Настоящее изобретение относится к ранее неизвестным ферментам метанолдегидрогеназам (MDH), идентифицированным в метилотрофных бактериях, и в частности касается новых MDH-кодирующих генов, идентифицированных в Bacillus methanolicus MGA3 и Bacillus methanolicus РВ1. Изобретение основано на неожиданном открытии того, что в этих штаммах В.methanolicus встречается множество изоформ MDH, которые отличаются своими биохимическими свойствами. Такие новые гены, кодирующие ранее неизвестные изоформы MDH, можно использовать в генной инженерии микроогранизмов-хозяев, например в контексте использования метанола и/или других С1-соединений в качестве субстрата для роста. Таким образом, новые гены/ферменты можно использовать для введения или модификации, например активации/усиления, активности MDH в микроорганизме-хозяине.
Метилотрофные микроорганизмы могут использовать одноуглеродные (С1) источники, такие как метан и метанол, в качестве своих единственных источников энергии и образования биомассы, и существует ряд различных ферментов и путей С1-метаболизма среди метилотрофов. Был выделен ряд грамположительных термостойких бацилл со способностью роста на метаноле при температурах вплоть до 60°С, которых классифицировали как Bacillus methanolicus. В.methanolicus представляет собой так называемый ограниченный метилотроф, что означает, что он может использовать незначительное число многоуглеродных источников энергии и роста. Научный интерес к этим организмам главным образом был сфокусирован на их потенциале в качестве клеточных фабрик для промышленного производства аминокислот, особенно L-лизина и L-глутамата, из метанола при повышенных температурах, но также предполагалось их возможное применение в качестве хозяев для продуцирования других полезных продуктов, включая витамины, цитохромы, коферменты и рекомбинантные белки.
В.methanolicus MGA3 (АТСС53907) был первоначально выделен из образцов почвы в Миннесоте (Schendel, Bremmon et al. (1990) Appl Environ Microbiol 56(4): 963-970), и он представлял собой главный модельный штамм, применяемый в метаболической инженерии этой бактерии (Brautaset, Jakobsen et al. (2007) Appl Microbiol Biotechnol 74(1): 22-34; Jakobsen, Brautaset et al. (2009) Appl Environ Microbiol 75(3): 652-661; Brautaset, Jakobsen et al. (2010) Appl Microbiol Biotechnol 87(3): 951-964). B. methanolicus имеет несколько уникальных особенностей, включая НАД(никотинамидадениндинуклеотид)-зависимую метанолдегидрогеназу (MDH) для окисления метанола (de Vries, Arfman et al. (1992) J Bacteriol 174(16): 5346-5353; Arfman, Hektor et al. (1997) Eur J Biochem 244(2): 426-433; Hektor, Kloosterman et al. (2002) J Biol Chem 277(49): 46966-46973). Активность метанолдегидрогеназы (MDH) является ключевой характеристикой метилотрофного роста и вовлечена в первую стадию ферментации метанола, а именно окисление метанола до формальдегида. Формальдегид представляет собой промежуточное соединение в метаболизме метанола, и следовательно детоксикация этого токсичного для клеток метаболита является очень важной. Формальдегид можно ассимилировать с помощью РМФ(рибулозомонофосфатного)-пути. Для непосредственного превращения формальдегида в CO2 также предполагался линейный диссимиляционный путь. Предполагают, что диссимиляционные пути являются важными для полного генерирования энергии в клетках при росте на метаноле. Вместе с РМФ-путем диссимиляционные пути могут также играть роль в поддержании количества формальдегида ниже токсических уровней в клетках. Поэтому эффективное окисление метанола и сопутствующая ассимиляция формальдегида имеет решающее значение для роста и потока энергии в первичном метаболизме и для продуцирования желаемых продуктов. Кроме того, все это должно быть тщательно сбалансировано для того, чтобы обеспечить эффективное превращение метанола и в то же время избежать токсической аккумуляции формальдегида в клетках. В этом смысле, MDH играет важную роль в бактериальной метилотрофии.
Бактериальные MDH можно разделить на группы согласно механизмам их взаимодействия и использования кофактора(ов). Наиболее изученной является состоящий из двух субъединиц пирролохинолинхинон(PQQ)-зависимый хинопротеин MDH, широко распространенный в грамотрицательных метилотрофных бактериях. Грамположительные метилотрофы обычно кодируют НАД(Ф)+-зависимые метанолдегидрогеназы, и в дополнение к MDH из штамма MGA3, рассмотренного выше, НАД+-зависимая MDH была идентифицирована в другом штамме В.methanolicus, штамме С1 (Vonck, Arfman et al. (1991) J Biol Chem 266(6): 3949-3954; de Vries, Arfman et al. (1992) J Bacteriol 174(16): 5346-5353). MDH В.methanolicus демонстрирует сходство первичной последовательности с железосодержащими алкогольдегидрогеназами и поэтому отнесена к семейству III НАД-зависимых алкогольдегидрогеназ. Фермент состоит из десяти идентичных субъединиц, каждая из которых содержит прочно, но нековалентно, связанную молекулу НАД(Н) в дополнении к иону Zn2+ и 1-2 ионам Mg2+.
Было обнаружено, что метилотрофия в В.methanolicus является плазмидозависимой и вовлекает согласованный набор как плазмидных, так и хромосомных генов. В результате работы с В.methanolicus MGA3 была идентифицирована природная плазмида рВМ19, несущая mdh и пять генов РМФ-пути; потеря рВМ19 приводит к утрате способности роста на метаноле. В работе, предшествовавшей настоящему изобретению, о которой ранее не сообщалось, была показана соответствующая аналогичная плазмида, обозначенная рВМ20, в физиологически очень непохожем альтернативном модельном штамме РВ1 (NCIMB 13113).
Было показано, что НАД-зависимые ферменты MDH каталитически активируются активаторным белком Act, который относят к семейству nudix гидролаз.
Окисление метанола является основным критическим элементом в попытках конструировать метилотрофию в микроорганизмах-хозяевах. Действительно, даже в контексте организмов-хозяев, которые представляют собой природные метилотрофы, например В.methanolicus, модификация активности или экспрессии MDH может быть выгодной для улучшения роста и/или выхода желаемых продуктов. Поэтому сохраняется потребность в ферментах MDH, и в частности в новых генах mdh, которые можно использовать в генетической инженерии организмов, особенно таких генах, которые кодируют новые ферменты, обладающие измененными или улучшенными свойствами относительно ферментов MDH из данной области техники, например улучшенной активностью или стабильностью, или которые могут так или иначе быть выгодными для применения в генетической модификации желаемых хозяев.
В целях лучшего понимания физиологии метилотрофной клетки-хозяина В.methanolicus авторы настоящего изобретения секвенировали геном MGA3 и альтернативного штамма дикого типа РВ1. В процессе этого секвенирования было неожиданно обнаружено, что оба штамма имеют множество изоформ MDH; в обоих штаммах были идентифицированы три гена, кодирующие три обособленных белка НАД-зависимой MDH. Таким образом, в В.methanolicus MGA3, в дополнение к описанному ранее кодируемому плазмидой гену mdh-MGA3, были идентифицированы два новых гена, обозначенных в данном описании изобретения mdh2-MGA3 и mdh3-MGA3. Интересно, что эти новые гены mdh локализованы в хромосомах. В В.methanolicus РВ1 были идентифицированы три новых гена, обозначенных в данном описании изобретения mdh-PB1, mdh1-PB1 и mdh2-PB1, где первый из них несет плазмида (плазмида рВМ20), а последние два являются хромосомными. Все эти гены были экспрессированы рекомбинантным путем, очищены и охарактеризованы биохимически in vitro. Несмотря на проявление некоторых сходств, становится ясно, что эти разные ферменты MDH могут иметь разные свойства, включая их активность. На основе этих исследований, и в частности анализа последовательностей, было идентифицировано два различных подсемейства MDH.
Первое подсемейство включает ранее описанный ген mdh, который несет плазмида, штамма MGA3 (mdh-MGA3) и два гена из штамма РВ1, mdh-PB1 и mdh1-РВ1 (mdn-PB1 несет плазмида, а mhd1-ΡΒ1 является хромосомным) и определено в данном описании изобретения как "семейство mdh/mdh1-типа". Второе подсемейство включает новые хромосомные гены mdh2-MGA3, mdh3-MGA3 и mdn2-PB1 и определено в данном описании изобретения как "семейство mdh2/mdh3-типа". Именно это последнее семейство образует объект настоящего изобретения.
Члены семейства mdh2/mdh3-типа обладают по меньшей мере 90%-ной идентичностью последовательности друг к другу на уровне ДНК (см. Фиг. 1) и на уровне аминокислотной последовательности относительно кодируемых белков (см. Фиг. 2). В частности, кодирующие последовательности mdn2-MGA3 (SEQ ID NO: 1) и mdh3-MGA3 (SEQ ID NO: 3) имеют 96%-ную идентичность последовательности ДНК, а предсказанные полипептиды Mdh2-MGA3 (SEQ ID NO: 2) и Mdh3-MGA3 (SEQ ID NO: 4) имеют 96%-ную идентичность аминокислот (см. Фиг. 2В). Предсказанный полипептид Mdh2-PB1 (SEQ ID NO: 6) на 91% идентичен предсказанному полипептиду Mdh2-MGA3 (SEQ ID NO: 2) и на 92% идентичен предсказанному полипептиду Mdh3-MGA3 (SEQ ID NO: 4) (см. Фиг. 2В).
С другой стороны, идентичность последовательности между членами двух разных подсемейств значительно ниже, приблизительно 60-66%. Например, кодирующая последовательность mdh2-MGA3 (SEQ ID NO: 1) на 65% идентична кодирующей последовательности mdh-MGA3 (SEQ ID NO: 7), a предсказанный полипептид Mdh2-MGA3 (SEQ ID NO: 2) на 61% идентичен предсказанному полипептиду Mdh-MGA3 (SEQ ID NO: 8). Кодирующая последовательность гена mdh3-MGA3 (SEQ ID NO: 3) на 66% идентична кодирующей последовательности mdh-MGA3 (SEQ ID NO: 7), а предсказанный полипептид Mdh3 (SEQ ID NO: 4) на 62% идентичен Mdh-MGA3 (SEQ ID NO: 8).
Как уже отмечалось, исследования биохимических характеристик выявили различия между ферментами MDH семейств mdh2/mdh3- и mdh/mdh1-типов. Например, Mdh3-MGA3 (SEQ ID NO: 4) и Mdh2-PB1 (SEQ ID NO: 6) обладают улучшенной термостабильностью. Также обнаружены различия в субстратной специфичности и уровне активности на субстратах различных спиртов. Это открывает возможность для применения таких ферментов в окислении разных спиртов (например этанола или пропанола), а не только метанола.
Также были выполнены исследования по гетерологичной экспрессии генов в различных неметилотрофных хозяевах. Эти исследования установили полезность новых последовательностей по изобретению семейства mdh2/mdh3-типа в генетической инженерии ряда различных клеток-хозяев для введения активности MDH и тем самым дают возможность использования метанола. Предполагается, что настоящее изобретение имеет широкую применимость, поскольку имеет отношение к разным клеткам-хозяевам, и в исследованиях, описанных в данном описании изобретения, были использованы два биотехнологически хорошо охарактеризованных и филогенетически очень различных бактериальных штамма-хозяина, т.е. грамотрицательная Escherichia coli и грамположительная Bacillus subtilis, и было показано, что каждый генетически модифицированный микроорганизм-хозяин демонстрирует повышенную активность MDH при модификации для экспрессии новых ферментов MDH по настоящему изобретению, особенно ферментов из семейства mdh2/mdh3-типа из В.methanolicus MGA3 и В.methanolicus РВ1.
В особенности, результаты, представленные в данном описании изобретения, показывают, что разные конкретные ферменты могут проявлять улучшенную активность в различных хозяевах. Например, для экспрессии активности MDH в хозяине Е.coli наилучшие результаты дает Mdh2-MGA3 (SEQ ID NO: 1). Выбор фермента MDH может также зависеть от условий экспрессии и точной природы клетки-хозяина и/или условий культивирования, например от того, коэкспрессируется ли ген act, и если да, то какой конкретно. Таким образом, новые ферменты по изобретению и их кодирующие последовательности преимущественно предоставляют новый и расширенный спектр ферментов MDH и кодирующих молекул нуклеиновой кислоты для применения в окислении спиртов, включая метанол, и в частности для применения в генетической модификации клеток-хозяев (например для продуцирования рекомбинантных клеток-хозяев), например для введения или модификации активности алкогольдегидрогеназы в клетку(е)-хозяина(е), особенно активности MDH, или для введения метилотрофии в клетку-хозяина. Как дополнительно описано ниже, молекулы нуклеиновой кислоты, кодирующие новые ферменты по настоящему изобретению, можно применять по отдельности или в комбинации.
Таким образом, в первом аспекте настоящего изобретения предложена молекула нуклеиновой кислоты, конкретно выделенная молекула нуклеиновой кислоты, которая кодирует полипептид (или белок), обладающий активностью алкогольдегидрогеназы, в частности активностью метанолдегидрогеназы, содержащая или имеющая (например состоящая из) нуклеотидную последовательность, выбранную из группы, состоящей из:
1) нуклеотидной последовательности, представленной в любой из SEQ ID NO: 1 (mdh2-MGA3), 3 (mdh3-MGA3) или 5 (mdh2-PB1);
2) нуклеотидной последовательности, обладающей по меньшей мере 90%-ной идентичностью последовательности, более конкретно по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99%-ной идентичностью последовательности с нуклеотидной последовательностью, представленной в любой из SEQ ID NO: 1, 3 или 5;
3) нуклеотидной последовательности, которая является вырожденной по отношению к любой нуклеотидной последовательности из SEQ ID NO: 1, 3 или 5;
4) нуклеотидной последовательности, которая представляет собой часть нуклеотидной последовательности любой из SEQ ID NO: 1, 3 или 5 или нуклеотидной последовательности, которая является вырожденной по отношению к последовательности SEQ ID NO: 1, 3 или 5;
5) нуклеотидной последовательности, кодирующей весь полипептид или часть полипептида, аминокислотная последовательность которого представлена в любой из SEQ ID NO: 2 (Mdh2-MGA3), 4 (Mdh3-MGA3) или 6 (Mdh2-PB1); и
6) нуклеотидной последовательности, кодирующей весь полипептид или часть полипептида, который имеет аминокислотную последовательность, обладающую по меньшей мере 90%-ной идентичностью последовательности, предпочтительно по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99%-ной идентичностью последовательности с аминокислотной последовательностью, представленной в любой из SEQ ID NO: 2, 4 или 6;
или молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, которая комплементарна нуклеотидной последовательности любой из (1)-(6).
В еще одном аспекте в настоящем изобретении предложен полипептид, обладающий активностью алкогольдегидрогеназы, в частности активностью метанолдегидрогеназы, и содержащий или имеющий (например, состоящий из) аминокислотную последовательность, выбранную из группы, состоящей из:
1) всей или части аминокислотной последовательности, представленной в любой из SEQ ID NO: 2, 4 или 6; и
2) всей или части аминокислотной последовательности, обладающей по меньшей мере 90%-ной идентичностью последовательности, предпочтительно по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99%-ной идентичностью последовательности с аминокислотной последовательностью, представленной в любой из SEQ ID NO: 2, 4 или 6.
Молекулы нуклеиновой кислоты по настоящему изобретению преимущественно дают возможность для введения или модификации активности алкогольдегидрогеназы, и в частности MDH, в организм(е)-хозяина(е). Это может быть достигнуто путем модификации организма для экспрессии одной или более чем одной молекулы нуклеиновой кислоты по изобретению. Как отмечено выше, указанные молекулы нуклеиновой кислоты могут быть получены или иметь происхождение из генов mdh штаммов В.methanolicus, в частности штаммов MGA3 и РВ1. В конкретном воплощении молекулы нуклеиновой кислоты, кодирующие различные ферменты MDH (например, различные изоферменты или ферменты из различных штаммов или различные варианты полипептидов и т.д.) или имеющие происхождение из молекул нуклеиновой кислоты, кодирующих различные ферменты MDH, можно использовать в комбинации. Таким образом, можно коэкспрессировать две или более различных молекул нуклеиновой кислоты.
Таким образом, в настоящем изобретении предложен способ введения или модификации активности MDH в организме-хозяине путем экспрессирования в указанном организме одной или более чем одной молекулы нуклеиновой кислоты по изобретению. В частности, молекула нуклеиновой кислоты может быть гетерологичной или ненативной по отношению к организму-хозяину. Ее можно экспрессировать под контролем нативного или ненативного промотора.
Соответственно, в еще одном аспекте настоящего изобретения предложен способ введения или модификации активности алкогольдегидрогеназы, и в частности активности MDH, в организм(е)-хозяина(е), включающий введение в указанный организм молекулы нуклеиновой кислоты по изобретению, как определено выше в данном описании изобретения, и выращивание (или культивирование) указанного организма в условиях, при которых указанная молекула нуклеиновой кислоты экспрессируется.
Легко заметить, что в этом аспекте в изобретении также может быть предложен способ продуцирования полипептида(ов), обладающего(их) активностью алкогольдегидрогеназы, и в частности активностью MDH, включающий введение в организм-хозяина молекулы нуклеиновой кислоты по изобретению, как определено выше в данном описании изобретения, и выращивание (или культивирование) указанного организма в условиях, при которых указанный(ые) полипептид(ы) продуцируе(ю)тся. Организм-хозяин может представлять собой организм, который изначально (например, в его диком типе) не обладает активностью MDH (т.е. не имеет или не обладает эндогенной MDH), и следовательно в таком воплощении изобретения предусмотрено введение активности MDH в хозяина. С альтернативной точки зрения в таком воплощении хозяина можно модифицировать для введения способности превращать метанол в формальдегид или, другими словами, модифицировать хозяина для обеспечения исходной стадии использования С1-углеродного источника, конкретно использования метанола.
В альтернативном воплощении организм-хозяин может иметь или обладать эндогенным ферментом MDH, и поэтому способ по изобретению может включать модификацию активности MDH в таком хозяине путем введения молекулы нуклеиновой кислоты, кодирующей еще один или дополнительный фермент MDH, который, например, может быть гетерологичным хозяину. Также охвачена сверхэкспрессия активности MDH в организме-хозяине путем введения в указанный организм молекулы нуклеиновой кислоты, кодирующей нативный фермент MDH (т.е. в котором введенная молекула нуклеиновой кислоты кодирует эндогенный фермент MDH).
Модифицированный организм-хозяин можно культивировать или выращивать, используя любой желаемый источник углерода в качестве субстрата, включая метанол или высший спирт, но не ограничиваясь ими. Таким образом, в одном воплощении способ по изобретению может включать культивирование или выращивание организма-хозяина, который содержит одну или более чум одну MDH-кодирующую молекулу нуклеиновой кислоты, введенную экзогенно, как определено в данном описании изобретения.
В другом аспекте настоящего изобретения предложен организм-хозяин, который модифицирован для введения молекулы нуклеиновой кислоты по изобретению, как определено выше в данном описании изобретения.
В частности, в этом аспекте изобретения молекула нуклеиновой кислоты, которую вводят, содержит нуклеотидную последовательность, которая гетерологична организму-хозяину. Гетерологичная последовательность может представлять собой нуклеотидную последовательность, кодирующую полипептид алкогольдегидрогеназу (например, MDH), или она может представлять собой гетерологичную последовательность контроля экспрессии или некоторую другую последовательность (например вектор или маркерная последовательность). В случае организма-хозяина, который эндогенно экспрессирует фермент алкогольдегидрогеназу, модифицированный хозяин может отличаться от немодифицированного организма-хозяина содержанием дополнительной копии молекулы нуклеиновой кислоты, кодирующей полипептид алкогольдегидрогеназу. Другими словами, он может содержать больше копий кодирующей нуклеотидной последовательности, чем немодифицированный хозяин.
