Настоящая заявка является частично продолжающей заявкой одновременно рассматриваемой заявки серийный номер 08/943608, поданной 3 октября 1997 года, которая полностью включена сюда в качестве ссылки.
1. Область изобретения
Изобретение относится к области клеточной биологии опухолей и лечения злокачественных опухолей. Более конкретно изобретение относится к специфической экспрессии гетерологичных генов, в частности генов, кодирующих цитотоксические продукты, в опухолевых клетках.
2. Предпосылки изобретения
2.1 Ген Н19
Ген H19 является одним из немногих генов, для которых известно, что они подвергаются импритингу у людей ("Hurst et al., 1996, Nature Genetics 12: 234-237). На самых ранних стадиях эмбриогенеза ген H19 экспрессируется обоими аллелями хромосом (DeGroot et al., 1994, Trofoblast 8:285-302). Чуть позже происходит подавление отцовского аллеля, и транскрибируется только унаследованный от матери аллель. Ген H19 является избыточно экспрессирующимся в ходе эмбриогенеза и впервые был идентифицирован как ген, который соответственно регулируется альфа-фетопротеином в печени с помощью трансвзаимодействия с локусом raf (Pachnis et al., 1984, Proc. Natl. Acad. Sci. USA 81: 5523-5527). Кроме того, ген Н19 был независимо клонирован рядом лабораторий, применяющих скрининг с целью выделения генов, экспрессируемых в ходе тканевой дифференцировки. Например, Davis et al. (1987, Cell 51:987-1000) идентифицировали гомолог гена Н19 у мыши при скрининге генов, активных на ранних этапах дифференцировки клеток С3Н10Т1/2. Pourier et al. (1991, Development 113:1105-1114) обнаружили, что ген Н19 мышей экспрессируется в ходе дифференцировки стволовых клеток и в период имплантации. Транскрипция человеческого гена Н19 также была обнаружена при дифференцировке цитотрофобластов плаценты человека (Rachmilewitz et al., 1992, Molec. Reprod. Dev.32: 196-202).
Несмотря на то что транскрипция РНК гена H19 происходит в ряде различных эмбриональных тканей в течение всего эмбрионального периода и развития плаценты, после рождения наблюдается даун-регуляция экспрессии гена Н19. Однако имеются данные об относительно низких уровнях транскрипции гена H19 в мышечной ткани и печени взрослых мышей (Brannan et al., 1990, Molec. Cell. Biol. 10: 28-36). Ген Н19 также постнатально активен в малигнизированных клетках. Ariel et al. (1997, Molec. Pathol. 50:34-44) показали экспрессию гена H19 в ряде опухолей, происходящих из тканей, в которых ген H19 экспрессирует пренатально. Кроме того, данные авторы обнаружили РНК гена H19 в опухолях, происходящих из нервных тканей, в частности в астроцитоме и ганглионейробластоме, связь которых с экспрессией гена H19 неизвестна. Получив данные по широкому кругу злокачественных опухолей, экспрессирующих РНК гена H19, эти авторы сделали предположение, что ген H19 экспрессирует онкофетальную РНК, и предложили исследование гена H19 как опухолевого маркера новообразований человека.
Как человеческий, так и мышиный гены H19 были клонированы и секвенированы (Brannan et al., 1990, Molec. Cell. Biol. 10:28-36). Сравнение генов Н19 человека и мыши позволило выявить в общей сложности 77%-ную идентичность нуклеотидных последовательностей. Несмотря на данную консервативность межвидовой гомологичности нуклеотидных последовательностей, могла быть предсказана очень низкая идентичность аминокислотных последовательностей с открытых рамок считывания двух данных генов (Id). Далее хотя РНК гена H19 транскрибируется с помощью РНК-полимеразы II, сплайсируется и полиаденилируется, она не подвергается трансляции. Вместо этого была найдена взаимосвязь РНК гена H19 с цитоплазматической 28S РНК, что позволило предположить, что РНК гена H19 может иметь функцию РНК-компонента рибонуклеопротеина (Id).
Настоящая физиологическая роль гена H19 понятна не до конца. Ген H19 может действовать как доминантный летальный ген; высокая эктопическая экспрессия гена Н19 у трансгенных мышей является причиной летальности незадолго перед рождением (Brunkow et al., supra). Данный летальный период совпадает со временем подавления транскрипции гена H19. С другой стороны, не было обнаружено повреждения ни у гетерозиготных, ни у гомозиготных нокаут-мышей по H19 аллелю (Leighton et al., 1995, Nature 375:34-39). Выбивание материнского наследственного аллеля мешает проведению импритинга генетически связанного и противоположно подверженного импритингу гена IGF-2; получившиеся мыши рождаются крупнее, чем остальное потомство, вследствие увеличения пренатальной экспрессии гена IGF-2 (Id). Поскольку данные два противоположно подверженных импритингу гена разделены цис-взаимодействующими регуляторными последовательностями, Leighton и коллеги сделали предположение, что ген H19 может быть вовлечен в импритинг гена IGF-2.
Другой предполагаемой функцией продукта гена H19 является РНК-супрессия опухоли. Нао et al. (1993, Nature 365:764-767) сообщили, что трансфекция в две эмбриональные опухолевые клеточные линии RD и G401 экспрессирующей конструкции гена H19 приводит к задержке клеточного роста, морфологическим изменениям и уменьшению туморогенности у мышей nude. Подобная опухолесупрессорная активность была замечена в случаях, согласующихся с наблюдаемой летальностью при эктопической экспрессии у мышей (Нао et al., supra) и также увеличением размера мышей с выключенным материнским аллелем Н19 (Leighton et al. , supra). Однако предположение о том, что ген H19 имеет функцию опухолевого супрессора является спорным. Некоторые из опубликованных результатов не подтвердились и возможно существование другого кандидата гена опухолевого супрессора, тесно связанного с геном H19 (Ariel et al., supra). Предположительная роль гена H19 как опухолевого супрессора также не согласуется с экспериментальными данными о том, что ген H19 активируется во множестве опухолевых клеток (смотри, например, Listig-Yariv et al., 1997, Oncogene 23: 169-177).
2.2 Гены инсулиноподобного фактора роста (IGF)
IGF-2 является другим подверженным импритингу геном, экспрессия которого зависит от его родительского источника происхождения. Однако, в отличие от гена Н19, ген IGF-2 является подверженным материнскому импритингу как у мышей, так и у человека, и поэтому экспрессируется с унаследованного от отца аллеля (Rainer et al., 1993, Nature 363:747-749). Ген IGF-2 человека проявляет сложный транскрипционный профиль. Существуют четыре промотора для гена IGF-2, активирующиеся в ткани на определенной стадии развития. Только три промотора Р2, Р3 и Р4 подвержены импритингу и активны в течение периода развития плода и в злокачественных опухолевых тканях. Четвертый промотор Р1 не подвержен импритингу и активируется только во взрослом состоянии в печени и сосудистом сплетении (смотри Holthuizen et al., 1993, Mol. Reprod. Dev. 35: 391-393). Промотор Р3 гена IGF-2 вовлекается при развитии цирроза печени и гепатоцеллюлярной карциномы (Kim and Park, 1998, J.Korean Med. sci. 13: 171-178).
Потеря импритинга гена IGF-2 наблюдается при нефробластоме (Ogawa et al. , 1993, Nature 363:749-751). Данное наблюдение привело многих исследователей к предположению, что потеря импритинга и экспрессия подверженных импритингу генов с двух аллелей может приводить к нарушениям роста и развитию злокачественных опухолей (смотри также Rainer et al., 1993, Nature 362:747-749 и Glassman et al., 1996. Cancer Genet. Cytogenet. 89:69-73).
2.3 Опухоль-специфическая генная терапия
Регуляторные последовательности опухольассоциированных генов применяли с целью селективной прицельной экспресии суицидного гена в клетках опухоли. Например, в гепатоцеллюлярной карциноме индуцируют экспрессию альфа-фетопротеина. Huber и др. (1991, Proc. Natl. Acad. Sc. USA 88:8039-8043) использовали регуляторные последовательности как гена альбумина, так и гена альфа-фетопротеина для прицельной экспрессии гена тимидинкиназы возбудителя ветряной оспы (VZV ТК), кодирующего последовательности в клетках гепатомы. Клетки гепатомы, инфицированные in vitro ретровирусным вектором, содержащим одну из данных конструкций, экспрессировали VZV TK и становились чувствительными к обычно нетоксичному пролекарственному препарату 6-метоксипуринарабинонуклеозиду (агаМ). Kaneko et al. (1995, Cancer Res. 55:5283-5287) создали аденовирусный вектор, экспрессирующий HSV ТК под контролем регуляторных последовательностей альфа-фетопротеина. Рекомбинантные аденовирусные частицы, содержащие данный вектор, непосредственно вводили в опухоли, возникшие из гепатоцеллюлярной карциномы, развившиеся у бестимусных мышей nude. Последовательные интраперитонеальные инъекции ганцикловира приводят к регрессии опухолей, развившихся из гепатоцеллюлярной карциномы.
Osaki et al. (1994, Cancer Res. 54;5258-5261) трансфицировали клетки легочной карциномы А549 экспрессирующей конструкцией, содержащей регуляторные последовательности гена легочного карциномоэмбрионального антигена, связанного с кодирующей последовательностью тимидинкиназы вируса простого герпеса (HSV ТК). Трансфицированные клетки были чувствительны к ганцикловиру. Кроме того, рост опухоли у мышей nude из подкожно введенных трансфицированных клеток ингибировали повторными интраперитониальными инъекциями ганцикловира. Однако ген карциномоэмбрионального антигена недавно был описан как экспрессирующийся в нормальной слизистой оболочке толстого кишечника, таким образом ограничивая пригодность данных регуляторных последовательностей в качестве специфических регуляторных областей (Osaki et al., supra). Таким образом, сохраняется потребность в разработке векторов для генной терапии, которые специфически экспрессируют генные продукты в опухолевых клетках.
3. Сущность изобретения
Изобретение относится к способам и композициям для индукции селективной экспрессии гетерологичных генов в опухолевых клетках. В частности, изобретение относится к полинуклеотидам, содержащим регуляторную транскрипционную последовательность, оперативно связанную с гетерологичными генами, что приводит к опухоль-специфической экспрессии гетерологичных генов. Более подробно, регуляторная транскрипционная последовательность состоит из подверженного геномному импритингу гена, который специфически экспрессируется в малигнизированных клетках, такого как ген Н19, и промоторов Р3 и Р4 гена IGF-2, и гетерологичного гена, кодирующего цитотоксический белок или цитостатический генный продукт. Другая реализация изобретения относится к промотору IGF-1, оперативно связанному с гетерологичным геном, что приводит к опухоль-специфической экспрессии гена. Регуляторные последовательности будут управлять экспрессией гена в ряде различных типов клеток злокачественных опухолей. Подобные способы и композиции применимы для лечения широкого спектра злокачественных опухолей и гиперпролиферативных состояний.
