ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к области ферментации, и более конкретно к получению L-аминокислот посредством ферментации микроорганизмов.
УРОВЕНЬ ТЕХНИКИ
L-аминокислоты получают промышленным способом посредством ферментации с использованием микроорганизмов, относящихся к роду Brevibacterium, Corynebacterium, Escherichia или тому подобных. В таких способах получения используют штаммы, которые выделяют из природы, или искусственно полученные варианты таких штаммов. Кроме того, могут быть использованы штаммы микроорганизмов, которые модифицированы способом, основанным на рекомбинации ДНК, так чтобы увеличить активность основного фермента биосинтеза L-аминокислоты, и т.д. (патентные документы 1-9).
Когда аминокислоты получают с использованием микроорганизмов, обычно используют сахара в качестве основного компонента субстрата, но также можно использовать глицерин в качестве субстрата, подобного сахарам (патентные документы 10 и 11).
Известно, что Escherichia coli имеет множество генов, которые принимают участие в метаболизме глицерина. Однако было выявлено, что мутантный штамм, дефицитный по glpK, который представляет собой ген, кодирующий глицеринкиназу, или glpD, который представляет собой ген, кодирующий глицерин-3-фосфатдегидрогеназу, не может расти на среде, когда глицерин является единственным источником углерода, основной путь ассимиляции глицерина у E. coli включает в себя глицеринкиназу и глицерин-3-фосфатдегидрогеназу (непатентный документ 1).
Известно, что глицериндегидрогеназа E. coli также является одним из ферментов, которые принимают участие в метаболизме глицерина, и указанный фермент спасает мутантный штамм, дефицитный по трем генам glpK, glpD и glpR, который является геном репрессора регулона glp, от гибели в среде, содержащей глицерин в качестве единственного источника углерода при скрининге с использованием такого штамма (непатентный документ 2).
Считается, что путь через глицерин-3-фосфат, в который вовлечены глицеринкиназа и глицерин-3-фосфатдегидрогеназа, является основным путем ассимиляции глицерина у микроорганизмов, относящихся к семейству Enterobacteriaceae, которые описаны выше, а путь ассимиляции глицерина через дигидроксиацетон является путем, не нужным для ассимиляции глицерина у микроорганизмов, относящихся к семейству Enterobacteriaceae.
Патентный документ 1: EP 0643135 B
Патентный документ 2: EP 0733712 B
Патентный документ 3: EP 1477565 A
Патентный документ 4: EP 0796912 A
Патентный документ 5: EP 0837134 A
Патентный документ 6: WO01/53459
Патентный документ 7: EP 1170376 A
Патентный документ 8: WO2005/010175
Патентный документ 9: WO96/17930
Патентный документ 10: EP 1715055 A
Патентный документ 11: EP 1715056 A
Непатентный документ 1: J. Bacteriol., 23 (2006) 8259-8271
Непатентный документ 2: J. Bacteriol., 131 (1977) 1026-1028
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Цель изобретения
Одним аспектом настоящего изобретения является способ получения L-аминокислоты ферментацией с использованием субстрата, содержащего глицерин, который представляет собой улучшенный способ по сравнению с традиционными способами.
Средства достижения цели
Для достижения указанной выше цели авторы настоящего изобретения тщательно проводили различные исследования. В результате было обнаружено, что усиление либо глицериндегидрогеназы, либо дигидроксиацетонкиназы, которые являются ферментами пути ассимиляции глицерина через дигидроксиацетон, не влияло на продукцию L-аминокислот из глицерина. Однако усиление обоих ферментов, глицериндегидрогеназы и дигидроксиацетонкиназы, заметно повышало продукцию L-аминокислот из глицерина, что позволило осуществить настоящее изобретение.
Таким образом, в настоящем изобретении предлагается следующее:
(1) Способ получения L-аминокислоты, включающий в себя:
культивирование микроорганизма, относящегося к семейству Enterobacteriaceae, обладающего способностью продуцировать L-аминокислоты и модифицированного так, чтобы повысить активности глицериндегидрогеназы и дигидроксиацетонкиназы, в среде, содержащей глицерин в качестве источника углерода, для продуцирования и накопления L-аминокислоты в среде или в клетках, и
сбор L-аминокислоты из среды или клеток.
(2) Способ по п. (1), в котором активности глицериндегидрогеназы и дигидроксиацетонкиназы повышены в результате увеличения количества копий генов, кодирующих глицериндегидрогеназу и дигидроксиацетонкиназу, или в результате модификации последовательностей регуляции экспрессии генов.
(3) Способ по п. (1) или (2), в котором дигидроксиацетонкиназа использует АТФ в качестве донора фосфата.
(4) Способ по любому из пп. (1)-(3), в котором микроорганизм дополнительно модифицирован, чтобы повысить активность поглощения глицерина.
(5) Способ по любому из пп. (1)-(4), в котором микроорганизм дополнительно модифицирован, чтобы повысить активность или активности одного или нескольких ферментов, выбранных из группы, состоящей из триозофосфатизомеразы, фруктозобисфосфатальдолазы, фруктозо-1,6-бисфосфатазы и фруктозо-6-фосфатальдолазы.
(6) Способ по любому из пп. (1)-(5), в котором микроорганизм дополнительно модифицирован, чтобы уменьшить активность или активности глицеринкиназы и/или глицерин-3-фосфатдегидрогеназы связанного с мембраной типа.
(7) Способ по любому из пп. (1)-(6), в котором микроорганизм, относящийся к семейству Enterobacteriaceae, представляет собой бактерию Escherichia или бактерию Pantoea.
(8) Способ по любому из пп. (1)-(7), в котором L-аминокислота выбрана из группы, состоящей из L-глутаминовой кислоты, L-лизина, L-лейцина, L-изолейцина, L-валина, L-триптофана, L-фенилаланина, L-тирозина, L-треонина, L-метионина, L-цистеина, L-аргинина, L-серина, L-пролина, L-аспарагиновой кислоты, L-аспарагина, L-глутамина и L-гистидина.
Наилучший способ осуществления изобретения
Далее настоящее изобретение будет объяснено подробно.
Микроорганизм согласно настоящему изобретению
Примеры микроорганизмов согласно настоящему изобретению включают микроорганизм, относящийся к семейству Enterobacteriaceae, который обладает способностью продуцировать L-аминокислоту и модифицирован, чтобы повысить активности глицериндегидрогеназы и дигидроксиацетонкиназы. Способность продуцировать L-аминокислоту (продуцирующая L-аминокислоту способность) может означать способность приведенных в качестве примера микроорганизмов согласно настоящему изобретению продуцировать и накапливать L-аминокислоту в среде или клетках при культивировании в среде. Типичный микроорганизм согласно настоящему изобретению может обладать способностью продуцировать два или более видов L-аминокислот. Хотя микроорганизм, обладающий способностью продуцировать L-аминокислоту, может обладать наследственным свойством продуцировать L-аминокислоту, такой микроорганизм может быть получен в результате модификации таких микроорганизмов, которые указаны ниже, с использованием методики рекомбинантной ДНК так, чтобы они обладали способностью продуцировать L-аминокислоту.
Хотя тип L-аминокислоты особо не ограничен, примерами являются основные аминокислоты, такие как L-лизин, L-орнитин, L-аргинин, L-гистидин и L-цитруллин, алифатические аминокислоты, такие как L-изолейцин, L-аланин, L-валин, L-лейцин и L-глицин, аминокислоты, которые являются гидроксимоноаминокарбоновыми кислотами, такие как L-треонин и L-серин, циклические аминокислоты, такие как L-пролин, ароматические аминокислоты, такие как L-фенилаланин, L-тирозин и L-триптофан, серосодержащие аминокислоты, такие как L-цистеин, L-цистин и L-метионин, кислые аминокислоты, такие как L-глутаминовая кислота и L-аспарагиновая кислота, и аминокислоты с амидной группой в боковой цепи, такие как L-глутамин и L-аспарагин. Типичный микроорганизм согласно настоящему изобретению может обладать способностью продуцировать два или более видов L-аминокислот.
Микроорганизмы, относящиеся к семейству Enterobacteriaceae, включают бактерии Escherichia и бактерии Pantoea. Другие примеры микроорганизмов, относящихся к семейству Enterobacteriaceae, включают микроорганизмы, относящиеся к γ-протеобактериям, такие как бактерии рода Enterobacter, Klebsiella, Serratia, Erwinia, Salmonella, Morganella или тому подобные.
В настоящем изобретении термин «глицериндегидрогеназа» означает фермент, который обратимо катализирует следующую реакцию окисления, которая превращает глицерин в дигидроксиацетон с использованием NAD в качестве кофермента (EC:1.1.1.6).
Глицерин+NAD=дигидроксиацетон+NADH+H+
В настоящем изобретении фраза «модифицирован, чтобы повысить активность глицериндегидрогеназы» может означать, что количество молекул глицериндегидрогеназы в клетке может быть увеличено по сравнению с количеством в штамме дикого типа или в немодифицированном штамме, или что активность глицериндегидрогеназы на молекулу может быть повышена по сравнению с активностью в штамме дикого типа или немодифицированном штамме. Кроме того, когда активность фермента не регистрируется в штамме дикого типа и ее повышают до регистрируемого уровня, такое состояние также включено в определение состояния «повышения активности». В настоящем изобретении активность глицериндегидрогеназы может быть на любом уровне, при условии, что она может быть выявлена, но модификацию предпочтительно осуществляют так, чтобы активность глицериндегидрогеназы составляла 0,05 ед./мг или выше, предпочтительно 0,25 ед./мг или выше, более предпочтительно 0,5 ед./мг или выше. Примеры штаммов дикого типа микроорганизма, относящегося к семейству Enterobacteriaceae, которые могут служить в качестве эталона для сравнения, включают штамм MG1655 (ATCC No. 47076) и штамм W3110 (ATCC No. 27325) Escherichia coli, штамм AJ13335 Pantoea ananatis (FERM BP-6615), и другие. Активность глицериндегидрогеназы может быть измерена согласно способу Ansis, R.E. et al. (J. Biol. Chem., 2-3, 153-159 (1953)).
В настоящем изобретении «дигидроксиацетонкиназа» представляет собой фермент, который обратимо катализирует следующую реакцию превращения дигидроксиацетона в дигидроксиацетонфосфат и использует АТФ в качестве донора фосфата (EC 2.7.1.29), и фермент, использующий PEP в качестве донора фосфата (EC 2.7.1.29) (Cell. Mol. Life Sci., 63 (2006) 890-900; Biochemistry, 43 (2004) 1337-13045):
АТФ+дигидроксиацетон=АДФ+дигидроксиацетонфосфат (EC 2.7.1.29)
Фосфоенолпируват+дигидроксиацетон=пируват+дигидроксиацетон-фосфат (EC2.7.1.29)
В настоящем изобретении особенно предпочтительно, чтобы дигидроксиацетонкиназа использовала АТФ в качестве донора фосфата.
Фраза «модифицирован, чтобы повысить активность дигидроксиацетонкиназы» может означать, что количество молекул дигидроксиацетонкиназы в клетке увеличено по сравнению с количеством в штамме дикого типа или в немодифицированном штамме или что активность дигидроксиацетонкиназы на молекулу может быть повышена по сравнению с активностью в штамме дикого типа или немодифицированном штамме. Модификацию предпочтительно осуществляют так, чтобы активность дигидроксиацетонкиназы на клетку была повышена до 150% или выше, более предпочтительно до 200% или выше, еще более предпочтительно до 300% или выше по сравнению с активностью штамма дикого типа или немодифицированного штамма. Примеры штаммов дикого типа микроорганизма, относящегося к семейству Enterobacteriaceae, которые могут служить в качестве эталона для сравнения, включают штамм MG1655 (ATCC No. 47076) и штамм W3110 (ATCC No. 27325) Escherichia coli, штамм AJ13335 Pantoea ananatis (FERM BP-6615) и другие. Активность дигидроксиацетонкиназы может быть измерена способом Johnson E.A. (J. Bacteriol., 1984 Oct; 160(1): 55-60).
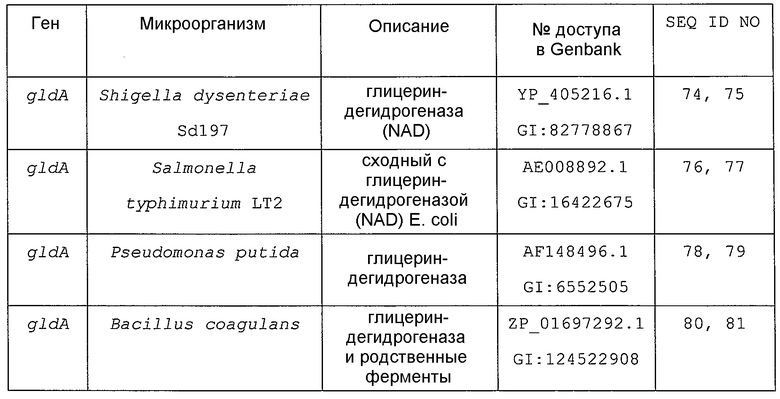
Примеры гена, кодирующего глицериндегидрогеназу, включают ген gldA, и предпочтительным примером является ген gldA, полученного из микроорганизма, относящегося к семейству Enterobacteriaceae. Примеры микроорганизма, относящегося к семейству Enterobacteriaceae, включают Escherichia coli. Примером гена Escherichia coli является, например, ген gldA с последовательностью SEQ ID NO: 1 (нить, комплементарная нуклеотидам с номерами 4135955..4137058 в GenBank с номером доступа NC_000913).
Кроме того, гомологами гена, кодирующего глицериндегидрогеназу, могут быть гены, клонированные на основе гомологии с приведенным выше в качестве примера геном из бактерии рода Escherichia, Enterobacter, Klebsiella, Serratia, Erwinia, Yersinia, Shigella, Salmonella, Vibrio, Aeromonas, Bacillus, Staphylococcus, Lactobacillus, Enterococcus, Clostridium, Pseudomonas, Agrobacterium, Citrobacter, Corynebacterium или тому подобной. Примеры генов, которые обладают высокой степенью гомологии с геном gldA Escherichia coli и могут быть использованы в настоящем изобретении в качестве гена, кодирующего глицериндегидрогеназу, приведены в таблице 1.
Гены, обладающие высокой степенью гомологии с геном gldA Escherichia coli и кодирующие глицериндегидрогеназу
Гомология (идентичность и т.д.) аминокислотных последовательностей и нуклеотидных последовательностей может быть определена, например, с использованием алгоритма BLAST согласно Karlin and Altschul (Pro. Natl. Acad. Sci. USA, 90, 5873 (1993)) или FASTA согласно Pearson (Methods Enzymol., 183, 63 (1990)). Программы, названные BLASTN и BLASTX, были разработаны на основе указанного алгоритма BLAST (смотри www.ncbi.nlm.nih.gov).
В качестве гена, кодирующего дигидроксиацетонкиназу согласно настоящему изобретению, можно использовать гены, названные геном dhaKLM, геном dak1, геном dhaK и геном dhbK. Примерами гена, кодирующего фермент, использующий PEP в качестве донора фосфата, являются гены, полученные из Escherichia coli, такие как ген dhaK с последовательностью SEQ ID NO: 34 (нить, комплементарная нуклеотидам с номерами 1248991..1250061 в GenBank с номером доступа No. NC_000913), ген dhaL с последовательностью SEQ ID NO: 36 (нить, комплементарная нуклеотидам с номерами 1248348..1248980 в GenBank с No. доступа NC_000913), и ген dhaM с последовательностью SEQ ID NO: 38 (нить, комплементарная нуклеотидам с номерами 1246919..1248337 в GenBank с No. доступа NC_000913).
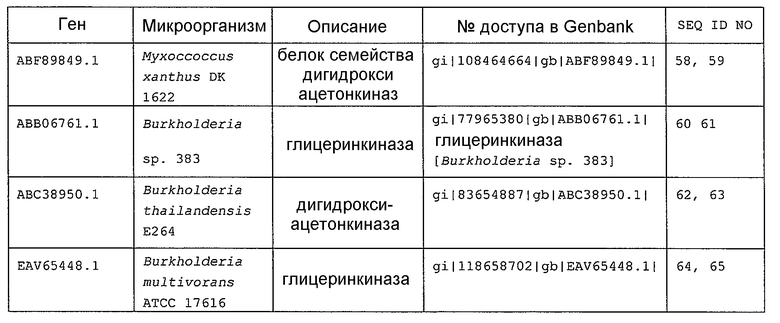
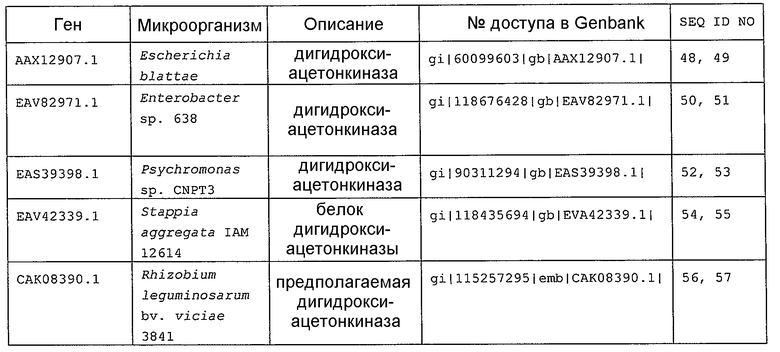
В настоящем изобретении предпочтительно может быть использован ген, кодирующий дигидроксиацетонкиназу, которая использует АТФ в качестве донора фосфата, и таким геном может быть ген dak1, полученный из дрожжей, ген dhbK, полученный из бактерий Agrobacterium, и ген dhaK, полученный из бактерий Citrobacter. Примером гена dak1, полученного из дрожжей, является ген dak1 с последовательностью SEQ ID NO: 3, полученный из Saccharomyces cerevisiae (No. доступа в GenBank NP_013641.1 GI: 6323570), примером гена dhbK, полученного из бактерий Agrobacterium, является ген dhbK с последовательностью SEQ ID NO: 5, полученный из Agrobacterium tumefaciens (No. доступа в GenBank. NP_357070.1 GI: 15891398), и примером гена dhaK, полученного из бактерий Citrobacter, является ген dhaK с последовательностью SEQ ID NO: 7, полученный из Citrobacter freundii (No. доступа в GenBank. U09771).
Кроме того, гомологи гена, кодирующего дигидроксиацетонкиназу, могут представлять собой гены, клонированные на основе гомологии с геном, приведенным в качестве примера выше, из бактерии, такой как бактерии рода Escherichia, Enterobacter, Klebsiella, Serratia, Erwinia, Yersinia, Shigella, Salmonella, Vibrio, Aeromonas, Bacillus, Staphylococcus, Lactobacillus, Enterococcus, Clostridium, Agrobacterium, Citrobacter и Mycobacterium, дрожжей, таких как дрожжи рода Saccharomyces, Schizosaccharomyces или Pichia, или тому подобных.
В частности, в качестве гена, кодирующего дигидроксиацетонкиназу, которая использует АТФ в качестве донора фосфата, можно применять следующие последовательности. Гены, кодирующие дигидроксиацетонкиназу и обладающие высокой степенью гомологии с геном dak1, полученным из Saccharomyces cerevisiae, показаны в таблице 2, гены дигидроксиацетонкиназы, обладающие высокой степенью гомологии с геном dhbK, полученным из Agrobacterium tumefaciens, показаны в таблице 3, и гены дигидроксиацетонкиназы, обладающие высокой степенью гомологии с геном dhaK, полученным из Citrobacter freundii, показаны в таблице 4.
Гены, кодирующие дигидроксиацетонкиназу и обладающие высокой степенью гомологии с геном dak1, полученным из Saccharomyces cerevisiae

