Область техники, к которой относится изобретение
Настоящее изобретение относится к способу получения α-L-аспартил-L-фенилаланин-β-эфира (также называемого "α-L-(β-o-замещенный аспартил)-L-фенилаланин" (сокращение: α-ARP)) и к способу получения α-L-аспартил-L-фенилаланин-α-метилового эфира (также называемого "α-L-аспартил-L-фенилаланин метилового эфира (сокращение: α-APM). Более конкретно, настоящее изобретение относится к способу получения α-L-аспартил-L-фенилаланин-β-эфира, который является важным промежуточным продуктом для получения α-L-аспартил-L-фенилаланин-α-метилового эфира (название продукта: аспартам), который пользуется большим спросом в качестве подсластителя, и к способу получения α-L-аспартил-L-фенилаланин-α-метилового эфира, используя способ получения α-L-аспартил-L-фенилаланин-β-эфира.
Предшествующий уровень техники
Общеизвестные способы получения α-L-аспартил-L-фенилаланин-α-метилового эфира (далее здесь "α-APM" для сокращения в некоторых случаях) включают способ химического синтеза и способ ферментативного синтеза. Что касается способа химического синтеза, то известен способ конденсации N-защищенного ангидрида L-аспарагиновой кислоты с метиловым эфиром L-фенилаланина с получением N-защищенного APM и удаления N-защитной группы с получением APM, а что касается способа ферментативного синтеза, то известен способ конденсации N-защищенной L-аспарагиновой кислоты с метиловым эфиром L-фенилаланина с получением N-защищенного APM и удаления N-защитной группы с получением APM. В обоих способах, однако, необходимы стадии введения и удаления защитной группы, и процессы являются очень затруднительными. С другой стороны, был разработан способ получения APM без использования N-защитной группы (смотри патентную публикацию Японии № H02-015196 Gazette). Однако этот способ не подходит для промышленного получения из-за очень низкого выхода продукта. Таким образом, в связи с этим желательно разработать недорогой способ промышленного получения аспартама.
Раскрытие изобретения
Целью настоящего изобретения является простой недорогой способ получения α-L-аспартил-L-фенилаланин-β-эфира, который является промежуточным продуктом α-L-аспартил-L-фенилаланин-α-метилового эфира, обеспечивающий высокий выход, не используя сложный способ синтеза. Кроме того, целью настоящего изобретения является разработка простого недорогого способа получения α-L-аспартил-L-фенилаланин-α-метилового эфира, обеспечивающего высокий выход.
В результате проведенных исследований в отношении вышеуказанных целей изобретателями настоящего изобретения было обнаружено, что недавно открытый фермент или вещество, содержащее фермент, способны к избирательному синтезу α-L-аспартил-L-фенилаланин-β-эфира из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина, что и привело к осуществлению настоящего изобретения.
Конкретно, настоящее изобретение описано ниже.
[1] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенный аспартил)-L-фенилаланина), включающий получение α-L-аспартил-L-фенилаланин-β-эфира из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина с помощью фермента или содержащего фермент вещества, которые способны избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[2] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [1] выше, где фермент или содержащее фермент вещество представляют собой один или два или несколько типов, выбранных из группы, включающей культуру микроорганизма, который обладает способностью избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь, клетку микроорганизма, выделенную из культуры, и обработанного продукта микроорганизмом клетки микроорганизма.
[3] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2] выше, в котором микроорганизм представляет собой микроорганизм, принадлежащий к роду, выбранному из группы, включающей Aeromonas, Azotobacter, Alcaligenes, Brevibacterium, Corynebacterium, Escherichia, Empedobacter, Flavobacterium, Microbacterium, Propionibacterium, Brevibacillus, Paenibacillus, Pseudomonas, Serratia, Stenotrophomonas, Sphingobacterium, Streptomyces, Xanthomonas, Williopsis, Candida, Geotrichum, Pichia, Saccharomyces, Torulaspora, Cellulophaga, Weeksell, Pedobacter, Persicobacter, Flexithrix, Chitinophaga, Cyclobacterium, Runella, Thermonema, Psychroserpens, Gelidibacter, Dyadobacter, Flammeovirga, Spirosoma, Flectobacillus, Tenacibaculum, Rhodotermus, Zobellia, Muricauda, Salegentibacter, Taxeobacter, Cytophaga, Marinilabilia, Lewinella, Saprospira, и Haliscomenobacter.
[4] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2] выше, где микрорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (A) или (B):
(A) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 23 по 616 аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей,
(B) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с 23 по 616 аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[5] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2], где микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (C) или (D):
(C) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 21 по 619 аминокислотной последовательности, представленной в SEQ ID NO: 12 перечня последовательностей,
(D) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с 21 по 619 аминокислотной последовательности, представленной в SEQ ID NO: 12 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[6] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2], в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (E) или (F), ниже:
(E) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 23 по 625 аминокислотной последовательности, представленной в SEQ ID NO: 18 перечня последовательностей,
(F) белок с аминокислотной последовательностью, включающей замену, делецию, исерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с 23 по 625 аминокислотной последовательности, представленной в SEQ ID NO: 18 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[7] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2] выше, в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (G) или (H), ниже:
(G) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 23 по 645 аминокислотной последовательности, представленной в SEQ ID NO: 23 перечня последовательностей,
(H) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с 23 по 645 аминокислотной последовательности, представленной в SEQ ID NO: 23 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[8] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2], в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (I) или (J), ниже:
(I) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 26 по 620 аминокислотной последовательности, представленной в SEQ ID NO: 25 перечня последовательностей,
(J) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с 26 по 620 аминокислотной последовательности, представленной в SEQ ID NO: 25 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[9] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2] выше, в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (K) или (L), ниже:
(K) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 18 по 644 аминокислотной последовательности, представленной в SEQ ID NO: 27 перечня последовательностей,
(L) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с 18 по 644 аминокислотной последовательности, представленной в SEQ ID NO: 27 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[10] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2], в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (M) или (N), ниже:
(M) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 6 перечня последовательностей,
(N) белок, содержащий область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[11] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2], в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (O) или (P), ниже:
(O) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 12 перечня последовательностей,
(P) белок, содержащий область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 12 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[12] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно п.[2], в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (Q) или (R), ниже:
(Q) белок, содержащий область зрелого белка с аминокислотной последовательностью, представленной в SEQ ID NO: 18 перечня последовательностей,
(R) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 18 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[13] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [3], в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (S) или (T), ниже:
(S) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 23 перечня последовательностей,
(T) белок, содержащий область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 23 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[14] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2], в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (U) или (V), ниже:
(U) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 25 перечня последовательностей,
(V) белок, содержащий область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 25 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[15] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [2] выше, в котором микроорганизм представляет собой трансформированный микроорганизм, который способен экспрессировать белок (W) или (X), ниже:
(W) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 27 перечня последовательностей,
(X) белок, содержащий область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 27 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[16] Способ получения α-L-аспартил-L-фенилаланин-β-эфира (т.е. α-L-(β-o-замещенного аспартил)-L-фенилаланина) согласно [1] выше, в котором фермент является, по меньшей мере, одним из белков, выбранных из группы, включающей (A)-(X), ниже:
(A) белка с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 23 по 616 аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей,
(B) белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 23 по 616 аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(C) белка с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 21 по 619 аминокислотной последовательности, представленной в SEQ ID NO: 12 перечня последовательностей,
(D) белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 21 по 619 аминокислотной последовательности, представленной в SEQ ID NO: 12 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(E) белка с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 23 по 625 аминокислотной последовательности, представленной в SEQ ID NO: 18 перечня последовательностей,
(F) белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 23 по 625 аминокислотной последовательности, представленной в SEQ ID NO: 18 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(G) белка с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 23 по 645 аминокислотной последовательности, представленной в SEQ ID NO: 23 перечня последовательностей,
(H) белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 23 по 645 аминокислотной последовательности, представленной в SEQ ID NO: 23 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(I) белка с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 26 по 620 аминокислотной последовательности, представленной в SEQ ID NO: 25 перечня последовательностей,
(J) белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 26 по 620 аминокислотной последовательности, представленной в SEQ ID NO: 25 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(K) белка с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 18 по 644 аминокислотной последовательности, представленной в SEQ ID NO: 27 перечня последовательностей,
(L) белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 18 по 644 аминокислотной последовательности, представленной в SEQ ID NO: 27 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(M) белка с аминокислотной последовательностью, представленной в SEQ ID NO: 6 перечня последовательностей,
(N) белка, содержащего область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(O) белка с аминокислотной последовательностью, представленной в SEQ ID NO: 12 перечня последовательностей,
(P) белка, содержащего область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 12 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(Q) белка с аминокислотной последовательностью, представленной в SEQ ID NO: 18 перечня последовательностей,
(R) белка, содержащего область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 18 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(S) белка с аминокислотной последовательностью, представленной в SEQ ID NO: 23 перечня последовательностей,
(T) белка, содержащего область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 23 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(U) белка с аминокислотной последовательностью, представленной в SEQ ID NO: 25 перечня последовательностей,
(V) белка, содержащего область зрелого белка с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 25 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(W) белка с аминокислотной последовательностью, представленной в SEQ ID NO: 27 перечня последовательностей, и
(X) белка, содержащего область зрелого белка, с аминокислотной последовательностью в аминокислотной последовательности, представленной в SEQ ID NO: 27 перечня последовательностей, и способного избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
[17] Способ получения α-L-аспартил-L-фенилаланин-α-метилового эфира (т.е. метилового эфира α-L-аспартил-L-фенилаланина), включающий: реакционную стадию синтеза α-L-аспартил-L-фенилаланин-β-метилового эфира (называемого также α-L-(β-о-метиласпартил)-L-фенилаланин (сокращение: α-AMP) c помощью способа получения α-L-аспартил-L-фенилаланин-β-эфира согласно любому из пунктов 1-16; и реакционную стадию преобразования α-L-аспартил-L-фенилаланин-β-метилового эфира (т.е. α-L-(β-о-метиласпартил)-L-фенилаланин) в α-L-аспартил-L-фенилаланин-α-метиловый эфир.
α-L-аспартил-L-фенилаланин-β-эфир может быть легко получен с помощью настоящего изобретения. α-L-аспартил-L-фенилаланин-β-эфир может быть легко получен и иметь высокий выход наряду со сниженным использованием сложных способов синтеза, таких как введение/удаление защитных групп с помощью способа настоящего изобретения.
Более того, с помощью настоящего изобретения α-L-аспартил-L-фенилаланин-α-метиловый эфир может быть легко получен и может иметь высокий выход при минимуме затрат.
Другие объекты, особенности и преимущества настоящего изобретения будут подробно описаны или станут очевидны из нижеследующего описания, сопровождающегося фигурами.
Краткое описание фигуры
На чертеже представлена диаграмма, отражающая количество ферментов, которое существует в цитоплазматической фракции (Cy) и фракции периплазмы (Pe).
Наилучший вариант осуществления изобретения
Далее настоящее изобретение будет описано в следующем порядке
<1> Способ получения α-L-аспартил-L-фенилаланин-β-эфира
1. Способ получения α-L-аспартил-L-фенилаланин-β-эфира
2. Микроорганизмы, используемые в настоящем изобретении
3. Ферменты, используемые в настоящем изобретении; и
<2> Способ получения α-L-аспартил-L-фенилаланин-α-метилового эфира.
<1> Способ получения α-L-аспартил-L-фенилаланин-β-эфира
1. Способ получения α-L-аспартил-L-фенилаланин-β-эфира
В способе получения α-L-аспартил-L-фенилаланин-β-эфира по настоящему изобретению (называемом здесь также «способом получения пептида по настоящему изобретению») L-фенилаланин и α,β-диэфир L-аспарагиновой кислоты взаимодействуют в присутствии фермента, обладающего известной пептид-образующей активностью. То есть способ получения пептида по настоящему изобретению заключается в том, что α-L-аспартил-L-фенилаланин-β-эфир образуется из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина с помощью фермента или содержащего фермент вещества, способных избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь. Фермент или содержащее фермент вещество, способные избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты, относятся к ферменту или содержащему фермент веществу, обладающим способностью или активностью катализировать реакцию, в которой по существу L-фенилаланин не способен осуществлять нуклеофильное воздействие на β-эфирный участок α,β-диэфира L-аспарагиновой кислоты, но осуществляет нуклеофильное воздействие только на его α-эфирный участок. Как показано здесь ниже в ссылочном примере, однако, получены также фермент или содержащее фермент вещество, которые обладают способностью катализировать реакцию, в которой по существу L-фенилаланин не способен осуществлять воздействие на α-эфирный участок α,β-диэфира L-аспарагиновой кислоты, но осуществляет нуклеофильное воздействие только на его β-эфирный участок в отличие от указанной выше способности, и что дает β-L-аспартил-L-фенилаланин-α-эфир (называемый также β-L-(α-o-замещенный аспартил)-L-фенилаланин (сокращение: β-ARP) из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина.
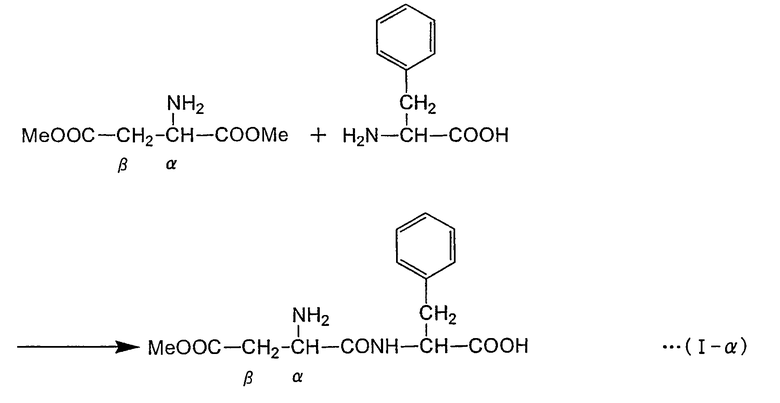
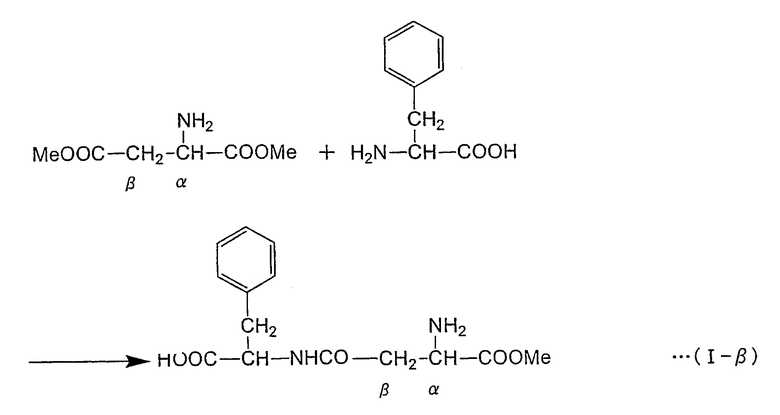
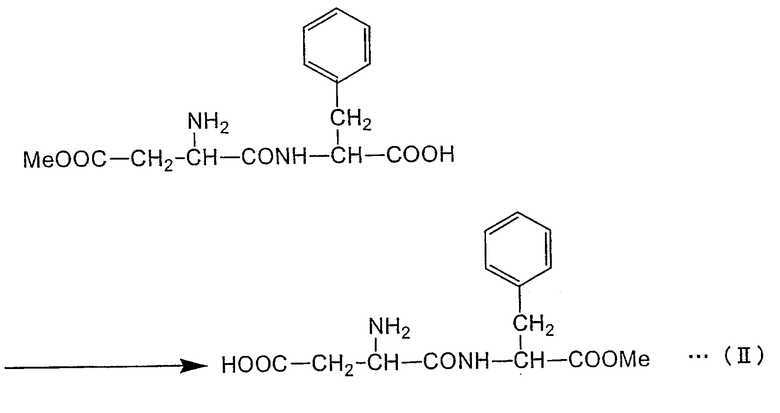
Формула реакции, в которой L-фенилаланин осуществляет нуклеофильное воздействие на α-эфирный участок α,β-диэфира L-аспарагиновой кислоты с получением α-L-аспартил-L-фенилаланин-β-эфира (сокращение: α-ARP), представлена в нижеследующей формуле (I-α) (где "Me" представляет собой метильную группу) путем цитирования случая, в котором в качестве α,β-диэфира L-аспарагиновой кислоты применяют α,β-диметиловый эфир L-аспарагиновой кислоты. Как показано в формуле (I-α), в способе получения пептида по настоящему изобретению аминогруппа L-фенилаланина взаимодействует с α-метиловым эфирным участком α,β-диметилового эфира L-аспарагиновой кислоты с образованием пептидной связи. С другой стороны, в нижепредставленной формуле (I-β) указана реакция, в которой β-метиловый эфирный участок α,β-диметилового эфира L-аспарагиновой кислоты подвергается нуклеофильному воздействию с образованием β-L-аспартил-L-фенилаланин-α-метилового эфира (называемого также β-L-(α-o-метил-аспартил)-L-фенилаланин (сокращение: β-AMP)). Пептидная связь в β-AMP образуется по β-метиловому эфирному участку α,β-диметилового эфира L-аспарагиновой кислоты. Фермент или содержащее фермент вещество, использующиеся в настоящем изобретении, ускоряют по существу только реакцию, как в формуле (I-α), но по существу не вызывают реакции как в формуле (I-β). α-APM может быть получен из α-AMP через простую стадию реакции (формула (II)), но α-APM не может быть получен непосредственно из β-AMP. Таким образом, способ по настоящему изобретению крайне эффективен в качестве способа получения промежуточного продукта α-APM и может использоваться для промышленного получения.
Формула реакции I-α
Формула реакции I-β
Формула реакции II
Способ, позволяющий ферменту или содержащему фермент веществу взаимодействовать с α,β-диэфиром L-аспарагиновой кислоты и L-фенилаланином, может быть осуществлен путем смешивания фермента или содержащего фермент вещества с α,β-диэфиром L-аспарагиновой кислоты и L-фенилаланином. Более конкретно, может быть использован способ, в котором фермент или содержащее фермент вещество добавляют к раствору, содержащему диэфир L-аспарагиновой кислоты и L-фенилаланин для воздействия на реакцию. Когда используют микроорганизм в качестве содержащего фермент вещества, который продуцирует фермент, реакция может быть осуществлена либо как описано выше, либо может быть использован способ, который включает культивирование микроорганизма, который продуцирует фермент для получения и аккумуляции фермента в микроорганизме или культуральной жидкости, в которой культивируется микроорганизм, и добавление к культуральной жидкости α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина, или сходный с ним способ. Полученный таким образом α-L-аспартил-L-фенилаланин-β-эфир выделяют общепринятым способом, и, при необходимости, он может быть очищен.
«Содержащее фермент вещество» может быть любым веществом, если оно содержит фермент, и конкретные его типы включают культуру микроорганизма, который продуцирует фермент, клетку микроорганизма, выделенную из культуры, и обработанный продукт клетки микроорганизма. Под культурой микроорганизма подразумевают вещество, полученное путем культивирования микроорганизма, и, конкретно, смесь клеток микроорганизма, среды, используемой для культивирования микроорганизма, и вещества, продуцируемого культивируемым микроорганизмом, и так далее. Кроме того, клетка микроорганизма может быть очищена для применения в качестве очищенной клетки микроорганизма. Более того, обработанный продукт клетки микроорганизма включает продукты, которые получают путем разрушения, лизиса или замораживания-оттаивания клетки микроорганизма и дальнейшего восстановления неочищенного фермента путем обработки клетки микроорганизма, и очистки фермента путем дальнейшей очистки. В качестве фермента, обработанного для очистки, может быть применен частично очищенный фермент, полученный с помощью различных способов очистки и так далее. Кроме того, могут быть использованы иммобилизованные ферменты, которые иммобилизованы с помощью метода образования ковалентных связей, метода адсорбции, метода улавливания и так далее. Кроме того, в случае некоторых используемых микроорганизмов часть клеток микроорганизма может подвергаться лизису в процессе культивирования и в таком случае в качестве содержащего фермент вещества также может быть использован супернатант культуральной жидкости.
Кроме того, в качестве микроорганизма, который содержит фермент, может быть использован штамм дикого типа или может быть использован рекомбинантный штамм, в котором экспрессируется фермент. Такой микроорганизм не ограничивается клеткой микроорганизма, продуцирующей фермент, а также могут использоваться обработанные продукты клетки микроорганизма, такие как обработанная ацетоном клетка микроорганизма и лиофилизованная клетка микроорганизма. Кроме того, могут быть использованы иммобилизованные клетки микроорганизма, полученные путем иммобилизации обработанного продукта клетки микроорганизма с применением метода образования ковалентных связей, метода адсорбции, метода улавливания или тому подобное, или иммобилизованный обработанный клеточный продукт микроорганизмов.
