Конструкция модифицированного нитевидного бактериофага для лечения или профилактики бактериальной инфекции, модифицированный бактериофаг М13, фармкомпозиция и способ лечения бактериальной инфекции, моноклональное антитело, гибридома (вариант)
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к бактериофагам, пригодным для лечения бактериальных инфекций, особенно бактериальных инфекций слизистых оболочек, таких как инфекции Helicobacter pylori.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Бактериофаги и антибиотикоустойчивость
Устойчивость к антибиотикам - это всеобщая проблема все возрастающего медицинского и экономического значения. Существует настоятельная необходимость в альтернативных способах уничтожения бактерий, которые смогут разрешить проблему такой устойчивости.
Бактериофаг или фаг - это вирус, который специфически инфицирует бактерии. Фаги прикрепляются к своим бактериям-хозяевам и переносят в них гены, кодирующие различные фаговые белки. Для своей репликации они используют белок-синтезирующий комплекс, аминокислоты и тому подобное, а также энергию, обеспечиваемую бактерией-хозяином (Maloy et al. (eds.): Microbial genetics. Jones and Bartlett Publishers, 1994).
Большинство фагов посредством лизиса или других механизмов поражают специфические штаммы бактерий. Настоящее изобретение исходит из положения о том, что генетическая модификация фагов, в частности нитевидных бактериофагов, предоставляет средство для конструирования новых бактерия-специфичных фагов, способных уничтожать определенные бактерии, например Helicobacter pylori, и позволяющих разрешить проблемы, связанные с антибиотикоустойчивостью.
Нитевидные фаги
Клетки Е. coli, несущие подобные волосу F-фимбрии, являются хозяевами для нитевидных фагов, таких как М13, fd и f1. Эти Ff (F фимбрии, нитевидные) фаги очень близки по последовательности и свойствам (Rashed & Oberer (1986) Microbiological reviews 50, 401-427; Kornberg & Baker, in: DNA Replication, p. 557-570, W.H. Freeman and Co., New York 1992). Ff фаги единственные среди бактериальных вирусов не продуцируют литическую инфекцию, они скорее индуцируют состояние, в котором инфицированные клетки-хозяева продуцируют и секретируют фаговые частицы, не подвергаясь лизису.
Однонитевой геном фага М13 кодирует 10 различных белков. ДНК заключена в белковую оболочку, состоящую из приблизительно 2700 копий белка гена 8 (g8p). Жизнеспособный фаг М13 также экспрессирует на своем конце пять копий белка 43 кДа гена 3 (g3p), причем этот белок ответственен за адсорбцию на фимбрии Е. соli. Белок гена 3 прикреплен к вирусной оболочке посредством С-терминальной части полипептидной цепи, в то время как N-терминальный глобулярный домен остается доступным и опосредует прикрепление фага к концу хозяйской F-фимбрии. При электронном микроскопировании комплекс адсорбции выглядит как структура "шишка-на-стебле" на одном конце фага. В ходе инфицирования лидерные последовательности g3p и g8p направляют транспорт этих белков во внутреннюю мембрану бактериальной клетки.
Ff фаги пользуются заслуженной популярностью в качестве векторов клонирования, поскольку они не имеют физических ограничений, лимитирующих длину ДНК, которая может быть упакована, и поэтому они позволяют осуществлять легкую очистку однонитевой ДНК. Фагмида - это вектор, который несет ориджины репликации как М13 (однонитевой), так и плазмиды (двунитевой). Фагмиды могут быть выращены как плазмиды или упакованы как рекомбинантный фаг М13 с помощью хелперного фага, такого как М13К07 (Veira & Messing (1987) Methods in Enzymol. 153, 3-11).
Получение рекомбинантного антитела
Молекулы антитела содержат дискретные фрагменты, которые могут быть выделены посредством расщепления протеазами или могут быть получены с помощью рекомбинантных методов. Одним из таких фрагментов является Fv (вариабельный фрагмент), который состоит только из VL и VH областей антитела. Известна (US 4946778) рекомбинантная версия Fv фрагмента, именуемая как одноцепочечный Fv (ScFv), в котором две вариабельных области искусственно соединены нейтральным линкером и экспрессируются как единственная полипептидная цепь.
Метод получения рекомбинантного антитела был разработан McCafferty с соавторами (McCafferty (1990) Nature 348, 552-554; Winter & Milstein (1991) Nature 349, 293). Этот подход базируется на системе фаг-воспроизведение, в которой VH (вариабельная тяжелая) и VL (вариабельная легкая) гены клонируют в фаговом векторе, после чего фрагменты антител экспрессируются как слитые белки, воспроизводимые на фаговой поверхности. При таком подходе из популяции могут быть отобраны антитела определенной специфичности и афинности. Было предложено называть антитела, выделенные и изготовленные в прокариотических системах, "коликлональными" антителами (Chiswell & McCafferty (1992) Trends in Biotechnology 10, 80-84).
