Область техники
Настоящее изобретение относится к способу получения спирта, такого как метанол, с использованием микроорганизма. Более конкретно, настоящее изобретение относится к способу превращения алкана в спирт при чрезвычайно мягких условиях биохимического окисления с использованием микроорганизма.
Уровень техники
Используемый в настоящее время способ получения метанола включает в себя «стадию получения синтез-газа» для реформинга метана в качестве сырья в смешанный газ моноксида углерода и водорода, и «стадию синтеза метанола» для реакции синтез-газа в присутствии катализатора для превращения этого газа в метанол. Стадию получения синтез-газа и стадию синтеза метанола проводят при высокой температуре и при высоком давлении, а именно, стадию получения синтез-газа проводят при 850-880°С, а стадию синтеза метанола проводят при 50-100 атм. Таким образом, желательным является способ синтеза метанола, который может проводиться при более мягких условиях. Хотя недавно был описан способ синтеза метанола из метана с высоким выходом с использованием сравнительно термостабильного катализатора типа комплекса металла (Science, vol. 280, 24, 560-563, April, 1998), этот способ все-таки требует температуры 100°С или выше.
Между тем, если используют фермент монооксигеназу (далее упоминаемую в виде аббревиатуры "ММО"), утилизирующей метан бактерии, метанол может быть синтезирован непосредственно из метана при обычной температуре и обычном давлении. Известно, что имеется ММО растворимого типа и ММО мембраносвязанного типа. Фермент ММО растворимого типа является комплексным белком, состоящим из трех компонентов: гидроксилазы, Компонента В и редуктазы (Компонента С). Реакцию окисления проводят гидроксилазой среди этих компонентов, и сообщалось, что, если эта гидроксилаза является химически восстановленной, вместо использования редуктазы, эта реакция окисления может катализироваться только гидроксилазой (J. Biol. Chem., 264(17) 10023-10033, 1989). Эту гидроксилазу называют также Компонентом А, и она состоит из трех типов белковых субъединиц, α, β и γ.
В патенте США № 5190870 описан ферментативный способ получения спирта с использованием гидроксилазы, очищенной из утилизирующей метан бактерии. Однако способ очистки этой гидроксилазы является сложным, и уменьшение активности этого фермента является значительным после его выделения, что приводит к нестабильной активности этого фермента. Кроме того, хотя в патенте США № 5192672 описан способ стабилизации активности гидроксилазы после очистки, фундаментальная проблема этого сложного способа очистки все еще остается нерешенной.
Кроме того, известен также способ микробиологического получения метанола с использованием утилизирующей метан бактерии. Однако клетки, содержащие ММО, содержат также метанолдегидрогеназу и т.д. и, следовательно, существует проблема, что метанол, производимый окислением метана, немедленно окисляется и посредством этого превращается в формальдегид, т.е. метанол метаболизируется в этих клетках. В патенте Японии Japanese Patent Laid-open (KOKAI) № 3-43090 описывает способ селективного ингибирования метанолдегидрогеназы циклопропаном. Однако этот способ является чрезвычайно сложным, т.е. он включает в себя замену циклопропаном воздушной фазы в суспензии клеток, содержащих ММО, и затем удаление циклопропана гелием и т.д., и этот способ страдает также от проблемы, заключающейся в том, что метанолдегидрогеназа может не инактивироваться в достаточной степени циклопропаном.
В Journal of General Microbiology (1992), 138, 1301-1307 описано, что активность ММО могла бы успешно экспрессироваться экспрессией Компонента В и редуктазы ММО растворимого типа, происходящей из Methylococcus capsulatus (Bath), в Escherichia coli и смешиванием их с природной гидроксилазой. Кроме того, в Arch. Microbiol. (1999) 171:364-370, сообщается, что активность ММО могла бы успешно экспрессироваться введением гена ММО растворимого типа в утилизирующую метан бактерию, имеющую только ММО мембраносвязанного типа. Однако, нет примера экспрессии активностей всех компонентов ММО в микроорганизме, который не утилизирует метан или метанол, и, следовательно, желательной является экспрессия этого сложного комплекса ферментов в гетерогенном организме.
Описание изобретения
Целью настоящего изобретения является экспрессия всех компонентов ММО растворимого типа (далее называемой также сокращенно "sMMO") в микроорганизме, который не утилизирует метан и метанол, и посредством этого продукция активного фермента sMMO и обеспечение способа получения спирта из алкана с использованием такого микроорганизма.