Применение штамма дикого типа, который может продуцировать фермент, образующий пептид и способный образовывать α-L-аспартил-L-фенилаланин-β-эфир, является предпочтительным, так как получение пептида может быть осуществлено более легко без прохождения стадии создания рекомбинантного штамма. С другой стороны, рекомбинантный штамм, который был трансформирован и экспрессирует фермент, образующий пептид и способный образовывать α-L-аспартил-L-фенилаланин-β-эфир, может быть модифицирован таким образом, чтобы фермент, образующий пептид, продуцировался в большем количестве. Таким образом, можно синтезировать α-L-аспартил-L-фенилаланин-β-эфир в большем количестве и с более высокой интенсивностью. Культивирование микроорганизма штамма дикого типа или рекомбинантного штамма в среде для аккумуляции в среде фермента, образующего пептид, и/или микроорганизма, и смешивание аккумулированного таким образом продукта с α,β-диэфиром L-аспарагиновой кислоты и L-фенилаланином может давать α-L-аспартил-L-фенилаланин-β-эфир.
Следует отметить, что, когда используют культивируемые продукты, культивируемые клетки микроорганизма, промытые клетки микроорганизма и обработанные продукты клетки микроорганизма, полученные путем разрушения или лизиса клеток микроорганизма, зачастую присутствует фермент, который разрушает образованный α-L-аспартил-L-фенилаланин-β-эфир, не будучи вовлеченным в образование α-L-аспартил-L-фенилаланин-β-эфира. В таком случае в некоторых случаях предпочтительно добавлять ингибитор металлопротеиназ, такой как этилендиаминтетрауксусная кислота (ЭДТА). Добавляемое количество находится в диапазоне от 0,1 миллимолярного (мМ) до 300 мМ, предпочтительно от 1 мМ до 100 мМ.
Используемое количество фермента или содержащего фермент вещества может быть достаточным, если это количество является количеством, при котором наблюдается требуемый эффект (эффективное количество). Несмотря на то что специалист в данной области техники может легко определить такое эффективное количество путем простого предварительного эксперимента, применяемое количество составляет, например, от приблизительно 0,01 до приблизительно 100 единиц ("Ед") в случае использования фермента и от приблизительно 0,1 до приблизительно 500 г/л в отмытых клеток микроорганизма. Следует отметить, что 1 Ед определяется как количество фермента, которое позволяет продуцировать 1 микромоль (мкмоль) α-L-аспартил-L-фенилаланин-β-метилового эфира из 100 мМ α,β-диметилового эфира L-аспарагиновой кислоты и 200 мМ L-фенилаланина при 25°С в одну минуту.
α,β-Диэфир L-аспарагиновой кислоты, используемый в реакции, может быть любым эфиром, конденсируемым с L-фенилаланином с получением α-L-аспартил-L-фенилаланин-β-эфира. Примеры α,β-диэфира L-аспарагиновой кислоты включают α,β-диметиловый эфир L-аспарагиновой кислоты и α,β-диэтиловый эфир L-аспарагиновой кислоты. Когда взаимодействуют α,β-диметиловый эфир L-аспарагиновой кислоты и L-фенилаланин, то образуется α-L-аспартил-L-фенилаланин-β-метиловый эфир (α-AMP), а когда взаимодействуют α,β-диэтиловый эфир L-аспарагиновой кислоты и L-фенилаланин, то образуется α-L-аспартил-L-фенилаланин-β-этиловый эфир (также называемый α-L-(β-o-этил аспартил)-L-фенилаланин (сокращение: α-AEP).
Тогда как концентрация α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина, которые являются исходными продуктами, в каждом случае составляют от 1 мМ до 10 мМ, предпочтительно от 0,05 М до 2 М, могут быть случаи, когда предпочтительно добавлять один субстрат в эквимолярном или большем количестве по отношению к другому субстрату и по необходимости делать выбор. Кроме того, в тех случаях, когда высокие концентрации субстратов вызывают ингибирование реакции, тогда субстраты добавляют в процессе реакции в концентрации, которая не вызывает ингибирования.
Температура реакции, при которой возможно получение α-L-аспартил-L-фенилаланин-β-эфира, составляет от 0 до 60°С, предпочтительно от 5 до 40°С. Кроме того, рН реакции, при котором возможно получение α-L-аспартил-L-фенилаланин-β-эфира, составляет от 6,5 до 10,5, предпочтительно от 7,0 до 10,0.
2. Микроорганизмы, используемые в настоящем изобретении
В качестве микроорганизмов, используемых в настоящем изобретении, могут быть использованы без особого ограничения те микроорганизмы, которые способны продуцировать α-L-аспартил-L-фенилаланин-β-эфир из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина. Микроорганизмы, которые способны продуцировать α-L-аспартил-L-фенилаланин-β-эфир из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина, включают, например, микроорганизмы, принадлежащие к родам
Aeromonas, Azotobacter, Alcaligenes, Brevibacterium, Corynebacterium, Escherichia, Empedobacter, Flavobacterium, Microbacterium, Propionibacterium, Brevibacillus, Paenibacillus, Pseudomonas, Serratia, Stenotrophomonas, Sphingobacterium, Streptomyces, Xanthomonas, Williopsis, Candida, Geotrichum, Pichia, Saccharomyces, Torulaspora, Cellulophaga, Weeksella, Pedobacter, Persicobacter, Flexithrix, Chitinophaga, Cyclobacterium, Runella, Thermonema, Psychroserpens, Gelidibacter, Dyadobacter, Flammeovirga, Spirosoma, Flectobacillus, Tenacibaculum, Rhodotermus, Zobellia, Muricauda, Salegentibacter, Taxeobacter, Cytophaga, Marinilabilia, Lewinella, Saprospira, Haliscomenobacter
Могут быть приведены следующие конкретные примеры
Aeromonas hydrophila ATCC 13136
Azotobacter vinelandii IFO 3741
Alcaligenes faecalis FERM P-8460
Brevibacterium minutiferuna FERM BP-8277
Corynebacterium flavescens ATCC 10340
Escherichia coli FERM BP-8276
Empedobacter brevis ATCC 14234
Flavobacterium resinovorum ATCC 14231
Microbacterium arborescens ATCC 4348
Propionibacterium shermanii FERM BP-8100
Brevibacillus parabrevis ATCC 8185
Paenibacillus alvei IFO 14175
Pseudomonas fragi IFO 3458
Serratia grimesii ATCC 14460
Stenotrophomonas maltophilia ATCC 13270
Sphingobacterium sp. PERM BP-8124
Streptomyces griseolus NRRL B-1305
(Streptomyces lavendulae)
Xanthomonas maltophilia FERM BP-5568
Williopsis saturnus IFO 0895
Candida magnoliae IFO 0705
Geotrichum fragrance CBS 152.25
(Geotrichum amycelium)
Geotrichum amycelium IFO 0905
Pichia ciferrii IFO 0905
Saccharomyces unisporus IFO 0724
Torulaspora delbrueckii IFO 0422
Cellulophaga lytica NBRC 14961
Weeksella virosa NBRC 16016
Pedobacter heparinus NBRC 12017
Persicobacter diffluens NBRC 15940
Flexithrix dorotheae NBRC 15987
Chitinophaga pinensis NBRC 15968
Cyclobacterium marinum ATCC 25205
Runella slithyformis ATCC 29530
Thermonema lapsum ATCC 43542
Psychroserpens burtonensis ATCC 700359
Gelidibacter algens ATCC 700364
Dyadobacter fermentans ATCC 700827
Flammeovirga aprica NBRC 15941
Spirosoma linguale DSMZ 74
Flectobacillus major DSMZ 103
Tenacibaculum maritimum ATCC 43398
Rhodotermus marinus DSMZ 4252
Zobellia galactanivorans DSMZ 12802
Muricauda ruestringensis DSMZ 13258
Salegentibacter salegens DSMZ 5424
Taxeobacter gelupurpurascens DSMZ 11116
Cytophaga hutchinsonii NBRC 15051
Marinilabilia salmonicolor NBRC 15948
Lewinella cohaerens ATCC 23123
Saprospira grandis ATCC 23119
Haliscomenobacter hydrossis ATCC 27775
Среди указанных выше штаммов микроорганизмов те микроорганизмы, которые обозначены номерами FERM, были депонированы независимой в управлении корпорацией, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов (Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), и могут быть предоставлены при ссылке на каждый номер.
Среди указанных выше штаммов микроорганизмов те микроорганизмы, которые обозначены номерами ATCC, были депонированы американской коллекцией типов культур (P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) и могут быть предоставлены при ссылке на каждый номер.
Среди указанных выше штаммов микроорганизмов те микроорганизмы, которые обозначены номерами IFO, были депонированы институтом ферментации Osaka (2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) и могут быть предоставлены при ссылке на каждый номер.
Среди указанных выше штаммов микроорганизмов те микроорганизмы, которые обозначены номерами NBRC, были депонированы NITE центром биологических источников национального института технологии и вычислений (5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) и могут быть предоставлены при ссылке на каждый номер.
Среди указанных выше штаммов микроорганизмов те микроорганизмы, которые обозначены номерами DSMZ, были депонированы Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур) (Mascheroder Weg 1b, 38124 Braunschweig, Германия) и могут быть предоставлены при ссылке на каждый номер.
Также как указанные выше штаммы, те микроорганизмы, которые обозначены номерами FERM, представляют собой микроорганизмы, которые были депонированы в независимой в управлении корпорации, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов (Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566 Япония. Alcaligenes faecalis FERM P-8460 представляет собой микроорганизм, который задепонирован 30 сентября 1985 г. и которому присвоен номер депозита FERM P-8460. Propionibacterium shermanii FERM P-9737 представляет собой микроорганизм, который был первоначально задепонирован 4 декабря 1987 г., и контроль над данным организмом был в последующем передан международному депозитарию по условиям Будапештского договора от 1 июля 2002 г., и ему присвоен номер депозита FERM BP-8100. Xanthomonas maltophilia FERM BP-5568 представляет собой микроорганизм, который был первоначально задепонирован 14 июня 1995 г., и контроль над данным организмом был в последующем передан международному депозитарию по условиям Будапештского договора от 14 июня 1996 г. Brevibacterium minutiferuna FERM BP-8277 был задепонирован на международное хранение по условиям Будапештского договора от 20 января 2002 г. Esherichia coli FERM BP-8276 был задепонирован на международное хранение по условиям Будапештского договора от 20 января 2002 г.
Empedobacter brevis, штамм ATCC 14234 (штамм FERM P-18545, штамм FERM BP-8113), был задепонирован в международный депозитарий запатентованных организмов независимой в управлении корпорации, национальный институт развития промышленной науки и технологии (Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), 1 октября 2001 г., и ему присвоен номер депозита FERM P-18545. Контроль над данным организмом был в последующем передан депозитарию по условиям Будапештского договора в международном депозитарии запатентованных организмов независимой в управлении корпорации, национальный институт развития промышленной науки и технологии 8 июля 2002 г., и ему был присвоен номер депозита FERM P-8113 (индекс микроорганизма: Empedobacter brevis strain AJ 13933).
Sphingobacterium sp., штамм AJ 110003, был задепонирован в международный депозитарий запатентованных организмов независимой в управлении корпорации, национальный институт развития промышленной науки и технологии (адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония), 22 июля 2002 г., и ему был присвоен номер депозита FERM BP-8124.
Следует обратить внимание, что штамм AJ 110003 (FERM BP-8124) был идентифицирован как указанный выше Sphingobacterium sp. в эксперименте по идентификации, описанном ниже. Штамм FERM BP-8124 представляет собой грамотрицательную палочку (от 0,7 до 0,8 х 1,5 до 2,0 мкм), которая не образует спор и является неподвижной. Ее колонии являются круглыми с совершенно гладкими границами, содержат мало выпячиваний и имеют блестящий, ярко желтый цвет. Организм растет при 30°С и является каталаза-позитивным, оксидаза-позитивным и негативным в отношении теста OF (на глюкозу) и идентифицирован на основе его свойств как бактерия, принадлежащая к роду Sphingobacterium. Более того, из-за того, что он является негативным в отношении восстановления нитратов, негативным в отношении продукции индолов, негативным в отношении продукции кислоты из глюкозы, негативным в отношении аргининдигидролазы, позитивным в отношении уреазы, позитивным в отношении гидролиза эскулина, негативным в отношении гидролиза желатина, позитивным в отношении β-галактозидазы, позитивным в отношении ассимиляции глюкозы, негативным в отношении ассимиляции L-арабинозы, позитивным в отношении ассимиляции D-маннозы, негативным в отношении ассимиляции D-маннита, позитивным в отношении ассимиляции N-ацетил-D-глюкозамина, позитивным в отношении ассимиляции мальтозы, негативным в отношении ассимиляции глюконата калия, негативным в отношении ассимиляции н-каприновой кислоты, негативным в отношении ассимиляции адипиновой кислоты, негативным в отношении ассимиляции dl-яблочной кислоты, негативным в отношении ассимиляции цитрата натрия, негативным в отношении ассимиляции фенилацетата и позитивным в отношении ассимиляции цитохромоксидазы, было установлено, что он имеет свойства, которые являются сходными с таковыми Sphingobacterium multivorum или Sphingobacterium spiritivorum. Более того, хотя результаты анализа гомологии последовательности оснований гена 16S рРНК указывают на то, что наивысшая степень гомологии обнаруживается с Sphingobacterium multivorum (98,8%), не существует штамма, с которым бы бактериальный штамм полностью совпадал. Соответственно, данный бактериальный штамм был, следовательно, идентифицирован как Sphingobacterium sp.
В качестве микроорганизмов могут быть использованы либо штаммы дикого типа, либо мутантные штаммы или могут быть использованы рекомбинантные штаммы, полученные слиянием клеток или генетическими способами, такими как генетические манипуляции.
Для получения клеток таких микроорганизмов микроорганизмы можно культивировать и выращивать в подходящей среде. Не существует особых ограничений в отношении среды, применяемой для данной цели, если она дает возможность роста микроорганизма. Данная среда, если необходимо, может быть обычной средой, содержащей обычные источники углерода, источники азота, источники фосфора, источники серы, неорганические ионы и источники органических питательных веществ.
Например, может быть применен любой источник углерода, если микроорганизмы могут его использовать. Конкретные примеры источника углерода, который может быть использован, включают сахара, такие как глюкоза, фруктоза, мальтоза и амилоза, спирты, такие как сорбит, этанол и глицерин, органические кислоты, такие как фумаровая кислота, лимонная кислота, уксусная кислота и пропионовая кислота и их соли, углеводороды, такие как парафин, а также их смеси.
Примеры источников азота, которые могут быть использованы, включают аммонийные соли неорганических кислот, такие как сульфат аммония и хлорид аммония, аммонийные соли органических кислот, такие как фумарат аммония и цитрат аммония, нитраты, такие как нитрат натрия и нитрат калия, органические соединения, содержащие азот, такие как пептоны, экстракт дрожжей, мясной экстракт и жидкость кукурузного экстракта, а также их смеси.
Кроме того, источники азота, применяемые в обычных средах, такие как неорганические соли, соли металлических микроэлементов и витамины, могут быть также подходящим образом смешаны и применены.
Не существует особого ограничения в отношении условий культивирования и культивирование может быть осуществлено, например, в течение от приблизительно 12 до приблизительно 48 часов при должным образом контролируемом рН и температуре с диапазоном рН от 5 до 8 и температурой в интервале от 15 до 40°С, соответственно, в аэробных условиях.
3. Ферменты, используемые в настоящем изобретении
В способе получения пептида согласно настоящему изобретению, описанном выше, используют фермент, который способен избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь. В способе получения пептида по настоящему изобретению фермент не ограничен своим происхождением и предлагаемым способом, если он обладает такой активностью. Далее будет описана очистка ферментов, применяемых в настоящем изобретении, и применение способов генетической инженерии.
(3-1)Микроорганизмы, содержащие фермент, которые могут быть использованы для способа получения настоящего изобретения
В качестве микроорганизмов, которые продуцируют фермент по настоящему изобретению, могут быть использованы все микроорганизмы, которые способны продуцировать α-L-аспартил-L-фенилаланин-β-эфир из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина. Микроорганизмы включают бактерии и тому подобное, которые принадлежат к родам, выбранным из группы, состоящей из
Aeromonas, Azotobacter, Alcaligenes, Brevibacterium, Corynebacterium, Escherichia, Empedobacter, Flavobacterium, Microbacterium, Propionibacterium, Brevibacillus, Paenibacillus, Pseudomonas, Serratia, Stenotrophomonas, Sphingobacterium, Streptomyces, Xanthomonas, Williopsis, Candida, Geotrichum, Pichia, Saccharomyces, Torulaspora, Cellulophaga, Weeksella, Pedobacter, Persicobacter, Flexithrix, Chitinophaga, Cyclobacterium, Runella, Thermonema, Psychmserpens, Gelidibacter, Dyadobacter, Flammeovirga, Spirosoma, Flectobacillus, Tenacibaculum, Rhodotermus, Zobellia, Muricauda, Salegentibacter, Taxeobacter, Cytophaga, Marinilabilia, Lewinella, Saprospira, Haliscomenobacter.
Более конкретно, микроорганизмы включают Empedobacter brevis ATCC 14234 (FERM штамм P-18545, FERM штамм BP-8113 (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002 г.)), Sphingobacterium sp. FERM штамм BP-8124 (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного депозита: 22 июля 2002 г.), Pedobacter heparinus IFO 12017 (институт-депозитарий: институт ферментации, Osaka; 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония), Taxeobacter gelupurpurascens DSMZ 11116 (институт-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес института-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), Cyclobacterium marinum ATCC 25205 (институт-депозитарий; Американская коллекция типов культур, адрес института-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) и Psychroserpens burtonensis ATCC 700359 (институт-депозитарий; Американская коллекция типов культур, адрес института-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) и так далее. Empedobacter brevis ATCC штамм 14234 (FERM штамм P-18545, FERM штамм BP-8113 (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002 г.)) и Sphingobacterium sp. FERM штамм BP-8124 (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного депозита: 22 июля 2002 г.), Pedobacter heparinus IFO штамм 12017 (институт-депозитарий: институт ферментации, Osaka; 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония), Taxeobacter gelupurpurascens DSMZ штамм 11116 (институт-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес института-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), Cyclobacterium marinum ATCC штамм 25205 (институт-депозитарий; Американская коллекция типов культур, адрес института-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) и Psyclloserpens burtonensis ATCC штамм 700359 (институт-депозитарий; Американская коллекция типов культур, адрес института-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) и тому подобное являются микроорганизмами, выбранными заявителями настоящего изобретения в результате поиска микроорганизмов, продуцирующих фермент, который производит α-L-аспартил-L-фенилаланин-β-эфир из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина с высоким выходом.
(3-2) Очистка фермента
Как указывалось выше, фермент, образующий пептид, используемый в настоящем изобретении, может быть очищен из бактерий, принадлежащих, например, к роду Empedobacter. Способ выделения и очистки фермента, образующего пептид, из Empedobacter brevis приведен в качестве примера очистки фермента.
Сначала получают экстракт клеток микроорганизма из клеток микроорганизма Empedobacter brevis, например, штамма FERM BP-8113 (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002 г.) путем разрушения клеток с применением физического способа разрушения, такого как ультразвуковое разрушение, или ферментативного способа разрушения с применением фермента, растворяющего клеточную стенку, и удаления нерастворимой фракции путем центрифугирования и так далее. Фермент, образующий пептид, может быть затем очищен путем фракционирования клеточного экстракта, полученного указанным выше способом, с помощью сочетания обычных способов очистки белков, таких как анионообменная хроматография, катионообменная хроматография или гель-фильтрационная хроматография.
Примером носителя для применения в анионообменной хроматографии является Q-Сефароза HP (производство Amersham). После прохождения клеточного экстракта, содержащего фермент, через колонку с носителем фермент восстанавливают в неадсорбированной фракции при pH 8,5.
Примером носителя, используемым в катионообменной хроматографии, является MonoS HR (производство Amersham). После прохождения клеточного экстракта, содержащего фермент, через колонку с носителем и адсорбции фермента на колонке колонку промывают и фермент элюируют буферным раствором с высокой концентрацией соли. На этом этапе концентрация соли может последовательно увеличиваться или может быть применен градиент концентрации. Например, в случае использования MonoS HR адсорбированный на колонке фермент элюируют при концентрации NaCl от приблизительно 0,2 до приблизительно 0,5 М.
Фермент, очищенный описанным выше способом, может быть затем дополнительно очищен стандартной гель-фильтрационной хроматографией и так далее. Примером носителя, используемым в гель-фильтрационной хроматографии, является Sephadex 200 пг (производство Amersham).