Имеющаяся в продаже фагмида pCANTAB5 сконструирована таким образом, что гены вариабельной области антитела могут быть клонированы между лидерной последовательностью и главным телом М13 гена 3. G3p лидерная последовательность направляет транспорт полученного слитого белка во внутреннюю мембрану и/или периплазму Е. соli, где главный g3р домен присоединяет слитый белок к концу собирающегося фага. Экспрессия конструкции антитело-g3р ген контролируется индуцибельным lас промотором на фагмиде.
Инфекция Helicobacter pylori
Считается, что бактерия Helicobacter pylori является главной причиной язвы желудка и двенадцатиперстной кишки, ответственной за 84% и 95% соответственно известных случаев (Kuipes, E.L. et al. (1995) Aliment. Pharmacol. Ther. 9 (suppl.2), 59-69). H. pylori колонизирует стенку желудка, защищенную от кислого окружения слоем слизи, который покрывает стенку желудка, а также метаболическим процессом, который дает возможность организму секретировать аммиак для нейтрализации кислоты.
По-видимому, традиционное лечение антибиотиками оказывает слабое действие на Н. pylori. Это возможно определяется следующими причинами: (1) ограниченный доступ антимикробного агента к организму, поскольку указанный агент непосредственно не попадает в кровообращение; и (2) быстрое прохождение многих пероральных антибиотиков через желудок или разложение таких антибиотиков в кислых условиях желудка.
ЗАДАЧА ИЗОБРЕТЕНИЯ
Задачей настоящего изобретения является предложение новых форм лечения для уничтожения бактерий, особенно уничтожения бактерий, ответственных за бактериальные инфекции слизистых оболочек, таких как Helicobacter pylori. В частности, в изобретении предложены нитевидные бактериофаги, генетически модифицированные таким образом, что они обладают специфичностью связывания по отношению к другому бактериальному хозяину, для использования в терапии.
Считается, что способы лечения бактериальных инфекций слизистых оболочек, основанные на рекомбинантных фагах, превосходят традиционное лечение антибиотиками по ряду причин, в том числе следующих:
- они делают возможным уничтожение бактерий, устойчивых к традиционным антибиотикам;
- высокая специфичность рекомбинантного фага в отношении специфических бактериальных видов;
- репродукция фага является самоограничивающейся;
- в случае инфекций, вызываемых Helicobacter pylori, подвижность Helicobacter pylori могла бы помочь распространить фаг по всем частям слизистой оболочки желудка.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем описании и примерах термины "стандартные протоколы" и "стандартные процедуры" следует понимать как протоколы и процедуры, имеющиеся в обычном лабораторном руководстве, например: Sambrook, J., Fritsch, E. F. and Maniatis, Т. (1989) Molecular Cloning: A laboratory manual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
Согласно первому аспекту в изобретении предложен модифицированный бактериофаг для использования в лечении или профилактике бактериальной инфекции, причем указанный бактериофаг имеет на своей поверхности рекомбинантный белок, содержащий
(1) первый компонент, полученный из поверхностного белка бактериофага; и
(2) второй компонент, содержащий последовательности вариабельной области антитела, для создания сайта связывания бактериального антигена, причем указанный второй компонент делает указанный бактериофаг способным связываться с бактериальными клетками, вовлеченными в этиологию указанной инфекции, и тем самым ингибировать их рост.
Указанный модифицированный бактериофаг может, например, представлять собой модифицированный нитевидный фаг, такой как модифицированный М13 фаг.
Указанная бактериальная инфекция может представлять собой, например, бактериальную инфекцию слизистой оболочки, такую как инфекция Helicobacter pylori. Однако настоящее изобретение не ограничено фагами, способными поражать клетки Helicobacter pylori, а скорее включает в себя фаги с измененными свойствами, которые могут быть использованы для поражения широкого круга бактерий. Следует понимать, что фаг по изобретению, который специфичен для некоторых бактериальных видов, может быть получен специалистом на основе настоящего изобретения. Фаги согласно изобретению пригодны для лечения любой бактериальной инфекции слизистой оболочки, доступной внешнему миру. Примерами таких эпителиев слизистых оболочек являются слизистая носа, легкого, желудочно-кишечного тракта, мочевого пузыря и вагины.