Авторы настоящего изобретения, поначалу ставили целью ввести кластер генов, кодирующий ММО растворимого типа, в микроорганизм, не имеющий какого-либо фермента, окисляющего метанол, такого как метанолдегидрогеназа, т.е. в микроорганизм, который не утилизирует метан или метанол, для конструирования микроорганизма, который экспрессирует этот кластер генов в виде активных белков, и тем самым обеспечить способ получения метанола из метана с использованием этого микроорганизма. Однако sMMO является комплексным белком, состоящим из 5 субъединиц, и, следовательно, нелегко экспрессировать его в активной форме, в частности, в гетерогенном микроорганизме. Таким образом, авторы изобретения провели широкое исследование на типах плазмид и промоторов, используемых для плазмиды, предназначенной для экспрессии ММО растворимого типа, и условий культивирования. В результате авторам изобретения удалось экспрессировать sMMO в клетках гетерогенного микроорганизма с сохранением активности этого фермента. Кроме того, авторам изобретения удалось также получить метанол из метана с использованием полученного микроорганизма и, следовательно, осуществить настоящее изобретение.
Настоящее изобретение обеспечивает следующее.
(1) Способ получения спирта, предусматривающий культивирование рекомбинанта микроорганизма, которому несвойственна способность утилизировать алкан и спирт, образующийся окислением этого алкана, причем указанный рекомбинант приобрел способность превращения алкана в спирт вследствие трансформации ДНК, кодирующей метаноксигеназу, и оставление полученной культуры, клеток, выделенных из этой культуры, или обработанного продукта этих клеток с алканом для продукции спирта.
(2) Способ получения спирта по пункту (1), где указанная метаноксигеназа является метаноксигеназой растворимого типа.
(3) Способ получения спирта по пункту (2), где указанная метаноксигеназа состоит из метангидроксилазы, Компонента В и редуктазы.
(4) Способ получения спирта по любому из пунктов (1)-(3), где указанная ДНК, кодирующая метаноксигеназу, является геном метаноксигеназы растворимого типа Methylococcus capsulatus.
(5) Способ получения спирта по любому из пунктов (1)-(4), где указанным микроорганизмом является бактерия Escherichia, коринеформная бактерия или бактерия Bacillus.
(6) Способ получения спирта по пункту (5), где указанный микроорганизм является бактерией Escherichia.
(7) Способ получения спирта по пункту (6), где указанный микроорганизм культивируют при 20-30°С.
(8) Способ получения спирта по любому из пунктов (1)-(7), где указанный алкан является алканом, имеющим 1-8 атомов углерода, а спирт является спиртом, который образуется окислением этого алкана.
(9) Способ получения спирта по пункту (8), где указанный алкан является метаном, а указанный спирт является метанолом.
ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ
Далее настоящее изобретение будет объяснено подробно.
Микроорганизм, используемый в настоящем изобретении, является микроорганизмом, которому несвойственна способность утилизировать алкан и спирт, образующийся окислением этого алкана, но этот микроорганизм приобрел способность превращения алкана в спирт вследствие трансформации его ДНК, кодирующей sMMO. Что касается свойства, заключающегося в том, что используемый в настоящем изобретении микроорганизм не утилизирует спирт, оно является несущественным, превращает ли этот микроорганизм алкан в спирт или не превращает, тогда как предпочтительным для накопления продуцируемого спирта является то, что данный микроорганизм не должен утилизировать спирт.
Микроорганизм, который не имеет природной способности утилизировать алкан и спирт, который образуется окислением этого алкана, не ограничивается особо, пока он является микроорганизмом, который может приобретать способность превращения алкана в спирт. Характерные примеры включают в себя бактерии Escherichia, такие как Escherichia coli, коринеформные бактерии, такие как Brevibacterium lactofermentum (Corynebacterium glutamicum), и бактерии Bacillus, такие как Bacillus subtilis, и т.д.
ДНК, кодирующая метаноксигеназу, предпочтительно является ДНК, кодирующей ММО растворимого типа (sMMO). sMMO состоит из Компонента А (метангидроксилазы), Компонента В и Компонента С (редуктазы). Компонент А состоит из субъединиц α, β и γ. Хотя эти компоненты могут вводиться в данный микроорганизм по отдельности, их предпочтительно вводят с использованием единого вектора, содержащего ДНК, кодирующую все эти компоненты. В дальнейшем описании кластер генов, кодирующий все эти компоненты sMMO, называют для удобства геном sMMO.
Ген sMMO может быть получен из хромосомной ДНК, утилизирующей метан бактерии, например, штамма Methylococcus capsulatus NCIMB 11132. Этот штамм может быть получен из BCIMB (The National Collections of Industrial, Food and Marine Bacteria Ltd., 23 St. Machar Drive, Aberdeen AB2 1RY, Scotland, UK).
Пример способа получения фрагмента, содержащего ген sMMO, из Methylococcus capsulatus в качестве утилизирующей метан бактерии Bacillus описан ниже. Однако такой фрагмент может быть также получен из других утилизирующих метан бактерий подобным образом.
