Настоящее изобретение относится к генетическому конструированию растений с помощью метода рекомбинантной ДНК и к ферментам, участвующим в биосинтезе цианогенных гликозидов, в частности, к генам, кодирующим эти ферменты. Протеины и гены по изобретению могут применяться для увеличения питательной ценности растений или их устойчивости к вредителям.
Цианогенные гликозиды представляют собой вторичные растительные метаболиты у более чем 2000 видов растений. В некоторых случаях они являются источником HCN, что может приводить к токсичности растений при их употреблении в пищу. Например, клубни цианогенного культурного растения маниока съедобного (Manihot esculenta) являются одним из главных продуктов питания в тропических зонах. Присутствующие в клубнях цианогенные гликозиды могут вызвать отравление цианидом у людей при недостаточной обработке продуктов из маниока. Другие виды растений, в которых ферментативным путем продуцируется HCN и которые обладают потенциальной токсичностью при избыточном потреблении в качестве пищи или при применении в качестве корма для скота, включают белый клевер (Trifolium repens), сорго (Sorghum bicolor), лен обыкновенный (Linum usitatissimum), триостренник (Triglochin maritima), фасоль лима (Phaseolus lunatus), миндаль (Amygdalus) и косточки абрикоса (р. Primus), вишен и яблок (р. Malus). Токсичность этих растений может быть уменьшена путем ингибирования биосинтеза цианогенных гликозидов в этих растениях.
Первичные предшественники встречающихся в естественных условиях цианогенных гликозидов ограничены пятью гидрофобными входящими в состав протеинов аминокислотами, такими, как валин, лейцин, изолейцин, фенилаланин и тирозин, а также одной не входящей в состав протеинов аминокислотой циклопентенглицином. Эти аминокислоты превращаются в результате серий реакций в циангидрины, которые в конечном итоге связываются с остатком сахара. Например, амигдалин содержит O-β-гентиобиозид и пруназин, т.е. O-β-гликозид (R)-манделонитрила. Другим примером цианогенных гликозидов, имеющих ароматические агликоны, является эпимерная пара цианогенных гликозидов дхуррин и таксифиллин, которые обнаружены в растениях родов Sorghum и Taxus соответственно. Например, пара-гидроксиманделонитрил превращается в дхуррин (β-D-глюкопиранозилокси-(S)-пара-гидроксиманделонитрил) с помощью УДФГ-гликозилтрансферазы. Вероятно, аналогичные гликозилтрансферазы присутствуют в большинстве растений. Вицианин и лукумин представляют собой дополнительные примеры производных дисахаридов, аналогичных амигдалину. Самбунигрин содержит (S)-мaндeлoнитpил в качестве агликона и, следовательно, представляет собой эпимер пруназина. Примерами цианогенных гликозидов, имеющих алифатические агликоны, являются линамарин и лотаустралин, обнаруженные в клевере, льне, маниоке и фасоли. Подробный обзор цианогенных гликозидов и путей их биосинтеза приведен в работе Conn, Naturwissenschaften 66: 28-34, 1979, которая включена в настоящее описание в качестве ссылки.
Путь биосинтеза цианогенного глюкозида дхуррина, являющегося производным тирозина, хорошо изучен (Halkier и др., "Cyanogenic glucosides: the biosynthetic pathway and the enzyme system" в "Cyanogenic compounds in biology", Wiley Chichester (Ciba Foundation Symposium 140), стр. 49-66, 1988; Halkier и Moller, Plant Physiol. 90: 1552-1559, 1989; Halkier и др., The J. of Biol. Chem. 264: 19487-19494, 1989; Halkier и Moller, Plant Physiol. 96: 10-17, 1990; Halkier и Moller, The J. of Biol. Chem. 265: 21114-21121, 1990; Halkier и др., Proc. Natl. Acad. Sci. USA 88: 487-491, 1991; Sibbesen и др., "Biochemistry and Biophysics of cytochrom P450. Structure and Function, Biotechnological and Ecological Aspects", под ред. Archakov A.I., 1991; Koch и др., Материалы 8-й Международной конференции по цитохрому P450 (8th Int. Conf. on Cytochrome P450), реферат PII.053; Sibbesen и др., Материалы 8-й Международной конференции по цитохрому P450 (8th Int. Conf. on Cytochrome P450), реферат PII.016). L-тирозин превращается в пара-гидроксиманделонитрил (предшественник дхуррина), при этом N-гидрокситирозин, N,N-дигидрокситирозин, (Е)- и (Z)-пара-гидроксифенилацетальдегидоксим и пара-гидроксифенилацетонитрил являются промежуточными продуктами. В этот путь метаболизма включены два типа (цитохром Р450)-монооксигеназ. У маниока аналогичный путь метаболизма, включающий P450-зависимые монооксигеназы, используется для синтеза линамарина и лотаустралина из валина и изолейцина соответственно (Koch и др., Archives of Biochemistry and Biophysics, 292: 141-150, 1992). Как установлено для Sorghum bicolor, для сложного пути превращения L-тирозина в пара-гидроксиманделонитрил необходимы только две многофункциональные P450-зависимые монооксигеназы. Первый фермент, обозначенный как P450TYR, превращает тирозин в пара-гидроксифенилацетальдегидоксим. Второй фермент, обозначенный как Р450ox, превращает альдоксим в пара-гидроксиманделонитрил. С точки зрения сходства путей биосинтеза цианогенных глюкозидов в различных растениях, как правило, считается, что указанные пути метаболизма включают две многофункциональные P450-зависимые монооксигеназы Р450I и Р450II, которые превращают аминокислоту, являющуюся предшественником, в соответствующий альдоксим, а альдоксим в соответствующий циангидрин соответственно. Р450I представляет собой определенный фермент, который обладает субстратной специфичностью и вследствие этого участвует в пути метаболизма гликозида определенного типа, в то время как Р450II, вероятно, является менее специфичным и осуществляет превращение широкого диапазона различных по строению альдоксимов в соответствующий циангидрин.
Глюкозинолаты представляют собой гидрофильные нелетучие тиогликозиды, обнаруженные в нескольких отрядах двудольных покрытосеменных растений (Cronquist, "The Evolution and Classification of Flowering Plants", New York Botanical Garden, Bronx, 1988). Встречаемость цианогенных глюкозинолатов и глюкозидов является взаимно исключающей. Наибольшее экономическое значение имеет наличие глюкозинолатов во всех представителях сем. Brassicaceae (каперсовые, порядок Capparales), многие сорта которых в течение многих столетий являются для человечества источником приправ, закусок, салатных культур и овощей, а также фуражных и кормовых культур. В настоящее время рапс (особенно Brassica napus и Brassica campestris) стал основной коммерческой масличной культурой. Известно примерно 100 различных глюкозинолатов, имеющих одинаковое общее строение, но отличающихся природой боковой цепи. Глюкозинолаты образуются из входящих в состав протеинов аминокислот либо непосредственно, либо после одного или после многократного удлинения цепи (Underbill и др., Biochem. Soc. Symp. 38: 303-326, 1973). N-гидроксиаминокислоты и альдоксимы, которые были выявлены в качестве промежуточных продуктов биосинтеза цианогенных гликозидов, также служат в качестве эффективных предшественников биосинтеза глюкозинолатов (Kindl и др., Phytochemistry 7: 745-756, 1968; Matsuo и др., Phytochemistry 11: 697-701, 1972; Underbill Eur. J. Biochem. 2: 61-63, 1967). Таким образом, цитохром Р450I, который участвует в синтезе цианогенных гликозидов, функционирует практически так же, как и соответствующий фермент биосинтеза глюкозинолатов, и, вероятно, является представителем этого же семейства Р450-ферментов. Так, авторами изобретения был выделен клон кДНК из Sinapis alba (горчица белая), кодирующий фермент Р450 (SEQ ID NO: 17), который на 54% идентичен Р450TYR (CYP79) и который катализирует первую стадию биосинтеза глюкозинолатов, в процессе которой из исходной аминокислоты образуется альдоксим. Этот клон кДНК оказался примерно на 90% идентичен последовательности EST Arabidopsis (T42902), что с высокой долей вероятности свидетельствует о том, что фермент цитохром Р450 является высоко консервативным у видов, содержащих глюкозинолаты.
Сведение описанного выше сложного пути биосинтеза циангидринов к каталитической активности только двух ферментов, цитохромов P450I и Р450II, позволяет управлять путем биосинтеза цианогенных глюкозидов в растениях. Путем трансфекции генных конструкций, кодирующих одну или обе монооксигеназы, путь биосинтеза цианогенных глюкозидов может быть либо модифицирован, либо восстановлен, либо вновь создан.
Модификация или интродукция пути биосинтеза цианогенных гликозидов в растениях известными в данной области методами представляет большой интерес, поскольку цианогенные гликозиды могут обладать токсичностью для насекомых, клещей и нематод. Таким образом, модификация, интродукция или восстановление пути биосинтеза цианогенных гликозидов в растениях или в определенных тканях растения должны сделать растения несъедобными для насекомых, клещей или нематод и, следовательно, способствовать уменьшению повреждения культурного растения вредителями. В комбинации с другими инсектицидными агентами, такими, как эндотоксины Bacillus thurigiensis, повреждение культурного растения вредителями может быть дополнительно уменьшено.
В альтернативном варианте последовательности генов, кодирующих монооксигеназы по изобретению, могут применяться для создания плазмидных ДНК, которые при трансфекции в растение, содержащее цианогенные гликозиды, такое, как маниок, сорго или ячмень, устраняют цианогенные гликозиды, обычно продуцируемые растениями дикого типа. Реализовать это можно путем экспрессии антисмысловой либо смысловой РНК или рибозимов согласно методу, описанному в ЕР-458367-А1, ЕР-240208-А2, US 5231020, WO 89/05852 и WO 90/11682, что позволяет ингибировать экспрессию монооксигеназ по изобретению. Разработка такого подхода представляет большой интерес, поскольку несмотря на многочисленные попытки методами традиционной селекции растений не удалось полностью удалить цианогенные гликозиды, например, из маниока или сорго. С другой стороны, установлено, что увеличенные количества цианогенных гликозидов в эпидермальных клетках разных культиваров ячменя обусловливают повышенную чувствительность к грибам Erysiphe graminis, являющимся возбудителями мучнистой росы (Pourmohensi, PhD thesis,  1989; Ibenthal и др., Angew. Bot. 67: 97-106, 1993). Аналогичное действие было обнаружено для цианогенного каучуконосного дерева гевеи (Hevea brasiliensis) в отношении поражения грибом Microcyclus ulei (Lieberei и др., Plant Phys. 90: 3-36, 1993) и для льна в отношении поражения грибом Colletotrichum lini (Ludtke и др., Biochem. Z. 324: 433-442, 1953). В этих примерах уровень устойчивости указанных выше растений и других растений, у которых цианогенные гликозиды обусловливают повышенную чувствительность к поражению микроорганизмами, может быть увеличен путем предупреждения образования цианогенных гликозидов в таких растениях. У ячменя цианогенные гликозиды локализованы в эпидермальных клетках. Таким образом, экспрессия антисмысловых, смысловых или рибозимных конструкций предпочтительно, но не обязательно, находится под контролем специфичного для эпидермиса промотора.
1989; Ibenthal и др., Angew. Bot. 67: 97-106, 1993). Аналогичное действие было обнаружено для цианогенного каучуконосного дерева гевеи (Hevea brasiliensis) в отношении поражения грибом Microcyclus ulei (Lieberei и др., Plant Phys. 90: 3-36, 1993) и для льна в отношении поражения грибом Colletotrichum lini (Ludtke и др., Biochem. Z. 324: 433-442, 1953). В этих примерах уровень устойчивости указанных выше растений и других растений, у которых цианогенные гликозиды обусловливают повышенную чувствительность к поражению микроорганизмами, может быть увеличен путем предупреждения образования цианогенных гликозидов в таких растениях. У ячменя цианогенные гликозиды локализованы в эпидермальных клетках. Таким образом, экспрессия антисмысловых, смысловых или рибозимных конструкций предпочтительно, но не обязательно, находится под контролем специфичного для эпидермиса промотора.
Присутствие даже минорных количеств цианогенных гликозидов в растениях также может создавать проблемы, связанные с их питательной ценностью, из-за образования нежелательных канцерогенов, что известно для ячменя. Ячменный солод, например, содержит небольшие количества цианогенного глюкозида эпигетеродендрина, который в процессе получения спиртов из зерна может превращаться в этилкарбамат, который считается канцерогеном. Вследствие этого делаются попытки ввести обязательное нормирование предельно допустимых концентраций этилкарбамата в сбраживаемом корме, напитках и спиртах (Food Chemicals News 29: 33.35, 1988).
В WO 95/16041 предложена молекула ДНК, кодирующая (цитохром Р450I)-монооксигеназу, которая катализирует превращение аминокислоты в соответствующую N-гидроксиаминокислоту, N,N-дигидроксиаминокислоту и превращение N,N-дигидроксиаминокислоты в соответствующий альдоксим. Аминокислоту-предшественника выбирают из группы, включающей тирозин, фенилаланин, триптофан, валин, лейцин, изолейцин и циклопентенилглицин.
Молекулы ДНК либо соответствуют встречающимся в естественных условиях генам, либо их функциональным гомологам, которые являются результатом мутации, делеции, усечения и т.д., но еще сохраняют способность кодировать (цитохром Р450I)-монооксигеназу, способную катализировать более одной реакции пути биосинтеза цианогенных гликозидов. Монооксигеназы предпочтительно содержат один каталитический центр.
Кроме того, в WO 95/16041 предложены молекулы ДНК, кодирующие (цитохром Р450II)-монооксигеназы, такие, как Р450ox растения Sorghum bicolor (L.) Moench. Они катализируют превращение альдоксима в нитрил и превращение нитрила в соответствующий циангидрин. Катализ превращения тирозина в пара-гидроксифенилацетонитрил двумя многофункциональными ферментами Р450 объясняет, почему путь метаболизма всех промежуточных продуктов, участвующих в этом превращении, кроме (Z)-пара-гидроксифенила-цетальдоксима, происходит в определенном направлении.
