Настоящее изобретение касается новых цитохром Р450-монооксигеназ с измененной специфичностью субстрата, пригодных для окисления органических субстратов, например N-гетероциклических ароматических соединений, кодированных для этого последовательностей нуклеотидов, содержащих эту последовательность экспрессионных конструктов и векторов, и тем самым трансформированных микроорганизмов, способа микробиологического окисления различных органических субстратов, таких как N-гетероциклических ароматических соединений и, в частности, способа изготовления индиго и индирубина.
Энзимы с новыми функциями и свойствами могут быть получены или путем отсеивания природных проб или путем изменения белка известных энзимов. При определенных обстоятельствах последний метод может быть более предпочтительным в получении свойств, которые невероятны на пути природной селекции. Несмотря на различные попытки по созданию энзимов, до сих пор мало удачных исследований по стимулированию каталитической активности мутантов энзимов относительно определенного субстрата (1-10). В этих известных случаях субстраты структурно тесно связаны с природными субстратами соответствующего энзима. До сих пор нет сообщений об успешном создании энзимов, которые после модификации катализировали бы преобразование соединения, которое полностью отличается от природного субстрата энзима.
Выделенная из бактерии Bacillius megaterium цитохром Р450-монооксигеназа обычно катализирует субтерминальное гидроксилирование длинноцепочечных насыщенных кислот и соответствующих амидов и спиртов из них или эпоксидирование ненасыщенных длинноцепочечных жирных кислот со средней длиной цепи (11-13). Оптимальная длина цепи насыщенных жирных кислот составляет от 14 до 16 атомов углерода. Жирные кислоты с длиной цепи менее 12 не гидроксилируются (11).
Структура гем-домена Р450 ВМ-3 определялась с помощью рентгеноструктурного анализа (14-16). Места связывания субстрата имеют форму длинного туннелевидного отверстия, которое от поверхности молекулы идет к гем-молекуле и ограничивается почти исключительно гидрофобными остатками аминокислот. Отдельные заряженные остатки на поверхности гем-домена являются остатками Arg47 и Туr51. Принимается, что они участвуют в связывании карбоксилатной группы субстрата путем образования водородной связи (14). Мутация Arg47 к Glu вызывает дезактивацию энзима арахидоновой кислоты (13), но повышает его активность в отношении С12-С14-соединений алкилтриметиламмиака. Использование субстрата для ароматических соединений, в частности для одно-, двух- или полициклических, в частности, гетероциклических ароматических углеводородов, алканов, алкенов, циклоалканов и циклоалкенов относительно этого энзима не было описано. До сих пор среди специалистов было принято, что иные, чем описанные здесь органические субстраты, как, например, индол, не могут быть субстратами по причине очевидных структурных различий с природными субстратами Р450 ВМ-3, в частности, из-за отсутствия функциональных групп, которые могли бы связывать вышеупомянутые остатки в кармане субстрата.
Поэтому задачей настоящего изобретения является подготовка новых цитохром Р450 монооксигеназ с измененной специфичностью субстрата или измененным профилем субстрата. В частности, должны быть подготовлены мутанты моно-оксигеназы, которые в сравнении с немутировавшим природным энзимом должны быть энзиматически активны со структурно существенно отличающимися субстратами.
