Область техники, к которой относится изобретение
Настоящее изобретение относится к новому ферменту, на основе которого может быть легко получен пептид с высоким выходом и недорого, без потребностей в сложном способе синтеза. Более конкретно настоящее изобретение относится к новому ферменту, который катализирует реакцию образования пептида из карбоксильного компонента и компонента с аминогруппой, к микробу, который продуцирует данный фермент, и к способу получения дипептида с использованием указанного фермента или микроба.
Предпосылки создания изобретения
Пептиды используются при производстве фармацевтических препаратов, в пищевой промышленности и в различных других областях. В частности, например, L-аланил-L-глутамин в связи с тем, что он обладает большей стабильностью и водорастворимостью, чем L-глутамин, широко используется в качестве компонента жидкостей, применяемых для инфузии, и в составе бессывороточных сред.
Способы химического синтеза, которые известны как способы образования пептидов, не всегда являются легко выполнимыми. Известные примеры таких способов включают способ, в котором используется N-бензилоксикарбонилаланин (далее в описании называемый как "Z-аланин") и защищенный L-глутамин (см. Bull. Chem. Soc. Jpn., 34, 739 (1961), Bull. Chem. Soc. Jpn., 35, 1966 (1962)), способ, в котором используется Z-аланин и защищенный сложный γ-метиловый эфир L-глутаминовой кислоты (см. Bull. Chem. Soc. Jpn., 37, 200 (1964)), способ, в котором используется сложный эфир Z-аланина и незащищенная глутаминовая кислота (см. Патентный документ 1), способ, который включает синтез производного N-(2-замещенного)пропионилглутамина в качестве промежуточного продукта из 2-замещенного пропионилгалогенида в качестве исходного материала (см. Патентный документ 2).
Однако, поскольку все указанные способы требуют введения, а затем удаления защитных групп или использования оптически активных промежуточных продуктов, они не являются достаточно удовлетворительными с точки зрения их промышленной применимости.
С другой стороны, широко известны примеры обычных способов производства пептидов с использованием ферментов, которые включают проведение реакции конденсации и в которых используют N-защищенный и C-незащищенный карбоксильный компонент, а также N-незащищенный, C-защищенный компонент с аминогруппой, или аминокомпонент (способ, обозначаемый далее как "реакция 1"), и реакцию замещения, в которой используют N-защищенный и C-защищенный карбоксильный компонент и N-незащищенный, C-защищенный аминокомпонент (способ, обозначаемый далее как "реакция 2"). Примером реакции 1 является способ получения сложного метилового эфира Z-аспартилфенилаланина из Z-аспарагиновой кислоты и сложного метилового эфира фенилаланина (см. Патентный документ 3), а примером реакции 2 является способ получения амида ацетилфенилаланиллейцина из сложного этилового эфира ацетилфенилаланина и амида лейцина (см. Biochemical J., 163, 531, (1977)). Имеются сообщения, относящиеся к примерам очень небольшого числа исследований, направленных на поиски способа, в котором использовался бы N-незащищенный, С-защищенный карбоксильный компонент. Пример реакции замещения, в которой используется N-незащищенный, С-защищенный карбоксильный компонент и N-незащищенный, С-защищенный аминокомпонент (способ, обозначаемый далее как "реакция 3"), описан в публикации по Международному патенту WO 90/01555 (Патентный документ 4). В частности, в этой связи можно указать на способ получения амида аргиниллейцина из сложного этилового эфира аргинина и амида лейцина. Примеры реакций замещения, в которых используют N-незащищенный, С-защищенный карбоксильный компонент и N-незащищенный, С-незащищенный аминокомпонент (способ, обозначаемый далее как "реакция 4") описаны в публикации по Европейскому патенту EP 278787A1 (Патентный документ 5) и в публикации по Европейскому патенту EP 359399В1 (Патентный документ 6). В частности, может быть указан способ получения тирозилаланина из сложного этилового эфира тирозина и аланина.
Патентный документ 1: Публикация выложенной заявки на патент Японии, No. Н1-96194 Gazette.
Патентный документ 2: Публикация выложенной заявки на патент Японии, No. Н6-234715 Gazette.
Патентный документ 3: Публикация выложенной заявки на патент Японии, No. S53-92729 Gazette.
Патентный документ 4: Публикация по международному патенту WO 90/01555 Gazette.
Патентный документ 5: Публикация по международному патенту EP 278787A1 Gazette.
Патентный документ 6: Публикация по международному патенту EP 359399B1 Gazette.
Документ 1, не относящийся к патентным публикациям: Biochemical J., 163, 531 (1977).
Описание изобретения
Самый недорогой способ получения среди указанных способов применения реакций 1-4 относится к классу реакции 4, которая включает наименьшее число защитных групп.
Однако вариант осуществления реакции 4 в уровне техники (см. публикацию по Европейскому патенту EP 278787A1) характеризуется следующими основными проблемами: (1) чересчур низкая скорость образования пептида, (2) низкий выход пептида, (3) получаемые пептиды ограничены теми пептидами, которые содержат аминокислоты со сравнительно высокой гидрофобностью, (4) количество добавляемого фермента чрезвычайно велико и (5) требуются относительно дорогие препараты карбоксипептидазы, получаемые из плесневых грибов, дрожжей или растений. В том что касается реакции 4, неизвестны способы, которые бы включали использование фермента, полученного из микробов или дрожжей, отличных от представителей рода Saccharomyces, и неизвестен способ получения аланилглутамина и других пептидов, которые бы характеризовались высокой гидрофильностью. В свете уровня техники в данной области, очевидна потребность в разработке подходящего для промышленного применения недорогого способа получения указанных пептидов.
Целью настоящего изобретения является разработка нового фермента, который позволял бы получать пептид легко, с высоким выходом и недорого, без применения сложного способа синтеза. Более конкретно, целью настоящего изобретения является обеспечение нового фермента, который катализирует реакцию образования пептида из карбоксильного компонента и компонента, содержащего аминогруппу (аминокомпонента), микроба, который продуцирует данный фермент, и способ получения недорогого пептида с использованием указанного фермента или микроба.
В результате проведенных обширных исследований в контексте указанной выше цели авторы настоящего изобретения обнаружили новый фермент, который эффективно образует пептид и который имеется в новых обнаруженных ими бактериях, принадлежащих к роду Empedobacter и т.п., при этом авторы определили последовательность гена, кодирующего данный фермент, что и составило суть настоящего изобретения.
А именно, настоящее изобретение описано ниже.
[1] ДНК, кодирующая белок, указанный в пунктах (А) или (В) ниже:
(А) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 23-616, в аминокислотной последовательности, описанной в виде SEQ ID NO:6 в перечне последовательностей,
(В) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 23-616 в аминокислотной последовательности, описанной в виде SEQ ID NO:6 в перечне последовательностей, и обладающий активностью по образованию пептида.
[2] ДНК, кодирующая белок, указанный в пунктах (С) или (D) ниже:
(С) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 21-619 в аминокислотной последовательности, описанной в виде SEQ ID NO:12 в перечне последовательностей,
(D) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 21-619 в аминокислотной последовательности, описанной в виде SEQ ID NO:12 в перечне последовательностей, и обладающий активностью по образованию пептида.
[3] ДНК, кодирующая белок, указанный в пунктах (E) или (F) ниже:
(E) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 23-625 в аминокислотной последовательности, описанной в виде SEQ ID NO:18 в перечне последовательностей,
(F) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 23-625 в аминокислотной последовательности, описанной в виде SEQ ID NO:18 в перечне последовательностей, и обладающий активностью по образованию пептида.
[4] ДНК, кодирующая белок, указанный в пунктах (G) или (H) ниже:
(G) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 23-645 в аминокислотной последовательности, описанной в виде SEQ ID NO:23 в перечне последовательностей,
(H) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 23-645 в аминокислотной последовательности, описанной в виде SEQ ID NO:23 в перечне последовательностей, и обладающий активностью по образованию пептида.
[5] ДНК, кодирующая белок, указанный в пунктах (I) или (J) ниже:
(I) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 26-620 в аминокислотной последовательности, описанной в виде SEQ ID NO:25 в перечне последовательностей,
(J) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 26-620 в аминокислотной последовательности, описанной в виде SEQ ID NO:25 в перечне последовательностей, и обладающий активностью по образованию пептида.
[6] ДНК, кодирующая белок, указанный в пунктах (К) или (L) ниже:
(К) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 18-644 в аминокислотной последовательности, описанной в виде SEQ ID NO:27 в перечне последовательностей,
(L) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 18-644 в аминокислотной последовательности, описанной в SEQ ID NO:27 в перечне последовательностей, и обладающий активностью по образованию пептида.
[7] ДНК, кодирующая белок, указанный в пунктах (M) или (N) ниже:
(М) белок, обладающий аминокислотной последовательностью, описанной в SEQ ID NO:6 в перечне последовательностей,
(N) белок, содержащий участок зрелого белка, который имеет аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:6 в перечне последовательностей, и который обладает активностью по образованию пептида.
[8] ДНК, кодирующая белок, указанный в пунктах (O) или (P) ниже:
(O) белок, обладающий аминокислотной последовательностью, описанной в SEQ ID NO:12 в перечне последовательностей,
(P) белок, содержащий участок зрелого белка, который имеет аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:12 в перечне последовательностей, и который обладает активностью по образованию пептида.
[9] ДНК, кодирующая белок, указанный в пунктах (Q) или (R) ниже:
(Q) белок, обладающий аминокислотной последовательностью, описанной в SEQ ID NO:18 в перечне последовательностей,
(R) белок, содержащий участок зрелого белка, который имеет аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:18 в перечне последовательностей, и который обладает активностью по образованию пептида.
[10] ДНК, кодирующая белок, указанный в пунктах (S) или (T) ниже:
(S) белок, обладающий аминокислотной последовательностью, описанной в SEQ ID NO:23 в перечне последовательностей,
(T) белок, содержащий участок зрелого белка, который имеет аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:23 в перечне последовательностей, и который обладает активностью по образованию пептида.
[11] ДНК, кодирующая белок, указанный в пунктах (U) или (V) ниже:
(U) белок, обладающий аминокислотной последовательностью, описанной в SEQ ID NO:25 в перечне последовательностей,
(V) белок, содержащий участок зрелого белка, который имеет аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:25 в перечне последовательностей, и который обладает активностью по образованию пептида.
[12] ДНК, кодирующая белок, указанный в пунктах (W) или (X) ниже:
(W) белок, обладающий аминокислотной последовательностью, описанной в SEQ ID NO:27 в перечне последовательностей,
(X) белок, содержащий участок зрелого белка, который имеет аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:27 в перечне последовательностей, и который обладает активностью по образованию пептида.
[13] ДНК, указанная в пунктах (a) или (b) ниже:
(a) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 127-1908 в последовательности оснований, описанной в SEQ ID NO:5 в перечне последовательностей,
(b) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 127-1908 в последовательности оснований, описанных в SEQ ID NO:5 в перечне последовательностей, и которая кодирует белок, обладающий активностью по образованию пептида.
[14] ДНК, указанная в пунктах (c) или (d) ниже:
(с) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 121-1917 в последовательности оснований, описанной в SEQ ID NO:11 в перечне последовательностей,
(d) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 121-1917 в последовательности оснований, описанной в SEQ ID NO:11 в перечне последовательностей, и которая кодирует белок, обладающий активностью по образованию пептида.
[15] ДНК, указанная в пунктах (e) или (f) ниже:
(e) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 127-1935 в последовательности оснований, описанной в SEQ ID NO:17 в перечне последовательностей,
(f) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 127-1935 в последовательности оснований, описанной в SEQ ID NO:17 в перечне последовательностей, и которая кодирует белок, обладающей активностью по образованию пептида.
[16] ДНК, указанная в пунктах (g) или (h) ниже:
(g) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 127-1995 в последовательности оснований, описанной в SEQ ID NO:22 в перечне последовательностей,
(h) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 127-1995 в последовательности оснований, описанной в SEQ ID NO:22 в перечне последовательностей, и которая кодирует белок, обладающий активностью по образованию пептида.
[17] ДНК, указанная в пунктах (i) или (j) ниже:
(i) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 104-1888 в последовательности оснований, описанной в SEQ ID NO: 24 в перечне последовательностей,
(j) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 104-1888 в последовательности оснований, описанной в SEQ ID NO:24 в перечне последовательностей, и которая кодирует белок, обладающий активностью по образованию пептида.
[18] ДНК, указанная в пунктах (k) или (l) ниже:
(k) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 112-1992 в последовательности оснований, описанной в SEQ ID NO:26 в перечне последовательностей,
(l) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 112-1992 в последовательности оснований, описанной в SEQ ID NO:26 в перечне последовательностей, и которая кодирует белок, обладающий активностью по образованию пептида.
[19] ДНК, указанная в пунктах (m) или (n) ниже:
(m) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 61-1908 в последовательности оснований, описанной в SEQ ID NO:5 в перечне последовательностей,
(n) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 61-1908 в последовательности оснований, описанной в SEQ ID NO:5 в перечне последовательностей, и которая кодирует белок, содержащий участок зрелого белка, который обладает активностью по образованию пептида.
[20] ДНК, указанная в пунктах (o) или (p) ниже:
(o) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 61-1917 в последовательности оснований, описанной в SEQ ID NO:11 в перечне последовательностей,
(p) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 61-1917 в последовательности оснований, описанной в SEQ ID NO:11 в перечне последовательностей, и которая кодирует белок, содержащий участок зрелого белка, который обладает активностью по образованию пептида.
[21] ДНК, указанная в пунктах (q) или (r) ниже:
(q) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 61-1935 в последовательности оснований, описанной в SEQ ID NO:17 в перечне последовательностей,
(r) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 61-1935 в последовательности оснований, описанной в SEQ ID NO:17 в перечне последовательностей, и которая кодирует белок, содержащий участок зрелого белка, который обладает активностью по образованию пептида.
[22] ДНК, указанная в пунктах (s) или (t) ниже:
(s) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 127-1995 в последовательности оснований, описанной в SEQ ID NO:22 в перечне последовательностей,
(t) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 127-1995 в последовательности оснований, описанной в SEQ ID NO:22 в перечне последовательностей, и которая кодирует белок, содержащий участок зрелого белка, который обладает активностью по образованию пептида.
[23] ДНК, указанная в пунктах (u) или (v) ниже:
(u) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 29-1888 в последовательности оснований, описанной в SEQ ID NO:24 в перечне последовательностей,
(v) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 29-1888 в последовательности оснований, описанной в SEQ ID NO:24 в перечне последовательностей, и которая кодирует белок, содержащий участок зрелого белка, который обладает активностью по образованию пептида.
[24] ДНК, указанная в пунктах (w) или (x) ниже:
(w) ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 61-1992 в последовательности оснований, описанной в SEQ ID NO:26 в перечне последовательностей,
(x) ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 61-1992 в последовательности оснований, описанной в SEQ ID NO:26 в перечне последовательностей, и которая кодирует белок, содержащий участок зрелого белка, который обладает активностью по образованию пептида.
[25] ДНК по любому из пунктов [13]-[24], где строгие условия включают такие условия, в которых промывка проводится при температуре 60°C с использованием концентрации соли, эквивалентной 1 × SSC и 0,1% ДСН.
[26] Рекомбинантная ДНК, включающая ДНК по любому из пунктов [1]-[25].
[27] Трансформированная клетка, включающая введенную в нее рекомбинантную ДНК, описанную в пункте [26].
[28] Способ получения пептидобразующего фермента, включающий:
культивирование трансформированной клетки по пункту [27] в среде и создание условий, способствующих накоплению пептидобразующего фермента в среде и/или в трансформированной клетке.
[29] Способ получения дипептида, включающий: культивирование трансформированной клетки по пункту [28] в среде с получением культуры и последующее смешивание такой культуры с карбоксильным компонентом и с аминокомпонентом для синтеза дипептида.
[30] Способ получения дипептида, включающий: получение дипептида из компонента с карбоксигруппой и компонента с аминогруппой, с использованием культуры микроба, принадлежащего к роду Sphingobacterium и обладающего способностью образовывать дипептид из компонента с карбоксигруппой и компонента с аминогруппой, с последующим отделением микробной клетки от культуры, продукта обработанной микробной клетки данного микроба или пептидобразующего фермента, полученного из данного микроба.
Кроме того, аминокислотная последовательность, описанная в SEQ ID NO:6, определяется природой ДНК, описанной в SEQ ID NO:5 в перечне последовательностей. Аминокислотная последовательность, описанная в SEQ ID NO:12, определяется природой ДНК, описанной в SEQ ID NO:11. Аминокислотная последовательность, описанная в SEQ ID NO:18, определяется природой ДНК, описанной в SEQ ID NO:17. Аминокислотная последовательность, описанная в SEQ ID NO:23, определяется природой ДНК, описанной в SEQ ID NO:22. Аминокислотная последовательность, описанная в SEQ ID NO:25, определяется природой ДНК, описанной в SEQ ID NO:24. Аминокислотная последовательность, описанная в SEQ ID NO:27, определяется природой ДНК, описанной в SEQ ID NO:26.
Краткое описание фигур
Фиг.1 представляет собой график, иллюстрирующий оптимальный pH для фермента из Empedobacter по настоящему изобретению.
Фиг.2 представляет собой график, иллюстрирующий оптимальную температуру для фермента из Empedobacter по настоящему изобретению.
Фиг.3 представляет собой график, иллюстрирующий образование с течением времени L-аланил-L-глутамина из сложного метилового эфира L-аланина и L-глутамина.
Фиг.4 представляет собой диаграмму, иллюстрирующую количество фермента, присутствующего во фракции цитоплазмы (Cy) и во фракции периплазмы (Pe).
Наилучший способ осуществления изобретения
Ниже приводится описание гена для нового фермента, образующего дипептид, и дипептидобразующего фермента, который является продуктом данного гена.
(1) Микробы, содержащие ДНК по настоящему изобретению
ДНК по настоящему изобретению кодирует белок, который обладает способностью образовывать пептид из компонента с карбоксильной группой и компонента с аминогруппой. В настоящем описании компонент с карбоксильной группой относится к компоненту, который обеспечивает карбонильный участок (СО) в пептидной связи (-CONH-), тогда как компонент с аминогруппой относится к компоненту, который обеспечивает аминный участок (NH) в пептидной связи. Кроме того, в настоящем описании, если особо не оговорено иное, термин "пептид" при использовании его без дополнительных определений относится к полимеру, имеющему по меньшей мере одну пептидную связь. Кроме того, в настоящем описании термин "дипептид" относится к пептиду, имеющему одну пептидную связь.
Примеры микробов, содержащих ДНК по настоящему изобретению, включают бактерии, принадлежащие к роду Empedobacter, к роду Sphingobacterium, к роду Pedobacter, к роду Taxeobacter, к роду Cyclobacterium или к роду Psycloserpens, тогда как более конкретные примеры их включают Empedobacter brevis, штамм ATCC 14234 (штамм FERM P-18545, штамм FERM BP-8113), Sphingobacterium sp. штамм FERM BP-8124, Pedobacterheparinus штамм IFO 12017, Taxeobacter gelupurpurascens штамм DSMZ 11116, Cyclobacterium marinum штамм АТСС 25205 и Psycloserpensburtonensis штамм АТСС 700359. Empedobacter brevis штамм АТСС 14234 (штамм FERM P-18545, штамм FERM BP-8113), Sphingobacterium sp. штамм FERM BP-8124, Pedobacterheparinus штамм IFO 12017, Taxeobacter gelupurpurascens штамм DSMZ 11116, Cyclobacterium marinum штамм АТСС 25205 и Psycloserpensburtonensis штамм АТСС 700359) представляют собой бактерии, которые были выбраны в результате поисков авторами настоящего изобретения тех бактерий, которые продуцируют фермент, способный образовывать с высоким выходом пептид на основе компонента с карбоксильной группой и компонента с аминогруппой.
В числе указанных штаммов микроорганизмов, те микробы, которые обозначены номерами с аббревиатурой FERM, депонированы независимой корпорацией, Национальным Институтом современной промышленной науки и технологии (National Institute of Advanced, Industrial Science and Technology), в Международном Депозитарии запатентованных организмов (International Patent Organism Depositary) (Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония) и могут быть найдены по ссылке на данный номер.
Среди указанных штаммов микроорганизмов, те микробы, наименования которых содержат номера АТСС, депонированы в Американской Коллекции типовых культур (American Type Culture Collection (P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) и могут быть найдены по ссылке на данный номер.
Среди указанных штаммов микроорганизмов, те микробы, наименования которых содержат номера IFO, депонированы в Институте ферментации (Institute of Fermentation) в Осаке (2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) и могут быть найдены по ссылке на данный номер.
Среди указанных штаммов микроорганизмов, те микробы, наименования которых содержат номера NBRC, депонированы в Центре Биологических Ресурсов Национального Института технологии и аналитической оценки (NITE Biological Resource Center of the National Institute of Technology and Evaluation) (5-8 Kazusa-Kamaashi, 2-Chome, Kisarazu-shi, Chiba-ken, Япония) и могут быть найдены по ссылке на данный номер.
Среди указанных штаммов микроорганизмов, те микробы, наименования которых содержат номера DSMZ, депонированы в Германском Институте микроорганизмов и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (German Collection of Microbes and Cell Cultures) (Mascheroder Weg, 1b, 38124 Braunschweig, Германия) и могут быть найдены по ссылке на данный номер.
Empedobacter brevis штамм АТСС 14234 (штамм FERM P-18545, штамм FERM BP-8113) был депонирован в Международном Депозитарии запатентованных организмов (International Patent Organism Depositary) независимой корпорацией, Национальным Институтом современной промышленной науки и технологии в Японии (National Institute of Advanced Industrial Science and Technology) (Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония) 1 октября 2001 года и ему был присвоен депозитный номер FERM P-18545. Функция контроля данного организма была перенесена впоследствии в Депозитарий, функционирующий в рамках Будапештского договора при Международном Депозитарии запатентованных организмов (International Patent Organism Depositary) независимой корпорации, Национального Института современной промышленной науки и технологии в Японии (National Institute of Advanced Industrial Science and Technology) 8 июля 2002 года и ему был присвоен депозитный номер FERM BP-8113 (наименование микроорганизма: Empedobacter brevis штамм AJ 13933).
Sphingobacterium sp. штамм AJ110003 был депонирован в Международном Депозитарии запатентованных организмов (International Patent Organism Depositary) независимой административной корпорации, Национального Института современной промышленной науки и технологии (National Institute of Advanced Industrial Science and Technology) 22 июля 2002 года и ему был присвоен депозитный номер FERM BP-8124. Следует отметить, что штамм AJ 110003 (FERM BP-8124) был идентифицирован, как указано выше, как Sphingobacterium sp. по результатам идентифицирующего эксперимента, описанного ниже. Штамм FERM BP-1824 представляет собой Грамотрицательные палочки (размером 0,7-0,8 × 1,5-2,0 мкм), которые не образуют спор и не являются подвижными. Его колонии имеют округлую форму с практически гладкими краями, содержат мало выступов и характеризуются блестящей светло-желтой окраской. Данный микроорганизм растят при температуре 30°C, он является положительным по каталазе, положительным по оксидазе и отрицательным по результатам OF теста (на глюкозу) и на основе указанных свойств был идентифицирован как бактерия, принадлежащая к роду Sphingobacterium. Кроме того, в связи с наличием таких свойств, как отрицательная реакция в отношении восстановления нитрата, отрицательная реакция в отношении образования индола, отрицательная реакция в отношении образования кислоты из глюкозы, отрицательная реакция на аргининдигидролазу, позитивная реакция на уреазу, позитивная реакция в отношении гидролиза эскулина, отрицательная реакция в отношении гидролиза желатина, положительная реакция на β-галактозидазу, положительная реакция в отношении ассимиляции глюкозы, отрицательная реакция в отношении ассимиляции L-арабинозы, положительная реакция в отношении ассимиляции D-маннозы, отрицательная реакция в отношении D-маннита, положительная реакция в отношении ассимиляции N-ацетил-D-глюкозамина, положительная реакция в отношении ассимиляции мальтозы, отрицательная реакция в отношении ассимиляции глюконата калия, отрицательная реакция в отношении ассимиляции н-капроновой кислоты, отрицательная реакция в отношении ассимиляции адипиновой кислоты, отрицательная реакция в отношении ассимиляции ди-яблочной кислоты, отрицательная реакция в отношении ассимиляции цитрата натрия, отрицательная реакция в отношении ассимиляции фенилацетата и положительная реакция на цитохромоксидазу, было показано, что они аналогичны свойствам, характерным для Sphingobacterium multivorum или Sphingobacterium spiritivorum. Кроме того, несмотря на то, что результаты анализа гомологии по последовательности оснований гена 16S рРНК указывают на наивысший уровень гомологии со Sphingobacterium multivorum (98,8%), не было штамма, с которым указанный бактериальный штамм совпадал бы полностью. Соответственно, данный бактериальный штамм был идентифицирован как Sphingobacterium sp.
(2) Культивирование микроорганизмов
Для получения микробных клеток микроорганизмов, включающих ДНК по настоящему изобретению, указанные микробы культивируют и растят в соответствующей среде. При этом отсутствуют какие-то специфические ограничения в отношении среды, используемой для данной цели, главное, чтобы она позволяла бактериям расти. Указанная среда может представлять собой обычную среду, содержащую обычные источники углерода, источники азота, источники фосфора, источники серы, неорганические ионы и органические питательные источники, по необходимости.
