Изобретение относится к способам получения L-аминокислот и используемым для этого бактериям. Более конкретно данное изобретение относится к бактериям, обладающим повышенной способностью продуцировать L-аминокислоту, и к способам получения L-аминокислот с использованием указанных бактерий.
Описание связанной области
Обычно L-аминокислоты, такие как L-глутаминовая кислота, главным образом получают ферментацией, используя так называемые коринеформные бактерии, относящиеся к роду Brevibacterium, Corynebacterium или Microbacterium ("Amino Acid Fermentation", Gakkai Shuppan Center, pp.195-215, 1986). Кроме того, для получения L-аминокислот также можно использовать микроорганизмы рода Bacillus, Streptomyces, Penicillium (патент США № 3220929), Pseudomonas, Arthrobacter, Serratia, Aerobacter, Candida (патент США № 3563857), Escherichia (выложенная заявка на выдачу патента Японии (Kokai) № 5-244970) и т.п. Кроме того, для получения L-аминокислот, таких как L-глутаминовая кислота, также можно использовать микроорганизмы, относящиеся к роду Enterobacter (EP1078989A2), Klebsiella, Erwinia или Pantoea (выложенная заявка на выдачу патента Японии № 2000-106869).
Кроме того, описаны различные способы повышения способности продуцировать L-аминокислоты путем увеличения ферментов, вовлеченных в биосинтез L-аминокислот, благодаря использованию технологии рекомбинантной ДНК. Например, описан способ получения L-глутаминовой кислоты с использованием бактерии, относящейся к роду Enterobacter или Klebsiella, в которую вводят ген цитратсинтазы (ЕР0999282А2), и способ получения L-глутаминовой кислоты с использованием бактерии, относящейся к роду Enterobacter, в которую вводят гены, кодирующие цитратсинтазу, фосфоенолпируваткарбоксилазу и глутаматдегидрогеназу (ЕР 1078989А2).
Кроме того, также известны способы усиления способности продуцировать L-аминокислоту введением генов, кодирующих гликолитические ферменты, такие как, глюкоза-6-фосфатизомераза (WO 01/02542 A1), фруктозофосфотрансфераза (WO 01/48146 A1) и енолаза (WO 01/02543 A1).
Между тем, многие грамотрицательные бактерии, включая энтеробактерии, имеют путь Энтнера-Дудорова в качестве одного из путей метаболизма глюкозы. В указанном пути принимают участие 6-осфоглюконатдегидратаза (в дальнейшем сокращенно "EDD"), которая катализирует реакцию получения 2-кето-3-дезокси-6-фосфоглюконата из 6-фосфоглюконовой кислоты, и 2-кето-3-дезокси-6-фосфоглюконатальдолаза (в дальнейшем сокращенно "EDA"), которая расщепляет 2-кето-3-дезокси-6-фосфоглюконат с получением глицеральдегид-3-фосфата и пировиноградной кислоты. Гены, кодирующие EDD и EDA, клонированы из Escherichia coli, Zymomonas mobilis и так далее, и их нуклеотидные последовательности описаны. Нуклеотидные последовательности гена, кодирующего EDD (edd), и гена, кодирующего EDA (eda), Escherichia coli зарегистрированы в GenBank с инвентарным номером L20897. Кроме того, нуклеотидная последовательность гена edaZymomonas mobilis зарегистрирована в GenBank с инвентарным номером Х58364 и нуклеотидная последовательность его гена edd зарегистрирована в GenBank с инвентарным номером М60615 М37982 в базе данных.
Однако взаимосвязь пути Энтнера-Дудорова и продуктивности L-аминокислот неизвестна.
Сущность изобретения
Целью данного изобретение является способ повышения продуктивности L-аминокислот у бактерий на основе точки зрения, отличающейся от известных способов.
Авторы данного изобретения сосредоточили свое внимание на пути Энтнера-Дудорова, который имеется у грамотрицательных бактерий. В метаболических путях от сахаридов до L-аминокислот, таких как L-глутаминовая кислота, диоксид углерода образуется в реакции получения рибулозо-5-фосфата из 6-фосфоглюконовых кислот 6-фосфоглюконатдегидрогеназой. В частности, в бактериальных штаммах, имеющих большой приток углерода в пентозофосфатный путь, также должно высвобождаться большое количество диоксида углерода в результате указанной выше реакции. Поэтому авторы предположили, что способность продуцировать L-аминокислоту, такую как L-глутаминовая кислота, можно повысить, прекратив приток в пентозофосфатный путь.
Предполагаются два способа снижения притока углерода в пентозофосфатный путь: (1) устранение или снижение активности глюкозо-6-фосфатдегидрогеназы или 6-фосфоглюконатдегидрогеназы; и (2) усиление пути Энтнера-Дудорова. Можно ожидать, что оба способа приведут к обходу пентозофосфатного пути. Однако в случае (2) предполагается, что так как распределение углерода по отношению к пентозофосфатному пути можно изменить, регулируя активности EDD и EDA, также может обеспечиваться поставка производного промежуточного вещества в пентозофосфатном пути, такого как нуклеиновая кислота. Кроме того, в результате различных исследований авторы обнаружили, что способность бактерий продуцировать L-аминокислоты можно улучшить посредством усиления пути Энтнера-Дудорова, и таким образом выполнили данное изобретение.
Итак, данное изобретение относится к следующему:
(1) Способ получения L-аминокислоты, включающий в себя культивирование микроорганизма, обладающего способностью продуцировать L-аминокислоту, в среде, чтобы продуцировать и накопить L-аминокислоту в среду, и сбор L-аминокислоты из среды, при этом микроорганизм является грам-отрицательной бактерией, имеющей путь Энтнера-Дудорова, которая была модифицирована так, чтобы активность 6-фосфоглюконатдегидратазы или активность 2-ето-3-дезокси-6-фосфоглюконатальдолазы или обе активности были усилены, а L-аминокислота выбрана из L-аминокислот, продуцируемых путем биосинтеза с использованием пировиноградной кислоты в качестве промежуточного соединения.
(2) Способ по п.(1), где бактерия является энтеробактерией.
(3) Способ по п.(2), где бактерия относится к роду Enterobacter.
(4) Способ по любому из пп.(1)-(3), где 6-осфоглюконатдегидратазная активность или 2-кето-3-дезокси-6-фосфоглюконатальдолазная активность усилены за счет увеличения количества копий гена, кодирующего 6-фосфоглюконатдегидратазу или 2-кето-3-дезокси-6-фосфоглюконатальдолазу, или модифицирования последовательности, регулирующей экспрессию гена, так, что экспрессия гена усилена в клетке бактерии.
