Область техники, к которой относится изобретение
Настоящее изобретение обеспечивает последовательности нуклеиновых кислоты и аминокислотные последовательности цитохрома Р450, способного окислять молекулы терпенов. Оно также обеспечивает способ окисления молекул терпенов, включающий взаимодействие цитохрома Р450 изобретения с молекулой терпена, предназначенной для окисления. В частности, указанный способ может быть осуществлен in vitro или in vivo для получения окисленных молекул терпенов, которые могут быть применены в различных областях техники, таких, например, как парфюмерия и получение ароматических веществ. Настоящее изобретение также обеспечивает вектор экспрессии, содержащий нуклеиновую кислоту. Организм-хозяин или клетка-хозяин, не представляющие собой организм или клетку человека, трансформированные нуклеиновой кислотой, также представляют собой объект изобретения.
Уровень техники
Терпены были обнаружены в большинстве организмов (микроорганизмы, животные и растения). Данные соединения состоят из пяти углеродных единиц, называемых звеньями изопрена, и их классифицируют по числу данных звеньев, присутствующих в их структурах. Таким образом, монотерпены, сесквитерпены и дитерпены представляют собой терпены, содержащие 10, 15 и 20 атомов углерода, соответственно. Дитерпены, например, широко распространены в царстве растений, и было описано свыше 2500 дитерпеновых структур (Connolly and Hill, Dictionary of terpenoids, 1991, Chapman & Hall, London). Молекулы терпенов и их окисленные производные вызывали интерес в течение тысяч лет, из-за их вкусовых и ароматических свойств и их косметических, лекарственных и противомикробных эффектов. Растительные экстракты, полученные с помощью различных подходов, таких как перегонка с водяным паром или экстракция растворителем, применяли в качестве источника окисленных производных молекул терпенов. Альтернативно, молекулы терпенов, обнаруженные в растительных экстрактах или полученные с помощью биосинтетических способов, окисляли, применяя химические и энзиматические способы.
В процесс энзиматического окисления терпенов часто вовлечены ферменты, называемые цитохромы Р450 (Р450), которые, как правило, способны катализировать превращение гидрофобного субстрата, такого как молекула терпена, в более гидрофильную молекулу. Ферменты цитохром Р450 образуют суперсемейство гемопротеинов, которое обнаруживают у бактерий, архей и эукариот. В одной из наиболее распространенных активностей цитохром Р450 действует как монооксигеназа, вставляя один атом кислорода молекулярного кислорода в молекулу субстрата, тогда как другой атом кислорода восстанавливается до воды.
Для протекания этой каталитической реакции необходимо два электрона для активации молекулярного кислорода. Р450 из эукариот используют NADPH в качестве внешнего восстановителя и источника электронов. Два электрона переносятся поочередно к активному центру цитохрома Р450, и для этого переноса необходима цитохром Р450 редуктаза (CPR), представляющая собой белок-донор электронов. Одна CPR не специфична для одного цитохрома Р450. CPR представляет собой белок-донор электронов для нескольких Р450 в данном организме. Кроме того, CPR из одного организма может действовать как белок-донор электронов для Р450 из других организмов. В некоторых случаях Р450 также может быть сопряжен с белком цитохром b5, который может действовать как белок-донор электронов или может улучшать эффективность переноса электронов от CPR к Р450. В эукариотических клетках и, в особенности, в растениях, Р450 и CPR обычно представляют собой мембранно-связанные белки, которые ассоциированы с эндоплазматическим ретикулумом. Данные белки заякорены в мембране с помощью N-концевой трансмембранной спирали.
Многие из Р450 имеют низкую субстратную специфичность и, следовательно, способны катализировать окисление многих разнообразных структур, таких, например, как различные молекулы терпенов. Большинство из данных ферментов имеют определенную регио- и стереоселективность для данного субстрата, но они часто продуцируют смесь из нескольких продуктов из определенного субстрата. Такие Р450 обычно вовлечены в процессы распада и детоксикации таких молекул, как ксенобиотики, и их обычно обнаруживают в бактериях и у животных. С другой стороны, Р450, вовлеченные в биосинтетические пути, обычно демонстрируют специфичность для субстратов определенных типов и регио- и стереоселективность. Это справедливо для большинства растительных Р450.
Большое число Р450 может быть обнаружено в природе и, в особенности, в растениях. Один растительный геном может содержать несколько сотен генов, кодирующих Р450. Многие растительные Р450 охарактеризованы, но считается, что множество функций очень большого числа присутствующих в растения Р450, остаются неизвестными.
Следовательно, целесообразно проводить поиск новых Р450, способных катализировать новые энзиматические реакции, с тем, чтобы обеспечить энзиматическую продукцию новых оксигенированных соединений или для получения оксигенированных соединений в реакциях различных типов, например, из различные субстратов, которые могут оказаться более легко доступными.
Некоторые из Р450 уже охарактеризованы. Сообщалось, что в особенности, цитохромы Р450, обладающие определенным процентом идентичности последовательности с цитохромом Р450 настоящего изобретения, используют молекулы терпенов в качестве субстратов.
Цитохромы Р450, наиболее близкие к Р450 настоящего изобретения, представляют собой Р450 из Sorghum bicolor, среди них наиболее близкая последовательность обладает 67% идентичностью с аминокислотными последовательностями, описанными в настоящем документе (номер доступа EER94164).
Некоторые из оксигенированных терпенов, продуцируемых цитохромом Р450 настоящего изобретения, очень полезны в областях производства парфюмерных изделий и ароматизаторов. В особенности кусимол, который получают путем гидроксилирования зизаена, представляет собой один из ключевых компонентов масла ветивера и сам по себе служит ценным ароматизирующим ингредиентом. Окисление зизаена с применением цитохрома Р450 настоящего изобретения обеспечивает выгодную альтернативу выделению кусимола из масла ветивера, представляющему собой трудный и дорогостоящий способ. Насколько нам известно, энзиматический способ производства кусимола не известен. Некоторые другие ценные парфюмерные и ароматизирующие ингредиенты, для которых в настоящее время не известен энзиматический синтез, могут быть получены с применением цитохрома Р450 настоящего изобретения как будет описано ниже.
Другие оксигенированные терпены, продуцируемые цитохромом Р450 настоящего изобретения, полезны для других целей, таких как производство лекарственных препаратов или агрохимических продуктов. Цитохром Р450 настоящего изобретения, следовательно, открывает новый биосинтетический путь к разнообразным молекулам, обладающим интересными свойствами, полезным в различных областях промышленности и которые трудно или даже невозможно изолировать из природных материалов и трудно или невозможно получить с помощью органического синтеза.
Цель настоящего изобретения заключается в обеспечении способов получения оксигенированных терпенов, в особенности, кусимола, экономичным способом. Соответственно, цель настоящего изобретения заключается в производстве оксигенированных терпенов, сопровождающемся образованием небольшого количества отходов, более энерго- и ресурсосберегающим способом и при одновременном снижении зависимости от ископаемых видов топлива. Дополнительная цель заключается в обеспечении ферментов, способных окислять молекулы терпенов, окисленные таким способом продукты полезны в качестве ингредиентов при производстве парфюмерии и/или ароматических веществ.
Примененные сокращения
Описание изобретения
Настоящее изобретение обеспечивает способ энзиматического окисления терпенов экономически выгодным, надежным и воспроизводимым образом.
Как предполагается в настоящей заявке, все соединения, приведенные в настоящей заявке, определяют по их формулам, как показано на фигуре 1.
Термины «цитохром Р450» или «полипептид, обладающий активностью цитохрома Р450» предназначен для целей настоящей заявки в качестве полипептида, способного катализировать окисление молекулы терпена с образованием оксигенированного соединения, такого как спирт, альдегид, кетон или карбоновая кислота. В соответствии с предпочтительным воплощением, цитохром Р450 действует как монооксигеназа путем добавления только одного атома кислорода соединению терпена. Способность полипептида катализировать окисление определенного терпена может быть просто подтверждена путем осуществления ферментного анализа, подробно описанного в примере 8.
В соответствии с настоящим изобретением, термин «полипептиды» также подразумевает включение усеченных полипептидов при условии, что они сохраняют свою цитохром Р450 активность, как определена в любом из воплощений изобретения, и что они обладают, по меньшей мере, определенным процентом идентичности с соответствующим фрагментом последовательностей SEQ ID NO: 1 или 2.
Процент идентичности между двумя пептидными или нуклеотидными последовательностями представляет собой функцию от числа остатков аминокислот или нуклеиновых кислот, которые идентичны в двух последовательностях, после выравнивания данных двух последовательностей. Идентичные остатки определяют как остатки, одинаковые в двух последовательностях в данном положении выравнивания. Процент идентичности последовательности, как применен в настоящем документе, рассчитывают из оптимального выравнивания, деля число остатков, идентичных между двумя последовательностями, на общее число остатков в самой короткой последовательности и умножая полученное значение на 100. Оптимальное выравнивание представляет собой выравнивание, в котором процент идентичности представляет собой максимально возможный. Для получения оптимального выравнивания в одну или в обе последовательности, в одном или в нескольких положениях выравнивания могут быть введены пробелы. Данные пробелы затем учитывают как неидентичные остатки для расчета процента идентичности последовательности.
Выравнивание с целью определения процента идентичности последовательности аминокислот или последовательности нуклеиновой кислоты может быть достигнуто разными путями с помощью компьютерных программ и, например, с помощью публично доступных компьютерных программ, доступных во всемирной сети. Предпочтительно, для получения оптимального выравнивания пептидной или нуклеотидной последовательности и для расчета процента идентичности последовательности может быть применена программа BLAST (Tatiana et al., FEMS Microbiol Lett., 1999, 174: 247-250, 1999) с установленными по умолчанию параметрами, доступная в Национальном центре биотехнологической информации (NCBI) на сайте http://www.ncbi.nlm.nih.gov/BLAST/bl2seq/wblast2.cgi.
Одним из объектов настоящего изобретения является полипептид, включающий аминокислотную последовательность, по меньшей мере, на 70% идентичную последовательностям SEQ ID NO: 1 или 2, и обладающий активностью цитохрома Р450.
В предпочтительном воплощении назначение полипептида, обладающего активностью цитохрома Р450, заключается в способности катализировать окисление, по меньшей мере, одного соединения терпена, которое выбирают из моно- или полициклических монотерпенов и сесквитерпенов. В предпочтительном воплощении, указанный сесквитерпен или монотерпен включает, по меньшей мере, одну метильную группу в качестве заместителя в циклической части молекулы. В соответствии с более предпочтительным воплощением, цитохром Р450 изобретения окисляет указанный метильный заместитель для обеспечения первичного спирта.
В соответствии с предпочтительным воплощением, соединение терпена представляет собой соединение, которое выбирают из группы, состоящей из зизаена, альфа-цедрена, альфа-лонгипинена, альфа-фунебрена, туйопсена, валенсена, бета-чамигрена, аллоаромадендрена, альфа-неокловенена, изосативена, ледена, s-лимонена, альфа-гумулена, альфа-гурюнена, альфа-пинена, бета-фунебрена, R-лимонена и бета-пинена. Более предпочтительно указанное соединение терпена представляет собой терпен, который выбирают из зизаена, альфа-цедрена, альфа-фунебрена, валенсена и туйопсена. Наиболее предпочтительно, указанное соединение терпена представляет собой зизаен.
В предпочтительном воплощении, один атом кислорода добавляют к метильной группе так, чтобы обеспечить первичный спирт, альдегид и/или карбоновую кислоту. В наиболее предпочтительном воплощении, зизаен окисляется до кусимола, зизаналя и/или зизановой кислоты.
В случае если формируется альдегид и/или карбоновая кислота, указанный альдегид и/или карбоновая кислота формируется путем дальнейшего окисления первичного спирта или под действием Р450 изобретения, или под действием одного или нескольких ферментов из других семейств, таких, например, как алкогольдегидрогеназы, альдегидредуктазы, альдегид оксидазы. Последние ферменты, например, присутствуют в любом организме-хозяине или в клетке-хозяине, в которых полипептид изобретения может быть экспрессирован.
В соответствии с предпочтительным воплощением, полипептид включает аминокислотную последовательность, по меньшей мере, на 75%, предпочтительно, по меньшей мере, на 80%, предпочтительно, по меньшей мере, на 85%, предпочтительно, по меньшей мере, на 89%, предпочтительно, по меньшей мере, на 90%, более предпочтительно, по меньшей мере, на 95% и еще более предпочтительно, по меньшей мере, на 98% идентичную последовательностям SEQ ID NO: 1 или 2. В соответствии с более предпочтительным воплощением, полипептид включает SEQ ID NO: 1 или 2. Еще более предпочтительно, он состоит из последовательностей SEQ ID NO: 1 и 2.
В предпочтительном воплощении изобретения последовательность также включает последовательность мембранного якоря. Последовательность, представленная последовательностями SEQ ID NO: 1 или 2, или их производными, и имеющая необходимый процент идентичности представляет собой часть полипептида, который обеспечивает Р450 активность. Последовательность мембранного якоря не вовлечена в каталитическую активность фермента. Якорная последовательность делает возможным связывание с мембраной. Подходящие якорные последовательности зависят от организма, в котором экспрессируется полипептид, и последовательности, сконструированные для распространенных типов организмов-хозяев, известны специалисту в этой области техники. Любые подходящие якорные последовательности могут быть применены в комбинации с полипептидом настоящего изобретения. Следовательно, в соответствии с одним из предпочтительных воплощений, полипептид включает последовательности SEQ ID NO: 1 или 2, скомбинированные с последовательностью мембранного якоря.
Более предпочтительно, полипептид изобретения состоит из последовательностей SEQ ID NO: 1 или 2, необязательно скомбинированных с последовательностью мембранного якоря.
Если полипептид не скомбинирован с якорной последовательностью, такой полипептид не связывается с клеточной мембраной. В этом случае, полипептидные последовательности SEQ ID NO: 1 или 2 предпочтительно могут быть модифицированы для того, чтобы улучшить его растворимость в цитоплазме.