В указанной выше процедуре очистки фракция, содержащая фермент, может быть идентифицирована тестированием каждой фракции на пептид-образующую активность в соответствии со способом, указанным в примерах, которые будут описаны далее. Внутренняя аминокислотная последовательность фермента, очищенного описанным выше способом, представлена в SEQ ID NO: 1 и SEQ ID NO: 2 перечня последовательностей.
(3-3) Выделение ДНК, получение трансформанта и очистка фермента, образующего пептид
(3-3-1) Выделение ДНК
Изобретателям настоящего изобретения сначала удалось выделить один тип ДНК фермента, образующего пептид, который может быть применен в способе получения пептида настоящего изобретения из штамма FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002 г.).
ДНК, содержащая последовательность оснований, состоящую из оснований с номера 61 по 1908 последовательности оснований, представленной в SEQ ID NO: 5, которая представляет собой ДНК по настоящему изобретению, была выделена из штамма FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002 г.). ДНК, содержащая последовательность оснований, состоящую из оснований с номера 61 по 1908, представляет собой кодирующую часть последовательности (CDS). В последовательности оснований, состоящей из оснований с номера 61 по 1908, содержатся область сигнальной последовательности и область зрелого белка. Область сигнальной последовательности представляет собой область, которая состоит из оснований с номера 61 по 126, в то время как область зрелого белка представляет собой область, которая состоит из оснований с номера 127 по 1908. Таким образом, настоящее изобретение относится и к гену ферментативного белка, образующего пептид, который содержит сигнальную последовательность, так и ген ферментативного белка, образующего пептид, в форме зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, представленной в SEQ ID NO: 5, представляет собой тип лидирующей последовательности. Основной функцией лидирующего пептида, кодируемого лидирующей последовательностью, как предполагается, является экскреция извне клетки. Белок, кодируемый основаниями с номера 127 по 1908, а именно с эфирным участком, за исключением лидирующего пептида, представляет собой, как предполагается, зрелый белок и проявляет высокую степень активности в отношении образования пептида.
ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 61 по 1917, представленной в SEQ ID NO: 11, которая представляет собой также ДНК по настоящему изобретению, была выделена из Sphingobacterium sp., штамм FERM BP-8124 (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного депозита: 22 июля 2002 г.). ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 61 по 1917, представляет собой кодирующую часть последовательности (CDS). В последовательности оснований, состоящей из оснований с номера 61 по 1917, содержатся область сигнальной последовательности и область зрелого белка. Область сигнальной последовательности представляет собой область, которая состоит из оснований с номера 61 по 120, в то время как область зрелого белка представляет собой область, которая состоит из оснований с номера 121 по 1917. Таким образом, настоящее изобретение относится как к гену ферментативного белка, образующего пептид, который содержит сигнальную последовательность, так и к гену ферментативного белка, образующего пептид, в форме зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, представленной в SEQ ID NO: 11, представляет собой тип лидирующей последовательности. Основная функция лидирующего пептида, кодируемого лидирующей последовательностью, заключается, как предполагается, в экскреции из клетки. Белок, кодируемый основаниями с номера 121 по 1917, а именно часть, за исключением лидирующего пептида, представляет собой, как предполагается, зрелый белок и проявляет высокую степень активности в отношении образования пептида.
ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 61 по 1935, представленной в SEQ ID NO: 17, которая представляет собой также ДНК настоящего изобретения, была выделена из Pedobacter heparinus, штамм IFO 12017 (институт-депозитарий: институт ферментации Osaka; 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония). ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 61 по 1935, представленной в SEQ ID NO: 17, представляет собой кодирующую часть последовательности (CDS). В последовательности оснований, состоящей из оснований с номера 61 по 1935, содержатся область сигнальной последовательности и область зрелого белка. Область сигнальной последовательности представляет собой область, которая состоит из оснований с номера 61 по 126, в то время как область зрелого белка представляет собой область, которая состоит из оснований с номера 127 по 1935. Таким образом, настоящее изобретение относится как к гену ферментативного белка, образующего пептид, который содержит сигнальную последовательность, так и к гену ферментативного белка, образующего пептид, в форме зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, представленной в SEQ ID NO: 17, представляет собой тип лидирующей последовательности. Основная функция лидирующего пептида, кодируемого лидирующей последовательностью, заключается, как предполагается, в экскреции изнутри наружу через мембрану клетки. Белок, кодируемый основаниями с номера 127 по 1935, а именно часть, за исключением лидирующего пептида, представляет собой, как предполагается, зрелый белок и проявляет высокую степень активности в отношении образования пептида.
ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 61 по 1995, представленной в SEQ ID NO: 22, которая представляет собой также ДНК настоящего изобретения, была выделена из Taxeobacter gelupurpurascens DSMZ 11116 (институт-депозитарий; Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес института-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия). ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 61 по 1995, описанная в SEQ ID NO: 22, представляет собой кодирующую часть последовательности (CDS). В последовательности оснований, состоящей из оснований с номера 61 по 1995, содержатся область сигнальной последовательности и область зрелого белка. Область сигнальной последовательности представляет собой область, которая состоит из оснований с номера 61 по 126, в то время как область зрелого белка представляет собой область, которая состоит из оснований с номера 127 по 1995. Таким образом, настоящее изобретение относится как к гену ферментативного белка, образующего пептид, который содержит сигнальную последовательность, так и к гену ферментативного белка, образующего пептид, в форме зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, представленной в SEQ ID NO: 22, представляет собой тип лидирующей последовательности. Основная функция лидирующего пептида, кодируемого лидирующей последовательностью, заключается, как предполагается, в экскреции из клетки. Белок, кодируемый основаниями с номера 127 по 1995, а именно часть, за исключением лидирующего пептида, представляет собой, как предполагается, зрелый белок и проявляет высокую степень активности в отношении образования пептида.
ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 29 по 1888, представленной в SEQ ID NO: 24, которая представляет собой также ДНК настоящего изобретения, была выделена из Cyclobacterium marinum ATCC 25205 (институт-депозитарий; Американская коллекция типов культур, адрес института-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки). ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 29 по 1888, описанная в SEQ ID NO: 24, представляет собой кодирующую часть последовательности (CDS). В последовательности оснований, состоящей из оснований с номера 291 по 1888, содержатся область сигнальной последовательности и область зрелого белка. Область сигнальной последовательности представляет собой область, которая состоит из оснований с номера 29 по 103, в то время как область зрелого белка представляет собой область, которая состоит из оснований с номера 104 по 1888. Таким образом, настоящее изобретение относится как к гену ферментативного белка, образующего пептид, который содержит сигнальную последовательность, так и к гену ферментативного белка, образующего пептид, в форме зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, представленной в SEQ ID NO: 24, представляет собой тип лидирующей последовательности. Основная функция лидирующего пептида, кодируемого лидирующей последовательностью, заключается, как предполагается, в экскреции из клетки. Белок, кодируемый основаниями с номера 104 по 1888, а именно часть, за исключением лидирующего пептида, представляет собой, как предполагается, зрелый белок и проявляет высокую степень активности в отношении образования пептида.
ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 61 по 1992, представленной в SEQ ID NO: 26, которая представляет собой также ДНК настоящего изобретения, была выделена из Psychroserpens burtonensis ATCC 700359 (институт-депозитарий; Американская коллекция типов культур, адрес института-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки). ДНК, состоящая из последовательности оснований, которая состоит из оснований с номера 61 по 1992, представленная в SEQ ID NO: 26, представляет собой кодирующую часть последовательности (CDS). В последовательности оснований, состоящей из оснований с номера 61 по 1992, содержатся область сигнальной последовательности и область зрелого белка. Область сигнальной последовательности представляет собой область, которая состоит из оснований с номера 61 по 111, в то время как область зрелого белка представляет собой область, которая состоит из оснований с номера 112 по 1992. Таким образом, настоящее изобретение относится как к гену ферментативного белка, образующего пептид, который содержит сигнальную последовательность, так и к гену ферментативного белка, образующего пептид, в форме зрелого белка. Сигнальная последовательность, содержащаяся в последовательности, представленной в SEQ ID NO: 26, представляет собой тип лидирующей последовательности. Основная функция лидирующего пептида, кодируемого лидирующей последовательностью, заключается, как предполагается, в экскреции из клетки. Белок, кодируемый основаниями с номера 112 по 1992, а именно часть, за исключением лидирующего пептида, представляет собой, как предполагается, зрелый белок и проявляет высокую степень активности в отношении образования пептида.
Более того, различные способы рекомбинации генов, указанные ниже, могут быть осуществлены в соответствии с описаниями в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989) и в других публикациях.
ДНК, кодирующая фермент, который может быть использован в настоящем изобретении, может быть получена с помощью полимеразной цепной реакцией (ПЦР, ссылка на White T.J. et al., Trends Genet., 5, 185 (1989)) или гибридизацией из хромосомной ДНК или библиотеки ДНК Empedobacter brevis, Sphingobacterium sp., Pedobacter heparinus, Taxeobacter gelupurpurascens, Cyclobacterium marinum или Psychroserpens burtonensis. Праймеры, применяемые в ПЦР, могут быть созданы на основе внутренних последовательностей аминокислот, определяемых на основе очищенного фермента, образующего пептид, как объяснялось в предыдущем разделе (3). Кроме того, так как в настоящем изобретении были идентифицированы последовательности оснований генов ферментов, образующих пептид (SEQ ID NO: 5, SEQ ID NO: 11, SEQ ID NO:17, SEQ ID NO:22, SEQ ID NO:24 и SEQ ID NO:26), праймеры или зонды для гибридизации могут быть созданы на основе данных последовательностей оснований и гены могут быть выделены с применением этих зондов. Если праймеры, имеющие последовательности, соответствующие 5'-нетранслируемой области и 3'-нетранслируемой области, соответственно, применяются в качестве праймеров для ПЦР, то может быть амплифицирована полноразмерная кодирующая область фермента. Используя в качестве примера случай амплификации области, содержащей как лидирующую последовательность, так и кодирующую область зрелого белка, как представлено в SEQ ID NO: 5, конкретные примеры праймеров включают праймер с последовательностью оснований области выше основания номер 61 в SEQ ID NO: 5 для праймера 5'-конца, и праймер, имеющий последовательность, комплементарную последовательности оснований области ниже основания номер 1908 для праймера 3'-конца.
Праймеры могут быть синтезированы, например, в соответствии с обычными способами, используя фосфоамидитный способ (ссылка на Tetrahedron Letters (1981), 22, 1859) с помощью синтезатора ДНК модели 380B, производимого Applied Biosystems. Реакция ПЦР может осуществляться, например, с применением Gene Amp PCR System 9600 (Perkin-Elmer) и Takara LA PCR In Vitro Cloning Kit (Takara Shuzo) в соответствии со способом, описанным поставщиком, таким как производитель.
ДНК, которая кодирует фермент, который может использоваться в способе получения пептида по настоящему изобретению, независимо от того, содержит ли ДНК лидирующую последовательность или нет, включает ДНК, которая по существу идентична ДНК, состоящей из CDS, представленной в SEQ ID NO: 5 перечня последовательностей. А именно, ДНК, по существу идентичная ДНК по настоящему изобретению, может быть получена путем выделения ДНК, которая гибридизуется с ДНК с последовательностью оснований, комплементарных CDS, представленной в SEQ ID NO: 5 перечня последовательностей, или с зондом, полученным из последовательности оснований в жестких условиях, и кодирующей белок с пептид-образующей активностью, из ДНК, кодирующей мутантный фермент или клетки, которые содержат эту ДНК.
ДНК по настоящему изобретению, независимо от того, содержит ли она лидирующую последовательность или нет, включает ДНК, которая по существу идентична ДНК, состоящей из CDS, представленной в SEQ ID NO: 11 перечня последовательностей. А именно, ДНК, по существу идентичная ДНК по настоящему изобретению, может быть получена путем выделения ДНК, которая гибридизуется с ДНК, состоящей из последовательности оснований, комплементарных CDS, представленной в SEQ ID NO: 11 перечня последовательностей, или с зондом, полученным из последовательности оснований в жестких условиях, и кодирующей белок, с пептид-образующей активностью из ДНК, кодирующей мутантный фермент, или из клеток, которые обладают данной ДНК.
ДНК по настоящему изобретению, независимо от того, содержит ли она лидирующую последовательность или нет, включает ДНК, которая по существу является идентичной ДНК, состоящей из CDS, представленной в SEQ ID NO: 17 перечня последовательностей. А именно, ДНК, по существу идентичная ДНК настоящего изобретения, может быть получена путем выделения ДНК, которая гибридизуется с ДНК, состоящей из последовательности оснований, комплементарных CDS, представленной в SEQ ID NO: 17 перечня последовательностей, или с зондом, полученным из последовательности оснований в жестких условиях, и кодирующей белок, с пептид-образующей активностью из ДНК, кодирующей мутантный фермент, или из клеток, которые содержат данную ДНК.
ДНК по настоящему изобретению, независимо от того, содержит ли она лидирующую последовательность или нет, включает ДНК, которая по существу идентична ДНК, состоящей из CDS, представленной в SEQ ID NO: 22 перечня последовательностей. А именно, ДНК, по существу идентичная ДНК по настоящему изобретению, может быть получена путем выделения ДНК, которая гибридизуется с ДНК, состоящей из последовательности оснований, комплементарных CDS, представленной в SEQ ID NO: 22 перечня последовательностей, или с зондом, полученным из последовательности оснований в жестких условиях, и кодирующей белок, с пептид-образующей активностью из ДНК, кодирующей мутантный фермент, или из клеток, которые обладают данной ДНК.
ДНК по настоящему изобретению, независимо от того, содержит ли она лидирующую последовательность или нет, включает ДНК, которая по существу идентична ДНК, состоящей из CDS, представленной в SEQ ID NO: 24 перечня последовательностей. А именно, ДНК, по существу идентичная ДНК настоящего изобретения, может быть получена путем выделения ДНК, которая гибридизуется с ДНК, состоящей из последовательности оснований, комплементарных CDS, представленной в SEQ ID NO: 24 перечня последовательностей, или с зондом, полученным из последовательности оснований в жестких условиях, и кодирующей белок, с пептид-образующей активностью из ДНК, кодирующей мутантный фермент, или из клеток, которые обладают данной ДНК.
ДНК по настоящему изобретению, независимо от того, содержит ли она лидирующую последовательность или нет, включает ДНК, которая по существу идентична ДНК, состоящей из CDS, представленной в SEQ ID NO: 26 перечня последовательностей. А именно, ДНК, по существу идентичная ДНК настоящего изобретения, может быть получена путем выделения ДНК, которая гибридизуется с ДНК, состоящей из последовательности оснований, комплементарных CDS, представленной в SEQ ID NO: 26 перечня последовательностей, или с зондом, полученным из последовательности оснований в жестких условиях, и кодирующей белок, с пептид-образующей активностью из ДНК, кодирующей мутантный фермент, или из клеток, которые обладают данной ДНК.
Зонд может быть получен, например, в соответствии с обычными способами, основанными, например, на последовательности оснований, представленной в SEQ ID NO: 5 перечня последовательностей. Кроме того, можно также использовать способ выделения интересующей ДНК, используя зонд для определения ДНК, которая гибридизуется с зондом, в соответствии с обычными методами. Например, ДНК зонд может быть получен амплификацией последовательности оснований, клонированной в плазмидном или фаговом векторе, отщеплением последовательности оснований, которую желательно использовать в качестве зонда, с помощью фермента рестрикции, и затем экстракцией желаемой последовательности оснований. Отщепляемая часть может быть скорректирована в зависимости от интересующей ДНК.
Используемый здесь термин «в жестких условиях» относится к условиям, при которых не образуется неспецифический гибрид, а так называемый специфический гибрид. Трудно в точности передать эти условия в цифровых значениях. Например, можно указать условия, при которых ДНК с высокой гомологией, например 50% или более предпочтительно 80% или более, более предпочтительно 90% или более, гибридизуются друг с другом, а ДНК с более низкой степенью гомологии, чем указанные, не гибридизуются друг с другом, или обычные условия при промывании для гибридизации по Саузерну, при которых гибридизацию осуществляют при 60°С при концентрации соли, соответствующей 1хSSC и 0,1% SDS, предпочтительно 0,1хSSC и 0,1% SDS. Хотя гены, которые гибридизуются в таких условиях, включают те гены, в последовательностях которых в определенных положениях находятся стоп-кодоны, или которые потеряли активность в результате мутации в активном центре, они могут быть легко удалены путем лигирования с коммерчески доступным вектором экспрессии, экспрессирующим их в подходящем хозяине, и тестировании ферментативной активности продукта экспрессии с применением способа, который будет описан позже.
Однако в случае последовательности оснований, которая гибридизуется в жестких условиях, как описано выше, предпочтительно, чтобы белок, кодируемый этой последовательностью оснований, содержал приблизительно половину или более, предпочтительно 80% или более и более предпочтительно 90% или более, ферментативной активности белка с аминокислотной последовательностью, кодируемой исходной последовательностью оснований, служащей в качестве основы, способной сохраняться в условиях 50°С и рН 8. Например, в качестве примера можно привести случай, когда последовательность оснований, которая гибридизуется в жестких условиях с ДНК с последовательностью оснований, комплементарную последовательности оснований, состоящей из оснований с номерами 127 по 1908 последовательности оснований, представленной в SEQ ID NO: 5, предпочтительно, чтобы белок, кодируемый этой последовательностью оснований, сохранял приблизительно половину или более, предпочтительно 80% или более и более предпочтительно 90% или более, ферментативной активности белка с аминокислотной последовательностью, которая состоит из аминокислотных остатков с номерами 23 по 616 аминокислотной последовательности, представленной в SEQ ID NO: 6, в условиях 50°С и рН 8.
Аминокислотная последовательность, кодируемая CDS, представленной в SEQ ID NO: 5 перечня последовательностей, показана в SEQ ID NO: 6 перечня последовательностей. Аминокислотная последовательность, кодируемая CDS, представленной в SEQ ID NO: 11 перечня последовательностей, показана в SEQ ID NO: 12 перечня последовательностей. Аминокислотная последовательность, кодируемая CDS, представленной в SEQ ID NO: 17 перечня последовательностей, показана в SEQ ID NO: 18 перечня последовательностей. Аминокислотная последовательность, кодируемая CDS, представленной в SEQ ID NO: 22 перечня последовательностей, показана в SEQ ID NO: 23 перечня последовательностей. Аминокислотная последовательность, кодируемая CDS, представленной в SEQ ID NO: 24 перечня последовательностей, показана в SEQ ID NO: 25 перечня последовательностей. Аминокислотная последовательность, кодируемая CDS, представленной в SEQ ID NO: 26 перечня последовательностей, показана в SEQ ID NO: 27 перечня последовательностей.
Полная аминокислотная последовательность, представленная в SEQ ID NO: 6, содержит лидирующий пептид и область зрелого белка с аминокислотными остатками с номерами с 1 по 22, составляющими лидирующий пептид, и аминокислотными остатками с номерами с 23 по 616, составляющими область зрелого белка.
Полная аминокислотная последовательность, представленная в SEQ ID NO: 11, включает лидирующий пептид и область зрелого белка с аминокислотными остатками с номерами с 1 по 20, составляющими лидирующий пептид, и аминокислотными остатками с номерами с 21 по 619, составляющими область зрелого белка.
Полная аминокислотная последовательность, представленная в SEQ ID NO: 18, содержит лидирующий пептид и область зрелого белка с аминокислотными остатками с номерами с 1 по 22, составляющими лидирующий пептид, и аминокислотными остатками с номерами с 23 по 625, составляющими область зрелого белка.
Полная аминокислотная последовательность, представленная в SEQ ID NO: 23, содержит лидирующий пептид и область зрелого белка с аминокислотными остатками с номерами с 1 по 22, составляющими лидирующий пептид, и аминокислотными остатками с номерами с 23 по 645, составляющими область зрелого белка.
Полная аминокислотная последовательность, представленная в SEQ ID NO: 25, содержит лидирующий пептид и область зрелого белка с аминокислотными остатками с номерами с 1 по 25, составляющими лидирующий пептид, и аминокислотными остатками с номерами с 26 по 620, составляющими область зрелого белка.
Полная аминокислотная последовательность, представленная в SEQ ID NO: 27, содержит лидирующий пептид и область зрелого белка с аминокислотными остатками с номерами с 1 по 17, составляющими лидирующий пептид, и аминокислотными остатками с номерами с 18 по 644, составляющими область зрелого белка.