Предлагаемая для выделения Р450 стратегия основана на применении методик, предложенных для выделения Р450TYR (CYP79, Sibbesen и др., Proc. Natl. Acad. Sci. USA 91: 9740-9744, 1994) из сорго. При использовании этого метода ионообменную колонку с ионообменником типа сефарозы с диэтиламиноэтилом (ДЭАЭ) применяют для связывания ферментов Р450, при этом желтые пигменты в образце не связываются. Удаление пигментов служит двум целям. Это является необходимым условием для связывания ферментов Р450 со следующими колонками и позволяет определить содержание Р450 спектрометрически (с использованием окиси углерода (СО) и связывания с субстратом). В отличие от этого согласно настоящему изобретению Р450ox обладает низким сродством к связыванию с ДЭАЭ-колонкой и практически полностью извлекается из фракций погона и промывочных фракций. С целью разделения активности Р450ox и желтых пигментов применяют процедуру распределения фаз с использованием Тритона Х-114. Установлено, что при использовании предпочтительно от 0,6 до 1% Тритона Х-114 Р450ox распределяется в обеих фазах, в отличие от Р450TYR, который переходит в верхнюю фазу с высоким содержанием детергента. При увеличении концентрации Тритона Х-114 до 6% основная часть Р450ox извлекается из нижней фазы с низким содержанием детергента, в то время как желтые пигменты находятся в верхней фазе. Недостатком применения 6%-ного Тритона Х-114 является повышение превращения Р450ox в его денатурированную форму Р420. Эти данные используют в настоящем изобретении прежде всего для очистки Р450II-монооксигеназ, таких, как Р450ox, с целью клонирования генов, кодирующих монооксигеназы, и для стабильной трансформации растений генами, кодирующими монооксигеназы. Выделение Р450ox и определение частей аминокислотных последовательностей способствует созданию олигонуклеотидных зондов и выделению кДНК, кодирующей Р450. Однако в этом случае клонирование проводят с использованием независимого подхода.
Изобретение прежде всего относится к молекулам ДНК, кодирующим (цитохром Р450II)-монооксигеназы, которые катализируют превращение альдоксима в нитрил и превращение этого нитрила в соответствующий циангидрин. Предпочтительно альдоксим представляет собой продукт превращения аминокислоты, выбранной из группы, включающей тирозин, фенилаланин, триптофан, валин, лейцин, изолейцин и циклопентенилглицин, или аминокислоты, выбранной из группы, включающей L-тирозин, L-валин и L-изолейцин, катализируемого Р450I-монооксигеназой, как описано в WO 95/16041. Молекулы ДНК по изобретению либо соответствуют встречающимся в естественных условиях генам, либо их гомологам, полученным в результате мутации, делеции, усечения и т.д., но еще сохраняющим способность кодировать (цитохром Р450II)-монооксигеназу, которая катализирует превращение альдоксима в нитрил и последующее превращение этого нитрила в соответствующий циангидрин. Монооксигеназы по изобретению катализируют более одной реакции пути биосинтеза цианогенных гликозидов и предпочтительно содержат один каталитический центр.
Ферменты системы цитохрома Р450II могут присутствовать в большинстве живых организмов. Молекулы ДНК по настоящему изобретению, кодирующие Р450II-монооксигеназы, структурно и функционально подобны молекулам ДНК, которые могут быть получены из различных растений, продуцирующих цианогенные гликозиды. В предпочтительном варианте осуществления изобретения молекулы ДНК гибридизуются с фрагментом молекулы ДНК, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1. Этот фрагмент имеет длину более 10 нуклеотидов и предпочтительно более 15, 20, 25, 30 или 50 нуклеотидов. Факторы, которые влияют на стабильность гибридов, определяют строгость условий гибридизации и могут быть определены на основе данных о температуре плавления (tпл) получаемых гибридов. Метод расчета tпл описан в нескольких руководствах. Например, у Keller и др. в "DNA Probes: Background, Applications, Procedures", Macmillan Publishers Ltd, 1993, на стр. 8-10 описаны факторы, которые следует учитывать при расчете значений tпл для реакций гибридизации. Молекулы ДНК по настоящему изобретению гибридизуются с фрагментом, имеющим последовательность, представленную в SEQ ID NO:1, при температуре на 30°С ниже рассчитанного значения tпл гибрида, который должен быть получен. Предпочтительно они гибридизуются при температурах на 25, 20, 15, 10 или 5°С ниже рассчитанного значения tпл.
Для целей манипуляции с генами с помощью метода рекомбинантной ДНК молекула ДНК по изобретению может в дополнение к гену, кодирующему монооксигеназу, включать ДНК, которая позволяет, например, осуществлять репликацию и отбор ДНК по изобретению в микроорганизмах, таких, как Е. Coli, Bacillus, Agrobacterium, Streptomyces или дрожжи. Она также может включать ДНК, которая позволяет осуществлять экспрессию и отбор генов монооксигеназ в гомологичных или в гетерологичных растениях. Такие последовательности включают (но не ограничены ими) гены, наиболее часто встречающиеся кодоны которых адаптированы к наиболее часто встречающемуся кодону гетерологичного растения, как описано в WO 93/07278, гены, обусловливающие устойчивость к неомицину, канамицину, метотрексату, гигромицину, блеомицину, стрептомицину или гентамицину, к аминоэтилцистеину, глиофосфату, сульфонилмочевинам или фосфинотрицину, пригодные для осуществления скрининга гены-маркеры, такие, как галактозидаза, их природные промоторы и сигналы терминации транскрипции, элементы промоторов, таких, как промоторы 35S и 19S CaMV, или тканеспецифичные растительные промоторы, такие, как промоторы, специфичные для корня (например, описанные в ЕР-452269-А2, WO 91/13992, US 5023179), специфичные для зеленых листьев, такие, как фосфоенолпируваткарбоксилаза (ФЕПК) кукурузы, для сердцевины или пыльцы (например, описанные в WO 93/07278), или индуцибельные растительные промоторы (ЕР-332104), и гетерологичные сигналы терминации транскрипции.
Настоящее изобретение также относится к Р450II-монооксигеназам, которые катализируют превращение альдоксима в нитрил и превращение этого нитрила в соответствующий циангидрин. В предпочтительном варианте осуществления изобретения монооксигеназы очищают, и они могут использоваться для создания моноклональных или поликлональных антител, которые специфично связываются с монооксигеназами. В частности, цитохром Р450ox, имеющий молекулярную массу 55 кДа, что установлено с помощью электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (ДСН-ПААГ), выделяют из Sorghum bicolor (L.) Moench. Его аминокислотная последовательность приведена в SEQ ID NO: 2.
Каталитические особенности Р450ox сходны с активностью цитохрома Р450, выявленной в микросомах печени крысы (DeMaster и др., J.Org.Chem. 5074-5075, 1992). Характерной особенностью цитохрома Р450ox и других представителей, принадлежащих к семейству цитохромов Р450ox, является то, что дегидратация альдоксима с получением соответствующего нитрила зависит от присутствия восстановленного никотинамид-аденин-динуклеотидфосфата (NADPH), но эта зависимость в некоторых случаях может быть преодолена добавлением дитионита натрия или других восстановителей.
Установлено, что из всех известных последовательностей ферментов системы цитохрома Р450 аминокислотная последовательность цитохрома Р450ox наиболее высоко идентична (44%) аминокислотной последовательности фермента CYP71A1 авокадо, а ее идентичность с аминокислотной последовательностью всех остальных представителей семейства CYP71 составляет менее 40%. Растения авокадо не продуцируют цианогенные гликозиды, а CYP71A1 не катализирует превращение альдоксима в нитрил и превращение этого нитрила в соответствующий циангидрин. Таким образом, согласно настоящему изобретению семейство (цитохром Р450II)-монооксигеназ может быть ограничено представителями, которые катализируют превращение альдоксима в соответствующий циангидрин и аминокислотные последовательности которых на 40% или более идентичны аминокислотной последовательности цитохрома Р450ox. Предпочтительно аминокислотная последовательность идентична аминокислотной последовательности цитохрома Р450^ более чем на 50% или более чем на 55%.
Можно предположить, что Р450ox является первым представителем нового подсемейства CYP71 (CYP71E1), поскольку он входит в один кластер с другими последовательностями CYP71 на дендрограммах, представляющих собой графическое изображение первичной структуры множества последовательностей. В целом, согласно положению Комитета по номенклатуре для того, чтобы цитохром Р450 был отнесен к новому семейству CYP, необходимо, чтобы на уровне аминокислотной последовательности идентичность составляла менее 40%, а при идентичности более чем 55% его относят к этому же подсемейству. При проведении сравнительного анализа множества последовательностей рассматривают не только идентичность последовательностей, но и сходство последовательностей, такое, как одинаковый чистый заряд или сопоставимая гидрофобность/гидрофильность отдельных аминокислот. При таких сравнительных анализах первичной структуры последовательностей Р450ox группируется с другими последовательностями CYP71 и поэтому должен быть включен в семейство CYP71, несмотря на тот факт, что он идентичен менее чем на 40% всем другим представителям семейства CYP71, за исключением CYP71A1 авокадо. Поскольку он имеет низкую идентичность последовательности с другими представителями, его следует отнести к новому подсемейству. Все другие представители семейства CYP71 выделены из видов, в которых не продуцируются цианогенные соединения, и их функция остается неизвестной. Каталитические особенности ранее выделенных цитохромов Р450, принадлежащих к семейству CYP71, остаются неясными. Вероятно, они участвуют в гидроксилировании терпена. Предполагается, что ни один из них не использует оксимы в качестве субстратов и не способен к многофункциональному превращению альдоксимов в нитрилы и циангидрины.
Еще одним объектом настоящего изобретения является способ получения кДНК, кодирующей (цитохром Р450II)-монооксигеназу, которая катализирует превращение альдоксима в нитрил и превращение этого нитрила в соответствующий циангидрин. Этот способ включает:
(а) выделение и солюбилизацию микросом из растительной ткани, продуцирующей цианогенные гликозиды,
(б) очистку (цитохром Р450)-монооксигеназы,
(в) выработку антител против очищенной монооксигеназы,
(г) зондирование библиотеки экспрессии кДНК растительной ткани, продуцирующей цианогенные гликозиды, с помощью указанного антитела и
(д) выделение клонов, которые экспрессируют монооксигеназу.
Микросомы могут быть выделены из растительных тканей, в которых установлена высокая активность ферментной системы, ответственной за биосинтез цианогенных гликозидов. Эти ткани могут варьироваться от одних видов растений к другим. Предпочтительным источником микросом является свежевыделенные проростки, собранные через 1-20 дней, предпочтительно через 2-10 дней и наиболее предпочтительно через 2-4 дня после прорастания. Предпочтительными являются этиолированные проростки растений, продуцирующих цианогенные гликозиды, но можно использовать и проростки, выращенные на свету. После выделения микросомы солюбилизируют в буфере, содержащем один или несколько детергентов. Предпочтительными детергентами являются RENEX 690 (фирма J.Lorentzen A/S, Kvistgard, Дания), восстановленный Тритон Х-100 (RTX-100), Тритон Х-114 и CHAPS.
(Цитохром Р450)-монооксигеназы могут быть очищены с использованием стандартных методов очистки протеинов, таких, как ультрацентрифугирование, фракционированное осаждение, диализ, электрофорез в ДСН-ПААГ и хроматография на колонках. Пригодные колонки включают (но не ограничены ими) ионообменные колонки с таким носителем, как ДЭАЭ-сефароза, колонки с реакционноспособным красителем, таким, как агароза с Cibacron желтым 3 (Cibacron- yellow 3 agarose), агароза с Cibacron голубым (Cibacron-blue agarose) и агароза с реакционноспособным красным 120 (Reactive red 120 agarose), и колонки для гель-фильтрации, такие, как Sephacryl S-1000. Содержание цитохрома Р450 в отдельных фракциях может быть определено на основе дифференциального спектра окиси углерода (СО-спектр). Определенная трудность при выделении Р450ox, которая также возникает при количественной оценке Р450ox, заключается в его совместной миграции с желтыми пигментами во время начальных стадий очистки, вместо связывания с ионообменной колонкой, которую, как правило, применяют для очистки ферментов Р450, таких, как, например, Р450TYR. Присутствие желтых пигментов создает помехи связыванию Р450ox с целым рядом различных носителей колонки и, таким образом, является основным препятствием для дальнейшей очистки. Кроме того, отделение Р450ox от желтых пигментов можно осуществлять индуцируемым температурой распределением фаз с помощью Тритона Х-114. Этот способ был оптимизирован применительно к выделению Р450ox и удалению желтых пигментов путем увеличения количества Тритона Х-114. При концентрации Тритона Х-114 6%, что в 6-10 раз превышает концентрацию, применяемую для других цитохромов Р450ox, примерно 80% активности Р450ox в результате распределения может быть обнаружено в прозрачной нижней фазе. Помимо удаления желтых пигментов, на этой стадии очистки также происходит некоторая очистка Р450. Однако, когда содержащую Р450ox нижнюю фазу вносят на колонку с красителем Cibacron голубой, элюция в градиенте соли позволяет получить практически гомогенный Р450ox, что подтверждается наличием основной полосы, окрашенной кумасси бриллиантовым голубым, с точной молекулярной массой 55 кДа в тех фракциях, которые при восстановлении проявили Р450ox-активность.
Выделенный Р450ox давал СО-спектр с пиком абсорбции при 450 нм, но относительно большая часть выделенного фермента присутствовала в денатурированной форме Р420. Количественному определению общего содержания и удельной активности Р450ox на различных стадиях процесса выделения препятствовало постоянное превращение Р450ox в денатурированную форму Р420. Кроме того, удельная активность Р450 зависит от ингибирующих воздействий различных применяемых детергентов. Таким образом, метод определения общего содержания Р450 во фракциях может рассматриваться как полуколичественный.
Очищенные протеины могут применяться для выработки антител, например, у мышей, коз, овец, кроликов или цыплят, в ответ на их инъекцию.
От 5 до 50 мкг протеина вводят несколько раз путем инъекции примерно с 14-дневными интервалами. В предпочтительном варианте осуществления изобретения инъецируют от 10 до 20 мкг 2-6 раз с 14-дневными интервалами. Инъекции могут быть сделаны с использованием адъювантов или без них. Иммуноглобулины, очищенные от антисывортки и селезенки, могут использоваться для слияния с гибридомой согласно методу, описанному в работе Hariow и Lane в "Antibodies A Laboratory Manual", Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1988, включенной в настоящее описание в качестве ссылки. Специфическое связывание антител с (цитохром Р450II)-монооксигеназой может также использоваться при селекции растений с целью выявления растений, продуцирующих измененные количества (цитохром Р450)-монооксигеназ и, как следствие, измененные количества цианогенных гликозидов.
