Изобретение относится к медицине, конкретно к экспериментальной гинекологии, и касается способов моделирования поликистозных яичников. (Здесь и далее термин "поликистозные яичники" следует рассматривать как собирательное понятие, служащее для обозначения всех яичников с кистами, возникающими, так или иначе, из фолликула [1].)
Известны способы моделирования поликистозных яичников путём субтотальной или частичной кастрации [1-5], введения стероидных гормонов [3, 6-13], воздействия сильными раздражителями [3, 6, 14-16].
Сущность моделей, полученных путём овариоэктомии, заключается в первичном снижении функции яичников, а именно в снижении продукции эстрогенов и определённом нарушении взаимоотношения между яичником и гипофизом: в ответ на падение уровня эстрогенов по механизму отрицательной обратной связи гипофиз начинает выделять повышенные количества гонадотропинов, причём соотношение между лютеинизирующим гормоном (ЛГ) и фолликулостимулирующим гормоном (ФСГ) меняется в пользу последнего. Повышенная, но недостаточная для овуляции, секреция ЛГ вместо овуляции вызывает процессы атрезии фолликулов, лютеинизацию тека-ткани и развитие множественных фолликулярных кист. Причём кисты яичников впервые обнаруживаются через 4-5, а по некоторым данным через 8-12 месяцев. Частота возникновения кист редко превышает 75%. Возникшие кисты чаще всего являются фолликулярными, реже лютеиновыми, иногда возникают другие типы кист [1].
Основными методиками с использованием половых гормонов являются неонатальная андрогенизация (введение новорождённым крысятам-самкам тестостерона пропионата) и экзогенная эстрогенизация (введение половозрелым крысам эстрадиола валерата). В первом случае ранняя андрогенизация самок не влияет на обратные отрицательные связи со стороны половых гормонов, но она резко отражается на реализации положительных обратных связей, так как гипоталамус становится рефракторным к стимулирующему действию половых гормонов. Вместе с тем, сохранившаяся тоническая секреция гонадотропинов вызывает различной степени созревание фолликулов, выделяющих эстрогены. В условиях повышенной, но без овуляторного подъёма концентрации ЛГ в яичниках могут развиться только поликистозные изменения [6-8].
Во втором случае экзогенное введение эстрогенов вызывает снижение секреции эндогенных эстрогенов, что ведёт к увеличению секреции ФСГ в течение длительного времени (многих потенциальных циклов). Повышенный уровень ФСГ гиперстимулирует рост фолликулов, причём без цикличности, непрерывно и при явном недостатке ЛГ. Следовательно, непрерывно растут фолликулы без овуляции, что закономерно приводит к развитию фолликулярных кист, непрерывно секретирующих эстрогены в течение длительного времени. Повышенная секреция эстрогенов угнетает секрецию ФСГ и приводит к стимуляции выработки ЛГ, замыкая порочный круг [3, 7, 17].
С использованием вышеперечисленных методов кисты яичников удаётся получить в 35-100% случаев по данным различных авторов, сроки формирования также колеблются в широких пределах - от 20 дней до 3,5 месяцев. При микроскопии превалируют мелкие атретические фолликулы, небольшие по величине кисты, чаще имеющие строение фолликулярных, реже встречаются другие типы кист.
Среди методов с использованием сильных раздражителей наиболее часто применяется круглосуточное электрическое освещение крыс-самок в течение 50-150 дней. В результате возникают гипоталамические изменения функционального характера, проявляющиеся нарушением цикличности выработки гормонов и отсутствием овуляции. В яичниках наступает поликистозная дегенерация [3, 6, 7, 14].
Кисты яичников впервые могут быть обнаружены только к 50-100 суткам электрического освещения. Частота образования достаточно высока - до 100%. В ранние сроки наиболее часто встречаются мелкие кистозно-атрезированные фолликулы, в более поздние формируются крупные многокамерные кисты, выстланные плоским эпителием.
Вышеперечисленные методики позволяют создать модель поликистозных яичников, оценить воздействие различных фармакологических препаратов и хирургического лечения. Однако данные методики не лишены недостатков. Так, при моделировании поликистозных яичников необходимо достаточно длительное время для формирования данной патологии, от 3-х недель до 12 месяцев. Большинство методик обладают низкой стабильностью воспроизведения, за исключением электрического освещения. Вместе с этим данные модели не учитывают все звенья патогенеза формирования данной патологии, в частности влияния гиперинсулинемии и действия инсулиноподобных факторов роста (ИПФР). Следовательно, необходимо разработать в опытах на животных такой метод индукции фолликулярных кист яичников, который позволил бы закономерно вызывать их развитие, и в то же время, чтобы условия возникновения таких кист в эксперименте были близки к таким, которые встречаются в организме человека.
Наиболее близким к предлагаемому является способ моделирования поликистозных яичников, предложенный К.Bogovich, J.demons, L.Poretsky [18,19]. В одной из серий экспериментов авторы используют крыс-самок в возрасте 85 дней, которым вводят субовуляторную дозу человеческого хорионического гонадотропина (ХГ) (1,5-6 ME) дважды в день и дозу инсулина, способную вызвать гиперинсулинемию (1,5-9 ЕД в сутки) в течение 22 дней. Двусторонние поликистозные яичники выявляются на 23 день эксперимента, преобладают фолликулярные кисты. Частота образования кист колеблется в пределах 64-80%.
Среди недостатков следует отметить относительно длительные сроки для формирования модели, невысокую стабильность воспроизведения. Кроме того, рядом авторов высказано предположение, что в формировании фолликулярных кист яичников у женщин имеет значение кратковременная гиперстимуляция тека-клеток суперовуляторными концентрациями ЛГ. Вместе с этим, доказано, что предельная гиперинсулинемия имеет парадоксальную способность уменьшать индукцию овариальных кист ХГ.
Задачей, решаемой данным изобретением, является приближение патоморфологических изменений, патогенетических особенностей (условий формирования) множественных фолликулярных кист яичников в эксперименте к таким, которые имеют значение в клинике, а также сокращение сроков моделирования, повышение стабильности воспроизведения и расширение области использования данной модели.
Поставленная задача решается новым способом моделирования поликистозных яичников с преобладанием фолликулярных кист путем внутримышечного введения беспородным половозрелым крысам-самкам, находящимся в фазах покоя эстрального цикла, ХГ и инсулина, причём вводят ХГ в дозе 300 ЕД, протафан НМ в дозе 2,5 ЕД ежедневно в первой половине дня в течение 7 дней.
Новым является то, что вводят ХГ в дозе 300 ЕД, протафан НМ в дозе 2,5 ЕД ежедневно в первой половине дня в течение 7 дней.
Данные отличительные признаки не вытекают явным образом для специалиста из уровня техники и позволяют достичь новых положительных эффектов, а именно - приближения патоморфологических изменений, патогенетических особенностей (условий формирования) множественных фолликулярных кист яичников в эксперименте к таким, которые имеют значение в клинике, а также сокращения сроков моделирования, повышения стабильности воспроизведения и расширения области использования данной модели.