Сначала средой для культивирования Methylococcus capsulatus может быть любая среда, в которой эта бактерия может пролиферировать в достаточной степени. Пример предпочтительной среды включает в себя среду Whittenbury et al. (J. Gen. Microbiol., 61, 205-208, 1970). Все пространство в культуральном сосуде, содержащем эту среду, заменяют смешанным газом и кислородсодержащим газом (воздухом и т.д.) и Methylococcus capsulatus инокулируют в эту среду, находящуюся в контакте с этим газом. Methylococcus capsulatus является аэробной бактерией, и культивирование проводят при 20-50°С в аэробных условиях в виде периодической культуры или непрерывной культуры.
ДНК-фрагмент, содержащий ген sMMO, может быть выделен и получен гибридизационным способом, как описано ниже, из библиотеки ДНК, полученной из хромосом штамма Methylococcus capsulatus, с использованием, например, зонда, ДНК-фрагмента, содержащего часть гена sMMO, полученного при помощи ПЦР (полимеразной цепной реакции) с применением олигонуклеотидов, приготовленных на основании известной нуклеотидной последовательности гена sMMO (GenBank Accession M90050 M32314 M58498 M58499, Stainthorpe, A.C., et al., Gene, 91 (1), 27-34, 1990, SEQ ID NO:5), в качестве праймеров и хромосомной ДНК Methylococcus capsulatus в качестве матрицы. Открытые рамки считывания, содержащиеся в гене sMMO, обозначены mmoX, mmoY, mmoB, mmoZ, OrfY и mmoC в этом порядке от 5'-конца.
Хромосомная ДНК может быть экстрагирована из культурального бульона штамма Methylococcus capsulatus NCIMB 11132 обычно используемым способом, известным per se (например, способом, описанным в Biochem. Biophys. Acta, 72, 619 (1963) и т.д.).
Библиотека ДНК может быть получена разрушением хромосомной ДНК с использованием подходящего рестрикционного фермента, такого как BamHI, лигированием полученных ДНК-фрагментов разного размера с плазмидным вектором, таким как pUC18 (производимым Takara Shuzo), и трансформацией подходящего хозяина, такого как Escherichia coli JM109, реакционной смесью лигирования.
Зонд для отбора клона, имеющего ДНК-фрагмент, содержащий ген sMMO, гибридизацией может быть получен при помощи ПЦР с использованием олигонуклеотидов, сконструированных подходящим образом на основании известной нуклеотидной последовательности гена sMMO, например, нуклеотидных последовательностей, показанных как SEQ ID NO:1 и 2, в качестве праймеров и хромосомной ДНК Methylococcus capsulatus в качестве матрицы.
Гибридизацию колоний выполняют в отношении библиотеки хромосомной ДНК штамма Methylococcus capsulatus NCIMB 11132 с использованием зонда, полученного, как описано выше. ДНК-фрагмент, содержащий ген sMMO или его часть, может быть получен экстракцией плазмидной ДНК из клона, который гибридизовался с этим частичным ДНК-фрагментом гена sMMO, используемым в качестве зонда в гибридизации, и получением инсертированного фрагмента расщеплением этой плазмидной ДНК подходящей рестриктазой.
Когда фрагмент клона, полученный, как описано выше, содержит часть гена sMMO, может быть получена другая область при помощи ПЦР-способа, способа гибридизации и т.п. Например, когда клонированный фрагмент не имеет 5'-концевой области гена sMMO, область выше от клонированного фрагмента может быть получен при помощи способа 5'-RACE (способа быстрой амплификации концов кДНК). Кроме того, область выше от гена sMMO может быть также получен при помощи ПЦР с использованием олигонуклеотидов, показанных в виде SEQ ID NO:3 и 4, в качестве праймеров и хромосомной ДНК штамма NCIMB в качестве матрицы. Полученный ДНК-фрагмент может быть лигирован с полученным ранее фрагментом гена sMMO с получением полноразмерного фермента sMMO.
Белки, не являющиеся белками дикого типа, каждый компонент или каждая субъединица sMMO могут иметь аминокислотную последовательность, которая включает в себя делецию, инсерцию или добавление одного или нескольких аминокислотных остатков, если сохраняется функция каждого компонента или каждой субъединицы. Хотя число «нескольких» аминокислот, указанных здесь, отличается в зависимости от положения аминокислотных остатков в трехмерной структуре белка или типа аминокислот, оно может быть предпочтительно 2-10, более предпочтительно 2-5, наиболее предпочтительно 2-3.
