КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области молекулярной фармакологии, и, в частности, к получению пептидов, пригодных для обработки эпителиальных опухолей, и в основном опухолей, ассоциированных с онкогенными типами HPV. Главная цель настоящего изобретения заключается в идентификации пептидов, структура которых позволяет блокировать фосфорилирующий домен казеинкиназы (CKII) в результате прямого взаимодействия с таким участком. В настоящем изобретении представлено одиннадцать циклических пептидов с разными аминокислотными последовательностями, которые ингибируют in vitro CKII-фосфорилирование, проявляют цитотоксичность в отношении трансформированных HPV-16 клеток (CaSki) и, кроме того, повышают чувствительность этих клеток к цитостатическому действию интерферона (ИФН). Кроме того, настоящее изобретение относится к применению указанных пептидов, конъюгированных или слитых с другими пептидами и химическими соединениями, которые проникают в клетки, а также к применению и пептидов, и химических молекул-миметиков.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области молекулярной фармакологии и, в частности, к получению пептидов, используемых для лечения ассоциированных с HPV эпителиальных опухолей, поскольку они позволяют блокировать фосфорилирующий домен казеинкиназы II (CKII) путем прямого их взаимодействия с таким участком.
CKII представляет собой треонин/сериновый фермент, участвующий в клеточной пролиферации, и в процессе малигнизации он локализован внутри клетки, главным образом, в ядре (Tawfic S., Yu S., Wang H., Faust R., Davis A., Ahmed K., 2001, Histol. Histopathol. 16:573-582).
На основании имеющихся данных о высоких уровнях CKII в разных эпителиальных солидных опухолях было высказано предположение, что фосфорилирование, вызываемое этим ферментом, является существенным признаком злокачественной трансформации и маркером опухолевой прогрессии (Seidin D.C., Leder P., 1995, Science 267:894-897) (Faust R.A., Gapany M., Tristani P., Davis A., Adams G.L., Ahmed K., 1996, Cancer Letters 101:31-35). С другой стороны, сверхэкспрессия CKII у трансгенных мышей приводит к опухолеобразованию в молочных железах из-за увеличения путей передачи Wnt/бета-катениновых сигналов в эпителиальных клетках этих молочных желез (Landesman-Bollag E., Romien-Mourez R., Song D.H., Sonenshein G.E., Cardiff R.D., Seldin D.C., 2001, Oncogene 20:3247-3257).
Среди эпителиальных опухолей те, которые возникают из-за HPV, составляют большую часть. Например, большинство опухолей мочеполовой системы ассоциируется с деятельностью этих онковирусов, и наличие ДНК-последовательностей HPV обнаруживается в 99,7% данных опухолей, возникающих из сквамозных клеток шейки матки (Walboormers J.M., Jacobs M.V., Manos M.M., Bosch F.X., Kummer J.A., Shah K.V., Snijders P.J., Peto J., Meijer C.J., Muňoz N., 1999, J. Pathol. 189:12-19). ВОЗ также сообщает о 500000 пациентах во всем мире, заболевающих ежегодно злокачественной опухолью шейки матки (Parkin D.M., Laara E., Muir C.S., 1980, Int. J. Cancer 41:184, 1972). На Кубе из-за этой болезни ежегодно умирает 370 женщин с диагнозом: Злокачественная опухоль шейки матки (Organization Panamericana de la Salud, 1999, Basic Country Health Profiles for the Americas, Куба, 206-219).
В зависимости от того, развивается ли малигнизация или нет, HPV делят на онкогенные и неонкогенные папилломавирусы (Lorincz A.T., Temple G.F., Kurman R.J., Jenson A.B., Lancaster W.D., 1987, J. Natl. Cancer Inst. 79:671-677). HPV-16 и HPV-18 ассоциированы с интраэпителиальной неоплазией, которая в большинстве случаев малигнизирует, но, кроме того, оба типа HPV ассоциированы более чем с 90% дисплазий и карциномами шейки матки (Fujinaga Y., Shimada M., Okasawa K., Fukushima M., Kato I., Fujinaga K., 1991 J. gen. Virol. 72:1039-1044).
Поскольку терапевтическая или профилактическая вакцина все еще недоступна для радикального лечения (удаления) HPV-ассоциированных злокачественных опухолей, использование ингибиторов вирусной транскрипции и вирусных онкогенных белков становится более привлекательным. Биомодуляторы, подобные IFN, используют с определенной эффективностью в отношении некоторых HPV-ассоциированных заболеваний, подобных кондиломе, подошвенным бородавкам и респираторным папилломатозам (Koromilas A.E., Li S., Matlashewski G., 2001, Cytokine & Growth Factor Reviews 12:157-170). В предыдущих экспериментах с HPV-трансформированными клетками (HeLa) нами показано, что длительное их выдерживание с альфа-ИФН возвращает их к исходному фенотипу и сопровождается ингибированием экспрессии мРНК HPV в этих клетках (López-Oceio O., Perea S.E., Reyes A., Vigoa L., López-Saura P., 1993. J. IFN Res 13:369-375). На этой же клеточной модели нами установлено, что альфа-ИФН модулирует мРНК HPV путем репрессии эндогенной вирусной транскрипции (Perea S.E., López-Oceio O., Garcia-Milián R., Araňa M.J., 1995, J. IFN & Cytokine Res. 15:495-501). В соответствии с данными результатами, полученными на клеточных линиях, в предварительном экспериментальном исследовании пациентов, страдающих злокачественной опухолью шейки матки, мы наблюдали, что обработка с помощью альфа-ИФН модулирует экспрессию мРНК (Garcia-Milián R., Rios M.A., Diaz D., Silveria M., Guilar O., Amigó M., Araňa M.J., Perea S.E., 1996, J. IFN and Cytokine Res. 16:709-713). Несмотря на многообещающие данные по использованию ИФН в качестве регулятора экспрессии мРНК HPV, установленные данные свидетельствуют о вариабельности ИФН-ответа и о