Предлагаемый способ апробирован на 24-х беспородных половозрелых белых крысах-самках в отделе экспериментальной физиологии и хирургии Центральной научно-исследовательской лаборатории Сибирского государственного медицинского университета. Эксперимент согласован с этическим комитетом ГОУВПО СибГМУ (протокол № 15 от 26.01.2004 г.). Таким образом, техническое решение соответствует критериям изобретения "новизна", "изобретательский уровень", "промышленная применимость".
Способ осуществляют следующим образом.
В качестве экспериментального материала используют половозрелых белых беспородных крыс-самок (массой 200-220 г), находящихся в фазах покоя (метэструс, диэструс) астрального цикла, который контролируется кольпоцитологически. Крыс фиксируют в положении на спине. При помощи инсулинового шприца внутримышечно в ягодичную область, исключая ранение магистральных сосудов и нервных пучков, вводят суперовуляторную дозу ХГ (300 ЕД), разведённого 0,3 мл раствора для инъекций, приложенного к ампуле ХГ, раствор приготавливают ex tempore. Аналогичным образом вводят 2,5 ЕД протафана НМ - дозу, необходимую для достижения умеренной гиперинсулинемии, препарат разводят 0,05 мл воды для инъекций, раствор приготавливают ex tempore. Указанные препараты вводят ежедневно в течение 7 дней в первой половине дня до 12 часов.
Экспериментальных животных содержат в стандартных условиях вивария на обычном питании с добавлением овощей и творога, при дозированном освещении (12:12, свет с 8 часов). Ежедневно всех животных осматривают, отмечают их общее состояние, внешний вид, поведение, пищевую возбудимость и двигательную активность, измеряют массу тела и проводят кольпоцитологическое исследование для диагностики фазы эстрального цикла. Животных выводят из эксперимента декапитацией под ингаляционным наркозом парами эфира в соответствии с правилами работы с экспериментальными животными в фазу непрерывного эструса, которую определяют кольпоцитологически, на 3-5-10-15-30-60-е сутки опыта. При аутопсии визуально оценивают состояние яичников, измеряют размеры, объём яичников, отмечают характер структуры коркового слоя, наличие фолликулов, кист, кровоизлияний. Гистологически обнаруживают различные по величине кистозные полости, кровоизлияния в просвет.
Следует отметить, что типичные для фолликулярных кист макро- и микроскопические признаки, обнаруживаемые на 3-и сутки после окончания введения гормонов свидетельствуют о формировании модели уже к 10-м суткам эксперимента.
Среди возможных механизмов кистообразования в данном случае следует отметить следующие. ХГ, являясь структурным аналогом ЛГ, реализует своё действие на уровне тека-клеток. Избыточная стимуляция ХГ вначале приводит к даун-регуляции ЛГ-рецепторов, а затем к гиперплазии тека-клеток, приводящей к гиперандрогении. С одной стороны гиперандрогения усиливает выработку эстрогенов (формируется гиперэстрогения), что проявляется непрерывным эструсом у подопытных животных. С другой, повышенный синтез андрогенов препятствует нормальному развитию фолликулов, нарушает процесс селекции доминантного фолликула и вызывает образование множества кистозно-атрезирующихся фолликулов. Гиперстимуляция ЛГ-рецепторов сопровождается также повреждением ФСГ-рецепторов в клетках гранулёзы, снижением эффективности воздействия ФСГ на растущие фолликулы. Кроме того, согласно современным представлениям об овариальном стероидогенезе большое значение уделяется ИПФР и инсулину. Так основным механизмом развития гиперчувствительности клеток теки к ХГ является способность инсулина и ИПФР I и II блокировать гомологическую десенситизацию ЛГ-рецепторов, что усиливает продукцию андрогенов тека-клетками, а также вызывает преждевременную лютеинизацию гранулёзы, способствуя остановке роста фолликулов и их атрезии. Кроме того, инсулин, вероятно, усиливает эффект гонадотропного рилизинг-гормона на продукцию эндогенного ЛГ гипофизом [18-20]. Таким образом, хроническая гиперстимуляция тека-клеток субовуляторными дозами ХГ в сочетании с выраженной гиперинсулинемией закономерно приводит к формированию множественных фолликулярных кист яичников в эксперименте.
Конкретный пример
В качестве экспериментального материала использовалась половозрелая белая беспородная крыса-самка массой 210 г, находящаяся в фазе диэструса эстрального цикла, полученная в Центральном виварии Сибирского государственного медицинского университета. Эксперимент выполнялся на базе отдела экспериментальной физиологии и хирургии центральной научно-исследовательской лаборатории Сибирского государственного медицинского университета.
Крысу фиксировали в положении на спине. При помощи инсулинового шприца внутримышечно в ягодичную область, исключая ранение магистральных сосудов и нервных пучков, вводили 300 ЕД хорионического гонадотропина, разведённого 0,3 мл раствора для инъекций, приложенного к ампуле хорионического гонадотропина, раствор приготавливали ex tempore. Аналогичным образом вводили 2,5 ЕД протафана НМ, разведённого 0,05 мл воды для инъекций, раствор приготавливали ex tempore. Указанные препараты вводили ежедневно в течение 7 дней в первой половине дня до 12 часов.
Экспериментальное животное содержали в стандартных условиях вивария на обычном питании с добавлением овощей и творога, при дозированном освещении (12:12, свет с 8 часов). Ежедневно животное осматривали, отмечали его общее состояние, внешний вид, поведение, пищевую возбудимость и двигательную активность, измеряли массу тела и проводили кольпоцитологическое исследование для диагностики фазы астрального цикла. Животное выводили из эксперимента декапитацией под ингаляционным наркозом парами эфира в соответствии с правилами работы с экспериментальными животными в фазу непрерывного эструса, которую определяли кольпоцитологически, на 3-и сутки эксперимента.
Тотчас после взятия яичники фиксировали в жидкости Карнуа и нейтральном формалине, заливали в парафин и после приготовления серийных срезов толщиной 4-5 мкм окрашивали гематоксилином и эозином и по Ван-Гизону. Гистологические исследования проводились на кафедре гистологии, эмбриологии и цитологии Сибирского государственного медицинского университета.
На 3-и сутки после введения гормонов поведение, двигательная активность животного заметно не изменялись, однако отмечалась повышенная пищевая возбудимость, проявлявшаяся достоверным увеличением массы тела животного. Визуально в брюшной полости у животного, выведенного из эксперимента на 3-и сутки, обращали на себя внимание: асимметричное увеличение в размерах яичников до 0,9-1,0 см, выраженный крупнокистозный рисунок коркового слоя, многочисленные фолликулы, кисты, содержащие прозрачный секрет (фиг.1, б). Кисты имели размеры от 0,5 до 2 и более миллиметров в диаметре. Обращало на себя внимание резкое полнокровие сосудов (при незначительном механическом воздействии на яичники возникало кровотечение), гиперемия и утолщение маточных рогов. Яичники были темно окрашены, свободно лежали в брюшной полости.