Например, может быть использован любой источник углерода, если указанные микробы могут его утилизировать. Конкретные примеры источников углерода, которые могут при этом использоваться, включают сахара, такие как глюкоза, фруктоза, мальтоза и амилоза, спирты, такие как сорбит, этанол и глицерин, органические кислоты, такие как фумаровая кислота, лимонная кислота, уксусная и пропионовая кислота, и их соли, углеводороды, такие как парафин, а также их смеси.
Примеры источников азота, которые могут при этом использоваться, включают аммониевые соли неорганических кислот, такие как сульфат аммония и хлорид аммония, аммониевые соли органических кислот, такие как фумарат аммония и цитрат аммония, нитраты, такие как нитрат натрия и нитрат калия, соединения органического азота, такие как пептоны, дрожжевой экстракт, мясной экстракт и жидкий кукурузный экстракт, а также их смеси.
Кроме того, могут быть соответствующим образом смешаны и далее использованы питательные источники, используемые в обычных средах, такие как неорганические соли, соли микроэлементов (металлов) и витамины.
При этом отсутствуют какие-либо специфические ограничения по условиям культивирования, так что культивирование может проводиться, например, в течение периода времени от примерно 12 до примерно 48 часов, при соответствующем контроле значений pH и температуры, которые должны оставаться в диапазоне значений pH от 5 до 8 и в диапазоне температур от 15°C до 40°C, соответственно, в аэробных условиях.
Очистка фермента
ДНК по настоящему изобретению кодирует пептидобразующий фермент. Указанный пептидобразующий фермент может быть выделен и очищен из бактерий, принадлежащих, например, к роду Empedobacter. Способ выделения и очистки пептидобразующего фермента из Empedobacter brevis поясняется ниже как пример очистки фермента.
Вначале получают экстракт из бактериальных клеток Empedobacter brevis, например, из штамма FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии в Японии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года) путем разрушения клеток с помощью физического способа, такого как ультразвуковое разрушение, или ферментативного способа с использованием фермента, растворяющего клеточную стенку, с последующим удалением нерастворимой фракции центрифугированием и т.п. Далее пептидобразующий фермент может быть подвергнут очистке путем фракционирования раствора экстракта бактериальных клеток, полученного указанным выше способом, комбинированием обычных способов очистки белков, таких как анионообменная хроматография, катионообменная хроматография или гель-фильтрационная хроматография.
Примером носителя, используемого в анионообменной хроматографии, является сефароза Q-Sepharose HP (производство компании Amersham). Фермент восстанавливают из неадсорбируемой фракции в условиях pH 8,5 путем пропускания содержащего фермент клеточного экстракта через колонку, заполненную носителем.
Примером носителя, используемого в катионообменной хроматографии, являются MonoS HR (производство компании Amersham). После адсорбции фермента на колонку пропускают содержащий фермент клеточный экстракт через колонку, заполненную носителем, после чего промывают колонку, тогда как фермент элюируется буферным раствором, обогащенным высокой концентрацией соли. В процессе элюирования концентрация соли может последовательно повышаться или может использоваться концентрационный градиент. Например, в случае использования MonoS HR адсорбированный на колонку фермент элюируют с использованием NaCl в концентрации от примерно 0,2 до примерно 0,5М.
Фермент, очищенный указанным выше способом, может быть затем подвергнут очистке методом гель-фильтрационной хроматографии и т.п. Примером носителя, используемого в качестве геля в фильтрационной хроматографии, является сефадекс (Sephadex 200pg) (производство компании Amersham).
В приведенных выше процедурах очистки фракция, содержащая фермент, может быть подвергнута анализу для подтверждения наличия пептидобразующей активности в каждой из фракций способом, указанным в описанных ниже примерах. Внутренняя аминокислотная последовательность очищенного таким образом фермента показана в виде SEQ ID NO:1 и SEQ ID NO:2 в перечне последовательностей.
(4) ДНК по настоящему изобретению и трансформанты
(4-1) ДНК по настоящему изобретению
ДНК по настоящему изобретению, имеющая последовательность оснований, состоящую из оснований с номерами 61-1908, и описанная в SEQ ID NO:5, была выделена из Empedobacter brevis штамм FERM BP-8113 (Депонирующий институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии в Японии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года). ДНК, состоящая из оснований с номерами 61-1908, описанная в SEQ ID NO:5, представляет собой последовательность кодирующей части (обозначаемая далее как "CDS"). Последовательность оснований, состоящая из оснований с номерами 61-1908, содержит участок сигнальной последовательности и участок зрелого белка. Участок сигнальной последовательности состоит из оснований с номерами 61-126, тогда как участок зрелого белка состоит из оснований с номерами 127-1908. Конкретно, настоящее изобретение относится как к гену белка с активностью пептидобразующего фермента, который содержит сигнальную последовательность, так и к гену белка, имеющего активность пептидобразующего фермента в виде зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, которая описана как SEQ ID NO:5, представляет собой тип лидерной последовательности, и основной функцией лидерного пептида, кодируемого лидерной последовательностью, является, прежде всего, выделение из внутренней части клеточной мембраны за пределы клеточной мембраны. Белок, кодируемый основаниями с номерами 127-1908, а именно, участок, за исключением лидерного пептида, представляет собой зрелый белок и, как считается, он демонстрирует высокий уровень пептидобразующей активности.
ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 61-1917, описанную в SEQ ID NO:11, которая также представляет ДНК по настоящему изобретению, выделяют из Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: Независимая административная корпорация, Национальный Институт современной промышленной науки и технологии в Японии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary, Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года). ДНК, имеющая последовательность оснований, состоящую из оснований с номерами 61-1917, представляет собой последовательность кодирующей части (CDS). Последовательность оснований, состоящая из оснований с номерами 61-1917, содержит участок сигнальной последовательности и участок зрелого белка. Участок сигнальной последовательности представляет собой участок, который состоит из оснований с номерами 61-120, тогда как участок зрелого белка представляет собой участок, который состоит из оснований с номерами 121-1917. Конкретно, настоящее изобретение относится как к гену белка, который содержит сигнальную последовательность, так и к гену белка, имеющего активность пептидобразующего фермента, в виде зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, которая описана в виде SEQ ID NO:11, представляет собой тип лидерной последовательности. Основной функцией лидерного пептида, кодируемого лидерной последовательностью, является, прежде всего, выделение из внутренней части клеточной мембраны за пределы клеточной мембраны. Белок, кодируемый основаниями с номерами 121-1917, а именно, часть его, выделяющая лидерный пептид, представляет собой зрелый белок и, как считается, он демонстрирует очень высокую степень пептидобразующей активности.
ДНК по настоящему изобретению, имеющая последовательность оснований, состоящую из оснований с номерами 61-1935, описанная в SEQ ID NO:17, была выделена из Pedobacter heparinus штамм IFO 12017 (Депонирующий Институт: Институт ферментации в Осаке (Depositary institution: Institute of Fermentation, Osaka), Адрес Депонирующего Института: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония). ДНК, состоящая из оснований с номерами 61-1935, описанная в SEQ ID NO:17, представляет собой часть CDS. Участок сигнальной последовательности и участок зрелого белка содержатся в последовательности оснований, состоящей из оснований с номерами 61-1935. Участок сигнальной последовательности состоит из оснований с номерами 61-126, тогда как участок зрелого белка состоит из оснований с номерами 127-1935. Конкретно, настоящее изобретение относится как к гену белка пептидобразующего фермента, который содержит сигнальную последовательность, так и к гену белка пептидобразующего фермента, который содержит сигнальную последовательность, так и к гену белка пептидобразующего ферментом в виде зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, которая описана в виде SEQ ID NO:17, представляет собой тип лидерной последовательности, а основной функцией лидерного пептида, кодируемого указанной лидерной последовательностью, является, прежде всего, выделение из внутренней части клеточной мембраны наружу, за пределы клеточной мембраны. Белок, кодируемый основаниями с номерами 127-1935, а именно, за исключением участка лидерного пептида, представляет собой зрелый белок и, как полагают, он демонстрирует высокий уровень пептидобразующей активности.
ДНК по настоящему изобретению, имеющая последовательность оснований, состоящую из оснований с номерами 61-1995, описанную в SEQ ID NO:22, была выделена из Taxeobacter gelupurpurascens штамм DSMZ 11116 (Депонирующий Институт: Германский Институт микроорганизмов и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (German Collection of Microbes and Cell Cultures), Адрес Депонирующего Института: (Mascheroder Weg, 1b, 38124 Braunschweig, Германия)). ДНК, состоящая из оснований с номерами 61-1995, описанная в SEQ ID NO:22, представляет собой часть CDS. Участок сигнальной последовательности и участок зрелого белка содержатся в последовательности оснований, состоящей из оснований с номерами 61-1995. Участок сигнальной последовательности состоит из оснований с номерами 61-126, тогда как участок зрелого белка состоит из оснований с номерами 127-1995. Конкретно, настоящее изобретение относится как к гену белка пептидобразующего фермента, который содержит сигнальную последовательность, так и к гену белка пептидобразующего фермента в форме зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, которая описана в виде SEQ ID NO:22, представляет собой тип лидерной последовательности, и основной функцией лидерного пептида, кодируемого указанным участком лидерной последовательности, является, прежде всего, выделение из внутренней части клеточной мембраны наружу, за пределы клеточной мембраны. Белок, кодируемый основаниями с номерами 127-1995, а именно участок, за исключением лидерного пептида, представляет собой зрелый белок и, как полагают, он демонстрирует очень высокий уровень пептидобразующей активности.
ДНК по настоящему изобретению, имеющая последовательность оснований, состоящую из оснований с номерами 29-1888, описанную в SEQ ID NO:24, была выделена из Cyclobacterium marinum штамм АТСС 25205 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки). ДНК, состоящая из оснований с номерами 29-1888, описанная в SEQ ID NO:24, представляет собой часть CDS. Участок сигнальной последовательности и участок зрелого белка содержатся в последовательности оснований, состоящей из оснований с номерами 29-1888. Участок сигнальной последовательности состоит из оснований с номерами 29-103, тогда как участок зрелого белка состоит из оснований с номерами 104-1888. Конкретно, настоящее изобретение относится как к гену белка пептидобразующего фермента, который содержит сигнальную последовательность, так и к гену белка, который содержит пептидобразующий фермент в виде зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, которая описана в виде SEQ ID NO:24, представляет собой тип лидерной последовательности, и основной функцией лидерного пептида, кодируемого указанной лидерной последовательностью, является, прежде всего, выделение из внутренней части клеточной мембраны наружу, за пределы клеточной мембраны. Белок, кодируемый основаниями с номерами 104-1888, а именно участок, за исключением лидерного пептида, представляет собой зрелый белок и, как полагают, он проявляет высокий уровень пептидобразующей активности.
ДНК по настоящему изобретению, имеющая последовательность оснований, состоящую из оснований с номерами 61-1992, описанную в SEQ ID NO:26, была выделена из Psycloserpens burtonensis штамм АТСС 700359 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки). ДНК, состоящая из оснований с номерами 61-1992, описанная в SEQ ID NO:26, представляет собой часть CDS. Участок сигнальной последовательности и участок зрелого белка содержатся в последовательности оснований, состоящей из оснований с номерами 61-1992. Участок сигнальной последовательности состоит из оснований с номерами 61-111, тогда как участок зрелого белка состоит из оснований с номерами 112-1992. Конкретно настоящее изобретение относится как к гену белка пептидобразующего фермента, который содержит сигнальную последовательность, так и к гену белка пептидобразующего фермента в виде зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, которая описана в виде SEQ ID NO:31, представляет собой тип лидерной последовательности, и основной функцией лидерного пептида, кодируемого указанной лидерной последовательностью, является, прежде всего, выделение из внутренней части клеточной мембраны наружу, за пределы клеточной мембраны. Белок, кодируемый основаниями с номерами 112-1992, а именно участок, за исключением лидерного пептида, представляет собой зрелый белок и, как полагают, он демонстрирует высокий уровень пептидобразующей активности.
Кроме того, различные методики работы с рекомбинантными генами, описанные ниже, могут использоваться в соответствии с приведенными в литературе описаниями, такими как в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
ДНК по настоящему изобретению может быть получена путем полимеразной цепной реакции (далее обозначаемой как "ПЦР") (что касается ПЦР, см. White T.J. et al., Trends Genet., 5, 185 (1989)) или путем гибридизации на основе хромосомной ДНК или библиотеки ДНК из Empedobacter brevis, Sphingobacterium sp, Pedobacterheparinus, Taxeobacter gelupurpurascens, Cyclobacterium marinum или Psycloserpensburtonensis. Праймеры для ПЦР могут быть созданы на основе известных внутренних аминокислотных последовательностей, определенных с использованием пептидобразующего фермента, очищенного по методике, указанной в приведенном выше разделе (3). Кроме того, поскольку последовательности оснований гена пептидобразующего фермента (SEQ ID NO:5, SEQ ID NO:11, SEQ ID NO:22, SEQ ID NO:24 и SEQ ID NO:26) были четко определены в настоящем изобретении, праймеры или зонды для гибридизации могут быть разработаны на основании указанных последовательностей оснований, а также указанный ген может быть выделен с использованием зонда. Если в качестве праймеров для ПЦР используют праймеры, имеющие последовательности, соответствующие 5'-нетранслируемому участку и 3'-нетранслируемому участку, полная длина кодирующего участка для настоящего фермента может быть амплифицирована. Например, участок для амплификации, содержащий кодирующий участок и для лидерной последовательности, и для зрелого белка, описан в виде SEQ ID NO:5, в частности, примером 5'-бокового праймера является праймер, имеющий последовательность оснований на участке против хода транскрипции основания с номера 61 в SEQ ID NO:5, тогда как примером 3'-бокового праймера является праймер, имеющий последовательность, комплементарную к последовательности оснований на участке по ходу транскрипции с основания номер 1908.
Праймеры могут быть синтезированы с использованием фосфоамидитного метода (см. Tetrahedron Letters (1981), 22, 1859) с использованием, например, синтезатора ДНК модели 380В (Model 380B DNA Synthesizer), производимого компанией Applied Biosystems, по общеизвестным методикам. ПЦР может быть проведена, например, в соответствии с методикой, приведенной поставщиком, таким как производитель, с использованием системы для проведения ПЦР (Gene Amp PCR System 9600) (производство Perkin-Elmer) и набора для клонирования in vitro (Takara LA PCR in vitro Cloning Kit) (производство Takara Shuzo).
Независимо от того, содержится или нет лидерная последовательность, ДНК, которая по существу идентична ДНК, состоящей из CDS, описанной в SEQ ID NO:5 в перечне последовательностей, также включается в число ДНК по настоящему изобретению. А именно, ДНК, по существу идентичная ДНК по настоящему изобретению, может быть получена путем выделения ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную CDS, описанной в SEQ ID NO:5 в перечне последовательностей, или с использованием зонда, полученного на основе той же последовательности оснований, и которая кодирует белок, обладающий пептидобразующей активностью, на основе ДНК, кодирующих данный фермент, и которые имеют мутацию, или из клеток, обладающих такой ДНК.
Независимо от того, содержится или нет лидерная последовательность, ДНК, по существу идентичная ДНК, состоящей из CDS, описанной в SEQ ID NO:11 в перечне последовательностей, также включается в число ДНК по настоящему изобретению. А именно, ДНК, по существу идентичная ДНК по настоящему изобретению, может быть получена путем выделения ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную CDS, описываемой в SEQ ID NO:11 в перечне последовательностей, или с использованием зонда, полученного на основе той же последовательности оснований, и которая кодирует белок, обладающий пептидобразующей активностью, на основе ДНК, кодирующих данный фермент, и которые имеют мутацию, или из клеток, обладающих такой ДНК.
Независимо от того, содержится или нет лидерная последовательность, ДНК, по существу идентичная ДНК, состоящей из CDS, описанной в SEQ ID NO:17 в перечне последовательностей, также включается в число ДНК по настоящему изобретению. А именно, ДНК, по существу идентичная ДНК по настоящему изобретению, может быть получена путем выделения ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную CDS, описываемой в SEQ ID NO:17 в перечне последовательностей, или с использованием зонда, полученного на основе той же последовательности оснований, и которая кодирует белок, обладающий пептидобразующей активностью, на основе ДНК, кодирующих данный фермент, и которые имеют мутацию, или из клеток, обладающих такой ДНК.
Независимо от того, содержится или нет лидерная последовательность, ДНК, по существу идентичная ДНК, состоящей из CDS, описанной в SEQ ID NO:22 в перечне последовательностей, также включается в число ДНК по настоящему изобретению. А именно, ДНК, по существу идентичная ДНК по настоящему изобретению, может быть получена путем выделения ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную CDS, описываемой в SEQ ID NO:22 в перечне последовательностей, или с использованием зонда, полученного на основе той же последовательности оснований, и которая кодирует белок, обладающий пептидобразующей активностью, на основе ДНК, кодирующих данный фермент, и которые имеют мутацию, или из клеток, обладающих такой ДНК.
Независимо от того, содержится или нет лидерная последовательность, ДНК, по существу идентичная ДНК, состоящей из CDS, описанной в SEQ ID NO:24 в перечне последовательностей, также включается в число ДНК по настоящему изобретению. А именно, ДНК, по существу идентичная ДНК по настоящему изобретению, может быть получена путем выделения ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную CDS, описываемой в SEQ ID NO:24 в перечне последовательностей, или с использованием зонда, полученного на основе той же последовательности оснований, и которая кодирует белок, обладающий пептидобразующей активностью, на основе ДНК, кодирующих данный фермент, и которые имеют мутацию, или из клеток, обладающих такой ДНК.
Независимо от того, содержится или нет лидерная последовательность, ДНК, по существу идентичная ДНК, состоящей из CDS, описанной в SEQ ID NO:26 в перечне последовательностей, также включается в число ДНК по настоящему изобретению. А именно, ДНК, по существу идентичная ДНК по настоящему изобретению, может быть получена путем выделения ДНК, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную CDS, описываемой в SEQ ID NO:26 в перечне последовательностей, или с использованием зонда, полученного на основе той же последовательности оснований, и которая кодирует белок, обладающий пептид-образующей активностью, на основе ДНК, кодирующих данный фермент, и которые имеют мутацию, или из клеток, обладающих такой ДНК.
Зонд может быть получен, например, в соответствии с установленными способами, основанными, например, на известной последовательности оснований, описанной в SEQ ID NO:5 в перечне последовательностей. Кроме того, способ выделения целевой ДНК с использованием зонда для нахождения ДНК, которая гибридизуется с зондом, также может быть осуществлен по известным методикам. Например, зонд ДНК может быть получен путем амплификации последовательности оснований, клонированной в плазмидном или фаговом векторе, расщепления рестрикционным ферментом последовательности оснований, которую предполагается использовать в качестве зонда, с последующей экстракцией желательной последовательности оснований. Участок, подвергаемый расщеплению, может быть откорректирован в зависимости от природы целевой ДНК.
Термин "в строгих условиях" в контексте настоящего описания относится к условиям, в которых образуется так называемый специфический гибрид и не образуется неспецифический гибрид. Трудно точно выразить данные условия в числовых показателях. Например, можно указать на условия, при которых ДНК, обладающие высокой гомологией, например 50% или более, предпочтительно 80% или более, или предпочтительно 90% или более, гибридизуются друг с другом, а ДНК, имеющие более низкий уровень гомологии, чем указанный выше, не гибридизуются друг с другом, или указать обычные условия, используемые для промывки в методе саузерн-гибридизации, при которых гибридизация проводится в условиях 60°C при концентрации соли, соответствующей 60°C, 1 × SSC и 0,1% ДСН, предпочтительно 0,1 × SSC и 0,1% ДСН. Хотя гены, которые гибридизуются в таких условиях, включают также те гены, в которых стоп-кодоны встречаются в определенных положениях последовательностей или которые потеряли активность в связи с мутацией в активном центре, но они могут быть легко удалены путем лигирования их с коммерчески доступным вектором экспрессии, экспрессирующим их в подходящей клетке-хозяине, с последующим анализом ферментативной активности продукта экспрессии описанным ниже способом.
Однако в случае последовательности оснований, которая гибридизуется в строгих условиях, как описано выше, предпочтительно, чтобы белок, кодируемый такой последовательностью оснований, сохранял примерно половину или более, предпочтительно 80% или более, и более предпочтительно 90% или более, от ферментативной активности белка, имеющего аминокислотную последовательность, кодируемую исходной последовательностью оснований, которая служит в качестве базового показателя и которая должна сохраняться в условиях 50°C и pH 8. Например, в том случае, когда говорится о том, что последовательность оснований гибридизуется в строгих условиях с ДНК, которая имеет последовательность оснований, комплементарную к последовательности оснований, состоящей из оснований с номерами 127-1908 в последовательности оснований, описанной в SEQ ID NO:5, предпочтительно, чтобы белок, кодируемый такой последовательностью оснований, сохранял примерно половину или более, предпочтительно 80% или более, и более предпочтительно 90% или более, от ферментативной активности белка, который состоит из аминокислотных остатков с номерами 23-616 в аминокислотной последовательности, описанной в SEQ ID NO:6, в условиях 50°C и pH 8.
Аминокислотная последовательность, кодируемая CDS, описанной в SEQ ID NO:5 в перечне последовательностей, показана в виде SEQ ID NO:6 в перечне последовательностей. Аминокислотная последовательность, кодируемая CDS, описанной в SEQ ID NO:11 в перечне последовательностей, показана в виде SEQ ID NO:12 в перечне последовательностей. Аминокислотная последовательность, кодируемая CDS, описанной в SEQ ID NO:17 в перечне последовательностей, показана в виде SEQ ID NO:18 в перечне последовательностей. Аминокислотная последовательность, кодируемая CDS, описанной в SEQ ID NO:22 в перечне последовательностей, показана в виде SEQ ID NO:23 в перечне последовательностей. Аминокислотная последовательность, кодируемая CDS, описанной в SEQ ID NO:24 в перечне последовательностей, показана в виде SEQ ID NO:25 в перечне последовательностей. Аминокислотная последовательность, кодируемая CDS, описанной в SEQ ID NO:26 в перечне последовательностей, показана в виде SEQ ID NO:27 в перечне последовательностей.
Полная аминокислотная последовательность, описанная в SEQ ID NO:6, содержит лидерный пептид и участок зрелого белка, при этом аминокислотные остатки с номерами 1-22 составляют лидерный пептид, а аминокислотные остатки с номерами 23-616 составляют участок зрелого белка. Кроме того, полная аминокислотная последовательность, описанная в SEQ ID NO:11, включает лидерный пептид и участок зрелого белка, при этом аминокислотные остатки с номерами 1-20 составляют лидерный пептид, а аминокислотные участки с номерами 21-619 составляют участок зрелого белка.
Полная аминокислотная последовательность, описанная в SEQ ID NO:18, содержит лидерный пептид и участок зрелого белка, при этом аминокислотные остатки с номерами 1-22 составляют лидерный пептид, а аминокислотные участки с номерами 23-625 составляют участок зрелого белка.
Полная аминокислотная последовательность, описанная в SEQ ID NO:23, содержит лидерный пептид и участок зрелого белка, при этом аминокислотные остатки с номерами 1-22 составляют лидерный пептид, а аминокислотные участки с номерами 23-645 составляют участок зрелого белка.
Полная аминокислотная последовательность, описанная в SEQ ID NO:25, содержит лидерный пептид и участок зрелого белка, при этом аминокислотные остатки с номерами 1-25 составляют лидерный пептид, а аминокислотные участки с номерами 26-620 составляют участок зрелого белка.
Полная аминокислотная последовательность, описанная в SEQ ID NO:27, содержит лидерный пептид и участок зрелого белка, при этом аминокислотные остатки с номерами 1-17 составляют лидерный пептид, а аминокислотные участки с номерами 18-644 составляют участок зрелого белка.