Одним аспектом изобретения являются экспрессирующие векторы, содержащие полинуклеотиды, такие как регуляторные области, оперативно связанные с гетерологичными генами. Особенно предпочтительными регуляторными областями являются участки, формирующие регуляторные области гена H19, такие как промоторная и энхансерная последовательности, промоторы Р3 и Р4 гена IGF-2 или промотор гена IGF-1. В связи с этим энхансер гена H19 и его активные участки могут быть использованы в любой комбинации с промотором гена H19, промотором гена IGF-1, промотором Р3 гена IGF-2 или промотором Р4 гена IGF-2. Также изобретение охватывает и клетки-хозяева, содержащие подобные векторы. В связи с этим экспрессирующая конструкция, содержащая гетерологичный ген, контролируемый промотором гена H19 в присутствии или в отсутствии энхансера гена H19, может быть введена в клетку совместно со второй конструкцией, включающей гетерологичный ген, контролируемый промотором гена IGF-1, промотором Р3 или промотором Р4 гена IGF-2 в сочетании с энхансером гена Н19. В другом аспекте изобретение обеспечивает способы применения подобных векторов для экспрессии гетерологичных генов в опухолевых клетках. Еще одним аспектом изобретения явления лечение злокачественных опухолей с применением векторов по настоящему изобретению методами генной терапии.
4. Краткое описание чертежей
Фиг.1А-1С - нуклеотидная последовательность промоторной области гена H19 человека. Промоторная область от -837 нуклеотидного положения до -7 (относительно точки начала транскрипции).
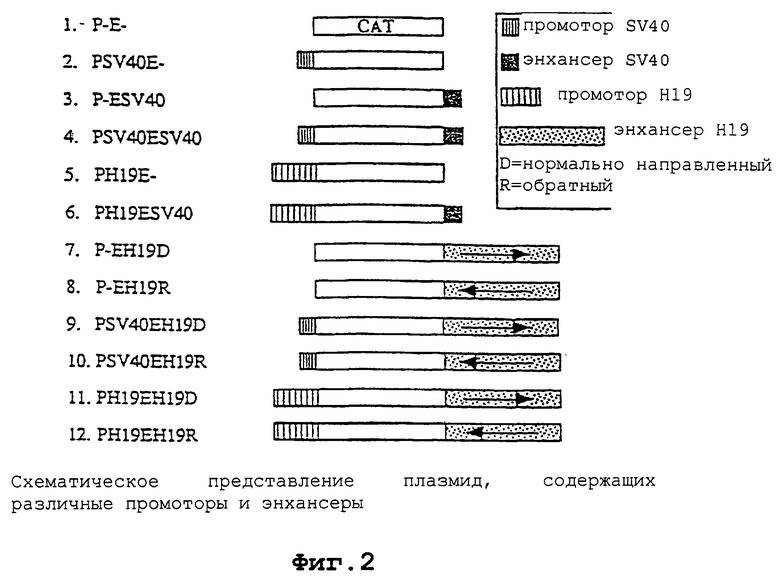
Фиг. 2 - схематическая диаграмма векторов, применяемых для экспрессии гетерологичного гена под контролем регуляторных последовательностей гена H19.
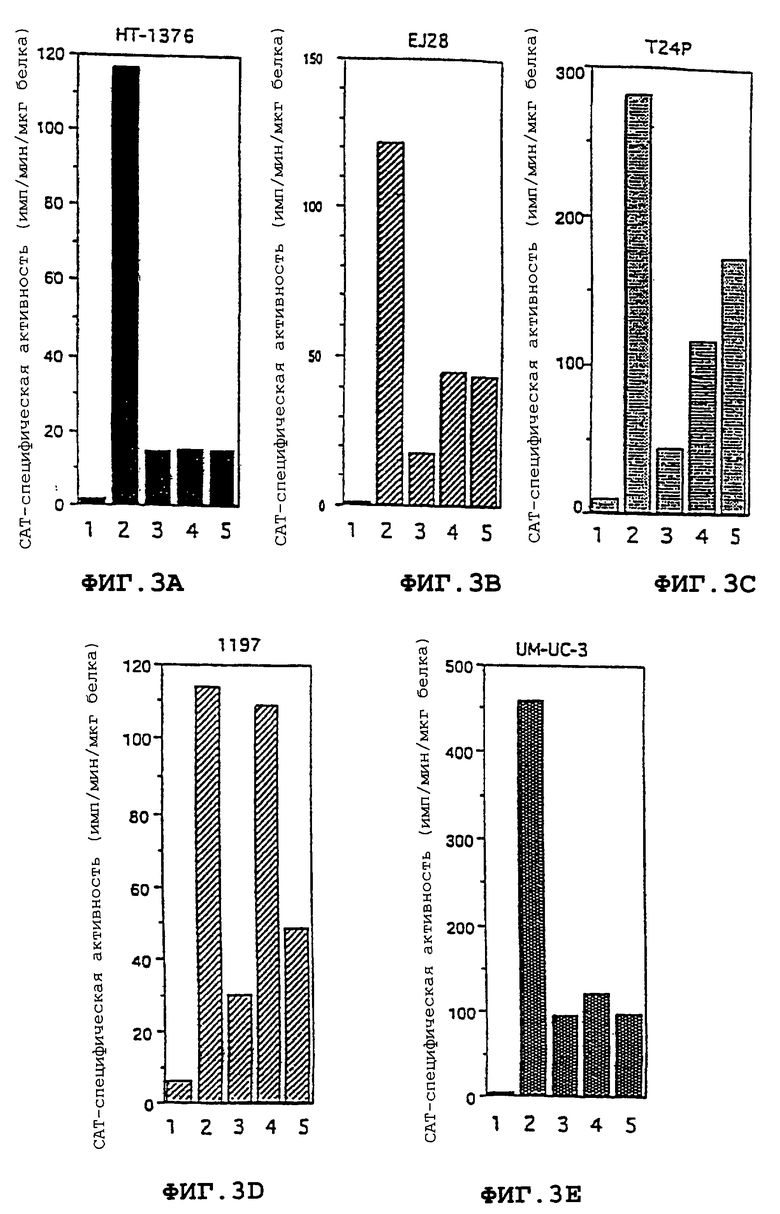
Фиг. 3А-3Е - регуляторные последовательности гена Н19 управляют экспрессией гетерологичного гена (CAT) в клеточных линиях злокачественной опухоли мочевого пузыря. Для пяти различных указанных клеточных линий CAT специфическая активность (импульс в минуту/мкг белка) изображена как функция используемого для трансфекции вектора. Фиг.3А: клетки НТ-1376. Фиг.3В: клетки EJ28. Фиг.3С: клетки Т24Р. Фиг.3Д: клетки 1197. Фиг.3Е: клетки UM-UC-3. Следующие векторы более подробно описаны ниже в части 6: (1) pCAT-Basic; (2) pCAT-Control; (3) рН19Е; (4)pHl9EH19D; (5) pH19EH19R.
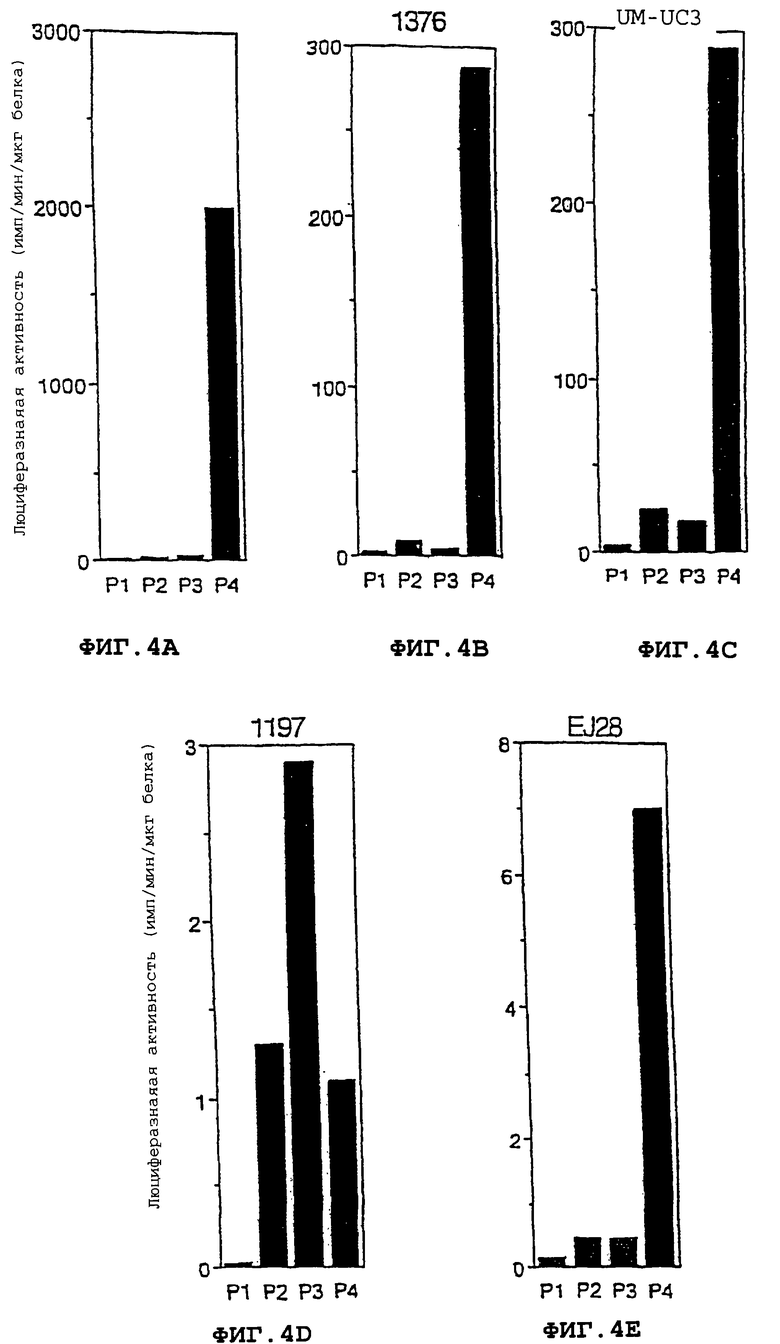
Фиг. 4А-4Е - промоторы Р3 и Р4 гена IGF-2 управляют экспрессией гетерологичного гена (люциферазы) в клеточных линиях злокачественной опухоли мочевого пузыря. Для пяти различных клеточных линий показана специфическая люцеферазная активность (импульсов мкг белка) представлена как функция промотора гена IGF-2, применяемого в трансфекционной конструкции для управления экспрессией люциферазы Фиг.4А: клетки Т24Р. Фиг.4В: клетки 1376. Фиг.4С: клетки UM-UC3. Фиг. 4D: клетки 1197. Фиг.4Е: клетки EJ28. Более подробно векторы описаны в части 10.
Фиг. 5 - нуклеотидная последовательность промоторного фрагмента гена H19 человека (SEQ ID NO:2).
Фиг. 6 - нуклеотидная последовательность энхансерного фрагмента гена H19 длиной 0,9 т.п.н. (SEQ ID NO:3).
Фиг. 7А и 7В - нуклеотидная последовательность энхансерного фрагмента гена Н19 длиной 2 т.п.н. (SEQ ID NO:4).
Фиг. 8А-8С - нуклеотидная последовательность энхансерного фрагмента гена H19 длиной 4 т.п.н. (SEQ ID NO:5).
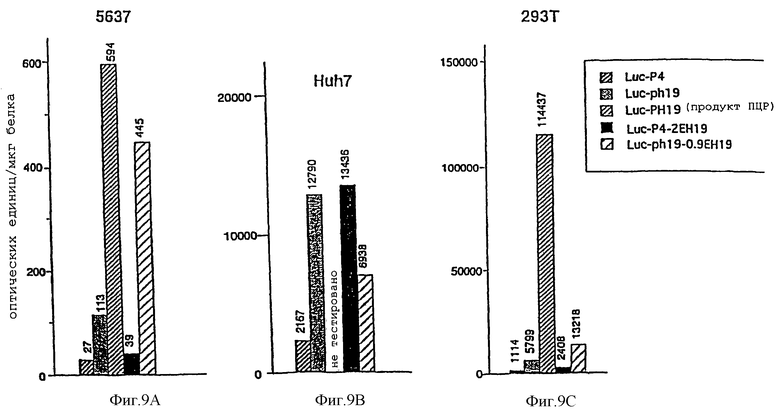
Фиг. 9А-9С - трансфекция векторами, содержащими различные комбинации регуляторной области гена H19 с промотором Р4, позволяет управлять экспрессией люциферазы в опухолевых клетках. Фиг.9А: клетки 5637. Фиг.9В: клетки Huh7. Фиг.9с: клетки 293Т.