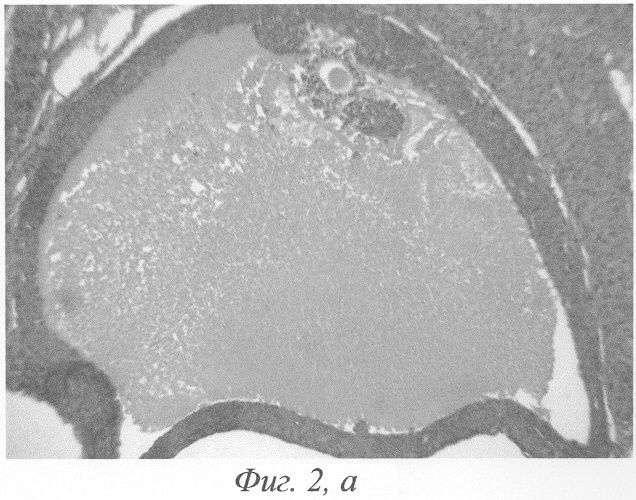
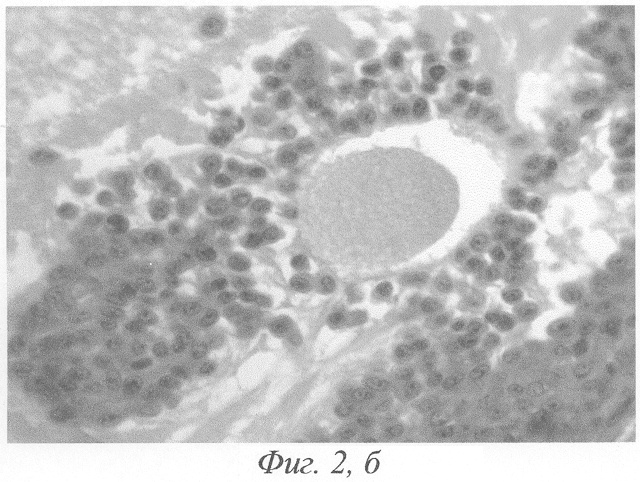
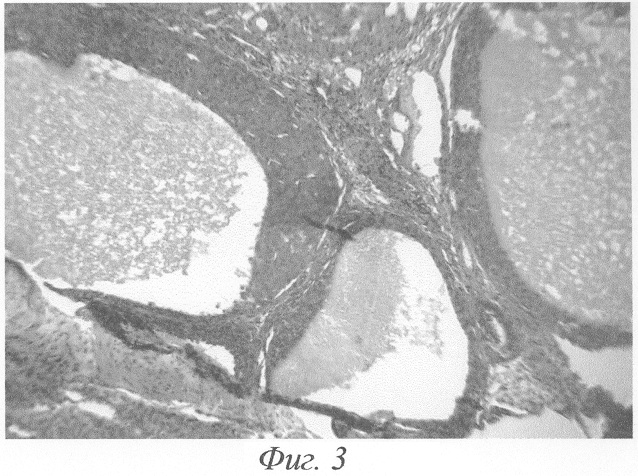
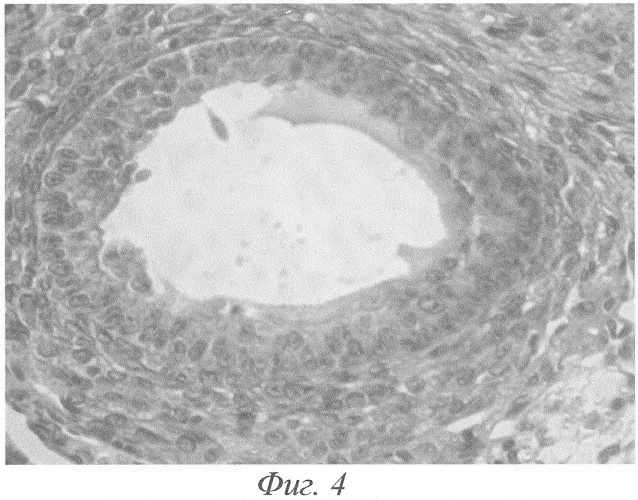
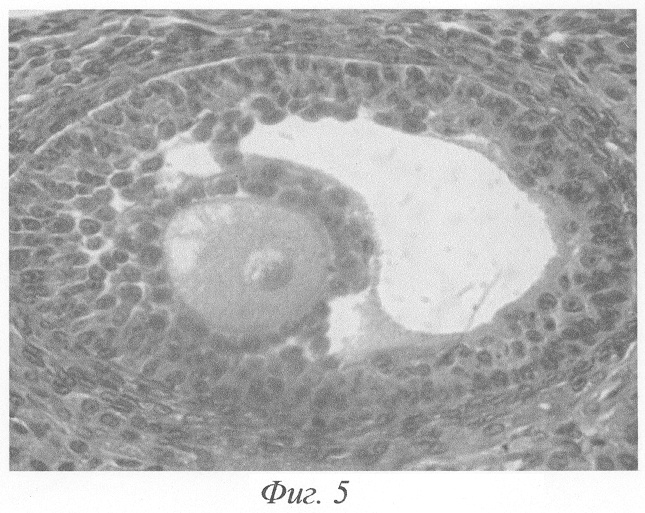
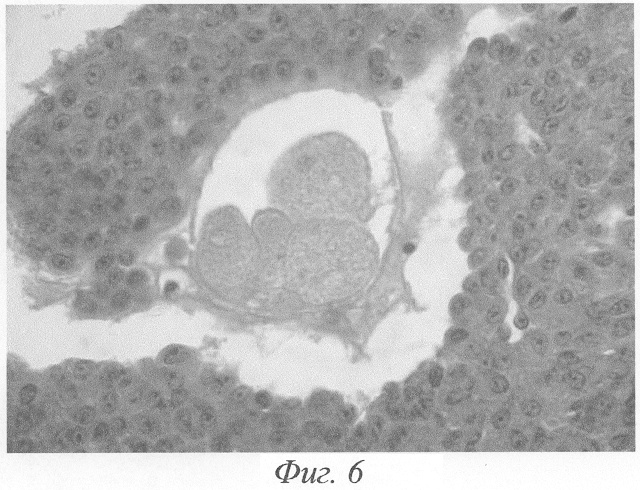
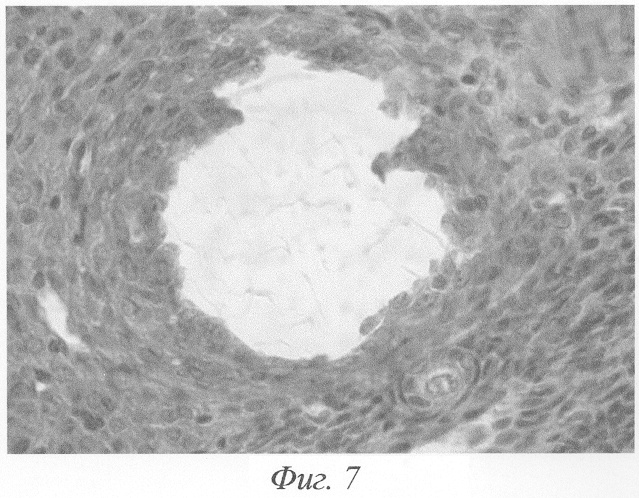
При гистологическом исследовании яичников животного на 3-и сутки после окончания введения данных гормонов формировались кисты на этапе граафова пузырька (фиг.2, а, б). Полость кисты содержала либо гомогенный, либо мелкозернистый бледно окрашивающийся секрет, не отличимый от секрета нормального фолликула. В тех кистах, в которых обнаруживались кровоизлияния, просвет содержал агглютинированные и подверженные частичному лизису эритроциты. В большей части кист отмечалась полная дегенерация и цитолиз овоцитов, дискомплексация фолликулярного эпителия, в части эпителиоцитов - фрагментация ядра и цитоплазмы (фиг.4). Обнаруживались вторичные фолликулы, где овоцит был сохранён, однако структура его оказывалась нарушенной: имели место очаговый отёк цитоплазмы, невыраженные явления дискомплексации фолликулярного эпителия (фиг.5). В других же кистах отмечалось явление псевдодробления овоцитов, при этом бластомеры, расположенные внутрифолликулярно, как правило, были неравномерны (фиг.6). Значительно увеличивалось количество атретических фолликулов. Атрезии подвергались фолликулы на разных стадиях развития, поэтому обнаруживались атретические фолликулы и кисты разной величины (фиг.3). Наряду с атретическими фолликулами обнаруживалось небольшое количество неизменённых фолликулов, вступающих в стадию роста. Выявлялись сохранившие целостность примордиальные фолликулы (фиг.7). Количество третичных фолликулов было снижено. В фолликулярном эпителии в большом количестве встречались апоптотические тельца.
