Настоящее изобретение относится к мутанту цитохром Р-45cam монооксигеназы и к способу окисления некоторых органических соединений с помощью этого мутанта.
Монооксигеназы катализируют селективное окисление нефункциональных углеводородов с использованием кислорода [1] и поэтому представляют большой интерес для возможного применения в органическом синтезе. Однако сложности возникают с выделением достаточных количеств фермента и связанных электронотранспортных белков. Несмотря на доступность аминокислотных последовательностей более чем 150 различных цитохром Р-450 монооксигеназ, современные структурные данные есть только для трех [2,3,4] и незначительное число успешно сверхэкспрессировано в бактериальных системах.
Одной из цитохром Р-450 монооксигеназ, которая растворима и может быть выделена в достаточных количествах, является высокоспецифичная Р-450cam из P. putida, которая катализирует регио- и стереоселективное гидроксилирование камфоры (1) в 5-экзо-гидроксикамфору [6]. Структура кристалла Р-450cam определена с высоким разрешением, и так как механизм действия этого бактериального фермента, как полагают, весьма сходен с механизмом действия его аналогов у млекопитающих, его используют в качестве основы, на которой базируются модели ферментов млекопитающих.
Описаны нуклеотидная последовательность и соответствующая аминокислотная последовательность Р-450cam [5] . Известно расположение активного участка фермента и исследована структурно-функциональная взаимосвязь [13,14]. Описаны мутанты Р-450cam в положениях 101, 185 и 247 [15] и в положении 87 [16]. Описан мутант, в котором тирозин 96 заменен на фенилаланин-96 [12,17,18]. Однако все случаи описывают влияние мутаций на механизм известных реакций окисления. Теории и предложения о том, как мутация могла бы быть использована для создания биокатализаторов для окисления различных субстратов, отсутствуют.
С целью нахождения новых биокатализаторов было осуществлено переконструирование активного участка Р-450cam таким образом, чтобы он был способен специфически окислять органические молекулы, которые не являются субстратами природных белков. Первоочередная задача заключалась во введении "ароматического кармана" в активный участок Р-450cam, что могло бы способствовать связыванию субстратов, содержащих ароматические боковые цепи.
Кроме того, был идентифицирован отдаленный от активного участка поверхностный остаток (цистеин-334), который влияет на обработку и стабильность белка. Цистеин приводит к нежелательной димеризации белка в процессе очистки и поэтому для избежания этих двух свойств цистеин был замещен на аланиновый остаток.
Трехмерная структура Р-450cam показывает, что активный участок обеспечивает близкое к ван-дер-Ваальсовому взаимодействие с гидрофобными группами камфоры, как это показано на фиг. 1. Три ароматических остатка (Y96, F87 и F98) сгруппированы вместе и образуют связывающий субстрат карман с водородной связью между тирозином 96 и карбонильным кислородом камфоры, поддерживая субстрат в ориентации, подходящей для того, чтобы гарантировать регио- и стереоспецифичность реакции. Замещение любого из этих ароматических остатков на меньшую, гидрофобную неароматическую боковую цепь может обеспечить получение требуемого "ароматического кармана".
Для исследования вероятных эффектов точечных мутаций в трех ароматических остатках использовали молекулярное моделирование. Для расчета энергии взаимодействия между ароматической пробой и возможными мутантами цитохрома Р-450cam, где эти остатки заменяют на аланин (F87A, Y96A и F98A), применяли программу GRID [7]. Эти результаты затем были оценены графически с использованием пакета молекулярного моделирования Quanta [6].
Мутант F98A, как оказалось, имеет самое сильное связывающее взаимодействие внутри впадины активного участка, доступного ароматической пробе, взаимодействие Y96A немного меньше, a F87A существенно меньше. Было решено в первом случае подвергнуть мутации тирозин 96 до аланина, так как он занимает в связывающем кармане более центральное положение, тогда как фенилаланин 98 находится в выемке к одной стороне. Кроме того, удаление тирозина 96 должно уменьшить специфичность фермента по отношению к камфоре из-за потери водородной связи с субстратом.