Примерами других бактериальных инфекций, которые могут быть излечены фагами по изобретению, являются:
- инфекции мочевых путей, вызываемые E.coli, Staphylococcus saprophyticus, Klebsiella spp, Proteus spp или Pseudomonas aeruginosa:
- вагинальные инфекции, вызываемые Clamydia:
- носовые/тонзиллярные/легочные инфекции, вызываемые Streptococcus, Staphylococcus, Haemophilus influence, Pneumococcus и Mycoplasma pneumonic:
- инфекции желудочно-кишечного тракта, вызываемые Salmonella, Shigella, Yersinia, Campylobacter jejuni, Campylobacter соli, Helicobacter, Vibrio cholera или E.coli.
Указанный первый компонент рекомбинантного белка, упомянутый выше, предпочтительно может быть получен из белка, ответственного за адсорбцию немодифицированной формы указанного бактериофага на бактериальной фимбрии, например белка g3р из М13 фага.
Указанный второй компонент указанного рекомбинантного белка может, в частности, содержать рекомбинантный одноцепочечный Fv (ScFv) полипептид. Следовательно, указанный рекомбинантный белок может представлять собой, например, g3р-ScFv слитый белок.
В предпочтительной форме бактериофаг согласно изобретению представляет собой бактериофаг для использования в лечении или профилактике инфекции Helicobacter pylori, в котором последовательности вариабельной области антитела указанного рекомбинантного полипептида являются последовательностями вариабельной области моноклонального антитела, выбранного из моноклональных антител гибридомных клеточных линий 5F8 (ЕСАСС 95121524), 2Н6 (ЕСАСС 95121526) и 5D8 (ЕСАСС 95121527).
Таким образом, бактериофаг согласно изобретению может, в частности, представлять собой модифицированный М13 бактериофаг, обозначенный как В8, депонированный в NCIMB под вступительным номером NCIMB 40779, или его производное, которое сохраняет способность связываться с Helicobacter pylori и инфицировать ее.
Фаги с желаемыми свойствами могут быть получены, например, одним из следующих способов:
(а) Скрининг встречающихся в природе фагов или фаговых библиотек, содержащих фаги, экспрессирующие множество вариабельных фрагментов антител. Фаговые библиотеки могут быть получены, например, из иммунных клеток от большого числа индивидуумов. Из-за огромной генетической вариабельности такие большие фаговые библиотеки, очевидно, содержат желаемые специфические фаги, нацеленные на бактерии, действию которых предварительно подвергались индивидуумы.
(б) Осуществление мутаций существующих фагов. Мутации происходят во всех живущих организмах, включая фаги. Частота мутаций может быть увеличена, например, химическим путем или посредством облучения коротковолновым электромагнитным излучением.
(в) Направленная генетическая модификация одной или более чем одной аминокислоты или другие модификации, например, углеводных или липидных компонентов области связывания фага для усиления желаемых свойств природного или рекомбинантного фага. Пример такого подхода описан далее в экспериментальном разделе. Бактериофаг согласно изобретению, таким образом, может быть получен способом, при котором (а) выделяют антитело против бактериальной клетки; (б) выделяют ДНК, кодирующую вариабельную область тяжелой и легкой цепей указанного антитела: и (в) вводят указанную ДНК в фаговую ДНК таким образом, что указанные области антитела будут экспрессироваться на поверхности фага.
Согласно другому аспекту в изобретении предложена фармацевтическая композиция, содержащая бактериофаг согласно изобретению в смеси с фармацевтически приемлемым носителем или эксципиентом.
Примеры пригодных средств введения фагов согласно изобретению включают в себя:
- спрей для назального и легочного применения:
- предварительная обработка омепразолом, а затем - фагами, суспендированными в бикарбонатных буферах, для лечения слизистой оболочки желудочно-кишечного тракта;
- смеси слизисто-адгезивных гелей (например гелей на основе целлюлозы, поликарбофила, полоксамера и так далее) для слизистой оболочки желудка и вагинальной слизистой оболочки;
- бикарбонатные буферы для использования в мочевом пузыре.
Количество фагов, подлежащее введению, может быть определено специалистом. В зависимости от типа инфекции количество фагов, подлежащих введению, может лежать в интервале от 104 до 1010.
Согласно еще одному аспекту в изобретении предложен способ лечения бактериальной инфекции у млекопитающего, при котором вводят бактериофаг или фармацевтическую композицию согласно изобретению. Указанные бактериальные инфекции могут представлять собой, в частности, бактериальные инфекции слизистых оболочек, такие как Helicobacter pylori инфекции. В изобретение включено также использование бактериофага согласно изобретению в изготовлении лекарственного средства для лечения или профилактики бактериальной инфекции слизистых оболочек, в частности инфекций Helicobacter pylori.