Методы получения библиотек кДНК растительной ткани подробно описаны у Sambrook и др. в "Molecular cloning: A Laboratory manual", Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989, при этом основные разделы этого руководства, касающиеся получения библиотек кДНК, включены в настоящее описание в качестве ссылки. Поли-(А+)-РНК выделяют из растительной ткани, которая обладает высокой активностью ферментной системы, ответственной за биосинтез цианогенных гликозидов. Эти ткани могут изменяться от одного вида растения к другому. Предпочтительной тканью для выделения поли-(А+)-РНК является ткань, выделенная из свежих проростков, собранных через 1-20 дней, предпочтительно через 2-10 дней и наиболее предпочтительно через 2-4 дня после прорастания. Полученные библиотеки кДНК могут зондироваться с помощью антител, специфически связывающихся с (цитохром Р450II)-монооксигеназой, и могут быть выделены клоны, экспрессирующие монооксигеназу.
Альтернативный способ получения кДНК, кодирующей (цитохром Р450II)-монооксигеназу, включает:
(а) выделение и солюбилизацию микросом из растительной ткани, продуцирующей цианогенные гликозиды,
(б) очистку (цитохром Р450II)-монооксигеназы,
(в) получение полной или частичной последовательности протеина монооксигеназы,
(г) создание олигонуклеотидной последовательности специфичной ДНК, кодирующей 4-15 аминокислот указанной последовательности протеина монооксигеназы,
(д) зондирование указанными олигонуклеотидами или молекулами ДНК, полученными с помощью ПЦР-амплификации кДНК с использованием указанных олигонуклеотидов, библиотеки кДНК растительной ткани, продуцирующей цианогенные гликозиды, и
(е) выделение клонов, которые кодируют (цитохром Р450II)-монооксигеназу.
Аминокислотные последовательности внутренних пептидов, которые образуются в результате расщепления протеазой, могут быть получены стандартными методами, такими, как расщепление по Эдману. Олигонуклеотиды, специфичные для ДНК, которая кодирует частичные последовательности протеинов монооксигеназ по изобретению, получают с помощью обратной относительно генетического кода трансляции частей последовательности протеина. Для обратной трансляции предпочтительны последовательности протеинов, кодируемые последовательностями ДНК, являющимися маловырожденными. Их длина состоавляет от 4 до 15, предпочтительно от 5 до 10 аминокислот. При необходимости кодоны, использующиеся в олигонуклеотидах, могут быть адаптированы к наиболее часто встречающемуся кодону растительного источника (Murray и др., Nucleic Acids Research 17: 477-498, 1989). Полученные нуклеотиды могут использоваться для зондирования библиотек кДНК согласно методу, описанному у Sambrook и др. ("Molecular cloning: A Laboratory manual", Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989), для клонов, которые способны образовывать парные основания с указанными олигонуклеотидами. В альтернативном варианте олигонуклеотиды могут использоваться в полимеразной цепной реакции (ПЦР), методология которой хорошо известна в данной области, с использованием растительной кДНК в качестве матрицы для амплификации. В этом случае полученные продукты амплификации используют для зондирования библиотек кДНК. Выделяют клоны, кодирующие (цитохром Р450II)-монооксигеназы.
Альтернативный метод клонирования генов основан на конструировании библиотеки генов, состоящей из векторов экспрессии. Согласно этому методу, аналогично описанным выше методам, сначала из клетки или ткани, способной экспрессировать Р450II-монооксигеназу, выделяют геномную ДНК, предпочтительно, однако, кДНК, а затем осуществляют сплайсинг в пригодном векторе экспрессии. Полученные таким путем библиотеки генов могут быть подвергнуты скринингу с использованием соответствующих методов, например, с помощью антител. Отбирают клоны, которые включают в качестве вставки требуемый ген или по крайней мере часть гена.
В альтернативном варианте молекулы кДНК, кодирующие (цитохром Р450)-монооксигеназу, которая катализирует превращение альдоксима в нитрил и превращение этого нитрила в соответствующий циангидрин, могут быть получены путем
(а) создания вырожденных олигонуклеоидов, соответствующих 3-10 аминокислотам консервативных областей цитохромов типа А,
(б) использования вырожденных олигонуклеотидов для амплификации одного или нескольких специфичных для цитохрома фрагментов ДНК с помощью полимеразной цепной реакции,
(в) скрининга библиотеки кДНК с помощью специфичных для цитохрома фрагментов с получением полноразмерной кДНК,
(г) экспрессии полноразмерной кДНК в микроорганизме-хозяине,
(д) идентификации хозяев, экспрессирующих (цитохром Р450)-монооксигеназу, которая катализирует превращение альдоксима в нитрил и превращение этого нитрила в соответствующий циангидрин, и
(е) выделения клонированной ДНК из хозяина.
Общая ДНК из библиотеки ДНК, предпочтительно из библиотеки кДНК, может использоваться в качестве матрицы в ПЦР с одним или с несколькими праймерами, характерными для консервативных областей цитохромов типа А (Durst и др., Grug Metabolism and Drug Interactions 12: 189-206, 1995), которые, вероятно, являются производными общего для растительного цитохрома Р450 прародителя. На основе сравнительного анализа множества последовательностей цитохромов Р450 типа А на аминокислотном уровне могут быть выделены три высококонсервативные области: область 1 (V/I)KEX(L/F)R, область 2 FXPERF и область 3 PFGXGRRXCXG. Могут быть созданы вырожденные инозин (I)-содержащие праймеры, каждый из которых перекрывает от 3 до 10, предпочтительно примерно 5 или 6 аминокислот, двух областей соответственно. ПЦР проводят, например, в виде трех последовательных серий реакций. В первой серии реакций с помощью праймера, перекрывающего консенсусную область FXPERF, и стандартного праймера фага Т7, перекрывающего промотор Т7, в библиотеке векторов амплифицируют кДНК, происходящие из мРНК, кодирующих цитохромы Р450 типа А. Во второй серии ПЦР с помощью праймеров, перекрывающих две консенсусные области, и с использованием в качестве матрицы амплифицированной ДНК, полученной в первой серии ПЦР, предпочтительно амплифицируют фрагмент размером 100 пар оснований, который затем встраивают в pBluescript и секвенируют. Специфичные для гена праймеры создают на основе полученной последовательности ДНК. Их используют в третьей серии ПЦР в комбинации с поли-А-хвостом (праймер dT+V) и ДНК, полученной в первой серии ПЦР, в качестве матрицы для амплификации фрагмента ДНК размером примерно 500 пар оснований, который может применяться в качестве специфичного для гена зонда для выделения полноразмерных кДНК. Этот подход, основанный на использовании серий ПЦР, не является уникальным для выделения Р450ox, а он является общим для выделения цитохромов Р450 типа А. Полученные цитохромы Р450 типа А необходимо подвергать гетерологичной экспрессии для определения их функции.
Клоны кДНК или ПЦР-продукты, полученные согласно описанному выше способу, или их фрагменты могут использоваться в качестве зонда для гибридизации в процессе идентификации дополнительных последовательностей ДНК, кодирующих продукт, представляющий собой протеин, обладающий активностью Р450II-зависимой монооксигеназы, из гомологичного или гетерологичного организма-источника, такого, как грибы или гетерологичные растения. Пригодным источником является ткань растений, содержащих цианогенные гликозиды.
Такие клоны или ПЦР-продукты также могут применяться в качестве RFLP-маркеров для определения, например, локализации гена (цитохром Р450)-монооксигеназы либо близко сцепленного с ним признака в растительном геноме или для селекции с помощью маркеров (ЕР-А-306139, WO 89/07647).
С помощью описанных выше методов возможно выделять различные гены, которые кодируют Р450II-монооксигеназу. Эти гены могут применяться в способе получения очищенной рекомбинантной (цитохром Р450II)-монооксигеназы, которая катализирует превращение альдоксима в нитрил и превращение этого нитрила в соответствующий циангидрин, причем этом способ включает:
(а) конструирование гена, кодирующего указанную монооксигеназу, которая может экспрессироваться в организме-хозяине, таком, как бактерии, дрожжи или клетки насекомых,
(б) трансформацию указанного организма-хозяина сконструированным геном и
(в) выделение протеина из организма-хозяина или из супернатанта культуры.
В предпочтительном варианте этот способ применяют для очищения рекомбинантного цитохрома Р450ox или цитохрома Р450ox, который модифицирован с использованием известных методик генной инженерии. Предпочтительно модификации приводят к увеличению экспрессии рекомбинантного протеина или к изменению субстратной специфичности.
Молекулы ДНК по изобретению могут применяться для получения трансгеннных растений, устойчивых к насекомым или к клещам. Их конкретные, но не ограничивающие, примеры перечислены в таблице В международной заявки WO 95/16041 (стр. 45), а также включают растения, устойчивые к указанным ниже нематодам. Для удобства эта таблица не повторена в данном описании, но подразумевается, что она включена в него посредством ссылки на описание к WO 95/16041. Предпочтительно трансгенные растения обладают устойчивостью к представителям отрядов Coleoptera и Lepidoptera, таких, как блошка длинноусая западная (Diabrotica virgifera virgifera), блошка длинноусая северная (Diabrotica longicornis barberi), блошка длинноусая южная (Diabrotica undecimpunctata howardi), совка хлопковая, мотылек кукурузный, огневка Crambus caliginosellus, розовый коробочный червь хлопчатника и совка Heliothis virescens.
Нематоды являются важными паразитами растений (которые относятся к классу первичнополостных червей), общие потери сельского хозяйства от которых составляют более 100 миллиардов долларов США в год. Некоторые виды нематод создают места кормежки, в которых происходит модификация растительной клетки, и питаются на одном месте в течение нескольких часов или существенно дольше. Они включают виды родов Meloidogyne, Globodera, Heterodera, Rotylenchulus, Tyienchulus, Naccobus, Xiphinema, Longidorus, Paralongidorus, Cryphodera, Trophotylenchulus, Hemicycliophora, Criconemella, Verutus и Heliocotylenchus. Виды нематод, которые, как считается, питаются в одном месте более ограниченный промежуток времени, включают представителей таких родов, как Pratylenchus, Radopholus, Hirschmanniella, Trichodorus, Paratrichodorus, Ditylenchus, Aphelenchoides, Scutellonema и Belonolaimus.
Трансгенные растения включают ДНК, кодирующую новые монооксигеназы, которые катализируют превращение альдоксима в нитрил и превращение этого нитрила в соответствующий циангидрин. Кроме того, трансгенные растения могут включать гены монооксигеназ, генетически связанные с генами устойчивости к гербицидам. Трансгенные растения предпочтительно являются представителями однодольных или двудольных растений. Их конкретные примеры перечислены в таблице А международной заявки WO 95/16041 (стр. 33-44). Для удобства эта таблица не повторена в данном описании, но подразумевается, что она включена в него посредством ссылки на описание к WO 95/16041. Предпочтительно растения выбирают из группы, включающей кукурузу, рис, пшеницу, ячмень, сорго, хлопчатник, сою, подсолнечник, травы, масличный рапс, сахарную свеклу, брокколи, цветную капусту, капусту, огурец, кукурузу сладкую, дайкон, горохи, салат-латук, дыню, перец, тыкву, томаты и арбуз. Растения могут быть получены способом, включающим
(а) интродукцию в растительную клетку или в растительную ткань, которая может быть регенерирована до полного растения, ДНК, включающую экспрессируемый в этом растении ген, кодирующий монооксигеназу по изобретению, и
(б) отбор трансгенных растений.
Аналогично этому молекулы ДНК по изобретению могут применяться для получения трансгенных растений, экспрессирующих антисмысловую либо смысловую РНК или рибозимы, для которых мишенью служат гены эндогенных Р450II-монооксигеназ. Экспрессия этих молекул в трансгенных растениях уменьшает экспрессию (цитохром Р450II)-монооксигеназ. Установлено, что такие растения обладают повышенной устойчивостью к болезням или большей питательной ценностью в результате снижения экспрессии цианогенных гликозидов. Растения могут быть получены с помощью способа, включающего
(а) интродукцию в растительную клетку или в растительную ткань, которая может быть регенерирована до полного растения, ДНК, кодирующей смысловую РНК, антисмысловую РНК или рибозим, экспрессия которых уменьшает экспрессию (цитохром Р450II)-монооксигеназ, и
(б) отбор трансгенных растений.
Целый ряд очень эффективных способов доступен для интродукции ДНК в растительные клетки, и эти способы основаны на использовании векторов для переноса гена или на непосредственном переносе гена.
Один из возможных методов встраивания генной конструкции в клетку основан на заражении растительной клетки бактерией Agrobacterium tumefaciens и/или Agrobacterium rhizogenes, трансформированной указанной генной конструкцией. Трансгенные растительные клетки затем культивируют с использованием приемлемых условий, известных специалисту в данной области, таким образом, чтобы они образовывали побеги и корни и в конечном итоге формировались в целые растения.
В объеме настоящего изобретения используется метод так называемой трансформации листовых дисков с помощью Agrobacterium (Horsch и др., Science 227: 1229-1231, 1985). Стерильные листовые диски соответствующего растения-мишени инкубируют с клетками Agrobacterium, включающими одну из конструкций химерных генов по изобретению, а затем переносят в пригодную питательную среду или на нее. Особенно пригодной и, следовательно, предпочтительной согласно изобретению является LS-среда, отвержденная путем добавления агара и дополненная одним или несколькими обычно применяемыми регуляторами роста растений, прежде всего регуляторами, выбранными из группы ауксинов, включающей α-нафтилуксусную кислоту, пиклорам, 2,4,5-трихлорфеноксиуксусную кислоту, 2,4-дихлорфеноксиуксусную кислоту, индол-3-масляную кислоту, индол-3-молочную кислоту, индол-3-янтарную кислоту, индол-3-уксусную кислоту и пара-хлорфеноксиуксусную кислоту, а также выбранными из группы цитокининов, включающей кинетин, 6-бензиладенин, 2-изопентениладенин и зеатин. Предпочтительные концентрации ауксинов и цитокининов составляют от 0,1 до 10 мг/л.