Белок, кодируемый ДНК по настоящему изобретению, представляет собой белок, в котором зрелый белок обладает пептид-образующей активностью, а ДНК, которая кодирует белок, по существу идентична белку с аминокислотной последовательностью, представленную в SEQ ID NO: 6, в SEQ ID NO: 12, SEQ ID NO: 18, SEQ ID NO: 23, SEQ ID NO: 25 или в SEQ ID NO: 27 перечня последовательностей, не зависимо от того, содержит ли он лидирующую последовательность или нет, также входит в состав ДНК по настоящему изобретению. (Следует отметить, что последовательности оснований описываются из аминокислотных последовательностей в соответствии с кодами универсальных кодонов). А именно, настоящее изобретение относится к ДНК, которые кодируют белки, указанные с (A) по (X) ниже:
(A) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 23 по 616 аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей,
(B) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 23 по 616 аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей, способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(C) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 21 по 619 аминокислотной последовательности, представленной в SEQ ID NO: 12 перечня последовательностей,
(D) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 21 по 619 аминокислотной последовательности, представленной в SEQ ID NO: 12 перечня последовательностей, способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(E) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 23 по 625 аминокислотной последовательности, представленной в SEQ ID NO: 18 перечня последовательностей,
(F) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 23 по 625 аминокислотной последовательности, представленной в SEQ ID NO: 18 перечня последовательностей, способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(G) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 23 по 645 аминокислотной последовательности, представленной в SEQ ID NO: 23 перечня последовательностей,
(H) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 23 по 645 аминокислотной последовательности, представленной в SEQ ID NO: 23 перечня последовательностей, способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(I) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 26 по 620 аминокислотной последовательности, представленной в SEQ ID NO: 25 перечня последовательностей,
(J) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 26 по 620 аминокислотной последовательности, представленной в SEQ ID NO: 25 перечня последовательностей, способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(K) белок с аминокислотной последовательностью, состоящей из аминокислотных остатков с номерами от 18 по 644 аминокислотной последовательности, представленной в SEQ ID NO: 27 перечня последовательностей,
(L) белок с аминокислотной последовательностью, включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, состоящей из аминокислотных остатков с номерами от 18 по 644 аминокислотной последовательности, представленной в SEQ ID NO: 27 перечня последовательностей, способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(M) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 6 перечня последовательностей,
(N) белок, содержащий область зрелого белка, имеющий аминокислотную последовательность, включающую замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(O) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 12 перечня последовательностей,
(P) белок, содержащий область зрелого белка, имеющий аминокислотную последовательность, включающую замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 12 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(Q) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 18 перечня последовательностей,
(R) белок, содержащий область зрелого белка, имеющий аминокислотную последовательность, включающую замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 18 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(S) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 23 перечня последовательностей,
(T) белок, содержащий область зрелого белка, имеющий аминокислотную последовательность, включающую замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 23 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(U) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 25 перечня последовательностей,
(V) белок, содержащий область зрелого белка, имеющий аминокислотную последовательность, включающую замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 25 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
(W) белок с аминокислотной последовательностью, представленной в SEQ ID NO: 27 перечня последовательностей, и
(X) белок, содержащий область зрелого белка, имеющий аминокислотную последовательность в аминокислотной последовательности, представленной в SEQ ID NO: 27 перечня последовательностей, и способный избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь.
Здесь, хотя значение термина «множество» и изменяется в зависимости от положений и типов аминокислотных остатков в трехмерной структуре белка, тем не менее оно находится в диапазоне, при котором не происходит существенного нарушения трехмерной структуры и активность белка с аминокислотными остатками составляет конкретно от 2 до 50, предпочтительно от 2 до 30 и более предпочтительно от 2 до 10. Однако в случае аминокислотных последовательностей, включающих замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотных последовательностях белков (B), (D), (F), (H), (J), (L), (N), (P), (R), (T), (V) или (X), предпочтительно, чтобы белки сохраняли приблизительно половину или более, более предпочтительно 80% или более и еще более предпочтительно 90% или более, ферментативной активности белков в состоянии, когда мутация отсутствует, в условиях 50°С и рН 8. В качестве примера приведено описание случая (B); в случае аминокислотной последовательности (B), включающей замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 6 перечня последовательностей, предпочтительно, чтобы этот белок сохранял приблизительно половину или более, более предпочтительно 80% или более и еще более предпочтительно 90% или более, ферментативной активности белка с аминокислотной последовательностью, представленной в SEQ ID NO: 6 перечня последовательностей, в условиях 50°С и рН 8.
Мутацию аминокислоты, такую как указанная в представленной выше последовательности (B) и так далее, получают путем модификации последовательности оснований так, чтобы в отношении аминокислоты специфического эфирного участка в гене фермента по настоящему изобретению была бы проведена замена, делеция, инсерция, добавление с помощью, например, мутагенеза, направленного на эфирный участок. Кроме того, модифицированную ДНК, подобную описанной выше, можно также получить с помощью мутагенного воздействия, известного в данной области. Мутагенное воздействие относится, например, к способу, при котором ДНК, кодирующую фермент по настоящему изобретению, обрабатывают in vitro гидроксиламином и так далее, а также к способу, при котором бактерии Escherichia, которые содержат ДНК, кодирующей фермент настоящего изобретения, обрабатывают мутагеном, обычно применяемым для искусственного мутагенеза, таким как ультрафиолетовое излучение, N-метил-N'-нитро-N-нитрозогуанидин (NTG) или азотистой кислотой.
Кроме того, замена, делеция, инсерция, добавление и/или инверсия, описанные выше, могут быть видовыми природными мутациями или мутациями, различающимися у различных штаммов микроорганизмов. Путем экспрессии ДНК, содержащей такую мутацию, в подходящих клетках и исследования ферментативной активности продукта экспрессии может быть получена ДНК, которая кодирует белок, по существу идентичный белку, описанному в SEQ ID NO: 6, 12, 18, 23, 25 или 27 перечня последовательностей.
(3-3-2) Получение трансформантов и продукция образующих пептид ферментов
Образующие пептид ферменты, которые могут использоваться в способе получения пептида по настоящему изобретению, могут быть получены введением ДНК, описанной в (3-3-1) выше, в подходящего хозяина и экспрессией ДНК в этом хозяине.
В отношении хозяев для экспрессии белка, определяемого ДНК, примеры хозяев, которые могут использоваться, включают различные прокариотные клетки, включая бактерии Escherichia, такие как Escherichia coli, бактерии Empedobacter, бактерии Sphingobacterium, бактерии Flavobacterium и Bacillus subtilis, а также различные эукариотические клетки, включая Saccharomyces cerevisiae, Pichia stipitis и Aspergillus oryzae.
Рекомбинантная ДНК, используемая для введения ДНК в хозяина, может быть получена путем инсерции вводимой ДНК в вектор, соответствующий типу хозяина, в котором должна быть экспрессирована ДНК, в такой форме, чтобы белок, кодируемый данной ДНК, мог быть экспрессирован. В случае, когда промотор уникален для гена образующего пептид фермента Empedobacter brevis и далее функционирует в клетках-хозяевах, промотор может быть применен в качестве промотора для экспрессии ДНК настоящего изобретения. Кроме того, другой промотор, который действует в клетках-хозяевах, может быть лигирован с ДНК настоящего изобретения, и ДНК может быть экспрессирована, если это необходимо, под контролем промотора.
Примеры способов трансформации для введения рекомбинантной ДНК в клетки-хозяева включают способ D.M. Morrison (смотри Methods in Enzymology, 68, 326 (1979)) или способ, в котором проницаемость для ДНК повышают обработкой рецептирующих клеток микроорганизма хлоридом кальция (смотри Mandel, H. and Higa, A., J. Mol. Biol., 53, 159 (1970)).
В случае крупномасштабной продукции белка с использованием способа рекомбинантной ДНК конъюгация белка в трансформанте, который продуцирует белок с образованием телец включения белка, также является предпочтительным способом осуществления настоящего изобретения. Преимуществами данного способа экспрессии и продукции является защита целевого белка от гидролиза протеазами, находящимися в клетках микроорганизма, и простую и легкую очистку целевого белка путем разрушения клеток микроорганизма с последующим разделением центрифугированием и так далее.
Тельца включения белка, полученные данным способом, солюбилизируют с помощью денатурирующего белок агента, и белок преобразуют в правильно уложенный, физиологически активный белок посредством процедуры восстановления активности, которая состоит прежде всего из удаления денатурирующего агента. Существует множество таких примеров, включая восстановление активности интерлейкина-2 человека (смотри японскую патентную заявку, вложенная публикация No. S61-257931).
Для получения активного белка из телец включения требуется ряд процедур, включая солюбилизацию и восстановление активности, и процедура является более сложной, чем в случае непосредственного получения активного белка. Однако в случае получения белка, который оказывает отрицательный эффект на рост микроорганизмов в больших объемах в клетках микроорганизма, этот эффект может быть подавлен путем аккумуляции белков в форме телец включения неактивного белка в клетках микроорганизма.
Примеры способов крупномасштабной продукции для получения интересующего белка в форме телец включения включают способ, в котором интересующий белок экспрессируют независимо под контролем сильного промотора, и способ, в котором интересующий белок экспрессируют в форме гибридного белка с белком, который известен как экспрессирующийся в большом количестве.
Далее настоящее изобретение будет описано более конкретно с рассмотрением в качестве примера способа получения трансформированной Escherichia coli и использование такого трансформированного микроорганизма для получения фермента, образующего пептид. Более того, в случае получения фермента, образующего пептид, в микроорганизм, такой как Escherichia coli, может быть введена ДНК, которая кодирует белок-предшественник, содержащий лидирующую последовательность, или может быть введена ДНК, которая состоит только из области зрелого белка, которая не содержит лидирующей последовательности, и ДНК может быть подходящим образом выбрана для последовательности, кодирующей белок, в зависимости от условий получения, формы, условий применения и так далее для продуцируемого фермента.
Промоторы, обычно применяемые для получения гетерологичных белков в Escherichia coli, могут использоваться в качестве промотора для экспрессии ДНК, кодирующей фермент, образующий пептид. Примеры таких промоторов включают промотор T7, промотор lac, промотор trp, промотор trc, промотор tac, промотор PR фага лямбда, промотор PL и другие сильные промоторы. Кроме того, примеры векторов, которые могут быть применены, включают pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219 и pMW218. Кроме того, могут использоваться векторы на основе ДНК фага. Более того, могут использоваться экспрессионные векторы, которые содержат промоторы и способны экспрессировать введенную последовательность ДНК.
Для получения фермента, образующего пептид, в виде телец включения гибридного белка, ген, кодирующий другой белок и предпочтительно гидрофильный пептид, лигируют выше или ниже гена фермента, образующего пептид, для получения гена гибридного белка. Ген, который кодирует другой белок в данном способе, может представлять собой любой ген, который увеличивает количество аккумулированного гибридного белка и увеличивает растворимость гибридного белка после стадий денатурации и восстановления. Примеры возможных генов включают ген 10 T7, ген β-галактозидазы, ген дегидрофолатредуктазы, ген γ-интерферона, ген интерлейкина-2 и ген прохимозина.
Если же гены лигируют с генами, которые кодируют ферменты, образующие пептид, гены лигируют так, чтобы рамки считывания кодонов совпадали. Рекомендуется, чтобы гены были лигированы у подходящего участка рестрикции фермента-эфира или использовалась ДНК, имеющая подходящую последовательность.
Кроме того, для увеличения количества продуцируемого фермента, образующего пептид, в некоторых случаях предпочтительно, чтобы терминатор, который представляет собой последовательность терминации транскрипции, был лигирован ниже гена гибридного белка. Терминатор включает, например, терминатор T7, терминатор фага fd, терминатор T4, терминатор гена устойчивости к тетрациклину и терминатор гена trpA Escherichia coli.
В качестве векторов для введения гена, который кодирует фермент, образующий белок, или гибридный белок фермента, образующего пептид, и другого белка, в Escherichia coli предпочтительны так называемые мультикопийные типы векторов, примеры которых включают плазмиду, имеющую репликатор, полученный из ColE1, например плазмиду на основе pUC и плазмиду на основе pBR322 или их производные. Применяемый здесь термин «производные» относится к тем плазмидам, которые подвергаются модификации путем замены, делеции, инсерции, добавления и/или инверсии оснований. Следует отметить, что применяемая здесь модификация включает модификации с помощью мутагенного воздействия мутагеном или УФ-облучением или модификации при спонтанной мутации.
Для скрининга трансформантов предпочтительно, чтобы векторы имели маркеры, такие как ген устойчивости к ампициллину. Такие плазмиды являются коммерчески доступными экспрессионными векторами с сильными промоторами (вектор на основе pUC (производство Takara Shuzo, Co., Ltd.), вектор на основе pRROK (производство Clonetech Laboratories, Inc.), pKK233-2 (производство Clonetech Laboratories, Inc.) и так далее.
Рекомбинантную ДНК получают лигированием фрагмента ДНК с вектором ДНК. В данном случае промотор, ген, кодирующий гидролазу амидов L-аминокислот, или гибридный белок, состоящий из гидролазы амидов L-аминокислот и другого белка, и, в зависимости от случая, терминатор, лигируют в таком порядке.
Если Escherichia coli трансформируют с использованием рекомбинантной ДНК и полученную Escherichia coli культивируют, то экспрессируется и продуцируется фермент, образующий пептид, или гибридный белок, состоящий из фермента, образующего пептид, и другого белка. Хотя в качестве хозяина, подлежащего трансформации, использован штамм, который обычно применяют при экспрессии гетерогенного гена, штамм JM109 Escherichia coli, например, является предпочтительным. Способы осуществления трансформации и способы скрининга трансформантов описаны в Molecular Cloning, 2nd Edition, Cold Spring Harbor Press (1989) и в других публикациях.
В случае экспрессии фермента, образующего пептид, в форме гибридного белка фермент, образующий пептид, может быть выщеплен с применением рестрикционной протеазы, такой как фактор Xa свертывания крови или калликреин, которая в качестве узнаваемой последовательности использует последовательность, не присутствующую в ферменте, образующем пептид.
В качестве среды для получения может быть использована среда, обычно применяемая для культивирования Escherichia coli, такая как M9-касаминокислотная среда или среда LB. Кроме того, условия культивирования и условия индукции продукции подбирают подходящим образом в соответствии с маркером применяемого вектора, промотором, типом микроорганизма-хозяина и так далее.
Для получения фермента, образующего пептид, или гибридного белка, состоящего из фермента, образующего пептид, и другого белка может быть использован следующий способ. Если фермент, образующий пептид, или его гибридный белок растворимы в клетках микроорганизма, после получения клеток микроорганизма, клетки микроорганизма разрушают или лизируют так, чтобы они могли быть использованы в качестве неочищенной жидкости, содержащей фермент. Более того, фермент, образующий пептид, или его гибридный белок могут быть очищены перед применением обычными способами, таких как осаждение, фильтрация или колоночная хроматография, как это необходимо. В данном случае может быть также использован способ очистки, при котором применяют антитело против фермента, образующего пептид, или его гибридного белка.
В случае, когда образуются тельца включения белка, тельца включения солюбилизируют денатурирующим агентом. Они могут быть солюбилизированы вместе с белком клетки микроорганизма. Однако при рассмотрении следующей процедуры очистки тельца включения предпочтительно получают и затем солюбилизируют. Для получения телец включения из клеток микроорганизма могут быть применены известные общепринятые способы. Например, тельца включения могут быть получены путем разрушения клеток микроорганизма с последующим разделением путем центрифугирования. Примеры денатурирующих агентов, способных солюбилизировать тельца включения, включают гуанидин гидрохлорид (например, 6 М, рН от 5 до 8) и мочевину (например, 8 М).
Белок, обладающий активностью, восстанавливают путем удаления данных денатурирующих агентов с помощью диализа. При диализе раствор трис-HCl буфера или раствор фосфатного буфера и так далее могут быть применены в качестве раствора для диализа и концентрации могут составлять, например, от 20 мМ до 0,5 М, в то время как рН может быть, например, от 5 до 8.
Концентрация белка в процессе стадии восстановления составляет предпочтительно приблизительно 500 мкг/мл или менее. Температура диализа составляет предпочтительно 5°С или ниже для торможения собственной поперечной сшивки восстановленного фермента, образующего пептид. Более того, в добавление к диализу разведение или ультрафильтрация могут быть применены для удаления денатурирующих агентов и ожидается, что активность может быть восстановлена не зависимо от того, какой денатурирующий агент применялся.
<2> Способ получения α-L-аспартил-L-фенилаланин-α-метилового эфира
Способ получения α-APM в соответствии с настоящим изобретением включает первую стадию синтеза α-L-аспартил-L-фенилаланин-β-метилового эфира в соответствии с «<1> методом получения α-L-аспартил-L-фенилаланин-β-метилового эфира» и второй стадией превращения α-L-аспартил-L-фенилаланин-β-метилового эфира в α-L-аспартил-L-фенилаланин-α-метиловый эфир.
Предпочтительные условия на первой стадии и тому подобное описаны в «<1> методе получения α-L-аспартил-L-фенилаланин-β-метилового эфира». Кроме того, вторая стадия может быть осуществлена в соответствии с известным способом и может быть сделана ссылка на способ и предпочтительные условия, описанные, например, в японской патентной заявке № H4-41155 и так далее. С помощью способа получения α-APM согласно настоящему изобретению α-APM, который важен в качестве подсластителя и тому подобное, может быть получен недорого и с высоким выходом.
Примеры
Далее здесь настоящее изобретение будет описано с помощью примеров. Однако настоящее изобретение не ограничено данными примерами. В добавление к подтверждению окраской нингидрином тонкослойных хроматограмм (качественной) были проведены количественные определения с помощью следующей жидкостной хроматографии высокого разрешения для анализа продуктов. Колонка: InertsiL ODS-2 (производства GL Science, Inc.), элюат: водный фосфатный раствор, содержащий смесь 5,0 мМ 1-октансульфоната натрия (pH 2,1):метанол=100:15 до 50, скорость потока 1,0 мл/мин, измерение 210 нанометров (нм).
Пример 1.
Микроорганизмы, которые продуцируют α-L-аспартил-L-фенилаланин-β-метиловый эфир
Для культивирования бактерий и актиномицетов, представленных в табл.1-1, применяли 50 миллилитров («мл») среды (pH 7,0), содержащей 20 грамм («г») глицерина, 5 г сульфата аммония, 1 г монокалийгидрофосфата, 3 г дикалийгидрофосфата, 0,5 г сульфата магния, 10 г дрожжевого экстракта и 10 г пептона в 1 литре (л), которую переносили в 500-мл сосуд Сакагуши и стерилизовали при 115°С в течение 15 минут (среда 1). Получали скошенный агар (pH 7,0), содержащий 5 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л NaCl и 20 г/л агара, и микроорганизмы, представленные в табл.1, культивировали в штамм скошенном агаре при 30°С в течение 24 часов. Затем собранные одной петлей микроорганизмы культивировали в среде 1 при 30°С в течение 24 часов и затем культивировали, встряхивая при 30°С и 120 встряхиваний/мин в течение 17 часов. После окончания культивирования клетки микроорганизма отделяли от культуральной жидкости центрифугированием и суспендировали в 0,1 М боратном буфере (pH 9,0), содержащем 10 мМ ЭДТА, до 100 г/л сырых клеток микроорганизма.
Для культивирования дрожжей, указанных в табл.1-1, применяли 50 мл среды (pH 6,0), содержащей 10 г/л глюкозы, 10 г глицерина, 5 г сульфата аммония, 1 г монокалийгидрофосфата, 3 г дикалийгидрофосфата, 0,5 г сульфата магния, 5 г дрожжевого экстракта 5 г экстракта солода и 10 г пептона в 1 л, перенесенной в 500-мл сосуд Сакагуши и стерилизованной при 115°С в течение 15 минут (среда 2). Получали приспособленную агаровую среду (pH 6,0), содержащую 5 г/л глюкозы, 5 г/л дрожжевого экстракта, 5 г/л экстракта солода, 10 г/л пептона, 5 г/л NaCl и 20 г/л агара в среде 2, и микроорганизмы, представленные в табл.1, культивировали в данной приспособленной среде агара при 30°С в течение 24 часов. Затем собранные одной петлей дрожжи культивировали при встряхивании при 30°С в течение 24 часов в среде 2 при 25°С и 120 встряхиваниях/мин в течение 17 часов. После окончания культивирования клетки микроорганизма отделяли от жидкостей данной культуры центрифугированием и суспендировали в 0,1 М боратном буфере (pH 9,0), содержащем 10 мМ ЭДТА, до 100 г/л сырых клеток микроорганизма.
Микроорганизмы, представленные в табл.1-2, культивировали следующим образом. Для культивирования Cellulophaga lytica NBRC 14961 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) или Flexithrix dorotheae NBRC 15987 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) применяли агаровую твердую среду (pH 7,2, стерилизованную при 120°С в течение 15 минут), содержащую 1 г триптона, 1 г дрожжевого экстракта и 15 г агара в 1 л искусственной морской воды Daigo SP. Высеянные клетки микроорганизма Cellulophaga lytica NBRC 14961 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) или Flexithrix dorotheae NBRC 15987 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала, после чего их наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов.