В соответствии с одним из аспектов настоящего изобретения предлагается мутант цитохром Р-450cam монооксигеназы, в котором тирозиновый остаток в положении 96 и/или остаток цистеина в положении 334 заменены на другой аминокислотный остаток, за исключением фенилаланина.
В соответствии с другим аспектом настоящего изобретения предлагается мутант цитохром Р-450cam монооксигеназы, в котором тирозиновый остаток в положении 96 и/или остаток цистеина в положении 334 заменены на другой аминокислотный остаток, и этот мутант обладает свойством катализировать окисление любого из следующих соединений: полициклические ароматические углеводороды, линейные или разветвленные алканы, соединения дифенила и бифенила, включая галогенированные варианты таких соединений, и галогенированные углеводороды.
В соответствии с еще одним аспектом настоящего изобретения предлагается способ окисления соединения, выбираемого из полициклического ароматического углеводорода, линейного или разветвленного алкана, соединения дифенила или бифенила, включая галогенированные варианты такого соединения или галогенированного углеводорода, причем способ включает контактирование одного из выбранных соединений в окисляющих условиях с цитохром Р-450cam монооксигеназой, в которой тирозиновый остаток в положении 96 и/или цистеиновый остаток в положении 334 заменен на другой аминокислотный остаток.
Предпочтительно аминокислоту выбирают из одной из следующих аминокислот: аланин, аргинин, аспаргин, аспаргиновая кислота, цистеин, глутаминовая кислота, глутамин, глицин, гистидин, изолейцин, лейцин, лизин, метионин, пролин, серин, треонин, триптофан, тирозин и валин, за исключением того, что в случае тирозинового остатка в положении 96, аминокислота не является тирозином и в случае цистеинового остатка в положении 334 аминокислота не является цистеином.
В качестве аминокислот, которые заменяют тирозин в положении 96, удобно использовать одну из небольших гидрофобных аминокислот, то есть аланин, глицин, валин, лейцин или изолейцин, причем аланин является предпочтительным, как это показано ниже.
С другой стороны, аминокислота, заменяющая тирозин в положении 96, может быть одной из заряженных аминокислот, то есть отрицательно заряженной кислотой, такой, как аспаргиновая кислота или глутаминовая кислота, для связывания посредством водородной связи с положительно заряженным субстратом; или положительно заряженным соединением, таким, как лизин, агринин или гистидин, для связывания посредством водородной связи с отрицательно заряженным субстратом, который не является членом семейства камфоры.
Полагают, что мутация в положении 96 является ключом, который дает возможность мутировавшему ферменту катализировать окисление относительно широкого спектра органических субстратов. С целью изменить форму и специфичность активного участка фермента также могут быть мутированы аминокислоты, примыкающие к активному участку фермента. Эти другие аминокислоты включают в себя аминокислоты в положениях 87, 98, 185, 244, 247, 295 и 297. Предполагается, что аминокислота в одном или нескольких из этих положений может быть заменена на небольшую гидрофобную аминокислоту, чтобы увеличить активный участок; или большую гидрофобную аминокислоту, чтобы уменьшить размер активного участка; или на аминокислоту, имеющую ароматическое кольцо для связывания с соответствующим ароматическим кольцом субстрата.
При рассмотрении реакций окисления приведены условия, описанные в литературных ссылках. Ферментная система обычно включает путидаредоксин и путидаредоксинредуктазу вместе с восстановленной формой никотинамидадениндинуклеотида (NADH) в качестве кофакторов в дополнение к мутированному ферменту. Рассмотрены различные классы органических соединений:
I) Органическое соединение представляет собой ароматическое соединение, либо углеводород, либо соединение, используемое в условиях, в которых оно не дезактивирует и не денатурирует фермент. Вследствие мутации фермента с целью создания на его поверхности кармана, связывающего ароматику, мутантный фермент приобретает способность катализировать окисление широкого спектра ароматических соединений. Примеры окисления ароматических и полиароматических соединений показаны ниже в экспериментальной части и было обнаружено, что природный фермент катализирует окисление только представителей семейства камфоры.