После инкубации в течение нескольких дней, предпочтительно в течение 2-3 дней, при температуре от 20 до 40°С, предпочтительно от 23 до 35°С и более предпочтительно при 25°С и в рассеянном свете, листовые диски переносят в приемлемую среду с целью индукции побегов. Особенно предпочтительной для отбора трансформантов является LS-среда, которая не содержит ауксин, но вместо этого содержит цитокинин и к которой добавлено вещество, позволяющее осуществлять отбор. Культуры выдерживают на свету и затем через определенные интервалы времени, предпочтительно с интервалом в одну неделю, переносят на свежую среду. Развивающиеся зеленые побеги срезают и дополнительно культивируют в среде, которая индуцирует у побегов развитие корней. Особенно предпочтительной согласно данному изобретению является LS-среда, которая не содержит ауксин или цитокинин, но которая содержит селектирующее вещество, добавленное для отбора трансформантов.
Для встраивания в растительный материал конструкций генов по изобретению помимо опосредуемой Agrobacterium трансформации согласно настоящему изобретению возможно использовать методы непосредственной трансформации.
Например, генетический материал, входящий в вектор, может быть непосредственно встроен в растительную клетку, например, с использованием чисто физических методов, например, путем микроинъекции с помощью микропипеток с тонкими оттянутыми концами (Neuhaus и др., Theoretical and Applied Genetics 74: 363-373, 1987), электропорации (D'Halluin и др., The Plant Cell 4: 1495-1505, 1992; WO 92/09696) или предпочтительно путем бомбардировки клеток микроснарядами, покрытыми трансформирующей ДНК (Whang и др., "Microprojectile Bombardment", Plant Molecular Biology 11: 433-439, 1988; Gordon-Kamm и др., The Plant Cell 2: 603-618, 1990; McCabe и др., Bio/Technology 11: 596-598, 1993; Christou и др., Plant Physiol. 87: 671-674, 1988; Koziel и др., Biotechnology 11: 194-200, 1993). Кроме того, растительный материал, подлежащий трансформации, необязательно может быть предварительно обработан осмотически активным веществом, таким, как сахароза, сорбит, полиэтиленгликоль, глюкоза или маннит.
Другие возможные способы непосредственного переноса генетического материала в растительную клетку включают обработку протопластов с помощью процедур, приводящих к модификации плазматической мембраны, например, обработку полиэтиленгликолем, обработку тепловым шоком или электропорацию либо различные сочетания указанных способов (Shillito и др. Biotechnology 3:1099-1103, 1985).
Еще один, основанный на чисто химических процедурах способ непосредственной интродукции генетического материала в растительные клетки, который позволяет очень эффективно и быстро осуществить трансформацию, описан у Negrutiu и др. в Plant Molecular Biology 8: 363-373, 1987.
Также пригодным для трансформации растительного материала является непосредственный перенос гена с помощью котрансформации (Schocher и др., Bio/Technology 4: 1093-1096, 1986).
Приведенный выше перечень возможных методов трансформации указан в качестве примера, и он не является полным и не ограничивает объем изобретения.
Генетические свойства, сконструированные у описанных выше трансгенных семенах и растениях, передаются с помощью полового размножения или вегетативного роста и, таким образом, могут сохраняться и передаваться по наследству потомству растений. Обычно указанное сохранение и передачу потомству осуществляют с использованием известных сельскохозяйственных методов, разработанных для этих конкретных целей, таких, как обработка почвы, посев или сбор урожая. Также могут применяться специализированные способы, такие, как гидропоника или выращивание в закрытом грунте. Поскольку выращиваемые культурные растения подвержены поражению инфекциями и повреждениям насекомыми, а также подвержены конкуренции со стороны сорных растений, для улучшения урожая проводят мероприятия по борьбе с сорняками, болезнями растений, насекомыми, нематодами и другими вредными факторами. Такие мероприятия включают механическую обработку, как, например, обработка почвы или удаление сорняков и зараженных растений, а также применение средств защиты растений, таких, как гербициды, фунгициды, гаметоциды, нематоциды, регуляторы роста, агенты, способствующие созреванию, и инсектициды.
Кроме того, обладающие рядом преимуществ генетические свойства трансгенных растений и семян по изобретению могут быть использованы при селекции растений с целью выведения растений с улучшенными свойствами, такими, как толерантность к пестицидам, гербицидам или стрессу, с улучшенной питательной ценностью, повышенной урожайностью или улучшенным строением, способствующим уменьшению потерь от полегания или осыпания. Для различных стадий селекции характерно целенаправленное вмешательство человека, такое, как отбор линий, подлежащих скрещиванию, непосредственное опыление родительских линий или отбор соответствующего потомства растений. В зависимости от требуемых свойств выбирают различные методы селекции. Соответствующие методы хорошо известны в данной области и включают (но не ограничены ими) гибридизацию, инбридинг, возвратное скрещивание, многолинейное скрещивание, смешивание сортов, межвидовую гибридизацию, методы анэуплодии и т.д. Методы гибридизации также включают стерилизацию растений с целью получения растений с мужской или женской стерильностью с помощью механических, химических или биохимических способов. Перекрестное опыление мужского стерильного растения пыльцой различных линий гарантирует, что геном мужского стерильного, но фертильного женского растения будет равномерно приобретать свойства обеих родительских линий. Таким образом, трансгенные семена и растения по настоящему изобретению могут применяться для селекции улучшенных линий растений, что, например, позволяет увеличить эффективность обычных методов, таких, как обработка гербицидом или пестицидом, или позволяет отказаться от применения этих методов благодаря модифицированным генетическим особенностям этих растений. Кроме того, могут быть получены новые культурные растения с улучшенной толерантностью к стрессу благодаря оптимизации генетического "аппарата", что позволяет получить урожай продуктов лучшего качества по сравнению с продуктами, полученными от растений, которые не обладают способностью сопротивляться аналогичным вредным условиям в процессе их развития.
При получении семян качество прорастания и однородность семян являются важными характеристиками продукта, в то время как качество прорастания и однородность семян, собираемых и продаваемых фермером, не являются важными. Поскольку трудно выращивать культурное растение вне контакта с семенами другого культурного и сорного растения, для борьбы с передающимися с семенами болезнями и для получения семян с хорошим прорастанием производителями, занимающимися выращиванием, кондиционированием и продажей чистых семян, разработаны довольно обширные и целенаправленные технологии получения семян. Так, обычно фермер практикует покупку сертифицированного семенного материала, удовлетворяющего определенным стандартам качества, а не использует семенной материал, полученный от его собственной культуры. Материал для размножения, который используют в качестве семенного материала, обычно обрабатывают защитным покрытием, включающим гербициды, инсектициды, фунгициды, бактерициды, нематоциды, моллюскициды или их смеси. Обычно применяемые защитные покрытия включают такие соединения, как каптан, карбоксин, тирам (TMTD®), металаксил (Apron®) и пиримифосметил (Actellic®). При необходимости эти соединения изготавливают в виде препаративной формы вместе с дополнительными носителями, поверхностно-активными веществами или способствующими нанесению адъювантами, обычно применяемыми в технологии приготовления препаративных форм, предназначенных для защиты от повреждения, вызванного бактериями, грибами или насекомыми-вредителями. Защитные покрытия могут быть нанесены путем пропики материала для размножения жидкой композицией или путем нанесения покрытия из объединенной влажной или сухой композицией. Также возможны другие способы нанесения, такие, как обработка почек или плодов.
Еще одним объектом настоящего изобретения являются новые сельскохозяйственные способы, такие, как приведенные выше, для которых характерно применение трансгенных растений, трансгенного растительного материала или трансгенного семенного материала по настоящему изобретению.
В приведенных ниже примерах дополнительно описаны материалы и методы, используемые при осуществлении изобретения, и полученные результаты. Эти примеры только иллюстрируют настоящее изобретение и не ограгичивают его объем.
Примеры
Пример 1: Получение микросом
Все стадии получения микросом проводили при 4°С, если не указано иное. Все буферы дегазировали перемешиванием в вакууме и продувкой аргоном.
Семена Sorghum bicolor (L.) Moench (гибрид SS1000 от фирмы AgriPro, Texas, США) выращивали в темноте в течение 40 ч при 28°С на металлических ситах, покрытых марлей. Микросомы получали из этиолированных проростков длиной примерно 3 см. Проростки собирали и гомогенизировали с использованием ступки и пестика в двух объемах (об./мас.) 250 мМ сахарозы, 100 мМ Трицина (рН 7,9), 2 мМ ЭДТК и 2 мМ ДТТ. Перед гомогенизацией добавляли поливинилполипирролидон (0,1 г/г массы сырой ткани). Гомогенат фильтровали через найлоновое полотно с размером пор 22 мкм и центрифугировали в течение 10 мин при 16500×g. Супернатант центрифугировали в течение 1 ч при 165000×g. Микросомальный дебрис повторно суспендировали и гомогенизировали в буфере для выделения, используя гомогенизатор Поттера-Элвехима, снабженный тефлоновым пестиком. После повторного центрифугирования и повторной гомогенизации гомогенат замораживали в жидком азоте и хранили при -80°С до использования.
Пример 2: Анализ фермента: определение общего содержания цитохрома Р450
Количественное определение общего цитохрома Р450 проводили с помощью дифференицальной спектроскопии, используя коэффициент затухания 91 мМ-1см-1 аддукта восстановленного цитохрома Р450 и окиси углерода (A450-490) (Omura и др., J.Biol.Chem. 239: 2370-2378, 1964).
Пример 3: Очистка цитохрома Р450
Все стадии очистки проводили при 4°С, если не указано иное.

Перед добавлением детергента и ДТТ буферы трижды дегазировали перемешиванием в вакууме. Между каждой дегазацией буфер продували аргоном. Способность различных полученных с колонок фракций преобразовывать в ходе обмена веществ радиоактивно меченный пара-гидроксифенилацетальдоксим определяли во время процесса очистки, выявляя присутствие во фракциях Р450ox.
Микросомы (400 мг протеина в 20 мл) разводили до 100 мл буфером, включающим 8,9% глицерина, 10 мМ KH2РО4/К2HРО4 (рН 7,9), 0,2 мМ ЭДТК, 2 мМ ДТТ, после чего при постоянном перемешивании медленно добавляли 100 мл 10 мМ КН2РO4/К2НРO4 (рН 7,9), 8,9% глицерина, 0,2 мМ ЭДТК, 2 мМ ДТТ, 0,1 об.% RTX-100, 2% Renex. После дополнительного перемешивания в течение 30 мин и последующего ультрацентрифугирования при 150000×g в течение 35 мин примерно 190 мл супернатанта вносили со скоростью потока 100 мл/ч на сефарозную колонку размером 5×5 см, заполненную ДЭАЭ-сефарозой FF/сефарозой S-100 (в отношении 20/80 объемов сырого продукта, фирма Pharmacia), уравновешенную буфером А. Ионообменную смолу типа ДЭАЭ-сефароза разбавляли материалом для гель-фильтрации сефарозой S-100 в соотношении 1:4 с целью избежать получения слишком высоких концентраций (цитохром Р450)-зависимых ферментов при связывании, что может привести к необратимой агрегации. Затем колонку промывали 150 мл буфера А. Р450ox слабо связывался с колонкой, и его главным образом получали из погона и промывочных фракций, которые содержат желтый пигмент. Объединяли содержащие Р450ox фракции (примерно 200 мл), идентифицированные по их абсорбции при 420 нм, их спектру связывания с СО и их способности преобразовывать в ходе обмена веществ оксим в экспериментах по восстановлению (см. пример 4). Их использовали для дополнительной очистки или их можно было заморозить.
Концентрацию объединенных фракций Р450ox доводили при постоянном перемешивании до 30 об.% глицерином и 6%-ным Тритоном Х-114, добавляя по каплям соответствующие количества глицерина и Тритона Х-114. Перемешивание продолжали в течение 20 мин, после чего немедленно смесь, не прерываясь, центрифугировали в течение 25 мин при 24500×g и при 25°С (индуцированное температурой распределение фаз с помощью Тритона Х-114). Образовывались две фазы: верхняя желтая фаза и нижняя прозрачная фаза. Нижнюю фазу, которая содержала основную часть активности цитохрома Р450ox, собирали, разбавляли в 2,5 раза до объема примерно 350 мл буфером В и вносили со скоростью потока 70 мл/ч на агарозную колонку размером 1,9×5 см, заполненную 3GА-агарозой с красителем Cibacron голубой, уравновешенную буфером В. Колонку промывали 50 мл буфера В и сохранившийся цитохром Р450ox элюировали примерно 60 мл элюента с линейным градиентом от 0 до 1,5М КСl в буфере В. Десять фракций, для которых с помощью электрофореза в ДСН-ПААГ подтверждено наличие одной полосы, соответствующей полипептиду с молекулярной массой в диапазоне 50-60 кДа, объединяли и подвергали диализу в атмосфере азота в противотоке 1 л буфера, включающего 8,9% глицерина, 10 мМ КН2РO4/К2НРO4 (рН 7,9), 5 мМ ЭДТК, 2 мМ ДТТ (буфер для диализа), с целью уменьшения содержания соли и детергента. Препарат фермента замораживали в жидком азоте и хранили при -80°С.
Пример 4: Определение характеристик цитохрома Р450ox, полученного путем выделения из микросом сорго
4.1. Молекулярная масса и данные об аминокислотной последовательности
Молекулярная масса Р450ox, определенная с помощью электрофореза в ДСН-ПААГ, составляет 55 кДа. Протеиновую полосу, соответствующую Р450ox, выделенную из колонки, заполненной 3GА-агарозой с красителем Cibacron голубой, вырезали из 8-25%-ных ДСН-полиакриламидных гелей и подвергали электроэлюции. Полученный в результате электроэлюции протеин расщепляли с помощью эндопротеиназы Glu-C (протеаза V8 с чистотой, необходимой для секвенирования, 18 ч, 23°С) согласно инструкции производителя (фирма Boehringer Mannheim), используя массовое соотношение между протеиназой и протеином примерно 1:100. Образец полученного в результате электроэлюции протеина и расщепленного протеина подвергали электрофорезу в ДСН-ПААГ и переносили на мембраны типа ProBlott (фирма Applied Biosystems). Области мембраны, окрашенные кумасси бриллиантовым голубым, вырезали и секвенировали N-концевые аминокислоты на секвенаторе фирмы Applied Biosystems, модель 470А, снабженном анализатором модели 120А, работающим в режиме "он-лайн", для идентификации фенилтиогудантионовых производных аминоксилот.
