Область изобретения
Данное изобретение относится к микробиологическому ферментативному производству, точнее к способу придания способности утилизировать метанол микроорганизму, который исходно не обладал такой способностью, или усиления такой способности в случае микроорганизма, обладающего такой способностью на низком уровне, и к способу производства целевого вещества утилизацией метанола с применением микроорганизма, полученного таким способом, который указан выше.
Вещества, полученные согласно данному изобретению, включают L-аминокислоты, нуклеиновые кислоты, антибиотики, витамины, факторы роста, физиологически активные вещества и так далее, которые традиционно получали с использованием микроорганизмов.
Описание связанной области
До настоящего времени большую часть сырья для ферментации, используемого в производстве полезных веществ посредством микробиологической ферментации, составляют сахара, полученные из сельскохозяйственных продуктов. Однако, так как считается, что цены на сахара, полученные из сельскохозяйственных продуктов, в будущем будут расти, требуется недорогой материал хорошего качества как альтернативное сырье для ферментации.
Метанол легко растворим в воде и недорог, и его можно получить с высокой степенью очистки. Более того, его можно сравнительно легко получить из метана, который является основным компонентом природного газа. Поэтому метанол привлекателен в качестве сырья для производства веществ. Если в качестве сырья для микробиологической ферментации используют метанол, можно не только снизить стоимость основного сырья, но также можно упростить очистку продуктов из растворов для ферментации и способ удаления отработанного раствора. Таким образом, можно снизить общую стоимость продукции.
В качестве способов производства веществ, особенно аминокислот, из метанола, используемого в качестве сырья, с применением микроорганизмов, уже был известен способ с применением микроорганизма рода Achromobacter или Pseudomonas (заявка на патент Японии (Kokoku) No. 45-25273), способ с применением микроорганизма рода Protaminobacter или Methanomonas (выложенная заявка на выдачу патента Японии (Kokai) No. 50-25790), способ с применением микроорганизма рода Methylobacillus (выложенная заявка на выдачу патента Японии No. 4-91793), способ с применением метилотрофной бактерии, относящейся к роду Bacillus (выложенная заявка на выдачу патента Японии No. 3-505284, патент США, No. 6083728), и так далее. Однако все бактериальные штаммы не обладали высокой продуктивностью в отношении аминокислот, приемлемой для практического применения бактерий.
Между тем способы с применением микроорганизмов рода Brevibacterium, Corynebacterium, Bacillus или Escherichia составляют широко распространенные способы производства аминокислот из глюкозы ("Amino Acid Fermentation", Ed. By H. Aida et al., Gakkai Shuppan Center). Указанные бактерии, продуцирующие аминокислоты, являются ценными бактериальными штаммами, выведенными путем введения различных мутаций для того, чтобы получить максимальную продуктивность в отношении аминокислот, в течение длительного исторического периода от их открытия до выведения практически пригодных штаммов. Однако указанные промышленно используемые штаммы являются микроорганизмами, которые не могут утилизировать метанол.
Сущность изобретения
Целью данного изобретения является предоставление нового микроорганизма, обладающего способностью продуцировать продукт ферментации, такой как аминокислота, из метанола как сырья для ферментации, путем придания способности утилизировать метанол микроорганизму, который исходно может утилизировать сахар, но не может утилизировать метанол, или путем усиления такой способности в случае микроорганизма, обладающего способностью на низком уровне, и способа производства целевого вещества из метанола с использованием такого микроорганизма.
Авторы данного изобретения проводили тщательные исследования для того, чтобы достичь вышеуказанных целей. В результате, авторы обнаружили, что введением в микроорганизм гена метанолдегидрогеназы и усиления активностей гексулозофосфатсинтазы и фосфогексулоизомеразы микроорганизма можно придать микроорганизму способность утилизировать метанол, или можно усилить способность микроорганизма, и таким образом выполнили данное изобретение.
Конкретно, данное изобретение относится к следующему.
(1) Способ получения целевого вещества посредством использования микроорганизма, включающий в себя культивирование микроорганизма, обладающего способностью продуцировать целевое вещество, в среде, чтобы продуцировать и накопить целевое вещество в среде или клетках микроорганизма, и сбор целевого вещества из среды или клеток микроорганизма, при этом микроорганизм является микроорганизмом, в который введен ген метанолдегидрогеназы и который модифицирован для того, чтобы были повышены активности гексулозофосфатсинтазы и фосфогексулоизомеразы и была придана или усилена способность утилизировать метанол, и среда содержит метанол в качестве источника углерода.
(2) Способ по п.(1), где целевым веществом является L-аминокислота.
(3) Способ по п.(2), где L-аминокислотой является L-лизин.
(4) Способ по п.(3), где микроорганизмом является бактерия, относящаяся к роду Bacillus.
(5) Микроорганизм, в который введен ген метанолдегидрогеназы, причем микроорганизм модифицирован для того, чтобы были усилены активности гексулозофосфатсинтазы и фосфогексулоизомеразы, и была придана или усилена способность утилизировать метанол.
(6) Микроорганизм по п.(5), который является грам-положительной бактерией.
(7) Микроорганизм по п.(6), который является бактерией, относящейся к роду Bacillus.
(8) Микроорганизм по п.(7), который является Bacillus subtilis.
Согласно данному изобретению способность утилизировать метанол можно придать микроорганизму, исходно не обладающему такой способностью, или такую способность для микроорганизма, обладающего этой способностью на низком уровне, можно усилить, и таким образом можно получить микроорганизм, который может утилизировать дешевый метанол в качестве источника углерода или источника энергии, используемого микроорганизмом. Кроме того, используя полученный микроорганизм, в среде с добавлением метанола из метанола можно получить различные продукты ферментации.
На чертеже показана структура плазмиды pDLT3 для введения гена в хромосому. Символ "cat" означает ген устойчивости к хлорамфениколу, "bla" означает ген устойчивости к ампициллину и "pBR ori" означает локус начала репликации pBR322.
Предпочтительные варианты изобретения
Далее будет дано подробное объяснение данного изобретения.
Микроорганизм согласно данному изобретению представляет собой микроорганизм, в который введен ген метанолдегидрогеназы и который модифицирован для того, чтобы были повышены активности гексулозофосфатсинтазы и фосфогексулоизомеразы и была придана или усилена способность утилизировать метанол. Хотя родительский штамм, используемый для конструирования микроорганизма согласно данному изобретению, может быть микроорганизмом, исходно обладающим слабой способностью утилизировать метанол, предпочтительно он является микроорганизмом, который может утилизировать сахар, но не может утилизировать метанол.
Микроорганизм, который может утилизировать метанол, имеет метанолоксидазу (например, метанолдегидрогеназу) и он диссимилирует и ассимилирует формальдегид, продуцируемый при окислении метанола посредством точной метаболической регуляции. Это происходит потому, что формальдегид является сильно токсичным для организма и поэтому клетки должны его быстро утилизировать в качестве источника углерода или источника энергии или удалять его посредством детоксикации. Поэтому, если имеет место предполагаемое придание способности утилизировать метанол микроорганизму, который не может утилизировать метанол, конечно обязательным является введение метанолоксидазы. Однако нет достаточно специфичных критериев надлежащего удаления формальдегида, продуцируемого в результате экспрессии метанолоксидазной активности, и поэтому считалось, что невозможно придать способность утилизировать метанол произвольно выбранному микроорганизму.
Однако авторы данного изобретения обнаружили, что способность утилизировать метанол можно придать даже микроорганизму, который исходно не мог утилизировать метанол, если получить фермент, обладающий способностью к окислению метанола, в клетках микроорганизма, и, кроме того, усилить гексулозофосфатсинтазную активность и фосфогексулоизомеразную активность микроорганизма.