Белок, кодируемый ДНК по настоящему изобретению, представляет собой белок, в котором зрелый белок обладает пептидобразующей активностью, а ДНК, которая кодирует белок, по существу идентичный белку, обладающему аминокислотной последовательностью, описанной в SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:18, SEQ ID NO:23, SEQ ID NO:25 или SEQ ID NO:27 в перечне последовательностей, независимо от того, содержит он лидерный пептид или нет, также включается в число ДНК по настоящему изобретению. (Следует отметить, что последовательности оснований описаны со ссылкой на аминокислотные последовательности, указываемые в соответствии с общепринятыми кодами универсальных кодонов). А именно, настоящее изобретение относится к ДНК, которые кодируют белки, показанные в пунктах (A)-(X) ниже:
(А) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 23-616 в аминокислотной последовательности, описанной в SEQ ID NO:6 в перечне последовательностей;
(B) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 23-616 в аминокислотной последовательности, описанной в SEQ ID NO:6 в перечне последовательностей, и обладающий пептидобразующей активностью;
(С) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 21-619 в аминокислотной последовательности, описанной в SEQ ID NO:12 в перечне последовательностей;
(D) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 21-619 в аминокислотной последовательности, описанной в SEQ ID NO:12 в перечне последовательностей, и обладающий пептидобразующей активностью;
(E) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 23-625 в аминокислотной последовательности, описанной в SEQ ID NO:18 в перечне последовательностей;
(F) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 23-625 в аминокислотной последовательности, описанной в SEQ ID NO:18 в перечне последовательностей, и обладающий пептидобразующей активностью;
(G) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 23-645 в аминокислотной последовательности, описанной в SEQ ID NO:23 в перечне последовательностей;
(H) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 23-645 в аминокислотной последовательности, описанной в SEQ ID NO:23 в перечне последовательностей, и обладающий пептидобразующей активностью;
(I) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 26-620 в аминокислотной последовательности, описанной в SEQ ID NO:25 в перечне последовательностей;
(J) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 26-620 в аминокислотной последовательности, описанной в SEQ ID NO:25 в перечне последовательностей, и обладающий пептидобразующей активностью;
(K) белок, имеющий аминокислотную последовательность, состоящую из аминокислотных остатков с номерами 18-644 в аминокислотной последовательности, описанной в SEQ ID NO:32 в перечне последовательностей;
(L) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами 18-644 в аминокислотной последовательности, описанной в SEQ ID NO:32 в перечне последовательностей, и обладающий пептидобразующей активностью;
(M) белок, имеющий аминокислотную последовательность, описанную в SEQ ID NO:6 в перечне последовательностей;
(N) белок, содержащий участок зрелого белка, имеющего аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:6 в перечне последовательностей, и обладающий пептидобразующей активностью;
(O) белок, имеющий аминокислотную последовательность, описанную в SEQ ID NO:12 в перечне последовательностей;
(P) белок, содержащий участок зрелого белка, имеющего аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:12 в перечне последовательностей, и обладающий пептидобразующей активностью;
(Q) белок, имеющий аминокислотную последовательность, описанную в SEQ ID NO:18 в перечне последовательностей;
(R) белок, содержащий участок зрелого белка, имеющего аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:18 в перечне последовательностей, и обладающий пептидобразующей активностью;
(S) белок, имеющий аминокислотную последовательность, описанную в SEQ ID NO:23 в перечне последовательностей;
(T) белок, содержащий участок зрелого белка, имеющего аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:23 в перечне последовательностей, и обладающий пептидобразующей активностью;
(U) белок, имеющий аминокислотную последовательность, описанную в SEQ ID NO:25 в перечне последовательностей;
(V) белок, содержащий участок зрелого белка, имеющего аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:25 в перечне последовательностей, и обладающий пептидобразующей активностью;
(W) белок, имеющий аминокислотную последовательность, описанную в SEQ ID NO:27 в перечне последовательностей; и
(X) белок, содержащий участок зрелого белка, имеющего аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:27 в перечне последовательностей, и обладающий пептидобразующей активностью.
В настоящем описании, несмотря на то, что значение термина "множество" варьирует, в зависимости от положения и типа аминокислотных остатков в трехмерной структуре белка, оно не выходит за пределы диапазона тех показателей, которые существенно не влияют на трехмерную структуру и активность белка с определенной аминокислотной последовательностью, и конкретно составляет от 2 до 50, предпочтительно от 2 до 30, и более предпочтительно от 2 до 10. Однако в случае аминокислотных последовательностей, включающих замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности белков (B), (D), (F), (H), (J), (L), (N), (P), ( R), (T), (V) или (X), предпочтительно, чтобы белки сохраняли примерно половину или более, более предпочтительно 80% или более, и еще более предпочтительно 90% или более, от ферментативной активности белков в том состоянии, когда не включается мутация, в условиях с температурой 50°C и pH 8. Например, ситуация может быть пояснена для случая варианта (B): если аминокислотная последовательность (В) включают замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, описанной в SEQ ID NO:6 в перечне последовательностей, предпочтительно, чтобы такой белок сохранял примерно половину или более, более предпочтительно 80% или более, и еще более предпочтительно 90% или более, от ферментативной активности белка, имеющего аминокислотную последовательность, описанную в SEQ ID NO:6 в перечне последовательностей, в условиях с температурой 50°C и pH 8.
Мутацию аминокислот, аналогичную указанной для белка в пункте (В) и т.п., получают путем модификации последовательности оснований таким образом, что аминокислота в конкретном сайте гена данного фермента была замещена, делетирована, встроена или добавлена, например, путем направленного мутагенеза. Кроме того, модифицированная ДНК, описанная выше, может быть также получена путем мутагенной обработки, известной в данной области. Мутагенез включает, например, обработку способом, при котором ДНК, кодирующую настоящий фермент, обрабатывают in vitro гидроксиламином и т.п., а также обработку способом, при котором бактерии, принадлежащие к роду Escherichia, которые содержат ДНК, кодирующую данный фермент, обрабатывают мутагеном, обычно используемым в искусственном мутагенезе, таким как ультрафиолетовое облучение, N-метил-N'-нитро-N-нитрозогуанидин (НТГ) или азотистая кислота.
Кроме того, естественные мутации, такие как различия, определяемые видом и штаммом микроба, также включаются в понятие замещение, делеция, вставка, добавление и/или инверсия основания, указанные выше. При экспрессии ДНК, имеющей такую мутацию, в подходящих клетках и при последующем исследовании ферментативной активности экспрессированного продукта, можно получить ДНК, которая кодирует белок, по существу идентичный белку, описанному SEQ ID NO:6 или 12 в перечне последовательностей.
(4-2) Получение трансформантов и образование пептидобразующих ферментов
Пептидобразующие ферменты могут быть получены при введении ДНК по настоящему изобретению в подходящую клетку-хозяина и при последующей экспрессии ДНК в организме-хозяине.
Примеры организмов-хозяев, которые подходят для экспрессии белка, определяемого ДНК по настоящему изобретению, и которые могут далее использоваться, включают различные прокариотические клетки, такие как бактерии, принадлежащие к роду Escherichia, такие как Escherichia coli, Empedobacter, Sphingobacterium, Flavobacterium и Bacillus, такие как Bacillus subtilis, а также эукариотические клетки, такие как Saccharomyces cerevisiae, Pichia stipitis и Aspergillus oryzae.
Рекомбинантная ДНК, используемая для введения ДНК в клетку-хозяина, может быть получена при встраивании ДНК, подлежащей введению, в вектор, соответствующий типу того организма-хозяина, в котором ДНК должна экспрессироваться, в такой форме, чтобы белок, кодируемый такой ДНК, мог экспрессироваться. В том случае, когда в клетках-хозяевах функционирует промотор, уникальный для гена пептидобразующего фермента из Empedobacter brevis и т.п., такой промотор может использоваться в качестве промотора для экспрессии ДНК по настоящему изобретению. Кроме того, другой промотор, который воздействует на клетки-хозяева, может быть лигирован с ДНК по настоящему изобретению и такая ДНК может экспрессироваться под контролем указанного промотора, при необходимости.
Примеры способов трансформации, применяемых для введения рекомбинантной ДНК в клетки-хозяева, включают способ Моррисона (см. D. M. Morrison, Methods in Enzymology, 68, 326 (1979)) или способ, в соответствии с которым повышают проницаемость клеток для ДНК путем обработки рецепторных микробных клеток хлоридом кальция (см. Mandel, H. and Higa, A., J.Mol. Biol., 53, 159 (1970)).
В случае масштабного производства белка с использованием методов рекомбинантной ДНК предпочтительным способом осуществления настоящего изобретения является также конъюгирование ДНК внутри трансформанта, который продуцирует белок, с образованием тела включения белка. Преимущества такого способа экспрессии продукции включают защиту целевого белка от расщепления протеазами, имеющимися в микробных клетках, а также простота и легкость очистки целевого белка путем разрушения микробных клеток с последующим центрифугированием и т.п.
Тела включения белка, получаемые таким способом, солюбилизируют с использованием средства, денатурирующего белок, и солюбилизированный белок превращают в физиологически активный белок соответствующей складчатой структуры путем обработки его в рамках процедуры регенерации активности, которая состоит в основном в лизировании белка средством, способствующим денатурации белка, с последующим удалением денатурирующего средства. Имеется множество примеров такой обработки, включающих регенерацию активности человеческого интерлейкина-2 (см. публикацию по выложенной заявке на патент Японии No. S61-257931).
Для получения активного белка из тел включения белка необходима серия операций, включающих солюбилизацию и регенерацию активности, и такие процедуры являются более сложными, чем в случае продукции активного белка непосредственно. Однако в случае продуцирования большого объема белка, что оказывает неблагоприятное воздействие на рост микробных клеток, указанный эффект может быть подавлен за счет аккумуляции белков в форме белков включения неактивного белка в микробных клетках.
Примеры способов масштабного производства, используемых для получения больших объемов целевого белка в форме тел включения, охватывают, например, способ, в соответствии с которым целевой белок экспрессируется независимо под контролем мощного промотора, а также способ, в соответствии с которым целевой белок экспрессируется в форме слитого белка с белком, в отношении которого известно, что он экспрессируется в большом объеме.
Далее в описании настоящее изобретение поясняется более конкретно на примере способа образования трансформированной Escherichia coli и использования трансформированных бактерий для получения пептидобразующего фермента. Кроме того, в случае продукции пептидобразующего фермента в микробных клетках, таких как Escherichiacoli, может быть включена ДНК, которая кодирует белок-предшественник, содержащий лидерную последовательность, или может быть включена ДНК, которая состоит только из участка зрелого белка и которая не содержит лидерной последовательности, при этом ДНК может быть соответствующим образом выбрана для того, чтобы получить кодирующую последовательность данного белка, в зависимости от условий продуцирования, формы, вариантов использования и т.п.
Промоторы, которые в норме используются для получения гетерогенных белков в Escherichiacoli, могут использоваться в качестве промоторов для экспрессии ДНК, кодирующей пептидобразующий фермент. Примеры таких промоторов включают Т7 промотор, lac промотор, trp промотор, trc промотор, tac промотор, PR промотор фага лямбда, PL промотор и другие мощные промоторы. Кроме того, примеры векторов, которые могут использоваться, включают pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219 и pMW218. Кроме того, могут использоваться также векторы фаговой ДНК. Вдобавок, могут использоваться векторы экспрессии, которые содержат промоторы и которые способны к экспрессии встроенной последовательности ДНК, включающей промотор.
Для получения пептидобразующего фермента в форме тела включения слитого белка, ген, который кодирует другой белок, и предпочтительно гидрофильный пептид лигируют против хода транскрипции или по ходу транскрипции гена пептидобразующего фермента с получением гена слитого белка. При этом ген, который кодирует другой белок, может быть любым геном, который повышает количество накапливаемого слитого белка и увеличивает растворимость слитого белка после осуществления стадий денатурации и регенерации. Примеры возможных вариантов для таких генов включают ген 10 Т7, ген β-галактозидазы, ген дегидрофолятредуктазы, ген γ-интерферона, ген интерлейкина-2 и ген прохимозина.
При лигировании указанных генов с образованием гена, который кодирует пептидобразующие ферменты, осуществляют лигирование обоих генов таким образом, что их рамки считывания кодонов становятся совместимыми. Гены могут быть лигированы в сайте соответствующего фермента рестрикции или может использоваться синтетическая ДНК, имеющая соответствующую последовательность.
Далее для повышения уровня продукции пептидобразующего фермента, предпочтительно в некоторых случаях, чтобы терминатор, который представляет собой последовательность терминации транскрипции, был лигирован по ходу транскрипции гена белка слияния. Терминатор включает, например, терминатор Т7 и терминатор фага fd, терминатор Т4, терминатор гена резистентности к тетрациклину и терминатор гена trpAиз Escherichiacoli.
В качестве векторов, подходящих для осуществления процедуры введения гена, который кодирует пептидобразующий фермент или слитый белок на основе пептидобразующего фермента и другого белка в Escherichiacoli, предпочтительны так называемые векторы мультикопийного типа, примеры которых включают плазмиду, имеющую ориджин репликации, полученный из ColE1, например, плазмиду на основе pUC и плазмиду на основе pBR322 или их производные. Термин "производные" в контексте настоящего описания относится к тем плазмидам, которые подверглись модификации путем замещения, делеции, вставки, добавления и/или инверсии основания. Следует отметить, что модификация в контексте настоящего описания включает модификации, полученные путем мутагенеза при обработке мутагеном или УФ-облучением, или модификации, достигаемые путем спонтанной мутации.
Для скрининга трансформантов предпочтительно, чтобы векторы имели маркеры, такие как ген резистентности к ампициллину. Такие плазмиды включают коммерчески доступные векторы экспрессии, имеющие мощные промоторы (вектор на основе pUC (производство Takara Shuzo, Co., Ltd), вектор на основе pRROK, (производство Clonetech Laboratories, Inc.), вектор на основе pKK233-2 (производство Clonetech Laboratories, Inc.)) и т.п.
Рекомбинантную ДНК получают путем лигирования фрагмента ДНК с векторной ДНК; во фрагменте ДНК, при этом лигирование осуществляют в следующем порядке: промотор, ген, кодирующий гидролазу амида L-аминокислоты или слитый белок, состоящий из гидролазы амида L-аминокислоты и другого белка, и в зависимости от конкретной ситуации, терминатор.
В случае трансформации E.coli с использованием рекомбинантной ДНК и культивирования полученной E.coli достигается экспрессия и продукция пептидобразующего фермента или слитого белка, состоящего из пептидобразующего фермента и другого белка. Несмотря на то что в качестве трансформируемого организма-хозяина может применяться штамм, который обычно используется для экспрессии гетерогенного гена, предпочтительным является Escherichiacoli штамм JM109. Способы осуществления трансформации и способы скрининга трансформантов описаны в Molecular Cloning, 2nd Edition, Cold Spring Harbor Press (1989) и других публикациях.
В случае экспрессии пептидобразующего фермента в форме белка слияния пептидобразующий фермент может быть расщеплен с использованием рестрикционной протеазы, которая использует последовательность, отсутствующую в пептидобразующем ферменте, такую как последовательность фактора Xa свертывания крови, или калликреина в качестве распознаваемой последовательности.
В качестве среды для продукции может использоваться среда, обычно применяемая для культивирования E. coli, такая как среда М9 с казаминовыми кислотами или LB среда. Кроме того, условия культивирования и условия индукции образования могут быть соответствующим образом выбраны в зависимости от маркера используемого вектора, промотора, типа микробной клетки-хозяина и т.п.
Приведенный ниже способ может использоваться для выделения пептидобразующего фермента или слитого белка, состоящего из пептидобразующего фермента и другого белка. Если пептидобразующий фермент или его белок слияния был солюбилизирован в микробных клетках, вначале выделяют микробные клетки, а затем разрушают или лизируют их, так чтобы использовать продукт такой обработки в качестве неочищенного жидкого препарата фермента. Кроме того, пептидобразующий фермент или его белок слияния могут быть очищены перед использованием с помощью обычных методик, таких как осаждение, фильтрование или хроматография на колонке при необходимости. При этом может быть также использован способ очистки, в котором применяется антитело против пептидобразующего фермента или его белка слияния.
В том случае, когда образуются тела включения данного белка, указанные тела включения солюбилизируют с использованием денатурирующего средства. Несмотря на возможность достичь их солюбилизации вместе с белком микробных клеток, предпочтительно, с точки зрения дальнейших процедур очистки, чтобы тела включения были выделены и солюбилизированы. Для выделения тел включения из микробных клеток могут использоваться общеизвестные методики. Например, тела включения могут быть выделены путем разрушения микробных клеток с последующим центрифугированием. Примеры денатурирующих средств, способных солюбилизировать тела включения, охватывают гидрохлорид гуанидина (например, 6М, pH 5-8) и мочевину (например, 8 М) и т.п.
Белок, который обладает активностью, регенерируют путем удаления указанных денатурирующих средств при проведении диализа или аналогичных процедур. В процедуре диализа может использоваться буферный раствор Трис-HCl, фосфатный буферный раствор или т.п. в качестве диализирующего раствора, концентрация которого может составлять, например, от 20 мМ до 0,5 М, тогда как его pH может иметь значения, например, от 5 до 8.
Концентрация белка на стадии восстановления предпочтительно поддерживается на уровне 500 мкг/мл или ниже. Температура диализа составляет предпочтительно 5°C или ниже для предотвращения самосшивки восстанавливаемого пептидобразующего фермента. Кроме того, способ удаления денатурирующих средств включает разбавление или ультрафильтрацию, в дополнение к диализу, и ожидается, что активность может быть восстановлена независимо от используемого денатурирующего средства.
(5) Свойства фермента, кодируемого ДНК по настоящему изобретению
Активность фермента, кодируемого ДНК по настоящему изобретению, может быть проанализирована, например, при помещении фермента в условия, способствующие реакции в боратном буферном растворе, содержащем в качестве субстратов эфир аминокислоты и амин, с последующим количественным определением образованного пептида. В более конкретном примере реакцию проводят при температуре 25°C в течение нескольких минут с использованием боратного буферного раствора (pH 9,0), содержащего 100 мМ сложного метилового эфира L-аланина и 200 мМ L-глутамина.
Активность в единицах фермента, используемого в процедуре настоящего изобретения, определяют, исходя из того, что 1 единица (Ед) представляет собой количество фермента, которое образует 1 микромоль (1 мкмоль) пептида за 1 минуту в условиях реакции при температуре 25°C с использованием 100 мМ боратного буферного раствора (pH 9,0), содержащего 100 мМ сложного метилового эфира L-аланина и 200 мМ L-глутамина.
Белок, кодируемый ДНК по настоящему изобретению, представляет собой белок пептидобразующего фермента. Пептидобразующая активность относится к активности, которая приводит к образованию пептида из карбоксильного компонента и аминокомпонента.
Далее предпочтительный вариант фермента, кодируемого ДНК по настоящему изобретению, будет пояснен со ссылкой на указанные свойства.
Один предпочтительный вариант фермента, кодируемого ДНК по настоящему изобретению, включает фермент, обладающий способностями, описанными ниже, для анализа которых скорость образования дипептида используют в качестве индикатора. А именно, один предпочтительный вариант фермента, кодируемого ДНК по настоящему изобретению, включает фермент, который обладает способностью образовывать пептид из карбоксильного компонента и аминокомпонента и характеризуется скоростью образования L-аланил-L-глутамина в реакции образования дипептида в условиях, соответствующих приведенным ниже пунктам (i)-(iv), предпочтительно на уровне 0,03 мМ/мин или более, более предпочтительно на уровне 0,3 мМ/мин или более, и особенно предпочтительно на уровне 1,0 мМ/мин или более. Ниже приведены условия проведения реакции образования дипептида:
(i) карбоксильный компонент представляет собой гидрохлорид сложного метилового эфира L-аланина (100 мМ);
(ii) аминокомпонент представляет собой L-глутамин (200 мМ);
(iii) pH составляет 9,0 и
(iv) количество добавляемого гомогенно очищенного фермента составляет менее чем 0,61 мг/мл, по количеству белка.
Указанная выше скорость образования существенно превышает обычную скорость образования при синтезе пептида с использованием фермента, и фермент по настоящему изобретению обладает способностью катализировать пептидный синтез с чрезвычайно высокой скоростью.
Указанное выше количество добавляемого фермента означает конечное количество фермента, который добавляют в реакционную систему, и добавление фермента в количестве 0,01 мг/мл или более и предпочтительно 0,02 мг/мл или более, по количеству белка, является желательным. Термин "количество белка" относится к значению, определяемому колориметрическим методом анализа с использованием красителя кумасси бриллиантового синего, раствора CBB для анализа белка (производство компании Nakarai) и бычьего сывороточного альбумина в качестве стандартного вещества.
В конкретном примере осуществления процедуры анализа ферментативной активности указанная ферментативная активность анализируется в ходе реакции фермента в боратном буферном растворе, содержащем в качестве субстратов эфир аминокислоты и амин, с последующим количественным определением полученного пептида. Более конкретно в качестве примера можно указать на способ, в соответствии с которым фермент вводят в реакцию для взаимодействия в течение нескольких минут при температуре 25°C с использованием 100 мМ боратного буферного раствора (pH 9,0), содержащего 100 мМ сложного метилового эфира L-аланина и 200 мМ L-глутамина.
Кроме того, предпочтительный вариант фермента, кодируемого ДНК по настоящему изобретению, включает фермент, обладающий свойством, благодаря которому и сложный эфир аминокислоты, и амид аминокислоты могут использоваться в качестве субстрата для карбоксильного компонента. Фраза "и сложный эфир аминокислоты, и амид аминокислоты могут использоваться в качестве субстрата" означает, что по меньшей мере один тип или более сложных эфиров аминокислоты и по меньшей один тип или более амидов аминокислоты могут использоваться в качестве субстрата. Кроме того, предпочтительный вариант фермента по настоящему изобретению включает фермент, обладающий свойством, в соответствии с которым аминокислота, С-защищенная аминокислота и амин, все, могут использоваться в качестве субстрата для аминокомпонента. Фраза "аминокислота, С-защищенная аминокислота и амин могут использоваться в качестве субстрата" означает, что по меньшей мере один тип или более аминокислот и по меньшей один тип или более С-защищенных аминокислот и по меньшей мере один тип или более аминов могут использоваться в качестве субстрата. В связи с широким диапазоном субстратной специфичности в отношении карбоксильного компонента или аминокомпонента, фермент по настоящему изобретению является предпочтительным в том смысле, что может быть выбран широкий перечень исходных материалов, что, в свою очередь, может быть благоприятным фактором с точки зрения стоимости и потребностей в производственном оборудовании в случае промышленного производства.
Конкретные примеры карбоксильного компонента включают сложные эфиры L-аминокислоты, сложные эфиры D-аминокислоты, амиды L-аминокислоты и амиды D-аминокислоты. Кроме того, сложные эфиры аминокислот включают не только сложные эфиры аминокислот, соответствующие природным аминокислотам, но также сложные эфиры, соответствующие неприродным аминокислотам или их производным. Более того, примеры сложных эфиров аминокислоты включают сложные эфиры α-аминокислоты, а также сложные эфиры β-, γ- и ω-аминокислоты и т.п., которые имеют различные сайты присоединения аминогруппы. Типичные примеры сложных эфиров аминокислоты включают сложные метиловые эфиры, сложные этиловые эфиры, сложные н-пропиловые эфиры, сложные изопропиловые эфиры, сложные н-бутиловые эфиры, сложные изобутиловые эфиры и сложные трет-бутиловые эфиры аминокислот.
Конкретные примеры аминокомпонента включают L-аминокислоты, С-защищенные L-аминокислоты, D-аминокислоты, С-защищенные D-аминокислоты и амины. Кроме того, примеры аминов включают не только природные амины, но также неприродные амины или их производные. Более того, примеры аминокислот включают не только природные аминокислоты, но также неприродные аминокислоты и их производные. В их число входят α-аминокислоты, а также β-, γ- и ω-аминокислоты и т.п., которые имеют различные сайты присоединения аминогруппы.
Далее, в другом аспекте предпочтительный вариант фермента, кодируемого ДНК по настоящему изобретению, включает фермент, характеризующийся диапазоном pH, в котором он может катализировать реакцию образования пептида, от 6,5 до 10,5. Способность фермента по настоящему изобретению катализировать данную реакцию в таком широком диапазоне pH, как указано выше, является предпочтительной в том смысле, что она дает возможность гибкой адаптации в процессе промышленного производства, которое может подвергаться действию разных ограничивающих факторов. Однако в фактическом процессе производства пептидов предпочтительно использовать фермент при корректировке условий для достижения оптимального значения pH, соответствующего получаемому ферменту, поскольку это позволяет максимально усилить каталитические характеристики фермента.
Кроме того, в другом аспекте один предпочтительный вариант фермента, кодируемого ДНК по настоящему изобретению, включает фермент, для которого характерен температурный диапазон, в котором он способен катализировать реакцию образования пептида, от 0°C до 60°C. Поскольку фермент по настоящему изобретению способен катализировать реакцию в таком широком температурном диапазоне, он предпочтителен в том плане, что позволяет достичь гибкой адаптации в процессе промышленного производства, которое может подвергаться воздействию разных ограничивающих факторов. Однако в действительности, при производстве пептидов предпочтительно использовать фермент при корректировке условий до оптимальной температуры, соответствующей получаемому ферменту, поскольку это позволяет максимально усилить каталитические характеристики фермента.
(6) Способ получения дипептида
Способ получения дипептида по настоящему изобретению включает реакцию между карбоксильным компонентом и аминокомпонентом в присутствии заданного фермента. Способ получения дипептида по настоящему изобретению включает условия, позволяющие ферменту или веществу, содержащему фермент, обладающему способностью образовывать пептид из карбоксильного компонента и аминокомпонента, воздействовать на карбоксильный компонент и аминокомпонент с образованием дипептида в процессе синтеза.
Способ, в соответствии с которым достигается воздействие фермента или вещества, содержащего фермент, по настоящему изобретению на карбоксильный компонент и аминокомпонент, может включать смешивание друг с другом фермента или вещества, содержащего фермент, карбоксильного компонента и аминокомпонента. Более конкретно, может использоваться способ добавления фермента или вещества, содержащего фермент, к раствору, содержащему карбоксильный компонент и аминокомпонент, при создании условий для их взаимодействия. Альтернативно, в случае использования микроба, который продуцирует данный фермент, может использоваться способ, который включает культивирование микроба, образующего данный фермент, продуцирование и накопление фермента в микробной клетке или в культуральном бульоне, с последующим добавлением карбоксильного компонента и аминокомпонента к культуральному бульону. Далее полученный таким образом дипептид может быть собран известными способами и очищен, при необходимости.
Термин "вещество, содержащее фермент" означает любое вещество, которое содержит фермент, и примеры его конкретных форм включают культуру микробов, которые продуцируют фермент, микробные клетки, выделенные из культуры, и продукт, получаемый при обработке микробных клеток (определяемый далее термином "продукт обработанных микробных клеток"). Под фразой "культура микробов" подразумевается результат культивирования микроорганизма, и более конкретно она относится к смеси микробных клеток, среде, используемой для культивирования микроба, и веществам, продуцируемым культивируемым микробом, и т.п. Кроме того, микробные клетки могут быть промыты и далее использованы в виде промытых микробных клеток. Кроме того, продукт обработанных микробных клеток включает продукты разрушенных, лизированных или лиофилизированных микробных клеток и т.п., а также включает неочищенный фермент, восстановленный путем обработки микробных клеток и т.п., а также очищенный фермент, полученный путем очистки необработанного фермента и т.п. Частично очищенный фермент, получаемый при использовании различных методов очистки, может использоваться для получения очищенного фермента, или могут использоваться иммобилизованные ферменты, которые были иммобилизованы методом ковалентного связывания, методом адсорбции, методом захвата молекул и т.п. Кроме того, поскольку некоторые микробы частично лизируются в ходе культивирования, в зависимости от используемых видов микроорганизмов, то в таких случаях в качестве вещества, содержащего фермент, может также использоваться культуральный супернатант.