Гены, кодирующие дигидроксиацетонкиназу и обладающие высокой степенью гомологии с геном dhbK, полученным из Agrobacterium tumefaciens
Гены, кодирующие дигидроксиацетонкиназу и обладающие высокой степенью гомологии с геном dhaK, полученным из Citrobacter freundii
Гомологи указанных выше генов могут представлять собой мутантные гены, полученные из других микроорганизмов, или природные или искусственные мутантные гены, которые имеют высокое структурное сходство с указанными выше генами и которые могут повышать активность глицериндегидрогеназы и активность дигидроксиацетонкиназы при введении в хозяина или при амплификации. Гомологи генов глицериндегидрогеназы и дигидроксиацетонкиназы могут представлять собой гены, кодирующие белок, имеющий гомологию 80% или выше, предпочтительно 90% или выше, более предпочтительно 95% или выше, особенно предпочтительно 98% или выше, с общей аминокислотной последовательностью SEQ ID NO: 2, 4, 6 или 8 или любой из аминокислотных последовательностей, кодируемых последовательностями, указанными в таблицах 1-4, и обладающими функцией глицериндегидрогеназы или дигидроксиацетонкиназы. Кодирует ли ген белок, обладающий активностью глицериндегидрогеназы или активностью дигидроксиацетонкиназы, можно подтвердить посредством экспрессии гена в клетке-хозяине и определения того, повышена ли ферментативная активность по сравнению с немодифицированным штаммом, согласно указанному выше способу измерения ферментативной активности. Кроме того, тот факт, является ли ген гомологом или не является, можно подтвердить посредством получения штамма с нарушенным геном, в котором соответствующий ген дикого типа нарушен, введения гена в штамм с нарушением и исследования того, комплементирует ли ген функцию гена дикого типа, например, восстанавливается ли ферментативная активность, сниженная в результате нарушения гена.
Кроме того, гены, кодирующие глицериндегидрогеназу и дигидроксиацетонкиназу, используемые в настоящем изобретении, не ограничены генами дикого типа, и гены могут представлять собой мутантные или искусственно модифицированные гены, кодирующие белок, имеющий аминокислотную последовательность SEQ ID NO: 2, 4, 6 или 8 или любую из аминокислотных последовательностей, указанных в таблице 1-4, которые могут включать в себя замену, делецию, инсерцию, добавление или тому подобное одного или нескольких аминокислотных остатков в одном или нескольких положениях, при условии, что функция кодируемой глицериндегидрогеназы или дигидроксиацетонкиназы не снижена. Хотя количество, определяемое как «один или несколько» аминокислотных остатков, может отличаться в зависимости от положений в трехмерной структуре или типов аминокислотных остатков белка, такое количество может составлять, в частности, от 1 до 20, предпочтительно от 1 до 10, более предпочтительно от 1 до 5, еще более предпочтительно от 1 до 3. Указанные замены предпочтительно являются консервативными заменами. Консервативная замена представляет собой мутацию, при которой замена происходит взаимно между аминокислотами Phe, Trp и Tyr в том случае, если местом замены является ароматическая аминокислота; между Leu, Ile и Val, если аминокислота представляет собой гидрофобную аминокислоту; между Gln и Asn, если аминокислота представляет собой полярную аминокислоту; между Lys, Arg и His, если аминокислота является основной аминокислотой; между Asp и Glu, если аминокислота является кислой аминокислотой; и между Ser и Thr, если аминокислота представляет собой аминокислоту, имеющую гидроксильную группу. Примерами консервативной замены являются замена Ala на Ser или Thr, замена Arg на Gln, His или Lys, замена Asn на Glu, Gln, Lys, His или Asp, замена Asp на Asn, Glu или Gln, замена Cys на Ser или Ala, замена Gln на Asn, Glu, Lys, His, Asp или Arg, замена Glu на Gly, Asn, Gln, Lys или Asp, замена Gly на Pro, замена His на Asn, Lys, Gln, Arg или Tyr, замена Ile на Leu, Met, Val или Phe, замена Leu на Ile, Met, Val или Phe, замена Lys на Asn, Glu, Gln, His или Arg, замена Met на Ile, Leu, Val или Phe, замена Phe на Trp, Tyr, Met, Ile или Leu, замена Ser на Thr или Ala, замена Thr на Ser или Ala, замена Trp на Phe или Tyr, замена Tyr на His, Phe или Trp и замена Val на Met, Ile или Leu. Вышеуказанная аминокислотная замена, делеция, инсерция, добавление, инверсия или тому подобное может быть результатом встречающейся в природе мутации вследствие индивидуального отличия или видового отличия (мутант или вариант) микроорганизма, имеющего гены, кодирующие глицериндегидрогеназу и дигидроксиацетонкиназу.
Гены, кодирующие глицериндегидрогеназу и дигидроксиацетонкиназу, также могут представлять собой ДНК, которая способна гибридизоваться с последовательностью, комплементарной нуклеотидной последовательности SEQ ID NO: 2, 4, 6 или 8 или любой из нуклеотидных последовательностей, указанных в таблицах 1-4, или зондом, который может быть получен из нуклеотидных последовательностей, в жестких условиях и кодирует белок, обладающий глицериндегидрогеназной активностью или дигидроксиацетонкиназной активностью. «Жесткие условия» представляют собой условия, при которых образуется так называемый специфичный гибрид и не образуется неспецифичный гибрид. Хотя трудно четко определить условие числовыми значениями, примеры жестких условий включают условия, при которых высокогомологичные ДНК гибридизуются друг с другом, например, ДНК, гомологичные не менее чем на 80%, предпочтительно гомологичные не менее чем на 90%, более предпочтительно гомологичные не менее чем на 95%, особенно предпочтительно гомологичные не менее чем на 98%, гибридизуются друг с другом, а DДНК с меньшей гомологией, чем указанная выше, не гибридизуются друг с другом, или условия промывки для типичной Саузерн-гибридизации, т.е. промывка один раз, предпочтительно 2 или 3 раза при концентрации соли и температуре, соответствующей 1×SSC, 0,1% SDS при 60°C, предпочтительно 0,1×SSC, 0,1% SDS при 60°C, более предпочтительно 0,1×SSC, 0,1% SDS при 68°C.
В настоящем изобретении фраза «внутриклеточная активность фермента возрастает» означает состояние, при котором внутриклеточная активность фермента повышена по сравнению со штаммом дикого типа (например, штаммами W3110 и MG1655 Escherichia coli) или родительским штаммом (штаммом, в котором внутриклеточные активности всех ферментов, указанных в настоящем изобретении, не повышены), а также включает состояние, при котором клетки обладают активностью, которой штамм дикого типа или родительский штамм не обладают.
Примеры способов повышения внутриклеточной активности включают указанные ниже способы и их сочетания. Однако способы не ограничены указанным. В качестве способов повышения активностей глицериндегидрогеназы и дигидроксиацетонкиназы можно использовать любой из способов (1)-(5), и можно применять один и тот же или разные способы.
(1) Увеличение количества копий гена, кодирующего каждый белок, в результате трансформации с использованием вектора, содержащего ген.
(2) Увеличение количества копий гена, кодирующего каждый белок, в результате интеграции гена в хромосому.
(3) Повышение уровня экспрессии гена, кодирующего каждый белок, посредством модификации области регуляции экспрессии гена.
(4) Повышение уровня экспрессии посредством модификации фактора, который влияет на регуляцию экспрессии.
(5) Увеличение ферментативной активности посредством введения мутации в кодирующую область гена, кодирующего каждый белок.
(6) Увеличение количества белка в результате повышения эффективности трансляции.
Далее гены, кодирующие глицериндегидрогеназу и дигидроксиацетонкиназу, могут быть названы целевыми генами.
(1) Увеличение количества копий гена, кодирующего каждый белок, посредством трансформации с использованием вектора, содержащего ген
Например, фрагмент ДНК, содержащий целевой ген, может быть лигирован в вектор, который функционирует в микроорганизме-хозяине, предпочтительно в вектор многокопийного типа, чтобы получить рекомбинантную ДНК, и рекомбинантная ДНК может быть введена в микроорганизм, чтобы его трансформировать. Целевой ген может быть получен в ПЦР (полимеразной цепной реакции, см. White, T.J. et al., Trends Genet., 5, 185 (1989)) с использованием в качестве матрицы хромосомной ДНК Escherichia coli, дрожжей, бактерии Citrobacter, бактерии Agrobacterium или тому подобной. Целевые гены из других микроорганизмов также можно получить из хромосомной ДНК или библиотеки хромосомной ДНК любого микроорганизма с помощью ПЦР, используя в качестве праймеров олигонуклеотиды, полученные на основе известного целевого гена микроорганизма или на основе информации о последовательности целевого гена или белка микроорганизма другого вида, или с помощью гибридизации с использованием в качестве зонда олигонуклеотида, полученного на основе информации о такой последовательности, которая указана выше. Хромосомная ДНК может быть получена из микроорганизма, который служит в качестве донора ДНК, способом, описанным Saito and Miura (см. Saito H. and Miura K., Biochem. Biophys. Acta, 72, 619 (1963); Experimental Manual for Biotechnology, edited by The Society for Biotechnology, Japan, pp.97-98, Baifukan Co., Ltd., 1992) или тому подобным.
Затем целевой ген, амплифицированный в ПЦР, может быть лигирован в векторную ДНК, которая может функционировать в клетке микроорганизма-хозяина, чтобы получить рекомбинантную ДНК. Примерами вектора, который может функционировать в клетке микроорганизма-хозяина, являются векторы, которые автономно реплицируются в клетках микроорганизма-хозяина.
Примерами векторов, которые автономно реплицируются в микроорганизмах, относящихся к семейству Enterobacteriaceae, являются pUC19, pUC18, pHSG299, pHSG399, pHSG398, pACYC184, (векторы серий pHSG и pACYC доступны из Takara Bio), RSF1010 (Gene, vol. 75(2), p. 271-288, 1989), pBR322, pMW219, pMW119 (векторы серий pMW доступны из Nippon Gene), pSTV28, pSTV29 (Takara Bio) и т.д. Можно использовать вектор на основе фаговой ДНК.
Чтобы получить рекомбинантную ДНК лигированием какого-либо гена с указанным выше вектором конструируют вектор так, чтобы рестрикционный фермент соответствовал концам фрагмента ДНК, содержащего целевой ген. Лигирование обычно осуществляют, используя лигазу, такую как ДНК-лигаза T4. В качестве способов расщепления и лигирования ДНК можно применять способы получения хромосомной ДНК, ПЦР, получения плазмидной ДНК, трансформации, конструирования олигонуклеотидов, используемых в качестве праймеров, и т.д., которые хорошо известны специалисту в данной области. Указанные способы описаны в Sambrook, J., Fritsch, E.F., and Maniatis, T., "Molecular Cloning A Laboratory Manual, Second Edition", Cold Sprig Harbor Laboratory Press, (1989), и других публикациях.
Рекомбинантная ДНК, полученная, как описано выше, может быть введена в бактерию согласно обычному известному способу трансформации. Примеры включают электропорацию (Canadian Journal of Microbiology, 43, 197 (1997)). Также можно применять способ увеличения проникающей способности ДНК обработкой реципиентных клеток хлоридом кальция, как описано для Escherichia coli K-12 (Mandel, M. and Higa, A., J. Mol. Biol., 53, 159 (1970), или способ введения ДНК в компетентную клетку, полученную из клетки на стадии пролиферации, как описано для Bacillus subtilis (Duncan, C.H., Wilson, G.A and Young, F.E, Gene, 1, 153 (1977)).
(2) Увеличение количества копий гена, кодирующего каждый белок, в результате интеграции гена в хромосому
Увеличение внутриклеточной активности каждого фермента может быть достигнуто в результате увеличения количества копий целевого гена посредством введения целевого гена в хромосомную ДНК микроорганизма. Введение целевого гена в хромосомную ДНК микроорганизма можно осуществить с помощью гомологичной рекомбинации, используя последовательность-мишень, присутствующую в хромосомной ДНК в многочисленных копиях. Так как такая последовательность присутствует в хромосомной ДНК в многочисленных копиях, то можно использовать повторяющуюся ДНК или инвертированный повтор, имеющийся на концах транспозируемого элемента. Альтернативно, как описано в выложенном патенте Японии (Kokai) No. 2-109985, целевой ген может быть введен в хромосомную ДНК путем встраивания гена в транспозон и его переноса так, чтобы ген был интегрирован в хромосомную ДНК. Кроме того, также можно ввести целевой ген в хромосому, используя способ зависимой от Red интеграции (WO2005/010175). Целевой ген также может быть введен в хромосому посредством трансдукции с использованием фага, такого как фаг P1, или с использованием вектора для конъюгативного переноса. Перенос гена в хромосому может быть подтвержден при осуществлении Саузерн-гибридизации с использованием части гена в качестве зонда. Увеличение количества копий может быть подтверждено Саузерн-гибридизацией с использованием зонда, комплементарного целевому гену. Хотя количество копий может быть увеличено в любой степени, при условии, что оно увеличено на одну или несколько копий, ген, кодирующий глицериндегидрогеназу, предпочтительно амплифицируют на две или более копий, более предпочтительно на три или более копий, еще более предпочтительно на пять или более копий, а ген, кодирующий дигидроксиацетонкиназу, предпочтительно амплифицируют на две или более копий, более предпочтительно на три или более копий, еще более предпочтительно на пять или более копий. В том случае, когда вводят ген, который не является нативным для выбранного микроорганизма-хозяина, может быть введено любое количество копий, при условии, что вводят одну или несколько копий.
(3) Повышение уровня экспрессии гена, кодирующего каждый белок, посредством модификации области регуляции экспрессии гена
Кроме того, помимо увеличения количества копий целевого гена, описанного выше, может быть достигнуто повышение внутриклеточной активности каждого фермента, посредством замены последовательности регуляции экспрессии, такой как промотор гена, в хромосомной ДНК или в плазмиде более сильным промотором способом, описанным в WO 00/18935. В качестве сильных промоторов известны, например, промотор lac, промотор trp, промотор trc, промотор PR фага лямбда, промотор PL, промотор lpp, промотор T7, промотор tet и другие. В настоящем изобретении, в частности, для того, чтобы амплифицировать глицериндегидрогеназу, предпочтительно используют промотор tacM (SEQ ID NO: 10). dhaK, dhaL и dhaM, кодирующие дигидроксиацетонкиназу Escherichia coli, составляют структуру оперона, и уровни экспрессии всех трех генов повышают, усиливая промотор, расположенный перед dhaK.
Кроме того, также возможно введение нуклеотидной замены или тому подобного в область промотора целевого гена, чтобы модифицировать его с получением более сильного промотора. Способы оценки силы промоторов и примеры сильных промоторов описаны в публикации Goldstein et al. (Prokaryotic promoters in biotechnology, Biotechnol. Annu. Rev., 1995, 1, 105-128) и других публикациях. Кроме того, известно, что замена нескольких нуклеотидов в области спейсера между сайтом связывания рибосом (RBS) и стартовым кодоном, в частности, в области, расположенной непосредственно перед стартовым кодоном, существенно влияет на эффективность трансляции мРНК, и такая область также может быть модифицирована. Экспрессия целевого гена усиливается при такой замене или модификации промотора.
Что касается замены промотора в хромосоме более сильным промотором, то промотор, расположенный перед целевым геном в геноме, может быть заменен более сильным промотором посредством трансформации микроорганизма, относящегося к семейству Enterobacteriaceae, ДНК, содержащей более сильный промотор, амплифицированный в ПЦР или т.п., чтобы вызвать рекомбинацию более сильного промотора и промотора дикого типа в геноме. Для такой замены в гене с использованием гомологичной рекомбинации можно применять способ, называемый зависимой от Red интеграцией (Datsenko, K.A, and Wanner, B.L., Proc. Natl. Acad. Sci. USA, 97: 6640-6645 (2000)), способ, основанный на использовании линейной ДНК, такой как способ с использованием зависимой от Red интеграции в сочетании с системой разрезания, полученной из фага λ (Cho, E.H., Gumport, R.I., Gardner, J.F., J. Bacteriol., 184: 5200-5203 (2002)) (см. WO2005/010175), способ с применением плазмиды, содержащей чувствительное к температуре начало репликации (Datsenko, K.A, and Wanner, B.L., Proc. Natl. Acad. Sci. USA, 97: 6640-6645 (2000), патента США No. 6303383, выложенный патент Японии No. 05-007491), и т.д.
(4) Повышение уровня экспрессии посредством модификации фактора, который влияет на регуляцию экспрессии
Повышение уровня экспрессии посредством модификации фактора, который влияет на регуляцию экспрессии, можно осуществить в результате амплификации гена, кодирующего активатор, который увеличивает экспрессию генов, кодирующих глицериндегидрогеназу и дигидроксиацетонкиназу, или в результате делеции или ослабления гена, кодирующего регулятор, который снижает экспрессию генов. Примером активатора dhaKLM, кодирующего дигидроксиацетонкиназу, является, например, dhaR (SEQ ID NO: 66, номера нуклеотидов 1250289..1252208 в GenBank с No. доступа NC_000913), и уровень экспрессии dhaKLM, кодирующего дигидроксиацетонкиназу, увеличивается при мутации гена dhaR (1: EMBO J., 2005 Jan. 26, 24(2): 283-93). Уровень экспрессии dhaKLM, кодирующего дигидроксиацетонкиназу, также увеличивают в результате нарушения гена ptsI (SEQ ID NO: 86, номера нуклеотидов 2532088..2533815 в GenBank с No. доступа NC_000913) (Microbiology, 147 (2001) 247-253).
(5) Увеличение ферментативной активности посредством введения мутации в кодирующую область гена, кодирующего каждый белок
Кроме того, увеличение активностей глицериндегидрогеназы и дигидроксиацетонкиназы также можно достичь в результате введения мутации, которая увеличивает удельные активности белков или улучшает субстратные специфичности ферментов в кодирующих областях целевых генов.
Такой ген, кодирующий какой-либо фермент, имеющий мутацию, может быть получен, например, посредством модификации нуклеотидной последовательности SEQ ID NO: 1, 3, 5 или 7, или кодирующей области в любой из нуклеотидных последовательностей, указанных в таблицах 1-4, так чтобы аминокислотные остатки конкретной части кодируемого белка включали в себя замену, делецию, инсерцию, добавление аминокислотных остатков или тому подобное. Кроме того, такой ген также может быть получен общеизвестными мутагенными обработками, описанными ниже. Что касается мутагенных обработок, то способом обработки нуклеотидной последовательности SEQ ID NO: 1, 3, 5 или 7, любой из нуклеотидных последовательностей, указанных в таблицах 1-4, или последовательности кодирующей области в любой из таких последовательностей гидроксиламином или подобным веществом in vitro, способом обработки микроорганизма, такого как микроорганизм, относящийся к семейству Enterobacteriaceae, содержащего ген, ультрафиолетовым излучением или мутагенным средством, используемым для обычных мутагенных обработок, таким как N-метил-N'-нитро-N-нитрозогуанидин (NTG) или этилметансульфонат (EMS), посредством склонной к ошибкам ПЦР (Cadwell, R.C., PCR Meth. Appl., 2, 28 (1992)), перетасовкой ДНК (Stemmer, W.P., Nature, 370, 389 (1994)), или StEP-ПЦР (Zhao, H., Nature Biotechnol., 16, 258 (1998)), мутация может быть искусственно введена в гены, кодирующие глицериндегидрогеназу и дигидроксиацетонкиназу, посредством рекомбинации генов, чтобы получить гены, кодирующие высокоактивные глицериндегидрогеназу и дигидроксиацетонкиназу. Кодируют ли такие мутантные ферменты глицериндегидрогеназу и дигидроксиацетонкиназу, можно подтвердить, например, введением генов в микроорганизм, относящийся к семейству Enterobacteriaceae и обладающий способностью продуцировать L-аминокислоту, его культивированием в среде, содержащей глицерин в качестве источника углерода, и определением того, повышена ли способность продуцировать L-аминокислоту, или измерением активностей ферментов указанными выше способами.
(6) Увеличение количества белка в результате повышения эффективности трансляции
Увеличение количества белка в результате повышения эффективности трансляции можно осуществить в результате увеличения количества тРНК, соответствующей кодонам, менее часто используемым в хозяине, или в результате модификации целевого гена так, чтобы он имел оптимальные кодоны в отношении частоты использования кодонов у хозяина (Gene 85, 109-114 (1989), Biochemistry, 31, 2598-2608 (1992), J. Bacteriol., 175, 716-722 (1993), Protein Expression and Purification, 50, 49-57 (2006)). Увеличение количества целевого белка по сравнению с немодифицированным штаммом или штаммом дикого типа можно подтвердить, например, регистрацией при Вестерн-блоттинге с использованием антител (Molecular Cloning (Cold Spring Harbor Laboratory Press, Cold spring Harbor (USA), 2001)).
Микроорганизм, используемый в способе получения согласно настоящему изобретению, может представлять собой микроорганизм, модифицированный так, чтобы повысить активность поглощения глицерина в добавление к усилению активности глицериндегидрогеназы и дигидроксиацетонкиназы. Активность поглощения глицерина означает активность включения глицерина в цитоплазму, и в указанный процесс также вовлечен посредник поглощения глицерина, который представляет собой мембранный белок. Примером гена, кодирующего посредник глицерина, является, например, ген glpF Escherichia coli (SEQ ID NO: 16, нить, комплементарная нуклеотидам с номерами 4115268..4116113 в GenBank с No. доступа NC_000913).
Ген, кодирующий посредник глицерина, может представлять собой ДНК, которая гибридизуется с последовательностью, комплементарной нуклеотидной последовательности SEQ ID NO: 16, или зондом, который может быть получен из комплементарной последовательности, в жестких условиях и кодирует белок, обладающий активностью в процессе поглощения глицерина. Примеры также включают ДНК, кодирующую белок с последовательностью SEQ ID NO: 17. Белок может представлять собой белок, имеющий гомологию, составляющую 80% или выше, предпочтительно 90% или выше, более предпочтительно 95% или выше, особенно предпочтительно 98% или выше с общей аминокислотной последовательностью SEQ ID NO: 17, при условии, что он повышает способность поглощать глицерин микроорганизмом, относящимся к семейству Enterobacteriaceae, при его введении в микроорганизм.
Кроме того, ген может представлять собой ДНК, кодирующую белок, имеющий аминокислотную последовательность SEQ ID NO: 17, но которая может включать в себя замену, делецию, инсерцию, добавление или тому подобное одного или нескольких аминокислотных остатков, при условии, что активность поглощения глицерина не снижена. Активность может быть увеличена способом, подобным вышеуказанным способам, указанным выше для усиления активности глицериндегидрогеназы и дигидроксиацетонкиназы.
Активность поглощения глицерина может быть измерена с использованием способа анализа транспорта с помощью мембранного белка (Voegele, R.T., Sweet, G.D., and Boos, W.J., Bacteriol., 175: 1087-1094 (1993)).
Микроорганизм, используемый в способе получения согласно настоящему изобретению, может быть модифицирован так, чтобы повысить активности одного или нескольких ферментов, выбранных из группы, состоящей из триозофосфатизомеразы, фруктозобисфосфатальдолазы, фруктозо-1,6-бисфосфатазы и фруктозо-6-фосфатальдолазы, в добавление к усилению глицериндегидрогеназы и дигидроксиацетонкиназы и усилению активности поглощения глицерина.
Триозофосфатизомераза является ферментом, который катализирует реакцию, в которой происходит обратимое превращение дигидроксиацетонфосфата в глицеральдегид-3-фосфат (EC:5.3.1.1).
Дигидроксиацетонфосфат=D-глицеральдегид-3-фосфат
Фраза «модифицирован так, чтобы повысить активность триозофосфатизомеразы» может означать, что количество молекул триозофосфатизомеразы в клетке может быть увеличено по сравнению с количеством в штамме дикого типа или в немодифицированном штамме или что активность триозофосфатизомеразы на молекулу может быть повышено по сравнению с активностью в штамме дикого типа или немодифицированном штамме. Модификацию предпочтительно осуществляют так, чтобы активность триозофосфатизомеразы была повышена до 150% или выше, более предпочтительно до 200% или выше, еще более предпочтительно до 300% или выше по сравнению с активностью штамма дикого типа или немодифицированного штамма. Примеры штаммов дикого типа микроорганизма, относящегося к семейству Enterobacteriaceae, которые могут служить в качестве эталона для сравнения, включают штамм MG1655 (ATCC No. 47076) и штамм W3110 (ATCC No. 27325) Escherichia coli, штамм AJ13335 Pantoea ananatis (FERM BP-6615), и другие.
Примером гена, кодирующего триозофосфатизомеразу, является ген tpiA, полученный из Escherichia coli (SEQ ID NO: 18, нить, комплементарная нуклеотидам с номерами 4108763..4109530 в GenBank с No. доступа NC_000913).