(5) Способ по п.(1), где L-аминокислота является L-глутаминовой кислотой или L-аминокислота, образуемой путем биосинтеза с использованием L-глутаминовой кислоты в качестве промежуточного соединения или донора аминогруппы.
(6) Способ по любому из пп.(1)-(5), где L-аминокислота выбрана из L-глутаминовой кислоты, L-аргинина, L-глутамина, L-ролина, L-лейцина, L-изолейцина, L-валина и L-аланина.
(7) Способ по п.(6), где L-аминокислотой является L-глутаминовая кислота.
Согласно данному изобретению посредством увеличения активности пути Энтнера-Дудорова можно повысить способность микроорганизма, имеющего данный путь, продуцировать L-аминокислоту.
Краткое описание чертежей
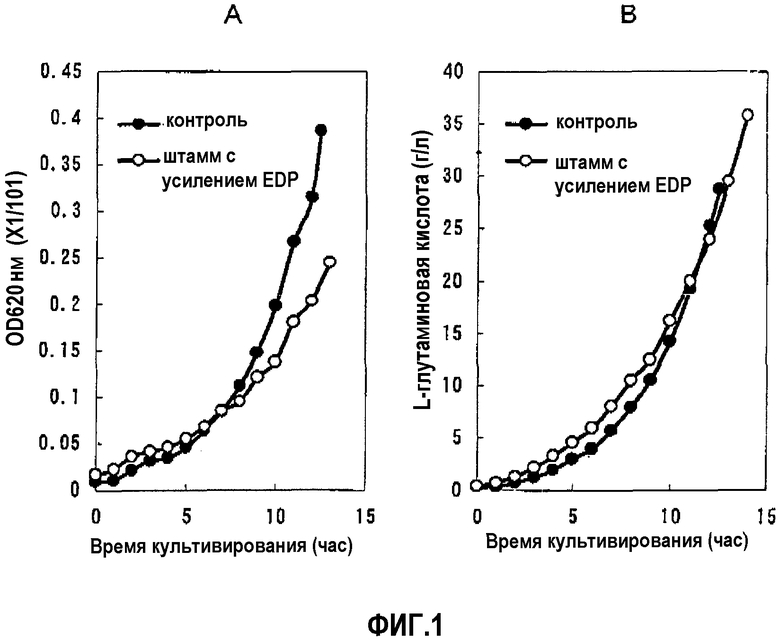
На фиг.1 показан рост штамма, в котором усилены ген edd и ген eda (А), и количество продуцируемой L-глутаминовой кислоты (В).
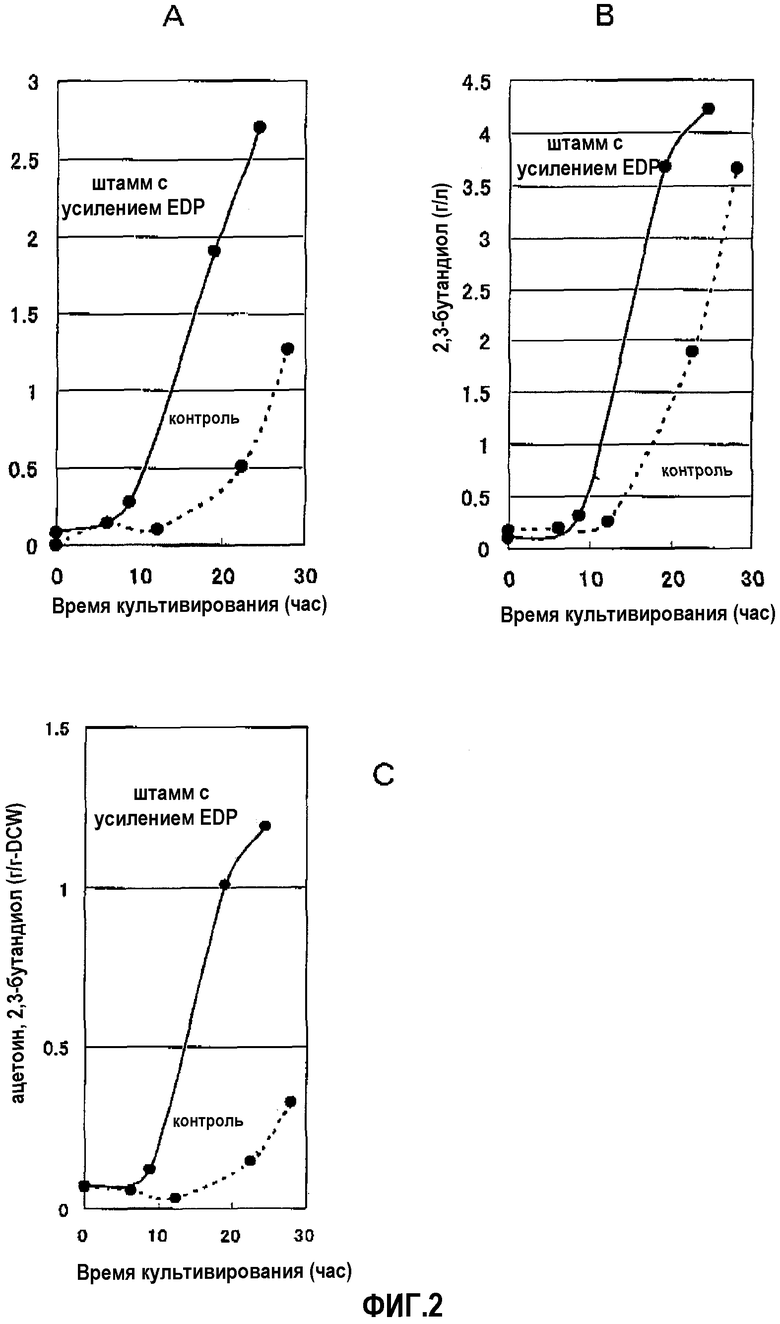
На фиг.2 показаны количества ацетоина и 2,3-бутандиола, продуцируемых штаммом, в котором усилены ген edd и ген eda: (А) количество ацетоина в среде, (В) количество 2,3-бутандиола в среде и (С) общее количество продуцируемого ацетоина и 2,3-утандиола на единицу бактериальных клеток (масса на единицу сухой массы клеток).
Подробное описание изобретения
Ниже будет дано подробное объяснение изобретения.
<1> Бактерия согласно данному изобретению
Грамотрицательная бактерия, используемая в данном изобретении, является грамотрицательной бактерией, обладающей способностью продуцировать L-аминокислоту и имеющей путь Энтнера-Дудорова.