Как отмечалось выше, молекулы нуклеиновой кислоты, кодирующие новые ферменты MDH по настоящему изобретению, могут быть получены, например выделены или клонированы, из В.methanolicus, в частности из штаммов MGA3 и РВ1. Таким образом, фермент MDH может представлять собой Mdh2 или Mdh3 из MGA3 (SEQ ID NO: 2 или 4, соответственно) либо Mdh2 из РВ1 (SEQ ID NO: 6). Однако, в дополнение к конкретным нативным ("дикого типа") последовательностям, указанным выше, также включены варианты этих последовательностей, которые обладают по меньшей мере 90%-ной идентичностью нуклеотидной последовательности с ними и которые сохраняют активность алкогольдегидрогеназы, и конкретно MDH. Такие варианты могут включать природные варианты, например различные варианты, которые могут встречаться в штаммах в природе или которые могут быть получены из других штаммов В.methanolicus и которые кодируют полипептиды MDH, функционально эквивалентные полипептидам MDH SEQ ID NO: 2, 4 или 6. Альтернативно, варианты могут представлять собой синтетические или искусственные варианты, например полученные или имеющие происхождение путем модификации (например мутации) аминокислотных последовательностей SEQ ID NO: 2, 4 или 6 или нуклеотидных последовательностей SEQ ID NO: 1, 3 или 5. Как отмечено выше, можно использовать комбинации двух или более различных молекул нуклеиновой кислоты по изобретению. Альтернативно, молекула нуклеиновой кислоты по изобретению может содержать две или более различных нуклеотидных последовательностей, кодирующих полипептид, обладающий активностью алкогольдегидрогеназы, как определено в данном описании изобретения, или их комплемент. Модификации могут быть выбраны исходя из улучшенной активности метанолдегидрогеназы соответствующего варианта или, альтернативно, могут быть сконструированы исходя из алгоритмов проектирования белка, используя молекулярные структуры или модели для предсказания улучшенной ферментативной активности.
Полипептид MDH по настоящему изобретению может также включать полипептид, кодируемый фрагментом (частью) нуклеотидной последовательности SEQ ID NO: 1, 3 или 5, или может содержать или состоять из фрагмента (или части) аминокислотной последовательности SEQ ID NO: 2, 4 или 6. "Часть" нуклеотидной или аминокислотной последовательности по изобретению может включать или содержать по меньшей мере 50, 55, 60, 65, 70, 75, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% или более смежных нуклеотидов или аминокислот последовательности.
Организм-хозяин может представлять собой любой подходящий организм-хозяин, но, в частности, будет представлять собой микробный организм-хозяин (т.е. микроорганизм). Он может представлять собой любой прокариотический организм, но, конкретно, будет представлять собой бактерию. Можно использовать любую грамположительную или грамотрицательную бактерию, но конкретно упомянуть можно следующие классы или роды: Escherichia, Corynebacterium и Bacillus. Типичные организмы-хозяева включают Е.coli, В.subtilis и С.glutamicum. Как отмечено выше, можно также использовать В.methanolicus или другие метилотрофные организмы-хозяева, например Methylomonas, Methylobacillus, Methylobacterium, Methylophilus или Methylococcus. Однако настоящее изобретение не ограничивается этими организмами и распространяется на любого микробного хозяина.
С.glutamicum представляет собой палочковидную непатогенную и грамположительную почвенную бактерию. Он растет в аэробных и анаэробных условиях и является ауксотрофом по биотину. С.glutamicum способен к росту на ряде субстратов в виде единственных или комбинированных источников углерода и энергии. Среди субстратов метаболизируемыми являются сахара, как глюкоза, фруктоза или сахароза, и органические кислоты, такие как L-лактат и ацетат. Кроме того, С.glutamicum способен к росту на этаноле в качестве единственного источника углерода. Его широко используют для крупномасштабного промышленного производства аминокислот L-глутамата и L-лизина. Недавние исследования в метаболической инженерии показали, что С.glutamicum также способен продуцировать ряд других интересных с коммерческой точки зрения соединений, например другие L-аминокислоты, D-аминокислоты, диамины, такие как кадаверин или путресцин, органические кислоты, такие как сукцинат, и биотопливо, такое как этанол или изобутанол.
Согласно настоящему изобретению одну или более чем одну молекулу нуклеиновой кислоты по изобретению можно экспрессировать в организме-хозяине, включая, в частности, по меньшей мере одну гетерологичную молекулу нуклеиновой кислоты (т.е. молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая является гетерологичной хозяину), и в частности содержащую гетерологичную последовательность, кодирующую полипептид MDH. Таким образом, организм-хозяин может быть модифицирован для экспрессии одной или более чем одной копии молекулы нуклеиновой кислоты или, альтернативно, может быть модифицирован для экспрессии одной или более чем одной копии ряда различных молекул нуклеиновой кислоты по изобретению.
Таким образом, микроорганизм, который модифицируют (или "конструируют") для экспрессии MDH согласно настоящему изобретению, будет содержать MDH-кодирующую молекулу нуклеиновой кислоты, введенную экзогенно, как определено в данном описании изобретения. Другими словами, организм можно трансформировать при помощи такой MDH-кодирующей молекулы нуклеиновой кислоты и его можно рассматривать как трансгенный или рекомбинантный организм. Как отмечено выше, молекула нуклеиновой кислоты может кодировать фермент MDH, который является гомологичным или гетерологичным (т.е. нативным или ненативным) этому хозяину. Таким образом, может быть введена еще одна (или более чем одна) копия гена, который является нативным хозяину. Молекула нуклеиновой кислоты, которую вводят, может содержать нуклеотидную последовательность, имеющую происхождение из нативного гена или из другого источника.
MDH можно экспрессировать в комбинации с другими ферментами, чтобы предоставить организму новые свойства.
"Экспрессия" при использовании в данном описании изобретения относится к транскрипции нуклеотидной последовательности в мРНК и последующей трансляции указанной мРНК в полипептидный продукт.
Как изложено в данном описании изобретения, "сверхэкспрессия" означает, что экспрессия нуклеотидной последовательности повышена по сравнению с или относительно уровнем(я) экспрессии, имеющем(го) место в организме, который не был модифицирован согласно изобретению. Экспрессию можно рассматривать в пересчете на количество продуцируемого полипептидного продукта (например фермента MDH), которое может быть определено посредством любого подходящего способа, известного в данной области техники. Например, экспрессию можно определять путем измерения активности белка (т.е. активности экспрессируемого полипептида MDH). Альтернативно, количество продуцируемого белка можно измерять для определения уровня экспрессии, например, вестерн-блоттингом или другими системами обнаружения антител или, действительно, посредством любого способа оценки или количественного определения белка. Можно также использовать ПЦР (полимеразную цепную реакцию) в реальном времени. Анализ может представлять собой анализ in vivo или in vitro.
Активность может быть определена путем анализа на активность алкогольдегидрогеназы посредством способов, известных в данной области техники и описанных в литературе, например, как подробно изложено в Примерах ниже. Активность MDH кодируемого белка может, например, катализировать превращение метанола в формальдегид, и указанную активность определяют в данном описании изобретения как количество фермента, необходимого для продуцирования 1 мкмоль НАДН в минуту, для которого можно использовать в качестве субстрата различные спирты, например этанол, метанол, пропанол, бутанол, пентанол, гексанол, изопропанол и 1,3-пропандиол. Активности алкогольдегидрогеназ можно измерять спектрофотометрически, как описано ранее в Hektor et al. (2002; Chem 277(49): 46966-46973).
Полипептид алкогольдегидрогеназу можно экспрессировать или сверхэкспрессировать посредством любого способа, известного в данной области техники, такого как введение молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую полипептид MDH, например копии нативного гена, например, экспрессирующейся из более сильного или нерегулируемого промотора относительно нативного гена, и/или введение множества копий MDH-кодирующей молекулы нуклеиновой кислоты.
Организм можно также конструировать для введения дополнительных или альтернативных регуляторных элементов.
В конкретном воплощении MDH-кодирующая молекула нуклеиновой кислоты может быть экспрессирована из ненативного или гетерологичного промотора (т.е. промотора, который является гетерологичным MDH-кодирующей нуклеотидной последовательности, т.е. не является нативным промотором гена MDH), и в частности сильного, ненативного или гетерологичного промотора. Таким образом, в конкретном воплощении MDH-кодирующий ген не применяют с его нативным промотором. Можно вводить MDH-кодирующий ген, который находится под контролем ненативного промотора. Как изложено в данном описании изобретения, сильный промотор представляет собой промотор, который экспрессирует ген с высоким уровнем или по меньшей мере с более высоким уровнем, чем под действием его нативного промотора. Термин "сильный промотор" представляет собой термин, хорошо известный и широко используемый в данной области техники, и множество сильных промоторов известно в данной области техники или может быть идентифицировано путем рутинного экспериментирования. Альтернативно, промотор представляет собой промотор mdh В.methanolicus. Однако выбор промотора конкретно не ограничен.
Альтернативно, ген MDH можно экспрессировать, используя нативный промотор. Изобретение охватывает применение микроорганизма, который может эндогенно экспрессировать ген mdh или который не делает этого. В первом случае, может быть введена одна или более чем одна дополнительная копия нативного гена или его варианта либо другой MDH или кодирующей молекулы нуклеиновой кислоты, и они могут быть введены под контролем нативного или ненативного промотора. С нативным промотором можно использовать, например, многокопийный вектор. Во втором случае, вводят MDH (или кодирующую молекулу нуклеиновой кислоты), которая является гетерологичной этому хозяину, но которая может находиться под контролем промотора, который является нативным или ненативным гену MDH, из которого имеет происхождение кодирующая молекула нуклеиновой кислоты.
Способы введения генов или молекул нуклеиновой кислоты хорошо известны в данной области техники и широко описаны в литературе, и можно использовать любой желаемый способ. Таким образом, ген (молекулу нуклеиновой кислоты) можно вводить, используя вектор, который может представлять собой автономно реплицирующийся вектор или вектор, который позволяет гену (молекуле нуклеиновой кислоте) интегрироваться в геном хозяина (например, хромосому). Таким образом, ген (молекулу нуклеиновой кислоты), подлежащий экспрессии, можно вводить в экспрессионный вектор и затем экспрессионный вектор можно вводить в клетку-хозяина. Способы конструирования экспрессионных векторов и введения их в клетки-хозяева хорошо известны в данной области техники. Подходящим образом, можно вводить ген, кодирующий MDH, используя плазмидный вектор, и микроорганизм-хозяина можно трансформировать при помощи плазмиды, например путем электропорации. Выбор способа может зависеть от используемого микроорганизма. Способы введения нуклеиновых кислот и векторов в микроорганизмы хорошо известны и широко описаны в литературе.
Молекула нуклеиновой кислоты предпочтительно кодирует полипептид или белок, представляющий собой MDH или ее часть, обладающую активностью MDH.
Предпочтительно, молекула нуклеиновой кислоты, как она определена в частях (1)-(6) выше, кодирует полипептид или белок, имеющий или сохраняющий функцию или активность или свойства полипептида MDH, как определено аминокислотными последовательностями любой из SEQ ID NO: 2, 4 или 6.
Термины "полипептид" и "белок" используют взаимозаменяемо в данном описании изобретения и включают аминокислотную цепь любой длины (т.е. любой полимер или олигомер аминокислот).
Как отмечено выше, изобретение распространяется на части или функциональные фрагменты нуклеотидных последовательностей, определенных выше, посредством чего подразумевают части или фрагменты, которые кодируют белок или полипептид, который имеет такую же или по существу такую же активность, что и полноразмерный белок, как определено выше. Тесты для определения того, имеет ли белок/полипептид, кодируемый такой частью или фрагментом, такую же или по существу такую же активность (например, каталитическую или ферментативную активность), что и полноразмерный полипептид/белок, как определено выше, включают тесты, рассмотренные выше. Обычно части или функциональные фрагменты молекул нуклеиновой кислоты имеют только небольшие делеции относительно полноразмерной молекулы нуклеиновой кислоты, например делеции менее 50, 40, 30, 20 или 10 нуклеотидов, например на 5'-конце, кодирующем N-конец белка, на 3'-конце, кодирующем С-конец белка, или внутри кодирующей области, хотя более значительные делеции, например по меньшей мере 60, 70, 80, 90, 100, 150, 200, 300, 400, 500, 600 или 700 нуклеотидов, или делеции менее 60, 70, 80, 90, 100, 150, 200, 300, 400, 500, 600 или 700 нуклеотидов также могут быть осуществлены, если фрагмент имеет такую же или по существу такую же активность (например, каталитическую или ферментативную активность), что и полноразмерный белок, как определено выше. Активность кодируемого полипептида или белка можно легко протестировать, чтобы определить, имеет ли он такую же активность, что и полноразмерный полипептид или белок, например как изложено выше.
Типичные части или фрагменты могут содержать по меньшей мере 50% и предпочтительно по меньшей мере 60, 70, 75, 80, 85, 90 или 95% смежных нуклеотидов нуклеотидной последовательности, представленной в SEQ ID NO: 1, 3 или 5. Типичные размеры части или фрагмента включают по меньшей мере 620, 700, 800, 850, 900, 950, 1000, 1050, 1100 и 1150 нуклеотидов.
Более короткие фрагменты молекулы нуклеиновой кислоты по изобретению можно использовать в качестве зондов, например для ПЦР или протоколов гибридизации. Более короткие фрагменты могут составлять, например, 10-30, 20-25 нуклеотидов в длину. Такие зонды полезны в протоколах идентификации дополнительных молекул нуклеиновой кислоты, которые имеют гомологию с молекулами нуклеиновой кислоты по изобретению.
Термин "молекула нуклеиновой кислоты" при использовании в данном описании изобретения относится к полимеру РНК или ДНК, который является одно- или двухцепочечным, возможно содержащему синтетические, неприродные или измененные нуклеотидные основания. Примеры таких полинуклеотидов включают среди прочего кДНК, геномную ДНК и дцРНК (двухцепочечную РНК). Предпочтительно, молекула нуклеиновой кислоты представляет собой ДНК.
Хотя последовательности нуклеиновой кислоты, относящиеся к данному описанию изобретения, содержат нуклеотид тимидин ("t"), понятно, что изобретение также относится к соответствующим последовательностям, где тимидин замещен уридином ("u").
Как отмечено выше, данное изобретение включает молекулы нуклеиновой кислоты, которые представляют собой варианты молекул нуклеиновой кислоты SEQ ID NO: 1, 3 или 5, конкретно функционально эквивалентные варианты. Таким образом, "вариантные" молекулы нуклеиновой кислоты могут иметь одно или множество нуклеотидных изменений по сравнению с молекулами нуклеиновой кислоты SEQ ID NO: 1, 3 или 5. Например, варианты могут иметь 1, 2, 3, 4 или 5 или более добавлений, замен, вставок или делеций нуклеотидов.
В еще одном аспекте изобретения предложен белок (или полипептид), обладающий активностью алкогольдегидрогеназы, конкретно MDH, как определено выше в данном описании изобретения.
Белок или полипептид предпочтительно представляет собой MDH или ее часть, обладающую активностью MDH. Более конкретно, эта часть сохраняет функцию или активность свойств MDH, из которой она имеет происхождение (как определено посредством ссылки на аминокислотную последовательность SEQ ID NO: 2, 4 или 6).
Белок или полипептид может быть альтернативно определен при помощи ссылки на кодирующие последовательности нуклеиновой кислоты, и как таковой белок или полипептид по изобретению может быть кодирован любыми молекулами нуклеиновой кислоты по изобретению, как описано выше.
Изобретение распространяется на функциональные части или фрагменты полноразмерных молекул белка, под которыми подразумевают части или фрагменты, которые обладают такой же или по существу такой же активностью, что и полноразмерные белки, как определено выше, т.е. их следует рассматривать как функционально эквивалентные варианты. Как отмечено в данном описании изобретения, свойство легко может быть проверено различными путями. Обычно эти функциональные фрагменты имеют лишь незначительные делеций относительно полноразмерной молекулы белка, например менее 50, 40, 30, 20 или 10 аминокислот, хотя, как отмечено выше в отношении молекул нуклеиновой кислоты, более значительные делеций, например вплоть до 60, 70, 80, 90, 100, 150, 200 аминокислот или по меньшей мере 60, 70, 80, 90, 100, 150, 200 аминокислот, могут быть подходящими. Во всех случаях фрагменты должны обладать такой же или по существу такой же активностью, что и полноразмерные белки, как определено выше, т.е. их следует рассматривать как функционально эквивалентные варианты. Эти делеций могут быть на N-конце, С-конце, или они могут представлять собой внутренние делеции.
Типичные части или фрагменты могут содержать по меньшей мере 50% и предпочтительно по меньшей мере 60, 70, 75, 80, 85, 90 или 95% смежных аминокислот аминокислотной последовательности, представленной в SEQ ID NO: 2, 4 или 6.
Таким образом, полипептид по изобретению, как определено выше, включает варианты последовательностей SEQ ID NO: 2, 4 или 6, например последовательности, обладающие конкретными уровнями идентичности последовательности с перечисленными последовательностями. Такие варианты могут представлять собой встречающиеся в природе варианты, такие как сопоставимые белки или гомологи, обнаруживаемые в других видах, или более конкретно варианты, обнаруживаемые в других микроорганизмах (которые имеют общие функциональные свойства кодируемого белка, как определено в данном описании изобретения).
Варианты встречающихся в природе полипептидов, как определено в данном описании изобретения, можно также получать синтетически, например, путем использования стандартных способов молекулярной биологии, которые известны в данной области техники, например стандартных способов мутагенеза, таких как сайт-направленный или случайный мутагенез (например используя перетасовку генов или допускающую ошибки ПЦР). Такие способы мутагенеза можно использовать для разработки ферментов, которые обладают улучшенными или другими каталитическими свойствами.
Также можно использовать производные полипептидов, как определено в данном описании изобретения. Под производным подразумевают полипептид, как он описан выше, или его вариант, который вместо встречающейся в природе аминокислоты содержит структурный аналог этой аминокислоты. Дериватизация или модификация (например, мечение, гликозилирование, метилирование аминокислот в белке) также может иметь место до тех пор, пока не оказывает нежелательного влияния на функцию белка.
Под "структурным аналогом" подразумевают нестандартную аминокислоту. Примерами таких нестандартных аминокислот или структурных аналогов, которые можно использовать, являются D-аминокислоты, изостеры амидов (такие как N-метиламид, ретро-инвертированный амид, тиоамид, тиоэфир, фосфонат, кетометилен, гидроксиметилен, фторвинил, (Е)-винил, метиленамино, метилентио или алкан), L-N-метиламинокислоты, D-α-метиламинокислоты, D-N-метиламинокислоты.
Идентичность последовательностей можно оценивать посредством любого подходящего способа. Однако для определения степени идентичности последовательности между последовательностями полезными являются компьютерные программы, которые выполняют множественные выравнивания последовательностей, например Clustal W (Thompson et al, (1994) Nucleic Acids Res., 22: 4673-4680). Программы, которые сопоставляют и выравнивают пары последовательностей, как ALIGN (Myers et al., (1988) CABIOS, 4: 11-17), FASTA (Pearson et al., (1988) PNAS, 85: 2444-2448; Pearson (1990), Methods Enzymol, 183: 63-98) и gapped BLAST (Altschul et al, (1997) Nucleic Acids Res., 25: 3389-3402), также являются полезными для этой цели. Кроме того, сервер Dali при Европейском институте биоинформатики предлагает выравнивания последовательностей белков на основе структур (Holm (1993) J. Mol. Biol, 233: 123-38; Holm (1995) Trends Biochem. Sci., 20: 478-480; Holm (1998) Nucleic Acids Res., 26: 316-9).
Множественные выравнивания последовательностей и вычисления процента идентичности могут быть осуществлены с использованием стандартных параметров BLAST (используя последовательности всех доступных организмов, матрицу Blosum 62, цена гэпов: наличие = 11, удлинение = 1). Альтернативно можно использовать следующие программу и параметры: Program: Align Plus 4, версия 4.10 (Sci Ed Central Clone Manager Professional Suite), сравнение ДНК: глобальное сравнение, стандартная линейная оценочная матрица, штраф за несовпадение = 2, штраф за открытие гэпа = 4, штраф за удлинение гэпа = 1. Сравнение аминокислот: глобальное сравнение, оценочная матрица BLOSUM 62.
В еще одном воплощении изобретения предложена конструкция, например рекомбинантная конструкция, содержащая молекулу нуклеиновой кислоты по изобретению, как определено в данном описании изобретения, функционально связанную с гетерологичной последовательностью контроля экспрессии. В этом контексте следует понимать, что последовательность контроля экспрессии будет гетерологичной (т.е. ненативной) молекуле нуклеиновой кислоты, более конкретно гетерологичной нуклеотидной последовательности, которая кодирует полипептид алкогольдегидрогеназу. В этой связи, когда кодирующая нуклеотидная последовательность не является встречающейся в природе последовательностью, последовательность контроля экспрессии будет гетерологична нуклеотидной последовательности, из которой она имеет происхождение. Как отмечено выше, можно использовать комбинации различных молекул нуклеиновой кислоты.
Такая последовательность контроля экспрессии обычно будет представлять собой промотор. Соответственно, конструкция предпочтительно будет содержать ненативный промотор, конкретно сильный ненативный промотор. Возможно, конструкция может дополнительно содержать еще один или более чем один ген и/или одну или более чем одну подходящую регуляторную последовательность. Возможные дополнительные один или более чем один ген могут находиться под контролем того же промотора, что и MDH-кодирующая молекула нуклеиновой кислоты по изобретению. Возможные одна или более чем одна регуляторная последовательность может представлять собой ненативную регуляторную последовательность (т.е. ненативную по отношению к кодирующей нуклеотидной последовательности или нуклеотидной последовательности).
В контексте данного изобретения термин "функционально связанный" относится к ассоциации двух или более молекул нуклеиновой кислоты в одном фрагменте нуклеиновой кислоты, так что функция одной подвергается воздействию другой. Например, промотор является функционально связанным с кодирующей последовательностью, когда он способен воздействовать на экспрессию этой кодирующей последовательности (т.е. кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующие последовательности могут быть функционально связанными с регуляторными последовательностями смысловой или антисмысловой ориентации.
Термин "регуляторные последовательности" относится к нуклеотидным последовательностям, расположенным "вверх по течению" (5' некодирующие последовательности), внутри или "вниз по течению" (3' некодирующие последовательности) кодирующей последовательности, и которые влияют на транскрипцию, процессинг или стабильность РНК или трансляцию ассоциированной кодирующей последовательности. Регуляторные последовательности могут включать промоторы, операторы, энхансеры и лидерные последовательности трансляции. При использовании в данном описании изобретения термин "промотор" относится к нуклеотидной последовательности, способной контролировать экспрессию кодирующей последовательности или РНК. В общем случае, кодирующую последовательность располагают в направлении 3' по отношению к последовательности промотора. Промоторы могут иметь происхождение во всей их полноте от нативного гена или состоять из разных элементов, имеющих происхождение от различных промоторов, встречающихся в природе, или даже содержать синтетические нуклеотидные сегменты. Кроме того, признается, что, так как в большинстве случаев точные границы регуляторных последовательностей не были определены полностью, фрагменты нуклеиновой кислоты различной длины могут обладать идентичной промоторной активностью.
В еще одном воплощении изобретения предложен вектор, содержащий молекулу нуклеиновой кислоты или конструкцию, как определено в данном описании изобретения.
Более конкретно, можно конструировать векторы, содержащие одну или более чем одну MDH-кодирующую молекулу нуклеиновой кислоты по изобретению (или конструкцию по изобретению). Выбор вектора может зависеть от микроорганизма-хозяина, способа, который будет использоваться для трансформации клеток-хозяев, способа, который используется для экспрессии белка, или от другого предполагаемого применения вектора. Специалисту хорошо известны генетические элементы, которые должны присутствовать в векторе для того, чтобы успешно трансформировать, подвергать селекции и размножать клетки-хозяева, содержащие MDH-кодирующую молекулу нуклеиновой кислоты или конструкцию по изобретению. Специалист также будет понимать, что различные независимые случаи трансформации будут приводить к различным уровням и паттернам экспрессии и, таким образом, что множественные случаи могут нуждаться в скрининге с целью получения клеток, проявляющих желаемый уровень и паттерн экспрессии. Такой скрининг можно осуществлять, среди прочего, саузерн-анализом ДНК, нозерн-анализом экспрессии мРНК, вестерн-анализом экспрессии белка.
В изобретении также предложен микроорганизм или хозяин, который может представлять собой любой организм-хозяин, как рассмотрено выше, например Е.coli, В.subtilis и С.glutamicum, содержащий одну или более чем одну молекулу нуклеиновой кислоты, конструкцию или вектор по изобретению. Хозяин подвергают генетической манипуляции, чтобы ввести или изменить экспрессию MDH. Это может быть достигнуто путем введения одной или более чем одной копии MDH-кодирующей молекулы нуклеиновой кислоты по изобретению под контролем ненативного, предпочтительно сильного, промотора. Таким образом, генетический материал присутствует в организме-хозяине, который не является встречающимся в природе организмом (т.е. присутствует экзогенный генетический материал).
В общем случае экзогенный генетический материал вводят, используя процесс трансформации. Трансформация обычно будет включать плазмиду или другой вектор, которые также будут содержать ген, позволяющий идентифицировать успешно трансформированные микроорганизмы, например ген устойчивости к антибиотику (например против ампициллина) или какой-нибудь другой маркер. Другие способы селекции трансформантов известны специалисту и включают использование светочувствительного вектора, гена lux, который вызывает свечение положительных колоний в темноте. Другие подходящие носители для трансформации бактерий включают космиды и молекулы бактериофагов.
Теперь изобретение будет дополнительно описано со ссылкой на следующие неограничивающие Примеры. Следует понимать, что эти Примеры, хотя и показывают воплощения изобретения, даны только в качестве иллюстрации. Из приведенного выше обсуждения и этих Примеров специалист в данной области техники сможет установить существенные характеристики этого изобретения и, без отклонения от сущности и объема изобретения, сможет выполнить различные изменения и модификации изобретения для адаптации его к различным применениям и условиям. Таким образом, различные модификации изобретения, в дополнение к уже показанным и описанным в данном описании изобретения, будут очевидны специалисту в данной области техники из предшествующего описания. Такие модификации также входят в объем прилагаемой формулы изобретения. Все документы, упоминаемые в данном описании изобретения, включены посредством ссылки.
В Примерах сделаны ссылки на следующие Фиг.:
Фиг. 1. Выравнивания нуклеотидных последовательностей для В.methanolicus mdh-MGA3, mdh2-MGA3, mdh3-MGA3, mdh-PB1, mdh1-PB1 и mdh2-PB1.
Фиг. 2. (А) Выравнивания первичных последовательностей предсказанных белков В.methanolicus MGA3 Mdh, Mdh2 и Mdh3 и PB1 Mdh, Mdh1 и Mdh2. (В) Выравнивания первичных последовательностей подсемейства mdh2/mdh3 (т.е. белков В.methanolicus MGA3 Mdh2 и Mdh3 и РВ1 Mdh2).
Фиг. 3. Каталитические активности очищенных Mdh (черный), Mdh2 (темно-серый) и Mdh3 (светло-серый) на различных спиртах (200 мМ), исследованные in vitro. (А) Субстратную специфичность анализировали in vitro для MDH из В.methanolicus MGA3. Спиртовые субстраты использовали в концентрациях 500 мМ, за исключением пентанола (300 мМ) и гексанола (50 мМ). Данные вычисляли по среднему значению из двух экспериментов, которые выполняли в трех повторностях. (В) Субстратную специфичность для MDH из В.methanolicus РВ1 анализировали in vitro. Спиртовые субстраты использовали в концентрациях 500 мМ, за исключением пентанола (300 мМ) и гексанола (50 мМ). Данные вычисляли по среднему значению из двух экспериментов, которые выполняли в трех повторностях.
Фиг. 4. (А) Определение in vitro температурного оптимума для каталитической активности Mdh, Mdh2 и Mdh3. (В) Определение оптимальных температурных условий для катализа посредством белков MDH из В.methanolicus РВ1 выполняли in vitro: удельную активность вычисляли для 500 мМ этанола; измерения выполняли в трех повторностях.
Фиг. 5. (А) Температурную стабильность MGA3 Mdh, Mdh2 и Mdh3 исследовали in vitro. Ферменты инкубировали при 45°С или при 60°С перед анализом фермента. (В) Температурную стабильность РВ1 Mdh, Mdh1 и Mdh2 исследовали in vitro. Ферменты инкубировали при 45°С или при 60°С перед анализом фермента.
Фиг. 6. Каталитическую активность Mdh, Mdh2 и Mdh3 исследовали в присутствии Act и сравнивали с уровнем активности, измеренном, когда Act не присутствовал. (А) Активацию MDH при помощи Act из В.methanolicus MGA3 исследовали in vitro. Исследования выполняли в трех повторностях с использованием 500 мМ спирта и 5 мкг/мл белков MDH и Act. (В) Активацию MDH при помощи Act из В.methanolicus РВ1 исследовали in vitro. Исследования выполняли в трех повторностях с использованием 500 мМ спирта и 5 мкг/мл белков MDH и Act.
Фиг. 7. (А) Стратегии клонирования. (В) Физическая карта плазмиды act-pHCMC04. ORF - открытая рамка считывания.
Фиг. 8. Активности in vitro рекомбинантных штаммов В.subtilis при исследовании с использованием этанола и метанола в качестве субстратов. Сокращения: Blanco - буфер для ресуспендирования; WT - хозяин В.subtilis 168 дикого типа; WT+Mdh2(EC) - лизат Е.coli с Mdh2, экспрессируемым из плазмиды рЕТ21а; рНСМС04 - В.subtilis 168 с пустым вектором рНСМС04; pHCMC04+Mdh2(Ec) - рНСМС04 с лизатом Е.coli с mdh2, экспрессируемым из плазмиды рЕТ21а; act-pHCMC04 - лизат В.subtilis 168 с Act, экспрессируемым из плазмиды рНСМС04; act-pHCMC04+Mdh2(Ec) - act-pHCMS04 с лизатом Е.coli с mdh2, экспрессируемым из плазмиды рЕТ21а; mdh1-pHCMC04 - лизат В.subtilis 168 с mdh1, экспрессируемым из плазмиды рНСМС04; mdhl-pHCMC04+Act(Ec) - mdh1-pHCMC04 с лизатом Ε.coli с act, экспрессируемым из плазмиды рЕТ21а; mdh2-pHCMC04 - лизат В.subtilis 168 с mdh2, экспрессируемым из плазмиды рНСМС04; mdh2-pHCMC04+Act(Ec) - mdh2-рНСМС04 с лизатом Е.coli c act, экспрессируемым из плазмиды рЕТ21а; mdh3-рНСМС04 - лизат В.subtilis 168 с mdh3, экспрессируемым из плазмиды рНСМС04; mdh3-pHCMC04+Act(Ec) - mdh3-pHCMC04 с лизатом Е.coli с act, экспрессируемым из плазмиды рЕТ21а; Amdh1-pHCMC04 - лизат В.subtilis 168 с act и mdh1, экспрессируемыми из плазмиды рНСМС04; Amdh2-pHCMC04 - лизат В.subtilis 168 с act и mdh2, экспрессируемыми из рНСМС04; Amdh3-рНСМС04 - лизат В.subtilis 168 с act и mdh3, экспрессируемыми из плазмиды рНСМС04.
Фиг. 9. Мас.%, фракции изотопомеров М1 различных метаболитов до (т.е. в нулевой точке отсчета) и после (т.е. в моменты времени 30 и 90 минут) добавления 13С-метанола. Три линии представляют результаты от трех независимых повторных биологических опытов. А: С.glutamicum delta aid pEKEX3; В: штамм С.glutamicum delta aid, который экспрессирует Mdh2 (pVWEx1-Mdh2), Hps и Phi (pEKEX3 - Hps + Phi). PEP: фосфоенолпируват; 2/3 Ρ: 2- и 3-фосфоглицерат; FBP: фруктозобисфосфат; R5P: рибозо-5-фосфат.
Фиг. 10. Метаболическое мечение с использованием 13С-метанола или 13С-формальдегида в качестве субстрата. (А) клетки ΔfrmA, экспрессирующие mdh2 и hps phi с использованием 13С-метанола в качестве источника углерода. (В) клетки ΔfrmA, экспрессирующие hps и phi с использованием 13С-формальдегида в качестве источника углерода.
Фиг. 11. Сборка синтетического оперона. Каждый последующий ген вводят в сайты рестрикции SwaI/BgIII. Последний ген в опероне содержит HiS6-метку. Для экспериментов 13С-мечения (см. Пример 18) использовали плазмиды AMAhx1B- и AMABGFTPrpe-pHCMC04. RBS: сайт связывания рибосомы.
Фиг. 12. Использовали SDS-PAGE очищенных белков, экспрессированных в В.subtilis 168. Использовали штаммы В.subtilis, которые содержали любую из конструкций, показанных на Фиг. 11. Белки очищали с использованием колонки HisTrap и концентрировали с использованием колонок Vivaspin. Белковые полосы отмечены в прямоугольниках. М: маркер молекулярной массы.
Фиг. 13. Мас.%, фракции изотопомеров М1 различных метаболитов до (т.е. в нулевой точке отсчета) и после (т.е. в моменты времени 30 и 90 минут) добавления 13С-метанола. Две линии представляют результаты от двух повторных независимых биологических опытов. (А): В.subtilis 168 рНСМС04; (В): В.subtilis 168 AM3Ahx1B-pHCMC04; (С): В.subtilis 168 АМ3 ABGFTPrpe-рНСМС04. PEP: фосфоенолпируват; 2/3 PG: 2- и 3-фосфоглицерат; FBP: фруктозобисфосфат.
ПРИМЕРЫ


Материалы и методы
Биологические материалы, манипуляции с ДНК и условия роста
Бактериальные штаммы и плазмиды, используемые в данном исследовании, перечислены в Таблице 1. В качестве стандартного хозяина для клонирования использовали Е.coli DH5, в то время как Е.coli ER2566 использовали в качестве хозяина для рекомбинантной экспрессии белков MDH, Act и NudF. Штаммы Ε.coli в основном выращивали при 37°С в жидкой или на твердой среде Луриа-Бертани (LB) (Sambrook (2001) Cold Spring Harbor Laboratory Press) с добавлением ампициллина (100 мкг/мл) или хлорамфеникола (10 мкг/мл) при необходимости. Процедуры рекомбинантной Е.coli выполняли, как описано в Sambrook and Russell (2001; Cold Spring Harbor Laboratory Press). ПЦР выполняли посредством применения системы для ПЦР Expand High Fidelity (Roche Applied Science, Indianapolis, IN), и секвенирование ДНК выполняли посредством Eurofins MWG Operon (Ebersberg, Germany, www.eurofinsdna.com). Выделение тотальной ДНК В.methanolicus MGA3 и РВ1 и рекомбинантное продуцирование белков MDH, Act и NudF в Е.coli ER2566 выполняли, как описано ранее (Brautaset et al, (2004) J Bacteriol 186(5): 1229-1238; Brautaset et al., (2010) Appl Microbiol Biotechnol 87(3): 951-964). Трансформацию В.methanolicus MGA3 выполняли путем электропорации (Jakobsen et al, (2006) J Bacteriol 188(8): 3063-3072). Клетки В.methanolicus выращивали при 50°C в 100 мл среды МеОН200, содержащей 200 мМ метанола, в среде Mann10, содержащей 10 г/л маннита, или в среде SOBsuc (Jakobsen, Benichou et al. (2006) J Bacteriol 188(8): 3063-3072) и при необходимости добавляли хлорамфеникол (5 мкг/мл).
Конструирование экспрессирующих векторов
рЕТ21а_mdh-MGA3, рЕТ21а_mdh2-MGA3, рЕТ21а_mdh3-MGA3 и рЕТ21а_act-MGA3:
Из-за высокого сходства последовательностей между кодирующим областями MGA3 mdh2 и mdh3, праймеры для ПЦР-амплификации и сопутствующего клонирования конструировали на основе уникальных последовательностей, представляющих окружающие области соответствующих генов, и являются следующими:
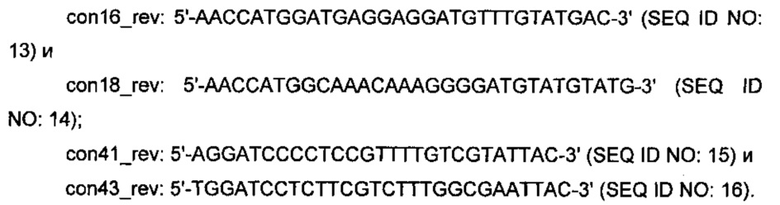
Соответствующие фрагменты ДНК расщепляли при помощи NcoI + BamHI (сайтов распознавания, подчеркнутых в праймерных последовательностях) и лигировали в соответствующие сайты pLITMUS28, получая в результате плазмиду рТМВ1, несущую mdh2, и рТМВ2, несущую mdh3. Затем клонированные гены MDH в обеих плазмидах секвенировали. Далее кодирующие области mdh и act ПЦР-амплифицировали из тотальной ДНК B.methanolicus MGA3, а кодирующие области mdh2 и mdh3 ПЦР-амплифицировали из плазмид рТМВ1 и рТМВ2, соответственно, путем использования следующих пар ПЦР-праймеров:

В прямых и обратных праймерах подчеркнуты сайты рестрикции для NdeI и XhoI, соответственно. Полученные продукты ПЦР mdh-MGA3 (1149 п.о. (пар оснований)), mdn2-MGA3 (1163 п.о.), mdh3-MGA3 (1165 п.о.) и act-MGA3 (570 п.о.) непосредственно А/T-лигировали в общий клонирующий вектор pGEM-T и соответствующие клонированные вставки подтверждали путем секвенирования ДНК. Затем полученные векторы расщепляли при помощи XhoI и NdeI и вставки лигировали в соответствующие сайты в рамке с использованием последовательности метки 6-His плазмиды рЕТ21а, получая плазмиды pET21a_mdh-MGA3, pET21a_mdh2-MGA3, pET21a_mdh3-MGA3 и pET21a_act-MGA3, соответственно.
pET21a_mdh-PB1, pET21a_mdh1-PB1, pET21a_mdh2-PB1 и pET21a_act-РВ1:
Кодирующие области генов mdh-PB1, mdh1-ΡΒ1 и mdh2-PB1 ПЦР-амплифицировали из тотальной ДНК РВ1 путем использования следующих пар праймеров:

Три полученных продукта ПЦР mdn-PB1 (1164 п.о.), mdh-PB1 (1164 п.о.) и mdh2-PB1 (1170 п.о.) А/Т-лигировали в плазмиду pGEM-T. Полученные плазмиды расщепляли при помощи XhoI и NdeI (сайты рестрикции, подчеркнутые в праймерах) и лигировали в соответствующие сайты плазмиды рЕТ21а, получая плазмиды pET21a_mdh-PB1, pET21a_mdh1-PB1 и pET21a_mdh2-PB1, соответственно. Кодирующую область act-PB1 ПЦР-амплифицировали из тотальной ДНК РВ1 путем использования пары праймеров:
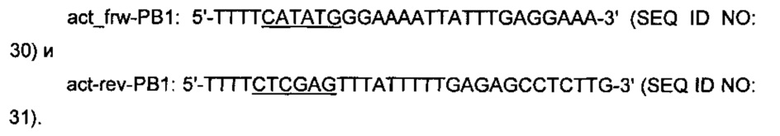
Продукт ПЦР act-PB1 расщепляли при помощи NdeI и XhoI (сайты рестрикции, подчеркнутые в праймерах) и лигировали в соответствующие сайты рЕТ21а, имея результатом плазмиду pET21a_act-PB1.
рЕТ21а_nudF:
Кодирующую область nudF ПЦР-амплифицировали из тотальной ДНК B.subtilis 168 путем использования следующей пары праймеров:

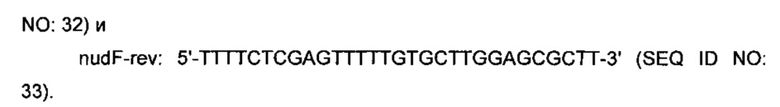
Полученный продукт (572 п.о.) ПЦР А/Т-лигировали в pGEM-T и клонированную вставку подтверждали посредством секвенирования ДНК. Полученный вектор расщепляли при помощи XhoI и NdeI (сайты рестрикции, подчеркнутые в праймерах) и вставку лигировали в соответствующие сайты рЕТ21а, получая в результате плазмиду pET21a_nudF.
Все сконструированные векторы трансформировали в хозяина экспрессии Е.coli ER2566.
Аффинная очистка рекомбинантных белков
Шесть различных белков MDH, два различных белка Act и NudF очищали от клеточных экстрактов соответствующих рекомбинантных штаммов Е.coli ER2566 путем применения аффинной хроматографии, по существу как описано ранее (Brautaset et al, (2010) Appl Microbiol Biotechnol 87(3): 951-964). Концентрации белка оценивали спектрофотометрически на спектрофотометре NanoDrop, (Nano Drop Technologies, Wilmington, Delaware) с настройками по молекулярной массе и коэффициенту экстинкции, вычисленными для белков MDH, Act и NudF (данные не показаны) с применением инструмента Expasy ProtParam (expasy.org/tools/protparam.html) (Gasteiger et al. (2003) Nucleic Acids Res. 31(13): 3784-3788). Чистоту очищенных белков анализировали посредством электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) (Sambrook и Russel, (2001) Cold Spring Harbor Laboratory Press) с последующим визуальным контролем полученных изображений. Очищенные белки быстро замораживали в жидком N2 и хранили при -80°С до тех пор, пока их не размораживали на льду и использовали в биохимических анализах.
Анализы фермента
Активности алкогольдегидрогеназ измеряли спектрофотометрически, по существу как описано ранее (Hektor, Kloosterman et al. (2002) Chem 277(49): 46966-46973), и реакционная смесь содержала: 100 мМ глицин-KOH рН 9,5 (если не указано иное), 5 мМ MgSO4, 0,5 мМ НАД+ и 500 мМ спирта (метанол, этанол, пропанол, 1,3-пропандиол или бутанол). НАД+ замещали равными концентрациями НАДФ+ (никотинамидадениндинуклеотидфосфат), ФМН+ (флавинмононуклеотид) и ФАД+ (флавинадениндинуклеотид), если указано. Реакционная смесь для измерения активности формальдегидредуктазы содержала: 50 мМ фосфатно-калиевый буфер рН 6,7; 0,15 мМ НАДН, 1 мМ ДТТ (дитиотреитол) и 11,6 мМ (0,1-116 мМ) формальдегид. Компоненты для анализа смешивали в кювете и предварительно нагревали до 45°С, если не указано иное. Реакцию начинали посредством добавления 5-40 мкг очищенных белков MDH и за продуцированием НАДН наблюдали при 340 нм в течение 4 минут. Одну единицу активности MDH определяли как количество фермента, необходимого для продуцирования 1 мкмоль NADH в минуту в условиях, описанных выше. Очищенные белки Act (0,1-40 мкг) или NudF (20 мкг) добавляли к реакционной смеси, как указано в тексте.
Биохимическая характеристика in vitro очищенных ферментов
Очищенные белки MDH и Act (20 мкг) использовали в кинетических экспериментах, выполняя анализы метанолдегидрогеназы и формальдегиддегидрогеназы, по существу как описано выше. Для определения Km для метанола (Km,MeOH) и Vмакс концентрацию НАД+ сохраняли на уровне насыщения (0,5 мМ или 0,15 мМ), в то время как концентрацию метанола варьировали (0,1-2000 мМ). Для определения Km для НАД+ (Km,НАД+) и Vмакс концентрацию метанола поддерживали постоянной на уровне насыщения (500 мМ), в то время как концентрацию НАД+ варьировали (5-1000 мкМ). Для определения Km для формальдегида (Km,FA) и Vмакс концентрацию НАДН поддерживали на уровне насыщения (0,5 мМ или 0,15 мМ), в то время как концентрацию формальдегида варьировали (0,1-40 мМ). Act (20 мкг) добавляли к реакционным смесям для определения значений Km,MeOH и Vмакс в присутствии этого активатора, как указано в тексте. В общем случае, наклоны зависимости активности от времени были линейными в течение измеряемого периода (данные не показаны). Значения Km и Vмакс вычисляли с применением нелинейной регрессии при помощи программы для решения уравнений Microsoft Excel для согласования измеренных данных с уравнением Михаэлиса-Ментен, как описано ранее (Jakobsen et al., (2009) Appl Environ Microbiol 75(3): 652-661). Значения, полученные из регрессии, сравнивали затем со значениями, полученными из графиков Лайнуивера-Берка и Hanes-Woolf, чтобы убедиться, что был обнаружен глобальный минимум, а не локальный минимум.
Выделение тотальной РНК, синтез кДНК и ПЦР в реальном времени
Эксперименты с ПЦР в реальном времени выполняли, по существу как описано ранее (Brautaset et al., (2010) Appl Microbiol Biotechnol 87(3): 951-964). Тотальную РНК выделяли из клеточных культур MGA3 и РВ1 во время экспоненциальной фазы роста (ОП600 (оптическая плотность при 600 нм) равна 1,0) с маннитом или метанолом в качестве единственного источника углерода, используя набор RNAqueous (Ambion). Концентрацию РНК определяли на спектрофотометре NanoDrop (Nano Drop Technologies, Wilmington, Delaware) и целостность тотальной РНК оценивали при помощи Agilent Bioanalyzer 2100 и RNA 6000 Nano LabChip Kit (Agilent Technologies, Palo Alto, CA). кДНК синтезировали из выделенной тотальной РНК, используя набор для синтеза однонитевой кДНК (Amersham) согласно инструкциям производителя, и использовали в качестве матриц для экспериментов с ПЦР в реальном времени. ПЦР-анализы в реальном времени выполняли, используя ABI PRISM 7700 Sequence Detection System с ее стандартными настройками (Applied Biosystems, Foster City, CA, USA). Используемые для ПЦР праймеры выбирали при помощи программы Primer Express 2.0 (Applied Biosystems), и они представляли собой следующие праймеры:

Относительную дискретизацию рассматриваемых генов выполняли посредством нормализации результатов относительно 16s РНК (эндогенный контроль) и выборки калибратора, используя сравнительный Ct способ (способ 2-AACt), как описано ранее (Heid, Stevens et al. (1996) Genome Res 6(10): 986-994; Jakobsen, Benichou et al. (2006) J Bacteriol 188(8): 3063-3072; Brautaset, Jakobsen et al. (2010) Appl Microbiol Biotechnol 87(3): 951-964). Относительные разности в уровнях транскриптов трех генов определяли посредством вычисления значений ACT, заданных следующим образом: mdh2 (Ct mdh2 - Ct mdh), и значения ACT из mdh3 (Ct mdh3 - Ct mdh). Эффективность праймеров трех генов испытывали до выполнения других экспериментов.
3D-моделирование предсказанных белков MDH
Структурные модели белков Mdh и Mdh2 MGA3 создавали, используя сервер полностью автоматизированного гомологичного моделирования структуры белка SWISS-MODEL (http://swissmodel.expasv.org/) (Peitsch (1995) Bio-Technology 13(7): 658-660; Arnold, Bordoli et al. (2006) Bioinformatics 22(2): 195-201; Kiefer, Arnold et al. (2009) Nucleic Acids Res 37(Database issue): D387-392). Из-за высокой гомологии между предсказанными первичными структурами белков Mdh2 и Mdh3 MGA3 поиск модели для Mdh3 не выполняли. Поиски с gapped BLAST (Altschul et al. (1997) J Mol Biol 215(3): 403-410; Schaffer et al. (2001) Nucleic Acids Res. 29(14): 2994-3005) имели результатом 9 удачных попыток с общими матрицами со значениями Е, варьирующимися от 1⋅e-98 (pdb:3bfj, 1,3-пропандиолоксидоредуктаза) до 1⋅е-17 (pdb: 1oj7, Ε.coli K12 YQHD) для Mdh и от 1⋅е-112 (pdb:3bfj) до 2⋅е-14 (pdb: 1oj7) для Mdh2. 3D-выравнивания файлов матриц с использованием программы просмотра Deep view/Swiss pdb (Guex and Peitsch (1997) Elerctrophoresis 18(15): 2714-2723) показали, что все они имели очень похожие складки, и структурные модели на основе матрицы 3bfj, которые имели самый высокий показатель сходства аминокислот как для Mdh, так и для Mdh2, выбирали для представления Mdh и Mdh2. Программу просмотра Deep view/Swiss pdb использовали также для визуализации структурных моделей MDH.
Пример 1: Генетическая организация генов метанолдегидрогеназы и активаторного белка в штаммах В.methanolicus дикого типа MGA3 и РВ1
Скрининг in silico геномной последовательности В.methanolicus MGA3 (Heggeset et al, 2011) идентифицировал mdh, кодируемый плазмидой рВМ19, обозначенный в данном описании изобретения mdh-MGAZ, и два более предполагаемых MDH-кодирующих гена в геноме MGA3, обозначенных в данном описании изобретения mdh2-MGA3 и mdh3-MGA3, локализованных в хромосоме вдали друг от друга. Кодирующие последовательности mdh2-MGA3 и mdh3-MGA3 были на 96% идентичны друг другу и на 65% и 66% идентичны, соответственно, кодирующей последовательности mdh-MGA3. Выравнивание первичных последовательностей предсказанных полипептидов Mdh2-MGA3 и Mdh3-MGA3 выявило, что они на 96% идентичны друг другу и на 61% и 62% идентичны, соответственно, Mdh-MGA3 (Фиг. 2).
Недавно авторами изобретения были получены результаты ферментации метанола с подпиткой субстратом, демонстрирующие, что два штамма В.methanolicus дикого типа MGA3 и РВ1 существенно отличаются в отношении метилотрофных свойств. Проверка геномной последовательности РВ1 подтвердила наличие трех различных MDH-кодирующих генов и одного гена act, аналогичного MGA3. Ген mdn-PB1, локализованный на плазмиде рВМ20, был на 92% идентичен гену mdh-MGA3 MGA3, и соответствующие генные продукты демонстрировали 93%-ную идентичность первичных последовательностей (Фиг. 2). В отличие от MGA3 последовательности двух хромосомных генов РВ1, обозначенных mdh1-PB1 и mdh2-PB1, были не очень похожими. Ген mdh1-PB1 кодировал предполагаемый белок Mdh1 с 92%-ной идентичностью первичной последовательности с белком Mdh MGA3, в то время как mdh2-PB1 кодировал предполагаемый белок Mdh2 с 91%-ной и 92%-ной идентичностью первичной последовательности с белками Mdh2 и Mdh3 MGA3, соответственно. На основе этих анализов последовательностей кажется, что MGA3 и РВ1 обладают двумя подтипами MDH-кодирующих генов: "mdh/mdh1-типом" и "mdh2/mdh3-типом". MGA3 имеет один ген mdh/mdh1-типа (рВМ19) и два гена mdh2/mdh3-типа (хромосома), в то время как РВ1 имеет два гена mdh/mdh1-типа (рВМ20 и хромосома) и один ген mdh/mdh3-типа (хромосома). Биологическое воздействие этих различий дополнительно исследовали ниже.
Пример 2: 3D-моделирование показывает, что В.methanolicus MDH принадлежат типу III суперсемейства Fe-НАД-зависимых алкогольдегидрогеназ
Предсказанные первичные последовательности Mdh, Mdh2 и Mdh3 MGA3 подвергали сравнениям последовательностей с белками в базах данных, используя BLAST (Altschul, Gish et al. (1990) J Mol Biol 215(3): 403-410), показывая, что они предположительно принадлежат типу III алкогольдегидрогеназ (ADH) (de Vries, Arfman et al. (1992) J Bacteriol 174(16): 5346-5353), который представляет собой суперсемейство железосодержащих ADH. Наиболее близким гомологом MDH с известной 3D-структурой являлась 1,3-пропандиолдегидрогеназа из Klebsiella pneumoniae (PDB ID: 3BF), которая демонстрировала 46%-ную идентичность первичной последовательности с Mdh и 52%-ную идентичность первичной последовательности с Mdh2 и Mdh3. Эта 1,3-пропандиолдегидрогеназа представляет собой тип III Fe-НАД-зависимых алкогольдегидрогеназ, которые катализируют превращение 3-гидроксипропиональдегида в 1,3-пропандиол (1,3-PD). Структура MDH В.methanolicus С1 была ранее анализирована посредством электронной микроскопии, и было сделано заключение, что она представляет собой декамер, в котором 10 субъединиц объединены в два кольца по 5 (Vonck, Arfman et al. (1991) J Biol Chem 266(6): 3949-3954). Что интересно, недавно экспериментально было продемонстрировано, что 1,3-PD-дегидрогеназа имеет аналогичную четвертичную структуру. Основываясь на этом, авторы изобретения решили использовать информацию о выясненной 3D-структуре 1,3-PD-дегидрогеназы, чтобы прогнозировать 3D-структуру Mdh В.methanolicus MGA3, больше узнать о сайтах каталитической активности в НАД-зависимой алкогольдегидрогеназе. Первичную аминокислотную последовательность Mdh MGA3 передавали в Swissmodel и конструировали модель. Мономеры 1,3-PD-дегидрогеназы складывались в два структурных домена, которых разделяет углубление. N-концевой домен содержит сайт связывания кофактора НАД+, а С-концевой домен включает остатки, вовлеченные в связывание железа. Консервативный мотив GGGSX2DX2K, вовлеченный в связывание кофактора НАД+, был обнаружен в N-концевой области MDH В.methanolicus С1. Этот мотив также присутствует в Mdh В.methanolicus MGA3 в положении 95-104, и он был также обнаружен в N-концевой области 1,3-PD-дегидрогеназы из K.pneumoniae. Область 258-290 MDH В.methanolicus С1 содержала несколько остатков His, и поэтому прогнозировали, что она вовлечена в связывание металла. Это хорошо согласуется с полученными данными о 1,3-PD-дегидрогеназе из K.pneumoniae, где обнаружили 4 остатка, ответственных за координационное положение металла железа. Эти остатки являются консервативными и соответствуют остаткам Asp193, His197, His262 и His276 в Mdh В.methanolicus MGA3 и наиболее вероятно являются остатками, ответственными за связывание цинка в этом ферменте. В заключение, эти данные указывают, что MDH В.methanolicus принадлежат к типу III Fe-НАД-зависимой алкогольдегидрогеназы, что подкреплено экспериментальными результатами, полученными в данном исследовании (см. ниже).
Пример 3: Все очищенные белки MDH из MGA3 и РВ1 демонстрировали in vitro активность НАД-зависимой MDH
Кодирующие области mdh-MGA3, mdh2-MGA3, mdh3-MGA3, mdh-PB1, mdh1-PB1 и mdn2-PB1 ПЦР-амплифицировали и клонировали в вектор рЕТ21а Е.coli, получая в результате экспрессионные плазмиды pET21a_mdh-MGA3, pET21a_mdh2-MGA3, pET21a_mdh3-MGA3, pET21a_mdh-PB1, pET21a_mdh-PB1 и pET21a_mdh2-PB1, соответственно. В полученных векторах рекомбинантные гены транскрибируют из сильного промотора Т7 и сливают в рамке с кодирующей последовательностью с меткой 6-His на их 3'-концах для упрощения очистки. Гены act MGA3 и PBI и аналогичный ген nudF В.subtilis клонировали аналогичным образом в рЕТ21а, получая в результате плазмиды pET21a_act-MGA, pET21a_act-PB1 и pET21a-nudF, соответственно (Таблица 1). Все сконструированные экспрессионные векторы трансформировали в Е.coli ER2566 и полученные рекомбинантные штаммы культивировали во встряхиваемых колбах для продуцирования соответствующих рекомбинантных белков. Белки очищали посредством аффинной хроматографии до чистоты выше 95%, что было оценено с помощью SDS-PAGE (данные не показаны), и белки Act и NudF хранили для более позднего использования (см. ниже).
Затем шесть очищенных белков МОН анализировали с использованием метанола в качестве субстрата, и результаты показали, что все ферменты являются каталитически активными (см. Фиг. 3). Для исключения того, что эти белки могут использовать альтернативные кофакторы, повторяли анализы с замещением НАД+ на ФАД+, ФМН+ и НАДФ+. Во всех случаях каталитическая активность обнаружена не была (данные не показаны), что подтверждает, что ни один из альтернативных кофакторов не может использоваться MDH в этих условиях. Эти результаты демонстрировали, что оба штамма В.Methanolicus, MGA3 и РВ1, имеют три различных гена, один из которых локализован на плазмиде, а два других локализованы в хромосоме, и все они кодируют активные и НАД-зависимые MDH.
Пример 4: Все MDH имеют in vitro широкие субстратные специфичности и различные предпочтения в отношения спиртов
Очищенные белки MDH исследовали на каталитические активности посредством использования нескольких альтернативных спиртов, и все ферменты демонстрировали активности на этаноле, пропаноле, бутаноле, пентаноле, гексаноле, изопропаноле и 1,3-пропандиоле в качестве субстратов (Фиг. 3). Неожиданно, относительные каталитические активности на большинстве этих альтернативных субстратов были существенно выше, чем по отношению к метанолу, для всех шести MDH. Относительные каталитические активности на каждом отличном спирте существенно варьировали среди трех MDH, указывая на различные субстратные предпочтения среди этих белков. Например, активности ферментов Mdh3-MGA3 и Mdh2-PB1 по отношению к пропанолу были примерно в 25-35 раз выше их активности на метаноле. Что интересно, эти два фермента демонстрировали значительно более высокую каталитическую активность, чем остальные ферменты, на всех этих субстратах в условиях испытания. Все шесть ферментов также демонстрировали активности формальдегид- и ацетальдегидредуктазы, которые дополнительно исследовали ниже. Основываясь на этих данных, было весьма заманчиво классифицировать эти белки в качестве ADH, а не MDH, способных катализировать превращение широкого диапазона различных первичных и вторичных спиртов в альдегиды или кетоны.
Пример 5: Белки MDH демонстрировали in vitro схожие рН и температурные оптимумы
С целью установления достоверных условий анализа для сравнительного определения биохимических характеристик шесть белков MDH анализировали в отношении рН и температурных оптимумов. Из-за более высоких по сравнению с метанолом каталитических активностей MDH на этаноле (см. Фиг. 3) авторы изобретения проводили эти эксперименты с этанолом в качестве субстрата для увеличения чувствительности данных. Ранее сообщалось, что белок Mdh из штамма В.methanolicus С1 имеет оптимум рН 9,5 (Kloosterman, Vrijbloed et al. (2002) J Biol Chem 277(38): 34785-34792), и поэтому шесть очищенных MDH (20 мкг) испытывали на активность при рН в диапазоне от 8,5 до 10,5. Все ферменты демонстрировали самую высокую каталитическую активность при рН от 9,5 до 10 (данные не показаны).
Далее MDH анализировали при рН 9,5 на активность при температурах в диапазоне от 25°С до 50°С, и результаты показали, что все они имели температурные оптимумы от 45°С до 50°С (Фиг. 4). Основываясь на этих данных, все дальнейшие анализы MDH проводили при рН 9,5 и 45°С.
Пример 6: Mdh3-MGA3 и Mdh2-PB1 демонстрируют in vitro более высокие температурные стабильности по сравнению с остальными MDH
Термостабильность шести MDH испытывали путем предварительной инкубации белков при 45°С и 60°С и образцы отбирали для анализов ферментов в различные моменты времени. Как и ожидали, все ферменты сохраняли по существу всю каталитическую активность при предварительных инкубациях при 45°С (Фиг. 5). Каталитические активности Mdh-MGA3, Mdh2-MGA3, Mdh-PB1 и Mdh1-PB1 сильно снижались (вплоть до 90%) при предварительных инкубациях при 60°С в течение 6 минут, в то время как эта обработка предположительно оказывала лишь умеренные отрицательные воздействия на каталитические активности Mdh-MGA3 и Mdh2-PB1 (Фиг. 5). Селекцию экспериментов повторяли в присутствии равных количеств очищенного Act, и это не оказывало влияния на температурную стабильность какого-либо из MDH (данные не показаны).
Пример 7: Все шесть белков MDH каталитически стимулируются in vitro посредством Act
Геномные последовательности как MGA3, так и РВ1 имели только один ген act, расположенный на хромосоме, схожий с геном act, клонированным ранее из MGA3 (Brautaset, Jakobsen et al. (2004) J Bacteriol 186(5): 1229-1238). Таким образом, было интересно исследовать, могут ли соответствующие белки Act стимулировать in vitro каталитическую активность всех белков MDH. Для установления достоверных условий сначала Mdh-MGA3 испытывали вместе с Act-MGA3 при различных относительных концентрациях белков (1:2-20:1) и с использованием метанола в качестве субстрата. Полная активация достигалась при относительной концентрации от 1:1 до 5:1, и ингибирования не наблюдали вследствие относительно высоких концентраций активатора (данные не показаны). Для дальнейшего испытания всегда использовали равные концентрации MDH и Act (1:1). Далее аналогичные анализы выполняли со всеми шестью MDH, используя метанол в качестве субстрата, и данные показали, что активности MDH индуцировались в 5-7 раз для MDH MGA3 (Фиг. 6А) и в 4-10 раз для MDH РВ1 (Фиг. 6В) в присутствии Act.
Затем авторы изобретения проводили аналогичные анализы, но с использованием этанола в качестве субстрата, и результаты показали, что каталитические активности повышались в 6-8 раз для MDH MGA3 (Фиг. 6В) и в 2-5 раз для MDH РВ1 (Фиг. 6В) в присутствии Act. Что интересно, при использовании формальдегида или ацетальдегида в качестве субстратов присутствие Act не вызывало значительного стимулирования каталитических активностей для какой-либо из MDH (данные не показаны). Таким образом, Act увеличивает in vitro отношение дегидрогеназной активности к редуктазной активности для всех шести белков MDH.
Пример 8: Mdh можно также каталитически стимулировать in vitro посредством белка NudF В.subtilis
Было обнаружено, что гены nudix гидролазы широко распространены в бактериальных геномах, и ген act В.methanolicus является единственным известным членом этого семейства, кодирующим регуляторный белок. Продукт гена nudF В.subtilis, NudF, демонстрирует в целом 33%-ную идентичность первичной последовательности с Act, и было подтверждено экспериментально, что NudF принадлежит к подсемейству АДФ-рибозопирофосфатаз. NudF и Act идентичны по остаткам, в отношении которых документально подтверждено, что они являются важными для связывания субстрата и/или ингибитора, связывания металла и для каталитического сайта. Было исследовано, можно ли NudF заменять Act при активации in vitro MDH В.methanolicus. Рекомбинантный штамм Ε.coli ER2566 (pET21a-nudF) культивировали для рекомбинантного продуцирования и сопутствующей очистки NudF. Mdh-MGA3 выбирали в качестве модельного белка и испытывали вместе с NudF, используя этанол в качестве субстрата, как описано выше, и результаты ясно демонстрировали, что активность Mdh стимулировалась одинаково хорошо (примерно 8-кратно) как при помощи NudF, так и при помощи Act в тех же условиях (данные не показаны). Этот результат впервые показывает, что гетерологичные Nudix гидролазы могут функционировать в качестве активаторного белка, и это также должно оказывать влияние на наше современное понимание биологической роли многообразного класса белков.
Пример 9: MDH имеют in vitro схожие значения Vмакс и Km,MeOH в отсутствие Act
MDH подвергали исследованию в отношении кинетических характеристик для определения значений Vмакс и значений Km, и для получения биологически релевантных данных эти эксперименты проводили с использованием метанола в качестве субстрата. Три белка MDH анализировали в отношении начальных скоростей реакции при оптимизированных условиях анализа, как описано выше, и с варьированием концентраций метанола (см. раздел "Материалы и методы"), и данные показали, что они демонстрировали схожую и нелинейную кинетику Михаэлиса-Ментен. Эти результаты согласуются с аналогичной биохимической характеристикой MDH из В.methanolicus С1 (Hektor, Kloosterman et al. (2002) Chem 277(49): 46966-46973; Kloosterman, Vrijbloed et al. (2002) J Biol Chem 277(38): 34785-34792). Эти авторы предполагали, что MDH в неактивированном состоянии демонстрирует механизм реакции типа "пинг-понг", при котором редокс-активный кофактор функционирует в качестве временного депо электронов, тогда как MDH в активированном состоянии катализирует независимую от кофактора реакцию, которая демонстрирует механизм реакции типа тройного комплекса. Значения Km и Vмакс вычисляли посредством применения нелинейной регрессии при помощи программы для решения уравнений Microsoft Excel для согласования измеренных данных с уравнением Михаэлиса-Ментен. Значения Кт для метанола были схожими и составляли от 150 мМ до 250 мМ для MDH MGA3 и от 160 мМ до 220 мМ для MDH РВ1. Соответствующие значения Vмакс составляли от 0,04 МЕ/мг до 0,09 МЕ/мг для MDH MGA3 и составляли от 0,013 до 0,065 МЕ/мг для MDH РВ1 (Таблица 2). Вместе эти данные указывают на то, что кинетические константы для всех шести MDH являются относительно похожими в условиях испытания. Авторы изобретения также выбрали три MDH MGA3 для испытания в отношении начальных скоростей реакции с варьированием концентраций НАД+, показывая, что они демонстрировали линейную кинетику Михаэлиса-Ментен (данные не показаны). Из нелинейного приближения уравнения Михаэлиса-Ментен было определяло, что значения Km,НАД+ составляют от 14 мкМ до 40 мкМ (Таблица 2).
Таблица 2: Кинетические константы очищенных MDH В.methanolicus in vitro в присутствии и в отсутствие Act. Анализы выполняли при 45°С и при рН 9,5.
MGA3:
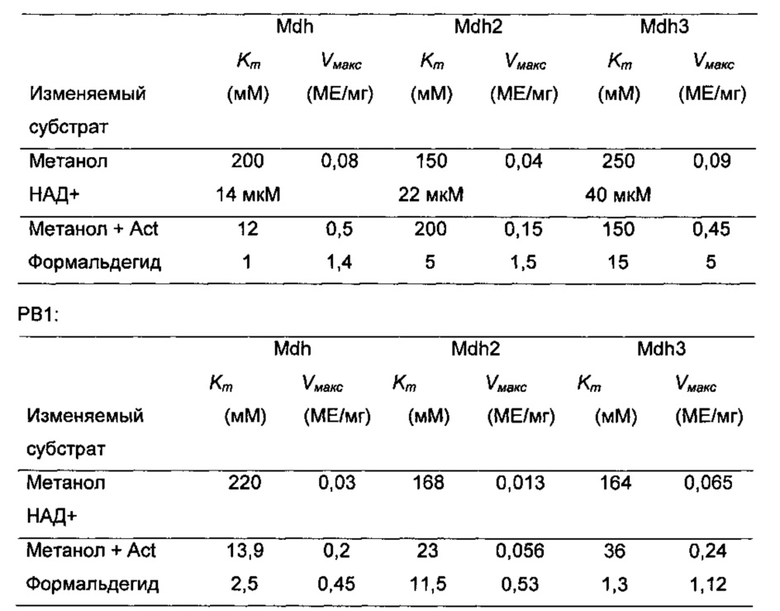
Пример 10: Значения Vмакс для MDH повышаются в 4-6 раз в присутствии Act
Далее осуществляли кинетические эксперименты с использованием равных концентраций MDH и Act в реакционных смесях (20 мкг MDH + 20 мкг Act). Значения Vмакс повышались в 4-6 раз по сравнению с полученными из анализов ферментов без Act (Таблица 2), подтверждая, что каталитические активности всех шести MDH стимулируются посредством Act. Эти данные согласовываются с данными, представленными на Фиг. 6.
Пример 11: Значения Km,MeOH для Mdh-MGA3, Mdh-ΡΒ1 и Mdh1-PB1 существенно (вплоть до в 70 раз) снижаются в присутствии Act
Что интересно, Km,MeOH резко снижалась (в 17 раз) до 12 мМ для Mdh-MGA3, когда Act добавляли в реакционную смесь, в то время как соответствующие значения Km,MeOH как для Mdh2-MGA3, так и для Mdh3-MGA3 оставались по существу такими же, как и при испытании без Act. Для трех ферментов РВ1 она была разной. Для Mdh-PB1 и Mdh1-PB1 значения Km,MeOH существенно снижались (в 16 раз и 70 раз, соответственно) в присутствии Act, и это значение умеренно (в 4 раза) снижалось посредством Act для Mdh2-PB1 (Таблица 2). Что интересно, белки MGA3-Mdh, PB1-Mdh и PB1-Mdh1 были внесены авторами изобретения в одну подгруппу MDH на основе выравнивания последовательностей (см. выше), и биологическое влияние этих результатов исследования было рассмотрено (см. ниже).
Пример 12: MDH обычно имеет более высокие значения Vмакс, и более низкие значения Km для формальдегида по сравнению с метанолом
Биологическое значение MDH для окисления метанола во время метилотрофного роста является однозначным, в то время как биологическая роль этого фермента в качестве системы детоксикации формальдегида в потребляющих метанол клетках исследована в меньшей степени. В описании изобретения было продемонстрировано, что все ферменты проявляли активности как формальдегид-, так и ацетальдегидредуктазы (см. выше), и авторы изобретения решили охарактеризовать это свойство кинетически. При использовании формальдегида в качестве субстрата значения Km равны 1 мМ, 5 мМ и 15 мМ, а соответствующие значения Vмакс равны 1,4 МЕ/мг, 1,5 МЕ/мг и 5 МЕ/мг для белков Mdh, Mdh2 и Mdh3 MGA3, соответственно (Таблица 2). Для белков РВ1 значения Km для формальдегида равнялись 2,5 мМ, 4 мМ и 1,3 мМ, соответственно, а соответствующие значения Vмакс равнялись 0,45 МЕ/мг, 0,53 МЕ/мг и 1,12 МЕ/мг, соответственно. Взятые вместе эти результаты показывают, что все шесть MDH обычно имеют более высокое сродство и более высокую Vмакс, когда субстратом является формальдегид, по сравнению с тем, когда субстратом является метанол.
Пример 13. Три гена mdh транскрибируют с различными уровнями в клетках В.methanolicus во время экспоненциальной Фазы роста
Ранее был продемонстрировано, что транскрипция mdh-MGA3 предположительно является очень высокой в клетках В.methanolicus и немного повышена (примерно в 3 раза) в клетках, растущих на метаноле по сравнению с растущими на манните, в то время как уровни транскрипта act были аналогичны при тех и других условиях роста (Jakobsen et al., (2006) J Bacteriol 188(8): 3063-3072). В данном описании все три MDH-кодирующих гена из MGA3 были включены в подобный анализ, и результаты показали, что относительные уровни транскрипции mdh-MGA3 и mdn2-MGA3 были в 2 раза и в 3 раза выше при росте на метаноле по сравнению с ростом на манните. Уровень транскрипта mdh3-MGA3 был по существу схожим при двух различных условиях роста. Для mdh этот результат был немного другим по сравнению с предшествующими данными (Jakobsen et al., (2006) J Bacteriol 188(8): 3063-3072), и причина этого неизвестна. Что интересно, соответствующие значения Ct, полученные в стандартизированных условиях для трех генов, были очень разными, причем mdh-MGA3 явно демонстрировал самое низкое значение, указывая на самые высокие уровни транскриптов. Было обнаружено, что Ct mdh2 - Ct mdh составляет 8, a Ct mdh3 - Ct mdh составляет 14 (принимая во внимание примерно 100%-ную эффективность праймера во всех этих экспериментах, эти числа должны означать, что уровни транскриптов mdh2-MGA3 и mdh3-MGA3 примерно в 250 раз и в 10000 раз ниже, чем уровень транскрипта mdh, соответственно, при условиях испытания).
Кодирующие последовательности mdh2-MGA3 и mdh3-MGA3 на 96% идентичны на уровне ДНК, и для исключения любой перекрестной гибридизации праймеров rt-ПЦР в этих экспериментах соответствующие пары праймеров rt-ПЦР (см. раздел "Материалы и методы") испытывали по отношению к плазмидным ДНК, рТМВ1 и рТМВ2, несущим соответствующие последовательности генов mdh2 и mdh3. Результаты ясно показывают, что определимых продуктов ПЦР получено не было, когда использовали mdh2-специфичные праймеры вместе с ДНК рТМВ2 или, альтернативно, когда использовали mdh3-специфичные праймеры вместе с матрицей ДНК рТМВ1 (данные не показаны). Эти данные подтвердили, что праймеры rt-ПЦР, применяемые для mdh2-MGA3 и mdn3-MGA3, являются специфичными по отношению к их соответствующим мишеням, подтверждая, что полученные данные должны быть достоверными.
Выполняли аналогичный анализ генов mdh из РВ1, и результаты показали, что уровни транскриптов mdn-PB1 и mdh2-PB1 были по существу подобными при росте на манните по сравнению с ростом на метаноле. Неожиданно, уровень транскрипта mdh1-PB1 на манните был в 14 раз выше, чем на метаноле, и биологическое влияние этого осталось неизвестным. Как и для MGA3, авторы изобретения выявили, что относительный уровень транскриптов этих трех генов в РВ1 предположительно был очень разным, и ген mdh-PB1 транскрибировался до более высоких уровней, чем mdh1-ΡΒ1 и mdh2-РВ1 (данные не показаны).
Пример 14: Экспрессия генов mdh В.methanolicus в Е.coli
Конструирование экспрессионных векторов
Гены, кодирующие Mdh и активаторный белок Act для Mdh, амплифицировали из плазмид рЕТ21а, несущих гены из штаммов В.methanolicus MGA3 (mdh-MGA3, mdn2-MGA3, mdn3-MGA3 и act-MGA3) и PB1 (mdh-PB1, mdh1-PB1 и mdh-РВ1). Затем гены клонировали или в плазмиду pSEVA424 (гены mdh) или в плазмиду pSEVA131 (ген act). Для клонирования mdn-MGA3, mdh-PB1, mdh1-PB1 и act-MGA3 использовали сайты рестрикции EcoRI и HindIII, в то время как mdn2-MGA3, mdn2-PB1 и mdh3-MGA3 клонировали с использованием сайтов рестрикции EcoRI и PstI. Полученные экспрессионные векторы трансформировали в электрокомпетентную Е.coli дикого типа K-12 (BW25113) и в Ε.coli K-12 (BW25113) с делетированным геном frmA
Эксперименты по экспрессии
Для экспериментов по экспрессии клетки культивировали или в среде Луриа-Бертани (LB) для анализов in vitro или в среде М9 для анализов in vivo, каждая из которых содержала 20 мкг/мл стрептомицина для pSEVA424. Когда коэкспрессировался Act, среду дополняли 100 мкг/мл ампициллином. Экспрессию индуцировали, когда клетки достигали ОП 0,5 (для испытаний in vitro) или ОП 1 (для испытаний in vivo), путем добавления 0,1 мМ IPTG (изопропилтиогалактозид) (конечная концентрация) в течение 6 часов. Затем клетки собирали посредством центрифугирования. Для анализов in vitro неочищенный экстракт клеток получали путем лизирования клеток во Френч-прессе после ультрацентрифугирования. Альтернативно, клетки ресуспендировали в среде М9 без глюкозы для измерения активности in vivo.
Анализы ферментов
Измерение активности Mdh in vitro: для определения активности Mdh в неочищенном экстракте клеток за Mdh-зависимым образованием НАДН следили при 340 нм. Анализы выполняли или при 37°С или 45°С в предварительно нагретых буферных растворах. Образец Mdh для анализа содержал 10-20 мкг фермента, 50 мМ буфер K2HPO4, рН 7,4; 2,5 мМ MgCl и 0,5 мМ НАД+ (конечные концентрации). После 5 мин предварительной инкубации реакцию начинали с 1 M метанола (конечная концентрация).
Измерение активности Mdh in vivo: для определения активности Mdh в суспензиях клеток клетки собирали после индукции посредством IPTG, промывали и ресуспендировали в среде М9 без глюкозы и IPTG. ОП600 устанавливали на 1 для нормализации. Анализ выполняли при 37°С или при 45°С в водяной бане-шейкере. Анализ начинали посредством добавления 1 Μ метанола и последующего измерения аккумуляции в супернатанте формальдегида, образующегося в результате катализируемого метанолдегидрогеназой окисления метанола. Рассчитанные активности были основаны на предположении, что 1 л культуры с ОП, равной 1, содержит 0,3 г биомассы, 50% которой составляет белок.
Результаты
In vivo и in vitro активности различных Mdh из штаммов В.methanolicus MGA3 и РВ1, рекомбинантно экспрессируемых в Е.coli, представлены в Таблице 3 ниже. Act клонировали из В.methanolicus MGA3.
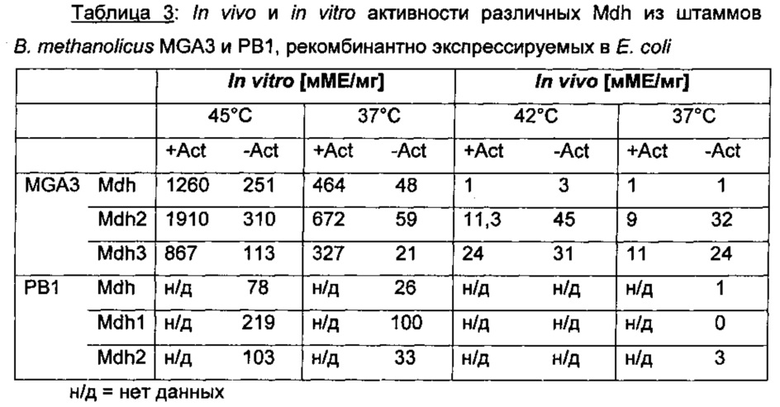
Все Mdh демонстрировали in vitro более высокую активность при 45°С, чем при 37°С. Кроме того, активности in vitro Mdh из MGA3 резко повышались, когда коэкспрессировался Act. Эффект температуры in vivo был значительно меньше по сравнению с эффектом in vitro, и благоприятного влияния Act обнаружено не было. MGA3-Mdh2 демонстрировал в общем самые высокие активности как in vitro, так и in vivo среди всех испытанных генов. Для Mdh из РВ1 ситуация выглядит по-другому. В данном исследовании Mdh1, который является структурно близкородственным в отношении Mdh из MGA3, показал самую высокую активность для большей части условий испытаний. Неожиданно все 3 Mdh из РВ1 продемонстрировали отсутствие активности или очень слабую активность при испытании in vivo. Эти данные являются неожиданными, так как активности in vitro при 37°С выглядят перспективно. Причина этого неясна. На основе доступных данных кажется, что mdh2-MGA3 является в целом самым лучшим выбором для получения максимальной метанолдегидрогеназной активности в Е.coli при испытаниях как in vitro, так и in vivo.
Пример 15: Экспрессия генов mdh из В.methanolicus в В.subtilis
Конструирование экспрессионных векторов
Все стадии клонирования выполняли, используя клетки Е.coli DH5. Ген act-MGA3 клонировали из геномной ДНК В.methanolicus MGA3 с использованием прямого праймера, который содержит сайт связывания рибосом (RBS) mntA В.subtilis, и обратного праймера, который содержит короткий линкер, содержащий сайты рестрикции SwaI и BgIII, и метку 6-His (Фиг. 7А). Ген включали в плазмиды рНВ201 и рНСМС04, используя сайты рестрикции SpeI и BamHI. Таким же образом клонировали ген mdn-MGA3 из плазмиды рВМ19 из В.methanolicus MGA3, а гены mdh2-MGA3 и mdh3-MGA3 клонировали из геномной ДНК В.methanolicus MGA3. Эти три гена также лигировали в плазмиды рНВ201 и рНСМС04.
Для конструирования векторов для коэкспрессии act и трех различных генов метанолдегидрогеназы гены метанолдегидрогеназы ПЦР-амплифицировали при помощи прямого праймера, который содержит стоп-кодон и RBS mntA В.subtilis, и обратного праймера, который содержит короткий линкер, содержащий сайты рестрикции SwaI и BgIII (Фиг. 7В). Затем расщепляли концы соответствующих генов при помощи StuI и BgIII и их лигировали в сайты SwaI и BgIII вектора act-pHB201. Таким образом, стоп-кодон вводят после гена act, и гены метанолдегидрогеназы теперь содержат метку 6-His. После секвенирования гены переносили в плазмиду рНСМС04 (Фиг. 7В), используя сайты рестрикции SpeI и BamHI. Вставки подтверждали посредством секвенирования.
Создание рекомбинантных клеток В.subtilis, экспрессирующих метанолдегидрогеназу
Клетки В.subtilis 168 трансформировали с использованием экспрессионных плазмид act-MGA3, mdh-MGA3, mdh2-MGA3 и mdh3-MGA3 и аналогичных векторов, коэкспрессирующих каждый из генов метанолдегидрогеназы с геном act-MGA3. Положительные колонии отбирали из чашки и плазмиду выделяли и проверяли посредством рестрикции на положительные клоны. Колонии отбирали из чашки и выращивали в течение ночи при 37°С (250 об/мин) и разбавляли до ОП600, равной 0,1, в 20 мл LB, содержащей хлорамфеникол (5 мкг/мл). После 3 ч роста при 37°С (250 об/мин) к культуре добавляли 500 мкл 40% ксилозы для индукции экспрессии. Культуру выращивали в течение еще 3 ч и отбирали образцы по 2 мл. Образец осаждали путем центрифугирования в течение 2 мин, 11000×g, и удаляли супернатант. Осадок ресуспендировали в 300 мкл Birnboim А для В.subtilis (10 мМ Трис-HCl рН 8,0; 20% сахарозы; 50 мМ NaCl; 0,25 мг/мл лизоцима; ингибитор протеазы) и инкубировали при 37°С в течение 30 мин. Образцы хранили при -80°С до использования. Кроме того, авторы изобретения воспользовались рекомбинантными клетками Е.coli, сконструированными в рамках экспрессии mdh-MGA3, mdh2-MGA3, mdh3-MGA3 и act-MGA3 из плазмиды рЕТ21а в анализах in vitro.
Анализы in vitro на активность метанолдегидрогеназы
Активности белков Mdh измеряли посредством отслеживания образования НАДН спектрофотометрически. Реакционная смесь содержала:
- 100 мкл смеси глицин-KOH (рН 9,5);
- 100 мкл 5 M метанола (или 5 M этанола);
- 5 мкл 1 M MgSO4;
- 10 мкл 50 мМ НАД+; -10 мкл образца;
- (10 мкл лизата Е.coli act-pET21a);
- 775 мкл H2O.
Кюветы и реакционную смесь без клеточного лизата предварительно инкубировали при 50°С. За образованием НАДН следили в течение 10 мин при 340 нм при 50°С. Общую активность вычисляли путем деления увеличения единиц абсорбции в минуту на коэффициент экстинкции (6,23 см-1 мм-1) и общую концентрацию белка в МЕ/мг общего белка. Все анализы выполняли с использованием как метанола, так и этанола в качестве альтернативных субстратов.
Результаты
Испытывали все три гена, mdh-MGA3, mdh2-MGA3, и mdh3-MGA3, экспрессирующие активные метанолдегидрогеназы в хозяине В.subtilis, в то время как один act-MGA3 не экспрессирует обнаружимую активность метанолдегидрогеназы (Фиг. 8). В целом активности были значительно выше при использовании этанола вместо метанола в качестве субстрата для всех трех испытываемых генов (это похоже на то, что наблюдали, когда эти ферменты были очищены от рекомбинантных клеток Е.coli). Во всех случаях активности метанолдегидрогеназы значительно стимулировались посредством Act. Кажется, что когда act и гены метанолдегидрогеназы коэкспрессируются из одной и той же плазмиды в В.subtilis 168, белок Mdh2 является наиболее активным. Однако авторы изобретения заметили, что когда Act поступает из лизатов Ε.coli, тогда гены mdh-MGA3 являются наиболее активными. Таким образом, ген mdh2-MGA3, коэкспрессированный вместе с act-MGA3, является в целом самым лучшим выбором для максимальной метанолдегидрогеназной активности в В.subtilis при испытании in vitro.
Пример 16: Включение метанола в генетически сконструированную С.glutamicum
Эксперименты 13С-мечения
Для экспериментов мечения авторы изобретения использовали штамм С.glutamicum delta aid, который экспрессирует Mdh2-MGA3 (pVWEx1-Mdh2), Hps и Phi (рЕКЕХ3 - Hps + Phi). В качестве отрицательного контроля использовали штамм С.glutamicum delta aid с отсутствующей плазмидой pEKEX3. Все штаммы С.glutamicum выращивали на среде М9, содержащей (в литре) 3,48 г Na2HPO4⋅12 H2O, 0,60 г KH2PO4, 0,51 г NaCl, 2,04 г NH4Cl, 0,10 г MgSO4, 4,38 мг CaCl2, 15 мг Na2EDTA⋅2 H2O, 4,5 мг ZnSO4⋅7 H2O, 0,3 мг CoCl2⋅6 H2O, 1 мг MnCl2⋅4 H2O, 1 мг H3BO3, 0,4 мг Na2MoO4⋅2 H2O, 3 мг FeSO4⋅7 H2O и 0,3 мг CuSO4⋅5 H2O. Для всех культивирований (в М9 или LB) использовали 1 мМ IPTG в качестве индуктора и добавляли в среду 100 мкг/мл спектиномицина и 25 мкг/мл канамицина в качестве маркеров устойчивости. Все культивирования выполняли при 30°С. Штаммы в виде клеток высевали из клеточной линии в глицерине на чашку с агаровой средой LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl и 16 г/л агара) и впоследствии выращивали в жидкой среде LB в течение 6 ч. Жидкие прекультуры, содержащие среду М9 с 3 г/л рибозы, инокулировали из культур в LB с конечной ОП600 0,6 в течение 12 ч. Для экспериментов мечения жидкие культуры, содержащие среду М9, инокулировали из культур в М9 (плюс рибоза) с конечной ОП600 0,8. Один образец культивирования отбирали перед добавлением 13С-метанола (т.е. в нулевой точке отсчета), затем добавляли 40 мМ 13С-метанол и два образца культивирования отбирали в моменты времени 30 и 90 мин. Для того чтобы гасить метаболическую активность и экстрагировать внутриклеточные метаболиты, образцы культивирования направляли в холодный (-20°С) раствор смеси ацетонитрил/метанол/0,1 M муравьиная кислота (40/40/20 об./об.). Паттерны мечения внутриклеточных метаболитов измеряли, используя систему Dionex ICS 2000 (Dionex, Sunnyvale, USA), соединенную с тройным квадрупольным масс-спектрометром QTrap 4000 (Applied Biosystems, Foster City, USA).
Результаты
Как и ожидалось, мечения в массовых фракциях изотопомеров М1 (т.е. молекул, которые имеют один меченый атом углерода) в штамме дикого типа после импульса 13С-метанола обнаружено не было (Фиг. 9А). Значительное включение меток в метаболиты наблюдали у мутанта, экспрессирующего две рекомбинантные реакции из РМФ-пути (Hps и Phi) и НАД-зависимую метанолдегидрогеназу из В.methanolicus MGA3 (Mdh2-MGA3) (Фиг. 9В). Хотя процент мечения повышался у фруктозобисфосфата и рибозо-5-фосфата в период времени от 30 до 90 минут, он оставался постоянным у фосфоенолпирувата и 2/3-фосфоглицерата. Эти данные ясно демонстрируют, что введенный метилотрофный путь функционирует in vivo, приводя к ассимиляции метанола в центральном метаболизме углерода.
Следует отметить, что в этих и дальнейших экспериментах, описанных ниже, технические ограничения означают, что невозможно было проверить 13С-мечение формальдегида непосредственно. Однако, благодаря одновременной экспрессии находящихся "вниз по течению" ферментов РМФ-пути можно было анализировать включение 13С в метаболиты и, таким образом, опосредованно обнаруживать ассимиляцию 13С-меченного метанола. Кроме того, активность рекомбинантно экспрессируемого Mdh была продемонстрирована in vitro в описанных выше экспериментах, см., например, Примеры 3-6 выше.
Пример 17: включение метанола в генетически сконструированную Е.coli
Результаты
Авторы изобретения использовали эксперименты метаболического мечения, чтобы доказать, что Mdh, Hps и Phi являются функциональными в живых клетках. Клеткам, экспрессирующим все три белка, давали или 13C-помеченный метанол или 13С-формальдегид, и можно было продемонстрировать включение обоих С1-соединений в некоторые метаболиты.
Для экспериментов авторы изобретения использовали клетки Е.coli, лишенные гена формальдегиддегидрогеназы (ΔfrmA), экспрессирующие mdh2, hps и phi или одни hps и phi из различных плазмид pSEVA (424 и 131). Все гены, используемые для экспериментов, имели происхождение из В.methanolicus MGA3. Прекультуры получали при 37°С в оптимизированной минимальной среде М9, содержащей рибозу в качестве единственного источника углерода. Для экспериментов клетки переносили в свежую среду М9 без рибозы. Эксперименты начинали путем добавления либо 13С-меченного метанола, либо формальдегида. Для контроля включения образцы отбирали в различные моменты времени, метаболизм останавливали путем холодного гашения и образцы впоследствии анализировали посредством анализа ЖХ-МС (жидкостная хроматография-масс-спектрометрия).
Когда метанол добавляли в качестве единственного субстрата (Фиг. 10А), мечение можно было обнаружить в нескольких метаболитах, таких как пентозо-5-фосфаты, гексозо-6-фосфаты, фосфоенолпируват и ацетил-СоА. Более глубокий анализ выявил то, что несколько метаболитов, необходимых для РМФ-цикла, например пентозо-5-фосфаты, продемонстрировали включение соединений с множественным мечением углеродов, что указывает на действие полного функционального цикла. Когда использовали формальдегид (Фиг. 10В), включение меченых С-атомов также происходило (фактически оно происходило быстрее по отношению к включению метанола, позволяя предположить, что Hps и Phi действуют быстрее, чем Mdh, или что количество молекул С5-предшественников, необходимых для включения формальдегида, или концентрация формальдегида, продуцируемого из метанола, была ограничена). В контрольном эксперименте с использованием 13С-меченного метанола в качестве субстрата было обнаружено, что экспрессия одних Hps и Phi не дает возможности для ассимиляции 13С-меченного метанола (данные не показаны).
Полученные данные ясно показывают, что три введенных метилотрофных модуля экспрессируются функционально. Авторы изобретения также показывают, что экспрессия трех белков приводит к включению метанола и формальдегида в биомассу посредством установленного РМФ-цикла. Используя метанол или формальдегид в качестве источника углерода, авторы изобретения смогли показать, что ассимиляция формальдегида с помощью Hps и Phi идет гораздо быстрее, чем ассимиляция метанола, но может быть ограничена доступностью молекул С5-предшественников.
Пример 18: включение метанола в генетически сконструированную В.subtilis
Конструирование экспрессионных векторов
Все стадии клонирования выполняли, используя клетки Е.coli DH5α. Ген act-MGA3 клонировали из геномной ДНК В.methanolicus MGA3 при помощи прямого праймера, который содержит сайт связывания рибосом (RBS) mntA В.subtilis, и обратного праймера, который содержит короткий линкер, содержащий сайты рестрикции SwaI и BgIII, и метку 6-His (Фиг. 7А). Ген включали в плазмиду рНВ201, используя сайты рестрикции SpeI и BamHI.
Для конструирования векторов для коэкспрессии act, mdh3, hxIA, hxIB, glpX, Iba, tkt, pfk и rpe гены ПЦР-амплифицировали при помощи прямого праймера, который содержит стоп-кодон и RBS mntA В.subtilis, и обратного праймера, который содержит короткий линкер, содержащий сайты рестрикции SwaI и BgIII (Фиг. 11). Для амплификации использовали act и mdh3 геномной ДНК В.methanolicus MGA3, для генов glpX, iba, tkt, pfk и rpe использовали плазмиду рВМ19 из В.methanolicus MGA3, а для генов hxIA и hxIB использовали геномную ДНК В.subtilis 168. Затем расщепляли концы соответствующих генов при помощи StuI и BgIII и их лигировали в сайты SwaI и BgIII вектора. Гены последовательно вводили в вектор, тем самым собирая синтетический оперон девяти генов (Фиг. 11). Таким образом, стоп-кодон вводят после каждого гена, и последний ген в синтетическом опероне теперь содержит метку His6. После введения каждого гена в синтетический оперон правильность вставки подтверждали посредством секвенирования. После секвенирования синтетические опероны переносили в плазмиду рНСМС04 (Фиг. 7В), используя сайты рестрикции SpeI и BamHI. Вставки подтверждали посредством секвенирования.
Экспрессия генов РМФ-пути в В.subtilis 168
Для создания рекомбинантных клеток В.subtilis, экспрессирующих ферменты РМФ-пути, клетки В.subtilis 168 трансформировали при помощи векторов, содержащих гены РМФ-пути (Фиг. 7А). Положительные колонии отбирали из чашки и плазмиду выделяли и проверяли посредством рестрикции на положительные клоны. Колонии отбирали из чашки и выращивали в течение ночи при 37°С (250 об/мин) и разбавляли до ΟΠ600, равной 0,1, в 100 мл среды MSR (25 г/л дрожжевого экстракта, 15 г/л триптона, 3 г/л K2HPO4, 1% глюкозы), дополненной витаминами и хлорамфениколом (5 мкг/мл). После 3 ч роста при 37°С (250 об/мин) к культуре добавляли 1,25 мл 40% ксилозы для индукции экспрессии. Культуру выращивали в течение еще 3 ч и клетки осаждали путем центрифугирования в течение 10 мин, 4000×g, и удаляли супернатант. Осадок промывали с использованием среды MSR, ресуспендировали в 4 мл Birnboim А для В.subtilis (буфер для лизиса, содержащий 2 мМ Трис-HCl (рН 7,4), 20% сахарозы, 50 мМ NaCl и 0,25 мг/мл лизоцима) и инкубировали при 37°С в течение 30 минут. Образец центрифугировали в течение 20 мин при 13000×g при 4°С и супернатант использовали для очистки на HisTrap.
Фракции очищенного белка объединяли и концентрировали, используя колонки Vivaspin (товарный знак) (GE Healthcare). Очищенные белки визуализировали посредством SDS-PAGE и окрашивания Кумасси (Фиг. 12). Авторы изобретения показали, что каждый белок, который содержит метку Hise, экспрессировался из синтетических оперонов (Фиг. 11).
Эксперименты 13С-мечения
Для экспериментов мечение авторы изобретения использовали штамм В.subtilis, который экспрессирует Act, Mdh3, HxIA и HxIB, и штамм, который экспрессирует Act, Mdh3, HxIA, HxIB, GIpX, Fba, Tkt, Pfk и Rpe. В качестве отрицательного контроля использовали штамм В.subtilis с отсутствующей плазмидой рНСМС04. Все штаммы В.subtilis выращивали на среде М9, содержащей (в литре) 3,48 г Na2HPO4⋅12 H2O, 0,60 г KH2PO4, 0,51 г NaCl, 2,04 г NH4Cl, 0,10 г MgSO4, 4,38 мг CaCl2, 15 мг Na2EDTA⋅2 H2O, 4,5 мг ZnSO4⋅7 H2O, 0,3 мг CoCl2⋅6 H2O, 1 мг MnCl2⋅4 H2O, 1 мг H3BO3, 0,4 мг Na2MoO4⋅2 H2O, 3 мг FeSO4⋅7 H2O и 0,3 мг CuSO4⋅5 H2O, 10 г ксилозы (т.е. индуктора) и 5 мг хлорамфеникола. Все культивирования выполняли при 37°С. Штаммы в виде клеток высевали из клеточной линии в глицерине на чашку с агаровой средой LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl и 16 г/л агара), содержащей 5 мкг/мл хлорамфеникола, и впоследствии выращивали в жидкой среде LB + хлорамфеникол в течение приблизительно 5 ч. Жидкие прекультуры, содержащие среду М9, инокулировали из культур в LB с конечной ΟΠ600 от 1,4 до 1,8. Один образец культивирования отбирали перед добавлением 13С-метанола (т.е. в нулевой точке отсчета), затем добавляли 40 мМ 13С-метанол и два образца культивирования отбирали в моменты времени 30 и 90 минут. Для того, чтобы гасить метаболическую активность и экстрагировать внутриклеточные метаболиты, образцы культивирования направляли в холодный (-20°С) раствор смеси ацетонитрил/метанол/0,1 M муравьиная кислота (40/40/20 об./об.). Паттерны мечения внутриклеточных метаболитов измеряли, используя систему Dionex ICS 2000 (Dionex, Sunnyvale, USA), соединенную с тройным квадрупольным масс-спектрометром QTrap 4000 (Applied Biosystems, Foster City, USA).
Результаты
Как и ожидалось, мечения в массовых фракциях изотопомеров М1 (т.е. молекул, которые имеют один меченый атом углерода) в штамме дикого типа после импульса 13С-метанола обнаружено не было (Фиг. 13А). Кроме того, мечение не было обнаружено у мутанта, экспрессирующего две рекомбинантные реакции из РМФ-пути (HxIA и HxIB), активаторный белок (Act) и НАД-зависимую метанолдегидрогеназу (Mdh3), но у которого ни один из генов пентозофосфатного пути (РРР) не сверхэкспрессировался (Фиг. 13В). Однако когда гены РРР сверхэкспрессировались в дополнение к предыдущим четырем генам, было обнаружено включение значительного мечения (Фиг. 13С). Эти данные ясно демонстрируют, что введенный метилотрофный путь функционирует in vivo, приводя к ассимиляции метанола в центральном метаболизме углерода. Эти результаты также показывают, что поступление молекул С5-предшественников посредством РРР является критическим элементом для включения метанола в В.subtilis. Однако это можно облегчить посредством сверхэкспрессии генов, кодирующих связанные с РРР ферменты.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПРОДУКЦИИ БЕЛКА | 2006 |
|
RU2435863C2 |
| БАКТЕРИЯ РОДА BACILLUS, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2002 |
|
RU2299907C2 |
| КОРИНЕФОРМНАЯ БАКТЕРИЯ, ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2004 |
|
RU2333247C2 |
| Микроорганизм, продуцирующий молочную кислоту, и способ продуцирования молочной кислоты с его использованием | 2016 |
|
RU2711983C2 |
| НАПРАВЛЕННАЯ МОДИФИКАЦИЯ МАЛАТДЕГИДРОГЕНАЗЫ | 2013 |
|
RU2658437C2 |
| МИКРООРГАНИЗМЫ ДЛЯ ПОВЫШЕННОГО ПРОДУЦИРОВАНИЯ АМИНОКИСЛОТ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2015 |
|
RU2688486C2 |
| ПОЛИНУКЛЕОТИД НА ОСНОВЕ ГЕНА MDH, ОБЛАДАЮЩИЙ ПРОМОТОРНОЙ АКТИВНОСТЬЮ, И ЕГО ПРИМЕНЕНИЕ | 2022 |
|
RU2825450C2 |
| ВЕКТОР ДЛЯ НОКАУТА ГЕНА АЦЕТАТКИНАЗЫ В Clostridium thermocellum, КЛЕТКА-ХОЗЯИН, ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЙ МИКРООРГАНИЗМ Clostridium thermocellum, СПОСОБ ПОЛУЧЕНИЯ ТАКОГО МИКРООРГАНИЗМА И СПОСОБ ПРЕОБРАЗОВАНИЯ ЛИГНОЦЕЛЛЮЛОЗНОЙ БИОМАССЫ В ЭТАНОЛ. | 2008 |
|
RU2541785C2 |
| НОВАЯ ФИТАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ЕЕ ПРИМЕНЕНИЕ | 2015 |
|
RU2712881C2 |
| СПОСОБ ПОЛУЧЕНИЯ 2,4-ДИГИДРОКСИБУТИРАТА | 2013 |
|
RU2645260C2 |
























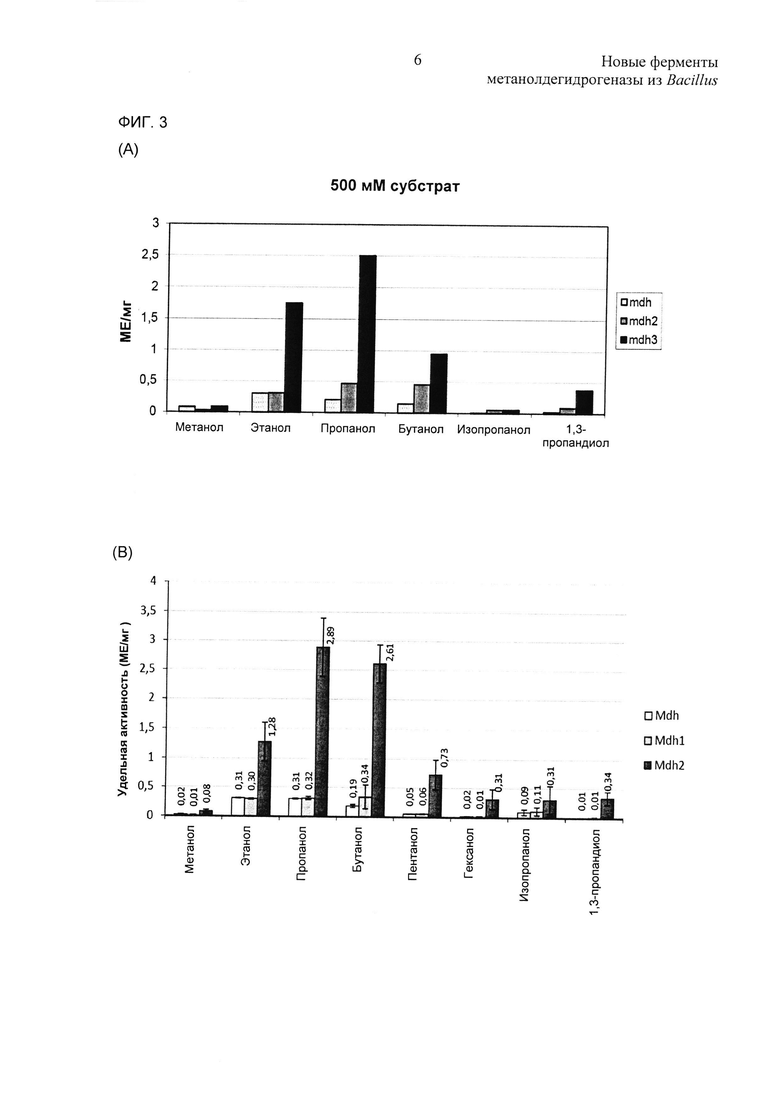
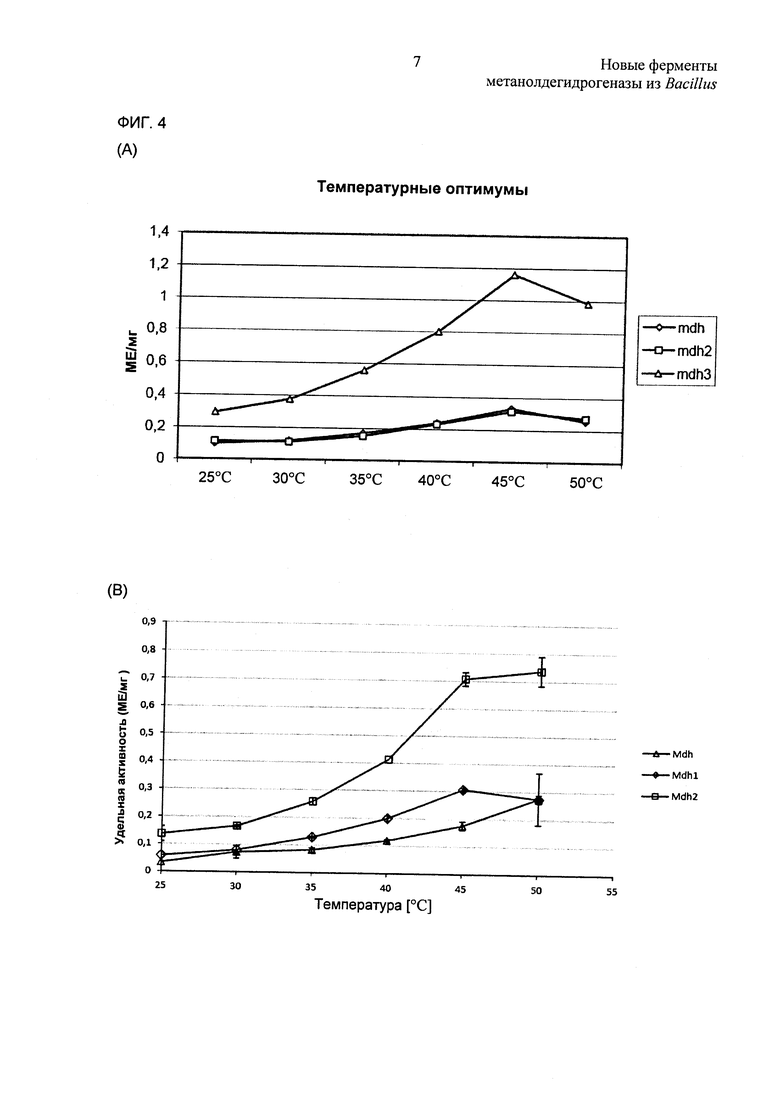
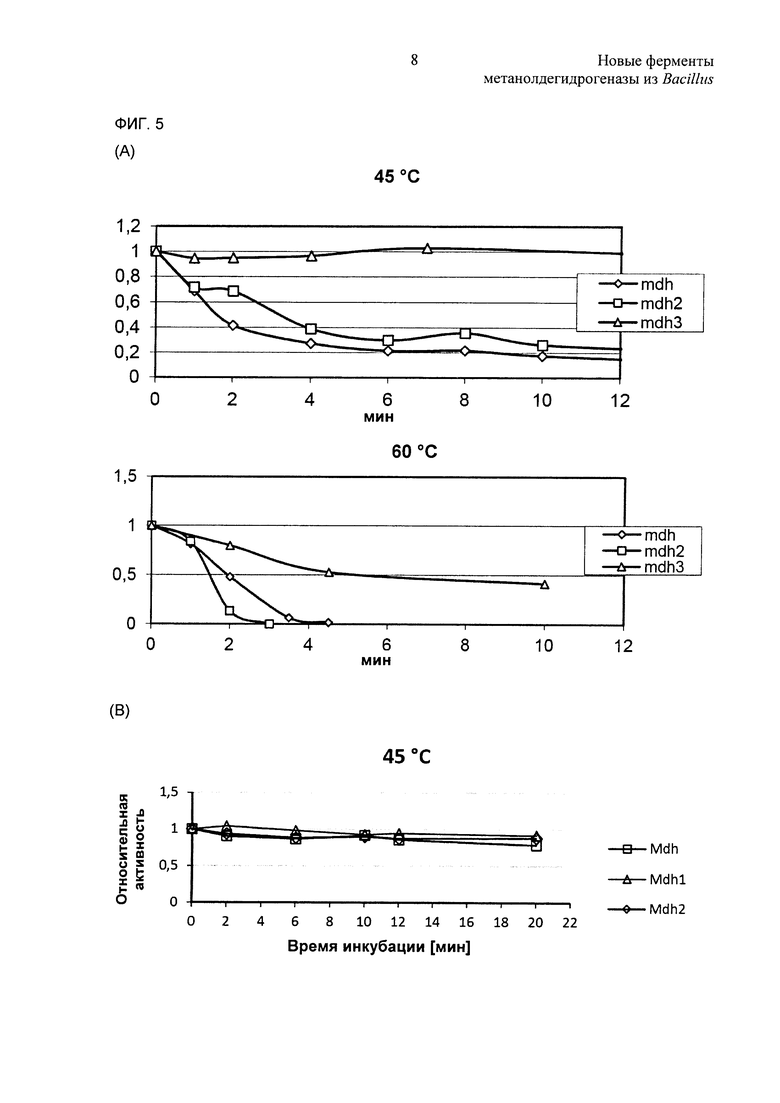
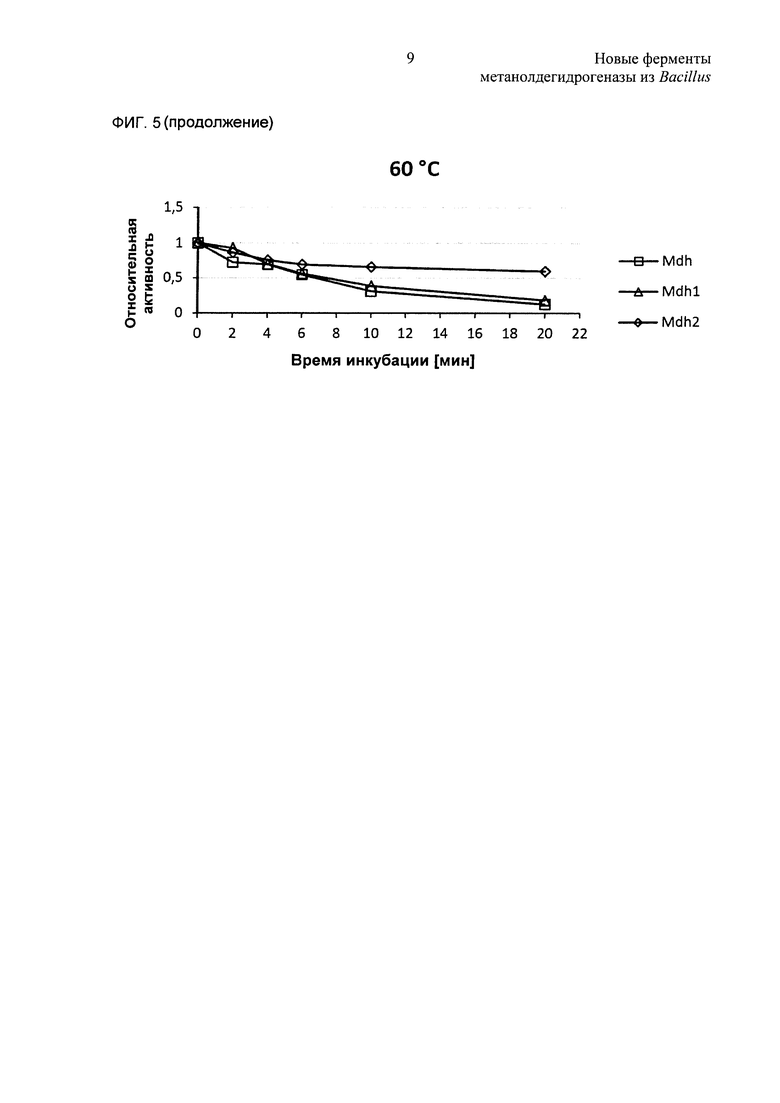
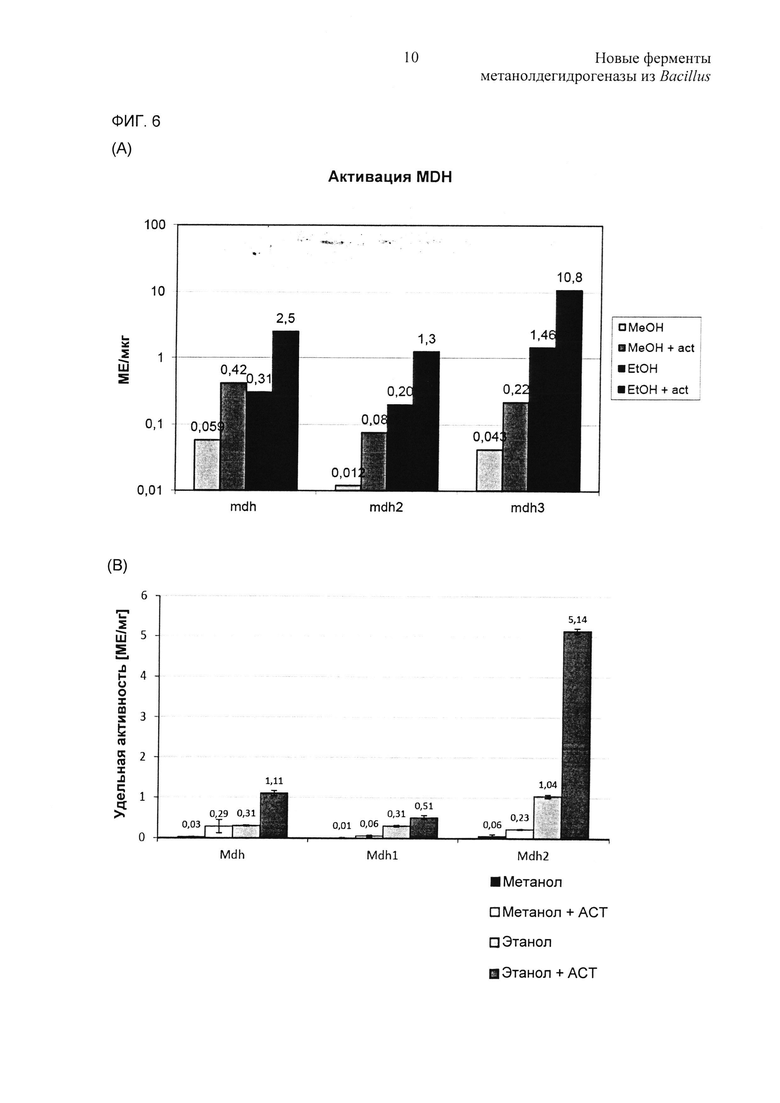
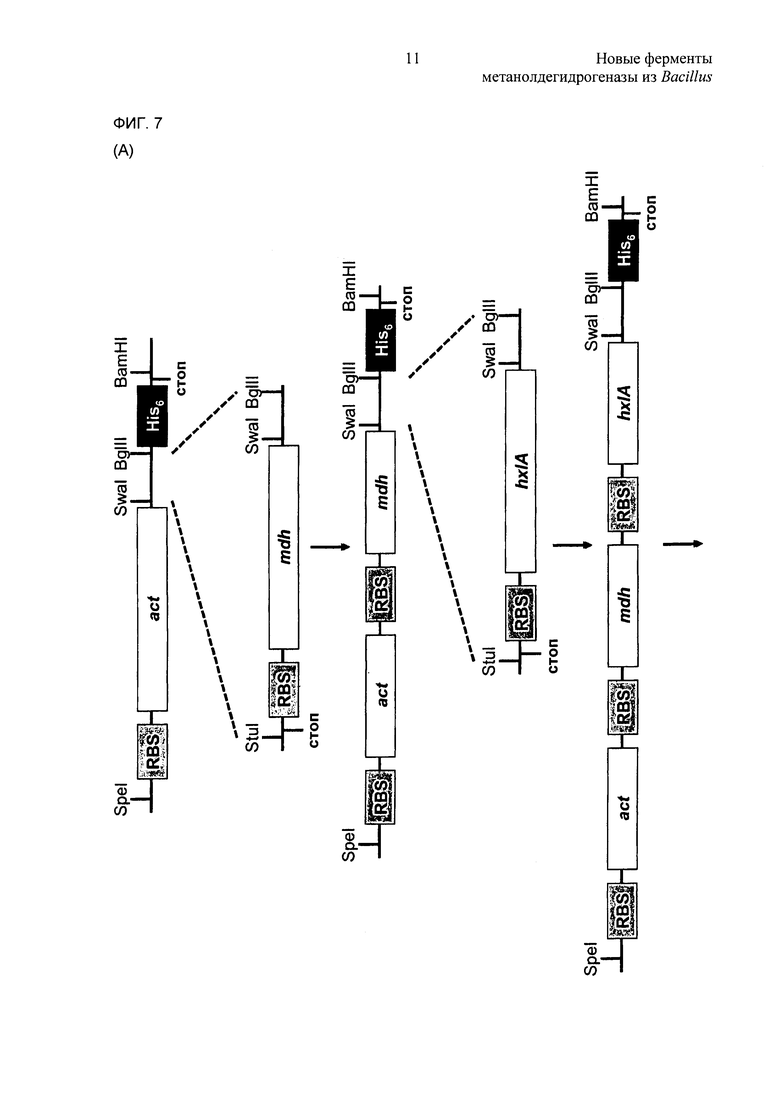
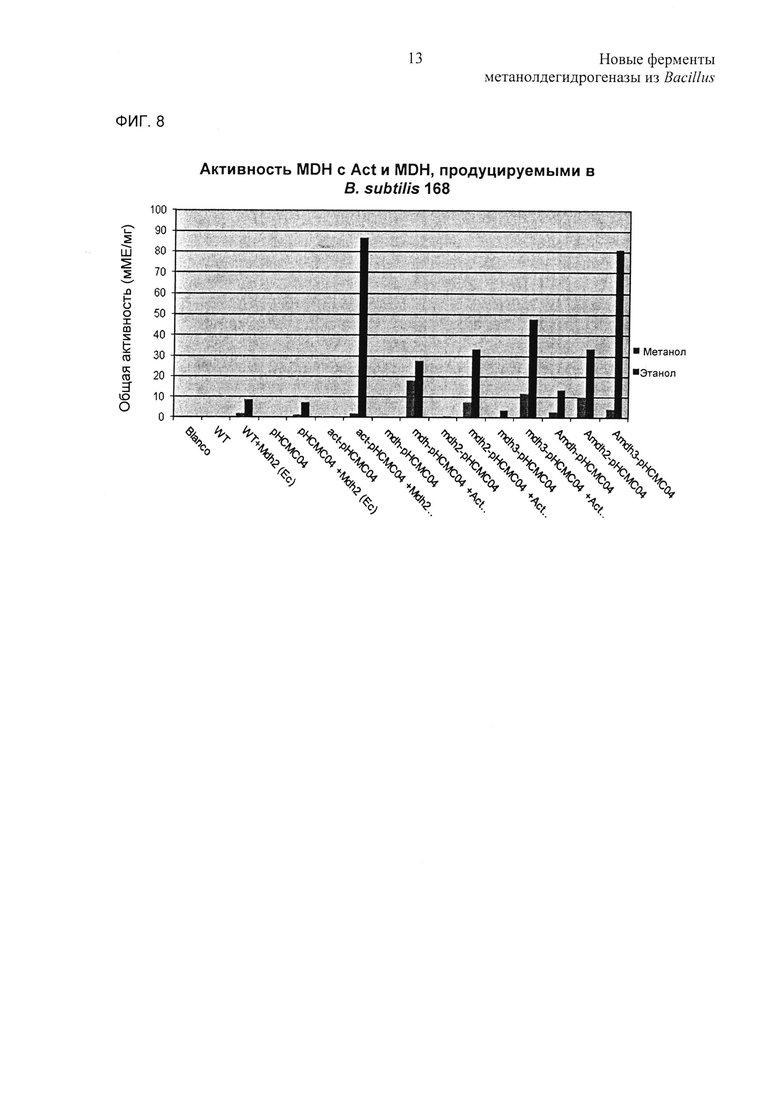
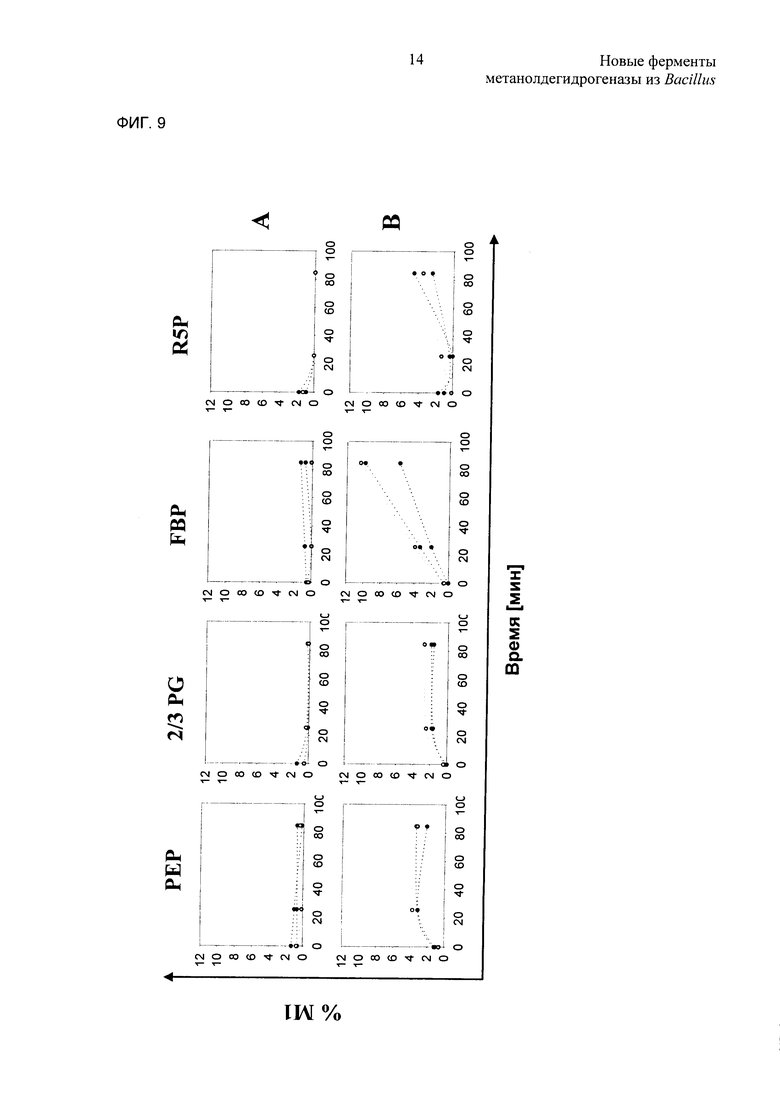
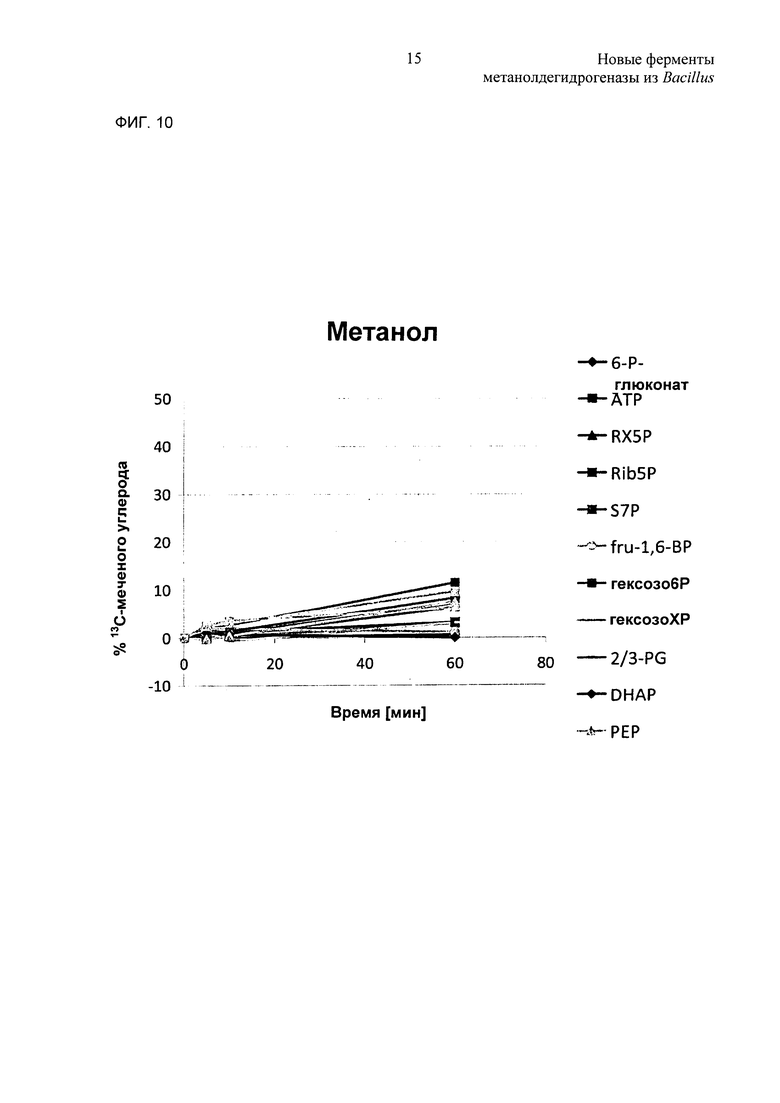
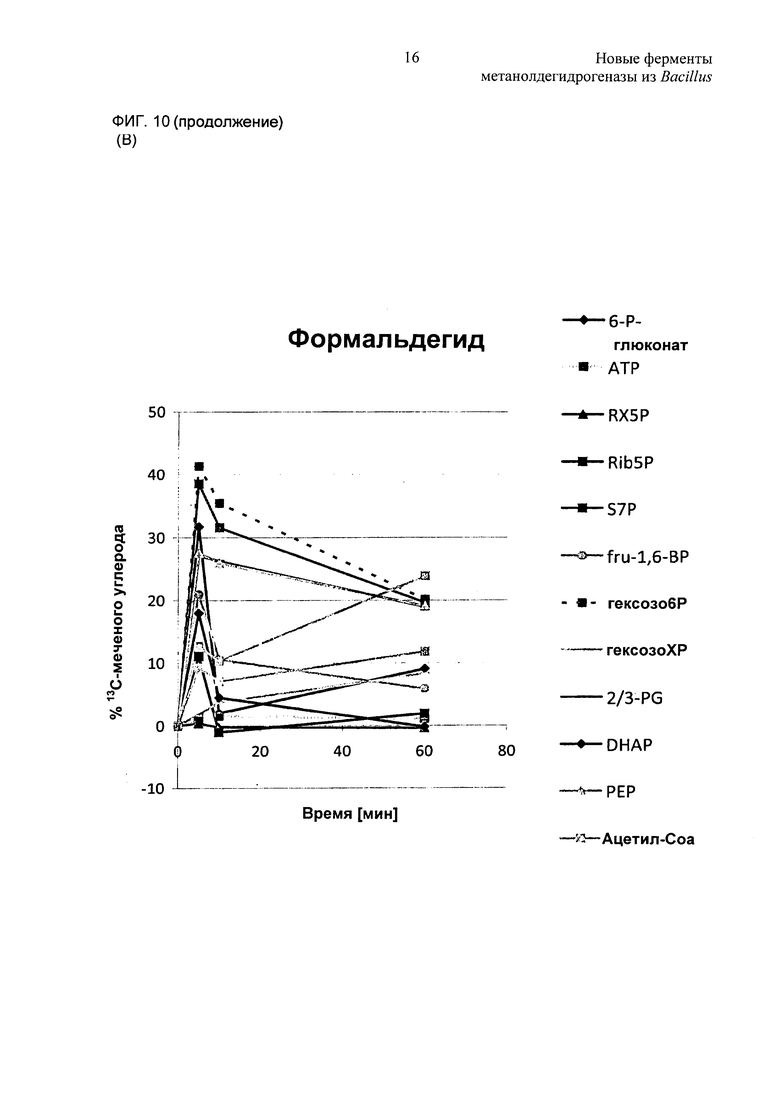
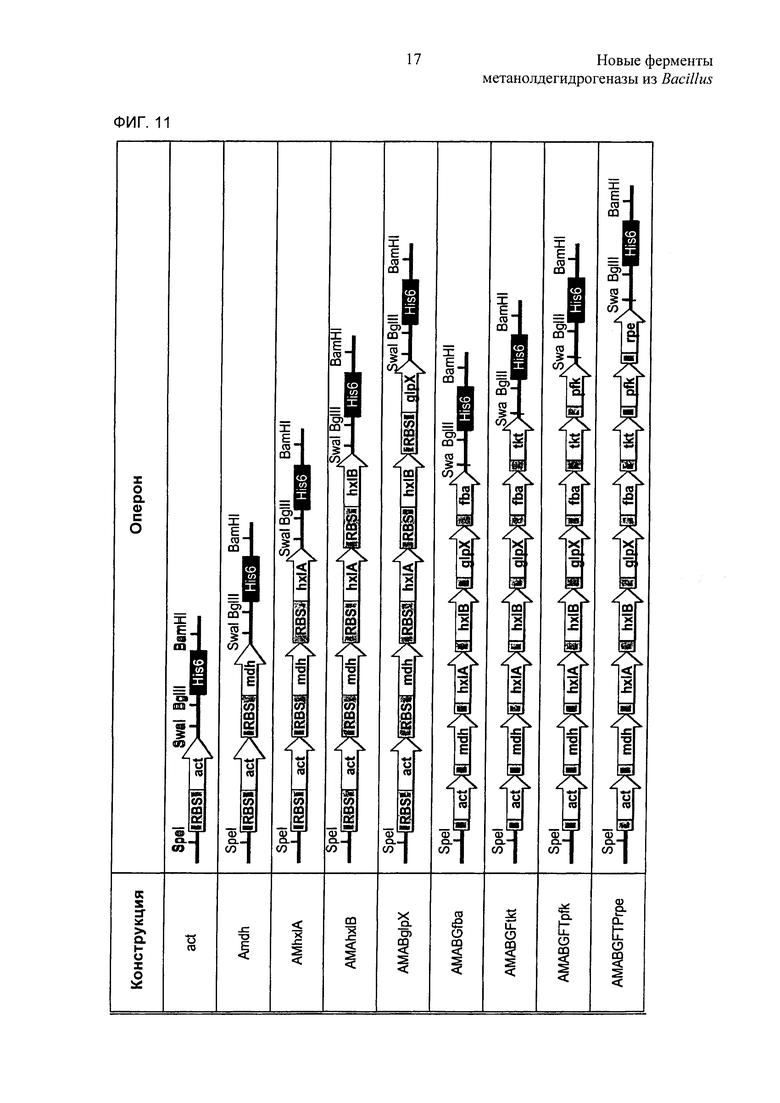
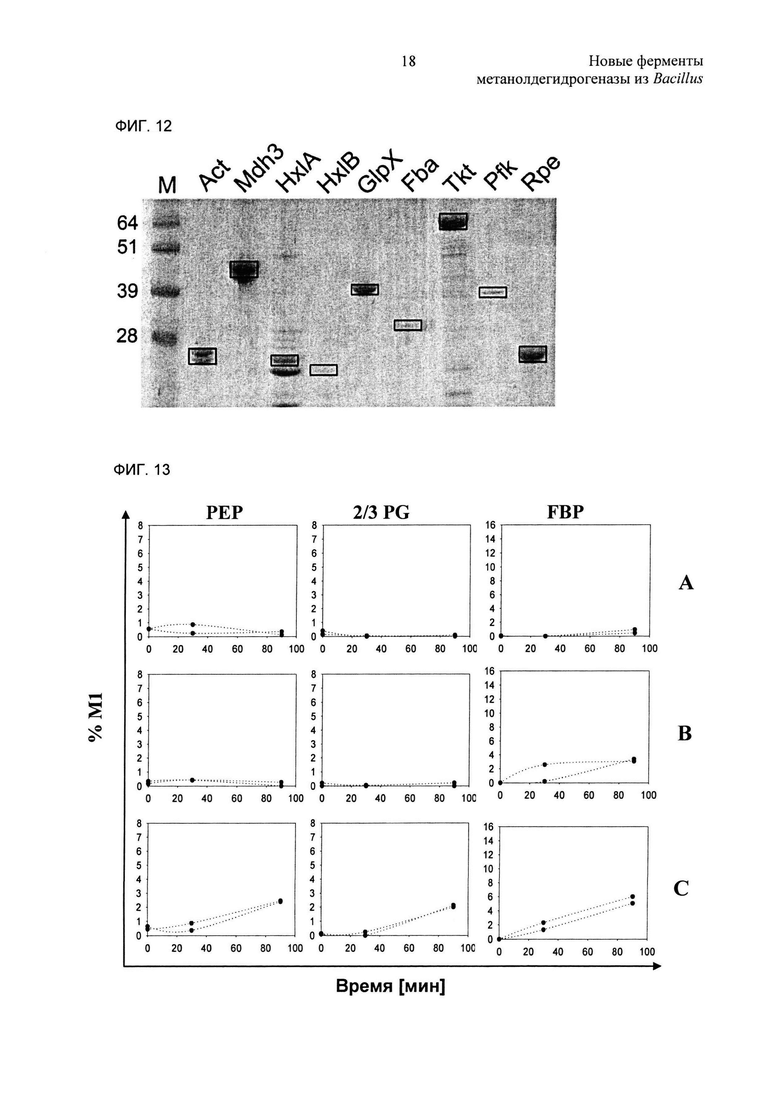
Изобретение относится к области биохимии, генной инженерии и биотехнологии, в частности к новым полипептидам из Bacillus methanolicus, обладающим активностью алкогольдегидрогеназы, в том числе метанолдегидрогеназы. Указанные полипептиды характеризуются аминокислотными последовательностями, представленными в SEQ ID NO: 2, 4, 6. Настоящее изобретение также относится к молекулам нуклеиновой кислоты, кодирующим эти полипептиды. Нуклеотидные последовательности указанных молекул нуклеиновой кислоты представлены в SEQ ID NO: 1, 3, 5. Изобретение относится к векторам экспрессии, содержащим эти молекулы нуклеиновой кислоты. Изобретение также относится к прокариотическим микроорганизмам-хозяинам для экспрессии указанных полипептидов алкогольдегидрогеназ. Настоящее изобретение позволяет получать новые полипептиды из Bacillus methanolicus с активностью алкогольдегидрогеназы, в том числе метанолдегидрогеназы. 4 н. и 5 з.п. ф-лы,13 ил., 3 табл., 18 пр.
1. Молекула нуклеиновой кислоты, которая кодирует полипептид, обладающий активностью алкогольдегидрогеназы, и имеющая нуклеотидную последовательность, выбранную из группы, состоящей из:
1) нуклеотидной последовательности, представленной в любой из SEQ ID NO: 1 (mdh2-MGA3), 3 (mdh3-MGA3) или 5 (mdh2-PB1);
2) нуклеотидной последовательности, обладающей по меньшей мере 90%-ной идентичностью последовательности, более конкретно по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99%-ной идентичностью последовательности с нуклеотидной последовательностью, представленной в любой из SEQ ID NO: 1, 3 или 5;
3) нуклеотидной последовательности, которая является вырожденной по отношению к любой нуклеотидной последовательности из SEQ ID NO: 1, 3 или 5;
4) нуклеотидной последовательности, кодирующей полипептид, аминокислотная последовательность которого представлена в любой из SEQ ID NO: 2 (Mdh2-MGA3), 4 (Mdh3-MGA3) или 6 (Mdh2-PB1); и
5) нуклеотидной последовательности, кодирующей полипептид, который имеет аминокислотную последовательность, обладающую по меньшей мере 90%-ной идентичностью последовательности, предпочтительно по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99%-ной идентичностью последовательности с аминокислотной последовательностью, представленной в любой из SEQ ID NO: 2, 4 или 6;
или молекула нуклеиновой кислоты, имеющая нуклеотидную последовательность, которая комплементарна нуклеотидной последовательности любой из (1)-(5).
2. Молекула нуклеиновой кислоты по п. 1, где указанный полипептид обладает активностью метанолдегидрогеназы
3. Полипептид, обладающий активностью алкогольдегидрогеназы, имеющий аминокислотную последовательность, выбранную из группы, состоящей из:
1) аминокислотной последовательности, представленной в любой из SEQ ID NO: 2, 4 или 6; и
2) аминокислотной последовательности, обладающей по меньшей мере 90%-ной идентичностью последовательности, предпочтительно по меньшей мере 91, 92, 93, 94, 95, 96, 97, 98 или 99%-ной идентичностью последовательности с аминокислотной последовательностью, представленной в любой из SEQ ID NO: 2, 4 или 6.
4. Полипептид по п. 3, обладающий активностью метанолдегидрогеназы.
5. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты, как определено в п. 1 или 2.
6. Прокариотический микроорганизм-хозяин для экспрессии алкогольдегидрогеназы, в который введена молекула нуклеиновой кислоты, как определено в п. 1 или 2, или вектор экспрессии, как определено в п. 5.
7. Прокариотический микроорганизм-хозяин по п. 6 для экспрессии метанолдегидрогеназы.
8. Прокариотический микроорганизм-хозяин по п. 6 или 7, представляющий собой бактерию, выбранную из рода Escherichia, Corynebacterium или Bacillus.
9. Прокариотический микроорганизм-хозяин по любому из пп. 6-8, представляющий собой Е. coli, С. glutamicum, В. subtilis или В. methanolicus.
| BRAUTASET T ET AL, Plasmid-dependent methylotrophy in thermotolerant Bacillus methanolicus, JOURNAL OF BACTERIOLOGY, 2004 | |||
| BRAUTASET T ET AL, Bacillus methanolicus: a candidate for industrial production of amino acids from methanol at 50 C, APPLIED MICROBIOLOGY AND BIOTECHNOLOGY, SPRINGER, BERLIN, DE, 2007 | |||
| КОРИНЕФОРМНАЯ БАКТЕРИЯ, ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2004 |
|
RU2333247C2 |