Дополнительными аспектами изобретения являются гибридома, выбранная из 5F8 (ЕСАСС 95121524), 2Н6 (ЕСАСС 95121526) и 5D8 (ЕСАСС 95121527), а также моноклональное антитело, выбранное из моноклональных антител, продуцируемых указанными гибридомами. Гибридомная технология, в которой продуцирующие антитело В-клетки от иммунизированных животных сливают с клетками миеломы и выбирают полученные в результате гибридомные клеточные линии, продуцирующие желаемое антитело, хорошо известны из уровня техники.
ПРИМЕРЫ
ПРИМЕР 1: Получение моноклональных антител против Н. Pylori
1.1 Препарат антигена
Штаммы Н. pylori 17874, 25, 66, 253 и 1139 (получены из Astra Hassle, Sweden) культивируют на колумбийском агаре, обогащенном 8,5% лошадиной крови, 10% лошадиной сыворотки, 1% изовиталекса, в микроаэрофильных условиях 1 в установке Anaerocult С при +37oС.
Для получения поверхностного белка Н. pylori следуют известной (Ма J-Y et al. (1994) Scand. J. Gastroenterol. 29, 961-965) методике. А именно общее количество, равное 4 г, пяти штаммов Н.pylori инкубируют в течение 15 минут при комнатной температуре в 100 мл 0,2 М глицин-НСl (рН 2,2). рН нейтрализуют 1 М NaOH. Препарат антигена центрифугируют при 10000хg в течение 10 минут при +4oС. Осадок отбрасывают, а супернатант диализируют в течение ночи против дистиллированной воды при +4oС и затем используют как "Н.pylori антигенный препарат" или "Н.pylori поверхностные белки".
1.2. Получение моноклонального антитела
Процедуру иммунизации проводят по существу так, как описано (Саbеrо, J. L. et al. (1992) Acta Physiol. Scand. 144, 369-378). А именно 2 мг/мл поверхностного белка Н. pylori эмульгируют с равным объемом полного адъюванта Фрейнда (Freund's) при +4oС. Двух самок DBA/1 мышей инъецируют в задние стопы разовой дозой 50 мкл эмульсии антигена. Через 11 дней после иммунизации лимфоциты из подколенных лимфатических узлов сливают с клетками мышиной миеломы (sp2 линия), используя 50% (мас.) ПЭГ 4000. Суспензию слитых клеток распределяют затем по пяти титрационным микропланшетам. Все клетки выращивают в DMEM (модифицированная по Дульбекко среда Игла) культуральной среде, содержащей 10% фетальной телячьей сыворотки плюс 50 мкг/мл гентамицина.
1.3. Твердофазный иммуноферментный анализ (ELISA)
Иммунологические планшеты сенсибилизируют 50 мкл 0,05 М Nа2СО3/NаНСО3 буфера, рН 9,8, содержащего указанный антиген (10 мкг/мл) и инкубируют в течение ночи при +4oС. Свободные сайты связывания блокируют с помощью ЗФР, содержащего 0,05% Твин-20 (ЗФР-Т) при 37oС в течение 1 часа. Добавляют первичный супернатант антитела и инкубируют при +37oС в течение 1 часа. Конъюгат козий анти-мышиный lgG-пероксидаза используют в качестве вторичного антитела. Связанную пероксидазу определяют с помощью 0,04% (мас./об.) OPD и 14 мМ перекиси водорода в смеси 0,1 М лимонная кислота/О,2 М NaHPO4, рН 5. Планшеты анализируют при 492 нм после остановки реакции добавлением 2 М H2SO4. Между каждой инкубацией проводят три промывания ЗФР-Т.
1.4. Начальный скрининг
45 гибридомных клонов, полученных после слияния клеток лимфатических узлов и клеток миеломы, были позитивными против Н. pylori поверхностных белков при проведении ELISA. 8 из них отчетливо окрашивали Н.pylori, взятый из культуры на агаровой пластинке, посредством иммуногистохимии. Гибридомные клоны, обозначенные как 2Н6 (ЕСАСС 95121526), 5D8 (ЕСАСС 95121527) и 5F8 (ЕСАСС 95121524), давали более сильную реакцию против Н.pylori, чем остальные клоны, и они были отобраны для дальнейших исследований.