В соответствии с другим предпочтительным воплощением, полипептид включает аминокислотную последовательность, которая представляет собой вариант последовательностей SEQ ID NO: 1 или 2, полученных с помощью генной инженерии. В других терминах, указанный полипептид включает аминокислотную последовательность, кодируемую нуклеотидной последовательностью, которая была получена с помощью модификации последовательностей SEQ ID NO: 3, 4 или их комплементарной цепи. В соответствии с более предпочтительным воплощением, полипептид, обладающий активностью цитохрома Р450, состоит из аминокислотной последовательности, которая представляет собой вариант последовательностей SEQ ID NO: 1 или 2, полученный с помощью генной инженерии, т.е. аминокислотную последовательность, кодируемую нуклеотидной последовательностью, которая была получена с помощью модификации любой из последовательностей SEQ ID NO: 3, 4 или их комплементарной цепи.
Полипептиды, кодируемые нуклеиновой кислотой, полученной с помощью природной или искусственной мутации нуклеиновой кислоты изобретения, как будет описано в дальнейшем, также охвачены изобретением.
Варианты полипептидов, полученные в результате слияния дополнительных пептидных последовательностей на амино- и на карбокси-терминальных концах, также охвачены полипептидами изобретения. В особенности, такое слияние может усилить экспрессию полипептидов, полезно при очистке белка, улучшает путь, с помощью которого полипептид может быть заякорен в мембране или улучшает энзиматическую активность полипептида в желаемом окружении или в системе экспрессии. Такие дополнительные пептидные последовательности могут представлять собой, например, сигнальный пептид. Соответственно, настоящее изобретение охватывает варианты полипептидов изобретения, такие как полипептиды, полученные с помощью слияния с другими олиго- или полипептидами, и/или такие как полипептиды, соединенные с сигнальными пептидами. Полипептиды, полученные в результате слияния с другим функциональным белком, таким как белок пути биосинтеза терпенов, предпочтительно, терпен-синтазой, также охвачены полипептидами изобретения. Один особенно предпочтительный пример полипептида изобретения, представляющий собой вариант, полученный в результате слияния с пептидной последовательностью, - это слитый полипептид, включающий как полипептид изобретения (обладающий активностью цитохрома Р450), так и CPR.
В соответствии с другим воплощением, полипептид выделяют из Vetiveria zizanioides (L.) Nash.
Нуклеиновая кислота, кодирующая полипептид в соответствии с любым из вышеописанных воплощений, также представляет собой цель настоящего изобретения.
В соответствии с предпочтительным воплощением, нуклеиновая кислота включает нуклеотидную последовательность, по меньшей мере, на 70%, предпочтительно, по меньшей мере, на 75%, предпочтительно, по меньшей мере, на 80%, предпочтительно, по меньшей мере, на 85%, предпочтительно, по меньшей мере, на 90%, предпочтительно, по меньшей мере, на 93%, более предпочтительно, по меньшей мере, на 95% и еще более предпочтительно, по меньшей мере, на 98% идентичную последовательностям SEQ ID NO: 3, 4 или их комплементарной цепи. В соответствии с более предпочтительным воплощением, нуклеиновая кислота включает SEQ ID NO: 3, 4 или их комплементарные цепи. В соответствии с еще более предпочтительным воплощением, нуклеиновая кислота состоит из последовательностей SEQ ID NO: 3, 4 или их комплементарных цепей, необязательно вместе с нуклеотидной последовательностью, кодирующей последовательность мембранного якоря.
В соответствии с другим воплощением, нуклеиновую кислоту выделяют из Vetiveria zizanioides (L.) Nash.
Нуклеиновая кислота изобретения может быть определена как включающая дезоксирибонуклеотидные или рибонуклеотидные полимеры как в одно- или в двухцепочечной форме (ДНК и/или РНК). Термины «нуклеотидная последовательность» также следует понимать как включающие молекулу полинуклеотида или молекулу олигонуклеотида в форме отдельного фрагмента или как составную часть более крупной нуклеиновой кислоты. Нуклеиновые кислоты изобретения также включают определенные изолированные нуклеотидные последовательности, включающие последовательности, которые практически не загрязнены эндогенным материалом. Нуклеиновая кислота изобретения может быть усеченной, при условии, что она кодирует полипептид, охваченный настоящим изобретением, как описано выше.
В соответствии с более предпочтительным воплощением, по меньшей мере, одна нуклеиновая кислота в соответствии с любым описанным выше воплощением включает нуклеотидную последовательность, которая была получена с помощью модификации SEQ ID NO: 3, 4 или их комплементарной цепи. Предпочтительно, указанная нуклеиновая кислота состоит из нуклеотидной последовательности, которая была получена с помощью модификации SEQ ID NO: 3, 4 или их комплементарной цепи.
Нуклеиновые кислоты, включающие последовательность, полученную с помощью мутации последовательности SEQ ID NO: 3, 4 или их комплементарной цепи, охвачены изобретением, при условии, что последовательности, которые они включают, обладают, по меньшей мере, определенным процентом идентичности с соответствующими фрагментами последовательности SEQ ID NO: 3, 4 или их комплементарной цепи, и при условии, что они кодируют полипептид, обладающий активностью цитохрома Р450, как определено любым из вышеописанных воплощений. Мутации могут представлять собой любой тип мутаций данных нуклеиновых кислот, такие как точечные мутации, делеционные мутации, вставочные мутации и/или мутации сдвига рамки считывания. Вариант нуклеиновой кислоты может быть получен с целью адаптации ее нуклеотидной последовательности к специфической системе экспрессии. Например, известно, что бактериальные системы экспрессии более эффективно экспрессируют полипептиды, если аминокислоты кодируются предпочтительным кодоном. Благодаря вырожденности генетического кода, в котором более чем один кодон может кодировать одну и ту же аминокислоту, множество последовательностей ДНК могут кодировать один и тот же полипептид, все данные последовательности ДНК охвачены изобретением.
Настоящее изобретение также обеспечивает способ окисления, по меньшей мере, одного соединения терпена, включающий
a) взаимодействие указанного соединения терпена, по меньшей мере, с одним полипептидом изобретения в присутствии цитохрома Р450 редуктаза (CPR);
b) необязательно, выделение окисленного терпена, полученного на стадии а).
Соединение терпена, окисленное полипептидом изобретения, и сам полипептид изобретения, представляют собой такое соединение терпена и такой полипептид как определено по любому из вышеописанных воплощений.
Способ может быть осуществлен как in vitro, так и in vivo, как будет объяснено подробно далее.
Если способ осуществляют in vitro, то полипептид изобретения, предназначенный для взаимодействия с соединением терпена и CPR, может быть получен с помощью экстракции из любого экспрессирующего его организма с помощью стандартных методик экстракции белков или ферментов. Если организм-хозяин представляет собой одноклеточный организм или клетку, высвобождающий полипептид изобретения в культуральную среду, например, если отсутствуют мембранные якори, то полипептид может быть просто собран из культуральной среды, например, центрифугированием, необязательно, с последующими стадиями промывки и повторного суспендирования в подходящих буферных растворах. Если организм или клетка накапливают полипептид внутри своих клеток, то полипептид может быть получен с помощью разрушения или лизиса клеток и последующей экстракции полипептида из клеточного лизата. Если Р450 и CPR, такие как природные Р450 и CPR в растениях, включают последовательность мембранного якоря, то они ассоциированы с мембранами и, следовательно, локализованы в мембранной фракции клеточных лизатов. Мембранная фракция (микросомы) может быть легко отделена от других белковых фракций с помощью различных методик центрифугирования грубого клеточного лизата известными способами.
Для способа in vitro полипептид изобретения и CPR могут быть обеспечены независимо в изолированной форме или как часть белкового экстракта и суспендированы в буферном растворе при оптимальном pH. Если необходимо, то для оптимизации ферментативной активности могут быть добавлены соли, DTT, NADPH, NADH, FAD, FMN и другие вид энзиматических кофакторов. Подходящие условия будут описаны более подробно в приведенных далее примерах.
Соединение терпена затем добавляют к суспензии или к раствору, которые затем инкубируют при оптимальной температуре, например, от 15 до 40°C, предпочтительно, от 25 до 35°C, более предпочтительно, при 30°C. После инкубации, полученный окисленный терпен может быть изолирован из инкубируемого раствора с помощью стандартных процедур выделения, таких как экстракция растворителем и перегонка, необязательно, после удаления полипептидов из раствора.
При взаимодействии Р450 и соединения терпена должна присутствовать CPR.
В соответствии с другим предпочтительным воплощением, способ окисления соединения терпена осуществляют in vivo. В этом случае, стадия а) описанного выше способа включает культивирование организма-хозяина или клетки-хозяина, не представляющих собой организм или клетку человека, трансформированных для экспрессии, по меньшей мере, одного полипептида изобретения в присутствии соединения терпена для окисления в условиях, способствующих окислению соединения терпена, указанный организм или клетка дополнительно экспрессирующие CPR.
Соединение терпена и полипептид представляют собой такие соединение терпена и полипептид как определено в любом из воплощений настоящего изобретения.
В одном из воплощений такого способа, соединение терпена, предназначенное для окисления, продуцируется в организме-хозяине или в клетке-хозяине, которые экспрессируют полипептид изобретения. В этом случае, соединение терпена продуцируется в организме-хозяине или в клетке-хозяине под действием терпен-синтазы, способной катализировать образование указанного соединения терпена из нециклического предшественника терпена. Указанная терпен-синтаза может быть или образована естественным образом организмом-хозяином или клеткой-хозяином, или если организм-хозяин или клетка-хозяин не экспрессируют такой терпен-синтазы естественным образом, то они могут быть трансформированы так, чтобы они могли делать это.
В альтернативном воплощении, в случае, если применяют клетку-хозяина или если организм-хозяин представляет собой микроорганизм, то соединение терпена, предназначенное для окисления, могут добавлять к культуральной среде указанной клетки или микроорганизма. Соединение терпена будет проникать через мембрану клетки или микроорганизма, и таким образом, получать возможность вступать в реакцию с полипептидом изобретения, экспрессируемым указанной клеткой-хозяином или микроорганизмом.
В соответствии с более предпочтительным воплощением, способ дополнительно включает, перед стадией а), трансформирование организма или клетки, не представляющих собой организм или клетку человека, по меньшей мере, одной нуклеиновой кислотой изобретения, так чтобы указанный организм или клетка экспрессировали, по меньшей мере, один полипептид изобретения. Полипептид и нуклеиновая кислота были такими, как определено в любом из вышеописанных воплощений.
Особенно выгодно осуществлять способ in vivo, поскольку возможно осуществлять способ без предварительного выделения полипептида. Реакция протекает непосредственно внутри организма или клетки, трансформированных для экспрессии полипептида.
Для проявления каталитической активности Р450 должен быть применен в комбинации с Р450-редуктазой (CPR), которая способна переносить электроны от NADPH (никотинамидадениндинуклеотид фосфат, восстановленная форма) к активному центру Р450, таким образом, чтобы реконструировать активность Р450. CPR должна присутствовать для осуществления как способа in vitro, так и способа in vivo. Если способ осуществляют in vivo, CPR может либо присутствовать естественным образом в организме-хозяине или клетке-хозяине, или такие организм или клетка могут быть трансформированы для экспрессии CPR перед трансформацией, одновременно с трансформацией или после трансформации, проведенной для экспрессии полипептида изобретения. В предпочтительном воплощении изобретения клетку-хозяина или организм трансформируют слитым полипептидом, включающим как полипептид изобретения, так и CPR.
В другом предпочтительном воплощении CPR представляет собой растительную CPR. Наиболее предпочтительно ее получают из CPR из Arabidopsis thaliana.
Организм или клетка, не относящиеся к человеку, могут быть успешно дополнительно трансформированы, по меньшей мере, одним геном, кодирующим полипептид, вовлеченный в метаболизм, приводящий к образованию ациклических предшественников терпенов, таких как геранилпирофосфат, фарнезилпирофосфат или геранилгеранилпирофосфат. Такие полипептиды включают, например, ферменты МЕР-пути, MVA-пути и/или пренилтрансферазы.
Трансформирование организма или клетки, не представляющих собой организм или клетку человека, полипептидом, обладающим активностью цитохрома Р450, и трансформирование CPR, или трансформирование слитым полипептидом, включающим оба полипептида, в присутствии предназначенного для окисления соединения терпена, как описано в любом из воплощений изобретения, достаточно для того, чтобы произошло окисление терпена. Тем не менее, дополнительная трансформация, по меньшей мере, одним ферментом, вовлеченным в продукцию ациклического предшественника терпена и/или изопентенилдифосфата (IPP) или диметилаллилдифосфата (DMAPP), имеет то преимущество, что увеличивает количество доступного для окисления соединения терпена.
Организм или клетка предназначены для «экспрессии» полипептида, при условии, что организм или клетка трансформируют для несения нуклеиновой кислотой, кодирующей указанный полипептид, данная нуклеиновая кислота транскрибируется в мРНК и полипептид обнаруживают в организме-хозяине или в клетке-хозяине. Термин «экспрессируют» охватывает термины «гетерологично экспрессируют» и «сверх-экспрессируют», последнее относится к уровням мРНК, полипептида и/или ферментативной активности превышающим таковые, измеренные в нетрансформированных организме или клетке. Более подробное описание подходящих способов трансформации организма-хозяина или клетки-хозяина, не представляющих собой организм или клетку человека, будет описано в дальнейшем в той части описания, которая посвящена таким трансформированным организмам-хозяевам или клеткам-хозяевам, не представляющим собой организм или клетки человека, как специфическим целям настоящего изобретения, и в примерах.
Способы трансформации организмов, например, микроорганизмов, такие чтобы они экспрессировали терпен-синтазу, также известны в этой области техники. Такие способы, например, могут быть найдены в WO 2010/134004, в которой описана трансформация различных организмов-хозяев и клеток-хозяев зизаен-синтазой, т.е. ферментом, способным катализировать образование зизаена из фарнезилпирофосфата.