Для культивирования Weeksella virosa NBRC 16016 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) применяли агаровую среду с кровью овец (Nissui Plate, Nissui Pharmaceutical). Клетки микроорганизма Weeksella virosa NBRC 16016 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония), которые культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов.
Для культивирования Pedobacter heparinus NBRC 12017 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) применяли агаровую твердую среду (pH 7,0, стерилизованную при 120°С в течение 15 минут), содержащую 10 г пептона, 2 г дрожжевого экстракта, 1 г MgSO4·7H2O и 15 г агара в 1 л дистиллированной воды. Клетки микроорганизма Pedobacter heparinus NBRC 12017 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония), которые культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов.
Для культивирования Persicobacter diffluens NBRC 15940 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) применяли агаровую твердую среду (pH 7,0, стерилизованную при 120°С в течение 15 минут), содержащую 0,5 г KNO3, 0,1 г глицерофосфата натрия, 1 г трисгидроксиметиламинометана, 5 г триптона, 5 г дрожжевого экстракта, 15 г агара и 1 мл раствора микроэлементов в 1 л искусственной морской воды Daigo SP. Следует отметить, что раствор микроэлементов содержал 2,85 г H3BO4, 1,8 г MnCl2·4H2O, 1,36 г FeSO4·7H2O, 26,9 мг CuCl2·2H2O, 20,8 мг ZnCl2, 40,4 мг CoCl2·6H2O, 25,2 мг Na2MoO4·2H2O и 1,77 г тартрата натрия. Клетки микроорганизма Persicobacter diffluens NBRC 15940 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония), которые культивировали в данной среде при 25°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 25°С в течение 48 часов.
Для культивирования Chitinophaga pinensis NBRC 15968 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) применяли агаровую твердую среду (pH 7,0, стерилизованную при 120°С в течение 15 минут), содержащую 3 г бактоказитона, 1 г дрожжевого экстракта, 1,36 г CaCl2·H2O и 15 г агара в 1 л дистиллированной воды. Клетки микроорганизма Chitinophaga pinensis NBRC 15968 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония), которые культивировали в данной среде при 25°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 25°С в течение 48 часов.
Для культивирования Cyclobacterium marinum ATCC 25205 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) применяли агаровую твердую среду (pH 7,0, стерилизованную при 120°С в течение 15 минут), содержащую 5 г пептона, 1 г дрожжевого экстракта, 0,2 г FeSO4·7H2O и 15 г агара в 1 л искусственной морской воды Daigo SP. Клетки микроорганизма Cyclobacterium marinum ATCC 25205 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), которые культивировали в данной среде при 25°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 25°С в течение 48 часов.
Для культивирования Runella slithyformis ATCC 29530 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) применяли агаровую твердую среду (pH 7,0, стерилизованную при 120°С в течение 15 минут), содержащую 1 г пептона, 1 г дрожжевого экстракта, 1 г глюкозы и 15 г агара в 1 л дистиллированной воды. Клетки микроорганизма Runella slithyformis ATCC 29530 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), которые культивировали в данной среде при 25°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 25°С в течение 48 часов.
Для культивирования Thermonema lapsum ATCC 43542 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) применяли агаровую твердую среду (pH 8,2, стерилизованную при 120°С в течение 15 минут), содержащую 0,2 г нитрилтриуксусной кислоты, 2 мл 0,03% раствора FeCl3, 0,12 г CaSO4·2H2O, 0,2 г MgSO4·7H2O, 0,016 г NaCl, 0,21 г KNO3, 1,4 г NaNO3, 0,22 г Na2HPO4, 2 мл раствора микроэлементов и 15 г агара в 1 л дистиллированной воды. Следует отметить, что раствор микроэлементов содержал 0,5 мл H2SO4, 2,2 г MnSO4, 0,5 г ZnSO4, 0,5 г H3BO3, 0,016 г CuSO4, 0,025 г Na2MoO4 и 0,046 г CoCl2. Клетки микроорганизма Thermonema lapsum ATCC 43542 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), которые культивировали в данной среде при 60°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 25°С в течение 48 часов.
Для культивирования Gelidibacter algens ATCC 700364 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), Lewinella cohaerens ATCC 23123 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), Psychroserpens burtonensis ATCC 700359 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) или Salegentibacter salegens DSMZ 5424 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) применяли морской агар (производства Difco). В случае Gelidibacter algens ATCC 700364 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) или Psychroserpens burtonensis ATCC 700359 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) применяли клетки микроорганизма Gelidibacter algens ATCC 700364 или Psychroserpens burtonensis ATCC 700359 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), которые культивировали на данной среде при 10°С в течение 72 часов для получения семенного материала, с последующим основным культивированием при 10°С в течение 72 часов. В случае Lewinella cohaerens ATCC 23123 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) клетки микроорганизма Lewinella cohaerens ATCC 23123 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), которые культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов. В случае Salegentibacter salegens DSMZ 5424 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) клетки микроорганизма Salegentibacter salegens DSMZ 5424 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), которые культивировали в данной среде при 25°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 25°С в течение 48 часов.
Для культивирования Dyadobacter fermentans ATCC 700827 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) применяли агаровую твердую среду (pH 7,0, стерилизованную при 120°С в течение 15 минут), содержащую 0,8 г NH4Cl, 0,25 г KH2PO4, 0,4 г K2HPO4, 0,505 г KNO3, 15 мг CaCl2·2H2O, 20 мг MgCl2·6H2O, 7 мг FeSO4·7H2O, 5 мг Na2SO4, 5 мг MnCl2·4H2O, 0,5 мг H3BO3, 0,5 мг ZnCl2, 0,5 мг CoCl2·6H2O, 0,5 мг NiSO4·6H2O, 0,3 мг CuCl2·2H2O, 10 мг Na2MoO4·2H2O, 0,5 г дрожжевого экстракта, 0,5 г пептона, 0,5 г казаминовой кислоты, 0,5 г декстрозы, 0,5 г растворимого крахмала, 0,5 г пирувата натрия и 15 г агара в 1 л дистиллированной воды. Клетки микроорганизма Dyadobacter fermentans ATCC 700827 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), которые культивировали в данной среде при 25°С в течение 48 часов для получения семенного материала, затем подвергали основному культивированию при 25°С в течение 48 часов.
Для культивирования Flammeovirga aprica NBRC 15941 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) применяли агаровую твердую среду (pH 7,2, стерилизованную при 120°С в течение 15 минут), содержащую 2 г триптона, 0,5 г мясного экстракта, 0,5 г дрожжевого экстракта, 0,2 г ацетата натрия и 15 г агара в 1 л искусственной морской воды Daigo SP. Клетки микроорганизма Flammeovirga aprica NBRC 15941 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония), которые культивировали в этой среде при 25°С в течение 48 часов для получения семенного материала, затем подвергали основному культивированию при 25°С в течение 48 часов.
Для культивирования Spirosoma linguale DSMZ 74 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) или Flectobacillus major DSMZ 103 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) (pH 7,0, стерилизованную при 120°С в течение 15 минут), содержащую 1 г глюкозы, 1 г пептона, 1 г дрожжевого экстракта и 15 г агара в 1 л дистиллированной воды. Клетки микроорганизма Spirosoma linguale DSMZ 74 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) или Flectobacillus major DSMZ 103 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), которые культивировали в данной среде при 25°С в течение 48 часов для получения семенного материала, затем подвергали основному культивированию при 25°С в течение 48 часов.
Для культивирования Tenacibaculum maritimum ATCC 43398 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) применяли агаровую твердую среду (pH 7,0, стерилизованную при 120°С в течение 15 минут), содержащую 0,5 г триптона, 0,5 г дрожжевого экстракта, 0,2 г экстракта мяса, 0,2 г ацетата натрия и 15 г агара в 300 мл дистиллированной воды и 700 мл искусственной морской воды Daigo SP. Клетки микроорганизма Tenacibaculum maritimum ATCC 43398 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), которые культивировали в данной среде при 25°С в течение 48 часов для получения семенного материала, затем подвергали основному культивированию при 25°С в течение 48 часов.
Для культивирования Rhodothermus marinus DSMZ 4252 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) применяли агаровую твердую среду (pH 7,2, стерилизованную при 120°С в течение 15 минут), содержащую 2,5 г дрожжевого экстракта, 2,5 г триптона, 100 мг нитрилтриуксусной кислоты, 40 мг CaSO4·H2O, 200 мг MgCl2·6H2O, 0,5 мл 0,01 М цитрата Fe, 0,5 мл раствора микроэлементов, 100 мл фосфатного буфера, 900 мл дистиллированной воды 28 г агара в 1 л. Следует отметить, что раствор микроэлементов содержал 12,8 г нитрилтриуксусной кислоты, 1 г FeCl2·4H2O, 0,5 г MnCl2·4H2O, 0,3 г CoCl2·4H2O, 50 мг CuCl2·2H2O, 50 мг Na2MoO4·2H2O, 20 мг H3BO3 и 20 мг NiCl2·6H2O. Клетки микроорганизма Rhodothermus marinus DSMZ 4252 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), которые культивировали в данной среде при 60°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 60°С в течение 48 часов.
Для культивирования Zobellia galactanivorans DSMZ 12802 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) применяли агаровую твердую среду (1,5% агар, pH 7,6, стерилизованную при 120°С в течение 15 минут), содержащую бульон Bacto Marine BROTH (Difco 2216). Данную среду применяли с клетками микроорганизма Zobellia galactanivorans DSMZ 12802 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), которые культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала и наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов.
Для культивирования Muricauda ruestringenesis DSMZ 13258 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) применяли агаровую твердую среду (pH 7,2, стерилизованную при 120°С в течение 15 минут), содержащую 1,5 г дрожжевого экстракта, 2,5 г пептона, 2 г гексадекана, 17,7 г NaCl, 0,48 г KCl, 3,4 г MgCl2·6H2O, 4,46 г MgSO4·7H2O, 0,98 г CaCl2 и 15 г агара в 1 л дистиллированной воды. Клетки микроорганизма Muricauda ruestringenesis DSMZ 13258 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), которые культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов.
Для культивирования Taxeobacter gelupurpurascens DSMZ 11116 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) применяли агаровую твердую среду (pH 7,2, стерилизованную при 120°С в течение 15 минут), содержащую 3 г казитона, 1 г дрожжевого экстракта, 1,36 г CaCl2·2H2O и 15 г агара в 1 л дистиллированной воды. Клетки микроорганизма Taxeobacter gelupurpurascens DSMZ 11116 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), которые культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов.
Для культивирования Cytophaga hutchinsonii NBRC 15051 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) применяли агаровую твердую среду (pH 7,2, стерилизованную при 120°С в течение 15 минут), содержащую 3 г казитона, 1 г дрожжевого экстракта, 1,36 г CaCl2·2H2O, 5 г целлобиозы и 15 г агара в 1 л дистиллированной воды. Клетки микроорганизма Cytophaga hutchinsonii NBRC 15051 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония), которые культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов.
Для культивирования Marinilabilia salmonicolor NBRC 15948 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония) применяли агаровую твердую среду (pH 7,2, стерилизованную при 120°С в течение 15 минут), содержащую 10 г пептона, 2 г дрожжевого экстракта, 0,5 г MgSO4·7H2O и 15 г агара в 200 мл дистиллированной воды и 750 мл искусственной морской воды Daigo SP. Клетки микроорганизма Marinilabilia salmonicolor NBRC 15948 (учреждение-депозитарий; the NITE Biological Resource Center of the National Institute of Technology and Evaluation, адрес учреждения-депозитария; 5-8 Kazusa-Kamaashi 2-Chome, Kisarazu-shi, Chiba-ken, Япония), которые культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов.
Для культивирования Saprospira grandis ATCC 23119 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) применяли агаровую твердую среду (pH 7,0, стерилизованную при 120°С в течение 15 минут), содержащую 0,5 г KNO3, 0,1 г глицерофосфата натрия, 1 г трисгидроксиметиламинометана, 2 г триптона, 2 г дрожжевого экстракта, 15 г агара и 1 мл раствора микроэлементов в 1 л искусственной морской воды Daigo SP. Следует отметить, что раствор микроэлементов содержал 2,85 г H3BO4, 1,8 г MnCl2·4H2O, 1,36 г FeSO4·7H2O, 26,9 мг CuCl2·2H2O, 20,8 мг ZnCl2, 40,4 мг CoCl2·6H2O, 25,2 мг Na2MoO4·2H2O и 1,77 г тартрата натрия. Клетки микроорганизма Saprospira grandis ATCC 23119 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), которые культивировали в данной среде при 30°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 30°С в течение 48 часов.
Для культивирования Haliscomenobacter hydrossis ATCC 27775 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) применяли агаровую твердую среду (pH 7,5, стерилизованную при 120°С в течение 15 минут), содержащую 27 мг KH2PO4, 40 мг K2HPO4, 40 мг Na2HPO4·2H2O, 50 мг CaCl2·2H2O, 75 мг MgCl2·7H2O, 5 мг FeCl3·6H2O, 3 мг MnSO4·H2O, 1,31 г глутаминовой кислоты, 2,5 мг триптиказного бульона сои без глюкозы, 0,4 мг тиамина, 0,01 мг витамина B12, 2 г глюкозы и 1 мл раствора микроэлементов в 1 л дистиллированной воды. Следует отметить, что раствор микроэлементов содержал 0,1 г ZnSO4·7H2O, 0,03 г MnCl2·4H2O, 0,3 г H3BO3, 0,2 г CoCl2·6H2O, 0,01 г CuCl2·2H2O, 0,02 г NiCl2·6H2O и 0,03 г Na2MoO4·H2O.Клетки микроорганизма Haliscomenobacter hydrossis ATCC 27775 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), которые культивировали в данной среде при 25°С в течение 48 часов для получения семенного материала, наносили на ту же среду с последующим основным культивированием при 25°С в течение 48 часов.
Полученные таким образом клетки микроорганизма в каждом случае собирали с агаровой среды и суспендировали в 0,1 М боратном буфере (pH 9,0), содержащем 10 мМ ЭДТА, в количестве 100 г/л в виде невысушенных клеток микроорганизма.
К 0,1 мл каждой суспензии клеток этих микроорганизмов добавляли 0,1 мл 100 мМ боратного буфера (pH 9,0), содержащей 10 мМ ЭДТА, 100 мМ гидрохлорида α,β-диметилового эфира L-аспарагиновой кислоты и 200 мМ L-фенилаланина с доведением суммарного объема до 0,2 мл. Затем проводили реакцию при 20°С в течение 3 часов, когда применяли микроорганизм, указанный в табл.1-1, или в течение 1 часа, когда применяли микроорганизм, указанный в табл.1-2. Полученное количество (мМ) β-метилового эфира α-L-аспартил-L-фенилаланина (α-AMP) представлено на табл.1-1 и 1-2, определяли во всех случаях, когда применяли микроорганизмы. Причем во всех случаях использования микроорганизмов β-AMP не был зафиксирован.
Таблица 1-1

Таблица 1-2
Справочный пример 1
Микроорганизм, который продуцирует α-метиловый эфир β-L-аспартил-L-фенилаланина
Микроорганизмы, представленные в табл.2, культивировали сходно с процедурой для бактерий табл.1 примера 1. После завершения культивирования клетки микроорганизма выделяли из данных бульонов культивирования с помощью центрифугирования и суспендировали в 0,1 М боратном буфере (pH 9,0), содержащем 10 мМ ЭДТА до 100 г/л сырых клеток микроорганизма. К 0,1 мл каждой из микроорганизмных суспензий данных микроорганизмов добавляли 0,1 мл 100 мМ боратного буфера (рН 9,0), содержащего 10 мМ ЭДТА, 100 мМ гидрохлорида α,β-диметилового эфира L-аспарагиновой кислоты и 200 мМ L-фенилаланина для получения суммарного объема 0,2 мл с последующей реакцией при 30°С в течение 2 часов. Количество (мМ) α-метилового эфира β-L-аспартил-L-фенилаланина (β-AMP), продуцированного в данном случае, представлено в табл.2. Следует отметить, что α-AMP выявлялся во всех микроорганизмах.
Пример 2
Очистка фермента из Empedobacter brevis
50 мл среды (pH 6,2), содержащей 5 грамм (г) глюкозы, 5 г сульфата аммония, 1 г монокалийфосфата, 3 г дикалийфосфата, 0,5 г сульфата магния, 10 г дрожжевого экстракта и 10 г пептона в 1 л, переносили в 500-мл сосуд Сакагуши и стерилизовали при 115°С в течение 15 минут. Данную среду затем инокулировали 2 миллилитрами (мл) штамма FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002 г.), который культивировали при 30°С в течение 16 часов в той же среде и затем культивировали при встряхивании при 30°С и 120 встряхиваниях/мин в течение 16 часов.
Последующую после отделения центрифугированием процедуру проводили либо на льду, либо при 4°С. Полученный культуральный бульон центрифугировали для сбора клеток микроорганизма. После промывки 16 г клеток микроорганизма 50 мМ трис-HCl буфером (pH 8,0) их суспендировали в 40 миллилитрах (мл) того же буфера и подвергали обработке разрушением ультразвуком в течение 45 минут при 195 Вт. Разрушенную ультразвуком жидкость затем центрифугировали (10000 об/мин, 30 минут) для удаления фрагментов разрушенных клеток и получения супернатанта разрушенной ультразвуком жидкости. Данный супернатант разрушенной ультразвуком жидкости диализовали в течение ночи против 50 мМ трис-HCl буфера (pH 8,0) с последующим удалением нерастворимой фракции ультрацентрифугированием (50000 об/мин, 30 минут) с получением растворимой фракции в форме надосадочной жидкости. Полученную растворимую фракцию наносили на колонку Q-сефарозы HP (произведенную Amersham), предварительно уравновешенную трис-HCl буфером (pH 8,0), и из неадсорбированной фракции собирали активную фракцию. Данную активную фракцию диализовали в течение ночи против 50 мМ ацетатного буфера (pH 4,5) с последующим удалением нерастворимой фракции с помощью разделения центрифугированием (10000 об/мин, 30 минут) с получением диализованной фракции в форме надосадочной жидкости. Данную диализованную фракцию затем наносили на колонку моно S (произведенную Amersham), предварительно уравновешенную 50 мМ ацетатным буфером (pH 4,5) для элюирования фермента линейным градиентом концентрации того же буфера, содержащего от 0 до 1 М NaCl. Из активных фракций фракцию, содержащую наименьший уровень загрязняющего белка, наносили на колонку Superdex 200 pg (произведенную Amersham), предварительно уравновешенную 50 мМ ацетатным буфером (pH 4,5), содержащим 1 М NaCl и гель-фракцию, осуществляли с тем же буфером (рН 4,5), содержащим 1М NaCl, который пропускали для свободного прохождения через колонку с получением раствора активной фракции. В результате проведения данных процедур с помощью экспериментальных данных электрофореза было подтверждено, что фермент, участвующий в образовании пептида, применяемый в настоящем изобретении, был очищен до однородного состояния. Степень выхода фермента в указанном выше процессе очистки составляла 12,2%, а степень очистки была 707-кратной.
Пример 3
Получение β-метилового эфира α-L-аспартил-L-фенилаланина с применением ферментативной фракции Empedobacter brevis
10 микролитров (мкл) фракции фермента Mono S (приблизительно 20 Е/мл), полученной в примере 2, добавляли к 190 мкл боратного буфера (pH 9,0), содержащего 105,3 мМ гидрохлорида α,β-диметилового эфира L-аспарагиновой кислоты, 210,5 мМ L-фенилаланина и 10,51 мМ ЭДТА, и реакцию проводили при 20°С. Ход получения β-метилового эфира α-L-аспартил-L-фенилаланина (α-AMP) показан в табл.3. Следует обратить внимание на то, что практически полное отсутствие образования β-метилового эфира α-L-аспартил-L-фенилаланина было подтверждено с партией без добавления фермента.
Более того, 10 мкл фракции фермента Mono S (приблизительно 20 Е/мл), полученной в примере 2, добавляли к 190 мкл боратного буфера (pH 9,0), содержащего 105,3 мМ гидрохлорида α-метилового эфира L-аспарагиновой кислоты, 210,5 мМ L-фенилаланина и 10,51 мМ ЭДТА, и реакцию проводили при 20°С. В результате не наблюдалось образования соответствующих пептидов.
Пример 4
Очистка фермента из Sphingobacterium sp.