II) Органическое соединение может представлять собой углеводород, то есть алифатический или алициклический, несущий функциональную группу. Перед окислением к функциональной группе присоединяется ароматическая защитная группа и удаляется с функциональной группы после реакции окисления. Подходящая ароматическая группа представляет собой фенильную группу. Предполагается, что ароматическая защитная группа удерживает субстрат на месте. Таким образом защитная группа, во-первых, делает субстрат более гидрофобным и, следовательно, увеличивает связывание с гидрофобным карманом фермента; во-вторых, она удерживает субстрат на месте на активном участке. Таким образом, с подходящей ароматической защитной группой может быть достигнуто и регио- и стереоселективное гидроксилирование субстрата. В качестве примера ниже в экспериментальной части описан циклогексилбензол.
Примерами монофункционализированных углеводородов являются циклогексильные, циклопентильные и алкильные производные (схема 1). Продукты окисления этих соединений используются в качестве исходных в органическом синтезе, в особенности при получении гомохиральной формы. Рассмотрен спектр ароматических защитных групп, то есть бензиловые или нафтиловые простые эфиры и сложные эфиры и амиды бензойной и нафтойной кислот (схема 1). Также представляют интерес бензоксазольные группы в качестве защитных групп для карбонильной группы и N- бензилоксазолидиновые группы в качестве защитных групп для альдегида. Обе могут быть легко сняты после ферментативного окисления и их использование описано в литературе ранее для микробного окисления альдегидов и кислот.
III) Органическое соединение представляет собой алифатический или алициклический углеводород от C5 до C12. Окисление циклогексана и линейных углеводородов показано ниже в экспериментальной части и было обнаружено, что природный фермент катализирует окисление только представителей семейства камфоры.
IV) Органическое соединение представляет собой галогенированный алифатический или алициклический углеводород. Ниже также описано окисление линдана (гексахлорциклогексана).
Фиг. 2 представляет собой газовую хроматограмму дифенилметана (А) и гидроксилированного продукта, образующегося при выдерживании с мутантом Y96A Р-450cam.
На основании вышеизложенного конструируют мутированные белки, которые содержат аланин, лизин, валин или фенилаланин вместо тирозина в положении 96 (Y96). Создают дополнительные мутанты, в которых эти замены в активном участке объединены с поверхностной мутацией цистеина в положении 334 на аланин. Наконец, в одном белке объединяют несколько мутаций активного участка и мутаций поверхности для составления фермента с несколькими мутациями. Гены, кодирующие цитохром Р-450cam и их природные переносящие электрон партнеры путидаредоксин и путидаредоксинредуктаза, амплифицируют из целой клеточной ДНК P. Putida с использованием цепьевой реакции полимеризации (PCR, ЦПР). Применяемые комбинации экспрессирующий вектор/носитель E.coli представляет собой pRH1091g в штамме JM109 для Р-450cam pUC118 в штамме JM109 для путидаредоксина и pGLW11 в штамме DH5oc для путидаредоксинредуктазы. Олигонуклеотиднаправленный сайт-специфичный мутагенез проводят с использованием субклона M13mp19 по методу Zoller и Smith [10], а селекцию мутанта проводят по методу Kunkel [11].
Показано, что мутант Y96A катализирует гидроксилирование камфоры (1), хотя по сравнению с природным ферментом реакция менее селективна, аналогично тому, как это сообщалось для мутанта Y96F [12]. Это уменьшение селективности может быть отнесено к потере водородной связи между Y96 и камфорой. Свойства белков дикого типа и Y96A в дальнейшем исследуют с помощью различных испытаний на связывание и активность.
Связывание потенциальных субстратов исследуют спектроскопическими методами. Природный фермент в отсутствие субстрата находится в 6-координированном низкоспиновом состоянии с слабосвязанной водой, занимающей шестое координационное положение, что проявляется в виде максимума Сорета (Soret) при 391 нм. Связывание аналогов субстрата адамантанона (2), адамантана (3) и норборнана (4) также полностью переводит гем в высокоспиновое состояние. Однако дифенилметан (5) не вызывает сдвига в спектре поглощения.