Термин "способность продуцировать L-аминокислоту", используемый в данном изобретении, означает способность накапливать L-аминокислоту в среде в том случае, когда бактерию согласно данному изобретению культивируют в среде. Указанная способность продуцировать L-аминокислоту может быть свойством дикого штамма грам-отрицательной бактерии или свойством, приобретаемым или усиливаемым при селекции. L-аминокислоты, по отношению к которым можно применять данное изобретение, являются L-аминокислотами, получаемые путем биосинтеза с использованием пировиноградной кислоты в качестве промежуточного соединения. Конкретные примеры таких L-аминокислот включают L-глутаминовую кислоту, L-аргинин, L-глутамин, L-пролин, L-лейцин, L-изолейцин, L-валин, L-аланин и т.д.
Как показано в описанных далее примерах в бактерии, имеющей путь Энтнера-Дудорова, усиленный благодаря увеличению активностей EDD и EDA, наблюдается повышенная продукция ацетоина и 2,3-бутандиола. Так как 2,3-бутандиол образуется из ацетоина, а ацетоин образуется из пировиноградной кислоты, то увеличение продукции ацетоина и 2,3-бутандиола свидетельствует об увеличении количества используемой пировиноградной кислоты. Таким образом, предполагается, что бактерия, имеющая усиленный путь Энтнера-Дудорова, обладает повышенной способностью продуцировать L-аминокислоту, образуемую путем биосинтеза с использованием пировиноградной кислоты в качестве промежуточного соединения.
Конкретные примеры грамотрицательных бактерий, имеющих путь Энтнера-Дудорова, включают бактерии, относящиеся к родам Enterobacter, Klebsiella, Serratia, Erwinia или Pantoea,Escherichia, Pseudomonas, Arthrobacter и Aerobacter и так далее. Имеет ли бактерия путь Энтнера-Дудорова или не имеет, можно, например, определить, смешивая суспензию разрушенных клеток с глицеральдегид-3-фосфатдегидрогеназой, 6-фосфоглюконовой кислотой и ацетилпиридинадениндинуклеотидом и детектируя глицеральдегид-3-фосфат, образуемый из 6-фосфоглюконовой кислоты в качестве субстрата, измеряя увеличение оптической плотности при 365 нм. Бактерия, для которой подтверждают образование глицеральдегид-3-фосфата, имеет путь Энтнера-Дудорова.
Бактерии, используемые для данного изобретения, можно соответствующим образом выбрать в зависимости от типа L-аминокислоты-мишени. Примеры бактерий, подходящих для получения L-глутаминовой кислоты, приведены ниже. Однако объем данного изобретения не ограничен указанными примерами.
Конкретные примеры бактерий Enterobacter включают следующие бактерии:
Enterobacter agglomerans
Enterobacter aerogenes
Enterobacter amnigenus
Enterobacter asburiae
Enterobacter cloacae
Enterobacter dissolvens
Enterobacter gergoviae
Enterobacter hormaechei
Enterobacter intermedius
Enterobacter nimipressuralis
Enterobacter sakazakii
Enterobacter taylorae
Более предпочтительными являются следующие бактериальные штаммы:
Enterobacter agglomerans ATCC 12287
Enterobacter agglomerans AJ13355
Enterobacter agglomerans AJ13356
Enterobacter agglomerans AJ13601
Enterobacter agglomerans AJ13355 и AJ13556 депонировали в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology (в настоящее время независимая административная корпорация, International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology, Адрес: Chuo Dai-6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Japan, почтовый индекс: 305-5466) 19 февраля 1998 г., штаммы получили инвентарные номера FERM P-16644 и FERM P-16645, соответственно. Затем депозиции перевели в международные депозиции по условиям Будапештского договора 11 января 1999 г., и им присвоили инвентарные номера FERM BP-6614 и FERM BP-6615. Enterobacter agglomerans AJ13601 депонировали в National Institute of Bioscience and Human-Technology, Agency of Bioscience and Human Technology 18 августа 1999 г., и ему присвоили инвентарный номер FERM P-17516. Затем депозицию перевели в международную депозицию по условиям Будапештского договора 6 июля 2000 г., и ей присвоили инвентарный номер FERM BP-7207. Enterobacter agglomerans ATCC 12287 можно получить из ATCC (Американской коллекции типов культур, 10801 University Boulevard, Manassas, VA 20110-2209, U.S.A.).
Примеры бактерий, относящихся к роду Klebsiella, включают следующие бактерии:
Klebsiella planticola
Klebsiella terrigena
Более предпочтительной является Klebsiella planticola AJ13399. Klebsiella planticola AJ13399 депонировали в National Institute of Bioscience and Human-Technology, Agency of Bioscience and Human Technology (в настоящее время независимая административная корпорация, International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology) 19 февраля 1998 г., и ему присвоили инвентарный номер FERM P-16646. Затем депозицию перевели в международную депозицию по условиям Будапештского договора 11 января 1999 г., и ей присвоили инвентарный номер FERM BP-6616.
Klebsiella planticola AJ13399 является штаммом, выделенным из почвы в Sapporo-shi, Hokkaido.
Примеры микроорганизмов, относящихся к роду Serratia, используемые для данного изобретения, включают следующие бактерии:
Serratia liquefacience
Serratia entomophila
Serratia ficaria
Serratia fonticola
Serratia grimesii
Serratia proteamaculans
Serratia odorifera
Serratia plymuthica
Serratia rubidaea
Более предпочтительными являются следующие бактериальные штаммы:
Serratia liquefacience ATCC 14460
Serratia liquefacience ATCC 14460 можно получить из ATCC.
Примеры микроорганизмов, относящихся к роду Erwinia, используемые в данном изобретении, включают следующие бактерии:
Erwinia herbicola (недавно классифицирована как Pantoea agglomerans)
Erwinia ananas
Erwinia cacticida
Erwinia chrysanthemi
Erwinia mallotivora
Erwinia persicinus
Erwinia psidii
Erwinia quercina
Erwinia rhapontici
Erwinia rubrifaciens
Erwinia salicis
Erwinia uredovora
Более предпочтительна Erwinia herbicola IAM1595 (Pantoeaagglomerans AJ2666). Erwinia herbicola IAM1595 можно получить из Institute of Molecular and Cellular Bioscience, the University of Tokyo.