Микроорганизм, используемый для данного изобретения, особым образом не ограничен при условии, что можно придать вышеуказанные свойства, характерные для микроорганизма согласно данному изобретению, и конкретно можно указать бактерии, относящиеся к роду Bacillus, такие как Bacillus subtilis, бактерии Escherichia, такие как Escherichia coli, коринеформные бактерии, такие как Brevibacterium lactofermentum. (Corynebacterium glutamicum), бактерии Serratia, такие как Serratia marcescens, и так далее. Из них предпочтительны грам-положительные бактерии, особенно бактерии, относящиеся к роду Bacillus.
Конкретно можно указать Escherichia coli VKRM B-3996 (RIA 1867, смотри патент США No. 5175107), Corynebacterium acetoacidophilum AJ12318 (FERM BP-1172, смотри патент США No. 5188949) и т.д. для L-треонина как продукта ферментации; Bacillus subtilis AJ11779 (FERM P-18453), Bacillus subtilis AJ13291 (FERM P-6722, смотри выложенную заявку на выдачу патента Японии No. 59-63193), Escherichia coli AJ11442 (NRRL B-12185, FERM BP-1543, смотри патент США No.4346170), Escherichia coli W3110 (tyrA) (указанный штамм можно получить посредством удаления плазмиды pHATerm из Escherichia coli W3110 (tyrA)/pHATerm (FERM BP-3653), смотри международную заявку на патент W095/16042), Brevibacterium lactofermentum AJ12435 (FERM BP-2294, патент США No. 5304476), Brevibacterium lactofermentum AJ3990 (ATCC 31269, смотри патент США No. 4066501) и т.д. для L-лизина как продукта ферментации; Escherichia coli AJ12624 (FERM BP-3853, смотри выложенную заявку на выдачу патента Франции No. 2680178), Brevibacterium lactofermentum AJ12821 (FERM BP-4172, выложенная заявка на выдачу патента Японии No. 5-26811, выложенная заявка на выдачу патента Франции No. 2701489), Brevibacterium lactofermentum AJ12475 (FERM BP-2922, смотри патент США No. 5272067), Brevibacterium lactofermentum AJ13029 (FERM BP-5189, смотри международную заявку на патент JP95/01586), и т.д. для L-глутаминовой кислоты как продукта ферментации; Escherichia coli AJ11478 (FERM Р-5274, смотри заявку на выдачу патента Японии No. 62-34397), Brevibacterium lactofermentum AJ3718 (FERM P-2516, смотри патент США No. 3970519), и т.д. для L-лейцина как продукта ферментации; Escherichia coli KX141 (VKPM В-4781, смотри выложенную Европейскую заявку на патент No. 519113), Brevibacterium flavum AJ12149 (FERM BP-759, смотри патент США No. 4656135), и т.д. для L-изолейцина как продукта ферментации; Escherichia coli VL1970 (VKPM B-4411, смотри выложенную Европейскую заявку на патент No. 519113), Brevibacterium lactofermentum AJ12341 (FERM BP-1763, смотри патент США No. 5188948), и т.д. для L-валина как продукта ферментации; Bacillus subtilis AJ12000 (FERM P-6895, смотри выложенную заявку на выдачу патента Японии No. 59-143596), Escherichia coli AJ12604 (FERM BP-3579, смотри выложенную заявку на выдачу патента Японии No. 5-236947, выложенную Европейскую заявку на выдачу патента No. 488424), Brevibacterium lactofermentum AJ12637 (FERM BP-4160, смотри выложенную заявку на выдачу патента Франции No. 2686898), и т.д. для L-фенилаланина как продукта ферментации; Bacillus subtilis AJ11488 (FERM P-5290, смотри патент Японии No. 1426798), Escherichia coli KB862 (DSM7196, смотри Европейский патент No. 662143), и т.д. для L-триптофана как продукта ферментации и т.д.
В результате проведенных детальных исследований авторы данного изобретения пришли к выводу, что основным условием для придания способности утилизировать метанол является получение достаточного проявления метанолдегидрогеназной активности в клетках и усиление функции ассимиляции формальдегида, получаемого при ферментативной реакции. Кроме того, авторы данного изобретения пришли к выводу, что усиление ферментативных активностей гексулозофосфатсинтазы (HPS) и фосфогексулоизомеразы (PHI), которые являются ключевыми ферментами рибулозомонофосфатного пути, было бы полезно для эффективной ассимиляции формальдегида. Таким образом, авторы данного изобретения обнаружили, что можно придать способность утилизировать метанол микроорганизму, который исходно не может утилизировать метанол, путем введения в микроорганизм гена метанолдегидрогеназы и усиления активностей HPS и PHI.
Метанолдегидрогеназа (MDH), используемая в данном изобретении, представляет собой фермент, обладающий ферментативной активностью, за счет которой может быть окислен метанол, с превращением его в формальдегид. Например, MDH зависимого от PQQ (пирролохинолинхинон) типа, которая главным образом наблюдается в грам-отрицательных микроорганизмах, может быть указана в качестве MDH, которую можно использовать для данного изобретения. В частности, можно указать MDH штамма AMI Methylobacterium extorquens (Biochem. Biophys. Acta, 1119:97-106 (1992)) и т.д. Кроме того, можно указать MDH NAD-зависимого (никотинамидадениндинуклеотид) типа, наблюдаемого в грам-положительных микроорганизмах, в частности MDH Bacillus methanoliocus (J. Bacteriol., 174: 5346-5353 (1992)), алькогольдегидрогеназу (ADH), полученную из Bacillus stearothermophilus штамма DSM 2334 (Biochem. J., 252: 661-666) и т.д. Кроме того, также можно указать ADH печени быка (Biochem. J., 100: 34-46 (1966)) и печени человека (Arch. Toxicol., 72: 604-607 (1998)). Кроме того, также можно заново создать и использовать алкогольдегидрогеназу мутантного типа, которая в значительной степени также действует на метанол, путем введения мутации в ген алкогольдегидрогеназы, которая исходно не действует на метанол, для того, чтобы модифицировать ее субстратную специфичность. Однако в качестве MDH, которую можно соответствующим образом использовать для данного изобретения, особо можно указать, MDH, полученную, например, из Bacillus brevis NCIMB No. 12524, которая является ассимилирующей метанол бактерией, относящейся к роду Bacillus.
Ген, кодирующий MDH (mdh), можно получить из микроорганизма, который продуцирует MDH, таким же способом, который используют в обычном методе клонирования генов. Например, ген MDH можно получить посредством ПЦР (полимеразной цепной реакции) с использованием хромосомной ДНК Bacillus brevis штамма SI (NCIMB 12524) в качестве матрицы и олигонуклеотидов, имеющих нуклеотидные последовательности, показанные как SEQ ID NO:1 и 2, в качестве праймеров. Способы получения библиотеки геномной ДНК, используемой для клонирования генов, гибридизации, ПЦР, получения плазмидной ДНК, расщепления и лигирования ДНК, трансформации и т.д. описаны в Sambrook, J., Fritsch, E.F., Maniatis, Т., Molecular Cloning, Cold Spring Harbor Laboratory Press, 1.21 (1989). К тому же функционирует ли ген MDH в микроорганизме, в который введен ген, можно подтвердить измерением активности MDH лизата микроорганизма. Активность MDH можно, например, измерить способом измерения восстановления NAD+ (никотинамидадениндинуклеотид), сопровождающего окисление метанола в формальдегид, посредством измерения поглощения света с длиной волны 340 нм.
В качестве конкретного примера гена mdh, используемого для данного изобретения, можно указать ген mdh штамма SI Bacillus brevis. Ген mdh Bacillus methanolicus штамма C1 (NCIMB 13114, Eur. J. Biochem., 244: 426-433 (1997)) зарегистрирован в GenBank под инвентарным номером М65004 (имя для ввода BACMDH).