Штамм FERM BP-8124 Sphingobacterium sp. (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 22 июля 2002 г.) культивировали тем же способом, что и в примере 2, с применением среды, указанной в примере 2. Последующую процедуру после разделения центрифугированием проводили либо на льду, либо при 4°С. Полученный культуральный бульон центрифугировали (10000 об/мин, 15 минут) для сбора клеток микроорганизма. После промывки 2 г клеток микроорганизма 20 мМ трис-HCl буфером (pH 7,6) их суспендировали в 8 мл того же буфера и подвергали обработке ультразвуковым разрушением в течение 45 минут при 195 Вт. Разрушенную ультразвуком жидкость затем центрифугировали (10000 об/мин, 30 минут) для удаления фрагментов разрушенных клеток и получения супернатанта разрушенной ультразвуком жидкости. Данный супернатант разрушенной ультразвуком жидкости диализовали в течение ночи против 20 мМ трис-HCl буфера (pH 7,6) с последующим удалением нерастворимой фракции ультрацентрифугированием (50000 об/мин, 30 минут) с получением растворимой фракции в форме надосадочной жидкости. Полученную растворимую фракцию наносили на колонку Q-сефарозы HP (произведенную Amersham), предварительно уравновешенную трис-HCl буфером (pH 7,6), и из неадсорбированной фракции собирали активную фракцию. Данную активную фракцию диализовали в течение ночи против 20 мМ ацетатного буфера (pH 5,0) с последующим удалением нерастворимой фракции с помощью разделения центрифугированием (10000 об/мин, 30 минут) с получением диализованной фракции в форме надосадочной жидкости. Данную диализованную фракцию затем наносили на колонку SP-сефарозы HP (произведенную Amersham), предварительно уравновешенную 20 мМ ацетатным буфером (pH 5,0) с получением активной фракции, в которой фермент элюировался линейным градиентом концентрации того же буфера, содержащего от 0 до 1 М NaCl.
Пример 5
Получение β-метилового эфира α-L-аспартил-L-фенилаланина и β-этилового эфира α-L-аспартил-L-фенилаланина с применением ферментативной фракции Sphingobacterium sp.
В случае получения β-метилового эфира α-L-аспартил-L-фенилаланина (α-AMP) 15 мкл концентрированного раствора фракции SP-сефарозы HP (приблизительно 15 Е/мл), полученной в примере 4, добавляли к 185 мкл боратного буфера (pH 9,0), содержащего 108,1 мМ гидрохлорида α,β-диметилового эфира L-аспарагиновой кислоты, 216,2 мМ L-фенилаланина и 10,8 мМ ЭДТА, и реакцию проводили при 20°С. Сходным образом, в случае получения β-этилового эфира α-L-аспартил-L-фенилаланина (α-AEP) 10 мкл концентрированного раствора фракции SP-сефарозы HP (приблизительно 15 Е/мл), полученной в примере 4, добавляли к 190 мкл боратного буфера (pH 9,0), содержащего 52,6 мМ гидрохлорида α,β-диэтилового эфира L-аспарагиновой кислоты, 105,2 мМ L-фенилаланина и 10,8 мМ ЭДТА, и реакцию проводили при 20°С. Ход получения AMP или AEP показан в табл.4. Следует обратить внимание на то, что практически полное отсутствие образования AMP или AEP было подтверждено с партией без добавления фермента. Для образования AEP описаны численные величины, полученные с применением стандартного продукта AMP.
Пример 6
Выделение образующего пептид фермента, генетически происходящего от Empedobacter brevis
Далее здесь будет объяснено выделение гена образующего пептид фермента. Штамм FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002 г.) применяли в качестве микроорганизма. При выделении гена в качестве хозяина применяли Escherichia coli JM-109, а в качестве вектора применяли pUC118.
(1) Получение праймера ПЦР на основе определенной внутренней аминокислотной последовательности
Смешанный праймер, имеющий последовательности оснований, указанных в SEQ ID NO.: 3 и SEQ ID NO: 4 соответственно, были получены на основании аминокислотных последовательностей (SEQ ID NOs: 1 и 2), определенных согласно способу разложения по Эдману, продукта гидролиза лизиловой эндопептидазы образующего пептид фермента, берущего начало от штамма FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002).
(2) Получение клеток микроорганизма
Штамм FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002) культивировали при 30°С в течение 24 часов на агаровой среде CM2G (содержащей 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну петлю полученных клеток микроорганизма инокулировали в 500-мл сосуд Сакагуши, содержащий 50 мл жидкой среды CM2G (упомянутой выше среды, исключая агар), с последующим культивированием со встряхиванием при 30°С.
(3) Получение хромосомной ДНК из клеток микроорганизма
50 мл культурального бульона центрифугировали (12000 об/мин, 4°С, 15 минут) для сбора клеток микроорганизма. Затем из клеток микроорганизма получали хромосомную ДНК с применением QIAGEN Genomic-Tip System (Qiagen) на основании процедуры, описанной ее производителем.
(4) Получение фрагмента ДНК, содержащего часть гена образующего пептид фермента с помощью ПЦР
Фрагмент ДНК, содержащий часть гена образующего пептид фермента, берущего начало от штамма FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002), получали с помощью способа ПЦР с применением LA-Taq (производства Takara Shuzo). Реакцию ПЦР проводили на хромосомной ДНК, полученной из штамма FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002), с применением праймеров, имеющих последовательности SEQ ID NOs: 3 и 4.
Реакцию ПЦР проводили на протяжении 30 циклов при следующих условиях с применением Tekara PCR Thermal Cycler PERSONAL (производства Takara Shuzo).
После завершения реакции 3 мкл реакционной жидкости наносили на 0,8% агарозу для электрофореза. В результате было подтверждено, что амплифицируется фрагмент ДНК из приблизительно 1,5 тысячи оснований (т.н.)
(5) Клонирование гена образующего пептид фермента из генной библиотеки
Для получения полноразмерного гена образующего пептид фермента была проведена гибридизация по Саузерну с применением в качестве зонда фрагмента ДНК, амплифицированного в процедуре ПЦР. Процедура для гибридизации по Саузерну объяснена в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Фрагмент ДНК приблизительно 1,5 т.н., амплифицированный с помощью процедуры ПЦР, выделяли с помощью электрофореза в 0,8% агарозе. Желаемую полосу затем вырезали и очищали. Данный фрагмент ДНК метили с помощью зонда дигоксинигена с применением DIG High Prime (производства Boehringer-Mannheim) на основе процедуры, описанной в руководстве для применения DIG High Prime (производства Boehringer-Mannheim).
После завершения гидролиза хромосомной ДНК Empedobacter brevis, полученной на стадии (3) настоящего примера 6, с помощью реакции при 37°С в течение 16 часов с ферментом рестрикции HindIII полученный продукт подвергали электрофорезу в 0,8% агарозном геле. Разделенную электрофорезом хромосомную ДНК переносили после электрофореза из агарозного геля на положительно заряженный фильтр нейлоновой мембраны (производства Roche Diagnostics) с последующей обработкой, состоящей из щелочной денатурации, нейтрализации и иммобилизации. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 50°С в течение 1 часа добавляли зонд, меченный дигоксинигеном, полученный, как описано выше, и гибридизацию проводили при 50°С в течение 16 часов. Затем фильтр промывали в течение 20 минут при комнатной температуре 2 x SSC, содержащим 0,1% SDS. Более того, фильтр дополнительно промывали дважды при 65°С в течение 15 минут 0,1 x SSC, содержащим 0,1% SDS.
Выявление полос, которые гибридизовались с зондом, производили с помощью DIG Nucleotide Detection Kit (производства Boehringer-Mannheim). В результате была выявлена полоса приблизительно 4 т.н., способная гибридизоваться с зондом.
После этого 5 мкг хромосомной ДНК, полученной на стадии (3) настоящего примера 6, полностью гидролизовали с помощью HindIII. ДНК приблизительно 4 т.н. отделяли с помощью электрофореза в 0,8% агарозном геле и затем ДНК очищали с помощью Gene Clean II Kit (производства Funakoshi) и растворяли в 10 мкл TE. 4 мкл данного продукта затем смешивали с pUC118 HindIII/BAP (производства Takara Shuzo) и проводили реакцию лигирования с помощью DNA Ligation Kit Ver. 2 (производства Takara Shuzo). 5 мкл смеси реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM109 (производства Toyobo) смешивали для трансформации Escherichia coli. Содержимое затем наносили на подходящую твердую среду для получения библиотеки хромосомной ДНК.
Для получения цельного полноразмерного гена образующего пептид фермента библиотеку хромосомной ДНК подвергали скринингу путем гибридизации колоний, как объяснено в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии библиотеки хромосомной ДНК переносили на фильтр нейлоновой мембраны (Nylon Membrane for Colony and Plaque Hybridization (производства Roche Diagnostics) с последующей обработкой, состоявшей из щелочной денатурации, нейтрализации и иммобилизации. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 37°С в течение 1 часа добавляли указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 50°С в течение 16 часов. Затем фильтр промывали в течение 20 минут при комнатной температуре 2 x SSC, содержащим 0,1% SDS. Более того, фильтр дополнительно промывали дважды при 65°С в течение 15 минут 0,1 x SSC, содержащим 0,1% SDS.
Выявление колоний, которые гибридизовались с меченым зондом, производили с помощью DIG Nucleotide Detection Kit (производства Boehringer-Mannheim) на основе объяснения, описанного в руководстве по применению DIG Nucleotide Detection Kit (производства Boehringer-Mannheim). В результате была выявлена окраска двух колоний, способных гибридизоваться с меченым зондом.
(6) Последовательность оснований гена образующего пептид фермента, происходящего от Empedobacter brevis
Плазмиды, содержащиеся в Escherichia coli JM109, получали из вышеупомянутых двух клонов клеток микроорганизма, которые, как было подтверждено с помощью Wizard Plus Minipreps DNA Purification System (производства Promega), гибридизовались с меченым зондом, и была определена последовательность оснований части, где имела место гибридизация, и поблизости от нее. Реакцию секвенирования проводили с применением CEQ DTCS-Quick Start Kit (производства Beckman-Coulter) на основе процедуры, описанной в руководстве к нему. Кроме того, проводили электрофорез с применением CEQ 2000-XL (производства Beckman-Coulter).
В результате было подтверждено, что рамка с открытым считыванием, которая кодирует белок, содержащий внутренние аминокислотные последовательности образующего пептид фермента (SEQ ID NOs: 1 и 2), действительно существует, что тем самым подтвердило то, что рамка с открытым считыванием представляет собой ген, кодирующий образующий пептид фермент. Последовательность оснований полноразмерных генов образующего пептид фермента, наряду с соответствующими аминокислотными последовательностями, показана в SEQ ID NO: 5 перечня последовательностей. По результатам анализа гомологии полученной открытой рамки считывания, проведенного с помощью программы BLASTP, была обнаружена гомология между двумя ферментами; он показал гомологию 34% на уровне аминокислотной последовательности, проявляемой гидролазой сложных эфиров α-аминокислот Acetobacter pasteurianus (смотри Appl. Environ. Microbiol., 68(1), 211-218 (2002), и 26% гомология на уровне аминокислотной последовательности, проявляемой глутарил-7ACA ацилазой Brevibacillus laterosporum (смотри J. Bacteriol., 173(24), 7848-7855 (1991).
(7) Экспрессия гена образующего пептид белка, берущего начало от Empedobacter brevis, в Escherichia coli
Область гена-мишени в промоторной области оперона trp хромосомной ДНК Escherichia coli W3110 амплифицировали проведением ПЦР с применением олигонуклеотидов, указанных в SEQ ID NOs: 7 и 8, в качестве праймеров и полученные фрагменты ДНК лигировали с вектором pGEM-Teasy (производства Promega). Затем E. coli JM109 трансформировали в данном растворе лигирования и из устойчивых к ампициллину штаммов отбирали клоны, содержащие желаемую плазмиду, в которой вставленный промотор trp находился в ориентации, противоположной ориентации промотора lac. Затем фрагмент ДНК, содержащий промотор trp, полученный путем обработки этой плазмиды EcoO109/EcoRI, лигировали с EcoO109/EcoRI, продуктом обработки pUC19 (производства Takara). Затем фрагмент ДНК, полученный обработкой данной плазмиды HindIII/PvuII, лигировали с фрагментом ДНК, содержащим терминатор rrnB, полученный обработкой pKK223-3 (производства Amersham Pharmacia) HindIII/HincII. Затем E. coli JM109 трансформировали данным раствором лигирования, штаммы, содержащие желаемую плазмиду, отбирали из устойчивых к ампициллину штаммов, и плазмида была обозначена как pTrpT.
Желаемый ген амплифицировали с помощью ПЦР с применением хромосомной ДНК штамма FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002 г.) в качестве матрицы и олигонуклеотидов, указанных в SEQ ID NO: 9 и 10, в качестве праймеров. Данный фрагмент ДНК затем обрабатывали NdeI/PstI и полученный фрагмент ДНК лигировали с продуктом обработки NdeI/PstI pTrpT. Затем Escherichia coli JM109 трансформировали данным раствором лигирования, штаммы, содержащие желаемую плазмиду, отбирали из устойчивых к ампициллину штаммов, и плазмида была обозначена как pTrpT_Gtg2.
Escherichia coli JM109, содержащую pTrpT_Gtg2, культивировали для получения семенного материала при 30°С в течение 24 часов в среде LB, содержащей 100 мг/л ампициллина. 1 мл полученного бульона культуры высевали в 500-мл сосуд Сакагуши, содержащий 50 мл среды (2 г/л D глюкозы, 10 г/л дрожжевого экстракта, 10 г/л казаминокислот, 5 г/л сульфата аммония, 3 г/л монокалийгидрофосфата, 1 г/л дикалийгидрофосфата, 0,5 г/л гептагидрата сульфата магния и 100 мг/л ампициллина), с последующим культивированием при 25°С в течение 24 часов. Культуральный бульон обладал активностью образования β-метилового эфира α-L-аспартил-L-фенилаланина 0,11 Е на 1 мл культурального бульона, и было подтверждено, что клонированный ген экспрессируется E. coli. Более того, активность не была обнаружена в трансформанте, в который была введена только pTrpT в качестве контроля.
Предсказание сигнальной последовательности
Когда аминокислотная последовательность SEQ ID NO: 6, описанная в перечне последовательностей, была проанализирована с помощью программы Signal P v 1.1 (смотри Protein Engineering, Vol. 12, No. 1, pp. 3-9, 1999), было предсказано, что аминокислоты с номера 1 по 22 действуют в качестве сигнала, который секретируется в периплазму, а зрелый белок, как было установлено, располагается ниже аминокислоты с номером 23.
Подтверждение секреции
Escherichia coli JM109, содержащую pTrpT_Gtg2, культивировали для получения семенного материала при 30°С в течение 24 часов в среде LB, содержащей 100 мг/л ампициллина. 1 мл полученного бульона культуры высевали в 500-мл сосуд Сакагуши, содержащий 50 мл среды (2 г/л D глюкозы, 10 г/л дрожжевого экстракта, 10 г/л казаминокислот, 5 г/л сульфата аммония, 3 г/л монокалийгидрофосфата, 1 г/л дикалийгидрофосфата, 0,5 г/л гептагидрата сульфата магния и 100 мг/л ампициллина), с последующим культивированием при 25°С в течение 24 часов для получения культуры клеток микроорганизма.
Культивированные клетки микроорганизма фракционировали на фракцию периплазмы и цитоплазматическую фракцию с помощью способа шокового осмотического давления с применением раствора сахарозы 20 грамм/децилитр (г/дл). Клетки микроорганизма, погруженные в раствор сахарозы 20 г/дл, погружали в 5 мМ водный раствор MgSO4. Супернатант после центрифугирования был назван периплазматической фракцией. ("Pe"). Кроме того, осадок после центрифугирования ресуспендировали и подвергали ультразвуковому разрушению. Полученное было названо цитоплазматической фракцией ("Cy"). Активность глюкозо-6-фосфатдегидрогеназы, которая, как известно, находится в цитоплазме, применяли в качестве индикатора для подтверждения отделения цитоплазмы. Данное измерение проводили добавлением подходящего количества фермента к реакционному раствору при 30°С, содержащему 1 мМ глюкозо-6-фосфат, 0,4 мМ НАДФ, 10 мМ MgSO4 и 50 мМ трис-HCl (pH 8), с последующим измерением поглощения при 340 нм для измерения образования НАДФН.
Количество ферментов в периплазматической фракции и цитоплазматической фракции, когда активность полученного бесклеточного экстракта была принята за 100%, показано на чертеже. То, что активность глюкозо-6-фосфатдегидрогеназы не смешивалась с периплазматической фракцией, указывает на то, что периплазматическая фракция не была смешана с цитоплазматической фракцией. Приблизительно 60% активности, образующей β-метиловый эфир α-L-аспартил-L-фенилаланина (α-AMP), было обнаружено в периплазматической фракции, и было подтверждено, что образующий Ala-Gln фермент секретируется в периплазму, как это было предсказано на основании аминокислотной последовательности с помощью программы Signal P v 1.1.
Пример 7
Выделение гена образующего пептид фермента, происходящего от Sphingobacterium sp.
Далее здесь описывается выделение гена образующего пептид фермента. Примененный микроорганизм представлял собой штамм FERM BP-8124 Sphingobacterium sp. (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 22 июля 2002 г.). Для выделения гена в качестве хозяина применяли Escherichia coli DH5α, а в качестве вектора использовали pUC118.
(1) Получение клеток микроорганизма
Штамм FERM BP-8124 Sphingobacterium sp. (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 22 июля 2002 г.) культивировали в течение 24 часов при 25°С на агаровой среде CM2G (содержащей 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну петлю полученных клеток микроорганизма инокулировали в 500-мл сосуд Сакагуши, содержащий 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара) с последующим культивированием со встряхиванием при 25°С.
(2) Получение хромосомной ДНК из клеток микроорганизма
50 мл культурального бульона центрифугировали (12000 об/мин, 4°С, 15 минут) для сбора клеток микроорганизма. Затем из клеток микроорганизма получали хромосомную ДНК с применением Qiagen Genomic-Tip System (Qiagen) на основании процедуры, описанной ее производителем.
(3) Получение фрагмента ДНК зонда с помощью ПЦР
Фрагмент ДНК, содержащий часть гена образующего пептид фермента, берущего начало от штамма FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002), получали с помощью способа ПЦР с применением LA-Taq (производства Takara Shuzo). Затем проводили реакцию ПЦР на хромосомной ДНК, полученной из штамма FERM BP-8113 Empedobacter brevis (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 8 июля 2002), с применением праймеров, имеющих последовательности оснований SEQ ID NOs: 3 и 4.
Реакцию ПЦР проводили с применением Takara PCR Thermal Cycler - PERSONAL (Takara Shuzo) на протяжении 30 циклов при следующих условиях.
После завершения реакции 3 мкл реакционной жидкости наносили на 0,8% агарозу для электрофореза. В результате было подтверждено, что амплифицируется фрагмент ДНК из приблизительно 1,5 т.н.
(4) Клонирование гена образующего пептид фермента из библиотеки генов
Для получения полноразмерного гена образующего пептид фермента была проведена гибридизация по Саузерну с применением в качестве зонда фрагмента ДНК, амплифицированного в указанной выше процедуре ПЦР. Операции гибридизации по Саузерну объяснены в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Фрагмент ДНК приблизительно 1,5 т.н., амплифицированный с помощью указанной выше процедуры ПЦР, выделяли с помощью электрофореза в 0,8% агарозе. Желаемую полосу затем вырезали и очищали. Данный фрагмент ДНК метили с помощью зонда дигоксинигена с применением DIG High Prime (производства Boehringer-Mannheim) на основе процедуры, описанной в руководстве.
После взаимодействия хромосомной ДНК Sphingobacterium sp., полученной на стадии (2) настоящего примера 7, с ферментом рестрикции SacI при 37°С в течение 16 часов до полного гидролиза ДНК полученное подвергали электрофорезу в 0,8% агарозном геле. После электрофореза разделенную электрофорезом ДНК из агарозного геля переносили на положительно заряженный фильтр нейлоновой мембраны (производства Roche Diagnostics) с последующей обработкой, состоящей из щелочной денатурации, нейтрализации и иммобилизации. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 37°С в течение 1 часа добавляли зонд, меченный дигоксинигеном, полученный, как описано выше, и гибридизацию проводили при 37°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление полос, которые гибридизовались с зондом, производили с помощью DIG Nucleotide Detection Kit (производства Boehringer-Mannheim) на основе процедуры, описанной в его руководстве. В результате была успешно выявлена полоса приблизительно 3 т.н., которая гибридизовалась с зондом.