Ген, кодирующий триозофосфатизомеразу, может представлять собой ДНК, которая гибридизуется с последовательностью, комплементарной нуклеотидной последовательности SEQ ID NO: 18, или зондом, который может быть получен из комплементарной последовательности, в жестких условиях и кодирует белок, обладающий активностью триозофосфатизомеразы. Примеры также включают ДНК, кодирующую белок с последовательностью SEQ ID NO: 19. Белок может представлять собой белок, имеющий гомологию, составляющую 80% или выше, предпочтительно 90% или выше, более предпочтительно 95% или выше, особенно предпочтительно 98% или выше с общей аминокислотной последовательностью SEQ ID NO: 19, при условии, что он проявляет повышенную активность триозофосфатизомеразы в микроорганизме, относящемся к семейству Enterobacteriaceae, при его введении в микроорганизм.
Кроме того, ген может представлять собой ДНК, кодирующую белок, имеющий аминокислотную последовательность SEQ ID NO: 19, но который может включать в себя замену, делецию, инсерция, добавление или тому подобного одного или нескольких аминокислотных остатков, при условии, что активность триозофосфатизомеразы не понижена.
Активность триозофосфатизомеразы может быть измерена с использованием способа Andersen and Cooper (FEBS Lett., 4, 19-20 (1969)). Активность может быть повышена способами, сходными со способами, указанными выше для усиления глицериндегидрогеназы и дигидроксиацетонкиназы.
В настоящем изобретении «фруктозобисфосфатальдолаза» означает фермент, который обратимо катализирует следующую реакцию, в которой происходит превращение дигидроксиацетонфосфата и глицеральдегид-3-фосфата в D-фруктозо-1,6-бисфосфат (EC:4.1.2.13).
Дигидроксиацетонфосфат (глицеринфосфат)+D-глицеральдегид-3-фосфат=D-фруктозо-1,6-бисфосфат
Фраза «модифицирован так, чтобы повысить активность фруктозобисфосфатальдолазы» может означать, что количество молекул фруктозобисфосфатальдолазы в клетке может быть увеличено по сравнению с количеством в штамме дикого типа или в немодифицированном штамме или что активность фруктозобисфосфатальдолазы на молекулу может быть повышена по сравнению с активностью в штамме дикого типа или немодифицированном штамме. Модификацию предпочтительно осуществляют так, чтобы активность фруктозобисфосфатальдолазы была повышена до 150% или выше, более предпочтительно до 200% или выше, еще более предпочтительно до 300% или выше по сравнению с активностью штамма дикого типа или немодифицированного штамма. Примеры штаммов дикого типа микроорганизма, относящегося к семейству Enterobacteriaceae, которые могут служить в качестве эталона для сравнения, включают штамм MG1655 (ATCC No. 47076) и штамм W3110 (ATCC No. 27325) Escherichia coli, штамм AJ13335 Pantoea ananatis (FERM BP-6615), и другие.
Примерами гена, кодирующего фруктозобисфосфатальдолазу, являются ген fbaA, полученный из Escherichia coli (SEQ ID NO: 20, нить, комплементарная нуклеотидам с номерами 3068187..3069266 в GenBank с No. доступа NC_000913), и ген fbaB, полученный из Escherichia coli (SEQ ID NO: 72, нить, комплементарная нуклеотидам с номерами 2175534..2176586 в GenBank с No. доступа NC_000913).
Ген, кодирующий фруктозобисфосфатальдолазу, может представлять собой ДНК, которая гибридизуется с последовательностью, комплементарной нуклеотидной последовательности SEQ ID NO: 20 или 72, или зондом, который может быть получен из комплементарной последовательности, в жестких условиях и кодирует белок, обладающий активностью фруктозобисфосфатальдолазы. Примеры также включают ДНК, кодирующую белок с последовательностью SEQ ID NO: 21 или 73. Белок может представлять собой белок, имеющий гомологию, составляющую 80% или выше, предпочтительно 90% или выше, более предпочтительно 95% или выше, особенно предпочтительно 98% или выше с общей аминокислотной последовательностью SEQ ID NO: 21, при условии, что он проявляет повышенную активность фруктозобисфосфатальдолазы в микроорганизме, относящемся к семейству Enterobacteriaceae, при его введении в микроорганизм.
Кроме того, ген может представлять собой ДНК, кодирующую белок, имеющий аминокислотную последовательность SEQ ID NO: 21 или 73, но которая может включать в себя замену, делецию, инсерция, добавление или тому подобного одного или нескольких аминокислотных остатков, при условии, что активность фруктозобисфосфатальдолазы не снижена.
Активность фруктозобисфосфатальдолазы может быть измерена с использованием способа Richard and Rutter (J. Biol. Chem., 236, 3177-3184). Активность может быть повышена способами, сходными со способами, указанными выше для усиления глицериндегидрогеназы и дигидроксиацетонкиназы.
Фруктозо-1,6-бисфосфатаза является ферментом, который обратимо катализирует следующую реакцию, в которой происходит превращение D-фруктозо-1,6-бисфосфата в D-фруктозо-6-фосфат (EC:3.1.3.11).
D-фруктозо-1,6-бисфосфат+H2O=D-фруктозо-6-фосфат+фосфат
Фраза «модифицирован так, чтобы повысить активность фруктозо-1,6-бисфосфатазы» может означать, что количество молекул фруктозо-1,6-бисфосфатазы в клетке может быть увеличено по сравнению с количеством в штамме дикого типа или в немодифицированном штамме или что активность фруктозо-1,6-бисфосфатазы на молекулу повышена по сравнению с активностью в штамме дикого типа или немодифицированном штамме. Модификацию предпочтительно осуществляют так, чтобы активность фруктозо-1,6-бисфосфатазы была повышена до 150% или выше, более предпочтительно до 200% или выше, еще более предпочтительно до 300% или выше по сравнению с активностью штамма дикого типа или немодифицированного штамма. Примеры штаммов дикого типа микроорганизма, относящегося к семейству Enterobacteriaceae, которые могут служить в качестве эталона для сравнения, включают штамм MG1655 (ATCC No. 47076) и штамм W3110 (ATCC No. 27325) Escherichia coli, штамм AJ13335 Pantoea ananatis (FERM BP-6615), и другие.
Примеры гена, кодирующего фруктозо-1,6-бисфосфатазу, включают ген glpX (SEQ ID NO: 22, нить, комплементарная нуклеотидам с номерами 4112592..4113602 в GenBank с No. доступа NC_000913), ген fbp (SEQ ID NO: 82, нуклеотиды с номерами 4452634..4453632 в GenBank с No. доступа NC_000913) и ген ybhA (SEQ ID NO: 84, нуклеотиды с номерами 796836..7976554 в GenBank с No. доступа NC_000913), которые получены из Escherichia coli. Ген, кодирующий фруктозо-1,6-бисфосфатазу, может представлять собой ДНК, которая гибридизуется с последовательностью, комплементарной нуклеотидной последовательности SEQ ID NO: 22, 82 или 84, или с зондом, который может быть получен из комплементарной последовательности в жестких условиях, и кодирует белок, обладающий активностью фруктозо-1,6-бисфосфатазы. Примером также является ДНК, кодирующая белок с последовательностью SEQ ID NO: 23, 83 или 85. Белок может представлять собой белок, имеющий гомологию 80% или выше, предпочтительно 90% или выше, более предпочтительно 95% или выше, еще более предпочтительно 98% или выше с общей аминокислотной последовательностью SEQ ID NO: 23, 83 или 85, при условии, что он обладает повышенной активностью фруктозо-1,6-бисфосфатазы в микроорганизме, относящемся к семейству Enterobacteriaceae, при введении в микроорганизм.
Кроме того, ген может представлять собой ДНК, кодирующую белок, имеющий аминокислотную последовательность SEQ ID NO: 23, 83 или 85, но которая может включать в себя замену, делецию, инсерция, добавление или тому подобного одного или нескольких аминокислотных остатков, при условии, что активность фруктозо-1,6-бисфосфатазы не снижена.
Активность фруктозо-1,6-бисфосфатазы может быть измерена с использованием способа Nakajima et al. (Protein Nucleic Enzyme, 22, 1585-1589). Активность может быть повышена способами, сходными со способами, указанными выше для усиления глицериндегидрогеназы и дигидроксиацетонкиназы.
В настоящем изобретении «фруктозо-6-фосфатальдолаза» означает фермент, который обратимо катализирует следующую реакцию, в которой происходит превращение дигидроксиацетона в фруктозо-6-фосфат.
D-фруктозо-6-фосфат=дигидроксиацетон+D-глицеральдегид-3-фосфат
Фраза «модифицирован так, чтобы повысить активность фруктозо-6-фосфатальдолазы» может означать, что количество молекул фруктозо-6-фосфатальдолазы в клетке может быть увеличено по сравнению с количеством в штамме дикого типа или в немодифицированном штамме или что активность фруктозо-6-фосфатальдолазы на молекулу может быть повышена по сравнению с активностью в штамме дикого типа или немодифицированном штамме. Модификацию предпочтительно осуществляют так, чтобы активность фруктозо-6-фосфатальдолазы была повышена до 150% или выше, более предпочтительно до 200% или выше, еще более предпочтительно до 300% или выше по сравнению с активностью, наблюдаемой в штамме дикого типа или немодифицированном штамме. Примеры штаммов дикого типа микроорганизма, относящегося к семейству Enterobacteriaceae, которые служат в качестве эталона для сравнения, включают штамм MG1655 (ATCC No. 47076) и штамм W3110 (ATCC No. 27325) Escherichia coli, штамм AJ13335 Pantoea ananatis (FERM BP-6615), и другие.
Примерами гена, кодирующего фруктозо-6-фосфатальдолазу, являются ген fsaA, кодирующий альдолазу типа I (SEQ ID NO: 68, номера нуклеотидов 862865..863527 в GenBank с No. доступа NC_000913), и ген fsaB (ген talC) (SEQ ID NO: 70, нить, комплементарная нуклеотидам с номерами 4137069..4137731 в GenBank с No. доступа NC_000913), кодирующий альдолазу типа II, которые получены из Escherichia coli.
Ген, кодирующий фруктозо-6-фосфатальдолазу, может представлять собой ДНК, которая гибридизуется с последовательностью, комплементарной нуклеотидной последовательности SEQ ID NO: 68 или 70, или с зондом, который может быть получен из комплементарной последовательности в жестких условиях, и кодирует белок, обладающий активностью фруктозо-6-фосфатальдолазы. Примером также является ДНК, кодирующая белок с последовательностью SEQ ID NO: 69 или 71. Белок может представлять собой белок, имеющий гомологию 80% или выше, предпочтительно 90% или выше, более предпочтительно 95% или выше, еще более предпочтительно 98% или выше с общей аминокислотной последовательностью SEQ ID NO: 69 или 71, при условии, что он обладает повышенной активностью фруктозо-6-фосфатальдолазы в микроорганизме, относящемся к семейству Enterobacteriaceae, при введении в микроорганизм.
Кроме того, ген может представлять собой ДНК, кодирующую белок, имеющий аминокислотную последовательность SEQ ID NO: 69 или 71, но которая включает в себя замену, делецию, инсерция, добавление или тому подобного одного или нескольких аминокислотных остатков, при условии, что активность фруктозо-6-фосфатальдолазы не снижена.
Активность фруктозо-6-фосфатальдолазы может быть измерена с использованием способа Schurmann M., Sprenger G.A. et al. (J. Biol. Chem., 2001 Apr. 6, 276 (14): 11055-61). Активность может быть повышена способами, сходными со способами, указанными выше для усиления глицериндегидрогеназы и дигидроксиацетонкиназы.
Микроорганизм, используемый в способе получения согласно настоящему изобретению, предпочтительно модифицируют так, чтобы снизить активности глицеринкиназы и/или глицерин-3-фосфатдегидрогеназы связанного с мембраной типа в добавление к усилению глицериндегидрогеназы и дигидроксиацетонкиназы, повышению активности поглощения глицерина и повышению активностей одного или нескольких видов ферментов, выбранных из группы, состоящей из триозофосфатизомеразы, фруктозобисфосфатальдолазы, фруктозо-1,6-бисфосфатазы и фруктозо-6-фосфатальдолазы.
В настоящем изобретении «глицеринкиназа» означает фермент, который обратимо катализирует следующую реакцию, которая приводит к созданию глицерин-3-фосфата и АДФ из глицерина и АТФ (EC2.7.1.30)
АТФ+глицерин=АДФ+sn-глицерин-3-фосфат
Фраза «модифицирован так, чтобы уменьшить активность глицеринкиназы» может означать, что количество молекул глицеринкиназы в клетке уменьшено по сравнению с количеством в штамме дикого типа или в немодифицированном штамме или что активность глицеринкиназы на молекулу снижена по сравнению с активностью в штамме дикого типа или немодифицированном штамме. Модификацию предпочтительно осуществляют так, чтобы активность глицеринкиназы была снижена до 70% или ниже, более предпочтительно до 50% или ниже, еще более предпочтительно до 30% или ниже, наиболее предпочтительно до 20% или ниже по сравнению с активностью штамма дикого типа или немодифицированного штамма и ферментативная активность может быть устранена. Ферментативная активность может быть снижена в результате снижения уровня экспрессии гена, кодирующего фермент. Снижение уровня экспрессии гена включает в себя снижение уровня транскрипции мРНК, транскрибируемой с гена, и снижение уровня трансляции такой мРНК.
Полное исключение продукции молекулы ферментного белка или уменьшение или исключение активности молекулы ферментного белка достигается в результате нарушения гена, кодирующего фермент. Примеры штаммов дикого типа микроорганизма, относящегося к семейству Enterobacteriaceae, которые могут служить в качестве эталона для сравнения, включают штамм MG1655 (ATCC No. 47076) и штамм W3110 (ATCC No. 27325) Escherichia coli, штамм AJ13335 Pantoea ananatis (FERM BP-6615), и другие.
Примером гена, кодирующего глицеринкиназу, является ген glpK (SEQ ID NO: 24, нить, комплементарная нуклеотидам с номерами 4113737..4115245 в GenBank с No. доступа NC_000913), полученный из Escherichia coli. Ферментативная активность глицеринкиназы может быть измерена способом Thorner and Paulus (The Enzymes, 3rd ed., 8, 487-508).
В настоящем изобретении «глицерин-3-фосфатдегидрогеназа связанного с мембраной типа» означает фермент, который катализирует реакцию окисления, в которой происходит превращение глицерин-3-фосфата в дигидроксиацетонфосфат, и является ферментом, который обратимо катализирует следующую реакцию.
sn-Глицерин-3P+убихинон=дигидроксиацетон-P+убихинол (EC:1.1.99.5)
Фраза «модифицирован так, чтобы уменьшить активность глицерин-3-фосфатдегидрогеназы связанного с мембраной типа» может означать, что количество молекул глицерин-3-фосфатдегидрогеназы связанного с мембраной типа в клетке уменьшено по сравнению с количеством в штамме дикого типа или в немодифицированном штамме, или что активность глицерин-3-фосфатдегидрогеназы связанного с мембраной типа на молекулу снижена по сравнению с активностью в штамме дикого типа или немодифицированном штамме. Модификацию предпочтительно осуществляют так, чтобы активность глицерин-3-фосфатдегидрогеназы связанного с мембраной типа на клетку была снижена до 70% или ниже, более предпочтительно до 50% или ниже, еще более предпочтительно до 30% или ниже по сравнению с активностью штамма дикого типа или немодифицированного штамма и ферментативная активность может быть устранена. Ферментативная активность может быть снижена в результате снижения уровня экспрессии гена, кодирующего фермент. Примеры штаммов дикого типа микроорганизма, относящегося к семейству Enterobacteriaceae, которые могут служить в качестве эталона для сравнения, включают штамм MG1655 (ATCC No. 47076) и штамм W3110 (ATCC No. 27325) Escherichia coli, штамм AJ13335 Pantoea ananatis (FERM BP-6615), и другие.
Глицерин-3-фосфатдегидрогеназа связанного с мембраной типа кодируется опероном glpABC и геном glpD, и примеры гена glpA Escherichia coli включают последовательность SEQ ID NO: 26 (нуклеотиды с номерами 2350669..2352297 в GenBank с No. доступа NC_000913), примером гена glpB Escherichia coli является последовательность SEQ ID NO: 28 (нуклеотиды с номерами 2352287..2353546 в GenBank с No. доступа NC_000913), примером гена glpC Escherichia coli является последовательность SEQ ID NO: 30 (нуклеотиды с номерами 2353543..2354733 в GenBank с No. доступа NC_000913), и примером гена glpD Escherichia coli является последовательность SEQ ID NO: 32 (нуклеотиды с номерами 3560036..3561541 в GenBank с No. доступа NC_000913).
Уменьшения активности целевого фермента, такого как глицеринкиназа и глицерин-3-фосфатдегидрогеназа, указанное выше, можно достичь в результате
(1) уменьшения или устранение ферментативной активности посредством введения мутации в кодирующую область гена, кодирующего целевой фермент, или
(2) уменьшения или устранения ферментативной активности посредством модификации последовательности регуляции экспрессии гена, кодирующего целевой фермент.
(1) Уменьшение или устранение ферментативной активности посредством введения мутации в кодирующую область гена, кодирующего целевой фермент
Введение мутации в кодирующую область гена, кодирующего целевой фермент, можно осуществить посредством введения мутации для аминокислотной замены (миссенс-мутация), стоп-кодона (нонсенс-мутация), или мутации со сдвигом рамки, которая добавляет или делетирует один или два нуклеотида в область целевого гена, кодирующего фермент, в хромосоме в результате генетической рекомбинации (Journal of Biological Chemistry, 272: 8611-8617 (1997); Proceedings of the National Academy of Sciences, USA, 95 5511-5515 (1998); Journal of Biological Chemistry, 266, 20833-20839 (1991)). Мутацию также можно осуществить посредством делетирования части или всей кодирующей области гена. В частности, мутацию можно осуществить посредством введения мутации в часть ДНК с последовательностью SEQ ID NO: 24, 26, 28, 30 или 32, или посредством делеции части или всей такой ДНК.
Что касается введения мутации, то ферментативная активность также может быть снижена или устранена в результате конструирования гена, кодирующего мутантный фермент, кодирующая область которого делетирована или в кодирующую область которого введена мутация, и в результате замены сконструированным геном нормального гена в хромосоме посредством гомологичной рекомбинации или тому подобным способом, или в результате введения транспозона или IS-фактора в ген.
Для введения таких мутаций с целью снижения или устранения активности фермента, который описан выше, в ген посредством генетической рекомбинации, используют, например, следующие способы. Посредством модификации части последовательности целевого гена для получения мутантного гена, сконструированного так, чтобы он не продуцировал фермент, который способен нормально функционировать, и трансформации микроорганизма, относящегося к семейству Enterobacteriaceae, с использованием ДНК, содержащей ген, чтобы вызвать рекомбинацию мутантного гена и соответствующего гена в хромосоме, целевой ген в хромосоме может быть заменен мутантным геном. Для такой замены гена с использованием гомологичной рекомбинации можно применять способ, названный зависимой от Red интеграцией (Datsenko, K.A, and Wanner, B.L., Proc. Natl. Acad. Sci. USA, 97: 6640-6645 (2000)), способ с использованием линейной ДНК, такой как способ с использованием зависимой от Red интеграции в сочетании с эксцизионной системой, полученной из фага λ (Cho, E.H., Gumport, R.I., Gardner, J.F., J. Bacteriol., 184:5200-5203 (2002)), способ, основанный на использовании плазмиды, содержащей чувствительное к температуре начало репликации (Datsenko, K.A, and Wanner, B.L., Proc. Natl. Acad. Sci. USA, 97: 6640-6645 (2000), патент США No. 6303383, выложенный патент Японии No. 05-007491), и другие. Кроме того, такой сайт-специфичный мутагенез, основанный на замене гена с использованием гомологичной рекомбинации, который описан выше, также можно осуществлять, используя плазмиду, которая не способна реплицироваться в хозяине. Кроме того, снижение или устранение ферментативной активности также можно осуществлять с помощью модификации с целью введения мутации в кодирующую область целевого гена посредством обычной мутагенной обработки на основе рентгеновского излучения или ультрафиолетового излучения или с использованием мутагена, такого как N-метил-N'-нитро-N-нитрозогуанидин.
(2) Уменьшение или устранение ферментативной активности посредством модификации последовательности регуляции экспрессии гена, кодирующего целевой фермент
Уменьшение или устранение ферментативной активности в результате модификации последовательности регуляции экспрессии гена, кодирующего целевой фермент, также можно осуществлять посредством снижения уровня экспрессии с помощью введения мутации в последовательность регуляции экспрессии, такую как промотор и последовательность SD в хромосомной ДНК, амплификации гена, кодирующего регулятор, который уменьшает экспрессию гена, или делеции или ослабления гена, кодирующего активатор, который усиливает экспрессию гена. Способы оценки силы промоторов и примеры сильных промоторов описаны в публикации Goldstein et al. (Prokaryotic promoters in biotechnology, Biotechnol. Annu. Rev., 1995, 1, 105-128) и других публикациях. Кроме того, известно, что замена нескольких нуклеотидов в области спейсера между сайтом связывания рибосом (RBS) и стартовым кодоном, в частности, в области, расположенной непосредственно перед стартовым кодоном, существенно влияет на эффективность трансляции мРНК, и такая область также может быть модифицирована. В частности, гены glpA, B и C образуют структуру оперона, и поэтому уровень их экспрессии может быть снижен посредством введения мутации в область регуляции экспрессии, такую как область промотора, расположенная выше glpA.
2. Способ получения согласно настоящему изобретению
Способ получения согласно настоящему изобретению представляет собой способ получения L-аминокислоты, который включает в себя культивирование микроорганизма, относящегося к семейству Enterobacteriaceae, обладающего способностью продуцировать L-аминокислоту и модифицированного так, чтобы увеличить активности глицериндегидрогеназы и дигидроксиацетонкиназы, в среде, содержащей глицерин в качестве источника углерода, чтобы получить и накопить L-аминокислоту в среде или клетках, и сбор L-аминокислоты из среды или клеток. В случае способа согласно настоящему изобретению можно использовать любое периодическое культивирование, культивирование с подпиткой или непрерывное культивирование. Глицерин, находящийся в среде, может входить в состав начальной среды, среды, используемой для подпитки или в обеих средах.
В настоящем изобретении упоминаемое выше культивирование с подпиткой относится к способу культивирования, при котором среду непрерывно или с определенными интервалами подают в сосуд для культивирования и среду не извлекают до конца культивирования. Непрерывное культивирование означает способ, при котором среду непрерывно или с определенными интервалами подают в сосуд для культивирования и одновременно среду извлекают из сосуда (обычно в объеме, равном объему подаваемой среды). Начальная среда означает среду, используемую в периодической культуре до подачи среды для подпитки при культивировании с подпиткой или при непрерывном культивировании (среда, используемая в начале культивирования). Среда для подпитки означает среду, которую подают в чан для ферментации при культивировании с подпиткой или при непрерывном культивировании. Периодическое культивирование означает способ, при котором для каждой культуры готовят свежую среду, штамм высевают в свежую среду и после этого среду не добавляю вплоть до сбора.
Глицерин, присутствующий в среде, используемой в настоящем изобретении, может быть единственным источником углерода, или можно использовать смешанную среду, которая содержит другие источники углерода, кроме глицерина. Предпочтительными являются сахариды, такие как глюкоза, фруктоза, сахароза, лактоза, галактоза, мелассы и раствор сахаров, полученный в результате гидролиза крахмального гидролизата или биомассы, спирты, такие как этанол, и органические кислоты, такие как фумаровая кислота, лимонная кислота и янтарная кислота. При использовании смешанной среды глицерин предпочтительно присутствует в среде в соотношении 50% или больше, предпочтительно 60% или больше, более предпочтительно 70% или больше, еще более предпочтительно 80% или больше, особенно предпочтительно 90% или больше. Особенно предпочтительно использование глицерина, полученного в виде побочного продукта при производстве биодизельного топлива (Mu Y., et al., Biotechnol. Lett., 28, 1755-91759 (2006); Haas M.J., et al., Bioresour. Technol., 97, 4, 671-8678 (2006)).
Что касается других компонентов, которые могут быть добавлены в среду, то можно использовать обычную среду, содержащую кроме источника углерода источник азота, неорганические ионы и другие органические компоненты, которые необходимы. В качестве источника азота, находящегося в среде, используемой для настоящего изобретения, можно использовать аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония, ацетат аммония и мочевину, нитраты и т.д. Газообразный аммиак и водный раствор аммиака, используемый для корректировки pH, также может быть использован в качестве источника азота. Кроме того, также можно использовать пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт, гидролизат сои и т.д. Среда может содержать один или несколько источников азота. Такие источники азота также можно использовать как в начальной среде, так и в среде для подпитки. Кроме того, можно использовать один и тот же источник азота как в начальной среде, так и в среде для подпитки, или источник азота в среде для подпитки может отличаться от источника азота в начальной среде.