К тому же сообщалось о существовании факторов, активирующих активность метанолдегидрогеназы (Amd: активатор метанолдегидрогеназы). Например, имеется активатор метанолдегидрогеназы Bacillus methanolicus штамма C1 (Eur. J. Biochem., 244:426-433 (1997)) и продукт гена YqkG Bacillus subtilis штамма 168 (выложенная заявка на выдачу патента Японии No. 2000-69976). Применение указанных активаторов является эффективным средством усиления активности MDH. При введении ДНК, кодирующей любой из указанных активаторов MDH (гена amd) в микроорганизм, содержащий ген MDH, можно усилить активность MDH в клетках микроорганизма. Ген, кодирующий Amd (amd), такой как ген YqkG, можно получить из хромосомной ДНК Bacillus subtilis, такой как Bacillus subtilis штамма 168, посредством ПЦР, используя хромосомную ДНК в качестве матрицы и праймеры, имеющие нуклеотидные последовательности, указанные в списке последовательностей в виде SEQ ID NO: 10 и 11.
В качестве конкретного примера гена YqkG, используемого в данном изобретении, можно указать ген YqkG штамма 168 Bacillus subtilis. Нуклеотидная последовательность и аминокислотная последовательность, кодируемая указанным геном, показаны как SEQ ID NO: 14 и 15.
Объяснение способов усиления активностей HPS и PHI микроорганизма будет дано ниже.
Для того чтобы усилить активности HPS или PHI в микроорганизме-мишени, можно получить рекомбинантную ДНК путем лигирования гена, кодирующего HPS (hps) или PHI (phi) с вектором, функционирующим в микроорганизме-мишени, предпочтительно вектором мультикопийного типа, и ввести в микроорганизм-мишень, чтобы его трансформировать. При этом увеличивают число копий гена hps или гена phi в клетке трансформированного штамма, и в результате усиливают ту или иную ферментативную активность.
Ген hps или phi можно получить из микроорганизма, который продуцирует HPS или PHI таким же способом, который используют в обычном методе клонирования генов, подобно гену MDH.
В качестве микроорганизма, который продуцирует HPS, известны Methylomonas capsulatus (J.R.Quayle, Methods in Enzymology, 188, p.314, 1990), штамм М15 Methylomonas (Methods in Enzymology, 188, p. 319, 1990), штамм 77а Methylomonas aminofaciens (Biochim. Biophys. Acta., 523, p. 236, 1978), Methylomonas gastri MB19 (Methods in Enzymology, 188, p.393, 1990), Acetobacter methanolicus MB58 (Methods in Enzymology, 188, p. 401), и т.д. Кроме того, в качестве микроорганизма, который продуцирует PHI, известны штамм 77 а Methylomonas aminofaciens (Agric. Biol. Chem., 41 (7), p.1133, 1977), Mycobacterium gastri (выложенная заявка на выдачу патента Японии No. 11-127869), которая является грам-положительной факультативной ассимилирующей метанол бактерией, и т.д. Кроме того, сообщалось о наличии как гена hps, так и гена phi для Bacillus subtilis (J. Bacteriol., 181:7154-7160 (1999)). К тому же сообщалось, что в штамме S1 Bacillus brevis, которая является ассимилирующей метанол бактерией, относящейся к роду Bacillus, ген hps и ген phi существуют в тандеме на хромосомной ДНК (Annual Meeting of the Society for Fermentation and Bioengineering Japan, Lecture Abstracts, p.113 (2000)). Фрагмент ДНК, содержащий гены hps и phi, можно получить посредством ПЦР, используя хромосомную ДНК штамма в качестве матрицы и олигонуклеотиды, имеющие нуклеотидные последовательности, показанные как SEQ ID NO: 12 и 13, в качестве праймеров.
В качестве конкретных примеров гена hps и гена phi, используемых для данного изобретения, можно указать ген hps и ген phi штамма 168 Bacillus subtilis. Нуклеотидная последовательность фрагмента ДНК, содержащего гены hps и phi штамма SI Bacillus brevis, показана как SEQ ID NO: 16. Аминокислотные последовательности, кодируемые генами, показаны как SEQ ID NO: 17 и 18.
Штамм SI Bacillus brevis (NCIMB 12524) можно получить из национальных коллекций промышленных и морских бактерий, адрес: NCIMB Lts., Torry Research Station 135, Abbey Road, Aberdeen AB9 8DG, United Kingdom).
Активность HPS можно измерить способом, описанным в Methods in Enzymology, 188, 397-401 (1990). Кроме того, активность PHI можно измерить способом, описанным в Journal of Bacteriology, 181, р. 7154-7160 (1999).
Усиления активности HPS или PHI также можно достичь путем введения множественных копий гена hps или гена phi в хромосомную ДНК микроорганизма-мишени. Для того чтобы ввести множественные копии гена hps или гена phi в хромосомную ДНК микроорганизма-мишени, выполняют гомологичную рекомбинацию, используя в качестве мишени последовательность, множественные копии которой присутствуют в хромосомной ДНК. В качестве последовательностей, множественные копии которых существуют в хромосомной ДНК, можно использовать повторяющуюся ДНК или инвертированный повтор, существующий на конце транспозируемого элемента. Кроме того, как описано в выложенной заявке на выдачу патента Японии No. 2-109985, также можно включать ген hps или ген phi в транспозон, и обеспечивать возможность для его переноса, чтобы ввести множественные копии генов в хромосомную ДНК. Согласно любому из указанных способов активность HPS или PHI усиливают в результате увеличения числа копий гена hps или гена phi в трансформированном штамме.
Усиления активности HPS или PHI также можно достичь не только способом, основанным на указанной выше амплификации генов, но и заменой последовательности регуляции экспрессии, такой как промотор гена hps или гена phi, более сильным промотором (смотри выложенную заявку на выдачу патента Японии No. 1-215280). Например, как сильные промоторы известны промотор lac, промотор trp, промотор trc, промотор tac, промотор PR и промотор PL фага лямбда, промотор tet, промотор amyE, промотор veg и так далее. Замена указанными промоторами усиливает экспрессию гена hps или гена phi, и, следовательно, усиливается активность HPS или PHI. Усиление последовательности регуляции экспрессии можно комбинировать с увеличением числа копий HPS или PHI.
Гены mdh, hps, phi и amd, используемые для данного изобретения, не ограничены генами дикого типа, и они могут быть мутантными или искусственно модифицированными генами, кодирующими генный продукт, включающий замену, делецию, инсерцию, присоединение или инверсию одной или нескольких аминокислот в одном или нескольких сайтах, при условии, что не понижается функция кодируемого белка MDH, HPS, PHI или Amd. Хотя количество указанных здесь "нескольких" аминокислот отличается в зависимости от положения или типа аминокислотных остатков в трехмерной структуре белка, оно в частности может составлять от 2 до 20, предпочтительно от 2 до 10, более предпочтительно от 2 до 5.
В качестве ДНК, кодирующей белок, в значительной степени идентичный вышеуказанному белку Amd, можно указать ДНК, которая способна гибридизоваться с нуклеотидной последовательностью, содержащей последовательность нуклеотидов с номерами от 1 до 555 в SEQ ID NO: 16, или зондом, который можно получить из нуклеотидной последовательности, в жестких условиях, и кодировать белок, обладающий активностью, подобной активности Amd.
Кроме того, в качестве ДНК, кодирующей белок, в значительной степени идентичный белку HPS, можно указать ДНК, которая способна гибридизоваться с нуклеотидной последовательностью, содержащей последовательность нуклеотидов с номерами от 508 до 1140 в SEQ ID NO: 16, или зондом, который можно получить из нуклеотидной последовательности, в жестких условиях, и кодировать белок, обладающий активностью, подобной активности HPS.