ДНК, кодирующая белок или пептид, по существу идентичный описанному выше ферменту sMMO, включает в себя ДНК, которая может гибридизоваться с открытой рамкой считывания в нуклеотидной последовательности, показанной в виде SEQ ID NO:4, или зондом, который может быть получен из этой последовательности при строгих условиях и кодирует белок, который может составлять активный фермент sMMO. Вышеупомянутые "строгие условия" включают в себя условия, в которых образуется так называемый специфический гибрид, а неспецифический гибрид не образуется. Трудно выразить эти строгие условия с использованием какой-либо числовой величины. Однако, например, строгие условия включают в себя условия, в которых ДНК, имеющие высокую гомологию, например, ДНК, имеющие гомологию 50% или более, гибридизуются друг с другом, но ДНК, имеющие гомологию, более низкую, чем указанная выше, не гибридизуются друг с другом. Альтернативно, строгие условия включают в себя условия, посредством которых ДНК гибридизуются друг с другом при концентрации соли, соответствующей типичным условиям промывания гибридизации по Саузерну, т.е. 1×SSC, 0,1% ДСН, предпочтительно 0,1×SSC, 0,1% ДСН, при 60°С.
Вектор, используемый для введения гена sMMO в микроорганизм-хозяин, может быть любым вектором, автономно реплицирующимся в клетке микроорганизма-хозяина, и характерные примеры включают в себя, для Escherichia coli, плазмидные векторы pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219, pMW218 и т.д. Кроме того, векторы для коринеформных бактерий включают в себя рАМ330 (см. выложенную публикацию патента Японии № 58-67699), pHM1519 (см. выложенную публикацию патента Японии № 58-77895), pAJ655, pFJ611, pAJ1844 (см. выложенную публикацию патента Японии № 58-192900 в отношении этих плазмид), pCG1 (см. выложенную публикацию патента Японии № 57-134500), pCG2 (см. выложенную публикацию патента Японии № 58-35197), pCG4, pCG11 (см. выложенную публикацию патента Японии № 57-183799 в отношении этих плазмид), pHK4 (см. выложенную публикацию патента Японии № 5-7491) и т.д. Кроме того, векторы для бактерий Bacillus включают в себя pUB110, pHY300PLK, pHV1248, pE194, pC194, pBС16, pSA0501, pSA2100, pAM77, pT181, pBD6, pBD8 и pBD64, pHV14 и т.д.
Промотор гена sMMO может быть заменен подходящим промотором в зависимости от микроорганизма, в который вводят этот ген. Примеры таких промоторов включают в себя промотор lac, промотор trp, промотор trc, промотор tac, промотор PR и промотор PL фага лямбда, промотор tet, промотор amyE и т.д. Если экспрессирующий вектор, содержащий промотор, используют в качестве вектора, лигирование гена sMMO, вектора и промотора может достигаться в единой операции лигирования. Примеры таких векторов включают в себя рКК233-3, содержащий промотор tac (производимый Pharmacia), и т.д.
В качестве способа трансформации для введения вектора с включенным в него геном sMMO в рассматриваемый микроорганизм может быть использован, например, способ обработки реципиентных клеток хлоридом кальция для увеличения проницаемости этих клеток для ДНК, который был сообщен для Escherichia coli К-12 (Mandel, M. and Higa, A., J. Mol. Biol., 53, 159 (1970)) и способ получения компетентных клеток из клеток, которые находятся в фазе роста, с последующим введением в них ДНК, который был сообщен для Bacillus subtilis (Duncan, C.H., Wilson, G.A. and Young, F.E., Gene, 1, 153 (1977)). Альтернативно может быть также использован способ превращения ДНК-реципиентных клеток в протопласты или сферопласты, которые могут легко поглощать рекомбинантную ДНК, с последующим введением рекомбинантной ДНК в ДНК-акцепторные клетки, который, как известно, применим к Bacillus subtilis, актиномицетам и дрожжам (Chang, S. and Choen, S.N., Molec. Gen. Genet., 168, 111 (1979); Bibb, M.J., Ward, J.M. and Hopwood, O.A., Nature, 274, 398 (1978); Hinnen, A., Hicks, J.B. and Fink, G.R., Proc. Natl. Sci., USA, 75, 1929 (1978)). Способ трансформации может быть подходящим образом выбран из этих способов в зависимости от клетки, используемой в качестве хозяина. Кроме того, рекомбинантная ДНК может быть введена в реципиентную бактерию, принадлежащую к бактериям Brevibacterium или Corynebacterium, способом электропорации (Sugimoto et al., выложенный патент Японии № 2-207791) (ОР1999).
Способы получения библиотеки геномной ДНК, гибридизации, ПЦР, получения плазмидной ДНК, расщепления и лигирования ДНК, трансформации и т.д. описаны в Sambrook, J., Fritsch, E.F., Maniatis, T., Molecular Cloning, Cold Spring Harbor Laboratory Press, 1.21 (1989).