Erwinia herbicola не упоминается Bergey в Manual of Determinative Bacteriology, 9th Ed., и микроорганизм, который был классифицирован как Erwinia herbicola, классифицируют как Pantoea agglomerans. Таким образом, микроорганизмы, относящиеся к роду Erwinia, и микроорганизмы, относящиеся к роду Pantoea, являются близко родственными по отношению друг к другу. Следовательно, микроорганизмы, относящиеся к роду Pantoea, можно использовать подобно микроорганизмам, относящимся к роду Erwinia. В качестве таких микроорганизмов, относящихся к роду Pantoea, можно назвать Pantoea agglomerans, Pantoea dispersa и Pantoea ananas. Erwinia herbicola IAM1595 назвали Pantoea agglomerans AJ2666 и депонировали в National Institute of Bioscience and Human-Technology, Agency of Bioscience and Human Technology (в настоящее время International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology) в качестве международного депонирования по условиям Будапештского договора 25 февраля 1999 г. и ему присвоили инвентарный номер FERM BP-6660.
Примеры микроорганизмов, относящихся к роду Escherichia, используемых для данного изобретения, включают Escherichia coli.
Более предпочтительна Escherichia coli, обладающая устойчивостью к валину, и конкретными примерами являются следующие штаммы:
Escherichia coli K-12 (ATCC 10798)
Escherichia coli W (ATCC 9637)
Escherichia coli K-12 (ATCC 10798) и Escherichia coli W (ATCC 9637) можно получить из ATCC.
Грамотрицательной бактерией согласно данному изобретению является грамотрицательная бактерия, которая обладает способностью продуцировать L-аминокислоту и имеет вышеуказанный путь Энтнера-Дудорова и которая была модифицирована так, чтобы активность EDD или EDA или обе активности были усилены. Предпочтительно бактерия согласно данному изобретению представляет собой грамотрицательную бактерию, которая была модифицирована так, чтобы обе активности EDD и EDA были усилены.
Выражение "модифицирована так, чтобы активность EDD или EDA усилена" означает, что активность EDD или EDA на клетку увеличена по сравнению с активностью бактерии дикого типа. Например, можно указать бактерии, в которых увеличено количество молекул EDD или EDA на клетку, бактерии, у которых повышена удельная активность EDD или EDA на молекулу EDD или EDA. Кроме того, бактерия дикого типа, с которой проводят сравнение, является бактерий, которую не подвергали какой-либо обработке для усиления активности EDD или EDA.
Усиление активности EDD и/или EDA в бактерии осуществляют посредством увеличения числа копий гена, кодирующего EDD и/или EDA. Например, можно получить рекомбинантную ДНК лигированием фрагмента гена, кодирующего EDD и/или EDA, с вектором, функционирующем в бактерии-мишени, предпочтительно вектором мультикопийного типа, и можно ввести в бактерию, чтобы ее трансформировать. В случае усиления обеих активностей EDD и EDA, фрагмент гена, кодирующий EDD, и фрагмент гена, кодирующий EDA, можно включить отдельно в разные векторы, но предпочтительно их можно включить в один и тот же вектор. Рекомбинантную ДНК можно ввести в бактерию, обладающую способностью продуцировать L-аминокислоту, альтернативно рекомбинантную ДНК можно ввести в бактерию дикого типа, чтобы получить трансформированный штамм, и затем трансформированному штамму можно придать способность продуцировать L-аминокислоту.
В качестве гена, кодирующего EDD, и гена, кодирующего EDA, можно использовать любой из генов, полученных из грамотрицательных бактерий, имеющих путь Энтнера-Дудорова. В частности, можно указать гены, полученные из бактерий Escherichia. Сообщалось, что ген, кодирующий EDD, (edd) и ген, кодирующий EDA, (eda), полученные из Escherichia coli, образуют оперон (J. Bacteriol., 174 (14): 4638-46, July 1992). В дальнейшем ген, кодирующий EDD, называют edd, а ген, кодирующий EDA, называют eda. Кроме того, также сообщали о генах бактерий рода Zymomonas, и ген edd, и ген eda можно получить посредством ПЦР (полимеразной цепной реакции, см. White, T.J. et al., Trends Genet. 5, 185 (1989)), используя праймеры, полученные на основе последовательностей указанных генов, или посредством гибридизации с использованием зонда, полученного на основе вышеуказанных последовательностей генов. Например, фрагмент оперона, содержащего ген edd и ген edaEscherichia coli, можно получить посредством ПЦР, используя праймеры edd-F (SEQ ID NO:1) и eda-R (SEQ ID NO:2), описанные далее. Подобным образом можно получить ген edd и ген eda других микроорганизмов. Примерами условий гибридизации являются условия, при которых промывку выполняют при концентрации соли, соответствующей 1ЧSSC, 0,1% SDS, предпочтительно 0,1ЧSSC, 0,1% SDS при 60°С.
Кроме того, ген edd и ген eda, используемые для данного изобретения, не ограничены генами дикого типа и могут быть мутантами или искусственно модифицированными генами, кодирующими генные продукты, включающими замену, делецию, инсерцию, присоединение и тому подобное одной или нескольких аминокислот в одном или нескольких участках, при условии, что функции кодируемых EDD и EDA не нарушаются. Хотя количество "несколько" аминокислот, упоминаемое в данном описании, различно в зависимости от положения и типа аминокислотных остатков в трехмерной структуре белка, оно более конкретно может составлять от 2 до 60, предпочтительно от 2 до 40, более предпочтительно от 2 до 20. Кроме того, в качестве ДНК, кодирующей белок в значительной степени идентичный вышеуказанным EDD и/или EDA, можно указать ДНК, которая гибридизуется с нуклеотидными последовательностями известного гена edd или гена eda (например, в GenBank с инвентарными номером L20897, X58364, M60615, M37982) или зондом, который можно получить из указанных нуклеотидных последовательностей, в жестких условиях и кодирует белок, обладающий активностью, сходной с активностью EDD или EDA. "Жесткие условия" в данном описании относятся к условиям, при которых образуются так называемые специфичные гибриды, а неспецифичные гибриды не образуются. Такие условия трудно четко выразить с использованием числовых значений. Однако жесткие условия, например, включают условия, при которых ДНК, имеющие высокую степень гомологии, например, ДНК, имеющие гомологию 50% или выше, гибридизуются друг с другом, а ДНК, имеющие гомологию ниже, чем указанная выше, не гибридизуются друг с другом. Альтернативно примером жестких условий являются условия, при которых ДНК гибридизуются друг с другом при концентрации соли, соответствующей обычным условиям промывки при Саузерн-гибридизации, т.е. 1ЧSSC, 0,1% SDS, предпочтительно 0,1ЧSSC, 0,1% SDS при 60°С.
Хромосомную ДНК можно получить из бактерии, являющейся донором ДНК, например, способом Saito и Miura (см. H. Saito and K. Miura, Biochem. Biophys. Acta, 72, 619 (1963), Text for Bioengineering experiments, Edited by the Society for Bioscience and Bioengineering, Japan, pp.97-98, Baifukan, 1992) или тому подобным.