феномене резистентности к данному цитокину, который в клинических испытаниях составляет у пациентов 40-50% (Arany I., Tyring S.K., Stanley M.A., Tomai M.A., Miller R.L., Smith M.H., McDermott D.J., Slade H.B., 1999, Antiviral Res. 43:55-63). Некоторые молекулярные и клинические факты свидетельствуют о том, что онкогенный белок Е7 играет ключевую роль в феномене ИФН-резистентности. К примеру, сообщалось, что Е7 связывается с индуцированным ИФН-фактором транскрипции (p48), вызывая тем самым ИФН-ответ в результате блокирования активации транскрипции (Barnard P. and McMillan N.A.J., 1999, Virology 259:305-313). Кроме того, сообщается также об изменении в присутствии Е7 регуляторного ИФН-фактора (IRF-1) (Park J.S., Kim E.J., Know H.J., Hwang E.S., Namkoong S.E., Um S.J., 2000, J. Biol. Chem. 275:6764-6769) (Perea S.E., Massimi P., Banks L., 2000, J. Mol. Med. 5:661-666). По результатам клинических испытаний считают, что ответ на ИФН зависит от экспрессии Е7 в HPV-содержащих очагах поражения (Frazer I.H., McMillan N.A.J., 1997, Papillomatosis and condyloma acuminate. Clinical Applications of the interferons (R. Stuart Harris and R.D. Penny, eds.) Pp 79-90. Chapman and Hall Medical, London). Онкогенный белок Е7 играет существенную роль в малигнизации, вызываемой данными онкогенными вирусами. Так, было показано, что индуцированная Е7 иммортализация первичных клеток приводит к мутациям и хромосомным аберрациям после начала процесса иммортализации (Mougin C., Humbey O., Gay C., Riethmuller D., 2000, J. Gynecol. Obstet. Biol Reprod. 29:13-20). С другой стороны, нами показано, что стабильная трансфекция с помощью гена Е7 приводит к созданию в чувствительных опухолевых клетках феномена ИФН-резистентности (Moro A., Calixto A., Suárez E., Araňa M.J., Perea S.E., 1998, Bioch. Biophys. Res. Comm. 245:752-756). Сообщается также, что онкогенный белок Е7 связывает и блокирует функцию опухолевых супрессорных генов, подобных генам ретинобластомы (Rb) и инсулинподобному фактору роста 3, связывающему белок (IGFBP-3), соответственно, через Cys 24 и С-концевой домен (Nevins J.R., 1992, Science 258:424-429) (Zwerscke W. and Jansen-Durr P., 2000, Advances in cancer Res. 78:1-29). Аналогично этому показано, что пары Ser31/Ser32 в белке Е7 являются субстратом для фермента CKII (Hashida T., Yasumoto S., 1990, Biochem. Biophys. Res. Comm., 172:958-964) и что этот домен существенен для трансформирующей способности данного онкогенного белка (Barbosa M.S., Edmonds C., Fisher C., Schiller J.T., Lowy D.R., Vousden K., 1990, EMBO J. 9:153-160) (Chien W.-M., Parker J.N., Schmidt-Grimminger D.-C., Broker T.R., Chow L.T., 2000, Cell Growth & Differentiation 11:425-435), а также для ингибирования сигнального каскада ИФН (Perea S.E., López-Ocejo O., Garcia Millan R., Banks L., Araňa M.J., 1996, Eur. Cytokine Net. 7:503).
Исходя из роли фосфорилирующего сайта CKII для резистентности HPV к ИФН и развития злокачественной опухоли, можно полагать, что разработка лекарственных средств, блокирующих такой домен, могла бы стать полезной в терапии злокачественных опухолей. Молекулы, ингибирующие CKII-фосфорилирующий участок в Е7 или в иных клеточных субстратах, до сих пор не описаны.
Относительно онкогенного белка Е7, описаны лишь пептиды, блокирующие Rb-связывающий сайт (Cys24) (Webster K.R., Koleman K.G., 1997, US 5625031) (Oliff A.I., Riemen M.W., EP 0412762 A2 910213) и другие С-концевые области (39-98) (Pidder J.-D., Zwerschkie W., 2000, EP 09699013).
В клинических или в доклинических испытаниях к настоящему времени описаны несколько вакцин-кандидатов, которые представляют интерес для создания CTL-ответа, специфичного в отношении Е7 HPV (Chen C., Wang C.C., Hung C., Pardoll D.M., Wu T., 2000, Vaccine 18:2015-2022) (Chen C.H., Ji H., Suh K.W., Chotl M.A., Pardoll D.M., Wu T.C., 1999, Gene Ther. 12:1972-1981). Однако попытки, сосредоточенные на получении CTL-ответа, встречают различные затруднения, связанные с биологией HPV. Например, онкогенные типы HPV подавляют антигены MHC класса I, которые существенны для CTL-ответа (Connor M.E., Stern P.L., 1990, Int. J. Cancer 46:1029-1034). Кроме того, экспрессия Е7 ассоциируется с локальной иммуносупрессией в окружении данной злокачественной опухоли, и это могло бы также влиять на соответствующее проявление CTL-ответа (Le Buanec H., D'Anna R., Lachgar A., Zagury J.F., Bemard J., Ittlele D., d'Alessio P., Hallez S., Giannouli C., Burny A., Bizzini B., Gallo R.C., Zagury D., 1999, Biomed. Pharmacother. 53:424-431) (Lee S.J., Cho Y.S., Shim J.H., Lee K.A., Ko K.K., Choe Y.K., Park S.N., Hoshino T., Kim S., Dinarello C.A., Yoon F.U., 2001, J. Immunol. 167:497-504). В соответствии с вышеуказанными соображениями, очевидно, что объединение CTL-вакцин и фармацевтических препаратов, направленных на Е7, могло бы иметь большие перспективы. Кроме того, данный подход к получению превентивных HPV-вакцин приносит большую пользу, а ценовой риск обусловлен различными биологическими и социальными аспектами, включая: 1) длительный латентный период после первичной инфекции HPV, 2) слабое понимание механизма HPV-инфекции, 3) отсутствие животной модели для подходящего размножения HPV, 4) видовую специфичность и 5) оценку социального влияния превентивной HPV-вакцины, которая может приниматься очень долго. Поэтому использование фармацевтических препаратов, специфически направленных на вирусные онкогенные белки, могло бы создать преимущества перед подходами, сосредоточенными на управлении иммунной системой.
ИЗЛОЖЕНИЕ СУЩЕСТВА ИЗОБРЕТЕНИЯ
Существо и новизна данного изобретения основаны на описании впервые циклических пептидов, позволяющих прямо ингибировать фосфорилирующий участок CKII, а также цитотоксически воздействовать in vivo на клетки карциномы шейки матки HPV-16. Кроме того, данные пептиды повышают чувствительность указанных клеток к цитостатическому действию ИФН.
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Настоящее изобретение относится главным образом к пептидам, способным связывать фосфорилирующий участок CKII, в котором обнаружены следующие последовательности:
(а) CSVRQGPVQKC (Id. Sec. No.1)
(b) CSSCQNSPALC (Id. Sec. No.2)
(c) CQIPQRTATRC (Id. Sec. No.3)
(d) CAKQRTDPGYC (Id. Sec. No.4)
(e) CWMSPRHLGTC (Id. Sec. No.5)
(f) CRNCTVIQFSC (Id. Sec. No.6)
(g) CHYIAGTVQGC (Id. Sec. No.7)
(h) CPLVSLRDHSC (Id. Sec. No.8)
(i) CKQSYLHHLLC (Id. Sec. No.9)
(j) CFQPLTPLCRC (Id. Sec. No.10)
(k) CQSYHELLLQS (Id. Sec. No.11)
Настоящее изобретение включает в себя также любой гомологичный вариант или миметик из вышеуказанных пептидов, которые получают путем синтеза или рекомбинантным способом, а также любой слитый пептид, содержащий пептиды, описанные в приведенном списке. Любой пептид, структура которого позволяет блокировать фосфорилирующий участок CKII для соответствующего субстрата, считается гомологичным вариантом. Кроме того, любую химическую молекулу (не пептидную), структура которой позволяет блокировать такой фосфорилирующий участок, считают вариантом миметика.
Другая цель настоящего изобретения заключается в создании фармацевтической композиции, которая включает в себя один или несколько пептидов, описанных в настоящем изобретении, а также подходящий носитель.
Кроме того, настоящее изобретение включает в себя применение вышеуказанных пептидов отдельно или в сочетании с любой иной подходящей молекулой, такой как цитокины и интерфероны для: 1) ингибирования пролиферации опухолевых клеток, 2) обработки HPV-ассоциированной и неассоциированной злокачественной опухоли и 3) обработки HPV-ассоциированных очагов поражения на предопухолевых стадиях.
Кроме того, пептиды согласно изобретению можно было бы использовать для лечения HPV-инфицированных пациентов, не чувствительных к лечению интерфероном.
В другом отношении настоящее изобретение включает в себя определенный экспрессирующий вектор для клеток млекопитающего, содержащий ДНК-последовательность, которая кодирует любой из вышеуказанных пептидов.
Пептиды согласно изобретению обладают циклической структурой и в основном характеризуются способностью связывать фосфорилирующий участок CKII и аннулировать такое биохимическое событие. Данные пептиды описаны в прилагаемом списке. С другой стороны, показаны также эффекты in vivo, оказываемые пептидами на HPV-трансформированные клетки.
Описанные пептиды характеризуют их способностью ингибировать фосфорилирование последовательности RRREEETEEE, ранее описанной в качестве оптимального консенсусного домена для CKII-фосфорилирования (Promega Cat:V5661), и участка фосфорилирования, заключенного в области 28-38 онкогенного белка Е7 HPV-16.
Чтобы охарактеризовать пептиды, описанные в настоящем изобретении, сконструировали единственную библиотеку 11-аминокислотого циклического пептида, которая экспрессируется в Р8-области нитевидных фагов. Скрининг данной библиотеки осуществляют с использованием синтетической 28-38-области Е7 в качестве мишени, которую также конъюгируют с биотином для ее фиксации на твердой поверхности. Селекцию фагов, связавшихся с 28-38-областью Е7, осуществляют путем иммунодетекции с использованием специфичного антитела к Р8-области в данном фаге. В конечном итоге ДНК, соответствующую отобранным одиннадцати фагам с высокой способностью связывания с 28-38-областью Е7, секвенируют, и соответствующие пептиды химически синтезируют твердофазным методом. Затем полученные синтетические пептиды выделяют, подвергая очистке с помощью ВЭЖХ, анализируют с помощью масс-спектрометрии, и, наконец, оценивают полученную in vitro- и in vivo-эффективность.
Согласно настоящему изобретению несмотря на различие аминокислотных последовательностей описанных здесь циклических пептидов они одинаково ингибируют CKII-фосфорилирование. Данный факт означает, что взаимодействие этих пептидов с фосфорилирующим участком CKII определяется преимущественно их структурой, а не самой последовательностью.
В настоящем изобретении также показано, что линейные пептиды проявляют более низкую способность к ингибированию фосфорилирующего участка CKII. Данный факт усиливает значение структуры для связывающей способности рассматриваемых пептидов с таким доменом. Кроме того, данный факт предполагает эффективность других миметических молекул, которые связываются с фосфорилирующим участком CKII.
Для того чтобы добиться осуществления внутриклеточного действия CKII на эндогенные субстраты, описанные пептиды можно химически конъюгировать или генетически срастить с проникающими в клетку пептидами, относящимся к белкам, подобным, в числе других, Tat 1 вируса иммунодефицита человека (ВИЧ-1) (Schwarze S.R., Dowdy S.F., 2000, Trends Pharmacol. 21:45-48), транскрипционному фактору, кодируемого геном Drosophila Antenapedia (Derossi D., и соавт., 1996, J. Biol. Chem. 271:18188-18193), белку VP22 вируса простого герпеса (HSV) (Lindgreen M., и соавт., 2000, Trends Pharmacol. Sci. 21:99-103), пенетратину (penetratin) и транспортану (transportan) (Gariepy J., Kawamura K., 2001, Trends Biotech. 19:21-28). Для проверки in vivo гипотезы настоящего изобретения данные циклические пептиды синтетически сливали с проникающим в клетку пептидом, приведенным для белка Tat 1 ВИЧ-1 (GRKKRRQRRRPPQC), и с пептидом для локализованного в ядре сигнала, принадлежащего большому антигену Т (KKKRKVE) SV40.