В то время как мутант Y96A дает те же результаты для соединений (3) и (4), он не полностью переводится в высокоспиновое состояние даже, когда соединения (1) и (2) добавляют в избытке. Однако наиболее интересным, в отличие от дикого типа, является то, что Y96A демонстрирует частичную конверсию в гем с высокой скоростью с дифенилметаном, что говорит о связывании этого соединения с мутировавшим белком.
Как и ожидалось, константа диссоциации (Kapp) для камфоры и адамантанона увеличиваются в Y96A. С другой стороны, величины Kapp для гидрофобных субстратов адамантана и норборнана уменьшаются, что говорит о том, что карман фермента становится более селективным по отношению к гидрофобным субстратам. Наибольшее изменение в связывании достигается в случае дифенилметана, который связывается с белком дикого типа слабо, но показывает сильно увеличенное сродство по отношению к мутанту Y96A (табл. 1).
После установления связывания дифениламина с белком Y96A, исследуют каталитическое превращение субстрата. Мутант белка переконструируют с помощью путидаредоксина и путидаредоксинредуктазы. Добавляют дифенилметан (5) и смесь выдерживают с восстановленной формой никотинамидадениндинуклеотида (NADH) и кислородом.
Раствор, содержащий 10 мкМ путидаредоксина, 2 мкМ путидаредоксинредуктазы, 1 мкМ цитохром Р-450cam монооксигеназы (дикий тип или мутанта) и 1 мМ дифенилметана в 100 мМ KCl, 20 мМ KH2PO4, pH 7,4, предварительно выдерживают при 25oC в шейкере в течение 5 минут. Ферментативную реакцию инициируют добавлением NADH до общей концентрации 2 мМ. Следующие четыре аликвоты NADH (для увеличения концентрации NADH на 1 мМ каждый раз) добавляют с интервалами 5 мин и через 30 минут реакцию останавливают добавлением 0,5 мл хлороформа. Слой хлороформа анализируют методом газовой хроматографии.
Органические экстракты сырой инкубационной смеси анализируют методом газовой хроматографии. Методом газовой хроматографии обнаруживают только один основной новый пик (см. фиг. 2), который имеет то же время удерживания, что и эталонный образец парагидроксидифенилметана (6). Другие ароматические продукты гидроксилирования, орто- и мета-изомеры, имеют другие времена удерживания. Идентичность продукта структуре (6) дополнительно подтверждена с помощью масс-спектрометрии, которая дает соответствующий массовый пик при 184.
С использованием приведенных выше экспериментальных методов исследовано большое число органических соединений в качестве субстратов как для фермента дикого типа Р-450can, так и для мутированной версии Y96A. Дополнительно используют мутанты, обозначенные Y96V; Y96L, Y96F; C334A; комбинированный мутант F87A, Y96G, F193A и комбинированные мутанты активного участка и поверхности Y96A, C334A; Y96V, C334A; Y96L, C334A; Y96F, C334A; F87A, Y96G, F193A, C334A.