В том случае, если рекомбинантную ДНК получают лигированием гена edd и/или гена eda, амплифицированных в ПЦР, с векторной ДНК, автономно реплицирующейся в клетке Escherichia coli или тому подобной, и вводят в Escherichia coli, последующие операции облегчаются. Примеры векторов, автономно реплицирующихся в клетке Escherichia coli, включают pMW219, pSTV28, pUC19, pUC18, pHSG299, pHSG399, pHSG398, RSF1010, pBR322, pACYC184 и т.д.
Для того чтобы ввести рекомбинантную ДНК, полученную как описано выше, в грамотрицательную бактерию, можно использовать способы трансформации, которые сообщались до настоящего времени. Например, можно указать способ D.A. Morrison (Methods in Enzymology, 68, 326 (1979)), способ обработки реципиентных клеток хлоридом кальция, чтобы увеличить проницаемость для ДНК (Mandel, M. and Higa, A., J. Mol. Biol., 53, 159 (1970)), способ электропорации (Miller J.H., "A Short Course in bacterial Genetics; Handbook", Cold Spring Harbor Laboratory Press, U.S.A., p.279, 1992) и т.д.
Количество копий гена edd и/или гена eda также можно увеличить, обеспечивая существование множественных копий указанных генов в хромосомной ДНК бактерии. Чтобы ввести множественные копии гена edd и/или гена eda в хромосомную ДНК бактерии, выполняют гомологичную рекомбинацию, используя в качестве мишени последовательность, множественные копии которой существуют в хромосомной ДНК. В качестве последовательностей, множественные копии которых существуют в хромосомной ДНК, можно использовать повторяющуюся ДНК или инвертированный повтор, существующий на конце транспозируемого элемента. Кроме того, как описано в выложенной заявке на выдачу патента Японии № 2-109985, ген edd и/или ген eda также можно ввести в состав транспозона и обеспечить возможность его переноса, чтобы ввести множественные копии генов в хромосомную ДНК.
Усиления активностей EDD и/или EDA также можно добиться, кроме способа, основанного на вышеуказанной амплификации генов, путем замены последовательности, регулирующей экспрессию, такой как промотор гена edd и/или гена eda в хромосомной ДНК или плазмиде более сильным промотором. Например, в качестве сильных промоторов известны промотор lac, промотор trp, промотор trc и так далее. Кроме того, как описано в международной заявке на выдачу патента WO00/18935, промотор можно модифицировать путем введения замены нескольких нуклеотидов в область промотора гена edd и/или гена eda так, чтобы промотор стал более сильным. Замена или модификация указанных промоторов усиливает экспрессию гена edd и/или гена eda, и таким образом активности EDD и/или EDA повышаются. Модификацию указанных регулирующих экспрессию последовательностей можно комбинировать с увеличением числа копий гена edd и/или гена eda.
Усиление активностей EDD и EDA можно подтвердить, смешивая суспензию разрушенных клеток с глицеральдегид-3-фосфатдегидрогеназой и 6-фосфоглюконовой кислотой и измеряя глицеральдегид-3-фосфат, образуемый из 6-фосфоглюконовой кислоты в качестве субстрата. В указанной реакции активность EDD можно измерить, оценивая количество 6-фосфоглюконовой кислоты, оставшейся после реакции с использованием 6-осфоглюконатдегидрогеназы, или оценивая количество пировиноградной кислоты, образованной в присутствии избыточного количества 2-кето-3-дезокси-6-фосфоглюконатальдолазы, используя лактатдегидрогеназу. Количество 6-фосфоглюконовой кислоты или пировиноградной кислоты можно определить по увеличению NADH в дегидрогеназной реакции. Кроме того, активность EDA также можно измерить, детектируя пировиноградную кислоту, образуемую из 2-ето-3-дезокси-6-фосфоглюконата в качестве субстрата, с использованием лактатдегидрогеназы.
В грамотрицательной бактерии согласно данному изобретению можно усилить активность фермента, катализирующего биосинтез L-минокислоты, отличного от EDD и EDA, при условии, что эффект усиления активностей EDD и EDA не снижается.
Например, в том случае, когда L-аминокислотой-мишенью являются L-глутаминовая кислота, примеры такого фермента включают глутаматдегидрогеназу (также называемую в дальнейшем "GDH"), глутаминсинтетазу, глутаматсинтазу, изоцитратдегидрогеназу, аконитатгидратазу, цитратсинтазу (также называемую в дальнейшем "CS"), фосфоенолпируваткарбоксилазу (также называемую в дальнейшем "PEPC"), фосфоенолпируватсинтазу, пируватдегидрогеназу, пируваткиназу, пируваткарбоксилазу, енолазу, фосфоглицеромутазу, фосфоглицераткиназу, глицеральдегид-3-фосфатдегидрогеназу, триозофосфатизомеразу, фруктозобифосфатальдолазу, фосфофруктокиназу, глюкозофосфатизомеразу и т.д. В том случае, когда бактерией, используемой для получения L-глутаминовой кислоты, является бактерия Enterobacter, среди вышеуказанных ферментов предпочтительным является любой из трех видов ферментов CS, PEPC и GDH. Более того, предпочтительно, когда усилены активности всех трех видов ферментов CS, PEPC и GDH. В частности, предпочтительна CS Brevibacterium lactofermentum, поскольку указанный фермент не подвергается ингибированию a-кетоглутаровой кислотой, L-глутаминовой кислотой и NADH.
В качестве организмов, которые могут служить источниками гена, кодирующего CS (gltA), гена, кодирующего PEPC (ppc), и гена, кодирующего GDH (gdhA), можно использовать любые организмы, при условии, что они обладают активностями CS, PEPC и GDH. В частности, предпочтительны бактерии, которые являются прокариотами, например, бактерии, относящиеся к роду Enterobacter, Klebsiella, Erwinia, Pantoea, Serratia, Escherichia, Corynebacterium, Brevibacterium или Bacillus. Их конкретные примеры включают Escherichia coli, Brevibacterium lactofermentum и так далее. Ген gltA, ген ppc и ген gdhA можно получить из хромосомной ДНК вышеуказанных микроорганизмов.