Для осуществления изобретения in vivo, организм-хозяин или клетку-хозяин культивируют в условиях, способствующих продукции окисленного терпена. Такие условия представляют собой любые условия, приводящие к росту организма-хозяина или клетки-хозяина. Предпочтительно, такие условия разрабатывают для оптимального роста организма-хозяина или клетки-хозяина. Соответственно, если хозяин представляет собой трансгенное растение, то обеспечивают условия оптимального роста, такие как, например, оптимальный свет, воду и условия питания. Если хозяин представляет собой одноклеточный организм, то условия, способствующие продукции окисленного терпена, могут включать добавление к культуральной среде хозяина подходящих кофакторов. Кроме того, культуральная среда может быть выбрана так, чтобы сделать окисление терпена максимальным. Оптимальные условия культивирования известны специалисту в этой области техники и не представляют собой специфические условия настоящего изобретения. Примеры подходящих условий описаны более подробно в следующих далее примерах.
Организмы-хозяева, не представляющие собой организм человека, подходящие для осуществления способа изобретения in vivo могут представлять собой любые многоклеточные или одноклеточные организмы, не представляющие собой организм человека. В предпочтительном воплощении, организм-хозяин, не представляющий собой организм человека, применяемый для осуществления изобретения in vivo, представляет собой растение, прокариота или гриб. Любое растение, прокариот или гриб могут быть применены. В особенности, полезны те растения, которые в природе продуцируют большие количества терпенов. В более предпочтительном воплощении, растение выбирают из семейств Solanaceae, Poaceae, Brassicaceae, Fabaceae, Malvaceae, Asteraceae или Lamiaceae. Например, растение выбирают из родов Nicotiana, Solanum, Sorghum, Arabidopsis, Brassica (рапс), Medicago (люцерна), Gossypium (хлопок), Artemisia, Salvia и Mentha. Предпочтительно, растение принадлежит к виду Nicotiana tabacum.
В более предпочтительном воплощении организм-хозяин, не представляющий собой человека, применяемый для осуществления способа изобретения in vivo, представляет собой микроорганизм. Может быть применен любой микроорганизм, но в соответствии с еще более предпочтительным воплощением указанный микроорганизм представляет собой бактерию или гриб. Предпочтительно указанный гриб представляет собой дрожжи. Наиболее предпочтительно, указанная бактерия представляет собой Е. coli и указанные дрожжи представляют собой Saccharomyces cerevisiae.
Некоторые из данных организмов не производят терпен подвергаемый окислению естественным образом. Для того чтобы подходить для осуществления способа изобретения, данные организмы должны быть трансформированы с целью продуцировать указанный терпен. Они могут быть трансформированы таким образом или перед трансформацией, одновременно с трансформацией или после трансформации нуклеиновой кислотой, описанной в соответствии с любым из описанных выше воплощений, как объяснено выше.
Также вместо полных организмов в качестве хозяев для осуществления способа изобретения in vivo, могут быть применены изолированные клетки высших эукариот. Подходящие эукариотические клетки могут представлять собой любую клетку, не представляющую собой клетку человека, но предпочтительно представляют собой растительные клетки.
Важный инструмент трансформирования организмов-хозяев или клеток-хозяев, подходящих для осуществления способа изобретения in vivo, представляет собой вектор экспрессии, включающий нуклеиновую кислоту в соответствии с любым из воплощений изобретения. Такой вектор, следовательно, представляет собой также цель настоящего изобретения.
«Вектор экспрессии», как применен в настоящем документе, включает любой линейный или кольцевой рекомбинантный вектор, включающий, без ограничений, вирусные векторы, бактериофаги и плазмиды. Специалист способен выбрать подходящий вектор в соответствии с системой экспрессии. В одном из воплощений, вектор экспрессии включает нуклеиновую кислоту изобретения, функционально связанную, по меньшей мере, с одной регуляторной последовательностью, которая контролирует транскрипцию, трансляцию, инициацию и терминацию, такой как промотор для транскрипции, оператор или энхансер, или сайт связывания рибосом в мРНК и, необязательно, включающей, по меньшей мере, один маркер отбора. Нуклеотидные последовательности представляют собой «функционально связанные» нуклеотидные последовательности, если регуляторная последовательность функционально связана с нуклеиновой кислотой изобретения.
Вектора экспрессии настоящего изобретения могут быть применены в способах получения генетически трансформированного организма-хозяина и/или клетки-хозяина, в организмах-хозяевах и/или клетках-хозяевах, несущих нуклеиновые кислоты изобретения, и в способах получения или изготовления полипептидов изобретения, как дополнительно раскрыто ниже.
Рекомбинантные организмы-хозяева и клетки-хозяева, не представляющие собой организм и клетки человека, трансформированные для несения, по меньшей мере, одной нуклеиновой кислоты в соответствии с любым из воплощений настоящего изобретения, также представляют собой очень полезное инструменты для осуществления способа изобретения. Следовательно, такие организмы-хозяева и клетки-хозяева, не представляющие собой организм и клетки человека, представляют собой еще один объект настоящего изобретения. В предпочтительном воплощении, такой организм-хозяин или такая клетка-хозяин гетерологически экспрессировали или сверхэкспрессировали полипептид в соответствии с любым воплощением настоящего изобретения.
В соответствии с предпочтительным воплощением, организм-хозяин или клетка-хозяин, не представляющий собой организм или клетку человека, дополнительно экспрессировали Р450-редуктазу (CPR), как описано выше. CPR может или присутствовать в организме-хозяине или в клетке-хозяине естественным образом, или такой организм или клетка могут быть трансформированы для экспрессии CPR перед трансформацией, одновременно с трансформацией или после трансформации для экспрессии полипептид изобретения. В предпочтительном воплощении изобретения клетку-хозяин или организм-хозяин трансформируют для экспрессии слитого полипептида, включающего как полипептид изобретения, так и CPR.
В другом предпочтительном воплощении, организм или клетка способны продуцировать предназначенный для окисления терпен. Это происходит в том случае, когда организм или клетка экспрессируют терпен-синтазу, способную катализировать образование указанного терпена. В случае если организм-хозяин или клетка-хозяин не экспрессируют такую терпен-синтазу естественным образом, то их можно трансформировать перед трансформацией, одновременно с трансформацией или после трансформации полипептидом, обладающим активностью Р450.
Организм или клетка, не относящиеся к организму или клетке человека, могут быть успешно дополнительно трансформированы, по меньшей мере, одним геном, кодирующим полипептид, вовлеченный в метаболизм продукции ациклического предшественника терпена, такого как геранилпирофосфат, фарнезилпирофосфат или геранилгеранилпирофосфат. Такие полипептиды включают, например, ферменты МЕР-пути, MVA-пути и/или пренилтрансферазы. Трансформированные организм или клетка, не представляющие собой организм или клетку человека, способны продуцировать соединение терпена с полипептидом изобретения и с CPR, или со слитым полипептидом, включающим и полипептид изобретения и CPR, как описано в любом из воплощений изобретения, достаточными для того, чтобы произошло окисление терпена. Тем не менее дополнительная трансформация, по меньшей мере, одним ферментом, вовлеченным в продукцию ациклического предшественника терпена и/или изопентенилдифосфата (IPP) или диметилаллилдифосфата (DMAPP), имеет то преимущество, что увеличивает количество доступного для окисления соединения терпена.
Типы организмов-хозяев и клеток-хозяев изобретения, не представляющие собой организм и клетки человека, представляют собой такие организм и клетки, как описано в любом из воплощений способа окисления соединения терпена.
Термин «трансформированный» относится к тому факту, что хозяин был подвергнут генно-инженерной обработке для включения одной, двух или более копий каждой их нуклеиновых кислот, необходимой для любого из вышеописанных воплощений. Предпочтительно термин «трансформированный» относится к хозяевам, которые гетерологически экспрессируют полипептиды, кодируемые нуклеиновой кислотой, которой они были трансформированы, а также сверх-экспрессируют указанные полипептиды. Соответственно, в воплощении, настоящее изобретение обеспечивает трансформированный организм, в котором полипептиды экспрессируются в более высоком количестве по сравнению с таким же организмом, не подвергавшемся подобной трансформации.
Существует несколько, известных в этой области техники, способов создания трансгенных организмов-хозяев или клеток-хозяев, таких как растения, грибы, прокариоты или культуры клеток высших эукариот. Подходящие векторы для клонирования и экспрессии для применения в бактериальных, грибковый, дрожжевых, растительных клетках-хозяевах и в клетках-хозяевах млекопитающих описаны, например, в руководствах Pouwels et al., Cloning Vectors: A Laboratory Manual, 1985, Elsevier, New York и Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd edition, 1989, Cold Spring Harbor Laboratory Press. Векторы клонирования и экспрессии для высших растений и/или, в особенности, растительных клеток доступны специалисту. Смотри, например, Schardl et al. Gene 61: 1-11, 1987.
Способы трансформирования организмов-хозяев или клеток-хозяев для несения трансгенных нуклеиновых кислот хорошо знакомы специалисту. Современные способы создания трансгенного растения, например, включают: электропорацию растительных протопластов, опосредованную липосомами трансформацию, опосредованную агробактериями трансформацию, опосредованную полиэтиленгликолем трансформацию, бомбардировку частицами, микроинъекцию в растительную клетку и трансформацию с применением вирусов.
В одном из воплощений, трансформированная ДНК встраивается в хромосому организма-хозяина и/или клетки-хозяина, не представляющих собой организм и/или клетку человека, таким образом, чтобы произошло образование стабильной рекомбинантной системы. Любой способ интеграции в хромосому, известный в этой области техники, может быть применен при осуществлении изобретения на практике, включая без ограничений опосредуемый рекомбиназой обмен кассетами (RMCE), вирусную сайт-специфическую хромосомную вставку, инъекцию аденовируса и пронуклеарную инъекцию.
Для осуществления способа окисления соединение терпена in vitro, как было описано в настоящем документе выше, очень выгодно обеспечить способ изготовления полипептида изобретения. Следовательно, изобретение обеспечивает способ получения полипептида настоящего изобретения, включающий
а) культивирование организма-хозяина или клетки-хозяина, не представляющих собой организм или клетку человека, трансформированных для несения, по меньшей мере, одной нуклеиновой кислоты в соответствии с изобретением и экспрессирование или сверх-экспрессирование, по меньшей мере, одного полипептида изобретения;
b) выделение полипептида изобретения из организма-хозяина или клетки-хозяина, не представляющих собой организм или клетку человека, культивированных на стадии а).
В соответствии с предпочтительным воплощением, указанный способ дополнительно включает, перед стадией а), трансформирование организма-хозяина или клетки-хозяина, не представляющих собой организм или клетку человека, по меньшей мере, одной нуклеиновой кислотой в соответствии с изобретением, так чтобы они экспрессировали или сверх-экспрессировали полипептид в соответствии с изобретение.
Трансформация и культивирование организма-хозяина или клетки-хозяина, не представляющих собой организм или клетку человека, могут быть осуществлены как описано выше для способа продуцирования окисленного терпена in vivo. Стадия b) может быть проведена с применением любой, хорошо известной в этой области техники, методики выделения определенного полипептида из организма или клетки.
Термин «полипептидный вариант», так как применен в настоящем документе, означает полипептид, способный катализировать окисление соединения терпена, имеющего формулу (I) и имеющего достаточный процент идентичности последовательности в соответствии с любым описанным выше воплощением. Такие варианты полипептида кодируются нуклеотидными последовательностями, которые претерпели одну или более делеций, вставок или замен.
Варианты могут включать консервативно замещенные последовательности, это означает, что данный аминокислотный остаток замещают остатком, имеющим сходные физико-химические характеристики. Примеры консервативных замен включают замену одного алифатического остатка, такого как Ile, Val, Leu или Ala на другой остаток, или замену одного полярного остатка на другой остаток, такие как замена между Lys и Arg; Glu и Asp; или Gln и Asn. Смотри Zubay, Biochemistry, Addison-Wesley Pub. Co., (1983). Эффекты таких замен могут быть рассчитаны с помощью матрицы замещения, такой как РАМ-120, РАМ-200 и РАМ-250, как обсуждалось в работе Altschul, (J. Mol. Biol. 219: 555-65, 1991). Другие такие консервативные замены, например, замена целых участков, имеющих сходные характеристики гидрофобности, хорошо известны. Полипептиды изобретения также может быть подвергнуты неконсервативным заменам, для того чтобы создавать более разнообразные варианты, при условии, что такие варианты сохраняют требуемую активность цитохрома Р450. Варианты также могут быть созданы путем делеции и вставки нуклеотида(нуклеотидов) в последовательность нуклеиновой кислоты, кодирующую вариант полипептида.
Варианты полипептидов изобретения могут быть применены для достижения, например, требуемого усиления или снижения энзиматической активности, модификации региохимии или стереохимии, или изменения утилизации субстрата или распределение продукта, увеличения сродства к субстрату, улучшения специфичности производства одного или нескольких требуемых соединений, увеличения скорости ферментативной реакции, повышения активности или стабильности в специфическом окружении (pH, температура, растворитель и т.д.), или улучшения уровня экспрессии в требуемой системе экспрессии. Вариант или мутант, полученный способом сайт-направленного мутагенеза, может быть изготовлен любым способом, известный в этой области техники. Варианты и производные природных полипептидов могут быть получены с помощью выделения природных вариантов, или нуклеотидной последовательности вариантов, из других или тех же линий или видов растений, или путем искусственного программирования мутаций нуклеотидных последовательностей, кодирующих полипептиды изобретения. Изменения природных аминокислотных последовательностей могут быть завершены с помощью любого числа общепринятых способов.
Варианты полипептидов, полученные в результате слияния дополнительных пептидных последовательностей с амино- и карбоксильного концов полипептидов изобретения, могут быть применены для усиления экспрессии полипептидов, полезны при очистке белка или могут быть применены для улучшения энзиматической активности полипептида в требуемом окружении или системе экспрессии. Такие дополнительные пептидные последовательности могут представлять собой, например, сигнальные пептиды. Соответственно, настоящее изобретение охватывает варианты полипептидов изобретения, такие как те, которые были полученные с помощью слияния с другими олиго- или полипептидами, и/или такие как те, которые были присоединены к сигнальным пептидам.