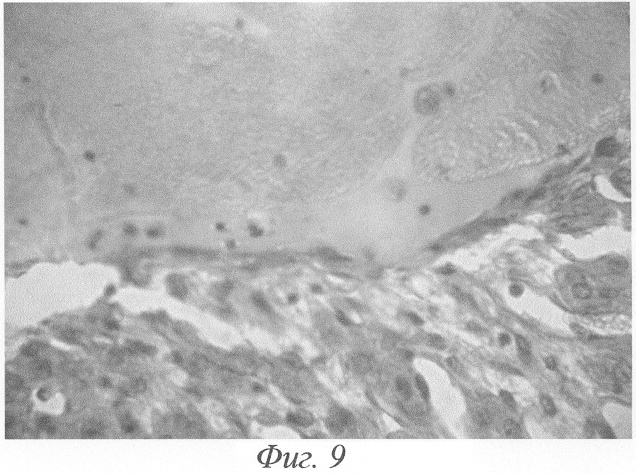
Наряду с фолликулярными кистами в яичниках подопытной крысы наблюдались жёлтые тела, кисты желтого тела. В отдельных случаях в центре жёлтых тел удавалось наблюдать резко расширенные кровеносные сосуды, просвет которых был заполнен агглютинированными эритроцитами или тромбами (фиг.9).
Размер кисты желтого тела обычно несколько превышал размер нормального сформировавшегося желтого тела. Полость кисты содержала гомогенный, интенсивно красящийся эозином секрет. Стенка кисты состояла из нескольких рядов (до 20-30 рядов) лютеиновых клеток. По своим размерам и морфологии эти клетки почти ничем не отличались от лютеиновых клеток нормального желтого тела: клетки располагались радиальными тяжами, разделенными тонкими прослойками соединительной ткани с хорошо развитой капиллярной сетью. На границе лютеиновых клеток с полостью кисты располагался слой вытянутых клеток.
Таким образом, 7-дневное введение ХГ в суперовуляторной дозе и умеренная гиперинсулинемия вызывают в 100% развитие поликистозных яичников с преобладанием фолликулярных кист с выраженными макро- и микроскопическими проявлениями данной патологии на 3-и сутки эксперимента.
Результаты исследований
Эксперимент был проведен на 24 половозрелых беспородных крысах-самках, у которых были созданы поликистозные яичники с преобладание фолликулярных кист.
Контролем служили интактные животные (12 особей) одного возраста с подопытными и содержавшиеся вместе с ними в условиях вивария. Взятие контрольного материала производили в динамике одновременно с экспериментальным для учета возрастных изменений. Выбор в качестве экспериментального материала крыс обусловлен тем, что среди лабораторных животных закономерности регуляции овариального цикла у них наиболее близки таковым у женщин [3].
При вскрытии брюшной полости визуально изучали состояние органов брюшной полости: наличие и характер выпота, состояние брюшины, измеряли размеры яичников, отмечали их цвет, характер структуры коркового слоя, наличие фолликулов, кист, кровоизлияний. Тотчас после взятия яичники фиксировали в жидкости Карнуа и нейтральном формалине, заливали в парафин и после приготовления серийных срезов толщиной 4-5 мкм окрашивали гематоксилином и эозином и по Ван-Гизону. Гистологические исследования проводились на кафедре гистологии, эмбриологии и цитологии Сибирского государственного медицинского университета.
При введении хорионического гонадотропина и протафана НМ в течение 7-и суток двигательная и пищевая возбудимость крыс были не изменены, по поведению они не отличались от контрольной группы животных. Максимальное увеличение массы тела отмечалось на 10-12-е сутки после введения гормонов. На 3-5-е сутки эксперимента при вскрытии брюшной полости животных отмечалось увеличение в размерах яичников до 0,9-1,0 см (фиг.1, а), выраженный крупнокистозный рисунок коркового слоя, многочисленные фолликулы, кисты, содержащие прозрачный секрет (фиг.1, б, в), в части кист имели место кровоизлияния (фиг.1, г). Кисты имели размеры от 0,5 до 2 и более миллиметров в диаметре. У одних крыс яичники были увеличены и состояли из крупных кист разной величины, у других они были меньше по величине и состояли из большого числа мелких кист. Обращало на себя внимание резкое полнокровие сосудов (при незначительном механическом воздействии на яичники возникало кровотечение), гиперемия и утолщение маточных рогов. Яичники свободно лежали в брюшной полости. В одних случаях яичники были темно окрашены, в других имели белесоватый оттенок. На 10-15-е сутки после введения гормонов описываемая картина сохранялась. Яичники имели прежние размеры и цвет, были бугристы. Экссудата, спаечного процесса в брюшной полости не было. Во многих фолликулах визуализировались кровоизлияния. Сохранялась гиперемия и утолщение маточных рогов. На 30-е и 60-е сутки эксперимента яичники несколько уменьшались в размере, но сохраняли бугристую поверхность, чаще имели белесоватый оттенок. Менее часто выявлялись фолликулы, кисты. В части фолликулов имелись кровоизлияния. Кроме того, наблюдались гиперемия и утолщение маточных рогов.
При детальном гистологическом изучении яичников, содержащих кисты, удалось установить, что в своём развитии фолликулярные кисты проходили ряд определённых стадий. Фолликулы в яичниках увеличивались в размерах, главным образом за счёт увеличения полости. Эти образования имели вид кистозно растянутых, так называемых персистирующих фолликулов. Затем размеры полости ещё более увеличивались, яйцеклетка подвергалась некробиозу, окончательному распаду, и персистирующий фолликул превращался в фолликулярную кисту.
