Область, к которой относится изобретение
Настоящее изобретение относится к кристаллическим структурам фосфодиэстеразы 5 (PDE5) и комплексов "PDE5/лиганд PDE5" и к их использованию для идентификации лигандов PDE5, включая соедиенния-ингибиторы PDE5. Настоящее изобретение также относится к способам идентификации указанных PDE5-ингибирующих соединений и к их использованию в медицине. В настоящем изобретении также рассматриваются кристаллы комплексов "PDE5/ингибитор PDE5".
На различные биологические процессы широкого ряда, включая сокращение сердечной мышцы, регуляцию кровообращения, передачу сигналов нейронами, гландулярную секрецию, дифференцировку клеток и экспрессию генов, влияют стационарные уровни циклических нуклеотидов, вторичных мессенджеров сАМР и cGMP. Внутриклеточными рецепторами этих молекул являются циклические нуклеотид-зависимые протеинкиназы (PGK) (Lohmann et al., 1997), каналы, открываемые циклическими нуклеотидами и фосфодиэстеразы класса I (PDE) (Charbonneau, 1990). PDE представляют собой большое семейство белков, о котором впервые сообщили Сазерленд (Sutherland) и сотрудники (Rall & Sutherland, Butcher & Sutherland, 1962). Ферменты семейства (циклические нуклеотиды)-фосфодиэстераз, катализируют гидролиз 3',5'-циклических нуклеотидов с образованием соответствующих 5'-монофосфатов. В современной литературе указывается, что существует одиннадцать родственных, но биохимически отличающихся групп, генов человеческих фосфодиэстераз, и что многие из этих групп включают более чем один подтип генов, состоящий, в целом, из двадцати генов. Некоторые из PDE являются в высокой степени специфичными в отношении гидролиза сАМР (PDE4, PDE7, PDE8), некоторые из них являются в высокой степени специфичными в отношении гидролиза cGMP (PDE5, PDE6, PDE9), а некоторые имеют смешанную специфичность (PDE1, PDE2, PDE3, PDE10, PDE11).
Все PDE представляют собой мультидоменные белки; при этом каждый PDE содержит домен примерно из 270 аминокислот, расположенный ближе к С-концу и имеющий в высокой степени консервативные аминокислотные последовательности для всех семейств (Charbonneau, 1986). Были проведены тщательные исследования этого домена, и было показано, что он этот домен является ответственным за общие каталитические функции (Francis, S.H. et al., 1994). Негомологичные сегменты остальной части белка обладают регуляторными функциями или наделяют этот белок специфическими свойствами связывания. Сообщалось, что PDE2, PDE5, PDE6 и PDE10 содержат предполагаемые домены GAF в своей регуляторной амино-концевой части (Aravind & Ponting 1997 & Soderling & Beavo 2000). Было показано, что эти домены GAF связываются с cGMP, однако их функция пока еще полностью не ясна. До сих пор считалось, что полноразмерные PDE млекопитающих характеризуются тем, что они образуют димеры в растворе, однако роль этой димерной структуры не ясна. Недавно была определена структура регуляторного сегмента PDE2А, связанного с cGMP, и был выявлен параллельный димер из четырех доменов GAF, причем, cGMP связывается только с одним из двух доменов GAF на каждом мономере (Martinez et al., 2001).
За последние годы было установлено, что PDE5, cGMP-специфический PDE, является важной терапевтической мишенью. Он состоит из консервативного С-концевого, цинк-содержащего каталитического домена, который катализирует расщепление cGMP, и N-концевой регуляторной части, которая содержит два повтора домена GAF. Каждый домен GAF содержит cGMP-связывающий сайт, один из которых обладает высокой аффинностью, а другой обладает более низкой аффинностью. Активность PDE5 регулируется посредством связывания cGMP с cGMP-связывающими сайтами с высокой и низкой степенью аффинности, и последующего фосфорилирования, которое происходит только в том случае, если оба эти сайта являются занятыми (Thomas et al., 1990). PDE5 присутствует в различных концентрациях в различных тканях, включая тромбоциты, гладкие мышцы сосудов и внутренних органов, и скелетную мышцу. Этот белок является ключевым регулятором уровней cGMP в гладкой мышце кавернозной ткани полового члена. Физиологический механизм эрекции заключается в высвобождении окиси азота (NO) в пещеристом теле во время полового возбуждения. Затем NO активирует фермент гуанилат-циклазу, что приводит к увеличению уровней cGMP, к релаксации гладкой мышцы в пещеристом теле и к притоку крови. Ингибирование PDE5 предотвращает разложение cGMP, что приводит к сохранению уровней cGMP, а следовательно, и к релаксации гладкой мышцы (Corbin & Francis, 1999). Силденафил (Sildenafil) (UK-092480), являющийся активным ингредиентом виагры (Viagra®) и сильным ингибитором PDE5, представляет особый интерес для эффективного лечения нарушения эрекции у мужчин.
Недавно были получены структурные данные для каталитического домена PDE4b, то есть сАМР-специфического PDE (Xu et al., 2000). Эта структура дает информацию о всей укладке каталитического домена белков семейства PDE, но до настоящего времени отсутствовали какие-либо структурные данные, указывающие на механизм связывания потенциальных ингибиторов с ферментом.
Была определена кристаллографическая структура рекомбинантной конструкции PDE5, содержащей каталитический домен в комплексе с силденафилом. Методами генной инженерии была также получена конструкция, которая давала более высокое качество при продуцировании кристаллов комплексов PDE5/ингибитор. Этот белок был также использован для расшифровки его структуры, связанной с силденафилом. Эти комплексы не только позволяют получить важную информацию о структуре этого нового семейства белков, но также позволяют сконструировать более сильные и специфические ингибиторы для лечения многих заболеваний, в которых определенную роль играют PDE.
Краткое описание изобретения
Было обнаружено, что PDE5 может кристаллизоваться. Было также обнаружено, что модификация аминокислотной последовательности PDE5 дикого типа может стимулировать кристаллизацию PDE5. В частности, было обнаружено, что модификация некоторых частей аминокислотной последовательности PDE5 дикого типа может стимулировать кристаллизацию PDE5.
Было показано, что модификация каталитического домена PDE5, а в частности области 657-682 PDE5 ("области петли"), может стимулировать кристаллизацию PDE5. Более конкретно, модификация аминокислотной последовательности области петли (HRGVNNSYIQRSEHPLAQLYCHSIME = SEQ ID NO:1) PDE5 может стимулировать кристаллизацию PDE5. Эта модификация может быть осуществлена посредством делеции, добавления или замены одного или более аминокислотных остатков "области петли" PDE5, либо она может быть осуществлена путем полной замены области петли PDE5 на область петли (или другую эквивалентную аминокислотную последовательность, например, субдомен), происходящую от другого белка, предпочтительно от другого PDE, более предпочтительно от PDE4, а наиболее предпочтительно от PDE4b.
Было обнаружено, что кристаллы PDE5 могут быть использованы для скрининга лигандов PDE5, а в частности ингибиторов PDE5 (например, путем совместной кристаллизации PDE5 с лигандом PDE5 (например, с ингибитором PDE5), либо путем погружения лиганда PDE5 (например, ингибитора PDE5) в кристалл PDE5).
Лиганды PDE5, а в частности, ингибиторы PDE5, идентифицированные способами настоящего изобретения, могут быть использованы для лечебной, паллиативной или профилактической терапии.
Таким образом, настоящее изобретение относится к следующим (пронумерованным) аспектам, таким как:
1. Кристалл фосфодиэстеразы 5 (PDE5).
2. Кристалл PDE5, в соответствии с аспектом 1, где указанный PDE5 происходит от млекопитающих.
3. Кристалл PDE5, в соответствии с аспектом 1 или с аспектом 2, где указанный PDE5 происходит от человека.
4. Кристалл PDE5, в соответствии с любым из аспектов 1-3, где указанный PDE5 представляет собой изоформу, выбранную из группы, состоящей из PDE5А1, PDE5А2, PDE5А3 и PDE5А4.
5. Кристалл PDE5, в соответствии с аспектом 3 или с аспектом 4, где указанный PDE5 содержит SEQ ID NO:1 или его гомолог, фрагмент, вариант, аналог или производное.
SEQ ID NO:1 представляет собой так называемую "область петли" PDE5. Эта область петли или ее гомолог, фрагмент, вариант, аналог или производное включает добавления, делеции или замены аминокислотных остатков, присутствующих в области петли.
Вариант, относящийся к аминокислотной последовательности кристалла PDE5 настоящего изобретения, предпочтительно, включает делецию или замену гистидинового (His/Н) остатка, показанного жирным шрифтом и подчеркнутого в SEQ ID NO:1 (HRGVNNSYIQRSEHPLAQLYCHSIME). Этот гистидин координирует атом цинка в PDE5 дикого типа. Замену указанного гистидинового (Н) остатка, предпочтительно, осуществляют путем введения одного или более аминокислотных остатков (но не гистидиновых), где указанные аминокислотные остатки, предпочтительно, являются нейтральными или неполярными.
Более предпочтительно, вариант, относящийся к аминокислотной последовательности кристалла PDE5 настоящего изобретения, включает область петли, полностью замененную на область петли (или другую аминокислотную последовательность, например, эквивалентный субдомен), происходящую от другого белка, предпочтительно от PDE, более предпочтительно от PDE4, а наиболее предпочтительно от PDE4b (см. ниже).
Альтернативно, вариант, относящийся к аминокислотной последовательности кристалла PDE5 настоящего изобретения, включает делецию или замену аминокислотных остатков PLAQ (пролина, лейцина, аланина и глутамина), показанных жирным шрифтом и подчеркнутых в SEQ ID NO:1 (HRGVNNSYIQRSEHPLAQLYCHSIME). Аминокислотная последовательность PLAQ представляет собой сайт протеолитического расщепления PDE5. Путем модификации этого сайта, например посредством делеции и/или замены одного или более аминокислотных остатков, может быть предотвращено нежелательное протеолитическое расщепление PDE5 или снижен его уровень. Предпочтительно, такую замену аминокислотных остатков осуществляют с использованием аминокислот с зарядом, аналогичным заряду замененных аминокислот.
Модификации "области петли" PDE5 могут быть осуществлены в соответствии с настоящим изобретением в целях стабилизации данной области. Аналогичные модификации могут быть осуществлены в PDE5-родственных белках, в других белках PDE и в PDE-родственных белках в целях стабилизации указанных белков.
Таким образом, настоящее изобретение относится к (пронумерованным) аспектам, таким как:
6. Кристалл PDE5 в соответствии с любым из аспектов 3-5, где указанный PDE5 содержит SEQ ID NO:2 или его гомолог, фрагмент, вариант, аналог или производное. Предпочтительно, указанный PDE5 состоит из SEQ ID NO:2 или его гомолога, фрагмента, варианта, аналога или производного.
7. Кристалл PDE5 в соответствии с любым из аспектов 3-6, где указанный PDE5 содержит SEQ ID NO:3 или его гомолог, фрагмент, вариант, аналог или производное. Предпочтительно, указанный PDE5 состоит из SEQ ID NO:3 или его гомолог, фрагмент, вариант, аналог или производное.
8. Кристалл PDE5 в соответствии с любым аспектом 3 или аспектом 4, где указанный PDE5 содержит SEQ ID NO:4 или его гомолог, фрагмент, вариант, аналог или производное.
SEQ ID NO:4 представляет собой так называемую "область петли" (или субдомен) PDE4 (PDE4b). Эта область петли или ее гомолог, фрагмент, вариант, аналог или производное включает добавления, делеции или замены аминокислотных остатков, присутствующих в области петли.
9. Кристалл PDE5 в соответствии с любым из аспектов 3, 4 или 8, где указанный PDE5 содержит SEQ ID NO:5 или его гомолог, фрагмент, вариант, аналог или производное. Предпочтительно, указанный PDE5 состоит из SEQ ID NO:5 или его гомолога, фрагмента, варианта, аналога или производного.
10. Кристалл PDE5 в соответствии с любым из аспектов 3, 4, 8 или 9, где указанный PDE5 содержит SEQ ID NO:6 или его гомолог, фрагмент, вариант, аналог или производное. Предпочтительно, указанный PDE5 состоит из SEQ ID NO:6 или его гомолога, фрагмента, варианта, аналога или производного.
11. Кристалл комплекса PDE5/лиганд PDE5.
12. Кристалл комплекса PDE5/лиганд PDE5 в соответствии с аспектом 11, где указанным лигандом PDE5 является ингибитор PDE5.
13. Кристалл комплекса PDE5/лиганд PDE5 в соответствии с аспектом 12, где указанным ингибитором лиганда PDE5 является силденафил.