Дополнительно могут использоваться дикие штаммы в качестве микробов, которые содержат фермент, а также штаммы с рекомбинантным геном, который экспрессирует фермент. Используемые микробы не ограничиваются интактными микробными клетками, а включают обработанные ацетоном микробные клетки, лиофилизированные микробные клетки или микробные клетки, обработанные другими способами. При этом могут также использоваться иммобилизованные микробные клетки и продукт иммобилизованных обработанных микробных клеток, получаемый при иммобилизации микробных клеток или продукта обработанных микробных клеток путем ковалентного связывания, адсорбции, захвата или с использованием других способов, а также обработанные иммобилизованные микробные клетки.
Кроме того, в случае использования культур, культивируемых микробных клеток, промытых микробных клеток или продукта обработанных микробных клеток, который получают путем разрушения или лизирования микробных клеток, нередки случаи наличия фермента, который разрушает образованные пептиды, не участвуя при этом в образовании пептидов. В ряде случаев в такой ситуации может быть предпочтительно добавлять ингибитор металлопротеазы типа этилендиаминтетрауксусной кислоты (ЭДТА). Добавляемое количество может находиться в диапазоне от 0,1 миллимолей (мМ) до 300 мМ и предпочтительно может составлять от 1 мМ до 100 мМ.
Предпочтительный вариант способа получения дипептида по настоящему изобретению представляет собой способ, в соответствии с которым трансформированные клетки, описанные в приведенном выше разделе (4-2), культивируют в среде, позволяя пептидобразующему ферменту накапливаться в среде и/или в трансформированных клетках. Поскольку пептидобразующий фермент может легко продуцироваться в больших объемах при использовании трансформанта, дипептиды могут образовываться быстро и в больших количествах.
Количество используемого фермента или вещества, содержащего фермент, может быть достаточным, если оно представляет собой то количество, при котором выявляется целевой эффект (эффективное количество), при этом эффективное количество может быть легко определено специалистом со средним уровнем знаний в данной области в ходе простого предварительного эксперимента. В случае использования фермента, например, его количество составляет от примерно 0,01 Ед до примерно 100 Ед, тогда как в случае использования промытых микробных бактериальных клеток используемое количество составляет от примерно 1 г/л до примерно 500 г/л.
Может использоваться любой карбоксильный компонент, если он способен образовывать пептид при конденсации с другим субстратом в виде аминокомпонента. Примеры карбоксильного компонента включают сложные эфиры L-аминокислоты, сложные эфиры D-аминокислоты, амиды L-аминокислоты и амиды D-аминокислоты, а также сложные эфиры органических кислот, не содержащие аминогруппу. Кроме того, примеры сложных эфиров аминокислоты включают не только сложные эфиры аминокислот, соответствующие природным аминокислотам, но также сложные эфиры аминокислот, соответствующие неприродным аминокислотам или их производным. Более того, примеры сложных эфиров аминокислоты включают сложные эфиры α-аминокислоты, а также сложные эфиры β-, γ- и ω-аминокислоты и т.п., которые имеют разные сайты присоединения аминогруппы. Типичные примеры сложных эфиров аминокислоты включают сложные метиловые эфиры, сложные этиловые эфиры, сложные н-пропиловые эфиры, сложные изопропиловые эфиры, сложные н-бутиловые эфиры, сложные изобутиловые эфиры и сложные трет-бутиловые эфиры аминокислот.
Может использоваться любой аминокомпонент, если он способен образовывать пептид при конденсации с другим субстратом в виде карбоксильного компонента. Примеры аминокомпонента включают L-аминокислоты, С-защищенные L-аминокислоты, D-аминокислоты, С-защищенные D-аминокислоты и амины. Дополнительно, примеры аминов включают не только природные амины, но и также неприродные амины или их производные. Вдобавок, примеры аминокислот включают не только природные аминокислоты, но также неприродные аминокислоты или их производные. В их число входят α-аминокислоты, а также β-, γ- и ω-аминокислоты и т.п., которые имеют разные сайты присоединения аминогруппы.
Концентрации карбоксильного компонента и аминокомпонента, служащих в качестве исходных материалов, составляют от 1 мМ до 10 мМ и предпочтительно от 0,05 мМ до 2 мМ соответственно; однако имеются случаи, при которых предпочтительно добавлять аминокомпонент в эквимолярном количестве или в избыточном молярном количестве относительно карбоксильного компонента. Кроме того, в ряде случаев, когда высокие концентрации субстратов ингибируют реакцию, они могут добавляться порциями в ходе реакции, после чего проводится корректировка до концентрации, которая не вызывает ингибирования.
Температура реакции, позволяющая осуществлять синтез пептида, составляет от 0°C до 60°C и предпочтительно от 5°C до 40°C. Кроме того, значение pH реакции, позволяющее осуществлять синтез пептида, составляет от 6,5 до 10,5 и предпочтительно от 7,0 до 10,0.
Примеры
Далее настоящее изобретение поясняется примерами. Однако настоящее изобретение не ограничивается приведенными примерами. Кроме того, для подтверждения результатов анализа на тонкослойных хроматограммах с окрашиванием нингидрином (качественная реакция), были проведены количественные определения с использованием высокоэффективной жидкостной хроматографии для анализа продуктов. Используют в процедуре анализа колонку: Inertsil OD-2 (производство GL Science, Inc.), элюат: водный фосфатный раствор, содержащий 5,0 мМ 1-октансульфонат натрия (pH 2,1): метанол = 100:15 до 50, скорость течения 1,0 мл/мин, обнаружение при длине волны 210 нанометров (далее "нм").
(Пример 1) Культивирование микроба (Empedobacter brevis штамм FERM BP-8113)
50 мл среды (pH 6,2), содержащей 5 грамм (г) глюкозы, 5 г сульфата аммония, 1 г монокалия фосфата, 3 г дикалия фосфата, 0,5 г сульфата аммония, 10 г дрожжевого экстракта и 10 г пептона в 1 литре (л), переносят в колбу Сакагучи на 500 мл и стерилизуют при 150°C в течение 15 минут. Затем в среду инокулируют полную петлю клеток Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года) и затем культивируют при 30°C в течение 16 часов в той же самой среде, с последующим культивированием на качалке при 30°C в течение 16 часов со скоростью 120 качаний/мин.
(Пример 2) Получение пептида с использованием микробных клеток
Микробные клетки собирают центрифугированием (10000 оборотов в минуту (об/мин), 15 минут) культурального бульона по процедуре примера 1 и затем суспендируют полученные клетки с достижением концентрации 100 г/л в 100 мМ боратном буфере (pH 9,0), содержащем 10 мМ ЭДТА. После добавления 1 мл суспензии к 1 мл каждого из 100 мМ растворов боратного буфера (pH 9,0), содержащих 10 мМ ЭДТА, 200 мМ приведенных ниже карбоксильных компонентов и 400 мМ приведенных ниже аминокислот до достижения конечного объема 2 мл, проводят реакцию при 18°C в течение 2 часов. Образованные в результате данной реакции пептиды показаны в таблице 1.
компонент
(Пример 3) Очистка фермента
Процедуру после центрифугирования проводят либо на льду, либо при 4°C. Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года) культивируют по процедуре примера 1 и микробные клетки собирают центрифугированием (10000 об/мин, 15 минут). После промывки 16 г микробных клеток с использованием 50 мМ Трис-HCl буфера (pH 8,0) клетки суспендируют в миллилитрах (мл) того же буфера и подвергают ультразвуковой обработке для разрушения в течение 45 минут при мощности 195 ватт. Полученную после ультразвуковой обработки жидкость центрифугируют (10000 об/мин, 30 минут) для удаления клеточного дебриса, получая супернатант обработанной ультразвуком жидкости. Указанный супернатант обработанной ультразвуком жидкости диализуют в течение ночи против 50 мМ Трис-HCl буфера (pH 8,0) с последующим удалением нерастворимой фракции ультрацентрифугированием (50000 об/мин, 30 минут) с получением растворимой фракции в виде супернатантной жидкости. Полученную растворимую фракцию наносят на колонку с сефарозой Q-Sepharose HP (производство Amersham), которая была предварительно уравновешена Трис-HCl буфером (pH 8,0), и активную фракцию собирают из неадсорбированной фракции. Указанную активную фракцию диализуют в течение ночи против 50 мМ ацетатного буфера (pH 4,5) с последующим удалением нерастворимой фракции центрифугированием (10000 об/мин, 30 минут) с получением диализованной фракции в виде супернатантной жидкости. Указанную диализованную фракцию затем наносят на колонку с Mono S (производство Amersham), которая была предварительно уравновешена 50 мМ ацетатным буфером (pH 4,5) и затем элюируют фермент линейным концентрационным градиентом того же буфера, содержащего от 0 до 1 М NaCl. Фракцию, которая характеризуется наименьшим уровнем загрязняющего белка из активных фракций, наносят на колонку Superdex 200pg (производство Amersham), которая была предварительно уравновешена 50 мМ ацетатным буфером (pH 4,5), содержащим 1 М NaCl, и проводят гель-фильтрацию с использованием того же буфера (pH 4,5), содержащего 1 М NaCl, с получением при прохождении через колонку раствора активной фракции. В результате проведения указанных операций было подтверждено, что пептидобразующий фермент, используемый в настоящем изобретении, характеризуется равномерной очисткой, что следует из результатов экспериментального электрофореза. Уровень восстановления фермента в указанном выше процессе очистки составляет 12,2% и степень очистки составляет 707 раз.
(Пример 4) Определение молекулярной массы фермента
(ДСН-гель электрофорез)
Эквивалент 0,3 микрограммов (мкг) фракции очищенного фермента, полученной по методике примера 3, используют для анализа методом электрофореза в полиакриламиде. Для электрофореза используют буферный раствор, содержащий 0,3% (вес/объем) Трис, 1,44% (вес/объем) глицина и 0,1% (вес/объем) лаурилсульфата натрия, в качестве полиакриламидного геля используют гель с концентрационным градиентом от 10 до 20% (Multigel 10-20, производство Daiichi Pure Chemicals), а в качестве маркеров молекулярной массы используют маркеры молекулярной массы, производимые компанией Pharmacia. После завершения электрофореза гель окрашивают красителем кумасси бриллиантовым синим R-250, выявляя в результате однородную полосу в месте, соответствующем молекулярной массе примерно 75 килодальтон (кДа).
(Гель-фильтрация)
Фракцию очищенного фермента, полученную по методике примера 3, наносят на колонку с Superdex 200pq (производство компании Amersham), которая была предварительно уравновешена 50 мМ ацетатным буфером (pH 4,5), содержащим 1 М NaCl, и проводят гель-фильтрацию, пропуская через колонку тот же буфер (pH 4.5), содержащий 1 М NaCl, для определения молекулярной массы. Используют маркеры молекулярной массы компании Pharmacia в качестве стандартных белков, имеющих известную молекулярную массу, для построения калибровочной кривой. В результате определяют молекулярную массу фермента, равную примерно 150 кДа.
На основании результатов анализа методом ДСН-электрофореза и гель-фильтрации определяют, что данный фермент является гомодимером с молекулярной массой примерно 75 кДа.
(Пример 5) Оптимальный pH для ферментативной реакции
Определяют влияние pH на реакцию, в ходе которой образуется L-аланил-L-глутамин из гидрохлорида сложного метилового эфира L-аланина и L-глутамина. В качестве буферов используют ацетатный буфер (pH 3,9-5,4), MES буфер (pH 5,4-6,4), фосфатный буфер (pH 6,0-7,9), боратный буфер (pH 7,8-9,3), CAPS буфер (pH 9,3-10,7) и K2HPO4-NaOH буфер (pH 10,8-11,6). Добавляют 1 микролитр (мкл) фракции фермента, полученной при элюировании с колонки MonoS в процедуре примера 3 (примерно 180 Ед/мл), к 100 мкл каждого из буферов (100 мМ), содержащих по 100 мМ сложного метилового эфира L-аланина, 200 мМ L-глутамина и 10 мМ ЭДТА, и проводят реакцию при 18°C в течение 5 минут с целью определения эффекта pH в реакции. Результаты выражают, принимая за 100% активность в случае использования боратного буфера (pH 9,3), как показано на фиг.1. В итоге определяют, что оптимальный pH составляет от 8 до 9,5.
(Пример 6) Оптимальная температура осуществления ферментативной реакции
Исследуют влияние температуры на реакцию, в ходе которой образуется L-аланил-L-глутамин из гидрохлорида сложного метилового эфира L-аланина и L-глутамина. Добавляют 1 мкл той же фракции фермента, что и в примере 5, к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего 100 мМ сложного метилового эфира L-аланина, 200 мМ L-глутамина и 10 мМ ЭДТА, и проводят реакцию в течение 5 минут при каждой из исследуемых температур для определения влияния температуры на реакцию. Результаты выражают, принимая за 100% активность в случае проведения реакции при температуре 34°C, как показано на фиг.2. В итоге определяют, что оптимальная температура составляет от 30 до 40°C.
(Пример 7) Ингибиторы фермента
Влияние ингибиторов на образование L-аланил-L-глутамина исследуют с использованием в качестве субстратов гидрохлорида сложного метилового эфира L-аланина и L-глутамина. Добавляют 2 мкл той же фракции фермента, что была использована в примере 5, к 50 мкл 100 мМ боратного буфера (pH 9,0), содержащего каждый из исследуемых ингибиторов фермента в концентрации 10 мМ, как показано в таблице 2, и проводят реакцию при температуре 25°C в течение 5 минут. Следует отметить, что о-фенантролин, фенилметилсульфонилфторид и п-нитрофенил-п'-гуанидинбензоат растворяют в метаноле до концентрации 50 мМ перед использованием. Ферментативную активность в указанных условиях выражают в виде относительной активности, принимая за 100 единиц уровень образования L-аланил-L-глутамина в отсутствие ингибитора фермента. В таблице 2 приведены результаты проведенного эксперимента. В итоге было показано, что в числе исследованных ингибиторов серинового фермента указанный фермент не ингибируется под действием фенилметилсульфонилфторида, но ингибируется под действием п-нитрофенил-п'-гуанидинбензоата.
L-Ala-L-Gln
(%)
(Пример 8) Образование L-аланил-L-глутамина из сложного метилового эфира L-аланина и L-глутамина
3 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего 100 мМ гидрохлорида сложного метилового эфира L-аланина, 200 мМ L-глутамина и 10 мМ ЭДТА, и проводят реакцию при 18°C. В результате, как видно из фиг.3, образуется 83 мМ L-аланил-L-глутамина (L-Ala-L-Gln) в случае добавления порции фермента, и концентрация побочного продукта L-Ala-L-Ala-L-Gln составляет 1,3 мМ. С другой стороны, наблюдается очень небольшой уровень образования L-Ala-L-Gln без добавления порции фермента, и концентрация фермента составляет лишь примерно 0,07 мМ после реакции в течение 120 минут.
(Пример 9) Влияние концентрации L-глутамина на уровень образования L-аланил-L-глутамина
1 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего 100 мМ гидрохлорида сложного метилового эфира L-аланина, L-глутамин в концентрациях, показанных в таблице 3, и 10мМ ЭДТА, и проводят реакцию при 18°C в течение 2 часов. Полученные результаты приведены в таблице 3.
(мМ)
(Пример 10) Субстратная специфичность фермента (1)
Специфичность в отношении сложного эфира исследуют в ситуации использования сложного эфира L-аминокислоты в качестве карбоксильного компонента. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего показанные в таблице 4 карбоксильные компоненты в количестве 100 мМ, 200 мМ L-глутамина и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 2 часов. Количества образованного L-Ala-L-Gln в данной реакции показаны в таблице 4 (HCl обозначает гидрохлорид в таблице 4).
L-Ala-L-Gln (мМ)
Сложный этиловый эфир L-аланина·HCl
Сложный изопропиловый эфир L-аланина·HCl
Сложный трет-бутиловый эфир L-аланина·HCl
91,5
78,9
7,5
(Пример 11) Субстратная специфичность фермента (2)
Исследуют уровень продукции пептида в случае использования сложного метилового эфира L-аланина в качестве карбоксильного компонента и использования различных L-аминокислот в качестве аминокомпонента. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего 100 мМ гидрохлорида сложного метилового эфира L-аланина, показанные в таблице 5 L-аминокислоты в количестве 150 мМ и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 3 часов. В таблице 5 показаны количества различных пептидов, образованных в данной реакции. (Знак "+" указывает на те пептиды, в отношении которых был подтвержден факт их образования, но определить количество которых не было возможности из-за отсутствия стандарта, тогда как знак "tr" указывает на наличие следовых количеств).
L-Ala
L-Val
L-Leu
L-Ile
L-Met
L-Phe
L-Trp
L-Ser
L-Thr
L-Ala-L-Ala
L-Ala-L-Val
L-Ala-L-Leu
L-Ala-L-Ile
L-Ala-L-Met
L-Ala-L-Phe
L-Ala-L-Trp
L-Ala-L-Ser
L-Ala-L-Thr
25,4
20,8
45,3
33,9
83,3
74,4
53,9
62,5
53,9
L-Gln
L-Tyr
L-CySH
L-Lys
L-Arg
L-His
L-Asp
L-Glu
L-Pro
L-Ala-L-Gln
L-Ala-L-Tyr
L-Ala-L-CySH
L-Ala-L-Lys
L-Ala-L-Arg
L-Ala-L-His
L-Ala-L-Asp
L-Ala-L-Glu
L-Ala-L-Pro
79,3
17,6
+
71,8
88,0
66,9
2,1
42,9
tr
(Пример 12) Субстратная специфичность фермента (3)
Образование пептида исследуют в случае использования различных типов сложных метиловых эфиров L-аминокислоты в качестве карбоксильного компонента, а также L-глутамина в качестве аминокомпонента. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего показанные в таблице 6 гидрохлоридные соли сложного метилового эфира L-аминокислот (AA-OMe·HCl) в количестве 100 мМ, 150 мМ L-глутамина и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 3 часов. В таблице 6 показаны количества различных пептидов, образованных в данной реакции. (Знак "+" указывает на те пептиды, в отношении которых был подтвержден факт их образования, но определить количество которых не было возможности из-за отсутствия стандарта, тогда как знак "tr" указывает на наличие следовых количеств). Затем к реакционной системе добавляют Твин-80 до конечной концентрации 0,1%, в случае использования L-Trp-OMe и L-Tyr-OMe.
(Пример 13) Субстратная специфичность фермента (4)
Исследуют образование пептида в случае использования сложных метиловых эфиров различных L-аминокислот в качестве карбоксильного компонента и различных L-аминокислот в качестве аминокомпонента. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего показанные в таблице 7 гидрохлоридные соли сложного метилового эфира L-аминокислот (AA-OMe·HCl) в количестве 100 мМ, L-аминокислоты, показанные в таблице 7, по 150 мМ и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 3 часов. В таблице 7 показаны количества различных пептидов, образованных в данной реакции. (Знак "tr" указывает на наличие следовых количеств). Затем к реакционной системе добавляют Твин-80 до конечной концентрации 0,1%, в случае использования L-Trp-OMe. (Знак "+" указывает на те пептиды, в отношении которых был подтвержден факт их образования, но определить их количество не было возможности из-за отсутствия стандарта).
(Пример 14) Субстратная специфичность фермента (5)
Исследуют образование пептида в случае использования L- или D- форм сложных метиловых эфиров различных аминокислот в качестве карбоксильного компонента и L- или D- форм различных аминокислот в качестве аминокомпонента. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего показанные в таблице 8 гидрохлоридные соли сложного метилового эфира различных аминокислот (AA-OMe·HCl) в количестве 100 мМ, показанные в таблице 8 различные аминокислоты в количестве 150 мМ и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 3 часов. Количество различных пептидов, образованных в данной реакции, показано в таблице 8 (Знак "tr" указывает на наличие следовых количеств).
(Пример 15) Субстратная специфичность фермента (6)
Исследуют образование пептида в случае использования различных амидов L-аминокислот в качестве карбоксильного компонента и различных L-аминокислот в качестве аминокомпонента. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего показанные в таблице 9 гидрохлоридные соли амида L-аминокислот (AA-NH2·HCl) в количестве 100 мМ, показанные в таблице 9 L-аминокислоты в количестве 150 мМ и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 3 часов. В таблице 9 показаны количества различных пептидов, образованных в данной реакции.
(Пример 16) Субстратная специфичность фермента (7)
Исследуют образование пептида в случае использования различных сложных метиловых эфиров L-аланина в качестве карбоксильного компонента и С-защищенных L-аминокислот в качестве аминокомпонентов. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего гидрохлоридную соль сложного метилового эфира L-аланина (Ala-OMe·HCl), как показано в таблице 10, в количестве 100 мМ и гидрохлоридные соли амида L-аминокислоты, как показано в таблице 10, в количестве 150 мМ и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 3 часов. В таблице 10 показаны количества различных пептидов, образованных в данной реакции.
(Пример 17) Субстратная специфичность фермента (8)
Исследуют образование пептида в случае использования сложных метиловых эфиров различных аминокислот в качестве карбоксильного компонента и метиламина в качестве аминокомпонента. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего показанные в таблице 11 гидрохлоридные соли сложного метилового эфира аминокислот (АА-OMe·HCl) в количестве 100 мМ, метиламин, как показано в таблице 11, в количестве 150 мМ и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 3 часов. В таблице 11 показаны количества различных пептидов, образованных в данной реакции.
(Пример 18) Субстратная специфичность фермента (9)
Исследуют образование пептида в случае использования сложного эфира β-аминокислоты в качестве карбоксильного компонента или β-аминокислоты в качестве аминокомпонента. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего показанные в таблице 12 карбоксильные компоненты в количестве 100 мМ, аминокомпоненты, показанные в таблице 12, в количестве 150 мМ и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 3 часов. В таблице 12 показаны количества различных пептидов, образованных в данной реакции. (Знак "tr" указывает на наличие следовых количеств).
(Пример 19) Субстратная специфичность фермента (10)
Исследуют образование олигопептида в случае использования сложного эфира L-аминокислот в качестве карбоксильного компонента и пептида в качестве аминокомпонента. 2 мкл той же фракции фермента, что использовалась в примере 5, добавляют к 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего показанные в таблице 13 карбоксильные компоненты в количестве 100 мМ и показанные в таблице 13 аминокомпоненты в количестве 150 мМ и 10 мМ ЭДТА, и проводят реакцию при 25°C в течение 3 часов. В таблице 13 показаны количества различных пептидов, образованных в данной реакции. В результате было четко показано, что настоящий фермент может образовывать не только дипептид, но также длинноцепочечные пептиды при использовании пептида в качестве аминокомпонента.
Как было показано в указанных выше примерах 9-20, данный фермент, полученный из Empedobacter brevis штамм FERM BP-18545, характеризуется чрезвычайно широкой субстратной специфичностью.
(Пример 20) Сравнение способности катализировать образование пептида с известными ферментами
Пептидобразующую способность фермента по настоящему изобретению сравнивают с соответствующей способностью известных ферментов. В качестве известных ферментов берут карбоксипептидазу Y, описанную в ЕР 278787А1, и тиоловые эндопептидазы (фицин, папаин, бромелайн и химопапаин), описанные в ЕР 359399B1, которые используют в виде очищенных ферментных препаратов (производство компании Sigma). Полученный по процедуре примера 3 однородно очищенный фермент используют в качестве источника фермента по настоящему изобретению. Указанные ферменты добавляют в реакционную систему в количестве по белку, указанному в таблице 14. Реакцию осуществляют путем добавления фермента к 100 мкл боратного буфера (pH 9,0), содержащего 100 мМ гидрохлорида сложного метилового эфира L-аланина и 200 мМ L-глутамина, и оставляют полученную смесь для реакции при 25°C. Следует отметить, что использованная карбоксипептидаза была единственным ферментом, который растворяют в 10 мМ ацетатном буфере (pH 5,0), содержащем 1 мМ ЭДТА, тогда как другие используемые ферменты, тиоловые эндопептидазы, были растворены в 10 мМ ацетатном буфере (pH 5,0), содержащем 2 мМ ЭДТА, 0,1 М KCl и 5 мМ дитиотрейтола. Соотношение уровней продукции L-аланил-L-глутамина за счет указанных ферментов приведено в таблице 14.
В результате было показано, что образование чрезвычайно малых следовых количеств L-аланил-L-глутамина происходит даже в отсутствие фермента, причем наблюдается незначительное повышение уровня образования, когда сравнивают образцы с добавлением карбоксипептидазы или тиоловой эндопептидазы с образцом, в который не добавлен фермент. И наоборот, отмечается чрезвычайное возрастание уровня образования L-аланил-L-глутамина при добавлении образца с рассматриваемым ферментом, при этом указанный уровень продукции превосходит примерно в 5000-100000 раз уровень образования при использовании карбоксипептидазы Y и тиоловой эндопептидазы. Как отмечалось выше, фермент по настоящему изобретению характеризуется чрезвычайно высоким уровнем образования пептидов, в отличие от любого известного на достигнутом уровне техники фермента.