Кроме того, в качестве ДНК, кодирующей белок, в значительной степени идентичный белку PHI, можно указать ДНК, которая способна гибридизоваться с нуклеотидной последовательностью, содержащей последовательность нуклеотидов с номерами от 1149 до 1700 в SEQ ID NO: 16, или зондом, который можно получить из нуклеотидной последовательности, в жестких условиях, и кодировать белок, обладающий активностью, подобной активности PHI.
Кроме того, в качестве ДНК, кодирующей белок, в значительной степени идентичный белку MDH, можно указать ДНК, которая способна гибридизоваться с нуклеотидной последовательностью, зарегистрированной в GenBank под инвентарным номером М65004 (имя для ввода BACMDH), или зондом, который можно получить из нуклеотидной последовательности, в жестких условиях, и кодировать белок, обладающий активностью, подобной активности MDH.
Вышеуказанные "жесткие условия" представляют собой условия, при которых образуется так называемый специфичный гибрид, и не образуется неспецифичный гибрид. Трудно ясно выразить такое условие, используя численное значение. Однако, например, примером жестких условий является условие, при котором ДНК, обладающие высокой степенью гомологии, например, обладающие 50% или большей гомологией, гибридизуются друг с другом, а ДНК, обладающие более низкой степенью гомологии, чем указанная выше, не гибридизуются друг с другом. Альтернативно, примером жестких условий является условие, при котором ДНК гибридизуются друг с другом при концентрации соли, соответствующей обычному условию промывки при Саузерн-гибридизации, т.е. 1×SSC, 0,1% SDS, предпочтительно 0,1×SSC, 0,1% SDS при 60°С.
Чтобы ввести в микроорганизм различные гены, которые можно получить, как описано выше, например, применим способ обработки реципиентных клеток хлоридом кальция, для того, чтобы увеличить проницаемость ДНК, который известен для Escherichia coli K-12 (Mandel, M. And Higa, A., J. Mol. Biol., 53, 159 (1970)), и способ получения компетентных клеток из клеток, которые находятся в фазе роста, с последующим введением в них ДНК, который известен для Bacillus subtilis (Duncan, C.H., Wilson, G.A. and Young, F.E., Gene, 1, 153 (1977)). В добавление к указанному также применим способ превращения ДНК-реципиентных клеток в протопласты или сферопласты, которые могут легко захватывать рекомбинантную ДНК, с последующим введением рекомбинантной ДНК в клетки, который, как известно, подходит для Bacillus subtilis, актиномицетов и дрожжей (Chang, S. and Choen, S.N., Molec. Gen. Genet., 168, 111 (1979); Bibb, M.J., Ward, J.M. and Hopwood, O.A., Nature, 274, 398 (1978); Hinnen, A., Hicks, J.B. and Fink, G.R., Proc. Natl. Acad. Sci., USA, 75, 1929 (1978)). Кроме того, можно использовать способ электропорации (Canadian Journal of Microbiology, 43, 197 (1997)). Любой из указанных способов можно соответственно выбрать в зависимости от клеток, используемых в качестве реципиента.
В микроорганизме, используемом для данного изобретения, можно усилить активность фермента, вовлеченного в биосинтез целевого вещества, в соответствии с типом целевого вещества. Кроме того, активность фермента, снижающего продукцию целевого вещества, можно уменьшить или элиминировать.
В том случае, когда требуется введение в микроорганизм генов mdh, hps, phi и amd, порядок введения генов особым образом не ограничен. Кроме того, микроорганизм согласно данному изобретению можно получить либо путем введения указанных генов в микроорганизм, обладающий способностью продуцировать целевое вещество, либо путем придания способности продуцировать целевое вещество микроорганизму, которому вводят указанные гены.
Целевое вещество можно получить на основе использования метанола, культивируя микроорганизм согласно данному изобретению, полученный как описано выше, в среде, чтобы продуцировать и накопить целевое вещество в среде или клетках микроорганизма, и собирая целевое вещество из среды или клеток микроорганизма.
Целевые вещества, по отношению к которым применим способ согласно данному изобретению, включают, например, вещества, получаемые при метаболизме метанола, и вещества, получаемые при использовании энергии, генерируемой в ходе метаболизма метанола. В частности, можно, например, указать аминокислоты, такие как глутаминовая кислота, лизин, треонин, фенилаланин и триптофан, витамины, такие как витамин С, макромолекулярные вещества, такие как различные виды ферментов, и так далее.
Термин "способность продуцировать целевое вещество", используемый в данном изобретении, означает способность микроорганизма согласно данному изобретению продуцировать и накапливать целевое вещество в среде или клетках микроорганизма в таком количестве, в котором вещество оттуда можно собрать, при культивировании микроорганизма в среде в соответствующих условиях.
Хотя среду и условия культивирования, используемые для культуры в способе согласно данному изобретению, можно соответственно выбрать в зависимости от типа используемого микроорганизма, можно использовать стандартные среды, содержащие источник азота, неорганические ионы и другие органические следовые питательные вещества при необходимости.
В качестве источника углерода используют метанол. Наряду с метанолом можно использовать сахариды, такие как глюкоза, лактоза, галактоза, фруктоза и гидролизат крахмала, спирты, такие как глицерин и сорбит, или органические кислоты, такие как фумаровая кислота, лимонная кислота и янтарная кислота.
В качестве источника азота можно использовать неорганические соли аммония, такие как сульфат аммония, хлорид аммония и фосфат аммония, органический азот, такой как гидролизат соевого белка, газообразный аммиак, водный раствор аммиака и так далее.
В небольших количествах добавляют неорганические ионы или их источники, фосфат калия, сульфат магния, ион железа, ион марганца и так далее. В качестве следовых органических питательных веществ желательно добавлять требуемые вещества, такие как L-гомосерин и витамин B1, дрожжевой экстракт и так далее в соответствующих требуемых количествах.
Культивирование предпочтительно выполняют в условиях, подходящих для используемого микроорганизма. Обычно культивирование предпочтительно проводят в аэробных условиях в течение 16-72 часов. Температуру культивирования предпочтительно регулируют так, чтобы она составляла от 20°С до 45°С, и значение рН предпочтительно регулируют так, чтобы оно в ходе культивирования составляло от 5 до 8,5. Для доведения рН можно использовать неорганические или органические, кислые или щелочные вещества, а также газообразный аммиак и так далее. Если в качестве хозяина используют термофильную бактерию, ее можно культивировать при температуре культивирования от 42°С до 60°С.
Для сбора продукта метаболизма из среды после завершения культивирования для данного изобретения не требуется никакой специальный способ. А именно, сбор можно осуществлять посредством комбинирования хорошо известных способов, таких как способы с использованием ионообменных смол, осаждения и другие способы. Кроме того, в том случае, когда в качестве источника углерода используют метанол, очистку целевого вещества и процесс удаления отработанного раствора можно упростить, по сравнению с тем случаем, когда используют сахара, полученные из сельскохозяйственных продуктов.
Примеры
Далее будет дано более конкретное объяснение данного изобретения со ссылкой на следующие примеры.
Пример 1: Клонирование гена метанолдегидрогеназы.
Хромосомную ДНК получали традиционным способом из Bacillus brevis штамма S1 (NCIMB 12524, полученного из NCIMB), которая является ассимилирующей метанол, резистентной к высокой температуре бактерией, относящейся к роду Bacillus. Затем ген MDH клонировали с использованием ПЦР, используя полученную ДНК в качестве матрицы (смотри выложенную заявку на выдачу патента Японии No. 2000-69976).
Использовали ДНК-праймеры MDH-BM-1 (SEQ ID NO: 1) и MDH-BM-2 (SEQ ID NO: 2). Указанные праймеры получали, обращаясь к уже известной нуклеотидной последовательности гена MDH Bacillus methanolicus штамма С1 (зарегистрированного в GenBank под инвентарным номером М65004, имя для ввода BACMDH). ПЦР выполняли, используя LA-Taq (Takara Shuzo), и ПЦР включала тепловую обработку при 94°С в течение 90 секунд, затем реакции при 98°С в течение 10 секунд, при 55°С в течение 30 секунд и 70°С в течение 4 минут, повторяемых в ходе 30 циклов, и затем следовала инкубация при 72°С в течение 10 минут. В ходе указанных реакций получали фрагмент ДНК требуемого размера. Данный фрагмент ДНК очищали и затем клонировали в коммерчески доступном векторе, pCR2.1, чтобы получить pCR-mdh24-1.
Для того чтобы ввести ген mdh, находящийся в вышеуказанном pCR-mdh24-1, в хромосому Bacillus subtilis, ген mdh включали в вектор для введения гена в хромосому, плазмиду pDLT3. Плазмиду pDLT3 получали традиционной технологией рекомбинантной ДНК для введения гена-мишени в сайт amyE хромосомы Bacillus subtilis, и ее полная нуклеотидная последовательность показана в виде SEQ ID NO: 3.
pDLT3 получали следующим образом (фиг.1). Сначала для того, чтобы удалить фрагмент гена lacZ из pMUTIN3 (описана в Molecular Microbiology, 29 (1), 179-187 (1998)), плазмиду расщепляли ферментами рестрикции Clal и Bpu1102I. Затем расщепленные концы затупляли ферментом Кленова, и больший по размеру фрагмент ДНК подвергали самолигированию, используя лигазу Т4. Затем pDLd (описана в Molecular Microbiology, 29(2), 505-513, (1998)) расщепляли сходным образом посредством обработки ферментом рестрикции, и выделяли больший по размеру фрагмент ДНК (Tthl111-BamHI). Фрагмент ДНК Tthl111-Bg1II плазмиды, содержащей промоторы lacI и spac лигировали с большим по размеру фрагментом ДНК, чтобы сконструировать pDLT3.
Плазмиду pDLT3 обрабатывали ферментами рестрикции BamHI и EcoRI обычным способом, получая больший фрагмент ДНК. Отдельно pCR-mdh24-1 сходным образом обрабатывали ферментами рестрикции BamHI и EcoRI, подобным образом получая фрагмент ДНК, содержащий ген mdh. Оба полученных фрагмента лигировали, используя лигазу Т4, чтобы сконструировать плазмиду pDLT-MD11, в которой ген mdh включен между районом первой половины и районом второй половины гена amyE в pDLT3.
Полученные кольцевые плазмиды, pDLT-MD11 и pDLT3, переводили в линейную форму обработкой ферментом рестрикции Bst1107I, и каждой из них трансформировали штамм 168 Bacillus subtilis традиционным способом. Отбор трансформантов проводили как отбор штаммов, устойчивых к хлорамфениколу. Поскольку в каждом случае получили много устойчивых к хлорамфениколу штаммов, в каждом случае отобрали шесть колоний, и исследовали, был ли введен в сайт amyE на хромосоме только ген mdh или вектор, как предполагалось. Можно ли ввести линеаризованный ген mdh в ходе дважды происходящего процесса рекомбинации, подтверждали посредством ПЦР колоний. Использовали праймеры ДНК N1, N2, N3, C1, C2 и С3 (SEQ ID NO: 4, 5, 6, 7, 8 и 9, соответственно, в указанном порядке). В результате анализа фрагментов ДНК, амплифицированных в ПЦР, отобрали штамм AM1 01 в качестве штамма, имеющего структуру, в которой ген mdh включен в предназначенный район. С другой стороны, штамм, в который был включен векторный район, отобрали в качестве контрольного штамма и назвали штаммом DT101.
Затем определяли активность MDH, экспрессированного в штамме АМ101, используя в качестве контроля штамм DT101. Оба штамма инокулировали по отдельности в 30 мл среды LB, содержащей 5 мкг/мл хлорамфеникола, и культивировали при 30°С примерно в течение 16 часов. Культуральный бульон центрифугировали (10000 g, 10 минут), чтобы собрать клетки в виде осадка, и затем клетки суспендировали в 0,8 мл буфера для суспендирования (состав: 5 мл 0,1 М фосфатного буфера (рН 7,6), 1 мл 50 мМ сульфата магния, 0,1 мл 0,2 М дитиотреитола, 3,3 мл 60% сахарозы и 0,6 мл стерильной воды на 10 мл) и разрушали ультразвуком при поддержании температуры суспензии на уровне температуры ниже 4°С. Затем твердое вещество, находящееся в суспензии, удаляли центрифугированием (17000 g, 10 минут) суспензии при 4°С, чтобы получить клеточный экстракт.
Ферментативную активность MDH измеряли следующим образом. Сначала 0,9 мл реакционного буфера (10 мл 0,1 М глицина/гидроксида натрия (рН 9,5), 2 мл 50 мМ сульфата магния, 0,1 мл 0,2 М дитиотреитола, 0,5 мл 40 мМ NAD, 7,4 мл стерильной воды на 20 мл) помещали в кварцевую кювету объемом 1,5 мл, и кювету вставляли в спектрометр. В спектрометре температуру реакционной смеси контролировали инкубацией при 47°С в течение 3 минут. Затем 50 мкл клеточного экстракта, полученного в ходе описанной процедуры, добавляли к реакционной смеси и перемешивали в достаточной мере. В это время начинали измерение оптической плотности реакционной смеси при 340 нм, при которой NADH проявляет максимум оптической плотности, чтобы исследовать изменение оптической плотности в условиях отсутствия метанола. Затем примерно через 60 секунд к реакционной смеси добавляли 50 мкл метанола и в достаточной степени перемешивали, и далее наблюдали и регистрировали увеличение оптической плотности при 340 нм, т.е. образование NADH. Увеличения оптической плотности при 340 нм из-за MDH не наблюдали в том случае, когда использовали клеточный экстракт контрольного штамма, штамма DT101. Однако в том случае, когда использовали экстракт штамма AM101, наблюдали увеличение оптической плотности при 340 нм в зависимости от добавления метанола, и это подтверждает, что в данном штамме присутствует MDH. Таким образом, обнаружено, что ген mdh, включенный в хромосому Bacillus subtilis, кодировал фермент MDH, который мог проявлять ферментативную активность. NAD и NADH представляют собой окисленный тип и восстановленный тип никотинамидадениндинуклеотида, соответственно.
Пример 2: Включение mdh в хромосому штамма Bacillus subtilis, продуцирующего L-лизин.
Штамм AJ11779 Bacillus subtilis, модифицированный так, что он может продуцировать L-лизин, представляет собой штамм, описанный в выложенной заявке на выдачу патента Японии No. 58-149689. Указанному штамму способность продуцировать L-лизин придавали посредством придания устойчивости к ингибированию роста АЕС, который является соединением-аналогом лизина, и треонином. Указанный штамм был депонирован в независимой административной организации National Institute of Advanced Industrial Science and Technology, International Patent Organism Depository (адрес: почтовый индекс 305-5466, Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Japan) 9 августа 2001 г., и получил инвентарный номер FERM P-18453. Затем, 19 августа 2002 г., депонирование заменили международным депонированием по условиям Будапештского договора, и получили инвентарный номер FERM BP-8155.