Следовательно, в воплощении, настоящее изобретение обеспечивает способ получения варианта полипептида, способного катализировать окисление соединения терпена, включающий следующие стадии:
(a) отбор нуклеиновой кислоты в соответствии с любым из раскрытых выше воплощений;
(b) модификация выбранной нуклеиновая кислота для получения, по меньшей мере, одной мутантной нуклеиновой кислоты;
(c) трансформирование клетки-хозяина или одноклеточного организма-хозяина последовательностью мутантной нуклеиновой кислоты для экспрессии полипептида, кодируемого последовательностью мутантной нуклеиновой кислоты;
(d) скрининг полипептид, по меньшей мере, на одну модифицированную активность цитохрома Р450; и,
(e) необязательно, если полипептид не имеет требуемого варианта активности цитохрома Р450, повторение стадий способа от (а) до (d) до тех пор, пока полипептид с требуемым вариантом активности цитохрома Р450 не будет получен;
(f) необязательно, если полипептид, имеющий требуемый вариант активности цитохрома Р450 идентифицируют на стадии d), выделение соответствующей мутантной нуклеиновой кислоты, полученной на стадии (с).
На стадии (b), может быть создано большое число последовательностей мутантной нуклеиновой кислоты, например, с помощью случайного мутагенеза, сайт-специфического мутагенеза или перестановки в ДНК. Подробное описание процедур генных перестановок можно найти в работе Stemmer, DNA shuffling by random fragmentation and reassembly: in vitro recombination for molecular evolution. Proc Natl Acad Sci U S A., 1994, 91 (22): 10747-1075. Коротко, перестановка в ДНК относится к способу случайной рекомбинации известной последовательности in vitro, включающему, по меньшей мере, две нуклеиновые кислоты, выбранные для рекомбинации. Например, мутации могут быть введены в определенные локусы путем синтеза олигонуклеотидов, содержащих мутантную последовательность, фланкированную сайтами рестрикции, позволяющими проводить лигирование с фрагментами природной последовательности. После лигирования полученная реконструированная последовательность кодирует аналог, имеющий требуемую аминокислотную вставку, замену или делецию. Альтернативно, можно применить методику олигонуклеотид-направленного сайт-специфического мутагенеза для обеспечения измененного гена, заранее определенные кодоны которого могут быть изменены в результате замены, делеции или вставки.
Соответственно, нуклеиновая кислота, кодирующая полипептид, включающий SEQ ID NO:3, 4 или их комплементарные цепи, может быть рекомбинирована с любой другой нуклеиновой кислотой, кодирующей цитохром Р450, например, изолированной из организма, отличного от Vetiveria zizanioides (L.) Nash. Таким образом, могут быть получены и выделены мутантные нуклеиновые кислоты, которые могут быть применены для трансформирования клетки-хозяина в соответствии со стандартными методиками, например, такими как методики, раскрытые в представленных примерах.
На стадии (d), полипептид, полученный на стадии (с), подвергают скринингу для обнаружения, по меньшей мере, одной модифицированной активности цитохрома Р450. Примеры требуемой модифицированной активности цитохрома Р450, для которой экспрессируемый полипептид может быть подвергнут скринингу, включают усиленную или сниженную энзиматическую активность, измеренную по отношению к величине Км или Vmax, модифицированную регио-химию или стереохимию и измененную утилизацию субстрата или распределение продукта. Скрининг энзиматической активности может быть выполнен в соответствии с методиками, знакомыми специалисту, и теми, которые раскрыты в представленных примерах.
Стадию (е) обеспечивают для повторений стадий способа (a)-(d), которые, возможно предпочтительно, выполняют параллельно. Соответственно, путем создания значительного числа мутантных нуклеиновых кислот, многие клетки-хозяева могут быть трансформированы различными мутантными нуклеиновыми кислотами одновременно, что позволяет проводить последующий скрининг увеличенного числа полипептидов. Вероятность получения требуемого варианта полипептида, таким образом, может быть увеличена по усмотрению специалиста.
Все публикации, упомянутые в настоящей заявке, включены путем отсылок для раскрытия и описания способов и/или материалов, в связи с которыми публикации были процитированы.
Описание чертежей
Фигура 1: Структуры приведенных соединений.
Фигура 2: Выравнивание аминокислотных последовательностей выбранных Р450 монооксигеназ с определенной терпенгидроксилазной активностью (известный уровень техники). Консервативные области, обнаруживаемые во всех Р450-ферментах, подчеркнуты. Шесть областей, примененных для конструирования терпен-гидроксилаза-специфических олигонуклеотидов, подчеркнуты стрелками. Направление стрелок указывает ориентацию олигонуклеотидов.
Фигура 3: N-концевые модификации (мембранный якорь), введенные в две Р450 ветивера для улучшенной гетерологичной экспрессии в Е. coli.
Фигура 4: Типичный СО-дифференциальный спектр, полученный с рекомбинантным белком VzP521-11.
Фигура 5: Последовательность области спейсера между Р450 и CPR в бицистронных конструктах. Показаны последовательность ДНК и аминокислотные последовательности на конце Р450 и в начале CPR.
Фигура 6: GCMS-анализ биопревращения (+)-зизаена с Е. coli, экспрессирующей Р450 ветивера VzP521-16 и CPR из Arabisdopsis (tcATR1). А. Общая ионная хроматограмма. В. Масс-спектры субстрата (1) и продуктов (от 2 до 4) с идентичностью и структурой соответствующих соединений.
Фигура 7: Схема, показывающая последовательные стадии энзиматического окисления зизаена.
Фигура 8: Общая ионная хроматограмма GCMS-анализа биопревращения некоторых молекул терпенов под действием Р450 ветивера. Указаны пики, соответствующие субстрату, и молекулярные массы продуктов.
Фигура 9: Схема, показывающая биопревращение под действием Р450 ветивера некоторых молекул терпенов, для которых идентифицированы продукты.
Фигура 10: Организация искусственного оперона, разработанного для совместной экспрессии в Е. coli Р450 ветивера, CPR и зизаен-синтазы.
Фигура 11: Общая ионная хроматограмма GCMS анализа сесквитерпена, продуцируемого клетками Е. coli, экспрессирующими (+)-зизаен-синтазу, Р450 ветивера VzP521-16-1 и CPR из Arabidopsis, вместе с ферментами для получения FPP в гетерологичном мевалонатном пути.
Конкретные воплощения изобретения или примеры
Изобретение теперь будет описано подробнее с помощью следующих примеров.
Пример 1
Экстракция РНК и создание библиотеки кДНК
Растения ветивера (Vetiveria zizanioides (L.) Nash) получали из питомника растений (Южная компания по производству растений (The Austral Plants Company), Les Avirons, остров Реюньон, Франция). Растения выращивали в горшках в теплице (Агрономическая научно-исследовательская станция Lullier, Женева, Швейцария) и размножали вегетативно путем разделения скоплений растений в возрасте от шести месяцев до одного года. Для сбора корней растения извлекали из горшков и промывали водопроводной водой.
Для экстракции РНК корни от нескольких растений объединяли: молодые растения (4-6 месяцев после размножения), старые растения с хорошо развитой плотной корневой системой (1-2 года после размножения) и молодые растения высушивали при комнатной температуре в течение 24-36 часов после извлечения из горшков. Корни отрезали от надземной части растений и замораживали в жидком азоте. Корни сначала грубо нарезали в жидком азоте с помощью блендера Waring (Waring Laboratory, Torrington, США) и затем перемалывали до тонкоизмельченного порошка с помощью ступки и пестика. Общую РНК экстрагировали, следуя методике, описанной в работе Kolosova et al. (Kolosova N, Miller В, Ralph S, Ellis BE, Douglas C, Ritland K, and Bohlmann J, Isolation of high-quality RNA from gymnosperm and angiosperm trees. J. Biotechniques, 36 (5), 821-4, 2004) со следующими модификациями. Объем буфера экстракции, равный 20 мл, применяли для 2 г измельченной ткани, и буфер экстракции дополняли 2% (масса/объем) PVP (поливинилпирролидон, «Sigma-Aldrich»). Для экстракции с помощью СТАВ (цетилтриметиламмония бромид, «Sigma-Aldrich»), осадок нуклеиновой кислоты ресуспендировали в 2 мл ТЕ-буфера (10 мМ Трис-HCl, pH 8, 1 мМ EDTA) и экстракцию проводили с 2 мл 5 М NaCl и 1 мл 10%-ного СТАВ. Для осаждения изопропанолом, осадок нуклеиновой кислоты растворяли в 500 мкл ТЕ. Конечный осадок РНК ресуспендировали в 50 мкл воды.
Библиотеку двухцепочечной кДНК с присоединенными адаптерами получали из 1 мкг мРНК с помощью набора для амплификации кДНК Marathonтм («Clontech», Takara Bio Europe), следуя протоколу производителя.
Пример 2
Конструирование Р450-специфических олигонуклеотидов
Для конструирования олигонуклеотидов, специфических для растительных Р450 с терпенгидроксилазной активностью, выбирали аминокислотные последовательности из известных терпен-гидроксилирующих Р450: лимонен 6-гидроксилазу из мяты (номер доступа в GenBank AAD44150), две лимонен-3-гидроксилазы из перечной мяты (номера доступа в GenBank AAD44152 и AAD44151), эии-аристолочен-гидроксилазу из табака (номер доступа в GenBank ААК62343), премнаспиродиен-гидроксилазу из Hyoscyamus muticus (номер доступа в GenBank ABS00393), две лимонен-гидроксилазы из шотландской мяты (номера доступа в GenBank AAQ18707 и AAQ18708), дитерпен-гидроксилазу из табака (номер доступа в GenBank AAD47832) и двух членов из семейства CYP71D, Cyp71D4 из картофеля (номер доступа в GenBank No. САС24711) и CYP71D6 из картофеля Чако (номер доступа в Genbank Р93530). Последовательности выравнивали с помощью программы ClustalW (Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994); CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice; Nucleic Acids Res. 22, 4673-4680). Выравнивание представлено на фигуре 2.
Для того чтобы сконструировать олигонуклеотиды, которые предполагается применять для ПЦР-амплификации фрагментов гомологических Р450 кДНК, в данном выравнивании выбирали консервативные области. При выборе данных областей рассматривали такие параметры, как консервативность аминокислот по всей последовательности и присутствие аминокислот с низкой вырожденностью кодона. Кроме того, поскольку геномы растений содержат большое число Р450, вовлеченных во множество различных метаболических процессов, намеренно избегали области, относящиеся к функциям, общим для всех Р450. Данные области включают, например, связывающий гем домен, фланкирующий абсолютно консервативный остаток цистеина, который ковалентно связывает железо в гемме через его тиолатную боковую цепь (мотив PFGxGRRICPG в нашем выравнивании), так называемый «меандр» (мотив FxPERF в нашем выравнивании), предположительно вовлеченный во взаимодействие с белком редокс-партнером и в стабилизацию комплекса гем-белок, и область спирали I, локализованную в активном центре на дистальной стороне гема (мотив (A/G)GTETSS) и вовлеченную в перенос протона и активацию кислорода. Таким образом, были выбраны шесть консервативных областей, предположительно характеризующие растительные терпен-монооксигеназы (подчеркнуты стрелками на фигуре 1).
Гибридные праймеры, содержащие 3' вырожденное ядро и 5' консенсусную часть, были сконструированы из данных областей, следуя стратегии гибридных консенсусно-вырожденных олигонуклеотидных праймеров (CODEHOP) (Rose Т.М., Schultz E.R., Henikoff J.G, Pietrokovski S., McCallum С.М., and Nenikoff S.; 1998; Consensus-degenerated hybrid oligonucleotide primers for amplification of distantly related sequences; Nucleic Acids Research 26 (7), 1628-1635) и используя веб-интерфейс компьютерной программы, доступной по адресу http://blocks.fhcrc.org/blocks/codehop.html. Олигонуклеотиды конструировали так, чтобы они имели вырожденное ядро, состоящее из 11-15 оснований, с максимальной вырожденностью, равной 192, и температурой отжига от 60 до 64°C. Применяя этот подход, три смысловых праймера (P450-Terp-F1 to F3 (SEQ ID NO: 5-7)) и четыре анти-смысловых праймера (P450-Terp-R1 to R4 (SEQ ID NO: 8-11) были сконструированы из шести консервативных областей, показанных на фигуре 1 (таблица 1).
Таблица 1. Терпен-гидроксилаза-специфические олигонуклеотиды
Последовательность вырожденного ядра каждого праймера указана строчными буквами и консенсусная часть указана заглавными буквами. Нуклеотидные последовательности указаны от 5' к 3' концу для прямых праймеров и от 3' к 5' концу для обратных праймеров. Вырожденность в нуклеотидных последовательностях указана с помощью однобуквенного кода IUPAC.

Пример 3
ПЦР-амплификация кДНК Р450 ветивера
Праймеры, описанные в примере 2, применяли для амплификации фрагментов кДНК Р450 с помощью ПЦР из библиотеки кДНК ветивера. ПЦР проводили, применяя смесь полимераз Advantage® 2 («Clontech», Takara Bio Europe). Каждая ПЦР-смесь содержала, в общем объеме, равном 50 мкл, 5 мкл буфера для ПЦР Advantage® 2, 200 мкМ дНТФэ, 200 нМ каждого олигонуклеотидного праймера, 5 мкл разведенной в 200 раз кДНК, 1 мкл смеси полимераз Advantage® 2. Для амплификации применяли следующие условия:
- 3 минуты денатурации при 94°C;
- 15 циклов, включающих
- 1 минуту денатурации при 94°C,
- 1 мин отжига при 65°C для первого цикла и минус один градус для каждого последующего цикла, и
- 2 минуты удлинения при 72°C;
- 20 циклов, включающих
- 1 минуту денатурации при 94°C,
- 1 мин отжига при 58°C и
- 2 минуты удлинения при 72°C; и
- наконец, 10 минут удлинения при 72°C.
Различные ПЦР проводили с возможными комбинациями специфических смысловых и анти-смысловых праймеров для терпен-гидроксилазы. Ампликоны с ожидаемым размером клонировали в вектор pCR® 2.1-ТОРО, применяя набор для клонирования ТОРО ТА («Invitrogen», Carlsbad, СА), вставки подвергали ДНК-секвенированию и последовательность сравнивали с базой данных нерезервированных белков в GenBank (NCBI), применяя алгоритм BLASTX (Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman, D.J. (1990) J. Mol. Biol. 215, 403-410).