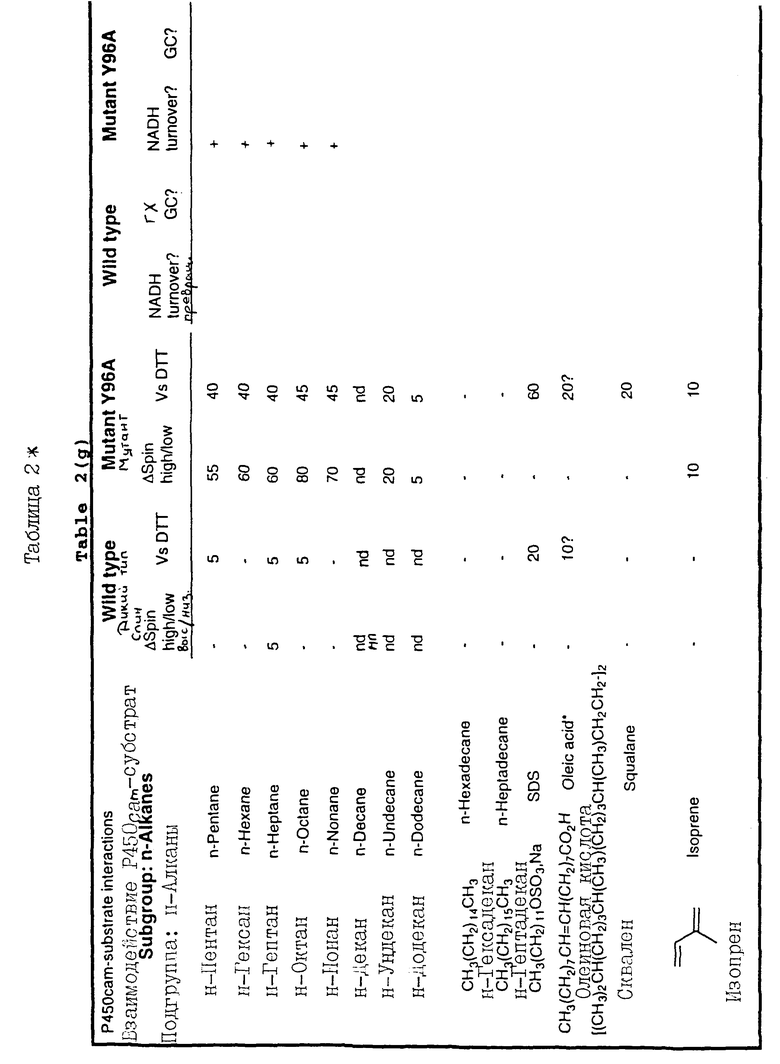
Результаты для мутанта Y96A приведены в табл. 2, в которой родственные по структуре молекулы объединены вместе. Те субстраты, окисление которых проводится путем превращения NADH, помечены знаком +.
Спин высокий/низкий: числа показывают процент Р-450 (OD417 0,2-0,4), переведенного из низко- в высокоспиновое равновесное состояние в присутствии 200 мкМ испытуемого соединения в фосфатном буфере (40 мМ фосфата, 68 мМ калия, pH 7,4). Спиновое равновесие оценивают с помощью УФ/видимого спектрофотометра: низкоспиновый при OD417 (оптическая плотность) и высокоспиновый при OD392; нп означает "не проводилось".
Относительно DTT (VsDTT): числа показывают процент замещения ВTT (200 мкМ), связанного с Р-450, конкурентным испытуемым соединением (200 мкМ) в фосфатном буфере. Связывание DTT с Р-450 приводит к появлению пиков поглощения OD347 и OD461, таким образом замещение измеряют с помощью УФ/видимого спектрофотометра.
Примеры приведены в таблицах от 2 (а) до 2(h) для каждого класса соединений, указанных в пунктах от I) до IV).
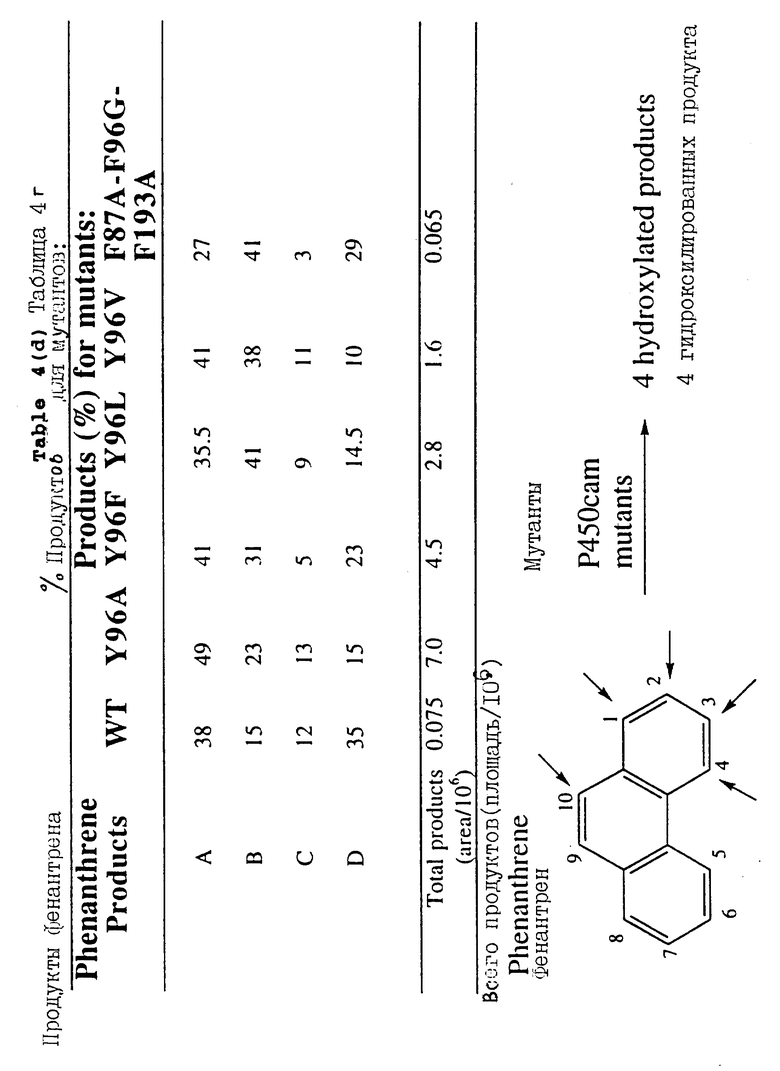
Продукты реакции некоторых субстратов очищают методом высокоэффективной жидкостной хроматографии и идентифицируют методом масс-спектроскопии, ядерного магнитного резонанса и/или со-элюирования. В таблице 3 представлено поглощение NADH при окислении небольших линейных, разветвленных и циклических углеводородов мутантом Y96A, C334A. В таблицах от 4 (а) до 4 (h) подробно представлены распределения продуктов для комбинаций субстрат/мутант, которые изучены на сегодняшний день.