"Измененный профиль субстрата" следует отличать у заявляемых мутантов по отношению к природным энзимам. Для соответствующих мутантов наблюдается, в частности, улучшение реакционной способности, например повышение удельной активности (выраженной в нмоль преобразованного субстрата/в минуту/к нмоль энзима Р450), и/или, как минимум, кинетического параметра, выбранного среди Kcat, Km или Kcat/Кm (например, как минимум, 1%, от 10% до 1000%, от 10% до 500% или от 10% до 100%) при преобразовании, как минимум, одного определенного в группах от а) до d) окисляемого соединения. Заявляемая реакция окисления охватывает энзимно-каталитическое окисление, как минимум, одного экзогенного (т.е. добавляемого в реакционную среду) или эндогенного (уже имеющегося в реакционной среде) соединения. В частности, заявляемая реакция окисления охватывает моно- или полигидроксилирование, как, например, моно- и/или дигидроксилирование по отношению к алифатической или ароматической С-Н-группе, или эпоксидирование по отношению к преимущественно неароматической С=С-группе. Возможны также комбинации вышеуказанных реакций. Непосредственный продукт реакции может преобразовываться, кроме того, в рамках неэнзиматической последовательной или побочной реакции. Комбинации такого рода энзиматических и неэнзиматических процессов также являются предметом изобретения.
Вышеупомянутые задачи неочевидным образом могут быть решены путем использования новых цитохром Р450-монооксигеназ, которые, например, способны окислять N-гетероциклические двух- или многозвенные ароматические соединения.
В частности, предметом изобретения являются такие монооксигеназы, у которых связывающая субстрат область способна путем местного специфичного мутагенеза функционально воспринимать новые, например, N-гетероциклические субстраты.
В предпочтительной форме выполнения изобретения новые монооксигеназы растворимы, т.е. существуют не в мембранно-связанной форме и в этой форме энзиматически активны.
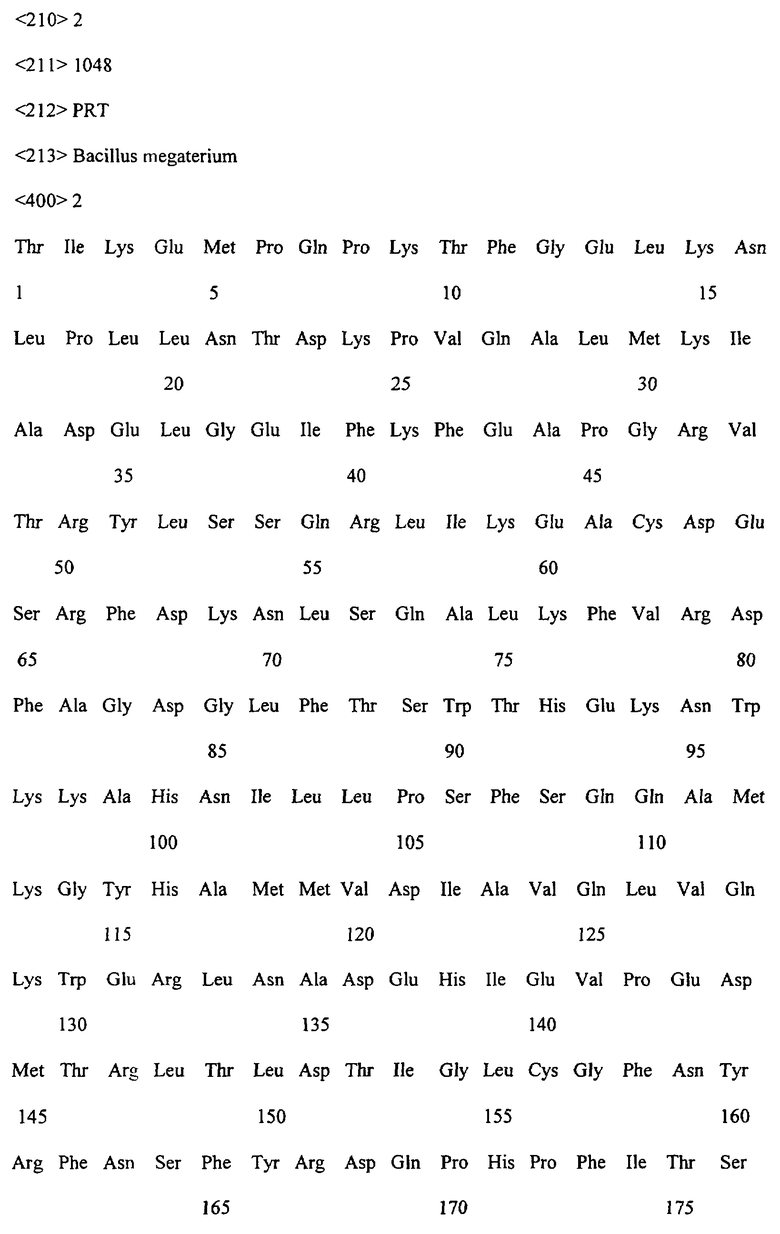
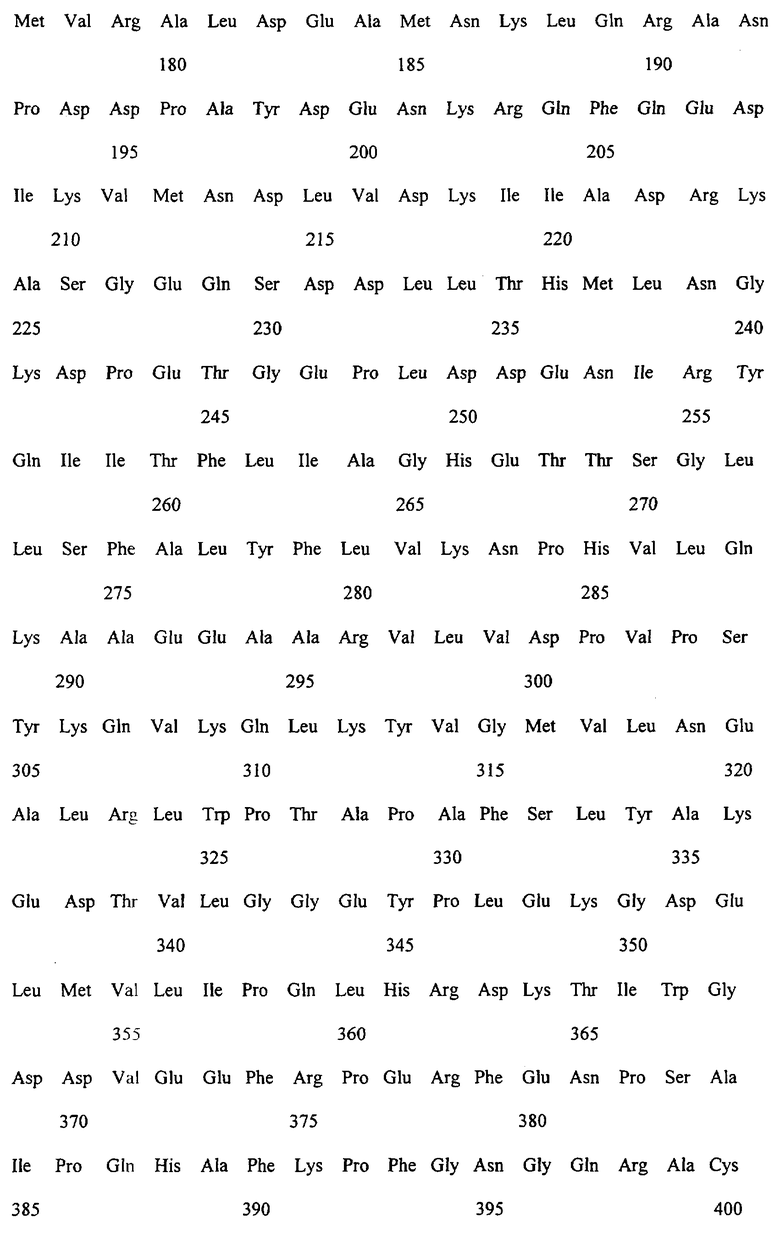
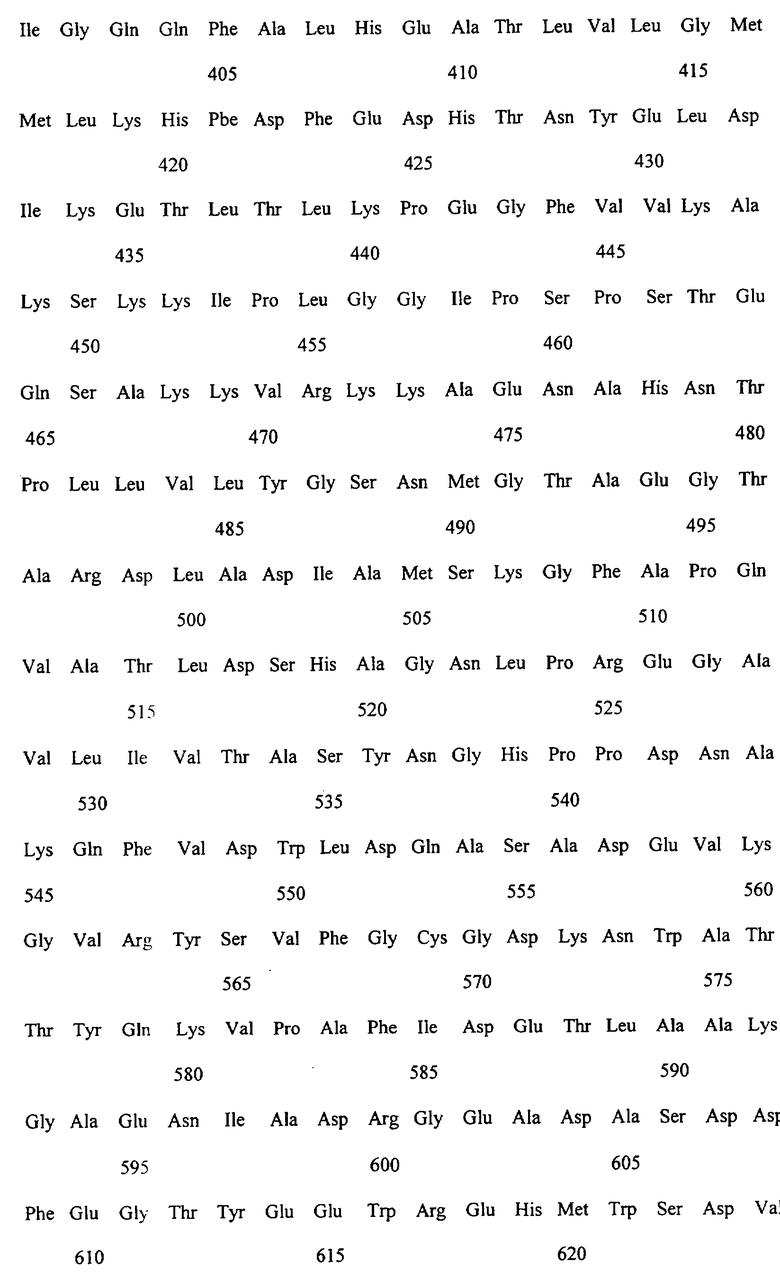
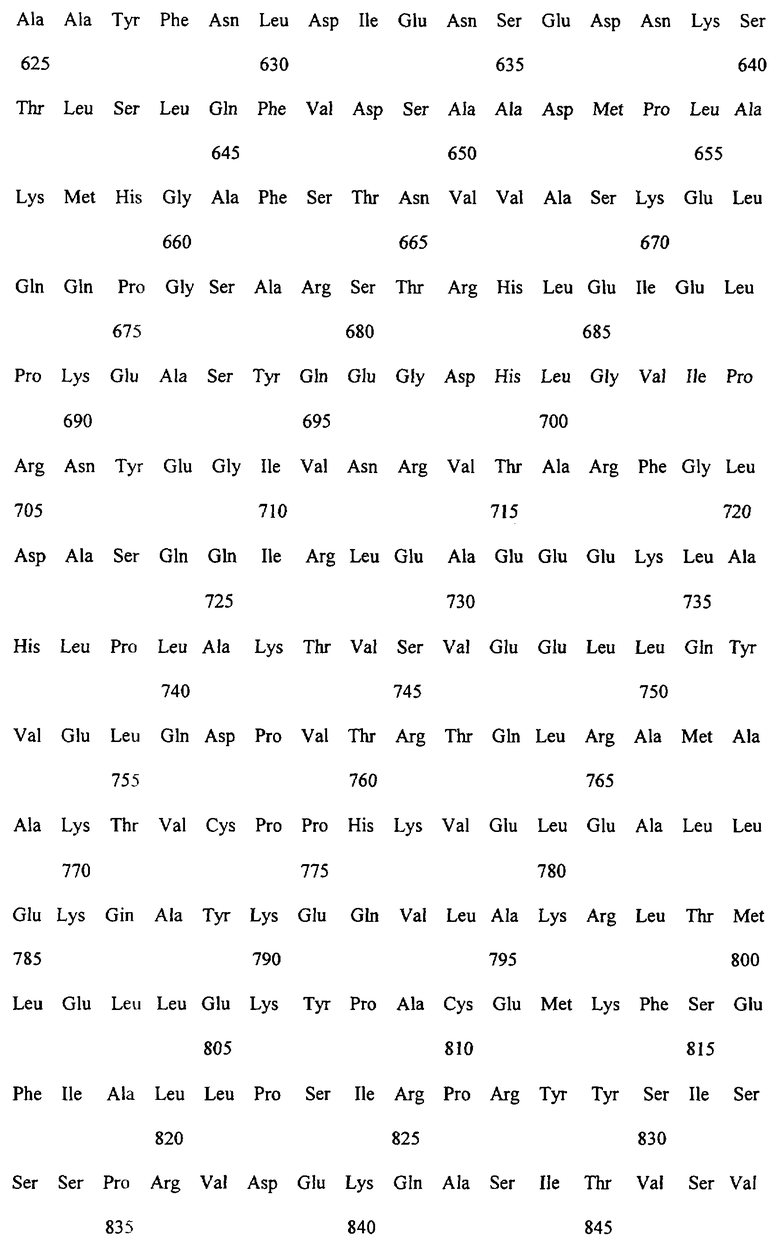
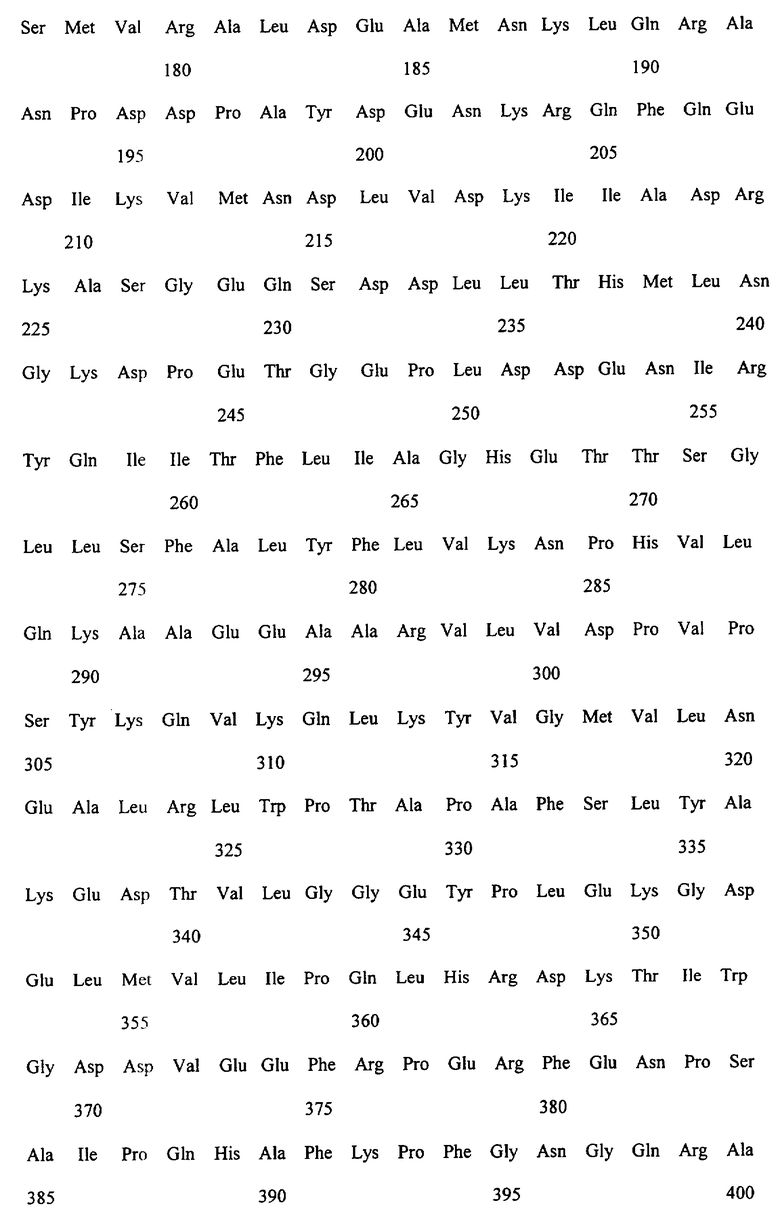
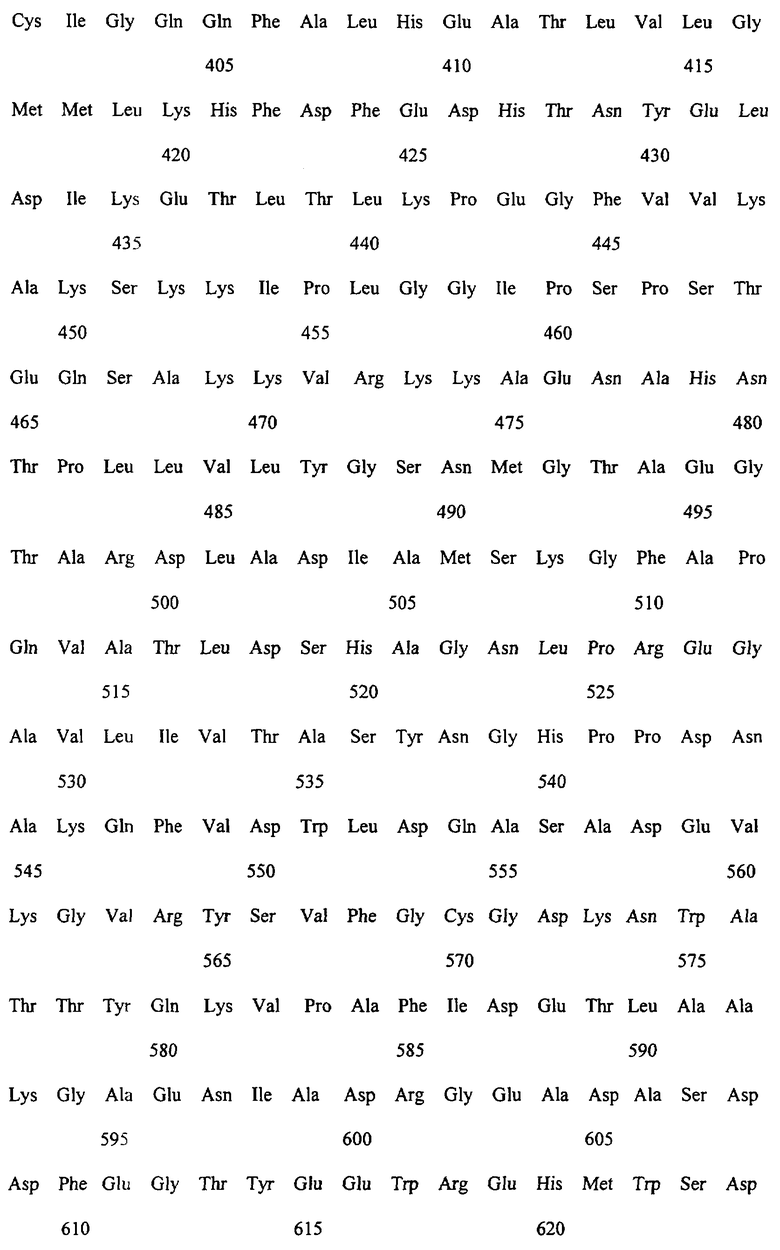
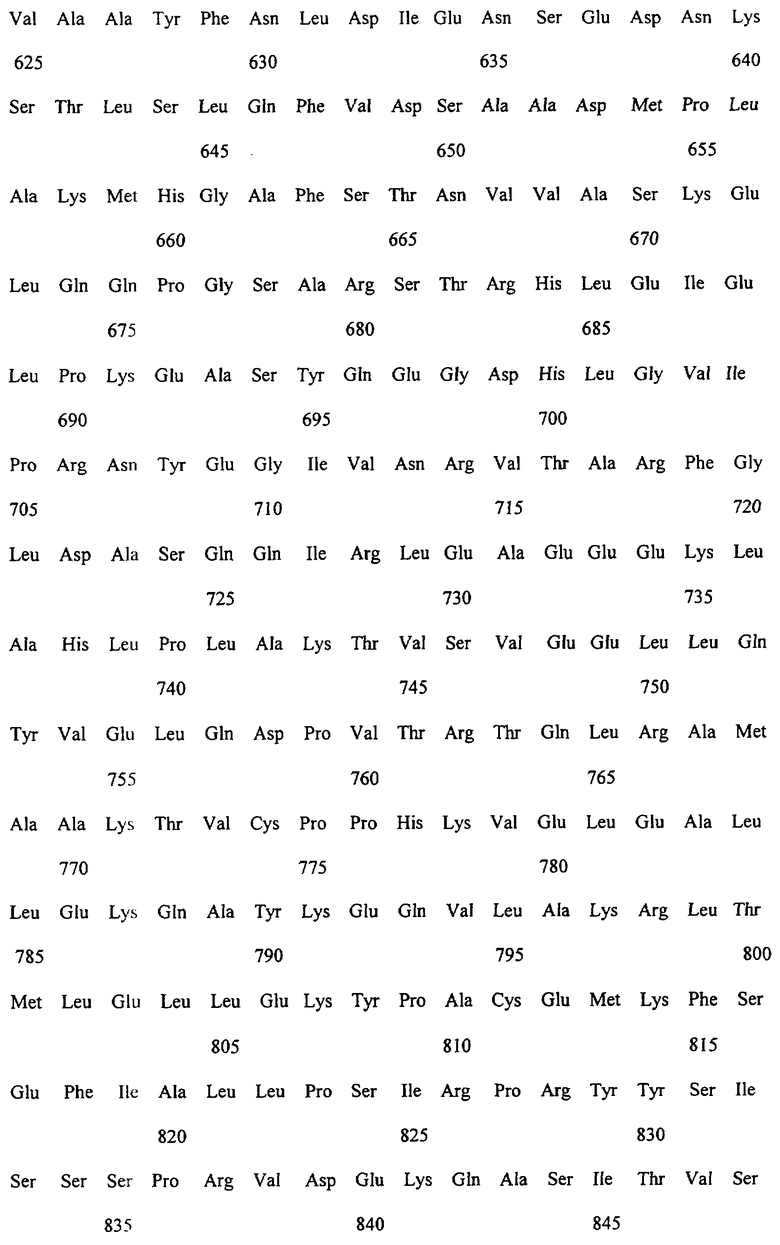
Заявляемые монооксигеназы преимущественно производны из цитохром Р450-монооксигеназы бактериального происхождения, как, в частности, производная из цитохром Р450-монооксигеназы ВМ-3 из Bacillicus megaterium с последовательностью аминокислот согласно SEQ ID NO:2, которая имеет, по меньшей мере, одну функциональную, т.е. способствующую окислению новых органических субстратов (см., в частности, определенные от а) до d) группы соединений), как, например, N-гетероциклические одно-, двух- или многозвенные ароматические соединения, мутацию в области последовательности аминокислот 172-224 (замкнутая область F/G), 39-43 (β-полоса 1), 48-52 (β-полоса 2), 67-70 (β-полоса 3), 330-335 (β-полоса 5), 352-356 (β-полоса 8), 73-82 (спираль 5), 86-88 (спираль 6).
Изготовленные согласно изобретению мутанты цитохром Р450-монооксигеназы предпочтительно, как минимум, способны к одной из следующих реакций:
a) окисление в случае необходимости замещенных N-, О-, или S-гетероциклических одно-, двух- или многозвенных ароматических соединений;
b) окисление в случае необходимости замещенных одно- или многозвенных ароматических углеводородов;
c) окисление неразветвленных или разветвленных алканов и алкенов и
d) окисление в случае необходимости замещенных циклоалканов и циклоалкенов.
Предпочтительные мутанты монооксигеназы имеют, по меньшей мере, одну функциональную мутацию, в частности, замещение аминокислоты, как минимум, в одной из областей последовательности 73-82, 86-88 и 172-224. Так, например, Phe87 может быть замещена аминокислотой с алифатической боковой цепью, как, например, Ala, Val, Leu, в частности Val, Leu188 может быть замещен аминокислотой с амидной боковой цепью, как, например, Asn или, в частности, Gln; а Аlа74 может замещаться другой аминокислотой с алифатической боковой цепью, как, например, Val и, в частности, Gly.
Особенно предпочтительные мутанты монооксигеназы этого типа отличаются тем, что они имеют, по меньшей мере, одно- или многократные замещения аминокислот:
1) Phe87Val;
2) Phe87Val, Leu188Gln; или
3) Phe87Val, Leu188Gln; Ala74Gly;