Среда, используемая для настоящего изобретения, предпочтительно содержит источник фосфорной кислоты и источник серы в добавление к источнику углерода, источнику азота и серы. В качестве источника фосфорной кислоты можно использовать первичный кислый фосфат калия, вторичный кислый фосфата калия, фосфатные полимеры, такие как пирофосфорная кислота, и т.д. Источником серы может быть любой источник серы, при условии, что он содержит атом серы, и предпочтительными являются соли серной кислоты, такие как сульфаты, тиосульфаты и сульфиты, и серосодержащие аминокислоты, такие как цистеин, цистин и глутатион. Из таких источников особенно предпочтителен сульфат аммония.
Кроме того, среда может содержать стимулирующий рост фактор (питательное вещество, обладающее стимулирующим рост действием) в добавление к источнику углерода, источнику азота и серы. В качестве стимулирующего рост фактора можно использовать металлические микроэлементы, аминокислоты, витамины, нуклеиновые кислоты, а также пептон, казаминовую кислоту, дрожжевой экстракт, продукт расщепления соевого белка и другие добавки, содержащие указанные выше вещества. Примеры металлических микроэлементов включают железо, марганец, магний, кальция и другие. Примеры витаминов включают витамин B1, витамин B2, витамин B6, никотиновую кислоту, амид никотиновой кислоты, витамин B12 и другие. Такие стимулирующие рост факторы могут находиться в начальной среде или в среде для подпитки.
Кроме того, при использовании ауксотрофного мутанта, для роста которого требуется аминокислота или тому подобное, предпочтительно добавление требуемого питательного вещества в среду. В частности, так как путь биосинтеза L-лизина усилен, а способность осуществлять распад L-лизина ослаблена у многих продуцирующих L-лизин бактерий, которые могут быть использованы для настоящего изобретения, как описано ниже, предпочтительно добавляют один или несколько типов веществ, выбранных из L-треонина, L-гомосерина, L-изолейцина и L-метионина.
Начальная среда и среда для подпитки могут иметь одинаковые или разные составы. Кроме того, начальная среда и среда для подпитки могут иметь одинаковые или разные концентрации серы. Кроме того, в том случае, когда среду для подпитки подают на нескольких стадиях, состав среды для подпитки может быть одним и тем же или разным.
Культивирование предпочтительно осуществляют в виде культивирования с аэрацией при температуре ферментации 20-45°C, особенно предпочтительно при 30-42°C. Концентрацию кислорода доводят до 5-50%, предпочтительно примерно до 10%. Кроме того, культивирование с аэрацией предпочтительно осуществляют при значении pH 5-9. Если pH снижается во время культивирования, то можно добавлять, например, карбонат кальция или щелочь, такую как газообразный аммиак и водный раствор аммиака, чтобы нейтрализовать культуру. Когда культивирование осуществляют в таких условиях предпочтительно в течение примерно от 10 до 120 часов, в культуральной среде накапливается заметное количество L-аминокислоты. Хотя концентрация L-аминокислоты, которая накапливается, не ограничена, при условии, что она не превышает концентрацию, которая наблюдается при использовании штаммов дикого типа, и L-аминокислота может быть выделена и собрана из среды, такая концентрация может составлять 50 г/л или выше, предпочтительно 75 г/л или выше, более предпочтительно 100 г/л или выше.
После культивирования L-аминокислота может быть собрана из среды известным способом. Например, L-аминокислота может быть собрана посредством удаления клеток из культуральной среды центрифугированием или тому подобным способом и затем кристаллизацией L-аминокислоты с помощью концентрирования.
В настоящем изобретении культивирование микроорганизма можно осуществить в виде посевной культуры и основной культуры, чтобы обеспечить накопление L-аминокислоты выше определенного уровня. Посевную культуру можно получать в виде встряхиваемой культуры, используя колбу или тому подобное, или в виде периодической культуры, а основную культуру можно получать в виде культуры с подпиткой или непрерывной культуры. Альтернативно и посевную и основную культуру можно получать в виде периодической культуры.
В том случае, когда осуществляют культивирование с подпиткой или непрерывное культивирование согласно настоящему изобретению, среда для подпитки может подаваться с определенными интервалами, так чтобы подача глицерина и другого источника углерода временно приостанавливалась. Подачу среды для подпитки предпочтительно останавливают на период времени, составляющий максимум 30% или меньше, предпочтительно 20% или меньше, особенно предпочтительно 10% или меньше от времени подпитки. Когда среду для подпитки подают с интервалами, среду для подпитки можно сначала добавлять в течение заранее определяемого промежутка времени, а второе и последующие добавления можно регулировать так, чтобы они начинались тогда, когда компьютер регистрирует повышение pH или концентрации растворенного кислорода при истощении источника углерода в среде для ферментации. Обычно это происходит в течение периода, когда среду не подают, и перед подачей среды, и, таким образом, концентрация субстрата в резервуаре для культивирования всегда автоматически поддерживается на низком уровне (патент США No. 5912113).
Среда для подпитки, используемая при культивировании с периодической подпиткой, предпочтительно представляет собой среду, содержащую глицерин или другой источник углерода и питательное вещество, оказывающее стимулирующие рост действие (стимулирующий рост фактор), и концентрацию глицерина и другого источника углерода в среде для ферментации можно регулировать так, чтобы она была на уровне предварительно определяемой концентрации или ниже. В качестве источника углерода предпочтительны глюкоза, сахароза и фруктоза. В качестве стимулирующего рост фактора предпочтительны источник азота, фосфорная кислота, аминокислоты и т.д. В качестве источника азота можно использовать аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония, ацетат аммония и мочевина, нитраты и т.д. Кроме того, в качестве источника фосфорной кислоты можно использовать первичный кислый фосфат калия и вторичный кислый фосфат калия. Что касается аминокислот в том случае, когда используют ауксотрофный мутантный штамм, то предпочтительно добавление требуемых питательных веществ. Кроме того, среда для подпитки может включать в себя один тип среды или смесь двух или более типов сред. Когда используют два или более типов сред для подпитки среда может быть смешанной и подаваться с использованием одного сосуда для подачи или подаваться с использованием двух или более сосудов для подачи.
При использовании для настоящего изобретения способа непрерывного культивирования среда может быть удалена и одновременно добавлена, или часть среды может быть удалена и затем среда может быть добавлена. Кроме того, способ также может представлять собой способ непрерывного культивирования, который включает в себя извлечение культуральной среды, содержащей L-аминокислоту и бактериальные клетки, и возвращение в ферментер только клеток для их повторного использования (патент Франции No. 2669935). В качестве способа непрерывной подпитки или подпитки с интервалами источником азота применяют такой же способ, как и способ, используемый при периодическом культивировании с подпиткой.
Способ непрерывного культивирования с многократным использованием бактериальных клеток представляет собой способ периодического или непрерывного извлечения среды для ферментации, когда концентрация аминокислоты достигает предварительно определяемого уровня, удаления только L-аминокислоты и возвращения остатков, полученных после фильтрования и содержащих бактериальные клетки, в ферментер, и способ может быть осуществлен согласно, например, патенту Франции No. 2669935.
В том случае, когда культуральную среду извлекают периодически, предпочтительно, чтобы извлечение части количества L-аминокислоты происходило при достижении концентрации L-аминокислоты предварительно определяемого уровня, и после этого осуществлялась подача свежей среды, чтобы продолжить культивирование. Кроме того, что касается объема добавляемой среды, то культивирование предпочтительно осуществляют так, чтобы конечный объем среды после добавления среды был равен объему культуральной среды перед извлечением. Термин «равен» может означать, что объем соответствует примерно 93-107% объема культуральной среды до извлечения.
В том случае, когда извлечение культуральной среды осуществляют постоянно, извлечение предпочтительно начинают одновременно или после добавления питательной среды. Например, начальное время извлечения составляет максимум 5 часов, предпочтительно 3 часа, более предпочтительно 1 час после начала подпитки. Кроме того, извлекаемый объем культуральной среды предпочтительно равен объему подаваемой среды.
3. Микроорганизмы, которые могут быть использованы в качестве родительских штаммов, чтобы получить типичные микроорганизмы согласно настоящему изобретению
В настоящем изобретении бактерия, относящаяся к семейству Enterobacteriaceae и обладающая способностью продуцировать L-аминокислоту, которая может метаболизировать глицерин в качестве источника углерода, может быть использована в качестве родительского штамма, и требуемое свойство в настоящем изобретении можно придать указанными выше способами.
Семейство Enterobacteriaceae охватывает бактерии, относящиеся к роду Escherichia, Enterobacter, Erwinia, Klebsiella, Pantoea, Photorhabdus, Providencia, Salmonella, Serratia, Shigella, Morganella, Yersinia и т.д. В частности, предпочтительны бактерии, классифицируемые как бактерии семейства Enterobacteriaceae согласно систематике, используемой в базе данных NCBI (National Center for Biotechnology Information) (http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=91347).
Выражение «бактерия, относящаяся к роду Escherichia» означает, что бактерия классифицирована как бактерия рода Escherichia согласно классификации, известной специалисту в области микробиологии, хотя конкретно бактерия не ограничена. Примером бактерии, относящейся в роду Escherichia, используемой в настоящем изобретении, является без ограничения Escherichia coli (E. coli).
Бактерия, относящаяся к роду Escherichia, которая может быть использована в настоящем изобретении, особым образом не ограничена. Однако примеры включают, например, бактерии филогенетических групп, описанных в работе Bachmann et al., в таблице 1 (Bachmann, B.J., 1996, pp. 2460-2488, In F.D. Neidhardt (ed.), Escherichia coli and Salmonella: Cellular and Molecular Biology/Second Edition, American Society for Microbiology Press, Washington, D.C.). Конкретные примеры включают Escherichia coli W3110 (ATCC 27325), Escherichia coli MG1655 (ATCC 47076) и другие штаммы, полученные из прототипного штамма дикого типа K12.
Такие штаммы доступны, например, из американской коллекции типов культур (адрес: 12301 Parklawn Drive, Rockville, Maryland 20852, United States of America). То есть, для каждого штамма приведены номера доступа, и штаммы можно заказать, используя указанные номера. Номера доступа штаммов перечислены в каталоге Американской коллекции типов культур.
Выражение «бактерия, относящаяся к роду Pantoea» означает, что бактерия классифицирована как бактерия рода Pantoea согласно классификации, известной специалисту в области микробиологии. Некоторые штаммы Enterobacter agglomerans недавно были классифицированы как Pantoea agglomerans, Pantoea ananatis, Pantoea stewartii или тому подобные на основании анализа нуклеотидных последовательностей рРНК 16S и других признаков (Int. J. Syst. Bacteriol., 43, 162-173 (1993)). В настоящем изобретении бактерии, относящиеся к роду Pantoea, могут охватывать такие бактерии, которые были переклассифицированы как бактерии рода Pantoea, которые описаны выше.
В настоящем изобретении бактерия, обладающая продуцирующей L-аминокислоту способностью (способность продуцировать L-аминокислоту), означает бактерию, которая может продуцировать и секретировать L-аминокислоту в среду при ее культивировании в среде. Предпочтительно указанный термин означает бактерию, которая может накапливать целевую L-аминокислоту в среде в количестве, составляющем не менее 0,5 г/л, более предпочтительно не менее 1,0 г/л. Термин «L-аминокислота» охватывает L-аланин, L-аргинин, L-аспарагин, L-аспарагиновую кислоту, L-цистеин, L-глутаминовую кислоту, L-глутамин, глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин. Особенно предпочтительны L-треонин и L-лизин.
В качестве родительского штамма, используемого для настоящего изобретения, можно использовать любые продуцирующие L-аминокислоты бактерии, описанные до настоящего времени, при условии, что выбран штамм, который может ассимилировать глицерин. Далее описаны бактерии, продуцирующие L-аминокислоты, применимые в способе согласно настоящему изобретению.
Бактерии, продуцирующие L-треонин
Примеры продуцирующих L-треонин бактерий и родительского штамма, который можно использовать для получения таких бактерий, включают без ограничения штаммы, относящиеся к роду Escherichia, такие как E. coli TDH-6/pVIC40 (VKPM B-3996) (патент США No. 5175107, патент США No. 5705371), E. coli 472T23/pYN7 (ATCC 98081) (патент США No. 5631157), E. coli NRRL-21593 (патент США No. 5939307), E. coli FERM BP-3756 (патент США No. 5474918), E. coli FERM BP-3519 и FERM BP-3520 (патент США No. 5376538), E. coli MG442 (Гусятинер и соавт., Генетика (Россия), 14, 947-956 (1978)), E. coli VL643 и VL2055 (EP 1149911 A) и другие.
Штамм TDH-6 является дефицитным по гену thrC, а также усваивает сахарозу, и ген ilvA имеет мутацию с просачиванием исходного фенотипа. Указанный штамм также имеет мутацию в гене rhtA, который придает резистентность к высокой концентрации треонина или гомосерина. Штамм B-3996 несет плазмиду pVIC40, полученную инсерцией оперона thrA*BC, содержащего мутантный ген thrA, в вектор, являющийся производным RSF1010. Указанный мутантный ген thrA кодирует аспартокиназу-гомосериндегидрогеназу I, которая по существу не чувствительна к ингибированию треонином по механизму обратной связи. Штамм B-3996 депонирован 19 ноября 1987 во Всесоюзном научном центре антибиотиков (ул. Нагатинская 3-A, 117105 Москва Россия) с номером доступа RIA 1867. Штамм также депонирован во Всероссийской национальной коллекции промышленных микроорганизмов (ВКПМ) 7 апреля 1987 с номером доступа VKPM B-3996.
E. coli VKPM B-5318 (EP 0593792 B) также можно использовать в качестве бактерии, продуцирующей L-треонин, или родительского штамма для ее получения. Штамм B-5318 является прототрофным по отношению к изолейцину, и в таком штамме чувствительный к температуре репрессор Cl фага лямбда и промотор PR заменяют регуляторную область треонинового оперона в плазмиде pVIC40. Штамм VKPM B-5318 депонирован во Всероссийской национальной коллекции промышленных микроорганизмов (ВКПМ) (1 Дорожный проезд, 1 Москва 117545, Россия) 3 мая 1990 с номером доступа VKPM B-5318.
Предпочтительно бактерию, используемую для настоящего изобретения, дополнительно модифицируют так, чтобы повысить экспрессию одного или нескольких из следующих генов:
- мутантный гена thrA, который кодирует аспартокиназу-гомосериндегидрогеназу I, резистентную к ингибированию треонином по механизму обратной связи;
- ген thrB, который кодирует гомосеринкиназу;
- ген thrC, который кодирует треонинсинтазу;
- ген rhtA, который кодирует предполагаемый трансмембранный белок;
- ген asd, который кодирует аспартат-β-полуальдегиддегидрогеназу; и
- ген aspC, который кодирует аспартатаминотрансферазу (аспартаттрансаминазу).
Ген thrA, который кодирует аспартокиназу-гомосериндегидрогеназу I Escherichia coli, был определен (номера нуклеотидов 337-2799, доступ в GenBank NC_000913.2, gi: 49175990). Ген thrA локализован между генами thrL и thrB в хромосоме E. coli K-12. Ген thrB, который кодирует гомосеринкиназу Escherichia coli, был определен (номера нуклеотидов 2801-3733, доступ в GenBank NC_000913.2, gi: 49175990). Ген thrB локализован между генами thrA и thrC в хромосоме of E. coli K-12. Ген thrC, который кодирует треонинсинтазу Escherichia coli, был определен (номера нуклеотидов 3734-5020, доступ в GenBank NC_000913.2, gi: 49175990). Ген thrC локализован между геном thrB и открытой рамкой считывания yaaX в хромосоме E. coli K-12. Все три указанных гена функционируют в виде одного треонинового оперона. Чтобы увеличить экспрессию треонинового оперона, область аттенюатора, который влияет на транскрипцию, предпочтительно удаляют из оперона (WO2005/049808, WO2003/097839).
Мутантный ген thrA, который кодирует аспартокиназу-гомосериндегидрогеназу I, резистентную к ингибированию треонином по механизму обратной связи, а также гены thrB и thrC, могут быть получены в виде одного оперона из хорошо известной плазмиды pVIC40, которая присутствует в продуцирующем треонин штамме E. coli VKPM B-3996. Плазмида pVIC40 подробно описана в патенте США No. 5705371.
Ген rhtA присутствует в положении 18 мин на хромосоме E. coli вблизи оперона glnHPQ, который кодирует компоненты системы транспорта глутамина. Ген rhtA идентичен ORF1 (ген ybiF, номера нуклеотидов 764-1651, номер доступа в GenBank AAA218541, gi: 440181) и расположен между генами pexB и ompX. Единица, экспрессирующая белок, кодируемый открытой рамкой считывания ORF1, была названа геном rhtA (rht: резистентность к гомосерину и треонину). Также было обнаружено, что мутация rhtA23 представляет собой замену G на A в положении -1 по отношению к стартовому кодону ATG (ABSTRACTS of the 17th International Congress of Biochemistry and Molecular Biology in conjugation with Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francisco, California August 24-29, 1997, abstract No. 457, EP 1013765 A).
Ген asd E. coli уже определен (номера нуклеотидов 3572511-3571408, доступ в GenBank NC_000913.1, gi:16131307) и может быть получен в ПЦР (смотри White, T.J., Arnheim, N., and Erlich, H.A., Trends Genet., 5, 185-189 (1989)) с использованием праймеров, полученных на основе нуклеотидной последовательности гена. Гены asd других микроорганизмов могут быть получены подобным образом.
Ген aspC E. coli уже также определен (номера нуклеотидов 983742-984932, доступ в GenBank NC_000913.1, gi:16128895) и может быть получен в ПЦР. Гены aspC других микроорганизмов могут быть получены сходным образом.
Бактерии, продуцирующие L-лизин
Примеры бактерий, продуцирующих L-лизин, относящихся к роду Escherichia, включают мутанты, обладающие резистентностью к аналогу L-лизина. Аналоги L-лизина ингибируют рост бактерий, относящихся к роду Escherichia, но чувствительность к такому ингибированию полностью или частично снижается, если L-лизин присутствует в среде. Примеры аналогов L-лизина включают без ограничения оксализин, лизингидроксамат, S-(2-аминоэтил)-L-цистеин (AEC), γ-метиллизин, α-хлоркапролактам и т.д. Мутанты, обладающие резистентностью к таким аналогам лизина, можно получить, подвергая бактерии, относящиеся к роду Escherichia, обычной искусственной мутагенной обработке. Конкретными примерами бактериальных штаммов, применимых для получения L-лизина, являются Escherichia coli AJ11442 (FERM BP-1543, NRRL B-12185; смотри патент США No. 4346170) и Escherichia coli VL611. У таких микроорганизмов снижена чувствительность к ингибированию аспартокиназы L-лизином по механизму обратной связи.
Штамм WC196 может быть использован в качестве продуцирующей L-лизин бактерии Escherichia coli. Указанный бактериальный штамм был выведен в результате придания AEC-резистентности штамму W3110, который получали из Escherichia coli K-12. Полученный штамм назван Escherichia coli AJ13069 и депонирован в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology (в настоящее время National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary, Central 6, 1-1, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan) 6 декабря 1994 и ему присвоен номер доступа FERM P-14690. Затем он был превращен в международный депозит по условиям Будапештского соглашения 29 сентября 1995, и ему присвоен номер доступа FERM BP-5252 (патент США No. 5827698).
Примеры L-лизин-продуцирующих бактерий и родительских штаммов, которые могут быть использованы для получения таких бактерий, также включают штаммы, в которых экспрессия одного или нескольких генов, кодирующих фермент биосинтеза L-лизина, повышена. Примерами таких генов без ограничения являются гены дигидродипиколинатсинтазы (dapA), ген аспартокиназы (lysC), ген дигидродипиколинатредуктазы (dapB), ген диаминопимелатдекарбоксилазы (lysA), ген диаминопимелатдегидрогеназы (ddh) патент США No. 6040160), ген фосфоенолпируваткарбоксилазы (ppc), ген аспартатполуальдегиддегидрогеназы (asd) и ген аспартазы (aspA) (EP 1253195 A). Кроме того, родительские штаммы могут иметь повышенный уровень экспрессии гена, влияющего на энергетическую эффективность (cyo) (EP 1170376 A), гена, кодирующего никотинамиднуклеотидтрансгидрогеназу (pntAB) (патент США No. 5830716), ген ybjE (WO2005/073390), ген, кодирующий глутаматдегидрогеназу (gdhA, Gene, 23: 199-209 (1983)) или их сочетания. Сокращенные названия генов указаны в круглых скобках.
Известно, что дигидродипиколинатсинтетаза дикого типа, полученная из Escherichia coli, подвержена ингибированию L-лизином по механизму обратной связи, тогда как аспартокиназа дикого типа из Escherichia coli подвержена супрессии и ингибированию по механизму обратной связи L-лизином. Следовательно при использовании генов dapA и lysC такие гены предпочтительно являются мутантными генами, кодирующими ферменты, которые не подвергаются ингибированию L-лизином по механизму обратной связи.
Примеры ДНК, кодирующей мутантную дигидродипиколинатсинтетазу с пониженной чувствительностью у ингибированию L-лизином по механизму обратной связи, включают ДНК, кодирующую белок, который имеет аминокислотную последовательность фермента, в котором гистидин в положении 118 заменен тирозином. Примеры ДНК, кодирующей мутантную аспартокиназу с пониженной чувствительностью к ингибированию L-лизином по механизму обратной связи, включают ДНК, кодирующую AKIII, имеющую аминокислотную последовательность, в которой треонин в положении 352, глицин в положении 323 и метионин в положении 318 заменены изолейцином, аспарагином и изолейцином, соответственно (патент США No. 5661012 и патент США No. 6040160). Такие мутантные ДНК могут быть получены сайт-специфичным мутагенезом с использованием ПЦР и тому подобного.
Плазмиды RSFD80, pCAB1 и pCABD2 для широкого круга хозяев известны как плазмиды, содержащие мутантный ген dapA, кодирующий мутантную дигидродипиколинатсинтетазу, и мутантный ген lysC, кодирующий мутантную аспартокиназу (патент США No. 6040160). Штамм JM109 Escherichia coli, трансформированный плазмидой RSFD80, назван AJ12396 (патент США No. 6040160), и штамм депонирован в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (в настоящее время International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology) 28 октября 1993, и ему присвоен номер доступа FERM P-13936, и затем депозит был превращен в международный депозит по условиям Будапештского договора 1 ноября 1994, и ему присвоен номер доступа FERM BP-4859. RSFD80 может быть получена из штамма AJ12396 известным способом.
Примерами L-лизин-продуцирующих бактерий и родительских штаммов, которые могут быть использованы для получения таких бактерий, также являются штаммы, которые обладают пониженной активностью или у которых устранена активность фермента, который катализирует реакцию образования другого соединения, отличного от L-лизина, в результате ответвления от пути биосинтеза L-лизина. Примеры ферментов, которые катализируют реакцию образования другого соединения, отличного от L-лизина, в результате ответвления от пути биосинтеза L-лизина, включают гомосериндегидрогеназу, лизиндекарбоксилазу (патент США No. 5827698) и декарбоксилирующую малатдегидрогеназу (WO2005/010175). Чтобы снизить или исключить активность лизиндекарбоксилазы, предпочтительно уменьшить экспрессию гена cadA и гена ldcC, кодирующего лизиндекарбоксилазу (международная публикация WO2006/038695).
Бактерии, продуцирующие L-цистеин
Примеры бактерий, продуцирующих L-цистеин, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы, относящихся к роду Escherichia, такие как E. coli JM15, который трансформирован разными аллелями cysE, кодирующими серинацетилтрансферазы, резистентные к ингибированию по механизму обратной связи (патент США No. 6218168, заявка на выдачу патента РФ No. 2003121601); E. coli W3110, имеющий сверхэкпсрессируемые гены, которые кодируют белки, подходящие для секреции веществ, токсичных для клеток (патент США No. 5972663); штаммы E. coli, обладающие пониженной активностью цистеиндесульфогидразы (выложенный патент Японии No. 11-155571); и E. coli W3110 с повышенной активностью позитивного регулятора транскрипции цистеинового регулона, кодируемого геном cysB (WO01/27307).
Бактерии, продуцирующие L-лейцин
Примеры бактерий, продуцирующих L-лейцин, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы, относящиеся к роду Escherichia, такие как штаммы E. coli, резистентные к лейцину (например, штамм 57 (VKPM B-7386, патент США No. 6124121)) или аналогам лейцина, включая β-2-тиенилаланин, 3-гидроксилейцин, 4-азалейцин и 5,5,5-трифторлейцин (публикация патента Японии (Kokoku) No. 62-34397 и выложенный патент Японии No. 8-70879); штаммы E. coli, полученные способом генной инженерии, описанным в WO96/06926; и E. coli H-9068 (выложенный патент Японии No. 8-70879).