Планировали включить ген mdh, описанный в примере 1, в сайт amyE хромосомы штамма AJ11779. Сначала, кольцевую плазмиду pDLT-MD11 переводили в линейную форму обработкой ферментом рестрикции Bst1107I, и использовали для трансформации штамма AJ11779 обычным способом. Трансформанты отбирали на основе устойчивости штаммов к хлорамфениколу.
Поскольку получили много устойчивых к хлорамфениколу штаммов, из них отбирали две или более колоний, и штамм, в котором ген mdh был введен в сайт хромосомы amyE, отбирали таким же образом, как в примере 1. Полученный штамм назвали штаммом L-217M1.
Из указанного штамма L-217M1 получали клеточный экстракт, и активность MDH измеряли таким же способом, как в примере 1. В результате можно было выявить активность MDH.
Пример 3: Клонирование гена, кодирующего активатор MDH (Amd), полученного из Bacillus subtilis.
Известно, что существуют факторы активации ферментативной активности метанолдегидрогеназы NAD-зависимого типа, полученной из ассимилирующей метанол бактерии, относящейся к роду Bacillus. В выложенной заявке на выдачу патента Японии No. 2000-69976 (ОР884) описано, что один из таких факторов имеется в Bacillus subtilis. Данный фактор назван Amd (активатор метанолдегидрогеназы).
Ген, кодирующий Amd (amd), клонировали из Bacillus subtilis уже известным способом. В частности, клонирование проводили следующим образом. Штамм 168 Bacillus subtilis культивировали в среде LB, и из полученных клеток экстрагировали хромосомную ДНК традиционным способом (Biochem. Biophys. Acta., 72, 619-629 (1963)). Полученную хромосомную ДНК использовали в качестве матрицы в ПЦР, используя такие олигонуклеотиды, чтобы фрагмент ДНК-мишени содержал сайты фермента рестрикции EcoRI на обоих концах (SEQ ID NO: 10 и 11), чтобы амплифицировать фрагмент ДНК гена, содержащего amd, который являлся геном-мишенью. Для амплификации 30 раз повторяли цикл, состоящий из стадии денатурации при 98°С в течение 10 секунд, стадии отжига при 55°С в течение 30 секунд и стадии элонгации при 72°С в течение 2 минут. Используемым ферментом была ДНК-полимераза Pyrobest (Takara Shuzo), и ее использовали согласно инструкции производителя.
Амплифицированный фрагмент ДНК очищали обработкой фенолом/хлороформом и осаждением этанолом, и затем расщепляли ферментом рестрикции EcoRI, чтобы получить фрагмент amd, имеющий сайты EcoRI на обоих концах. Отдельно pHY300PLK (Takara Shuzo), которая является челночным вектором для Escherichia coli и Bacillus subtilis, сходным образом обрабатывали ферментом рестрикции EcoRI. После удаления фосфатных групп на концах с использованием щелочной фосфатазы вектор лигировали с вышеуказанным фрагментом amd. Используя полученную реакционную смесь для лигирования, компетентные клетки Е. coli штамма JM109 (Takara Shuzo) трансформировали согласно протоколу производителя, и отбирали несколько устойчивых к тетрациклину колоний.
Из полученных колоний экстрагировали плазмидную ДНК и анализировали ее структуру. Затем плазмиды, содержащие ген amd в таком же направлении, как и направление гена устойчивости к тетрациклину вектора, и в обратном направлении, обозначали соответственно pHY-A2 и pHY-A1, и использовали в последующих экспериментах.
Активность Amd, которая усиливает активность MDH, исследовали следующим образом. Штамм АМ101 Bacillus subtilis с включенным геном mdh, который сконструирован в примере 1, трансформировали обычным способом с помощью каждой из плазмид pHY-A1 и pHY-A2, а также pHY300PLK, которая является векторной плазмидой. Затем из каждого трансформанта получали клеточный экстракт следующим образом.
Каждый штамм трансформанта инокулировали в 30 мл среды LB, содержащей 5 мкг/мл хлорамфеникола и 10 мкг/мл тетрациклина, и культивировали при 30°С примерно в течение 16 часов. Каждый культуральный бульон центрифугировали (при 10000 g в течение 10 минут), чтобы собрать клетки в виде осадка, и затем клетки суспендировали в 0,8 мл буфера для суспендирования (5 мл 0,1 М фосфатного буфера (рН 7,6), 1 мл 50 мМ сульфата магния, 0,1 мл 0,2 М дитиотреитола, 3,3 мл 60% сахарозы и 0,6 мл стерильной воды на 10 мл) и разрушали ультразвуком при поддержании температуры суспензии на уровне температуры, равной 4°С, чтобы получить клеточный экстракт.
Усиление ферментативной активности MDH под действием Amd измеряли с помощью описанного ниже анализа. Реакционный буфер в объеме 0,9 мл (10 мл 0,1 М глицина/гидроксид а натрия (рН 9,5), 2 мл 50 мМ сульфата магния, 0,1 мл 0,2 М дитиотреитола, 0,5 мл 40 мМ NAD, 7,4 мл стерильной воды на 20 мл) инкубировали в кварцевой кювете при 37°С в течение 3 минут. Затем 50 мкл клеточного экстракта, полученного как описано выше, добавляли к реакционной смеси и перемешивали. В это время начинали измерение временного хода изменения оптической плотности реакционной смеси при 340 нм, при которой NADH проявляет максимум оптической плотности, и регистрировали изменение оптической плотности при 340 нм, которое не связано с добавлением метанола. Затем примерно через 60 секунд к реакционной смеси добавляли 50 мкл метанола и перемешивали, и в это время исследовали увеличение оптической плотности при 340 нм, т.е. образование NADH. В результате, по сравнению со скоростью увеличения оптической плотности при 340 нм из-за наличия MDH, наблюдаемой в случае использования клеточного экстракта штамма, в который вводили pHY300PLK, наблюдали по существу эквивалентную скорость увеличения оптической плотности при 340 нм в случае использования штамма, в который вводили pHY-A1, и в 5 раз более высокую скорость образования NADH наблюдали в случае использования штамма, в который вводили pHY-A2. Таким образом, доказали, что Amd, кодируемый pHY-A2, несомненно, будет обладать активностью в усилении активности MDH.
Пример 4: Клонирование гена hps и гена phi из ассимилирующей метанол бактерии, относящейся к роду Bacillus.
Получали хромосомную ДНК из штамма S1 Bacillus brevis, которая является ассимилирующей метанол бактерией, относящейся к роду Bacillus, таким же способом, как способ, описанный выше. Полученную хромосомную ДНК использовали в качестве матрицы в ПЦР, чтобы амплифицировать район ДНК мишени. Последовательности олигонуклеотидов для ПЦР (SEQ ID NO: 12 и 13) конструировали так, что сайты фермента рестрикции HindIII были введены в оба конца амплифицированного фрагмента ДНК. Условия ПЦР были такими же, как условия, используемые выше. Затем полученный фрагмент ДНК очищали стандартным способом, и затем обрабатывали ферментом рестрикции HindIII, чтобы получить ДНК мишени, имеющие расщепляемые HindIII концы на обоих концах.
Отдельно pHY300PLK обрабатывали ферментом рестрикции HindIII, затем обрабатывали щелочной фосфатазой и лигировали с вышеуказанным фрагментом ДНК, используя лигазу Т4 (Takara Shuzo). Полученный продукт использовали для того, чтобы трансформировать Е. coli штамма JM109 таким же способом, как описанный выше, чтобы получить множество устойчивых к тетрациклину колоний. Из них отбирали несколько колоний, и исследовали находящиеся в них плазмиды, чтобы отобрать одну, в которой гены-мишени, ген hps и ген phi, существовали в такой же ориентации, что и ориентация гена устойчивости к тетрациклину в векторе. Полученную плазмиду назвали pHY-H4.