а также их функциональные эквиваленты. Числовое значение здесь обозначает позицию мутации; перед числовым значением стоит первоначальная, за числовым значением - нововведенная аминокислота.
"Функциональные эквиваленты" или аналоги конкретно проявляющихся мутантов в этой связи - различные мутанты, которые в дальнейшем обладают желательной специфичностью субстрата в рамках, как минимум, одной вышеназванной реакции окисления а)-d), т.е., например, по отношению к гетероциклическим ароматическим углеводородам и, например, гидроксилируют индол, или в дальнейшем по сравнению с природными энзимами обнаруживают "измененный профиль субстрата".
Под "функциональными эквивалентами" понимают в соответствии с изобретением также мутанты, которые обнаруживают, как минимум, в одной из вышеназванных позиций последовательности иное, чем конкретно названное замещение аминокислоты, но, несмотря на это, приводящее к мутанту, который также, как конкретно названные мутанты, по сравнению с природными энзимами демонстрируют "измененный профиль субстрата" и катализируют, как минимум, одну из вышеназванных реакций окисления. Функциональный эквивалент задается, в частности, также тогда, когда изменения в профиле субстрата качественно совпадают, т.е., например, когда одинаковые субстраты преобразуются с различной скоростью.
"Функциональные эквиваленты" охватывают, конечно, также мутанты Р450-монооксигеназы, которые доступны путем мутации энзимов Р450 из других организмов тем же способом, как конкретно названные Р450 ВМ-3. Например, путем последовательного сравнения областей можно установить гомологичные регионы последовательностей. С помощью современных методов молекулярного моделирования тогда можно, основываясь на конкретных задачах изобретения, приняться за эквивалентные, влияющие на образец реакции мутации.
"Функциональные эквиваленты" охватывают также мутанты, полученные путем одного или многих добавок, замещений, удалений и/или преобразований аминокислот, причем названные дополнительные изменения могут появляться в каждой позиции последовательности, пока они приводят к мутанту с измененным профилем субстрата в вышеозначенном смысле.
Окисляемые согласно изобретению субстраты группы а) в случае необходимости являются замещенными гетероциклическими одно-, двух- или многозвенными ароматическими соединениями; в частности, окисляемыми или гидроксилируемыми N-, О-, или S-гетероциклическими одно-, двух- или многозвенными ароматическими соединениями. Они охватывают преимущественно двух или трех, в частности двух, от четырех до семизвенных, в частности шести или пятизвенные конденсированные кольца, причем, как минимум, одно, преимущественно все кольца имеют ароматический вид и, как минимум, одно из ароматических колец несет в себе от одного до трех, преимущественно один N-, О- или S-гетероатом. В общей кольцевой структуре могут быть в случае необходимости один или два других одинаковых или различных гетероатома. Ароматические соединения могут нести также от 1 до 5 заместителей кольцевого углерода или гетероатомов. Примерами подходящих заместителей являются от С1- до С4-алкил, как метил, этил, n- или i-пропил, или n-, i- или t-бутил, или от С2- до С4-алкенил, как этенил, 1-пропенил, 2-пропенил, 1-бутенил, 2-бутенил или 3-бутенил, гидроксилы и галогены, как F, Cl или Br. Названные алкиловые или алкениловые заместители в случае необходимости могут иметь также кето- или альдегидную группу, как, например, пропан-2-он-3-ил, бутан-2-ол-4-ил, 3-бутен-2-ол-4-ил. Не ограничивающими примерами пригодных гетероциклических субстратов являются, в частности, двухзвенные гетероциклы, как индол, N-метилиндол и их аналоги, замещенные от одного до трех вышеопределенными заместителями атомов углерода, как, например, 5-хлор- или 5-бром-индол; а также хинолин и производные хинолина, как, например, 8-метилхинолин, 6-метилхинолин и хинальдин; бензотиофен и его аналоги, замещенные от одного до трех вышеопределенными заместителями атомов углерода.