Некоторые комбинации праймеров (P450-terp-F1 (SEQ ID NO: 5) с P450-terp-R2 (SEQ ID NO: 9), P450-terp-F3 (SEQ ID NO: 7) с P450-terp-R3 (SEQ ID NO: 10) и P450-terp-F1 (SEQ ID NO: 5) с P450-terp-R4 (SEQ ID NO: 11)) обеспечили фрагменты ДНК с ожидаемым размером и с последовательностями, продемонстрировавшими гомологию с последовательностями Р450. Сохранили только фрагменты, показавшие гомологию с охарактеризованными терпен-монооксигеназами (приблизительно 50% отсеквенированных фрагментов). Выбранные последовательности ДНК выравнивали и выводили 1167-по консунсусную последовательность ДНК (СА521 (SEQ ID NO: 12). Аминокислотная последовательность, выведенная из СА521, показала идентичность, достигавшую 45%, с известными растительными терпен-монооксигеназами.
Полноразмерные последовательности были получены с помощью методики быстрой амплификации концов кДНК (RACE). Набор для амплификации кДНК Marathonтм («Clontech», Takara Bio Europe) применяли для всех RACE-экспериментов. Типичные реакционные смеси RACE содержат, в конечном объеме, равном 50 мкл, 5 мкл Advantage® 2 буфера для ПЦР («Clontech», Takara Bio Europe), 200 мкМ каждого из дНТФ, 1 мкл смеси полимераз Advantage® 2 («Clontech», Takara Bio Europe), 200 мкМ праймера, специфического к адаптеру, 200 мкМ кДНК-специфического праймера и 5 мкл разведенной в 200 раз кДНК из корней ветивера с лигированными адаптерами. Амплификацию проводили в термоциклере Eppendorf Mastercycler Gradiant. Условия термоциклирования были следующими: 1 мин при 94°C, 5 циклов, включающих 30 сек при 94°C и 3 мин при 72°C, 5 циклов, включающих 30 сек при 94°C и 3 мин при 70°C, 20 циклов, включающих 30 сек при 94°C и 3 мин при 68°C. При необходимости проводили второй раунд амплификации, применяя внутренние олигонуклеотиды (для гнездовой ПЦР). Ампликоны с ожидаемым размером клонировали в вектор pCR® 2.1-ТОРО («Invitrogen», Carlsbad, СА), вставки подвергали ДНК-секвенированию и последовательность сравнивали с базой данных нерезервированных белков в GenBank (NCBI), применяя алгоритм BLASTX (Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman, D.J. (1990) J. Mol. Biol. 215, 403-410).
Для амплификации концов CA-521, смысловые и анти-смысловые олигонуклеотиды выводили из полученных с помощью ПЦР-фрагментов ДНК и применяли в 3'RACE и 5'RACE: CA521-F1 (SEQ ID NO: 13), CA521-F2 (SEQ ID NO: 14), CA521-R1 (SEQ ID NO: 15) и CA521-R2 (SEQ ID NO: 16). Применяя смысловые олигонуклеотиды, мы получили фрагмент, размером в 500 по (СА635 (SEQ ID NO: 17)), охватывающий перекрывание из 176 идентичных остатков с фрагментом СА521. Данный фрагмент СА635 содержал дополнительные 138 по, кодировавшие область, включавшую стоп-кодон, за которой следовала 3' нетранслируемая область. 5'RACE обеспечили 426 по фрагмент (СА884 (SEQ ID NO: 18)), содержавший недостающие 243 по, кодирующие область на 5'-конце.
Олигонуклеотиды конструировали из стартовой области и стоп-области реконструированной полноразмерной последовательности, СА521-старт (SEQ ID NO: 19), СА521-стоп (SEQ ID NO: 20), и применяли в качестве праймеров для амплификации полноразмерных кДНК. Данную амплификацию проводили, применяя ДНК-полимеразу Pfu («Promega», Madison, WI, США), в конечном объеме, равном 50 мкл, содержавшем 5 мкл Pfu ДНК-полимеразного 10Х буфера, 200 мкМ каждого из дНТФ, 0,4 мкМ каждого из праймеров, 2,9 единиц ДНК полимеразы Pfu и 2,5 мкл разбавленной в 200 раз кДНК ветивера. Условия термоциклирования были следующими: 1,5 мин при 95°C; 30 циклов, включающих 45 сек при 95°C, 30 сек при 64°C и 4 мин при 72°C; и 10 мин при 72°C. ПЦР-продукты клонировали в вектор pCR®2.1-TOPO, применяя набор для клонирования ТОРО ТА («Invitrogen», Carlsbad, СА) и контролировали последовательность ДНК. Из секвенирования некоторых клонов, были сохранены две различные последовательности ДНК (VzP521-11 (SEQ ID NO: 21) и VzP521-16 (SEQ ID NO: 22)), обладавшие 93% идентичности последовательности. Выведенные аминокислотные последовательности, состоявшие из 513 и 514 остатков, соответственно, обладали 89% идентичности. Идентичности аминокислотных последовательностей VzP521-11 (SEQ ID NO: 23) и VzP521-16 (SEQ ID NO: 24) с самым близким соответствием в базах данных последовательностей приведены ниже.

Идентичность аминокислотных последовательностей с ближайшими общедоступными и функционально охарактеризованными белками приведена в нижеследующей таблице:

Полипептиды VzP521-11 (SEQ ID NO: 23) и VzP521-16 (SEQ ID NO: 24) включают одну часть, которая представляет собой мембранный якорь, и активную область, ответственную за каталитическую активность Р450. Активные области VZP521-11 и VzP521-16 представлены в последовательностях SEQ ID NO: 1 и 2, соответственно. Последовательности нуклеиновой кислоты, кодирующие данные активные последовательности, представлены в SEQ ID NO: 3 и 4, соответственно.
Пример 4
Гетерологичная экспрессия цитохромов Р450 ветивера в бактериях
В эукариотических Р450-монооксигеназах, N-концевая последовательность белка представляет собой мембранный якорь, необходимый для мембранной локализации данных ферментов. Эта часть белка, ограниченная богатым пролином доменом (PPGP в 521-11 (SEQ ID NO: 23) и 521-16 (SEQ ID NO: 24)), не существенна для контроля специфичности энзиматической активности. Следовательно, данная область может быть модифицирована путем делеции, вставки или мутации без влияния на каталитическую активность. Однако было показано, что специфическая модификация N-концевой области эукариотических Р450, включая растительные Р450, оказывает положительный эффект на уровни детектируемых рекомбинантных белков, если их экспрессируют в микроорганизмах (Halkier et al. (1995) Arch. Biochem. Biophys. 322, 369-377; Haudenschield et al. (2000) Arch. Biochem. Biophys. 379, 127-136). Таким образом, на основе данных предыдущих наблюдений область мембранного якоря в Р450 VzP521-11 и VzP521-16 была перестроена для введения модификации, показанной на фигуре 3.
Модифицированные кДНК получали с помощью ПЦР следующим образом. Первый фрагмент, соответствующий области мембранного якоря, амплифицировали, применяя праймеры Pfus-NdeI (SEQ ID NO: 25) и 521_fus_r (SEQ ID NO: 26) (с плазмидой P2-2-48 (Haudenschield et al. (2000) Arch. Biochem. Biophys. 379, 127-136) в качестве матрицы. Два других фрагмента амплифицировали, применяя праймеры 521-fus-f (SEQ ID NO: 27) и 521-Hind (SEQ ID NO: 28) и или кДНК VzP521-11 (SEQ ID NO: 21) или VzP521-16 (SEQ ID NO: 22) в качестве матрицы. Второй раунд ПЦР проводили, применяя в качестве матрицы первый ПЦР-продукт и или два последних ПЦР-продукта и в качестве праймеров Pfus-NdeI (SEQ ID NO: 25) и 521-Hind (SEQ ID NO: 28). Все ПЦР проводили с ДНК-полимеразой Pfu («Promega», Madison, WI, США), в конечном объеме, равном 50 мкл, содержавшем 5 мкл 10Х буфера ДНК-полимеразы Pfu, 200 мкМ каждого из дНТФ, 0,4 мкМ каждого из праймеров, 2,9 единиц ДНК-полимеразы Pfu и 2,5 мкл разбавленной в 200 раз кДНК ветивера. Условия термоциклирования были следующими: 1,5 мин при 95°C; 30 циклов, включавших 45 сек при 95°C, 30 сек при 64°C и 4 мин при 72°C; и конечная стадия - 10 мин при 72°C. Два ПЦР-продукта, VzP521-11-1 (SEQ ID NO: 37) и VzP521-16-1 (SEQ ID NO: 38), расщепляли с помощью рестриктаз NdeI и HindIII и лигировали в экспрессионную плазмиду pCWori (Barnes H.J (1996) Method Enzymol. 272, 3-14), обеспечивая плазмиды pCW-218-521-11 и pCW-218-521-16, содержавшие, соответственно кДНК, кодирующую N-концевые модифицированные VzP521-11 и VzP521-16 Р450 (аминокислотные последовательности VzP521-11-1 (SEQ ID NO: 35) и VzP521-16-1 (SEQ ID NO: 36)).
Для гетерологичной экспрессии, клетки JM109 Е. coli трансформировали плазмидами экспрессии 218-521-11 или 218-521-16. Единичные колонии трансформантов применяли для инокуляции культур в 5 мл ЛБ-среды, содержавшей 50 мкг/мл ампициллина. Клетки растили в течение 10-12 часов при 37°C. Затем культуры применяли для инокуляции в 250 мл ТВ-среды (Terrific Broth), дополненный 50 мкг/мл ампициллина и 1 мМ тиамина-HCL. Культуры инкубировали при 28°C в течение 3-4 ч при умеренном перемешивании (200 об/мин) перед добавлением 75 мг/л δ-аминолевулиновой кислоты («Sigma») и 1 мМ IPTG (изопропил-β-D-1-тиогалактопиранозида), и культуры поддерживали при 28°C в течение 24-48 ч при перемешивании при 200 об/мин.
Экспрессия Р450-ферментов может быть оценена качественно и количественно путем измерения спектра связывания СО (Omura, Т. & Sato, R. (1964) J. Biol. Chem. 239, 2379-2387) в белковых фракциях Е. coli. Для экстракции белка, клетки центрифугировали (10 мин, 5000 g, 4°C) и ресуспендировали в 35 мл ледяного буфера 1 (100 мМ Трис-HCl pH 7,5, 20% глицерин, 0,5 мМ EDTA). Добавляли один объем 0,3 мг/мл лизоцима (из куриного яичного белка, «Sigma-Aldrich») в воде, и суспензию оставляли на 10-15 мин при 4°C при перемешивании. Суспензию центрифугировали 10 мин при 7000 g и 4°C и осадок ресуспендировали в 20 мл буфера 2 (25 мМ КРO4 pH 7,4, 0,1 мМ EDTA, 0,1 мМ DTT, 20% глицерина). Суспензию подвергали одному циклу замораживания оттаивания при -80°C, добавляли 0,5 мМ PMSF (фенилметилсульфонил фторид, «Sigma-Aldrich»), и суспензию обрабатывали ультразвуком, 3 раза в течение 20 сек. Суспензию центрифугировали 10 мин при 10000 g (для удаления клеточного дебриса), супернатант собирали и центрифугировали 2 часа при 100000 g. Осадок (фракция мембранных белков) ресуспендировали в 2-3 мл буфера 3 (50 мМ Трис-HCl pH 7,4, 1 мМ EDTA, 20% глицерин). Для измерения СО-спектра, белковую фракцию разбавляли (1/10) в буфере 3 до конечного объема, равного 2 мл. Добавляли несколько кристаллов дитионита натрия (Na2S2O4), образец разделяли между двумя кюветами и регистрировали базовую линию от 370 до 500 нм. Затем кювету с образцом насыщали монооксидом углерода и регистрировали разностный спектр. Концентрация Р450-фермента можно определить по амплитуде пика при 450 нм, применяя коэффициент экстинкции для восстановленного комплекса CO, равный 91 мМ-1⋅см-1 (Omura, Т. & Sato, R. (1964) J. Biol. Chem. 239, 2379-2387).
Следуя этой процедуре, можно измерять типичные СО-спектры с максимумом поглощения при 450 нМ для рекомбинантных белков VzP521-11-1 (SEQ ID NO: 35) и VzP521-16-1 (SEQ ID NO: 36), подтверждающие правильное сворачивание в функциональные Р450-ферменты (фигура 4).
Пример 5
Гетерологичная экспрессия растительных Р450-редуктаз в бактериях
Для реконструкции активности растительных Р450, важно присутствие второго мембранного белка. Данный белок, Р450-редуктаза (CPR), вовлечен в перенос электронов от NADPH (никотинамидадениндинуклеотид фосфата, восстановленная форма) к активному центру Р450. Было показано, что CPR из одного растения может дополнять активность Р450 фермента из другого растения (Jensen и Moller (2010) Phytochemsitry 71, 132-141).
Сообщалось о нескольких CPR-кодирующих нуклеотидных последовательностях из различных растительных источников. Например, две различные CPR, ATR1 и ATR2 (номера доступа в NCBI САА46814.1 и САА46815), были идентифицированы в Arabidopsis thaliana (Urban et al. (1997) J. Biol. Chem. 272 (31) 19176-19186). Было показано, что данные CPR дополняют некоторые Р450-ферменты из разных видов растений. кДНК (последовательность tcATR1-opt (SEQ ID NO: 29), кодирующую усеченную версию ATR1 (делеция 17-ти аминокислот с N-конца), синтезируют применяя частоту использования кодона, оптимальную для экспрессии в Е. coli (DNA 2.0, Menlo Park, СА, США), и включая сайты рестрикции NcoI и BamHI на 5'-конце и 3'-конце, соответственно. Данную кДНК лигировали в плазмиду pJ206 (DNA2.0, Menlo Park, СА, США), обеспечивая плазмиду pJ206-tcATR1-opt. Вставку вырезали из плазмиды pJ206-tcATR1-opt с помощью рестриктаз NcoI и BamHI и между соответствующими сайтами рестрикции лигировали экспрессионную плазмиду pACYCDuet-1 («Novagen», Merck Chemicals), обеспечивая плазмиду pACYC-tcATR1-opt. Функциональная экспрессия CPRs в клетках Е. coli может быть оценена последующим энзиматическим восстановлением цитохрома С. Плазмиду pACYC-tcATR1-opt применяли для трансформации клеток Е. coli B121 (DE3) («Novagen») или JM109 (DE3) («Promega», Madisson, WI, США). Условия культивирования, экспрессия белка и получение бесклеточного препарата белка выполняли, как описано в примере 4. Белки разводили в 1 мл Трис pH 7,4, дополненном 5 мкМ FAD, 5 мкМ FMN, 40 мМ цитохрома С (««Sigma»-Aldrich»), 1 мМ MgCl2. Реакцию начинали добавлением 0,12 ммоль NADPH («Sigma»). Восстановление цитохрома С регистрировали путем измерения увеличения оптической плотности OD при 550 нМ в течение 0,5-2 минут. Рассчитывали специфическую редуктазную активность (в мЕд./мкл), применяя следующую формулу: (ODконечная-ODначальная)/21/время (сек)/Объем (мкл)Х60000 (в мЕд./мкл). Типичная активность, измеренная с рекомбинантной ATR1, варьировала от 7 до 10 мЕд./мл.