14. Кристалл комплекса PDE5/лиганд PDE5 в соответствии с любым из аспектов 11-13, где указанный лиганд PDE5 определен в любом из аспектов 1-10.
15. Кристалл комплекса PDE5/лиганд PDE5 в соответствии с любым из аспектов 11-13, где указанный лиганд PDE5 определен в любом из аспектов 5-7.
16. Кристалл комплекса PDE5/лиганд PDE5 в соответствии с любым из аспектов 11-13, где указанный лиганд PDE5 определен в любом из аспектов 8-10.
17. Кристалл PDE5 в соответствии с любым из аспектов 1-10, или кристалл комплекса PDE5/лиганд PDE5 в соответствии с любым из аспектов 11-16, выращенный в растворе, содержащем буфер и/или преципитирующий агент.
18. Кристалл PDE5 в соответствии с любым из аспектов 5-7, выращенный в растворе, содержащем буфер и/или фосфат.
19. Кристалл PDE5 в соответствии с аспектом 18, где указанным фосфатным буфером является фосфат натрия/калия, фосфат натрия или фосфат аммония. Предпочтительно, указанный фосфатный буфер представляет собой 1,8-2,3 М фосфат натрия, рН 3,4-5,0, и содержит или не содержит 0,1 М Hepes, рН 7,0-8,0, либо он представляет собой 1,8-2,3 М фосфат натрия/калия, рН 3,4-5,0, и содержит или не содержит 0,1 М Hepes, рН 7,0-8,0.
20. Кристалл PDE5 в соответствии с любым из аспектов 8-10, или кристалл комплекса PDE5/лиганд PDE5 в соответствии с любым из аспектов 11-16, выращенный в растворе, содержащем:
(i) Трис- или MES-буфер, фосфат аммония и/или PEG2KMME. Предпочтительно, указанный трис- или MES-буфер имеет рН 6,0-8,4. Более предпочтительно, указанный раствор содержит 0,1 М трис, рН 8,0, 50 мМ фосфат аммония, рН 7,0; 16-26% мас./об. PEG2KMME. Альтернативно, указанный раствор содержит 0,1 М MES, рН 6,0-6,5, 50 мМ фосфат аммония, рН 7,5; 22-34% мас./об. PEG2KMME; или
(ii) 0,16 М ацетат натрия, 80 мМ трис-гидрохлорид, рН 8,5, 24% мас./об. полиэтиленгликоля 8000 (или PEG8KMME).
21. Кристалл PDE5 в соответствии с любым из аспектов 8-10, или кристалл комплекса PDE5/лиганд PDE5 в соответствии с аспектом 16, выращенные в растворе, содержащем:
(i) Трис-буфер, ацетат натрия и/или PEG4K. Предпочтительно, указанный трис-буфер имеет рН 6,5-8,6. Альтернативно, указанный трис-буфер имеет рН 8,2-8,6. Более предпочтительно, указанный раствор содержит 0,1 М трис, рН 8,2-8,6, 0,2М ацетат натрия и 26-30% мас./об. PEG4K; или
(ii) 0,16 М ацетат натрия, 80 мМ трис-гидрохлорид, рН 8,5, 24% мас./об. полиэтиленгликоля 8000 (или PEG8KMME).
22. Кристалл PDE5, определенный в любом из аспектов 5-7 и имеющий одну или более из нижеследующих характеристик:
(а) спейсерную группу Р62;
(b) размеры элементарной ячейки а˜95 ű1%, b˜95 ű1%, c˜82 ű1%, α=β=90°, γ=120°;
(с) 1 молекулу на один асимметричный элемент:
(d) содержит PDE5 с молекулярной массой приблизительно 40 кДа ± 2 кДа;
(е) вычисленное содержание растворителя приблизительно 43±5%; и
(f) гексагональную кристаллическую систему.
23. Кристалл комплекса PDE5/лиганд PDE5 в соответствии с аспектом 15, имеющий одну или более из нижеследующих характеристик:
(а) спейсерную группу Р212121;
(b) размеры элементарной ячейки а˜94 ű1%, b˜104 ű1%, c˜142 ű1%, α=β=γ=90°;
(с) 4 молекулы на один асимметричный элемент:
(d) содержит PDE5 с молекулярной массой приблизительно 40 кДа ± 2 кДа;
(е) вычисленное содержание растворителя составляет приблизительно 43±5%; и
(f) орторомбическую кристаллическую систему.
24. Кристалл PDE5, определенный в соответствии с одним из аспектов 8-10, или кристалл комплекса PDE5/лиганд PDE5 в соответствии с аспектом 16, имеющий одну или более из нижеследующих характеристик:
(а) спейсерную группу Р21;
(b) размеры элементарной ячейки а˜55 ű1%, b˜78 ű1%, c˜82 ű1%, α=γ=90°, β˜101°±2°;
(с) 2 молекулы на один асимметричный элемент:
(d) содержит PDE5 с молекулярной массой приблизительно 38 кДа ± 2 кДа;
(е) вычисленное содержание растворителя составляет приблизительно 46±5%; и
(f) моноклинную кристаллическую систему.
25. Кристалл PDE5 в соответствии с одним из аспектов 5-7, где указанный PDE5 имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 3, или производное, выраженное в любой системе отсчета.
26. Кристалл комплекса PDE5/лиганд PDE5 в соответствии с аспектом 15, где указанный комплекс PDE5/лиганд PDE5 имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 4, или производное, выраженное в любой системе отсчета.
27. Кристалл PDE5 в соответствии с одним из аспектов 8-10, где указанный PDE5 имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 5, или производное, выраженное в любой системе отсчета.
28. Кристалл комплекса PDE5/лиганд PDE5 в соответствии с аспектом 16, где указанный комплекс PDE5/лиганд PDE5 имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 6, или производное, выраженное в любой системе отсчета.
29. Использование атомных координат, определенных исходя из кристалла PDE5 в соответствии с аспектом 25 или из кристалла комплекса PDE5/лиганд PDE5 в соответствии с аспектом 26, для определения трехмерной структуры (i) полноразмерного PDE5 дикого типа или его мутанта, производного, фрагмента, варианта, аналога или гомолога, или (ii) субдомена PDE5 дикого типа или его мутанта, производного, фрагмента, варианта, аналога или гомолога.
30. Использование в соответствии с аспектом 29, где указанный субдомен PDE5 представляет собой каталитический домен.
31. Использование атомных координат, определенных для кристалла PDE5 в соответствии с аспектом 27 или кристалла комплекса PDE5/лиганд PDE5 в соответствии с аспектом 28, для определения трехмерной структуры (i) полноразмерного PDE5 дикого типа или его мутанта, производного, фрагмента, варианта, аналога или гомолога, или (ii) субдомена PDE5 дикого типа или его мутанта, производного, фрагмента, варианта, аналога или гомолога.
32. Использование в соответствии с аспектом 31, где указанный субдомен PDE5 представляет собой каталитический домен.
33. Использование трехмерной структуры (i) полноразмерного PDE5 дикого типа или его мутанта, производного, фрагмента, варианта, аналога или гомолога, или (ii) субдомена PDE5 дикого типа или его мутанта, производного, фрагмента, варианта, аналога или гомолога, определенных в соответствии с любым из аспектов 29, 30, 31 или 32 для компьютерной оценки или оценки каким-либо иным способом, взаимодействий связывания лиганда PDE5 с активным сайтом на PDE5.
34. Использование в соответствии с аспектом 33, где указанный лиганд PDE5 представляет собой ингибитор PDE5.
35. Использование в соответствии с аспектом 34, где указанным ингибитором PDE5 является Силденафил.
36. Использование по любому из аспектов 33-35, где указанный активный центр на PDE5 находится в третьем субдомене белка и ограничен спиралями 15 (Н15 813-824) и 14 (Н14 772-797), С-концом спирали 13 (Н13 749-765) и С-концом спирали 11 (Н11 706-721) вместе с областью петли, расположенной между спиралями 11 и 12а (Н12а 725-731), как показано на фигуре 2.
37. Использование по любому из аспектов 33-36, где указанный активный сайт на PDE5 содержит Leu 765, Ala 767 и Ile 768 и один или более Phe 820, Val 782, Phe 786, Tyr 612, Leu 804, Ala 779, Ala 783, Ile 813, Met 816 и Gln 817.
38. Использование по любому из аспектов 33-37 для конструирования соединения, способного связываться с PDE5.
39. Использование по любому из аспектов 33-38 для конструирования соединения, способного связываться с любым активным сайтом PDE5.
40. Использование в соответствии с аспектом 38 или с аспектом 39, где указанным соединением является лиганд PDE5.
41. Использование в соответствии с аспектом 40, где указанным лигандом PDE5 является ингибитор PDE5.
42. Способ идентификации соединения, способного связываться с PDE5, включающий совместную кристаллизацию или погружение указанного соединения в кристалл PDE5 в соответствии с одним из аспектов 1-10 и определение трехмерной структуры в целях установления факта связывания указанного соединения с PDE5. Что касается термина "погружение", следует отметить, что соединение может быть добавлено к кристаллу, и таким образом, соединение погружается в этот кристалл. Альтернативно, кристалл может быть добавлен к соединению (например, в растворе), и в этом случае соединение погружается в кристалл.
43. Способ идентификации соединения, способного связываться с любым активным сайтом PDE5, включающий совместную кристаллизацию или погружение указанного соединения в кристалл PDE5 в соответствии с одним из аспектов 1-10 и определение трехмерной структуры в целях установления факта связывания указанного соединения с PDE5.
44. Соединение, сконструированное с использованием, описанным в любом из аспектов 33-41, или идентифицированное способом, описанным в аспекте 42 или в аспекте 43.
45. Соединение в соответствии с аспектом 44, которое является ингибитором PDE5.
46. Способ отбора лиганда PDE5 из группы возможных лигандов PDE5, включающий следующие стадии:
(а) компьютерного моделирования трехмерного представления структуры PDE5, выведенной исходя из атомных координат, определенных в соответствии с любым из аспектов 29, 30, 31 или 32, и трехмерного представления структуры потенциального лиганда PDE5;
(b) совместного отображения трехмерного представления потенциального лиганда PDE5 вместе с трехмерным представлением структуры PDE5; и
(с) установления факта соответствия трехмерного представления потенциального лиганда PDE5 и трехмерного представления активного сайта структуры PDE5.
47. Способ в соответствии с аспектом 46, дополнително включающий следующие стадии:
(d) включения потенциального лиганда PDE5 в анализе на биологическую активность PDE5; и
(е) установления факта модуляции активности PDE5 потенциальным лигандом PDE5 в указанном анализе.
48. Способ в соответствии с аспектом 46 или аспектом 47, где указанным потенциальным лигандом PDE5 является потенциальное соединение-ингибитор PDE5, и указанное потенциальное соединение-ингибитор PDE5 ингибирует активность PDE5.
49. Лиганд PDE5, выбранный способом, определенным в аспекте 46 или в аспекте 47, или соединение-ингибитор PDE5, выбранное способом, определенным в аспекте 48.
50. Фармацевтическая композиция, содержащая один или более лигандов PDE5 или соединений-ингибиторов PDE5 в соответствии с аспектом 49 и один или более фармацевтически приемлемых наполнителей.
51. Использование лиганда PDE5 или соединения-ингибитора PDE5 в соответствии с аспектом 49 в качестве фармацевтического средства.
52. Использование лиганда PDE5 или соединения-ингибитора PDE5 в соответствии с аспектом 49 для производства лекарственного средства для профилактики или лечения состояния, заболевания, расстройства или дисфункции, в случае, когда ингибирование PDE5 оказывает профилактически или терапевтически благоприятное действие.
53. Использование в соответствии с аспектом 52, где указанным расстройством является сексуальное расстройство млекопитающего.