Затем штамм AM101, указанный в примере 1, трансформировали обычным способом каждой из плазмид - pHY-H4, сконструированной как описано выше, и pHY300PLK, которая является вектором, используемым для конструирования pHY-H4, чтобы, соответственно, получить трансформанты, штамм pHY-H4/AM101 и штамм pHY300PLK/AM101. Собирали колонии каждого штамма, инокулировали в 5 мл среды LB, содержащей 10 мкг/мл тетрациклина и 5 мкг/мл хлорамфеникола, и затем культивировали при 30°С в течение 16 часов. Затем 0,1 мл культурального бульона переносили в 5 мл свежей среды LB, содержащей такие же антибиотики, которые указаны выше, и далее культивировали при 37°С в течение 8 часов. Культуральный бульон центрифугировали (10000 g, 10 минут), чтобы собрать клетки.
Полученные клетки суспендировали в 0,9 мл буфера для суспендирования (состав на 10 мл: 5 мл 0,1 М буфера на основе фосфата калия (рН 7,6), 0,3 мл 0,1 М хлорида магния и 50 мкл 0,2 М дитиотреитола, дополненный до 10 мл стерильной водой), подвергали обработке ультразвуком при поддержании температуры на уровне 4°С и центрифугировали при 4°С (при 20000 g в течение 10 минут), и собирали надосадочную часть, чтобы получить клеточный экстракт. Затем в каждом экстракте исследовали активность HPS и активность PHI, как описано ниже.
Реакционный буфер в объеме 0,9 мл (состав указан ниже) инкубировали в кварцевой кювете при 47°С в течение 3 минут. Затем 50 мкл клеточного экстракта, полученного как описано выше, добавляли к реакционному буферу и перемешивали в достаточной степени. В это время начинали измерение временного хода изменения оптической плотности реакционной смеси при 340 нм, при которой NADH проявляет максимум оптической плотности. Через 60 секунд после добавления клеточного экстракта к реакционной смеси добавляли 50 мкл 1М водного раствора формальдегида и перемешивали в достаточной степени. Затем регистрировали увеличение оптической плотности реакционной смеси при 340 нм, т.е. образование NADH. В то время как увеличение оптической плотности при 340 нм вследствие добавления формальдегида по существу не наблюдали в случае использования экстракта из контрольного штамма, штамма AM101, несущего pHY300PLK, быстрое увеличение оптической плотности наблюдали в случае использования штамма, несущего pHY-H4. Это свидетельствует о том, что каждый из продуктов гена hps и гена phi, клонированных в pHY-H4, проявлял ферментативную активность в Bacillus subtilis, в конечном итоге продуцируя фруктозо-6-фосфат из формальдегида и риболозо-5-фосфата. Таким образом, подтверждено, что были клонированы ген hps и ген phi (см. табл.1).
Состав буфера реакции
Пример 5: Конструирование плазмиды, содержащей hps, phi и amd.
Каждую из плазмид pHY-A2 и pHY-H4, полученных в примерах 3 и 4, обрабатывали двумя видами ферментов рестрикции, Bg1II и ЕсоТ22I. Из pHY-A2 получали меньший фрагмент ДНК, содержащий ген amd. С другой стороны, из pHY-H4 обычным способом получали больший фрагмент ДНК, содержащий гены hps и phi, и лигировали с фрагментом гена amd, используя лигазу Т4.
Используя указанную выше реакционную смесь, трансформировали компетентные клетки Е. coli JM109. Трансформанты отбирали, используя в качестве показателя устойчивость к тетрациклину. Из нескольких десятков колоний, появившихся на чашке с агаром, произвольно отбирали 6 колоний, и анализировали структуры находящихся в них плазмид. В результате подтвердили, что все плазмиды имели запланированную структуру, т.е. структуру, при которой в векторе рНY300РLК имелось три вида генов, amd, hps и phi. Полученную плазмиду назвали pHY-A2H4.
Пример 6: Получение штамма Bacillus subtilis, которому придана способность утилизировать метанол, и анализ способности полученного штамма утилизировать метанол.
Штамм L-217M1, сконструированный в примере 2, культивировали в течение ночи при 30°С в жидкой среде LB, содержащей 5 мкг/мл хлорамфеникола, при встряхивании. Культуральный бульон в объеме 0,1 мл добавляли к 5 мл среды CI (состав указан ниже), содержащей 5 мкг/мл хлорамфеникола, и перемешивали, и клетки культивировали при 37°С со встряхиванием. Измеряли оптическую плотность культурального бульона (660 нм) с соответствующими интервалами, и через 60 минут после того, как оптическая плотность превышала 1, культуральный бульон центрифугировали (3000 g, 10 минут), чтобы собрать клетки.
Затем собранные клетки суспендировали в 10 мл среды CII (содержащей 5 мкг/мл хлорамфеникола, состав указан ниже) и далее культивировали в течение 40 минут в данном культуральном бульоне, содержащем среду CII, при встряхивании. Затем извлекали 500 мкл культурального бульона, к нему добавляли 1 мкг pHY-A2H4, и клетки культивировали при 37°С в течение 90 минут при встряхивании. Полученный культуральный бульон соответствующим образом разбавляли и затем инокулировали в агаровую среду LB, содержащую 5 мкг/мл хлорамфеникола и 10 мкг/мл тетрациклина, и клетки культивировали в течение ночи при 37°С как обычную культуру, чтобы отобрать колонии, проявляющие устойчивость к обоим антибиотикам. Одной из них был штамм pHY-A2H4/L-217M1.
Штамм pHY-A2H4/L-217M1 был назван AJ13855 и депонирован в независимой административной организации National Institute of Advanced Industrial Science and Technology, International Patent Organism Depository (почтовый индекс 305-5466, Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Japan) 2 августа 2001 г., и получил инвентарный номер FERM Р-18447. Затем, 19 августа 2002 г., депонирование заменили международным депонированием по условиям Будапештского договора и получили инвентарный номер FERM BP-8154.
Среду CI, в объеме 5 мл, получали следующим образом. Сначала к 2,5 мл 2 × раствора А (К2НРО4: 2,8%; КН2PO4: 1,2%; (NH4)2SO4: 0,4%) добавляли 5 мкл раствора микроэлементов (CaCl2: 0,55 г; ZnCl2: 0,17 г; CuCl2·2H2O: 0,043 г; CoCl2·6H2O: 0,06 г; Na2MoO4·2Н2О: 0,06 г на 100 мл), 1 мкл раствора FeCl3 и 1 мкл раствора MnSO4, перемешивали и затем добавляли 2,5 мл 2 × раствора В (MgSO4·7H2O: 10 мМ; глюкоза: 1%), 50 мкл 5% (мас./об.) дрожжевого экстракта, 10 мкл 10% (мас./об.) казаминовой кислоты, 50 мкл раствора L-триптофана с концентрацией 5 мг/мл, 50 мкл раствора L-метионина с концентрацией 5 мг/мл и 50 мкл раствора тимина с концентрацией 2,5 мг/мл. Можно использовать К2HPO4 и КН2РО4 от Nakarai Tesque, (HN4)2SO4, MgSO4·7H2O и глюкозу от Junsei Chemical, и дрожжевой экстракт и казаминовую кислоту от DIFCO. CaCl2, ZnCl2, CuCl2·2H2O, CoCl2·6Н2О и Na2MoO4·2Н2O можно использовать от Wako Pure Chemical Industries.