Далее, фермент по настоящему изобретению представляет собой димер с молекулярной массой примерно 75000. Напротив, молекулярная масса карбоксипептидазы Y составляет, по имеющимся данным, примерно 61000, а молекулярная масса тиоловой эндопептидазы, также по имеющимся данным, составляет примерно 23000-36000. Таким образом, уровень образования L-аланил-L-глутамина ферментом по настоящему изобретению в сравнении с уровнем его образования под действием карбоксипептидазы Y и тиоловой эндопептидазы будет даже выше, если указанный уровень выражать в расчете на молекулярную массу, чем в случае его расчета на единицу веса, как указано в примерах.
(мг белка/мл)
L-Ala-L-Gln в расчете на единицу веса фермента
(Пример 21) Получение L-аланил-L-глутамина с использованием микробных клеток Sphingobacterium sp.
50 мл среды (pH 7,0), содержащей 5 г глюкозы, 5 г сульфата аммония, 1 г монокалия фосфата, 3 г дикалия фосфата, 0,5 г сульфата магния, 10 г дрожжевого экстракта и 10 пептона в 1 литре (л), переносят в колбу Сакагучи на 500 мл и стерилизуют при 115°C в течение 15 минут для последующего культивирования Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года). Затем в указанную среду инокулируют клетки Sphingobacterium sp. штамм FERM BP-8124 в количестве одной полной петли (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года), культивируют при 30°C в течение 24 часов на агаровой косой среде (агар: 20 г/л, pH 7,0), содержащей 5 г глюкозы, 10 г дрожжевого экстракта, 10 г пептона и 5 г NaCl в 1 л, с последующим культивированием при встряхивании при температуре 30°C в течение 20 часов и со скоростью качания 120 качаний в минуту. Затем 1 мл данного культурального бульона добавляют к указанной выше среде (50 мл/колба Сакагучи на 500 мл) при 30°C в течение 18 часов. После завершения культивирования микробные клетки отделяют от культурального бульона путем центрифугирования и суспендируют в 1 М боратном буфере (pH 9,0), содержащем 10 мМ ЭДТА, до концентрации 100 г/л по сырому весу бактериальных клеток. Затем добавляют 0,1 мл 10 мМ боратного буфера (pH 9,0), содержащего 10 мМ ЭДТА, 200 мМ гидрохлорида сложного метилового эфира L-аланила и 400 мМ L-глутамина, к 0,1 мл суспензии указанных микробных клеток. Полученную среду в количество 0,2 мл оставляют для реакции при температуре 25°C в течение 120 минут. Концентрация образовавшегося к указанному моменту времени L-аланил-L-глутамина составляет 62 мМ.
(Пример 22) Очистка фермента из клеток Sphingobacterium sp.
Описанную далее процедуру после центрифугирования проводят либо на льду, либо при 4°C. Клетки Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес депонирующего института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года) культивируют таким же образом, как и в примере 21, и затем микробные клетки собирают центрифугированием (10000 об/мин, 15 минут). После промывки 2 г микробных клеток 20 мМ буфером Трис-HCl (pH 7,6) полученные клетки суспендируют в 8 мл того же буфера и подвергают ультразвуковому разрушению путем обработки в течение 45 минут при мощности 195 ватт. Жидкость, полученную после ультразвукового разрушения клеток, центрифугируют (10000 об/мин, 30 минут) для удаления клеточного дебриса, получая супернатантную жидкость разрушенных ультразвуком клеток. Указанную супернатантную жидкость разрушенных ультразвуком клеток диализуют в течение ночи против 20 мМ Трис-HCl буфера (pH 7,6) с последующим удалением нерастворимой фракции ультрацентрифугированием (50000 об/мин, 30 минут) с получением растворимой фракции в виде супернатантной жидкости. Полученную растворимую фракцию наносят на колонку с сефарозой Q-Sepharose HP (производство компании Amersham), которая была предварительно уравновешена Трис-HCl буфером (pH 7,6), и собирают активную фракцию из неадсорбированной фракции. Указанную активную фракцию диализуют в течение ночи против 20 мМ ацетатного буфера (pH 5,0) с последующим удалением нерастворимой фракции центрифугированием (10000 об/мин, 30 минут) с получением диализованной фракции в виде супернатантной жидкости. Полученную диализованную фракцию наносят на колонку с сефарозой Q-Sepharose HP (производство компании Amersham), которая была предварительно уравновешена 20 мМ ацетатным буфером (pH 5,0), с получением активной фракции при проведении элюирования фермента линейным концентрационным градиентом того же буфера, содержащего от 0 до 1 М NaCl.
(Пример 23) Получение L-аланил-L-глутамина с использованием фракции фермента
10 мкл фракции, полученной после элюирования с сефарозы Q-Sepharose HP (примерно 27 Ед/мл) и очищенной по процедуре примера 22, добавляют к 90 мкл 111 мМ боратного буфера (pH 9,0), содержащего 111 мМ гидрохлорида сложного метилового эфира L-аланина, 222 мМ L-глутамина и 11 мМ ЭДТА, и проводят реакцию при 25°C в течение 120 минут. В результате реакции получают 73 мМ L-аланил-L-глутамина, образованного при добавлении образца с ферментом. С другой стороны, отмечалась чрезвычайно низкая продукция L-Ala-L-Glu в образце без добавления фермента, и указанное образованное количество составляло лишь примерно 0,07 мМ после проведения реакции в течение 120 минут.
(Пример 24) Субстратная специфичность фермента (11)
Исследуют субстратную специфичность фермента, полученного из клеток Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года). 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего разные карбоксильные компоненты в конечной концентрации 100 мМ и разные аминокомпоненты в конечной концентрации 150 мМ, показанные в таблицах (15-1)-(15-4), фракцию фермента, элюированного с сефарозы Q-Sepharose HP после очистки по процедуре примера 22 (при добавлении 0,33 единиц в реакционную жидкость), и 10 мМ ЭДТА, оставляют для реакции при температуре 25°C в течение 1,5 часов. В таблице 15 показаны количества различных пептидов, образованных в данной реакции. (Знак "+" указывает на те пептиды, для которых был подтвержден факт образования, но определить количество которых было невозможно из-за отсутствия стандарта, тогда как знак "tr" указывает на наличие следовых количеств). Дополнительно, в реакционную систему добавляют Твин-80 до конечной концентрации 0,1%, в случае использования L-Tyr-OMe. Следует отметить, что для всех карбоксильных компонентов используют гидрохлоридные соли.
(Пример 25) Субстратная специфичность фермента (12)
Субстратную специфичность в отношении продукции олигопептида исследуют для фермента, полученного из клеток Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года). Проводят реакцию 100 мкл 100 мМ боратного буфера (pH 9,0), содержащего разные карбоксильные компоненты в конечной концентрации 100 мМ и разные аминокомпоненты в концентрации 150 мМ, показанные в таблице 16, очищенной по методике примера 22 фракции фермента, элюированной с сефарозы SP-Sepharose HP (добавляют 0,33 единицы в реакционную жидкость), и 10 мМ ЭДТА при температуре 25°C в течение 1,5 часов. В таблице 16 показаны количества каждого из олигопептидов, образованных в данной реакции. Для всех карбоксильных компонентов используют гидрохлоридные соли.
(Пример 26) Субстратная специфичность фермента (13)
Дополнительно оценивают субстратную специфичность с использованием той же ферментативной фракции, что и в методике примера 5.
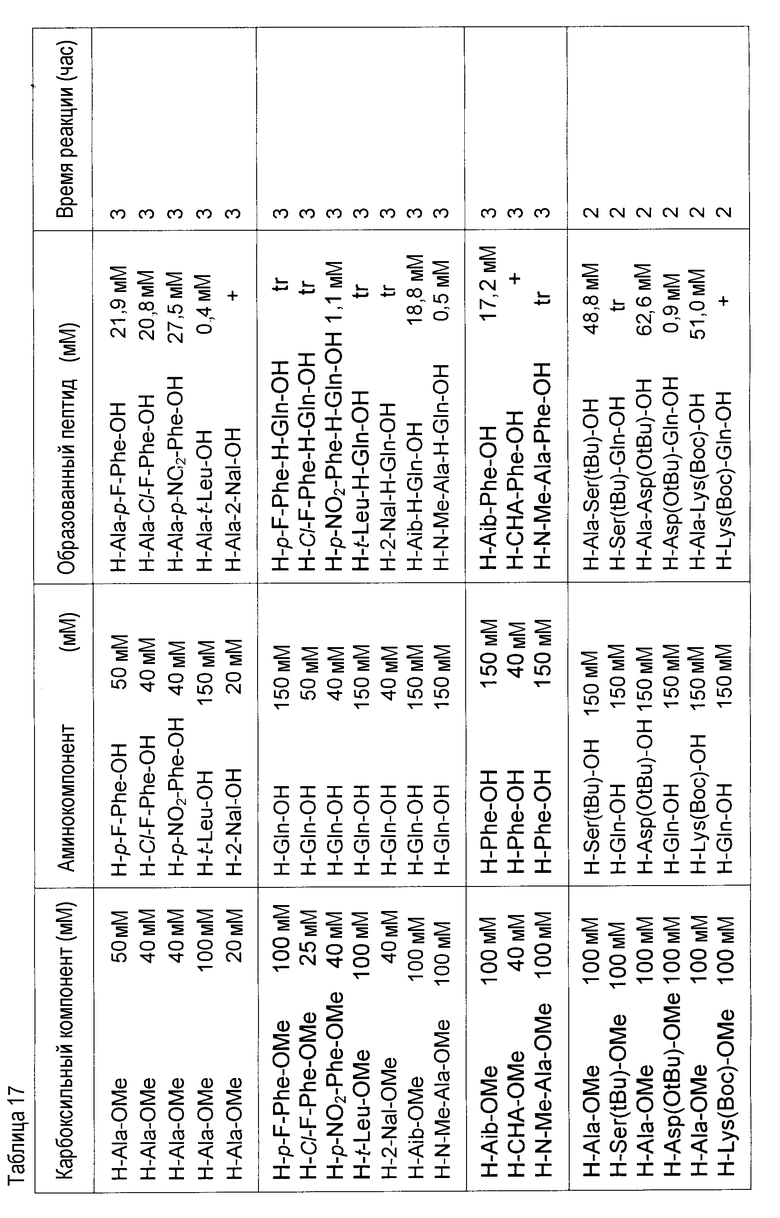
100 мкл реакционных растворов, содержащих 100 мМ боратного буфера (pH 9,0), содержащего каждый из карбоксильных компонентов и аминокомпонентов в конечных концентрациях, приведенных в таблице 17, фермент (добавляют 0,1 единицы в реакционную жидкость) и 10 мМ ЭДТА, объединяют для проведения реакции при температуре 25°C в течение времени реакции различной длительности, как показано в таблице 17. Количества различных пептидов, образованных в данной реакции, показаны в таблице 17. (Знак "+" указывает на те пептиды, для которых был подтвержден факт образования, но определить количество которых было невозможно из-за отсутствия стандарта, тогда как знак "tr" указывает на наличие следовых количеств).
(Сокращения)
H-Ala-OMe: гидрохлорид сложного метилового эфира L-аланина
H-p-F-Phe-OMe: гидрохлорид сложного метилового эфира п-фтор-L-фенилаланина
H-Cl-F-Phe-OMe: гидрохлорид сложного метилового эфира п-хлор-L-фенилаланина
H-p-NO2-Phe-OMe: гидрохлорид сложного метилового эфира п-нитро-L-фенилаланина
H-t-Leu-OMe: гидрохлорид сложного метилового эфира трет-L-лейцина
H-2-Nal-OMe: гидрохлорид сложного метилового эфира 3-(2-нафтил)-L-аланина
H-Aib-OMe: гидрохлорид сложного метилового эфира α-аминомасляной кислоты
H-N-Me-Ala-OMe: гидрохлорид сложного метилового эфира N-метил-L-аланина
H-CHA-OMe: гидрохлорид сложного метилового эфира β-циклогексил-L-аланина
H-Ser-(tBu)-OMe: гидрохлорид сложного метилового эфира О-трет-бутил-L-серина
H-Asp(OtBu)-OMe: гидрохлорид сложного α-метилового эфира сложного β-трет-бутилового эфира L-аспарагиновой кислоты
H-Lys(Boc)-OMe: гидрохлорид сложного метилового эфира N-ε-трет-бутоксикарбонил-L-лизина
H-p-F-Phe-OH: п-фтор-L-фенилаланин
H-Cl-F-Phe-OH: п-хлор-L-фенилаланин
H-p-NO2-Phe-OH: п-нитро-L-фенилаланин
H-t-Leu-OH: трет-L-лейцин
H-2-Nal-OH: 3-(2-нафтил)-L-аланин
H-Gln-OH:: L-глутамин
H-Phe-H: L-фенилаланин
H-Ser(tBbu)-OH: О-трет-бутил-L-серин
H-Asp(OtBu)-OH: сложный β-трет-бутиловый эфир L-аспарагиновой кислоты
H-Lys(Boc)-OH: N-ε-трет-бутоксикарбонил-L-лизин
(Пример 27) Субстратная специфичность фермента (14)
Субстратную специфичность в отношении образования олигопептида оценивают с использованием той же фракции фермента, что и в методике примера 5 (полученной из Empedobacter brevis). 100 мкл реакционных растворов, содержащих 100 мМ боратного буфера (pH 9,0), содержащего каждый из карбоксильных компонентов и аминокомпонентов в конечных концентрациях, приведенных в таблице 18, фермент (количества единиц, добавленных к раствору приведены в таблице 18) и 10 мМ ЭДТА, оставляют для реакции при температуре 25°C в течение 3 часов. В таблице 18 показаны количества различных образованных в реакции олигопептидов. (Знак "+" указывает те пептиды, для которых образование было подтверждено наличием продукции, но в случае которых не представилось возможным определить количество в связи с отсутствием стандарта, тогда как знак "tr" указывает на наличие следовых количеств). Следует отметить, что для всех карбоксильных компонентов использовались гидрохлоридные соли.
(Пример 28) Выделение гена пептидобразующего фермента из Empedobacter brevis
Далее следует описание методики выделения гена пептидобразующего фермента. В качестве микроорганизма используют Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года). При выделении гена в качестве клетки-хозяина используют Escherichia coli JM-109, и в качестве вектора используют pUC118.
(1) Получение праймера для ПЦР на основе определения внутренней аминокислотной последовательности
Получают смешанный праймер, имеющий последовательность оснований, указанную в SEQ ID NO:3 и SEQ ID NO:4 соответственно, на основе аминокислотных последовательностей (SEQ ID NO:1 и SEQ ID NO:2), определенных по методу разложения Эдмана для продукта расщепления лизилэндопептидазой пептидобразующего фермента, полученного из Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года), расщепленного лизилэндопептидазой.
(2) Получение микробных клеток
Клетки Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года) культивируют при 30°C в течение 24 часов на агаровой среде CM2G (содержащей 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну полную петлю полученных бактериальных клеток инокулируют в колбу Сакагучи на 500 мл, содержащую 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара) и далее проводят культивирование при встряхивании при 30°C.
(3) Получение хромосомной ДНК из микробных клеток
Вначале центрифугируют 50 мл культурального бульона (12000 об/мин, 4°C, 15 минут) для сбора микробных клеток. Затем из микробных клеток выделяют хромосомную ДНК с использованием системы QIAGEN Genomic-Tip System (Qiagen) по методике, описанной в руководстве для пользователя.
(4) Получение путем ПЦР фрагмента ДНК, содержащего часть гена пептидобразующего фермента
Фрагмент ДНК, содержащий часть гена пептидобразующего фермента, полученный из Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года), получают методом ПЦР с использованием LA-Taq (производство Takara Shuzo). Затем проводят реакцию ПЦР на основе хромосомной ДНК, полученной из клеток Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года), с использованием праймеров, имеющих последовательности оснований SEQ ID NO:3 и SEQ ID NO:4.
ПЦР проводят в течение 30 циклов с использованием системы термической циклической обработки (Takara PCR Thermal Cycler PERSONAL) (производство Takara Shuzo) в следующих условиях:
94°C в течение 30 секунд
52°C в течение 1 минуты
72°C в течение 1 минуты
После проведения реакции 3 мкл реакционной жидкости вносят в аппарат для проведения электрофореза на 0,8% агарозе. В результате подтверждают, что произошла амплификация фрагмента ДНК размером примерно 1,5 тысячи нуклеотидных пар (т.н. (kb)).
(5) Клонирование гена пептидобразующего фермента из генной библиотеки
Для получения гена пептидобразующего фермента полной длины проводят саузерн-гибридизацию с использованием фрагмента ДНК, амплифицированного по методике ПЦР в качестве зонда. Процедура саузерн-гибридизации описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Фрагмент ДНК размером примерно 1,5 т.н, амплифицированный методом ПЦР, отделяют электрофорезом в 0,8% агарозе. Целевую полосу вырезают и очищают фрагмент ДНК. Фрагмент ДНК метят с использованием зонда дигоксинигена с использованием набора DIG High Prime (производство компании Boehringer-Mannheim) по методике, описанной в руководстве к набору.
После полного расщепления хромосомной ДНК из клеток Empedobacter brevis, полученных на стадии (3) примера 28(3) путем обработки при 37°C в течение 16 часов рестрикционным ферментом HindIII, полученную ДНК подвергают электрофорезу в 0,8% геле агарозы. После электрофореза хромосомную ДНК подвергают блоттингу на положительно загруженном мембранном фильтре из найлона (Nylon) (производство компании Roche Diagnostics) из агарозного геля после электрофореза с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 50°C в течение 1 часа добавляют зонд, меченный дигоксинигеном, полученным по указанной выше методике, и проводят гибридизацию при 50°C в течение 16 часов. Далее фильтр последовательно промывают в течение 20 минут при комнатной температуре 2 × SSC, содержащим 0,1% ДСН. Кроме того, фильтр дополнительно промывают два раза при температуре 65°C в течение 15 минут в 0,1 × SSC.
Выявление полос, которые гибридизовались с зондом, проводят с использованием набора для выявления нуклеотидов (DIG Nucleotide Detection Kit) (производство компании Boehringer-Mannheim) по методике, описанной в инструкции к набору. В результате было показано, что полоса размером примерно 4 т.н. гибридизовалась с данным зондом.
Затем хромосомную ДНК, полученную на стадии (3) данного примера 28(3), полностью расщепляют с использованием HindIII. ДНК размером приблизительно 4 т.н. разделяют электрофорезом в 0,8% геле агарозы и далее проводят очистку ДНК с использованием набора Gene Clean II Kit (производство компании Funakoshi) и растворяют ДНК в 10 мкл ТЭ. Затем 4 мкл указанного продукта смешивают с pUC118 HindIII/BAP (производство компании Takara Shuzo) и проводят реакцию лигирования с использованием набора для лигирования ДНК (DNA Ligation Kit Ver.2) (производство компании Takara Shuzo). 5 мкл смеси для реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM-109 (производство компании Toyobo) смешивают для трансформации клеток Escherichia coli. Полученные таким образом трансформанты затем вносят в соответствующую твердую среду для получения библиотеки хромосомной ДНК.
Для получения гена пептидобразующего фермента полной длины библиотеку хромосомной ДНК подвергают скринингу по методике гибридизации колоний с использованием указанного выше зонда. Процедура гибридизации колоний описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии из библиотеки хромосомной ДНК переносят на мембранный фильтр из найлона (Nylon Membrane for Colony and Plaque Hybridization, производство компании Roche Diagnostics) с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 37°C в течение 1 часа добавляют указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 50°C в течение 16 часов. Далее фильтр последовательно промывают в течение 20 минут при комнатной температуре 2 × SSC, содержащим 0,1% ДСН. Кроме того, фильтр дополнительно промывают два раза при температуре 65°C в течение 15 минут в 0,1 × SSC.
Выявление колоний, которые гибридизовались с меченым зондом, проводят с использованием набора для выявления нуклеотидов (DIG Nucleotide Detection Kit) (производство компании Boehringer-Mannheim) по методике, описанной в инструкции к набору. В результате было подтверждено, что произошла гибридизация двух колоний с меченым зондом.
(6) Последовательность оснований гена пептидобразующего фермента, полученного из Empedobacter brevis
Плазмиды, содержащиеся в клетках Escherichia coli JM-109, получают из указанных двух колоний, для которых был подтвержден факт гибридизации с меченым зондом, с использованием системы очистки Wizard Plus Minipreps DNA Purification System (производство компании Promega) и определяют последовательность оснований в той части, где произошла гибридизация зонда, и рядом с нею. Реакцию секвенирования проводят с использованием набора CEQ DTCS-Quick Start Kit (производство компании Beckman-Coulter) по методике, описанной в инструкции к набору. Кроме того, проводят электрофорез с использованием CEQ 2000-XL (производство компании Beckman-Coulter).
В результате было подтверждено существование открытой рамки считывания, которая кодирует белок, содержащий внутренние аминокислотные последовательности пептидобразующего фермента (SEQ ID NO:1 и 2). Таким образом, было подтверждено, что открытая рамка считывания является геном, кодирующим пептидобразующий фермент. Последовательность оснований генов пептидобразующих ферментов полной длины и соответствующие им аминокислотные последовательности приведены в SEQ ID NO:5. В результате анализа уровня гомологии в отношении полученной рамки считывания по программе BLASTP была обнаружена гомология между двумя ферментами; указанная программа продемонстрировала наличие гомологии на уровне 34% для аминокислотных последовательностей при использовании для сравнения гидролазы сложного эфира α-аминокислоты из Acetobacter pasteurianus (см. Appl. Environ. Microbiol., 68(1), 211-218 (2002)) и гомологии на уровне 26% для аминокислотных последовательностей при использовании для сравнения ацилазы глутарил-7-АСА из Brevibacillus laterosporum (см. J. Bacteriol., 173(24), 7848-7855 (1991)).
(Пример 29) Экспрессия гена пептидобразующего фермента, полученного из Empedobacter brevis, в Escherichia coli
Участок целевого гена в промоторной области trp оперона на хромосомной ДНК Escherichia coli W3110 амплифицируют путем ПЦР с использованием в качестве праймеров олигонуклеотидов, указанных в SEQ ID NO:7 и 8, и полученные фрагменты ДНК лигируют с вектором pGEM-Teasy (производство компании Promega). Затем клетки E.coli JM109 трансформируют указанным лигирующим раствором и отбирают из ампициллин-резистентных штаммов штаммы, имеющие целевую плазмиду, в которой направление введенного trp промотора противоположно ориентации lac промотора. Затем фрагмент ДНК, содержащий trp промотор, полученный путем обработки данной плазмиды с использованием EcoO109I/EcoRI, лигируют с продуктом обработки pUC19 с использованием EcoO109I/EcoRI (производство компании Takara). Затем клетки Escherichia coli JM-109 трансформируют указанным лигирующим раствором и штаммы, несущие целевую плазмиду, отбирают из ампициллин-резистентных штаммов. Далее фрагмент ДНК, полученный при обработке данной плазмиды HindIII/PvuII, лигируют с фрагментом ДНК, содержащим терминатор rrnB, полученный при обработке pKK223-3 (производство компании Amersham Pharmacia) с использованием HindIII/HincII. Затем клетки E.coli JM109 трансформируют указанным лигирующим раствором, штаммы, несущие целевую плазмиду, отбирают из ампициллин-резистентных штаммов и обозначают полученную плазмиду как pTrpT.
Целевой ген амплифицируют путем ПЦР с использованием хромосомной ДНК из клеток Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo No Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года) в качестве матрицы и олигонуклеотиды, указанные в SEQ ID NO:9 и 10, в качестве праймеров. Далее указанный фрагмент ДНК обрабатывают NdeI/PstI и полученный фрагмент ДНК лигируют с продуктом обработки pTrpT при использовании NdeI/PstI. Далее Escherichia coli JM-109 трансформируют данным лигирующим раствором и штаммы, несущие целевую плазмиду, отбирают из ампициллин-резистентных штаммов и обозначают данную плазмиду как pTrpT_Gtg2.
Escherichia coli JM-109, несущую pTrpT_Gtg2, подвергают предварительному культивированию при 30°C в течение 24 часов в среде LB, содержащей 100 мг/л ампициллина. 1 мл полученного культурального бульона инокулируют в колбу Сакагучи на 500 мл, содержащую 50 мл среды (D-глюкоза в количестве 2 г/л, дрожжевой экстракт в количестве 10 г/л, казаминовые кислоты в количестве 10 г/л, сульфат аммония в количестве 5 г/л, дигидрофосфат калия в количестве 3 г/л, дикалий гидрофосфат в количестве 1 г/л, гептагидрат сульфата магния в количестве 0,5 г/л и ампициллин в количестве 100 мг/л) и далее культивируют при 25°C в течение 24 часов. Культуральный бульон обладает активностью по образованию L-аланил-L-глутамина с активностью 0,44 Ед в расчете на 1 мл культурального бульона, и было подтверждено, что клонированный ген экпрессируется в E. coli. Кроме того, не было обнаружено никакой активности у трансформанта, который служил в качестве контроля и в который был введен только pTrpT.
(Прогнозирование сигнальной последовательности)
В случае анализа аминокислотной последовательности SEQ ID NO:6, описанной в перечне последовательностей, с помощью программы Signal P v 1.1 (см. Protein Engineering, Vol. 12, No. 1, pp. 3-9, 1999), было предположено, что участок аминокислотной последовательности от 1 до 22 номера функционирует в качестве сигнала для секреции пептида в периплазму, тогда как зрелый белок считывается по ходу транскрипции, начиная с аминокислоты номер 23.
(Подтверждение секреции)
Клетки Escherichia coli JM-109, несущие pTrpT_Gtg2, подвергают предварительному культивированию при 30°C в течение 24 часов в среде LB, содержащей 100 мг/л ампициллина. 1 мл полученного культурального бульона инокулируют в колбу Сакагучи на 500 мл, содержащую 50 мл среды (глюкоза в количестве 2 г/л, дрожжевой экстракт в количестве 10 г/л, казаминовые кислоты в количестве 10 г/л, сульфат аммония в количестве 5 г/л, дигидрофосфат калия в количестве 3 г/л, дикалий гидрофосфат в количестве 1 г/л, гептагидрат сульфата магния в количестве 0,5 г/л и ампициллин в количестве 100 мг/л) и далее подвергают окончательному культивированию при 25°C в течение 24 часов с получением микробных клеток.
Культуру микробных клеток фракционируют с получением фракции периплазмы и фракции цитоплазмы после разрушения клеток путем осмотического шока с использованием раствора сахарозы в концентрации 20 грамм/децилитр (г/дл). Разрушенные микробные клетки погружают в раствор сахарозы с концентрацией 20 г/дл в 5 мМ водном растворе MgSO4. Полученный после центрифугирования супернатант представляет собой фракцию периплазмы ("Pe"). Дополнительно, полученный после центрифугирования осадок суспендируют и подвергают ультразвуковому разрушению. Полученный продукт представляет собой фракцию цитоплазмы ("Cy"). Активность глюкозо-6-фосфатдегидрогеназы, которая, как известно, присутствует в цитоплазме, использовалась в качестве индикатора для подтверждения отделения цитоплазмы. Указанное измерение проводят при добавлении соответствующего количества фермента при 30°C к реакционному раствору, содержащему 1 мМ глюкозо-6-фосфата, 0,4 мМ НАДФ, 10 мМ MgSO4 и 50 мМ Трис-HCl (pH 8), с последующим измерением величины поглощения при длине волны 340 нм для измерения уровня образования НАДФН.
На фиг.4 показано количество ферментов во фракции периплазмы и во фракции цитоплазмы, где активность полученного бесклеточного экстракта принимают за 100%. Во фракции периплазмы не была обнаружена активность глюкозо-6-фосфатдегидрогеназы. Это указывает на то, что фракция периплазмы не содержится в качестве примеси во фракции цитоплазмы. Примерно 60% активности образования Ala-Gln восстанавливается во фракции периплазмы, и было подтверждено, что Ala-Gln-образующий фермент секретируется в периплазму, как и было предположено на основе аминокислотной последовательности с использованием программы Signal P v 1.1.
(Пример 30) Получение L-аланил-L-глутамина с использованием микробных клеток Sphingobacterium sp.
50 мл (pH 7,0) среды, содержащей 5 г глюкозы, 5 г сульфата аммония, 1 г монокалия фосфата, 3 г дикалия фосфата, 0,5 г сульфата магния, 10 г дрожжевого экстракта и 10 г пептона в 1 л, переносят в колбу Сакагучи на 500 мл и стерилизуют при 115°C в течение 15 минут для последующего культивирования Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года). Затем в указанную среду инокулируют одну петлю клеток Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года), культивируют при 30°C в течение 24 часов на косой агаровой среде (агар: 20 г/л, pH 7,0), содержащей 5 г глюкозы, 10 г дрожжевого экстракта, 10 г пептона и 5 г NaCl в 1 л, с последующим культивированием при встряхивании при 30°C в течение 20 часов и со скоростью 120 встряхиваний/минуту. Затем 1 мл указанного культурального бульона добавляют к приведенной выше среде (50 мл/колбу Сакагучи на 500 мл) и культивируют при 30°C в течение 18 часов. После завершения культивирования микробные клетки отделяют от культурального бульона центрифугированием и суспендируют в 0,1 М боратном буфере (pH 9,0), содержащем 10 мМ ЭДТА, в концентрации 100 г/л в расчете на сырой вес микробных клеток. Затем 0,1 мл 100 мМ боратного буфера (pH 9,0), содержащего 10 мМ ЭДТА, 200 мМ гидрохлорида сложного метилового эфира L-аланина и 400 мМ L-глутамина, добавляют к 0,1 мл указанной суспензии микробных клеток. Полученную смесь в количестве 0,2 мл оставляют для реакции при температуре 25°C в течение 120 минут. Концентрация полученного за указанный период времени L-аланил-L-глутамина составляет 62 мМ.
(Пример 31) Очистка фермента из Sphingobacterium sp.
Описанную ниже процедуру после центрифугирования проводят на льду или при температуре 4°C. Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года) культивируют по методике примера 21 и далее микробные клетки собирают центрифугированием (10000 об/мин, 15 минут). После промывки 2 г микробных клеток с использованием 20 мМ Трис-HCl буфера (pH 7,6) полученные клетки суспендируют в 8 мл того же буфера и подвергают ультразвуковому разрушению путем обработки в течение 45 минут при мощности 195 ватт. Полученную после разрушения ультразвуком суспензию далее центрифугируют (10000 об/мин, 30 минут) для удаления клеточного дебриса, получая супернатантную жидкость разрушенных ультразвуком клеток. Указанную супернатантную жидкость разрушенных ультразвуком клеток диализуют в течение ночи против 20 мМ Трис-HCl буфера (pH 7,6) с последующим удалением нерастворимой фракции ультрацентрифугированием (50000 об/мин, 30 минут), получая растворимую фракцию в виде супернатантной жидкости. Полученную растворимую фракцию наносят на колонку с сефарозой Q-Sepharose HP (производство компании Amersham), которая была предварительно уравновешена Трис-HCl буфером (pH 7,6), и активную фракцию собирают из неадсорбированной фракции. Указанную активную фракцию диализуют в течение ночи против 20 мМ ацетатного буфера (pH 5,0) с последующим удалением нерастворимой фракции путем центрифугирования (10000 об/мин, 30 минут) с получением диализованной фракции в виде супернатантной жидкости. Диализованную фракцию далее наносят на колонку с сефарозой Q-Sepharose HP (производство компании Amersham), которая была предварительно уравновешена 20 мМ ацетатным буфером (pH 5,0) с получением активной фракции, при проведении элюирования фермента линейным концентрационным градиентом того же буфера, содержащего от 0 до 1 М NaCl.
(Пример 32) Получение L-аланил-L-глутамина с использованием активной фракции
10 мкл фракции, полученной после элюирования с сефарозы SP-Sepharose HP (примерно 27 Ед/мл) и очищенной по процедуре примера 31, добавляют к 90 мкл боратного буфера (pH 9,0), содержащего 111 мМ гидрохлорида сложного метилового эфира L-аланина, 222 мМ L-глутамина и 11 мМ ЭДТА, и оставляют для реакции при 25°C в течение 120 минут. В результате реакции получают 73 мМ L-аланил-L-глутамина в варианте с добавлением фермента. С другой стороны, отмечается ничтожный уровень образования L-Ala-L-Glu в образце без добавления фермента, и указанное образованное количество составляет лишь примерно 0,07 мМ после реакции в течение 120 минут.
(Пример 33) Выделение гена пептид-образующего фермента из Sphingobacterium sp.
Ниже приводится описание методики выделения гена пептид-образующего фермента, в которой в качестве микроорганизма используют Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года). Выделение гена проводят с использованием Escherichia coli DH5α в качестве клетки-хозяина и pUC118 в качестве вектора.
(1) Получение микроба
Клетки Sphingobacterium sp. штамм FERM BP-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года) культивируют при 25°C в течение 24 часов на агаровой среде CM2G (содержит 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну полную петлю полученных микробных клеток инокулируют в колбу Сакагучи на 500 мл, содержащую 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара), и затем культивируют при встряхивании при 25°C.
(2) Получение хромосомной ДНК из микробных клеток
50 мл культурального бульона центрифугируют (12000 об/мин, 4°C, 15 минут) для сбора микробных клеток. Затем получают хромосомную ДНК из бактериальных клеток с использованием системы Qiagen Genomic-Tip System (Qiagen).
(3) Получение зонда к фрагменту ДНК путем ПЦР
Фрагмент ДНК, содержащий часть гена пептидобразующего фермента, полученного из Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года), получают путем ПЦР с использованием LA-Taq (производство компании Takara Shuzo). Затем проводят реакцию ПЦР на основе хромосомной ДНК, полученной из клеток Empedobacter brevis штамм FERM BP-8113 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 8 июля 2002 года), с использованием праймеров, имеющих последовательности оснований SEQ ID NO:3 и SEQ ID NO:4.
Проводят ПЦР в течение 30 циклов с использованием набора для циклической термической обработки (Takara PCR Thermal Cycler PERSONAL) (производство компании Takara Shuzo) в указанных ниже условиях:
94°C в течение 30 секунд
52°C в течение 1 минуты
72°C в течение 1 минуты
После проведения реакции 3 мкл реакционной жидкости наносят на 0,8% гель агарозы для проведения электрофореза. В результате было подтверждено, что произошла амплификация фрагмента ДНК размером примерно 1,5 т.н.
(4) Клонирование гена пептидобразующего фермента из генной библиотеки
Для получения гена пептидобразующего фермента полной длины проводят саузерн-гибридизацию с использованием в качестве зонда фрагмента ДНК, амплифицированного по указанной выше методике ПЦР. Процедура саузерн-гибридизации описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Фрагмент ДНК размером примерно 1,5 т.н., амплифицированный по методике ПЦР, отделяют электрофорезом в 0,8% геле агарозы. Затем вырезают целевую полосу и очищают фрагмент ДНК. Указанный фрагмент ДНК метят с использованием зонда дигоксинигена с использованием набора DIG High Prime (производство компании Boehringer-Mannheim) по методике, описанной в руководстве к набору.
После полного расщепления хромосомной ДНК из клеток Sphingobacterium sp., полученных на стадии (2) данного примера 33, в процессе реакции при 37°C в течение 16 часов рестрикционным ферментом SacI, полученную ДНК подвергают электрофорезу в 0,8% геле агарозы. После электрофореза хромосомную ДНК подвергают блоттингу на положительно загруженном мембранном фильтре из найлона (Nylon) (производство компании Roche Diagnostics) из агарозного геля после электрофореза с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 37°C в течение 1 часа добавляют зонд, меченный дигоксинигеном, полученный по указанной выше методике, и проводят гибридизацию при 37°C в течение 16 часов. Далее фильтр последовательно промывают два раза при температуре 60°C в 1 × SSC, содержащем 0,1% ДСН.
Выявление полос, которые гибридизовались с зондом, проводят с использованием набора для выявления нуклеотидов DIG Nucleotide Detection Kit (производство компании Boehringer-Mannheim) по методике, описанной в инструкции к набору. В результате было показано, что полоса размером примерно 3 т.н. гибридизовалась с данным зондом.
Хромосомную ДНК, полученную на стадии (2) данного примера 33, подвергают полному расщеплению с использованием SacI. ДНК размером примерно 3 т.н. отделяют электрофорезом в 0,8% геле агарозы и далее очищают ДНК с использованием набора Gene Clean II Kit (производство компании Funakoshi) и растворяют ДНК в 10 мкл ТЭ. После проведения реакции 4 мкл указанного продукта с SacI при температуре 37°C в течение 16 часов для полного расщепления, продукт смешивают с pUС118, обработанным щелочной фосфатазой (E. coli C75) при 37°C в течение 30 минут и при 50°C в течение 30 минут и затем проводят реакцию лигирования с использованием набора для лигирования ДНК (DNA Ligation Kit Ver.2) (производство компании Takara Shuzo). Смешивают 5 мкл указанной жидкости для реакции лигирования и 100 мкл компетентных клеток Escherichia coli DH5α (производство компании Takara Shuzo) для трансформации клеток Escherichia coli. Полученные таким образом трансформанты затем вносят в подходящую твердую среду для получения библиотеки хромосомной ДНК.
Для целей получения гена пептидобразующего фермента полной длины библиотеку хромосомной ДНК подвергают скринингу по методике гибридизации колоний с использованием указанного выше зонда. Процедура гибридизации колоний описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии из библиотеки хромосомной ДНК переносят на мембранный фильтр из найлона (Nylon Membrane for Colony and Plaque Hybridization) (производство компании Roche Diagnostics) с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 37°C в течение 1 часа добавляют указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 37°C в течение 16 часов. Далее фильтр последовательно промывают дважды при температуре 60°C в 1 × SSC, содержащем 1% ДСН.
Выявление колоний, которые гибридизовались с меченым зондом, проводят с использованием набора для выявления нуклеотидов DIG Nucleotide Detection Kit (производство компании Boehringer-Mannheim) по методике, описанной в инструкции к набору. В результате было подтверждено, что шесть штаммов колоний гибридизовались с меченым зондом.
(5) Последовательность оснований гена пептидобразующего фермента, полученного из Sphingobacterium sp.
Получают плазмиды, содержащиеся в клетках Escherichia coli DH5α, из указанных выше шести штаммов микробных клеток, для которых был подтвержден факт гибридизации с меченым зондом, с использованием системы очистки ДНК Wizard Plus Minipreps DNA Purification System (производство компании Promega) для определения соседних последовательностей оснований, которые гибридизовались зондом. Реакцию секвенирования проводят с использованием набора CEQ DTCS-Quick Start Kit (производство компании Beckman-Coulter) по методике, описанной в инструкции к набору. Дополнительно, проводят электрофорез с использованием CEQ 2000-XL (производство компании Beckman-Coulter).
В результате было показано наличие открытой рамки считывания, которая кодирует пептидобразующий фермент. Последовательность оснований гена пептидобразующего фермента полной длины, полученного из Sphingobacterium sp., а также соответствующая аминокислотная последовательность приведены в SEQ ID NO:11. Пептидобразующий фермент, полученный из Sphingobacterium sp., демонстрирует гомологию на уровне 63,5% по аминокислотной последовательности с пептидобразующим ферментом, полученным из указанного выше Empedobacter brevis (при определении с использованием программы BLASTP).
(Пример 34) Экспрессия гена пептидобразующего фермента, полученного из Sphingobacterium sp., в Escherichia coli
Целевой ген подвергают амплификации в ходе реакции ПЦР с использованием хромосомной ДНК из Sphingobacterium sp. штамм FERM ВР-8124 (Депонирующий Институт: независимая административная корпорация, Национальный Институт современной промышленной науки и технологии, Международный Депозитарий запатентованных организмов (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary), Адрес Депонирующего Института: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), дата поступления в Международный Депозитарий: 22 июля 2002 года) в качестве матрицы и олигонуклеотидов, показанных в SEQ ID NO:13 и 14, в качестве праймеров. Далее указанный фрагмент ДНК обрабатывают NdeI/XbaI и лигируют полученный фрагмент ДНК и продукт обработки pTrpT с использованием NdeI/XbaI. Затем Escherichia coli JM-109 трансформируют указанным лигирующим раствором, из ампициллин-резистентных штаммов отбирают штаммы, несущие целевую плазмиду, и обозначают данную плазмиду как pTrpT_Sm_aet.
Клетки Escherichia coli JM-109, несущие pTrpT_Sm_aet, культивируют при 25°C в течение 20 часов после инокуляции петли с клетками данного штамма в обычную опытную пробирку, содержащую 3 мл среды (глюкоза в количестве 2 г/л, дрожжевой экстракт в количестве 10 г/л, казаминовые кислоты в количестве 10 г/л, сульфат аммония в количестве 5 г/л, дигидрофосфат калия в количестве 3 г/л, дикалий гидрофосфат в количестве 1 г/л, гептагидрат сульфата магния в количестве 0,5 г/л и ампициллин в количестве 100 мг/л). Получение клонированного гена, содержащего активность продуцирования L-аланил-L-глутамина на уровне 2,1 Ед в расчете на 1 мл культурального бульона, подтверждают путем экспрессии в E. coli. Кроме того, не было обнаружено никакой активности у трансформанта, который служит в качестве контроля и который содержит только pTrpT.
(Прогнозирование сигнальной последовательности)
В случае анализа аминокислотной последовательности SEQ ID NO:12, описанной в перечне последовательностей, с использованием программы Signal P v 1.1 (см. Protein Engineering, Vol. 12, No. 1, pp. 3-9, 1999), было предположено, что участок аминокислотной последовательности от 1 до 20 номера функционирует в качестве сигнала для секреции пептида в периплазму, тогда как зрелый белок считывается по ходу транскрипции, начиная с аминокислоты номер 21.
(Подтверждение сигнальной последовательности)
Клетки Escherichia coli JM-109, содержащие pTrpT_Sm_aet, культивируют при 25°C в течение 20 часов после инокулирования петли с клетками данного штамма в обычную опытную пробирку, содержащую 50 мл среды (глюкоза в количестве 2 г/л, дрожжевой экстракт в количестве 10 г/л, казаминовые кислоты в количестве 10 г/л, сульфат аммония в количестве 5 г/л, дигидрофосфат калия в количестве 3 г/л, дикалий гидрофосфат в количестве 1 г/л, гептагидрат сульфата магния в количестве 0,5 г/л и ампициллин в количестве 100 мг/л).
Описанную ниже процедуру после центрифугирования проводят либо на льду, либо при температуре 4°C. После завершения культивирования микробные клетки отделяют от культурального бульона путем центрифугирования и после промывания с использованием 100 мМ боратного буфера (pH 7) суспендируют в том же буфере. Затем бактериальные клетки подвергают ультразвуковому разрушению в течение 20 минут при мощности 195 ватт, жидкость после ультразвукового разрушения центрифугируют (12000 об/мин, 30 минут) для удаления клеточного дебриса и получают растворимую фракцию. Полученную растворимую фракцию наносят на колонку CHT-II (производство компании Biorad), которая была предварительно уравновешена 100 мМ фосфатным буфером (pH 7) и фермент элюируют 500 мМ фосфатным буфером с линейным градиентом концентрации. Раствор, полученный при смешивании активной фракции с 5-кратным объемом 2 М сульфата аммония и 100 мМ фосфатного буфера, наносят на колонку Resource-PHE (производство компании Amersham), которая была предварительно уравновешена 2М сульфатом аммония и 100 мМ фосфатным буфером, и фермент элюируют линейным градиентом концентрации от 2 до 0 М сульфата аммония с получением раствора активной фракции. В результате указанных процедур было подтверждено, что пептидобразующий фермент очищается однородно, с точки зрения поведения его при электрофорезе.
В том случае, когда аминокислотную последовательность указанного выше пептидобразующего фермента определяют по методу разложения Эдмана, получают аминокислотную последовательность SEQ ID NO:15, подтверждая, что зрелый белок считывается по ходу транскрипции от аминокислоты 21, как и было предположено при проведении анализа по программе SignalP v 1.1.
(Пример 35) Выделение гена пептидобразующего фермента, полученного из Pedobacter heparinus IFO 12017
Далее описывается методика выделения гена пептидобразующего фермента. Используемый микроб представляет собой Pedobacter heparinus IFO 12017 (Депонирующий Институт: Институт ферментации (Institute of Fermentation), Адрес Депонирующего Института: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония). Клетки Escherichia coli JM-109 используют в качестве клеток-хозяев, тогда как pUC118 используют в качестве вектора для выделения гена.
(1) Получение микроба
Клетки Pedobacter heparinus штамм IFO 12017 (Депонирующий Институт: Институт ферментации (Institute of Fermentation), Адрес Депонирующего Института: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) культивируют при 25°C в течение 24 часов на среде с агаром CM2G (содержит 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну полную петлю полученных бактериальных клеток инокулируют в колбу Сакагучи на 500 мл, содержащую 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара) и затем культивируют при встряхивании при 25°C.
(2) Получение хромосомной ДНК из микробных клеток
50 мл культурального бульона подвергают центрифугированию (12000 об/мин, 4°C, 15 минут) для сбора бактериальных клеток. Затем из микробных клеток выделяют хромосомную ДНК с использованием системы Qiagen Genomic-Tip System (Qiagen) с использованием методики, описанной в руководстве для пользователя.
(3) Получение фрагмента зондовой ДНК путем ПЦР
Фрагмент ДНК, содержащий часть гена пептидобразующего фермента, полученного из Pedobacter heparinus штамм IFO 12017 (Депонирующий Институт: Институт ферментации (Institute of Fermentation), Адрес депонирующего института: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония), получают методом ПЦР с использованием La-Taq (производство компании Takara Shuzo). ПЦР проводят на основе хромосомной ДНК, полученной из клеток Pedobacter heparinus штамм IFO 12017 (Депонирующий Институт: Институт ферментации (Institute of Fermentation), Адрес Депонирующего Института: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония), с использованием праймеров, имеющих последовательности оснований SEQ ID NO:15 и 16. Фрагмент ДНК размером примерно 1 т.н., амплифицированный методом ПЦР, разделяют электрофорезом в 0,8% агарозном геле. Затем вырезают целевую полосу и очищают полученный таким образом фрагмент ДНК. Указанную ДНК метят зондом с дигоксинигеном с использованием набора DIG High Prime по методике, описанной в инструкции, прилагаемой к набору (производство компании Boehringer-Mannheim).
(4) Клонирование гена пептидобразующего фермента из генной библиотеки
Для получения гена пептидобразующего фермента полной длины проводят саузерн-гибридизацию с использованием в качестве зонда фрагмента ДНК, амплифицированного методом ПЦР. Процедура саузерн-гибридизации описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
После завершения расщепления хромосомной ДНК из Pedobacter heparinus штамм IFO 12017 (Депонирующий Институт: Институт ферментации (Institute of Fermentation), Адрес Депонирующего Института: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) в ходе реакции при 37°C в течение 16 часов с использованием рестрикционного фермента HindIII далее проводят электрофорез в 0,8% геле агарозы. После электрофореза хромосомную ДНК подвергают блоттингу на положительно загруженном мембранном фильтре из найлона (Nylon) (производство компании Roche Diagnostics) из агарозного геля после электрофореза с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После проведения предварительной гибридизации на фильтре при температуре 50°C в течение 1 часа добавляют зонд, меченный дигоксинигеном, полученный по указанной выше методике, и проводят гибридизацию при 50°C в течение 16 часов. Далее фильтр промывают дважды при 60°C в 1 × SSC, содержащем 0,1% ДСН.
Выявление полос, которые гибридизовались с зондом, проводят с использованием набора для выявления нуклеотидов (DIG Nucleotide Detection Kit) (производство компании Boehringer-Mannheim) по методике, описанной в инструкции к набору. В результате было показано, что полоса размером примерно 5 т.н. гибридизовалась с данным зондом.
Хромосомную ДНК из Pedobacter heparinus штамм IFO 12017 (Депонирующий Институт: Институт ферментации (Institute of Fermentation), Адрес Депонирующего Института: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) полностью расщепляют с помощью HindIII. ДНК размером примерно 5 т.н. разделяют электрофорезом в 0,8% геле агарозы с последующей очисткой ДНК с использованием набора Gene Clean II Kit (производство компании Funakoshi) и растворяют ДНК в 10 мкл ТЭ. Затем 4 мкл указанного продукта смешивают с pUC118 HindIII/BAP (производство компании Takara Shuzo) и проводят реакцию лигирования с использованием набора для лигирования ДНК (DNA Ligation Kit Ver.2) (производство компании Takara Shuzo). 5 мкл указанной реакционной жидкости для реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM-109 (производство компании Takara Shuzo) смешивают для трансформации клеток Escherichia coli. Затем полученные таким образом трансформанты вносят в соответствующую твердую среду для получения библиотеки хромосомной ДНК.
Для получения гена пептидобразующего фермента полной длины библиотеку хромосомной ДНК подвергают скринингу на гибридизацию колоний с использованием указанного выше зонда. Процедура гибридизации описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии из библиотеки хромосомной ДНК переносят на мембранный фильтр из найлона (Nylon Membrane for Colony and Plaque Hybridization) (производство компании Roche Diagnostics) и затем проводят обработку, включающую щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После проведения предварительной гибридизации на фильтре при температуре 37°C в течение 1 часа добавляют зонд, меченный дигоксинигеном, и проводят гибридизацию при 37°C в течение 16 часов. Далее фильтр промывают дважды при 60°C в 1 × SSC, содержащем 1% ДСН.
Выявление колоний, который гибридизовались с меченым зондом, проводят по методике, описанной в руководстве, с использованием набора для выявления нуклеотидов (DIG Nucleotide Detection Kit) (производство компании Boehringer-Mannheim). В результате было показано, что 1 штамм колоний гибридизовался с меченым зондом.
(5) Последовательность оснований гена пептидобразующего фермента, полученного из Pedobacter heparinus штамм IFO-12017
Плазмиды, имеющиеся в Escherichia coli JM-109, получают из указанного выше штамма микробных клеток, в отношении которых было подтверждено, что они гибридизуются с меченым зондом, и была определена соответствующая последовательность оснований, которая гибридизуется с зондом. Реакцию секвенирования проводят с использованием набора CEQ DTCS-Quick Start Kit (производство компании Beckman-Coulter) по методике, описанной в инструкции к набору. Дополнительно проводят электрофорез с использованием CEQ 2000-XL (производство компании Beckman-Coulter).
В результате было показано существование открытой рамки считывания, которая кодирует пептидобразующий фермент. Последовательность оснований гена пептидобразующего фермента, полученного из Pedobacter heparinus штамм IFO-12017 (Депонирующий Институт: Институт ферментации (Institute of Fermentation), Адрес Депонирующего Института: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония), а также соответствующая аминокислотная последовательность показаны в SEQ ID NO:17 в перечне последовательностей.