Бактерия, используемая для настоящего изобретения, может быть улучшена благодаря усилению экспрессии одного или нескольких генов, вовлеченных в биосинтез L-лейцина. Предпочтительными примерами таких генов являются гены оперона leuABCD, типичным примером которых является мутантный ген leuA, кодирующий изопропилмалатсинтазу с пониженной чувствительностью к ингибированию L-лейцином по механизму обратной связи (патент США No. 6403342). Кроме того, бактерия, используемая для настоящего изобретения, может быть улучшена посредством усиления экспрессии одного или нескольких генов, кодирующих белки, которые экскретируют L-аминокислоту их бактериальных клеток. Примерами таких генов являются гены b2682 и b2683 (гены ygaZH) (EP 1239041 A2).
Бактерии, продуцирующие L-гистидин
Примеры бактерий, продуцирующих L-гистидин, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы, относящиеся к роду Escherichia, такие как штамм 24 E. coli (VKPM B-5945, РФ 2003677); штамм 80 E. coli (VKPM B-7270, РФ 2119536); E. coli NRRL B-12116 - B12121 (патент США No. 4388405); E. coli H-9342 (FERM BP-6675) и H-9343 (FERM BP-6676) (патент США No. 6344347); E. coli H-9341 (FERM BP-6674) (EP 1085087) и E. coli AI80/pFM201 (патент США No. 6258554).
Примерами продуцирующих L-гистидин бактерий и родительских штаммов, которые могут быть использованы для получения таких бактерий, также включают штаммы, в которых экспрессия одного или нескольких генов, кодирующих фермент биосинтеза L-гистидина, повышена. Примерами таких генов являются ген АТФ-фосфорибозилтрансферазы (hisG), ген фосфорибозил-АМФ-циклогидролазы (hisI), ген фосфорибозил-АТФ-пирофосфогидролазы (hisI), ген фосфорибозилформимино-5-аминоимидазолкарбоксамидриботидизомеразы (hisA), ген амидотрансферазы (hisH), ген гистидинолфосфатаминотрнасферазы (hisC), ген гистидинолфосфатазы (hisB), ген гистидинолдегидрогеназы (hisD), и т.д.
Известно, что ферменты биосинтеза L-гистидина, кодируемые hisG и hisBHAFI, ингибируются L-гистидином, и поэтому способность продуцировать L-гистидин также может быть эффективно повышена при введении мутации, которая придает резистентность к ингибированию по механизму обратной связи гену АТФ-фосфорибозилтрансферазы (hisG) (патенты РФ No. 2003677 и 2119536).
Конкретными примерами штаммов, обладающих способностью продуцировать L-гистидин, являются штаммы E. coli FERM-P 5038 и 5048, в которые введен вектор, несущий ДНК, кодирующую фермент биосинтеза L-гистидина (выложенный патент Японии No. 56-005099), штаммы E. coli, в которые введен ген для экспорта аминокислот (EP 1016710 A), штамм 80 E. coli, которому придана резистентность к сульфагуанидину, DL-1,2,4-триазол-3-аланину и стрептомицину (VKPM B-7270, патент РФ No. 2119536), и т.д.
Бактерии, продуцирующие L-глутаминовую кислоту
Примеры бактерий, продуцирующих L-глутаминовую кислоту, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы относящиеся к роду Escherichia, такие как E. coli VL334thrC+ (EP 1172433). E. coli VL334 (VKPM B-1641) является ауксотрофным по L-изолейцину и L-треонину штаммом, имеющим мутации в генах thrC и ilvA (патент США No. 4278765). Аллель дикого типа гена thrC переносили способом общей трансдукции с использованием бактериофага P1, выращиваемого на клетках штамма E. coli K12 дикого типа (VKPM B-7). В результате получали ауксотрофный по L-изолейцину штамм VL334thrC+ (VKPM B-8961), продуцирующий L-глутаминовую кислоту.
Примеры бактерий, продуцирующих L-глутаминовую кислоту, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы, в которых экспрессия одного или нескольких генов, кодирующих фермент биосинтеза L-глутаминовой кислоты, повышена. Примеры таких генов включают гены, кодирующие глутаматдегидрогеназу (gdhA), глутаминсинтетазу (glnA), глутаматсинтетазу (gltAB), изоцитратдегидрогеназу (icdA), аконитатгидратазу (acnA, acnB), цитратсинтазу (gltA), фосфоенолпируваткарбоксилазу (ppc), пируватдегидрогеназу (aceEF, lpdA), пируваткиназу (pykA, pykF), фосфоенолпируватсинтазу (ppsA), енолазу (eno), фосфоглицеромутазу (pgmA, pgmI), фосфоглицераткиназу (pgk), глицеральдегид-3-фосфатдегидрогеназу (gapA), триозофосфатизомеразу (tpiA), фруктозобисфосфатальдолазу (fbp), фосфофруктокиназу (pfkA, pfkB), глюкозофосфатизомеразу (pgi) и другие.
Примеры штаммов, модифицированных так, чтобы повысить экспрессию гена цитратсинтетазы, гена фосфоенолпируваткарбоксилазы и/или гена глутаматдегидрогеназы, включают штаммы, раскрытые в EP 1078989 A, EP 955368 A и EP 952221 A.
Примеры бактерий, продуцирующих L-глутаминовую кислоту, и родительских штаммов, которые могут быть использованы для получения таких бактерий, также включают штаммы, в которых понижена или устранена активность фермента, который катализирует синтез другого соединения, отличного от L-глутаминовой кислоты, в результате ответвления от пути биосинтеза L-глутаминовой кислоты. Примерами таких ферментов являются изоцитратлиаза (aceA), α-кетоглутаратдегидрогеназа (sucA), фосфотрансацетилаза (pta), ацетаткиназа (ack), синтаза ацетогидроксикислоты (ilvG), ацетолактатсинтаза (ilvI), формиатацетилтрансфераза (pfl), лактатдегидрогеназа (ldh), глутаматдекарбоксилаза (gadAB) и другие. Бактерии, относящиеся к роду Escherichia, дефицитные по активности α-кетоглутаратдегидрогеназы или имеющие пониженную активность α-кетоглутаратдегидрогеназы, и способы их получения описаны в патентах США No. 5378616 и 5573945.
Конкретными примерами таких штаммов являются следующие штаммы:
E. coli W3110sucA::Kmr
E. coli AJ12624 (FERM BP-3853)
E. coli AJ12628 (FERM BP-3854)
E. coli AJ12949 (FERM BP-4881)
E. coli W3110sucA::Kmr представляют собой штамм, полученный в результате нарушения гена α-кетоглутаратдегидрогеназы (также называемого далее «геном sucA») of E. coli W3110. Указанный штамм полностью лишен α-кетоглутаратдегидрогеназы.
Другими примерами бактерий, продуцирующих L-глутаминовую кислоту, являются бактерии, которые относятся к роду Escherichia и обладают резистентностью к антиметаболиту аспарагиновой кислоты. Такие штаммы также могут быть дефицитными по α-кетоглутаратдегидрогеназе, и примеры включают, например, штамм E. coli AJ13199 (FERM BP-5807) (патент США No. 5908768), FFRM P-12379), который дополнительно имеет пониженную способность к разложению L-глутаминовой кислоты (патент США No. 5393671); AJ13138 (FERM BP-5565) (патент США No. 6110714), и т.д.
Примерами бактерий, продуцирующих L-глутаминовую кислоту, являются мутантные штаммы, относящиеся к роду Pantoea, которые дефицитны по активности α-кетоглутаратдегидрогеназы или имеют пониженную активность α-кетоглутаратдегидрогеназы, и такие бактерии могут быть получены, как описано выше. Такие штаммы включают Pantoea ananatis AJ13356 (патент США No. 6331419). Штамм Pantoea ananatis AJ13356 депонирован в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology (в настоящее время National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary, Central 6, 1-1, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan) 19 февраля 1998 с номером доступа FERM P-16645. Затем он был превращен в международный депозит по условиям Будапештского соглашения 11 января 1999, и ему присвоен номер доступа FERM BP-6616. Штамм Pantoea ananatis AJ13356 имеет дефицит активности α-кетоглутаратдегидрогеназы в результате нарушения гена субъединицы αKGDH-E1 (sucA). Указанный штамм идентифицирован как Enterobacter agglomerans при его выделении и депонирован как Enterobacter agglomerans AJ13356. Однако недавно штамм заново классифицировали как Pantoea ananatis на основе нуклеотидного секвенирования рРНК 16S и другим признакам. Хотя AJ13356 депонирован в указанном выше депозитарии как Enterobacter agglomerans, он описан в настоящей публикации как Pantoea ananatis.
Бактерии, продуцирующие L-фенилаланин
Примеры бактерий, продуцирующих L-фенилаланин, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы, относящиеся к роду Escherichia, такие как E. coli AJ12739 (tyrA::Tn10, tyrR) (VKPM B-8197); E. coli HW1089 (ATCC 55371), несущий мутантный ген pheA34 (патент США No. 5354672); E. coli MWEC101-b (KR 8903681); E. coli NRRL B-12141, NRRL B-12145, NRRL B-12146 и NRRL B-12147 (патент США No. 4407952). В качестве родительских штаммов также можно использовать E. coli K-12 [W3110 (tyrA)/pPHAB] (FERM BP-3566), E. coli K-12 [W3110 (tyrA)/pPHAD] (FERM BP-12659), E. coli K-12 [W3110 (tyrA)/pPHATerm] (FERM BP-12662) и E. coli K-12 [W3110 (tyrA)/pBR-aroG4, pACMAB], названный AJ12604 (FERM BP-3579) (EP 488424 B1). Кроме того, также можно использовать бактерии, продуцирующие L-фенилаланин, относящиеся к роду Escherichia, с повышенной активностью белка, кодируемого геном yedA или геном yddG (опубликованные заявки на выдачу патента США No. 2003/0148473 A1 и 2003/0157667 A1).
Бактерии, продуцирующие L-триптофан
Примеры бактерий, продуцирующих триптофан, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы, относящиеся к роду Escherichia, такие как E. coli JP4735/pMU3028 (DSM10122) и JP6015/pMU91 (DSM10123), с дефицитом триптофанил-тРНК-синтетазы, кодируемой мутантным геном trpS (патент США No. 5756345); E. coli SV164 (pGH5), имеющий аллель serA, кодирующий фосфоглицератдегидрогеназу, не чувствительную к ингибированию серином по механизму обратной связи, и аллель trpE, кодирующий антранилатсинтазу, не чувствительную к ингибированию триптофаном по механизму обратной связи (патент США No. 6180373); E. coli AGX17(pGX44) (NRRL B-12263) и AGX6(pGX50)aroP (NRRL B-12264) с дефицитом триптофаназы (патент США No. 4371614); и штамм E. coli AGX17/pGX50,pACKG4-pps, в котором повышена способность продуцировать фосфоенолпируват (WO97/08333, патент США No. 6319696). Также можно использовать продуцирующие L-триптофан бактерии, относящиеся к роду Escherichia, с повышенной активностью белка, кодируемого геном yedA или геном yddG (опубликованные заявки на выдачу патента США No. 2003/0148473 A1 и 2003/0157667 A1).
Примерами бактерий, продуцирующих L-триптофан, и родительских штаммов, которые могут быть использованы для получения таких бактерий, также являются штаммы, в которых одна или несколько активностей ферментов, выбранных из антранилатсинтазы (trpE), фосфоглицератдегидрогеназы (serA) и триптофансинтазы (trpAB), повышены. Антранилатсинтаза и фосфоглицератдегидрогеназа подвергаются ингибированию по механизму обратной связи L-триптофаном и L-серином, и поэтому в такие ферменты может быть введена мутация, вызывающая снижение их чувствительности к ингибированию по механизму обратной связи. Конкретными примерами штаммов, имеющих такую мутацию, являются штамм E. coli SV164, который имеет десенсибилизированную антранилатсинтазу, и трансформированный штамм, полученный введением плазмиды pGH5 (WO94/08031), которая содержит мутантный ген serA, кодирующий фосфоглицератдегидрогеназу с пониженной чувствительностью к ингибированию по механизму обратной связи, в E. coli SV164.
Примерами L-триптофан-продуцирующих бактерий и родительских штаммов, которые могут быть использованы для получения таких бактерий, также являются штаммы, в которые введен триптофановый оперон, содержащий ген, кодирующий антранилатсинтазу с пониженной чувствительностью к ингибированию (выложенные патенты Японии No. 57-71397, 62-244382, патент США No. 4371614). Кроме того, способность продуцировать L-триптофан можно придать посредством увеличения экспрессии гена, который кодирует триптофансинтазу в триптофановом опероне (trpBA). Триптофансинтаза состоит из α- и β-субъединиц, которые кодируются генами trpA и trpB соответственно. Кроме того, способность продуцировать L-триптофан также может быть повышена в результате увеличения экспрессии оперона изоцитратлиазы-малатсинтазы (WO2005/103275).
Бактерии, продуцирующие L-пролин
Примеры бактерий, продуцирующих L-пролин, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы, относящиеся к роду Escherichia, такие как E. coli 702ilvA (VKPM B-8012), который дефицитен по гену ilvA и способен продуцировать L-пролин (EP 1172433).
Бактерия, используемая для настоящего изобретения, может быть улучшена посредством усиления экспрессии одного или нескольких генов, вовлеченных в биосинтез L-пролина. Предпочтительным примером таких генов является ген proB, кодирующий глутаматкиназу, десенсибилизированную в отношении ингибирования L-пролином по механизму обратной связи (DE 3127361). Кроме того, бактерия, используемая для настоящего изобретения, может быть улучшена посредством усиления экспрессии одного или нескольких генов, кодирующих белки, экскретирующие L-аминокислоту из бактериальных клеток. Примерами таких генов являются гены b2682 и b2683 (гены ygaZH) (EP 1239041 A2).
Примеры бактерий, относящихся к роду Escherichia и обладающих способностью продуцировать L-пролин, включают следующие штаммы E. coli штаммы: NRRL B-12403 и NRRL B-12404 (патент Великобритании No. 2075056), VKPM B-8012 (заявка на выдачу патента РФ No. 2000124295), плазмидные мутанты, описанные в патенте Германии No. 3127361, плазмидные мутанты, описанные Bloom F.R. et al. (The 15th Miami winter symposium, 1983, p. 34), и другие.
Бактерии, продуцирующие L-аргинин
Примеры бактерий, продуцирующих L-аргинин, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы, относящиеся к роду Escherichia, такие как штамм E. coli 237 (VKPM B-7925) (опубликованная заявка на выдачу патента США 2002/058315A1) и его производные, несущие мутантную N-ацетилглутаматсинтазу (заявка на выдачу патента РФ No. 2001112869), штамм E. coli 382 (VKPM B-7926) (EP 1170358 A1) и аргинин-продуцирующий штамм, в который введен ген argA, кодирующий N-ацетилглутаматсинтетазу (EP 1170361 A1).
Примерами L-аргинин-продуцирующих бактерий и родительских штаммов, которые могут быть использованы для получения таких бактерий, также являются штаммы, в которых экспрессия одного или нескольких генов, кодирующих фермент биосинтеза L-аргинина, повышена. Примерами таких генов являются ген N-ацетилглутамилфосфатредуктазы (argC), ген орнитинацетилтрансферазы (argJ), ген N-ацетилглутаматкиназы (argB), ген ацетилорнитинтрансаминазы (argD), ген орнитинкарбамоилтрансферазы (argF), ген синтетазы аргининянтарной кислоты (argG), ген лиазы аргининянтарной кислоты (argH) и ген карбамоилфосфатсинтетазы (carAB).
Бактерии, продуцирующие L-валин
Примеры бактерий, продуцирующих L-валин, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения штаммы, которые были модифицированы так, что они сверхэкспрессируют оперон ilvGMEDA (патент США No. 5998178). Желательно удаление области оперона ilvGMEDA, которая необходима для ослабления, чтобы не происходило ослабление экспрессии продуцируемым L-валином. Кроме того, предпочтительно нарушение гена ilvA в опероне, чтобы была снижена активность треониндезаминазы.
Примерами L-валин-продуцирующих бактерий и родительских штаммов, которые могут быть использованы для получения таких бактерий, также являются мутанты, имеющие мутацию аминоацил-тРНК-синтетазы (патент США No. 5658766). Например, можно использовать штамм E. coli VL1970, который имеет мутацию в гене ileS, кодирующем изолейцин-тРНК-синтетазу. E. coli VL1970 депонирована во Всероссийской национальной коллекции промышленных микроорганизмов (ВКПМ) (1 Дорожный проезд, 1 Москва 117545, Россия) 24 июня 1988 с номером доступа VKPM B-4411.
Кроме того, мутанты, требующие для роста липоевую кислоту и/или не имеющие H+-АТФазы (WO96/06926), также можно использовать в качестве родительских штаммов.
Бактерии, продуцирующие L-изолейцин
Примеры бактерий, продуцирующих L-изолейцин, и родительских штаммов, которые могут быть использованы для получения таких бактерий, включают без ограничения мутанты, обладающие резистентностью к 6-диметиламинопурину (выложенный патент Японии No. 5-304969 A), мутанты, обладающие резистентностью к аналогу изолейцина, такому как тиаизолейцин и изолейцингидроксамат, и такие мутанты, дополнительно обладающие резистентностью к DL-этионину и/или аргинингидроксамату (выложенный патент Японии No. 5-130882). Кроме того, рекомбинантные штаммы, трансформированные генами, кодирующими белки, вовлеченные в биосинтез L-изолейцина, такие как треониндезаминаза и ацетогидроксатсинтаза, также можно использовать в качестве родительских штаммов (выложенный патент Японии No. 2-458, FR 0356739, и патент США No. 5998178).
Примеры
[Пример 1] Конструирование бактерии, продуцирующей L-лизин, с повышенными активностями фруктозо-6-фосфатальдолазы, глицериндегидрогеназы и дигидроксиацетонкиназы
<1-1> Конструирование плазмиды для экспрессии гена dak1
Общая нуклеотидная последовательность хромосомы Saccharomyces cerevisiae уже определена (Science, 25 (1996)). На основе нуклеотидной последовательности гена dak1, описанной в указанном литературном источнике, получали синтетический олигонуклеотид с последовательностью SEQ ID NO: 14 в качестве 5'-праймера и получали синтетический олигонуклеотид с последовательностью SEQ ID NO: 15 в качестве 3'-праймера. ПЦР осуществляли, используя полученные синтетические олигонуклеотиды и хромосомную ДНК штамма Saccharomyces cerevisiae JCM7255 в качестве матрицы. Продукт ПЦР очищали и лигировали в вектор pMW119 (Takara Bio), расщепленный HindIII и SalI, конструируя плазмиду для экспрессии dak1, названную pMW-dak1. Штамм JCM7255 хранится в независимом административном ведомстве, RIKEN, «Japan Collection of Microorganisms», 2-1, Hirosawa, Wako-shi, Saitama-ken.
<1-2> Конструирование штамма с повышенной активностью глицериндегидрогеназы
Конструировали штамм WC196ΔcadAΔldcC, модифицированный так, чтобы он имел структуру, показанную в последовательности SEQ ID NO: 11. Для конструирования штамма, имеющего такую структуру, использовали последовательность SEQ ID NO: 9 (продукт ПЦР). В последовательности SEQ ID NO: 9 последовательность нуклеотидов с номерами 1-172 представляет собой последовательность attR фага λ phage, последовательность нуклеотидов с номерами 324-983 представляет собой ген резистентности к хлорамфениколу (cat), последовательность нуклеотидов с номерами 1540-1653 является последовательностью attL фага λ, и последовательность нуклеотидов с номерами 1654-1733 представляет собой промотор tacM.
Промотор tacM (SEQ ID NO: 10) может быть сконструирован заменой последовательности TTGACA промотора tac (Gene, 25 (2-3), 167-178 (1983)) в области -35 последовательностью TTCACA. Последовательность SEQ ID NO: 9 может быть сконструирована в соответствии с конструированием pMW118-attL-Cm-attR (WO2005/010175).
Последовательность SEQ ID NO: 9 в качестве матрицы амплифицировали в ПЦР, используя праймеры с последовательностями SEQ ID NO: 12 и 13, и полученный продукт амплификации встраивали в хромосому штамма WC196ΔcadAΔldcC (смотри международную публикацию WO2006/038695) способом λ-RED (WO2005/010175), чтобы сконструировать штамм, в котором промоторная последовательность выше gldA заменена. Таким образом получили штамм с повышенной активностью глицериндегидрогеназы, WC196ΔcadAΔldcCPtacMgldA::Cm штамм.
<1-3> Конструирование бактерии, продуцирующей L-лизин, с повышенными активностями фруктозо-6-фосфатальдолазы и глицериндегидрогеназы
Конструировали штамм WC196ΔcadAΔldcC, модифицированный так, чтобы он имел структуру, показанную в последовательности SEQ ID NO: 92. Для конструирования штамма, имеющего такую структуру, использовали последовательность SEQ ID NO: 9 (продукт ПЦР). В последовательности SEQ ID NO: 9 последовательность нуклеотидов с номерами 1-172 представляет собой последовательность attR фага λ, последовательность нуклеотидов с номерами 324-983 представляет собой ген резистентности к хлорамфениколу (cat), последовательность нуклеотидов с номерами 1540-1653 является последовательностью attL фага λ и последовательность нуклеотидов с номерами 1654-1733 является промотором tacM.
Промотор tacM (SEQ ID NO: 10) может быть сконструирован заменой последовательности TTGACA промотора tac (Gene, 25 (2-3), 167-178 (1983)) в области -35 последовательностью TTCACA. Последовательность SEQ ID NO: 9 может быть сконструирована в соответствии с конструированием pMW118-attL-Cm-attR (WO2005/010175).
Последовательность SEQ ID NO: 9 в качестве матрицы амплифицировали в ПЦР, используя праймеры с последовательностями SEQ ID NO: 93 и 94, и полученный продукт амплификации встраивали в хромосому штамма WC196ΔcadAΔldcC (смотри международную публикацию WO2006/038695) способом λ-RED (WO2005/010175), чтобы сконструировать штамм, в котором промоторная последовательность выше оперона fsaB-gldA, заменена. Таким образом получали штамм с повышенными активностями фруктозо-6-фосфатальдолазы и глицериндегидрогеназы, штамм WC196ΔcadAΔldcCPtacM fsaB-gldA::Cm.
<1-4> Конструирование бактерий, продуцирующих L-лизин, с повышенными активностями фруктозо-6-фосфатальдолазы, глицериндегидрогеназы и дигидроксиацетонкиназы
Штамм WC196ΔcadAΔldcC (смотри международную публикацию WO2006/038695), штамм WC196ΔcadAΔldcCPtacMgldA::Cm и штамм WC196ΔcadAΔldcCPtacM fsaB-gldA::Cm трансформировали плазмидой pCABD2 для продуцирования Lys, несущей гены dapA, dapB и lysC (Международная публикация WO01/53459) обычным образом, чтобы получить штамм WC196ΔcadAΔldcC/pCABD2, штамм WC196ΔcadAΔldcCPtacMgldA::Cm/pCABD2 и штамм WC196ΔcadAΔldcCPtacM fsaB-gldA::Cm/pCABD2. Кроме того, штамм WC196ΔcadAΔldcC/pCABD2, штамм WC196ΔcadAΔldcCPtacMgldA::Cm/pCABD2 и штамм WC196ΔcadAΔldcCPtacM fsaB-gldA::Cm/pCABD2 трансформировали экспрессирующей dak1 плазмидой pMW-dak1 обычным способом, чтобы получить штамм WC196ΔcadAΔldcC/pCABD2,pMW-dak1, штамм WC196ΔcadAΔldcCPtacMgldA::Cm/pCABD2,pMW-dak1 и штамм WC196ΔcadAΔldcCPtacM fsaB-gldA::Cm/pCABD2,pMW-dak1.
Каждый из полученных штаммов культивировали в среде L, содержащей 20 мг/л стрептомицина или 20 мг/л стрептомицина и 50 мг/л ампициллина, при 37°C до достижения конечного значения OD600 около 0,6, затем в каждую культуральную среду добавляли 40% раствор глицерина в объеме, равном объему культуральной среды, и смесь перемешивали, затем делили на подходящие объемы и хранили при -80°C. Такие культуры называли исходными глицериновыми культурами.
[Пример 2] Оценка бактерий, продуцирующих L-лизин, с повышенными активностями фруктозо-6-фосфатальдолазы, глицериндегидрогеназы и дигидроксиацетонкиназы
Указанные выше исходные глицериновые культуры штаммов размораживали, по 100 мкл каждой исходной культуры равномерно наносили на L-чашку, содержащую 20 мг/л стрептомицина или 20 мг/л стрептомицина и 50 мг/л ампициллина, и проводили культивирование при 37°C в течение 24 часов. Полученные клетки на чашке суспендировали в 1 мл физиологического раствора соли, суспензию инокулировали в объеме (V), рассчитываемом делением константы 50 на оптическую плотность при 600 нм (n) суспензии, разбавленной в 101 раз (V=50/n) в 20 мл среды для ферментации, содержащей 20 мг/л стрептомицина или 20 мг/л стрептомицина и 50 мг/л ампициллина, находящейся в колбе Сакагучи объемом 500 мл, и культивирование осуществляли при 37°C в течение 48 часов на качалке для культивирования с возвратно-поступательным движением. После культивирования количество лизина, накопленного в среде, измеряли известным способом (Biotec Analyzer AS210, SAKURA SEIKI).
Состав среды для ферментации показан ниже (единицы: г/л).
Значение pH среды доводили до pH 7,0, используя KOH, и автоклавировали при 115°C в течение 10 минут при условии, что глицерин и MgSO4·7H2O стерилизовали отдельно, и добавляли 30 г/л CaCO3 Japanese Pharmacopoeia, подвергнутый стерилизации горячим воздухом при 180°C в течение 2 часов.
В качестве антибиотиков добавляли 20 мг/л стрептомицина или 20 мг/л стрептомицина и 50 мг/л ампициллина. Культивирование осуществляли в условиях температуры 37°C и перемешивания при 115 об/мин в течение 48 часов.
Результаты показаны в таблице 5 (OD означает оптическую плотность при 600 нм, отображающую количество клеток, Lys (г/л) означает количество L-лизина, накопленного в колбе, и выход (%) означает выход L-лизина на основе субстрата). В то время как в штамме, в котором повышена активность только глицериндегидрогеназы, штамме, в котором повышена активность только дигидроксиацетонкиназы, и штамме, в котором повышены активности фруктозо-6-фосфатальдолазы и глицериндегидрогеназы, не обнаружено изменения выхода и продуктивности по сравнению с немодифицированным штаммом, в штамме WC196ΔcadAΔldcCPtacMgldA::Cm/pCABD2,pMW-dak1, в котором были повышены активности и глицериндегидрогеназы и дигидроксиацетонкиназы, использующей АТФ в качестве донора фосфата, накапливалось большее количество L-лизина по сравнению с другими штаммами. Кроме того, в случае штамма WC196ΔcadAΔldcCPtacM fsaB-gldA::Cm/pCABD2,pMW-dak1, в котором повышены активности фруктозо-6-фосфатальдолазы, глицериндегидрогеназы и дигидроксиацетонкиназы, использующей АТФ в качестве донора фосфата, накапливало еще большее количество L-лизина.
[Таблица 5]
Накопление L-лизина штаммами с повышенными активностями фруктозо-6-фосфатальдолазы (fsaB), глицериндегидрогеназы (gldA) и дигидроксиацетонкиназы (dak1)