ПРИМЕР 2: Получение рекомбинантного фага М13 против Н.pylori
2.1. Материалы
QuickPrep mRNA purification kitТМ, Mouse ScFv Module kitТМ, Expression Module kitТМ, Detection Module KitТМ, Sfil, Notl, T4 ДНК лигаза и анти-М13 овечье антитело получили от Pharmacia Biotech (Uppsala, Sweden). DNTPs смесь, 10хПЦР буфер и AmpliTaq ДНК полимераза были закуплены у Perkin Elmer. Бакто-дрожжевой экстракт, бакто-триптон, бакто-агар были закуплены в Difco Laboratories (Detroit, Michigan USA). Пластины колумбийского агара и бруцеллярный бульон были получены из отдела микробиологии (Linkoping University, Sweden). Anaerocult® был получен от Merck (Germany). SlowFadeТМ набор против обесцвечивания был получен из Milecular Probes Inc. (U.S.A).
2.2. Конструирование библиотеки фаговых антител
Для экспрессии фрагментов антител как слитых белков, проявляющихся на фаговой поверхности, использовали the Recombinant Phage Antibody SystemТМ (Pharmacia).
Общую мРНК выделяли из гибридомных клеточных линий (2Н6, 5D8 и 5F8) и очищали посредством афинной хроматографии на олигo(dТ)-целлюлозе, используя QuickPrep mRNA Purification KitТМ (Pharmacia).
С помощью Mouse ScFv Module KitТМ осуществляют следующие стадии:
- Первую нить кДНК синтезируют с гибридомной мРНК с помощью обратной транскриптазы и смесей праймеров из Mouse ScFv Module KitТМ.
- К-ДНК, соответствующую вариабельным областям тяжелых и легких цепей mAbs (моноклональных антител), амплифицируют с различными праймерами (VH и VL праймеры цепи, из набора) посредством полимеразной цепной реакции (ПЦР). VH и VL цепи анализируют посредством электрофореза в 1,5%-ном агарозном геле. Получали отдельные полосы правильного размера для VH (340 пн) и VL (325 пн) цепи.
- Амплифицированные VH и VL цепи очищают и выделяют посредством электрофореза в 1%-ном агарозном геле, а затем собирают в одноцепочечный Fv (ScFv) ген с помощью ДНК линкерного фрагмента из набора. Линкерный фрагмент сконструирован таким образом, что один конец отжигается к 3'-концу тяжелой цепи, в то время как другой конец гибридизуется с 5'-концом легкой цепи. После электрофореза наблюдали отдельную полосу правильного размера для ScFv гена (750 пн).
- ДНК фрагмент собранного антитела ScFv амплифицируют с набором олигонуклеотидных праймеров (из набора), посредством чего вводят Notl и Sfil сайты. Рестрикции. Фрагмент очищают на стеклянной колонке (из набора) для удаления линкеров, dNTPs и Taq ДНК полимеразы. ScFv фрагмент расщепляют с помощью Notl и Sfil для получения липких концов для лигирования с pCANTAB5 вектором.
С помощью Expression Module KitТМ осуществляют следующие стадии:
- ScFv фрагмент лигируют с фагмидным вектором pCANTAB5 (из набора), предварительно обработанным Notl и Sfil для получения липких концов. Для соединения концов фрагмента с соответствующими концами фагмиды используют Т4 ДНК лигазу. Затем ScFv фрагмент ориентируют в правильном направлении так, чтобы он примыкал к М13 гену 3 и был с ним в одной рамке считывания, для экспрессии ScFv-g3р слитого белка.
- Клетки E.coli TG1 клетки (из набора) делают компетентыми и трансформируют рекомбинантной фагмидой, содержащей lас промотор и маркер устойчивсти к ампициллину. Трансформированные клетки выращивают при +30oС в среде, содержащей глюкозу и ампициллин. Получили 3,2 х 104 ScFv клонов. Колонии, устойчивые к ампициллину, переносят в среду для получения библиотечной биомассы.
- Клетки, устойчивые к ампициллину, инфицируют хелперным фагом М13К07 (из набора), несущим маркер устойчивости к канамицину, и выращивают в глюкозо-дефицитной среде, содержащей ампициллин и канамицин. В отсутствие глюкозы lас промотор, присутствующий в фагмиде, больше не репрессируется. В результате продуцируются и высвобождаются из клеток фаговые частицы, несущие фрагменты рекомбинантного антитела на своих концах.