В яичниках белых крыс при создании экспериментальной модели функциональных кист путём 7-дневного введения хорионического гонадотропина и протафана ИМ на 3-и сутки после окончания введения данных гормонов формировались кисты на этапе граафова пузырька (фиг.2, а, б). Полость кисты содержала либо гомогенный, либо мелкозернистый бледно окрашивающийся секрет, не отличимый от секрета нормального фолликула. В тех кистах, в которых обнаруживались кровоизлияния, просвет содержал агглютинированные и подверженные частичному лизису эритроциты. Стенка кисты на этой стадии развития была выстлана хорошо сохранившимися гранулёзными клетками. Признаков атрофии, дегенерации или распада гранулёзы, как правило, не наблюдалось. Чаще всего гранулезная выстилка имела одинаковую толщину и состояла из 6-12 рядов клеток. Ядра клеток были несколько более компактными, чем в норме, имели округлую или овальную форму и были окружены узким ободком слабо базофильной цитоплазмы. Внутренняя тека утолщалась, клетки её увеличивались в размерах, ядра набухали, приобретали округлую форму и становились светлыми; цитоплазма клеток подвергалась вакуолизации. Наружная оболочка заметно не изменялась. Выше описанная картина свидетельствовала о гормональной активности таких кист.
Дальнейшая судьба фолликулярных кист могла быть различной. В одних кистах появлялись различного типа дегенеративные и атрофические изменения гранулезных клеток. Они сводились к вакуолизации цитоплазмы, пикнозу или распаду ядер на мелкие глыбки и слущиванию клеток в просвет кисты. Толщина атрофичной гранулёзы уменьшалась до одного-двух рядов клеток. В дальнейшем исчезали последние остатки гранулёзы, и киста превращалась в ретенционную полость, резко растянутую гомогенным веществом и выстланную одним-двумя рядами резко уплощенных клеток. Снаружи эти полости были окружены узким кольцом вытянутых фибробластоподобных клеток с небольшим количеством цитоплазмы, которые представляли собой резко атрофичные theca extema и interna. Такие кисты, потерявшие свои специфические секреторные элементы - гранулезу и теку, - соответствовали гормонально неактивным кистам. Содержание описанных кист на 3-и сутки эксперимента было незначительным.
Значительно увеличивалось количество атретических фолликулов. Атрезии подвергались фолликулы на разных стадиях развития, поэтому обнаруживались атретические фолликулы и кисты разной величины (фиг.3). В большей части кист отмечалась полная дегенерация и цитолиз овоцитов, дискомплексация фолликулярного эпителия, в части эпителиоцитов - фрагментация ядра и цитоплазмы (фиг.4). Обнаруживались вторичные фолликулы, где овоцит был сохранён, однако структура его оказывалась, нарушенной: имели место очаговый отёк цитоплазмы, невыраженные явления дискомплексации фолликулярного эпителия (фиг.5). В других же кистах отмечалось явление псевдодробления овоцитов, при этом бластомеры, расположенные внутрифолликулярно, как правило, были неравномерны (фиг.6). Наряду с атретическими фолликулами обнаруживалось небольшое количество неизменённых фолликулов, вступающих в стадию роста. Выявлялись сохранившие целостность примордиальные фолликулы (фиг.7). Количество третичных фолликулов было снижено. В фолликулярном эпителии в большом количестве встречались апоптотические тельца.
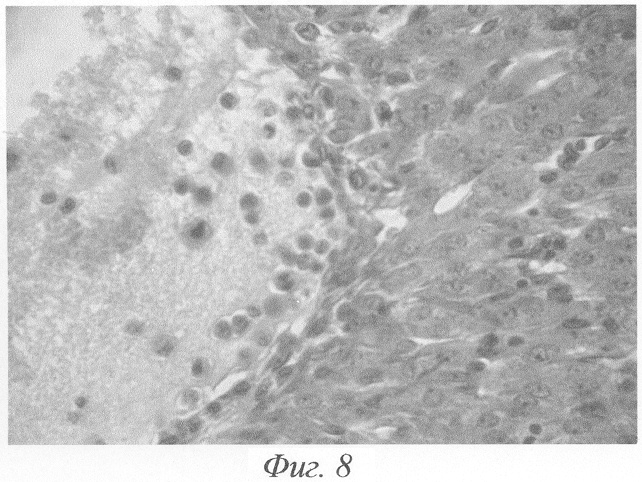
В клетках гранулезы и внутренней оболочки других кист очень рано происходило накопление липоидных веществ. В просвете кист, наряду с оксифильными массами, выявлялся клеточный детрит. Такие кисты были выстланы одним слоем плоских клеток, в части кист имело место выселение мононуклеаров в полость фолликула. Также отмечались выраженная гипертрофия клеток внутренней теки и железистый метаморфоз (фиг.8). Процесс лютеинизации заключался в образовании больших эпителиоидных клеток с округлым пузырькообразным ядром и вакуолизированной цитоплазмой. Обычно лютеинизация начиналась с клеток theca intema, богатой сосудами, а затем уже распространялась на гранулезные клетки. Иногда лютеинизации подвергались лишь отдельные участки стенки кисты, и тогда киста имела смешанное строение. Однако чаще эти изменения отмечались на всем протяжении ее стенки. В некоторых кистах, наряду с лютеинизацией, наблюдались распад и десквамация части гранулезных клеток, непосредственно примыкающих к полости кисты. В результате всех вышеуказанных процессов развивались кисты с лютеинизированной стенкой. Дальнейшая эволюция таких кист сводилась к процессам дегенерации и атрофии, вследствие чего толщина стенки прогрессивно уменьшалась и они превращались в гормонально неактивные кисты, морфологически неотличимые от вышеописанных неактивных кист, образовавшихся непосредственно из фолликулярных, минуя лютеинизацию. При далеко зашедшей атрофии эпителиальной выстилки определить происхождение старой, неактивной запустевшей кисты было невозможно. Иногда по небольшим остаткам гранулезных или лютеинизированных клеток можно было установить, произошла ли такая старая киста непосредственно из фолликулярной или из кисты с лютеинизированной стенкой.
Обычно в одном и том же яичнике крысы можно было видеть одновременно несколько кист, находившихся на разных стадиях развития: типичные гормонально активные фолликулярные кисты с многослойной гранулезной выстилкой, кисты, подвергающиеся лютеинизации или полностью лютеинизированные, подвергающиеся атрофии, и, наконец, запустевшие - неактивные кисты. Это являлось доказательством того, что фолликулярная киста проходила определенный путь развития. По-видимому, вначале киста обладала гормональной активностью, которая постепенно падала. Другие фолликулярные кисты начинали развиваться позже и своей активностью замещали выпадающую функцию более старых кист. Подтверждением сказанного являлось и то обстоятельство, что в яичниках подопытных крыс с увеличением сроков эксперимента незначительно уменьшалось число активных кист и нарастало число неактивных кист.