Окисляемые согласно изобретению субстраты группы b) в случае необходимости являются замещенными одно- или многозвенными ароматическими углеводородами, как бензол и нафталин. Ароматические соединения в случае необходимости могут быть однократно или многократно замещенными и, например, иметь от 1 до 5 заместителей атомов углерода кольца. Примерами подходящих заместителей являются от C1- до С4-алкил, как метил, этил, н- или изопропил, или н-, изо- или трет-бутил, или от С2- до С4-алкенил, как этенил, 1-пропенил, 2-пропенил, 1-бутенил, 2-бутенил или 3-бутенил, гидроксилы и галогены, как F, Cl или Br. Названные алкиловые или алкениловые заместители в случае необходимости могут иметь также кето- или альдегидную группу, как, например, пропан-2-он-3-ил, бутан-2-ол-4-ил, 3-бутен-2-ол-4-ил. Ароматический углеводород в случае необходимости может быть конденсирован четырех-семичленным неароматическим кольцом. Неароматическое кольцо в случае необходимости может обнаруживать одну или две двойных связи С=С, одно или многократно быть замещено вышеупомянутыми заместителями и в случае необходимости нести один или два кольцевых гетероатома. Примерами особо употребительных ароматических углеводородов являются однозвенные, как кумол, а также двухзвенные субстраты, как инден и нафталин, а также замещенные от одного до трех вышеопределенными заместителями атомов углерода.
Окисляемые согласно изобретению субстраты группы с) - неразветвленные или разветвленные алканы и алкены с 4-15, преимущественно 6-12 атомами углерода. В качестве примеров могут быть названы н-бутан, н-пентан, н-гексан, н-гептан, н-октан, н-нонан, н-декан, н-ундекан и н-додекан, а также одно- или многократно разветвленные аналоги этих соединений, как, например, аналоги соединений с 1-3 боковыми метильными группами; или однократно или многократно, например, однократно ненасыщенные аналоги вышеназванных алканов.
Окисляемыми согласно изобретению субстратами группы d) являются в случае необходимости замещенные циклоалканы и циклоалкены с 4-8 кольцевыми атомами углерода. Примеры такого рода представляют циклопентан, циклопентен, циклогексан, циклогексен, циклогептан и циклогептен. Кольцевая структура при этом может содержать один или несколько, например, от 1 до 5 заместителей согласно вышеуказанному определению для соединений группы а) и b). Неограничивающими примерами для этого являются иононы, как α-, β- и γ-иононы, а также соответствующие метилиононы и изометилиононы. Особенно предпочтительны α- и β-иононы.
Предметом изобретения являются также последовательности нуклеиновых кислот, кодированные для заявляемой монооксигеназы. Предпочтительные последовательности выведены из SEQ ID NO:1, которая, как минимум, имеет одно нуклеотидное замещение, ведущее к одной из вышеописанных функциональных аминокислотных мутаций. Предметом изобретения являются, кроме того, функциональные аналоги нуклеиновых кислот, получающиеся путем добавления, замещения, вставки и/или удаления одиночных или нескольких нуклеотидов, которые в дальнейшем кодируют монооксигеназу с желаемой специфичностью субстрата, как, например, с окисляющей индол активностью.
В соответствии с изобретением охватываются также такие последовательности нуклеотидов, которые включают так называемые немые мутации, или изменены соответственно использованию для кодировки специального организма происхождения или организма хозяина по сравнению с конкретно названной последовательностью, также как и предшествующие природные варианты. Изобретение охватывает, кроме того, также разновидности последовательностей нуклеиновых кислот, полученные путем дегенерации генетического кода (т.е. без изменения корреспондирующей последовательности аминокислот) или консервативного замещения нуклеотидов (т.е. корреспондирующие аминокислоты заменяются другими аминокислотами того же заряда, величины, полярности и/или растворимости), а также измененные путем добавления, вставки, инверсии или удаления нуклеотидов последовательности, которые кодируют заявляемую монооксигеназу "с измененным профилем субстрата", а также корреспондирующие комплементарные последовательности.
Предметом изобретения являются, кроме того, экспрессионные конструкты, содержащие при генетическом контроле регулятивных последовательностей нуклеиновых кислот, как минимум, одну кодирующую заявляемые мутанты последовательность нуклеиновых кислот, а также векторы, охватывающие, как минимум, один из этих экспрессионных конструктов.
Соответствующие изобретению конструкты охватывают промотор 5'-вверх по длине соответствующей кодирующей последовательности и терминаторную последовательность 3'-вниз по длине, а также, в случае необходимости, другие обычные регулятивные элементы, а именно оперативно связанные с кодирующей последовательностью. Под оперативным связыванием понимают последовательное расположение промотора, кодирующей последовательности, терминатора и, в случае необходимости, других регулятивных элементов такого рода, чтобы каждый из регулятивных элементов мог согласованно выполнять свою функцию при экспрессии кодирующей последовательности. Примерами оперативно связанных последовательностей являются последовательности мишеней, а также трансляционный усилитель, умножитель, сигналы полиаденилирования и другие. Другие регулятивные элементы охватывают селективный маркер, сигналы усиления, реплики и тому подобное.
Дополнительно к искусственным регуляционным последовательностям перед первоначальным структурным геном могут быть еще природная регуляционная последовательность. Путем генетического изменения эта природная регулировка может выключаться и повышать или понижать экспрессию гена. Конструкт гена может быть построен проще, что означает, что перед структурным геном не могут быть вставлены дополнительные регуляционные сигналы, а природный промотор со своей регулировкой не удаляется. Вместо этого природная регулировочная последовательность мутируется так, чтобы больше не нужно было регулировки, а экспрессия гена повышалась или понижалась. Последовательности нуклеиновых кислот могут содержаться в генном конструкте в одной или многих копиях.
Примерами употребительных промоторов являются: cos-, tac-, trp-, tet-, trp-tet-, lpp-, lac-, lpp-lac-, lacIg-, T7-, T5-, T3-, gal-, trc-, ara-, SP6-, 1-PR или 1-PL-промоторы, которые находят применение преимущественно в грамотрицательных бактериях, а также грамположительные промоторы amy и SPO2, дрожжевые промоторы ADCl, MFa, AC, P-60, CYC1, GAPDH или растительные промоторы CaMV/35S, SSU, OCS, lib4, usp, STLS1, B33, nos или убиквитиновые или фазеолиновые промоторы. Особенно предпочтительно применение индуцируемых промоторов, как, например, свето- и, особенно, термоиндуцированных промоторов, как PrPl. Принципиально, все природные промоторы могут применяться с их регулирующими последовательностями. Исходя из этого, могут также с выгодой применяться синтетические промоторы.
Названные регуляторные последовательности должны обеспечивать целенаправленную экспрессию последовательностей нуклеиновых кислот и экспрессию протеинов. Это может, например, в зависимости от организма хозяина означать, что ген экспримируется или переэкспримируется только после индукции, или он тотчас экспримируется или переэкспримируется.
Регуляторные последовательности, соответственно, факторы могут при этом преимущественно позитивно влиять на экспрессию и тем самым повышать или понижать ее. Таким образом, усиление регуляторных элементов может осуществляться преимущественно в плоскости транскрипции, причем применяются сильные транкскрипционные сигналы, как промоторы и/или "усилители". Наряду с этим возможно также усиление трансляции, причем, например, улучшается стабильность мРНК.
Изготовление экспрессионной кассеты осуществляется путем слияния подходящего промотора с подходящей последовательностью нуклеотидов моноокисигеназы, а также сигналом терминатора или полиаденилирования. Для этого применяют доступную технику рекомбинации и клонирования, описанную в T.Maniatis, E.F.Frinsch, J.Sambrook, Manual Cloning: A Labaratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, Нью-Йорк (1989), а также в T.J SIlhavy, M.L.Berman, L.W.Enquist, Experiments with Gene fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, Нью-Йорк (1984), и в Ausubel, F.M. et al.. Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley Interscience (1987).
Рекомбинантный конструкт нуклеиновой кислоты, соответственно, генный конструкт вводится для расширения в пригодный для этого организм хозяина преимущественным образом в специфическом для хозяина векторе, который делает возможным оптимальную экспрессию гена в хозяине. Векторы известны специалистам и могут быть взяты из "Cloning Vectors" (Pouwels P.H. et al., Hrsg, Elsevier, Амстердам - Нью-Йорк - Оксфорд, 1985). Под этими векторами кроме плазмидов следует понимать также все другие известные специалисту векторы, как, например, фаги, вирусы, как SV40, CMV (цитомегаловирус), бакуловирус и аденовирус, транспозонсы, IS-элементы (транспосабельные элементы), плазмиды, космиды и линейные или циркулярные ДНК. Эти векторы могут реплицироваться или автономно в организме хозяина, или хромосомно.
С помощью описанных в изобретении векторов производятся рекомбинантные микроорганизмы, которые трансформируются, например, как минимум, одним описанным в изобретении вектором и могут применяться для производства мутантов. Преимущественно вышеописанные описанные заявляемые рекомбинантные конструкты вводятся в пригодную для этого систему хозяина, затем экспримируются. При этом специалистами применяются преимущественно известные методы клонирования и трансфекции, чтобы вставить названные нуклеиновые кислоты в соответствующую экспрессионную систему для экспрессии. Пригодные системы описываются, например, в Carrent Protocols in Molecular Biology, F. Ausubel et al., Hrsg., Wiley Interscience, Нью-Йорк 1997.
В качестве организмов хозяина принципиально пригодны все организмы, которые делают возможной экспрессию заявляемых нуклеиновых кислот, вариантов их аллелей, их функциональных эквивалентов или производных. Под организмами хозяина следует понимать, например, бактерии, грибки, дрожжи, растительные или животные клетки. Предпочтительными организмами хозяина являются бактерии, как бактерии из рода Escherichia (например, Escherichia coli), стрептомицеты, бациллы или псевдомонады, эукариотические микроорганизмы, как Saccharimyces cerevisiae, Aspergillus, эукариотические клетки более высокого порядка животного или растительного происхождения, например, Sf9 или СНО.
При желании генный продукт можно вносить для экспрессии также в трансгенные организмы, как трансгенные животные, в частности, в мышей, овец или в трансгенные растения. В случае трансгенных организмов речь может идти также о так называемых Knock-Out животных или растениях, в которых был выключен корреспондирующий эндогенный ген, например, путем мутации или частичного или полного удаления.