В названиях штаммов, указанных в таблице, «LC» является сокращением «ΔcadAΔldcC» и «::Cm» не указано.
[Пример 3] Конструирование бактерий, продуцирующих L-треонин, с повышенными активностями глицериндегидрогеназы и дигидроксиацетонкиназы
<3-1> Конструирование штамма с повышенной активностью глицериндегидрогеназы
Конструировали штаммы B5318, модифицированные так, чтобы они имели структуры, показанные в последовательностях SEQ ID NO: 90 и 91. Для конструирования штаммов, имеющих такие структуры, использовали последовательности SEQ ID NO: 88 и 89 (продукты ПЦР). В последовательностях SEQ ID NO: 88 и 89, последовательности нуклеотидов с номерами 1-72 представляют собой последовательности attR фага λ, последовательности нуклеотидов с номерами 324-983 представляют собой гены резистентности к хлорамфениколу (cat), последовательности нуклеотидов с номерами 1540-1653 являются последовательностями attL фага λ и последовательности нуклеотидов с номерами 1654-1733 представляют собой промоторы tacM2 и tacM3.
Промоторы tacM2 и tacM3 являются конститутивными промоторами, которые могут быть сконструированы заменой последовательности TTGACA промотора tac (Gene, 25 (2-3), 167-178 (1983)) в области -35 последовательностями TGTACA и TTGGCA (Molecular Biology 39 (5) 719-726 (2005)). Последовательности SEQ ID NO: 88 и 89 могут быть сконструированы в соответствии с конструированием pMW118-attL-Cm-attR (WO2005/010175).
Последовательности SEQ ID NO: 88 и 89 в качестве матриц амплифицировали в ПЦР, используя праймеры с последовательностью SEQ ID NO: 12 и 13, и каждый из полученных продуктов амплификации встраивали в хромосому штамма B5318 (VKPM B-5318) способом λ-RED (WO2005/010175), чтобы получить штаммы, в которых промоторная последовательность выше gldA заменена. Таким образом получали штаммы с повышенной активностью глицериндегидрогеназы, штамм B5318PtacM2gldA::Cm и штамм B5318PtacM3gldA::Cm.
<1-3> Конструирование бактерий, продуцирующих L-треонин, с повышенными активностями глицериндегидрогеназы и дигидроксиацетонкиназы
Штамм B5318PtacM2gldA::Cm и штамм B5318PtacM3gldA::Cm трансформировали экспрессирующей dak1 плазмидой pMW-dak1 обычным способом, получая штамм B5318PtacM2gldA::Cm/pMW-dak1 и штамм B5318PtacM3gldA::Cm/pMW-dak1.
Каждый из полученных штаммов культивировали в среде L, содержащей 20 мг/л стрептомицина или 20 мг/л стрептомицина и 50 мг/л ампициллина, при 37°C до достижения конечного значения OD600 около 0,6, затем в каждую культуральную среду добавляли 40% раствор глицерина в объеме, равном объему культуральной среды, и смесь перемешивали, затем делили на подходящие объемы и хранили при
-80°C. Такие культуры называли исходными глицериновыми культурами.
[Пример 4] Оценка бактерий, продуцирующих L-треонин, с повышенными активностями глицериндегидрогеназы и дигидроксиацетонкиназы
Указанные выше исходные глицериновые культуры штаммов размораживали, по 100 мкл каждой исходной культуры равномерно наносили на L-чашку, содержащую 20 мг/л стрептомицина или 20 мг/л стрептомицина и 50 мг/л ампициллина, и проводили культивирование при 37°C в течение 24 часов. Полученные клетки на чашке суспендировали в 1 мл физиологического раствора соли, суспензию инокулировали в объеме (V), рассчитываемом делением константы 50 на оптическую плотность при 600 нм (n) суспензии, разбавленной в 101 раз (V=50/n) в 20 мл среды для ферментации, содержащей 20 мг/л стрептомицина или 20 мг/л стрептомицина и 50 мг/л ампициллина, находящейся в конической колбе с перегородкой объемом 500 мл, и культивирование осуществляли при 40°C в течение 24 часов на роторном устройстве для культивирования. После культивирования количество треонина, накопленного в среде, измеряли известным способом (колонка для жидкостной хроматографии Hitachi ODS-2 Column).
Состав среды для ферментации показан ниже (единицы: г/л).
Значение pH среды доводили до pH 7,0, используя KOH, и автоклавировали при 115°C в течение 10 минут при условии, что глицерин и MgSO4·7H2O стерилизовали отдельно, и добавляли 30 г/л CaCO3 Japanese Pharmacopoeia, подвергнутый стерилизации горячим воздухом при 180°C в течение 2 часов.
В качестве антибиотиков добавляли 20 мг/л стрептомицина или 20 мг/л стрептомицина и 50 мг/л ампициллина. Культивирование осуществляли в условиях температуры 40°C и перемешивания при 144 об/мин в течение 24 часов.
Результаты показаны в таблице 6 (OD означает оптическую плотность при 600 нм, отображающую количество клеток, Thr (г/л) означает количество L-треонина, накопленного в колбе, и выход (%) означает выход L-треонина на основе субстрата). В то время как в случае штамма, в котором повышена активность только глицериндегидрогеназы, не обнаружено изменения выхода и продуктивности по сравнению с немодифицированным штаммом, штамм B5318PtacM2gldA::Cm/pMW-dak1 и штамм B5318PtacM3gldA::Cm/pMW-dak1, в котором были повышены активности и глицериндегидрогеназы и дигидроксиацетонкиназы, использующей АТФ в качестве донора фосфата, накапливали большее количество L-треонина по сравнению с другими штаммами.
[Таблица 6]
Накопление L-треонина штаммами с повышенными активностями глицериндегидрогеназы (gldA) и дигидроксиацетонкиназы (dak1)
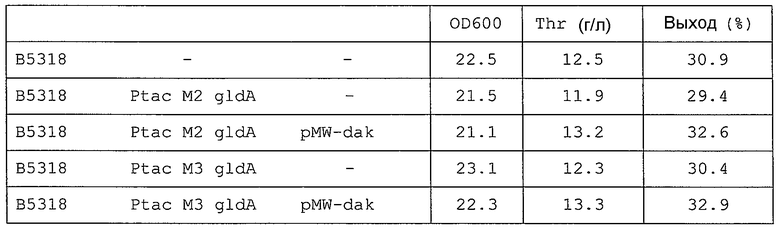
В названиях штаммов, указанных в таблице, «pMW-dak1» сокращено до «pMW-dak», и «::Cm» не указано.
[Пояснения к списку последовательностей]
SEQ ID NO: 1: последовательность гена gldA Escherichia coli (1104 п.н.)
SEQ ID NO: 2: аминокислотная последовательность GldA Escherichia coli (367 а/к)
SEQ ID NO: 3: последовательность гена dakA1 Saccharomyces cerevisiae (1755 п.н.)
SEQ ID NO: 4: аминокислотная последовательность DakA Saccharomyces cerevisiae (584 а/к)
SEQ ID NO: 5: последовательность гена dhbK1 Agrobacterium tumefaciens (1695 п.н.)
SEQ ID NO: 6: аминокислотная последовательность Dhbk1 Agrobacterium tumefaciens (564 а/к)
SEQ ID NO: 7: последовательность гена dhaK Citrobacter freundii (1659 п.н.)
SEQ ID NO: 8: аминокислотная последовательность DhaK Citrobacter freundii (552 а/к)
SEQ ID NO: 9: последовательность attR-cat-attL-ptacM-SD-спейсер (1740 п.н.)
SEQ ID NO: 10: промотор tacM (80 п.н.)
SEQ ID NO: 11: последовательность PtacMgldA::Cm
SEQ ID NO: 12: atL-Ptac-gldA (ПЦР-праймер для усиления gldA в хромосоме)
SEQ ID NO: 13: atR-Ptac-fsaB1 (ПЦР-праймер для усиления gldA в хромосоме)
SEQ ID NO: 14: pMW-dak1F (праймер для клонирования dakA)
SEQ ID NO: 15: pMW-dak1R (праймер для клонирования dakA)
SEQ ID NO: 16: последовательность гена glpF Escherichia coli (846 п.н.)
SEQ ID NO: 17: аминокислотная последовательность GlpF Escherichia coli (281 а/к)
SEQ ID NO: 18: последовательность гена tpiA Escherichia coli (768 п.н.)
SEQ ID NO: 19: аминокислотная последовательность TpiA Escherichia coli (255 а/к)
SEQ ID NO: 20: последовательность гена fbaA Escherichia coli (1080 п.н.)
SEQ ID NO: 21: аминокислотная последовательность FbaA Escherichia coli (359 а/к)
SEQ ID NO: 22: последовательность гена glpX Escherichia coli (1011 п.н.)
SEQ ID NO: 23: аминокислотная последовательность GlpX Escherichia coli (336 а/к)
SEQ ID NO: 24: последовательность гена glpK Escherichia coli (1509 п.н.)
SEQ ID NO: 25: аминокислотная последовательность GlpK Escherichia coli (502 а/к)
SEQ ID NO: 26: последовательность гена glpA Escherichia coli (1629 п.н.)
SEQ ID NO: 27: аминокислотная последовательность GlpA Escherichia coli (542 а/к)
SEQ ID NO: 28: последовательность гена glpB Escherichia coli (1260 п.н.)
SEQ ID NO: 29: аминокислотная последовательность GlpB Escherichia coli (419 а/к)
SEQ ID NO: 30: последовательность гена glpC Escherichia coli (1191 п.н.)
SEQ ID NO: 31: аминокислотная последовательность GlpC Escherichia coli (396 а/к)
SEQ ID NO: 32: последовательность гена glpD Escherichia coli (1506 п.н.)
SEQ ID NO: 33: аминокислотная последовательность GlpD Escherichia coli (501 а/к)
SEQ ID NO: 34: последовательность гена dhaK Escherichia coli (1071 п.н.)
SEQ ID NO: 35: аминокислотная последовательность DhaK Escherichia coli (356 а/к)
SEQ ID NO: 36: последовательность гена dhaL Escherichia coli (633 п.н.)
SEQ ID NO: 37: аминокислотная последовательность DhaL Escherichia coli (210 а/к)
SEQ ID NO: 38: последовательность гена dhaM Escherichia coli (1419 п.н.)
SEQ ID NO: 39: аминокислотная последовательность DhaM Escherichia coli (472 а/к)
SEQ ID NO: 40: ген дигидроксиацетонкиназы Schizosaccharomyces pombe (1776 п.н.)
SEQ ID NO: 41: дигидроксиацетонкиназа Schizosaccharomyces pombe (591 а/к)
SEQ ID NO: 42: ген дигидроксиацетонкиназы Pichia angusta (1830 п.н.)
SEQ ID NO: 43: дигидроксиацетонкиназа Pichia angusta (609 а/к)
SEQ ID NO: 44: ген дигидроксиацетонкиназы Pichia pastoris (1827 п.н.)
SEQ ID NO: 45: дигидроксиацетонкиназа Pichia pastoris (608 а/к)
SEQ ID NO: 46: ген дигидроксиацетонкиназы Debaryomyces hansenii (1824 п.н.)
SEQ ID NO: 47: дигидроксиацетонкиназа Debaryomyces hansenii (607 а/к)
SEQ ID NO: 48: ген дигидроксиацетонкиназы Escherichia blattae (1752 п.н.)
SEQ ID NO: 49: дигидроксиацетонкиназа Escherichia blattae (583 а/к)
SEQ ID NO: 50: ген дигидроксиацетонкиназы Enterobacter sp. 638 (1647 п.н.)
SEQ ID NO: 51: дигидроксиацетонкиназа Enterobacter sp. 638 (548 а/к)
SEQ ID NO: 52: ген дигидроксиацетонкиназы Psychromonas sp. CNPT3 (1695 п.н.)
SEQ ID NO: 53: дигидроксиацетонкиназа Psychromonas sp. CNPT3 (564 а/к)
SEQ ID NO: 54: ген дигидроксиацетонкиназы Stappia aggregata (1647 п.н.)
SEQ ID NO: 55: дигидроксиацетонкиназа Stappia aggregata (548 а/к)
SEQ ID NO: 56: ген дигидроксиацетонкиназы Rhizobium leguminosarum bv. viciae 3841 (1641 п.н.)
SEQ ID NO: 57: дигидроксиацетонкиназа Rhizobium leguminosarum bv. viciae 3841 (546 а/к)
SEQ ID NO: 58: ген дигидроксиацетонкиназы Myxococcus xanthus DK 1622 (1701 п.н.)
SEQ ID NO: 59: дигидроксиацетонкиназа Myxococcus xanthus DK 1622 (566 а/к)
SEQ ID NO: 60: ген дигидроксиацетонкиназы Burkholderia sp. 383 (1701 п.н.)
SEQ ID NO: 61: дигидроксиацетонкиназа Burkholderia sp. 383 (566 а/к)
SEQ ID NO: 62: ген дигидроксиацетонкиназы Burkholderia thailandensis E264 (1704 п.н.)
SEQ ID NO: 63: дигидроксиацетонкиназа Burkholderia thailandensis E264 (567 а/к)
SEQ ID NO: 64: ген дигидроксиацетонкиназы Burkholderia multivorans ATCC 17616 (1851 п.н.)
SEQ ID NO: 65: дигидроксиацетонкиназа Burkholderia multivorans ATCC 17616 (616 а/к)
SEQ ID NO: 66: ген dhaR Escherichia coli (1920 п.н.)
SEQ ID NO: 67: аминокислотная последовательность DhaR Escherichia coli (639 а/к)
SEQ ID NO: 68: ген fsaA Escherichia coli (663 п.н.)
SEQ ID NO: 69: аминокислотная последовательность FsaA Escherichia coli (220 а/к)
SEQ ID NO: 70: ген fsaB Escherichia coli (663 п.н.)
SEQ ID NO: 71: аминокислотная последовательность FsaB Escherichia coli (220 а/к)
SEQ ID NO: 72: ген fbaB Escherichia coli (1053 п.н.)
SEQ ID NO: 73: аминокислотная последовательность FbaB Escherichia coli (350 а/к)
SEQ ID NO: 74: ген gldA Shigella dysenteriae Sd197 (1143 п.н.)
SEQ ID NO: 75: аминокислотная последовательность GldA Shigella dysenteriae Sd197 (380 а/к)
SEQ ID NO: 76: ген gldA Salmonella typhimurium LT2 (1104 п.н.)
SEQ ID NO: 77: аминокислотная последовательность GldA Salmonella typhimurium LT2 (367 а/к)
SEQ ID NO: 78: ген gldA Pseudomonas putida (1098 п.н.)
SEQ ID NO: 79: аминокислотная последовательность GldA Pseudomonas putida (365 а/к)
SEQ ID NO: 80: ген gldA Bacillus coagulans 36D1 (1104 п.н.)
SEQ ID NO: 81: аминокислотная последовательность GldA Bacillus coagulans 36D1 (367 а/к)
SEQ ID NO: 82: ген fbp Escherichia coli (999 п.н.)
SEQ ID NO: 83: аминокислотная последовательность fbp Escherichia coli (322 а/к)
SEQ ID NO: 84: ген ybhA Escherichia coli (819 п.н.)
SEQ ID NO: 85: аминокислотная последовательность YbhA Escherichia coli (272 а/к)
SEQ ID NO: 86: ген ptsI Escherichia coli (1782 п.н.)
SEQ ID NO: 87: аминокислотная последовательность PtsI Escherichia coli (575 а/к)
SEQ ID NO: 88: последовательность attR-cat-attL-PtacM2-SD-спейсер
SEQ ID NO: 89: последовательность attR-cat-attL-PtacM3-SD-спейсер
SEQ ID NO: 90: последовательность PtacM2gldA::Cm
SEQ ID NO: 91: последовательность PtacM3gldA::Cm
SEQ ID NO: 92: последовательность PtacM fsaB-gldA::Cm
SEQ ID NO: 93: atL-Ptac-fsaB (ПЦР-праймер для усиления fsaB+gldA в хромосоме)
SEQ ID NO: 94: atR-Ptac-fsaB (ПЦР-праймер для усиления fsaB+gldA в хромосоме)
Промышленная применимость
Использование микроорганизма согласно настоящему изобретению обеспечивает возможность эффективного продуцирования L-аминокислоты из глицерина при ферментации.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |
| МУТАНТНАЯ АДЕНИЛАТЦИКЛАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ СЕМЕЙСТВА ENTEROBACTERIACEAE, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2010 |
|
RU2471868C2 |
| МИКРОБИОЛОГИЧЕСКИЙ СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА | 2009 |
|
RU2521502C2 |
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ ESCHERICHIA, - ПРОДУЦЕНТ L-ГИСТИДИНА И СПОСОБ ПОЛУЧЕНИЯ L-ГИСТИДИНА | 2003 |
|
RU2276687C2 |
| НОВЫЙ МИКРООРГАНИЗМ И СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА, ОСНОВАННЫЙ НА НАДФН-ЗАВИСИМОЙ АЦЕТОЛРЕДУКТАЗЕ И УЛУЧШЕННОЙ ДОСТАВКЕ НАДФН | 2015 |
|
RU2711870C2 |
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ ESCHERICHIA - ПРОДУЦЕНТ L-ТРЕОНИНА И СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 2004 |
|
RU2288264C2 |
| ГЕН ПЕПТИДООБРАЗУЮЩЕГО ФЕРМЕНТА, ПЕПТИДООБРАЗУЮЩИЙ ФЕРМЕНТ И СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА | 2002 |
|
RU2280077C2 |
| СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА (ВАРИАНТЫ) | 2005 |
|
RU2316596C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ L-ЛИЗИНА ИЛИ L-ТРЕОНИНА С ПОМОЩЬЮ БАКТЕРИЙ ESCHERICHIA, ИМЕЮЩИХ АТТЕНУИРОВАННУЮ АКТИВНОСТЬ МАЛЕИНОВОГО ФЕРМЕНТА | 2004 |
|
RU2337140C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |





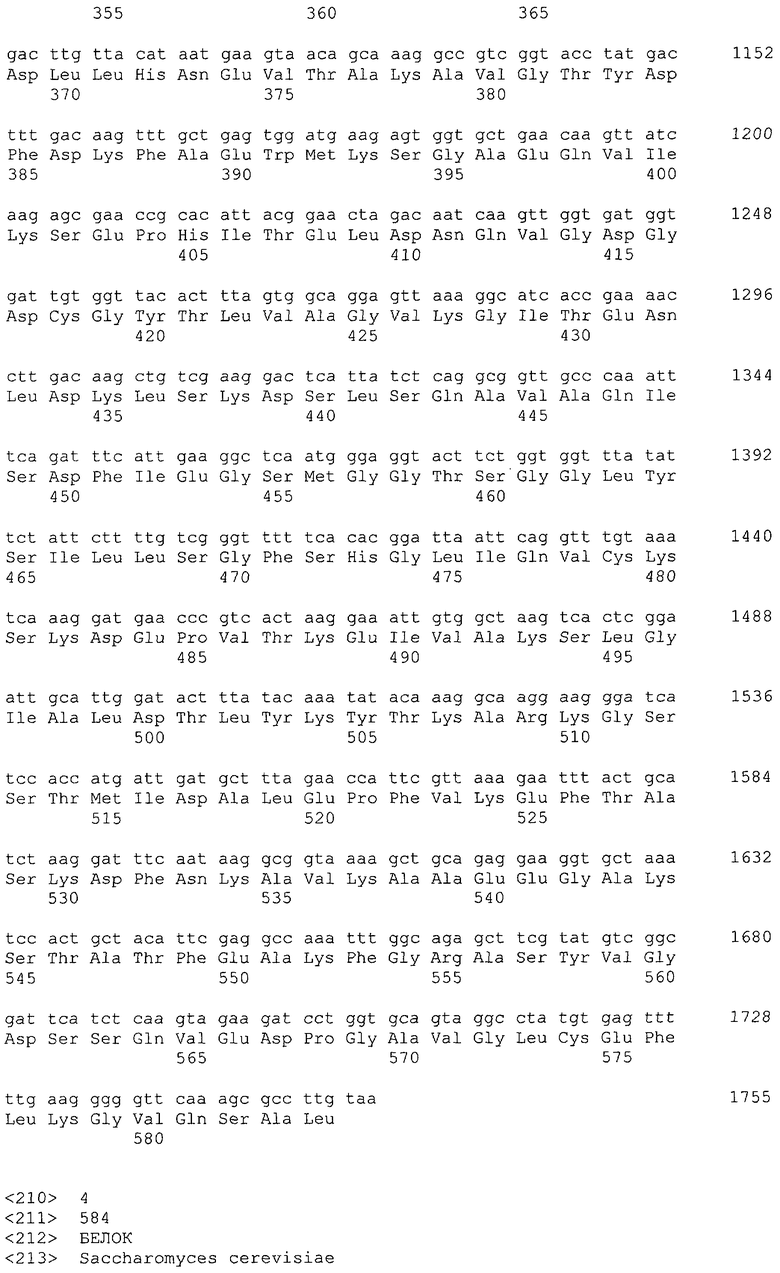








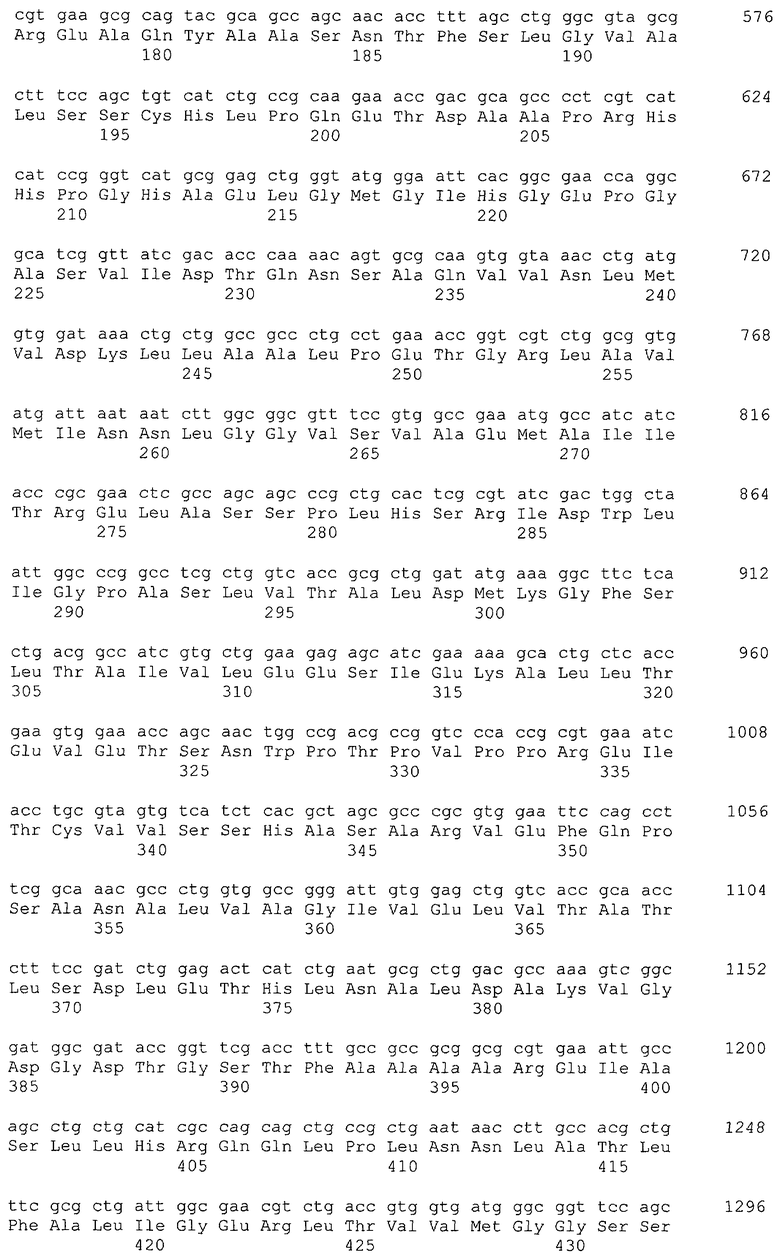







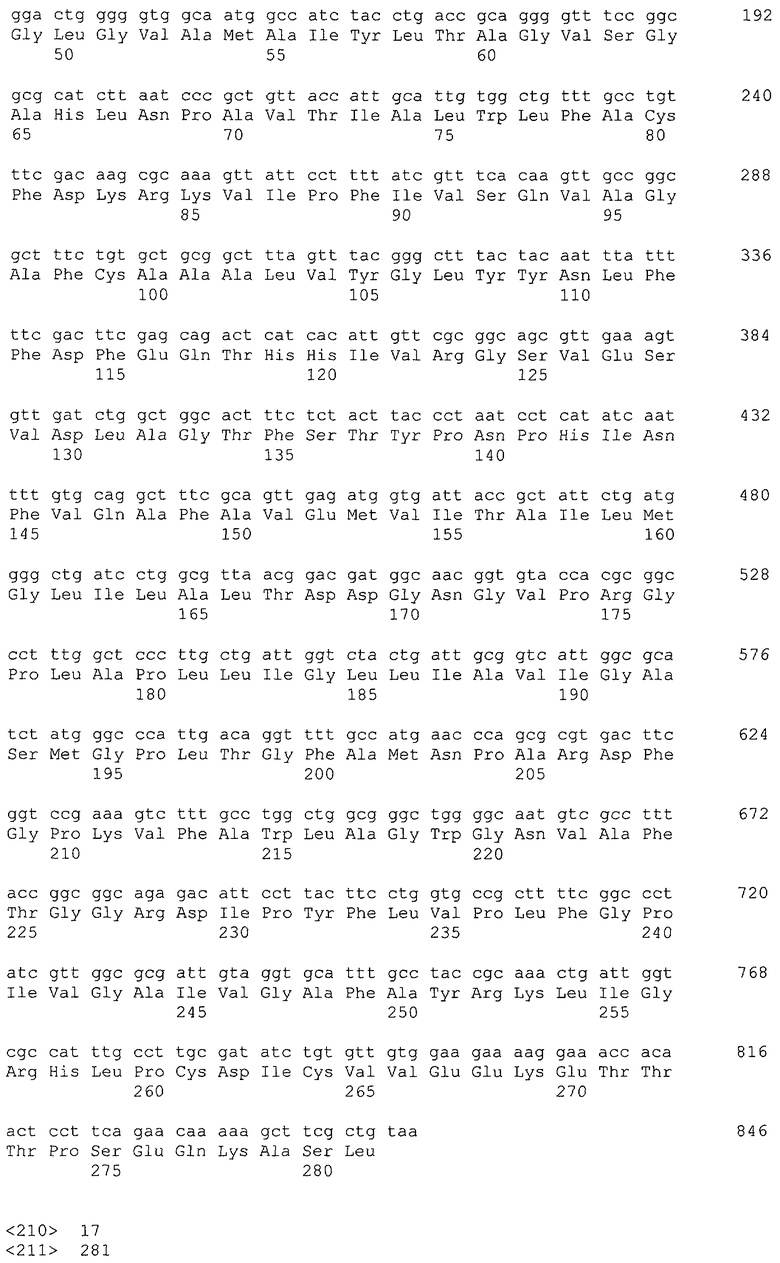







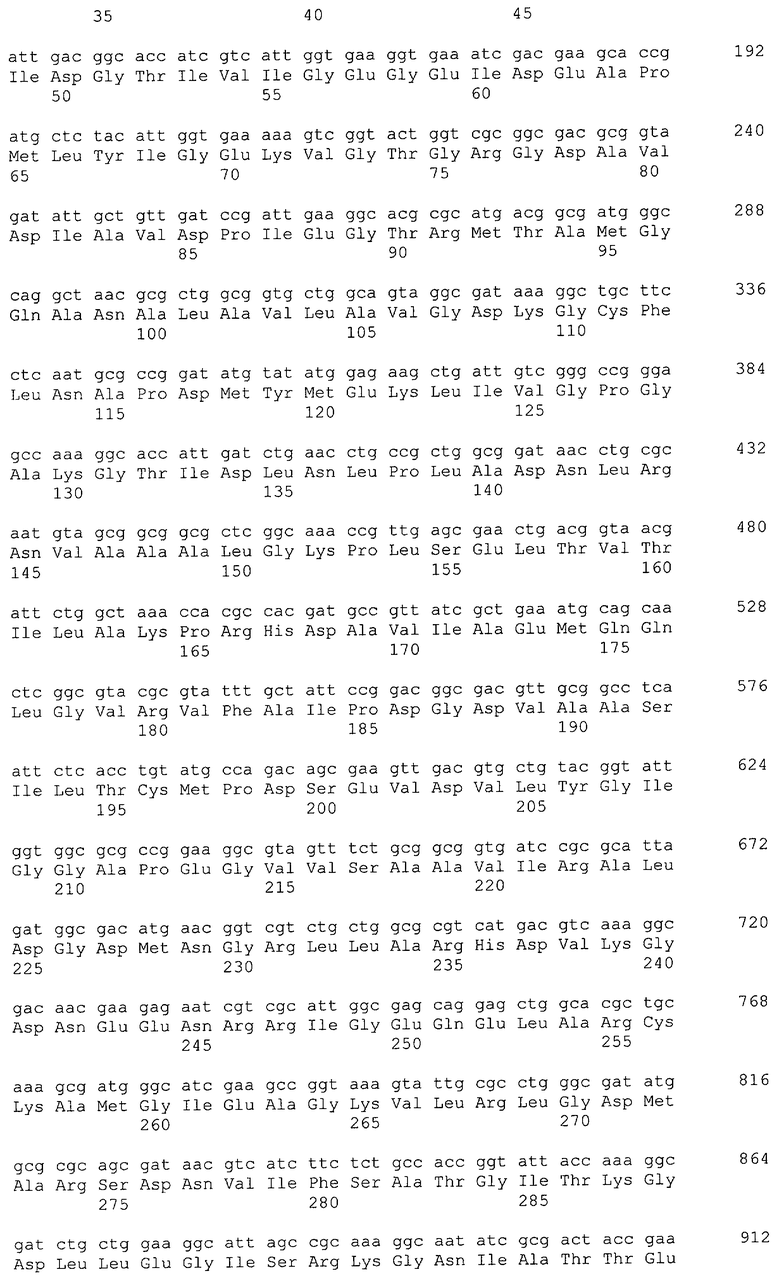


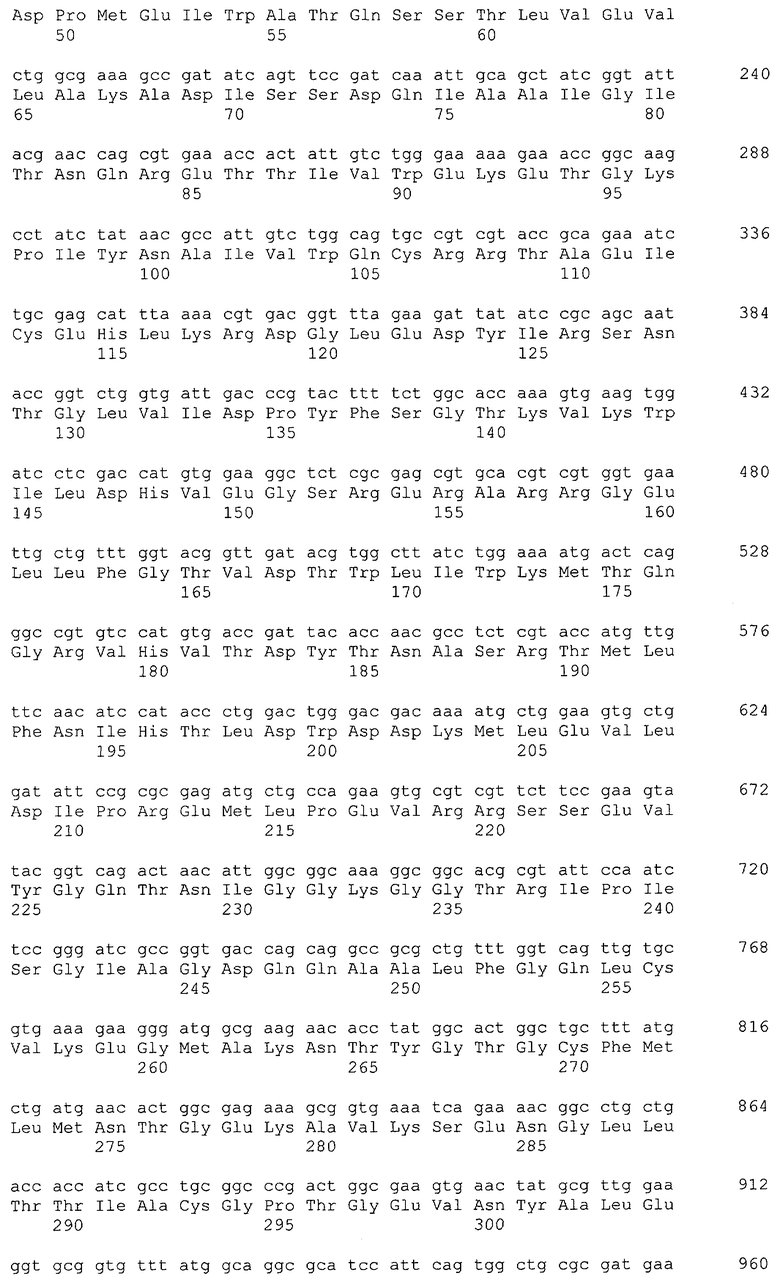

























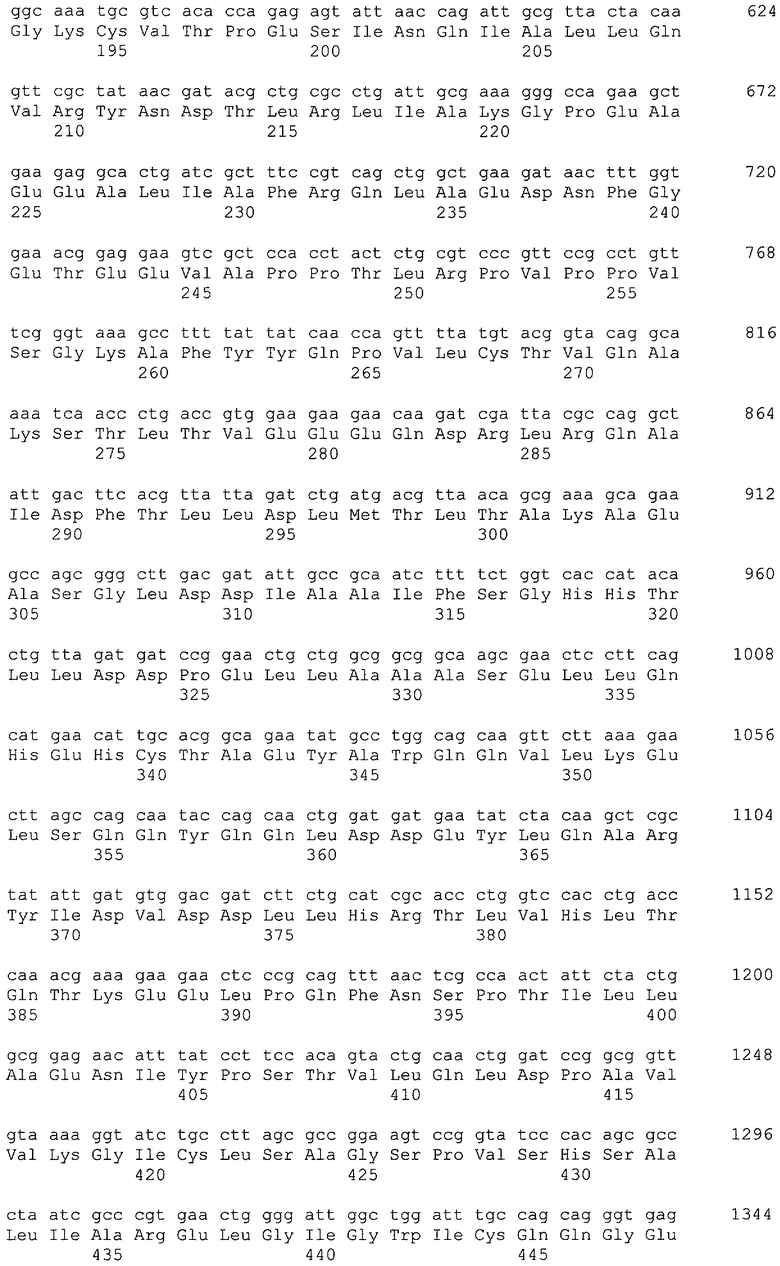



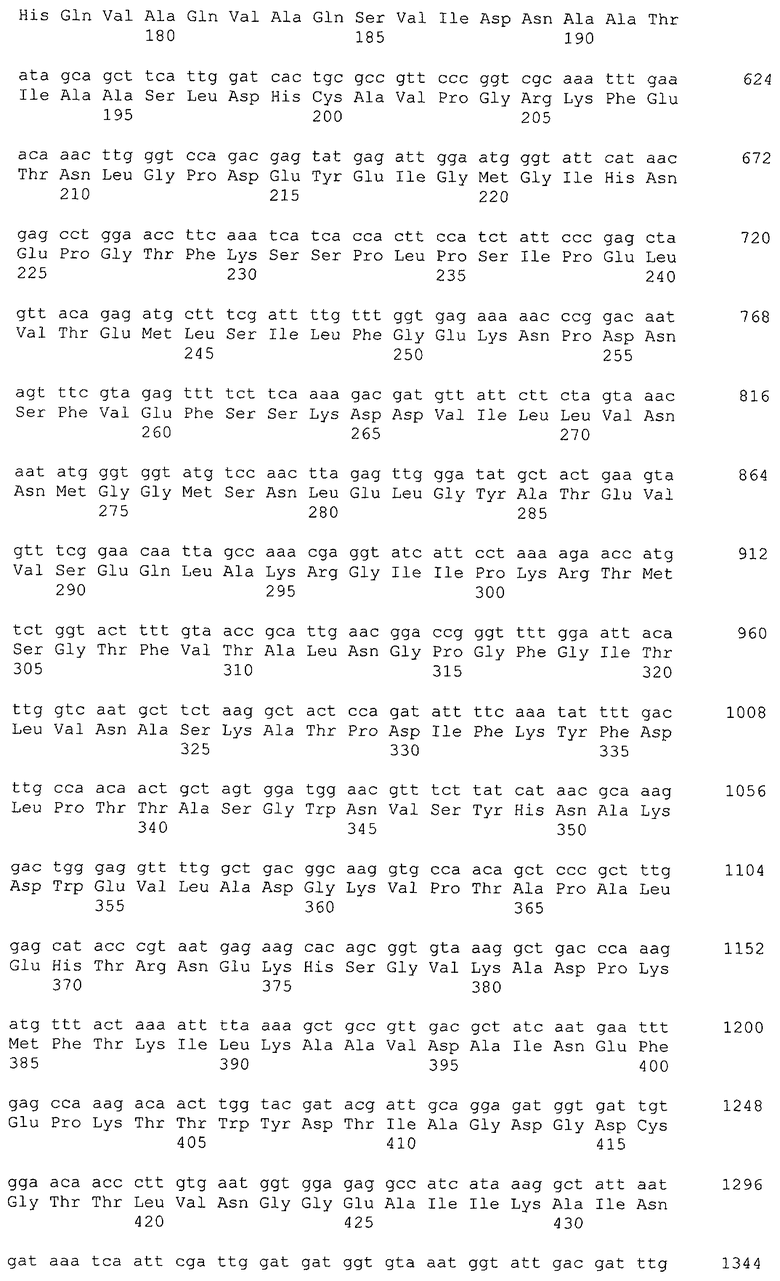





























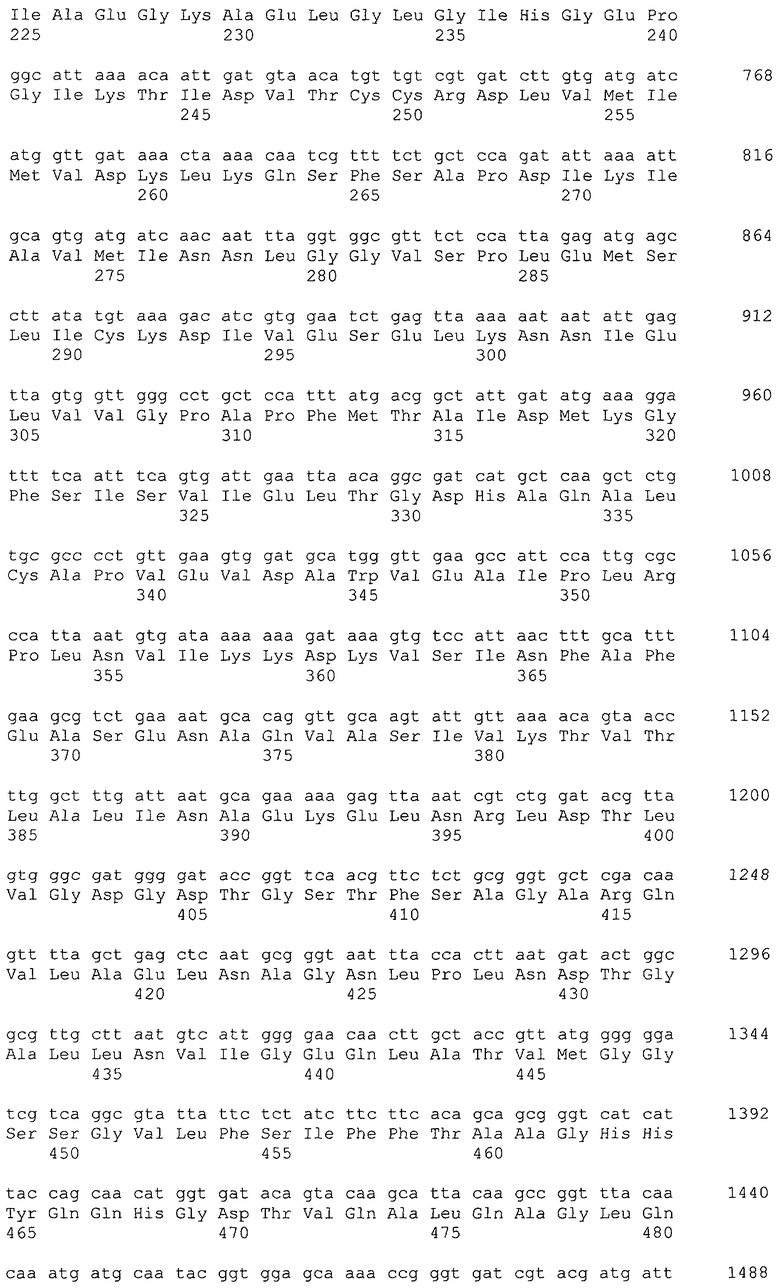



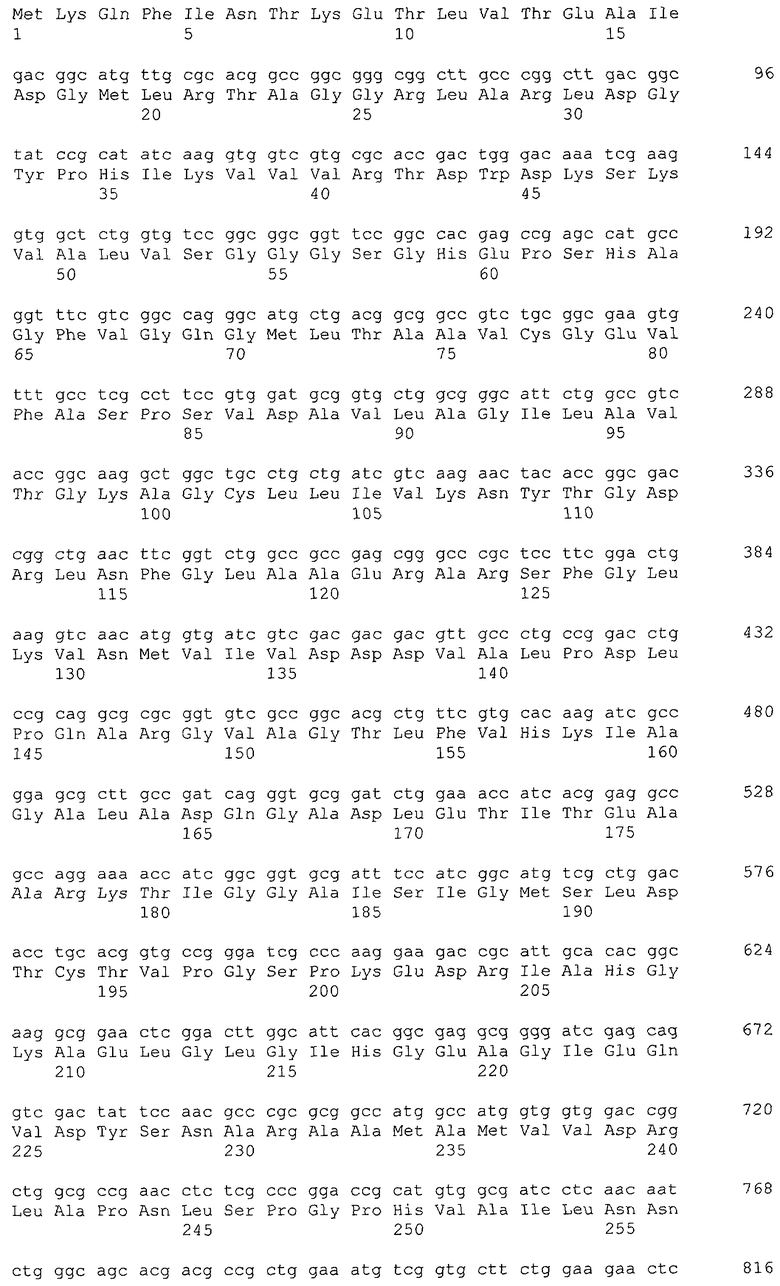
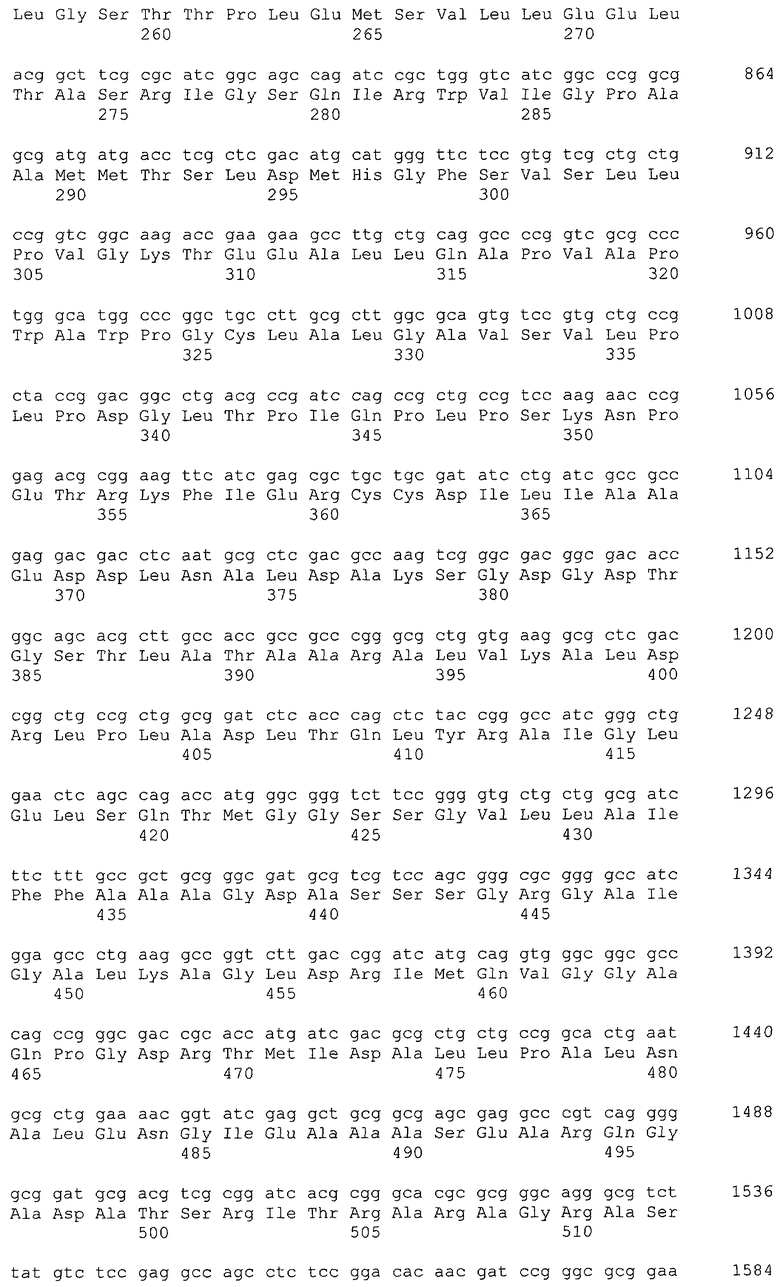