Штамм E. coli JM109, трансформированный экспрессирующим вектором pRSsMMOTB, был депонирован в независимом административном агентстве National Institute of Advanced Industrial Depository (postal code 305-5466, Tsukuba Central 6, 1-1 Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, Japan) on August 2, 2001 и получил номер доступа FERM P-18446. Затем этот депозит был превращен в международный депозит согласно условиям Будапештского договора 19 августа 2002 года и получил номер доступа FERM BP-8153.
Культивированием микроорганизма, который приобрел способность превращать алкан в спирт благодаря трансформации ДНК, кодирующей ДНК, содержащую ген sMMO, и обеспечением возможности полученной культуре или клеткам, выделенным из этой культуры, существовать с алканом для продуцирования спирта может быть получен этот спирт.
Среда, используемая для культивирования микроорганизма с введенным геном sMMO, может быть подходящим образом выбрана в зависимости от используемого микроорганизма. Например, она может быть обычной средой, которая содержит источник углерода, источник азота, неорганические ионы и другие органические ингредиенты, если требуется.
В качестве источника углерода могут быть использованы сахариды, такие как глюкоза, сахароза, лактоза, галактоза, фруктоза или гидролизат крахмала, спирты, такие как глицерин или сорбит, или органические кислоты, такие как фумаровая кислота, лимонная кислота или янтарная кислота.В качестве источника азота могут быть использованы неорганические соли аммония, такие как сульфат аммония, хлорид аммония или фосфат аммония, органический азот, такой как гидролизат белка сои, газообразный аммиак, водный аммиак и т.д.
Желательным является добавление необходимых веществ, таких как витамин В1, дрожжевой экстракт и т.д. к среде в подходящих количествах в виде органических питательных веществ, требующихся в незначительных количествах. Кроме них фосфат калия, сульфат магния, ионы железа, ионы марганца и т.д. добавляют в небольших количествах, если требуется.
Культивирование проводят предпочтительно в аэробных условиях в течение 16-72 часов. Температуру культуры предпочтительно регулируют таким образом, чтобы она составляла 25°С-45°С, а рН предпочтительно регулируют таким образом, чтобы он был равен 5-8, во время культивирования. Для коррекции рН могут быть использованы неорганические или органические, кислотные или щелочные вещества, а также газообразный аммиак и т.д.
При применении в качестве микроорганизма-хозяина бактерии Escherichia культивирование предпочтительно проводят при 20-30°С для получения субъединиц, составляющих sMMO, в виде растворимых пептидов.
В качестве катализатора для превращения алкана в спирт, кроме клеток и культуры, содержащей клетки, может быть также использован обработанный продукт этих клеток. Обработанным продуктом клеток могут быть клетки, обработанные ацетоном, лиофилизированные клетки, бесклеточный экстракт, полученный из обработанных ацетоном или лиофилизированных клеток, или живых клеток, продукт фракционирования, такой как мембранная фракция, фракционированная из бесклеточного экстракта, и иммобилизованный продукт этих клеток, бесклеточный экстракт или продукт фракционирования. Приведением таких клеток или обработанного продукта клеток в контакт с алканом и обеспечением возможности реакции может быть получен спирт в этом реакционном растворе. Используемый микроорганизм может состоять из одного типа микроорганизма или из смеси двух или нескольких типов микроорганизмов. Если алкан не ингибирует рост данного микроорганизма, алкан может быть добавлен к среде, используемой для культивирования данного микроорганизма для одновременного проведения культивирования микроорганизма и получения спирта. В этом случае алкан может быть введен в среду добавлением в газообразную фазу, находящуюся в контакте со средой, или барботированием среды алканом. Кроме того, с использованием микроорганизма, обнаруживающего сильную восстанавливающую активность, такого как бактерии Escherichia, в качестве микроорганизма, используемого в настоящем изобретении, или добавлением НАДН к реакционной смеси, реакция, катализируемая sMMO, может быть эффективно усилена.
Алкан, к которому применяют настоящее изобретение, предпочтительно является алканом, имеющим 1-8 атомов углерода, более предпочтительно 1-6 атомов углерода, особенно предпочтительно 1-5 атомов углерода. В частности, наиболее предпочтительным является метан. Кроме того, положение атома водорода в алкане, окисляемого sMMO, особо не ограничивается, и он может быть водородом, связывающимся с концевым углеродом, или водородом, связывающимся с углеродом, с которым связаны два или более углеродов. Алкан может быть линейным, разветвленным или циклическим алканом. Алкан может иметь заместитель, такой как галоген.
Исследовали субстратную специфичность sMMO, полученного из штамма Methylococcus capsulatus Bath, и было установлено, что этот штамм способен окислять алканы, в том числе по меньшей мере от метана до октана, с образованием спирта.