Пример 6
Совместная экспрессия Р450 и Р450 редуктазы с помощью двух плазмид
Для биотрансформации целой клетки, с помощью растительных Р450, необходима совместная экспрессия белков Р450 и CPR в одной клетке-хозяине. Данная совместная экспрессия может быть получена с применением двух плазмид. Например, клетки Е. coli BL21 Starтм (DE3) («Invitrogen», Carlsbad, СА) были совместно трансформированы плазмидой pACYC-tcATR1-opt и плазмидой pCW-218-521-11 или pCW-218-521-16. Трансформированным клетки отбирали на ЛБ-агарозных чашках с карбенициллином (50 мкг/мл) и хлорамфениколом (34 мкг/мл). Единичные колонии применяли для инокуляции в 5 мл жидкой ЛБ-среды, дополненной теми же антибиотиками. Культуру инкубировали в течение ночи при 37°C. На следующий день в 2-250 мл ТВ-среды, дополненной теми же антибиотиками и 1 мМ тиамином-HCL, инокулировали 0,2 мл ночной культуры. После 6-ти часов инкубации при 37°C, культуру охлаждали до 28°C и добавляли 1 мМ IPTG и 75 мг/л δ-аминолевулиновой кислоты. Культуру поддерживали от 24 до 36 часов. Белковые фракции получали, как описано в примере 4, и экспрессию рекомбинантных Р450 и CPR оценивали, применяя методику, описанную в примерах 4 и 5, соответственно.
Пример 7
Совместная экспрессия Р450 и Р450 редуктазы из единственной плазмиды
Получали экспрессионную плазмиду с бицистронным конструктом, включавшим кДНК, кодирующую Р450 ветивера и кДНК, кодирующую CPR. Конструкты были разработаны для вставки между двумя кодирующими областями последовательности спейсера, включавшей сайт связывания рибосом (RBS) (смотри фигуру 5).
кДНК tcATR1-opt (SEQ ID NO: 29), синтезированная с оптимальной частотой использования кодона Е. coli (DNA 2.0, Menlo Park, СА, США), была модифицирована для добавления на 5'-конец, перед старт-кодоном, удлиняющего сегмента, размером в 30 по, содержавшего последовательность спейсера (SEQ ID NO: 30) и последовательность RBS. кДНК tcATR1-opt амплифицировали, применяя праймеры 2390-CPR-F2 (SEQ ID NO: 31) и 2390-CPR-R2b (SEQ ID NO: 32), применяя ДНК-полимеразу Pfu («Promega», Madison, WI, США), в конечном объеме, равном 50 мкл, содержавшим 5 мкл 10Х буфера ДНК полимеразы Pfu, 200 мкМ каждого из дНТФ, 0,4 мкМ каждого из праймеров, 2,9 Ед. ДНК полимеразы Pfu и 50 нг плазмиды pJ206-tcATR1-opt в 2,5 мкл. Условия термоциклирования были следующими: 1,5 мин при 95°C; 30 циклов, включающих 45 сек при 95°C, 30 сек при 60°C и 4 мин при 72°C; и 10 мин при 72°C. После очистки на агарозном геле, ПЦР-продукт лигировали в плазмиды pCW-218-521-11 и pCW-218-521-16, расщепленные с помощью HindIII, применяя набор для клонирования In-Fusion® Dry-Down PCR Cloning Kit («Clontech», Takara Bio Europe) и следуя инструкциям производителя. Полученные две плазмиды pCW-2391-521-11 и pCW-2392-521-16 содержали бицистронные конструкты, состоявший из последовательностей VzP521-11-1 и VzP521-16-1, за которыми, соответственно, следовала последовательность tcATR1-opt.
Е. coli клетки трансформировали одной из двух данных плазмид, и получали фракции мембранного белка, как описано в примере 4. Экспрессию Р450 и CPR подтверждали с помощью СО-связывающих спектров и экспериментов по восстановлению NADPH, как описано в примерах 4 и 5.
Пример 8
Биопревращение зизаена в кусимол с применением целой клетки Е. coli, экспрессирующей Р450 ветивера и CPR
Окисление (+)-зизаена может быть выполнено с применением целой клетки Е. coli, экспрессирующей Р450 ветивера и CPR (биопревращение). Зизаен получали, применяя сконструированные клетки Е. coli, следуя способу, описанному в патенте WO 2010/134004 и, применяя сесквитерпен-синтазу с последовательностью, имеющую номер доступа HI931369.
Кратко, клетки Е. coli BL21Starтм (DE3) («Invitrogen» Ltd) трансформировали плазмидой pACYC-4506 и плазмидой pETDuet-VzZS-opt. Плазмида pACYC-4506 содержит гены, кодирующие пять ферментов биосинтетического пути превращения мевалоновой кислоты в FPP: мевалонаткиназу (MvaK1), фосфомевалонаткиназу (MvaK2), мевалонатдифосфат-декарбоксилазу (MvaD), изопентенилдифосфат-изомеразу (idi) и фарнезилдифосфат-синтазу (FPS). Для конструирования данной плазмиды, ген FPS амплифицировали из геномной ДНК S. cerevisiae и лигировали в первый сайт множественного клонирования (MCS) в pACYCDuet-1, оперон, кодирующий гены для MvaK1, MvaK2, MvaD и idi, амплифицировали из геномной ДНК Streptococcus pneumoniae (АТСС ВАА-334) и лигировали во второй MCS. pETDuet-VzZS-opt содержал кодон-оптимизированную версию (+)-зизаен-синтазы ветивера (как описано в SEQ ID NO: 11 в WO 2010/134004).
Единичные колонии трансформированных клеток применяли для инокуляции в 5 мл ЛБ-среды, дополненной карбенициллином (50 мг/мл) и хлорамфениколом (34 мг/мл). Культуры инкубировали в течение ночи при 37°C. На следующий день в 1 л Terrific Broth (ТВ)-среды, дополненный теми же антибиотиками, инокулировали 1/100 объема ночной культуры. Через 6 ч инкубации при 37°C, культуры охлаждали до 28°C и к культурам добавляли 1 мМ IPTG, 2 г/л мевалоновой кислоты, полученной путем разведения мевалонолактона (««Sigma»-Aldrich») в 0,5N NaOH в концентрации 1 г/мл и инкубацией раствора в течение 30 мин при 37°C), и 100 г/л смолы Amberliteтм XADтм-4 (Rhom и Haas). Через 48 ч инкубации, смолу извлекали, промывали водой и элюировали 3-мя объемами диэтилового эфира. Растворитель удаляли, и продукт очищали на силикагеле флэш-хроматографией, применяя н-гексан в качестве растворителя. Фракции, содержащие (+)-зизаен объединяли, растворитель удаляли дистилляцией, и остатки применяли в качестве субстрата для анализа по окислению.
Клетки Е. coli (BL21 Starтм (DE3) («Invitrogen» Ltd) или E. coli клетки JM109(DE3) («Promega»)) трансформировали плазмидами pCW-2391-521-11 или pCW-2392-521-16 или клетки были совместно трансформированы плазмидами pCW-218-521-11 или pCW-218-521-16 и pACYC-tcATR1-opt и растили на ТВ-среде, дополненной 3% глицерина, или на ЛБ-среде, дополненной 1% глюкозой. Культуры инкубировали при 37°C до тех пор, пока величина оптической плотности не достигала 1. Температуру культур затем снижали до 28°C, добавляли 1 мМ IPTG и 74 мкг/мл δ-аминолевулиновой кислоты и культуры инкубировали в течение 24 часов.
Клетки собирали на экспоненциальной фазе роста, центрифугировали и ресуспендировали в 0,5 объема калий-фосфатного буфера 50 мМ pH 7,0, дополненного 5% глицерин или 3% глюкозы. Субстрат ((+)-зизаен) добавляли до конечной концентрации 0,5 мг/мл в виде смеси, состоявшей из 10 мг Tween® 20 (««Sigma»-Aldrich»), 10 мг противовспенивателя (Erol DF, PMC Ouvrie, Lesquin, Франция), 20 мг (+)-зизаена и 1 мл воды. Превращение протекало в течение 24-х часов при 20°C при умеренном перемешивании. Среду экстрагировали 2 объемами этилацетата, и экстракт анализировали путем GCMS с помощью системы Agilent 6890 Series GC, соединенной масс-детектором Agilent 5975. GC была оборудована 30 м капиллярной колонкой SPB-1 с 0,25 мм внутренним диаметром (Supelco, Bellefonte, РА). Газ-носитель представлял собой He, который поступал с постоянной скоростью, равной 1 мл/мин. Исходная температура печи была равна 50°C (поддерживали 1 мин), затем температуру увеличивали со скоростью 10°C/мин до 300°C. Идентификация продуктов была основана на сравнении масс-спектров и индексов удерживания с аутентичными стандартами и внутренними базами данных.
В данных условиях окисление (+)-зизаена наблюдали в опытах с клетками, содержавшими рекомбинантные белки VzP521-11-1 и VzP521-16-1. В результате успешного окисления (+)-зизаена наблюдали и идентифицировали с помощью GCMS-анализа три продукта: кусимол, зизаналь и зизановую кислоту (фигура 6). Ферменты VzP521-11-1 и VzP521-162-1 катализировали, таким образом, окисление (+)-зизаена до кусимола. Дополнительное окисление кусимола до зизаналя и зизановой кислоты могло катализироваться рекомбинантными Р450 или эндогенными энзиматическими активностями Е. coli (фигура 7).
Пример 9
Биопревращение других молекул монотерпена и сесквитерпена с применением целой клетки Е. coli, экспрессирующей Р450 ветивера и CPR
Получали и растили клетки Е. coli, экспрессировавшие Р450 ветивера и CPR, и затем осуществляли биопревращения, применяя покоящиеся клетки в калий-фосфатном буфере, как описано в примере 8.
Анализ проводили с некоторыми молекулами терпенов и образование оксигенированных молекул терпенов оценивали, применяя GCMS-анализ, как описано в примере 8. Помимо (+)-зизаена, биопревращение наблюдали со следующими молекулами: (+)-лимонен, (-)-лимонен, альфа-пинен, альфа-цедрен, альфа-лонгипинен, альфа-фунебрен, туйопсен, валенсен, бета-чамигрен, аллоаромадендрен, альфа-неокловенен, изосативен, леден, альфа-гумулен, альфа гурюнен, гамма-гурюнен, бета-фунебрен, альфа-копаен, альфа-гурюнен и бета-пинен. Структуры данных молекул показаны на фигуре 1. Примеры хроматограмм, полученных с помощью GCMS-анализа данных биопревращений, показаны на фигуре 8. Для некоторых протестированных субстратов, продукты, которые могли быть идентифицированы, показаны на фигуре 9.
Пример 10
In-vitro окисление соединений с помощью Р450 ветивера
Окисление сесквитерпена с помощью Р450 из ветивера также может быть проведено in-vitro с применением клеточных лизатов или частично очищенного белка.
Клетки Е. coli (BL21 Starтм(DE3) («Invitrogen» Ltd) или клетки Е. coli JM109 (DE3) («Promega»)) трансформировали плазмидами pCW-2391-521-11 или pCW-2392-521-16 или клетки совместно трансформировали плазмидами pCW-218-521-11 или pCW-218-521-16 и pACYC-tcATR1-opt. Условия культивирования клеток, экспрессия белков и получение мембранных белков были такими же, как описано в примерах 4 и 5. Данные фракции белков применяли для in-vitro превращения (+)-зизаена или молекул терпенов, перечисленных в примере 9. Типичные анализы состояли из 20-50 мкл белка, 0,4 мг NADPH («Sigma»), 5 мкМ FAD (флавинадениндинуклеотид, «Sigma»), 5 мкМ FMN (флавинмононуклеотид, «Sigma»), 0,05 мг (+)-зизаена в общем объеме, равном 1 мл 100 мМ Трис буфера, pH 7,4. В некоторых анализах, добавляли NADPH-регенерирующую систему, состоявшую из 25 мМ глюкозо-6-фосфата («Sigma») и 6 мЕд. глюкозо-6-фосфат дегидрогеназы («Sigma»). Пробы инкубировали 2-12 часов при 30°C.Затем образцы дважды экстрагировали одним объемом этилацетата и анализировали с помощью GCMS, как описано в примере 8.
Применяя этот подход, получали такие же продукты, как были получены при применении целых клеток Е. coli, как описано в примерах 8 и 9.
Пример 11
In-vivo продукция кусимола в сконструированных клетках
Окисленные продукты (+)-зизаена также могут быть продуцированы в клетках Е. coli, сконструированных для получения сесквитерпенов из такого источника углерода, как глюкоза или глицерин. Полученные плазмиды состояли из pCWori плазмид (Barnes H.J (1996) Method Enzymol. 272, 3-14)), содержавших оперон, состоявший из Р450, Р450 редуктазы и терпен-синтазы. Таким образом, путем вставки последовательности RBS и оптимизированной последовательности, кодирующей (+)-зизаен-синтазу (VzZS), после стоп-кодона последовательности CPR в pCW-2391-521-11 или в pCW-2392-521-16 были получены две плазмиды.