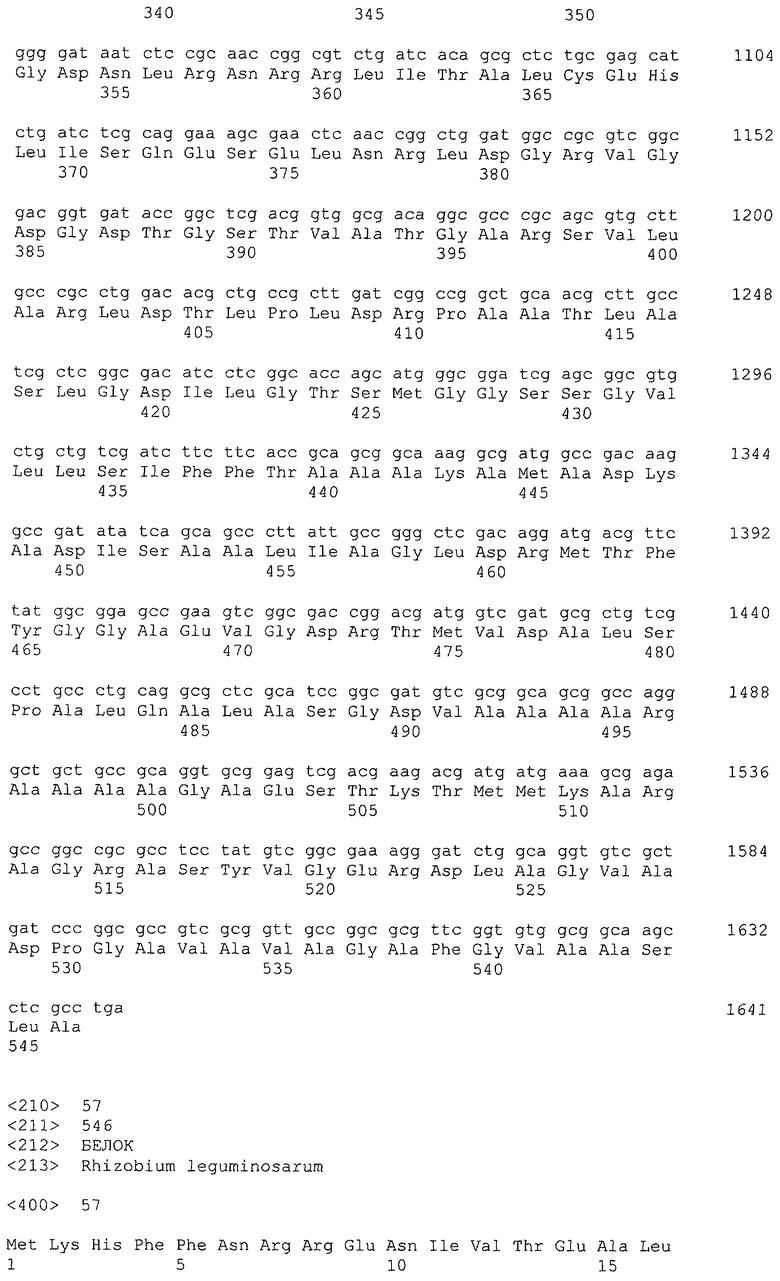



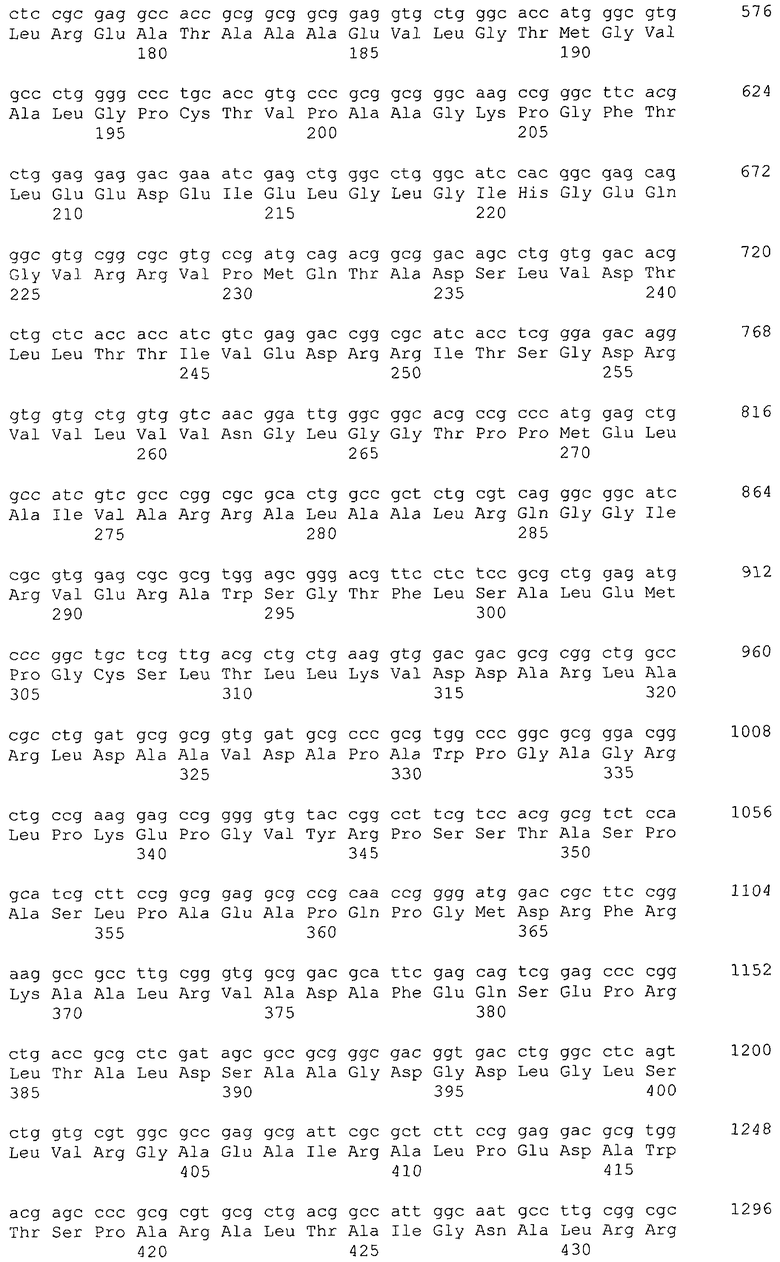









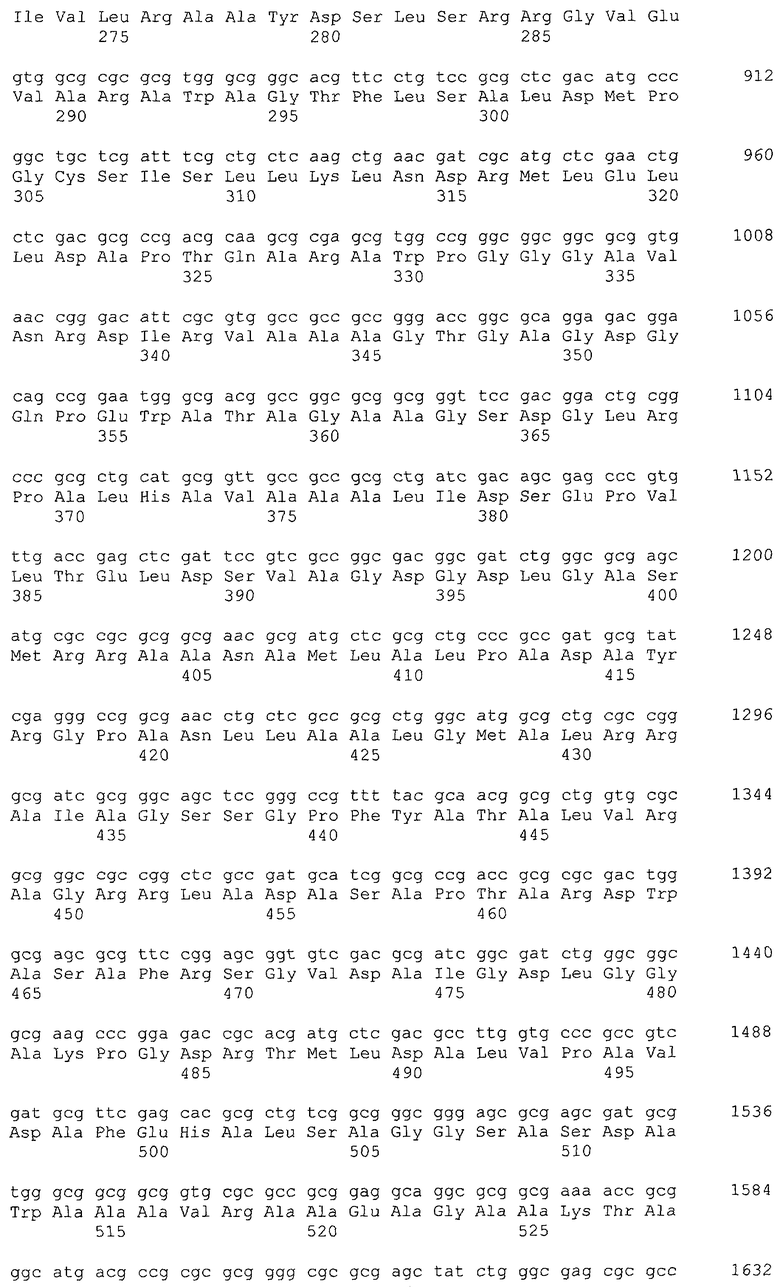





































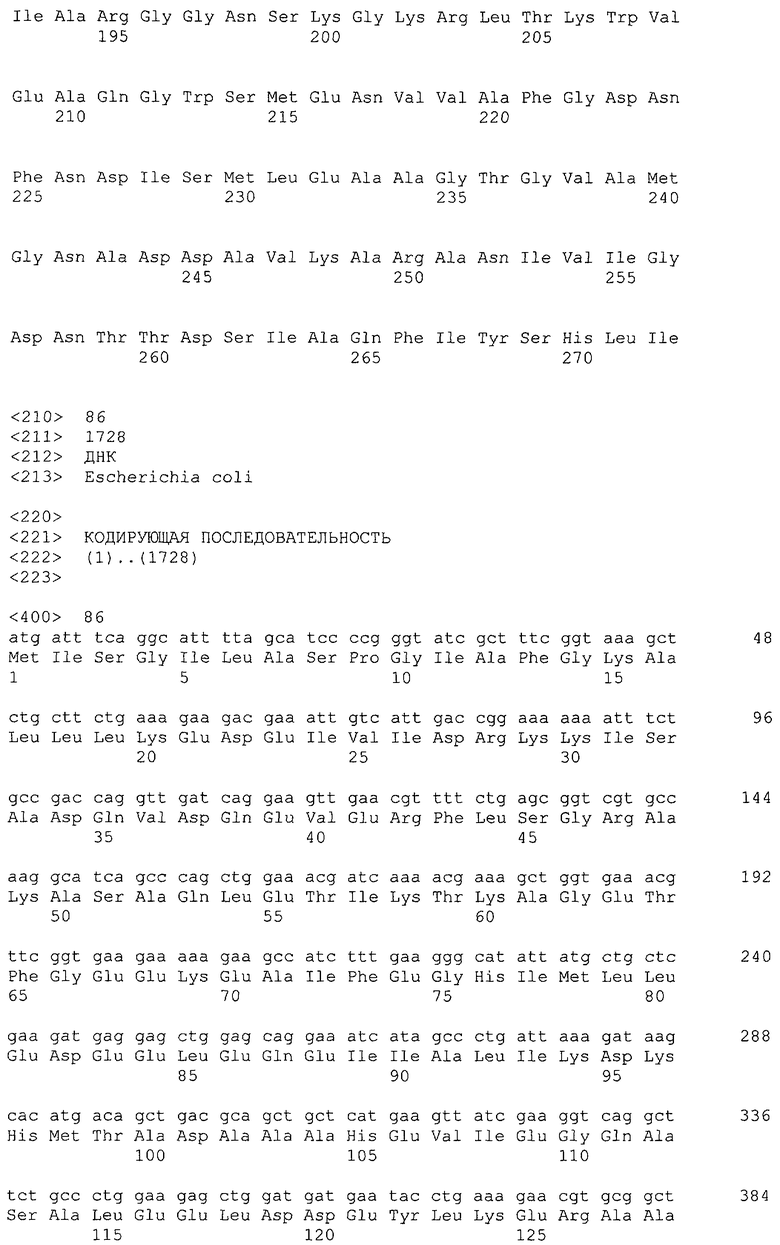
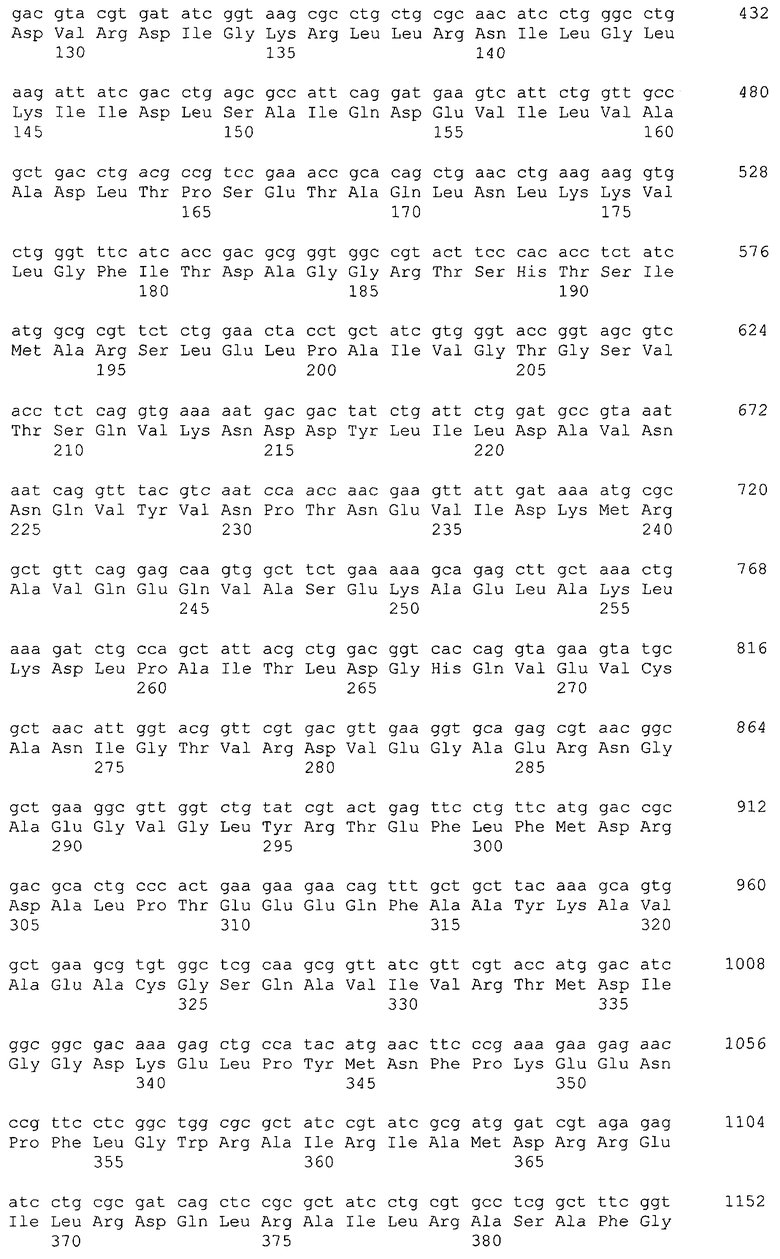











Изобретение относится к области биотехнологии и представляет собой способ получения L-аминокислоты, включающий культивирование микроорганизма, относящегося к семейству Enterobacteriaceae, обладающего способностью продуцировать L-аминокислоту и модифицированного так, чтобы повысить активности глицериндегидрогеназы и дигидроксиацетонкиназы, в среде, содержащей глицерин в качестве источника углерода, для продуцирования и накопления L-аминокислоты в среде или в клетках, и сбор L-аминокислоты из среды или клеток, причем активности глицериндегидрогеназы и дигидроксиацетонкиназы повышают путем увеличения количества копий генов, кодирующих указанные ферменты, или замещения промотора для этих генов более сильным промотором, и ген дигидроксиацетонкиназы кодирует белок, выбранный из группы, состоящей из: а) белка, содержащего аминокислотную последовательность SEQ ID NO: 4; и б) белка, содержащего аминокислотную последовательность, гомологичную не менее чем на 95% аминокислотной последовательности SEQ ID NO: 4, и белок имеет активность дигидроксиацетонкиназы. Изобретение позволяет получать на повышенном уровне L-аминокислоты при использовании указанного микроорганизма, в котором дигидроксиацетонкиназа использует АТФ в качестве донора фосфата. 13 з.п. ф-лы, 6 табл., 4 пр.
1. Способ получения L-аминокислоты, включающий культивирование микроорганизма, относящегося к семейству Enterobacteriaceae, обладающего способностью продуцировать L-аминокислоту и модифицированного так, чтобы повысить активности глицериндегидрогеназы и дигидроксиацетонкиназы, в среде, содержащей глицерин в качестве источника углерода, для продуцирования и накопления L-аминокислоты в среде или в клетках, и
сбор L-аминокислоты из среды или клеток,
где активности глицериндегидрогеназы и дигидроксиацетонкиназы повышают путем увеличения количества копий генов, кодирующих глицериндегидрогеназу и дигидроксиацетонкиназу, или замещения промотора для этих генов более сильным промотором,
где ген дигидроксиацетонкиназы кодирует белок, выбранный из группы, состоящей из:
(A) белка, содержащего аминокислотную последовательность SEQ ID NO: 4; и
(B) белка, содержащего аминокислотную последовательность, гомологичную не менее чем на 95% аминокислотной последовательности SEQ ID NO: 4, и белок имеет активность дигидроксиацетонкиназы.
2. Способ по п. 1, в котором количества копий генов, кодирующих глицериндегидрогеназу и дигидроксиацетонкиназу, повышены путем трансформирования микроорганизма вектором многокопийного типа, содержащим эти гены, и/или введения этих генов в хромосому микроорганизма.
3. Способ по п. 1, в котором дигидроксиацетонкиназа использует АТФ в качестве донора фосфата.
4. Способ по п. 1, в котором микроорганизм дополнительно модифицирован так, чтобы повысить активность посредника глицерина,
где активность посредника глицерина повышают путем увеличения количества копий гена, кодирующего посредник глицерина, или замещения промотора для этого гена более сильным промотором.
5. Способ по п. 1, в котором микроорганизм дополнительно модифицирован так, чтобы повысить активность или активности одного или более ферментов, выбранных из группы, состоящей из триозофосфатизомеразы, фруктозобисфосфатальдолазы, фруктозо-1,6-бисфосфатазы и фруктозо-6-фосфатальдолазы, где активность или активности ферментов, выбранных из группы, состоящей из триозофосфатизомеразы, фруктозобисфосфатальдолазы, фруктозо-1,6-бисфосфатазы и фруктозо-6-фосфатальдолазы и их комбинаций, повышают путем увеличения количества копий генов, кодирующих этот фермент (эти ферменты), или замещения промотора для этого гена (этих генов) более сильным промотором.
6. Способ по п. 1, в котором микроорганизм дополнительно модифицирован так, чтобы уменьшить активность или активности глицеринкиназы и/или глицерин-3-фосфатдегидрогеназы связанного с мембраной типа, где активность или активности глицеринкиназы и/или глицерин-3-фосфатдегидрогеназы связанного с мембраной типа понижают путем модифицирования контрольных последовательностей гена, кодирующего фермент.
7. Способ по п. 1, в котором микроорганизм, относящийся к семейству Enterobacteriaceae, представляет собой бактерию Escherichia или бактерию Pantoea.
8. Способ по п. 1, в котором L-аминокислота представляет собой один или более видов L-аминокислот, выбранных из группы, состоящей из L-глутаминовой кислоты, L-лизина, L-лейцина, L-изолейцина, L-валина, L-триптофана, L-фенилаланина, L-тирозина, L-треонина, L-метионина, L-цистеина, L-аргинина, L-серина, L-пролина, L-аспарагиновой кислоты, L-аспарагина, L-глутамина и L-гистидина.
9. Способ по п. 7, в котором бактерия представляет собой Escherichia coli.
10. Способ по п. 4 или 5, в котором количества копий генов, кодирующих белок (белки), повышают путем трансформирования микроорганизма вектором многокопийного типа, содержащим эти гены, и/или введения этих генов в хромосому микроорганизма.
11. Способ по п. 1, в котором ген глицериндегидрогеназы происходит из Escherichia coli.
12. Способ по п. 11, в котором ген глицериндегидрогеназы кодирует белок, выбранный из группы, состоящей из:
(a) белка, содержащего аминокислотную последовательность SEQ ID NO: 2; и
(b) белка, содержащего аминокислотную последовательность, гомологичную не менее чем на 95% аминокислотной последовательности SEQ ID NO: 2,
и белок имеет активность глицериндегидрогеназы.
13. Способ по п. 1, в котором промотор, под который помещен ген глицериндегидрогеназы, представляет собой промотор, полученный из промотора tac.
14. Способ по п. 1, в котором промотор, под который помещен ген дигидроксиацетонкиназы, представляет собой промотор, полученный из промотора tac.
| EP 1715055 A2, 25.10.2006 | |||
| DATABASE: UniProtKB/Swiss-Prot, P54838 (DAK1_YEAST), 01.10.1996 | |||
| MIKAEL MOLIN, et al., "Fragmentation of dihydroxyacetone kinase 1 from Saccharomyces cerevisiae indicates a two-domain structure", Proteomics 2003, 3, pp | |||
| Патрон для электрической лампочки накаливания с цоколем Свана | 1923 |
|
SU752A1 |
| WO 9928480 A1, 10.06.1999 | |||
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 1999 |
|
RU2188236C2 |
| EP 1715056 A2, 25.10.2006 | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |