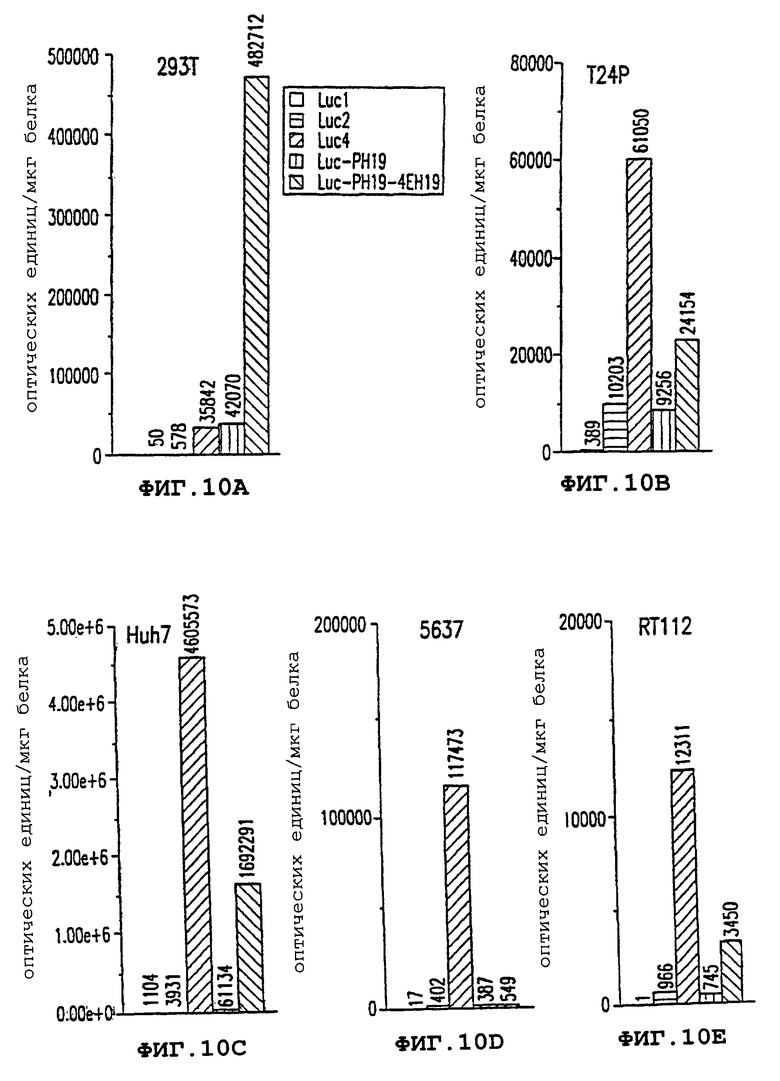
Фиг. 10А-10Е - трансфекция векторами, содержащими регуляторную область гена Н19, позволяет управлять экспрессией в опухолевых клетках. Фиг.10А: клетки 293Т. Фиг. 10В: клетки Т24Р. Фиг.10С: клетки Huh7. Фиг.10D: клетки 5637. Фиг.10Е: клетки RT112.
5. Подробное описание изобретения
Изобретение частично основывается на открытии того, что регуляторные области подверженных геномному импритингу генов, экспрессирующихся в малигнизированных клетках, могут быть использованы для прицельной экспрессии кодирующих последовательностей в интересующих малигнизированных клетках. В частности, было установлено, что экспрессия гена H19 активируется в целом ряде карцином, включая, но не ограничиваясь, карциному мочевого пузыря, гепатоцеллюлярную карциному, гепатобластому, рабдомиосаркому, карциному яичника, карциному шейки матки, карциному легких, карциному молочной железы, плоскоклеточную карциному головы и шеи, карциному пищевода, карциному щитовидной железы, астроцитому, ганглиобластому и нейробластому. Далее было обнаружено, что конструкции, содержащие промоторные области гена H19, оперативно связанные с гетерологичным геном, или промоторы Р3 или Р4 гена IGF-2, оперативно связанные с гетерологичным геном, или конструкции, содержащие подобный промотор в комбинации с ниже расположенным энхансером гена H19, специфически активируются в опухолевых клетках. В другом аспекте изобретения для управления экспрессией гетерологичного гена применяют промотор IGF-1.
Соответственно в одном из своих аспектов изобретение обеспечивает способы и композиции для изменения фенотипа или селективного уничтожения малигнизированных клеток. Данная цель выполняется с помощью внесения в клетки полинуклеотида, содержащего регуляторные области из подверженных геномному импритингу генов, которые экспрессируются в малигнизированных клетках, будучи оперативно связаны с гетерологичным геном. Гетерологичный ген, например, способен кодировать цитостатический или цитотоксический агент (например, токсин, антисмысловую РНК или рибозим).
Регуляторные области подверженных геномному импритингу гена, который экспрессируется в малигнизированных клетках, включают, не ограничиваясь, промотор и энхансер гена Н19 и промоторы Р3 и Р4 гена IGF-2.
В контексте настоящего изобретения, описанного здесь, термин "оперативно связанный" означает, что нуклеотидная последовательность связана с регуляторной последовательностью таким образом, что позволяет регуляторной последовательности управлять экспрессией нуклеотидной последовательности.
В контексте настоящей заявки термин "последовательность гетерологичного гена" относится к последовательности гена, которая обычно оперативно не связанна с регуляторными последовательностями гена H19. Как правило, последовательности гетерологичного гена включают последовательности, кодирующие цитостатические или цитотоксические генные продукты.
Термин "экспрессия", употребляемый здесь, относится к транскрипции интересующей ДНК, сплайсингу, процессингу, стабилизации и необязательно трансляции соответствующего мРНК-транскрипта. В зависимости от структуры доставляемой молекулы ДНК экспрессия может быть кратковременной или длительной.
5.1 Регуляторные последовательности гена Н19, пзомоторы Р3 и Р4 гена IGF-2 и промотор гена IGF-1
Описанные здесь регуляторные последовательности гена H19 могут быть использованы для управления специфической экспрессией в опухолевой клетке гетерологичной кодирующей последовательности. Данные регуляторные последовательности гена Н19 включают выше расположенную промоторную область гена H19 и/или ниже расположенную энхансерную область гена H19. Нуклеотидная последовательность одной промоторной области гена Н19 показана на фигуре 1А-1С (SEQ ID NO: 1). Данная 830-нуклеотидная последовательность располагается от -837 до -7 нуклеотида от сар-сайта (как описано Brannan et al., supra). Консенсусная ТАТА последовательность наблюдается в положении от -27 до -35 нуклеотида. Два консенсусных АР2 связывающих сайта (8/9 совпадений) располагаются приблизительно на -500 и -40 нуклеотидов выше сайта инициации транскрипции. Будучи расположены выше кодирующей области гетерологичного гена, что обсуждается более подробно ниже, приблизительно 830 пар нуклеотидов регуляторной области достаточны для управления экспрессией оперативно связанного гетерологичного гена в малигнизированных клетках, которые также экспрессируют эндогенный ген Н19. Кроме того, другая промоторная область гена H19 между нуклеотидами -819 до +14 (фигура 5, SEQ ID NO:2) также достаточна для управления экспрессией оперативно связанного гетерологичного гена в малигнизированных клетках.
Ниже расположенная энхансерная область гена H19 человека может быть необязательно встроена в конструкцию промотор гена Н19/гетерологичный ген для того, чтобы обеспечить повышение уровня опухоль-специфической клеточной экспрессии. Согласно более подробному описанию, приведенному ниже, и иллюстрации на примере в части 6 ниже расположенная энхансерная область входит в состав SacI-рестрикционного фрагмента, располагающегося от +6 т.п.н. до +11 т.п.н. относительно сайта начала транскрипции. Исходя из свойств энхансерной последовательности, ниже расположенный энхансер способен оказывать влияние в случае расположения как в обратном, так и в прямом направлении (относительно ориентации энхансера H19 в эндогенном гене H19) ниже кодирующей области гетерологичного гена под контролем промотора гена Н19. Кроме того, фрагменты данного энхансера, содержащие последовательности, изображенные на фигурах 6, 7А, 7В и 8А-8С (SEQ ID NOS:3-5), также могут быть использованы для облегчения экспрессии гена. Экспрессия гена IGF-1 взаимосвязана со злокачественными опухолями легких и молочной железы. Промотор гена IGF-1 является нуклеотидной последовательностью, расположенной между и 1630 нуклеотидами в последовательности гена IGF-1 человека (номер доступа в Genbank M12659 М77496, включенный здесь в качестве ссылки; Rotwein et al., 1986, J.Biol. Chem. 261:4828-4832).
Продукт гена IGF-2 экспрессируется с помощью одной из четырех различных промоторных областей. Три из четырех данных промоторов подвержены импринтингу и экспрессируются в эмбриональных тканях; однако промотор Р1 активируется только в тканях взрослого организма (Sussenbach et al., 1992, Growth Reg. 2: 1-9). Промотор Р3 является вовлеченным в развитие карциномы печени. Также установлено, что подверженные импритингу промотор Р4 (последовательность от -546 до +102 нуклеотидов гена IGF-2) и промотор Р3 (последовательность от -1229 до +140 нуклеотидов гена IGF-2) активируются у человека в клетках злокачественной опухоли мочевого пузыря и могут быть использованы для управления экспрессией оперативно связанного гетерологичного гена опухолевых клеток. Промоторы Р3 и Р4 гена IGF-2 могут применяться в комбинации с энхансером гена H19 или его активными фрагментами.
Данные регуляторные последовательности подверженных и неподверженных геномному импринтингу генов, которые экспрессируются в малигнизированных клетках, могут далее для определения быть изображены в виде минимальных регуляторных последовательностей, необходимых для достижения желаемой опухоль-специфической экспрессии. Например, промоторная область может быть изменена с помощью добавлений, замен и делеций и проанализирована на предмет снижения функции опухоль-специфической экспрессии. Различные части ниже расположенного энхансера гена H19 могут быть протестированы индивидуально на предмет способности усиливать транскрипцию с промотора гена H19.
Изменения в регуляторных последовательностях могут быть произведены с помощью химических и ферментативных способов, хорошо известных специалистам в данной области. Например, могут быть удалены области последовательностей, ограниченные сайгами рестрикции. Олигонуклеотидный направленный мутагенез может быть применен для изменения последовательности определенным образом и/или для введения сайтов рестрикции в определенные области последовательности. Кроме того, могут быть получены мутанты с делецией с помощью ДНК-нуклеаз, таких как Ваl31 или ЕхоIII и S1-нуклеаза. Более обширные делеции в регуляторных последовательностях создают инкубированием ДНК с нуклеазами в течение более продолжительных периодов времени (смотри Ausubel et al., 1989 Current protocols for Molecular biology, для обзора методик мутагенеза).