Представленные в настоящем изобретении данные ясно свидетельствуют о том, что циклические пептиды проявляют дозозависимую цитотоксичность в отношении трансформированных HPV-16 (CaSki) клеток карциномы шейки матки. Эти результаты предполагают использование данных пептидов в качестве терапевтического средства для обработки злокачественных опухолей одного и того же гистологического происхождения, а также предопухолевых стадий интраэпителиальной неоплазии шейки матки. Кроме того, экспериментальные данные in vivo свидетельствуют, что циклические пептиды оказываются более эффективными, чем их соответствующая линейная форма, подкрепляя тем самым положение о существенном значении структуры пептидов для их действия.
Кроме того, циклические пептиды, описанные в настоящем изобретении, эффективны в отношении клеток HeLa, содержащих HPV-18, а также в отношении клеток H-82, полученных из клеток немелкоклеточного рака легкого, негативных по HPV. Эти результаты коррелируют с результатами, полученными in vitro в настоящем изобретении и свидетельствующими о том, что пептиды блокируют не только участок CKII, фосфорилирующий Е7 HPV-16, но они блокируют его также и в других белках, содержащих такой участок. Тот факт, что описанные здесь пептиды эффективны в отношении HPV-негативных опухолевых клеток, является аргументом в пользу их возможного использования для других эпителиальных опухолей. Другие результаты настоящего изобретения свидетельствуют о том, что обработка клеток CaSki описанными здесь циклическими пептидами повышает чувствительность клеток к цитостатическому действию альфа-ИФН. С учетом предыдущих доказательств, свидетельствующих о том, что участок CKII, фосфорилирующий Е7 HPV-16, необходим для блокирования сигнального каскада ИФН (Perea S.E., López-Ocejo O., Garcia Millan R., Banks L., Araňa M.J., 1996, Eur. Cytokine Net. 7:503), описанные здесь пептиды можно использовать, чтобы компенсировать обычную ИФН-резистентность, наблюдаемую при HPV-инфекции.
Настоящее изобретение можно отнести также к ДНК, кодирующей каждый из описанных здесь пептидов. Данную ДНК можно вводить в экспрессирующий вектор млекопитающего и затем трансфицировать и в трансформированные HPV-16 клетки, и в нетрансформированные клетки. Данный вектор, содержащий олигонуклеотид, который кодирует каждый пептид, можно также использовать в качестве альтернативы генной терапии HPV-ассоциированной злокачественной опухоли.
В принципе описанные здесь пептиды можно использовать при HPV-ассоциированных заболеваниях вместе с другими агентами, а также с терапевтическими вакцинами, созданными на основе клеточного ответа против HPV.
Настоящее изобретение иллюстрируется нижеследующими примерами.
Пример 1: Влияние пептидов на фосфорилирующий участок CKII: Данный анализ основан на in vitro реакции фосфорилирования с использованием субстратной последовательности RRREEETEEE, которая представляет собой оптимизированный консенсусный домен CKII-фосфорилирования. Данную реакцию осуществляют в 50 мкл 25 мМ рН 7,5 Трис:HCl, 1 мкКи 32Р-γАТФ, 100 мкМ АТФ, 2 мг/мл субстратного пептида, 0,2 М NaCl, 10 мМ MgCl и 1 единицы фермента CKII (Promega). Реакцию инкубируют при 37°С в течение 10 минут. Затем на хроматографическую бумагу PE-81 (Whatmann) наносят 5 мкл реакционной смеси в виде пятна и осуществляют четыре промывки с помощью 10 мМ H3PO4. В заключение измеряют связанную с фильтрами радиоактивность и по уровням cpm определяют ферментативную активность CKII в каждом образце. Одновременно в каждый анализ в качестве внутреннего контроля включают специфический ингибитор CKII, подобный гепарину. Представленные на Фиг. 1 данные показывают, что циклические пептиды ингибируют CKII-фосфорилирование на 80%. Линейные пептиды также ингибируют CKII-фосфорилирование области 28-38 в Е7, хотя и в меньшей мере по сравнению с циклической формой. Эти данные указывают, что описанные здесь пептиды ингибируют фосфорилирующий участок CKII и дают основание полагать, что их структура играет существенную роль в их взаимодействии с последовательностями-мишенями.
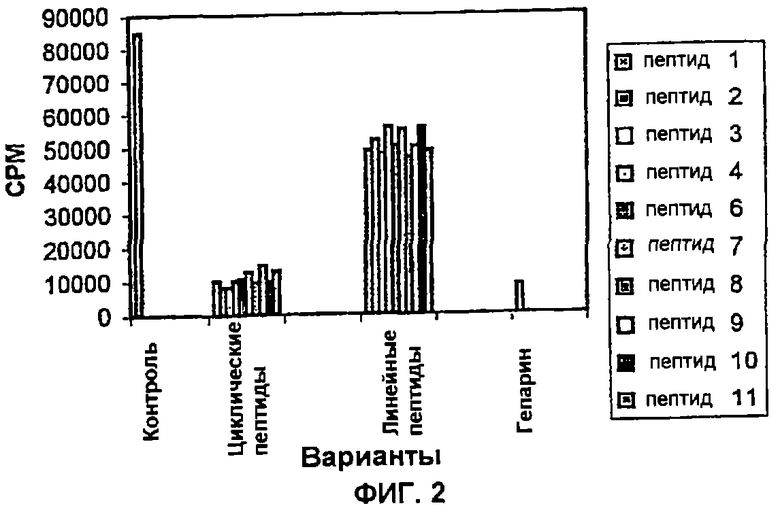
Пример 2: Влияние пептидов на фосфорилирование Е7 HPV-16: Данный анализ основан на in vitro реакции фосфорилирования онкогенного белка Е7 HPV-16, экспрессируемого в E.coli в виде слитого белка с глутатион-S-трансферазой (GST). Перед проведением ферментативной реакции слитый белок Е7-GST выделяют путем очистки с помощью хроматографии по сродству с использованием гранул глутатион-сефарозы (Pharmacia). Данную реакцию осуществляют в 50 мкл 25 мМ рН 7,5 Трис:HCl-буфера, 1 мкКи 32P-γАТФ, 100 мкМ АТФ, 40 мкл указанных гранул, содержащих Е7-GST, 0,2 М NaCl, 10 мМ MgCl и 1 единицы CKII (Promega). Указанную реакционную смесь инкубируют при 37°С в течение 40 мин. После чего гранулы промывают три раза с помощью 0,5 мл буфера и в конечном итоге уровень фосфорилирования Е7-GST анализируют с помощью 10%-го SDS-ПААГ-электрофореза. Визуализацию фосфорилированных белков осуществляют путем проявления рентгеновской пленки, экспонированной ранее с высушенными гелями. Количество фосфорилированного белка Е7 определяют денситометрированием. Данные на Фиг. 2 указывают, что описанные здесь пептиды одинаково эффективны в пределах ингибирования участка CKII, фосфорилирующего Е7 HPV-16.