Селекция успешно трансформированных организмов осуществляться маркерным геном, который также содержится в векторе или экспрессионной кассете. Примером таких маркерных генов являются гены стойкости к антибиотикам и к энзимам, которые катализируют цветную реакцию, вызывающую окраску трансформированных клеток. Последние могут тогда отбираться автоматически. Микроорганизмы, успешно трансформированные вектором, которые несут ген стойкости к антибиотикам (например, G418 или гидромицин), можно селектировать путем сред, содержащих антибиотики, или питательных почв. Маркерные белки, которые представлены на поверхности клеток, могут использоваться для селекции посредством аффинной хроматографии.
Комбинация организмов хозяев с векторами, подходящими для организмов, как то плазмиды, вирусы или фаги, например плазмиды с системой полимераза/промотор РНК, фаги λ, μ, или другие умеренные фаги или транспозонсы и/или другие имеющие преимущество регуляторные последовательности образует экспрессионную систему. Например, под понятием "экспрессионная система" следует понимать комбинацию из клеток млекопитающих, как СНО-клетки, и векторов, как вектор pcDNA3neo.
Как уже упоминалось, генный продукт может преимущественно вводиться для экспресии в трансгенные животные, например в мышей, в овец или в трансгенные растения. Также возможно программировать свободные от клеток трансляционные системы с полученной из нуклеиновой кислоты РНК.
Дальнейшим предметом изобретения является способ изготовления заявляемой монооксигеназы, когда культивируют микроорганизм, производящий монооксигеназу, в случае необходимости индуцируют экспрессию монооксигеназы и выделяют монооксигеназу из культуры. Заявляемая монооксигеназа при необходимости может производиться в промышленных масштабах.
Микроорганизмы могут культивироваться и ферментироваться известными способами. Бактерии могут размножаться, например, в ТВ- или LB-среде и при температуре от 20°С до 40°С и значении рН от 6 до 9. Подробно подходящие условия культивации описываются в T.Maniatis, E.F. Frinsch, J.Sambrook, Manual Cloning: A Labaratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, Нью-Йорк (1989).
В том случае, когда монооксигеназа не выделяется в культуральную среду, клетки разлагаются, а монооксигеназа добывается из лизата способом изоляции белка. Клетки могут разлагаться на выбор: высокочастотным ультразвуком, высоким давлением, как, например, в ячейке давления Френча, путем осмолиза, путем воздействия детергентов, литических энзимов или органических растворителей, с помощью гомогенизаторов или путем комбинации нескольких приведенных способов. Очистка монооксигеназы может быть достигнута такими известными хроматографическими способами, как хроматографией с молекулярным ситом (гелевая фильтрация), как хоматография Q-Sepharose, ионообменная хроматография и гидрофобная хроматография, а также другими обычными способами, как ультрафильтрование, кристаллизация, высаливание, диализ и нативный гелевый электрофорез. Подходящие способы описаны, например, в Cooper F.G., Biochemische arbeitsmenhoden, Verlag Walter de Gruyter, Berlin, New York или в Scopes R., Protein Purification, Springer Verlag, Нью-Йорк, Хайдельберг, Берлин.
Особенно выгодно применять для изоляции рекомбинантного белка векторную систему или олигонуклеотиды, которые продлевают кДНК на определенную последовательность нуклеотидов и тем самым кодируют удлиненные полипептиды или смешанные белки, которые служат для более простой очистки. Соответствующие модификации такого рода являются, например, действующими как иммобилизатор так называемыми "тагами", как, например, известная как гексагистидиновый иммобилизатор модификация или эпитоп, который может распознаваться как антиген антител (описан, например, в Harlow E. and Lane D., 1988, Antibodies: A Laboratory Manual. Cold Spring Harbor (N.Y.) Press). Эти иммобилизаторы могут служить для связывания белка на твердых носителях, например полимерных матрицах, которые, например, могут быть засыпаны в хроматографическую колонку, или могут применяться на микротитровальной пластине, или на прочих носителях.
Одновременно эти иммобилизаторы могут применяться для распознавания белков. Для распознавания белков могут применяться, кроме того, такие обычные маркеры, как флуоресцентные красители, энзимные маркеры, которые после реакции с субстратом образуют детектируемый продукт реакции, или радиоактивные маркеры самостоятельно или в комбинации с иммобилизаторами для дериватизации белков.
Изобретение касается, кроме того, способа микробиологического окисления органических соединений, как, например, N-гетероциклических одно-, двух- или многозвенных вышеопределенных ароматических соединений, которые отличаются тем, что
а1) культивируют вышеопределенный рекомбинантный микроорганизм в культуре в присутствии экзогенного (внесенного извне) или образованного промежуточно окисляющего субстрата заявляемой монооксигеназы, преимущественно в присутствии кислорода (т.е. аэробно); или
а2) инкубируют содержащую субстрат реакционную среду с заявляемым энзимом, преимущественно в присутствии кислорода и донора электронов; и
b) выделяют образовавшийся продукт окисления или последовательный продукт этого из среды.
Требуемый для преобразования кислород поступает в реакционную среду из окружающего воздуха или может вводиться, если требуется, известным способом.
Предпочтительно окисляемый субстрат выбирается среди
a) в случае необходимости замещенных N-гетероциклических одно-, двух- или многозвенных вышеопределенных ароматических соединений;
b) в случае необходимости замещенных одно- или многозвенных ароматических углеводородов;
c) неразветвленных или разветвленных алканов и алкенов;
d) в случае необходимости замещенных циклоалканов и циклоалкенов.
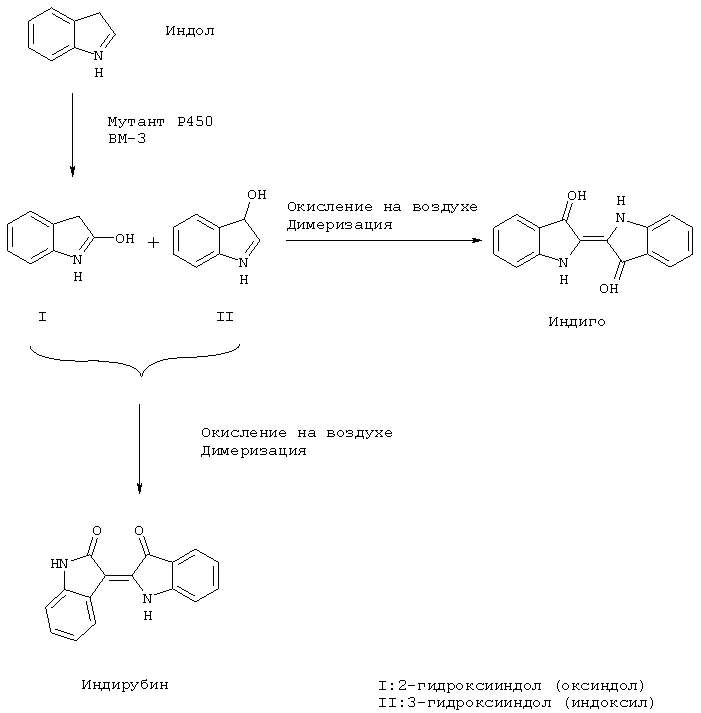
Предпочтительный вариант способа направлен на образование индиго/индирубина и отличается тем, что субстрат является промежуточно образованным в культуре индолом, и из культуральной среды выделяют образующийся индиго и/или индирубин, который был получен путем окисления промежуточно образующихся гидроксииндолов.
Если окисление по настоящему изобретению проводится с помощью рекомбинантного микроорганизма, то культивация микроорганизмов осуществляется преимущественно в присутствии кислорода в комплексной среде, как, например, ТВ- или LB-среда при температуре культивации примерно 20-40°С и значении рН от 6 до 9, пока не будет достигнута достаточная плотность клеток. Добавка экзогенного индола обычно не требуется, поскольку он образуется микроорганизмом как промежуточный продукт. При преобразовании других субстратов может однако требоваться добавка экзогенного субстрата. Чтобы можно было лучше управлять реакцией окисления, предпочтительно использование индуцируемого, в особенности, индуцируемого температурой промотора. При этом температуру повышают до точки индукции, например, 42°С при промоторе PrPl, поддерживают ее в течение достаточного времени, например от 1 до 10 или от 5 до 6 часов, для экспрессии активной монооксигеназы, а затем уменьшают температуру до 30-40°С. При этом культивация продолжается в присутствии кислорода от 12 часов до 3 дней. В частности, при окислении индола значение рН за счет добавки NaOH может повышаться, например, до 9-10, благодаря чему дополнительно способствуется образование индиго, соответственно, индирубина путем окисления на воздухе энзиматически образованных продуктов окисления 2- и 3-гидроксииндола.
Образование индиго/индирубина согласно настоящему изобретению поясняется следующей схемой реакции:
Если окисление согласно изобретению, напротив, проводится с помощью очищенных или обогащенных мутантов энзима, то заявляемый энзим растворяют в содержащей экзогенный субстрат, например индол, среде (примерно 0,01-10 мМ, или 0,05-5 мМ) и проводят преобразование преимущественно в присутствии кислорода, при температуре примерно от 10°С до 50°С, например от 30°С до 40°С, и значении рН примерно от 6 до 9 (какое, например, устанавливается при 100-200 мМ фосфатного или ТРИС-буфера), а также в присутствии восстановителя, причем содержащая субстрат среда содержит, кроме того, по отношению к окисляемому субстрату примерно 1-100-кратный или 10-100-кратный молярный избыток восстановительных эквивалентов. Предпочтительным восстановителем является НАДФ. Добавка восстановителя может, в случае необходимости, осуществляться порциями.
Аналогичным способом в качестве субстратов предпочтительны: н-гексан, н-октан, н-декан, н-додекан, кумол, 1-метилиндол, 5-Cl или Br-индол, инден, бензотриофен, α, β и γ-ионон, акридин, нафталин, 6-метил или 8-метилхинолин, хинолин и хинальдин.
Например, заявляемая энзиматическая реакция окисления может проводиться при следующих условиях:
Перед началом реакции, например, путем добавки донора электронов (например, НАДФ) может проводиться краткосрочная (1-5 минут) предварительная инкубация (примерно 20-40°С). Преобразование осуществляется аэробно, в случае необходимости при дополнительном введении кислорода.
При заявляемом способе окисления получающийся в реакционной среде или добавленный кислород расщепляется энзиматически и редуктивно. Требуемый восстановительный эквивалент поставляется добавляемым восстановительным средством (донором электронов).
Образующийся продукт окисления далее отделяется от среды и очищается обычным способом, как, например, путем экстракции или хроматографии.
Дальнейшими предметами изобретения являются биореакторы, включающие заявляемый энзим или заявляемый рекомбинантный организм в иммобилизированной форме.
Последний предмет изобретения касается применения заявляемой цитохром Р450-монооксигеназы или заявляемого вектора или микроорганизма для биологического окисления субстрата из групп a)-d), в частности, N-гетероциклических одно, двух или многозвенных ароматических соединений и, предпочтительно, для образования индиго и/или индирубина.