Наряду с фолликулярными кистами в яичниках подопытных крыс наблюдались жёлтые тела, кисты желтых тел. Как правило, такие кисты были единичными; иногда в одном яичнике, наряду с фолликулярными кистами, обнаруживались 2-3 кисты желтых тел. В отдельных случаях в центре указанных жёлтых тел удавалось наблюдать резко расширенные кровеносные сосуды, просвет которых был заполнен агглютинированными эритроцитами или тромбами (фиг.9). Размер кисты желтого тела обычно несколько превышал размер нормального сформировавшегося желтого тела. Полость кисты содержала гомогенный, интенсивно красящийся эозином секрет. Стенка кисты состояла из нескольких рядов (до 20-30 рядов) лютеиновых клеток. По своим размерам и морфологии эти клетки почти ничем не отличались от лютеиновых клеток нормального желтого тела: клетки располагались радиальными тяжами, разделенными тонкими прослойками соединительной ткани с хорошо развитой капиллярной сетью. На границе лютеиновых клеток с полостью кисты располагался слой вытянутых клеток. В некоторых кистах можно было наблюдать атрофию лютеиновой выстилки, в связи с чем толщина ее резко уменьшалась.
Образование кистозно-атрезированных фолликулов наблюдалось также и в яичниках контрольных крыс. Морфологически этот процесс происходил в общем так же, как и у подопытных животных. Атретические фолликулы также проходили стадии персистирующего фолликула, фолликула с наличием гранулёзной выстилки, сохранившего гормональную активность, и, наконец, запустевшего кистозно-атрезирующегося фолликула. Особенностью морфологических изменений у контрольных крыс являлось то, что превращение фолликула с гормональной активностью в запустевающий происходило у них чаще путем непосредственной атрофии гранулезы, без предварительной лютеинизации. Кроме того, у контрольных крыс значительно реже наблюдались кистозно-расширенные желтые тела.
Кровеносные сосуды коркового и мозгового вещества были расширены и гиперемированы. Наблюдалось прикраевое расположение лейкоцитарных элементов, миграция части лейкоцитов через сосудистую стенку и, как следствие, слабо выраженная лейкоцитарная инфильтрация периваскулярной соединительной ткани.
Гистологическая картина яичников крыс на 5-10-е сутки эксперимента, в целом, мало чем отличалась от описанной на 3-и сутки опыта. Прежде всего, обращало на себя внимание довольно значительное содержание в яичнике кистозно-атрезированных фолликулов. В них наблюдался деструктивно изменённый в виде кариопикноза и кариорексиса фолликулярный эпителий. Овоциты в подобных фолликулах были дегенеративно изменены, либо подвергнуты гомогенизации и распаду, в отдельных срезах отмечался отрыв яйценосного бугорка. Выявлялось достаточно много мелких атретических фолликулов и тел. При этом отмечалась выраженная пролиферация гормонопродуцирующей ткани. Обнаруживалось большое количество гормонально активных кист.
Вместе с тем обнаруживались единичные растущие фолликулы на стадии первичных и вторичных фолликулов, реже третичных, отмечалось формирование кист на этапе граафова пузырька. Выявлялись примордиальные фолликулы, имеющие обычное строение. В отдельных случаях белочная оболочка была локально истончена. Поверхностный эпителий имел очаговые дефекты. Встречались желтые тела и кисты жёлтых тел.
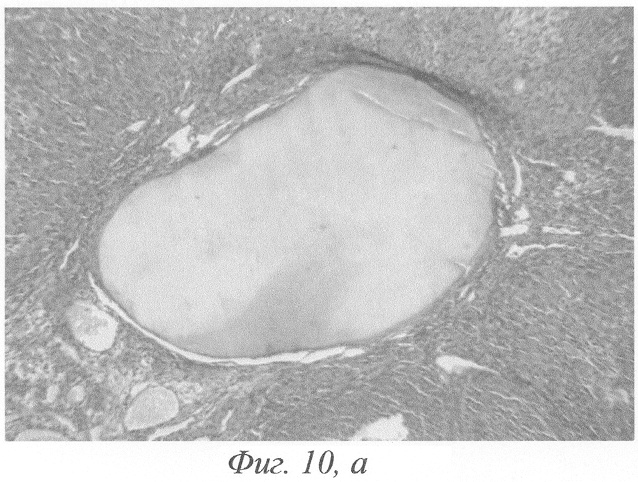
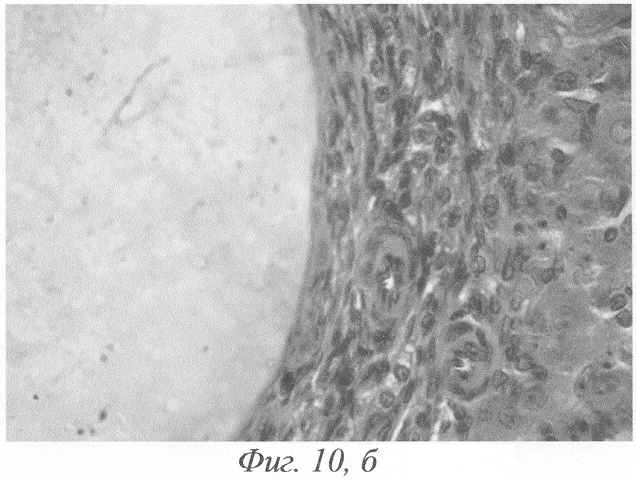
Через 15-30 суток после введения гормонов морфологическая картина яичников была сходной с описанной в предыдущие сроки эксперимента. Сходство заключалось главным образом в наличии большого количества кистозно-атрезирующихся фолликулов, различных по величине кист с кровоизлияниями в просвет (фиг.10, а, б), уменьшении количества растущих фолликулов, пролиферации гормонопродуцирующей ткани.
По сравнению с предыдущими сроками несколько возрастало содержание первичных, вторичных и третичных фолликулов. Однако зернистый слой, особенно у вторичных фолликулов, в отличие от контроля (интактные животные) весьма часто содержал клетки, характеризующиеся повышенной конденсацией хроматина, гиперхромией кариоплазмы, деформацией клеточных контуров. Часть клеток была пикноморфна. Наряду с этим в фолликулярном эпителии были нередки картины митоза.
Часть овоцитов описываемых растущих фолликулов была дегенеративно изменена. А в тех фолликулах, где овоцит сохранялся, структура его была нарушена, что проявлялось очаговым отёком цитоплазмы и маловыраженными явлениями дискомплексации фолликулярного эпителия. В фолликулярном эпителии встречались апоптотические тельца. Интерстиций яичников характеризовался мелкоочаговой лютеинизацией. Встречались единичные, небольшие по величине кисты с лютеинизированной стенкой.
На 60-е сутки описываемой модели наблюдалась гиперемия сосудов мозгового вещества. Больших кистозных полостей не было, но сохранялись мелкие кистозно-атрезирующиеся фолликулы. Процессы образования новых атретических фолликулов шли на убыль. Чаще, чем в предыдущие сроки обнаруживались растущие фолликулы. Однако значительная часть эпителиоцитов зернистого слоя в них была подвержена деструктивным изменениям. Овоциты также характеризовались различной степени выраженности нарушениями: локальным отёком цитоплазмы, повышенной конденсацией хроматина, деформацией ядерной оболочки. В большом количестве встречались жёлтые тела. Содержание интерстициальной железистой ткани, гормонопродуцирующих клеток атретических тел понижалось по сравнению с предшествующими сроками.