Измененные последовательности оценивают на предмет их способности управлять опухоль-специфической экспрессией гетерологичных кодирующих последовательностей в подходящих клетках-хозяевах, в частности в экспрессирующих ген H19 клетках карциномы (например, в клетках карциномы мочевого пузыря, приводимые). В объеме настоящего изобретения любые измененные регуляторные последовательности, которые сохраняют свою способность управлять опухоль-специфической экспрессией, встраиваются в рекомбинантные экспрессирующие векторы для дальнейшего применения.
Большое разнообразие гетерологичных генов способно экспрессироваться под контролем данных регуляторных последовательностей, таких как гены, кодирующие токсические генные продукты, потенциально токсические генные продукты и антипролиферативные или цитостатические генные продукты. Также способны экспрессироваться маркерные гены, включающие гены ферментов (например CAT, бета-галактозидазы, люциферазы), флюоресцентных белков, таких как зеленый флюоресцентный белок, или антигенные маркеры.
Цитотоксические генные продукты широко определяют токсины и индуцирующие апоптоз агенты. Кроме того, в контексте настоящего изобретения цитотоксические генные продукты включают ферменты, метаболизирующие лекарства, которые превращают пролекарство в цитотокоический продукт. Примеры цитотоксических генных продуктов, которые могут использоваться в методах данного изобретения, включают в себя дифтерийный токсин, токсин Pseudomonas, рицин, холерный токсин, РЕ40 и гены опухолевых супрессоров, такие как ген ретинобластомы и р53. Кроме того, могут применяться последовательности, кодирующие апоптотические белки, которые индуцируют клеточный апоптоз. Подобные апоптотические белки включают бета-пептид А Альцгеймера (смотри LaFerla et al., 1995, Nat. Genet. 9:21-30), предсердный натрийуретический фактор (смотри Wu et al., 1997, J. Biol. Chem. 272:14860-14866), пептид, связанный с геном кальцитонина (смотри Sacuta et al., 1996, J. Neuroiminunol. 67:103-109), так же, как и другие апоптотические белки, уже известные или которые будут открыты в будущем.
Ферменты, метаболизирующие лекарства, которые превращают пролекарство в цитотоксический продукт, включают в себя тимидинкиназу (вирусов простого герпеса или ветряной оспы), цитозиндезаминазу, нитроредуктазу, цитохром Р-450 2В1, тимидинфосфорилазу, пуриннуклеозидфосфорилазу, щелочную фосфатазу, карбоксипептидазы А и G2, линамаразу, лактамазу и ксантиноксидазу (смотри Rigg Sikora, August 1997, Mol. Med. Today, pp. 359-366 для ознакомления).
Кроме того, антисмысловые, антигенные или аптамерные олигонуклеотиды могут доставляться в малигнизированные клетки с помощью описанных здесь экспрессирующих конструкций. Рибозимы или одноцепочечные РНК также могут экспрессироваться в малигнизированных клетках для ингибирования экспрессии отдельного интересующего гена. Генами-мишенями для данных антисмысловых или рибозимных молекул должны являться гены, кодирующие продукты, которые существенны для поддержания функций клетки или для поддержания фенотипа малигнизированной клетки. Подобные гены-мишени включают гены cdk2, cdk8, cdk21, cdk25A, циклин D1, циклин Е, циклин А и cdk4, не ограничиваясь данным.
Например, экспрессирующие векторы под контролем регуляторных последовательностей подверженных импринтингу генов или промотора IGF-1, которые экспрессируются в малигнизированных клетках, антисмысловые РНК или рибозимы, специфичные по отношению к транскриптам онкогенных образований Р53, c-fos, c-jun, kr-ras и/или Her2/neu, вводятся в клетки для того, чтобы обеспечить даун-регуляцию экспрессии эндогенных генов. Опухолевые клетки, которые экспрессируют ген H19, и способны активировать регуляторные последовательности гена H19 (или которые специфически активируют промотор IGF-1, промоторы Р3 и Р4 IGF-2), могут являться специфической мишенью для экспрессии антисмысловой РНК или рибозимной РНК.
Антисмысловые подходы включают в себя создание олигонуклеотидов (в данном случае мРНК), которые комплементарны мРНК-мишени. Антисмысловые олигонуклеотиды будут связываться с комплементарными транскриптами мРНК-мишени и препятствовать трансляции. Абсолютная комплемент арность, хотя и предпочтительна, но не требуется. В данном случае последовательность, "комплементарная" части РНК, означает последовательность, обладающую достаточной комплементарностью для гибридизации с РНК, образуя стабильный дуплекс. Способность гибридизоваться будет зависеть как от степени комплементарности, так и от длины антисмысловой нуклеиновой кислоты. Как правило, чем длиннее гибридизующаяся нуклеиновая кислота, тем больше несовпадений нуклеотидов с РНК, она может содержать и все же формировать стабильный дуплекс (или триплекс в некоторых случаях). Специалисты в данной области могут установить допустимую степень несовпадения с помощью стандартных способов определения точки плавления гибридного комплекса.
Олигонуклеотиды, которые комплементарны 5'-концу последовательности-мишени, например 5'-нетранслируемая последовательность вплоть до и включая инициирующий кодон AUG, должны быть наиболее эффективны в ингибировании трансляции. Однако недавно было показано, что последовательности, комплементарные 3'-нетранслируемым последовательностям мРНК, также эффективны в ингибировании трансляции (смотри Wagner, R., 1994, Nature 372:333-335). Таким образом, олигонуклеотиды, комплементарные как 5'-, так и 3'-нетранслируемым, некодирующим областям транскриптов гена-мишени, могут использоваться в антисмысловом подходе для ингибирования трансляции эндогенных генов. Олигонукдеотиды, комплементарные 5'-нетранслируемой области мРНК, будут включать комплемент стартовому кодону AUG. Антисмысловые олигонуклеотиды, комплементарные кодирующим областям мРНК, являются менее эффективными ингибиторами трансляции, но могут быть использованы согласно данному изобретению. Чтобы создаваемая конструкция была способна к гибридизации с 5'-, 3'- или кодирующей областью мРНК-мишени, антисмысловая нуклеиновая кислота должна иметь по крайней мере длину в шесть нуклеотидов, и предпочтительными являются олигонуклеотиды длиной в пределах от 6 до приблизительно 50 нуклеотидов. В определенных случаях требуются олигонуклеотиды длиной по крайней мере 10 нуклеотидов, по крайней мере 17 нуклеотидов, по крайней мере 25 нуклеотидов или по крайней мере 50 нуклеотидов.
Невзирая на выбор последовательности мишени, предпочтительно, чтобы сначала проводились исследования in vitro для количественной оценки способности антисмысловых нуклеотидов ингибировать экспрессию гена. Данные исследования должны использовать контроль, чтобы разграничить антисенсорное ингибирование гена и неспецифические биологические эффекты олигонуклеотидов. Также предпочтительно, чтобы в данных исследованиях сравнивалось содержание РНК- или белка-мишени с РНК или белком внутреннего контроля.
Молекулы рибозима, предназначенные для каталитического расщепления исходного гена мишени, также могут применяться для предотвращения трансляции мРНК-мишени (смотри, например, РСТ International Publication WO 90/11364, published October 4, 1990; Sarver et al., 1990, Science 247:1222-1225). Когда рибозим специфичен для транскрипции гена, кодирующего белок, необходимый для роста малигнизированной клетки, подобные рибозимы могут приводить к изменению фенотипа малигнизированной клетки. Хотя рибозимы, расщепляющие мРНК в сайте специфического узнавания последовательностей, могут применяться для разрушения мРНК-мишеней, предпочтительным является применение молоткообразных рибозимов. Молоткообразные рибозимы расщепляют мРНК в положениях, определяемых фланкирующими областями, что создает комплементарные пары оснований с мРНК-мишенью. Единственным требованием является наличие в мРНК-мишени следующей последовательности из двух оснований: 5'-UG-3'. Создание и получение молоткообразных рибозимов технически хорошо известно и более подробно описано в Haseloff Gerlach, 1988, Nature, 334:585-591. Рибозим предпочтительно конструируют таким образом, чтобы сайт узнавания расщепления располагался вблизи 5'-конца мРНК мишени; т.е. для повышения эффективности и минимизации и внутриклеточного накопления нефункциональных транскриптов мРНК.
Применяемые в настоящем изобретении рибозимы также включают РНК-эндорибонуклеазы (здесь и далее называемые "рибозимы Сесh-типа"), подобно рибозиму естественно встречающемуся Tetrahymena thermophila (известные как IVS, L-19 IVS РНК), который подробно описан Thomas Cech и коллегами (Zaug et al. , 1984, Science, 224:574-578; Zaug and Cech, 1986, Science, 231:470-475; Zaug et al., 1986, Nature, 324-129-433; International Patent Application No. WO 88/04300 by University Patents Inc.; Been and Cech, 1986, Cell, 47:207-216). Рибозимы Cech типа содержат активный сайт, состоящий из восьми пар оснований, который гибридизуется с последовательностью РНК-мишени, после чего происходит расщепление РНК-мишени. Изобретение рассматривает применение данных рибозимов Cech типа, в которых мишенью для активного сайта, состоящего из восьми пар оснований, являются последовательности, присутствующие в генах-мишенях.
5.2 Активация генов в опухолевых клетках
Клетки, в которых восстанавливается экспрессия подверженного импринтингу гена, будут также способны к специфической активации экспрессии конструкций, содержащих подобные регуляторные области подверженного импритингу гена, оперативно связанного с гетерологичным геном. Подобные клетки, в частности опухолевые клетки, являются подходящими мишенями для способов генной терапии данного изобретения. Специфическая экспрессия гена Н19 и промоторов Р3 и Р4 гена IGF-2 как в опухолях, так и клеточных линиях может быть определена с помощью способов анализов РНК, гибридизации in situ и репортерных генных конструкций. Кроме того, опухолевые клетки с активированной экспрессией гена IGF-1 могут быть сходным образом определены и являться мишенью для генной терапии с помощью промотора гена IGF-1, управляющего экспрессией гетерологичного гена.
Для большинства применений анализа РНК меченый зонд, который специфически гибридизуется с интересующим генным транскриптом, создают любым из многочисленных хорошо известных в данной области методов. Меченый зонд может содержать по крайней мере 15-30 нуклеотидов, комплементарых нуклеотидной последовательности гена H19, и, как правило, содержит по крайней мере от 50 до 150 нуклеотидов, комплементарных транскрипту гена H19. Обычно предпочитаемым гибридизационным зондом для экспрессии гена H19 является полинуклеотид, комплементарный 3' - концу транскрипту гена H19 от приблизительно 800 пар нуклеотидов выше сайта полиаденилирования до сайта полиаденилирования.
Специфическим осуществлением изобретения, что проиллюстрировано ниже на рабочем примере, является создание меченой антисмысловой РНК с помощью Т7 или Т3 экспрессирующей плазмиды. Зонды для гена H19 также могут быть меченными с помощью случайного прайминга присутствующих меченых нуклеотидов, например, используя Prime-It kit (Stratagene, La Jolla, CA; Catalog NO. 300392). Альтеративно меченные зонды могут быть созданы в реакции ПЦР, применяя клон кДНК кодирующей области гена H19 и праймеры, предназначенные для амплификации региона кодирующей области, или с помощью стандартной реакции ник-трансляции.
Подходящими метками для полинуклеотидных зондов являются нуклеотиды, включающие радиоактивные изотопы (такие как 35S и 32P), флуоресцентные, люминисцентные, цветные метки и ферментативные структуры.
