельный домен тяжелой цепи иммуноглобулина, соответствующей аминокислотам 1-167 SEQ ID NO: 14;
(c) последовательности SEQ ID NO: 13, кодирующей тяжелую цепь иммуноглобулина; и
(d) нуклеотидной последовательности 1-501 SEQ ID NO: 13, кодирующей вариабельный домен тяжелой цепи иммуноглобулина; или полинуклеотидной последовательности, содержащей последовательность, выбранную из группы, состоящей из
(а) полинуклеотидной последовательности, кодирующей последовательность легкой цепи иммуноглобулина SEQ ID NO: 20;
(b) полинуклеотидной последовательности, кодирующей вариабельный домен легкой цепи иммуноглобулина, соответствующей аминокислотам 1-136 SEQ ID NO: 20;
(c) последовательности SEQ ID NO: 19; кодирующей легкую цепь иммуноглобулина; и
(d) нуклеотидной последовательности 1-408 SEQ ID NO: 19, кодирующей вариабельный домен легкой цепи иммуноглобулина.
Приоритет по пунктам:
| WO 9913910 A1, 25.03.1999 | |||
| Устройство для улавливания рыбьих чешуи и жира | 1933 |
|
SU35936A1 |
| РОЙТ А | |||
| и др | |||
| ИММУНОЛОГИЯ | |||
| -М.: Мир, 2000, с.153-155. | |||