Краткое объяснение чертежа
чертеж показывает калибровочные кривые, полученные построением с концентрацией метанола (мМ) на абсциссе и оптической плотности на ординате. Результаты для E. coli JM109/pRS показаны ◆, а результаты для E. coli JM109/pRSsMMOTB (AJ 13852) показаны ▪.
Наилучший способ осуществления изобретения.
Далее настоящее изобретение будет объясняться более конкретно со ссылкой на следующие примеры.
<1> Получение библиотеки хромосомной ДНК, утилизирующей метан бактерии, Methylococcus capsulatus
Пространство в культуральном сосуде, содержащем среду Whittenbury et al. (J. Gen. Microbiol., 61, 205-208, 1970), заменяли смешанным газом метана и воздуха. Утилизирующую метан бактерию, штамм Methylococcus capsulatus NCIMB 11132, инокулировали в эту среду, находящуюся в контакте с газом, и культивировали в аэробных условиях в виде периодической культуры при продолжении замены газа. Хромосомную ДНК экстрагировали из клеток Methylococcus capsulatus NCIMB 11132, культивируемых, как описано выше, по способу, описанному в Biochem. Biophys. Acta, 72, 619 (1963). Хромосомные ДНК полностью расщепляли рестриктазой BamHI. Полученные ДНК-фрагменты различных размеров встраивали в плазмидный вектор pUC18 (производимый Takara Shuzo) в сайте BamHI. Escherichia coli JM109 трансформировали полученными рекомбинантными плазмидами с получением библиотеки хромосомной ДНК.
<2> Клонирование гена sMMO гибридизацией колоний
Клон, содержащий фрагмент гена sMMO, отбирали из вышеупомянутой библиотеки хромосомной ДНК гибридизацией колоний. Гибридизационный зонд получали амплификацией фрагмента гена sMMO с использованием ПЦР-способа. Нуклеотидная последовательность гена sMMO Methylococcus capsulatus сообщалась (Gene, 91, 27-34 (1990)), и олигонуклеотиды, имеющие нуклеотидные последовательности, показанные как SEQ ID NO:1 и 2, синтезировали на основе этой последовательности.
Хромосомные ДНК Methylococcus capsulatus, полученные, как описано выше, использовали в качестве матрицы, а вышеупомянутые олигонуклеотиды использовали в качестве праймеров для проведения ПЦР. Для ПЦР проводили 30 циклов, причем каждый цикл состоял из стадии денатурации (94°С, 10 секунд), стадии отжига (55°С, 30 секунд) и стадии удлинения (72°С, 1 минута и 30 секунд).
Гибридизацию колоний проводили в отношении вышеупомянутой библиотеки хромосомной ДНК с использованием частичного фрагмента гена sMMO, амплифицированного, как описано выше, в качестве зонда. Мечение этого зонда и реакцию гибридизации выполняли с использованием набора для детектирования DIG-High Prime DNA Labeling and Detection Kit I (купленного из Boehringer Mannheim) в соответствии с прилагаемым протоколом.
Рекомбинантные плазмидные ДНК экстрагировали из клонов, которые показали положительные результаты в отношении гибридизации, и плазмидные ДНК расщепляли рестриктазой BamHI для подтверждения инсертированных фрагментов. В результате, кроме ДНК-фрагмента, имеющего длину приблизительно 2,3 т.п.н., соответствующий длине плазмиды PUC18, был подтвержден инсертированный ДНК-фрагмент, имеющий размер приблизительно 6 т.п.н.
Эту инсертированную ДНК расщепляли различными рестриктазами для подтверждения, что она была фрагментом, содержащим большую часть рассматриваемого гена sMMO. В результате стало ясным, что часть гена, кодирующего субъединицу α, являющуюся компонентом sMMO, была делетирована в вышеупомянутом фрагменте. Рекомбинантная плазмида, содержащая этот инсертированный ДНК-фрагмент приблизительно 6 т.п.н., была названа pUC6K.
<3> Конструирование плазмиды, содержащей полноразмерный ген sMMO
Затем с использованием ПЦР была получена область, расположенная выше гена sMMO, включающего делецию, как описано ниже. Нуклеотидные последовательности синтезированных праймеров показаны в Списке последовательностей в виде SEQ ID NO:3 и 4. Хромосомные ДНК штамма NCIMB 11132 использовали в качестве матрицы вместе с вышеупомянутыми олигонуклеотидами в качестве праймеров для проведения ПЦР. Для ПЦР проводили 30 циклов, причем каждый цикл состоял из стадии денатурации (98°С, 10 секунд), стадии отжига (60°С, 30 секунд) и стадии удлинения (72°С, 180 секунд). Полученный амплифицированный ДНК-фрагмент приблизительно 1,5 т.п.н. расщепляли рестриктазами EcoRI и BamHI и лигировали в сайте множественного клонирования pUC118. Фрагмент приблизительно 6 т.п.н., вырезанный из pUC6K с использованием BamHI, включали в сайт BamHI вышеупомянутой плазмиды с получением плазмиды pUCsMMO, содержащей ген sMMO полной длины.