Меченый зонд гибридизуют in situ в клетке или образце ткани с помощью стандартных технологий, подобных описанным далее в рабочем примере, и в совместно рассматриваемой патентной заявке США серийный номер 08/704,786, включенной сюда в качестве ссылки. Альтернативно, если может быть получено достаточное количество подходящих клеток, можно провести стандартные анализы РНК (такие как Нозерн-анализ, рибонуклеазная защита или удлинение праймера) для определения уровня экспрессии интересующего гена.
Кроме того, возможно провести подобный анализ экспрессии гена in situ, например непосредственно в срезах ткани (фиксированных и/или замороженных) из тканей плаценты, полученной биопсией или резекцией, таким образом не нуждаясь в проведении выделения нуклеиновой кислоты. Реагенты нуклеиновой кислоты, аналогичные описанным выше, смогут использоваться в качестве зондов и/или праймеров для подобных процедур in situ (смотри, например, Nuovo, G.J. , 1992, PCR In Situ Hybridization: Protocols And Applications, Raven Press, NY).
Альтернативным способом определения в случае, если опухоль или клетка будут способны специфически активировать экспрессирующие конструкции, содержащие частичные регуляторные области, оперативно связанные с гетерологичным геном, действительно является трансфекция подобных экспрессирующих конструкций в клетку. Для данных целей гетерологичный ген предпочтительно является продуктом маркерного гена. Положительным результатом при оценке продукта маркерного гена является то, что клетка или клеточная линия оказывается способной активировать экспрессию регуляторных областей.
Используя данные технологии, образцами типов опухолей, с активацией экспрессии гена H19 являются следующие типы:
А. Солидные опухоли у детей
1. Нефробластома
2. Гепатобластома
3. Эмбриональная рабдомиосаркома
В. Зародышевоклеточные опухоли и трофобластные опухоли
1. Тестикулярная зародышевая опухоль
2. Незрелая тератома яичника
3. Сакрококцигеальная опухоль
4. Хориокарцинома
5. Плацентарно-расположенная трофобластная опухоль.
С. Эпителиальные опухоли у взрослых
1. Карцинома мочевого пузыря
2. Гепатоцеллюлярная карцинома
3. Карцинома яичника
4. Карцинома шейки матки
5. Карцинома легких
6. Карцинома молочной железы
7. Плоскоклеточная карцинома головы и шеи
8. Карцинома пищевода
9. Карцинома щитовидной железы
D. Нейрогенные опухоли
1. Астроцитома
2. Ганглиобластома
3. Нейробластома
Соответственно вышеуказанные злокачественные опухоли считаются поддающимися лечению способами по настоящему изобретению. В самом деле, любые опухоли, способные активировать экспрессию гена H19, могут подвергаться лечению способами по настоящему изобретению.
Кроме того, вышеупомянутые технологии смогут применяться для определения опухолей, которые активируют промоторы гена IGF-1, Р3 и Р4 гена IGF-2. Подобные опухоли также могут подвергаться лечению способами данного изобретения. Например, ген IGF-2 активируется в опухолях детей, таких как нефробластома, рабдомиосаркомы, нейробластомы и гепатобластомы.
5.3. Способы введения полинуклеотидов под контролем регуляторных последовательностей в клетки-хозяева
Изобретение также имеет отношение к клетке-хозяину, трансфицированной полинуклеотидом, содержащим регуляторные области, оперативно связанным с гетерологичным геном. Подобные клетки-хозяева могут поддерживаться в культуре или могут быть частью животного, предпочтительно млекопитающего. Необходимые полинуклеотиды вставляют в любой из широкого ряда векторов, которые впоследствии доставляются с помощью раскрытых здесь методов и материалов. Подобные векторы могут быть получены с помощью хорошо отработанных технологий в молекулярной биологии (смотри Sambrook et al., (1989) Molecular Cloning Vols. I-III, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, Current Protocols in Molecular Biology (1989) John Wiley@Sons, all Vols. и периодику, до настоящего момента включенную сюда в качестве в ссылки). Обычно, если необходимо проведение трансляции интересующий гетерологичный ген также создают с включением подходящей 3'-полиаденилированной последовательности.
5.3.1. Культивируемые клетки.
Клетками-хозяевами, которые трансфицируют с помощью полинуклеотидов, содержащих регуляторные области подверженного импритингу гена, оперативно связанного с гетерологичным геном, могут служить любая прокариотическая или эукариотическая клетка. Встраивание полинуклеотида в генную конструкцию, такую как вектор, и трансформация или трансфекция в клетки-хозяева как эукариотические (дрожжей, птиц, насекомых или млекопитающих), так и прокариотические (бактериальные клетки), являются стандартными приемами, широко используемыми в микробиологических или тканевокультуральных технологиях.
Векторы, подходящие для культивирования подлежащего полинуклеотида в бактериальных клетках, такие как E.coli, включают плазмиды типа: pBR322 плазмиды, pEMBL плазмиды, рЕХ плазмиды, рВТас плазмиды и pUC плазмиды. Для репликации у дрожжей, YEP24 плазмиды, YIP5 плазмиды, YEP51 и YRP17 плазмиды клонируют и экспрессирующие частицы, способные транспортироваться, используют для введения генетических конструкций в S.cervisiae (Broach et al., 1993, in Experimental Manipulation of Gene Expression, ed. M.Inouye, Academic press, p.83). Данные векторы могут реплицироваться как в E.coli благодаря присутствию точки начала репликации плазмиды pBR322, так и в дрожжах, благодаря наличию детерминант репликации 2 m кольцевой плазмиды дрожжей. Кроме того, применяют маркеры устойчивости к лекарственным веществам, таким как ампицилин.
Также предпочтительны векторы млекопитающих, включающие полинуклеотиды данного изобретения, содержащие обе прокариотические последовательности для облегчения проникновения вектора в бактериальную клетку. Подобные векторы после трансфекции в клетки млекопитающих могут быть предназначены для объединения с хромосомой млекопитающих с продолжительным периодом стабильности, используя связанный выбранный маркерный ген. Альтернативно производные вирусов, таких как бычий папиломавирус (BPV-1) или вирус Эпштейн-Барра, могут быть использованы для временной экспрессии. Различные способы, применяемые для подготовки к плазмидной трансформации организмов-хозяев, технически хорошо известны. Другие подходящие системы векторов так же, как и основные рекомбинантные приемы, смотри Sambrook et al., supra.
5.3.2 Генная терапия
Изобретение также охватывает применение полинуклеотидов, содержащих генные регуляторные области, оперативно связанные с гетерологичным геном, для использования в генной терапии лечения злокачественных опухолей и гиперпролиферативных заболеваний. С целями генной терапии экспрессирующие конструкции данного изобретения могут применяться с любым биологически эффективным носителем, например любой системой или структурой, способной эффективно доставлять рекомбинантный ген в клетки in vivo. Подходы заключаются во вставке подлежащего гена в вирусные векторы, включающие рекомбинантные ретровирус, аденовирус, аденоассоциированый вирус, вирус простого герпеса-1, рекомбинантные бактериальные или эукариотические плазмиды. Вирусные векторы непосредственно трансфицируются в клетки; плазмидная ДНК может доставляться с помощью, например, катионных полимеров, катионных липосом (например, липофектин, производных холестерола, таких как D.D.A.B. и катионных фосфолипидов) или производных (например, связанных с антителом), соединений полилизина, грамицидина S, искуственных вирусных оболочек или других подобных внутриклеточных носителей, так же, как с помощью непосредственного впрыскивания генной конструкции без носителя, электропорации или преципитации фосфатом кальция, проводимой in vivo. Недавним обзором переноса гена и экспрессирующих систем для генной терапии злокачественных опухолей является Cooper, 1996, Seminars in Oncology 23:172-187.
Данный факт будет оценен потому, что трансдукция в подходящие клетки-мишени представляет собой первый важный шаг в генной терапии, выбор системы доставки частичного гена будет зависеть от таких факторов, как фенотип намеченной мишени и пути управления, например, локально или системно. Более того, будет признано, что частичная генная конструкция, обеспечивающая трансдукцию экспрессирующих конструкций in vivo, также применима для трансдукции клеток in vitro так же, как и для использования в тканях, культуральных системах ех vivo, описанных выше.
Предпочтительным подходом для введения нуклеиновой кислоты
в клетку in vivo является применение вирусного вектора, содержащего нуклеиновую кислоту, например частичного цитотоксического гена под контролем регуляторных последовательностей гена H19. Заражение клеток вирусным вектором имеет преимущество в том, что большая часть клеток-мишеней может получить нуклеиновую кислоту. Кроме того, молекулы, кодирующие вирусный вектор, например содержащаяся в вирусном векторе кДНК, эффективно экспрессируются в клетках, включивших нуклеиновую кислоту вирусного вектора. Подходящие векторы, которые способны доставляться с помощью настоящих открытых способов и компонентов, включают векторы вируса простого герпеса, аденовирусные векторы, аденоассоциированные векторы, ретровирусные векторы, вирус pseudorbies, векторы вируса альфа-герпеса и подобные вирусы, но не ограничиваются данным. Полный обзор вирусных векторов, в частности вирусных векторов, подходящих для модификации нереплицирующих клеток, и способы применения подобных векторов в связи с экспрессией интересующих полинуклеотидов можно найти в книге "Viral Vectors: Gene Therapy and Neuroscience Applications" Ed. Caplitt and Loewy, Academic Press, San Diego (1995).
Показано, что возможно ограничить инфекционный спектр вирусов и, следовательно, векторов на основе данных вирусов с помощью модификации упаковывающих белков на поверхности вирусной частицы (смотри, например, РСТ публикации WO 93/25234 и WO 94/06920). Например, стратегии для модификации инфекционного спектра ретровирусных векторов включают: присоединение антител, специфических для антигенов клеточной поверхности, к вирусному env белку (Roux et al. , 1989, Proc. Nat. Acad. Sci. USA 86:9079-9083; Julan et al., 1992, J. Gen. Virol. u3:3251-3255: Goud et al., 1983, Virology 163:251-254); или присоединение лигандов рецептора клеточной поверхности к вирусным env белкам (Neda et al., 1991. J. Biol. Chem 266:14143-14146). Присоединение может проводиться в виде химического связывания с белком или другими вариациями (например, с лактозой для превращения env белка в оксигликопротеин) так же, как и с помощью произведения слияния белков (например, одноцепочечное антитело/env объединенные белки). Например, малигнизированные клетки могут быть мишенью для данной технологии, например, в случае присоединения антител против опухольассоциированных молекул или поверхностных белков малигнизированной клетки к поверхности рекомбинантного вируса. Хотя данный способ пригоден для ограничения или в противном случае для направления инфицирования основных типов ткани, также может использоваться для превращения эктотропного вектора в амфотропный вектор.