- Антитела, присутствующие на фаге, способные связываться с Н. pylori антигеном, отбирают путем пэннинга против антигена. Культуральную колбу сенсибилизируют 5 мл поверхностного белка Н.pylori (15 мкг/мл в 50 мМ буфере из карбоната натрия, рН 9,6) в течение ночи. После трех промывок ЗФР колбу, содержащую 10 мл 1%-ного БСА (мас./об.) в ЗФР, инкубируют при +37oС в течение 2 часов в фаговом супернатанте (среда, содержащая фаг). Затем колбу промывают 20 раз ЗФР, содержащим 0,1% (мас./об.) Твин-20, и 20 раз - ЗФР. Связанные фаговые частицы затем элюируют добавлением 1 мл 100 мМ триэтиламина при осторожном перемешивании в течение 10 минут и немедленно нейтрализуют 0,5 мл 1 М Трис-HCI, рН 7,4.
Элюированный фаг используют для инфицирования клеток E.coli TG1 в лог-фазе на SOBAG агаре, содержащем 2% бакто-триптона, 0,2% бакто-дрожжевого экстракта, 0,05% NaCl, 0,1 М MgCl2, 0,01% глюкозы и 0,01% ампицилина. Колонии отбирают, переносят, выращивают и снова "спасают" с помощью М13К07.
После первого раунда селекции путем подсчета 100 клонов 6% "спасенных" клонов из титрационного микропланшета были позитивными против антигенного препарата Н. pylori в тесте ELISA. В третьем раунде селекции из "спасенных" на титрационном микропланшете 95% фаговых антител из отдельных клонов реагировали с Н.pylori антигенами.
В фаговом тесте ELISA с использованием Н. pylori антигенного препарата в качестве антигена рекомбинантный фаг В8 имеет титр, в 10 раз более высокий, чем хелперный фаг (широкий фаг). Фаг В8 (NCIMB 40779) был выбран для дальнейших анализов.
ПРИМЕР 3: ELISA
Фаго-проявляемые рекомбинантные антитела выявляют и идентифицируют в твердофазном иммуноферментном анализе (ELISA) с использованием Detection Module KitТМ.
96-луночные титрационные микропланшеты сенсибилизируют 200 мкл Н.pylori антигена (10 мкг/мл в 50 мМ Nа2СО3/NаНСОз, рН 9,6) и инкубируют в течение ночи при +4oС. Лунки три раза промывают ЗФР, содержащим 0,05% Твин 20 (ЗФР-Т) и затем блокируют 300 мкл ЗФР, содержащего 1% БСА, в течение 1 часа при +37oС. Антитела рекомбинантного фага разбавляют равным объемом 1% БСА/ЗФР и инкубируют в течение 20 минут при комнатной температуре. После промывки добавляют 5х1010 фаговых трансдуцирующих единиц (200 мкл/лунку) и инкубируют 2 часа при +37oC.
Лунки три раза промывают ЗФР-Т и затем добавляют НRР(пероксидаза из хрена)/анти М13 конъюгат из Detection Module kit, разведенный 1:5000 в 1% БСА/ЗФР, и инкубируют в течение 1 часа при +37oС. Лунки три раза промывают ЗФР-Т и затем добавляют 2'2'-азино-бис(3-этилбензтиазолин-6-сульфоновая кислота) диаммоний (ABTS) и H2O2 в качестве субстрата для пероксидазы и инкубируют при комнатной температуре в течение 30 минут. Регистрируют абсорбцию при 405 нм с помощью компьютеризированного ELISA датчика. В качестве контрольного антигена добавляют овальбумин (10 мкг/мл в ЗФР). Хелперный фаг используют в качестве негативного контроля. Результаты подтвердили, что рекомбинантный фаг В8 специфически связан с Н.pylori поверхностным антигеном.
ПРИМЕР 4: Иммуноблоттинг
Белки Н.pylori антигенного препарата разделяют посредством электрофореза в полиакриламидном геле в ДДС-ПААГ (10 мкг белков/ лунку) и затем переносят на нитроцеллюлозную бумагу в мини транс-блот электрофоретическом переносчике клеток (BioRad, Richmond, CA, USA). Нитроцеллюлозную бумагу блокируют 10%-ным БСА в ЗФР, содержащем 0,1% Твин 20, в течение 1 часа при комнатной температуре при осторожном перемешивании. Затем добавляют фаг В8 (1011 трансдуцирующих единиц/мл) и инкубируют вместе с нитроцеллюлозной бумагой в течение ночи при осторожном перемешивании при +4oС. Отсутствие первичного антитела используют в качестве негативного контроля. После промывания ЗФР-0,1% Твин 20 нитроцеллюлозную бумагу инкубируют с конъюгатом НRТ/анти-М13 (1:5000 разведение в блокирующем буфере) в течение 1 часа при перемешивании. Выявление связывания осуществляли с помощью ELC Detection Kit (Amersham, UK).
После окрашивания нитроцеллюлозной бумаги амидо черным главные полосы соответствовали белкам 64 кДа, 36 кДа, 31 кДа и 27 кДа. Объединенные MAbs (2H6, 5D8 и 5F8, соответствующие гибридомам, использованным для конструирования фагов) взаимодействуют с полосами 32 кДа и 64 кДа. Аналогичное окрашивание было получено при иммуноблоттинге с фаговым антителом В8. Этот результат свидетельствует о том, что гены вариабельных тяжелых и легких цепей, соответствующих H. pylori специфическим моноклональным антителам, правильно экспрессировались на фаге.
ПРИМЕР 5: Действие рекомбинантного фага на бактерии
В следующих экспериментах бактерии культивируют при +37oС в бруцеллярном бульоне, содержащем 5% фетальной телячьей сыворотки, в атмосфере, содержащей 10% СО2 и 5% O2.
Эксперимент начинают ("время 0"), когда 20 мкл из бактериального запаса смешивают с 10 мл бульона. В указанные моменты времени определяют количество КОЕ (колониеобразующие единицы) на 1 мл культуры путем разведения аликвот культур в ЗФР и распыления разведений по поверхности агаровых пластинок. Пластинки инкубируют два дня при +37oC и подсчитывают количество колоний на каждой пластинке.
5.1. Зависящее от времени действие
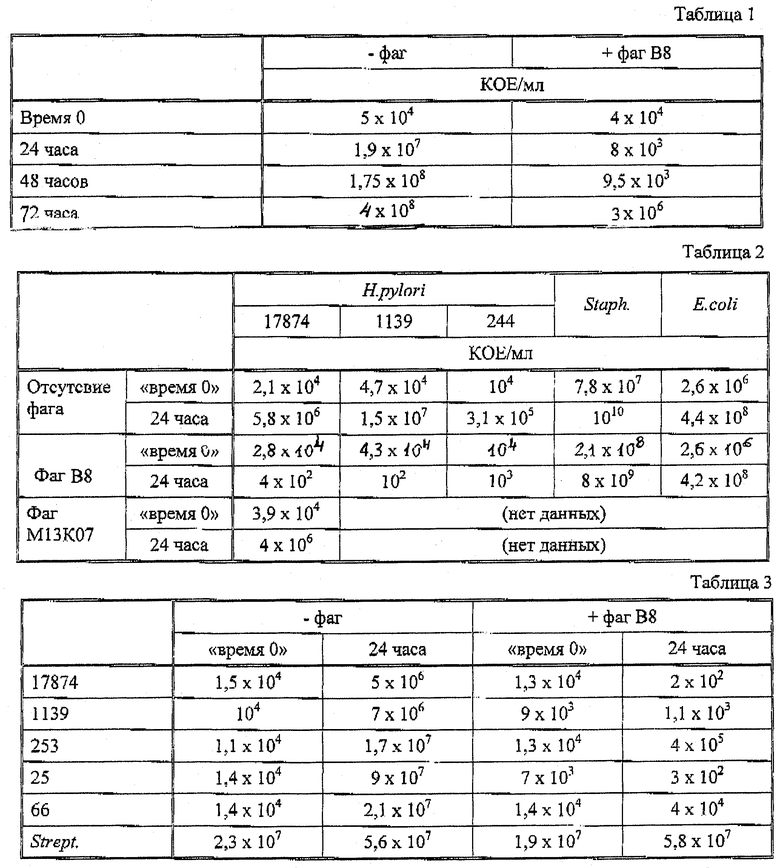
Зависящее от времени действие рекомбинантного фага В8 на рост штамма 17874 H.pylori исследуют путем измерения КОЕ в течение трех дней при наличии фага и без него (Таблица 1). К 10 мл среды в момент "время 0" добавляют 106 фагов ("+фаг"). В контроле фаги не добавляют ("-фаг").
Спустя 3 дня, КОЕ увеличивалось примерно на 5 порядков величины в отсутствие рекомбинантного фага. Спустя сутки в присутствии фага, имело место снижение КОЕ примерно на один порядок величины, и культуральный бульон изменял свой вид от мутного до менее мутного раствора.
5.2. Действие на различные бактериальные штаммы
Лабораторные штаммы Н. pylori 17874, 1139 и 244, а также Staphylococcus aureus, ATCC 29213 и E.coli TG1 культивируют с фагом (106 фагов на 10 мл среды) или без него в течение 24 часов. КОЕ анализируют в момент "время 0" и через 24 часа (Таблица 2). Рекомбинантный фаг снижал КОЕ трех Н.pylori тестировавшихся штаммов, но не влиял на Staphylococcus или e.coll. Хелперный фаг М13К07, используемый в качестве контроля, не влиял на H.pyldri 17874.