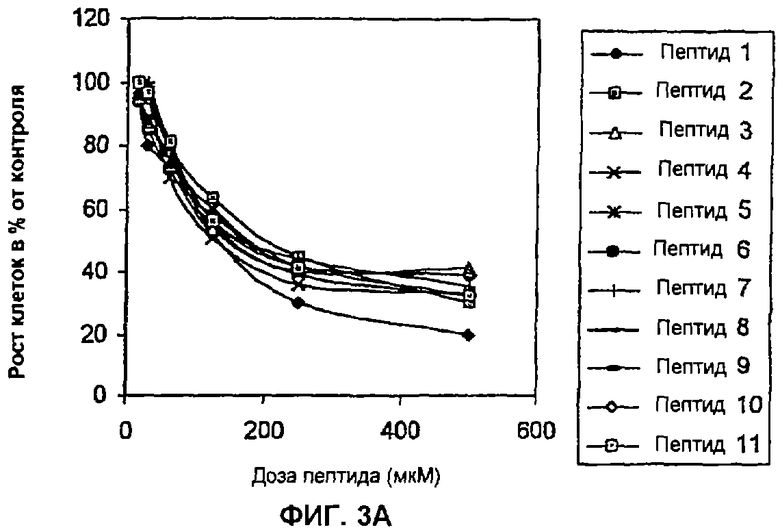
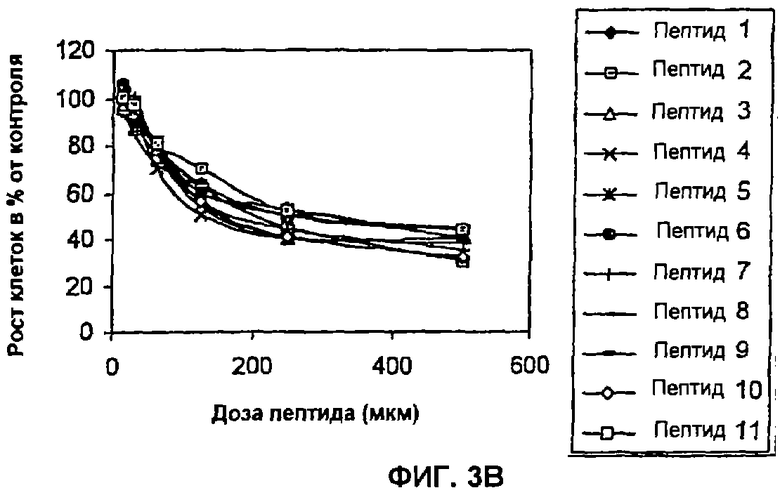
Пример 3: Влияние пептидов на пролиферацию клеток, трансформированных HPV-16 и HPV-18 (соответственно, CaSki и HeLa). В данном анализе клетки CaSki или HeLa в концентрации 2×104 клеток/мл высевают в 96-луночные планшеты (Costar) с использованием DMEM с добавлением 10% околоплодной сыворотки теленка (FCS) (Gibco). Через 24 часа в культуральную среду вносят пептиды в дозах, варьирующих между 15 и 500 мкМ. Инкубацию осуществляют в течение 96 часов в 5% СО2, после чего в каждую лунку приливают по 20 мкл раствора MTS (1,90 мг/мл) (Promega). Планшеты последовательно выдерживают в течение одного часа в одних и тех же инкубационных условиях и в заключение рассчитывают поглощение при 490 нм. Результаты выражают в виде процента роста относительно контроля без пептидов. С этой целью и циклические, и линейные пептиды сливают путем химического синтеза с проникающим в клетку пептидом Tat-1 ВИЧ-1, который способен проникать в цитоплазму и ядро (Schwarze S.R., Dowdy S.F., 2000, Trends Pharmacol. 21:45-48). Полученные в данном эксперименте данные показывают, что описанные здесь пептиды оказывают зависимый от дозы эффект и на клетки CaSki (HPV-16), и на клетки HeLa (HPV-18) (Фиг. 3А и 3В). Данный пример показывает, что пептиды согласно изобретению эффективны не только в отношении HPV-16, но также и в отношении HPV-18.
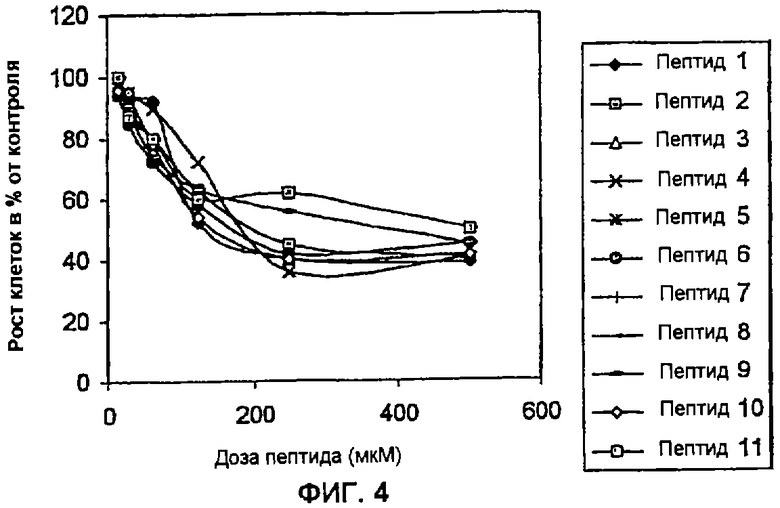
Пример 4: Влияние пептидов на пролиферацию HPV-негативных опухолевых клеток. В данном анализе клетки H-82 (мелкоклеточный рак легкого) в концентрации 2×104 клеток/мл высевают в 96-луночные планшеты (Costar) с использованием DMEM с добавлением 10% околоплодной сыворотки теленка (Gibco). Через 24 часа в данную культуральную среду вносят пептиды в дозах, варьирующих между 15 и 500 мкМ. Инкубацию осуществляют в течение 96 часов в 5% СО2, после чего в каждую лунку приливают 20 мкл раствора MTS (1,90 мг/мл) (Promega). Планшеты последовательно выдерживают в течение одного часа в одних и тех же инкубационных условиях и в заключение рассчитывают поглощение при 490 нм. Результаты выражают в виде процента роста относительно контроля без пептидов. В данном анализе описанные в настоящем изобретении циклические пептиды, слитые с проникающим в клетку пептидом Tat-1 ВИЧ-1, используют, как указано выше. Полученные в данном эксперименте результаты показывают, что пептиды согласно изобретению оказывают зависимый от дозы эффект на пролиферацию клеток H-82. На Фиг. 4 показано, что пептиды согласно изобретению эффективны не только в отношении HPV-трансформированных клеток, но также и в отношении опухолевых клеток другой локализации и гистологических типов, подобных мелкоклеточному раку легкого.