Предпочтительная система доставки вирусного гена, применяемая в настоящем изобретении, использует аденовирусные векторы. Геномом аденовируса можно манипулировать подобным образом, поскольку он кодирует и экспрессирует интересующий генный продукт, но инактивирован в отношении своей способности реплицироваться при нормальном литическом жизненном цикле вируса. Смотри, например, Bercner et al. , 1988, Bio-Techniques 6:616; Rosenfeld et al., 1991, Science 252:431-434; Rosenfeld et al., 1992, Cell 68:143-155. Подходящие аденовирусные векторы, происходящие из аденовирусного штамма AD типа 5 dl324 или других штаммов аденовирусов (например, Ad2, Ad3, Ad7), являются хорошо известными специалистам в данной области. Рекомбинантные аденовирусы могут быть выгодны в определенных обстоятельствах, поскольку они могут применяться для заражения широкого ряда типов клеток, включая эпителий дыхательных путей (Rosenfeld et al., 1992, cited supra), эндотелиальные клетки (Lemarchand et al., 1992, Proc. Nat. Acad. Sci. USA 89:6482-6486), гепатоциты (Herz and Gerard, 1993, Proc. Nat. Acad. Sci. USA 90:2812-2816) и мышечные клетки (Quantin et al., 1992, Proc. Nat. Acad. Sci. USA 89:2581-2584). Более того, вирусная частица является относительно стабильной, поддающейся очищению и концентрированию, и может подвергаться модифицированию для воздействия на спектр заражения. Кроме того, введенная аденовирусная ДНК (и содержащаяся там чужеродная ДНК) не интегрируются с геномом клетки-хозяина, а остаются в виде эписомы, посредством чего устраняя потенциальные проблемы, которые могут наблюдаться в результате вставочного мутагенеза в ситуациях, когда введенная ДНК интегрируется с геномом хозяина (например, ретровирусная ДНК). Более того, емкость аденовирусного генома для внесения чужеродной ДНК оказывается большой (до 8 тысяч пар нуклеотидов) относительно других векторов, доставляющих ген (Bercner et al., cited supra, Haj-Ahmand and Graham, 1986, J. Virol. 57:267). Большинство аденовирусных векторов с недостаточной репликацией применяются в настоящее время, и поэтому предпочитаемые настоящим изобретением векторы лишены всех или части вирусных Е1 и Е3 генов, но сохраняют более 80% аденовирусного генетического материала (смотри, например, Jones et al., 1979, Се1116:683; Bercner et al., supra; and Graham et al. in Methods in Molecular Biology, E.J. Murray, Ed(Humana, Clifton NJ, 1991) vol.7, pp.109-127).
Другой вирусной векторной системой, подходящей для доставки одной из подлежащих экспрессирующих конструкций, является аденоассоциированный вирус (AAV). Аденоассоциированный вирус является естественно встречающимся несамостоятельным вирусом, который требует присутствия другого вируса, такого как аденовирус или вирус герпеса, в качестве помощника для эффективной репликации и продуктивного жизненного цикла (смотри обзор Muzuczka et al., 1992, Curr. Topics in Micro, and Immunol. 158:97-129). Также он является одним из немногих вирусов, способных встраивать свою ДНК в неделящиеся клетки, и проявляет высокую частоту стабильного взаимодействия (смотри, например, Flotte et al. 1992, Аm. J. Respir. Cell. Mol. Biol. 7:349-354; Samulski et al. , 1989, J. Virol. 63:3822-3828; McLaughlin et al., 1989, J. Virol. 63: 1963-1973). Векторы, содержащие по крайней мере 300 пар оснований из AAV, способны упаковываться и способны встраиваться. Место для эндогенной ДНК является ограниченным до приблизительно 4,5 т.п.н. AAV вектор, подобный описанному Tratschin et al., 1985, Mol. Cell. Biol. 5:3251-3260, может применяться для введения ДНК в клетки. Целый ряд нуклеиновых кислот вводят в различные типы клеток, используя AAV векторы (смотри, например, Hermonat et al. , 1984, Proc. Nat. Acad. Sci. USA 81:6466-6470; Tratschin et al., 1985, Mol. Cell. Biol. 4:2072-2081; Wondisford et al., 1988, Mol. Endocrinol. 2: 32-39; Tratschin et al. , 1984, J. Virol. 51:611-619; and Flotte et al., 1993, J. Biol. Chem. 268:3781-3790).
В дополнение к способам вирусного переноса, подобным проиллюстрированным выше, могут также применяться невирусные методы для обеспечения управляемой экспрессии желаемого гетерологичного гена в ткани животного. Большинство невирусных методов переноса гена основываются на обычных механизмах, используемых клетками млекопитающих для захвата и внутриклеточного транспорта макромолекул. Предпочтительно невирусные системы доставки гена настоящего изобретения основываются на внутриклеточных путях захвата подлежащих экспрессирующих конструкций клеткой-мишенью. Образцы систем доставки гена данного типа включают липосомальные системы доставки, полилизиновые коньюгаты и искуственные вирусные оболочки.
В клиническом отношении системы доставки гена для терапевтической экспрессирующей конструкции могут быть введены в пациента с помощью любого из ряда методов, каждый из которых хорошо известен в данный области. Например, фармацевтический препарат, включающий системы доставки гена, может вводиться систематически, например, с помощью венозных инъекций и преобладание специфической экспрессии конструкции в клетках-мишенях наблюдается при специфичном переносе, обеспечиваемой экспрессией в клетке или ткани, благодаря регуляторным последовательностям, контролирующим экспрессию гетерологичного гена, или регуляторным последовательностям в сочетании со структурой доставки гена, направленной на определенные типы клеток. С другой стороны, первоначальная доставка рекомбинантной экспрессирующей конструкции является более ограниченной существованием локального места введения у животных. Например, структура доставки гена может быть введена с помощью катетера (смотри US Patent 5328470) или стереостатической инъекцией (например, Chen et al., 1994, Proc. Nat. Acad. Sci. USA 91:3054-3057). Экспрессирующая конструкция данного изобретения может быть доставлена в конструкцию для генной терапии с помощью электропарации, применяя технологии, описанные, например, Dev et al. , 1994, Cancer Treat. Rev. 20:105-115.
Фармацевтический препарат, содержащий конструкцию для генной терапии, может в основном состоять из системы доставки гена в подходящем растворителе или может включать в себя медленно высвобождающийся матрикс, в котором находится структура доставки гена. Альтернативно в случае, когда вся система доставки гена может продуцироваться целиком рекомбинантными клетками, например ретровирусные векторы, фармацевтический препарат может содержать одну или более клетку, продуцирующую систему доставки гена.
5.3.3. Терапевтические дозы
Со стороны практикующих врачей или перспектив для пациентов будет оценено, что фактически любое облегчение или предотвращение нежелательного симптома, связанного с развитием рака (например, боли, чувствительности, потери веса и подобного), может быть желаемым. Кроме того, является желаемым любое уменьшение массы опухоли или скорости роста так же, как и доказательство гистопатологической картины опухоли. Таким образом, для целей данной заявки термины "лечение", "терапевтическое применение" или "медицинское применение", используемые здесь, могут относится к любому и ко всем используемым компонентам формулы изобретения, которые корректируют состояние болезни, или симптомы, или, с другой стороны, предотвращают, препятствуют, задерживают или обращают развитие болезни или других нежелательных симптомов по любому пути.
Эффективная доза и протокол лечения могут быть определены с помощью обычных способов, начиная с низкой дозы на лабораторных животных, и затем увеличивая дозу, при этом наблюдая за эффектом и также систематически варьируя режим дозирования. Исследования на животных, предпочтительно исследование на млекопитающих, обычно используют для определения максимальной переносимой дозы или MTD, биоактивного вещества на килограмм веса. Специалисты, имеющие навыки в данной области, регулярно экстраполируют дозы для эффективности и избегания токсичности на другие виды, включая человека.
Перед исследованиями эффективности на человеке проводится первая фаза клинических испытаний на здоровых добровольцах, что помогает установить безопасные дозы. Клиницистом берется во внимание множество факторов при определении оптимальной дозы, даваемой добровольцу. Первостепенным среди данных факторов является токсичность и время полужизни выбранного продукта гетерологичного гена. Дополнительные факторы включают массу пациента, возраст пациента, основное состояние пациента и часть ракового заболевания, способного подвергаться лечению, тяжесть заболевания, прием других препаратов пациентом, активность генного продукта in vivo и другие подобные факторы. Дозы при испытании выбирают после изучения результатов испытаний на животных и клинической литературы.
Например, обычная доза для человека аденовирусного вектора, содержащего регуляторную область гена Н19, оперативно связанную с гетерологичным геном, кодирующим цитотоксическое вещество тимидинкиназу, составляет от 1•107 БОЕ до 1•1010 БОЕ, непосредственно вводимую в опухолевую массу в день. Более предпочтительно, чтобы суточная доза подобного аденовирусного вектора, непосредственно вводимого в опухоль, составляла от 1•108 БОЕ до 1•1010 БОЕ в зависимости от размера опухоли. Для аденовирусного вектора, содержащего регуляторную область гена H19, оперативно связанную с цитотоксическим генным продуктом с другим уровнем токсичности, данные значения должны, конечно, быть соответственно изменены. Сходные дозы аденовирусного вектора, содержащего промотор Р4 гена IGF-2, оперативно связанного с гетерологичным геном, кодирующим цитотоксическое вещество, подобное тимидинкиназе, также могут применяться в качестве предположительной начальной точки.
В частности, когда рассматривается применение in vivo, различные биохимические компоненты настоящего изобретения предпочтительно должны быть высоко очищенные и достаточно освобождены от потенциально вредных загрязнений (например, по крайней мере, National Food (NF) качества, обычно, по крайней мере, аналитического качества и предпочтительно фармацевтического качества). Учитывая, что данный компонент должен быть синтезирован перед использованием, подобные синтезы или последовательная очистка будут выражаться в получении продукта, который достаточно освобожден от любых потенциально токсичных веществ, которые могли использоваться в течение синтеза или процедур очистки.
Для применения в лечении раковых состояний на добровольцах настоящее изобретение также обеспечивает в одном из своих аспектов кит или упаковку в виде стерильной заполненной пробирки или ампулы, которая содержит полинуклеотидный вектор, включающий регуляторную область гена H19, оперативно связанного с гетерологичным геном, кодирующим цитотоксическое вещество, или клетку с высвобождающимся вектором. В одном случае кит содержит полинуклеотидный вектор, содержащий регуляторную область гена H19, оперативно связанного с гетерологичным геном, кодирующим цитотоксическое вещество, в качестве готового соединения как в одноразовой дозе, так и в мультидозовом количестве, где упаковка содержит инструкцию использования компонентов для лечения рака. Альтернативно и согласно другой реализации изобретения упаковка обеспечивает стерильную заполненную пробирку или ампулу, содержащую подобную клетку с высвобождающимся вектором или клеточную линию. Для транспортировки и хранения клетка с высвобождением вектора или клеточная линия должны быть замороженные. Факультативно упаковка также может содержать среду и реагенты для культивирования клетки с высвобождением вектора или клеточную линию.
Следующие примеры являются иллюстрацией к описанному изобретению.
6. Пример: регуляторные последовательности гена Н19, способствующие экспрессии гетерологичного гена в линиях опухолевых клеток.
В данной части описывается устройство различных экспрессирующих конструкций, содержащих CAT, продуцирующий ген, находящихся под контролем регуляторных последовательностей гена H19, и их переносу - некоторые различные клеточные линии рака мочевого пузыря.
6.1. Материалы и методы
6.1.1. Клеточные линии и трансфекции
Клеточные линии рака мочевого пузыря НТ-1376, EJ28, Т24Р, 1197 UM-UC-3 были получены из American Type Culture Collection (ATCC) и поддерживались согласно рекомендациям АТСС.
Кратковременные трансфекции проводили с помощью метода трансфекции преципитацией фосфатом кальция. Преципитаты (содержащие 7 г плазмид) вносили в 1 мл среды до 0,3•106 клеток в 30 мм чашки. После 14 часов трансфекционную среду удаляли и добавляли свежую среду, клетки собирали спустя 24-96 часов после трансфекции и определяли активность CAT с помощью бутирил-СоА органической фазовой экстракции (Sambrook et al., 1989). Аликвота из верхней органической фазы (100 мкл) помещали в сцинтилятор, хорошо вмещающий 3 мл сцинтиляционной жидкости, и измеряли.