Таким образом, для приближения патоморфологических изменений, патогенетических особенностей (условий формирования) множественных фолликулярных кист яичников в эксперименте к таким, которые имеют значение в клинике, а также сокращения сроков моделирования, повышения стабильности воспроизведения и расширения области использования данной модели, предложено комбинировать введение ХГ в суперовуляторной дозе с дозой инсулина, достаточной для формирования умеренной гиперинсулинемии.
Как показали экспериментальные исследования, умеренная гиперинсулинемия и суперовуляторные дозы ХГ вызывают в 100% развитие множественных фолликулярных кист яичников в эксперименте. Изучение морфологии яичников в динамике свидетельствует об обратимости описанных выше явлений - регрессе кист в течение 60-и суток после введения ХГ и инсулина, что подтверждает их функциональный характер. Яичники с экспериментальными множественными кистами имеют характерные морфологические особенности: размеры яичников увеличены за счёт различных по величине кист, поверхность которых гладкая, блестящая, имеет серо-розовую окраску, стенки тонкие, чаще однокамерные, содержащие прозрачный секрет, реже секрет коричневого цвета (кровоизлияние). При гистологическом изучении яичников выявляются кисты на разных стадиях развития. Наиболее часто встречаются фолликулярные кисты, реже кисты жёлтого тела, кисты с лютеинизированной стенкой. Описанные кисты морфологически соответствуют таковым у женщин. Следует заметить, что процесс образования кист проходит ряд последовательных стадий.
Предлагаемый способ позволяет создать модель поликистозных яичников с преобладанием фолликулярных кист у экспериментальных животных (беспородных белых половозрелых крыс-самок массой 200-220 г). Необходимо отметить относительную простоту воспроизведения данной модели, отсутствие летальных исходов у подопытных животных, стабильное воспроизведение полученных результатов, небольшие сроки моделирования, морфологическое сходство экспериментальных фолликулярных кист с таковыми у женщин.
Данная модель поликистозных яичников позволяет расширить область своего применения, более детально изучить звенья патогенеза данной патологии, провести апробацию терапевтического действия ряда новых препаратов.
Фиг.1. Макроскопический вид яичников крыс (после фиксации в жидкости Карнуа) на 3-и сутки после окончания введения ХГ и инсулина: а - увеличение яичника в размере; б, в -выраженный крупнокистозный рисунок коркового слоя, многочисленные фолликулы, кисты; г - кровоизлияния в кисты.
Фиг.2. а - формирование кисты на этапе граафова пузырька на 3-и сутки после окончания введения ХГ и инсулина; б - (фрагмент предыдущего) гомогенизация овоцита, дискомплексация фолликулярного эпителия в области яйценосного бугорка, отрыв части эпителиоцитов в полость фолликула. Окраска гематоксилином-эозином. Ув. а. - 120; б. -600.
Фиг.3. Три кисты на стадии граафова пузырька на 3-и сутки после окончания введения ХГ и инсулина. Окраска гематоксилином-эозином. Ув. 120.
Фиг.4. Кистозно-атрезирующийся фолликул. Полная дегенерация и цитолиз овоцита, дискомплексация фолликулярного эпителия, гипертрофия внутренней теки на 3-и сутки после окончания введения ХГ и инсулина. Окраска гематоксилином-эозином. Ув. 600.
Фиг.5. Вторичный фолликул. Структура овоцита нарушена: очаговый отёк цитоплазмы, невыраженные явления дискомплексации фолликулярного эпителия на 3-и сутки после окончания введения ХГ и инсулина. Окраска гематоксилином-эозином. Ув. 600.
Фиг.6. Псевдодробление овоцита в фолликуле яичника. Бластомеры внутрифолликулярно неравномерны. Выраженная гипертрофия и железистый метаморфоз внутренней теки на 3-и сутки после окончания введения ХГ и инсулина. Окраска гематоксилином-эозином. Ув. 600.
Фиг.7. Фолликулярная киста на месте мелкого растущего фолликула. Рядом примордиальный фолликул, сохранивший свою целостность, вступает в стадию роста на 3-и сутки после окончания введения ХГ и инсулина. Окраска гематоксилином-эозином. Ув. 600.
Фиг.8. Киста с лютеинизированной стенкой. Полость выстлана одним слоем плоских клеток. Выраженная гипертрофия внутренней теки. Выселение мононуклеаров в полость фолликула на 3-и сутки после окончания введения ХГ и инсулина. Окраска гематоксилином-эозином. Ув. 600.
Фиг.9. Массивное кровоизлияние в полость кисты жёлтого тела с формированием тромбов на 3-и сутки после окончания введения ХГ и инсулина. Окраска гематоксилином-эозином. Ув. 600.
Фиг.10. а - кровоизлияние в фолликулярную кисту, гемолиз эритроцитов на 15-е сутки после окончания введения ХГ и инсулина; б - фрагмент предыдущего. Окраска гематоксилином-эозином. Ув. а. - 120; б. - 600.
Источники информации
1. Ирд Е.А. Фолликулярные кисты яичников и дисгормональные опухоли. - М.: Медицина, 1966. - 119 с.
2. Convery M., McCarthy G.F., Brawer J.R. Remission of the polycystic ovarian condition (PCO) in the rat following hemiovariectomy //Anat. Rec. - 1990. - Mar. - Vol. 226, № 3. -P.328-336.
3. Лазарев Н.И, Ирд Е.А., Смирнова И.О. Экспериментальные модели эндокринных гинекологических заболеваний. - М., 1976. - 117 с.
4. Singh K.B. Induction of polycystic ovarian disease in rats by continuous light. III. Mechanism of ovarian compensatory hypertrophy and ovulation after unilateral cophorectomy // Am. J. Obstet. Gynecol. - 1969. - Aug. - Vol. 104, № 7. - P.1008-1011.
5. Farookhi R., Hemmings R., Brawer J.R. Unilateral ovariectomy restores ovulatory cyclicity in rats with a polycystic ovarian condition //Biol. Reprod. - 1985. - Apr. - Vol. 32, № 3. - P.530-540.
6. Арсеньева М.Г. Кольпоцитологические исследования в диагностике и терапии эндокринных гинекологических заболеваний. - М.: Медицина, 1977. - 365 с.
7. Гайфулина Ж.Ф. Влияние имплантатов с термомеханической памятью формы на морфофункциональное состояние нормальных и поликистозных яичников (экспериментальное исследование): Дисс. ... канд. мед. наук. - Томск, 1997. - 133 с.
8. Jones H.M., Vemon M.W., Rush M.E. Systematic studies invalidate the neonatally androgenized rat as a model for polycystic ovary disease //Biol. Reprod. - 1987. - June. -Vol. 36, № 5. - P.1253-1265.
9. Amirikia H., Savoy-Moore R.T., Sundareson A.S., Moghissi K.S. The effect of long-term androgen treatment on the ovary // Fertil. Steril. - 1986. - Feb. - P.202-208.
