Область изобретения
Данное изобретение относится к фармацевтической композиции, вызывающей специфический ответ цитотоксических Т-лимфоцитов (цТЛ) против по меньшей мере одного Т-клеточного эпитопа в организме млекопитающего, которая включает
- первое соединение, вызывающее лимфопению,
- в качестве второго соединения - молекулу, обладающую селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), где указанная молекула связана с указанным Т-клеточным эпитопом; и
- возможно, фармацевтически приемлемый носитель.
Изобретение также относится к фармацевтической композиции, включающей
- первое соединение, вызывающее лимфопению,
- в качестве второго соединения - молекулу, обладающую селективной аффинностью к АПК, где указанная молекула связана с по меньшей мере одним Т-клеточным эпитопом антигена, полученного из инфекционного агента; и
- возможно, фармацевтически приемлемый носитель в виде комбинированного препарата для одновременного, раздельного или последовательного применения в лечении и/или предупреждении инфекционных заболеваний пациентов.
Изобретение также относится к фармацевтическим композициям, включающим
- первое соединение, вызывающее лимфопению,
- в качестве второго соединения - молекулу, обладающую селективной аффинностью к АПК, где указанная молекула связана с по меньшей мере одним Т-клеточным эпитопом антигена, полученного из злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки; и
- возможно, фармацевтически приемлемый носитель в виде комбинированного препарата для одновременного, раздельного или последовательного применения в лечении и/или предупреждении соответственно злокачественных патологий, клеточной дисплазии, опухолевых процессов или рака у пациентов.
Предшествующий уровень техники
Общепризнано, что противораковые препараты обладают способностью ускорять приобретение противоопухолевого иммунитета у носителей опухоли. На различных моделях опухолей животных и у пациентов с прогрессирующей меланомой (2-4) и прогрессирующей почечной карциномой (7) было продемонстрировано, что противораковые препараты, такие как циклофосфамид (1-7), L-PAM4 (8), 1,3-бис(2-хлороэтил)-1-нитрозомочевина (9), винбластин (10), флударабин и блеомицин (11) усиливают развитие опосредованных Т-клетками противоопухолевых ответов.
Исследования механизмов, по которым противораковые препараты усиливают приобретение Т-зависимого иммунитета, обеспечивающего эрадикацию опухоли у носителей опухоли обнаружили, что химиотерапия вызывает сдвиг в цитокинном профиле: от противовоспалительных цитокинов (таких как фактор роста опухоли-β (TGF-β) и интерлейкин-10), обладающих ингибиторной активностью в отношении образования цТЛ, в сторону провоспалительных цитокинов (например, фактора некроза опухолей-α (ФНО-α), интерферона-γ (ИФН-γ) и гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ)), который способствует развитию клеточного противоопухолевого иммунитета.
В частности, флударабин является иммуносупрессирующим пуриновым аналогом, который применялся в основном в лечении лимфопролиферативных заболеваний низкой степени злокачественности (12). Повторяющиеся циклы терапии флударабином вызывают глубокое истощение Т-клеток, особенно CD4+ Т-клеток (13). В отличие от других иммуносупрессирующих цитотоксических лекарственных средств, таких как циклофосфамид, флударабин вызывает апоптоз лимфоцитов как в делящихся клетках, так и в клетках, находящихся в фазе G0-G1 клеточного цикла. Такая не зависящая от клеточного цикла активность может быть отнесена на счет подавления лекарственным препаратом сигналов от STAT1 (от англ. Signal Transducer and Activator of Transcription) (14).
До настоящего времени иммунотерапия рака и хронических инфекционных заболеваний имела ограниченный успех. В данной заявке изобретатели показывают, что комбинация лимфо-аблативных лекарственных препаратов с вакцинами на основе рекомбинантых белков показывают гораздо более многообещающие результаты.
Поскольку лимфо-аблативные лекарственные препараты убивают лимфоциты (вызывают апоптоз) или делают их неактивными (анергичными), можно предположить, что они будут снижать эффективность вакцин, доставляющих антигены к специализированным антиген-презентирующим клеткам.
В частности, аденилатциклаза (CyaA) Bordetella pertussis обладает способностью доставлять свой каталитических домен в цитозоль эукариотических клеток (15). Таким образом, эпитопы CD4+ и CD8+ Т-клеток, помещенные в каталитический сайт CyaA, подвергаются процессингу и презентируются молекулами главного комплекса гистосовместимости МНС (от англ. Major Histocompatibility Complex) II и I класса, соответственно, на поверхности антиген-презентирующих клеток (АПК; (16)).
Более того, было показано, что CyaA специфически связывается с интегрином αMβ2 (CD11b/CD18; (17)), и таким образом обеспечивает таргетинг, т.е. выполнение Т-клеточными эпитопами роли мишеней для субпопуляции CD11b+ дендритных клеток (18).
Иммунизация мышей рекомбинантной CyaA, несущей определенные Т-клеточные эпитопы, приводила к индукции выраженных Т-цитотоксических ответов, полной защиты на летальное введение вируса и вызывала эффективный профилактический и терапевтический противоопухолевый иммунитет (19-21).
Несмотря на большое количество многообещающих результатов на доклиническом уровне, внедрение в клиническую практику адоптивной клеточной иммунотерапии встречает множество трудностей. По данным Rosenberg (22), объективный уровень ответа в иммунотерапевтических исследованиях у больных раком остается ниже 3%, а наилучшие результаты получают при использовании технологий лечения на основе аутологичного адоптивного переноса клеток. Такой подход является трудоемким, дорогостоящим и трудным для безопасного введения во всеобщее употребление. Таким образом, представляет интерес уточнение на доклиническом уровне подходов к улучшению адоптивной клеточной иммунотерапии.
Сущность изобретения
Изобретение относится к способу, вызывающему у млекопитающих специфический ответ Т-цитотоксических лимфоцитов (цТЛ) против по меньшей мере одного Т-клеточного эпитопа, причем этот способ включает введение нуждающемуся млекопитающему
- соединения, вызывающего лимфопению и
- молекулы, обладающей селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), которая содержит чужеродный антиген, несущий по меньшей мере один указанный Т-кпеточный эпитоп.
В воплощении изобретение относится к способу, вызывающему терапевтический иммунный ответ у млекопитающих, страдающих хроническим инфекционным заболеванием или злокачественной клеточной патологией, дисплазией, опухолевым процессом или раком, и способ включает введение нуждающимся пациентам
- композиции, включающей соединение, вызывающее лимфопению и
- композиции, включающей молекулу с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), несущей антиген соответственно инфекционного агента или злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки.
В дополнительном воплощении изобретение относится к фармацевтической композиции, вызывающей у млекопитающих специфичный ответ цитотоксических Т-лимфоцитов (цТЛ) против по меньшей мере одного Т-клеточного эпитопа, или вызывающей терапевтический иммунный ответ у млекопитающего, страдающего хроническим инфекционным заболеванием или злокачественной клеточной патологией, дисплазией, опухолевым процессом или раком, где указанная композиция включает
- соединение, вызывающее лимфопению
- молекулу с селективной аффинностью к специфическим антиген-презентирующим клеткам (АПК), которая связана с по меньшей мере одним указанным Т-клеточным эпитопом или которая связана с антигеном соответственно инфекционного агента или злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки и
- возможно, фармацевтически приемлемый носитель.
В дополнительном воплощении изобретение относится к набору реагентов, вызывающих у млекопитающих специфический ответ цитотоксических Т-лимфоцитов (цТЛ) против по меньшей мере одного Т-клеточного эпитопа или вызывающих терапевтический иммунный ответ у млекопитающих, страдающих хроническим инфекционным заболеванием или злокачественной клеточной патологией, дисплазией, опухолевым процессом или раком, где указанный набор реагентов включает
- композицию, включающую соединение, вызывающее лимфопению и
- композицию, включающую молекулу, обладающую селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающую чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, несущую антиген соответственно инфекционного агента или злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки.
В дополнительном воплощении изобретение относится к способу предупреждения или лечения инфекций млекопитающих, где агент, ответственный за указанную инфекцию, экспрессирует специфический антиген, содержащий по меньшей мере один Т-клеточный эпитоп, и указанный способ включает введение нуждающимся пациентам композиции, включающей
- соединение, вызывающее лимфопению и
- молекулу, обладающую селективной аффинностью к специфическим антиген-презентирующим клеткам (АПК), включающую чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, включающую указанный антиген, специфичный для указанного агента, ответственного за указанную инфекцию, или по меньшей мере один Т-клеточный эпитоп, содержащийся в указанном антигене.
В дополнительном воплощении изобретение относится к способу предупреждения или лечения рака у млекопитающих, особенно рака дыхательных и пищеварительных путей, при котором опухолевые клетки экспрессируют белок человеческого раково-эмбринонального антигена (СЕА), и указанный способ включает введение нуждающемуся пациенту
- композиции, содержащей поликлональные анти-лимфоцитарные или антитимоцитарные антитела кролика или лошади (АТГ), вызывающие лимфопению и, в дополнение,
- композиции, включающей рекомбинантную аденилатциклазу, несущую участок A3-B3 белка человеческого СЕА.
В дополнительном воплощении изобретение относится к способу предупреждения или лечения рака, вызванного HPV у млекопитающих, при котором опухолевые клетки экспрессируют онкоген Е7 HPV, и указанный метод включает введение нуждающимся пациентам
- композиции, содержащей поликлональные АТГ кролика или лошади, вызывающие лимфопению, а затем
- композиции, включающей рекомбинантную аденилатциклазу, несущую онкоген Е7 HPV.
Краткое описание графических материалов
Фигура 1 (пример 1): SDS-электрофорез в ПААГ рекомбинантных белков СЕА.
Три микрограмма очищенных белков разделяли в полиакриламидном геле (ПААГ) с градиентом от 4% до 15% и окрашивали Кумасси синим. Дорожка 1, CyaA дикого типа; дорожка 2, CyaA - CEAA3B3; дорожка 3, CyaAΔ-CEAA3B3; дорожка 4, CEAA3B3.
Фигура 2 (пример 1): Асимметричный ответ Th1, вызванный двукратной иммунизацией рекомбинантными белками СЕА.
После первичной иммунизации путем внутривенного введения 50 мкг рекомбинантных белков СЕА через 7 и 14 дней проводили реиммунизацию путем интрадермального введения 10 мкг аналогичного материала. Клетки, продуцирующие СЕА-специфичный интерферон-γ, идентифицировали с помощью проточной цитометрии (FACS) и внутриклеточного окрашивания на цитокины спустя 7 дней после последней иммунизации. Результаты представлены как медиана процентного соотношения CD4+ и CD8+ клеток, экспрессирующих интерферон-γ (n=3) после рестимуляции. Фоновые результаты, полученные на нестимулированных спленоцитах, вычитали.
Фигура 3 (пример 1): Истощение лимфоцитов периферической крови, вызванное иммуносупрессивным лечением.
Кинетический анализ числа лимфоцитов в периферической крови после интраперитонеального введения 200 мг/кг/день флударабина в течение 6 последовательных дней или после однократного интраперитонеального введения 20 мг/кг АТГ (верхняя левая панель). Три другие панели отображают кинетический анализ процентного содержания CD4+, CD8+ и В220+ лимфоцитов в периферической крови после вышеописанного иммуносупрессивного лечения.
Фигура 4 (пример 1): Увеличение выраженности СЕА-специфичного ответа Th1 как результат иммуносупрессии и двукратной иммунизации.
За 10 дней до двукратной иммунизации рекомбинантными белками СЕА животным проводилось иммуносупрессивное лечение на основе флударабина или АТГ. Клетки, продуцирующие СЕА-специфичныий интерферон-γ, идентифицировали с помощью проточной цитометрии (FACS) и внутриклеточного окрашивания на цитокины через 7 дней после последней иммунизации. Результаты представлены как медиана процентного соотношения CD4+ и CD8+ клеток, экспрессирующих интерферон-γ (n=3) после рестимуляции. Фоновые результаты, полученные на нестимулированных спленоцитах, вычитали.
Фигура 5 (пример 1): Повышение медианы выживаемости после двукратной иммунизации CyaA-CEAA3B3 в результате или до лимфоаблации.
За 5 дней до подкожной инъекции 2×104 клеток B16F0-CEA-GFP животным проводилось иммуносупрессивное лечение на основе флударабина или АТГ. (А) Через 3 дня после инъекции опухолевых клеток животным проводили двукратную иммунизацию рекомбинантными белками СЕА. Затем начинали регулярный мониторинг роста опухоли. Мышей забивали согласно требованиям санитарных норм. Приведены кривые выживаемости. Животные из контрольной группы (Контроль) не подвергались иммуносупрессии. (В) Так же, как в А, но иммунотерапию проводили через 5 дней после инъекции опухолевых клеток.
Фигура 6 (пример 2): SDS-электрофорез в ПААГ рекомбинантных белков СЕА.
Три микрограмма очищенных белков разделяли в полиакриламидном геле (ПААГ) с градиентом от 4% до 15% и окрашивали Кумасси синим. Дорожка 1, CyaA-HPV16E7Δ; дорожка 2, CyaA-CEAA3B3; дорожка 3, CyaAΔ-CEAA3B3; дорожка 4, СЕА His-tag.
Фигура 7 (пример 2): Ответ Th1, вызванный иммунизацией рекомбинантными белками СЕА.
После первичной иммунизации путем внутривенного введения эквимолярного количества указанных рекомбинантных белков (50 мкг для CyaA, 19,2 мкг для СЕА His-tag) в присутствии 10 мкг CpG, через 7 и 14 дней проводили реиммунизацию путем интрадермального введения 10 мкг аналогичного материала (3,8 мкг для СЕА His-tag) и 2 мкг CpG. Через 7 дней после последней иммунизации помощью анализа ELISpot ex vivo с использованием 1 мкг/мл пептидного пула (pepscan) СЕА идентифицировали CD8+ клетки, продуцирующие СЕА-специфичный интерферон-γ. Результаты представлены как медиана числа колоний, вырабатывающих интерферон-γ, SFC (n=3).
Фигура 8 (пример 2): Истощение лимфоцитов периферической крови, вызванное иммуносупрессивным лечением.
Кинетика числа лимфоцитов (CD4+, CD8+, CD49d+ и В220+) в периферической крови после интраперитонеального введения 100 мг/кг/день флударабина в течение 6 дней или после однократного интраперитонеального введения 20 мг/кг АТГ. Контрольным животным вводили PBS или 20 мг/кг нормальной кроличьей сыворотки (НКС).
Фигура 9 (пример 2): Увеличение выраженности СЕА-специфичного ответа Th1 как результат иммуносупрессии и двукратной иммунизации.
За 7 дней до двукратной иммунизации рекомбинантными белками СЕА животным проводилось иммуносупрессивное лечение на основе АТГ или флударабина. После первичной иммунизации путем внутривенного введения эквимолярного количества рекомбинантных белков (50 мкг для CyaA, 19,2 мкг для СЕА His-tag) в присутствии 10 мкг CpG через 7 и 14 дней проводили реиммунизацию путем интрадермального введения 10 мкг аналогичного материала (3,8 мкг для СЕА His-tag) и 2 мкг CpG. Через 7 дней после последней иммунизации помощью анализа ELISpot ex vivo с использованием 1 мкг/мл пептидного пула СЕА *** идентифицировали CD8+ клетки, продуцирующие СЕА-специфичный интерферон-γ. Результаты представлены как медиана числа колоний, вырабатывающих интерферон-γ, SFC (n=2).
Фигура 10 (пример 2): Повышение медианы выживаемости после двукратной иммунизации CyaA-CEAA3B3 в результате или до лимфоаблации.
Инокуляцию опухоли проводили путем введения 5×105 клеток Мса32А. Через 3 дня проводили иммуносупрессивное лечение на основе флударабина или АТГ. Контрольные животные получали нормальную кроличью сыворотку. Через 10 дней после инъекции опухолевых клеток (через 7 дней после иммуносупрессии) животным проводили двукратную иммунизацию рекомбинантными белками. После первичной иммунизации путем внутривенного введения эквимолярного количества указанных рекомбинантных белков (50 мкг для CyaA, 19,2 мкг для СЕА His-tag) в присутствии 10 мкг CpG через 7 дней проводили реиммунизацию путем интрадермального введения 10 мкг аналогичного материала (3,8 мкг для СЕА His-tag) и 2 мкг CpG. Затем начинали регулярный мониторинг роста опухоли. Мышей забивали согласно требованиям санитарных норм. Приведены кривые выживаемости.
Подробное описание изобретения.
Авторы изобретения обнаружили, что потенциирующий эффект лимфо-аблативных агентов, таких как антитимоцитарный иммуноглобулин (АТГ) или флударабин, способствующий приобретению терапевтического иммунитета пациентом, существенно усиливается, если антиген (антигены), входящий в состав препарата, при введении in vivo направляются к специализированным антиген-презентирующим клеткам (АПК).
Векторные вакцины, направленно воздействующие на специализированные антиген-представляющие клетки (АПК), такие как макрофаги, нейтрофилы и дендритные клетки, обладают преимуществом селективно доставлять антигены к этим АПК благодаря специфичной аффинности связывания АПК с факторами на клеточной поверхности, что приводит к усилению антиген-специфичного Т-клеточного ответа (хелперного и/или цитотоксического). Однако в комбинации с лимфо-аблативными соединениями такой эффект, как правило, недостижим, поскольку целевая популяция лимфоцитов подвергается воздействию указанных лимфо-аблативных соединений, сокращается или становится анергичной (нефункциональной).
Авторы изобретения продемонстрировали, что в случае, когда антиген в условиях in vivo направляется к АПК с помощью белкового вектора, аденилатциклазы (CyaA), имеющей высокую аффинность к молекуле CD11b (специфичной для таких АПК, как дендритные клетки, макрофаги, а также нейтрофилы), неожиданно происходит гораздо более сильная индукция цТЛ по сравнению с тем, если бы АТГ-опосредованное лимфо-аблативное лечение не проводилось. Такой эффект оказался неожиданным.
Авторы изобретения показали, что при АТГ-опосредованном лимфо-аблативном лечении такая более сильная индукция цТЛ ассоциирована с их способностью лучше предотвращать рост опухоли in vivo.
Авторы изобретения также показали, что флударабин-опосредованное лимфо-аблативное лечение также подавляет рост опухоли. Такая особенность облегчает способность иммунотерапии с применением CyaA более эффективно предотвращать рост опухоли in vivo.
Кроме того, авторы изобретения показали, что такое усиление связано не только с использованием иммуно-аблативных соединений, таких как АТГ или флударабин, но и со способностью векторов на основе CyaA нацеливаться на CD11b, и может быть экстраполировано на другие АПК-нацеленные системы, такие как белки теплового шока. Данный вывод основан на следующих 3 причинах:
1. Вектор CyaA с делецией участка, связывающего CD11b на АПК, не направлялся к АПК, и являлся не только менее активным, чем исходный вектор CyaA, но и не усиливал в значительной степени индукцию иммунного ответа против встроенного чужеродного антигена (CEAA3B3), когда экспериментальные животные предварительно получали иммунодепрессанты: УСИЛЕНИЕ (ИММУННОГО ОТВЕТА) БЛАГОДАРЯ CD11b-НАПРАВЛЯЮЩИМ СВОЙСТВАМ вакцины.
2. Антиген СЕА в отсутствие молекулы, имеющей селективную аффинность к АПК, в сочетании с CpG или с Poly-ICLC или с Монофосфорилом А, вызывает выраженный иммунный ответ, направленный против СЕА, в зависимости от способа определения иммуногенности. Напротив, когда подопытные животные предварительно получали иммунодепрессанты, индукция иммунного ответа против чужеродного антигена (СЕА) значительно не усиливалась: ОТСУТСТВИЕ УСИЛЕНИЯ (ИММУННОГО ОТВЕТА) АДЪЮВАНТОМ, НЕ НАЦЕЛЕННЫМ НА СПЕЦИФИЧЕСКИЕ КЛЕТКИ.
3. Результаты предварительных исследований позволили предположить, что сопоставимого потенциирования иммунного ответа можно добиться при использовании белка теплового шока Hsp65 (от англ. Heat Shock Protein), гибридизованного с CEAA3B3. Белковый вектор Hsp65, несущий A3B3, направляется к АПК за счет распознавания Toll-подобных рецепторов (TLR4, Toll-Like Receptors) на поверхности: УСИЛЕНИЕ (ИММУННОГО ОТВЕТА) БЛАГОДАРЯ АПК-НАПРАВЛЯЮЩИМ СВОЙСТВАМ вакцины.
Таким образом, в одном аспекте данное изобретение относится к фармацевтической композиции, вызывающей специфичный ответ цитотоксических Т-лимфоцитов (цТЛ) против по меньшей мере одного Т-клеточного эпитопа в организме млекопитающего, включающей
- первое соединение, вызывающее лимфопению,
- в качестве второго соединения - молекулу, обладающую селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), где указанная молекула связана с указанным Т-клеточным эпитопом; и
- возможно, фармацевтически приемлемый носитель.
Предпочтительно, первое соединение способно вызывать транзиторное истощение Т-клеток. Более предпочтительно, первое соединение выбрано из группы, состоящей из поликлональных антилимфоцитарных и антитимоцитарных иммуноглобулинов (АТГ), таких как антилимфоцитарные и антитимоцитарные иммуноглобулины (АТГ) кролика или лошади, пуриновые, пиримидиновые аналоги, алкилирующие агенты, моноклональные и поликлональные антитела, способные вызывать периферическое и/или центральное истощение Т, В и NK лимфоцитов. Это может быть флударабин или циклофосфамид. Антитела, способные вызывать истощение лимфоцитов, выбраны из группы, состоящей из анти-CD8, анти-CD4, анти-CD25, анти-CD3 и анти-CD52 моноклональных антител.
В частном воплощении молекула, обладающая селективной аффинностью к АПК, выбрана из группы, состоящей из аденилатциклаз, белков теплового шока (HSP), шигатоксина и LAG-3 (от англ. Lymphocyte-activating gene - 3, ген активации лимфоцитов - 3). Предпочтительно, аденилатциклаза получена из Bordetella pertussis, HSP выбран из группы, состоящей из hsp65 и hsp70, указанные hsp65 и hsp70 получены предпочтительно из Mycobacterium bovis, а указанный шигатоксин получен из Shigella dysenteriae.
В предпочтительном воплощении фармацевтическая композиция согласно данному изобретению также содержит адъювант. Адъюванты хорошо известны специалистам в данной области и могут применяться в данном изобретении индифферентно. Предпочтительно им может быть лиганд Toll-подобного рецептора (TLR) класса 3 (например, poly-ICLC), лиганд TLR-9 (например, CpG) или лиганд TLR-4 (например, монофосфорил-А), и более предпочтительно, адъювант CpG с последовательностью SEQ ID №16, где основаниями является фосфоротиоат. Указанный адъювант предпочтительно находится в растворе со вторым соединением.
В дальнейшем предпочтительном воплощении молекула, обладающая селективной аффинностью к АПК, ковалентно связана с по меньшей мере одним указанным Т-клеточным эпитопом. Предпочтительно, молекула, обладающая селективной аффинностью к АПК, и указанный по меньшей мере один Т-кпеточный эпитоп являются полипептидами, которые закодированы в последовательностях ДНК, полученных с помощью технологий рекомбинантных ДНК.
В фармацевтической композиции согласно изобретению указанный Т-клеточный эпитоп принадлежит антигену, происходящему из инфекционного агента или злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки. Предпочтительно, инфекционный агент выбран из группы, состоящей из вируса папилломы человека (HPV), вируса иммунодефицита человека (HIV), вируса гепатита В (HVB), вируса гепатита С (HCV), Chlamydia trachomatis или Mycobacterium tuberculosis. Также предпочтительно, антиген, происходящий из злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки, выбран из группы, состоящей из раково-эмбрионального антигена (СЕА), антигена меланомы MAGE-A3, теломеразы (TERT), онкогена Е7 вируса папилломы человека (HPV) и проапоптотического белка Р53.
Изобретение также относится к фармацевтическим композициям, как описано выше, для применения в качестве профилактической или терапевтической вакцины.
В предпочтительном воплощении млекопитающим является человек.
Фармацевтическая композиция согласно изобретению предпочтительно предназначена для внутримышечного, внутривенного, внутридермального, кожного, подкожного, интраперитонеального, орального введения или введения в слизистую.
Изобретение также относится к фармацевтической композиции, включающей
- первое соединение, вызывающее лимфопению,
- в качестве второго соединения - молекулу, обладающую селективной аффинностью к АПК, где указанная молекула связана с по меньшей мере одним Т-клеточным эпитопом антигена, полученного из инфекционного агента; и
- возможно, фармацевтически приемлемый носитель, в виде комбинированного препарата для одновременного, раздельного или последовательного применения в лечении и/или предупреждении инфекционных заболеваний пациентов.
В предпочтительном воплощении комбинированного препарата первое соединение вводится перед введением второго соединения. Предпочтительно, второе соединение находится в растворе с адъювантом. Предпочтительно, адъювант представляет собой лиганд Toll-подобного рецептора (TLR), выбранный из группы, состоящей из лигандов TLR 3 класса, таких как poly-ICLC, лигандов TLR 9 класса, таких как CpG и лигандов TLR 4 класса, таких как монофосфорил-А.
Изобретение также относится к фармацевтическим композициям, включающим
- первое соединение, вызывающее лимфопению,
- в качестве второго соединения - молекулу, обладающую селективной аффинностью к АПК, где указанная молекула связана с по меньшей мере одним Т-клеточным эпитопом антигена, полученного из злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки; и
- возможно, фармацевтически приемлемый носитель в виде комбинированного препарата для одновременного, раздельного или последовательного применения в лечении и/или предупреждении соответственно злокачественных патологий, клеточной дисплазии, опухолевых процессов или рака у пациентов.
В предпочтительном воплощении комбинированого препарата первое соединение вводится перед введением второго соединения. Предпочтительно, второе соединение находится в растворе с адъювантом. Предпочтительно, адъювант представляет собой лиганд Toll-подобного рецептора (TLR), выбранный из группы, состоящей из лигандов TLR 3 класса, таких как poly-ICLC, лигандов TLR 9 класса, таких как CpG, и лигандов TLR 4 класса, таких как монофосфорил-А.
Предпочтительно, фармацевтическая композиция согласно изобретению применяется для лечения и/или предупреждения рака, при котором раковые клетки экспрессируют белок СЕА, предпочтительно рака дыхательных и пищеварительных путей, где первое соединение представляет собой поликлональный АТГ кролика или лошади, а второе соединение представляет собой аденилатциклазу, несущую А3-B3 участок человеческого белка СЕА.
В другом предпочтительном воплощении фармацевтическая композиция согласно изобретению применяется для лечения и/или предупреждения рака, при котором раковые клетки экспрессируют белок СЕА, такой как белок человеческого СЕА, предпочтительно, рака дыхательных и пищеварительных путей, где первое соединение представляет собой флударабин, а второе соединение представляет собой аденилатциклазу, несущую участок А3-В3 белка человеческого СЕА.
В другом предпочтительном воплощении фармацевтическая композиция согласно изобретению применяется для лечения и/или предупреждения рака, вызванного HPV, при котором раковые клетки экспрессируют онкоген Е7 HPV, где первое соединение представляет собой поликлональный АТГ кролика или лошади, а второе соединение представляет собой рекомбинантную аденилатциклазу, несущую онкоген Е7 HPV.
В альтернативном варианте фармацевтическая композиция согласно изобретению применяется для лечения и/или предупреждения рака, вызванного HPV, при котором раковые клетки экспрессируют онкоген Е7 HPV, где первое соединение представляет собой флударабин, а второе соединение представляет собой рекомбинантную аденилатциклазу, несущую онкоген Е7 HPV.
Изобретение также относится к способу индукции у млекопитающего специфичного ответа цитотоксических Т-лимфоцитов против по меньшей мере одного Т-клеточного эпитопа, способ включает введение млекопитающему
- соединения, вызывающего лимфопению и
- молекулы, обладающей селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), которая содержит чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп.
Согласно данному изобретению указанный Т-клеточный эпитоп может являться Т-клеточным эпитопом антигена, полученного из инфекционного агента или злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки.
Согласно данному изобретению, соединение, вызывающее лимфопению, и молекула, обладающая селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающая чужеродный антиген, несущий по меньшей мере один указанный Т-кпеточный эпитоп, могут быть введены пациенту одновременно, раздельно или последовательно, предпочтительно при последовательном введении пациенту указанного соединения и указанной молекулы, указанное соединение, вызывающее лимфопению, может быть введено перед введением указанной молекулы с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающей чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп.
Согласно данному изобретению, при одновременном введении пациенту соединения, вызывающего лимфопению, и молекулы с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающей чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, указанные соединение и молекула могут быть введены в одной и той же или в различных композициях.
В другом воплощении данное изобретение относится к способу выработки терапевтического иммунного ответа в организме млекопитающего, страдающего хроническим инфекционным заболеванием или злокачественной клеточной патологией, дисплазией, опухолевым процессом или раком, и способ включает введение нуждающимся пациентам
- композиции, включающей соединение, вызывающее лимфопению и
- композиции, включающей молекулу с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающей чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, несущей антиген соответственно инфекционного агента или злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки.
Согласно данному изобретению, композиция, включающая соединение, вызывающее лимфопению, и композиция, включающая молекулу с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающую чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, могут быть введены пациенту одновременно, раздельно или последовательно, предпочтительно при последовательном введении пациенту указанного соединения и указанной молекулы, указанное соединение, вызывающее лимфопению, вводится перед введением указанной молекулы с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающей чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп.
Согласно данному изобретению, при одновременном введении пациенту соединения вызывающего лимфопению, и композиции, содержащей молекулу с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающую чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, указанные композиции могут быть введены в одной и той же или в различных композициях.
Согласно данному воплощению указанным млекопитающим может быть человек.
В следующем воплощении данное изобретение относится к фармацевтической композиции, вызывающей у млекопитающих специфичный ответ цитотоксических Т-димфоцитов (цТЛ), направленный против по меньшей мере одного Т-клеточного эпитопа, или вызывающей терапевтический иммунный ответ у млекопитающего, страдающего хроническим инфекционным заболеванием или злокачественной клеточной патологией, дисплазией, опухолевым процессом или раком, где указанная композиция включает
- соединение, вызывающее лимфопению и
- молекулу с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), связанную с указанным Т-клеточным эпитопом или связанную с антигеном соответственно инфекционного агента или злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки и
- возможно, фармацевтически приемлемый носитель.
В следующем воплощении данное изобретение относится к набору реагентов для выработки у млекопитающих специфичного ответа цитотоксических Т-лимфоцитов (цТЛ) против по меньшей мере одного Т-клеточного эпитопа или вызывающих терапевтический иммунный ответ у млекопитающих, страдающих хроническим инфекционным заболеванием или злокачественной клеточной патологией, дисплазией, опухолевым процессом или раком, где указанный набор реагентов включает
- композицию, включающую соединение, вызывающее лимфопению и
- композицию, включающую молекулу с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающую чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, несущую антиген соответственно инфекционного агента или злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки.
Согласно данному изобретению, фармацевтическая композиция или набор могут быть вакциной.
Согласно данному изобретению, композиция может быть разработана для внутримышечного, внутривенного, внутридермального, кожного, подкожного, интраперитонеального, орального введения или введения в слизистую.
Согласно данному изобретению, молекула, обладающая селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающая чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, состоит из молекулы с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), которая может быть ковалентно связана с указанным антигеном, несущим по меньшей мере один Т-клеточный эпитоп.
Согласно данному изобретению, молекула, обладающая селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), и указанный антиген, несущий по меньшей мере один Т-клеточный эпитоп, могут быть полипептидами, которые кодируются последовательностями ДНК, полученными с помощью технологии рекомбинантной ДНК.
Согласно данному изобретению, соединение, вызывающее лимфопению, может быть соединением, способным вызывать транзиторное истощение Т-клеток.
Согласно данному изобретению, соединение, вызывающее лимфопению, может быть выбрано из группы, состоящей из АТГ и флударабина.
Согласно данному изобретению, молекула, обладающая селективной аффинностью к специализированным антиген-презентирующим клеткам, может быть выбрана из группы, состоящей из аденилатциклазы (CyaA), белков теплового шока (HSP), шигатоксина и LAG-3.
В воплощении изобретение касается способа применения вектора аденилатциклазы (CyaA), выделенной из Bordetella pertussis, где вектор CyaA при применении в комбинации с лимфо-аблативными соединениями имеет улучшенные характеристики в качестве векторной вакцины по сравнению с вакцинами, не нацеленными на специализированные антиген-презентирующие клетки.
В воплощении изобретение касается фармацевтической композиции вещества, состоящего из ингредиента, включающего АТГ, и ингредиента, включающего CyaA, несущую чужеродные антигенные последовательности. Указанные антигенные последовательности могут быть получены из любого инфекционного агента, вызывающего хронический инфекционный процесс, или из любой аномально делящейся клетки млекопитающего, диспластически измененной клетки, опухолевой или раковой клетки.
В воплощении изобретение касается фармацевтической композиции вещества, состоящего из ингредиента, включающего флударабин, и ингредиента, включающего CyaA, несущую чужеродные антигенные последовательности. Указанные антигенные последовательности могут быть получены из любого инфекционного агента, вызывающего хронический инфекционный процесс, или из любой аномально делящейся клетки млекопитающего, диспластически измененной клетки, опухолевой или раковой клетки.
Согласно данному изобретению, указанная аденилатциклаза может быть получена из Bordetella pertussis или указанный HSP может быть hsp65 и hsp70, полученный из Mycobacterium bovis, более предпочтительно hsp65, или указанный шигатоксин может быть получен из Shigella dysentehae.
Согласно данному изобретению, указанный Т-клеточный эпитоп выбран из группы, состоящей из:
- Т-клеточного эпитопа опухолевого антигена, выбранного из группы, состоящей из раково-эмбрионального антигена (СЕА), особенно несущего участок А3-В3, MAGE-A3, TERT, онкогена Е7 вируса папилломы человека (HPV) и Р53; и
- Т-клеточного эпитопа инфекционного агента, выбранного из группы, состоящей из HPV, HVB, HCV, Chlamydia trachomatis и Mycobacterium tuberculosis.
В другом воплощении данное изобретение относится к способу предупреждения или лечения опухолей, в частности злокачественных опухолей у млекопитающих, где опухолевые клетки экспрессируют специфический ассоциированный с опухолью антиген, указанный ассоциированный с опухолью антиген, презентирующий Т-клеточный эпитоп, указанный способ включает введение нуждающемуся пациенту композиции, включающей
- соединение, вызывающее лимфопению; и
- молекулу с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающую чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, включая указанный ассоциированный с опухолью антиген, специфичный для указанной опухоли, или Т-клеточный эпитоп, имеющийся в указанном ассоциированном с опухолью антигене.
Данное изобретение также относится к способу предупреждения или лечения опухолей, в особенности, злокачественных опухолей у млекопитающих, при которой опухолевые клетки экспрессируют специфический ассоциированный с опухолью антиген, указанный ассоциированный с опухолью антиген, презентирует Т-клеточный эпитоп, а указанный способ включает введение нуждающемуся пациенту композиции, включающей
- соединение, вызывающее лимфопению; и
- молекулу с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающую чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, включая указанный ассоциированный с опухолью антиген, специфичный для указанной опухоли, или Т-клеточный эпитоп, имеющийся в указанном ассоциированном с опухолью антигене.
В другом воплощении данное изобретение относится к способу предупреждения или лечения инфекционных заболеваний млекопитающих, при которых агент, ответственный за указанное инфекционное заболевание, экспрессирует специфический антиген, представляющий Т-клеточный эпитоп, и указанный способ включает введение нуждающемуся пациенту композиции, содержащей
- соединение, вызывающее лимфопению; и
- молекулу с селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), включающую чужеродный антиген, несущий по меньшей мере один указанный Т-клеточный эпитоп, включая указанный антиген, специфичный для указанного агента, ответственного за указанное инфекционное заболевание, или, по меньшей мере один Т-клеточный эпитоп, содержащийся в указанном антигене.
В другом воплощении данное изобретение относится к способу предупреждения или лечения рака у млекопитающих, в особенности рака дыхательных и пищеварительных путей, при котором опухолевые клетки экспрессируют СЕА, и указанный способ включает введение нуждающемуся пациенту
- композиции, содержащей соединение АТГ, вызывающее лимфопению; и, затем
- композиции, содержащей рекомбинантную аденилатциклазу, несущую участок А3-В3 белка СЕА.
В другом воплощении данное изобретение относится к способу предупреждения или лечения рака, вызванного HPV у млекопитающих, при котором опухолевые клетки экспрессируют онкоген Е7 HPV, и указанный способ включает введение нуждающемуся пациенту
- композиции, содержащей поликлональный АТГ кролика или лошади, вызывающей лимфопению; и, затем
- композиции, содержащей рекомбинантную аденилатциклазу, несущую онкоген Е7 HPV.
Данное изобретение можно понять лучше при прочтении описаний экспериментальных исследований, проведенных в контексте работы, выполненной заявителем, которые не следует расценивать как лимитирующие.
Промышленным применением данного изобретения может быть
Улучшенная адаптивная иммунотерапия рака, направленная на злокачественные или неоплазированные клетки, специфически экспрессирующие опухолевые антигены, предпочтительно полипептидные последовательности СЕА, Е7, MAGE-A3, TERT и Р53, которые независимо друг от друга находятся в составе нацеленной на АПК векторной вакцины, предпочтительно в составе рекомбинантного белкового вектора, такого как вектор аденилатциклазы (CyaA), векторы белков теплового шока (Hsp65), шигатоксина или рекомбинантных белковых векторов, содержащих последовательности лиганда АПК, например LAG-3.
Улучшенная адаптивная иммунотерапия латентных или хронических инфекционных заболеваний, направленно воздействующая на инфицированные клетки пациента, которые специфически экспрессируют антигены инфекционного агента, предпочтительно, вирусов, и особенно HPV, HIV, HVB и HCV или внутриклеточных бактерий, таких как Chlamydia trachomatis, Mycobacterium tuberculosis, помещенных в нацеленную на АПК векторную вакцину, предпочтительно, вектор CyaA или белка теплового шока (hsp), шигатоксина или рекомбинантный белковый вектор, содержащий последовательности лиганда АПК, например, LAG-3. Улучшенные вакцины содержат нацеленный на АПК вектор, такой как CyaA, Hsp65, Hsp70, шигатоксин, LAG-3, антигенные последовательности выбора, полученные из CyaA, Е7, MAGE-A3, TERT, P53, HPV, HIV, HVB, HCV, Chlamydia trachomatis, Mycobacterium tuberculosis, связанные или помещенные в вектор, и молекулу, вызывающую иммуно-деплецию, такую как флударабин, АТГ, где компонент, вызывающий иммуно-деплецию, предпочтительно вводится пациенту перед введением других компонентов или совместно, но будучи смешанным ex temporanis.
Примеры
Приведенные примеры 1 и 2 схожи между собой, за исключением того, что в Примере 2 введение молекулы, обладающей селективной аффинностью к АПК, производится в присутствии адъюванта CpG.
Пример 1.
Данный пример касается применения лимфоаблативного режима до проведения иммунотерапии по технологии прайм-буст (проводится двукратная иммунизация, вначале - первичная иммунизация, или прайминг, а затем реиммунизация, или буст) с применением аденилатциклазы B. pertussis в модели на грызунах. Иммуносупрессия, опосредованная низкими дозами антитимоцитарных глобулинов кролика или флударабином, была транзиторной и приводила к индукции большего количества антиген-специфичных Т-клеток после иммунотерапевтического лечения. Этот эффект коррелировал с усилением способности мышей контролировать рост / отторгать агрессивные опухолевые клетки, экспрессирующие определенный антиген. Данный подход представляет возможность улучшить потенциал использования данного вектора в терапии рака или инфекционных заболеваний.
СЕА представляет собой опухоле-ассоциированный антиген, который усиленно экспрессируется при различных злокачественных патологиях, особенно пищеварительного тракта и легких. При помещении суб-фрагментов СЕА в каталитический домен CyaA удалось создать конструкцию, продемонстрировавшую иммуногенный эффект у мышей C57/BL6. Лимфодеплеция, опосредованная АТГ и флударабином, приводила к значительному повышению числа CyaA-специфичных Т-лимфоцитов после двукратной иммунизации CyaA-CEAA3B3. Данный феномен оказался специфичным в том, что обнаруживался только при векторизации СЕА с CyaA, но не наблюдался для CD8+ лимфоцитов при вакцинации белком CEAA3B3 в сочетании с адъювантом CpG. В экспериментах, определяющих эффект адъювантов на вакцины на основе вектора CyaA, исследователи продемонстрировали, что частота эпитоп-специфичных Т-клеточных ответов может быть значительно увеличена при добавлении в вакцину на основе CyaA, несущей указанный эпитоп, любого из следующих адъювантов: липид А (монофосфорил-А), poly-ICLC или CpG, которые являются лигандами соответствующих Toll-подобных рецепторов класса 3, 4 или 9 (результаты не приведены).
Препараты АТГ, описанные в данном примере, являются поликлональными иммуноглобулинами G кролика, полученными при иммунизации лимфоцитами или тимоцитами. Они применяются на человеке в трансплантации с 1960-х годов (23) по следующим показаниям: профилактика (индукционная терапия) и борьба с острым отторжением аллотрансплантатов органов, включая отторжение, устойчивое к стероидам, борьба с реакцией трансплантат-против-хозяина при трансплантации костного мозга (24), лечение апластической анемии (25) и подготовка реципиентов костного мозга от неродственных HLA-совместимых (26) или гаплоидентичных родственных доноров (27).
1.1 Материалы и методы.
1.1.1 Мыши, клеточные линии
Самки мышей линии C57BL/6 возрастом от 6 до 10 недель, не имеющие специфичных патогенов, были получены в компании CER Janvier (Le Gesnet St-lsle, France). Эксперименты на животных проводились в соответствии с правилами института по обращению с животными. Инъекции при иммуносупрессивной терапии проводили интраперитонеально, инъекции опухолевых клеток - подкожно, а иммунизацию - внутривенно (ретро-орбитально) и интрадермально в дермис уха. Кровь забирали из хвостовой вены. Сплекэктомию и выделение лимфоузлов проводили у умерщвленных животных (CO2). Подмышечные, брыжеечные и паховые лимфоузлы выделяли и собирали для фенотипического анализа.
Клетки линии B16F0, экспрессирующие белок СЕА, получили путем ко-трансдукции плазмидой pIRES-EPI, кодирующей СЕА и GFP (от англ. green fluorescent protein - зеленый флуоресцентный белок), и плазмидой rMLV (Vectalys). Клетки B16F0-CEA-GFP поддерживали в RPMI 1640, содержащей Glutamax, с добавлением 10% инактивированной нагреванием сыворотки эмбрионов телят, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина и 5×10-5 моль/л 2-меркаптоэтанола (Life Technologies).
1.1.2 Подсчет и фенотипирование клеток
Для подсчета и фенотипирования с помощью проточной цитометрии клетки крови, лимфоузлов и спленоцитов обрабатывали согласно методикам, описанным ранее (28, 29). Лимфоциты крови подсчитывали с использованием раствора Тюрка.
1.1.3 Получение в организме кролика глобулинов против тимоцитов мыши Антитимоцитарные глобулины вырабатывали путем инъекции подкожно кролику 5×108 тимоцитов, полученных у 6-недельных мышей линии C57BL/6. Через 14 дней проводили повторную иммунизацию путем внутривенного введения аналогичного материала. Через 7 дней у животного забирали кровь и выделяли сывороточный иммуноглобулин (Ig) G путем хроматографии на колонках с иммобилизованным протеином G (Pierce). Степень чистоты контролировали путем SDS-электрофореза в полиакриламидном геле, а количество белка оценивали по методу Брэдфорда с известной концентрацией IgG кролика в качестве стандарта. Кроличий АТГ использовали в концентрации 20 мг/кг для однократной инъекции. В качестве контроля использовали нормальную сыворотку кролика.
1.1.4 Реагенты, пептиды и олигонуклеотиды
Флударабин (Sigma) использовали в концентрации 200 мг/кг/день в течение 6 последовательных дней. Моноклональные антитела: анти-CD4-АРС, анти-CD8-PerCP, анти-В220-FITC и анти-ИФН-γ-ПТС были из BD Biosciences.
Пептидный скан - пептиды длиной 15 аминокислот, перекрывающиеся по 11 аминокислот и охватывающие домен A3 и B3 белка СЕА - был разработан и получен в Mimotope™ (Australia). Пептиды разводили в DMSO и использовали в конечной концентрации 1 мкг/мл. CpG ODN 1826 был приобретен в Proligo (Paris, France) и использован в конечной дозе 10 мкг на инъекцию. Указанный CpG имеет последовательность SEQ ID №16, где основаниями является фосфоротиоат (устойчивый к нуклеазам).
1.1.5 Молекулярное клонирование рекомбинантной аденилатциклазы В. pertussis, несущей домены A3 и В3 СЕА. Получение и очистка рекомбинантной CyaA-CEAA3B3
Рекомбинантную аденилатциклазу, использованную в данном исследовании, экспрессировали в Eschehchia coil, используя производные плазмиды pkTRAC-HPV16E7Δ30-42, кодирующей CyaA, не обладающую ферментативной активностью (21).
CyaA-CEAA3B3 была сконструирована в 2 этапа. Первый фрагмент ДНК, кодирующий аминокислотные остатки СЕА с 492 по 557, амплифицировали в ПЦР с использованием синтетического гена СЕА (Sequentia) и праймеров СЕА1 (SEQ ID №1: 5'-accatcaccgtctctgcg-3') и СЕА2 (SEQ ID №2: 5'-gggcactagtggtcagggtacggttgcc-3'). Второй фрагмент ДНК, кодирующий аминокислотные остатки СЕА с 629 по 687, амплифицировали в ПЦР с использованием синтетического гена и праймеров СЕА3 (SEQ ID №3: 5'-gggcaccggtaatggtatcccgcagcaacac-3') и СЕА4 (SEQ ID №4: 5'-cgcagagacggtgatggtgttaacggcacccgcagacagacc-3'). Эти два фрагмента ДНК (которые частично перекрываются) были очищены и скомбинированы с помощью праймеров СЕА2 и СЕА3 в третьей ПЦР, в которой был получен фрагмент ДНК длиной 395 пн. Этот фрагмент был обработан рестриктазами NheI и KpnI и помещен между соответствующими сайтами pkTRAC-HPV16E7Δ30-42, что позволило получить плазмиду pkTRAC-СЕАВ3. Затем фрагмент ДНК, кодирующий аминокислотные остатки СЕА с 545 по 647, амплифицировали в ПЦР с использованием синтетического гена СЕА и праймеров СЕА5 (SEQ ID №5: 5'-gggcgctagccgtctgcagctgtccaatg-3') и СЕА6 (SEQ ID №6: 5'-cccgggtacccggcgtgattttggcgata-3'). Очищенный фрагмент ПЦР был обработан AgeI и SpeI и лигирован в плазмиду pkTRAC-СЕАВ3, обработанную теми же рестриктазами.
CyaAΔ-РЭАА3В3 была сконструирована в 3 этапа. Вначале фрагмент ДНК, соответствующий аминокислотной последовательности CyaA с 1149 по 1230, была амплифицирована в ПЦР с использованием pTRACE5 (30) в качестве ДНК и праймеров CyaAΔ1 (SEQ ID №9: 5'-gggcgagctctggggccacgat-3') и CyaAΔ2 (SEQ ID №10: 5'-actagtgcctcgatcccgaagccg-3'). Второй фрагмент ДНК, кодирующий аминокислотные остатки СЕА с 1300 по 1356 был амплифицирован в ПЦР с использованием той же плазмидной ДНК и праймеров CyaAΔ3 (SEQ ID №11: 5'-actagtcatgctgtatggcgacgc-3') и CyaAΔ4 (SEQ ID №12: 5'-cccggcatgcgcgccggtctgg-3'). Эти частично перекрывающиеся фрагменты ДНК были очищены и скомбинированы с помощью праймеров CyaAΔ1 и CyaAΔ4 в третьей ПЦР для амплификации фрагмента ДНК длиной 427 пн. Этот фрагмент был обработан SacI и SphI и лигирован в плазмиду pkTRAC-СЕАА3В3, обработанную теми же рестрикционными ферментами. Полученная плазмида pKTRACΔ-СЕАА3В3, кодировала CyaA, лишенную способности связывать молекулу CD11b (31).
Все конструкции проверили с помощью ДНК-секвенирования (Genome Express). Рекомбинантные CyaA продуцировали в штамме E.coli BLR (Novagen) как описано ранее (21). Рекомбинантные белки очищали из телец включений до гомогенности путем 2-ступенчатой процедуры, включающей хроматографию на ДЭАЭ-Сефарозе и фенил-Сефарозе. Дополнительный этап отмывки с помощью 60% изопропанола был проведен (21) для удаления большинства полисахаридных примесей. Очищенные рекомбинантные белки анализировали при помощи SDS-электрофореза в ПААГ с окрашиванием по Кумасси. Концентрацию белка определяли спектрофотометрически по абсорбции при 280 нм и коэффициенту молярной экстинкции 142000 моль/л при длине оптического пути 1 см.
1.1.6 Молекулярное клонирование доменов A3 и В3 рекомбинантного СЕА. Продукция и очистка рекомбинантного белка CEAA3B3.
Оптимизированная для E.coli кДНК, кодирующая белок CEAA3B3, была субклонирована в вектор pTriEx-1.1 hygro (Novagen) между рестрикционными сайтами NotI и XhoI после амплификации в ПЦР с использованием праймеров СЕА7 (SEQ ID №7: 5'-gcggccgcaccatcaccgtctctgcg-3') и СЕА8 (SEQ ID №8: 5'-cccgctcgagggcacccgcagacagacc-3'). Затем полученной плазмидой трансформировали штамм E.coli BL21ADE3 (Novagen). Белок CEAA3B3 с гексагистидиновым хвостом (His-Tag-CEAA3B3) был экспрессирован после индукции 0,5 ммоль изопропил-h-D-тиогалактопиранозидом (Euromedex) и очищен на Ni-NTA агарозе (Qiagen).
1.1.7 Внутриклеточное окрашивание на цитокины
Спленоциты стимулировали in vitro в течение 36 ч в присутствии (или отсутствие) полного пула пептидов, сканирующих РЭАА3В3 (pepscan). Через 1 час инкубации добавляли Brefeldin-A. Клетки пермеабилизовали с помощью FACSPerm2 (BD Biosciences) и окрашивали моноклональными антителами анти-CD4-APC, анти-CD8-PerCP и анти-ИФН-γ-FITC. Клетки анализировали на проточном цитометре FACScalibur® (BD Biosciences) и определяли процентное содержание клеток, секретирующих цитокины, после предварительного построения гейта для CD4+ или CD8+ клеток.
1.2 Результаты
1.2.1 SDS-электрофорез в полиакриламидном геле очищенных рекомбинантных белков
Для изучения способности вектора CyaA индуцировать СЕА-специфичные Т-клеточные ответы были сконструированы 3 различные рекомбинантные молекулы, содержащие домены A3 и В3 молекулы СЕА. Было показано, что этот участок СЕА содержит множество эпитопов человеческих цТЛ и хелперов (32), а также два эпитопа Н-2b, описанных к настоящему времени (33, 34). Для возможности проведения опытов in vivo и in vitro конструкции продуцировали в E.coli и очищали до гомогенности (Фиг.1). В протокол очистки (21) была добавлена процедура удаления липополисахарида для получения рекомбинантных белков, содержащих <300 единиц эндотоксина на мг (результаты не приводятся).
1.2.2. Умеренное усиление клеточного иммунного ответа, индуцированное двукратной иммунизацией рекомбинантными белками СЕА.
Предыдущие исследования показали, что рекомбинантная CyaA, несущая интересующие нас антигены, способна индуцировать Т-клеточные ответы типа Th1 при использовании различных способов инъекции (21, 35). Исследователи выясняли, приведет ли сочетание различных способов инъекций по методике прайм-буст к увеличению выраженности клеточного иммунного ответа. Как показано на Фиг.2, количество специфичных к СЕА CD4+ и CD8+ спленоцитов, секретирующих ИФН-γ, было достаточно высоким после однократного внутривенного введения 50 мкг рекомбинантных белков СЕА. Следует отметить, что CyaAΔ-CEAA3B3, лишенная домена, взаимодействующего с CD11b, значительно теряла свои векторные свойства. Напротив, белок CEAA3B3, в сочетании с адъювантом (CpG), был способен вызвать клеточный иммунный ответ типа Th1. СЕА-специфичные IL-5 CD4+ Т-клетки обнаруживались в значительном количестве только среди спленоцитов мышей, иммунизированных CyaAΔ-CEAA3B3 (результаты не приведены).
Эксперименты по иммунизации, описанные выше, повторяли с изменением способа введения вакцины: первичная вакцинация проводилась внутривенно, и через 7 и 14 дней следовала интрадермальная инъекция 10 мкг материала в дермис уха (вакцинация с праймингом и двукратной реиммунизацией). Первый буст (первая реиммунизация) приводил к умеренному повышению количества СЕА-специфичных Т-клеток у мышей, вакцинированных CyaA-CEAA3B3 или CyaA-CEAA3B3+CpG. Второй буст (вторая реиммунизация) не оказывал значительного влияния на этот показатель (Фиг.2).
Поскольку ключевой целью клеточной иммунотерапии опухолей является развитие выраженного и специфичного Т-клеточного ответа против определенных антигенов в ситуации, когда пациент часто подвергается иммуносупрессии за счет воздействия других способов лечения (химио- и лучевой терапии), важно определить условия проведения вакцинации на фоне лимфодеплеции. По этой причине исследователи провели эксперименты с иммуносупрессивными лекарственными препаратами, вызывающими умеренное и/или регулируемое истощение Т-клеток.
1.2.3. Истощение лимфоцитов периферической крови, вызванное иммуносупрессивным лечением
АТГ кролика являются поликлональными IgG, которые применялись в трансплантации органов в течение достаточно длительного времени. Исследования, проведенные на обезьянах, обнаружили, что при использовании кроличьего АТГ можно добиться дозо-зависимого истощения лимфоцитов (28). Флударабин представляет собой флуорированный аналог аденина, который часто используется для лечения хронического лимфолейкоза (39). Флударабин вызывает лимфопению за счет более выраженного истощения Т-лимфоцитов по сравнению с В-лимфоцитами (40).
Было проанализировано изменение субпопуляций лимфоцитов крови при проведении иммуносупрессивной терапии с помощью АТГ или флударабина. В предыдущих исследованиях умеренную лимфодеплецию у мышей C57BL/6 можно было вызвать однократной интреперитонеальной инъекцией 20 мг/кг веса тела (результаты не приведены). В таких условиях наблюдалось 2-кратное снижение числа циркулирующих лимфоцитов через 2 часа после инъекции АТГ (Фиг.3). Эти показатели медленно возвращались близко к нормальному уровню в течение 8 дней. Практически полное исчезновение CD4+ и CD8+ Т-лимфоцитов наблюдалось через 2 ч после инъекции АТГ, что сопровождалось относительным повышением процентного содержания В-лимфоцитов. Вслед за этим начиналось восстановление пула Т-лимфоцитов, с достижением значений близких к нормальным в течение 15 дней. По данным литературы (41, 42) режим на основе флударабина 200 мг/кг/день интраперитонеально в течение 6 дней оказывался также необходимым для индукции у мышей умеренного истощения лимфоцитов. В таких условиях истощение пула Т-лимфоцитов периферической крови оказывалось менее выраженным, чем при использовании АТГ, возвращение к нормальному уровню происходило по окончании химиотерапии.
1.2.4. Истощение лимфоцитов в лимфоидных органах, вызванное иммуносупрессивным лечением.
На кровь приходится всего от 1 до 2% общей массы лимфоидной ткани организма. Истощение лимфоцитов мониторировали путем фенотипирования Т- и В-клеток в селезенке и лимфоузлах через 10 дней от начала иммуносупрессии. Так же, как и в крови, иммуносупрессивная терапия на основе АТГ и флударабина вызывала деплецию пула Т-лимфоцитов в лимфоидных органах (Таблица 1).
В таблице 1 показана деплеция периферических лимфоидных органов, вызванная однократной инъекцией АТГ. Периферические лимфоидные органы выделяли через 10 дней от начала применения иммуносупрессивной терапии. Данные представлены как процентное содержание CD4+, CD8+ и В220 клеток, наблюдавшееся в селезенке и лимфоузлах.
При использовании АТГ эффект был более выражен, чем при использовании флударабина. Он сопровождался относительным увеличением пула В-лимфоцитов.
В заключение, в иммуносупрессивных условиях, использовавшихся в данном исследовании, наблюдалось заметное, но умеренное и преходящее уменьшение пула Т-лимфоцитов.
1.2.5. Увеличение выраженности СЕА-специфичного ответа Th1 как результат иммуносупрессии и двукратной иммунизации.
Далее требовалось определить, способна ли лимфодеплеция, проводимая в условиях, описанных выше, повысить выраженность СЕА-специфичного Т-клеточного ответа в использованной модели иммунизации. За 10 дней до двукратной иммунизации (50 мкг внутривенно и 10 мкг интрадермально через 7 дней) животные получали АТГ или флударабин, как описано выше. Выраженность СЕА-специфичного клеточного иммунного ответа была значительно усилена по сравнению с контролем у получавших АТГ и флударабин животных, иммунизированных CyaA-CEAA3B3. На фигуре 4 показано, что у животных, проходивших предварительную лимфоаблативную терапию, количество СЕА-специфичных CD4+ и CD8+ Т-клеток, секретирующих ИФН-γ, удваивалось. У животных, иммунизированных CyaAΔ-CEAA3B3, подобный эффект отсутствовал, что подчеркивает важность таргетинга CD11b для терапии с применением CyaA. Следует отметить, что предварительная лимфодеплеция приводила к выраженному увеличению количества СЕА-специфичных CD4+ Т-лимфоцитов, вырабатывающих ИФН-γ при иммунизации CEAA3B3 совместно с CpG, однако подобный эффект не наблюдался для СЕА-специфичных CD8+ Т-клеток.
1.2.6. Увеличение медианы выживаемости после двукратной иммунизации CyaA-CEAA3B3 как результат предварительной лимфоаблации.
Важность данного наблюдения была проверена на модели с отторжением опухоли. Исследователи установили клеточные линии В16, стабильно экспрессирующие белок СЕА. При инъекции 2×104 опухолевых клеток у всех животных, получавших плацебо и CyaAΔ-CEAA3B3, развились опухоли, умирающим животным требовалась эвтаназия (Фиг.5). В данных условиях медиана выживаемости животных составила соответственно 15 и 17 дней. Напротив, у животных, иммунизированных CyaA-CEAA3B3 и CEAA3B3+CpG через 3 дня после инъекции опухолевых клеток, медианы выживаемости были выше (35 и 33 дня, соответственно). У небольшой части этих животных опухолей не было и через 50 дней после инъекции клеток B16F0-CEA-GFP (Фиг.5А). При лимфодеплеции с помощью АТГ или флударабина медиана выживаемости мышей, получавших CyaA-CEAA3B3, еще более увеличилась до 45 и 41 дня соответственно, тогда как у животных, иммунизированных СЕА-A3B3+CpG, она не изменилась. Доля животных, оставшихся без опухолей, не изменилась. Как показано на Фиг.5Б, у животных, у которых опухоли имели возможность развиваться в течение 5 дней до выполнения терапевтической иммунизации CyaA-CEAA3B3, выживаемость после лимфодеплеции резко увеличивалась по сравнению с контролем. Действительно, в данной подгруппе медианы выживаемости у мышей, получавших CyaA-CEAA3B3 и CEAA3B3+CpG, снижались до 27 и 23 дней, соответственно, а на 50 день не оставалось ни одного животного, не имеющего опухоли. Лимфодеплеция не повлияла на медианы выживаемости мышей, иммунизированных CEAA3B3+CpG. Напротив, выживаемость мышей, получавших АТГ или флударабин, резко увеличивалась при иммунизации CyaA-CEAA3B3, так что выживаемость на 50 день повышалась до 80% и 60% соответственно.
1.3. Обсуждение
Множество нерешенных вопросов в области адоптивной клеточной иммунотерапии опухолей затрудняет внедрение результатов доклинических исследований в клиническую практику. Одним из них является выраженность индуцированного адоптивного ответа на определенный антиген, ассоциированный с опухолью. Действительно, у людей нарастание антиген-специфичных Т-клеток и их эффективность зависят от множества факторов, связанных с развитием опухоли. Они включают отсутствие индукции Т-клеточных клонов с высокой аффинностью к опухоле-ассоциированным антигенам, ингибирующие сигналы регуляторных клеток, опухолевой стромы и гомеостатическую регуляцию Т-клеток. В моделях адоптивного переноса Т-клеток на грызунах было показано, что лимфодеплеция увеличивает приживаемость и устойчивость пересаженных клеток (43-46). Такие технологии совсем недавно были достаточно успешно внедрены в клинике (36-38). Было решено определить роль лимфодеплеции в активной иммунизации с целью индукции специфичных Т-клеточных ответов, используя мощный инструмент для индукции специфичного ответа CD8+ и CD4+ Т-клеток, аденилатциклазу S. pertussis. В качестве мишени был выбран раково-эмбриональный антиген из-за его усиленной экспрессии при различных видах рака.
Отсутствие иммуногенности у CyaAΔ-CEAA3B3 подчеркнуло важность таргетинга CD11b для способности CyaA индуцировать специфичные Т-клеточные ответы. Как показало предыдущее исследование (21), CyaA-CEAA3B3 оказалась такой же эффективной, как CEAA3B3+CpG в отношении иммуногенности (Фиг.2) и иммунотерапии опухолей (Фиг.5).
Было показано, что предшествующий иммунитет не влияет на эффективность CyaA как вектора (20, 21), что обосновало стратегию гомологичного прайм-буста. Используя два различных способа инъекции, исследователи смогли продемонстрировать увеличение степени СЕА-специфичного иммунного ответа, хотя и не очень значительное. При повторной интрадермальной реиммунизации дальнейшее усиление клеточного иммунного ответа оказалось невозможным. Исследователи предположили, что транзиторная умеренная лимфодеплеция создаст необходимое пространство для дальнейшей амплификации специфичных Т-клеточных клонов. Внимание было сосредоточено на соединениях, которые широко применялись на человеке и вызывали истощение преимущественно Т-лимфоцитов. Профили лимфодеплеции, полученные при лечении АТГ, были схожи с теми, которые наблюдались в модели с низшими приматами (28). Общий уровень лимфодеплеции, полученный с флударабином, был ниже того, что показал АТГ, однако известно, что флударабин лучше работает в сочетании с циклофосфамидом (42).
Предварительная лимфодеплеция позволила практически удвоить количество СЕА-специфичных Т-лимфоцитов. Предстоит узнать, является ли это результатом нарушения регуляции гомеостаза Т-клеток или разрушением регуляторных Т-клеток. Нельзя исключить, что лимфодеплеция приводит к тому, что большее количество цитокинов становится доступным для оставшихся активированных лимфоцитов, что способствует их развитию и амплификации.
Лимфодеплеция не приводила к увеличению количества СЕА-специфичных CD8+ Т-клеток при иммунизации CEAA3B3+CpG в отличие от иммунизации CyaA-CEAA3B3. Это подчеркивает важность таргетинга АПК для описываемого здесь феномена. Вектор CyaA направленно воздействует на дендритные клетки за счет своей способности транслоцировать свой каталитический домен в цитозоль CD11b+антиген-презентирующих клеток (17, 18). Интересно, что похожее увеличение количества СЕА-специфичных CD8+ лимфоцитов наблюдалось на фоне лимфодеплеции в другой системе, с использованием белка hsp65 M.bovis, пришитого к CEAA3B3 (результаты не показаны). Однако нельзя исключить, что в условиях эксперимента состояние воспаления, вызванное CpG, подавляет образование достаточного для усиления иммунного ответа пула Т-клеток памяти, как это было продемонстрировано на другой модели (47).
Количество СЕА-специфичных Т-лимфоцитов было связано со способностью животных выживать при появлении опухоли, вызванном клетками B16F0-CEA-GFP, что предполагало наличие корреляции между количеством СЕА-специфичных лимфоцитов и способностью мышей контролировать и/или отторгать опухолевые клетки, экспрессирующие СЕА. Подобное влияние предварительной лимфодеплеции на эффективность двукратной иммунизации CyaA-CEAA3B3 было более выраженным в группе Д+5 по сравнению с группой Д+3. Подобное наблюдение может свидетельствовать о необходимости точного соблюдения временного интервала между лимфодеплецией и лимфостимуляцией для получения максимального эффекта, т.к. эффект лимфоаблативных препаратов может продолжаться после их исчезновения из организма. В этом отношении аналог АТГ, применяющийся у человека, оказался способным снижать количество поверхностных рецепторов лимфоцитов, что вызывало снижение их ответа на антиген-индуцированную пролиферацию (28). Нельзя исключить, что этот феномен был причиной отсутствия явного эффекта в группе Д+3, т.к. в этом случае промежуток между лимфодеплецией и иммунизацией составлял всего 8 дней.
Данное исследование показывает, что основанная на использовании CyaA активная клеточная иммунотерапия совместима с лимфодеплецией, вызванной АТГ или флударабином. В этих условиях двукратная иммунизация с применением двух различных способов инъекции (внутривенно, интрадермально) значительно увеличивала количество СЕА-специфичных Т-хелперов и, что особенно важно, СЕА-специфичных Т-цитотоксических лимфоцитов. Такое увеличение числа СЕА-специфичных Т-цитотоксических лимфоцитов коррелировало с улучшенной способностью контролировать и/или отторгать опухолевые клетки, экспрессирующие СЕА. Исходя из представленных результатов, для получения подобного эффекта более предпочтительно использовать вектор, направленный на антиген-презентирующие клетки, такой как CyaA или hsp65. Действительно, CyaAΔ, которая лишена способности связывать CD11b, теряла свою способность индуцировать СЕА-специфичные Т-клетки. Более того, ответ СЕА-специфичных CD8+ Т-клеток, индуцированный CEAA3B3 в сочетании с адъювантом CpG, не усиливался при состоянии лимфодеплеции в условиях эксперимента.
2. Пример 2.
2.1. Материалы и методы.
2.1.1. Мыши, клеточные линии
Самки мышей линии C57BL/6 возрастом от 6 до 10 недель, не имеющие специфичных патогенов, были получены в компании CER Janvier (Le Gesnet St-lsle, France). Эксперименты на животных проводились в соответствии с правилами института по обращению с животными. Инъекции при иммуносупрессивной терапии проводили интраперитонеально, инъекции опухолевых клеток - подкожно, а иммунизацию - внутривенно (ретро-орбитально) и интрадермально в дермис уха. Кровь забирали из хвостовой вены. Сплекэктомию и выделение лимфоузлов проводили у умерщвленных животных (CO2). Верхнечелюстные лимфоузлы выделяли и собирали для фенотипического анализа.
Клетки линии МС32аСЕА, экспрессирующие белок СЕА, получили у проф. W. Zimermann (LTI, Munich). Эти клетки были произведены из линии МС38 аденокарциномы толстого кишечника грызунов (48). Клетки поддерживали в RPMI 1640, содержащей Glutamax, с добавлением 10% инактивированной нагреванием сыворотки эмбрионов телят, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина и 5×10-5 моль/л 2-меркаптоэтанола в присутствии 0.3 мг/мл G418 (Life Technologies).
2.1.2. Подсчет и фенотипирование клеток
Для подсчета и фенотипирования с помощью проточной цитометрии клетки крови, лимфоузлов и спленоцитов обрабатывали согласно методикам, описанным ранее (28, 29). Лимфоциты крови подсчитывали с использованием Трипана синего.
2.1.3. Получение в организме кролика глобулинов против тимоцитов мыши
Антитимоцитарные глобулины вырабатывали путем инъекции подкожно кролику 5×108 тимоцитов, полученных у 6-недельных мышей линии C57BL/6. Через 14 дней проводили повторную иммунизацию путем внутривенного введения аналогичного материала. Через 7 дней у животного забирали кровь и выделяли сывороточный IgG путем хроматографии на колонках с иммобилизованным протеином G (Pierce). Степень чистоты контролировали путем SDS-электрофореза в полиакриламидном геле, а количество оценивали по методу Брэдфорда с известной концентрацией IgG кролика в качестве стандарта. Кроличий АТГ использовали в концентрации 20 мг/кг для однократной инъекции. В качестве контроля использовали нормальную сыворотку кролика.
2.1.4. Реагенты, пептиды и олигонуклеотиды
Флударабин (Sigma) использовали в концентрации 100 мг/кг/день в течение 6 дней. Моноклональные антитела: анти-CD4-АРС, анти-CD8-PerCP, анти-В220-FITC и анти-CD49a (Pan NK)-PE были из BD Biosciences.
Пептидный скан - пептиды длиной 15 аминокислот, перекрывающиеся по 11 аминокислот и охватывающие домен A3 и B3 белка СЕА, был разработан и получен в Mimotope™ (Australia). Пептиды разводили в DMSO и использовали в конечной концентрации 1 мкг/мл. CpG ODN 1826 был приобретен в Sigma. Указанный CpG имеет последовательность SEQ ID №16, где основаниями является фосфоротиоат (устойчивый к нуклеазам).
2.1.5. Молекулярное клонирование рекомбинантной аденилатциклазы В. pertussis, несущей домены A3 и B3 СЕА. Получение и очистка рекомбинантной CyaA.
Рекомбинантную аденилатциклазу, использованную в данном исследовании, экспрессировали в Escherichia coli, используя производные плазмиды pkTPAC-HPV16E7Δ30-42, кодирующей CyaA, не обладающую ферментативной активностью (21).
CyaA-CEAA3B3 была сконструирована в 2 этапа. Первый фрагмент ДНК, кодирующий аминокислотные остатки СЕА с 492 по 557, амплифицировали в ПЦР с использованием синтетического гена СЕА (Sequentia) и праймеров СЕА1 (SEQ ID №1: 5'-accatcaccgtctctgcg-3') и СЕА2 (SEQ ID №2: 5'-gggcactagtggtcagggtacggttgcc-3'). Второй фрагмент ДНК, кодирующий аминокислотные остатки СЕА с 629 по 687, амплифицировали в ПЦР с использованием синтетического гена и праймеров СЕАЗ (SEQ ID №3: 5'- gggcaccggtaatggtatcccgcagcaacac-3') и СЕА4 (SEQ ID №4: 5'-cgcagagacggtgatggtgttaacggcacccgcagacagacc-3'). Эти два фрагмента ДНК (которые частично перекрываются) были очищены и скомбинированы с помощью праймеров СЕА2 и СЕА3 в третьей ПЦР, в которой был получен фрагмент ДНК длиной 395 пн. Этот фрагмент был обработан рестриктазами NheI и KpnI и помещен между соответствующими сайтами pkTRAC-HPV16E7Δ30-42, что позволило получить плазмиду pkTRAC-CEAB3. Затем фрагмент ДНК, кодирующий аминокислотные остатки СЕА с 545 по 647, амплифицировали в ПЦР с использованием синтетического гена СЕА и праймеров СЕА5 (SEQ ID №5: 5'-gggcgctagccgtctgcagctgtccaatg-3') и СЕА6 (SEQ ID №6: 5'-cccgggtacccggcgtgattttggcgata-3'). Очищенный фрагмент ПЦР был обработан AgeI и SpeI и лигирован в плазмиду pkTRAC-CEAB3, обработанную теми же рестриктазами.
CyaAΔ-РЭАА3В3 была сконструирована в 2 этапа. Вначале был синтезирован фрагмент ДНК, соответствующий аминокислотной последовательности CyaA с 1149 по 1356 (SEQ ID №13 5'-caacgagctctggggccacgatggcaacgacacgatacgcggccggggcggcgacgacatcctgcgcggcggcctgggcctggacacgctgtatggcgaggacggcaacgacatcttcctgcaggacgacgagaccgtcagcgatgacatcgacggcggcgcggggctggacaccgtcgactactccgccatgatccatccaggcaggatcgttgcgccgcatgaatacggcttcgggatcgaggccatgctgtatggcgacgccggcaacgacaccctctacggggggctgggcgacgatacccttgaaggcggcgcgggcaacgattggttcggccagacgcaggcgcgcgagcatgacgtgctgcgcggcggagatggggtggataccgtcgattacagccagaccggcgcgcatgccggcattgccgc-3'), не имеющий аминокислот, соответствующих последовательности 1230-1300 (Genecust, France). Затем этот фрагмент был обработан рестриктазами Sad и SphI и лигирован в плазмиду pkTRAC-CEAA3B3, обработанную теми же рестрикционными ферментами. Полученная плазмида pKTRACΔ-CEAA3B3 кодировала CyaA, лишенную способности связывать молекулу CD11b (31).
Все конструкции проверили с помощью ДНК-секвенирования (Genome Express). Рекомбинантные CyaA продуцировали в штамме E.coli BLR (Novagen) как описано ранее (21). Рекомбинантные белки очищали из телец включений до гомогенности путем 2-ступенчатой процедуры, включающей хроматографию на ДЭАЭ-Сефарозе и фенил-Сефарозе. Дополнительный этап отмывки с помощью 60% изопропанола был проведен (21) для удаления большинства полисахаридных примесей. Очищенные рекомбинантные белки анализировали при помощи SDS-электрофореза в ПААГ с окрашиванием по Кумасси. Концентрацию белка определяли спектрофотометрически по абсорбции при 280 нм и коэффициенту молярной экстинкции 142000 моль/л при длине оптического пути 1 см.
2.1.6. Молекулярное клонирование рекомбинантного СЕА. Продукция и очистка рекомбинантного белка СЕА.
Оптимизированная для E.coli кДНК, кодирующая белок CEAA3B3, была субклонирована в вектор plvex 2.4b Nde (Roche) между рестрикционными сайтами NcoI и XhoI после амплификации в ПЦР с использованием праймеров СЕА7 (SEQ ID №14: 5'-aatctcgaggatcagcgcaacaccaacca-3') и СЕА8 (SEQ ID №15: 5'-atatccatggagtctccgtctgcc-3'). Затем полученной плазмидой трансформировали штамм E.coli BL21λDE3 (Novagen). Белок CEA-His-Tag был экспрессирован после индукции 1 ммоль изопропил-h-D- тиогалактопиранозидом (Euromedex) и очищен на Ni-NTA агарозе (Qiagen). Для удаления примесей липополисахарида использовали смолу Endotrap (Profos, Germany).
2.1.7. Тест ELISpot для определения интерферона γ
На поверхности фильтрационных планшетов Multiscreen (96 лунок, Millipore, France) в течение ночи при комнатной температуре иммобилизовали антитела крысы против мышиного интерферона гамма (ИФН-γ) (клон R4-6A2; PharMingen, San Diego, CA) в концентрации 4 мкг/мл. Клетки селезенки иммунизированных мышей вносили в лунки и инкубировали в течение 20 ч в присутствии 1 мкг/мл или в отсутствие пептидного пула (pepscan) СЕА. После тщательной отмывки планшетов проводили инкубацию 5 мкг/мл биотинилированного антитела крысы против мышиного ИФН-γ (клон XMG 1.2; PharMingen) с последующей инкубацией со стрептавидин-конъюгированной щелочной фосфатазой (PharMingen). Затем, с применением BCIP/NBT в качестве субстрата, отслеживали появление ореолов. Число клеток, продуцирующих ИФН-γ, определяли путем подсчета в каждой лунке (C.T.L, Germany) количества колоний, образующих ореолы (от англ. spot-forming colonies, SFC), и выражали результаты как число SFC на 1×106 спленоцитов (20).
2.2. Результаты
2.2.1. SDS-электрофорез в полиакриламидном геле очищенных рекомбинантных белков
Для изучения способности вектора CyaA индуцировать СЕА-специфичные Т-клеточные ответы исследователи сконструировали 2 различные рекомбинантные молекулы, содержащие субфрагменты молекулы СЕА: домены A3 и В3. Было показано, что этот участок СЕА содержит множество эпитопов человеческих цТЛ и хелперов (32). В качестве контроля был использован белок СЕА полной длины, содержащий все возможные эпитопы, с пришитым гексагистидиновым хвостом. Для проведения опытов in vivo и in vitro конструкции продуцировали в E.coli и очищали до гомогенности (Фиг.6). В протокол очистки (21) была добавлена процедура удаления липополисахарида для получения рекомбинантных белков, содержащих <300 единиц эндотоксина на мг (результаты не приводятся).
2.2.2. Умеренное усиление клеточного иммунного ответа, индуцированное двукратной иммунизацией рекомбинантными белками СЕА.
Предыдущие исследования показали, что рекомбинантная CyaA, несущая интересующие нас антигены, способна индуцировать Т-клеточные ответы типа Th1 при использовании различных способов инъекции (21, 35). Исследователи выясняли, приведет ли комбинирование различных способов инъекций по методике прайм-буст к увеличению выраженности клеточного иммунного ответа. Как показано на Фиг.7, количество специфичных к СЕА CD8+ спленоцитов, секретирующих ИФН-γ, было достаточно высоким после однократного внутривенного введения 50 мкг рекомбинантных белков СЕА. Напротив, ни CyaAΔ-CEAA3B3, ни белок CEA-His-tag в сочетании с адъювантом, таким как CpG, не были способны вызвать клеточный иммунный ответ с секрецией ИФН-γ.
Исследователи повторили эти эксперименты с изменением способа введения вакцины: первичная вакцинация проводилась внутривенно, как описано выше, и через 7 и 14 дней следовала интрадермальная инъекция 10 мкг материала в дермис уха (вакцинация с праймингом и двукратной реиммунизацией). Соответствующие инъекции были сделаны с адъювантом; первый буст (первая реиммунизация) приводил к умеренному повышению количества СЕА-специфичных Т-клеток у мышей, вакцинированных CyaA-CEAA3B3+CpG. Второй буст (вторая реиммунизация) не оказал значительного влияния на этот показатель (Фиг.7).
2.2.3. Истощение лимфоцитов периферической крови, вызванное иммуносупрессивным лечением
Флударабин представляет собой флуорированный аналог аденина, который часто используется для лечения хронического лимфолейкоза (39). Флударабин вызывает лимфопению за счет более выраженного истощения Т-лимфоцитов по сравнению с В-лимфоцитами (40). АТГ кролика являются поликлональными IgG, которые применялись в трансплантации органов в течение достаточно длительного времени. Исследования, проведенные на обезьянах, обнаружили, что с помощью данного препарата можно добиться дозо-зависимого истощения лимфоцитов (28).
Исследователи проанализировали изменение субпопуляций лимфоцитов крови при проведении иммуносупрессивной терапии с помощью флударабина или АТГ кролика.
Флударабин 100 мкг/кг/день интраперитонеально в течение 6 дней (41, 42) оказывал истощающий эффект строго на Т-клетки. Он был транзиторным, т.к. через 7 дней от начала лечения показатели возвращались к значениям контрольной группы. АТГ кролика при однократном введении 20 мг/кг интраперитонеально вызывал значительное снижение числа циркулирующих CD4+ и CD8+ лимфоцитов через 2 часа после инъекции АТГ (Фиг.8). Максимальный истощающий эффект наблюдался между 2 и 4 днями после инъекции. Впоследствии количество CD4+ и CD8+ лимфоцитов начинало повышаться, но не достигало нормальных значений. Снижение CD8+ Т-лимфоцитов было в 10 раз более выражено по сравнению с CD4+ Т-клетками. NK клетки также истощались при введении АТГ в соотношении, сопоставимом с CD4+ клетками. Количество В-клеток при лечении АТГ, напротив, не изменялось.
2.2.4. Истощение лимфоцитов в лимфоидных органах, вызванное иммуносупрессивным лечением.
На кровь приходится всего от 1 до 2% общей массы лимфоидной ткани организма. Исследователи мониторировали истощение лимфоцитов путем фенотипирования Т- и В-клеток в селезенке и лимфоузлах через 14 дней от начала иммуносупрессии. В сравнении с контролем (PBS и НКС) иммуносупрессивная терапия на основе АТГ и флударабина вызывала изменение соотношения лимфоцитов в лимфоидных органах (Таблица 2).
В таблице 2 показана деплеция периферических лимфоидных органов, вызванная однократной инъекцией АТГ. Периферические лимфоидные органы выделяли через 14 дней от начала применения иммуносупрессивной терапии. Данные представлены как процентное содержание CD4, CDS, CD49d и В220 клеток, наблюдавшееся в селезенке и верхнечелюстных лимфоузлах, и как абсолютные значения CD4, CD8, CD49d и В220 лимфоцитов, содержащихся в каждом органе.
Процентное содержание Т и NK клеток в селезенке увеличилось через 6 дней после окончания лечения флударабином. Это было наиболее очевидно в отношении CD4+ Т-клеток и в меньшей степени в отношении CD8+ и NK клеток. Соответственно, процентное содержание В220+клеток значительно снизилось через 6 дней после лечения. После лечения АТГ процентное содержание Т-клеток в селезенке и лимфоузлах понизилось, что выражалось в значительном падении числа CD4+ и CD8+. CD8+ лимфоциты были затронуты в большей степени, чем CD4+ Т-клетки. Следует отметить, что общее число лимфоцитов, восстановившихся в селезенке животных, леченых флударабином, было значительно выше, чем у контрольных животных. Кроме того, процентное содержание 0-клеток в селезенке животных, леченых АТГ и флударабином, превышало 15%. На уровне лимфоузлов подобный эффект не наблюдался.
2.2.5. Увеличение выраженности СЕА-специфичного ответа Th1 как результат иммуносупрессии и двукратной иммунизации.
Далее исследователи изучили роль лимфодеплеции, проводимой в описанных выше условиях, в отношении выраженности СЕА-специфичного Т-клеточного ответа в использованной модели иммунизации. За 7 дней до двукратной иммунизации животные получали флударабин или АТГ, как описано выше. Выраженность СЕА-специфичного клеточного иммунного ответа была снижена у иммунизированных CyaA-CEAA3B3 в сочетании с адъювантом CpG животных, получавших флударабин, по сравнению с контрольными животными, получавшими PBS. В группе животных, получавших АТГ, выраженность СЕА-специфичного клеточного иммунного ответа неожиданно оказалась в 2 раза выше по сравнению с контролем. СЕА-специфичные клеточные иммунные ответы определялись также после иммунизации с иррелевантной рекомбинантной CyaA, несущей антиген HPV16E7. Более низкие СЕА-специфичные клеточные иммунные ответы были обнаружены при иммунизации CEA-His-tag и CyaAΔ-CEAA3B3 в сочетании с CpG, что подчеркивало важность таргетинга CD11b для получения у животных упомянутого эффекта.
2.2.6. Увеличение медианы выживаемости после двукратной иммунизации CyaA-CEAA3B3 как результат предварительной лимфоаблации.
Исследователи проверили уместность данного наблюдения на модели с отторжением опухоли. Они использовали клетки линии МС38 аденокарциномы толстого кишечника грызунов, стабильно экспрессирующие белок СЕА (48). При инъекции 5×105 опухолевых клеток у животных, получавших плацебо, CyaAΔ-CEAA3B3 и CEA-His-tag развилась опухоль, умирающим животным требовалась эвтаназия (Фиг.10). В данных условиях медиана выживаемости животных составила соответственно 13-15 дней. Напротив, у животных, иммунизированных CyaA-CEAA3B3+CpG через 10 дней после инъекции опухолевых клеток, медиана выживаемости увеличилась (27 дней). При лечении АТГ медиана выживаемости мышей, получавших плацебо, увеличилась до 19-21 дня. В этой группе у 80% животных, получавших CyaA-CEAA3B3+CpG, опухоли полностью подверглись обратному развитию и животные оставались живы на 60 день. У животных, получавших флударабин, характер роста опухоли полностью отличался. Рост МСа32А частично подавлялся при лечении флударабином, длившимся в течение 6 дней. Вследствие этого медиана выживаемости животных, получавших плацебо, увеличивалась до 21-25 дней. У 60% животных, получавших CyaA-CEAA3B3+CpG, отмечалась полная регрессия опухоли и они оставались живы на 60 день.
2.3. Обсуждение
Множество нерешенных вопросов в области адоптивной клеточной иммунотерапии опухолей затрудняет внедрение результатов доклинических исследований в клиническую практику. Одним из них является выраженность индуцированного адоптивного ответа на определенный антиген, ассоциированный с опухолью. Действительно, у людей нарастание антиген-специфичных Т-клеток и их эффективность зависят от множества факторов, связанных с развитием опухоли. Они включают отсутствие индукции Т-клеточных клонов с высокой аффинностью к опухоле-ассоциированным антигенам, ингибирующие сигналы регуляторных клеток, опухолевой стромы и гомеостатическую регуляцию Т-клеток. В моделях адоптивного переноса Т-клеток на грызунах было показано, что лимфодеплеция увеличивает приживаемость и устойчивость пересаженных клеток (43-46). Такие технологии совсем недавно были достаточно успешно внедрены в клинике (36-38). Используя мощный инструмент для индукции специфичного ответа CD8+ и CD4+ Т-клеток, аденилатциклазу В. pertussis, исследователи проанализировали роль лимфодеплеции в активной иммунизации с целью индукции специфичных Т-клеточных ответов. В качестве мишени они выбрали раково-эмбриональный антиген из-за его усиленной экспрессии при различных видах рака.
Было показано, что предшествующий иммунитет не влияет на эффективность CyaA как вектора (20, 21), что обосновало стратегию гомологичного прайм-буста. При использовании двух различных способов инъекции исследователям удалось продемонстрировать увеличение степени СЕА-специфичного иммунного ответа. Им не удалось добиться дальнейшего усиления клеточного иммунного ответа при повторной интрадермальной иммунизации. Далее исследователи изучили роль лимфодеплеции в условиях активной иммунизации, опосредованной CyaA в сочетании с адъювантом CpG. Исследователи сосредоточили внимание на соединениях, которые широко применялись на человеке и вызывали истощение преимущественно Т-лимфоцитов. Общий уровень лимфодеплеции, полученный с флударабином, был ниже того, что показал АТГ.
Предварительная иммуносупрессия, вызванная флударабином, оказывала негативное воздействие на количество СЕА-специфичных Т-лимфоцитов при проведении двукратной вакцинации с помощью CyaA-CEAA3B3+CpG. Предстоит узнать, связано ли это с нарушением регуляции гомеостаза Т-клеток или разрушением регуляторных Т-клеток. Исследователи не исключают, что лимфодеплеция приводит к тому, что большее количество цитокинов становится доступным для оставшихся активированных лимфоцитов, что способствует их развитию и амплификации. Описанный эффект является достаточно примечательным и неожиданным, если принять во внимание степень лимфодеплеции на момент иммунизации, что отображено на фиг.8 и в таблице 2. Во время первичной иммунизации в контрольной группе количество циркулирующих лимфоцитов было в 20-13 раз выше по сравнению с группой АТГ. Во время реиммунизации в контрольной группе количество циркулирующих лимфоцитов было в 6 раз выше по сравнению с группой АТГ. Лимфодеплеция, зарегистрированная в лимфоидных органах, была сопоставима по выраженности.
Лимфодеплеция вызывала менее выраженное увеличение количества СЕА-специфичных CD8+ Т-клеток при иммунизации CEA-His-tag+CpG или CyaAΔ-CEAA3B3+CpG. Это подчеркивает важность таргетинга АПК для описываемого здесь феномена. Вектор CyaA направленно воздействует на дендритные клетки за счет своей способности транслоцировать свой каталитический домен в цитозоль CD11b+ антиген-презентирующих клеток (17, 18). Интересно, что похожее увеличение количества СЕА-специфичных CD8+ лимфоцитов исследователи наблюдали на фоне лимфодеплеции в другой системе, с использованием в качестве вектора белка hsp65 M. bovis, пришитого к CEAA3B3 (результаты не показаны). За исключением животных, леченых флударабином, количество СЕА-специфичных CD8+ Т-клеток было связано со способностью животных выживать при провоцирующем развитие опухоли введении клеток МСа32А, что отражало корреляцию между числом СЕА-специфичных лимфоцитов и способностью мышей контролировать и/или отторгать опухолевые клетки, экспрессирующие СЕА. Флударабин, являясь антиметаболитом, замедлял рост опухоли при введении животным МСа32А. Действительно, при иммунотерапии с применением CyaA размер опухолей в группе флударабина был в 20 раз меньше по сравнению с другими группами. Возможно, это объясняет, почему иммунотерапия CyaA-CEAA3B3 в сочетании с адъювантом CpG была настолько эффективна по сравнению с контрольной группой.
Данное исследование показывает, что основанная на использовании CyaA иммунотерапия совместима с лимфодеплецией, вызванной АТГ. В этих условиях двукратная иммунизация с применением двух различных способов инъекции (внутривенно, интрадермально) значительно увеличивала количество СЕА-специфичных CD8+ Т-клеток. Такое увеличение числа СЕА-специфичных Т-цитотоксических лимфоцитов коррелировало с улучшенной способностью контролировать и/или отторгать опухолевые клетки, экспрессирующие СЕА. На основании представленных результатов, для получения подобного эффекта необходимо использовать вектор, направленный на антиген-презентирующие клетки, такой как CyaA или hsp65. Действительно, CyaAΔ, которая лишена способности связывать CD11b, полностью теряла свою способность индуцировать СЕА-специфичные Т-клетки. Иммунотерапия с применением CyaA также совместима с лимфодеплецией, вызванной флударабином, которая приводит к увеличению выживаемости при провоцировании появления опухоли.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ БЕЛОК, НЕСУЩИЙ ЭПИТОПЫ ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА, ВСТРОЕННЫЕ В БЕЛОК АДЕНИЛАТЦИКЛАЗУ ИЛИ ЕГО ФРАГМЕНТ, ЕГО ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2005 |
|
RU2441022C2 |
| ПОЛИПЕПТИД, ПЕРЕНОСИМЫЙ CyaA (ВАРИАНТЫ), И ПРИМЕНЕНИЕ ДЛЯ ИНДУЦИРОВАНИЯ КАК ТЕРАПЕВТИЧЕСКОГО, ТАК И ПРОФИЛАКТИЧЕСКОГО ИММУННОГО ОТВЕТА | 2012 |
|
RU2668795C2 |
| МУТАНТНЫЕ ПОЛИПЕПТИДЫ СУАА И ПРОИЗВОДНЫЕ ПОЛИПЕПТИДОВ, ПОДХОДЯЩИЕ ДЛЯ ДОСТАВКИ ИММУНОГЕННЫХ МОЛЕКУЛ В КЛЕТКУ | 2010 |
|
RU2585216C2 |
| Способ получения in vitro популяций активированных антигенспецифических противоопухолевых цитотоксических Т-лимфоцитов, специфичных к эпитопам опухоль-ассоциированного антигена | 2016 |
|
RU2619186C1 |
| Полиэпитопная противоопухолевая вакцинная конструкция, содержащая эпитопы опухоль-ассоциированных антигенов, фармацевтическая композиция и ее применение для стимуляции специфического противоопухолевого иммунного ответа | 2016 |
|
RU2684235C2 |
| ПЕПТИДЫ ТЕМ8 И СОДЕРЖАЩИЕ ИХ ВАКЦИНЫ | 2008 |
|
RU2498993C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ КОМПОЗИЦИИ | 2016 |
|
RU2732118C2 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК, ОБЛАДАЮЩИХ АКТИВНОСТЬЮ ПРОТИВ КЛЕТОК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2013 |
|
RU2596920C2 |
| СПОСОБЫ ПРОГНОЗИРОВАНИЯ ПРИМЕНИМОСТИ СПЕЦИФИЧНЫХ ДЛЯ ЗАБОЛЕВАНИЯ АМИНОКИСЛОТНЫХ МОДИФИКАЦИЙ ДЛЯ ИММУНОТЕРАПИИ | 2018 |
|
RU2799341C2 |
| СЛИТЫЕ БЕЛКИ ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ ИММУНОГЕННЫХ УСИЛИВАЮЩИХ АГЕНТОВ ДЛЯ ИНДУЦИРОВАНИЯ АНТИГЕНСПЕЦИФИЧЕСКОГО Т-КЛЕТОЧНОГО ОТВЕТА | 2013 |
|
RU2631002C2 |




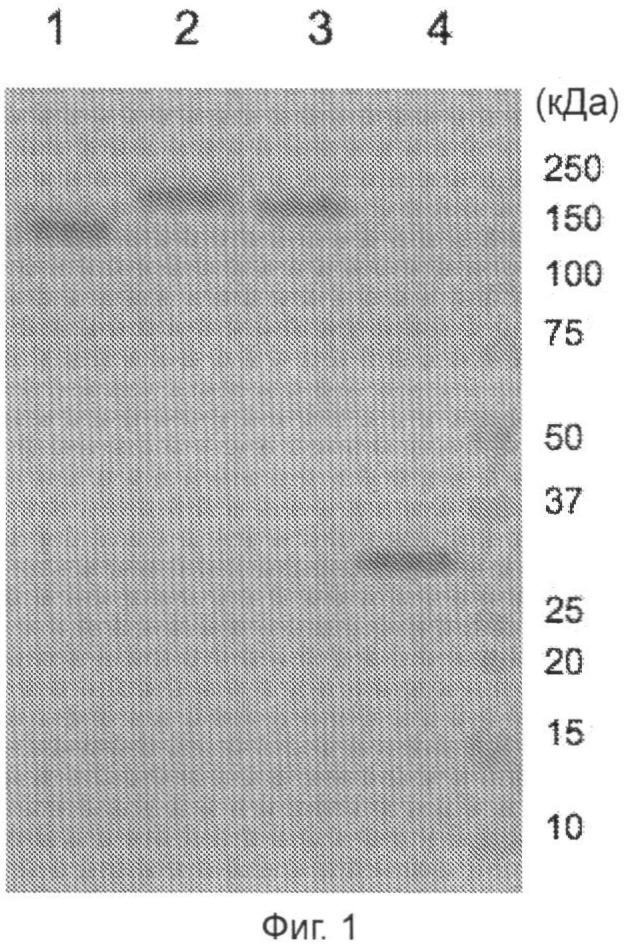
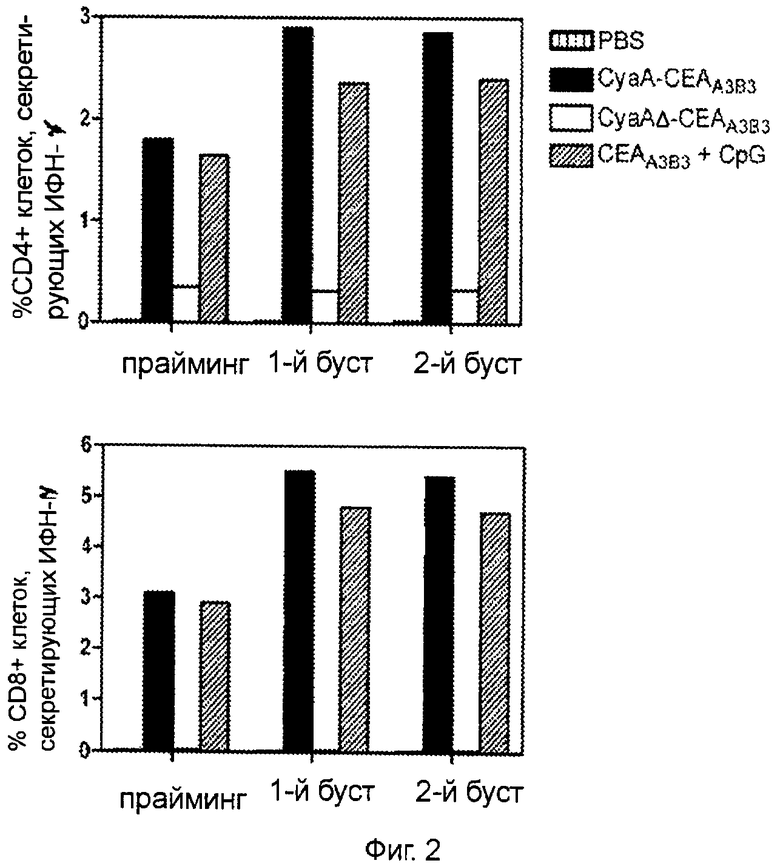
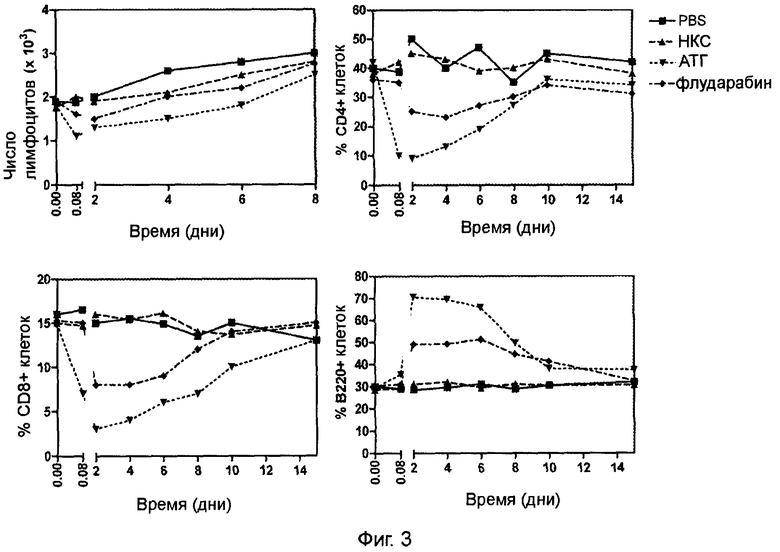
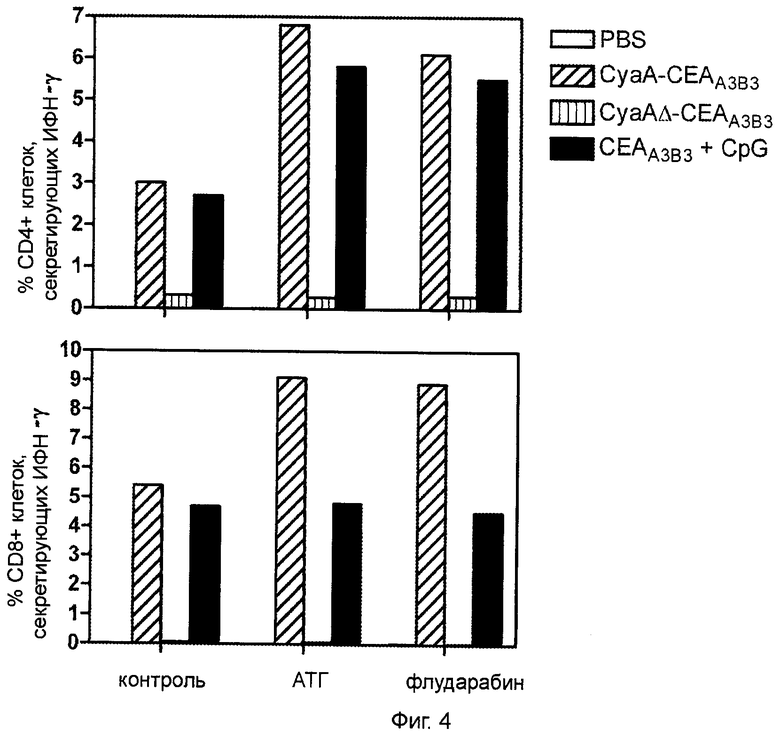
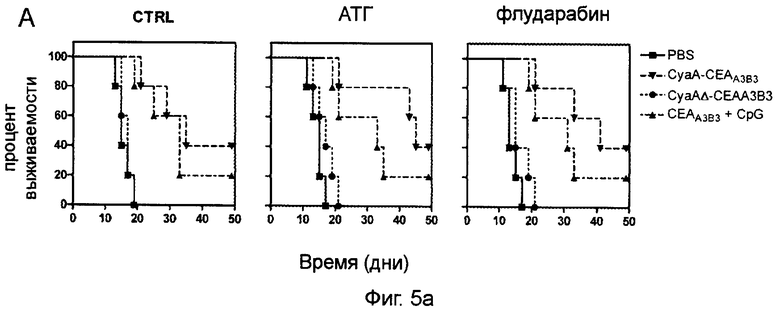
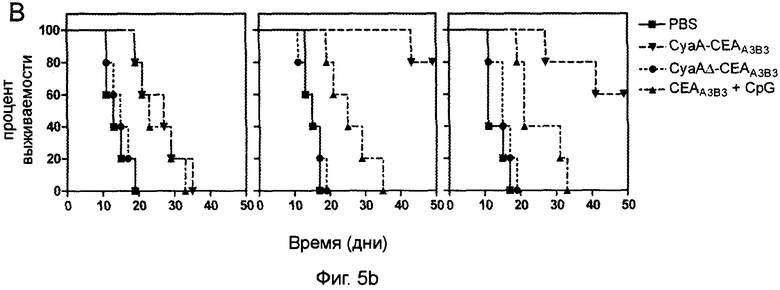

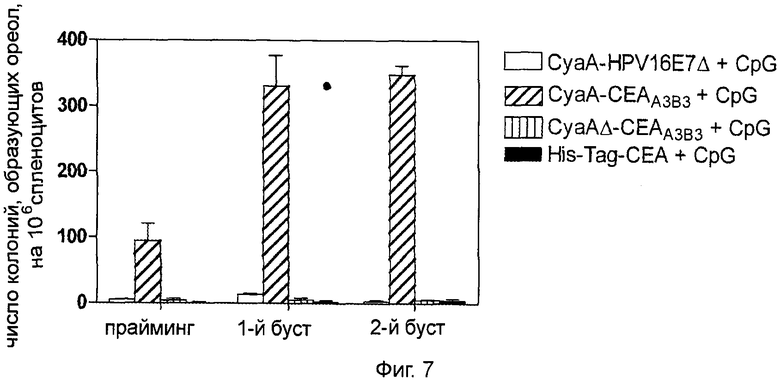
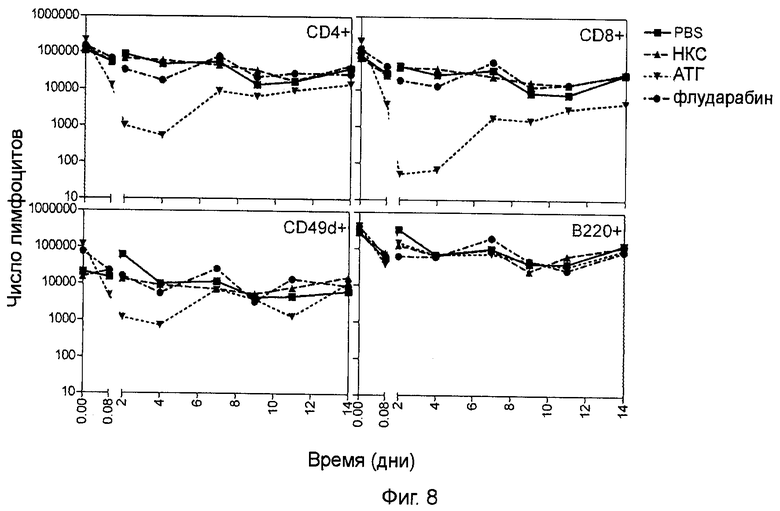
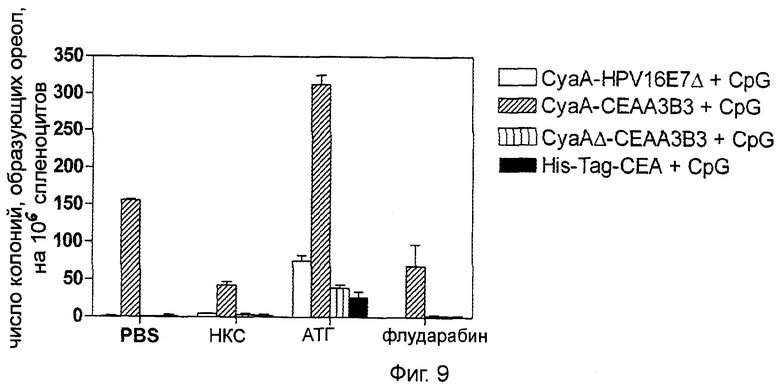
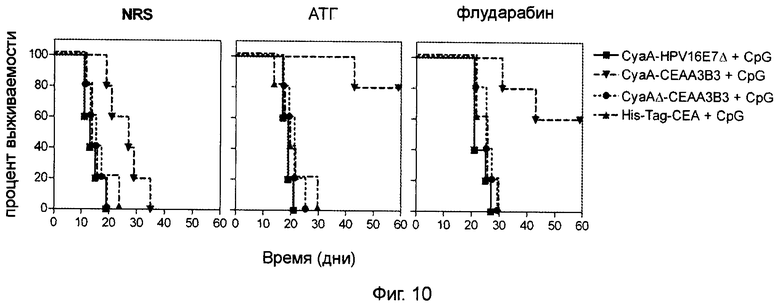
Группа изобретений относится к композициям, вызывающим специфический ответ цитотоксических Т-лимфоцитов (цТЛ) против Т-клеточных эпитопов в организме млекопитающего, включающим соединение, вызывающее лимфопению, молекулу, обладающую селективной аффинностью к специализированным антиген-презентирующим клеткам (АПК), где указанная молекула связана с указанным Т-клеточным эпитопом, и, возможно, фармацевтически приемлемый носитель. Композиции также могут содержать адъювант. С помощью композиций достигаются эффективные противоинфекционное или противоопухолевое действия. 3 н. и 17 з.п. ф-лы, 10 ил., 2 табл., 2 пр.
1. Фармацевтическая композиция, вызывающая специфический ответ цитотоксических Т-лимфоцитов (цТЛ) против по меньшей мере одного Т-клеточного эпитопа в организме млекопитающего, включающая:
- первое соединение, вызывающее лимфопению, выбранное из группы, включающей поликлональные анти-лимфоцитарные и анти-тимоцитарные иммуноглобулины (АТГ), пуриновые, пиримидиновые аналоги, алкилирующие агенты, моноклональные и поликлональные антитела, способные вызывать периферическое и/или центральное истощение Т, В и NK лимфоцитов, выбранные из группы, состоящей из анти-CD8, анти-CD4, анти-СD25, анти-СD3 и анти-СD52 моноклональных антител,
- в качестве второго соединения молекулу, имеющую селективную аффинность к специализированным антиген-презентирующим клеткам (АПК), где указанная молекула связана с указанным по меньшей мере одним Т-клеточным эпитопом антигена, полученного из инфекционного агента или злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки, где указанная молекула выбрана из группы, состоящей из аденилатциклаз (СуаА), белков теплового шока (HSP), шигатоксина и LAG-3,
- возможно, адъювант и
- возможно, фармацевтически приемлемый носитель.
2. Фармацевтическая композиция по п.1, где указанное первое соединение способно вызывать транзиторное истощение Т-клеток.
3. Фармацевтическая композиция по п.1, где аденилатциклаза происходит из Bordetella pertussis, белок теплового шока выбран из группы, состоящей из hsp65 и hsp70, а указанные hsp65 и hsp70 предпочтительно происходят из Mycobacterium bovis, а указанный шигатоксин происходит из Shigella dysenteriae.
4. Фармацевтическая композиция по п.1, где адъювант представляет собой лиганд Toll-подобного рецептора (TLR), выбранный из группы, состоящей из лигандов TLR класса 3, таких как poly-ICLC, лигандов TLR класса 9, таких как CpG, и лигандов TLR класса 4, таких как монофосфорил-А.
5. Фармацевтическая композиция по пп.1-4, где адъювант находится в растворе со вторым соединением.
6. Фармацевтическая композиция по п.1, где молекула, обладающая селективной аффинностью к АПК, ковалентно связана с указанным по меньшей мере одним Т-клеточным эпитопом.
7. Фармацевтическая композиция по п.6, где молекула, обладающая селективной аффинностью к АПК, и указанный по меньшей мере один Т-клеточный эпитоп являются полипептидами, кодирующимися последовательностями ДНК, полученными с помощью технологии рекомбинантной ДНК.
8. Фармацевтическая композиция по п.1, где инфекционный агент выбран из группы, состоящей из вируса папилломы человека (HPV), вируса иммунодефицита человека (HIV), вируса гепатита В (HVB), вируса гепатита С (HCV), Chlamydia trachomatis или Mycobacterium tuberculosis.
9. Фармацевтическая композиция по п.1, где антиген, полученный из злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки, выбран из группы, состоящей из раково-эмбрионального антигена (СЕА), MAGE-A3, теломеразы (TERT), E7 онкогена вируса папилломы человека (HPV) и Р53.
10. Фармацевтическая композиция по п.1 для применения в качестве профилактической вакцины или терапевтической вакцины.
11. Фармацевтическая композиция по п.1, где млекопитающим является человек.
12. Фармацевтическая композиция по п.1, которая предназначена для внутримышечного, внутривенного, интрадермального, кожного, подкожного, интраперитонеального, орального введения или введения в слизистую.
13. Фармацевтическая композиция, включающая:
- первое соединение, вызывающее лимфопению, выбранное из группы, включающей поликлональные анти-лимфоцитарные и анти-тимоцитарные иммуноглобулины (АТГ), пуриновые, пиримидиновые аналоги, алкилирующие агенты, моноклональные и поликлональные антитела, способные вызывать периферическое и/или центральное истощение Т, В и NK лимфоцитов, выбранные из группы, состоящей из анти-CD8, анти-CD4, анти-CD25, анти-СD3 и анти-СD52 моноклональных антител,
- в качестве второго соединения молекулу, имеющую селективную аффинность к АПК, где указанная молекула связана с по меньшей мере одним Т-клеточным эпитопом антигена, полученного из инфекционного агента, где указанная молекула выбрана из группы, состоящей из аденилатциклаз (СуаА), белков теплового шока (HSP), шигатоксина и LAG-3; и
- возможно, фармацевтически приемлемый носитель в виде комбинированного препарата для одновременного, раздельного или последовательного применения в лечении и/или предупреждении инфекционных заболеваний пациентов, где первое соединение вводится предпочтительно перед введением второго соединения.
14. Фармацевтическая композиция по п.13, где первое соединение вводится перед введением второго соединения.
15. Фармацевтическая композиция по п.14, где второе соединение находится в растворе с адъювантом.
16. Фармацевтическая композиция, включающая:
- первое соединение, вызывающее лимфопению, выбранное из группы, включающей поликлональные анти-лимфоцитарные и анти-тимоцитарные иммуноглобулины (АТГ), пуриновые, пиримидиновые аналоги, алкилирующие агенты, моноклональные и поликлональные антитела, способные вызывать периферическое и/или центральное истощение Т, В и NK лимфоцитов, выбранные из группы, состоящей из анти-CD8, анти-CD4, анти-СD25, анти-СD3 и анти-СD52 моноклональных антител,
- в качестве второго соединения молекулу, имеющую селективную аффинность к АПК, где указанная молекула связана с по меньшей мере одним Т-клеточным эпитопом антигена, полученного из злокачественной клетки, диспластически измененной клетки, опухолевой или раковой клетки, где указанная молекула выбрана из группы, состоящей из аденилатцикпаз (СуаА), белков теплового шока (HSP), шигатоксина и LAG-3; и
- возможно, фармацевтически приемлемый носитель в виде комбинированного препарата для одновременного, раздельного или последовательного применения в лечении и/или предупреждении соответственно злокачественных патологий, клеточной дисплазии, опухолевых процессов или рака у пациентов, где первое соединение вводится предпочтительно перед введением второго соединения.
17. Фармацевтическая композиция по п.16, где первое соединение вводится перед введением второго соединения.
18. Фармацевтическая композиция по п.17, где второе соединение находится в растворе с адъювантом.
19. Фармацевтическая композиция по любому из пп.16-18 для применения в лечении и/или предупреждении рака, когда раковые клетки экспрессируют белок СЕА, предпочтительно рак дыхательного или пищеварительного тракта, где первое соединение представляет собой поликлональный АТГ кролика или лошади, а второе соединение представляет собой рекомбинантную аденилатциклазу, несущую участок А3-В3 человеческого белка СЕА.
20. Фармацевтическая композиция по любому из пп.16-18 для применения в лечении и/или предупреждении рака, вызванного HPV, при котором раковые клетки экспрессируют онкоген Е7 HPV, где первое соединение представляет собой поликлональный АТГ кролика или лошади, а второе соединение представляет собой рекомбинантную аденилатциклазу, несущую онкоген Е7 HPV.
| WO 9947166 A1, 23.09.1999 | |||
| WENDELBO ⌀, BRUSERUD ⌀, Functional evaluation of proliferative Т cell responses in patients with severe Т lymphopenia: characterization of optimal culture conditions and standardized activation signals for a simple whole blood assay | |||
| J Hematother Stem Cell Res | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Телескоп | 1920 |
|
SU525A1 |
| COR H.J | |||
| ET AL. | |||