6.1.2. Создание экспрессирующих векторов
Плазмиды pCAT-Basic (содержащая CAT передающий ген, предшествующий множественному сайту клонирования), рСАТ промоторная (содержащая CAT передающий ген под контролем SV-40 промотора), рСАТ-энхансерная (содержащая SV-40 энхансер, расположенный ниже CAT передающего гена, и множественный сайт клонирования для вставки промотора, расположенный выше CAT передающего гена) были получены коммерчески от Promega (Madison/ WT).
Для создания плазмиды рН19 Е, содержащей CAT передающий ген под контролем промотора гена H19, область промотора гена H19 (SEQ ID NO:1) сначала клонировали pBluescript II SK + (Promega). Полинуклеотид, содержащий последовательность промотора гена H19, амплифицировали из ДНК плаценты человека с помощью праймеров
ESPCR21: CGGTTCCCCACTTCCCCAGTTT (SEQ ID NO:6) и ESPCR22: CGGAAGTCGACAACCCTCACCAAAGGCCAAGGT (SEQ ID NO:7).
ПЦР продукт обработали ферментом Кленова и клонировали в EcoRV сайт pBluescript II SK +. Вставленную ДНК определяли с помощью обработки внутренне разрезающими ферментами PvuII, EcoRI и Apal. Направление промоторов было противоположным направлению LacZ кодирующей области вектора. Область промотора затем удаляли с помощью вырезания ферментами HindIII и PstI, и полученный фрагмент длиной приблизительно 0,9 т.п.н. вставляли в HindIII-PstI сайты pCAT-Basic плазмиды для получения плазмиды рН19Е.
Экспрессирующие плазмиды, содержащие область энхансера гена H19, вставленную в обоих направлениях ниже расположенного промотора гена Н19/САТ передающего гена, создавали следующим образом. Фрагмент Sас I длиной 5 т.п. н. , содержащий ниже расположенный энхансер гена H19 (от +6,0 т.п.н. до +11 т. п. н. относительно начала транскрипции гена H19), клонировали в SасI сайт плазмиды pUC19. Данный энхансерный фрагмент затем вырезали с помощью рестриктаз EcoRI и HindIII, и лигировали в EcoRI-HindIII сайты плазмиды pBluescript для создания плазмиды pBhH19En-Sa. Плазмиду pBhH19En-Sa частично обрабатывали рестриктазой BamHI, и фрагмент длиной 5 т.п.н., содержащий энхансер гена H19 (и внутренний BamHI сайт), клонировали по BamHI сайту, расположенному ниже промотора гена Н19/САТ передающего гена в плазмиде рН19Е. Таким образом, получали плазмиды, содержащие энхансер гена H19 как в прямом (плазмида pH19EH19D), так и в обратном (плазмида pHl9EH19R) направлениях.
6.2 Результаты и обсуждение
Каждую из пяти различных клеточных линий злокачественной опухоли мочевого пузыря НТ-1376, EJ28, Т24Р, 1197 и UM-UC-3 трансфицировали pCAT-basic (обозначенной как Р-Е на фигуре 2), pCAT-control (обозначенной как pSV40ESV40 на фигуре 2), рН19Е, pH19EH19D и pHl9EH19R. Результаты экспрессии каждого конструкта представлены на фигуре 3А-3Е. В каждой клеточной линии наивысший уровень САТ-активности наблюдался с плазмидой pCAT-control, содержащей как энхансер SV40, так и промотор SV40. Данный конструкт служил в качестве положительного контроля, поскольку было установлено, что регуляторные последовательности SV40 являются индукторами генной экспрессии. Однако регуляторные последовательности SV40 не являются специфическими для опухолевых клеток в своей способности индуцировать генную экспрессию. Клеточные линии, трансфицированные рН19Е, содержащие репортерный ген CAT под контролем промотора H19, также проявляли значительно повышенные уровни экспрессии CAT по сравнению с фоновыми значениями. Уровень индукции САТ-активности промотором H19 изменялся в интервале от пятикратного в клеточной линии НТ-1376 до десятикратного в клеточной линии UM-UC-3. Добавление энхансера H19 в конструкты промотор Н19/репортерный ген CAT еще более увеличивало уровни экспрессии в определенных клеточных линиях. Например, в клеточных линиях EJ28, Т24Р и 1197 энхансер H19 значительно повышал уровень экспрессии промотора Н19/репортерного гена CAT. Однако ориентация энхансера давала различные результаты в различных клеточных линиях. В клеточных линиях НТ-1376 и UM-UC-3 энхансер оказывал небольшое влияние на экспрессию или не оказывал его вовсе.
Результаты показывают, что участок промотора человеческого H19 управляет экспрессией оперативно присоединенного гетерологичного репортерного гена в широком круге клеточных линий злокачественных опухолей мочевого пузыря. В некоторых клеточных линиях злокачественных опухолей мочевого пузыря энхансер H19 способен еще более повысить уровень экспрессии репортерного гена под контролем H19.
Пример 7: Ген токсина под контролем регуляторных последовательностей H19
7.1 Материалы и методы
Экспрессирующие конструкты, описанные выше в разделе 6, модифицируют таким образом, чтобы они экспрессировали последовательность, кодирующую токсичный продукт или пролекарство вместо CAT. Например, последовательность, кодирующую продукт гена CAT, удаляют и заменяют последовательностью, кодирующей тимидинкиназу вируса простого герпеса (HSV-TK), применяя стандартные методы клонирования, которые хорошо известны в данной области.
Экспрессирующими Н19/пролекарство плазмидами трансфицируют клеточные линии злокачественных опухолей мочевого пузыря, как описано в разделе 6. Будучи трансфицирована в клеточные линии злокачественных опухолей мочевого пузыря экспрессирующая H19/HSV-TK плазмида индуцирует специфичную цитотоксичность в отношении клеток злокачественной опухоли мочевого пузыря в присутствии ганцикловира.
Пример 8: Экспрессия H19 в мышиной модели химически индуцированной карциномы мочевого пузыря
8.1 Материалы и методы
Самок мышей С3Н/Не в возрасте семидесяти пяти недель (Charles River) содержали по 6 особей в одной клетке и им позволяли акклиматизироваться в кондиционируемом помещении со световым циклом 12-часового дня/12-часовой ночи. По достижении мышами возраста 8 недель начинали эксперимент и мышей произвольно подразделяли на контрольную группу (10 мышей) и экспериментальную группу (60 мышей). Экспериментальной группе мышей давали 0,05% N-бутил-N-(4-гидроксибутил)нитроз амин (ВВМ) (Tokyo Kasei Kogyo Co. Ltd., Tokyo, Japan), растворенный в питьевой воде ad libitum. Контрольным мышам давали питьевую воду. Животных из обеих групп забивали на 4, 8, 12, 16, 20 и 26 неделях от начала эксперимента. Мочевые пузыри иссекали, фиксировали и помещали в парафиновые блоки, применяя стандартные процедуры.
8.1.1 Получение зонда
Фрагмент длиной 2,1 т.п.н., содержащий кодирующую область мышиного Н19, субклонировали в плазмиду pBluescript II KS (Stratagene, La Jolla, CA) за сайтами связывания РНК-полимеразы Т7 и Т3. [35S]-меченую антисмымловую РНК Н19 получали in vitro из HindIII-линеаризованной плазмидной ДНК с применением полимеразы Т7 (Boehringer Mannheim) и набора Amersham RPN 2006. Полученные in vitro транскрипты обладали специфичной активностью 107 имп/мин/мкг. Смысловую мРНК Н19, полученную с помощью полимеразы Т3 (Boehringer Mannheim) и EcoRI-линеаризованной матрицы, применяли в качестве контроля.
8.1.2 Гибридизация in situ
Твердые парафиновые срезы (5 мкм) зафиксированных формалином тканей помещали на покрытые 3-аминопропилтриэтоксиланом (Tespa, Sigma) предметные стекла и сушили в течение ночи при 37oC. Срезы депарафинизировали ксилолом, фиксировали 4%-ным параформальдегидом и затем обрабатывали протеиназной К (Sigma). Срезы ацетилировали в целях снижения неспецифического связывания зонда и обезвоживали с помощью промывок этанолом.
[35S]-меченые РНК-зонды (специфическая активность 50000 имп/мин/мкл) гибридизовали, как описано Rangini et al. 1991, Mech. Dev. 35:13-24, опуская стадию тио-АМФ. Срезы наносили на пленку на 10 суток и обратно окрашивали гематоксилином и эозином. Срезы исследовали и фотографировали, применяя микроскоп Polyvar (Reichert Jung) в светло- и темнопольном освещении. Контроли включали гибридизацию со смысловым РНК-зондом и предгибридизационную обработку РНКазой. Кроме того, в качестве отрицательных и положительных контролей служили срезы мочевых пузырей взрослых здоровых мышей (которые не экспрессируют H19) и эмбриональных мочевых пузырей соответственно.
8.2 Результаты и обсуждение
К возрасту 26 недель у всех выживших мышей из экспериментальной группы развились пальпируемые опухоли мочевого пузыря. Интенсивная экспрессия H19 наблюдалась в химически индуцированных опухолях мочевого пузыря. Напротив, в нормальном зрелом мочевом пузыре экспрессии H19 не отмечалось. Соответственно данная мышиная модель химически индуцированной опухоли мочевого пузыря может быть применена в качестве животной модели для демонстрации опухоль-специфической цитотоксичности in vivo конструктов, содержащих регуляторные последовательности H19, оперативно присоединенные к гену токсина.
Пример 9: Генная терапия с применением регуляторных последовательностей Н19 для экспрессии гетерологичного гена в мышиной модели карциномы мочевого пузыря
Экспрессирующие Н19/токсин или пролекарство плазмиды включают в липосомы (как описано в Takashita et al., 1993, J. Clin. Invest. 93:652-651, включенной сюда в качестве ссылки) для доставки в мочевой пузырь мыши in vivo. Мыши, используемые в данном эксперименте, страдают химически индуцированными опухолями мочевого пузыря, как описано в разделе 8.
Вкратце, 5 мкг плазмидной ДНК, растворенной в 500 мкл бессывороточной среды Optimen (BRL Life Technоlogies, Gaithersburg, MD), добавляют в 250 мкл липофектамина и 250 мкл воды. Смесь инкубируют в течение 30 минут при комнатной температуре, затем разбавляют 10 мл сбалансированного солевого раствора (BSS(-): 140 мМ NaCl, 5,4 мМ KCl, 10 мМ трис-HCl, рН 7,6). После осаждения путем центрифугирования раствора при 15000 об/мин в течение 30 минут липосомы ресуспендируют в 1 мл BSS(-), содержащего 1 мМ CaCl2. Приблизительно 0,2 мл концентрированных липосом вводят мышам, страдающим химически индуцированными опухолями мочевого пузыря, посредством катетера. Контрольная группа мышей, страдающих опухолями мочевого пузыря, получает липосомы, не содержащие ДНК или содержащие конструкт, содержащий не относящийся к предмету изобретения ген под контролем регуляторных последовательностей H19. В определенные моменты времени мышей из каждой группы забивали, мочевые пузыри иссекали, фиксировали и помещали в парафиновые блоки, применяя стандартные процедуры. Чередующиеся срезы исследовали на предмет гибридизации in situ, применяя либо зонд Н19, как описано выше, или зонд кодирующей последовательности гена токсина Pseudomonas. Кроме того, между контрольной и экспериментальной группами сравнивают размер, количество и некротизирование опухолей. Было обнаружено, что экспрессия токсина Pseudomonas колокализована с экспрессией Н19 в опухолях мочевого пузыря из экспериментальной группы мышей. Кроме того, опухоли мочевого пузыря из экспериментальной группы мышей уменьшаются в размере и меньше некротизированы по сравнению с опухолями мочевого пузыря из контрольной группы мышей.
Пример 10: Экспрессия промоторов Р3 и Р4 IGF-2 в опухолевых клеточных линиях
10.1 Материалы и методы
В данном эксперименте конструировали множество экспрессирующих конструктов, содержащих люциферазный репортерный ген, находящийся под контролем одного из четырех различных промоторов IGF-2, и переносили их в несколько различных клеточных линий злокачественных опухолей мочевого пузыря. Конструировали конструкты промотора человеческого IGF-2/люциферазы (см. таблицу).
Промоторные последовательности IGF-2 описаны в Sussenbach et al., 1992, Growth Reg. 2:1-9, включенной сюда в качестве ссылки. Люциферазный репортерный вектор коммерчески доступен от Promega, Madison, WI (каталожный Е 1641).
10.2 Результаты и обсуждение
Полученными экспрессирующими плазмидами транфицировали клеточные линии злокачественных опухолей мочевого пузыря НТ-1376, EJ28, Т24Р, 1197 и UM-UC-3, как описано выше в разделе 6. Люциферазную активность анализировали с применением коммерческого набора (Promega, Madison, WI, каталожный Е1500). Результаты, показанные на фигурах 4А-4Е, свидетельствуют о том, что промотор Р4 IGF-2 направлял экспрессию люциферазного репортерного гена в каждой протестированной клеточной линии злокачественных опухолей мочевого пузыря. В клеточной линии 1197 промотор Р3 IGF-2 также направляет экспрессию люциферазного репортерного гена. В последующих экспериментах было показано, что промоторы Р3 и Р4 IGF-2 направляют экспрессию гена люциферазы в других опухолевых клеточных линиях, включая клетки хориокарциномы и рабдомиосаркомы.
Пример 11: Промотор Н19 и промотор IGF-2 функционируют с энхансером H19, обеспечивая экспрессию гетерологичного гена
11.1 Материалы и методы
Четыре люциферазных репортерных вектора, pGL3-Basic, pGL3-Promoter, pGLS-Enhancer и pGLS-Control получали из Promega. Данными векторами трансфицировали культивируемые клеточные линии с применением ряда различных реактивов для трансфекции, включая липофетамин (Gibco/BRL), fugene (Boehringer), набор для трансфекции Perfect из 8 различных липидных реактивов (Invitrogen), TFX-10, TFX-20, transfast (Promega) и кальций-фосфатного метода (Gorman et al., 1982, Mol. Cell. Biol. 2:1044-1051).
Промотор H19, клонированный в EcoRV-сайт pBluesbript II SK (pbhl9p #1), описан в разделе 6.1 выше. Промотор Н19 вырезали путем расщепления Sma I и Hind III, и полученный фрагмент длиной 0,9 т.п.н. встраивали в Sma I-Hind III-сайты вектора pGL3-Basic с получением конструкта Luc-pbhl9.
Участок промотора H19 от nt -819 до +14 амплифицировали с помощью ПЦР с плазмиды pbh19p # 1, применяя праймеры 5'-ATATGGTACCGACAACCCTCACCAAAG-3' (левее, SEQ ID NO:8) и 5'-ATATAAGCTTCTTCTCCCTCACCCTGCTC-3' (правее, SEQ ID NO: 9). Полученный продукт ПЦР расщепляли Kpnl и Hind III и лигировали в Крn I-Hind III-сайты вектора pGL3-Basic с получением конструкта Luc-PBH19. Данный полученный с помощью ПЦР промотор H19 секвенировали по обоим направлениям по методу автоматического dye terminator cycle секвенирования (ДНК-секвенатор АВТ Prism 377, Perkin Elmer). На фигуре 5 приведена нуклеотидная последовательность промотора Н19 (SEQ ID NO:2), полученная с помощью ПЦР.
Расположенный левее энхансер H19 длиной 5 т.п.н., описанный в разделе 6 выше, расщепляли DamH с получением двух фрагментов длиной 4,1 т.п.н. и 0,9 т.п.н. на 3'-конце. Конструкты Luc-PBH19-0.9EH19 и Luc-PBH19-4EH19 конструировали путем встраивания ВаmН 1-фрагментов длиной 4,1 т.п.н. и 0,9 т.п.н. энхансера H19 в BamH 1-сайт плазмиды Luc-PBH19 соответственно. Последовательности энхансера располагались правее гена промотора Н19/люциферазного репортера.
ВаmН I-фрагмент энхансера длиной 0,9 т.п.н. лигировали в BamH I-сайт вектора pGL-Basic с получением вектора Luc-0.9ЕН19. Промотор Н19 плазмиды pbhl9p #1 вырезали Kph I-BamH I и лигировали в Kpn I-Bgl 11-сайты конструкта Luc-0.9EH19 с получением экспрессирующего конструкта Luc-pbhl9-0.9ЕН19, который содержит клоны промотора, как описано в разделе 6 выше, и энхансер длиной 0,9 т.п.н. правее репортерного гена H19/LUC.
Экспрессирующие векторы, обозначенные как Нuр-1, Нuр-2, Нuр-3 и Нuр-4, содержащие ген люциферазы под контролем промоторов Р1, Р2, Р3 и Р4 человеческого IGF-2 соответственно, конструировали, как описано в Sussenbach et al. , 1992, Growth Reg. 2:1-9. Участок Р4 длиной 512 п.н. амплифицировали с помощью ПЦР с конструкта Нuр-4, применяя праймеры 5'-ACAGGTACCTCTAGAGTCGACCT-3' (левее, SEQ ID NO: 10) и 5'-ATATAAGCTTGCTCCCATCCTGCA-3' (правее, SEQ ID NO: 11). Полученный продукт ПЦР расщепляли Kpn I-Hind III и лигировали в Kpn I-Hind 111-сайты вектора pGL3-Basic репортерного гена с получением вектора репортерного гена Luc-P4.
Также получали экспрессирующие векторы, содержащие промотор Р4 и энхансер Н19 IGF-2. ВаmН I-фрагмент энхансера длиной 2 т.п.н., полученный из ранее описанного фрагмента длиной 4,1 т.п.н., клонировали в ВаmН I-сайт конструкта Luc-Р4 с получением экспрессирующего вектора Luc-P4-2EH19.
Энхансеры H19 длиной 0,9 т.п.н., 2 т.п.н. и 4,1 т.п.н. секвенировали с помощью автоматического ДНК-секвенирования. Нуклеотидная последовательность энхансера длиной 0,9 т. п.н. приведена на фигуре 6 (SEQ ID NO:3). Нуклеотидная последовательность энхансера длиной 2 т.п.н. приведена на фигурах 7А и 7В (SEQ ID NO:4). Нуклеотидная последовательность энхансера длиной 4,1 т. п.н. приведена на фигурах 8А-8С (SEQ ID NO:5).
11.2 Результаты и обсуждение
Тогда как для внедрения четырех содержащих ген люциферазы векторов в культивируемые клеточные линии применяли несколько реактивов для трансфицирования, кальций-фосфатная преципитация обеспечивала наивысшую эффективность трансфекции для большинства протестированных клеточных линий. Таким образом, кальций-фосфатную преципитацию последовательно применяли для трансфекции различными экспрессирующими векторами. Кроме того, повышенная концентрация плазмидной ДНК не ингибировала эффективность трансфекции даже при применении в концентрациях выше плато.
Каждую из клеточной линии злокачественной опухоли мочевого пузыря 5637, клеточной линии гепатоклеточной карциномы (ГКК) Huh7 и клеточной линии опухоли почки 293Т трансфицировали различными конструктами, содержащими люциферазный репортерный ген под контролем промотора H19 или промотора Р4 IGF-2 в сочетании с энхансером Н19.
Клетки, трансфицированные Luc-phl9 и LUC-PH19, содержащими репортерный ген и промотор H19, проявляли повышенный уровень экспрессии по сравнению с фоновыми значениями (фигуры 9А-9С). Конструкт Luc-PH19, содержащий полученный с помощью ПЦР промотор, проявляет более высокую активность, чем Luc-phl9, в каждой протестированной клеточной линии. Добавление фрагмента энхансера H19 длиной 0,9 т.п.н. в репортерный вектор Luc-phl9 (Luc-phl9-0.9ЕН19) далее способствовало повышению уровня экспрессии в 2-4 раза в клеточных линиях 5637 и 293Т соответственно.
Промотор Р4 IGF-2 также способствовал повышению уровня экспрессии люциферазы во всех клеточных линиях по сравнению с фоновыми значениями. Добавление фрагмента энхансера H19 длиной 2 т.п.н. в экспрессирующий вектор Luc-P4 усиливало активность промотора Р4. Уровень индукции люциферазной активности фрагментом энхансера длиной 2 т.п.н. находился в интервале от двукратного в клеточной линии 293Т до шестикратного в клеточной линии Huh7, тогда как в клетках 5673 энхансер усиливал активность промотора лишь в небольшой степени.
На фигурах 10А-10Е показана экспрессия конструкта Luc-phl9-4ЕН19, содержащего как полученный с помощью ПЦР промотор H19, так и фрагмент энхансера H19 длиной 4,1 т.п.н. Энхансер сильно повышал активность промотора в 3-28 раз в клеточных линиях за исключением клеточной линии 5637.
12. Депонирование клона
Следующая плазмида была депонирована в American Type Culture Collection (ATCC), Manassas, VA, в соответствии с Будапештским договором о международном признании банка микроорганизмов в целях патентования (the Budapest Treaty on the International Recognition of the Deposit Microorganisms for Purposes of Patent Procedure):
Клон - рН19УР19;
доступа АТСС - 209322;
Дата депонирования - 2 октября 1997 года.
Эквиваленты
Приведенное выше описание является достаточным для того, чтобы специалист в данной области смог осуществить настоящее изобретение на практике. Действительно подразумевается, что различные модификации описанных выше средств осуществления настоящего изобретения, которые очевидны для специалистов в области молекулярной биологии, медицины или связанных с ними областях, подпадают под следующие притязания.
Все публикации, процитированные здесь, включены сюда в полном объеме в качестве ссылки (список последовательностей см. в конце описания).
















Изобретение относится к области медицинской генетики. Сущность изобретения: способ экспрессии гетерологичной последовательности в опухолевой клетке путем внедрения в нее гена, подвергаемого импринтингу. Ген содержит регуляторную последовательность, например H19 с промотором Р3 или Р4 IGF-2 и др. , способную управлять экспрессией цитотоксических продуктов. Гены могут использоваться для лечения различных раковых, вирусных и аутоиммунных заболеваний. Описание изобретения содержит сведения о соответствующих векторах экспрессии и методах лечения. Технический результат - расширение арсенала средств генной терапии. 5 с. и 35 з.п.ф-лы, 10 ил., 1 табл.
| Подогреватель катодов электровакуумных приборов | 1981 |
|
SU1018540A1 |
| ОСНАСТКА ДЛЯ ФОРМОВАНИЯ ИЗДЕЛИЙ ИЗ ПОЛИМЕРНЫХ КОМПОЗИЦИОННЫХ МАТЕРИАЛОВ И СПОСОБ ЕЕ ИЗГОТОВЛЕНИЯ | 2014 |
|
RU2576303C1 |
| Ogawa I | |||
| et al | |||
| Nature | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