5 мкг хромосомной ДНК, полученной на стадии (2) настоящего примера 7, полностью гидролизовали с помощью SacI. ДНК приблизительно 3 т.н. отделяли с помощью электрофореза в 0,8% агарозном геле, ДНК очищали с помощью Gene Clean II Kit (производства Funakoshi) и растворяли в 10 мкл TE. Смешивали 4 мкл полученного раствора и pUC118, обработанной щелочной фосфатазой (E. coli C75) при 37°С в течение 30 минут и 50°С в течение 30 минут после полного гидролиза с помощью SacI при 37°С в течение 16 часов, и проводили реакцию лигирования с помощью DNA Ligation Kit Ver. 2 (производства Takara Shuzo). 5 мкл жидкости реакции лигирования и 100 мкл компетентных клеток Escherichia coli DH5α (производства Takara Shuzo) смешивали для трансформации Escherichia coli. Содержимое затем наносили на подходящую твердую среду для получения библиотеки хромосомной ДНК.
Для получения полноразмерного гена образующего пептид фермента библиотеку хромосомной ДНК подвергали скринингу путем гибридизации колоний с применением указанного выше зонда. Процедура для гибридизации колоний объяснена в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии библиотеки хромосомной ДНК переносили на нейлоновый мембранный фильтр (Nylon Membrane for Colony and Plaque Hybridization, производства Roche Diagnostics) с последующей обработкой щелочной денатурацией, нейтрализацией и иммобилизацией. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 37°С в течение 1 часа добавляли указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 37°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление колоний, которые гибридизовались с меченым зондом, производили с помощью DIG Nucleotide Detection Kit (производства Boehringer-Mannheim) на основе объяснения, описанного в его руководстве. В результате было выявлено шесть штаммов колоний, способных гибридизоваться с меченым зондом.
(5) Последовательность оснований гена образующего пептид фермента, происходящего от Sphingobacterium sp.
Плазмиды, содержащиеся в Escherichia coli DH5α, получали из шести клонов клеток микроорганизма, которые, как было подтверждено с помощью Wizard Plus Minipreps DNA Purification System (производства Promega), гибридизовались с меченым зондом, для определения последовательности оснований части, где имела место гибридизация с зондом и поблизости от нее. Реакцию секвенирования проводили с применением CEQ DTCS-Quick Start Kit (производства Beckman-Coulter) на основе процедуры, описанной в руководстве к нему. Кроме того, проводили электрофорез с применением CEQ 2000-XL (производства Beckman-Coulter).
В результате было подтверждено, что рамка с открытым считыванием, которая кодирует образующий пептид фермент, действительно существует. Полноразмерная последовательность оснований гена образующего пептид фермента из Sphingobacterium sp., наряду с соответствующей аминокислотной последовательностью, показана в SEQ ID NO: 11. Образующий пептид фермент, происходящий от Sphingobacterium sp., проявлял гомологию 63,5 % на уровне аминокислотной последовательности с происходящим от Empedobacter brevis образующим пептид ферментом (как определено с помощью программы BLASTP).
(6) Экспрессия гена образующего пептид фермента, берущего начало от Sphingobacterium sp., в Escherichia coli
Желаемый ген амплифицировали с помощью ПЦР с применением хромосомной ДНК Sphingobacterium sp. FERM BP-8124 (институт-депозитарий: независимая в управлении корпорация, национальный институт развития промышленной науки и технологии, международный депозитарий запатентованных организмов, адрес института-депозитария: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Япония, дата международного перевода депозита: 22 июля 2002) в качестве матрицы и олигонуклеотидов, указанных в SEQ ID NOs: 13 и 14, в качестве праймеров. Данный фрагмент ДНК обрабатывали NdeI/XbaI и полученный фрагмент ДНК лигировали с продуктом обработки NdeI/XbaI pTrpT. Затем Escherichia coli JM109 трансформировали данным раствором лигирования, и штаммы, содержащие желаемую плазмиду, отбирали из устойчивых к ампициллину штаммов. Плазмида была обозначена как pTrpT_Sm_aet.
Escherichia coli JM109, содержащую pTrpT_Sm_aet, культивировали при 25°С в течение 20 часов путем инокуляции одной ее петли в обычную пробирку для тестирования, содержащую 3 мл среды (2 г/л D глюкозы, 10 г/л дрожжевого экстракта, 10 г/л казаминокислот, 5 г/л сульфата аммония, 3 г/л монокалийгидрофосфата, 1 г/л дикалийгидрофосфата, 0,5 г/л гептагидрата сульфата магния и 100 мг/л ампициллина). Было подтверждено, что клонированный ген, обладающий активностью в отношении образования α-AMP 0,53 Е на мл культурального бульона, экспрессируется Escherichia. coli. Более того, активность не была обнаружена в трансформанте, содержащем только pTrpT в качестве контроля.
Предсказание сигнальной последовательности
Когда аминокислотная последовательность SEQ ID NO: 12, описанная в перечне последовательностей, была проанализирована с помощью программы Signal P v 1.1 (смотри Protein Engineering, Vol. 12, No. 1, pp. 3-9, 1999), было предсказано, что аминокислоты с номера 1 по 20 действуют в качестве сигнала, который секретируется в периплазму, а зрелый белок, как было установлено, располагается ниже аминокислоты с номером 21.
Подтверждение сигнальной последовательности
Одну петлю Escherichia coli JM109, содержащую pTrpT_Sm_aet, инокулировали в обычные пробирки для тестирования, содержащие 50 мл среды (2 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л казаминокислот, 5 г/л сульфата аммония, 3 г/л монокалийгидрофосфата, 1 г/л дикалийгидрофосфата, 0,5 г/л гептагидрата сульфата магния и 100 мг/л ампициллина), и основное культивирование проводили при 25°С в течение 20 часов.
Далее здесь процедуры после разделения центрифугированием проводили либо на льду, либо при 4°С. После завершения культивирования клетки микроорганизма отделяли от культурального бульона центрифугированием, промывали 100 мМ фосфатным буфером (pH 7) и затем суспендировали в том же буфере. Клетки микроорганизма затем подвергали обработке ультразвуковым разрушением в течение 20 минут при 195 Вт, разрушенную ультразвуком жидкость центрифугировали (12000 об/мин, 30 минут) для удаления фрагментов разрушенных клеток и получения растворимой фракции. Полученную растворимую фракцию наносили на колонку CHT-II (производства Biorad), предварительно уравновешенную 100 мМ фосфатным буфером (pH 7), и фермент элюировали линейным градиентом концентрации 500 мМ фосфатного буфера. Раствор, полученный смешиванием активной фракции с 5-кратными объемами 2 М сульфата аммония и 100 мМ фосфатного буфера, наносили на колонку Resource-PHE (производства Amersham), предварительно уравновешенную 2 М сульфатом аммония и 100 мМ фосфатным буфером, и фермент элюировали линейным градиентом концентрации от 2 до 0 М сульфата аммония с получением раствора активной фракции. В результате данных процедур было подтверждено, что образующий пептид фермент был очищен до электрофоретической однородности.
Когда аминокислотная последовательность упомянутого выше образующего пептид фермента была определена с помощью способа разложения по Эдману, была получена аминокислотная последовательность SEQ ID NO: 15 и было подтверждено, что зрелый белок располагается после аминокислоты номер 21, как это было предсказано с помощью программы Signal P v 1.1.
Пример 8
Выделение гена образующего пептид фермента из Pedobacter heparinus IFO 12017
Далее здесь описывается выделение гена образующего пептид фермента. Примененный микроорганизм представлял собой Pedobacter heparinus IFO 12017 (институт-депозитарий: Institute of Fermentation, Osaka, адрес института-депозитария: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония). Для выделения гена в качестве хозяина применяли Escherichia coli JM109, а в качестве вектора использовали pUC118.
(1) Получение клеток микроорганизма
Pedobacter heparinus IFO 12017 (институт-депозитарий: Institute of Fermentation, Osaka, адрес института-депозитария: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) культивировали в течение 24 часов при 25°С на агаровой среде CM2G (содержащей 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну петлю полученных клеток микроорганизма инокулировали в 500-мл сосуд Сакагуши, содержащий 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара) с последующим культивированием со встряхиванием при 25°С.
(2) Получение хромосомной ДНК из клеток микроорганизма
50 мл культурального бульона центрифугировали (12000 об/мин, 4°С, 15 минут) для сбора клеток микроорганизма. Затем из клеток микроорганизма получали хромосомную ДНК с применением Qiagen Genomic-Tip System (Qiagen) на основании процедуры, описанной ее производителем.
(3) Получение фрагмента ДНК зонда с помощью ПЦР
Фрагмент ДНК, содержащий часть гена образующего пептид фермента, берущего начало от Pedobacter heparinus IFO 12017 (институт-депозитарий: Institute of Fermentation, Osaka, адрес института-депозитария: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония), получали с помощью способа ПЦР с применением LA-Taq (производства Takara Shuzo). Затем проводили реакцию ПЦР на хромосомной ДНК, полученной из Pedobacter heparinus IFO 12017 (институт-депозитарий: Institute of Fermentation, Osaka, адрес института-депозитария: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония), с применением праймеров, имеющих последовательности оснований SEQ ID NOs: 15 и 16. С помощью электрофореза в 0,8% агарозе был выделен фрагмент ДНК приблизительно 1 т.н., амплифицированный с помощью ПЦР. Затем желаемую полосу вырезали и очищали. Фрагмент ДНК метили с помощью зонда дигоксинигена с применением DIG High Prime (производства Boehringer-Mannheim) на основе процедуры, описанной в его руководстве.
(4) Клонирование гена образующего пептид фермента из генной библиотеки
Для получения полноразмерного гена образующего пептид фермента была проведена гибридизация по Саузерну с применением в качестве зонда фрагмента ДНК, амплифицированного в указанной выше процедуре ПЦР. Операции гибридизации по Саузерну объяснены в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
После взаимодействия хромосомной ДНК Pedobacter heparinus IFO 12017 (институт-депозитарий: Institute of Fermentation, Osaka, адрес института-депозитария: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) с ферментом рестрикции HindIII при 37°С в течение 16 часов до полного гидролиза ДНК полученное подвергали электрофорезу в 0,8% агарозном геле. После электрофореза разделенную электрофорезом ДНК из агарозного геля переносили на положительно заряженный фильтр нейлоновой мембраны (производства Roche Diagnostics) с последующей обработкой, состоящей из щелочной денатурации, нейтрализации и иммобилизации. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 50°С в течение 1 часа добавляли зонд, меченный дигоксинигеном, полученный, как описано выше, и гибридизацию проводили при 50°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление полос, которые гибридизовались с зондом, производили с помощью DIG Nucleotide Detection Kit (Boehringer-Mannheim) на основе процедуры, описанной в руководстве к нему. В результате была успешно выявлена полоса приблизительно 5 т.н., которая гибридизовалась с зондом.
5 мкг хромосомной ДНК Pedobacter heparinus IFO 12017 полностью гидролизовали с помощью HindIII. ДНК приблизительно 5 т.н. отделяли с помощью электрофореза в 0,8% агарозном геле, ДНК очищали с помощью Gene Clean II Kit (производства Funakoshi) и растворяли в 10 мкл TE. 4 мкл полученного раствора и pUC118 HindIII/BAP смешивали и проводили реакцию лигирования с помощью DNA Ligation Kit Ver. 2 (производства Takara Shuzo). 5 мкл смеси данной жидкости реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM109 (производства Takara Shuzo) смешивали для трансформации Escherichia coli. Содержимое затем наносили на подходящую твердую среду для получения библиотеки хромосомной ДНК.
Для получения полноразмерного гена образующего пептид фермента библиотеку хромосомной ДНК подвергали скринингу с применением указанного выше зонда. Процедура гибридизации колоний объяснена в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии библиотеки хромосомной ДНК переносили на фильтр нейлоновой мембраны, Nylon Membrane for Colony and Plaque Hybridization (производства Roche Diagnostics), с последующей обработкой щелочной денатурацией, нейтрализацией и иммобилизацией. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 37°С в течение 1 часа добавляли указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 37°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление колоний, которые гибридизовались с меченым зондом, производили с помощью DIG Nucleotide Detection Kit (производства Boehringer-Mannheim) на основе объяснения, описанного в руководстве к нему. В результате был выявлен один штамм, чья колония гибридизовалась с меченым зондом.
(5) Последовательность оснований гена образующего пептид фермента, происходящего от Pedobacter heparinus IFO 12017
Плазмиды, содержащиеся в Escherichia coli JM109, получали из штамма, который, как было подтверждено, гибридизуется с меченым зондом, и была определена последовательность оснований части, где имела место гибридизация с зондом и поблизости от нее. Реакцию секвенирования проводили с применением CEQ DTCS-Quick Start Kit (производства Beckman-Coulter) на основе процедуры, описанной в руководстве к нему. Кроме того, проводили электрофорез с применением CEQ 2000-XL (производства Beckman-Coulter).
В результате было обнаружено, что рамка с открытым считыванием, которая кодирует образующий пептид фермент, действительно существует. Полноразмерная последовательность оснований гена образующего пептид фермента, происходящего от Pedobacter heparinus IFO 12017 (институт-депозитарий: Institute of Fermentation, Osaka, адрес института-депозитария: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) наряду с соответствующей аминокислотной последовательностью показана в SEQ ID NO: 17.
Пример 9
Экспрессия гена образующего пептид фермента, происходящего от Pedobacter heparinus IFO 12017, в Escherichia coli
Желаемый ген амплифицировали с помощью ПЦР с применением хромосомной ДНК Pedobacter heparinus IFO 12017 (институт-депозитарий: Institute of Fermentation, Osaka, адрес института-депозитария: 2-17-85 Jusanbon-cho, Yodogawa-ku, Osaka-shi, Япония) в качестве матрицы и олигонуклеотидов, показанных в SEQ ID NOs: 19 и 20, в качестве праймеров. Фрагмент ДНК обрабатывали NdeI/HindIII и полученный фрагмент ДНК и продукт обработки NdeI/HindIII pTrpT лигировали. Затем Escherichia coli JM109 трансформировали данным раствором лигирования, и из устойчивых к ампициллину штаммов были отобраны штаммы, содержащие желаемую плазмиду. Плазмида была обозначена как pTrpT_Ph_aet.
Одну петлю Escherichia coli JM109, содержащую pTrpT_Ph_aet, инокулировали в обычную пробирку для тестирования, содержащую 3 мл среды (2 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л казаминокислот, 5 г/л сульфата аммония, 3 г/л монокалийгидрофосфата, 1 г/л дикалийгидрофосфата, 0,5 г/л гептагидрата сульфата магния и 100 мг/л ампициллина), и основное культивирование проводили при 25°С в течение 20 часов. Было подтверждено, что культуральный бульон обладает продуцирующей α-AMP активностью на уровне 0,01 Е на мл культурального бульона, что подтвердило экспрессию клонированного гена в Escherichia coli. Более того, активность не была выявлена в трансформанте, содержащем лишь pTrpT, примененной в качестве контроля.
Пример 10
Выделение гена образующего пептид фермента, происходящего от Taxeobacter gelupurpurascens DSMZ 11116
Далее здесь описывается выделение гена образующего пептид фермента. Примененный микроорганизм представлял собой Taxeobacter gelupurpurascens DSMZ 11116 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур); адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия). Для выделения гена в качестве хозяина применяли Escherichia coli JM109, а в качестве вектора использовали pUC118.
(1) Получение клеток микроорганизма
Taxeobacter gelupurpurascens DSMZ 11116 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) культивировали в течение 24 часов при 25°С на агаровой среде CM2G (содержащей 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну петлю полученных клеток микроорганизма инокулировали в 500-мл сосуд Сакагуши, содержащий 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара) с последующим культивированием со встряхиванием при 25°С.
(2) Получение хромосомной ДНК из клеток микроорганизма
50 мл культурального бульона центрифугировали (12000 об/мин, 4°С, 15 минут) для сбора клеток микроорганизма. Затем из клеток микроорганизма получали хромосомную ДНК с применением Qiagen Genomic-Tip System (Qiagen) на основании процедуры, описанной ее производителем.
(3) Получение фрагмента ДНК зонда с помощью ПЦР
Фрагмент ДНК, содержащий часть гена образующего пептид фермента, берущего начало от Taxeobacter gelupurpurascens DSMZ 11116 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), получали с помощью способа ПЦР с применением LA-Taq (производства Takara Shuzo). Затем проводили реакцию ПЦР на хромосомной ДНК, полученной из Taxeobacter gelupurpurascens DSMZ 11116 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия), с применением праймеров, имеющих последовательности оснований SEQ ID NOs: 21 и 16. Фрагмент ДНК приблизительно 1 т.н., амплифицированный с помощью процедуры ПЦР, выделяли с помощью электрофореза в 0,8% агарозе. Желаемую полосу затем вырезали и очищали. Фрагмент ДНК метили с помощью зонда дигоксинигена с применением DIG High Prime (производства Boehringer-Mannheim) на основе процедуры, описанной в его руководстве.
(4) Клонирование гена образующего пептид фермента из библиотеки генов
Для получения полноразмерного гена образующего пептид фермента была проведена гибридизация по Саузерну с применением в качестве зонда фрагмента ДНК, амплифицированного в указанной выше процедуре ПЦР. Операции гибридизации по Саузерну объяснены в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
После взаимодействия хромосомной ДНК Taxeobacter gelupurpurascens DSMZ 11116 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) с ферментом рестрикции PstI при 37°С в течение 16 часов до полного гидролиза ДНК полученное подвергали электрофорезу в 0,8% агарозном геле. После электрофореза разделенную электрофорезом хромосомную ДНК из агарозного геля переносили на положительно заряженный фильтр нейлоновой мембраны (производства Roche Diagnostics) с последующей обработкой, состоящей из щелочной денатурации, нейтрализации и иммобилизации. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 50°С в течение 1 часа добавляли зонд, меченный дигоксинигеном, полученный, как описано выше, и гибридизацию проводили при 50°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление полос, которые гибридизовались с зондом, производили с помощью DIG Nucleotide Detection Kit (Boehringer-Mannheim) на основе процедуры, описанной в его руководстве. В результате была успешно выявлена полоса приблизительно 5 т.н., которая гибридизовалась с зондом.
5 мкг хромосомной ДНК Taxeobacter gelupurpurascens DSMZ 11116 (учреждение-депозитарий; the Deutche Sammlung von Mikroorganismen und Zellkulturen GmbH (Германская коллекция микроорганизмов и клеточных культур, адрес учреждения-депозитария; Mascheroder Weg 1b, 38124 Braunschweig, Германия) полностью гидролизовали с помощью PstI. ДНК приблизительно 5 т.н. отделяли с помощью электрофореза в 0,8% агарозном геле, ДНК очищали с помощью Gene Clean II Kit (производства Funakoshi) и растворяли в 10 мкл TE. 4 мкл полученного раствора смешивали с pUC118 PstI/BAP (производства Takara Shuzo) и проводили реакцию лигирования с помощью DNA Ligation Kit Ver. 2 (производства Takara Shuzo). 5 мкл данной жидкости реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM109 (производства Toyobo) смешивали для трансформации Escherichia coli. Содержимое затем наносили на подходящую твердую среду для получения библиотеки хромосомной ДНК.
Для получения полноразмерного гена образующего пептид фермента библиотеку хромосомной ДНК подвергали скринингу путем гибридизации колоний с применением указанного выше зонда. Процедура для гибридизации колоний объяснена в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии библиотеки хромосомной ДНК переносили на фильтр нейлоновой мембраны, Nylon Membrane for Colony and Plaque Hybridization (производства Roche Diagnostics) с последующей обработкой щелочной денатурацией, нейтрализацией и иммобилизацией. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 37°С в течение 1 часа добавляли указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 37°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление колоний, которые гибридизовались с меченым зондом, производили с помощью DIG Nucleotide Detection Kit (производства Boehringer-Mannheim) на основе объяснения, описанного в его руководстве. В результате был выявлен один штамм, колония которого гибридизовалась с меченым зондом.
(5) Последовательность оснований гена образующего пептид фермента, происходящего от Taxeobacter gelupurpurascens DSMZ 11116
Плазмиды, содержащиеся в Escherichia coli JM109, получали из штамма, который, как было подтверждено, гибридизуется с меченым зондом, и была определена последовательность оснований части, где имела место гибридизация с зондом и поблизости от нее. Реакцию секвенирования проводили с применением CEQ DTCS-Quick Start Kit (производства Beckman-Coulter) на основе процедуры, описанной в руководстве к нему. Кроме того, проводили электрофорез с применением CEQ 2000-XL (производства Beckman-Coulter).
В результате было обнаружено, что рамка с открытым считыванием, которая кодирует образующий пептид фермент, действительно существует. Полноразмерная последовательность оснований гена образующего пептид фермента из Taxeobacter gelupurpurascens DSMZ 11116, наряду с соответствующей аминокислотной последовательностью, показана в SEQ ID NO: 22.
Пример 11
Выделение гена образующего пептид фермента, происходящего от Cyclobacterium marinum ATCC 25205
Далее здесь описывается выделение гена образующего пептид фермента. Примененный микроорганизм представлял собой Cyclobacterium marinum ATCC 25205 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки). Для выделения гена в качестве хозяина применяли Escherichia coli JM109, а в качестве вектора использовали pUC118.