Предлагаемое изобретение более подробно описывается с учетом следующих примеров.
Пример 1
Рандомизация специальных кодонов Р450 ВМ-3
Опыты в существенной степени были проведены так, как описано в (19). Три позиции (Phe87, Leu188 и Аlа74) были рандомизированы с помощью местноспецифического мутагенеза с применением набора Stratagene QuikChange kit (Ла Джолла, Калифорния, США). В отдельных позициях были использованы следующие ПЦР-примеры:
Phe87: 5'-gcaggagacgggttgnnnacaagctggacg-3' (SEQ ID NO:3)
5'-cgtccagcttgtnnncaacccgtctcctgc-3' (SEQ ID NO:4)
Leu188: 5'-gaagcaatgaacaagnnncagcgagcaaatccag-3' (SEQ ID NO:5)
5'-ctggatttgctcgctgnnncttgttcattgcttc-3' (SEQ ID NO:6)
Ala74: 5'-gctttgataaaaacttaaagtcaannncttaaatttgtacg-3' (SEQ ID NO:7)
5'-cgtacaaatttaagnnnttgacttaagtttttatcaaagc-3' (SEQ ID NO:8)
Условия для ПЦР были для всех трех позиций идентичны. В частности, применялось по 50 мкл реакционного объема, 17,5 пмоль каждого примера, 20 пмоль шаблона плазмида ДНК, 3 ед. Pfu полимеразы и 3,25 нмоль для каждого dNTP (дезоксинуклеозид-5'-трифосфат). Реакция ПЦР была начата при 94°С/ 1 мин, а затем был 20 раз повторен следующий цикл: 94°С, 1 мин; 46°С, 2,5 мин; 72°С, 17 мин. После 20 циклов реакция была продолжена 15 мин при 72°С. После ПЦР шаблон ДНК реагировал с 20 ед. Dpni при 37°С в течение 3 часов. Далее трансформировались клетки E.coli DH5α. Трансформированные клетки E.coli DH5α размещались на агаровых LB-платах, которые содержали 150 мг/мл ампициллина. После этого 18 часов при 37°С производилась инкубация.
Пример 2
Экспрессия и очистка Р450 ВМ-3 и ее мутантов и производство синего пигмента
Ген Р450 ВМ-3 и его мутанты был экспримированы под контролем сильного температурно-индуцированного промотора PRPL плазмида pCYTEXP1 в E.coli DH5α, как уже описано (20). Колонии были взяты стерильным зубчатым отборником и перенесены на микротитровальные пластины с 96 ячейками, содержащими 200 мкл среды ТВ и 100 мкг/мл ампициллина на ячейку. После этого всю ночь при 37°С производилась инкубация. 40 мкл клеточной культуры каждой ячейки были затем переведены в культуральную пробирку, которая содержала 2 мл ТВ-среды со 100 мкг/мл ампициллина. После этого в течение 2 часов проводилась культивация при 37°С. Затем для индукции температура повышалась до 42°С на 6 часов. После этого культивация продолжалась всю ночь при 37°С, причем был получен голубой пигмент.
Препаративное изготовление энзима или голубого пигмента проводилось исходя из 300 мл клеточной культуры (OD578 нм=0,8-1,0). Для изоляции энзима клетки подвергались центрифугированию 10 мин при 4000 об/мин, затем ресуспендировались в 0,1 М КхPO4-буфере, рН 7,4. Охлажденные до льда клетки осторожно разлагались с помощью прибора Branson Sonifiers W25 (Дайтцебах, Германия) при выходной мощности 80 Вт путем трехкратного облучения по 2 мин. Суспензии были центрифугированы 20 мин при 32750 g. Сырой экстракт был использован для определения активности, соответственно, для очистки энзима. Очистка энзима осуществлялась, как описано в (21), на что здесь производится настоятельная ссылка. Концентрация очищенного энзима определялась с учетом разницы в экстинкции при 450 и 490 нм, как уже описано в (11), с применением коэффициента экстинкции ε=91 мМ-1см-1.
Пример 3
Выделение мутантов, которые производят большие количества голубого пигмента
Из каждой позиции были выделены соответственно 100 колоний мутантов, которые были получены путем рандомизации кодона соответствующей позиции. Эти колонии были культивированы в культуральных пробирках для производства голубого пигмента. После промывки клеток водой и многих медленных шагов центрифугирования (500 об/мин) голубой пигмент был экстрагирован диметилсульфоксидом (ДМСО). Растворимость голубого пигмента в ДМСО была наибольшей. Абсорбция экстракта была определена в области 677 нм. Те мутанты, которые производили наибольшее количество пигмента по отношению ко всем мутантам определенной позиции, были использованы для секвентирования ДНК (набор ABI DNA Sequenzierungs-Kit; ABI PrismTM 377 DNA Sequencer) и, кроме того, применялись как шаблон для местноспецифического рандомизированного мутагенеза.
Пример 4
Тест на активность для гидроксилирования индола
Активность гидроксилирования индола проверялась в растворе, который содержал 8 мкл раствора индола 10-500 мМ в ДМСО, 850 мкл буфера ТРИС/HCl (0,1 М, рН 8,2) и 0,6 нмоль Р450 ВМ-3 природного типа или мутантов в конечном объеме 1 мл. Смесь была предварительно инкубирована в течение 9 мин, прежде чем запускалась реакция путем добавки 50 мкл водного 1 мМ раствора НАДФ. Реакция прекращалась через 20 с путем добавления 60 мкл 1,2 М КОН. В течение 5-30 с (при аэробных условиях) энзимные продукты были полностью переведены в индиго ([Δ2,2'-бииндолин]-3,3'-дион) и индирубин ([Δ2,2'-бииндолин]-3,3'-дион). Продукты индиго были определены путем их абсорбции в области 670 нм. Градуировочный график для чистого индиго на этой длине волны дает коэффициент экстинкции 3,9 мМ-1см-1. Линейный ход градуировочного графика получен для производства индиго через время реакции в 40 с при применении 0,6 нмоль природного типа, соответственно, мутанта Р450 ВМ-3 и от 0,05 до 5,0 мМ индола. Индирубин имеет очень слабую абсорбцию при 670 нм, а образовавшиеся количества индирубина значительно меньше, чем количество идиго. При определении кинетических параметров образование индирубина не учитывалось. Потребление НАДФ определялось путем измерения в области 340 нм и расчета с использованием коэффициента экстинкции 6,2 мМ-1см-1, как описано в (17).
Пример 5
Очистка индиго и индирубина
После промывки клеток водой и повторяющимся центрифугированием при 500 g образовавшийся голубой остаток экстрагировался тетрагидрофураном (ТГФ). Экстракт выпаривался почти до сухого состояния, и красный пигмент многократно экстрагировался абсолютным этанолом. Оставшееся голубое твердое вещество было растворено в ТГФ и анализировалось методом тонкослойной хроматографии (TLC). Раствор этанола был испарен и очищен с помощью силикагелевой хроматографии (DC 60, Мерк, Дармштадт, Германия; 2 см × 30 см), прежде чем она была промыта ТГФ и петролейным эфиром в соотношении 1:2. Полученный красный раствор был выпарен и анализировался методом TLC. Абсорбционные спектры голубого и красного пигмента были получены с помощью спектрофотометра Ultraspec 3000 (Фармасия, Уппсала, Швеция) в области от 400 до 800 нм. Кроме того, голубой и красный краситель анализировался с помощью масс-спектроскопии и ЯМР-спектроскопии.
Результаты экспериментов
1. Повышение производства голубого пигмента путем мутагенеза Р450 ВМ-3
Природный Р450 ВМ-3 не обладает способностью производства голубого содержащего индиго пигмента, соответственно, промежуточных продуктов - 2-, соответственно, 3-гидроксихинола. Чтобы получить достаточное количество голубого пигмента, Р450 ВМ-3 был подвергнут целенаправленному преобразованию. Все мутанты, которые производят голубой пигмент, были секвенсированы. Было установлено, что мутировала, как минимум, одна из трех следующих позиций: Phe87, Leu188 и Аlа74. Поэтому принималось, что эти три позиции играют решающую роль для активности Р450 ВМ-3 при производстве голубого пигмента. Из структуры гем-домена цитохрома Р450 ВМ-3, комплексированного пальмитоолеиновой кислотой, видно, что Phe87 предохраняет субстрат от ближнего сдвига гем-группы (14). Мутант Phe87Val обнаруживает высокую регио- и стереоселективность при эпоксидации (14S, 15R)-арахидоновой кислоты (13), а мутант Phe87Ala перемещает позицию гидроксилирования ω-1, ω-2 и ω-3 (22). Позиция 87 была поэтому выбрана первой для местно-специфического рандомизированного мутагенеза с помощью ПЦР. В пробирочных культурах было получено 7 колоний, которые после индукции производили незначительное количество голубого пигмента. Колония, которая производила наибольшее количество голубого пигмента, была выбрана для секвенсирования ДНК. Данные секвенции давали замещение Phe87 на Val. Мутант Phe87Val был в итоге применен как шаблон на втором круге местно-специфического рандомизированного мутагенеза в позиции Leu188. Структура гем-домена, комплексированного пальмитоолеиновой кислотой показывает, что репозиционирование F- и G-спирали приводит остаток Leu188 в прямой контакт с субстратом (14). Поэтому эта позиция может играть важную роль в связывании или ориентировании субстрата. После проведения второго скрининга наблюдалась 31 колония, которая производила голубой пигмент. Мутант, который производил наибольшее количество пигмента, содержал замещения Phe87Val и Leu188Gln. Этот мутант был в итоге мутирован в позиции Аlа74 в третьем проходе местно-специфического рандомизированного мутагенеза. При этом получали трехкратные мутанты F87L188A74 (Phe87Val, Leu188Gln и Ala74Gly), которые производили многие мг голубого пигмента в двухлитровой колбе, содержащей 300 мл ТВ-среды. Этого количества было достаточно для выделения и определения голубого пигмента.
2. Выделение и определение голубого пигмента
После промывки клеток голубой остаток был экстрагирован ТГФ и проанализирован методом тонкослойной хроматографии. Голубой пигмент был разделен на быстро продвигающиеся голубые компоненты и медленно продвигающиеся красные компоненты. Оба компонента имеют такие же параметры подвижности, как компоненты пробы коммерческого индиго.