10. Brawer J.R., Munoz M., Farookhi R. Development of the polycystic ovarian condition in the estradiol-valerate - treated rat // Biol. Reprod. - 1986. - Oct. - Vol. 35. - P.647-655.
11. OtaH, Wakizaka A., Fukushima M., Maki M. Enhanced ovarian gonadotropin receptors in the testosterone-induced polycystic ovary in rats // Tohoku J. Exp. Med. - 1996. - Mar. - Vol. 148, № 3.-Р3 13-325.
12. Sawada T. Effects of multiple injection of LH on the secretion ofpregnane compounds from polycystic ovaries of androgen-sterilized rats // J. Endocrin. - 1986. - Sep.- Vol. 110, №3. - Р.507-510.
13. Grosser P.M., McCarthy G.F., Robaire B. et al. Plasma patterns of LH, FSH and prolactin in rats with a polycystic ovarian condition induced by oestradiol valerate // J. Endocrin. - 1987. - Jul. - Vol. 114, № 1. - P.33-39.
14. Singh K.B. Induction of polycystic ovarian disease in rats by continuous light. I. The reproductive cycle, organ weights, and histology of the ovaries // Am. J. Obstet. Gynecol. - 1969. - Apr. - Vol. 103, № 8. - P.1078-1083.
15. Sawada T., Kasaka T., Ichikawa S. Effect of multiple injections ofluteinizing hormone on secretion of pregnane compounds from polycystic ovaries of rats exposed to constant light //Acta Endocrinol. (Copenh.). - 1987. - Nov. - Vol. 116, № 3. - P.390-394.
16. Mahesh V.B., Mills T.M., Bagnell C.A., Conway B.A. Animal models for study of polycystic ovaries and ovarian atresia //Adv. Exp. Med. Biol. - 1987. - № 219. - P.237-257.
17. Иваненко О.Г. Биологическая активность эстрогенов на репродуктивный тракт самцов и самок крыс //Проблемы эндокринологии. - 1988. - Т. 34. - № 4. - С.72-74.
18. Bogovich К., Clemons J., Poretsky L. Insulin has a biphasic effect on the ability of human chorionic gonadotropin to induce ovarian cysts in the rat // Metab. - 1999. - Aug. - Vol. 48. № 8. - P.995-1002.
19. Poretsky L., Clemons J., Bogovich K. Hyperinsulinemia and human chorionic gonadotropin synergistically promote the growth of ovarian follicular cysts in rats // Metab. - 1992. - Aug. - Vol. 41, № 8. - P.903-910.
20. Тихомиров А.Л., Лубнин Д.М. Синдром поликистозных яичников: факты, теории, гипотезы // Фарматека. - 2003. - Т. 74, № 11. - С.8-19.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ МОДЕЛИРОВАНИЯ АУТОИММУННОГО ООФОРИТА | 2010 |
|
RU2439712C1 |
| СРЕДСТВО И СПОСОБ ПРОФИЛАКТИКИ РЕЦИДИВА КИСТ ЯИЧНИКОВ | 2008 |
|
RU2364411C1 |
| СПОСОБ ЛЕЧЕНИЯ ФУНКЦИОНАЛЬНЫХ КИСТ ЯИЧНИКОВ | 2008 |
|
RU2357733C1 |
| СПОСОБ ПРОФИЛАКТИКИ РАЗВИТИЯ СПАЕЧНОГО ПРОЦЕССА В МАЛОМ ТАЗЕ И НАРУШЕНИЙ ОВАРИАЛЬНО-МЕНСТРУАЛЬНОГО ЦИКЛА У ЖЕНЩИН, ПЕРЕНЕСШИХ ЭНДОСКОПИЧЕСКИЕ ОПЕРАЦИИ НА ЯИЧНИКАХ С ИСПОЛЬЗОВАНИЕМ ЭЛЕКТРОКОАГУЛЯЦИИ | 2003 |
|
RU2238687C1 |
| СПОСОБ МОДЕЛИРОВАНИЯ ХРОНИЧЕСКОГО ВОСПАЛЕНИЯ ПРИДАТКОВ МАТКИ | 2002 |
|
RU2224297C1 |
| СПОСОБ МОДЕЛИРОВАНИЯ ПОЛИКИСТОЗНЫХ ЯИЧНИКОВ | 2007 |
|
RU2337411C1 |
| СПОСОБ И СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ ДОБРОКАЧЕСТВЕННЫХ ГИПЕРПЛАСТИЧЕСКИХ ПРОЦЕССОВ ЭНДОМЕТРИЯ | 2004 |
|
RU2275194C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СИНДРОМА ПОЛИКИСТОЗНЫХ ЯИЧНИКОВ | 2020 |
|
RU2757220C1 |
| Способ прогнозирования исходов вспомогательных репродуктивных технологий при трубном бесплодии | 2017 |
|
RU2672267C1 |
| СПОСОБ ЛЕЧЕНИЯ ОСТРЫХ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ ЖЕНСКИХ ПОЛОВЫХ ОРГАНОВ | 1997 |
|
RU2141367C1 |
Изобретение относится к экспериментальной медицине, а именно к экспериментальной гинекологии, и может быть использовано для моделирования поликистозных яичников с преобладанием фолликулярных кист. Для этого беспородным половозрелым крысам-самкам, находящимся в фазах покоя эстрального цикла, путем внутримышечного введения вводят хорионический гонадотропин в дозе 300 ЕД, протафан НМ в дозе 2,5 ЕД ежедневно в первой половине дня в течение 7 дней. Способ обеспечивает создание фолликулярных кист яичников, близких по патоморфологии к таковым в клинике, при сокращении сроков моделирования и повышении стабильности воспроизведения модели. 10 ил.
Способ моделирования поликистозных яичников с преобладанием фолликулярных кист путем внутримышечного введения беспородным половозрелым крысам-самкам, находящимся в фазах покоя эстрального цикла, хорионического гонадотропина и инсулина, отличающийся тем, что вводят хорионический гонадотропин в дозе 300 ЕД, протафан НМ в дозе 2,5 ЕД ежедневно в первой половине дня в течение 7 дней.
| PORETSKY L | |||
| et al | |||
| Hyperinsulinemia and human chorionic gonadotropin synergistically promote the growth of ovarian follicular cysts in rats | |||
| Metabolism | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| СПОСОБ ЛЕЧЕНИЯ ХРОНИЧЕСКОЙ АНОВУЛЯЦИИ И ПРОФИЛАКТИКИ СИНДРОМА ГИПЕРСТИМУЛЯЦИИ ЯИЧНИКОВ | 1999 |
|
RU2201225C2 |
| WO 9428128 08.12.1994 | |||
| Способ облагораживания сульфит целлюлозных экстрактов | 1944 |
|
SU65492A1 |
| АНТОНИЧЕВ А.В | |||
| и др | |||
| Динамика тиреоидных гормонов у крыс при моделировании синдрома поликистозных яичников | |||
| Проблемы эндокринологии | |||
| - М., 1998, №3, с.33-35 | |||
| KUSCU N.K | |||
| et al | |||
| Insulin: does it induce follicular arrest in the rat ovary? Gynecol | |||
| Endocrinol | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |

