Область техники
Настоящее изобретение относится к способу получения пуриновых нуклозидов, являющихся важным исходным материалом для синтеза 5'-инозиновой кислоты и 5'-гуаниловой кислоты. Настоящее изобретение также раскрывает новые микроорганизмы, используемые в указанном способе. Настоящее изобретение также раскрывает новые ДНК и виды белков, придающие бактерии, экспрессирующей новую ДНК, устойчивость к аналогам пуриновых оснований и к пуриновым нуклеозидам.
Предшествующий уровень техники
Традиционно, промышленное производство нуклеозидов, таких как инозин, осуществляется с помощью ферментации штаммов, ауксотрофных по аденину, или штаммов, которым сообщили устойчивость к ряду веществ, таких как аналоги пурина и сульфагуанидина. Примерами таких штаммов могут являться штаммы, принадлежащие к роду Bacillus (патент Японии №54-17033 (1979), 55-2956 (1980), и 55-45199 (1980) выложенная патентная заявка Японии №56-162998 (1981), патент Японии №57-14160 (1982) и 57-41915 (1982), и выложенная патентная заявка Японии №59-42895 (1984)), или роду Brevibacterium (патент Японии №51-5075 (1976) и 58-17592 (1972), и Agric. Biol. Chem, 42, 399 (1978)), или роду Escherichia (WO 9903988) и подобные им.
Процесс конструирования таких мутантных штаммов обычно включает в себя мутагенез интересующих микроорганизмов посредством УФ излучения или обработкой нитрозогуанидином (N-метил-N'-нитро-N-нитрозогуанидин) и последующий отбор штамма с заданными характеристиками на соответствующей селективной среде. С другой стороны, для конструирования таких мутантных штаммов также применялись методы генной инженерии, при этом указанные штаммы принадлежали к роду Bacillus (выложенная патентная заявка Японии №58-158197 (1983), 58-175493 (1983), 59-28470 (1984), 60-156388 (1985), 1-27477 (1989), 1-174385 (1989), 3-58787 (1991), 3-164185 (1991), 5-84067 (1993) и 5-192164 (1993)), к роду Brevibacterium (выложенная патентная заявка Японии №63-248394 (1988)) или к роду Escherichia (WO 9903988).
Продуктивность таких штаммов-продуцентов инозина может быть в дальнейшем повышена за счет усиления способности штамма к экскреции инозина. На сегодняшний день общепринятым является тот факт, что прохождение метаболитов через цитоплазматическую клеточную мембрану обычно осуществляется благодаря специфичным эффлюксным белкам-транспортерам (Рао, S.S. et al., Microbiol. Mol. Biol. Rev., 62 (1), 1-34, (1998); Paulsen, I.T. et al., J. Mol. Biol, 277, 573-592 (1998); Saier, M.H. et al., J. Mol. Microbiol. Biotechnol., 257-279 (1999)). Авторы настоящего изобретения ранее показали, что штаммы-продуценты инозина или ксантозина, принадлежащие к родам Escherichia или Bacillus, а также обладающие повышенной активностью белка RhtA, кодируемого геном rhtA (ybiF), повышенной активностью белка YijE, кодируемого геном yijE, и повышенной активностью белка YdeD, кодируемого геном ydeD, продуцируют больше инозина или ксантозина, чем соответствующие родительские штаммы (патентные заявки России 2002101666, 2002104463 и 2002104464 соответственно). Недавно было обнаружено, что белок YdhL из В.subtilis снижает чувствительность бактериальных клеток к аналогу пурина - 2-флуороаденину. С другой стороны, существуют косвенные доказательства того, что белок YdhL понижает клеточный пул гипоксантина (Johansen et al., J. Bacteriol., 185, 5200-5209,2003). Однако не было показано эффекта влияния суперэкспрессии гена ydhL на экскрецию нуклеозидов и/или продукцию нуклеозидов.
Более того, не существует публикаций ни о белке YdhL из Bacillus amyloliquefaciens, ни о влиянии суперэкспрессии ydhL на экскрецию нуклеозидов и/или продукцию нуклеозидов.
Краткое описание чертежей.
На Фиг.1 изображено сравнение белка YdhL из В.amyloliquefaciens (ydhL_B.am), кодируемого фрагментом ДНК, входящего в состав плазмиды pYDHL4, и белка YdhL из В.subtilis (ydhL_Bac.su), кодируемого фрагментом ДНК, входящего в состав плазмиды pYDHL
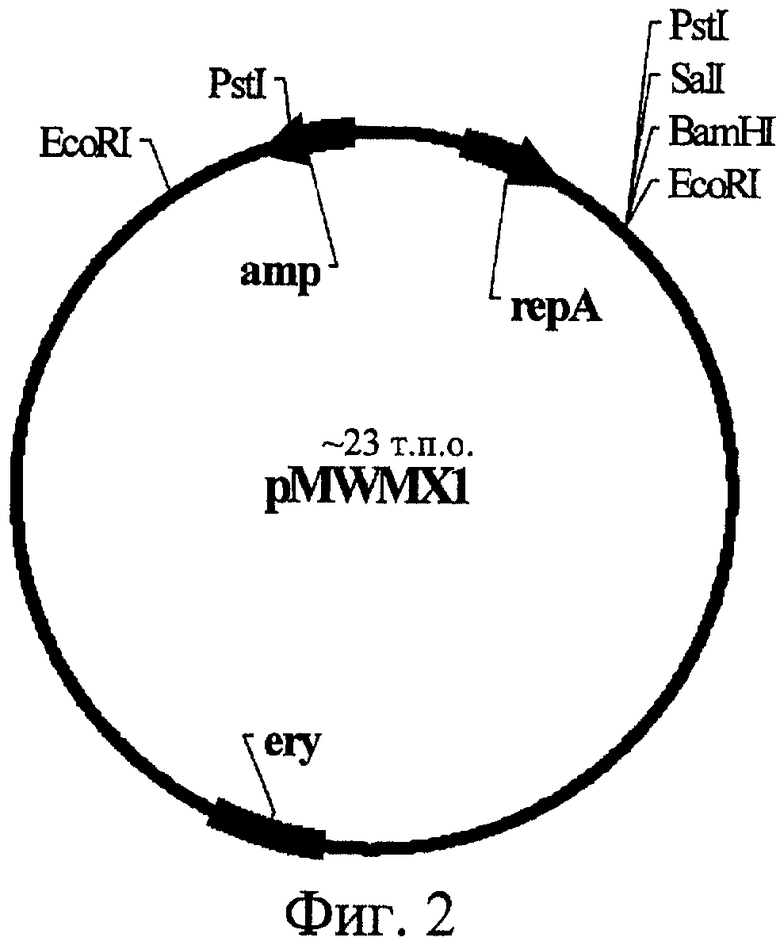
На Фиг.2 изображена структура плазмиды pMWMX1.
Описание изобретения
Целью настоящего изобретения является увеличение продуктивности штаммов-продуцентов инозина, а также предоставление способа получения инозина и 5'-инозиновой кислоты с использованием указанных штаммов. Эта цель была достигнута путем идентификации того факта, что ген ydhL из В.subtilis, кодирующий гипотетический мембранный белок, сообщает устойчивость к аналогам пуриновых оснований, таким как 8-азааденин, 6-метилпурин, 2,6-диаминопурин, 6-меркаптопурин, 8-азагуанин, и пуриновым нуклеозидам, таким как инозин и гуанозин, в случае, когда природная аллель указанного гена, клонированная на низкокопийном векторе, вводится в штамм Е.coli. Также был выделен ранее неизвестный ген ydhL из Bacillus amyloliquefaciens, и определена его нуклеотидная последовательность. Далее было установлено, что ген ydhL из Bacillus subtilis или Bacillus amyloliquefaciens может повысить продукцию нуклеозидов, когда дополнительные копии указанного гена введены в клетки соответствующего штамма-продуцента инозина, принадлежащего к роду Bacillus или роду Escherichia. Таким образом было совершено настоящее изобретение.
Следовательно, настоящее изобретение предоставляет микроорганизм, принадлежащий к роду Bacillus или роду Escherichia и обладающий способностью к продукции инозина.
Целью настоящего изобретения является предоставление белка YdhL из Bacillus amyloliquefaciens, выбранного из группы, состоящей из следующих белков:
(А) белок, включающий последовательность аминокислот, приведенную в списке последовательностей под номером 2 (SEQ ID NO:2); и
(В) белок, включающий последовательность аминокислот, содержащую делеции, замены, вставки или добавление одной или нескольких аминокислот в последовательности аминокислот, приведенной в списке последовательностей под номером 2 (SEQ ID NO:2), и который обладает активностью придавать бактерии, принадлежащей к роду Bacillus или к роду Escherichia, устойчивость к пурину, выбранному из группы, состоящей из аналогов основания, инозина и гуанозина, когда указанная активность указанного белка в указанной бактерии повышена.
Также целью настоящего изобретения является предоставление ДНК, кодирующей описанный выше белок.
Также целью настоящего изобретения является предоставление бактерии, принадлежащей к роду Bacillus или к роду Escherichia, при этом указанная бактерия обладает способностью к продукции пуринового нуклеозида, и указанная бактерия обладает повышенной активностью белка, выбранного из группы, состоящей из (А) белка, кодируемого геном ydhL из Bacillus amyloliquefaciens, (В) белка, кодируемого геном ydhL из Bacillus subtilis, и (С) белка, кодируемого ортологом любого из указанных генов ydhL, описанных в пункте (А) или (В).
Также целью настоящего изобретения является предоставление описанной выше бактерии, где пуриновым нуклеозидом является инозин или гуанозин.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой активность указанного белка повышена путем трансформации указанной бактерии с помощью ДНК, кодирующей данный белок, или путем изменения нуклеотидной последовательности, контролирующей экспрессию вышеупомянутой ДНК в хромосоме указанной бактерии.
Также целью настоящего изобретения является предоставление описанной выше бактерии, в которой трансформация осуществлена в составе низкокопийного вектора.
Также целью настоящего изобретения является предоставление способа продукции пуриновых нуклеозидов, включающего в себя выращивание описанной выше бактерии в питательной среде, позволяющее накапливать в указанной питательной среде указанные пуриновые нуклеозиды, и выделение из культуральной жидкости указанных пуриновых нуклеозидов.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором пуриновым нуклеозидом является инозин, ксантозин или гуанозин.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором указанная бактерия модифицирована таким образом, что экспрессия генов пути биосинтеза пуриновых нуклеозидов усилена.
Также целью настоящего изобретения является предоставление способа продукции пуриновых нуклеотидов, включающего в себя стадии выращивания описанной выше бактерии в питательной среде, позволяющего накапливать в питательной среде указанные пуриновые нуклеотиды, фосфорилирования пуриновых нуклеозидов и выделения пуриновых нуклеотидов.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором пуриновым нуклеотидом является 5'-инозиновая кислота, 5'-ксантозиновая кислота или 5'-гуаниловая кислота.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором пуриновым нуклеотидом является инозиновая кислота.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором указанная бактерия модифицирована таким образом, что экспрессия генов пути биосинтеза пуриновых нуклеотидов усилена.
Настоящее изобретение более детально будет описано ниже.
Наилучший способ осуществления изобретения.
I. Белки согласно настоящему изобретению.
Белки согласно настоящему изобретению включают в себя белок, аминокислотная последовательность которого приведена в списке последовательностей под номером 2 (SEQ ID NO:2), и белок, аминокислотная последовательность которого содержит делеции, замены, вставки или добавление одной или нескольких аминокислот в последовательности аминокислот, приведенной в списке последовательностей под номером 2 (SEQ ID NO:2), и который обладает активностью, придающей бактерии, принадлежащей к роду Bacillus или к роду Escherichia, устойчивость к аналогам пуринового основания или к пуриновым нуклеозидам, таким как инозин и гуанозин.
Белком, аминокислотная последовательность которого приведена в списке последовательностей под номером 2 (SEQ ID NO:2), является белок YdhL из Bacillus amyloliquefaciens. Аминокислотная последовательность указанного белка и нуклеотидная последовательность гена ydhL были неизвестны до настоящей публикации.
Белок YdhL из Bacillus subtilis не участвует в пути биосинтезапуриновых нуклеотидов и принадлежит к суперсемейству транспортных белков Major Facilitator Superfamily (Pao, S.S., et al, Microbiol. Mol. Biol. Rev. 62 (I): 1-32, (1998)) и к группе ортологов кластера пермеазы эффлюкса арабинозы (COG) 2814 (Tatusov, R.L. et al, Nucleic Acids Res., 29: 22-28 (2001)). Другие белки Е.coli, принадлежащие к этому семейству - AraJ и YdeA, также участвуют в экспорте арабинозы (Bost, S. et al, J. Bacteriol, 181: 2185-2191 (1999)). Геном Bacillus subtilis содержит еще 5 генов, кодирующих гомологи гена: ydhL: ybcL, yceJ, yfhI, ytbD, ywfA. Белок YdhL из В.subtilis - это высокогидрофобный белок с неизвестной функцией, состоящий из 388 аминокислот и содержащий 12 гипотетических трансмембранных сегментов. Белок YdhL кодируется геном ydhL. Ген ydhL из Bacillus subtilis subsp. subtilis str.168 (номера нуклеотидов с 624675 по 625838 в последовательности с инвентарным номером NC_000913.1 в GenBank; GL32468705) расположен на хромосоме между генами ydhK и ydhM в положении 53.50°.
Количество «нескольких» аминокислот, которое может быть увеличено в белках согласно настоящему изобретению, различается в зависимости от положения и/или типа аминокислотного остатка в трехмерной структуре белка. Это количество может составлять от 2 до 40, предпочтительно от 2 до 20 и более предпочтительно от 2 до 5 для белка, приведенного под номером 2 (SEQ ID No. 2). Этот факт объясняется тем, что некоторые аминокислоты обладают высокой гомологией друг к другу и различия между такими аминокислотами не оказывает существенного влияния на трехмерную структуру белка и его активность. Таким образом, белком, аминокислотная последовательность которого содержит делеции, замены, вставки или добавление одной или нескольких аминокислот в последовательности аминокислот, приведенной в списке последовательностей под номером 2 (SEQ ID NO:2), может являться белок, имеющий гомологию не меньшую, чем от 30% до 50%, предпочтительно от 50% до 70% и наиболее предпочтительно от 70% до 90% по отношению к общему количеству аминокислотных остатков, входящих в состав белка YdhL, и сохраняющий активность указанного белка.
Белок, обладающий гомологией как минимум 90%, более предпочтительно гомологией как минимум в 95%, еще более предпочтительно гомологией как минимум в 98% к белку, представленному в списке последовательностей под номером 2 (SEQ ID NO:2), и сохраняющий активность белка YdhL, является белком согласно настоящему изобретению и наиболее предпочтителен.
Для оценки степени гомологии между ДНК возможно использование нескольких способов расчета, таких как BLAST search, FASTA search и CrustalW.
BLAST (Basic Local Alignment Search Tool) это самообучающийся алгоритм поиска, используемый программами BLASTP, BLASTN, BLASTX, MEGABLAST, TBLASTN и TBLASTX; эти программы оценивают значимость найденных результатов с использованием статистических методов Karlin, Samuel и Stephen F. Altschul ("Methods for assessing the statistical significance of molecular sequence features by using general scoring schemes". Proc. Natl. Acad. Sci. USA, 1990, 87:2264-68; "Applications and statistics for multiple high-scoring segments in molecular sequences". Proc. Natl. Acad. Sci. USA, 1993, 90:5873-7). Способ поиска FASTA описан W. R. Pearson ("Rapid and Sensitive Sequence Comparison with FASTP and FASTA", Methods in Enzymology, 1990 183:63-98). Способ ClustalW описан у Thompson J.D., Higgins D.G. и Gibson T.J. ("CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice", Nucleic Acids Res. 1994, 22:4673-4680).
Термин "активность" по отношению к белкам согласно настоящему изобретению обозначает, что указанный белок, находясь в бактерии, придает этой бактерии устойчивость к аналогам пуринового основания, таким как 8-азааденин, 2,6-диаминопурин, 6-меркаптопурин, 6-тиогуанин, и устойчивость к нуклеозидам, таким как инозин и гуанозин.
Фраза "повышает устойчивость к аналогам пуринового основания и к пуриновым нуклеозидам" означает способность бактерии расти на минимальной среде, содержащей аналоги пуринового основания, такие как 8-азааденин, 6-метилпурин, 2,6-диаминопурин, 6-меркаптопурин, 8-азагуанин, или пуриновые нуклеозиды, такие как инозин и гуанозин, в концентрациях, при которых природный или родительский штамм не смог бы расти в тех же условиях, или способность бактерии быстрее расти на среде, содержащей аналоги пуринового основания или пуриновые нуклеозиды, по сравнению с природным или родительским штаммом указанной бактерии. Вышеупомянутые концентрации аналогов пуринового основания составляют 300 μг/мл для 8-азааденина; 400 μг/мл в случае 2,6-диаминопурина; 400 μг/мл для 6-меркаптопурина; и для пуриновых нуклеозидов эти концентрации составляют 300 μг/мл, предпочтительно 1000 μг/мл для инозина и 10 μг/мл для гуанозина.
II. Бактерия согласно настоящему изобретению.
Бактерией согласно настоящему изобретению является бактерия, в которой продукция пуриновых нуклеозидов указанной бактерией повышена за счет повышения активности указанного белка. Более конкретно, бактерией согласно настоящему изобретению является бактерия, где продукция пуриновых нуклеозидов указанной бактерией повышена за счет повышения активности белка согласно настоящему изобретению в этой бактерии. Еще более конкретно, бактерия согласно настоящему изобретению содержит ДНК гена ydhL или его ортолог в хромосоме или на плазмиде. ДНК гена ydhL или его ортолог суперэкспрессируются, и в результате указанная бактерия способна продуцировать большие количества пуриновых нуклеозидов.
Бактерией согласно настоящему изобретению является бактерия, принадлежащая к любому из родов Bacillus или Escherichia. В качестве примера микроорганизма, принадлежащего к роду Escherichia, который может быть использован в настоящем изобретении, можно привести Escherichia coli (Е.coli). Примеры микроорганизмов, принадлежащих к роду Bacillus, которые могут быть использованы в настоящем изобретении, включают в себя Bacillus subtilis subsp.subtilis str.168 (В.subtilis 168) или Bacillus amyloliquefaciens (В.amyloliquefaciens).
Примеры бактерий, принадлежащих к роду Bacillus, также включают в себя следующие микроорганизмы:
Bacillus licheniformis;
Bacillus pumilis;
Bacillus megaterium;
Bacillus brevis;
Bacillus polymixa;
Bacillus stearothermophilus.
В дополнение к уже упомянутым характеристикам бактерия согласно настоящему изобретению может обладать другими специфическими особенностями, такими как потребность в различных питательных веществах, устойчивость к антибиотикам, чувствительность к антибиотикам и зависимость от антибиотиков, не выходя при этом за рамки настоящего изобретения.
Используемый здесь термин "пуриновый нуклеозид" включает в себя инозин, ксантозин, гуанозин и аденозин, предпочтительно инозин.
Используемый здесь термин "способность к продукции пуринового нуклеозида", обозначает способность к продукции и к накоплению пуринового нуклеозида в культуральной жидкости. Термин "обладающий способностью к продукции пурина" обозначает, что микроорганизм, принадлежащий к роду Escherichia или Bacillus, способен продуцировать и вызывать накопление пуринов, таких как пуриновые нуклеозиды, в культуральной жидкости в количествах бóльших, чем природные штаммы Е.coli, такие как штаммы Е.coli W3110 и MG1655, или природные штаммы В.subtilis, такие как В.subtilis 168. Предпочтительно этот термин обозначает, что указанный микроорганизм способен продуцировать и вызывать накопление в культуральной жидкости инозина, ксантозина, гуанозина и/или аденозина в концентрациях не менее чем 10 мг/л; более предпочтительно в концентрациях не менее чем 50 мг/л.
Используемый здесь термин также означает, что если в бактерии активность белка согласно настоящему изобретению повышена, то повышено количество молекул белка в пересчете на клетку или повышена активность самой молекулы белка.
Термин "ген-ортолог" или "ортолог" обозначает ген из одного вида микроорганизма, соответствующий гену из другого вида, при этом оба эти гена являются родственными, поскольку имеют общего предка (ген-гомолог), но в процессе эволюции стали отличаться между собой в разных видах организмов. Гены-ортологи могут кодировать белки, обладающие одной или разными функциями. Если известны полные нуклеотидные последовательности хромосом организмов, для поиска ортологов может быть использован следующий алгоритм. Поиск с использованием BLAST проводится таким образом, что для гена из одного организма ищется лучший гомолог в другом организме. Затем проводится второй, обратный поиск BLAST, в котором для лучшего гомолога из другого организма, найденного на первом этапе поиска BLAST, находится лучший гомолог в первом организме. Если ген, для которого ищется гомолог на первом этапе поиска, совпадает с геном, найденным на втором этапе поиска, то ген из первого организма и его лучший гомолог в другом организме являются ортологами. Также в процессе поиска BLAST определяется степень гомологии белков, кодируемых найденными генами.
Для целей настоящего изобретения белки, кодируемые генами-ортологами, являются гомологичными белками и обладают одинаковой функцией. Поиск таких генов ортологов производится путем определения сиквенса этих генов и белков, кодируемых этими генами, а также функциями этих белков (смотри, например. Фиг.1 и Пример 2).
Нуклеотидная последовательность гена, кодирующего белок YdhL из Bacillus amyloliquefaciens, была определена авторами настоящего изобретения (SEQ ID NO:1). Нуклеотидная последовательность гена, кодирующего белок YdhL из Bacillus subtilis, ген ydhL из Bacillus subtilis subsp. subtilis str.168 (номера нуклеотидов с 13042 по 14208 в базе данных EMBL, инвентарный номер последовательности - Z99107) (SEQ ID NO:3) известен. Ген ydhL из Bacillus subtilis расположен на хромосоме между генами ydhK a ydhM в положении 53.50°. Следовательно, вышеупомянутые гены ydhL могут быть синтезированы с помощью ПЦР (полимеразная цепная реакция; по White, T.J. et al., Trends Genet., 5,185 (1989)) с использованием праймеров, полученных на основе нуклеотидной последовательности целевого гена. Гены, кодирующие белок YdhL в других микроорганизмах, могут быть получены аналогичным способом (смотри Пример 2).
Примером гена ydhL, полученного из Bacillus amyloliquefaciens, также может служить фрагмент ДНК, который кодирует белки согласно настоящему изобретению, такие как белок с аминокислотной последовательностью, приведенной под номером 2 (SEQ ID NO:2) и/или белок с аминокислотной последовательностью, включающей в себя делеции, замены, вставки или добавление одной или нескольких аминокислот в последовательности аминокислот, приведенной в списке последовательностей под номером 2 (SEQ ID NO:2), и который обладает активностью, придающей бактерии, принадлежащей к роду Bacillus или к роду Escherichia, устойчивость к аналогам пуриновых оснований, инозину и гуанозину, при этом активность указанного белка в указанной бактерии повышена.
Ген ydhL, полученный из Bacillus subtilis, также может быть представлен ДНК, которая кодирует белки согласно настоящему изобретению, такие как белок с аминокислотной последовательностью, приведенной под номером 4 (SEQ ID NO:4) и/или белок с аминокислотной последовательностью, включающей в себя делеции, замены, вставки или добавление одной или нескольких аминокислот в последовательности аминокислот, приведенной в списке последовательностей под номером 4 (SEQ ID NO:4), и который обладает активностью придавать бактерии, принадлежащей к роду Bacillus или к роду Escherichia, устойчивость к аналогам пуриновых оснований, инозину и гуанозину.
ДНК, кодирующие практически такие же белки, как описанные выше белки YdhL, могут быть получены, например, путем модификации соответствующей нуклеотидной последовательности с использованием сайт-направленного мутагенеза таким образом, что один или несколько аминокислотных остатков будут удалены, заменены, введены или добавлены. Модифицированная подобным образом ДНК может быть получена традиционными методами, вызывающими мутации. К подобного рода обработкам относятся обработка in vitro ДНК, кодирующей белки согласно настоящему изобретению, с помощью гидроксиламина, или обработка бактерии, содержащей указанную ДНК, с помощью УФ излучения или химического реагента, такого как N-метил-N'-нитро-N-нитрозогуанидин или азотистая кислота.
ДНК, кодирующая практически такой же белок, как белок YdhL, выделенный из Bacillus amyloliquefaciens, может быть обнаружена путем экспрессии указанной ДНК, содержащей описанные выше мутации, в подходящей для этой цели клетке и изучения активности всех экспрессируемых продуктов. ДНК, кодирующая практически такой же белок, как белок YdhL, может быть получена путем выделения ДНК из мутантной ДНК, кодирующей белок YdhL, или из клетки, которая содержит мутантную ДНК, при этом указанная ДНК гибридизуется с используемой в качестве зонда ДНК, содержащей нуклеотидную последовательность, приведенную под номером 1 (SEQ ID NO:1), в жестких условиях и кодирует белок, обладающий активностью белка YdhL. Термин «жесткие условия», упомянутый здесь, означает условия, при которых образуются так называемые специфические гибриды, а неспецифические - не образуются. Трудно четко описать такие условия в каких-либо численных выражениях. Тем не менее, например, к жестким условиям относятся условия, при которых гибридизуются ДНК, обладающие высокой степенью гомологии, к примеру ДНК, обладающие гомологией не менее 50% относительно друг друга, но ДНК, обладающие гомологией менее 50% относительно друг друга, не гибридизуются. С другой стороны, примером жестких условий являются условия, при которых ДНК гибридизуются друг с другом, при концентрации солей, соответствующей стандартным условиям отмывки при гибридизации по Саузерну, например 1×SSC, 0.1% SDS, предпочтительно 0.1×SSC, 0.1% SDS при 60°С. Продолжительность отмывки зависит от вида фильтра, используемого для блоттинга, и, как правило, рекомендована изготовителем. Например, рекомендованная продолжительность отмывки нейлонового фильтра Hybond™ N+(Amersham) в жестких условиях составляет 15 минут.
Часть нуклеотидной последовательности, приведенной под номером 1 (SEQ ID NO:1), также может быть использована в качестве зонда. Зонды могут быть получены с помощью ПЦР, с использованием праймеров, синтезированных на основе нуклеотидной последовательности, приведенной под номером 1 (SEQ ID NO:1), и фрагмента ДНК, содержащего нуклеотидную последовательность, приведенную под номером 1 (SEQ ID NO:1), в качестве матрицы. Когда фрагмент ДНК, длина которого составляет примерно 300 п.о., используется в качестве зонда, то условия отмывки при гибридизации могут быть следующими, например: 50°С, 2×SSC и 0.1% SDS.
Все, описанное выше, применимо к белку YdhL, полученному из Bacillus subtilis.
Способы повышения активности белка согласно настоящему изобретению и, прежде всего, способы повышения количества молекул указанного белка в бактериальной клетке, включают в себя: изменение последовательности, регулирующей экспрессию ДНК, кодирующей белок согласно настоящему изобретению, и увеличение количества копий гена, но не ограничиваются ими.
Изменение последовательности, регулирующей экспрессию ДНК, кодирующей белок согласно настоящему изобретению, может быть достигнуто помещением ДНК, кодирующей белок согласно настоящему изобретению, под контроль сильного промотора. В качестве сильных промоторов известны lac промотор, trp промотор, trc промотор, PL промотор фага лямбда. Кроме того, промотор может быть усилен, например, введением мутации в последовательность промотора для повышения уровня транскрипции гена, расположенного за промотором. Более того, уровень трансляции мРНК может быть повышен за счет введения мутации в спейсерный участок между сайтом связывания рибосомы (RBS) и старт-кодоном. Например, была обнаружена 20-кратная разница в уровне экспрессии гена, в зависимости от состава последовательности из трех нуклеотидов, находящихся перед старт-кодоном (Gold et al., Annu. Rev. Microbiol., 35, 365-403, 1981; Hui et al., EMBO J., 3, 623-629, 1984).
Кроме того, для повышения уровня транскрипции указанного гена, в хромосому может быть введен энхансер. Введение в хромосому ДНК, содержащей либо ген, либо промотор, описано, например, в выложенной патентной заявке Японии №1-215280 (1989).
Также количество копий указанного гена может быть увеличено благодаря клонированию указанного гена на векторах разной копийности с образованием рекомбинантной ДНК, которой затем трансформируют целевой микроорганизм. Более предпочтительно использование низкокопийных векторов. Примерами низкокопийных векторов являются, но не ограничиватся ими, векторы pSC101, pMW118, pMW119 и подобные им. Термин "низкокопийный вектор" используется для обозначения векторов, количество копий которых на клетку не превышает 5. Способы трансформации включают в себя все возможные способы, известные специалисту в данной области. Например, опубликован способ обработки клеток-реципиентов Escherichia coli К-12 хлоридом кальция для повышения проницаемости клеточных стенок для ДНК (Mandel, M. and Higa, A., J. Mol. BioL, 53, 159 (1970)).
Усиление экспрессии гена может также быть достигнуто путем введения нескольких копий гена в бактериальную хромосому, например, методом гомологичной рекомбинации, Mu интеграцией или подобными им. К примеру, один цикл Mu интеграции позволяет ввести в бактериальную хромосому до 3-х копий указанного гена.
Описанные выше способы, основанные на использовании сильного промотора или энхансера, могут быть совмещены со способами, основанными на увеличении числа копий гена или усилении экспрессии указанного гена.
В настоящем изобретении могут быть использованы традиционные методы получения хромосомной ДНК, гибридизации, ПЦР, получения плазмидной ДНК, разрезания и лигирования ДНК, трансформации, выбора олигонуклеотидов в качестве праймеров и подобные им. Эти методы описаны, например, в книге Sambrook, J., and Russell D., "Molecular Cloning A Laboratory Manual, Third Edition", Cold Spring Harbor Laboratory Press (2001) и подобным ей.
Для конструирования микроорганизма, принадлежащего к роду Bacillus, обладающего повышенной экспрессией указанного гена или генов, кодирующего(их) белок или белки согласно настоящему изобртению, необходимые области указанного гена или генов могут быть получены с помощью ПЦР, синтезированные, в первую очередь, на основе доступной информации о генах В.subtilis. Например, ген ydhL, возможно являющийся геном, кодирующим транспортный белок, может быть клонирован из хромосомной ДНК штаммов В.subtilis с помощью ПЦР. Хромосомная ДНК, используемая для этой цели, может быть получена из любого другого штамма В.subtilis.
Белок согласно настоящему изобретению включает в себя также мутанты и варианты белка YdhL, которые существуют ввиду природного разнообразия, а также гомологичные белку YdhL белки, которые могут быть получены из различных штаммов бактерий, при условии, что эти белки-мутанты, варианты и гомологи обладают функциональными характеристиками белка YdhL, а именно придают клетке устойчивость к аналогам пуриновых оснований, инозину или гуанозину.
Бактерия согласно настоящему изобретению может быть получена путем введения описанной выше ДНК, кодирующей белки согласно настоящему изобретению, в бактерию, обладающей наследуемой способностью к продукции пуриновых нуклеозидов, таких как инозин. Кроме того, бактерия согласно настоящему изобретению может быть получена путем придания бактерии, уже содержащей указанные ДНК, способности к продукции пуриновых нуклеозидов, таких как инозин.
Примером родительского штамма, принадлежащего к роду Bacillus, который мог быть использован в настоящем изобретении, может являться штамм В.subtilis KMBS16-1. Этот штамм является производным штамма-продуцента инозина В.subtilis KMBS16, в котором мутация purA::erm была заменена на мутацию purA::cat. В свою очередь, штамм В.subtilis KMBS 16 является производным от известного штамма В.subtilis trpC2, несущего мутации в гене purA, кодирующем сукцинил-АМФ синтазу (purA::erm), в гене purR, кодирующем пуриновый репрессор (purR::spc), и в гене deoD, кодирующем фосфорилазу пуриновых нуклеозидов (deoD::kan) (патентная заявка России 2002101666). Также примеры родительских штаммов, принадлежащих к роду Bacillus, которые могут быть использованы в настоящем изобретении, включают в себя штамм В.subtilis АЛ 2707 (FERM Р-12951) (патентная заявка Японии JP6113876А2), штамм В.subtilis AJ3772 (FERM P-2555) (патентная заявка Японии JP62014794A2) и подобные им.
Примером штамма-продуцента инозина, принадлежего к виду Е.coli, является FADRaddedd (pMWKQ). Этот штамм является производным от известного штамма W3110, несущего мутации в гене purF, кодирующем PRPP амидотрансферазу, в гене purr, кодирующем пуриновый репрессор, в гене deoD, кодирующем фосфорилазу пуриновых нуклеозидов, в гене purA, кодирующем кодирующем сукцинил-АМФ синтазу, в гене add, кодирующем аденозиндеаминазу, в гене edd, кодирующем 6-фосфоглюконатдегидразу (WO 9903988), и содержит плазмиду pMWKQ, несущую ген purFKQ, кодирующий PRPP амидотрансферазу, и является нечувствительным к ГМФ (WO 9903988).
Для увеличения удельной активности молекулы белка у белка согласно настоящему изобретению также возможно введение мутации в структурный ген, кодирующий указанный белок, для повышения активности данного белка. Для введения мутации в ген могут быть использованы: сайт-специфический мутагенез (Kramer, W. and Frits, H.J., Methods in Enzymology, 154, 350 (1987)), рекомбинантная ПЦР (PCR Technology, Stockton Press (1989)), химический синтез определенной части ДНК, обработка целевого гена гидроксиламином, обработка штаммов микроорганизмов, содержащих целевой ген, УФ излучением или химическим реагентом, таким как нитрозогуанидин или азотистая кислота и подобные им. Микроорганизмы, в которых активность указанного белка повышена, могут быть отобраны по росту на минимальной среде, содержащей 8-азааденин, или 6-метилпурин, или 2,6-диаминопурин, или 6-меркаптопурин, или 8-азагуанин, или инозин в концентрациях, указанных в Таблице 1 (смотри ниже).
Бактерия согласно настоящему изобретению может быть в дальнейшем модифицирована таким образом, что экспрессия одного или нескольких генов, участвующих в биосинтезе пуринов, будет усилена. Такие гены могут быть представлены pur регулоном из Е.coli (Escherichia coli and Salmonella, Second Edition, Editor in Chief: F.C.Neidhardt, ASM Press, Washington D.C., 1996). Был описан штамм-продуцент инозина Е.coli, содержащий мутантный ген purF, кодирующий PRPP амидотрансферазу, не подверженную ингибированию АДФ и ГМФ по принципу обратной связи, и инактивированный ген purR, кодирующий репрессор системы биосинтеза пуриновых нуклеотидов (WO 9903988).
III. Способ получения пуриновых нуклеозидов.
К способу согласно настоящему изобретению относится способ продукции пуриновых нуклеозидов, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, позволяющего указанной бактерии продуцировать и накапливать пуриновые нуклеозиды, и сбора пуриновых нуклеозидов из культуральной жидкости. Далее, к способу согласно настоящему изобретению относится способ продукции инозина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, позволяющего указанной бактерии продуцировать и накапливать инозин, и сбора инозина из культуральной жидкости. Далее, к способу согласно настоящему изобретению относится способ продукции ксантозина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, позволяющего указанной бактерии продуцировать и накапливать ксантозин, и сбора ксантозина из культуральной жидкости. Далее, к способу согласно настоящему изобретению относится способ продукции гуанозина, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, позволяющего указанной бактерии продуцировать и накапливать гуанозин, и сбора гуанозина из культуральной жидкости.
Согласно настоящему изобретению выращивание, сбор и очистка пуриновых нуклеозидов из культуральной или подобной ей жидкости могут быть осуществлены способом, подобным традиционным способам ферментации, в которых пуриновые нуклеозиды продуцируются с использованием микроорганизма. Питательная среда, используемая для получения пуриновых нуклеозидов, может быть традиционной питательной средой, содержащей источники углерода, азота, неорганические ионы и другие необходимые органические вещества. В качестве источников углерода могут быть использованы различные сахара, такие как глюкоза, лактоза, сахароза, галактоза, фруктоза, арабиноза, мальтоза, ксилоза, трегалоза, рибоза и гидролизаты крахмала;
спирты, такие как глицерин, маннитол и сорбитол; органические кислоты, такие как глюконовая кислота, фумаровая кислота, лимонная кислота и янтарная кислота, и подобные им.
В качестве источника азота могут использоваться различные неорганические соли аммония, такие как сульфат аммония, хлорид аммония и фосфат аммония, и органические соединения азота, такие как гидролизат соевых бобов, газобразный аммиак и раствор аммиака в воде, и подобные им. Желательно, чтобы в питательную среду входили в необходимых количествах в качестве следовых питательных веществ витамины, такие как витамин В1, необходимые вещества, например нуклеиновые кислоты, такие как аденин и РНК, или дрожжевой экстракт или подобные им. Кроме указанных компонентов, при необходимости, в питательную среду могут быть добавлены небольшие количества фосфата калия, сульфата магния, ионы железа и ионы марганца.
Выращивание осуществляется предпочтительно в аэробных условиях, от 16 до 72 часов, при поддерживаемой температуре культивирования от 30 до 45°С и рН питательной среды находится в пределах от 5 до 8. рН среды может регулироваться различными органическими и неорганическими кислотами, основаниями и в том числе газообразным аммиаком.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем целевой пуриновый нуклеозид может быть получен из культуральной жидкости любым из традиционных методов или любой их комбинацией, таких как метод ионообменной хроматографии и высушивания.
IV. Способ получения пуриновых нуклеотидов.
К способу согласно настоящему изобретению относится способ продукции пуриновых нуклеотидов, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, позволяющего указанной бактерии продуцировать и накапливать пуриновые нуклеозиды в культуральной жидкости, фосфорилирования полученного пуринового нуклеозида, и сбора пуринового нуклеотида. Далее, к способу согласно настоящему изобретению относится способ продукции 5'-инозиновой кислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, позволяющего указанной бактерии продуцировать и накапливать 5'-инозиновую кислоту в культуральной жидкости, фосфорилирования полученной 5'-инозиновой кислоты, и сбора 5'-инозиновой кислоты. Далее, к способу согласно настоящему изобретению относится способ продукции 5'-ксантиловой кислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, позволяющего указанной бактерии продуцировать и накапливать ксантозин в культуральной жидкости, фосфорилирования ксантозина, и сбора 5'-ксантиловой кислоты. Далее, к способу согласно настоящему изобретению относится способ продукции 5'-гуаниловой кислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, позволяющего указанной бактерии продуцировать и накапливать гуанозин в культуральной жидкости, фосфорилирования гуанозина, и сбора 5'-гуаниловой кислоты. Также к способу согласно настоящему изобретению относится способ продукции 5'-гуаниловой кислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, позволяющего указанной бактерии продуцировать и накапливать ксантозин в культуральной жидкости, фосфорилирования ксантозина, аминирования 5'-ксантиловой кислоты и сбора 5'-гуаниловой кислоты.
Согласно настоящему изобретению выращивание, сбор и очистка инозина из культуральной или подобной ей жидкости могут быть осуществлены способом, подобным традиционным способам ферментации, в которых инозин продуцируется с использованием микроорганизма. Далее, согласно настоящему изобретению стадии фосфорилирования инозина, позволяющие указанной бактерии выделять 5'-инозиновую кислоту в культуральную жидкость, и сбора 5'-инозиновой кислоты могут быть осуществлены способом, подобным традиционным способам ферментации, в которых пуриновые нуклеотиды, такие как 5'-инозиновую кислоту получают из пуриновых нуклеозидов, таких как инозин.
Фосфорилирование пуриновых нуклеозидов может быть осуществлено ферментативно, с использованием различных фосфатаз, нуклеозидных киназ или нуклеозидных фосфотрансфераз; или химически, с использованием фосфорилирующих агентов, таких как РОС1з или подобные ему. Может быть использована фосфатаза, способная катализировать селективный перенос фосфорной группы пирофосфата на С-5'-позицию нуклеозидов (Mihara et. al., Phosphorylation of nucleosides by the mutated acid phosphatase from Morganella morganii. Appl. Environ. Microbiol., 66:2811-2816 (2000)) или кислотная фосфатаза, использующая в качестве донора фосфорной кислоты полифосфорную кислоту (соли), фенилфосфорную кислоту (соли) или карбамил фосфорную кислоту (соли) (WO 9637603A1) или подобные им. Также примером фосфатазы является фосфатаза, которая способна к катализу реакции переноса фосфорной группы на С-2', 3', или 5'-позиции нуклеозида с р-нитрофенилфосфата (Mitsugi, К., et al, Agric. Biol. Chem., 28, 586-600 (1964)), с неорганического фосфата (выложенная патентная заявка Японии JP42-1186) или ацетилфосфата (выложенная патентная заявка Японии JP61-41555) в качестве донора, или подобные ей. Также примером нуклеотидной киназы является гуанозин/инозин киназа из Е.coli (Mori, H. et. al. Cloning of a guanosine-inosine kinase gene of Escherichia coli and characterization of the purified gene product. J. Bacteriol. 177:4921-4926 (1995); WO 9108286) или подобные ей. Также примером нуклеозидной фосфотрансферазы являются нуклеозидные фосфотрансферазы, описанные у Hammer-Jespersen, К. (Nucleoside catabolism, p.203-258. In A Munch-Petesen (ed.), Metabolism ofnucleotides, nucleosides, and nucleobases in microorganism. 1980, Academic Press, New York) или подобные им. Химическое фосфорилирование нуклеозидов может осуществляться при помощи фосфорилирующего агента, такого как POCl3 (Yoshikawa, К. et. al. Studies of phosphorylation. III. Selective phosphorylation of unprotected nucleosides. Bull. Chem. Soc. Jpn. 42:3505-3508 (1969)) или подобного ему.
Аминирование 5'-ксантиловой кислоты может осуществляться ферментативно, с использованием, например, ГМФ синтетазы из Е.coli (Fujio et. al. High level of expression of XMP aminase in Escherichia coli and its application for the industrial production of 5'-guanylic acid. Biosci. Biotech. Biochem. 1997,61:840-845; EP 0251489 B1).
Примеры
Более детально настоящее изобретение будет разъяснено ниже со ссылкой на следующие Примеры, не ограничивающие область применения настоящего изобретения.
Пример 1. Клонирование гена ydhL из В.subtilis.
Полная нуклеотидная последовательность генома штамма В.subtilis subsp.subtilis 168 определена (Kunst et al., Nature 390, 249-256 (1997)). На основе заявленной нуклеотидной последовательности были синтезированы праймеры ydhL_N (SEQ ID No. 5) и ydhL_C (SEQ ID NO:6), которые в дальнейшем использовались для амплификации гена ydhL из В.subtilis с помощью ПЦР. Праймер ydhL_N комплементарен нуклеотидной последовательности с 489 до 467 п.о. перед старт кодоном гена ydhL и содержит введенный в 5'-конец сайт узнавания рестриктазы Sail. Праймер ydhL_C комплементарен нуклеотидной последовательности с 58 по 80 п.о. после стоп кодона гена ydhL и содержит введенный в 5'-конец сайт узнавания рестриктазы BglII.
Хромосомная ДНК штамма В.subtilis 168 была получена традиционными методами. ПЦР осуществлялся на амплификаторе "Perkin Elmer GeneAmp PCR System 2400" при следующих условиях: 30 сек при 94°С, 45 сек при 61°С, 3 мин при 72°С, 25 циклов с использованием PfuI полимеразы (Fermentas). Полученный ПЦР фрагмент, содержащий ген ydhL под собственным промотором, был обработан смесью рестриктаз BglI и SalI и клонирован в низкокопийный вектор pMW118, также предварительно обработанный смесью рестриктаз BamHI и SalI. Таким образом была получена плазмида pYDHL1.
Для трансформации штамма Е.coli TG1 с помощью стандартных методов были использованы плазмида pYDHLl и соответствующий вектор. Трансформанты отбирали на чашках с L-агаром с добавлением 100 мкг/мл ампициллина. Таким образом были получены штаммы Е.coli TG1 (pYDHL1) и TG1 (pMW118). Затем оценивалась способность этих штаммов к росту в присутствии аналогов пуриновых оснований на чашках с минимальной глюкозосодержащей средой М9 с добавлением различных концентраций ингибитора роста. На чашки высевали от 105 до 106 клеток из ночной культуры, выращенной на минимальной среде (с добавлением 100 мг/мл ампициллина). Рост оценивался через 44 часа инкубирования при температуре 37°С.
Результаты представлены в Таблице 1.
Как видно из Таблицы 1, амплификация гена ydhL повышает устойчивость бактерий к 2,6-диаминопурину, 6-меркаптопурину и 8-азааденину.
Плазмида pYDHL1 и вектор pMW118 также были введены в штамм Е.coli TG1 deoD gsk3 путем трансформации. Штамм Е.coli TG1 deoD gsk3 является специально сконструированным родительским штаммом, используемым для оценки устойчивости к инозину и гуанозину. Этот штамм был получен путем последовательного введения мутации deoD, как описано в патентной заявке РСТ WO 9903988, и мутации gsk3, описанной у Petersen С. (J. Biol. Chem., 274, 5348-5356, 1999) с использованием трансдукции фагом PI в известный штамм TG1 (ВКПМ В-5837), важной характеристикой которого является его устойчивость к инозину и гуанозину, благодаря способности штамма деградировать пурин. Штамм TG1 deoD gsk3 не способен к деградации гуанозина и инозина за счет введенной мутации deoD (WO 9903988) и чувствителен к нуклеозидам за счет мутации gsk3 (Petersen С.J. Biol. Chem., 274, 5348-5356, 1999). Таким образом были получены штаммы TG1 deoD gsk3 (pYDHL1) и TG1 deoD gsk3 (pMW118). Способность указанных штаммов расти в присутствии инозина и гуанозина определялась, как описано выше. Результаты представлены в Таблице 2.
Из Таблицы 2 видно, что повышенная экспрессия гена ydhL придает штаммам повышенную устойчивость к инозину и некоторую устойчивость к гуанозину. Таким образом, можно предположить, что ген ydhL участвует в экскреции нуклеозидов и, более конкретно, в экскреции инозина.
Пример 2. Клонирование гена ydhL из Bacillus amyloliquefaciens.
Данные, приведенные в Примере 1, показывают, что повышенная экспрессия гена ydhL из В.subtilis (далее: ydhLBs) придает клеткам устойчивость к 2,6-диаминопурину и 6-меркаптопурину, одновременно добавленным в агаризованную М9 минимальную глюкозосодержащую среду (Таблица 1). На основе этого факта было осуществлено клонирование ортолога гена ydhLBs из В.amyloliquefaciens (далее: ydhLBa). Хромосомная ДНК из штамма В.amyloliquefaciens BA1 была получена традиционными методами. Хромосомную ДНК обработали рестриктазой EcoRI и лигировали с вектором pMW118, также предварительно обработанным той же рестриктазой. Полученной ДНК трансформировали штамм Е.coli TG1. Трансформанты были отобраны на минимальной среде с добавлением 400 мкг/мл 2,6-диаминопурина, 400 мкг/мл 6-меркаптопурина и 100 мкг/мл ампициллина. Плазмидную ДНК выделили из трансформантов и проанализировали. В этом эксперименте была получена гибридная плазмида pYDHL2, содержащая минимальный размер вставки ДНК - примерно 1.6 т.п.о. В отличие от гена ydhLBs, этот фрагмент не содержит сайта узнавания рестриктазы EcoRI. Полученная вставка была секвенирована с использованием универсальных праймеров, что показало, что фрагмент обладает высокой степенью гомологии (более 90%) с геном ydhLBs (Фиг.1). Новый ген из В.amyloliquefaciens был назван геном ydhL (здесь и далее обозначающийся как ген ydhLBa). Будучи введенной в штаммы TG1 и TG1 deoD gsk3, плазмида pYDHL2 придает клеткам устойчивость к аналогам пуриновых оснований и к нуклеозидам, соответственно, точно так же как и плазмида pYDHLI. Таким образом был сделан вывод, что плазмида pYDHL2 действительно содержит ген ydhLBa.
Пример 3. Эффект амплификации генов ydhLBs и ydhLBa на продукцию инозина штаммом-продуцентом инозина Е.coli.
Штамм-продуцент инозина Е.coli FADRaddedd (pMWKQ) трансформировали отдельно вектором pMW18, и каждой из плазмид YDHL1 и pYDHL2. Трансформанты отбирали на среде L-агар, содержащей 100 мг/л ампициллина и 75 мг/л канамицина. Таким образом были получены штаммы FADRaddedd (pMWKQ, pMW18), FADRaddedd (pMWKQ, pYDHL1) и FADRaddedd (pMWKQ, pYDHL2). Все эти штаммы выращивались при 37°С в течение 18 часов в L-бульоне со 100 мг/л ампициллина и 75 мг/л канамицина, после чего 0.3 мл полученной культуры вносили в 20×200 мм пробирки с 3 мл ферментационной среды, содержащей 100 мг/л ампициллина и 75 мг/л канамицина, и выращивали при 37°С в течение 72 часов на роторной качалке.
Состав ферментационной среды (г/л):
Глюкозу и сульфат магния стерилизовали отдельно. CaCO3 стерилизовали сухим жаром при температуре 180°С в течение 2 часов. рН доводили до 7.0. Антибиотики добавляли в среду после стерилизации.
После выращивания количество накопленного в среде инозина определяли с помощью ВЭЖХ. Пробу культуральной жидкости (500 мкл) центрифугировали при 15,000 об/мин в течение 5 минут, после чего супер разводили в 4 раза водой (Н2О) и анализировали с помощью ВЭЖХ.
Условия ВЭЖХ анализа:
Колонка: Luna C18(2) 250×3 мм, 5 мкм (Phenomenex, США). Буфер: 2% v/v C2H5OH; 0.8% v/v триэтиламин; 0.5% v/v уксусная кислота (ледяная); рН 4.5. Температура: 30°С. Скорость потока: 0.3 мл/мин. Объем пробы: 5-10 мкл. Длина волны: UV 250 нм. Время удерживания (мин):
Результаты представлены в Таблице 3.
Как видно из Таблицы 3, повышенная экспрессия генов ydhLBs и ydhLBa повышает продукцию инозина у штамма FADRaddedd (pMWKQAp).
Пример 4. Клонирование гена ydhLBs в низкокопийный челночный вектор pMWMX1.
ПЦР фрагмент, содержащий ген ydhLBs под собственным промотором (полученный, как описано в Примере 1), был обработан рестриктазами BglI и SalI и клонирован в низкокопийный челночный вектор pMWMX1, также предварительно обработанный рестриктазами BamHI и SalI. Таким образом была получена плазмида pYDHL3. Вектор pMWMX1 (Фиг.2) является производным от двух низкокопийных плазмид: pMW118 и рМХ30. Плазмида рМХ30 (Rabinovich et al., Mol. Biol (Moscow), 18, 189-196,1984), в свою очередь, является делеционным вариантом плазмиды pSM19035, вектора с широким спектром хозяев семейства inc18 (Brantl et al., Nucleic Acids Res., 18, 4783-4790). Обе плазмиды pMW118 и рМХ30 были обработаны рестриктазой EcoRI и затем дотированы. Таким образом был получен вектор pMWMX1.
Пример 5. Конструирование низкокопийной челночной плазмиды pYDHL3, содержащей ген vdhLBa.
Плазмида pYDHL2 была обработана рестриктазой EcoRI с недорестрикцией, для получения одного фрагмента, и дотирована в плазмиду рМХ30, обработанную той же рестриктазой. Таким образом была получена челночная плазмида pYDHL4, содержащая ген ydhLBa. Таким образом, плазмида pYDHL4 эквивалентна плазмиде pYDHL3, за исключением замены вектора pMWMX1, содержащего гены ydhLBs и ydhLBa.
Пример 6. Эффект амплификации гена ydhLBs или гена ydhLBa на устойчивость штамма В.subtilis 168 к аналогам пуриновых оснований.
Плазмиды pYDHL3, pYDHL4 и вектор pMWMX1 были использованы для трансформации природного штамма В.subtilis 168 с помощью стандартных методов. Трансформанты были отобраны на чашках с L-агаром, с добавлением 10 мкг/мл эритромицина. Таким образом были получены штаммы В.subtilis 168(pMWMX1), В.subtilis 168(pYDHL3) и В.subtilis 168(pYDHL4). Затем способность этих штаммов к росту в присутствии аналогов пуриновых оснований оценивали на чашках с минимальной агаризованной средой с добавлением глюкозы М9 (минимальная среда М9, глюкоза - 2%, витаминная смесь казаминовых кислот (Difco) - 0,1%, триптофан - 100 мкг/мл), содержащей различные концентрации пуриновых аналогов. На чашки наносили от 105 до 106 клеток из ночной культуры, выращенной на чашках с описанной выше минимальной агаризованной средой без аналогов. Рост оценивался через 48 часов инкубации при 34°С.
Результаты представлены в Таблице 4.
Как видно из Таблицы 4, амплификация гена ydhLBs или гена ydhLBa повышает устойчивость бактерий к 8-азагуанину, 6-метилпурину и 8-азааденину.
Пример 7: Эффект амплификации гена ydhLBs или гена ydhLBa на продукцию инозина штаммом-продуцентом инозина В.subtilis.
Штамм-продуцент инозина В.subtilis KMBS16-1 трансформировали вектором pMWMX1, плазмидой pYDHL3 или плазмидой pYDHL4. Таким образом были получены штаммы В.subtilis KMBS16-1 (pMWMX1), В.subtilis KMBS16-1 (pYDHL3) и В.subtilis KMBS16-1 (pYDHL4). Каждый из этих штаммов выращивали в L-бульоне с добавлением эритромицина (10 мг/л) при 37°С в течение 18 часов. Затем по 0.3 мл полученной культуры инокулировали в пробирку 20×200 мм с 3 мл ферментационной среды Bacillus, содержащей эритромицин (10 мг/л), и выращивали при 37°С в течение 72 часов на роторной качалке.
Состав ферментационной среды Bacillus: (г/л)
После выращивания количество накопленного в среде инозина определяли с помощью ВЭЖХ, как описано выше. Результаты представлены в Таблице 5.
Как видно из Таблицы 5, амплификация любого из генов ydhLBs или ydhLBa повышает продукцию инозина штаммом В.subtilis KMBS16-1.
Пример 8. Модификация регуляторной области гена ydhL из В.amyloliquefaciens.
Известно, что модификация регуляторной области гена ydhL из В.subtilis, a именно модификация в G-боксе 5'-нетранслируемой области РНК, может повысить экспрессию указанного гена (Mandal, M. and Breaker, R.R., Nat. Struct. Mol. BioL, 11 (1), 29-35 (2004)). В связи с этим, с целью усиления уровня экспрессии гена ydhL такая же мутация (замена Т на С) была введена в регуляторную область гена ydhL из В.amyloliquefaciens (положение "-77" от старт-кодона указанного гена) с использованием набора для сайт-направленного мутагенеза QuickChange™ (Stratagene). Для этой цели на основе последовательности ДНК регуляторной области гена ydhL из В.amyloliquefaciens были синтезированы два праймера 1 и 2 (SEQ ID NO:7 и 8, соответственно). В качестве матрицы использовали ДНК плазмиду pYDHL2 (pMW118-ydhLBA). Указанная плазмида была амплифицирована целиком с использованием вышеописанных праймеров и получено несколько плазмид, содержащих ген ydhL. Нуклеотидные последовательности указанных плазмид были определены с использованием праймера 3 (SEQ ID NO:9) и наличие желаемой замены Т на С было подтверждено. Новая плазмида, содержащая ген ydhL из В.amyloliquefaciens с мутацией в регуляторной области, была названа pYDHL5. Плазмида pYDHL5 была линеаризована рестриктазой EcoRI и затем дотирована с плазмидой рМХ30, предварительно линеаризованной той же рестриктазой. Таким образом была получена челночная плазмида pYDHL6, содержащая ген ydhLBa с мутацией, усиливающей уровень экспрессии этого гена.
Пример 9. Эффект усиления экспрессии гена ydhLBa на продукцию инозина штаммом-продуцентом инозина В.amyloliquefaciens.
Штамм В.subtilis 168 был трансформирован плазмидой pYDHL6 или челночным вектором pMWMX1 с использованием хорошо известного метода (Spizizen, J. Proc. Natl. Acad. Sci. USA, 44, 1072-1078 (1958)). Полученные штаммы В.subtilis (pYDHL6) и В.subtilis (pMWMX1) были использованы в качестве доноров для трансдукции с использованием фага Е40 с целью введения плазмид pYDHL6 и pMWMX1 в штамм В. subtilis G1136A (AJ1991) - продуцент инозина (патент США 3,575,809). Указанный штамм В.subtilis G1136A был переклассифицирован Всероссийской коллекцией промышленных микроорганизмов (ВКПМ) (РФ, 117545 Москва, 1-й Дорожный проезд, 1) в штамм Bacillus amyloliquefaciens и депонирован под инвентарным номером (VKPM В-8994). Поэтому в дальнейшем указанный штамм называется Bacillus amyloliquefaciens G1136A. Фаг Е40 является PBS1-подобным фагом (Takanishi, R., J. Gen. Microbiol., 31,211-217, (1963)), способным к пропагации на клетках В.subtilis и В.amyloliquefaciens. После трансдукции были получены штаммы В.amyloliquefaciens G1136A (pMWMX1) и В.amyloliquefaciens G1136A(pYDHL6).
Каждый из указанных штаммов выращивали при 37°С в течение 18 часов в L-бульоне, содержащем 10 мг/л эритромицина. Затем 0.3 мл полученной культуры переносили в 3 мл ферментационной среды, содержащей 10 мг/л эритромицина, в 20×200 мм пробирках, и выращивали при 37°С в течение 72 часов на роторной качалке.
Состав ферментационной среды, (г/л):
После выращивания количество накопленного в среде инозина определяли с помощью ВЭЖХ, как описано выше. Результаты представлены в Таблице 6.
Как видно из Таблицы 6, усиление экспрессии гена ydhLBa, вызванное наличием мутации, повышает продукцию инозина штаммом В.amyloliquefaciens G1136A.
Хотя указанное изобретение описано в деталях со ссылкой на конкретные Примеры, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
Каждый из упомянутых выше документов включен в описание в качестве ссылки во всей полноте.

Изобретение относится к биотехнологии. Предложен способ получения пуриновых нуклеозидов и пуриновых нуклеотидов, в частности инозина и 5'-инозиновой кислоты. Способ предусматривает использование бактерии, принадлежащей к роду Bacillus или к роду Escherichia. Продукция пуриновых нуклеозидов и нуклеотидов указанной бактерии повышена за счет повышения активности белка YdhL. Также предоставляется аминокислотная последовательность белка YdhL из Bacillus amyloliquefaciens и кодирующий его ген. Изобретение может быть использовано в промышленном производстве нуклеозидов и нуклеотидов. 7 н. и 18 з.п. ф-лы, 2 ил., 6 табл.
(А) белка, характеризующегося последовательностью аминокислот, приведенной в списке последовательностей под номером 2 (SEQ ID NO:2); и
(Б) белка, характеризующегося последовательностью аминокислот, приведенной в списке последовательностей под номером 2 (SEQ ID NO:2), содержащей делеции, замены, вставки или добавление одной или нескольких аминокислот в указанной последовательности аминокислот, не приводящие к потере активности указанного белка.
(а) фрагмента ДНК, характеризующегося нуклеотидной последовательностью, приведенной в списке последовательностей под номером 1 (SEQ ID NO:1); и
(б) фрагмента ДНК, гибридизующегося в жестких условиях с последовательностью нуклеотидов, приведенной в списке последовательностей под номером 1 (SEQ ID NO:1), или с зондом, приготовленным на основании указанной последовательности нуклеотидов, где жесткими являются условия, при которых отмывка осуществляется при 60°С, концентрация соли соответствует 1×SSC и 0,1% SDS, а продолжительность отмывки составляет 15 мин.
(в) фрагмента ДНК по п.3,
(г) фрагмента ДНК, характеризующегося нуклеотидной последовательностью, приведенной в списке последовательностей под номером 3 (SEQ ID NO:3); и
(д) фрагмента ДНК, являющегося ортологом любого из фрагментов ДНК, указанных в (в) или (г).
(в) фрагмента ДНК по п.3,
(г) фрагмента ДНК, характеризующегося нуклеотидной последовательностью, приведенной в списке последовательностей под номером 3 (SEQ ID NO:3); и
(д) фрагмента ДНК, являющегося ортологом любого из фрагментов ДНК, указанных в (в) или (г).
(а) выращивание бактерии по п.4 или 9 в питательной среде и
(б) сбор пуринового нуклеозида из культуральной жидкости.
(а) выращивание бактерии по п.4 или 9 в питательной среде и выделение полученного пуринового нуклеозида,
(б) фосфорилирование полученного пуринового нуклеозида и
(в) сбор пуринового нуклеотида.
(а) выращивание бактерии по п.4 или 9 в питательной среде и выделение полученного ксантозина,
(б) фосфорилирование ксантозина,
(в) аминирование 5'-ксантиловой кислоты и
(г) сбор 5'-гуаниловой кислоты.
| WO 9903988, 28.01.1999 | |||
| Способ чистовой обработки шаров | 1982 |
|
SU1060428A1 |
| Б.ГЛИК, ДЖ.ПАСТЕРНАК «Молекулярная биотехнология», М: «Мир», 2002, с.168-169. | |||