После очистки были зарегистрированы абсорбционные спектры обоих компонентов в ДМСО. Голубой компонент имеет такой же спектр, как проба коммерческого индиго. Очищенные голубые и красные компоненты, соответственно, были проанализированы с помощью масс-спектрометрии. Масс-спектры обоих пигментов имеют сильный ионно-молекулярный пик при m/z=262 и 2 фрагментарных пика при m/z=234 и 205 (относительная интенсивность, соответственно, 10%). Этот образец типичен для индигоидных соединений. Элементарный состав этих ионов был определен с помощью масс-спектроскопии высокого разрешения как C16H10N2O2, C15H10N2O, соответственно, С14H9N2. Это также характерно для структур индигоидных типов. Тем самым голубой пигмент был определен как индиго, а красный - как индирубин. Для подтверждения структуры был проверен ЯМР обоих пигментов в растворе ДМСО-D6. Результаты совпадают с литературными данными.
3. Производство индиго с изолированными энзимами
Известно, что индиго может быть получен из индола путем микробиологической трансформации (24-26). Однако ни одна из этих микробиологических систем не содержит монооксигеназы Р450. Согласно изобретению, прежде всего, должна быть определена каталитическая активность чистого энзима по отношению к индолу. Мутант F87L188A74 был смешан с индолом. Не наблюдалось никакой цветной реакции. Только после добавки НАДФ в реакционную смесь примерно через 20 мин образовался голубой пигмент. Путем доведения значения рН реакционной смеси до значения 11, через 30 с после добавления НАДФ, в течение нескольких секунд стала видна синяя окраска. Контрольные опыты с применением природного Р450 ВМ-3 были всегда отрицательны, даже при применении повышенных концентраций энзима, индола и НАДФ. Голубой пигмент был экстрагирован этилацетатом и проанализирован методом тонкослойной хроматографии. Голубой пигмент снова разделился на быстро продвигающийся голубой и медленно продвигающийся красный компонент. Рентгенофлуоресцентные значения и абсорбционные спектры были идентичны тем же значениям, что и в экстракте ферментационного бульона. Мутанты F87L188A74 Р450 ВМ-3 представляют собой гидроксилазу индола.
До сих пор было описано только 2 пути энзиматической трансформации индола в индиго. Один путь катализирует с помощью диоксигеназы, второй - стирол-монооксигеназы (24, 25). Стехиометрия НАДФ в обоих случаях составляет 2. Поэтому было принято, что в противоположность диоксигеназе заявляемые мутанты F87L188A74 индола гидроксилируются только в одной позиции, чтобы образовать оксиндол (2 гидроксииндол) или индоксил (3-гидроксииндол).
4. Кинетические параметры гидроксилирования индола
Для определения кинетических параметров гидроксилирования индола использовались чистые пробы природного энзима Р450 ВМ-3 и мутантов Leu188Gln, Phe87Val, F87L188 и F87L188A74. Результаты представлены в нижеследующей Табл.1.
Кинетические параметры гидроксилирования индола мутантов Р450 ВМ-3
b) не определено (активность была слишком незначительной чтобы ее можно было измерить)
Даже при избытке очищенного энзима и высокой концентрации индола природный энзим не в состоянии окислить индол. Мутант Leu188Gln обнаруживает незначительную активность. Мутант Phe87Val имеет каталитическую активность 119 М-1с-1 для гидроксилирования индола. Каталитическая эффективность двойных мутантов F87L188 (Phe87Val, Leu188Gln) повышается до 543 М-1с-1 и была повышена за счет введения других замещений Ala74Gly до 1365 М-1с-1. Значения Kcat повышаются от Phe87Val к трехкратным мутантам на, примерно, 35%, в то время как значения Кm уменьшаются примерно в 7 раз. Это указывает на то, что Ala74Gly и Leu188Gln участвуют в связывании субстрата.
Доля обратимости индола (Kcat=2,73 с-1) для трехкратных мутантов F87L188A74 более чем в 10 раз выше, чем для большинства энзимов Р450 (18).
Пример 6
Гидроксилирование н-октана модифицированной цитохром Р450-монооксигеназы
Преобразование было проведено с помощью мутанта монооксигеназы Р450 ВМ-3, который содержит следующие мутации: Phe87Val, Leu188Gln, Ala74Gly.
В качестве субстрата был выбран н-октан. Для гидроксилирования н-октана применялся следующий реакционный набор:
Лиофилизат энзима был растворен в 500 мкл буфера и сразу с субстратом и буфером был инкубирован 5 мин при комнатной температуре. Затем последовало добавление 300 мкл раствора НАДФ (5 мг/мл). Добавление НАДФ повторялось дважды. Ход реакции контролировался путем измерения абсорбции на 340 нм, благодаря чему можно было наблюдать уменьшение НАДФ. НАДФ при этом добавляли по 300 мкл, поскольку высокая концентрация НАДФ ведет к дезактивации энзима. Затем для выделения продукта реакционный раствор был экстрагирован 3 раза по 5 мл диэтилэфира. Объединенные органические фазы высушивались с помощью MgSO4 и собирались. Затем продукты были проанализированы методами тонкослойной хроматографии (DC), газовой хроматографии с масс-спектроскопическим окончанием (GC/MS) и ЯМР.
GC/MS - анализ реакционной смеси дал следующие результаты:
Продукт извлечения не был найден.
Пример 7
Гидроксилирование ароматических углеводородов, гетероароматических углеводородов, соединений триметилциклогексенила
а) Пример 6 был повторен, однако при этом вместо н-октана в качестве субстрата был взят нафталин. Продуктами оказались 1-нафтанол и цис-1,2-дигидрокси-1,2-дигидронафталин. Введенный нафталин преобразовывался на 88%.
Анализ реакций нафталина
GC:
Аппаратура: Carlo Erba Strumentazion тип HRGC 4160 на колоночном инжекторе; колонка: DB5 30 м × 0,2 мм; материал: 5% дифенил - 95% диметилполисилоксан; несущий газ: 0,5 бар Н2; температурная программа: 40°С 1 мин изотерм/10°С/мин до 300°С; Rt (1-нафтанол)=16,68.
ЯМР:
Были идентифицированы 1-нафтанол и цис-1,2-дигидрокси-1,2-дигидронафталин.
b) Пример 6 был повторен, однако при этом вместо н-октана в качестве субстрата был взят 8-метилхинолин. В качестве главного продукта был идентифицирован 5-гидрокси-8-метилхинолин, кроме других производных (соотношение продукта 5:1). Введенный продукт преобразовывался на 35%.
c) Пример 6 был повторен, однако при этом вместо н-октана в качестве субстрата был взят α-ионон. В качестве главного продукта был идентифицирован 3-гидрокси-α-ионон, кроме других производных (соотношение продукта 76:24). Введенный продукт преобразовывался на 60%.
d) Пример 6 был повторен, однако при этом вместо н-октана в качестве субстрата был взят кумол (изо-пропилбензол). Были идентифицированы 5 продуктов моногидроокиси и один продукт дигидроокиси. Введенный продукт преобразовывался на 70%.
Литература
1. Yano, Т., Oue, S. и Kagamiyama, H. (1998) Proc. Natl. Acad. Sci. USA 95, 5511-5515.
2. Zhang, J.-H., Dawes, G. и Stenner, W.P.C. (1997) Proc. Natl. Acad. Sci. USA 94, 4504-4509.
3. Wan, L., Twitchett, M.B., Eitis, L.D., Mauk, А.G. и Smith, M. (1998) Proc. Natl. Acad. Sci USA 95, 12825-12831.
4. Cronin, C.N. (1998) J. Biol. Chem. 273, 24465-24469.
5. Wilks, H.M., Hart, К-W., Feeney, R., Dunn, C.R., Muirhead, H., Chia, W.N., Barstow, D.A., Atkinson, Т., Clarke, A.R., Holbrook, I.J. (1988) Science 242, 1541-1544.
6. Hedstrom, L., Szilagyi, L., Rutter, W.J. (1992) Science 255, 1249-1253.
7. Tucker, C.L., Hurley, J.H., Miller, T.R. и Hurley, I.B. (1998) Proc. Natl. Acad. Sci. USA 95, 5993-5997.
8. Quemeneur, E., Moutiez, J.-B.C. и Menez, A. (1998) Mature (London) 391, 301-303.
9. Marsden, A-F.A., Wilkinson, В., Cortes, J., Dunster, N.J., Staunton, I Leadlay, P.F. (1998) Science 279, 199-201.
10. Chen, R., Greer, А., и Dean, A.M. (1998) Proc. Natl. Acad. Sci. USA 95, 11666-11670.
11. Boddupalli, S.S., Estabrook, R.W. и Peterson, J.A. (1990) J. Biol. Chem. 265, 4233-4239.
12. Capdevila, J.H., Wie, S., Helvig, C., Faick, J.R., Belosludtsev, Y., Truan, G., Graham-Lorence, S.E. и Peterson, J.A. (1996) J. Biol. Chem. 271, 22663-22671.
13. Graham-Lorence, S., Truan, G., Peterson, J.A., Flack, J.R., Wel S., Helvig, C., Capdevilla, J.H. (1997) J. Biol. Chem. 272, 1127-1135.
14. Li, H, Poulos, T.L. (1997) Nat. Structural Biol., 4, 45 140-146.
15. Ravichandran, K.G., Sekhar, S., Boddupalli, S., Hasemann, C.A., Peterson, J.A., Deisenhofer, 1 (1993) Science 261, 731-736.
16. Modi S., Sutcliffe, M.J, Primrose, W.U., Lian, L.-Y., Roberts, G.C.K (1996) Nat. Structural Biol. 3, 414-417.
17. Oliver, C.F., Modl S., Primrose, W.U., Lian, L.Y. и Roberts, G.C. K (1997) Bio-chem. J 327, 537-544.
18. Guengerich, F.G. (1991) J. Biol. Chem. 266, 10019-10022.
19. Cherry, J.R., Lamsa, M.H., Schneider, P., Vind, J., Svendsen, A., Jones, А. и Pedersen, A.H. (1999) Nature Biotechnology 17, 379-384.
20. Schwaneberg, U., Schmidt-Dannert, C., Schmitt, J. и Schmid, R.D. (1999) Anal. Biochem. 269, 359-366.
21. Schwaneberg, U., Sprauer, A.L., Schmidt-Dannert, С. и Schmid, R.D. J of Chromatogr. A, in press.
22. Oliver, C.F., Modi, S., Sutcliffe, M.J., Primrose, W.U., Lian, L.Y. и Roberts, G.C.K (1997) Biochemistry 36, 25 1567-1572.