VzZS амплифицировали из плазмиды pETDuet-VzZS-opt (как описано в WO 2010/134004), применяя праймеры 2401-VzZS-F (SEQ ID NO: 33) и 2401-VzZS-R (SEQ ID NO: 34). ПЦР проводили, применяя ДНК-полимеразу Pfu («Promega», Madison, WI, США), в конечном объеме, равном 50 мкл, содержавшем 5 мкл 10Х буфера ДНК-полимеразы Pfu, 200 мкМ каждого из дНТФ, 0,4 мкМ каждого из праймеров, 2,9 Ед. ДНК-полимеразы Pfu и 50 нг матрицы. Условия термоциклирования были следующими: 1,5 мин при 95°C; 30 циклов, включающих 45 сек при 95°C, 30 сек при 60°C и 4 мин при 72°C; и 10 мин при 72°C. ПЦР-продукт очищали и лигировали в плазмиды pCW-2391-521-11 или pCW-2392-521-16, расщепленные рестриктазами HindIII и EcorI. Данное лигирование проводили, применяя набор для клонирования In-Fusion® Dry-Down PCR Cloning Kit («Clontech», Takara Bio Europe), следуя инструкциям производителя. Полученные плазмиды pCW-2401-521-11 и pCW-2402-521-16 содержали вставку, состоящую из последовательности VzP521-11-1 или VzP521-16-l, соответственно, последовательности tcATR1-opt (Р450 редуктаза) и последовательности VzZS ((+)-зизаен-синтаза) (фигура 10).
Получали еще одну экспрессионную плазмиду, содержавшую два оперона, состоявших из генов, кодирующих ферменты для полного мевалонатного пути. Первый синтетический оперон, состоявший из генов ацетоацетил-СоА тиолазы Escherichia coli (atoB), HMG-CoA-синтазы Staphylococcus aureus (mvaS), HMG-CoA редуктазы Staphylococcus aureus (mvaA) и FPP-синтазы Saccharomyces cerevisiae (ERG20), был химически синтезирован (DNA2.0, Menlo Park, CA, США) и лигирован в расщепленный с помощью NcoI-BamHI вектор pACYCDuet-1 («Invitrogen») с получением pACYC-29258. Гены данной плазмиды кодируют ферменты, необходимые для превращения ацетил-СоА в мевалонат, и FPP-синтазу. Второй оперон содержал гены, кодирующие четыре фермента, необходимые для превращения мевалоната в IPP и DMAPP, и его амплифицировали из плазмиды pACYC-4506 (Пример 8), применяя праймеры MVAup1-старт (SEQ ID No: 39) и MVAup2-стоп (SEQ ID No: 40). Полученный ампликон клонировали в вектор pACYC-29258, который расщепляли с помощью NdeI-XhoI, применяя набор для клонирования In-Fusion 2.0 Dry-Down PCR Cloning Kit («Clontech»), и получали вектор pACYC-29258-4506.
Клетки E.coli OverExpressтм C43(DE3) (Lucigen® Corporation) были совместно трансформированы плазмидой pACYC-29258-4506 и плазмидой pCW-2401-521-11 или плазмидой pCW-2402-521-16. Единичные колонии трансформированных клеток применяли для инокуляции в 5 мл ЛБ-среды, дополненной карбенициллином (100 мкг/мл) и хлорамфениколом (17 мкг/мл). Культуры инкубировали в течение ночи при 37°C и, применяли для инокуляции в М9-среду, дополненную 2 г/л дрожжевым экстрактом, 3% глицерином, 10 мкМ FeSO4, 100 мкг/мл карбенициллина и 17 мкг/мл хлорамфеникола. Культуры инкубировали 6 ч при 37°C, охлаждали до 25°C и добавляли 0,1 мМ IPTG, 74 мкг/мл δ-аминолевулиновой кислоты и 1/10 объема до декана. Через 48 часов, культуры экстрагировали 2-мя объемами трет-бутил метилового эфира (МТВЕ), и экстракт анализировали с помощью GCMS, как описано в примере 8, за исключением того, что температуру печи исходно выставляли на 80°C на одну минуту, затем постепенно увеличивали со скоростью 10°C/мин до 300°C.
Были зарегистрированы сесквитерпен (+)-зизаен и произведенные из него спирт и альдегид (кусимол и зизаналь). Зизановая кислота также была обнаружена в виде минорного продукта (фигура 11). Этот эксперимент показал, что в сконструированных клетках, сесквитерпен (+)-зизаен продуцируется и окисляется под действием гетерологичного комплекса Р450 и CPR.
Данный пример показывает, что клетка Е. coli, трансформированная для экспрессии полипептида в соответствии с изобретением, способна окислять соединения терпена, такие как зизаен, при условии, что его применяют в комбинации с Р450-редуктазой. Другие ферменты, которыми трансформируют клетку Е. coli, не представляют собой ферменты, необходимые для такого окисления. Действительно, окисленный терпен также продуцируется, если клетку Е. coli трансформируют только цитохромом Р450, редуктазой и терпен-синтазой, но в меньшем количестве. Такие другие ферменты, которыми трансформируют клетку Е. coli, добавляли с единственной целью увеличения количества терпена, доступного в качестве субстрата для цитохрома Р450.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКИХ СПИРТОВ | 2014 |
|
RU2654670C2 |
| НОВЫЙ ПОЛИПЕПТИД СИСТЕМЫ ЦИТОХРОМА Р450 С ПОВЫШЕННОЙ ФЕРМЕНТНОЙ АКТИВНОСТЬЮ | 2015 |
|
RU2677997C2 |
| НОВЫЕ ФЕРМЕНТЫ МЕТАНОЛДЕГИДРОГЕНАЗЫ ИЗ Bacillus | 2013 |
|
RU2624017C2 |
| Микроорганизм для продуцирования L-аминокислоты, обладающий повышенной активностью цитохрома С, и способ получения L-аминокислоты с его использованием | 2020 |
|
RU2819270C1 |
| ГЕНЫ И БЕЛКИ ДЛЯ БИОСИНТЕЗА ЛАНТИБИОТИКА 107891 | 2007 |
|
RU2441915C9 |
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ ДЕФЕНСИНОВ В МИЦЕЛИАЛЬНЫХ ГРИБАХ | 2006 |
|
RU2404990C2 |
| НОВЫЕ ЛЮЦИФЕРАЗЫ И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2017 |
|
RU2674894C2 |
| СПОСОБЫ ФЕРМЕНТАТИВНОГО СИНТЕЗА МОГРОЗИДНЫХ СОЕДИНЕНИЙ | 2012 |
|
RU2633644C2 |
| БЫСТРЫЙ СПОСОБ КЛОНИРОВАНИЯ И ЭКСПРЕССИИ СЕГМЕНТОВ ГЕНА РОДСТВЕННОЙ ВАРИАБЕЛЬНОЙ ОБЛАСТИ АНТИТЕЛА | 2012 |
|
RU2612910C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ, КОТОРЫЕ ПРИДАЮТ УСТОЙЧИВОСТЬ К НАСЕКОМЫМ-ВРЕДИТЕЛЯМ ОТРЯДА ЖЕСТКОКРЫЛЫХ | 2011 |