Лечебная, паллиативная или профилактическая терапия, рассматриваемая в настоящем изобретении, включает лечебную, паллиативную или профилактическую терапию сексуальных расстройств у млекопитающих, в частности лечение половых расстройств у млекопитающих, таких как нарушение эрекции у мужчин (MED), импотенция, сексуальная дисфункция у женщин (FSD), дисфункция клитора, расстройство, связанное с пониженным сексуальным влечением у женщин, расстройство, связанное с половым возбуждением у женщин (FSAD), расстройство, связанное с возникновением боли во время полового акта у женщин, или нарушение наступления оргазма у женщин (FSOD), а также сексуальная дисфункция, вызванная поражением спинного мозга, или сексуальная дисфункция, индуцированная ингибитором селективного обратного захвата серотонина, но, как очевидно, такая терапия может быть использована и для лечения других клинических состояний, при которых показан ингибитор PDE5. Такими состояниями являются преждевременные роды, дисменорея, доброкачественная гиперплазия предстательной железы (ВРН), обструкция отверстия мочевого пузыря, недержание мочи, стабильная, нестабильная и вариантная стенокардия (болезнь Принцметала), гипертензия, легочная гипертензия, хроническое обструктивное заболевание легких, ишемическая болезнь сердца, застойная сердечная недостаточность, атеросклероз, состояния, связанные с пониженной проходимостью кровеносных сосудов, например, пост-чрезкожная катетерная коронарная ангиопластика (пост-РТСА), заболевания периферических сосудов, инсульт, индуцированная нитратом толерантность, бронхит, аллергическая астма, хроническая астма, аллергический ринит, глазные болезни и связанные с ними расстройства, такие как глаукома, невропатия зрительного нерва, дегенерация пятна, повышенное внутриглазное давление, окклюзия сетчатки и артерий глаза и заболевания, характеризующиеся нарушениями кишечной перистальтики, например синдром раздражения кишечника (СРК).
Другими клиническими состояниями, для которых показан ингибитор PDE5 и к которым может быть применено лечение соединениями настоящего изобретения, являются преэклампсия, синдром Кавазаки, рассеянный склероз, диабетическая нефропатия, невропатия, включая автономную и периферическую невропатию, а в частности, диабетическую невропатию и ее симптомы, гастропарез, периферическая диабетическая невропатия, болезнь Альцгеймера, острая респираторная недостаточность, псориаз, некроз кожи, рак, метастазы, облысение, эзофагит типа «щелкунчик», трещина заднего прохода, геморрой, синдром инсулинорезистентности, диабет, вазоконстрикция при гипоксии, а также стабилизация кровяного давления во время гемодиализа.
Особенно предпочтительными состояниями являются MED и FSD (предпочтительно, FSAD).
Другими (пронумерованными) аспектами настоящего изобретения являются:
54. Использование атомных координат, определенных для кристалла PDE5 в соответствии с аспектом 25 или аспектом 27, или для кристалла комплекса PDE5/лиганд PDE5 в соответствии с аспектом 26 или аспектом 28, для определения кристаллической структуры мутанта, производного, фрагмента, варианта, аналога, гомолога или комплекса PDE-родственного белка.
55. Использование в соответствии с аспектом 54, где указанным PDE-родственным белком является PDE.
56. Использование в соответствии с аспектом 55, где указанным PDE является PDE-родственный белок.
57. Использование в соответствии с аспектом 56, где указанным PDE5-родственным белком является PDE5.
58. Использование атомных координат, определенных для кристалла PDE5 в соответствии с аспектом 25 или аспектом 27, или кристалла комплекса PDE5/лиганд PDE5 в соответствии с аспектом 26 или аспектом 28, для получения модели трехмерной структуры PDE-родственного белка.
59. Использование в соответствии с аспектом 58, где указанным PDE-родственным белком является PDE.
60. Использование в соответствии с аспектом 59, где указанным PDE является PDE5-родственный белок.
61. Использование в соответствии с аспектом 60, где указанным PDE5-родственным белком является PDE5.
62. Использование трехмерной структуры PDE5 в соответствии с любым из аспектов 29, 30, 31 или 32 для сайт-направленного конструирования мутантов, имитирующих другие изоформы PDE5 или его варианты.
63. Кристалл PDE5 или кристалл комплекса PDE5/лиганд PDE5, где указанный активный сайт на PDE5 находится в третьем субдомене белка и ограничен спиралями 15 (Н15 813-824) и 14 (Н14 772-797), С-концом спирали 13 (Н13 749-765) и С-концом спирали 11 (Н11 706-721) вместе с областью петли, расположенной между спиралями 11 и 12а (Н12а 725-731), как показано на фигуре 2.
64. Кристалл PDE5 или кристалл комплекса PDE5/лиганд PDE5, где указанный активный сайт на PDE5 содержит Leu 765, Ala 767 и Ile 768 и один или более Phe 820, Val 782, Phe 786, Tyr 612, Leu 804, Ala 779, Ala 783, Ile 813, Met 816 и Gln 817.
65. Кристалл PDE5, где кристаллическая система указанного кристалла характеризуется тем, что она является моноклинной, орторомбической или гексагональной.
66. Кристалл комплекса PDE5/лиганд PDE5, где кристаллическая система указанного кристалла характеризуется тем, что она является моноклинной или орторомбической.
67. Способ продуцирования структурно стабилизированного PDE-родственного белка, включающий:
(а) сопоставление аминокислотной последовательности PDE-родственного белка с аминокислотной последовательностью (i) PDE4, (ii) каталитического домена PDE4, показанного на фигуре 1, или (iii) SEQ ID NO:4;
(b) идентификацию структурно-эквивалентного субдомена PDE-родственного белка, который соответствует SEQ ID NO:4; и
(с) осуществление методами генной инженерии замены структурно-эквивалентного субдомена PDE-родственного белка или его части последовательностью SEQ ID NO:4 или ее гомологом, фрагментом, вариантом, аналогом или производным.
68. Способ в соответствии с аспектом 67, дополнительно включающий:
(d) экспрессию сконструированного PDE-родственного белка со стадии (с), в клетке-хозяине.
69. Способ в соответствии с аспектом 68, дополнительно включающий:
(е) очистку экспрессированного сконструированного PDE-родственного белка со стадии (d).
70. Способ в соответствии с аспектом 69, дополнительно включающий:
(f) кристаллизацию очищенного сконструированного PDE-родственного белка со стадии (е).
71. Способ в соответствии с любым из аспектов 67-70, где указанным PDE-родственным белком является PDE.
72. Способ в соответствии с аспектом 71, где указанным PDE является PDE5-родственный белок.
73. Способ в соответствии с аспектом 72, где указанным PDE5-родственным белком является PDE5.
74. Способ в соответствии с аспектом 73, где указанный PDE5 определен в любом из аспектов 5-7.
Соединение настоящего изобретения (то есть соединение в соответствии с аспектом 44 или аспектом 45, или лиганд PDE5, или соединение-ингибитор PDE5 в соответствии с аспектом 49, называемые далее одним общим термином "соединение") может быть введено отдельно, но при лечении человека его обычно вводят в смеси с подходящим фармацевтическим наполнителем, разбавителем или носителем, выбранным в соответствии с нужным способом введения и в соответствии со стандартной фармацевтической практикой. Таким образом, фармацевтические композиции, фармацевтические средства и лекарственные средства, рассматриваемые в настоящем изобретении, могут быть приготовлены и введены различными методами, хорошо известными специалистам.
Так, например, для быстрого, замедленного, модифицированного или регулируемого высвобождения, такого как пролонгированная, двухразовая или периодическая доставка соединения настоящего изобретения, это соединение может быть введено перорально, трансбуккально или подъязычно в форме таблеток, капсул (включая мягкие гелевые капсулы), овулей, эликсиров, растворов или суспензий, которые могут содержать отдушки или подкрашивающие агенты. Соединение может быть также введено посредством инъекции вовнутрь пещеристого тела. Указанное соединение может быть также введено в виде быстро диспергирующихся или быстро растворяющихся лекарственных форм, либо в виде высокоэнергетической дисперсии, либо в виде частиц с оболочкой. Подходящие фармацевтические препараты данного соединения, при необходимости, могут иметь или не иметь оболочки.
Указанные таблетки могут содержать наполнители, такие как микрокристаллическая целлюлоза, лактоза, цитрат натрия, карбонат кальция, двухосновный фосфат кальция, глицин и крахмал (предпочтительно, кукурузный крахмал, картофельный крахмал или крахмал из тапиоки), дезинтегрирующие агенты, такие как натрийсодержащий гликолят крахмала, натрийсодержащая кроскармилоза и некоторые комплексные силикаты, и связующие агенты для грануляции, такие как поливинилпирролидон, гидроксипропилметилцеллюлоза (НРМС), гидроксипропилцеллюлоза (НРС), сахароза, желатин и аравийская камедь. Кроме того, могут быть включены замасливающие агенты, такие как стеарат магния, стеариновая кислота, глицерилбегенат и тальк.
Твердые композиции аналогичного типа могут быть также использованы в качестве наполнителей в желатиновых капсулах. При этом предпочтительными наполнителями являются лактоза, крахмал, целлюлоза, молочный сахар и высокомолекулярные полиэтиленгликоли. Для водных суспензий и/или эликсиров, данное соединение может быть объединено с различными подслащивающими или ароматизирующими агентами, с подкрашивающим веществом или красителями, с эмульгирующими и/или суспендирующими агентами и с разбавителями, такими как вода, этанол, пропиленгликоль и глицерин и их комбинации.
Лекарственные формы с модифицированным и периодическим высвобождением могут содержать наполнители, такие как наполнители, конкретно указанные при описании лекарственных форм быстрого высвобождения, вместе с дополнительными наполнителями, которые действуют как модификаторы скорости высвобождения и которые могут быть нанесены на корпус устройства и/или включены в него. Модификаторами скорости высвобождения являются, но не ограничиваются исключительно ими, гидроксипропилметилцеллюлоза, метилцеллюлоза, натрийсодержащая карбоксиметилцеллюлоза, этилцеллюлоза, ацетат целлюлозы, полиэтиленоксид, ксантановая камедь, карбомер, сополимер аммиака и метакрилата, гидрогенизированное касторовое масло, воск карнаубы, парафиновый воск, ацетат-фталат целлюлозы, фталат гидроксипропилметилцеллюлозы, сополимер метакриловой кислоты и их смеси. Лекарственные формы с модифицированным и периодическим высвобождением могут содержать один наполнитель, модифицирующий скорость высвобождения, или комбинацию таких наполнителей. Наполнители, модифицирующие скорость высвобождения, могут присутствовать в лекарственной форме, то есть в матриксе, и/или на лекарственной форме, то есть на ее поверхности или оболочке.
Быстро диспергирующиеся или растворяющиеся лекарственные препараты (FDDF) могут содержать следующие ингредиенты: аспартам, ацесульфам-калий, лимонную кислоту, натрийсодержащую кроскармелозу, кросповидон, двухосновную аскорбиновую кислоту, этилакрилат, этилцеллюлозу, желатин, гидроксипропиметилцеллюлозу, стеарат магния, маннит, метилметакрилат, мятную отдушку, полиэтиленгликоль, дымящую двуокись кремния, двуокись кремния, натрийсодержащий гликолят крахмала, стеарилфумарат натрия, сорбит, ксилит. Термины "диспергирующийся" или "растворяющийся", используемые здесь для описания FDDF, применяются в зависимости от степени растворимости используемого лекарственного вещества, то есть, если данное лекарственное вещество является нерастворимым, то на его основе может быть получена быстро диспергирующаяся лекарственная форма, а если данное лекарственное вещество является растворимым, то на его основе может быть получена быстро растворяющаяся лекарственная форма.
Соединение может быть также введено парентерально, например вовнутрь пещеристого тела, внутривенно, внутриартериально, внутрибрюшинно, интратекально, интравентрикулярно, интрауретрально, интрастернально, интракраниально, внутримышечно или подкожно, либо оно может быть введено путем вливания или путем инъекции с использованием безыгольного шприца. Для такого парентерального введения наиболее подходящей формой является стерильный водный раствор, который может содержать другие вещества, например достаточное количество соли или глюкозы, которые придают такому раствору изотоничность к крови. Если это необходимо, то водные растворы должны быть соответствующим образом забуферены (предпочтительно до рН от 3 до 9). Получение подходящих парентеральных композиций в стерильных условиях может быть легко осуществлено стандартными фармацевтическими методами, хорошо известными специалистам.
Для перорального и парентерального введения человеку ежедневная доза такого соединения обычно составляет от 10 до 500 мг (в виде разовой дозы или дробных доз).
Так, например, таблетки или капсулы данного соединения могут содержать от 5 мг до 250 мг активного соединения и могут быть введены один раз, два раза или более раз через определенные интервалы времени, когда это необходимо. В любом случае, врач может самостоятельно определить фактическую дозу, которая является наиболее подходящей для данного конкретного пациента, и эта доза будет варьироваться в зависимости от возраста, веса и восприимчивости конкретного пациента. Вышеуказанные дозы даны в качестве примеров для среднего пациента. Само собой разумеется, что отдельные случаи, где могут оказаться подходящими интервалы с более высокими или более низкими дозами, также входят в объем настоящего изобретения. Для каждого специалиста очевидно, что для лечения некоторых состояний (включая MED и FSD), соединение может быть введено в виде разовой дозы, определенной исходя из "потребностей" (то есть необходимости или желательности).