23. Hart, S., Koch. K.R. и Woods, D.R. (1992) J. Gen. Microbiol. 138, 211-216.
24. Murdock, D., Ensley, B.D., Serdar, С. и Thalen, M. (1993) Bio/Technology 11, 381-385.
25. O'Connor, ICE., Dobson, A-W. и Hartmans, S. (1997) Appl. Environ. Microbiol. 63, 4287-4291.
26. Eaton, R.W. и Chapman, P.J. (1995) J. Bacteriol. 177, 6983-6988.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЙ ЦИТОХРОМ Р-450 МОНООКСИГЕНАЗЫ (ВАРИАНТЫ), СПОСОБ ОКИСЛЕНИЯ СОЕДИНЕНИЯ, ВЫБИРАЕМОГО ИЗ ГРУППЫ, СОДЕРЖАЩЕЙ ПОЛИЦИКЛИЧЕСКИЙ АРОМАТИЧЕСКИЙ УГЛЕВОДОРОД, ЛИНЕЙНЫЙ ИЛИ РАЗВЕТВЛЕННЫЙ АЛКАН И СОЕДИНЕНИЕ ДИФЕНИЛА И БИФЕНИЛА, ВКЛЮЧАЯ ГАЛОГЕНПРОИЗВОДНЫЕ И ГАЛОГЕНУГЛЕВОДОРОДЫ | 1995 |
|
RU2133774C1 |
| (ЦИТОХРОМ Р450)-МОНООКСИГЕНАЗЫ | 1998 |
|
RU2237714C2 |
| ЦИТОХРОМ P450 И ЕГО ПРИМЕНЕНИЕ ДЛЯ ЭНЗИМАТИЧЕСКОГО ОКИСЛЕНИЯ ТЕРПЕНОВ | 2012 |
|
RU2625224C2 |
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКИХ СПИРТОВ | 2014 |
|
RU2654670C2 |
| МУТАНТНАЯ ГЛЮКОЗОИЗОМЕРАЗА С ИЗМЕНЕННОЙ СУБСТРАТНОЙ СПЕЦИФИЧНОСТЬЮ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 1991 |
|
RU2096457C1 |
| ПРЯМАЯ ЭЛЕКТРОХИМИЯ ФЕРМЕНТОВ | 1997 |
|
RU2157521C2 |
| СПОСОБ БИОКАТАЛИТИЧЕСКОГО ОКИСЛЕНИЯ С ПРИМЕНЕНИЕМ ПРОДУКТА ГЕНА alkL | 2011 |
|
RU2573900C2 |
| МОДИФИЦИРОВАННАЯ АЦИЛ-АСР ДЕСАТУРАЗА | 1997 |
|
RU2228356C2 |
| НОВЫЙ ПОЛИПЕПТИД СИСТЕМЫ ЦИТОХРОМА Р450 С ПОВЫШЕННОЙ ФЕРМЕНТНОЙ АКТИВНОСТЬЮ | 2015 |
|
RU2677997C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ, ПОЛОЖИТЕЛЬНОЙ ПО МУТАЦИИ FLT3, ИНГИБИТОР МУТАНТНОГО FLT3 И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2727195C2 |













Изобретение относится к биотехнологии. Предложен способ микробиологического окисления N-, О- или S-гетероциклических одно- или многоядерных ароматических соединений, включающий культивирование рекомбинантного микроорганизма, который экспримирует цитохром Р450-монооксигеназу ВМ-3 из Bacillus megaferium с аминокислотной последовательностью, представленной в SEQ ID NO:2, которая имеет по меньшей мере одну функциональную мутацию в области 86-88 и, при необходимости, по меньшей мере, одну функциональную мутацию в одной из областей 73-82, 172-224, 39-43, 48-52, 67-170, 300-335 и 352-356. Полученный продукт окисления выделяют из среды. Заявленное изобретение позволяет проводить окисление органических соединений с высокой степенью эффективности. 4 н. и 13 з.п. ф-лы, 1 табл.
а1) культивируют рекомбинантный микроорганизм, который экспримирует цитохром Р450-монооксигеназу бактериального происхождения в культуральной среде в присутствии экзогенного или промежуточно образующегося субстрата; или
а2) инкубируют содержащую субстрат реакционную среду с цитохром Р450-монооксигеназой бактериального происхождения;
b) выделяют образовавшийся продукт окисления или его производный продукт из среды,
причем монооксигеназа является производной от цитохром Р450-монооксигеназы ВМ-3 из Bacillus megaterium с аминокислотной последовательностью согласно SEQ ID NO:2, которая имеет, по меньшей мере, одну функциональную мутацию в области аминокислотной последовательности 86-88 и, при необходимости, дополнительно имеет, по меньшей мере, одну функциональную мутацию в одной из областей аминокислотной последовательности 73-82, 172-224, 39-43, 48-52, 67-70, 330-335 и 352-356.
1) Phe87Val;
2) Phe87Val, Leu188Gln; или
3) Phe87Val, Leu188Gln; Ala74Gly.
а1) культивируют рекомбинантный производящий цитохром Р450-монооксигеназу микроорганизм в культуральной среде в присутствии экзогенного, или промежуточного образующегося субстрата; или
а2) инкубируют содержащую субстрат реакционную среду с цитохром Р450-монооксигеназой, способной к окислению вышеупомянутых соединений; и
b) выделяют образовавшийся продукт окисления или его производный продукт из среды,
причем монооксигеназа является способной к окислению вышеупомянутых соединений и является производной от цитохром Р450-монооксигеназы ВМ-3 из Bacillus megaterium с аминокислотной последовательностью согласно SEQ ID NO:2, которая имеет, по меньшей мере, одну функциональную мутацию в области аминокислотной последовательности 86-88 и, при необходимости, дополнительно имеет, по меньшей мере, одну функциональную мутацию в одной из областей аминокислотной последовательности 73-82, 172-224, 39-43, 48-52, 67-70, 330-335 и 352-356.
1) Phe87Val;
2) Phe87Val, Leu188Gln; или
3) Phe87Val, Leu188Gln; Ala74Gly.
1) Phe87Val;
2) Phe87Val, Leu188Gln; или
3) Phe87Val, Leu188Gln; Ala74Gly.
причем монооксигеназа является производной от цитохром Р450-монооксигеназы ВМ-3 из Bacillus megaterium с аминокислотной последовательностью согласно SEQ ID NO:2, которая имеет, по меньшей мере, одну функциональную мутацию в области аминокислотной последовательности 86-88 и, по меньшей мере, одну функциональную мутацию в одной из областей аминокислотной последовательности 73-82 и 172-224 и, при необходимости, дополнительно имеет одну функциональную мутацию в одной из областей аминокислотной последовательности 39-43, 48-52, 67-70, 330-335 и 352-356.
a) Phe87Val, Leu188Gln; или
b) Phe87Val, Leu188Gln; Ala74Gly,
а также их функциональные эквиваленты, которые способны, как минимум, к одной из вышеуказанных реакций окисления.
Приоритет по пунктам:
п.16 формулы изобретения в части Цитохром Р450-монооксигеназы, которая способна, как минимум, к одной из реакций по подпунктам a)-d) п.15 формулы, кроме окисления О-гетероциклических ароматических соединений, и которая имеет функциональную мутацию в позиции аминокислотной последовательности 87 и, по меньшей мере, одну функциональную мутацию в одной из позиций аминокислотной последовательности 74 и 188;
п.17 формулы изобретения в части Цитохром Р450-монооксигеназы согласно подпункту а) или b), которая способна, как минимум, к одной из реакций по подпунктам a)-d) пункта 15 формулы, кроме окисления O-гетероциклических ароматических соединений.
пп.11-14 формулы изобретения;
п.15 формулы изобретения в части Цитохром Р450-монооксигеназы, которая способна к окислению N-гетероциклических двух- или многоядерных ароматических соединений по подпункту а), за исключением Цитохром Р450-монооксигеназы, которая имеет функциональную мутацию в позиции аминокислотной последовательности 87 и, по меньшей мере, одну функциональную мутацию в одной из позиций аминокислотной последовательности 74 и 188 и, при необходимости, дополнительно имеет функциональную мутацию в позиции аминокислотной последовательности 354;
п.16 формулы изобретения в части Цитохром Р450-монооксигеназы, которая способна, к окислению N-гетероциклических двух- или многоядерных ароматических соединений по подпункту а) п.15 формулы, за исключением Цитохром Р450-монооксигеназы, которая имеет функциональную мутацию в позиции аминокислотной последовательности 87 и, по меньшей мере, одну функциональную мутацию в одной из позиций аминокислотной последовательности 74 и 188.
пп.6-10 формулы изобретения;
п.15 формулы изобретения в части Цитохром Р450-монооксигеназы, которая способна, как минимум, к одной из реакций по подпунктам a)-d), за исключением Цитохром Р450-монооксигеназы, которая способна, как минимум, к одной из реакций по подпунктам а)-d), кроме окисления О-гетероциклических ароматических соединений, и которая имеет функциональную мутацию в позиции аминокислотной последовательности 87 и, по меньшей мере, одну функциональную мутацию в одной из позиций аминокислотной последовательности 74 и 188 и, при необходимости, дополнительно имеет функциональную мутацию в позиции аминокислотной последовательности 354 и за исключением Цитохром Р450-монооксигеназы, которая способна, к окислению N-гетероциклических двух- или многоядерных ароматических соединений по подпункту а);
п.16 формулы изобретения в части Цитохром Р450-монооксигеназы, которая способна, как минимум, к одной из реакций по подпунктам a)-d) п.15 формулы, за исключением Цитохром Р450-монооксигеназы, которая способна, как минимум, к одной из реакций по подпунктам а)-d) п.15 формулы, кроме окисления О-гетероциклических ароматических соединений, и которая имеет функциональную мутацию в позиции аминокислотной последовательности 87 и, по меньшей мере, одну функциональную мутацию в одной из позиций аминокислотной последовательности 74 и 188 и за исключением Цитохром Р450-монооксигеназы, которая способна, к окислению N-гетероциклических двух- или многоядерных ароматических соединений по подпункту а);
п.17 формулы изобретения в части Цитохром Р450-монооксигеназы согласно подпункту а) или b), которая способна, к окислению О-гетероциклических ароматических соединений.
| СПОСОБ ОПРЕДЕЛЕНИЯ БИОЛОГИЧЕСКОГО ВОЗРАСТА ЧЕЛОВЕКА | 2005 |
|
RU2294692C1 |
| ЗОЛОУЛОВИТЕЛЬ | 2006 |
|
RU2306485C1 |
| DE 19507546, 12.09.1996 | |||
| МОДИФИЦИРОВАННЫЙ ЦИТОХРОМ Р-450 МОНООКСИГЕНАЗЫ (ВАРИАНТЫ), СПОСОБ ОКИСЛЕНИЯ СОЕДИНЕНИЯ, ВЫБИРАЕМОГО ИЗ ГРУППЫ, СОДЕРЖАЩЕЙ ПОЛИЦИКЛИЧЕСКИЙ АРОМАТИЧЕСКИЙ УГЛЕВОДОРОД, ЛИНЕЙНЫЙ ИЛИ РАЗВЕТВЛЕННЫЙ АЛКАН И СОЕДИНЕНИЕ ДИФЕНИЛА И БИФЕНИЛА, ВКЛЮЧАЯ ГАЛОГЕНПРОИЗВОДНЫЕ И ГАЛОГЕНУГЛЕВОДОРОДЫ | 1995 |
|
RU2133774C1 |