|
RU2639549C2 |

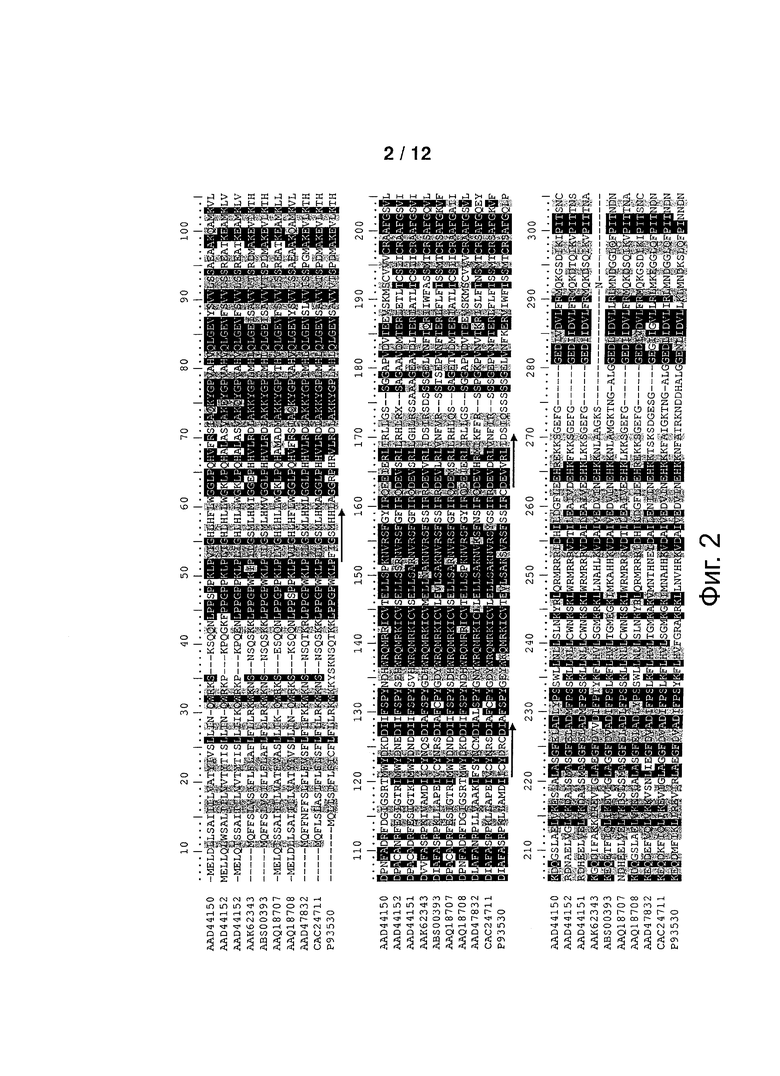


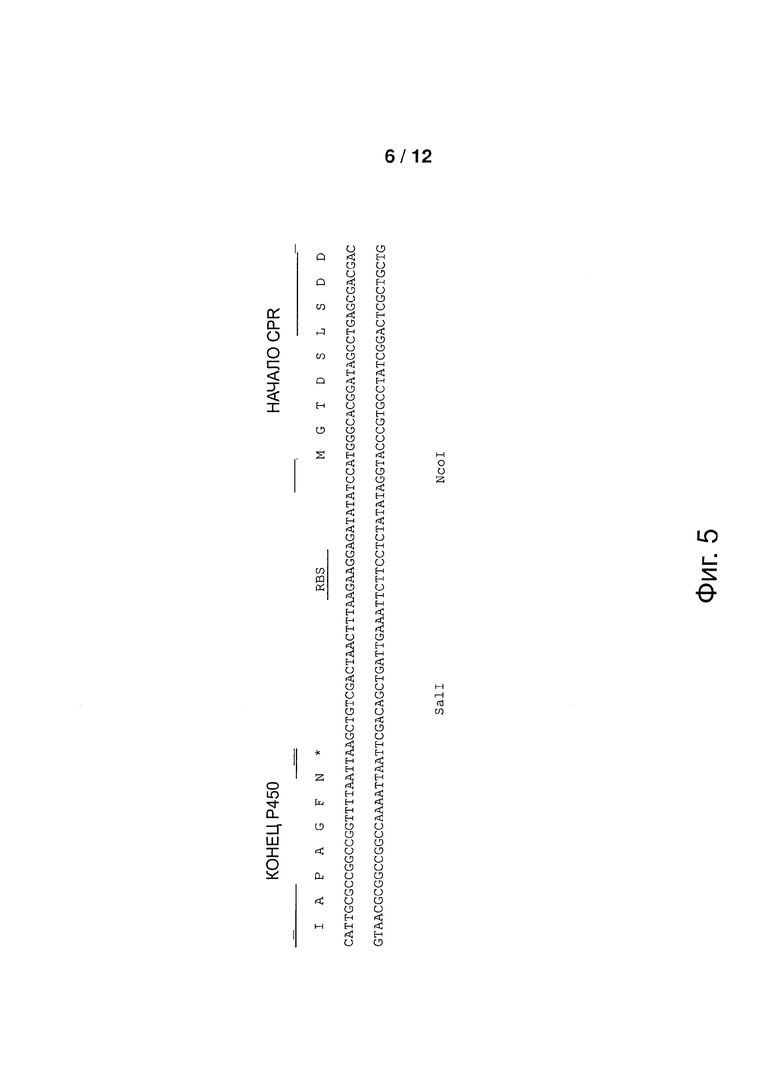
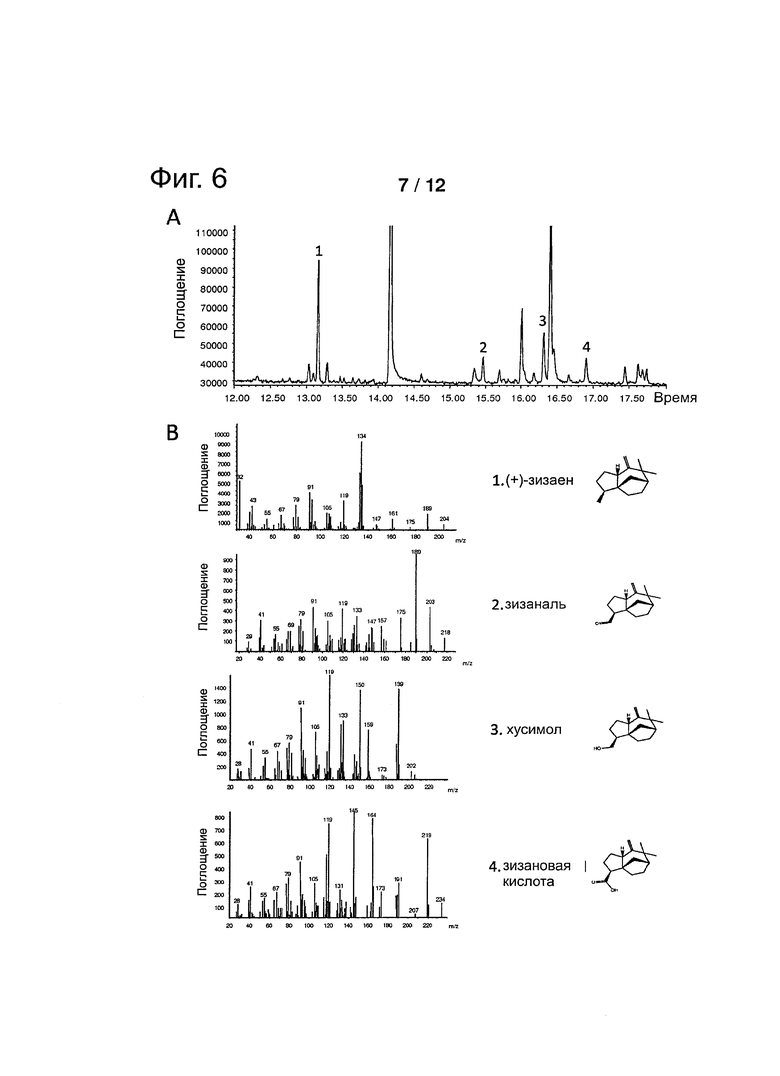
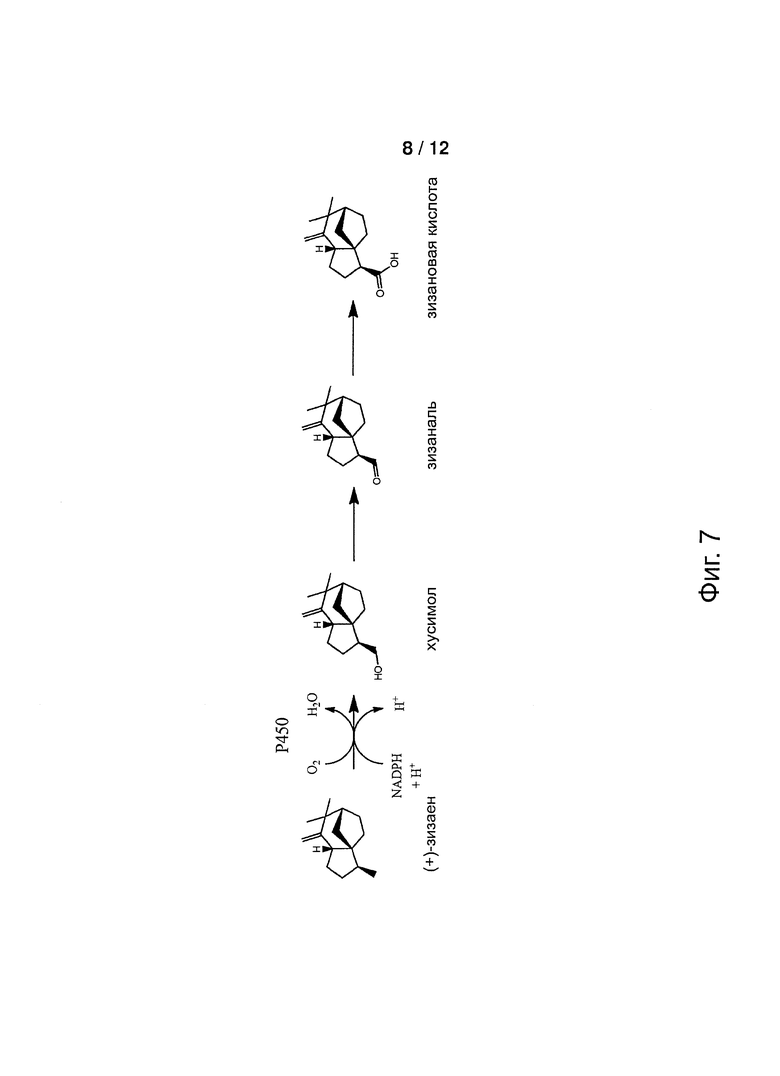

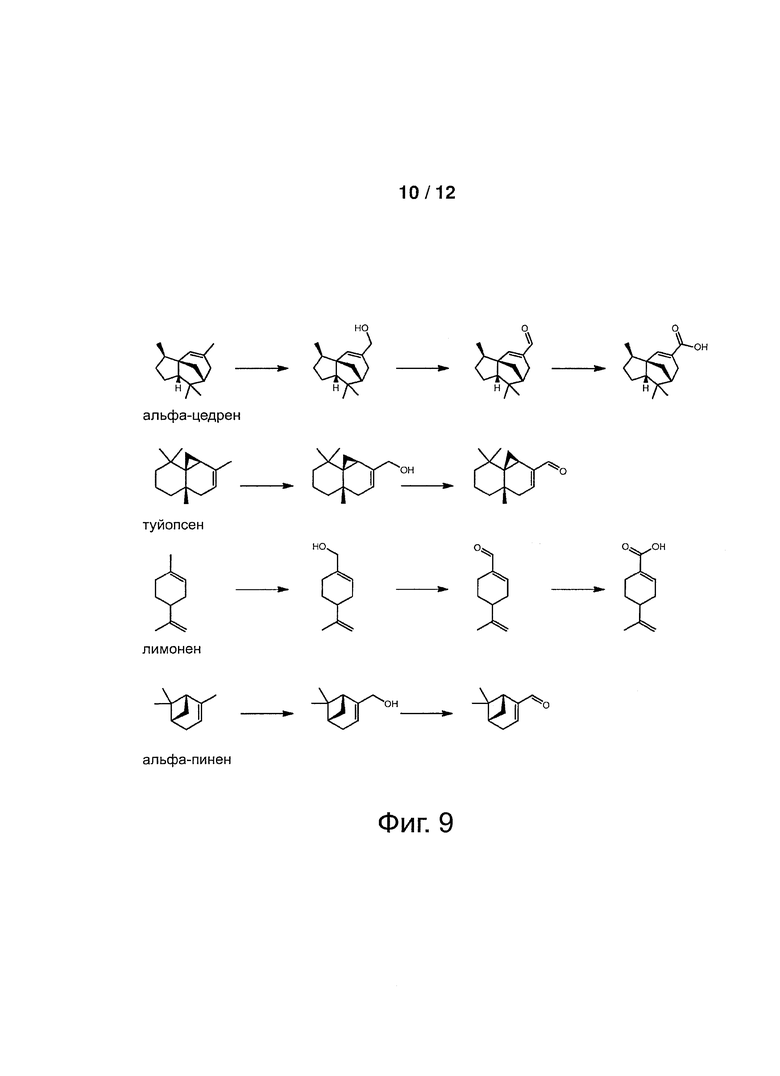
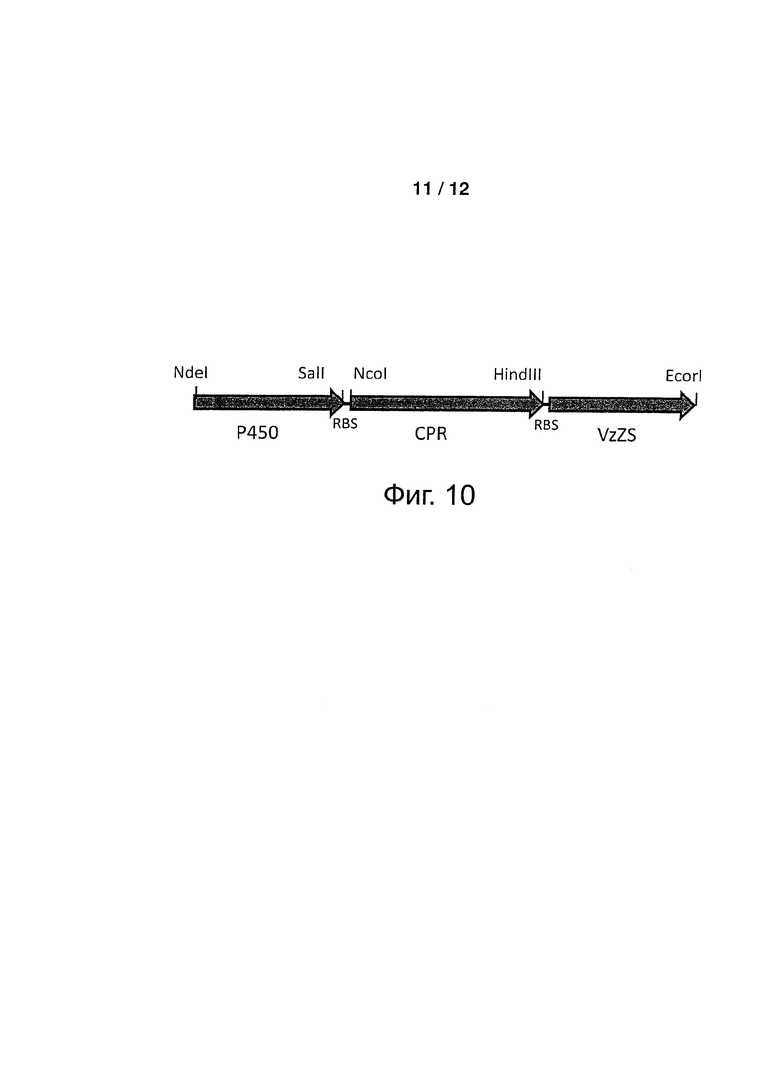

Настоящее изобретение относится к биохимии, в частности к полипептиду, обладающему активностью цитохрома P450. Указанный полипептид способен окислять такие соединения терпена, как моно- или полициклические монотерпены и сесквитерпены. Настоящее изобретение обеспечивает нуклеиновую кислоту, кодирующую указанный полипептид, вектор экспрессии, включающий указанную нуклеиновую кислоту, а также различные организмы-хозяины и клетки-хозяины, не относящиеся к человеку, и способ окисления терпенов с использованием указанного полипептида, обладающего активностью цитохрома P450. Настоящее изобретение позволяет расширить арсенал средств для окисления различных соединений терпена. 6 н. и 15 з.п. ф-лы, 11 ил., 3 табл., 11 пр.
1. Полипептид, включающий аминокислотную последовательность, по меньшей мере, на 85% идентичную последовательностям SEQ ID NO: 1 или 2, и обладающий активностью цитохрома Р450, характеризуемый тем, что полипептид способен окислять, по меньшей мере, одно соединение терпена, которое выбирают из моно- или полициклических монотерпенов и сесквитерпенов.
2. Полипептид по п. 1, включающий аминокислотную последовательность, по меньшей мере, на 90% идентичную последовательностям SEQ ID NO: 1 или 2.
3. Полипептид по п. 1, характеризуемый тем, что моно- или полициклический монотерпен или сесквитерпен включает, по меньшей мере, одну метильную группу в качестве заместителя в циклической части молекулы.
4. Полипептид по п. 3, характеризуемый тем, что моно- или полициклический монотерпен или сесквитерпен выбирают из группы, состоящей из зизаена, альфа-цедрена, альфа-лонгипинена, альфа-фунебрена, туйопсена, валенсена, бета-чамигрена, аллоаромадендрена, альфа-неокловена, изосативена, ледена, S-лимонена, альфа-гумулена, альфа-гурюнена, альфа-пинена, бета-фунебрена, R-лимонена и бета-пинена.
5. Полипептид по п. 4, характеризуемый тем, что моно- или полициклический монотерпен или сесквитерпен выбирают из группы, состоящей из зизаена, альфа-цедрена, альфа-фунебрена, валенсена и туйопсена.
6. Нуклеиновая кислота, кодирующая полипептид по любому из пп. 1-5.
7. Нуклеиновая кислота по п. 6, включающая нуклеотидную последовательность, по меньшей мере, на 90%, идентичную последовательностям SEQ ID NO: 3, 4.
8. Вектор экспрессии, включающий нуклеиновую кислоту по любому из пп. 6 и 7, где вектор имеет форму вирусного вектора, бактериофага или плазмиды.
9. Организм-хозяин, не представляющий собой человека, для экспрессии полипептида, где организм-хозяин не представляет собой организм человека, и трансформирован для несения, по меньшей мере, одной нуклеиновой кислоты по любому из пп. 6 и 7.
10. Организм-хозяин, не представляющий собой человека, по п. 9, где указанный организм-хозяин, не представляющий собой человека, представляет собой растение, прокариота или гриб.
11. Организм-хозяин, не представляющий собой человека, по п. 10, где указанный организм-хозяин, не представляющий собой человека, представляет собой микроорганизм.
12. Организм-хозяин, не представляющий собой человека, по п. 11, где указанный микроорганизм представляет собой бактерии или дрожжи.
13. Организм-хозяин, не представляющий собой человека, по п. 12, где указанные бактерии представляют собой Е. coli и указанные дрожжи представляют собой Saccharomyces cerevisiae.
14. Клетка-хозяин для экспрессии полипептида, где клетка-хозяин не представляет собой клетку человека, и трансформирована для несения, по меньшей мере, одной нуклеиновой кислоты по любому из пп. 6 и 7.
15. Клетка-хозяин, не представляющая собой клетку человека, по п. 14, которая представляет собой растительную клетку.
16. Способ окисления, по меньшей мере, одного соединения терпена, включающий:
a) взаимодействие указанного соединения терпена, по меньшей мере, с одним полипептидом по любому из пп. 1-5;
b) необязательно, выделение окисленного терпена, полученного на стадии а).
17. Способ по п. 16, в котором указанное соединение терпена выбирают из моно- или полициклических монотерпенов и сесквитерпенов.
18. Способ по п. 17, в котором указанное соединение терпена выбирают из группы, состоящей из зизаена, альфа-цедрена, альфа-лонгипинена, альфа-фунебрена, туйопсена, валенсена, бета-чамигрена, аллоаромадендрена, альфа-неокловена, изосативена, ледена, S-лимонена, альфа-гумулена, альфа-гурюнена, альфа-пинена, бета-фунебрена, R-лимонена и бета-пинена.
19. Способ по п. 18, характеризуемый тем, что соединение терпена выбирают из группы, состоящей из зизаена, альфа-цедрена, альфа-фунебрена, валенсена и туйопсена.
20. Способ по п. 18, в котором соединение терпена далее окисляют до первичного спирта, альдегида и/или карбоновой кислоты.
21. Способ по п. 19, в котором зизаен далее окисляют до кусимола, зизанала и/или зизаноевой кислоты.
| WO 2009047498 A2, 16.04.2009 | |||
| EP 1131440 B1, 12.09.2001 | |||
| НОВЫЕ ЦИТОХРОМ Р450-МОНООКСИГЕНАЗЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ОКИСЛЕНИЯ ОРГАНИЧЕСКИХ СОЕДИНЕНИЙ | 2000 |
|
RU2285044C2 |