Соединение может быть также введено интраназально, или путем ингаляции, и его доставку обычно осуществляют с помощью ингалятора для сухого порошка (инсуффлятора) или аэрозольного спрея, подаваемого из аэрозоля под давлением, насоса, распылителя или аэрозольного ингалятора с использованием подходящего пропеллента, например дихлордифторметана, трихлорфторметана, дихлортетрафторэтана, гидрофторалкана, такого как 1,1,1,2-тетрафторэтан (HFA 134ATM) или 1,1,1,2,3,3,3-гепта-фторпропан (HFA 227EATM), диоксида углерода или другого подходящего газа. В случае использования аэрозоля под давлением разовая лекарственная форма может быть введена посредством дозирующего клапана для доставки. Аэрозоль под давлением, насос, спрей или аэрозольный ингалятор могут содержать раствор или суспензию активного соединения, например смеси этанола и пропеллента в качестве растворителя, который может дополнительно содержать замасливатель, например сорбитантриолеат. Капсулы и картриджи (изготовленные, например, из желатина), предназначенные для использования в ингаляторе или в инсуффляторе, могут быть приготовлены так, чтобы они содержали порошкообразную смесь соединения настоящего изобретения и подходящую порошкообразную основу, такую как лактоза или крахмал.
Аэрозольные препараты или препараты в виде сухого порошка, предпочтительно, приготавливают так, чтобы каждая отмеренная доза или "впрыск" содержали от 1 до 50 мг соединения настоящего изобретения, предназначенного для введения пациенту. Общая суточная доза, доставляемая в виде аэрозоля, может составлять в пределах от 1 до 50 мг и может быть введена в виде разовой дозы или, более предпочтительно, в виде дробных доз, вводимых в течение дня.
Данное соединение может быть также приготовлено для доставки с помощью аэрозольного ингалятора. Композиции, вводимые с помощью аэрозольных ингаляторов, могут содержать такие ингредиенты, как солюбилизаторы, эмульгирующие или суспендирующие агенты: воду, этанол, глицерин, пропиленгликоль, низкомолекулярные полиэтиленгликоли, хлорид натрия, фторуглероды, эфиры полиэтиленгликоля, сорбитантриолеат, олеиновую кислоту.
Альтернативно, соединение может быть введено в форме суппозиториев или пессариев, либо оно может быть введено путем местного применения в виде геля, гидрогеля, лосьона, раствора, крема, мази или присыпки. Соединение может быть также нанесено на кожу. Соединение может быть также введено чрезкожно, например, с использованием кожного пластыря. Соединение может быть также введено офтальмически, через легкие или ректально.
Для офтальмического применения данное соединение может быть приготовлено в виде микросуспензий в изотоническом стерильном физиологическом растворе со скорректированным рН либо, предпочтительно, в виде растворов в изотоническом стерильном физиологическом растворе со скорректированным рН, необязательно, в комбинации с консервантом, таким как хлорид бензилалкония. Альтернативно, соединение может быть приготовлено в виде мази, такой как вазелиновая мазь.
Для местного нанесения на кожу соединение настоящего изобретения может быть приготовлено в виде подходящей мази, содержащей активное соединение, суспендированное или растворенное, например, в смеси с одним или более из нижеследующих компонентов, таких как минеральное масло, жидкое вазелиновое масло, белое вазелиновое масло, пропиленгликоль, полиоксиэтиленовое-полиоксипропиленовое соединение, эмульгирующийся воск и вода. Альтернативно, это соединение может быть приготовлено в виде подходящего лосьона или крема, в котором это соединение суспендиндировано или растворено, например, в смеси с одним или более ингредиентами, такими как минеральное масло, сорбитанмоностеарат, полиэтиленгликоль, жидкое вазелиновое масло, полисорбат 60, воск на основе цетиловых эфиров, цетеариловый спирт, 2-октилдодеканол, бензиловый спирт и вода.
Соединение может быть также использовано в комбинации с циклодекстрином. Известно, что циклодекстрины образуют включаемые и невключаемые комплексы с молекулами лекарственного средства. Образование комплекса "лекарственное средство-циклодекстрин" может приводить к изменению растворимости, скорости растворения, биологической доступности и/или стабильности молекулы лекарственного средства. В основном, комплексы "лекарственное средство-циклодекстрин" могут быть использованы во многих лекарственных формах и в способах введения. В качестве альтернативы прямому образованию комплекса с лекарственным средством, циклодекстрин может быть использован в качестве вспомогательной добавки, например, в качестве носителя, разбавителя или солюбилизатора. Широко используемые альфа-, бета- и гамма-циклодекстрины и подходящие примеры их использования описаны в WO-А-91/11172, WO-А-94/02518 и WO-А-98/55148.
Для человека, в основном, предпочтительным и более удобным является пероральное введение соединения, которое, например, в случае MED, позволяет избежать хорошо известных проблем, связанных с введением вовнутрь пещеристого тела (i.с.). Предпочтительной схемой порорального введения пациенту с типичным MED является введение 25-250 мг соединения, если это необходимо. В случаях, когда реципиент страдает нарушением глотания или нарушением абсорбции лекарственного средства после перорального введения, то такое лекарственное средство может быть введено парентерально, подъязычно или трансбуккально.
Для лечения животных соединение или его ветеринарно приемлемая соль или его ветеринарно приемлемый сольват или пролекарство могут быть введены в виде подходящего приемлемого препарата в соответствии с обычно ветеринарной практикой, и ветеринарный врач может самостоятельно определить схему и способ введения лекарственного средства, которые будут наиболее подходящими для данного конкретного животного.
Подробное описание изобретения
Используемый здесь термин "апо" относится к любому белку (или упомянутому белку), который отделяется от своего лиганда(ов) и/или простетической группы(групп).
Используемый здесь термин "активный сайт" означает любой сайт (например, специфические группы), который находится в молекуле (и связывается с ионами металла и/или гидратированными молекулами) и который обладает специфической активностью. Такой активностью может быть связывание лиганда с указанным сайтом, катализ связывания субстратов данных молекул с указанным сайтом, распознавание лиганда указанным сайтом и т.п.
Используемый здесь термин "буфер" означает любой раствор, содержащий слабую кислоту и конъюгат основания с этой кислотой (или, иногда, слабое основание и его конъюгат с кислотой). Таким образом, используемый здесь "буфер" обеспечивает сохранение рН при добавлении в него кислоты или основания, поскольку кислота нейтрализует добавленное основание (или, иногда, основание нейтрализует добавленную кислоту).
Используемый здесь термин "преципитирующий агент" означает любое вещество, которое при добавлении к раствору (обычно макромолекул) приводит к осаждению с образованием или ростом кристаллов.
Используемый здесь термин "комплекс" означает белок вместе со связанным с ним лигандом(ами) и может образовываться до, во время или после кристаллизации белка.
Используемый здесь термин "погружение" означает добавление раствора, содержащего (обычно) небольшую молекулу (например, ингибитор), к кристаллам белка с образованием комплекса белок-лиганд.
Используемый здесь термин "совместная кристаллизация" означает кристаллизацию предварительно образованного комплекса "белок/малая молекула".
Термины "мутант", "вариант", "гомолог", "аналог", "производное" или "фрагмент", используемые по отношению к аминокислотной последовательности кристалла PDE5 настоящего изобретения, включают любое замещение, изменение, модификацию, замену, делецию или добавление одной (или более) аминокислоты в последовательности при условии, что полученный PDE5 способен к кристаллизации.
Термины "мутант", "вариант", "гомолог", "аналог", "производное" или "фрагмент", используемые по отношению к нуклеотидной последовательности, кодирующей PDE5 кристалла PDE5 настоящего изобретения, включают любое замещение, изменение, модификацию, замену, делецию или добавление одной (или более) нуклеиновой кислоты в последовательности при условии, что полученная нуклеотидная последовательность кодирует или может кодировать PDE5, который способен к кристаллизации.
Вообще говоря, что касается "мутанта", "варианта", "гомолога", "аналога", "производного" или "фрагмента", используемых по отношению к аминокислотной последовательности кристалла PDE5 настоящего изобретения, то вводимые аминокислотные замены должны быть сделаны с использованием аминокислот таких типов, чтобы при этом сохранялась гидрофобность/гидрофильность аминокислотной последовательности. Аминокислотные замены могут быть сделаны, например, в 1, 2 или 3 - 10, 20 или 30 аминокислотах при условии, что модифицированный PDE5 будет сохранять способность к кристаллизации в соответствии с настоящим изобретением. Аминокислотные замены могут включать использование неприродных аналогов.
Что касается аминокислотных последовательностей, то используемый здесь термин "вариант" означает добавления, делеции или замены аминокислотных остатков, находящихся в аминокислотной последовательности дикого типа или в ее фрагменте. Предпочтительно, вариант, относящийся к аминокислотной последовательности кристалла PDE5 настоящего изобретения, должен включать делецию или замену гистидинового остатка (His/Н), показанного жирным шрифтом и подчеркнутого в SEQ ID NO:1 (HRGVNNSYIQRSEHPLAQLYCHSIME), где указанная последовательность находится в молекуле PDE5 кристалла PDE5 настоящего изобретения. Замену указанного гистидинового (Н) остатка осуществляют предпочтительно путем введения одного или более аминокислотных остатков (но не гистидиновых), где указанные аминокислотные остатки, предпочтительно, являются нейтральными или неполярными.
Более предпочтительно, вариант, относящийся к аминокислотной последовательности кристалла PDE5 настоящего изобретения, включает полную замену области петли на область петли (или другую эквивалентную аминокислотную последовательность, например субдомен), происходящий от другого белка, предпочтительно от PDE, более предпочтительно от PDE4, а наиболее предпочтительно от PDE4b.
Альтернативно, вариант, относящийся к аминокислотной последовательности кристалла PDE5 настоящего изобретения, включает делецию или замену аминокислотных остатков PLAQ (пролина, лейцина, аланина и глутамина), показанных жирным шрифтом и подчеркнутых в SEQ ID NO:1 (HRGVNNSYIQRSEHPLAQLYCHSIME). Предпочтительно, такую замену аминокислотных остатков осуществляют с использованием аминокислот с зарядом, аналогичным заряду замененных аминокислот.
Что касается нуклеотидных последовательностей, то используемый здесь термин "вариант" означает добавления, делеции или замены нуклеотидов, находящихся в нуклеотидной последовательности дикого типа или в ее фрагменте.
Используемый здесь термин "фрагмент" означает любую часть PDE5, определенную в настоящем изобретении, при условии, что полученный PDE5, содержащий указанную часть PDE5, способен кристаллизоваться. Таким образом, термин "фрагмент" также включает PDE5, которые содержит любую часть последовательности SEQ ID NO:1, 2, 3, 4, 5 или 6.
Так, например, специфическим фрагментом SEQ ID NO:3 (полноразмерной последовательности PDE5 дикого типа) настоящего изобретения может быть последовательность SEQ ID NO:2 (каталитический домен PDE5 дикого типа). Примером специфического фрагмента SEQ ID NO:2 (каталитического домена PDE5 дикого типа) настоящего изобретения может служить SEQ ID NO:1 ("область петли" PDE5; HRGVNNSYIQRSEHPLAQLYCHSIME).
Примером специфического фрагмента SEQ ID NO:6 (полноразмерной последовательности с "замененной петлей" PDE5 дикого типа) настоящего изобретения может быть последовательность SEQ ID NO:5 (каталитический домен PDE5 с "замененной петлей"). Кроме того, примером специфического фрагмента SEQ ID NO:5 (каталитического домена PDE5 с "замененной петлей") настоящего изобретения может служить SEQ ID NO:4 ("область петли" PDE4; HPGVSNQFLINTNSELALMYNDESVLE).
Используемый здесь термин "аналог" означает последовательность, которая аналогична аминокислотной последовательности кристалла PDE5 настоящего изобретения или любой из последовательностей SEQ ID NO:1, 2, 3, 4, 5 или 6, но в которой отсутствуют аминокислотные замены или делеции, оказывающие негативное действие (то есть негативное действие на способность PDE5 к кристаллизации).
Термин "производное", используемый здесь по отношению к аминокислотной последовательности кристалла PDE5 настоящего изобретения или к любой из последовательностей SEQ ID NO:1, 2, 3, 4, 5 или 6, означает PDE5, который включает химическую модификацию. Примерами таких модификаций являются замены водорода алкильной, ацильной или аминогруппой.
Используемый здесь термин "делеция" означает изменение в нуклеотидной или аминокислотной последовательности, где один или более нуклеотидов или аминокислотных остатков, соответственно, отсутствуют.
Используемый здесь термин "инсерция" или "добавление" означает изменение в нуклеотидной или аминокислотной последовательности, которое приводит к добавлению одного или более нуклеотидов или аминокислотных остатков, соответственно, по сравнению с природным PDE5.