Среду CII, объемом 10 мл, получали следующим образом. Сначала к 5 мл 2 × раствора А добавляли 10 мкл раствора микроэлементов, 2 мкл раствора FeCl3 и 2 мкл раствора MnSO4 и перемешивали. Затем к полученному раствору добавляли 5 мл 2 × раствора В, 10 мкл 5% (мас./об.) дрожжевого экстракта, 10 мкл 10% (мас./об.) казаминовой кислоты, 10 мкл раствора L-триптофана с концентрацией 5 мг/мл, 10 мкл раствора L-метионина с концентрацией 5 мг/мл и 10 мкл раствора тимина с концентрацией 2,5 мг/мл.
Затем, используя метанол, меченый стабильным изотопом 13С, проверяли, может ли сконструированный штамм AJ13855 (pHY-A2H4/L-217M1) утилизировать метанол. Штамм AJ13855 культивировали в течение ночи при 30°С в жидкой среде LB, содержащей 5 мкг/мл хлорамфеникола и 10 мкг/мл тетрациклина, при встряхивании. Полученный культуральный бульон инокулировали в жидкую среду LB, содержащую 5 мкг/мл хлорамфеникола и 10 мкг/мл тетрациклина в соотношении 1% (об./об.), культивирование проводили при 37°С в течение 6 часов при встряхивании, и культуральный бульон инокулировали в каждую из следующих сред, а именно среду А, содержащую 13С-меченый метанол, и среду А, содержащую обычный метанол, при отношении 3% (об./об.).
Состав среды А указан в таблице 2. Использовали среду, полученную добавлением 13С-меченого метанола к 50 мл среды А в конечной концентрации 0,4% (об./об.), и среду, полученную добавлением немеченого 13С обычного метанола к 50 мл среды А в конечной концентрации 0,4% (об./об.). После инокуляции клетки культивировали при 47°С в течение 30 часов при встряхивании. После культивирования оптическая плотность при 660 нм обоих культуральных бульонов, которая указывает степень роста, достигала примерно 2, 3, и таким образом, не было существенного различия в росте бактерий в обеих средах. Затем оба культуральных бульона центрифугировали (при 8000 об/мин в течение 15 мин), чтобы получить надосадки, которые лиофилизировали.
Лиофилизированный порошок, полученный из каждого культурального бульона, в количестве 62 мг растворяли в 500 мкл тяжелой воды. Когда измеряли количество L-лизина в обоих растворах, оно составляло примерно 0,55 мг в обоих растворах, и таким образом, было обнаружено, что количества L-лизина в обоих растворах были почти одними и теми же. Затем растворы анализировали посредством 13С-ЯМР (устройство для анализа ядерного магнитного резонанса), чтобы определить долю 13С в продуцированных молекулах L-лизина. В результате при сравнении с сигналом атомов углерода L-лизина, полученного при культивировании с использованием добавления немеченого метанола, примерно в 3,8-13 раз более сильный сигнал регистрировали для L-лизина, полученного при культивировании с использованием добавления 13С-меченого метанола. Это свидетельствует о том, что сконструированный штамм АJ1 3855 (pHY-A2H4/L-217M1) заново приобрел способность поглощать 13С-меченый метанол, добавленный в среду, и даже использовать его для продукции L-лизина.
Состав среды А (на 130 мл)
До общего объема 200 мл доводили стерильной водой.
Список последовательностей
<110> Ajinomoto Co., Inc.
<120> Способ получения целевого вещества
<130> ОР1386
<140>
<141> 2002-09-06
<140> JP 2001-270902
<151> 2001-09-06
<160> 18
<170> Patent In Версия 2.0
<210> 1
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: праймер MDH-BM-1
<400> 1
taaaaaggat ccccgatgat acaacaccaa acgg 34
<210> 2
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: праймер MDH-BM-2
<400> 2















| название | год | авторы | номер документа |
|---|---|---|---|
| КОРИНЕФОРМНАЯ БАКТЕРИЯ, ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2004 |
|
RU2333247C2 |
| НОВЫЕ ФЕРМЕНТЫ МЕТАНОЛДЕГИДРОГЕНАЗЫ ИЗ Bacillus | 2013 |
|
RU2624017C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2351653C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2368659C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2250266C2 |
| СПОСОБ ПОЛУЧЕНИЯ СПИРТА С ИСПОЛЬЗОВАНИЕМ МИКРООРГАНИЗМА | 2002 |
|
RU2294370C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2008 |
|
RU2515044C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПУРИНОВЫХ РИБОНУКЛЕОЗИДОВ И РИБОНУКЛЕОТИДОВ | 2008 |
|
RU2422510C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2003 |
|
RU2307165C2 |
Изобретение относится к биотехнологии и представляет собой способ получения L-аминокислоты посредством культивирования бактерии рода Bacillus, обладающей способностью продуцировать L-аминокислоту, в среде, содержащей метанол в качестве источника углерода, и сбора L-аминокислоты из среды или клеток указанной бактерии. При этом в качестве бактерии используют бактерию, в которую введен ген метанолдегидрогеназы, а активности гексулозофосфатсинтазы и фосфогексулоизомеразы усилены. Изобретение позволяет получать L-аминокислоты с высокой степенью эффективности. 2 н. и 4 з.п. ф-лы, 1 ил., 2 табл.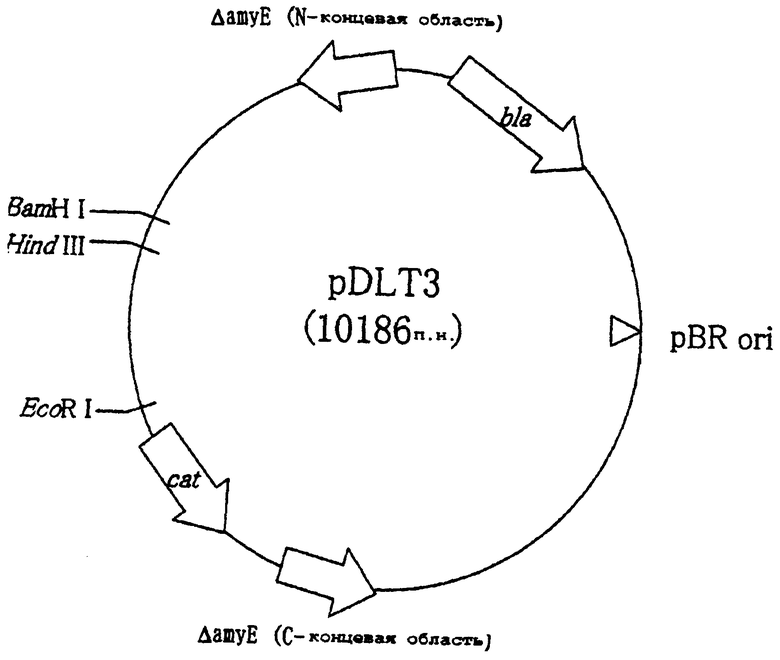
| US 5830716, 03.11.1998 | |||
| US 6083728, 04.07.2000 | |||
| МОДИФИЦИРОВАННАЯ ФОСФОЕНОЛПИРУВАТКАРБОКСИЛАЗА, ФРАГМЕНТ ДНК, ШТАММ ESCHERICHIA COLI, РЕКОМБИНАНТНАЯ ДНК, СПОСОБ ПОЛУЧЕНИЯ АМИНОКИСЛОТЫ (ВАРИАНТЫ), СПОСОБ СЕЛЕКЦИИ КЛЕТОК ESCHERICHIA COLI, СПОСОБ ПОЛУЧЕНИЯ МОДИФИЦИРОВАННОЙ ФОСФОЕНОЛПИРУВАТКАРБОКСИЛАЗЫ, СПОСОБ ПОЛУЧЕНИЯ ФРАГМЕНТА ДНК И СПОСОБ ПОЛУЧЕНИЯ МИКРООРГАНИЗМА | 1994 |
|
RU2133772C1 |