В результате N-концевого аминоксилотного секвенирования получено две последовательности, которые могли считываться независимо друг от друга благодаря различиям в их относительном содержании. Поиск в базе данных (в программе BLAST) показал, что последовательность -GLVKEGVDMEEGTL отличается только по одному положению от N-концевой последовательности субъединицы В вакуольной АТФазы ячменя (Hordeum vulgare), которая имеет следующую последовательность: MGLVKEGADMEEGTL (регистрационный номер L11862). Субъединица В ячменя имеет предсказанную молекулярную массу 54 кДа (20). Присутствие субъединицы В вакуольной АТФазы ячменя в качестве загрязнителя в препарате Р450ox далее было точно подтверждено Вестрен-блоттингом, который показал наличие одной полосы, соответствующей 55 кДа, при использовании моноклонального антитела к субъединице В вакуольной АТФазы из корней овса, предоставленного доктором Heven Sze.
Субъединица В может быть удалена из препарата Р450ox иммобилизацией на покрытых антителами лунках микротитрационного планшета. Этот подход позволял точно установить, что N-концевая аминокислотная последовательность Р450ox представляет собой -ATTATPQLLGGSSVPEQ, а также установить, что последовательность одного внутреннего фрагмента пептида Р450ox представляет собой MDRLVADLDRAAA. Попытки удалить остаточные количества субъединицы В вакуольной АТФазы привели к образованию СО-дифференциального спектра, в котором количество компонента с пиком абсорбции при 420 нм, представляющего собой неактивную денатурированную Р420-форму Р450ox, сильно возросло и также привело к потере или к существенному снижению способности к восстановлению активности Р450ox в полученных фракциях. Это свидетельствует о лабильности, присущей Р450ox. Маловероятно, чтобы субъединица В вакуольной АТФазы обладала какой-либо каталитической активностью, связанной с Р450ox. Таким образом, в приведенных далее исследованиях метаболизма Р450ox допускалось присутствие субъединицы В в качестве загрязнителя.
N - концевая последовательность:
--ATTATPQLLGGSVPEQ-- (SEQ ID NO:3)
Внутренняя последовательность:
--MDRLVADLDRAAA--(SEQ ID NO:4)
4.2. Выделение NADPH-P450-oкcидopeдyктaзы
NADPH-P450-oкcидopeдyктaзa связывалась с колонкой, заполненной ДЭАЭ-сефарозой FF/сефарозой S-100, и ее элюировали с помощью буфера А с добавлением 0,5М КСl. Затем редуктазу очищали до гомогенного состояния на колонке с 2',5'-ADP-ceфapoзoй 4В (фирма Pharmacia) согласно известному методу (Halkier и Moller, Plant Physiol. 96: 10-17, 1990) и концентрировали примерно до 15 ед./мл.
4.3. Получение растворимой УДФГ-глюкозилтрансферазы
Глюкозилтрансферазу частично очищали с помощью фракционирования сульфатом аммония супернатанта, полученного центрифугированием в процессе выделения микросом. Содержащая глюкозилтрансферазу фракция осаждалась при использовании концентрации (NH4)2SO4 от 40% до 60% и ее растворяли в 5 мл 50 М Трицина (рН 7,9), 2 М ДТТ и подвергали диализу в противотоке 2 л этого же буфера в течение ночи.
4.4. Восстановление активности цитохрома Р450ox
Восстановление ферментативной активности микросомального цитохрома Р450 осуществляли включением фермента цитохрома Р450 и соответствующей NADPH-(цитохром Р450)-оксидоредуктазы в липидные мицеллы. Можно использовать смесь липидов, однако в случае цитохрома Р450ox наиболее выраженная ферментативная активность, вероятно, достигается при использовании дилауроилфосфатидилхолина (ДЛФХ). Количество правильно сформированных комплексов на основе цитохрома Р450 и NADPH-(цитохром Р450)-оксидоредуктазы является фактором, ограничивающим скорость. Для получения достаточного количества активных комплексов использовали избыточные количества оксидоредуктазы и концентрированные растворы ферментов.
Функционально активный восстановленный фермент получали с помощью следующих компонентов:

5 мкл суспензии липида смешивали в пробирке Эппендорфа с 5 мкл NADPH-(цитохром Р450)-оксидоредуктазы (0,075 ед.), 10 мкл раствора цитохрома Р450ox (примерно 0,4 пмоля) и 0,5 мкл 14С-оксима (0,014 мкКи/мкл, 394 мКи/ммоль). Конечный объем доводили до 30 мкл с помощью 50 мМ Трицина (рН 7,9) и ферментативную реакцию инициировали, добавляя 1 мкл раствора NADPH. Контрольные образцы получают путем удаления либо NADPH-(цитохром Р450)-оксидоредуктазы, либо NADPH из реакционной смеси. Пробирки инкубировали при постоянной температуре 30°С при легком перемешивании в течение 1 ч. После инкубации реакционные смеси наносили на пластинки для тонкослойной хроматографии (ТСХ), покрытые двуокисью кремния (силикагель 60 F254, фирма Merck), и производили разделение с использованием в качестве подвижной фазы смеси этилацетат/толуол (в объемном соотношении 1:5). Пластинки помещали на экраны с нанесенным фосфором и выдерживали в течение ночи и полученные в результате продукты пара-гидроксифенилацетонитрил и пара-гидроксибензальдегид визуализировали с помощью томографа для выявления фосфора типа STORM 840 фирмы Molecular Dynamics.
После восстановления в липидных мицеллах цитохром Р450ox катализировал превращение пара-гидроксифенилацетальдегидоксима в пара-гидроксиманделонитрил, который диссоциировал с получением пара-гидроксибензальдегида и HCN. Это свидетельствует о том, что цитохром Р450 представляет собой многофункциональный протеин, катализирующий как превращение пара-гидроксифенилацетальдегидоксима в пара-гидроксифенилацетонитрил, так и превращение пара-гидроксифенилацетонитрила в пара-гидроксиманделонитрил. Активность Р450ox в значительной степени зависит от присутствия NADPH-P450-oкcидopeдyктaзы и NADPH. Дитионит натрия (10 мМ) не поддерживал метаболизм пара-гидроксифенилацетальдегидоксима. Исключение стадии диализа фермента перед его восстановлением приводило к относительному увеличению количества пара-гидроксифенилацетонитрила по сравнению с пара-гидроксибензальдегидом.
4.5. Восстановление in vitro полного пути синтеза дхуррина из его предшественника аминокислоты тирозина
Полная реакционная смесь содержала 3 мкл выделенного рекомбинантного P450TYR (6 пмолей, полученного в результате гетерологичной экспрессии в Е. coli и выделенного согласно методу, описанному Halkier и др. в Arch. Biochem. Biophys. 322: 369-377, 1995), 10 мкл выделенного и подвергнутого диализу Р450ox (примерно 0,4 пмоля), 5 мкл NADPH-P450-оксидоредуктазы (0,075 ед.), 1 мкл частично очищенной УДФГ-глюкозилтрансферазы из Sorghum, 5 мкл ДЛФХ (10 мг/мл в 50 мМ Крi (рН 7,0)), 0,25 мкл [U-14C] -тирозина (0,05 мкКи/ммоль, 443 мКи/ммоль, фирма Amersham), 3 мкл УДФГ (33 мг/мл в 50 мМ Крi (рН 7,0)) и 3 мкл кастаноспермина (2 мМ в 50 мМ Крi (рН 7,0)). Компоненты смешивали повторным суспендированием и при необходимости конечный объем доводили до 30 мкл с помощью 50 мМ Крi (рН 7,0)).
Ферментативную реакцию инициировали, добавляя 1 мкл NADPH (25 мг/мл). Дхуррин также синтезировали посредством восстановления Р450ox с использованием пара-гидроксифенилацетальдегидоксима (удаляли Р450TYR и тирозин из реакционных смесей, остальные компоненты оставляли без изменения). В этих опытах в качестве субстрата для Р450ox использовали либо 0,5 мкл [U-14С]-пара-гидроксифенилацетальдегидоксима (0,014 мкКи/мкл, 394 мКи/ммоль), либо 3 мкл немеченого пара-гидроксифенилацетальдегидоксима (20 мМ). В последнем случае радиоактивная метка представляла собой 1 мкл [U-14С]-УДФГ (0,025 мкКи/мкл, 287 мКи/ммоль, фирма Amersham). Все реакционные смеси готовили с дублированием. После инкубации в течение 1 ч при 30°С каждый набор реакционных смесей наносили на пластинки для ТСХ. Первый набор реакционных смесей анализировали, используя в качестве растворителя смесь этилацетат/толуол, описанную в примере 4.5. Второй набор реакционных смесей анализировали, используя систему растворителей, состоящую из этилацетата/ацетона/дихлорметана/метанола/воды (в объемном соотношении 20/15/6/5/4), с целью отделить гидрофильный продукт дхуррин от тирозина и от гидрофобных промежуточных продуктов. Радиоактивно меченные субстраты и продукты визуализировали с помощью томографа для выявления фосфора типа STORM 840.
Совместное использование выделенных P450TYR и Р450ox в экспериментах по восстановлению с использованием радиоактивно меченного тирозина в качестве субстрата привело к получению пара-гидроксифенилацетонитрила и пара-гидроксибензальдегида. Это свидетельствует о том, что Р450TYR и Р450ox обладают способностью действовать совместно in vitro. Таким образом, продуцируемый с помощью Р450TYR пара-гидроксифенилацетальдоксим эффективно используется в качестве субстрата цитохромом Р450ox. Никакой активности не обнаружено в отсутствие NАDРН-Р450-оксидоредуктазы или в отсутствие NADPH.
Получение in vitro дхуррина с использованием пара-гидроксифенилацетальдоксима в качестве субстрата осуществляли восстановлением Р450ox вместе с частично очищенной растворимой УДФГ-глюкозилтрансферазой в присутствии NADPH и УДФГ. Клон кДНК из сорго, кодирующий УДФГ-глюкозилтрансферазу, который специфично используется в качестве субстрата пара-гидроксиманделонитрил, не был доступен. Поэтому в настоящем исследовании неочищенный экстракт растворимой УДФГ-глюкозилтрансферазы из сорго использовали для глюкозилирования пара-гидроксиманделонитрила и для демонстрации in vitro восстановления полного пути биосинтеза дхуррина.
Внесенный радиоактивно меченный пара-гидроксифенилацетальдоксим оказался полностью преобразованным в ходе обмена веществ. Кастаноспермин добавляли для ингибирования глюкозидазной активности, присутствующей в препарате УДФГ-глюкозилтрансферазы. Кроме описанной выше ТСХ-системы, предназначенной для разделения гидрофобных соединений, вводили дополнительную ТСХ-систему для разделения гидрофильных соединений типа дхуррина. Образованный в опыте по восстановлению пара-гидроксиманделонитрил частично превращался в дхуррин, что подтверждалось образованием радиоактивно меченного соединения, мигрирующего совместно с чистым дхуррином, используемым в качестве стандарта. Дополнительное доказательство того, что это радиоактивно меченное соединение представляло собой дхуррин, получено по его разложению в отсутствие кастаноспермина и по образованию мигрирующего совместно радиоактивно меченного продукта, когда эксперимент повторяли с радиоактивно меченным УДФГ вместо радиоактивно меченного пара-гидроксифенилацетальдоксима. Радиоактивно меченный УДФГ неспецифично метил целый ряд относительно гидрофильных соединений. Вследствие лабильности пара-гидроксиманделонитрила его превращение в дхуррин экспериментально выявляли по исчезновению пара-гидроксибензальдегида. Когда радиоактивно меченный пара-гидроксифенилацетальдоксим использовали в качестве субстрата, то кроме дхуррина получали целый ряд неидентифицированных гидрофобных радиоактивно меченных соединений.
Образование этих соединений происходило в отсутствие УДФГ, но требовало присутствия растворимого экстракта, свидетельствуя о том, что экстракт УДФГ-глюкозилтрансферазы содержал дополнительные ферментативные активности. Глюкозилирование фенольной группы пара-гидроксиманделонитрила должно привести к образованию пара-глюкопиранозилоксиманделонитрила. Не было обнаружено никакого радиоактивно меченного продукта, мигрирующего совместно со стандартным чистым пара-глюкопиранозилоксиманделонитрилом. Глюкозидазная активность, присутствующая в экстракте УДФГ-глюкозилтрансферазы, эффективно ингибировалась кастаноспермином.
При восстановлении in vitro коэффициент обратного превращения для P450TYR (CYP79) составляет 230 мин-1 (Sibbesen и др., J.Biol.Chem. 270: 3506-3511, 1995). Частичное превращение Р450ox в его денатурированную форму Р420 препятствует определению его коэффициента обратного превращения. При использовании микросомальной системы значения Кm и Vmax для процесса получения пара-гидроксиманделонитрила из тирозина, пара-гидроксифенилацетальдоксима и пара-гидроксифенилацетонитрила составляют 0,03, 0,05 и 0,10 мМ и 145, 400 и 50 нмолей на мг протеина-1 ч-1 соответственно (Moller и др., J. Biol. Chem. 254: 8575-8583, 1979).
Полный путь биосинтеза дхуррина, начинающийся с его аминокислоты - предшественника тирозина, реконструировали in vitro, объединяя Р450TYR, Р450ox, NADPH-P450-оксидоредуктазу в ДЛФХ-мицеллах с УДФГ-глюкозилтрансферазой, тирозином, NADPH, УДФГ и кастаноспермином. Тирозин превращался с помощью Р450TYR в пара-гидроксифенилацетальдоксим, который далее с помощью Р450ox превращался в пара-гидроксифенилацетонитрил и пара-гидроксибензальдегид. При этом происходило накопление небольших количеств пара-гидроксифенилацетонитрила, в то время как весь образовавшийся пара-гидроксиманделонитрил превращался в дхуррин и в несколько неидентифицированных соединений. В этой серии экспериментов стехиометрическое отношение P450TYR к Р450ox составляло примерно 15. Таким образом, не является неожиданным обнаружение накопления пара-гидроксифенилацетальдоксима в опыте по восстановлению. Обнаруженное накопление пара-гидроксифенилацетонитрила является неожиданным, поскольку предыдущие эксперименты с микросомами сорго показали, что пара-гидроксифенилацетонитрил мало накапливается и трудно улавливается. Накопление пара-гидроксифенилацетонитрила в экспериментах по восстановлению с выделенным Р450ox можно объяснить частичной денатурацией или инактивацией выделенного Р450ox.