Используемый здесь термин "замена" означает замену одного или более нуклеотидов или аминокислот другими нуклеотидами или аминокислотами, соответственно.
При этом могут быть сделаны консервативные замены, например, в соответствии с нижеследующей таблицей. Аминокислоты в одном и том же блоке во втором столбце, а предпочтительно в одной и той же строке в третьем столбце могут быть заменены одна на другую.
Термин "гомологичный" охватывает понятие "гомология", а в частности, относится к структуре и означает любой структурный гомолог PDE5, который способен к кристаллизации.
Что касается гомологии подробно описанных здесь аминокислотных последовательностей, то эти аминокислотные последовательности предпочтительно, по крайней мере, на 70%, более предпочтительно, по крайней мере, на 75%, еще более предпочтительно, по крайней мере, на 80%, еще более предпочтительно, по крайней мере, на 85% и еще более предпочтительно, по крайней мере, на 90% гомологичны последовательностям SEQ ID NO:1, 2, 3, 4, 5 или 6. Более предпочтительно, если эти последовательности, по крайней мере, на 95%, а наиболее предпочтительно, по крайней мере, на 98% гомологичны последовательностям SEQ ID NO:1, 2, 3, 4, 5 или 6.
Что касается гомологии нуклеотидных последовательностей, кодирующих подробно описанные здесь аминокислотные последовательности, то эти нуклеотидные последовательности предпочтительно, по крайней мере, на 70%, более предпочтительно, по крайней мере, на 75%, еще более предпочтительно, по крайней мере, на 80%, более предпочтительно, по крайней мере, на 85% и еще более предпочтительно, по крайней мере, на 90% гомологичны нуклеотидным последовательностям, кодирующим SEQ ID NO:1, 2, 3, 4, 5 или 6. Более предпочтительно, если указанные последовательности, по крайней мере, на 95%, а наиболее предпочтительно, по крайней мере, на 98% гомологичны нуклеотидным последовательностям, кодирующим SEQ ID NO:1, 2, 3, 4, 5 или 6.
Термин "гомолог" по отношению к нуклеотидной последовательности PDE5, определенной в настоящей заявке, и аминокислотной последовательности PDE5, определенной в настоящей заявке, может быть синонимом термину "аллельные варианты" данных последовательностей.
В частности, используемый здесь термин "гомология" может быть синонимом термину идентичность. В настоящем описании гомология последовательности, например аминокислотной последовательности кристалла PDE5 настоящего изобретения, может быть определена путем простого "визуального" сравнения (то есть прямого сравнения) любой одной или более последовательностей с другой последовательностью, проводимого для того, чтобы убедиться, что эта другая последовательность, по крайней мере, на 70% идентична данной последовательности(ям). Относительная гомология последовательностей (то есть идентичность последовательностей) может быть также определена с помощью коммерчески доступных компьютерных программ, которые позволяют вычислить процент гомологии (%) между указанными двумя или более последовательностями. Типичным примером такой компьютерной программы является программа CLUSTAL.
% Гомологии может быть вычислен по непрерывным последовательностям, то есть путем сопоставления одной последовательности с другой последовательностью и непосредственного сравнения каждой аминокислоты в одной последовательности с соответствующей аминокислотой в другой последовательности, по одному аминокислотному остатку шаг за шагом. Это называется сопоставление "без пробелов". Обычно такое сопоставление без пробелов осуществляют только для относительно небольшого числа остатков (например, менее чем 50 смежных аминокислот).
Хотя этот метод является очень простым и удобным, однако он не учитывает то, что, например, в какой-либо идентичной паре последовательностей одна инсерция или делеция будет нарушать сопоставление следующих аминокислотных остатков, что может приводить в значительному снижению % гомологии при осуществлении глобального сопоставления первичных последовательностей. Следовательно, были разработаны методы сравнения наибольших частей последовательности для достижения оптимальных сопоставлений, которые учитывали бы возможные инсерции и делеции без излишнего наложения "штрафа" при общей оценке гомологии. Это может быть достигнуто путем введения "пробелов" при сопоставлении последовательностей в целях максимизации локальной гомологии.
Однако в этих более сложных методах предусматривается присвоение "штрафа на пробел" каждому пробелу, который используется при таком сопоставлении, так, чтобы для одного и того же числа идентичных аминокислот сопоставление последовательностей с использованием, по возможности, меньшего числа пробелов, отражающих более высокое сходство между двумя сравниваемыми последовательностями, была достигнута более высокая оценка, чем с использованием множества пробелов. При этом обычно используется понятие "цена аффинных пробелов", которая назначает относительно высокую "цену" на существование пробела и меньший штраф для каждого последующего остатка в данном пробеле. Такая система оценки с использованием "пробелов" является наиболее распространенной. Разумеется, что высокие штрафы на пробелы позволяют оптимизировать сопоставление с меньшим числом пробелов. Большинство программ по сопоставлению последовательностей дают возможность модифицировать штрафы на пробелы. Однако при применении таких программ для сравнения последовательностей предпочтительно использовать параметры по умолчанию. Так, например, при использовании пакета программ GCG Wisconsin Bestfit (см. ниже) "штраф на пробелы" по умолчанию для аминокислотных последовательностей составляет -12 для пробела и -4 для каждого удлинения.
Поэтому для вычисления максимального % гомологии сначала необходимо провести оптимальное сопоставление с учетом штрафов на пробелы. Подходящей компьютерной программой для осуществления такого сопоставления является пакет программ GCG Wisconsin Bestfit (University of Wisconsin, USA; Devereux et al., 1984, Nucleic Acids Research 12:387). Примерами других программ, которые могут быть использованы для проведения сравнения последовательностей, являются, но не ограничиваются ими, пакет программ BLAST (см. Ausubel et al., 1999, там же - глава 18), FASTA (Atschul et al., 1990, J. Mol. Biol. 403-410) и пакет программ GENEWORKS, используемых в качестве аппаратного средства для сравнения. Обе программы BLAST и FASTA являются доступными в режиме поиска "офф-лайн" и "он-лайн" (см. Ausubel et al., там же, стр.7-58 - 7-60). Однако в некоторых случаях предпочтительнее использовать программу GCG Bestfit.
Хотя конечный % гомологии может быть определен термином "идентичность", однако, в некоторых случаях, сам способ сопоставления обычно не основывается на сравнении пар по типу "все или ничего". Вместо этого обычно используют оценочную матрицу с масштабированным сходством, которая назначает оценки для каждого попарного сравнения исходя из химического сходства или эволюционной отдаленности. Примером такой матрицы, которую обычно используют, является матрица BLOSUM62 - матрица по умолчанию, имеющаяся в пакете программ BLAST. В программах GCG Wisconsin обычно используются либо общие параметры по умолчанию, либо сравнительная таблица специальных символов, если она прилагается (более подробно см. руководство пользователя). Для пакета программ GCG предпочтительно использовать общие параметры по умолчанию либо, при использовании других программ, матрицу по умолчанию, такую как BLOSUM62.
После осуществления оптимального сопоставления с использованием указанного пакета программ можно вычислить % гомологии, а предпочтительно % идентичности последовательностей. Этот пакет программ обычно осуществляет это вычисление в процессе сравнения последовательностей и дает численный результат.
Как уже указывалось, в некоторых случаях гомология (или идентичность) последовательностей может быть определена с использованием любого подходящего алгоритма гомологии, например с использованием параметров по умолчанию. Обсуждение основных проблем, связанных с поиском сходства последовательностей в базе данных, можно найти у Altschul et al. (1994) Nature Genetics 6:119-129. В некоторых случаях используется алгоритм BLAST с установленными параметрами по умолчанию. Алгоритм BLAST подробно описан на сайте http://www.ncbi.nih.gov/BLAST/blast_help.htlm. Преимущественно, "значительная гомология" при ее оценке с помощью программы BLAST означает, что последовательности соответствуют величине EXPECT, равной, по крайней мере, 7, предпочтительно, по крайней мере, 9, а более предпочтительно, по крайней мере, 10 или более. Для величины EXPECT в программе поиска BLAST порог по умолчанию обычно составляет 10.
Другими компьютерными методами определения идентичности и сходства между двумя последовательностями являются, но не ограничиваются ими, методы с использованием пакета программ GCG (Devereux et al., 1984, Nucleic Acids Research 12:387) и FASTA (Atschul et al., 1990, J. Mol. Biol. 403-410).
Аминокислотная последовательность PDE5 настоящего изобретения может быть продуцирована путем экспрессии нуклеотидной последовательности, кодирующей ту же последовательность в подходящей экспрессионной системе.
Кроме того, или альтернативно, сам белок может быть продуцирован методами химического синтеза аминокислотной последовательности PDE5, либо целиком или частично. Так, например, пептиды могут быть синтезированы твердофазными методами, могут быть отщеплены от смолы и очищены с помощью препаративной высокоэффективной жидкостной хроматографии (например, Creighton (1983) Proteins Structures and Molecular Principles, WH Freeman & Co., New York, NY, USA). Состав синтетических пептидов может быть подтвержден с помощью анализа или секвенирования аминокислотной последовательности (например, в соответствии с процедурой расщепления по Эдману).
Прямой пептидный синтез может быть осуществлен различными твердофазными методами (Roberge J.Y. et al., Science, Vol. 269, 1995, pp.202-204), а автоматический синтез может быть проведен, например, с использованием пептидного синтезатора ABI 431A (Perkin Elmer, Boston, MA, USA) в соответствии с инструкциями производителя. Кроме того, аминокислотная последовательность PDE5 или любая ее часть может быть модифицирована в процессе прямого синтеза и/или объединена химическими методами с последовательностью, происходящей от других субъединиц или любых их частей, с продуцированием варианта полипептида.
Структура каталитического домена PDE5 дикого типа, образующего комплекс с силденафилом
Экспрессировали рекомбинантную конструкцию каталитического домена (Е534-N975) человеческого PDE5, белок кристаллизовался в комплексе с силденафилом, и его структуру определяли методом многоволнового аномального рассеяния (Hendrickson et al., 1989).
В настоящее время определение этой структуры указывало на новую укладку, однако впоследствии была опубликована структура каталитического домена PDE4b (Xu et al., 2000). Топологическое сравнение каталитического домена PDE5 со структурами, имеющимися в банке данных белковых структур (Protein Data Bank, PDB), не выявило значительной гомологии с какими-либо известными структурами белков, за исключением структуры PLЕ4. Было проведено сравнение этих двух структур (на фигуре 1 показано сопоставление первичный последовательностей и вторичных структур для этих двух белков). Структура и расположение доменов PDE4b являются фактически такими же, как в PDE4, за исключением того, что второй субдомен, выявленный в PDE4, лишь частично присутствует в структуре PDE5, как подробно описано ниже.
Эта структура состоит из одного домена из 15 a-спиралей, образующих компактную укладку (фигура 2). Внутри целого домена могут также находиться три субдомена. Спирали 1 (Н1 539-545) и 2 (Н2 551-554) находятся на внешней стороне белка и составляют N-концевую область данной конструкции. Указанные две спирали не перекрываются с эквивалентными спиралями (Н0, Н1 и Н2) в структуре PDE4. Эта область не является абсолютно консервативной для всего семейства белков PDE. Спирали 3 (Н3 568-582), 4 (Н4 584-588), 5 (Н5 592-604), 6 (Н6 615-631) и 7 (Н7 640-652) образуют первый субдомен этого белка и находятся в сердцевине белка. Какой-либо заметной электронной плотности для спиралей 8 и 9 не наблюдалось исходя из номенклатуры PDE4. Спираль 10 (Н10 684-694) также находится на внешней части белка и образует димер в пограничной области структуры. Спирали 10 и 11 (Н11 706-721) представляет собой видимую часть второго субдомена. Спирали 12 (Н12а 725-731, Н12b 733-741), 13 (Н13 749-765), 14 (Н14 772-797), 15 (Н15 813-824), 16 (Н16 826-836) и 17 (Н17 841-861) образуют третий субдомен белка. При этом следует отметить, что в PDE5 спираль Н12 не является непрерывной спиралью, как в PDE4, и она состоит из двух коротких спиралей с изгибом в середине, а спираль Н15 представляет собой непрерывную спираль в PDE5, но не в PDE4.
Сборка димера для комплекса "PDE5 дикого типа - силденафил": каталитический домен
В асимметричном звене присутствуют четыре молекулы, причем каждая молекула содержит разрывы в цепи, а плотность в С-концевой части конструкции не определяется визуально (подробней см. ниже). Эти четыре молекулы могут быть определены как две копии димера. Молекула А (электронная плотность для остатков 534-536, 665-681, 863-875 не наблюдается) связывается с молекулой D (электронная плотность для остатков 534, 667-681, 865-875 не наблюдается), а молекула В (электронная плотность для остатков 534-536, 667, 865-875 не наблюдается) ассоциируется с молекулой С (электронная плотность для остатков 534-53, 663-678, 863-875 не наблюдается).