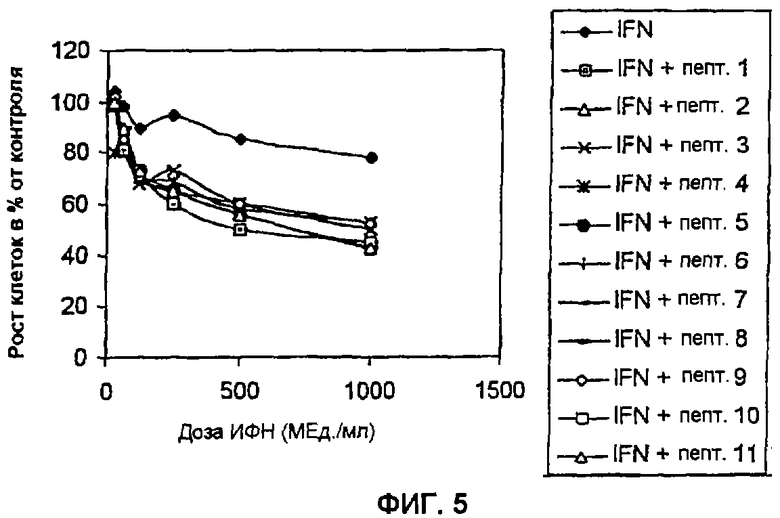
Пример 5: Влияние пептидов на HPV-16-ответ на обработку ИФН в клетках CaSki. В данном анализе клетки CaSki высевают в концентрации 2×104 клеток/мл в 96-луночные планшеты (Costar) с использованием DMEM с добавлением 10% FCS (Gibco). Через 24 часа в данную культуральную среду вносят 120 мкМ каждого пептида. Через 24 часа добавляют ИФН в количестве между 1000 и 31,5 Ед./мл. Инкубацию осуществляют в течение 96 часов в 5% СО2, после чего приливают 20 мкл MTS, 1,90 мг/мл. Кроме того, планшеты выдерживают в течение одного часа в одних и тех же условиях, и в заключение считывают поглощение при 490 нм. Данные представлены в виде процента роста относительно контроля. В данных экспериментах описанные в настоящем изобретении пептиды используют в их циклическом варианте, слитыми с проникающим в клетку пептидом, принадлежащим Tat-1-белку ВИЧ, как указано выше. Представленные на Фиг. 5 результаты показывают, что предварительная инкубация клеток CaSki с описанными в настоящем изобретении пептидами, делает эти клетки чувствительными к антипролиферативному воздействию альфа-ИФН. Эти данные дают основание полагать, что описанные в настоящем изобретении пептиды полезны для лечения HPV-инфицированных пациентов, устойчивых к ИФН-терапии.
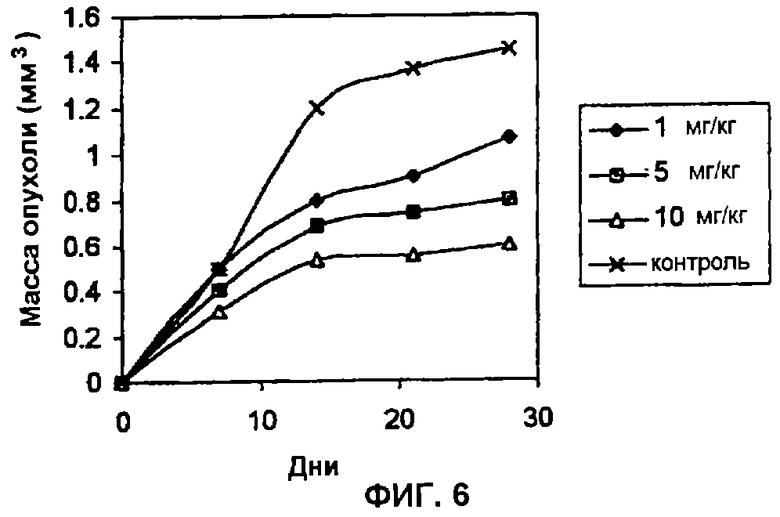
Пример 6: Противоопухолевое действие пептида, ингибирующего CKII-фосфорилирование в человеческих опухолях, имплантированных моделям голых мышей. В этих экспериментах используют 6-8-недельных самок голых мышей BalbC. Имплантацию злокачественной опухоли осуществляют с использованием клеток Н-125 (немелкоклеточный рак легкого) в концентрации 1000000 клеток/мл, которые ресуспендируют в солевом растворе (PBS). Клеточную суспензию инокулируют подкожно на брюхе. Введение пептида (последовательность 1 в приведенном списке) осуществляют вместе с указанными клетками и продолжают осуществлять через день вплоть до завершения одномесячной обработки. В данном анализе дозы варьируют между 1 и 10 мг/кг массы тела. Для исследования противоопухолевого действия оценивали такие параметры, как масса опухоли и выживаемость животных. Как показано на Фиг. 6, пептид во всех трех испытываемых дозах эффективно ингибировал опухолевую прогрессию. Эти данные демонстрируют противоопухолевую эффективность пептида, который ингибирует СКII-фосфорилирование на модели злокачественной опухоли человека, имплантированной подопытным животным.
Пример 7: Структурно-функциональный анализ гомологичных вариантов пептида CWMSPRHLGTC (SEQ. ID. No.5). Авторы изобретения исследовали биологическую активность различных вариантов гомологов и миметиков одного из описанных в данном изобретении пептидов. С этой целью было предпринято «аланиновое сканирование» этого пептида, относящегося к последовательности SEQ. ID. No.5 согласно изобретению.
Варианты, полученные при аланиновом сканировании:
CWMSPRHLGTC (SEQ. ID. No.5) (дикого типа, F20-16)
CAMSPRHLGTC (F2140)
CWASPRHLGTC (F2141)
CWMAPRHLGTC (F2142)
CWMSARHLGTC (F2143)
CWMSPAHLGTC (F2144)
CWMSPRALGTC (F2145)
CWMSPRHAGTC (F2146)
CWMSPRHLATC (F2147)
CWMSPRHLGAC (F2148)
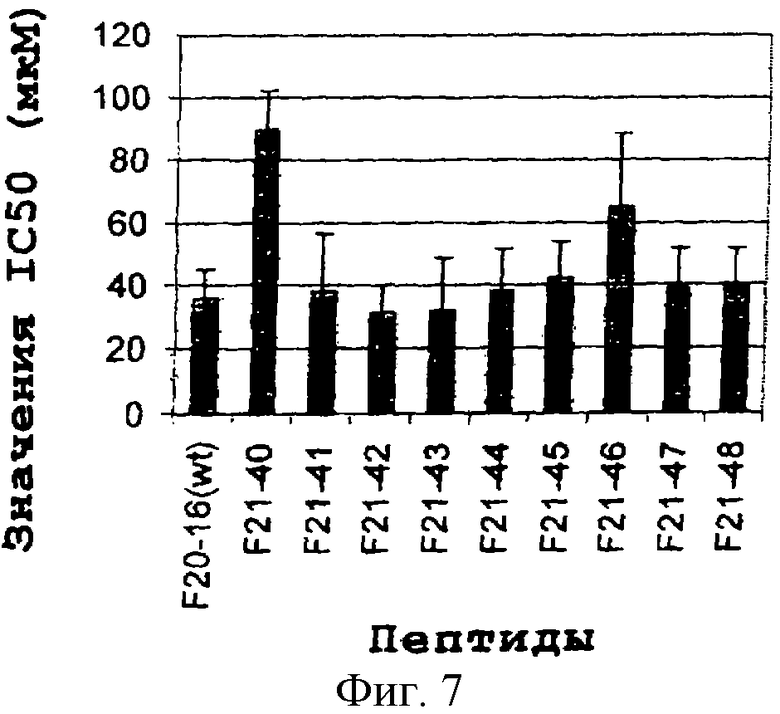
Каждый аланиновый мутант для проведения биологического анализа искусственно сливали с проникающим в клетки пептидом Tat-1. Для эксперимента in vitro клетки линии Н-125 немелкоклеточного рака легких (NSCLC) человека в концентрации 2×104 клеток/мл высевали в 96-луночных планшетах (Costar) с использованием среды DMEM, обогащенной 10% FCS (Gibco). Через 24 часа в каждую лунку добавляли пептиды, полученные в результате аланинового сканирования, в интервале доз от 0 до 200 мкМ. Инкубацию осуществляли в течение 72 часов при 37°С в 5% CO2, и в конце в каждую лунку добавляли по 20 мкл раствора MTS (1,90 мг/мл). Планшеты дополнительно выдерживали в течение одного часа в одних и тех же инкубационных условиях, затем определяли поглощение при 490 нм, используя считывающее устройство для микропланшетов Amersham-Pharmacia). Результаты (Фиг. 7) выражали в виде значений ингибирующей концентрации (IC50), полученных из соответствующих кривых доза-ответ. В настоящем изобретении IC50 определяли как концентрацию каждого химерного пептида, которая вызывает 50%-ное ингибирование клеточного роста. Как показано на Фиг. 7, все из вариантов, содержащих единичную замену аланина, были способны ингибировать клеточную пролиферацию в степени, сходной с таковой, наблюдаемой в случае пептида дикого типа. Интересно отметить, что варианты с заменой на W и L проявляли более высокие уровни значений IC50 в отношении этих клеток. То есть оба таких мутанта обладали меньшей эффективностью в отношении ингибирования роста клеток. Однако большая часть указанных пептидных вариантов, наподобие пептида дикого типа, сохраняла способность ингибировать клеточную пролиферацию.
Пример 8: Получение фармацевтической композиции, содержащей пептиды. Была получена стабильная лиофилизированная композиция, имеющая по меньшей мере 95% чистоты в отношении пептида. Эта композиция, по существу, состоит из различных химерных пептидов, которые содержат единственный из описанных в настоящем изобретении пептидов, слитый с проникающим в клетки пептидом Tat-1 (от 10 до 700 мг/мл). Композиция содержит также буферную систему для поддержания рН в области от 4,5 до 7,4 на основе фосфата, ацетата, цитрата или фосфат-цитрата при ионной силе от 0,1 до 100 мМ. Композиция включает в себя эффективное количество ЭДТА в качестве хелатобразующего средства в концентрации примерно от 0,1 до 10 мг/мл, а также некоторое количество сахара, сахарного спирта, неионного детергента или аминокислоты, или их смесей для стабилизации пептидов с целью сохранения степени их чистоты и биологической активности. Такими ингредиентами могут служить сахароза, лактоза, глюкоза, фруктоза, маннит, сорбит, полисорбат 20, полисорбат 80, цистеин, аланин, глицин и гистидин в концентрации приблизительно от 0,01 до 100 мг/мл. В конце в композицию добавляют эффективное количество наполнителя, выбранного из полиэтиленгликоля, маннита, декстрана и глицина в концентрации приблизительно от 1 до 100 мг/моль. В таких композициях пептиды, описанные в настоящем изобретении, обладают стабильностью в течение по меньшей мере 12 месяцев как при температуре 4°С, так и при температуре 28°С, что было показано методами ВЭЖХ и масс-спектрометрии.
Пример описанной выше композиции
Пептид (SEQ. ID. No.5), слитый с проникающим в клетки пептидом Tat-1: 350 мг/мл
ЭДТА Na2×2H2O: 0,5 мг/мл
Сорбит: 10 мг/мл
ПЭГ 6000: 5 мг/мл
Полисорбат 80: 0,02 мг/мл
Na2HPO4: 6,34 мг/мл
NaH2PO4×2H2O: 1,72 мг/мл
Преимущества настоящего изобретения
1. Созданы фармацевтические средства с широким спектром применения, которые не только используют при HPV-ассоциированных заболеваниях, но также и при солидных опухолях с высоким уровнем эндогенной активности CKII.
2. Тот факт, что 28-38-область консервативна у HPV, дает возможность использовать данное фармацевтическое средство при заболеваниях, ассоциированных с различными HPV-типами.
3. Пептиды в виде терапевтических молекул проявляют низкую антигенность при введении человеку.
4. Рассматриваемое терапевтическое средство является легким в изготовлении.
Краткое описание чертежей
Фиг. 1. Влияние пептидов на СКII-фосфорилирование.
Фиг. 2. Влияние пептидов на HPVE7-СКII-фосфорилирование.
Фиг. 3А. Влияние пептидов на пролиферацию CaSki-клеток.
Фиг. 3В. Влияние пептидов на пролиферацию HeLa-клеток.
Фиг. 4. Влияние пептидов на пролиферацию клеток опухоли легкого.
Фиг. 5. Влияние пептидов на ответную реакцию HPV-16-трансформированных клеток на действие ИФН.
Фиг. 6. Ингибирование опухолевой прогрессии под действием пептидов.
Фиг. 7. Структурно-функциональный анализ гомологичных вариантов пептида CWMSPRHLGTC (SEQ. ID. No.5). Клетки линии Н-125 рака легких человека в концентрации 2×104 клеток/мл высевали в 96-луночные планшеты. Каждый пептид, полученный в результате «аланинового сканирования» пептида дикого типа, искусственно сливали с проникающим в клетки пептидом Tat-1. Концентрация различных химерных пептидов варьировала от 0 до 200 мкМ. Антипролиферативный эффект в каждом случае выражали в виде значения IC50.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА, АССОЦИИРОВАННОГО С ИНФЕКЦИЕЙ HPV | 2013 |
|
RU2636003C2 |
| СЛИТЫЕ БЕЛКИ ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ ИММУНОГЕННЫХ УСИЛИВАЮЩИХ АГЕНТОВ ДЛЯ ИНДУЦИРОВАНИЯ АНТИГЕНСПЕЦИФИЧЕСКОГО Т-КЛЕТОЧНОГО ОТВЕТА | 2013 |
|
RU2631002C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА, АССОЦИИРОВАННОГО С ИНФЕКЦИЕЙ HPV | 2017 |
|
RU2664466C1 |
| СЛИТЫЕ БЕЛКИ ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ ИММУНОГЕННЫХ УСИЛИВАЮЩИХ АГЕНТОВ ДЛЯ ИНДУЦИРОВАНИЯ АНТИГЕНСПЕЦИФИЧЕСКОГО Т-КЛЕТОЧНОГО ОТВЕТА | 2013 |
|
RU2619187C2 |
| Иммуногенная композиция на основе рекомбинантных белков E6-CBD И E7-CBD для лечения злокачественных новообразований, ассоциированных с вирусом папилломы человека 16 типа | 2023 |
|
RU2805482C1 |
| ВЕКТОРЫ ДЛЯ МНОЖЕСТВЕННОЙ ГЕННОЙ ЭКСПРЕССИИ | 2008 |
|
RU2462513C2 |
| ПОЛИПЕПТИДЫ Е2 ПАПИЛЛОМАВИРУСА, ПРИМЕНЯЕМЫЕ ДЛЯ ВАКЦИНАЦИИ | 2008 |
|
RU2482189C2 |
| ВАКЦИНЫ ПРОТИВ HPV | 2012 |
|
RU2644201C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМБИНАЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ХЕМОСЕНСИБИЛИЗАЦИИ ОПУХОЛЕЙ, УСТОЙЧИВЫХ К ПРОТИВОРАКОВЫМ СРЕДСТВАМ | 2007 |
|
RU2396974C2 |
| КОМПОЗИЦИИ, ВЫЗЫВАЮЩИЕ СПЕЦИФИЧЕСКИЙ ОТВЕТ ЦИТОТОКСИЧЕСКИХ Т-ЛИМФОЦИТОВ, ВКЛЮЧАЮЩИЕ ЛИМФО-АБЛАТИВНОЕ СОЕДИНЕНИЕ И МОЛЕКУЛУ, СОДЕРЖАЩУЮ АНТИГЕННЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ И НАЦЕЛЕННУЮ НА СПЕЦИАЛИЗИРОВАННЫЕ АНТИГЕН-ПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ | 2007 |
|
RU2448729C2 |




Изобретение относится к области медицинской биохимии и может быть использовано в терапии некоторых видов злокачественных опухолей. Идентифицирована группа 11-членных пептидов, способных формировать циклическую структуру и ингибировать фосфорилирующую активность казеинкиназы II в результате прямого связывания с ее фосфорилирующим доменом. Новые пептиды проявляют цитотоксичность в отношении HPV-содержащих и некоторых HPV-негативных опухолевых клеток, в связи с чем предлагается использовать их, предпочтительно, в форме слитых полипептидов с повышенной способностью проникать в клетку для получения фармацевтических композиций и лечения ассоциированных с HPV и других эпителиальных опухолей. 6 с. и 6 з.п-лы, 7 ил.
a. CSVRQGPVQKC;
b. CSSCQNSPALC;
с.CQIPQRTATRC;
d. CAKQRTDPGYC;
е. CWMSPRHLGTC;
f. CRNCTVIQFSC;
g. CHYIAGTVQGC;
h. CPLVSLRDHSC;
i. CKQSYLHHLLC;
j. CFQPLTPLCRC;
k. CQSYHELLLQS;
или гомологичным вариантом любой из указанных последовательностей, сохраняющим свойства немодифицированной формы пептида.
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| ЕР 0969013 А, 05.01.2000 | |||
| WO 9501374 А, 12.01.1995. | |||