Источники информации:
1. "Cytochrome P-450: Structure, Mechanism, and Biochemistry", ed. P. R. Ortiz de Montellano, Plenum Press, New York, 1986.
2. T. L. Poulos, В. С. Finzel and A. J. Howard, J. Mol. Biol., 1987, 195, 687-700.
3. J. A. Peterason, U.-Y. Lu, J. Geisselsoder, S. Graham-Lorence, С. Carmona, F. Witney and M. С. Lorence, J. Biol. Chem., 1992, 267, 14193-14203.
4. К. G. Ravichandran, S. S. Boddupali, С. A. Hasemann, J. A. Peterson, and J. Deisenhofer, Science, 1993, 261, 731-736.
5. В. P. Unger, I. С. Gunsalus and S. G. Sligar, J. Biol. Chem., 1986, 261, 1158-1163; J. S. Miles, A. W. Munro, В. N. Rospendowski, W. E. Smith, J. McKnight and A. J. Thomson, Biochem. J., 1992, 288, 503-509; T. H. Richardson, M. J. Hsu, T. Kronbach, H. J. Barnes, G. Chan, M. R. Waterman, В. Kemper and E. F. Johnson, Arch. Biochem. Biophys., 1993, 300, 510-516; S. S. Boddupalli, T. Oster, R. W. Estabrook and J. A. Peterson, J. Biol. Chem., 1992, 267, 10375-10380; H. Li, К. Darish and T. L. Poulos, J. Biol. Chem., 1991, 266-11909-11914.
6. I. С. Gunsalus and G. С. Wagner, Methods Enzymol., 1978, 52, 166-188.
7. P. J. Goodford, J. Med. Chem., 1985, 28, 849-857.
8. Quanta 4.0, Molecular Simulations inc., 16 New England Executive Park, Burlington, MA 01803-5297.
9. J. E. Baldwin, J. M. Blackburn, R. J. Heath and J. D. Sutherland, Bioorg, Med. Chem. Letts. 1992, 2, 663-668.
10. M. J. Zoller and M. Smith, Nucleic Acids Res., 1982, 10, 6487.
11. T. A. Kunkel, Proc. Natl. Acad. Sci., 1985, 82, 488-492.
12. С. Di Primo, G. Hui Bin Hoa, P. Douzou and S. Sligar, J. Biol. Chem. , 1990, 265, 5361-5363.
13. D Filipovic, Biochemical and Biophysical Research Communications, Vol. 189, No. 1, 1992, November 30, 1992, p. 488-495.
14. S. G. Sligar, D Filipovic and P. S. Stayton, Methods in Enzymology, Vol. 206, p. 31-49.
15. P. J. Loida and S. G. Sligar, Protein Engineering, Vol. 6, No. 2, p. 207-212, 1993.
16. S. F. Tuck et al., The Journal of Biological Cemistry, Vol. 268, No. 1, January 5, 1993, p. 269-275.
17. W. M. Atkins and S. G. Sligar, The Journal of Biological Chemistry, Vol. 263, No. 35, 15 December 1988, p. 18842-18849.
18. W. M. Atkins and S. G. Sligar, Biochemistry 1990, 29, 1271- 1275.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ ЦИТОХРОМ Р450-МОНООКСИГЕНАЗЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ОКИСЛЕНИЯ ОРГАНИЧЕСКИХ СОЕДИНЕНИЙ | 2000 |
|
RU2285044C2 |
| РЕКОМБИНАНТНАЯ ТЕРМОСТАБИЛЬНАЯ ЛЮЦИФЕРАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ИЗОЛИРОВАННАЯ НУКЛЕИНОВАЯ КИСЛОТА, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, НАБОР ДЛЯ ИСПОЛЬЗОВАНИЯ В БИОЛЮМИНЕСЦЕНТНОМ АНАЛИЗЕ, АНАЛИТИЧЕСКИЙ ТЕСТ ДЛЯ ОПРЕДЕЛЕНИЯ ПРИСУТСТВИЯ В ОБРАЗЦЕ СоА | 2000 |
|
RU2251571C2 |
| ПРЯМАЯ ЭЛЕКТРОХИМИЯ ФЕРМЕНТОВ | 1997 |
|
RU2157521C2 |
| МУТАНТЫ ИНДУЦИРУЕМОЙ АКТИВАЦИЕЙ ЦИТИДИНДЕАМИНАЗЫ (AID) И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2537264C2 |
| (ЦИТОХРОМ Р450)-МОНООКСИГЕНАЗЫ | 1998 |
|
RU2237714C2 |
| ТЕРМОСТАБИЛЬНЫЕ МУТАНТЫ ФЕРМЕНТОВ БИОСИНТЕЗА КРАХМАЛА | 2002 |
|
RU2303633C2 |
| СТАБИЛИЗИРОВАННЫЕ ЖИДКИЕ ФЕРМЕНТНЫЕ КОМПОЗИЦИИ | 2009 |
|
RU2510662C2 |
| СТАБИЛИЗИРОВАННЫЕ ЖИДКИЕ ФЕРМЕНТНЫЕ КОМПОЗИЦИИ | 2013 |
|
RU2668563C2 |
| МУТАНТНЫЕ ОКСИДАЗЫ D-АМИНОКИСЛОТ | 2007 |
|
RU2362806C2 |
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ (ВАРИАНТЫ) | 2012 |
|
RU2507262C1 |



















Модифицированный цитохром P-450cam, в котором тирозиновый остаток в положении 96 заменен на остаток любой аминокислоты. Дополнительно могут быть заменены аминокислотные остатки по меньшей мере в одном из положений 87, 98, 185, 244, 247, 295 и 298. Модификация позволяет использовать цитохром Р-450cam в качестве катализатора при окислении полициклических ароматических углеводородов, линейных или разветвленных алканов, соединений дифенила и бифенила, включающих галогенпроизводные, а также галогенированные углеводороды. 3 с. и 6 з.п. ф-лы, 3 ил., 18 табл.
| W.M.Atkins et al | |||
| Двигатель внутреннего горения | 1921 |
|
SU450A1 |
| Journal of Bid | |||
| Chem., 1988, 263, N 35, p.18842-18849 | |||
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| Способ получения цитохрома Р-450 | 1989 |
|
SU1693043A1 |