Ген gltA, ген ppc и ген gdhA можно получить, используя мутант, дефицитный по активности CS, PEPC или GDH, и выделяя фрагмент ДНК, который комплементирует их ауксотрофность, из хромосомной ДНК вышеуказанных микроорганизмов. Кроме того, так как нуклеотидные последовательности указанных генов бактерий Escherichia и указанных генов бактерий Corynebacterium уже выяснены (Biochemistry, 22, pp.5243-5249, (1983); J. Biochem., 95 pp.909-916, (1984); Gene, 27, pp.193-199, (1984); Microbiology, 140, pp.1817-1828, (1994); Mol. Gen. Genet., 218, pp.330-339, (1989); Molecular Microbiology, 6, pp.317-326, (1992)), эти гены также можно получить, синтезируя праймеры на основании соответствующих нуклеотидных последовательностей и выполняя ПЦР с использованием хромосомной ДНК в качестве матрицы. Введение указанных генов в грамотрицательную бактерию, такую как бактерии Enterobacter, подробно писано в EP 0670370 A2, патенте США № 6197559, EP 0999282 A2 и EP 1078989 A2.
Активности CS, PEPC и GDH, а также другие указанные выше ферментативные активности можно усилить таким же образом, как указанное выше усиление активностей EDD и EDA.
Кроме того, в бактерии согласно данному изобретению также можно снизить или элиминировать активность фермента, катализирующего реакцию получения другого соединения в результате ответвления от пути биосинтеза L-аминокислоты-мишени, при условии, что эффект усиления активностей EDD и/или EDA не снижается. Например, в том случае, когда L-аминокислотой-мишенью является L-глутаминовая кислота, примеры такого фермента включают a-кетоглутаратдегидрогеназу (далее называемую в дальнейшем "aKGDH"), изоцитратлиазу, фосфатацетилтрансферазу, ацетаткиназу, синтазу ацетогидроксикислоты, ацетолактатсинтазу, формиатацетилтрансферазу, лактатдегидрогеназу, L-лутаматдекарбоксилазу, 1-пирролиндегидрогеназу и т.д.
Для того чтобы снизить или исключить активности вышеуказанных ферментов можно использовать способ обработки микроорганизма ультрафиолетовым излучением или мутагенным агентом, используемым при обычной мутагенной обработке, таким как N-метил-N'-нитро-N-нитрозогуанидин (NTG) или азотистая кислота, и отбора мутантного штамма, в котором снижена активность фермента-мишени, способ нарушения гена с использованием замены в гене на основе гомологичной рекомбинации, и тому подобные. Нарушение гена, кодирующего aKGDH, описано в патенте США № 5977331.
Если при конструировании бактерии согласно данному изобретению вводят гены, отличные от гена edd и гена eda, предпочтительно использовать меньше видов векторов. То есть, обычно вектор имеет маркерный ген и необходимо, чтобы в среду был добавлен агент, соответствующий маркерному гену, или ему подобный. Поэтому, если используется много видов векторов, то в среду надо добавлять большое количество агентов. Это может привести к плохому росту бактерий. Поэтому обычно предпочтительно использовать меньше видов векторов. Предпочтительно использовать два или меньше видов, более предпочтителен один вид вектора.
Кроме того, в том случае, когда используют два или больше видов векторов, каждый из которых имеет разное количество копий, предпочтительно определить распределение генов между вектором с большим количеством копий и вектором с малым количеством копий в зависимости от видов генов, которые необходимо ввести.
Для осуществления процесса выделения гена, введения гена в бактерию-хозяина, нарушения гена и т.д. можно использовать обычные способы, хорошо известные специалистам в данной области, а именно способы получения хромосомной ДНК, конструирования библиотеки хромосомной ДНК, гибридизации, ПЦР, получения плазмидной ДНК, расщепления и лигирования ДНК, трансформации, конструирования олигонуклеотидов, используемых в качестве праймеров, и т.д. Указанные способы описаны в Sambrook, J. Fritsh, E.F. and Maniatis, T. "Molecular Cloning. A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press, (1989) и т.д.
<2> Получение L-аминокислоты с использованием бактерии согласно данному изобретению
L-аминокислоту можно получить, культивируя бактерию согласно данному изобретению, полученную как описано выше, в среде, чтобы продуцировать и накопить L-аминокислоту в среде, и собирая L-аминокислоту из среды.
Чтобы получить L-аминокислоту, используя бактерию согласно данному изобретению, можно использовать обычные среды, содержащие источник углерода, источник азота, неорганические соли и органические питательные вещества в следовых количествах, такие как аминокислоты и витамины, которые необходимы в определенной степени. Можно использовать либо синтетическую, либо естественную среду. В среде можно использовать любые виды источника углерода и источника азота при условии, что они могут утилизироваться бактериальными штаммами, которые необходимо культивировать.
В качестве источника углерода используют сахариды, такие как глюкоза, глицерин, фруктоза, сахароза, мальтоза, манноза, галактоза, гидролизат крахмала и меласса. Также используют органические кислоты, такие как уксусная кислота и лимонная кислота, и спирты, такие как этанол, в отдельности или в комбинации с другими источниками углерода. Из них предпочтительны глюкоза и сахароза.
В качестве источника азота можно использовать аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония и ацетат аммония, нитраты и т.д.
В качестве органических питательных веществ, используемых в небольших количествах, можно использовать аминокислоты, витамины, жирные кислоты и нуклеиновые кислоты, а также пептон, казаминовую кислоту, дрожжевой экстракт и продукт разложения соевого белка, содержащий указанные вещества, и так далее. В том случае, когда используют ауксотрофный мутантный штамм, для роста которого требуются аминокислоты или тому подобные вещества, предпочтительно добавлять такое требуемое питательное вещество.
В качестве неорганических солей можно использовать фосфаты, соли магния, соли кальция, соли железа, соли марганца и т.д.
Что касается способа культивирования, то хотя это зависит от типа используемой бактерии, обычно выполняют культивирование в условиях аэрации, контролируя при этом температуру ферментации от 20 до 52°С и значение рН от 5 до 9. Если во время культивирования значение рН снижается, добавляют карбонат кальция, или культуру нейтрализуют щелочью, такой как газообразный аммиак. При культивировании таким образом в течение примерно от 10 до 120 часов в культуральном бульоне накапливается заметное количество L-глутамина.
В качестве способа для сбора L-аминокислот из культурального бульона после завершения культивирования можно использовать известные способы сбора, например, способы с использованием ионообменных смол, осаждения и т.д.
ПРИМЕРЫ
Далее будет дано более конкретное объяснение данного изобретения со ссылкой на следующие примеры.
<1> Клонирование генов ферментов, участвующих в пути Энтнера-Дудорова
Ген edd и ген eda, которые кодируют ферменты EDD и EDA, соответственно, вовлеченные в путь Энтнера-Дудорова, клонировали из Escherichia coli, Zymomonas mobilis и так далее. Enterobacter agglomerans таксономически относится к группе энтеробактерий и считается близко родственной Escherichia coli. Кроме того, известно, что гены Escherichia coli могут экспрессироваться в Enterobacter agglomerans. Поэтому решили клонировать ген edd и ген eda из Escherichia coli.
Указанные два гена образуют у Escherichia coli оперон (J. Bacteriol., 174 (14): 4638-46, July 1992). Поэтому сконструировали edd-F (SEQ ID NO:1) и eda-R (SEQ ID NO:2) в качестве праймеров, которые могут одновременно амплифицировать оба гена, чтобы с помощью ПЦР амплифицировать фрагмент ДНК, включающий в себя оба гена. ПЦР выполняли, используя ДНК-полимеразу Pyrobest (Takara Shuzo), и ПЦР состояла из реакции при 94°С в течение 1 минуты, затем реакций при 94°С в течение 30 секунд, 60°С в течение 30 секунд и 72°С в течение 3 минут, повторяемых в ходе 30 циклов.
Затем полученный амплифицированный фрагмент полностью расщепляли ферментами рестрикции SalI и BamHI, лигировали с плазмидой pMW219, полностью расщепленной ферментами рестрикции SalI и BamHI, и использовали для того, чтобы трансформировать Escherichia coli JM109 (приобретенную из Takara Shuzo). Из полученных трансформантов отобрали пять штаммов клонов, содержащих фрагмент требуемого размера, и из указанных штаммов экстрагировали плазмиды.
Каждую плазмиду вводили в штамм Enterobacter agglomerans AJ13601 способом электропорации (Miller, J.H., "A Short Course in Bacterial Genetics; Handbook", Cold Spring Harbor Laboratory Press, U.S.A., p.279, 1992) и измеряли активности EDD и EDA, чтобы отобрать клон, в котором экспрессировались ген edd и ген eda.
Штамм AJ13601 является бактериальным штаммом, полученным следующим образом. Штамм Enterobacter agglomerans AJ13355 выделяли из почвы в виде штамма, проявляющего резистентность к L-глутаминовой кислоте в кислых условиях среды и высокую скорость роста. Затем из штамма AJ13355 получали мутантный штамм, продуцирующий небольшое количество слизи, и разрушали ген aKGDH, чтобы получить штамм AJ13356. Штамм AJ13356 был дефицитным по активности aKGDH в результате нарушения гена субъединицы aKGDH-E1 (sucA). Затем в штамм AJ13356 вводили плазмиду RSFCPG, имеющую ген цитратсинтазы (gltA), ген фосфоенолпируваткарбоксилазы (ppc) и глутаматдегидрогеназы (gdhA) Escherichia coli, и плазмиду pSTVCB, имеющую ген gltA, полученный из Brevibacterium lactofermentum, чтобы получить штамм SC17sucA/RSFCPG+pSTVCB. Из полученного штамма отобрали штамм AJ13601 как бактериальный штамм, проявляющий повышенную резистентность к L-глутаминовой кислоте при низком значении рН окружающей среды и наилучшую скорость роста (EP 1078989 A2).
Штаммы, отобранные случайным образом из трансформантов, в которые вводили плазмиду, содержащую фрагменты гена edd и eda, как описано выше, культивировали в течение 15 часов в жидкой среде LBGM9 (среда, содержащая 10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl и 5 г/л глюкозы с добавлением 1/10 объема отдельно стерилизованной 10ЧМ9 (128 г/л Na2HPO4H2O, 30 г/л KH2PO4, 5 г/л NaCl, 10 г/л NH4Cl), содержащей тетрациклин, хлорамфеникол и канамицин, каждый в количестве 12,5, 25 или 25 мг/л. Клетки из указанных культуральных бульонов собирали центрифугированием, дважды промывали 50 мМ Трис-HCl-буфером (рН 7,6) и 10 мМ MaCl2 и затем суспендировали в таком же буфере. Клетки разрушали ультразвуковой обработкой и центрифугировали при 15000 об/мин в течение 30 минут и надосадок использовали в качестве раствора неочищенного фермента.
Активности EDD и EDA измеряли одновременно посредством измерения продуктов реакции, полученных благодаря двум ферментам, используя фотометрический способ. А именно, смешивали 50 мМ Трис-HCl (рН 8,0), 10 мМ MgCl2, 1 мМ EDTA, 1 мМ APAD (ацетилпиридинадениндинуклеотид), 5 мМ K2HPO4, 20 единиц глицеральдегид-3-фосфатдегидрогеназы, 6-фосфоглюконовую кислоту и раствор неочищенного фермента, и измеряли увеличение оптической плотности при 365 нм, чтобы измерить глицеральдегид-3-фосфат, образуемый из 6-фосфоглюконовой кислоты в качестве субстрата. Такое же измерение выполняли для штамма, в который вводили только вектор. Результаты показаны в таблице 1.
Подтвердили, что все штаммы обладали повышенными активностями. В штамме с самыми высокими активностями активности были усилены примерно в 7,2 раза. Плазмида данного штамма названа pMW-EDDA.
<2> Получение L-глутаминовой кислоты с использованием штамма, в котором усилен путь Энтнера-Дудорова
Затем исследовали влияние усиления пути Энтнера-Дудорова на продукцию L-глутаминовой кислоты.
Штамм Enterobacter agglomerans AJ13601, использованный в указанном выше разделе, содержал два вида плазмид, а штаммы, в которые, кроме того, вводили ген edd и ген eda, содержали три вида плазмид. Поэтому к культуре необходимо было добавить три вида агентов, и поэтому рост был очень плохим, т.е. в системе оценки культуры, продуцирующей L-глутаминовую кислоту, почти не наблюдалось роста. Поэтому использовали только два вида плазмид посредством введения гена цитратсинтазы (также называемой в дальнейшем "CS") Brevibacterium lactofermentum, гена edd и гена eda в одну плазмиду.
При сравнении вектора pSTV 28, используемого для конструирования плазмиды pSTVCB, содержащей ген CS Brevibacterium lactofermentum, и вектора pMW219, используемого для клонирования гена edd и гена eda, в случае первого вектора наблюдали большее число копий. Было сделано предположение, что в то время как в штамме AJ13601 ген CS Brevibacterium lactofermentum усилен вектором pSTV28, необходимо увеличить экспрессию в случае введения указанного гена с использованием pMW219. Поэтому сконструировали ген, в котором область промотора в гене CS заменили промотором гена CS Escherichia coli.
А именно область промотора в гене CS амплифицировали, используя праймеры GLTES1 (SEQ ID NO:3) и GLTEBO (SEQ ID NO:4) и хромосому штамма Escherichia coli W3110 в качестве матрицы. Затем фрагмент, содержащий область ОРС гена CS, амплифицировали с использованием праймеров GLTBBO (SEQ ID NO:5) и GLTBA1 (SEQ ID NO:6) и хромосомы Brevibacterium lactofermentum штамма 2256 в качестве матрицы. Выполняли перекрестную ПЦР, используя оба фрагмента в качестве матриц и праймеры GLTES2 (SEQ ID NO:7) и GLTBA2 (SEQ ID NO:8), чтобы получить фрагмент-мишень. Полученный фрагмент расщепляли ферментами рестрикции SmaI и HindIII и вводили в тот же самый сайт pSTV28, чтобы получить pSTV-CB (*). Полученную плазмиду расщепляли KpnI и HindIII и отбирали фрагмент слитого гена, содержащий промотор гена CS Escherichiacoli и кодирующую область гена CS Brevibacterium lactofermentum, и концы затупляли ДНК-полимеразой Т4. Указанный фрагмент слитого гена вводили в SmaI-сайт pMW219, чтобы получить pMW-CB(*). Затем pMW-EDDA обрабатывали BamHI и лигировали с указанным выше фрагментом слитого гена и концы продукта лигирования затупляли ДНК-полимеразой Т4, чтобы получить pMW-CB(*)ED.
Затем AJ13601 выращивали на качалке в течение ночи в жидкой среде LBGM9 при 31,5°С, соответствующим образом разводили так, чтобы получить от 100 до 200 колоний на чашку, и наносили на чашки с LBGM9, содержащей 12,5 мг/л тетрациклина. Реплики появившихся колоний переносили на чашку с LBGM9, содержащей 12,5 мг/л тетрациклина и 25 мг/л хлорамфеникола, и отобрали штамм, который стал чувствительным к хлорамфениколу, и назвали G106S. Штамм G106S содержал только RSFCPG и был дефицитным по pSTVCB. Штаммы, полученные введением pMW-CB(*) или pMW-CB(*) в указанный штамм, назвали G106S pMWB(*) и G106S pMW-CB(*), соответственно.
Чтобы оценить способность указанных штаммов продуцировать L-глутаминовую кислоту проводили оценку культуры, используя сосуд-ферментер. Объем используемой среды составлял 300 мл среды, содержащей 50 г/л сахарозы, 0,4 г/л MgSO4, 0,1 мл/л GD-113 (противопенный агент), 4 г/л (NH4)2SO4, 2 г/л KH2PO4, 4 г/л дрожжевого экстракта, 10 мг/л FeSO4H2O, 10 мг/л MnSO4-5H2O, 0,4 г/л L-лизина, 0,4 г/л DL-метионина, 0,4 г/л диаминопимелиновой кислоты, 12,5 мг/л тетрациклина и 25 мг/л хлорамфеникола. Культивирование выполняли в условиях аэрации 1/1 VVM, перемешивая при 1300 об/мин, и при значении рН 6,0, которое регулировали аммиаком, вплоть до того, как сахароза была израсходована. Изменения оптической плотности при 660 нм и количества продуцируемой L-глутаминовой кислоты в среде во времени показаны на Фиг.1. Кроме того, конечные количества продуцируемой L-глутаминовой кислоты показаны в таблице 2.
(х 1/101)
Обнаружено, что способность продуцировать L-глутаминовую кислоту можно повысить путем усиления пути Энтнера-Дудорова, хотя рост культуры был замедленным.
<3> Исследование продукции ацетоина и 2,3-бутандиола штаммом с усиленным путем Энтнера-Дудорова
Вышеуказанные штаммы G106S pMWCB(*)ED и G106S pMW-CB(*) культивировали таким же образом, как при оценке способности продуцировать L-глутаминовую кислоту в примере <2>, и в течение времени измеряли количества ацетоина и 2,3-бутандиола в среде и количество клеток. Измерения проводили газовой хроматографией (Shimadzu Corporation, GC-1700A) при следующих условиях.
Используемая колонка: Varian Poraploto Plot FS25х32 (0,32 ммЧ25 М).
Температура: испарительная камера: 250°С, колонка: 240°С, FID: 250°С.
Давление на входе в колонку: 180 кПа.
Скорость потока газа-носителя: 1,6062 мл/мин.
Результаты показаны на фиг.2А (количество ацетоина в среде), фиг.2В (количество 2,3-бутандиола в среде) и фиг.2С (общее количество продуцируемого ацетоина и 2,3-бутандиола на единичную клетку). Обнаружено, что продукция ацетоина и 2,3-бутандиола увеличивалась при усилении пути Энтнера-Дудорова.



| название | год | авторы | номер документа |
|---|---|---|---|
| Микроорганизм рода Escherichia, продуцирующий L-триптофан, и способ продуцирования L-триптофана с использованием данного микроорганизма | 2015 |
|
RU2678139C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 1999 |
|
RU2194076C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 1999 |
|
RU2188236C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2002 |
|
RU2282662C2 |
| ОРГАНИЧЕСКАЯ АЗОТСОДЕРЖАЩАЯ КОМПОЗИЦИЯ И УДОБРЕНИЕ, ВКЛЮЧАЮЩЕЕ ЕЕ | 2002 |
|
RU2291139C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2002 |
|
RU2288271C2 |
| ШТАММ ENTEROBACTER AGGLOMERANS, СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ МИКРООРГАНИЗМА ДЛЯ ВЫРАБОТКИ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2000 |
|
RU2261272C2 |
| СПОСОБ ПРОИЗВОДСТВА L-АМИНОКИСЛОТ | 2007 |
|
RU2422530C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН b2383 | 2006 |
|
RU2333952C2 |
Изобретение относится к биотехнологии и представляет собой способ получения L-аминокислоты. L-аминокислоту получают посредством культивирования микроорганизма, обладающего способностью продуцировать L-аминокислоту в среде, с дальнейшим сбором L-аминокислоты из среды. В качестве микроорганизма используют грамотрицательную бактерию, имеющую путь Энтнера-Дудорова и модифицированную так, что активность 6-фосфоглюконатдегидратазы или активность 2-кето-3-дезокси-6-фосфоглюконатальдолазы, или активности обоих ферментов усилены. Изобретение позволяет получать L-аминокислоты с высокой степенью эффективности. 6 з.п. ф-лы, 2 ил., 2 табл.
| Теплоприемник к радиационным пирометрам и радиометрам | 1954 |
|
SU102542A1 |
| US 6040160, 21.03.2000 | |||
| ЕР 1078989, 28.02.2001 | |||
| Штамм бактерий ЕSснеRIснIа coLI - продуцент L-треонина | 1987 |
|
SU1694643A1 |