(Пример 36) Экспрессия гена пептидобразующего фермента, полученного из Pedobacter heparinus штамм IFO-12017, в E. coli
Целевой ген амплифицируют методом ПЦР с использованием хромосомной ДНК из Pedobacter heparinus штамм IFO-12017 (Депонирующий Институт: Институт ферментации (Institute of Fermentation), Адрес Депонирующего Института: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) в качестве матрицы и олигонуклеотидов, приведенных в SEQ ID NO:19 и 20, в качестве праймеров. Указанный фрагмент ДНК обрабатывают NdeI/HindIII и лигируют полученный фрагмент ДНК и продукт обработки pTrpT с использованием NdeI/HindIII. Затем Escherichia coli JM-109 трансформируют указанным лигирующим раствором, отбирают из ампициллин-резистентных штаммов штаммы, несущие целевую плазмиду, и указанную плазмиду обозначают как pTrpT_Ph_aet.
Клетки Escherichia coli JM-109, несущие pTrpT_Ph_aet, подвергают предварительному культивированию при 25°C в течение 20 часов при инокуляции полной петли с клетками данного штамма в обычную опытную пробирку, содержащую 3 мл среды (глюкоза в количестве 2 г/л, дрожжевой экстракт в количестве 10 г/л, казаминовые кислоты в количестве 10 г/л, сульфат аммония в количестве 5 г/л, дигидрофосфат калия в количестве 3 г/л, дикалий гидрофосфат в количестве 1 г/л, гептагидрат сульфата магния в количестве 0,5 г/л и ампициллин в количестве 100 мг/л). Подтверждают наличие экспрессии в E. coli клонированного гена, обладающего активностью образования L-аланил-L-глутамина на уровне 0,3 Ед на мл культуральной жидкости. Более того, не было обнаружено никакой активности у трансформанта, который содержал только pTrpT и служил в качестве контроля.
(Пример 37) Выделение гена пептидобразующего фермента, полученного из Taxeobacter gelupurpurascens штамм DSMZ 11116
Далее в описании приводится методика выделения гена пептидобразующего фермента. Используемый микроорганизм представляет собой Taxeobacter gelupurpurascens штамм DSMZ 11116 (Депонирующий Институт: Германский Институт микробных и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (German Collection of Microbes and Cell Cultures), Адрес Депонирующего Института: Mascheroder Weg 1b, 38124 Braunschweig, Германия). Клетки Escherichia coli JM-109 используют в качестве организма-хозяина, тогда как pUC118 используют в качестве вектора при выделении гена.
(1) Получение микроба
Клетки Taxeobacter gelupurpurascens штамм DSMZ 11116 (Депонирующий Институт: Германский Институт микробных и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (German Collection of Microbes and Cell Cultures), Адрес Депонирующего Института: Mascheroder Weg 1b, 38124 Braunschweig, Германия) культивируют при 25°C в течение 24 часов на среде с агаром CM2G (содержит 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну полную петлю полученных микробных клеток инокулируют в колбу Сакагучи на 500 мл, содержащую 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара) и культивируют при встряхивании при 25°C.
(2) Получение хромосомной ДНК из микробных клеток
50 мл культуральной жидкости подвергают центрифугированию (12000 об/мин, 4°C, 15 минут) для сбора микробных клеток. Затем из указанных микробных клеток выделяют хромосомную ДНК с использованием системы Qiagen Genomic-Tip System (Qiagen) по методике, описанной в руководстве для пользователя.
(3) Получение фрагмента зондовой ДНК методом ПЦР
Фрагмент ДНК, содержащий часть гена пептидобразующего фермента, полученного из Taxeobacter gelupurpurascens штамм DSMZ 11116 (Депонирующий Институт: Германский Институт микробных и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (German Collection of Microbes and Cell Cultures), Адрес Депонирующего Института: Mascheroder Weg 1b, 38124 Braunschweig, Германия), выделяют методом ПЦР с использованием La-Taq (производство компании Takara Shuzo). ПЦР проводят на хромосомной ДНК, выделенной из клеток Taxeobacter gelupurpurascens штамм DSMZ 11116 (Депонирующий Институт: Германский Институт микробных и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (German Collection of Microbes and Cell Cultures), Адрес Депонирующего Института: Mascheroder Weg 1b, 38124 Braunschweig, Германия), с использованием праймеров, имеющих последовательности оснований SEQ ID NO:21 и 16. Фрагмент ДНК размером примерно 1 т.н., амплифицированный методом ПЦР, отделяют электрофорезом в 0,8% геле агарозы. Затем вырезают целевую полосу и очищают полученный таким образом фрагмент ДНК. Указанный фрагмент ДНК метят зондом с дигоксинигеном с использованием набора DIG High Prime по методике, описанной в инструкции, прилагаемой к набору (производство компании Boehringer-Mannheim).
(4) Клонирование гена пептидобразующего фермента из генной библиотеки
Для получения гена пептидобразующего фермента полной длины проводят саузерн-гибридизацию с использованием в качестве зонда фрагмента ДНК, амплифицированного методом ПЦР. Процедура саузерн-гибридизации описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
После завершения расщепления хромосомной ДНК из Taxeobacter gelupurpurascens штамм DSMZ 11116 (Депонирующий Институт: Германский Институт микробных и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (German Collection of Microbes and Cell Cultures), Адрес Депонирующего Института: Mascheroder Weg 1b, 38124 Braunschweig, Germany) в ходе реакции при 37°C в течение 16 часов с использованием рестрикционного фермента PstI, проводят электрофорез в 0,8% геле агарозы. Разделенную электрофорезом хромосомную ДНК переносят блоттингом на положительно загруженный мембранный фильтр из найлона (Nylon) (производство компании Roche Diagnostics) из агарозного геля после электрофореза с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 50°C в течение 1 часа добавляют зонд, меченный дигоксинигеном, полученный по указанной выше методике, и проводят гибридизацию при 50°C в течение 16 часов. Далее фильтр промывают дважды при 60°C в 1 × SSC, содержащем 0,1% ДСН.
Выявление полос, которые гибридизовались с зондом, проводят по описанной в руководстве методике с использованием набора для выявления нуклеотидов DIG Nucleotide Detection Kit (производство компании Boehringer-Mannheim). В результате было показано, что полоса размером примерно 5 т.н. гибридизовалась с данным зондом.
Хромосомную ДНК из Taxeobacter gelupurpurascens штамм DSMZ 11116 (Депонирующий Институт: Германский Институт микробных и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (German Collection of Microbes and Cell Cultures), Адрес Депонирующего Института: Mascheroder Weg 1b, 38124 Braunschweig, Germany) полностью расщепляют HindIII. ДНК размером приблизительно 5 т.н. разделяют электрофорезом в 0,8% геле агарозы и далее очищают ДНК с использованием набора Gene Clean II Kit (производство компании Funakoshi) и растворяют ДНК в 10 мкл ТЭ. Затем 4 мкл указанного продукта смешивают с pUC118 PstI/BAP (производство компании Takara Shuzo) и проводят реакцию лигирования с использованием набора для лигирования ДНК (DNA Ligation Kit Ver.2) (производство компании Takara Shuzo). Смешивают 5 мкл указанной жидкости для реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM-109 (производство компании Takara Shuzo) для трансформации клеток Escherichia coli. Далее полученные таким образом трансформанты вносят в соответствующую твердую среду для получения библиотеки хромосомной ДНК.
Для получения гена пептидобразующего фермента полной длины подвергают скринингу библиотеку хромосомной ДНК по методике гибридизации колоний с использованием указанного выше зонда. Процедура гибридизации колоний описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии из библиотеки хромосомной ДНК переносят на мембранный фильтр из найлона (Nylon Membrane for Colony and Plaque Hybridization) (производство компании Roche Diagnostics) с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 37°C в течение 1 часа добавляют указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 37°C в течение 16 часов. Далее фильтр дважды промывают при 60°C в 1 × SSC, содержащем 0,1% ДСН.
Выявление колоний, которые гибридизовались с меченым зондом, проводят с использованием набора для выявления нуклеотидов DIG Nucleotide Detection Kit (производство компании Boehringer-Mannheim) по методике, описанной в инструкции к набору. В результате было показано, что один штамм колоний гибридизовался с меченым зондом.
(5) Последовательность оснований гена пептидобразующего фермента, полученного из Taxeobacter gelupurpurascens штамм DSMZ 11116
Плазмиды, имеющиеся в Escherichia coli JM 109, получают из указанного выше штамма микробных клеток, в отношении которых показано, что они гибридизуются с меченым зондом, и определяют соответствующую последовательность оснований, которая гибридизовалась зондом. Проводят реакцию секвенирования с использованием набора CEQ DTCS-Quick Start Kit (производство компании Beckman-Coulter) по методике, описанной в инструкции к набору. Дополнительно проводят электрофорез с использованием CEQ 2000-XL (производство компании Beckman-Coulter).
В результате было показано наличие открытой рамки считывания, которая кодирует пептидобразующий фермент. Последовательность оснований гена пептидобразующего ферментов полной длины, полученного из Taxeobactergelupurpurascens штамм DSMZ 11116 (Депонирующий Институт: Германский Институт микробных и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (German Collection of Microbes and Cell Cultures), Адрес Депонирующего Института: Mascheroder Weg 1b, 38124 Braunschweig, Германия), а также соответствующая им аминокислотная последовательность показаны в SEQ ID NO:22 в перечне последовательностей.
(Пример 38) Выделение гена пептидобразующего фермента, полученного из Cyclobacterium marinum штамм АТСС 25205
Далее в описании приводится методика выделения гена пептидобразующего фермента. Используемый микроорганизм представляет собой Cyclobacterium marinum штамм АТСС 25205 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки). Клетки Escherichia coli JM-109 используют в качестве организма-хозяина, тогда как pUC118 используют в качестве вектора при выделении гена.
(1) Получение микроба
Клетки Cyclobacterium marinum штамм АТСС 25205 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) культивируют при 25°C в течение 24 часов на среде с агаром CM2G (содержит 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну полную петлю полученных микробных клеток инокулируют в колбу Сакагучи на 500 мл, содержащую 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара) и затем культивируют при встряхивании при 25°C.
(2) Получение хромосомной ДНК из микробных клеток
50 мл культурального бульона подвергают центрифугированию (12000 об/мин, 4°C, 15 минут) для сбора микробных клеток. Затем из указанных микробных клеток получают хромосомную ДНК с использованием системы Qiagen Genomic-Tip System (Qiagen) по методике, описанной в руководстве для пользователя.
(3) Получение фрагмента зондовой ДНК методом ПЦР
Фрагмент ДНК, содержащий часть гена пептидобразующего фермента, выделенного из Cyclobacterium marinum штамм АТСС 25205 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), получают методом ПЦР с использованием La-Taq (производство компании Takara Shuzo). Затем проводят ПЦР на основе хромосомной ДНК, выделенной из клеток Cyclobacterium marinum штамм АТСС 25205 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), с использованием праймеров, имеющих последовательности оснований SEQ ID NO:15 и 16. Фрагмент ДНК размером примерно 1 т.н., амплифицированный методом ПЦР, отделяют электрофорезом в 0,8% геле агарозы. Затем вырезают целевую полосу и очищают полученный таким образом фрагмент ДНК. Указанный фрагмент ДНК метят зондом с дигоксинигеном с использованием набора DIG High Prime по методике, описанной в инструкции, прилагаемой к набору (производство компании Boehringer-Mannheim).
(4) Клонирование гена пептидобразующего фермента из генной библиотеки
Для получения гена пептидобразующего фермента полной длины проводят саузерн-гибридизацию с использованием в качестве зонда фрагмента ДНК, амплифицированного по методике ПЦР. Процедура саузерн-гибридизации описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
После завершения полного расщепления хромосомной ДНК из Cyclobacterium marinum штамм АТСС 25205 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) в ходе реакции при 37°C в течение 16 часов с использованием рестрикционного фермента HincII, затем проводят электрофорез в 0,8% геле агарозы. Разделенную электрофорезом хромосомную ДНК переносят блоттингом на положительно загруженный мембранный фильтр из найлона (Nylon) (производство компании Roche Diagnostics) из агарозного геля после электрофореза с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Затем проводят гибридизацию с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 50°C в течение 1 часа добавляют зонд, меченный дигоксинигеном, полученный по указанной выше методике, и проводят гибридизацию при 50°C в течение 16 часов. Далее фильтр промывают дважды при 60°C в 1 × SSC, содержащем 0,1% ДСН.
Выявление полос, которые гибридизовались с зондом, проводят по описанной в руководстве методике с использованием набора для выявления нуклеотидов DIG Nucleotide Detection Kit (производство компании Boehringer-Mannheim). В результате было показано, что в продукте расщепления PstI полоса размером примерно 7 т.н. гибридизовалась с данным зондом, тогда как, по данным анализа, в продукте расщепления HincII полоса размером 2 т.н. гибридизовалась с зондом.
Хромосомную ДНК из Cyclobacterium marinum штамм АТСС 25205 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) полностью расщепляют с использованием PstI или HincII. ДНК размером приблизительно 7 т.н. или 2 т.н. разделяют электрофорезом в 0,8% геле агарозы и далее очищают ДНК с использованием набора Gene Clean II Kit (производство компании Funakoshi) и растворяют ДНК в 10 мкл ТЭ. Затем смешивают 4 мкл указанного продукта с pUC118 PstI/BAP (производство компании Takara Shuzo) или pUC118 HincII/BAP (производство компании Takara Shuzo) и проводят реакцию лигирования с использованием набора для лигирования ДНК (DNA Ligation Kit Ver.2) (производство компании Takara Shuzo). 5 мкл указанной жидкости для реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM-109 (производство компании Takara Shuzo) смешивают, соответственно, для трансформации клеток Escherichia coli. Далее полученные таким образом трансформанты вносят в соответствующую твердую среду для получения библиотеки хромосомной ДНК.
Для получения гена пептидобразующего фермента полной длины библиотеку хромосомной ДНК подвергают скринингу по методике гибридизации колоний с использованием указанного выше зонда. Процедура гибридизации колоний описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии из библиотеки хромосомной ДНК переносят на мембранный фильтр из найлона (Nylon Membrane for Colony and Plaque Hybridization), (производство компании Roche Diagnostics) с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 37°C в течение 1 часа добавляют указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 37°C в течение 16 часов. Затем фильтр дважды промывают при 60°C в 1 × SSC, содержащем 0,1% ДСН.
Выявление колоний, которые гибридизовались с меченым зондом, проводят с использованием набора для выявления нуклеотидов DIG Nucleotide Detection Kit (производство компании Boehringer-Mannheim) по методике, описанной в инструкции к набору. В результате было показано, что один штамм колоний гибридизовался с меченым зондом.
(5) Последовательность оснований гена пептидобразующего фермента, полученного из Cyclobacterium marinum штамм АТСС 25205
Плазмиды, имеющиеся в Escherichia coli JM 109, получают из указанного выше штамма микробных клеток, в отношении которых было показано, что они гибридизуются с меченым зондом, и определяют соответствующую последовательность оснований, которая гибридизовалась с зондом. Проводят реакцию секвенирования с использованием набора CEQ DTCS-Quick Start Kit (производство компании Beckman-Coulter) по методике, описанной в инструкции к набору. Дополнительно проводят электрофорез с использованием CEQ 2000-XL (производство компании Beckman-Coulter).
В результате было показано наличие открытой рамки считывания, которая кодирует пептидобразующий фермент. Последовательность оснований гена пептидобразующего фермента полной длины, полученная из Cyclobacterium marinum штамм АТСС 25205 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), а также соответствующая им аминокислотная последовательность показаны в SEQ ID NO:24 в перечне последовательностей.
(Пример 39) Выделение гена пептидобразующего фермента, полученного из Psycloserpens burtonensisштамм АТСС 700359
Далее в описании приводится методика выделения гена пептидобразующего фермента. Используемый микроорганизм представляет собой Psycloserpens burtonensis штамм АТСС 700359 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки). Клетки Escherichia coli JM-109 используют в качестве организма-хозяина, тогда как pUC118 используют в качестве вектора при выделении гена.
(1) Получение микроба
Psycloserpens burtonensis штамм АТСС 700359 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) культивируют при 10°C в течение 24 часов на среде с агаром CM2G (содержит 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну полную петлю полученных микробных клеток инокулируют в колбу Сакагучи на 500 мл, содержащую 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара) и культивируют при встряхивании при 10°C.
(2) Получение хромосомной ДНК из микробных клеток
50 мл культуральной жидкости подвергают центрифугированию (12000 об/мин, 4°C, 15 минут) для сбора микробных клеток. Затем из полученных микробных клеток выделяют хромосомную ДНК с использованием системы Qiagen Genomic-Tip System (Qiagen) по методике, описанной в руководстве для пользователя.
(3) Получение фрагмента зондовой ДНК методом ПЦР
Фрагмент ДНК, содержащий часть гена пептидобразующего фермента, полученного из Psycloserpens burtonensis штамм АТСС 700359 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), получают методом ПЦР с использованием La-Taq (производство компании Takara Shuzo). Затем проводят ПЦР на основе хромосомной ДНК, выделенной из клеток Psycloserpens burtonensis штамм АТСС 700359 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: (P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), с использованием праймеров, имеющих последовательности оснований SEQ ID NO:15 и 16. Фрагмент ДНК размером примерно 1 т.н., амплифицированный методом ПЦР, разделяют электрофорезом в 0,8% геле агарозы. Затем вырезают целевую полосу и очищают полученный таким образом фрагмент ДНК. Указанный фрагмент ДНК метят зондом с дигоксинигеном с использованием набора DIG High Prime по методике, описанной в инструкции, прилагаемой к набору (производство компании Boehringer-Mannheim).
(4) Клонирование гена пептидобразующего фермента из генной библиотеки
Для получения гена пептидобразующего фермента полной длины проводят саузерн-гибридизацию с использованием в качестве зонда фрагмента ДНК, амплифицированного методикой ПЦР. Процедура саузерн-гибридизации описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
После завершения полного расщепления хромосомной ДНК из Psycloserpens burtonensis штамм АТСС 700359 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) в ходе реакции при 37°C в течение 16 часов с использованием рестрикционного фермента EcoRI, затем проводят электрофорез в 0,8% геле агарозы. После электрофореза хромосомную ДНК переносят блоттингом на положительно загруженный мембранный фильтр из найлона (Nylon) (производство компании Roche Diagnostics) из агарозного геля с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Затем проводят гибридизацию с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 50°C в течение 1 часа добавляют зонд, меченный дигоксинигеном, полученный по указанной выше методике, и проводят гибридизацию при 50°C в течение 16 часов. Далее фильтр промывают дважды при 60°C в 1 × SSC, содержащем 0,1% ДСН.
Выявление полос, которые гибридизовались с зондом, проводят по методике, описанной в руководстве, с использованием набора для выявления нуклеотидов (DIG Nucleotide Detection Kit) (производство компании Boehringer-Mannheim). В результате было показано, что полоса размером примерно 7 т.н. гибридизовалась с данным зондом.
Хромосомную ДНК из Psycloserpens burtonensis штамм АТСС 700359 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) полностью расщепляют с использованием EcoRI. ДНК размером примерно 7 т.н. разделяют электрофорезом в 0,8% геле агарозы, затем очищают ДНК с использованием набора Gene Clean II Kit (производство компании Funakoshi) и растворяют ДНК в 10 мкл ТЭ. Далее смешивают 4 мкл указанного продукта с pUC118 EcoRI/BAP (производство компании Takara Shuzo) и проводят реакцию лигирования с использованием набора для лигирования ДНК (DNA Ligation Kit Ver.2) (производство компании Takara Shuzo). Смешивают 5 мкл указанной жидкости для реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM-109 (производство компании Takara Shuzo) для трансформации клеток Escherichia coli. Полученные таким образом трансформанты затем вносят в соответствующую твердую среду для получения библиотеки хромосомной ДНК.
Для получения гена пептидобразующего фермента полной длины библиотеку хромосомной ДНК подвергают скринингу по методике гибридизации колоний с использованием указанного выше зонда. Процедура гибридизации колоний описана в руководстве Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии из библиотеки хромосомной ДНК переносят на мембранный фильтр из найлона (Nylon Membrane for Colony and Plaque Hybridization) (производство компании Roche Diagnostics) с последующей обработкой, включающей щелочную денатурацию, нейтрализацию и иммобилизацию. Гибридизацию проводят с использованием набора EASY HYB (производство компании Boehringer-Mannheim). После предварительной гибридизации на фильтре при 37°C в течение 1 часа добавляют указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 37°C в течение 16 часов. Затем фильтр дважды промывают при 60°C в 1 × SSC, содержащем 0,1% ДСН.
Выявление колоний, которые гибридизовались с меченым зондом, проводят с использованием набора для выявления нуклеотидов (DIG Nucleotide Detection Kit) (производство компании Boehringer-Mannheim) по методике, описанной в инструкции к набору. В результате было показано, что один штамм колоний гибридизовался с меченым зондом.
(5) Последовательность оснований гена пептидобразующего фермента, полученного из Psycloserpens burtonensis штамм АТСС 700359
Плазмиды, имеющиеся в Escherichia coli JM 109, получают из указанного выше штамма микробных клеток, в отношении которых было показано, что они гибридизуются с меченым зондом, и определяют соответствующую последовательность оснований, которая гибридизовалась с зондом. Проводят реакцию секвенирования с использованием набора CEQ DTCS-Quick Start Kit (производство компании Beckman-Coulter) по методике, описанной в инструкции к набору. Дополнительно проводят электрофорез с использованием CEQ 2000-XL (производство компании Beckman-Coulter).
В результате было показано наличие открытой рамки считывания, которая кодирует пептидобразующий фермент. Последовательность оснований гена пептидобразующего фермента полной длины, полученная из Psycloserpens burtonensis штамм АТСС 700359 (Депонирующий Институт: Американская Коллекция типовых культур (American Type Culture Collection), Адрес Депонирующего Института: P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), а также соответствующая им аминокислотная последовательность показаны в SEQ ID NO:31 в перечне последовательностей.
Промышленное применение
В соответствии с настоящим изобретением предлагается новый фермент, который может использоваться для простого, недорогого получения пептида с высоким выходом, за счет снижения сложности методов синтеза, таких как введение и удаление защитных групп. Применение фермента по настоящему изобретению позволяет осуществлять эффективное промышленное получение пептида.
Перечень последовательностей
SEQ ID NO:3: Синтетический праймер 1
SEQ ID NO:4: Синтетический праймер 2
SEQ ID NO:5: Ген, кодирующий пептидобразующий фермент
SEQ ID NO:7: Синтетический праймер для получения pTrpT
SEQ ID NO:8: Синтетический праймер для получения pTrpT
SEQ ID NO:9: Синтетический праймер для получения pTrpT_Gtg2
SEQ ID NO:10: Синтетический праймер для получения pTrpT_Gtg2
SEQ ID NO:11: Ген, кодирующий пептид-образующий фермент
SEQ ID NO:13: Синтетический праймер для получения pTrpT_Sm_aet
SEQ ID NO:14: Синтетический праймер для получения pTrpT_Sm_aet
SEQ ID NO:15: Смешанный праймер 1 для Aet
SEQ ID NO:16: Смешанный праймер 2 для Aet
SEQ ID NO:19: Праймер 1 для конструирования векторов экспрессии aet, полученных из Pedobacter
SEQ ID NO:20: Праймер 2 для конструирования векторов экспрессии aet, полученных из Pedobacter
SEQ ID NO:21: Смешанный праймер 3 для Aet
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АЛЬФА-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИН-БЕТА-ЭФИРА И СПОСОБ ПОЛУЧЕНИЯ АЛЬФА-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИН-АЛЬФА-МЕТИЛОВОГО ЭФИРА | 2004 |
|
RU2312149C2 |
| ГЕН ПЕПТИДООБРАЗУЮЩЕГО ФЕРМЕНТА, ПЕПТИДООБРАЗУЮЩИЙ ФЕРМЕНТ И СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА | 2002 |
|
RU2280077C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПРОИЗВОДНОГО ГЛЮТАМАТА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ МОНАТИНА | 2002 |
|
RU2280078C2 |
| СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА (ВАРИАНТЫ) | 2005 |
|
RU2316596C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИНДОЛ-3-ПИРОВИНОГРАДНОЙ КИСЛОТЫ И ЕЕ ПРОИЗВОДНЫХ | 2006 |
|
RU2325442C2 |
| ПОЛИПЕПТИД С АКТИВНОСТЬЮ ФОСФОЛИПАЗЫ И ЕГО ПРИМЕНЕНИЕ | 2004 |
|
RU2363731C2 |
| СПОСОБ ПРОДУКЦИИ 2-АМИНО-3-МЕТИЛ-4-КЕТОПЕНТАНОАТА | 2007 |
|
RU2392323C2 |
| Микроорганизм для получения L-аминокислоты с повышенной активностью α-глюкозидазы и способ получения L-аминокислоты с его использованием | 2019 |
|
RU2732338C1 |
| ДНК, КОДИРУЮЩАЯ ДИПЕПТИД-СИНТЕЗИРУЮЩИЙ ФЕРМЕНТ (ВАРИАНТЫ), БАКТЕРИЯ РОДА Escherichia И СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДОВ С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2013 |
|
RU2560980C2 |
| СПОСОБ КОНСТРУИРОВАНИЯ БАКТЕРИИ-ПРОДУЦЕНТА (2S,3R,4S)-4-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА, БАКТЕРИЯ-ПРОДУЦЕНТ (2S,3R,4S)-4-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА И СПОСОБ ПРОДУКЦИИ (2S,3R,4S)-ГИДРОКСИ-L-ИЗОЛЕЙЦИНА ИЛИ ЕГО СОЛИ | 2007 |
|
RU2395580C2 |
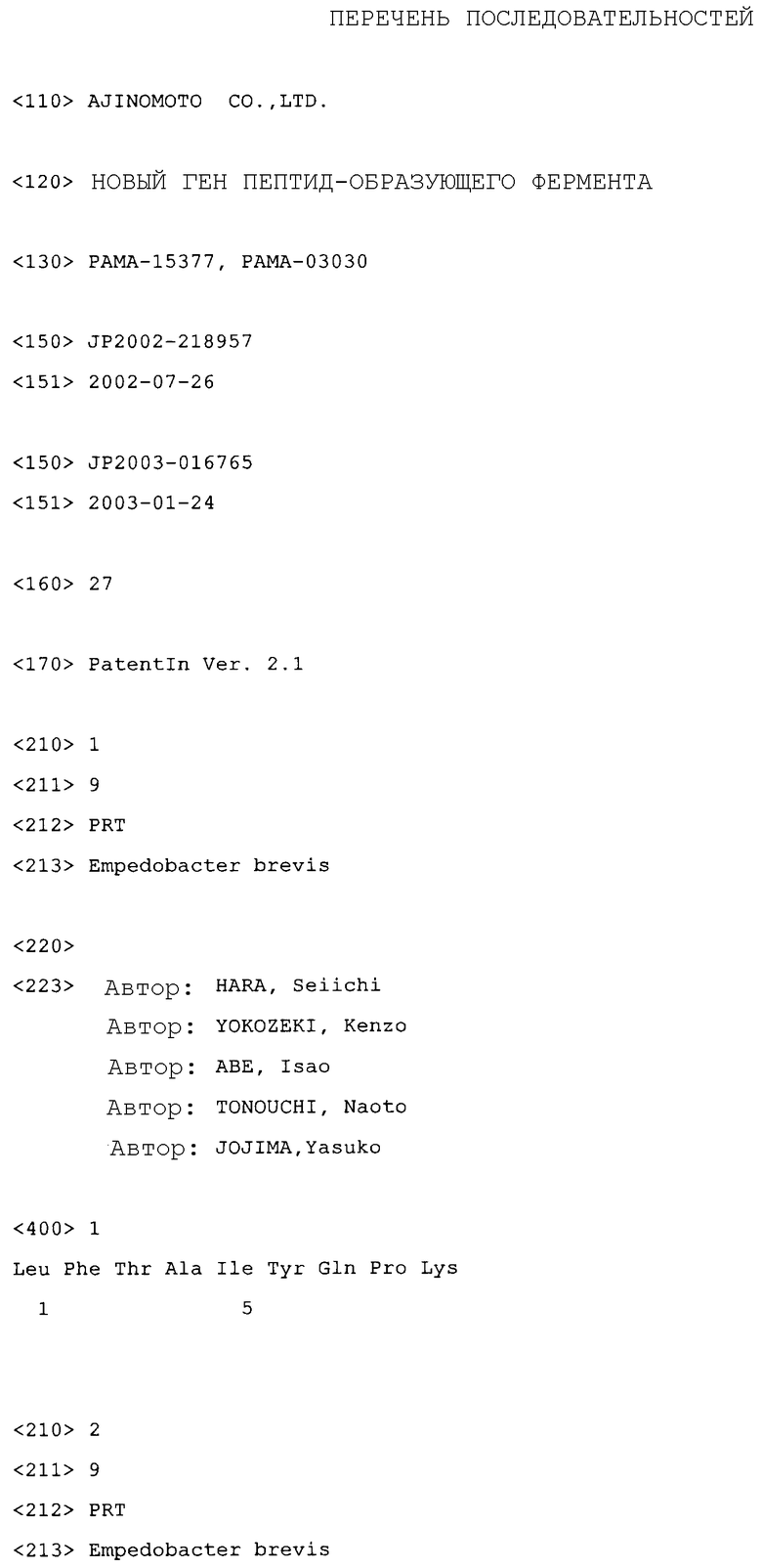







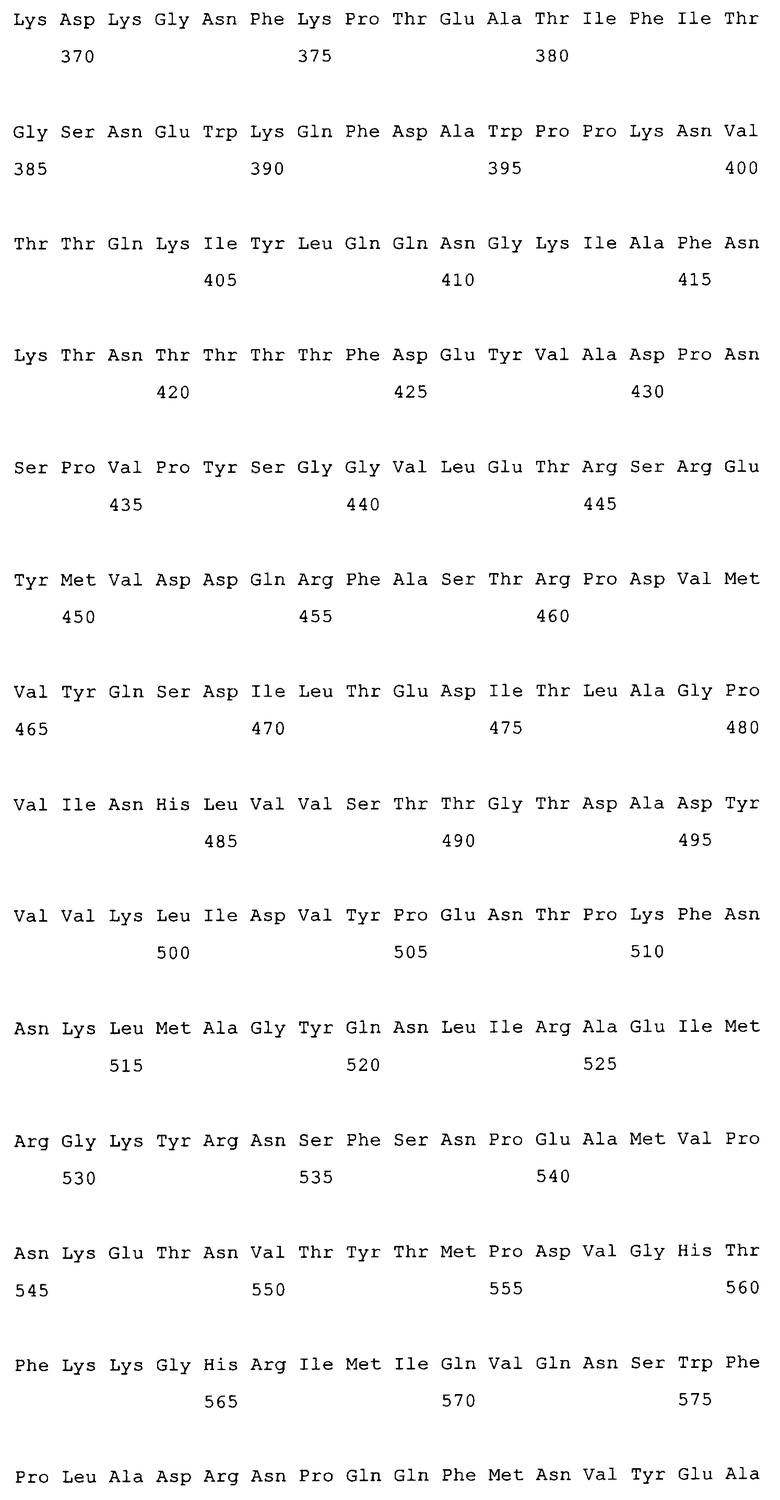







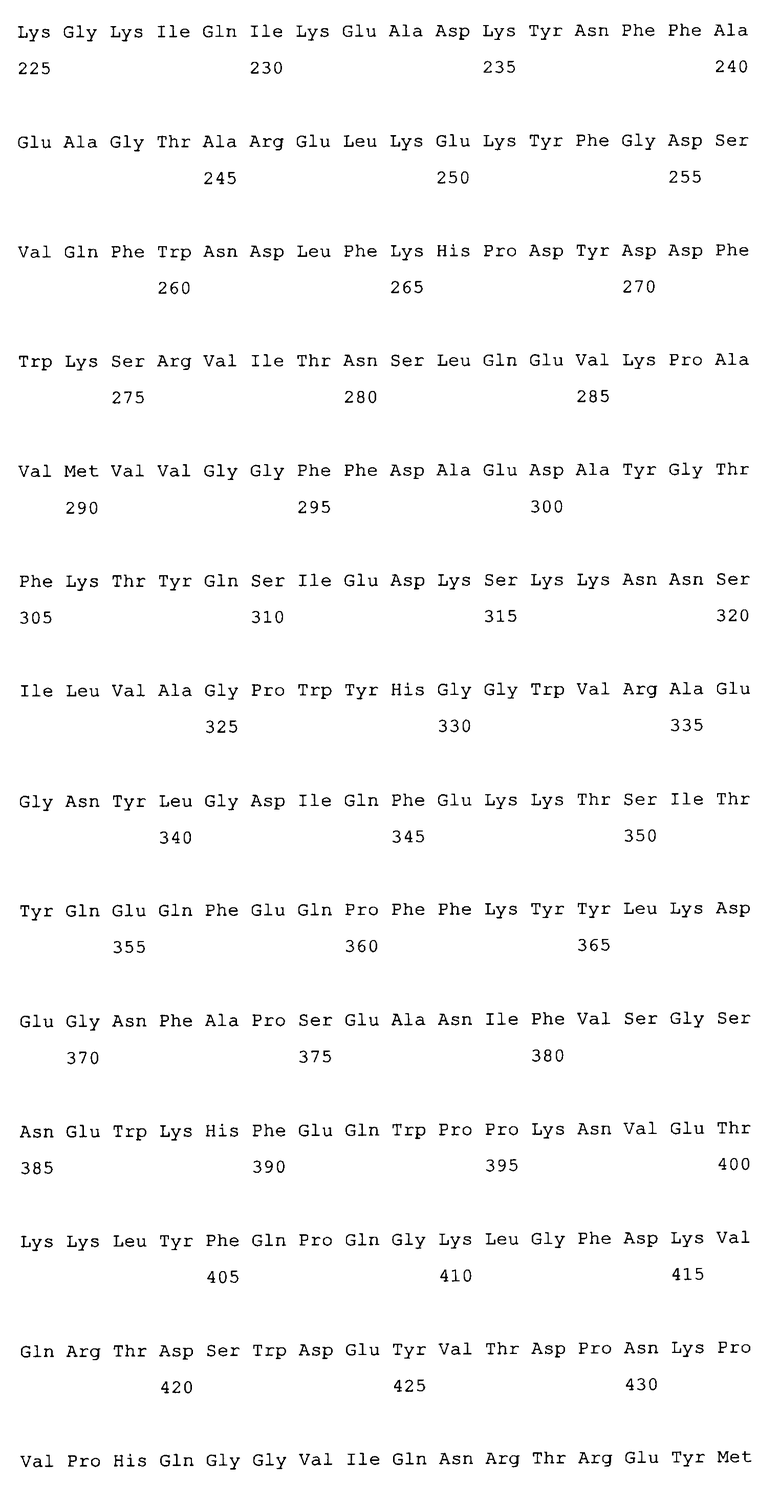






































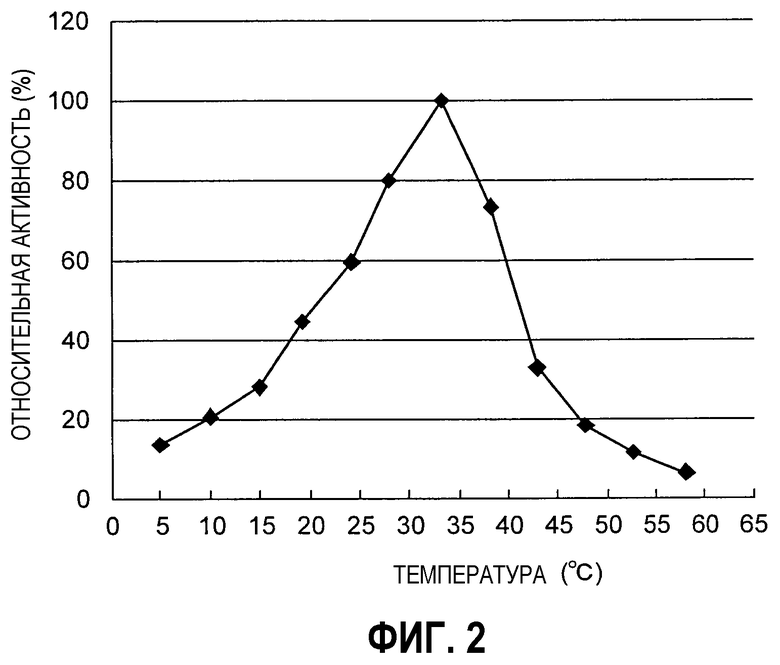
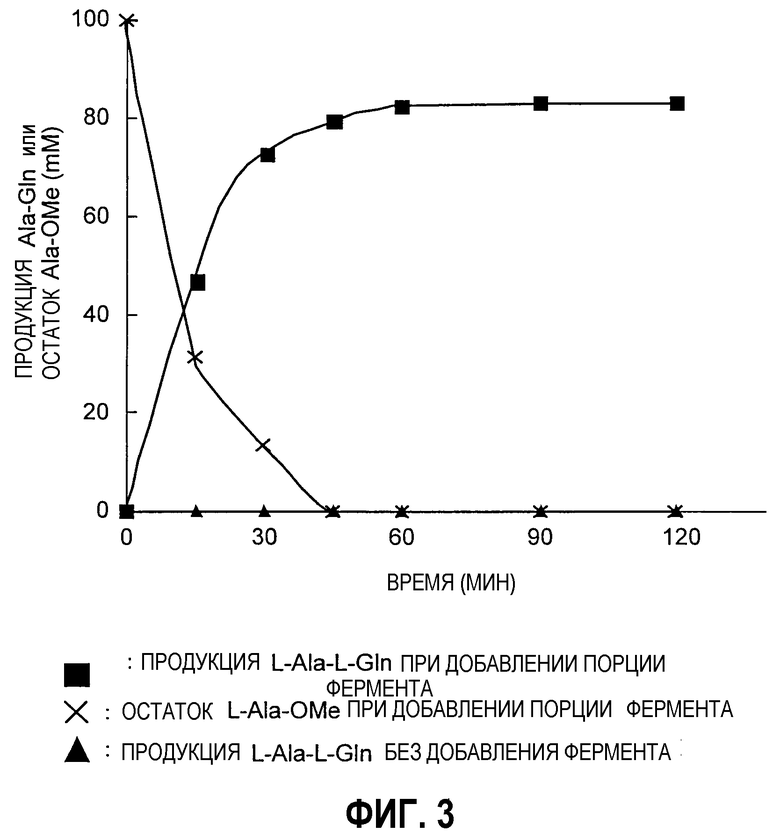
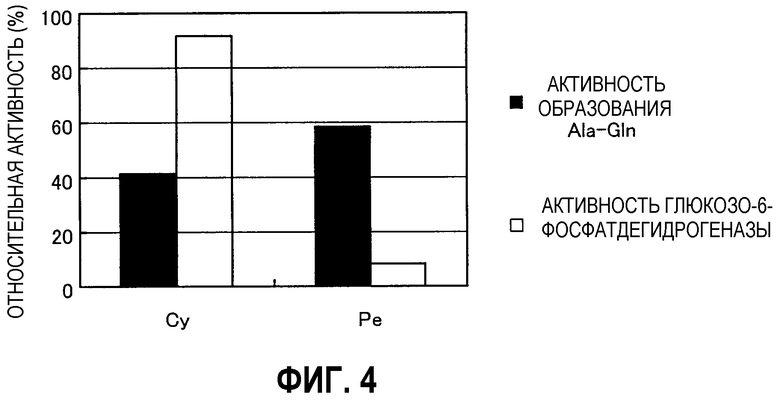
Изобретение относится к биотехнологии и представляет собой новый фермент, способный катализировать реакцию пептидного синтеза из карбоксильного компонента и аминокомпонента. Данный фермент получают культивированием бактериальной клетки-хозяина, которая трансформирована ДНК, кодирующей пептидобразующий фермент, с последующим накоплением данного фермента. Для получения дипептида полученную культуру смешивают с карбоксильным компонентом и аминокомпонентом. Данное изобретение позволяет получать пептиды с высоким выходом без осуществления сложной методики синтеза. 24 н. и 14 з.п. ф-лы, 4 ил, 18 табл.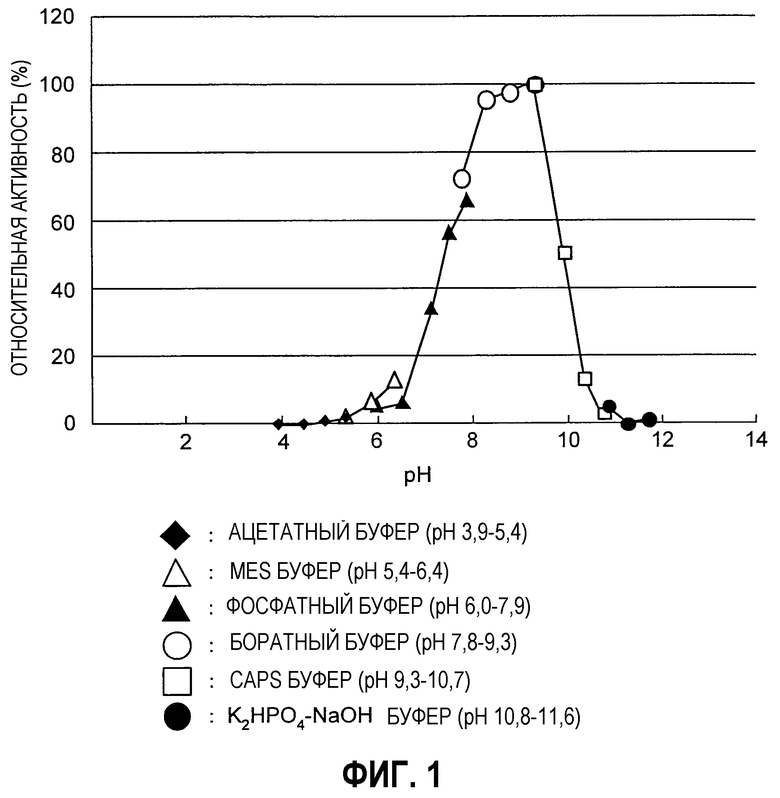
(А) аминокислотная последовательность, состоящая из аминокислотных остатков с номерами 23-616 в SEQ ID NO:6,
(С) аминокислотная последовательность, состоящая из аминокислотных остатков с номерами 21-619 в SEQ ID NO:12,
(Е) аминокислотная последовательность, состоящая из аминокислотных остатков с номерами 23-625 в SEQ ID NO:18,
(G) аминокислотная последовательность, состоящая из аминокислотных остатков с номерами 23-645 в SEQ ID NO:23,
(I) аминокислотная последовательность, состоящая из аминокислотных остатков с номерами 26-620 в SEQ ID NO:25,
(К) аминокислотная последовательность, состоящая из аминокислотных остатков с номерами 18-644 в SEQ ID NO:27,
(М) аминокислотная последовательность, состоящая из SEQ ID NO:6,
(О) аминокислотная последовательность, состоящая из SEQ ID NO:12,
(Q) аминокислотная последовательность, состоящая из SEQ ID NO:18,
(S) аминокислотная последовательность, состоящая из SEQ ID NO:23,
(U) аминокислотная последовательность, состоящая из SEQ ID NO:25, или
(W) аминокислотная последовательность, состоящая из SEQ ID NO:27.
(В) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотных остатках с номерами 23-616 в SEQ ID NO:6, и который обладает пептид-образующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированному участку с аминокислотными остатками 23-616 в SEQ ID NO:6 при 50°С и рН 8,
(D) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотных остатках с номерами 21-619 в SEQ ID NO:12, и который обладает пептид-образующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированному участку с аминокислотными остатками 21-619 в SEQ ID NO:12 при 50°С и рН 8,
(F) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотных остатках с номерами 23-625 в SEQ ID NO:18, и который обладает пептид-образующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированному участку с аминокислотными остатками 23-625 в SEQ ID NO: 18 при 50°С и рН 8,
(Н) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотных остатках с номерами 23-645 в SEQ ID NO:23, и который обладает пептид-образующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированному участку с аминокислотными остатками 23-645 в SEQ ID NO:23 при 50°С и рН 8,
(J) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотных остатках с номерами 26-620 в SEQ ID NO:25, и который обладает пептид-образующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированному участку с аминокислотными остатками 26-620 в SEQ ID NO:25 при 50°С и рН 8,
(L) белок, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотных остатках с номерами 18-644 в SEQ ID NO:27, и который обладает пептид-образующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированному участку с аминокислотными остатками 18-644 в SEQ ID NO:27 при 50°С и рН 8,
(N) участок зрелого белка, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из SEQ ID NO:6, и который обладает пептидобразующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированной SEQ ID NO:6 при 50°С и рН 8,
(Р) участок зрелого белка, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из SEQ ID NO:12, и который обладает пептидобразующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированной SEQ ID NO:12 при 50°С и рН 8,
(R) участок зрелого белка, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из SEQ ID NO:18, и который обладает пептидобразующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированной SEQ ID NO:18 при 50°С и рН 8,
(Т) участок зрелого белка, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из SEQ ID NO:23, и который обладает пептидобразующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированной SEQ ID NO:23 при 50°С и рН 8,
(V) участок зрелого белка, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из SEQ ID NO:25, и который обладает пептидобразующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированной SEQ ID NO:25 при 50°С и рН 8,
(X) участок зрелого белка, имеющий аминокислотную последовательность, включающую замещение, делецию, вставку, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из SEQ ID NO:27, и который обладает пептидобразующей активностью по меньшей мере на уровне 50% относительно белка, соответствующего немутированной SEQ ID NO:27 при 50°С и рН 8.
(а) последовательность оснований, состоящая из оснований с номерами 127-1908 в SEQ ID NO:5,
(с) последовательность оснований, состоящая из оснований с номерами 121-1917 в SEQ ID NO:11,
(е) последовательность оснований, состоящая из оснований с номерами 127-1935 в SEQ ID NO:17,
(g) последовательность оснований, состоящая из оснований с номерами 127-1995 в SEQ ID NO:22,
(i) последовательность оснований, состоящая из оснований с номерами 104-1888 в SEQ ID NO:24,
(k) последовательность оснований, состоящая из оснований с номерами 112-1992 в SEQ ID NO:26,
(m) последовательность оснований, состоящая из оснований с номерами 61-1908 в SEQ ID NO:5,
(о) последовательность оснований, состоящая из оснований с номерами 61-1917 в SEQ ID NO:11,
(q) последовательность оснований, состоящая из оснований с номерами 61-1935 в SEQ ID NO:17,
(s) последовательность оснований, состоящая из оснований с номерами 61-1995 в SEQ ID NO:22,
(u) последовательность оснований, состоящая из оснований с номерами 29-1888 в SEQ ID NO:24, или
(w) последовательность оснований, состоящая из оснований с номерами 61-1992 в SEQ ID NO:26.
культивирование трансформированной клетки по п.20 в среде в течение периода времени и в условиях, подходящих для продуцирования пептидобразующего фермента, и накопление пептидобразующего фермента в среде и/или в трансформированной клетке.
(b) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 127-1908 в SEQ ID NO:5, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 127-1908 в SEQ ID NO:5,
(d) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 121-1917 в SEQ ID NO:11, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 121-1917 в SEQ ID NO:11,
(f) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 127-1935 в SEQ ID NO:17, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 127-1935 в SEQ ID NO:17,
(h) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 127-1995 в SEQ ID NO:22, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 127-1995 в SEQ ID NO:22,
(i) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 104-1888 в SEQ ID NO:24, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°C и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 104-1888 в SEQ ID NO:24,
(l) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 112-1992 в SEQ ID NO:26, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 112-1992 в SEQ ID NO:26,
(n) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 61-1908 в SEQ ID NO:5, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 61-1908 в SEQ ID NO:5,
(р) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 61-1917 в SEQ ID NO:11, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 61-1917 в SEQ ID NO:11,
(г) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 61-1935 в SEQ ID NO:17, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 61-1935 в SEQ ID NO:17,
(t) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 61-1995 в SEQ ID NO:22, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 61-1995 в SEQ ID NO:22,
(v) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 29-1888 в SEQ ID NO:24, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 29-1888 в SEQ ID NO:24, или
(х) последовательность оснований, которая гибридизуется в строгих условиях с ДНК, имеющей последовательность оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 61-1992 в SEQ ID NO:26, и которая кодирует белок, обладающий пептидобразующей активностью на уровне по меньшей мере 50% при 50°С и рН 8 относительно белка, кодируемого немутированной последовательностью оснований с номерами 61-1992 в SEQ ID NO:26.
Приоритет:
| Многоярусная сушилка шкафного типа | 1938 |
|
SU58478A1 |
| ДИПЕПТИДИЛАМИНОПЕПТИДАЗА И СПОСОБ ФЕРМЕНТАТИВНОГО ОТЩЕПЛЕНИЯ N-КОНЦЕВЫХ ДИПЕПТИДОВ MET-TYR, MET-ARG ИЛИ MET-ASP ОТ ПОЛИПЕПТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ | 1993 |
|
RU2096455C1 |