4.6. Связывание с субстратами
Установление того факта, что Р450ox представляет собой многофункциональный фермент, превращающий пара-гидроксифенилацетальдоксим в пара-гидроксиманделонитрил с использованием в качестве промежуточного продукта пара-гидроксифенилацетонитрила, явилось предпосылкой к изучению способности Р450ox связываться с субстратами. Обратный спектр типа I с минимумом абсорбции при 381 нм и с максимум абсорбции при 418 нм был получен с использованием пара-гидроксифенилацетальдоксима, а это позволило предположить, что при добавлении субстрата происходит сдвиг от высокой величины спина к его низкой величине. Амплитуда увеличивалась при инкубации и достигала стабильного максимума после примерно 45 мин. При добавлении пара-гидроксифенилацетонитрила не обнаружено никакого спектра, характерного для связывания с субстратом.
Установлено, что Р450ox является существенно более лабильным по сравнению с другими ферментами Р450, выделенными из сорго. Выделенный Р450ox продуцировал обратный спектр типа I, характерный для связывания с субстратом, при инкубации с пара-гидроксифенилацетальдоксим. Коэффициент затухания Е420-390, соответствующий полному переходу от одной величины спина к другой, составлял 130 мМ-1см-1. В полученном при связывании с субстратом спектре максимальные амплитуды оказались примерно в 2 раза больше теоретически рассчитанных, даже с учетом полного сдвига от высокой величины спина к низкой величине спина. Это расхождение свидетельствует о том, что концентрация Р450ox была заниженной при количественной оценке пика, полученного при 450 нм в спектре связывания монооксида углерода. Альтернативно этому Р420-форма Р450ox обладает способностью связывать оксим и, таким образом, оказывает влияние на размер полученного при связывании с субстратом спектра. Последнее предположение может объяснить, почему максимальные амплитуды были получены только после пролонгированной инкубации.
Опосредованное Р450 расщепление альдоксимов с получением нитрилов ранее установлено с использованием микросом печени (DeMaster и др., J.Org.Chem. 5074-5075, 1992). Основное различие между Р450 из микросомальной системы печени и Р450ox состоит в том, что первый нуждается в строго анаэробных условиях, в то время как последний в аэробных условиях катализирует последующую реакцию С-гидроксилирования и преобразует в ходе обмена веществ как (Е)-, так и (Z)-изомер. В анаэробных условиях слабый пик спектра типа I получали при использовании микросом печени. При добавлении NADPH или дитионита возникал выраженный Soret-пик при 442-444 нм. Отсюда сделано заключение о том, что наиболее важные активные виды Р450 находятся в Fe (II)-состоянии. Исследования спектров Р450ox в анаэробных условиях не показали образования абсорбционного комплекса при 442 нм, но показали, что присутствие NADPH необходимо для каталитической активности, а это свидетельствует о том, что Р450ox также должен находиться в Fe (II-сосотоянии для того, чтобы опосредовать реакцию дегидратации.
Пример 5: Получение зонда цитохрома Р450 типа А
ПЦР проводили с использованием плазмидной ДНК, выделенной из однонаправленной библиотеки плазмидной кДНК (фирма Invitrogen), созданной из сильно этиолированных проростков Sorghum bicolor (L.) Moench высотой 1-2 см, с использованием в значительной степени вырожденных инозин (I)-содержащих праймеров, предпочтительно отобранных для цитохрома Р450 типа A (Nelson и Durst, Dgug Metabolism and Drug Interaction 12: 189-206 (1995)).
Праймер 1 (смысловая цепь) с последовательностью
5'-GCGGAATTCTTYIIICCNGARMGNTT-3' (SEQ ID NO: 5) перекрывает консенсусную аминокислотную последовательность FXPERF (SEQ ID NO: 6), где Х обозначает любую аминокислоту. Праймер 2 (антисмысловая цепь) с последовательностью 5'-GCGGATCCIIIRCAIIINCKNCKNCC-3' (SEQ ID NO: 7) перекрывает консенсусную аминокислотную последовательность GRRXCXG (SEQ ID NO: 8). Праймер 1 и праймер 2 были присоединены к сайтам EcoRl и BamHI соответственно для того, чтобы только ПЦР-продукты, полученные под контролем обоих праймеров, были клонированы в векторе pBluescript II SK (фирма Stratagene), расщепленном с помощью EcoRI/BamHI. ПЦР проводили в виде двух последовательных серий реакций. В первой серии реакций с помощью праймера 1 и стандартного праймера фага Т7, 5'-AATACGACTCACTATAG-3' (SEQ ID NO: 9), увеличивали пул кДНК, кодирующей цитохромы Р450 типа А. Во второй серии реакций с использованием праймера 1 и праймера 2 получали в основном одну полосу размером примерно 100 пар оснований, специфичную для цитохромов Р450 типа А. Первую серию ПЦР проводили в общем объеме 100 мкл, включающем 5% ДМСО, 200 мкМ дНТФ, 200 пмолей праймера 1, 100 пмолей стандартного праймера Т7, 2,5 ед. ДНК-полимеразы Taq в ПЦР-буфере и 1 мкл разбавленной в 100 раз плазмидной ДНК из библиотеки кДНК. Вторую серию ПЦР проводили в общем объеме 100 мкл, включающем 5% ДМСО, 200 мкМ дНТФ, 200 пмолей праймера 1 и праймера 2, 2,5 ед. ДНК-полимеразы Taq в ПЦР-буфере и 1 мкл продукта, полученного в первой серии ПЦР. В обеих сериях ПЦР за одним циклом продолжительностью 5 мин при 95°С следовало 35 циклов по 30 с при 95°С, 1 мин при 50°С и 30 с при 72°С. Полученный во второй серии ПЦР продукт размером примерно 100 пар оснований вырезали из 2%-ного агарозного геля и повторно амплифицировали перед клонированием в pBluescript. Из 19 секвенированных клонов 10 имели очень высокую идентичность на уровне аминокислотной последовательности с гидроксилазой коричной кислоты (CYP 74) и в дальнейшем не изучались. Сравнительный анализ остальных 9 последовательностей позволил разделить их на 2 группы, состоящие из 8 и 1 последовательности, которые были обозначены как "12" и "7" соответственно. Последовательность специфичного для гена "12" праймера, локализованного между праймером 1 и праймером 2:
5'-GCGGATCCGACTACTACGGCTCGC-3' (SEQ ID NO: 10), и праймер 5'-GCGGATCCTTTTTTTTTTTTTTTTV-3' (SEQ ID NO: 11), оба присоединены к BamHI, использовали для амплификации специфичного для гена "12" фрагмента размером примерно 500 пар оснований, полученного в первой серии ПЦР и клонированного в pBluescript. Аналогично этому специфичный для гена "7" фрагмент получали, используя специфичный для гена "7" праймер 5'-GCGGATCCGACATCAAGGGCAGCG-3' (SEQ ID NO: 12) и праймер 5'-GCGGATCCTTTTTTTTTTTTTTTTV-3' (SEQ ID NO: 11). Вставки метили с помощью дигоксигенина-11-дУТФ (фирма Boehringer Mannheim) путем ПЦР-амплификации с использованием стандартных праймеров Т7 и Т3 согласно инструкциям производителя и применяли для скрининга библиотеки кДНК.
Пример 6: Скрининг библиотеки и секвенирование ДНК
Все гибридизации на фильтре проводили с помощью DIG-системы (фирма Boehringer Mannheim). Получали съемы с колоний, используя найлоновые мембраны (фирма Boehringer Mannheim), и гибридизовали в течение ночи при 68°С в пятикратном растворе хлорида и цитрата натрия (5×РХЦН), дополненном 0,1% N-лауроилсаркозина, 0,02% ДСН, 1% блокирующего реагента (фирма Boehringer Mannheim). Фильтры перед исследованием промывали дважды в течение 15 мин в ОД×ЗХЦН, дополненном 0,1% ДСН, при 65°С. Как для "12", так и для "7" были получены полноразмерные клоны, что подтверждено анализом последовательностей. Секвенирование проводили с помощью набора для секвенирования Thermo Sequenase Fluorescent labelled Primer cycle sequencing kit (7-деаза-дГТФ) (фирма Amersham) и анализировали на устройстве ALF-Express (фирма Pharmacia). Компьютерный анализ последовательности проводили с помощью программ, входящих в пакет программ GCG Wisconsin SequenceAnalysis Package. Полноразмерная последовательность кДНК Р450ox и аминокислотная последовательность кодирующей области, полученной путем определения нуклеотидной последовательности "12", приведены в SEQ ID NO: 1 и SEQ ID NO: 2 соответственно.
Пример 7: Экспрессия в Е. coli
Экспрессирующий вектор pSP19g10L (Barnes, Method in Enzymology 272: 3-14, 1996) был получен от доктора Henry Barnes (Synthetic Genetics/Immune Complex Incorporation, San Diego, CA). Эта плазмида содержит промотор lacZ, слитый с короткой лидерной последовательностью гена 10 из бактериофага Т7, т.е. g10L, которая официально признана самой лучшей лидерной последовательностью для экспрессии различных гетерологичных протеинов (Olin и др., 1988). "7" и "12" модифицировали с помощью ПЦР с участием Pwo-полимеразы (фирма Boehringer Mannheim) для интродукции сайта NdeI в стартовый кодон и для замены стоп-кодона на стоп-кодон ochre непосредственно за сайтом HindlII. Для получения экспрессирующего клона "7" использовали праймер 3 (смысловая цепь) 5'-CGCGGATCCATATGGACGCATCATTACTCCTCTCCGTCGCGCTC-3' (SEQ ID NO: 13) и праймер 4 (антисмысловая цепь)
5'-CGCAAGCTTATTACATCTCAACGGGGACCCT-3' (SEQ ID NO: 14).
Праймер 3 интродуцирует молчащие мутации в кодоны 3, 4 и 5, уменьшая содержание G/C вокруг сайта инициации трансляции и сайта BamHI непосредственно против хода транскрипции от сайта NdeI. Полученный ПЦР-фрагмент расщепляли с помощью BamHI и HindIII, встраивали в pBluescript, расщепленном с помощью BamHI и HindIII, и контролировали посредством секвенирования с целью исключения ошибок ПЦР. Аналогично этому "12" интродуцировали в pBluescript, используя праймер 5 (смысловая цепь) 5'-CGCGGATCCATATGGCAACAACAGCAACCCCGCAGCTCCTC-3' (SEQ ID NO: 15) и праймер 6 (антисмысловая цепь) 5'-CGCAAGCTTATTATGCTGCGCGGCGGTTCTTGTATTTGG-3' (SEQ ID NO: 16). Праймер 5 интродуцирует молчащие мутации в кодоны 2, 3, 4 и 5, а праймер 6 интродуцирует молчащие мутации в 2 последних кодона, уменьшая содержание G/C. Вставки отщепляли с помощью NdeI и HindIII и встраивали в pSP19g10L, расщепленную с помощью NdeI и HindIII. Экспрессирующими плазмидами трансформировали клетки Е. coli JM109. Отдельные колонии выращивали в течение ночи при 37°С в LB-среде, содержащей 100 мг/мл ампициллина, и 5 мл культуры, выращенной в течение ночи, использовали для инокуляции 500 мл ТВ-среды, содержащей 50 мг/мл ампициллина, 1 мМ тиамин, 1 мМ изопропил-β-тиогалактопиранозид и 1 мМ β-аминолевулилиную кислоту. Клетки выращивали при 28°С в течение 48 ч при 125 об/мин. 1 мл культуры Е. coli, трансформированной экспрессирующими конструкциями "7", "12" и pSP19g10L, пеллетировали посредством центрифугирования (2000×g, 10 мин), промывали и концентрировали в 10 раз в 50 мМ Трицине, рН 7,9, и инкубировали с 14 нКи [U-14С]-пара-гидроксифенилацетальдегидоксимом с удельной активностью 394 мКи/ммоль при 30°С в течение 30 мин. Инкубационные смеси экстрагировали этилацетатом, наносили на пластинку для ТСХ (силикагель 60 F254, фирма Merck), проводя разделение с использованием смеси этилацетат/толуол (в объемном соотношении 1:5) в качестве подвижной фазы и визуализируя с помощью томографа типа STORM 840 фирмы Molecular Dynamics. Клон Е. coli, трансформированный конструкцией, экспрессирующей "12", оказался способен превращать пара-гидроксифенилацетальдегидоксим в пара-гидроксифенилацетонитрил.
СО-дифференциальные спектры солюбилизированных сферобластов Е. coli, экспрессирующих Р450ox, имели основной пик при 417 нм и минорный пик при 457 нм. В целом, СО-спектр с пиком абсорбции при длине волны, близкой к 420 нм, характерен для цитохрома Р450 в функционально неактивной конформации (Imai и др., Eur.J.Biochem. 1: 419-426, 1964). Присутствие основного пика при 417 нм позволяет предположить, что основная часть экспрессируемого цитохрома Р450 присутствовала в функционально неактивной конформации. Заметный сдвиг пика абсорбции с 450 нм к 457 нм может быть следствием присутствия больших количеств цитохрома Р450 в неконформационном состоянии. На основании величины пика при 457 нм установлено, что в течение 65 ч получено 50 нмолей Р450ox на 1 л культуры Е. coli. 5 мкл мембран, выделенных согласно методу, описанному у Halkier и др. в Archives of Biochemistry and Biophysics 322: 369-377, 1995, из клона Е. coli, экспрессирующего "12", восстанавливали с помощью 0,225 ед. NADPH-(цитохром Р450)-редуктазы, 50 мкг NADPH, 42 нКи пара-гидроксифенилацетальдегидоксима и 100 мкг дилаурилфосфатидилхолина в общем объеме 100 мкл 30 мМ Трицина, рН 7,9. После инкубации при 30°С в течение 30 мин реакционную смесь наносили на пластинку для ТСХ и анализировали согласно описанному выше методу. Восстановление мембран из клона Е. coli, экспрессирующего "12", привело к накоплению пара-гидроксибензальдегида, который представляет собой стабильный продукт диссоциации пара-гидроксиманделонитрила, последнего промежуточного продукта в пути биосинтеза цианогенного глюкозида дхуррина. Это свидетельствует о том, что "12" представляет собой цитохром Р450, который катализирует превращение пара-гидроксифенилацетальдегидоксима в пара-гидроксиманделонитрил. кДНК соответствует Р450ox. Клон, включающий описанную выше кДНК цитохрома Р450ox, был депонирован 10 января 1997 г. в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ, Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Mascheroder Weg Ib, D-38124 Braunschweig) под регистрационным номером DSM 11367.