5.3. Действие на штаммы Н. pylori
Во втором эксперименте (Таблица 3) штаммы Н.pylori 17874, 1139, 253, 25 и 66 тестируют вместе со Streptococcus (Raf M87). Без фага КОЕ возрастало в течение 24 часов инкубации у всех тестировавшихся бактерий. Однако в культуре с присутствующим рекомбинантным фагом (106 фагов на 10 мл среды в момент "время 0") значения КОЕ штаммов Н.pylori 17874, 1139 и 25 снижались по количеству, а скорость роста штаммов 253 и 66 была сильно понижена по сравнению с контролем. Таким образом, тестировавшиеся штаммы Н.pylori были подвержены действию фагов.
ПРИМЕР 6: Иммунофлуоресцентное окрашивание Н.pylori
Фаговое антитело В8 концентрируют ПЭГ осаждением и проводят флуоресцентное окрашивание на Н.pylori (17874 штамм), взятый из культуры.
Для проведения иммунофлуоресцентного окрашивания готовят следующие запасные реагенты:
Реагент А: 1012 трансдуцирующих единиц/мл фаговых антител разбавляют 1:1 с помощью 1% БСА/ЗФР;
Реагент Б: овечий анти-М13 IgG, разведенный 1% БСА/ЗФР;
Реагент В: анти-овечий IgФИТЦ конъюгат, разведение 1:100 в БСА/ЗФР;
Реагент Г: 1 мг (мас./об.) проназы в ЗФР.
Через три дня культивирования Н. pylori (5 различных штаммов), Staphylococcus aureus (ATCC 29213) и Streptococcus (Raf M 87) по-раздельности суспендируют в ЗФР, содержащем 1% БСА. На стеклянные предметные стекла наносят по 20 мкл суспензии каждой бактерии. Бактерии сушат на воздухе и фиксируют в нейтральном формалине в течение 2 минут, затем промывают путем шестикратного погружения предметных стекол в воду и сушат на воздухе при комнатной температуре. Короткая обработка в реагенте Г увеличивала соотношение сигнала к помехам при сравнении предметных стекол с образцами с контролем. Предметные стекла последовательно инкубируют с 30 мкл реагентов А, Б и В в течение 30 минут каждый раз, и между инкубациями промывают в ЗФР. Предметные стекла сушат на воздухе и перед покрытием добавляют 1 каплю SlowFade. Все инкубации проводят при комнатной температуре. Негативные контроли готовят путем исключения первичного антитела.
Результаты показывают, что Н.pylori позитивно окрашивается фаговым антителом. Результаты свидетельствуют о хорошем согласовании со значениями КОЕ, полученными из культуральных экспериментов. Таким образом, экспрессия специфического антигена на поверхности бактерий, очевидно, является предпосылкой для осуществления биологического действия фагом В8.
ХРАНЕНИЕ БИОЛОГИЧЕСКОГО МАТЕРИАЛА
Согласно Будапештскому договору в Европейской коллекции культур животных клеток (ЕСАСС), Salisbury, Wiltshire, U.K., 15 декабря 1995 года были депонированы следующие гибридомные клоны:
2Н6 (ЕСАСС No. 95121526);
5D8 (ЕСАСС No. 95121527);
5F8 (ECACC No. 95121524).
Рекомбинантный фаг В8 был депонирован согласно Будапештскому договору в Национальных коллекциях промышленных и морских бактерий (NCIMB), Aberdeen, Scotland, 20 декабря 1995 года под вступительным номером NCIMB 40779.
Изобретение относится к бактериофагам для использования в лечении или профилактике бактериальных инфекций, особенно бактериальных инфекций слизистых оболочек. С этой целью была получена конструкция бактериофага, несущего на своей поверхности рекомбинантный белок, содержащий поверхностный белок бактериофага и последовательности вариабельной области ScFv полипептида. Штамм модифицированного бактериофага М13 и конструкция пригодны для лечения или профилактики бактериальной инфекции в составе фармкомпозиции, а штамм, конструкция и фармкомпозиция используются в способе лечения бактериальной инфекции; также были получены моноклональные антитела из гибридом 5F8, 2Н6 и 5D8, которые обеспечивают связывание модифицированного бактериофага М13 с Helicobacter pylori. 8 с. и 9 з.п. ф-лы, 3 табл.
| US 5478731 С1, 26.12.1995. |