(1) Получение клеток микроорганизма
Cyclobacterium marinum ATCC 25205 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) культивировали в течение 24 часов при 25°С на агаровой среде CM2G (содержащей 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну петлю полученных клеток микроорганизма инокулировали в 500-мл сосуд Сакагуши, содержащий 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара), с последующим культивированием со встряхиванием при 25°С.
(2) Получение хромосомной ДНК из клеток микроорганизма
50 мл культурального бульона центрифугировали (12000 об/мин, 4°С, 15 минут) для сбора клеток микроорганизма. Затем из клеток микроорганизма получали хромосомную ДНК с применением Qiagen Genomic-Tip System (Qiagen) на основании процедуры, описанной в руководстве к ней.
(3) Получение фрагмента ДНК зонда с помощью ПЦР
Фрагмент ДНК, содержащий часть гена образующего пептид фермента, берущего начало от Cyclobacterium marinum ATCC 25205, получали с помощью способа ПЦР с применением LA-Taq (производства Takara Shuzo). Затем проводили реакцию ПЦР на хромосомной ДНК, полученной из Cyclobacterium marinum ATCC 25205 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), с применением праймеров, имеющих последовательности оснований SEQ ID NOs: 15 и 16. Фрагмент ДНК приблизительно 1 т.н., амплифицированный с помощью процедуры ПЦР, выделяли с помощью электрофореза в 0,8% агарозе. Желаемую полосу затем вырезали и очищали. Фрагмент ДНК метили с помощью зонда дигоксинигена с применением DIG High Prime (производства Boehringer-Mannheim) на основе процедуры, описанной в его руководстве.
(4) Клонирование гена образующего пептид фермента из библиотеки генов
Для получения полноразмерного гена образующего пептид фермента была проведена гибридизация по Саузерну с применением в качестве зонда фрагмента ДНК, амплифицированного в указанной выше процедуре ПЦР. Операции гибридизации по Саузерну объяснены в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
После взаимодействия хромосомной ДНК Cyclobacterium marinum ATCC 25205 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) с ферментом рестрикции PstI или HincII при 37°С в течение 16 часов до полного гидролиза ДНК полученное подвергали электрофорезу в 0,8% агарозном геле. После электрофореза разделенную электрофорезом хромосомную ДНК из агарозного геля переносили на положительно заряженный фильтр нейлоновой мембраны (производства Roche Diagnostics) с последующей обработкой, состоящей из щелочной денатурации, нейтрализации и иммобилизации. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 50°С в течение 1 часа добавляли зонд, меченный дигоксинигеном, полученный, как описано выше, и гибридизацию проводили при 50°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление полос, которые гибридизовались с зондом, производили с помощью DIG Nucleotide Detection Kit (Boehringer-Mannheim) на основе процедуры, описанной в его руководстве. В результате была успешно выявлена полоса 7 т.н., которая гибридизовалась с зондом, для гидролизованного PstI продукта и успешно выявлена полоса 2 т.н., которая гибридизовалась с зондом, для гидролизованного HincII продукта.
5 мкг хромосомной ДНК Cyclobacterium marinum ATCC 25205 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) полностью гидролизовали с помощью PstI или HincII. ДНК приблизительно 7 т.н. или 2 т.н. отделяли с помощью электрофореза в 0,8% агарозном геле. ДНК очищали с помощью Gene Clean II Kit (производства Funakoshi) и растворяли в 10 мкл TE. 4 мкл полученного раствора смешивали с pUC118 PstI/BAP (производства Takara Shuzo) или pUC118 HincII/BAP (производства Takara Shuzo) и проводили реакцию лигирования с помощью DNA Ligation Kit Ver. 2 (производства Takara Shuzo). 5 мкл данной жидкости реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM109 (производства Takara Shuzo) смешивали для трансформации Escherichia coli. Содержимое затем наносили на подходящую твердую среду для получения библиотеки хромосомной ДНК.
Для получения полноразмерного гена образующего пептид фермента библиотеку хромосомной ДНК подвергали скринингу путем гибридизации колоний с применением указанного выше зонда. Процедура для гибридизации колоний объяснена в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии библиотеки хромосомной ДНК переносили на фильтр нейлоновой мембраны, Nylon Membrane for Colony and Plaque Hybridization (производства Roche Diagnostics) с последующей обработкой щелочной денатурацией, нейтрализацией и иммобилизацией. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 37°С в течение 1 часа добавляли указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 37°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление колоний, которые гибридизовались с меченым зондом, производили с помощью DIG Nucleotide Detection Kit (производства Boehringer-Mannheim) на основе объяснения, описанного в его руководстве. В результате был выявлен один штамм, колония которого гибридизовалась с меченым зондом.
(5) Последовательность оснований гена образующего пептид фермента, происходящего от Cyclobacterium marinum ATCC 25205
Плазмиды, содержащиеся в Escherichia coli JM109, получали из каждого штамма, который, как было подтверждено, гибридизуется с меченым зондом, и была определена последовательность оснований части, где имела место гибридизация с зондом и поблизости от нее. Реакцию секвенирования проводили с применением CEQ DTCS-Quick Start Kit (производства Beckman-Coulter) на основе процедуры, описанной в руководстве к нему. Кроме того, проводили электрофорез с применением CEQ 2000-XL (производства Beckman-Coulter).
В результате было обнаружено, что рамка с открытым считыванием, которая кодирует образующий пептид фермент, действительно существует. Полноразмерная последовательность оснований гена образующего пептид фермента, происходящего от Cyclobacterium marinum ATCC 25205 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), наряду с соответствующей аминокислотной последовательностью, показана в SEQ ID NO: 24.
Пример 12
Выделение гена образующего пептид фермента, происходящего от Psychroserpens burtonensis ATCC 700359
Далее здесь описывается выделение гена образующего пептид фермента. Примененный микроорганизм представлял собой Psychroserpens burtonensis ATCC 700359 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки). Для выделения гена в качестве хозяина применяли Escherichia coli JM109, а в качестве вектора использовали pUC118.
(1) Получение клеток микроорганизма
Psychroserpens burtonensis ATCC 700359 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки) культивировали в течение 24 часов при 25°С на агаровой среде CM2G (содержащей 50 г/л глюкозы, 10 г/л дрожжевого экстракта, 10 г/л пептона, 5 г/л хлорида натрия и 20 г/л агара, pH 7,0). Одну петлю полученных клеток микроорганизма инокулировали в 500-мл сосуд Сакагуши, содержащий 50 мл жидкой среды CM2G (указанная выше среда, за исключением агара), с последующим культивированием со встряхиванием при 10°С.
(2) Получение хромосомной ДНК из клеток микроорганизма
50 мл культурального бульона центрифугировали (12000 об/мин, 4°С, 15 минут) для сбора клеток микроорганизма. Затем из клеток микроорганизма получали хромосомную ДНК с применением Qiagen Genomic-Tip System (Qiagen) на основании процедуры, описанной в руководстве к ней.
(3) Получение фрагмента ДНК зонда с помощью ПЦР
Фрагмент ДНК, содержащий часть гена образующего пептид фермента, берущего начало от Psychroserpens burtonensis ATCC 700359 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), получали с помощью способа ПЦР с применением LA-Taq (производства Takara Shuzo). Затем проводили реакцию ПЦР на хромосомной ДНК, полученной из Psychroserpens burtonensis ATCC 700359 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), с применением праймеров, имеющих последовательности оснований SEQ ID NOs: 15 и 16. Фрагмент ДНК приблизительно 1 т.н., амплифицированный с помощью процедуры ПЦР, выделяли с помощью электрофореза в 0,8% агарозе. Желаемую полосу затем вырезали и очищали. Фрагмент ДНК метили с помощью зонда дигоксинигена с применением DIG High Prime (производства Boehringer-Mannheim) на основе процедуры, описанной в его руководстве.
(4) Клонирование гена образующего пептид фермента из библиотеки генов
Для получения полноразмерного гена образующего пептид фермента была проведена гибридизация по Саузерну с применением в качестве зонда фрагмента ДНК, амплифицированного в указанной выше процедуре ПЦР. Операции гибридизации по Саузерну объяснены в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
После взаимодействия хромосомной ДНК Psychroserpens burtonensis ATCC 700359 с ферментом рестрикции EcoRI при 37°С в течение 16 часов до полного гидролиза ДНК полученное подвергали электрофорезу в 0,8% агарозном геле. После электрофореза разделенную электрофорезом хромосомную ДНК из агарозного геля переносили на положительно заряженный фильтр нейлоновой мембраны (производства Roche Diagnostics) с последующей обработкой, состоящей из щелочной денатурации, нейтрализации и иммобилизации. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 50°С в течение 1 часа добавляли зонд, меченный дигоксинигеном, полученный, как описано выше, и гибридизацию проводили при 50°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление полос, которые гибридизовались с зондом, производили с помощью DIG Nucleotide Detection Kit (Boehringer-Mannheim) на основе процедуры, описанной в его руководстве. В результате была успешно выявлена полоса приблизительно 7 т.н., которая гибридизовалась с зондом.
5 мкг хромосомной ДНК Psychroserpens burtonensis ATCC 700359 полностью гидролизовали с помощью EcoRI. ДНК приблизительно 7 т.н. отделяли с помощью электрофореза в 0,8% агарозном геле, ДНК очищали с помощью Gene Clean II Kit (производства Funakoshi) и растворяли в 10 мкл TE. 4 мкл полученного раствора смешивали с pUC118 EcoRI/BAP (производства Takara Shuzo) и проводили реакцию лигирования с помощью DNA Ligation Kit Ver. 2 (производства Takara Shuzo). 5 мкл данной жидкости реакции лигирования и 100 мкл компетентных клеток Escherichia coli JM109 (производства Takara Shuzo) смешивали для трансформации Escherichia coli. Содержимое затем наносили на подходящую твердую среду для получения библиотеки хромосомной ДНК.
Для получения полноразмерного гена образующего пептид фермента библиотеку хромосомной ДНК подвергали скринингу путем гибридизации колоний с применением указанного выше зонда. Процедура для гибридизации колоний объяснена в Molecular Cloning, 2nd edition, Cold Spring Harbor Press (1989).
Колонии библиотеки хромосомной ДНК переносили на фильтр нейлоновой мембраны, Nylon Membrane for Colony and Plaque Hybridization (производства Roche Diagnostics) с последующей обработкой щелочной денатурацией, нейтрализацией и иммобилизацией. Гибридизацию проводили с применением EASY HYB (производства Boehringer-Mannheim). После предварительной гибридизации фильтра при 37°С в течение 1 часа добавляли указанный выше зонд, меченный дигоксинигеном, с последующей гибридизацией при 37°С в течение 16 часов. Затем фильтр промывали дважды при 60°С 1 x SSC, содержащим 0,1% SDS.
Выявление колоний, которые гибридизовались с меченым зондом, производили с помощью DIG Nucleotide Detection Kit (производства Boehringer-Mannheim) на основе объяснения, описанного в его руководстве. В результате был выявлен один штамм, колония которого гибридизовалась с меченым зондом.
(5) Последовательность оснований гена образующего пептид фермента, происходящего от Psychroserpens burtonensis ATCC 700359
Плазмиды, содержащиеся в Escherichia coli JM109, получали из штамма, который, как было подтверждено, гибридизуется с меченым зондом, и была определена последовательность оснований части, где имела место гибридизация с зондом и поблизости от нее. Реакцию секвенирования проводили с применением CEQ DTCS-Quick Start Kit (производства Beckman-Coulter) на основе процедуры, описанной в руководстве к нему. Кроме того, проводили электрофорез с применением CEQ 2000-XL (производства Beckman-Coulter).
В результате было обнаружено, что рамка с открытым считыванием, которая кодирует образующий пептид фермент, действительно существует. Полноразмерная последовательность оснований гена образующего пептид фермента, происходящего от Psychroserpens burtonensis ATCC 700359 (учреждение-депозитарий; американская коллекция типов культур, адрес учреждения-депозитария; P.O. Box 1549, Manassas, VA 20110, Соединенные Штаты Америки), наряду с соответствующей аминокислотной последовательностью, показана в SEQ ID NO: 26.
Хотя изобретение было описано в отношении конкретного осуществления для полного и ясного раскрытия, прилагаемая формула изобретения не ограничена этим, и ее следует трактовать как включающую все модификации и альтернативные конструкции, которые могут быть осуществлены специалистом в данной области техники и которые четко подпадают в изложенную здесь базовую доктрину.
Перечень последовательностей в свободном доступе
SEQ ID NO: 3: Синтетический праймер 1
SEQ ID NO: 4: Синтетический праймер 2
SEQ ID NO: 5: Ген, кодирующий образующий пептид фермент
SEQ ID NO: 7: Синтетический праймер для получения pTrpT
SEQ ID NO: 8: Синтетический праймер для получения pTrpT
SEQ ID NO: 9: Синтетический праймер для получения pTrpT_Gtg2
SEQ ID NO: 10: Синтетический праймер для получения pTrpT_Gtg2
SEQ ID NO: 11: Ген, кодирующий образующий пептид фермент
SEQ ID NO: 13: Синтетический праймер для получения pTrpT_Sm_aet
SEQ ID NO: 14: Синтетический праймер для получения pTrpT_Sm_aet
SEQ ID NO: 15: Смешанный праймер 1 для Aet
SEQ ID NO: 16: Смешанный праймер 2 для Aet
SEQ ID NO: 19: Праймер 1 для конструирования экспрессионных векторов, берущих начало от pPedobacter
SEQ ID NO: 20: Праймер 2 для конструирования экспрессионных векторов, берущих начало от pedobacterPedobacter
SEQ ID NO: 21: Смешанный праймер 3 для Aet
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКОЙ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ydiB, СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА НИЗШИХ АЛКИЛОВ АЛЬФА-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИНА | 2007 |
|
RU2355759C1 |
| СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА (ВАРИАНТЫ) | 2005 |
|
RU2316596C2 |
| ГЕН ПЕПТИДООБРАЗУЮЩЕГО ФЕРМЕНТА, ПЕПТИДООБРАЗУЮЩИЙ ФЕРМЕНТ И СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА | 2002 |
|
RU2280077C2 |
| НОВЫЙ ГЕН ПЕПТИДОБРАЗУЮЩЕГО ФЕРМЕНТА | 2003 |
|
RU2296798C2 |
| МУТАНТНЫЙ БЕЛОК, КОДИРУЕМЫЙ ГЕНОМ yddG, И СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКИХ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ РОДА Escherichia | 2012 |
|
RU2530171C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2008 |
|
RU2515044C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-треонина С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2351646C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ РОДА Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ОПЕРОН astCADBE | 2010 |
|
RU2482188C2 |
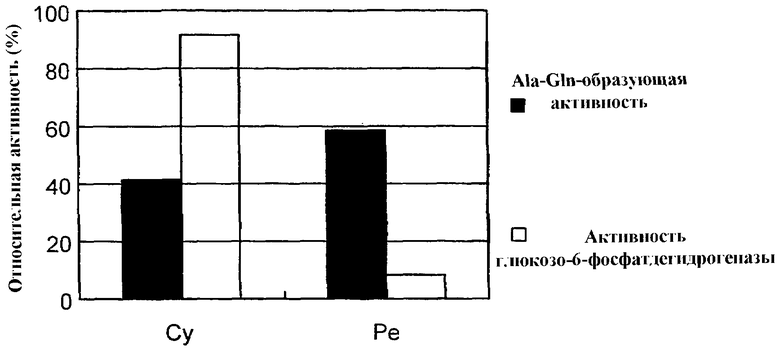
Изобретение относится к области биотехнологии и может быть использовано в пищевой промышленности. Предложен способ ферментативного получения α-L-аспартил-L-фенилаланин-β-эфира (α-L-(β-о-замещенный аспартил)-L-фенилаланин) из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина при 5-40°С и рН 7-10 в течение времени, достаточного для образования α-L-аспартил-L-фенилаланин-β-эфира из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина. Указанный способ используют в способе получения α-L-аспартил-L-фенилаланин-α-метилового эфира, включающего стадию синтеза α-L-аспартил-L-фенилаланин-β-метилового эфира и стадию превращения указанного эфира в α-L-аспартил-L-фенилаланин-α-метиловый эфир. Применение изобретения обеспечивает получение α-L-аспартил-L-фенилаланин-β-эфира с последующим получением α-L-аспартил-L-фенилаланин-α-метилового эфира по упрощенной и дешевой схеме с повышенным выходом. 2 н. и 3 з.п. ф-лы, 1 ил., 4 табл.
а) последовательности, образованной остатками с 23 по 616 аминокислотной последовательности, представленной в SEQ ID NO: 6;
б) последовательности, образованной остатками с 21 по 619 аминокислотной последовательности, представленной в SEQ ID NO: 12;
в) последовательности, образованной остатками с 23 по 625 аминокислотной последовательности, представленной в SEQ ID NO: 18;
г) последовательности, образованной остатками с 23 по 645 аминокислотной последовательности, представленной в SEQ ID NO: 23;
д) последовательности, образованной остатками с 26 по 620 аминокислотной последовательности, представленной в SEQ ID NO: 25;
е) последовательности, образованной остатками с 18 по 644 аминокислотной последовательности, представленной в SEQ ID NO: 27;
ж) последовательности, представленной в SEQ ID NO: 6;
з) последовательности, представленной в SEQ ID NO: 12;
и) последовательности, представленной в SEQ ID NO: 18;
к) последовательности, представленной в SEQ ID NO: 23;
л) последовательности, представленной в SEQ ID NO: 25;
м) последовательности, представленной в SEQ ID NO: 27;
н) последовательностей с (а) по (м), включающих такую замену, делецию, инсерцию, добавление и/или инверсию одной или множества аминокислот, при которой фермент способен избирательно присоединять L-фенилаланин к α-эфирному участку α,β-диэфира L-аспарагиновой кислоты через пептидную связь,
либо средства, содержащего указанный фермент, где контактирование проводят при 5-40°С и рН 7-10 в течение времени, достаточного для образования α-L-аспартил-L-фенилаланин-β-эфира из α,β-диэфира L-аспарагиновой кислоты и L-фенилаланина.
Aeromonas, Azotobacter, Alcaligenes, Brevibacterium, Corynebacterium, Escherichia, Empedobacter, Flavobacterium, Microbacterium, Propionibacterium, Brevibacillus, Paenibacillus, Pseudomonas, Serratia, Stenotrophomonas, Sphingobacterium, Streptomyces, Xanthomonas, Williopsis, Candida, Geotrichum, Pichia, Saccharomyces, Tomlaspora, Cellulophaga, Weeksella, Pedobacter, Persicobacter, Flexithrix, Chitinophaga, Cyclobacterium, Runella, Thermonema, Psychroserpens, Gelidibacter, Dyadobacter, Flammeovirga, Spirosoma, Flectobacillus, Tenacibaculum, Rhodotermus, Zobellia, Muricauda, Salegentibacter, Taxeobacter, Cytophaga, Marinilabilia, Lewinella, Saprospira, и Haliscomenobacter.
Приоритет по пп.1-5 в части (а), (б), (ж), (з), (н) в части SEQ ID NO: 6 и 12, а также по п.З в части Aeromonas, Azotobacter, Alcaligenes, Brevibacterium, Corynebacterium, Escherichia, Empedobacter, Flavobacterium, Microbacterium, Propionibacterium, Brevibacillus, Paenibacillus, Pseudomonas, Serratia, Stenotrophomonas, Sphingobacterium, Streptomyces, Xanthomonas, Williopsis, Candida, Geotrichum, Pichia, Saccharomyces, Tomlaspora установлен от 24.01.2003 (по дате подачи заявки JP 2003-016764). Приоритет по пп.1-5 в части (в), (г), (д), (е), (и), (к), (л), (м), (н) в части SEQ ID NO: 18, 23, 25 и 27, а также по п.3 в части Cellulophaga, Weeksella, Pedobacter, Persicobacter, Flexithrix, Chitinophaga, Cyclobacterium, Runella, Thermonema, Psychroserpens, Gelidibacter, Dyadobacter, Flammeovirga, Spirosoma, Flectobacillus, Tenacibaculum, Rhodotermus, Zobellia, Muricauda, Salegentibacter, Taxeobacter, Cytophaga, Marinilabilia, Lewinella, Saprospira, и Haliscomenobacter установлен от 25.07.2003 (по дате подачи заявки JP 2003-201819).
| RU 99110745 A, 27.03.2002 | |||
| Многоярусная сушилка шкафного типа | 1938 |
|
SU58478A1 |
| СПОСОБ ПОЛУЧЕНИЯ ПРЕПАРАТА ДЛЯ ЖИРОВАНИЯ КОЖ | 0 |
|
SU269390A1 |
| JP 4311389, 04.11.1992. | |||