Когда радиоактивно меченный пара-гидроксифенилацетальдоксим вводили в клетки Е. coil, трансформированные Р450ox, в клетках Е. coli, экспрессирующих Р450ox, накапливался пара-гидроксифенилацетонитрил. Установлено, что в клетках Е. coli, которые не содержат эндогенных цитохромов Р450 или NADPH-(цитохром Р450)-редуктазы, поддерживается каталитическая активность гетерологично экспрессируемых цитохромов Р450. Два растворимых флавопротеина Е. coli, а именно, флаводоксин и NADPH-флаводоксинредуктаза, отдают свои восстановительные эквиваленты рекомбинантным цитохромам Р450. Восстановление выделенных мембран Е. coli или очищенного рекомбинантного фермента с NАDРН-(цитохром Р450)-редуктазой из сорго приводило к превращению пара-гидроксифенилацетальдоксима в пара-гидроксиманделонитрил, в то время как экспрессирующие Р450ox клетки Е. coli преобразуют в ходе обмена веществ пара-гидроксифенилацетальдоксим только до пара-гидроксифенилацетонитрила. Отсутствие у Е. coli способности поддерживать превращение пара-гидроксифенилацетонитрила в пара-гидроксиманделонитрил является первым примером того, что система флаводоксин/NADPH-флаводоксинредуктаза Е. coli не способна поддерживать каталитическую активность в реакции с участием микросомального цитохрома Р450. Предварительные эксперименты по восстановлению с мембранами и растворимыми фракциями экспрессирующих Р450ox клеток Е. coli показали, что растворимая система флaвoдoкcин/NADPH-флaвoдoкcинpeдyктaзa поддерживают каталитическую активность Р450ox только в отношении превращения гидроксифенилацетальдоксима в пара-гидроксифенилацетонитрил и что ингибирующий фактор, препятствующий последующей реакции гидроксилирования, отсутствовал в растворимой фракции. Отсутствие у Е. coli способности поддерживать полную активность Р450ox может отражать нетипичную каталитическую реактивность Р450ox. В клетках Е. coli, трансформированных Р450ox или вектором экспрессии, накапливается соединение, обладающее несколько более низкой мобильностью по сравнению с пара-гидроксифенилацетальдоксимом. Это свидетельствует о том, что Е. coli может преобразовывать в ходе обмена веществ пара-гидроксифенилацетальдоксим независимо от присутствия Р450ox.
Пример 8: Очистка рекомбинантного Р450 (CYP71E1)
Сферобласты из 2 л культуры Е. coli, экспрессирующей Р450ox, подвергали индуцируемому температурой фазовому распределению с 1%-ным Тритоном X-114 согласно известному методу (Halkier и др., Archives of Biochemistry and Biophysics 322: 369-377, 1995). Содержащую Р450ox верхнюю фазу разбавляли в 100 раз с помощью 10 мМ КРi, рН 7,0, 0,05% восстановленного Тритона X-100, 1 мМ ДТТ, 0,5 мМ ФМСФ (фенилметилсульфонилфторид), значение рН доводили до 7,0 с помощью уксусной кислоты и вносили с большой скоростью потока на 16-миллилитровую колонку с СМ-сефарозой (фирма Pharmacia), уравновешенную буфером Г (10 мМ КРi, рН 7,0, 0,02% Тритона Х-114, 0,05% восстановленного Тритона Х-100, 10% глицерина, 1 мМ ДТТ, 0,5% ФМСФ). Колонку промывали буфером Г и Р450ox элюировали с помощью линейного градиента (0-1М) КСl (350 мл) в буфере Г. Объединенные содержащие Р450ox фракции (43 мл) использовали для экспериментов по восстановлению и электроэлюированный рекомбинантный Р450ox использовали для получения антител у цыплят.
Очищенный рекомбинантный Р450ox, восстановленный с помощью NADPH-(цитохром Р450)-редуктазы, в ДЛФХ катализировал превращение пара-гидроксифенилацетальдегидоксима в пара-гидроксиманделонитрил в присутствии NADPH, как описано в примере 4, раздел 4.2, касающемся очистки фермента растений.
Пример 9: Экспрессия дхуррина в трансгенных растениях Arabidopsis и табака
9.1. Конструирование векторных плазмид
Получали три бинарных вектора для опосредуемой Agrobacterium tumefaciens трансформации, а именно, pPZP111.79, pPZP221.71E1 и pPZP111.79.71E1.
Для конструирования pPZP111.79 клон кДНК Р450TYR (WO 95/16041; Koch и др., Arch. Biochem. Biophys. 323: 177-186, 1995) сначала отщепляли с помощью EcoRI и интродуцировали в сайт EcoRI pRT101 (Kopfer и др., Nucleic Acids Research 15: 5890, 1987) для функционального связывания кДНК с промотором 35S и сигналом полиаденилирования CaMV, получая плазмиду pRT101.79. Перед интродукцией кДНК Р450TYR часть полилинкера pRT101 удаляли путем расщепления с помощью SacI и XbaI с последующим повторным лигированием "затупленных" концов, полученных в результате обработки полимеразой Кленова, оставляя тем самым доступными только сайты EcoRI и XhoI. Из pRT101 с помощью SphI отщепляли клон Р450TYR, включающий промотор 35S и сигнал полиаденилирования CaMV. Для создания плазмиды pPZP111.79 концы "затупляли" с помощью полимеразы Кленова и полученный фрагмент встраивали в расщепленную с помощью EcoRI плазмиду PZP111 (Hajdukiewicz и др., Plant Mol. Biol. 25: 989-994, 1994), концы которой уже также были "затуплены" с помощью полимеразы Кленова и дефосфорилированы.
Для конструирования pPZP221.71E1 клон кДНК Р450 сначала отщепляли с помощью KpnI и XbaI и встраивали в сайты КрnI и XbaI pRT101, получая pRT101.71E1. Затем из pRT101.71E1 с помощью HindIII отщепляли клон Р450ox, включающий промотор 35S и сигнал полиаденилирования CaMV, и встраивали в сайт HindIII pZP221 (Hajdukiewicz и др., Plant Mol. Biol. 25: 989-994, 1994), получая таким образом pPZP221.71E1.
Для конструирования pPZP111.79.71E1 кДНК клона Р450ox, включающего промотор 35S и сигнал полиаденилирования CaMV, отщепляли от pPZP221.71E1 с помощью HindIII, "затупляли" концы с помощью полимеразы Кленова и лигировали в сайт SmaI pPZP111.79.
9.2. Трансформация Arabidopsis thaliana
Бинарные векторы pPZP111, pPZP221, pPZP111.79, pPZP221.71E1 и pPZP111.79.71El интродуцировали в штамм Agrobacterium tumefaciens C58C1/pGV3850 путем электропорации согласно методу, описанному Wenjun и Forde, Nucleic Acids Research 17: 8385, 1989. Экотип Arabidopsis thaliana Colombia трансформировали вакуумной фильтрацией в основном согласно методу, описанному у Bechtold и др., Molecular Biology and Genetics 316: 1194-1199. Трансформантов отбирали на MS-планшетах, содержащих либо 50 мкг/мл канамицина для векторов серий pPZP111, либо 200 мкг/мл гентамицинсульфата для векторов серий pPZP221. Через 4-6 недель после прорастания устойчивые к канамицину или гентамицину растения переносили в почву.
9.3. Трансформация Nicotiana tabacum cv Xhanti
Табак Nicotiana tabacum cv Xhanti в основном трансформировали методом листовых дисков, описанным у Svab и др. (Methods in Plant Molecular Biology, Cold Spring Harbor, стр. 55-60, 1995), используя штамм Agrobacterium tumefaciens C58C1/pGV3850, трансформированный либо pPZP111.79, либо pPZP221.71Е1, либо pPZP111.79.71E1. Для отбора клонов, трансформированных вектором pPZP111.79, использовали 100 мкг/мл канамицина, для отбора клонов, трансформированных вектором pPZP221.71Е1, использовали 100 мкг/мл гентамицинсульфата, и для отбора клонов, трансформированных вектором pPZP111.79.71E1, использовали 50 мкг/мл G-418. После образования корней растения табака переносили в почву и выращивали в теплице.
9.4. Определение дхуррина
Содержание дхуррина оценивали количественно с помощью метода спектрофотометрического определения цианида, описанного у Halkier и др. (Plant Physiol. 90: 1552-1559, 1989), за исключением того, что 5-10 мг листовой ткани замораживали и трижды подвергали оттаиванию перед добавлением 0,1 мг β-d-глюкозидазы типа II (фирма Sigma).
9.5. Анализ трансгенных растений Arabidopsis thaliana
В срезанные листья A. thaliana вводили радиоактивную метку, используя 2 мкл [U-14C]-тирозина (0,05 мкКи/мкл, 443 мКи/ммоль, фирма Amersham), и выдерживали в течение ночи в 100 мкл воды в закрытых пробирках Эппендорфа. Метаболиты экстрагировали с помощью кипящего 90%-ного метанола в течение 5 мин, экстракты концентрировали и наносили на пластинки для ТСХ (силикагель 60 F254). Метаболиты разделяли в трех различных системах растворителей в зависимости от их гидрофобности/гидрофильности. Система растворителей этилацетат/ацетон/дихлорметан/метанол/вода (в объемном соотношении 20/15/6/5/4) является предпочтительной для разделения различных цианогенных глюкозидов. Система растворителей изопропанол/этилацетат/вода (7/1/2) позволяет разделить различные глюкозинолаты. Система растворителей толуол/этилацетат (5/1) позволяет разделить гидрофобные промежуточные продукты. Радиоактивно меченные субстраты и продукты визуализировали с помощью томографа для выявления фосфора типа STORM 840 фирмы Molecular Dynamics.
Анализ с помощью ТСХ метанольных экстрактов листьев A. thaliana, трансформированных как Р450TYR, так и Р450ox с использованием A. tumefaciens C58C1/pGV3850/pPZP111.79.71E1, подтвердил присутствие нового соединения, которое мигрировало совместно с цианогенным глюкозидом дхуррином. Введение в срезанные листья радиоактивно меченного тирозина показало, что радиоактивная метка содержится в полосе, которая мигрирует вместе с дхуррином. Анализ всей листовой ткани трансформированных растений с помощью колориметрического метода определения цианидов (Lambert и др., Analytic Chemistry 47: 917-919, 1975) показал, что трансформанты Т1 содержали от 1 до 6 нмолей цианида/мг массы сырой ткани. Для сравнения: в контрольных растениях, трансформированных только nptII с использованием A. tumefaciens C58C1/pGV3850/pPZP111, содержание цианида составляло 1,2±0,35 нмоля цианида/мг массы сырой ткани. В литературе отсутствуют данные о том, что растения дикого типа A. thaliana содержат цианогенные глюкозиды, и заметные уровни цианида, обнаруженные в контрольных растениях, наиболее вероятно отражают присутствие тиоцианатов. Тиацианаты представляют собой продукты разложения глюкозинолатов и известно, что они дают искаженную реакцию при колориметрическом определении цианидов (Epstein, Analytic Chemistry 19: 272-274, 1974).
Обнаружение продуцирования больших количеств цианогенного глюкозида дхуррина в результате интродукции как Р450TYR, так и Р450ox, что подтверждено на примере A. thaliana, свидетельствует о присутствии соответствующей УДФГ-глюкозилтрансферазы, которая катализирует правильное глюкозилирование образованного пара-гидроксиманделонитрила. Поскольку отсутствуют данные о стереоспецифичности глюкозилтрансферазы, существует вероятность того, что полученный цианогенный глюкозид в действительности представляет собой таксифиллин, который является эпимером (оптический изомер) дхуррина.
Когда P450TYR интродуцировали с помощью A. tumefaciens C58C1/pGV3850/pPZP111.79, в A. thaliana накапливались большие количества соединения, которое мигрировало совместно с являющимся производным тирозина глюкозинолатом, пара-гидроксибензилглюкозинолатом. Это свидетельствует о том, что интродукция Р450TYR приводит к получению пара-гидроксифенилацетальдоксима из тирозина. Оксим, являющийся производным тирозина, затем далее превращается с помощью ферментов пути метаболизма глюкозинолата в пара-гидроксибензилглюкозинолат. Это является убедительным доказательством того, что ферменты, расположенные в прямом направлении на пути превращения оксима в глюкозинолат, обладают низкой субстратной специфичностью в отношении строения боковой цепи и что профиль глюкозинолата в целом определяется субстратной специфичностью первого цитохрома Р450 на пути биосинтеза.
Субстратная специфичность Р450ox не является столь узкой, как у P450TYR. Р450ox может преобразовывать в ходе обмена веществ оксимы, полученные из других аминокислот, примером которых является фенилацетальдоксим, т.е. оксим, являющийся производным фенилаланина, в то время как P450TYR участвует в метаболизме только производных тирозина. Следовательно, при интродукции Р450ox в растения, продуцирующие глюкозинолаты, можно ожидать, что в пути биосинтеза глюкозинолатов будет происходить накопление цианогенных глюкозидов, полученных из оксимов, являющихся производными аминокислот.
Поскольку P450TYR и Р450ox взаимодействуют с ферментами и предшественниками пути биосинтеза глюкозинолатов, можно ожидать, что глюкозинолатный профиль также будет измененным.