<4> Конструирование экспрессионной плазмиды гена sMMO
Затем этот участок гена sMMO вырезали из вышеупомянутой плазмиды pUCsMMO с использованием рестриктаз EcoRI и HindIII и включали в экспрессионный вектор рКК233-3 с получением экспрессионной плазмиды pKKsMMO, которую конструировали таким образом, что транскрипция от промотора tac должна была продолжаться в ген sMMO.
Ген sMMO, включающий в себя область промотора tac, полученную из рКК3-233, вырезали из pKKsMMO расщеплением рестриктазами NdeI и DraI, концы затупляли, расщепляли рестриктазой PstI и затем лигировали с вектором с затупленными концами pRS, подходящим для широкого круга хозяев (описанным в Международной патентной публикации на японском языке (Kohyo) № 3-501682), с получением экспрессионнной плазмиды sMMO pRSsMMOTB.
<5> Подтверждение экспрессии sMMO
Штамм AJ13852 инокулировали в жидкую среду LB (Luria-Bertani), содержащую 20 мг/мл стрептомицина, предварительно культивировали при 25°С, затем инокулировали в таким же образом приготовленную среду в количестве 1% (об./об.) и культивировали при 25°С в качестве основной культуры. Во время выращивания основной культуры, когда достигалась оптическая плотность OD660 0,6, к среде добавляли IPTG (изопропил-β-D-тиокалактопиранозид) в конечной концентрации 1 мМ и эти клетки дополнительно культивировали в течение 2,5 часов. Клетки, полученные с использованием описанных выше условий, разрушали обработкой ультразвуком и фракционировали на растворимую фракцию и нерастворимую фракцию центрифугированием. Каждую фракцию анализировали с использованием антител, направленных на каждый из компонентов-пептидов, составляющих sMMO. В результате, было подтверждено, что все из этих субъединиц, α, β и γ и оба Компонента В и С sMMO, находились в растворимой фракции.
При выполнении всех вышеупомянутых стадий культивирования при 37°С субъединица α полностью образовывала тела включения и не существовала в растворимой фракции. Кроме того, большая часть субъединиц β и γ также были нерастворимыми. Кроме того, при проведении культивирования при 30°С, количество субъединицы α, находящееся в растворимой фракции, становилось меньше в сравнении со случаем, когда культивирование проводили при 25°С.
<6> Получение метанола из метана
Штамм AJ13852 культивировали таким же образом, как описано выше, с использованием 100 мл среды (температура была 25°С) и клетки собирали и затем промывали 50 мМ калийфосфатным буфером (рН 7,0). Затем эти клетки суспендировали в том же самом буфере (рН 7,0) и бесклеточный экстракт получали обработкой ультразвуком. Концентрацию общего белка бесклеточного экстракта доводили до 15 мг/мл. В объеме 1 мл вышеупомянутый бесклеточный экстракт вводили в герметизируемый алюминием флакон с объемом 5 мл, смешивали с НАДН (восстановленным никотинамидадениндинуклеотидом) до конечной концентрации 1 мМ и герметизировали. Затем 5 мл (объем под давлением) метана вводили в этот флакон и давали происходить реакции. Реакции давали происходить при 37°С в течение 60 минут при встряхивании. Кроме того, штамм AJ13852 и E. coli JM109/pRS, в которые был введен только вектор pRS, также культивировали таким же образом, как описано выше, за исключением того, что не вводили метан.
Концентрацию метанола определяли количественно с использованием системы для количественного определения, имеющей три стадии ферментативных реакций. Составы реакционных смесей показаны в таблице 1. Сначала брали 600 мкл реакционной смеси после вышеупомянутой реакции образования метанола, смешивали с 50 мкл 5 н. гидроксида калия, перемешивали в течение достаточного времени и оставляли на 5 минут. Затем реакционную смесь смешивали с 50 мкл 5 М хлорида натрия и доводили до рН 7,5 1 М калийфосфатным буфером. После этого каждую реакционную смесь использовали для следующей ферментативной реакции, после того как денатурированные белки удаляли центрифугированием.
2 мкл
2 мкл
Ферментативную реакцию проводили в соответствии со следующими процедурами. Метанол превращали в формальдегид алкогольоксидазой и получали муравьиную кислоту из формальдегида с использованием формальдегиддегидрогеназы. НАДН, образующийся в вышеупомянутой реакции, определяли количественно с использованием цветной реакции диафоразы в виде оптической плотности при 550 нм. Алкогольдегидрогеназу и формальдегиддегидрогеназу приобретали из Sigma, диафоразу приобретали из Toyobo и иоднитротетразолий приобретали из Nikalai Tesque.
Реакционные смеси, полученные с использованием штамма AJ13852 и E. coli JM109/pRS, в которые вводили только вектор pRS без введения метана, подвергали вышеупомянутой трехстадийной ферментативной реакции, и считали, что измеренные величины оптической плотности соответствуют концентрации метанола 0 М. На основании этих величин метанол добавляли к вышеупомянутой системе определения количества в различных концентрациях и количества метанола строили в зависимости от величин оптической плотности для получения калибровочных кривых (см. чертеж). В результате было обнаружено, что величина концентрации метанола линейно соответствует величине оптической плотности в диапазоне концентраций метанола 200-1000 мкМ в этой системе измерения.
Количество полученного метанола для каждого типа клеток определяли количественно с использованием вышеупомянутых калибровочных кривых. В результате продуцирование метанола не детектировали для E. coli JM109/pRS, тогда как продуцирование метанола детектировали для штамма AJ13852, и концентрация была 240 мкМ. Удельная активность в этой реакции была равна 0,33 нмоль/мин/1 мг белка в растворе фермента.
Промышленная применимость
Согласно настоящему изобретению, способность превращать алкан в спирт могла быть сообщена микроорганизму, которому несвойственна способность утилизировать алкан и спирт, который образуется окислением этого алкана, и с использованием этого полученного микроорганизма может быть получен спирт из алкана.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОРИНЕФОРМНАЯ БАКТЕРИЯ, ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2004 |
|
RU2333247C2 |
| БАКТЕРИЯ РОДА BACILLUS, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2002 |
|
RU2299907C2 |
| СПОСОБ ПРОДУКЦИИ БЕЛКА | 2006 |
|
RU2435863C2 |
| МИКРООРГАНИЗМЫ ДЛЯ ПОВЫШЕННОГО ПРОДУЦИРОВАНИЯ АМИНОКИСЛОТ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2015 |
|
RU2688486C2 |
| СИСТЕМА, СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ БИОПЕРЕРАБОТКИ | 2013 |
|
RU2723620C2 |
| ФЕРМЕНТАЦИОННЫЙ СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 1994 |
|
RU2204605C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2351653C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2368659C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2003 |
|
RU2307165C2 |
Изобретение относится к биотехнологии и представляет собой способ получения метанола. Данный способ включает культивирование рекомбинанта E.coli при температуре 20-30°С, а полученную культуру, клетки, выделенные из этой культуры, или обработанный продукт этих клеток оставляют с метаном для продукции метанола. При этом указанный рекомбинант приобрел способность превращать метан в спирт вследствие трансформации ДНК, кодирующей метаноксигеназу растворимого типа. Согласно данному изобретению экспрессируются все компоненты метаноксигеназы, что позволяет осуществить способ получения метанола в более мягких условиях. 2 з.п. ф-лы, 1 ил., 1 табл.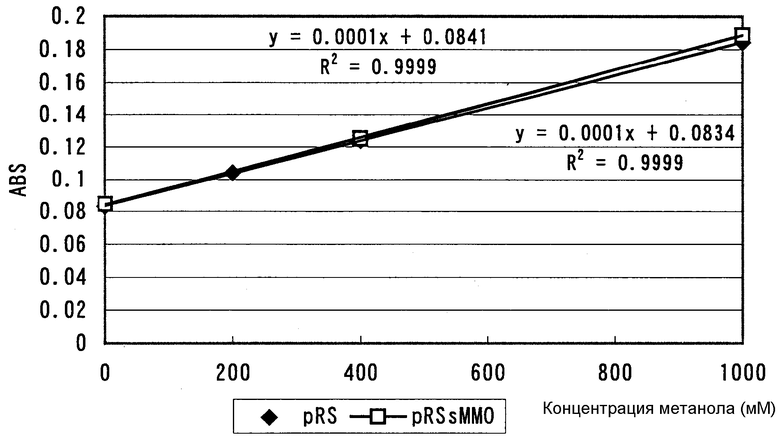
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| STAINTHORPE A.C | |||
| et al | |||
| The methane monooxygenase gene cluster of Methylococcus capsulatus (Bath) Gene, 1990, vol 91, no 1 p.27-34 | |||
| WEST CA et al | |||
| Functional expression in Escherichia coli of proteins B and С from soluble methane monooxygenase of Methylococcus capsulatus (Bath), Journal of General Microbiology, 1992, vol 138, p.1301-1307 | |||
| СПОСОБ ПОЛУЧЕНИЯ МЕТАНОЛА | 1993 |
|
RU2057745C1 |