9.6. Анализ трансгенных Nicotiana tabacum cv Xhanti
Микросомы, выделенные из растений табака, трансформированных pPZP111.79 и функционально экспрессирующие CYP79, катализируют превращение тирозина в пара-гидроксифенилацетальдоксим. Метанольные экстракты собранных листьев табака, экспрессирующих CYP79, с введенным радиоактивным тирозином содержали три дополнительные меченые полосы по сравнению с растениями табака дикого типа при анализе с помощью ТСХ с использованием системы растворителей изопропанол/этилацетат/вода (7/1/2). Три дополнительные полосы на пластинках для ТСХ совместно мигрировали с мечеными полосами, обнаруженными в растениях дикого типа, в которые вводили радиоактивно меченный пара-гидроксифенилацетальдоксим. При анализе с использованием системы растворителей толуол/этилацетат (5/1) установлено, что одна из полос соответствовала пара-гидроксифенилацетальдоксиму, что видно по совместной миграции с чистыми стандартами. Идентифицировать две другие дополнительные полосы пока не удалось, но на основании данных о их подвижности в двух системах растворителей можно заключить, что они являются менее гидрофобными, чем пара-гидроксифенилацетальдоксим, и наиболее вероятно они представляют собой гликозилированные производные пара-гидроксифенилацетальдоксима. Анализ CYP79 в растениях показал, что CYP79 может функционально экспрессироваться в табаке и при этом образуется некоторое количество свободного пара-гидроксифенилацетальдоксима, но основная часть образовавшегося пара-гидроксифенилацетальдоксима далее преобразуется в ходе обмена веществ растениями.
Растения, экспрессирующие как Р450TYR так и Р450ox, могут быть получены путем скрещивания растения, экспрессирующего Р450TYR, с растениями, экспрессирующими Р450ox. В альтернативном варианте растения, экспрессирующие P450TYR или Р450ox, могут быть повторно трансформированы с помощью A. tumefaciens C58C1/pGV3850/pPZP221.71E1 или A. tumefaciens C58C1/pGV3850/pPZP111.79, используя то преимущество, что две конструкции цитохромов Р450 связаны с двумя различными, но не единственными маркерами, а именно, nptII и aacCI, позволяющими производить отбор по признаку устойчивости.
Пример 10: Экспрессия дхуррина в трансгенной кукурузе
10.1. Конструирование векторных плазмид
Для биобаллистической трансформации кукурузы с помощью Р450TYR или Р450ox создавали два следующих бинарных вектора pCIB 9842 и pCIB 9833.
pCIB 9842: Клон кДНК, кодирующей Р450TYR, клонированный в сайте EcoRI вектора pBluescript II SK, согласно методу, описанному в WO 95/16041, использовали для создания с помощью ПЦР сайта BamHI на стартовом кодоне ATG и сайта BglII на стоп-кодоне. BamHI-BglII-фрагмент, содержащий ген Р450TYR, клонировали в расщепленной с помощью BamHI и BglII плазмиде pCIB 9805, которая является производной растительного вектора экспрессии pUC19, сконструированного с помощью сайтов AflII/NotI/AscI, расположенных на расстоянии 256 пар оснований против хода транскрипции от сайта HindIII и на расстоянии 778 пар оснований по ходу транскрипции от сайта BglII, который содержит металлотионинподобный промотор, описанный в ЕР-А-452269, и терминатор 35S.
pCIB 9833: Клон кодирующей Р450ox кДНК из примера, клонированный в сайте NotI-BstXI вектора pcDNAII (фирма Invitrogen), использовали для создания сайта BamHI на стартовом кодоне ATG и сайта BglII на стоп-кодоне. BamHI-BglII-фрагмент, содержащий ген Р450, также клонировали в расщепленной с помощью BamHI и BglII плазмиде pCIB 9805.
10.2. Способы трансформации кукурузы
Инициировали развитие культур эмбриогенного каллюса типа I (Green и др., 1983; Wan и др., 1994) инбредной линии типа Lancaster из незрелых зародышей длиной 1,5-2,5 мм. Примерно через 14 дней после опыления выделяли в стерильных условиях зародыши из выращенных в теплице початков, поверхность которых предварительно стерилизовали, помещали на среду Дункана для инициации каллюса с 2% сахарозы и 5 мг/л хлорамбена и выращивали в темноте. Примерно через 14 дней из эксплантатов выделяли зародыши, дающие эмбрионогенные реакции, и помещали на поддерживающую среду Дункана с 2% сахарозы и 0,5 мг/л 2,4-Д. После выращивания в течение 4-8 недель с ежеденельными пересевами на свежую поддерживающую среду получали компактные эмбриогенные культуры высокого качества. Отбирали кусочки активно растущего эмбрионогенного каллюса в качестве ткани-мишени для введения генов. Примерно за 4 ч до введения генов кусочки каллюса помещали на пластины-мишени, содержащие поддерживающую среду с 12% сахарозы. Кусочки каллюса располагали по окружностям на расстоянии 8 и 10 мм от центра пластины-мишени. Плазмидную ДНК осаждали на золотые микроносители согласно методу, описанному в руководстве фирмы DuPont Biolistics. Для каждого из 6 обстрелов препаратом микроносителя использовали по 2-3 мкг каждой плазмиды. Гены вводили в ткань-мишень с помощью устройства PDS-1000He Biolistics. Характеристики настройки устройства типа Biolistics были следующими: 8 мм между разрушающимся диском и макроносителем, 10 мм между макроносителем и задерживающим экраном и 7 см между задерживающим экраном и мишенью. Каждую пластину-мишень обстреливали дважды, используя диски, разрушающиеся при давлении 650 фунтов/кв.дюйм. Между задерживающим экраном и тканью-мишенью помещали сетку из нержавеющей стали размером 200х200 (фирма McMaster-Carr, New Brunswick, NJ). Через 7 дней после введения генов кусочки ткани-мишени переносили из среды с высоким осмотическим давлением на среду для селекции, содержащую 100-120 мг/л глуфосината аммония (гербицид Баста). Из среды для селекции удаляли все аминокислоты. После выдерживания в течение 5-8 недель на среде с высоким уровнем селектирующего агента все выжившие колонии переносили на среду, содержащую 20 мг/л Баста. Эмбриогенный каллюс пересевали каждые 2 недели в течение 4-8 недель и затем переносили на модифицированную MS-среду без селектирующего агента, содержащую 3% сахарозы, 0,25 мг/л анцимидола и 0,5 мг/л кинетина, и помещали на свет. Анцимидол и кинетин удаляли через 2 недели. Регенерированные проростки с корнями или без корней переносили в контейнеры типа Magenta, содержащие MS-среду с 3% сахарозы, и в конечном счете получали небольшие растения с корнями, которые затем переносили в почву в теплицу.
Пример 11: Идентификация с помощью ПЦР гомологов P450TYR в видах, содержащих глюкозинолаты
Основываясь на компьютерном анализе последовательностей гомолога EST Р450TYR из Arabidopsis (регистрационный номер Т42902) и гомолога Р450TYR из Sinapis, создавали два вырожденных олигонуклеоидных праймера, которые позволяли амплифицировать ПЦР-фрагменты геномной ДНК из гомологов P450TYR из видов, содержащих глюкозинолаты. Праймер смысловой цепи 5'-GCGGAATTCAARCCIGARMGICAYYT-3' перекрывает консервативную аминокислотную последовательность KPERHL (SEQ ID NO: 18) и включает сайт клонирования EcoRI. Праймер 2 антисмысловой цепи 5'-GCGGATCCRCAICCICKYTTICCNGT-3' перекрывает консервативную аминокислотную последовательность TGKRGC (SEQ ID NO: 19) и включает сайт клонирования BamHI. ПЦР проводили с использованием геномной дНК, полученной с помощью набора для экстракции растительной ДНК Nucleon Phytopure Plant DNA Extraction Kit фирмы Amersham. ПЦР проводили в общем объеме 100 мкл, включающем 5% ДМСО, 200 мкМ дНТФ, по 200 пмолей каждого праймера, 2,5 ед. полимеразы Taq в ПЦР-буфере (50 мМ КСl, 10 мМ Трис-HCl (рН 8,8), 1,5 мМ MgCl2 и 0,1% Тритона Х-100), используя 1 мкг геномной ДНК из любого следующего вида растений: Sinapis alba, A. thaliana, Brassica napus, Tropaeolum majus или N. tabacum cv Xhanti. Проводили ПЦР, включающую 4 последовательные стадии:
стадия 1: 1 цикл 5 мин при 95°С;
стадия 2: 5 циклов по 30 с при 95°С, 30 с при 55°С, 30 с при 72°С;
стадия 3: 30 циклов по 30 с при 95°С, 30 с при 60°С, 30 с при 72°С;
стадия 4: 1 цикл 5 мин при 72°С.
С целью получения достаточных количеств продукта, дающего полосу, соответствующую примерно 100 парам оснований из Т. majus, стадия 2 может быть модифицирована и состоять из 5 циклов по 30 с при 95°С, 30 с при 50°С, 30 с при 71°С.
ПЦР-продукты очищали с помощью набора QIAquick PCR Purification Kit (фирма Qiagen), расщепляли с помощью рестриктаз EcoRI и BamHI и разделяли на 3%-ном ТАЕ-агарозном геле. Вырезали основную полосу длиной примерно 100 пар оснований и встраивали в линеаризованный с помощью EcoRI/BamHI вектор pBluescript II SK (фирма Stratagene). Приблизительно 10 клонов каждого из 5 видов растений секвенировали, используя набор Thermo Sequenase Fluorescent labelled Primer cycle kit (7-деаза-дГТФ (фирма Amersham)) и анализировали на устройстве ALF-Express (фирма Pharmacia).
Из четырех содержащих глюкозинолаты видов S. alba, A. thaliana, В. napus и Т.majus выделены ПЦР-фрагменты, кодирующие консервативную аминокислотную последовательность:
KPERH(L/F)NECSEVTLTENDLRFISFSTGKRGC (SEQ ID NO: 20 и 21 соответственно). Эта консенсусная аминокислотная последовательность идентична ранее идентифицированным гомологичным Р450TYR последовательностям из S. alba и A. thaliana и имеет выраженное сходство с аминокислотной последовательностью Р450TYR из сорго. Из не содержащего глюкозинолаты растения N. tabacum cv Xhanti не был выделен ПЦР-фрагмент, кодирующий эту консенсусную последовательность. Присутствие этой консенсусной аминокислотной последовательности, гомологичной Р450TYR, в приведенных в качестве примера четырех видах растений, содержащих глюкозинолаты, свидетельствует о том, что аминокислотная N-гидроксилаза цитохрома Р450 из семейства Р450TYR в содержащих глюкозинолаты видах превращает аминокислоту-предшественник или аминокислоты-предшественники с удлиненной цепью в соответствующие оксимы. Получение ПЦР-фрагментов, специфичных для гомологов Р450TYR, позволяет выделить гомологичные кДНК или геномные клоны из соответствующих библиотек.
Хотя выше изобретение подробно рассмотрено на примерах конкретных вариантов его осуществления, следует отметить, что в него могут быть внесены различные изменения и модификации, не выходящие за объем формулы изобретения.

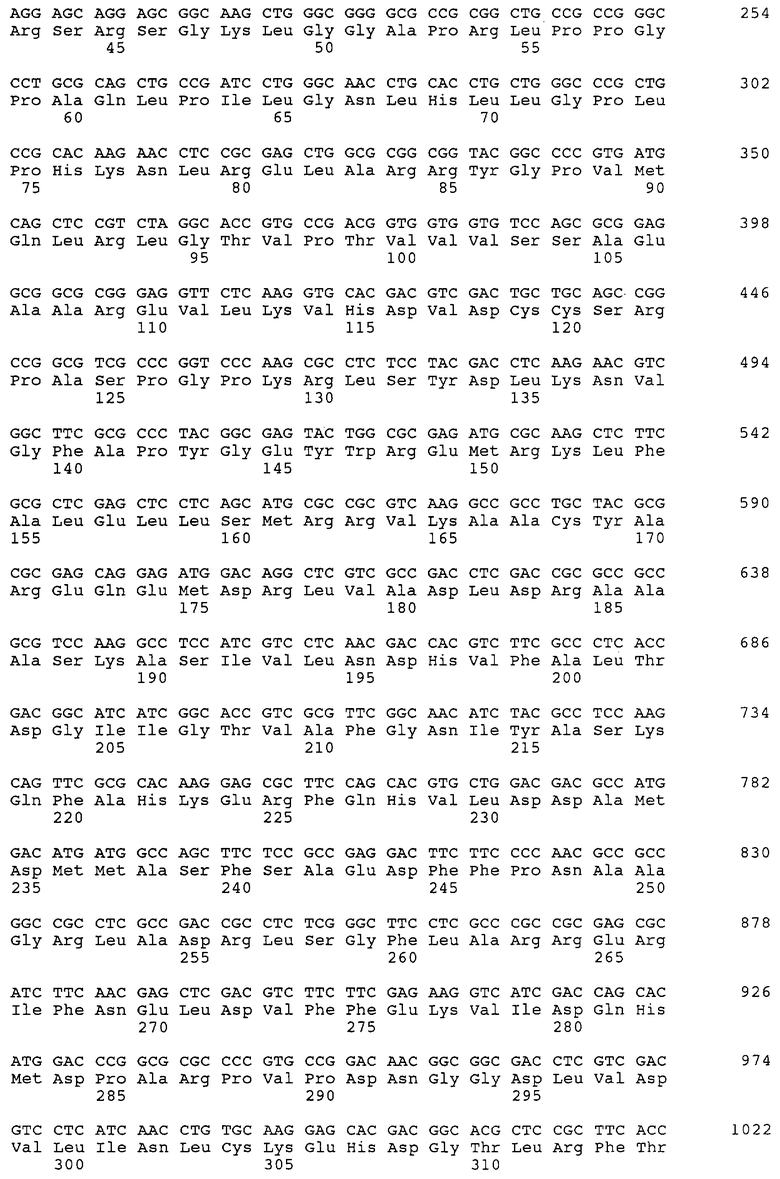










Изобретение относится к генетической инженерии и может быть использовано в селекции растений. Трансформация растительной клетки или ткани молекулой ДНК, кодирующей монооксигненазу, которая катализирует превращение альдоксима в нитрил и этого нитрила в соответствующий циангидрин, позволяет получить растение с измененным уровнем экспрессии цианогенных гликозидов. Уровень экспрессии этих веществ влияет на питательную ценность растения или их устойчивость к вредителям. 5 н. и 4 з.п. ф-лы.
где эти клоны включают молекулу кДНК, которая кодирует фермент, катализирующий превращение альдоксима в нитрил и превращение этого нитрила в соответствующий циангидрин.
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| RU 2070930 С, 27.12.1996. | |||