Молекулы в этом димере связаны посредством двойного вращения, при этом область раздела образуется посредством ассоциации спиралей Н10, происходящих от молекулы А и D. Ключевым фактором для такой ассоциации димеров является присутствие 2 ионов цинка (один из которых ассоциируется с каждым мономером). Остаток His 683 от одной молекулы и остаток His 684 и Asp 687 от другого димерного партнера координируют каждый ион цинка. Следует отметить, что координированная металлом димеризация является артефактом кристаллизации. Отсутствие определенных структурных областей в каждой молекуле, очевидно, является следствием высокой гибкости этой части структуры. Кроме того, очевидно, что в этой области происходит значительное расщепление белка, которое приводит к еще большей гибкости. Эта область соответствует спиралям Н8 и Н9 во втором субдомене структуры PDE4.
Комплекс "PDE5 дикого типа - силденафил": взаимодействие активного сайта и белка-ингибитора
Каждая из молекул с независимо уточненной структурой содержит одну молекулу силденафила, связанную в активном сайте. Активный сайт находится, главным образом, в третьем субдомене белка и ограничен спиралями Н15, Н14, С-концом Н13 и С-концом Н11 вместе с областью петли, расположенной между Н11 и Н12а. Большинство взаимодействий между ингибитором и белком по своей природе являются гидрофобными, при этом наблюдаются только две прямые водородные связи (фигура 3). Первая связь находится между N17 пуринового кольца ингибитора и Оe1 Gln 817 (2,8 Е), а вторая связь связывает смежный атом кислорода О16 ингибитора с Ne2 того же самого остатка Gln 817 (3,1Е).
Атом углерода С12 ингибитора указывает на расположение небольшого гидрофобного кармана, образованного Leu 765, Ala 767 и Ile 768. Эти остатки вместе с Phe 820 образуют плоскую поверхность для связывающего сайта, против которого располагается пуриновое кольцо ингибитора. Противоположная сторона пурина расположена напротив Val 782. С5-пропильный заместитель обеспечивает хорошие ван-дер-ваальсовские взаимодействия с Val 782 и Phe 786 и Tyr 612. Phe 786 и Leu 804 обеспечивают дополнительные гидрофобные взаимодействия с фенильной частью ингибитора. О-алкильная часть занимает небольшой карман, ограниченный Ala 779, Phe 786, Ala 783, Val 782, Leu 804, Ile 813, Met 816 и Gln 817. Сульфонамидная группа обращена в направлении растворителя, а пиперазиновое кольцо ограничено выступающими остатками 662-665, хотя пока еще не известно, влияет ли разрыв цепи на конформацию данной части структуры. При этом какого-либо прямого взаимодействия между ингибитором и ионом цинка в активном сайте не обнаруживалось. Структура подтверждает конкурентную природу такого типа ингибирования силденафила посредством связывания в активном сайте и, тем самым, блокирования доступа к субстрату cGMP (который также был смоделирован - данные не приводятся).
Комплекс "PDE5 дикого типа - силденафил": ионы металла в активном сайте
В активном сайте этой структуры присутствует лишь один атом цинка. Он может быть четко идентифицирован как атом цинка, поскольку фазы, используемые для определения структуры, были получены в эксперименте, проводимом методом трехволнового аномального рассеяния на цинке (MAD). Наблюдаемый аномальный сигнал явно указывал на присутствие иона цинка. Координация иона внутри активного сайта также совпадала с координацией, ожидаемой для цинка. Металл координирован остатками His 653 (Ne2-Zn 2,0Е), His 617 (Ne2-Zn 2,1Е), Asp 764 (OD2-Zn 2,2Е), а также Asp 654 (OD2-Zn 2,2Е). Указанные остатки являются полностью консервативными для всего семейства генов PDE. Каких-либо данных, свидетельствующих о присутствии второго иона металла в активном сайте, получено не было. Возможной причиной отсутствия любого второго иона металла в активном сайте является секвестрация иона металла (в данном случае цинка, как было также подтверждено по аномальному сигналу) и образование области раздела димера. Кроме того, возможно, что остатки, которые, по всей вероятности, участвуют в координации второго иона металла в активном сайте, не присутствуют в нативной конформации, что обусловлено близостью к разупорядоченной области белка и димерной области раздела.
Конструирование белка PDE5*
Анализ белка с каталитическим доменом, проводимый с помощью масс-спектрометрии и электрофореза в ДСН-ПААГ (данные не приводятся), показал, что белок расщепляется в области, которая не видна в этой структуре (остатки 664-682). Высокие концентрации ингибиторов протеазы обеспечивают некоторую стабилизацию белка, что позволяет определить вышеуказанную структуру.
С помощью генной инженерии была сконструирована форма каталитического домена PDE5, где область 657-682 PDE5 была заменена той же самой областью в PDE4, в результате чего была получена химерная конструкция (см. фигуру 1 для сопоставления первичных последовательностей в этой области). С-конец этой конструкции также был усечен (С-концев. 858) по сравнению со структурой дикого типа (С-концев. 875). Далее, эта сконцентрированная конструкция будет обозначаться "PDE5*".
Было показано, что этот сконструированный белок является стабильным в отношении расщепления, на что указывала масс-спекторометрия и электрофорез в ДСН-ПААГ (данные не приводятся). Этот белок обнаруживает улучшенные биофизические свойства, что позволяет разработать альтернативный протокол очистки. Этот новый протокол предусматривает связывание на колонке с сефарозой Blue и специфическое элюирование cGMP. Было показано, что белок дикого типа не связывается с этой колонкой, что, вероятно, обусловлено нарушением структуры возле сайта расщепления протеазой. Этот белок PDE5* был использован для продуцирования кристаллов с силденафилом, которые дают дифракционную картину с более высоким разрешением и не имеют разупорядоченных областей. Белок также был использован для репродуцирования кристаллов с другими ингибиторами, которые обычно дифрагируют с разрешением 1,8Е или выше, что делало этот улучшенный белок подходящим для разработки лекарственных средств на основе его структуры.
Структура каталитического домена PDE5* с силденафилом
Структуру каталитического домена белка PDE5* определяли методом молекулярного замещения с использованием структуры белка дикого типа в качестве исходной модели для поиска. Эта структура содержит 17 a-спиралей, и ее полная укладка имеет большое сходство со структурой дикого типа, но она также имеет ряд важных отличий. Главным отличием в этой структуре является присутствие спиралей Н8 и Н9, состоящих из замененной части, происходящей от PDE4, остатков 657-682. Эти спирали имеют такую же укладку, как и спирали, наблюдаемые в структуре PDE4, и образуют полный второй субдомен белка. Вся С-концевая область этой конструкции также может вносить свой вклад в электронную плотность, при этом на N-конце этой структуры остаются только три разупорядоченных остатка. С-область, очевидно, обеспечивает повышенную способность к кристаллизации. Каталитический домен PDE5* кристаллизуется как мономер с двумя молекулами, присутствующими в асимметричном элементе вследствие трансляционного сдвига. (PDE5* был также кристаллизован с другими ингибиторами PDE5 в спейсерной группе р21, с одной молекулой в асимметричном элементе. Кристаллы имеют элементарную кристаллическую ячейку размером приблизительно: а=56, b=77, с=83 Å, α=γ=90°, β=103°).
PDE5*: Взаимодействия активного сайта и белка-ингибитора
Каждая из молекул с независимо уточненной структурой содержит одну молекулу силденафила в активном сайте. Силденафил занимает ту же самую область активного сайта, которая наблюдалась для структуры дикого типа, обеспечивая, в основном, аналогичные гидрофобные взаимодействия с белком (фигура 4). Те же самые две простые водородные связи образуются между Gln 817 белка и ингибитором (Оe1- N17 2,8 Å и Ne2-О16 3,1 Å). Остальная часть ингибитора обеспечивает такие же взаимодействия с сульфонилпиперазином, также ориентированным в направлении растворителя. Эта часть близка к сконструированной области белка, но пиперазиновое кольцо не взаимодействует с упорядоченной замененной областью конструкции каталитического домена. Этот факт имеет очень важное значение при рассмотрении возможности использования данного химерного каталитического домена для получения лекарственных средств.
PDE5*: Ионы металла в активном сайте
Другим заметным отличием в структуре PDE5*, по сравнению со структурой PDE5 дикого типа, является присутствие двух ионов металла в активном сайте. Как наблюдалось для комплекса дикого типа, прямого взаимодействия между ингибитором и ионом цинка, присутствующим в активном сайте, не происходит. В этом комплексе также не наблюдалось прямого взаимодействия между силденафилом и вторым ионом металла. Этот второй ион металла координирован к Asp 764 (OD1 2,15 Å) и к водной структуре, которая стабилизирует окружение металла. Исходя из координационной геометрии и наблюдаемой относительной электронной плотности, этот второй ион металла был точно определен как Mg2+, что соответствует аналогичному наблюдению при разрешении структуры PDE4. Это структурное расположение соответствует предполагаемому механизму, где ион ОН- происходит от молекулы H2O, ионизованной в присутствии атомов двухвалентного металла, связанных в активном сайте (Goldberg et al., 1980, Francis et al., 1994). Фосфодиэфирная связь между атомами фосфора и кислорода в 3'-положении cGMP затем гидролизуется посредством нуклеофильной атаки ОН-.
Настоящее изобретение подробно описано на нижеследующих примерах со ссылками на список последовательностей и графический материал, где:
SEQ ID NO:1 представляет собой аминокислотную последовательность области петли PDE5.
SEQ ID NO:2 представляет собой аминокислотную последовательность каталитического домена PDE5 дикого типа.
SEQ ID NO:3 представляет собой аминокислотную последовательность полноразмерной последовательности PDE5 дикого типа.
SEQ ID NO:4 представляет собой аминокислотную последовательность области петли PDE4.
SEQ ID NO:5 представляет собой аминокислотную последовательность каталитического домена PDE5 с замененной петлей = PDE5*.
SEQ ID NO:6 представляет собой аминокислотную последовательность полноразмерной последовательности PDE5, содержащей PDE5*.
SEQ ID NOS:7-14 представляют собой олигонуклеотидные праймеры.
На фигуре 1 показано сопоставление каталитических доменов PDE5 (верхней последовательности) и PDE4b (нижней последовательности). Положения и номера спиралей помечены для каждой структуры. Остатки, помеченные жирным шрифтом, относятся к последовательности, сопоставляемой с последовательностью сконструированной области. Последовательность PDE4 была использована для замены соответствующей области в PDE5. Эта замена приводит к инсерции остатка в данной области. Подчеркивание означает, что С-концевая область отсутствует в PDE5*.
На фигуре 2 дано ленточное представление полной укладки белков с указанием элементов вторичной структуры. Ингибитор показан во всех атомах в виде палочек, а ионы металла показаны в виде сфер. (А)=PDE4b, (В)=PDE5 дикого типа + силденафил, (С)=PDE5 с "замененной петлей" (PDE5*) + силденафил. Спирали пронумерованы с использованием структуры PDE4 в качестве эталона. Спирали НО-Н7 образуют субдомен 1, спирали Н8-Н11 образуют субдомен 2, а спирали Н12-Н16 образуют субдомен 3.
На фигуре 3 показан вид соединения силденафила, связанного с PDE5 дикого типа.
На фигуре 4 показан вид соединения силденафила, связанного с PDE5 с "замененной петлей" (PDE5*).
На фигуре 5 показана химическая структура ингибитора силденафила.
Экспериментальная часть
Пример 1 - Конструирование и экспрессия каталитического домена PDE5 дикого типа (Е534-N875)
Олигонуклеотидные праймеры конструировали исходя из последовательности человеческого PDE5 (рег.№ АВ001635). ДНК-фрагменты генерировали путем ПЦР-амплификации из полноразмерного клона PDE5. При этом были использованы следующие олигонуклеотиды:
5'-немеченый олигонуклеотид PDE5
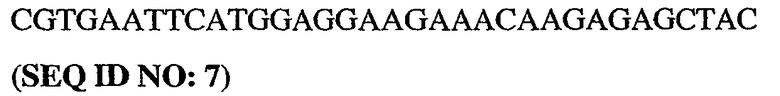
3'-длинный олигонуклеотид PDE5
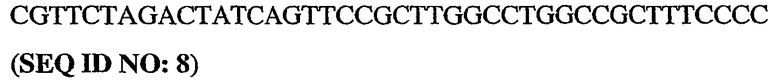
ПЦР-реакцию осуществляли за 30 циклов в полном объеме, составляющем 50 мкл, в растворе, содержащем 1,5 мМ MgCl2, 200 мкМ dNTP, 50 пмоль каждого праймера и 2,5 единиц ДНК-полимеразы Expand (Roche, East Sussex, UK). Каждый цикл проводили в следующем режиме: 94°С, 1 мин; 50°С, 1 мин; и 72°С, 2 минуты.
Конечные амлифицированные ДНК-фрагменты для обеих конструкций разделяли на 1% агарозном геле и очищали с использованием набора для экстракции геля QIAquick (Qiagen, West Sussex, UK). Затем фрагмент гидролизовали ферментами EcoRI и XbaI и лигировали в EcoRI/XbaI-расщепленный вектор pFastbac1 (Life Technologies, Paisley, UK). Лигирование осуществляли при 12°С в течение 16 часов. Лигированную смесь подвергали электропорации в E.coli (TOP10)(Invitrogen, Gronigen, The Netherlands).
Клоны, содержащие нужную вставку, отбирали с использованием планшетов со средой 2YT, содержащей 100 мкг/мл ампициллина, и оценивали на присутствие вставки нужного размера путем эндонуклеазного расщепления. Анализ ДНК-последовательности осуществляли как описано Lark (Saffron Waldon, UK).
Рекомбинантную бакмидную ДНК продуцировали путем трансформации DH10BACTME.coli плазмидной ДНК pFastbac1::каталитический домен PDE5 (534-875). Это продуцирование осуществляли методом, описанным в руководстве по бакуловирусной экспрессии "Вас to BacTM baculovirus expression manual" (Life Technologies, Paisley, UK). Для подтверждения успешного переноса в бакмиду проводили ПЦР-анализ с использованием праймеров для амплификации pUC19/М13 (Invitrogen, Gronigen, The Netherlands).
Генерирование первичных бакуловирусных штаммов осуществляли путем трансфекции с использованием клеток насекомых Sf-9. Бакмидную ДНК, содержащую нужную вставку, смешивали с трансфецирующим реагентом CELLFECTINTM (Life Technologies, Paisley, UK) и добавляли в монослой клеток насекомых Sf-9 с использованием бессывороточной среды SF-900II (Invitrogen, Gronigen, The Netherlands). После инкубирования в течение 72 часов при 27°С супернатант собирали в виде исходного бакуловирусного штамма. Этот штамм амплифицировали путем добавления исходного вирусного штамма в суспензию клеток насекомых Sf-9 при плотности 1х106 клеток/мл в 1-литровых колбах Эрленмейера (Corning Life Sciences, New York, USA) с перемешиванием при 125 об/мин и при 27°С. Через 6 дней после инфицирования супернатант собирали центрифугированием и хранили при 4°С в виде рабочего вирусного штамма. Титр этого рабочего штамма определяли с помощью стандартного анализа методом бляшек, как описано в руководстве по бакуловирусной экспрессии "Вас to BacTM baculovirus expression manual" (Invitrogen, Gronigen, The Netherlands).
Экспрессию белка оптимизировали в культурах в колбах Эрленмейера с использованием культур клеток насекомых Sf-9 и High5 T.ni, скринируя при различных множественностях заражения (m.o.i.) и временах сбора, после чего установленные оптимальные условия корректировали в соответствии с масштабами ферментеров.
Используемыми ферментерами были 3-литровые автоклавы Applikon с мешалкой, регулируемые с использованием биоконтроллеров Applikon 1030. Сначала из культур в шейкерных колбах получали инокулят клеток High5 T.ni. Ферментер инокулировали клетками при плотности 5х105 клеток/мл, при этом исходный рабочий объем в бессывороточной среде Excel 405 (JRH Biosciences, Kansas, USA) составлял 1,8 литров. Температуру поддерживали при 27°С, концентрацию растворенного кислорода поддерживали на уровне 60%, а рН измеряли, но не корректировали. Концентрацию кислорода регулировали в течение всей процедуры. Перемешивание проводили в культуре при 150 об/мин с использованием системы судовых мешалок с двойными лопастями и мешалки Раштона (Rushton) на границе раздела жидкость/"пустая зона". Аэрирование непрерывно проводили до "пустой зоны" при 0,5 л/мин.
Когда жизнеспособные клетки достигали плотности 2х106 клеток/мл, культуру инфицировали титрованным рабочим штаммом бакуловируса при m.o.i.=1. Данную культуру собирали через 48 часов после инфицирования. Сбор осуществляли путем центрифугирования при 2000 g в течение 15 минут, после чего осадок клеток насекомых хранили при -80°С до очистки.
Пример 2 - Конструирование и экспрессия каталитического домена His6-меченого PDE5 дикого типа (Е534-N875)
Олигонуклеотидные праймеры конструировали на основе из последовательности человеческого PDE5 (изоформы=PDE5A1; рег.№ =АВ001635). ДНК-фрагменты генерировали путем ПЦР-амплификации из полноразмерного клона PDE5. При этом были использованы следующие олигонуклеотиды:
5'- His6-меченый олигонуклеотид PDE5
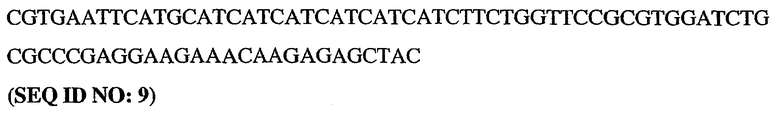
3'-длинный олигонуклеотид PDE5
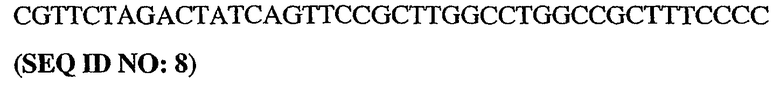
ПЦР-реакцию осуществляли за 30 циклов в полном объеме, составляющем 50 мкл, в растворе, содержащем 1,5 мМ MgCl2, 200 мкМ dNTP, 50 пмоль каждого праймера и 2,5 единиц ДНК-полимеразы Expand (Roche, East Sussex, UK). Каждый цикл проводили в следующем режиме: 94°С, 1 мин; 50°С, 1 мин; и 72°С, 2 минуты.
Конечные амлифицированные ДНК-фрагменты для обеих конструкций разделяли на 1% агарозном геле и очищали с использованием набора для экстракции геля QIAquick (Qiagen, West Sussex, UK). Затем фрагмент расщепляли ферментами EcoRI и XbaI и лигировали в EcoRI/XbaI-расщепленный вектор pFastbac1 (Life Technologies, Paisley, UK). Лигирование осуществляли при 12°С в течение 16 часов. Затем лигированную смесь подвергали электропорации в E.coli (TOP10) (Invitrogen, Gronigen, The Netherlands).
Клоны, содержащие нужную вставку, отбирали с использованием планшетов со средой 2YT, содержащей 100 мкг/мл ампициллина, и оценивали на присутствие вставки нужного размера посредством эндонуклеазного расщепления. Анализ ДНК-последовательности осуществляли как описано Lark (Saffron Waldon, UK).
Рекомбинантный бакуловирус генерировали методами, описанными для каталитического домена PDE5 дикого типа (см. пример 1).
В данном случае, оптимизация экспрессии указывала на то, что клетки насекомых High5 T.ni давали наилучшую экспрессию. Поэтому бакуловирусную экспрессию в ферментерах осуществляли с использованием тех же самых процедур, которые были описаны для предыдущей конструкции.
Пример 3 - Конструирование и экспрессия PDE5* (Е534-Е858) в бакуловирусе
Конструкцию PDE5* получали посредством ПЦР с перекрывающимся удлинением, где были использованы следующие олигонуклеотиды:
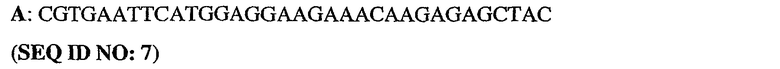
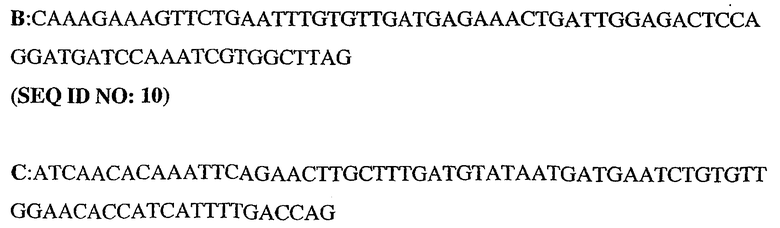
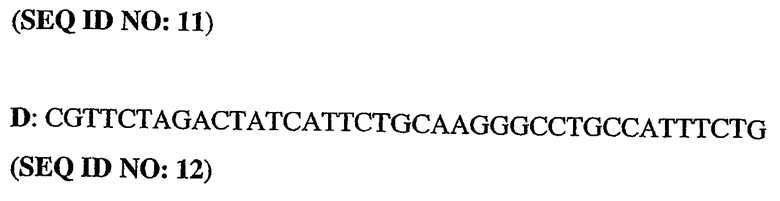
Начальные ДНК-фрагменты конструировали с использованием олигонуклеотидов А+В и С+D с помощью той же матричной ДНК, которая была использована для конструирования каталитического домена PDE5 дикого типа. ПЦР-реакцию осуществляли за 30 циклов в полном объеме 50 мкл в растворе, содержащем 1,5 мМ MgCl2, 200 мкМ dNTP, 50 пмоль каждого праймера и единицы ДНК-полимеразы Expand (Roche, East Sussex, UK). Каждый цикл проводили в следующем режиме: 94°С, 1 мин; 50°С, 2 мин; и 72°С, 3 минуты. Во втором раунде ПЦР, ДНК-продукты от нуклеотидов А+В и С+D для ПЦР использовали в качестве матричной ДНК вместе с олигонуклеотидами А+D для амплификации полноразмерной конструкции. ПЦР-условия были аналогичны условиям исходной ПЦР-реакции. Эта реакция генерировала конструкцию с замененной областью PDE4 и с С-концевым усечением (С-концев. 858), в отличие от каталитического домена PDE5 (С-концев. 875).
Рекомбинантный бакуловирус генерировали методами, описанными для каталитического домена PDE5 дикого типа (см. пример 1).
И в данном случае оптимизация экспрессии указывала на то, что клетки насекомых High5 T.ni давали наилучшую экспрессию. Поэтому бакуловирусную экспрессию в ферментерах осуществляли с использованием тех же самых процедур, которые были описаны для предыдущей конструкции.
Пример 4 - Конструирование и экспрессия PDE5* (Е534-Е858) в E.coli
Конструкцию PDE5* продуцировали в E.coli посредством ПЦР, где были использованы нижеследующие олигонуклеотиды, а в качестве матричной ДНК была использована плазмидная ДНК pFastbac1::PDE5* (подтвержденная последовательность), продуцированная как описано в примере 3.
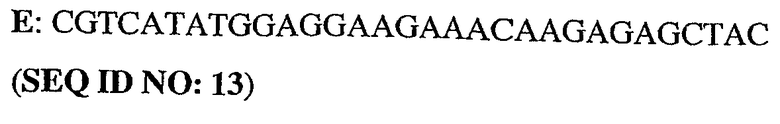
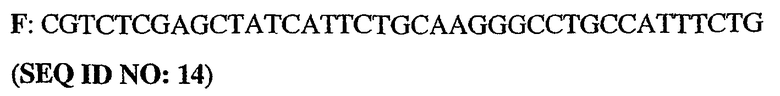
ПЦР-реакцию осуществляли за 30 циклов в полном объеме 50 мкл в растворе, содержащем 1,5 мМ MgCl2, 200 мкМ dNTP, 50 пмоль каждого праймера и 2,5 единиц ДНК-полимеразы Expand (Roche, East Sussex, UK). Каждый цикл проводили в следующем режиме: 94°С, 1 мин; 50°С, 1 мин; и 72°С, 2 минуты.
Конечный амлифицированный ДНК-фрагмент выделяли на 1% агарозном геле и очищали с использованием набора для экстракции геля QIAquick (Qiagen, West Sussex, UK). Затем фрагмент расщепляли ферментами NdeI и XhoI и лигировали в NdeI/XhoI-расщепленный вектор pЕТ21С (Novagen, Nottingham, UK). Лигирование осуществляли при 12°С в течение 16 часов. Затем лигированную смесь подвергали электропорации в E.coli (TOP10) (Invitrogen, Gronigen, The Netherlands).
Клоны, содержащие нужную вставку, отбирали с использованием планшетов со средой 2YT, содержащей 100 мкг/мл ампициллина. Плазмидную ДНК также оценивали на присутствие вставки нужного размера посредством эндонуклеазного расщепления. Анализ ДНК-последовательности осуществляли, как описано у Lark (Saffron Waldon, UK).
Затем правильно секвенированную плазмидную ДНК подвергали электропорации в E.coli BL21 (DE3) (Novagen, Nottingham, UK) для экспрессии. Экспрессию осуществляли в 7-литровых ферментерах Applikon с использованием 5 литров бульона 2YT, содержащего 100 мкг/мл карбенициллина в качестве среды. Перемешивание осуществляли при 1000 об/мин с использованием системы мешалок Раштона с двойными лопастями и аэрировали с помощью барботера со скоростью 2 литра/мин. Ферментер инокулировали ночной культурой, выращенной в шейкерных колбах при 37°С и при 200 об/мин, при этом плотность инокуляции составляла 1% об./об. рН среды для ферментации поддерживали при 7,2 с использованием 20% об./об. раствора NH4ОН, а начальную температуру устанавливали на 37°С. После достижения OD600нм=1,5, установленную температуру снижали до 25°С, и культуру индуцировали IPTG при конечной концентрации 1 мМ. Через 4 часа после индуцирования, ферментационную культуру собирали путем периодического центрифугирования (8000 об/мин в течение 10 минут). Затем полученный осадок замораживали (-80°С) и хранили до последуюшей очистки.
Пример 5 - Очистка каталитического домена PDE5 дикого типа
Осадок, полученный в результате ферментации, ресуспендировали в 10 мл буфера для лизиса на один грамм сырой клеточной массы и механически разрушали с использованием клеточного дезинтегратора непрерывного действия (Constant Systems, Warwickshire, UK) под давлением 20 к.фунт/кв.дюйм. Буфер для лизиса состоял из 50 мМ Трис-HCl (рН 7,2), 100 мМ NaCl, 1 мМ DL-дитиотреитола (DTT) и содержал таблетки из полной смеси ингибиторов протеазы без EDTA (Roche, East Sussex, UK) и 10 мкМ эпоксисукцинил-1-лейциламидо-(4-гуанидино)бутан (Е-64) (Sigma, Dorset, UK; Catalogue № Е-3132). Лизат охлаждали и центрифугировали при 14000 g в течение 45 минут для удаления клеточного дебриса. Все процедуры очистки последовательно осуществляли с использованием системы очистки Dkta Explorer (Amersham Pharmacia, Buckinghamshire, UK). Супернатант наносили на проточную 50 мл-колонку с Q-сефарозой (Amersham Pharmacia, Buckinghamshire, UK) со скоростью потока 5 мл/мин, где выходящий поток непосредственно подавали на 20 мл колонку с никелевым хелатным комплексом (Amersham Pharmacia, Buckinghamshire, UK), предварительно загруженную 0,1 М NiSO4. Эту колонку с никелевым хелатным комплексом промывали 5 колоночными объемами буфера для лизиса. После этого колонку поэтапно элюировали буфером для лизиса, содержащего 50 мМ имидазола. Полученную элюированную фракцию непосредственно вводили в 2-литровую колонку для сверхтонкого обессоливания G-25 (Amersham Pharmacia, Buckinghamshire, UK), уравновешенную в SP-сефарозном буфере А (25 мМ Бис-трис (рН 6,5), 50 мМ NaCl, 1 мМ DTT и 2 мкМ Е-64). Белок элюировали в этом буфере при 50 мл/мин. Затем элюированную фракцию загружали на высокоразрешающую 20 мл колонку с SP-сефарозой (Amersham Pharmacia, Buckinghamshire, UK) при скорости потока 5 мл/мин. Проходящий поток собирали и диализовали в течение ночи при 4°С в гепариновом буфере А (25 мМ Бис-трис (рН 6,5), 1 мМ DTT и 2 мкМ Е-64). Объем диализа в 50 раз превышал объем образца белка, а в качестве диализной трубки использовали 10 кДа SnakeskinTM (Pierce, Cheshire, UK).
Затем диализованный образец загружали на 20 мл колонку с гепарин-сефарозой (Amersham Pharmacia, Buckinghamshire, UK), уравновешенную гепариновым буфером А. Колонку элюировали с использованием 10 колоночных объемов линейного градиента гепаринового буфера А, содержащего 300 мМ NaCl при скорости потока 3 мл/мин. Фракции, содержащие каталитический домен PDE5 (534-875), собирали и концентрировали до 2 мг/мл с использованием центрофужных концентраторов для белка (Vivascience, Gloucestershire, UK), а затем загружали при скорости потока 1,5 мл/мин в колонку 26/60 с Superdex-200 Prep, предварительно уравновешенную 50 мМ Бис-трис (рН 6,8), 500 мМ NaCl, 1 мМ DTT и 2 мкМ Е-64. Элюированные фракции анализировали с помощью электрофореза в трис-глицин-ДСН-ПААГ-гелях.
Пример 6 - Очистка каталитического домена PDE5 дикого типа (для апо-кристаллизации)
Осадок, полученный в результате ферментации, ресуспендировали в 5 мл буфера для лизиса на один грамм сырой клеточной массы и механически разрушали с использованием клеточного дезинтегратора непрерывного действия (Constant Systems, Warwickshire, UK) под давлением 20 к.фунт/кв.дюйм. Буфер для лизиса состоял из 50 мМ Бис-трис (рН 6,8), 10 мМ имидазола, 10% глицерина, 50 мМ хлорида натрия и 3 мМ b-меркаптоэтанола (b-МЕ) и содержал таблетки из полной смеси ингибиторов протеазы без EDTA (Roche, East Sussex, UK). Лизат охлаждали и центрифугировали при 13000 х g в течение 30 минут для удаления клеточного дебриса, а затем пропускали через 0,2 мкм фильтр. Все процедуры очистки последовательно осуществляли с использованием системы очистки ЖЭХБ (Amersham Pharmacia, Buckinghamshire, UK). Супернатант наносили на 20 мл колонку с никелевым хелатным комплексом (Amersham Pharmacia, Buckinghamshire, UK), предварительно загруженную 0,1 М NiSO4. Эту колонку с никелевым хелатным комплексом промывали 10 колоночными объемами буфера А (буфер для лизиса с таблетками из полной смеси ингибиторов протеазы не использовали), а затем 10 колоночными объемами буфера А, содержащего 50 мМ имидазола. Затем колонку элюировали градиентом 100-500 мМ имидазола в буфере А. Эти элюированные фракции анализировали с использованием электрофореза в трис-глицин-ДСН-ПААГ-геле и хранили в течение ночи при 4°С. Фракции, содержащие каталитический домен PDE5, концентрировали до 1,5 мг/мл с использованием центрифужных концентраторов Centriprep с отсечкой молекулярной массы 10 кДа (Amicon Bioseparations, Maine, USA) при скорости 3000 об/мин, 4°С. Половину концентрированной фракции загружали на 320 мл колонку с Сефакрилом S300НR (Amersham Pharmacia, Buckinghamshire, UK), предварительно уравновешенную в 50 мМ Бис-трис, рН 6,8, 10% глицерина, 50 мМ NaCl, 1 мМ DL-дитиотрейтола (DTT), при скорости потока 2 мл/мин. Элюированные фракции анализировали с помощью электрофореза в трис-глицин-ДСН-ПААГ-гелях, и фракции, содержащие каталитический домен PDE5, хранили при -80°С.
Пример 7 - Очистка каталитического домена PDE5*
Осадок, полученный в результате ферментации в E.coli и в бакуловирусе, ресуспендировали в 10 мл буфера для лизиса на один грамм сырой клеточной массы и механически разрушали с использованием клеточного дезинтегратора непрерывного действия (Constant Systems, Warwickshire, UK) под давлением 20 к.фунт/кв.дюйм. Буфер для лизиса состоял из 50 мМ Трис-HCl (рН 7,5), 100 мМ NaCl, 1 мМ DTT и содержал таблетки из полной смеси ингибиторов протеазы без EDTA (Roche, East Sussex, UK) и 10 мкМ Е-64. Лизат охлаждали и центрифугировали при 14000 g в течение 45 минут для удаления клеточного дебриса. Все процедуры очистки последовательно осуществляли с использованием системы очистки Äkta Explorer (Amersham Pharmacia, Buckinghamshire, UK). Супернатант наносили на проточную 50 мл колонку с Q-сефарозой (Amersham Pharmacia, Buckinghamshire, UK) со скоростью потока 5 мл/мин и выходящий поток собирали. Затем образец проходящего потока непосредственно подавали со скоростью 50 мл/мин на 2-литровую колонку для сверхтонкого обессоливания G-25 (Amersham Pharmacia, Buckinghamshire, UK), предварительно уравновешенную в буфере А с сефарозой Blue (50 мМ Бис-трис (рН 6,4), 50 мМ NaCl, 2 мМ EDTA, 2 мМ EGTA и 1 мМ DTT). Белковую фракцию элюировали в буфере А с сефарозой Blue.
Следующий этап колоночной очистки проводили последовательно, то есть сначала образец загружали в высокоразрешающую 20 мл колонку с SP-сефарозой (Amersham Pharmacia, Buckinghamshire, UK), а затем поток, выходящий из колонки, непосредственно загружали в проточную 10 мл колонку с сефарозой Blue (Amersham Pharmacia, Buckinghamshire, UK) при скорости потока 2 мл/мин. После завершения загрузки, колонку с SP-сефарозой удаляли из линии, а колонку с сефарозой Blue промывали 5 колоночными объемами буфера А с сефарозой Blue. Колонку промывали буфером А с сефарозой Blue, содержащим 1 М NaCl, до тех пор, пока оптическая плотность при 280 нм не достигала своего фонового значения, после чего колонку промывали 5 колоночными объемами буфера А с сефарозой Blue. Белок PDE5* поэтапно элюировали с использованием буфера А с сефарозой Blue, содержащего 20 мМ cGMP (Na-соль) (Sigma, Dorset, UK). Фракции анализировали на гелях с трис-глицин-ДСН (Invitrogen, Gronigen, The Netherlands) и соответствующим образом собирали. Эти фракции концентрировали до 2,5 мг/мл с использованием центрифужных концентраторов (Vivascience, Gloucestershire, UK) и загружали при скорости 1,5 мл/мин в колонку 26/60 с Superdex-200 Prep, предварительно уравновешенную 50 мМ Бис-трис (рН 6,8), 50 мМ NaCl, 1 мМ DTT и 2 мкМ Е-64. Элюированные фракции анализировали с помощью электрофореза в трис-глицин-ДСН-ПААГ-гелях.
Пример 8 - Кристаллизация his6-меченого каталитического домена (апо) PDE5 дикого типа
Фракции PDE5 из последней гель-фильтрационной колонки, которые были заморожены при -80°С, оттаивали и измеряли концентрацию белка. Раствор концентрировали до 5,8 мл/мл с использованием центрифужного концентратора Centriprep с отсечкой молекулярной массы 10 кДа (Amicon Bioseparations, Maine, USA) при скорости 3000 об/мин, 20°С, а затем переносили в центрифужный концентратор Centricon с отсечкой молекулярной массы 10 кДа (Amicon Bioseparations, Maine, USA) и концентрировали до 12,8 мг/мл при скорости 4000 об/мин, 20°С. После этого раствор белка разводили до 10 мг/мл с использованием ультрафильтрата, полученного в конечной стадии концентрирования, и замораживали до -80°С. Перед кристаллизацией раствор белка оттаивали и центрифугировали в течение 2 минут при 14000 об/мин в центрифуге Эппендорфа.
Эксперименты по кристаллизации осуществляли путем диффузии из паровой фазы при 20°С методом "висячей" капли. Капли, состоящие из 2 мкл резервуарного буфера, смешанного с 2 мкл раствора белка, суспендировали на силиконизированных покровных стеклах в 950 мкл резервуарных растворах, содержащих 50 мМ HEPES, рН 7,6, 1,1 М одноосновного фосфата натрия и 1,1 М одноосновного фосфата калия (все они были получены от Sigma, Dorset, UK). Перед использованием, кристаллизаторы и резервуарные растворы охлаждали до 4°С. Комплект кристаллизоторов помещали в холодное помещение при 4°С. Через 1-2 недели из осадка вырастали палочкообразные кристаллы, которые в своем наибольшем измерении достигали 400 мкм. Кристаллы постепенно переносили в помещение при 4°С, с использованием растворов с возрастающей концентрацией глицерина, в раствор, содержащий 0,1 М HEPES, рН 7,6, 2,3 М одоосновный фосфат натрия и 20% глицерин, используемый в качестве криопротектанта. Затем образцы быстро замораживали и проводили сбор данных для рентгеноструктурного анализа.
Пример 9 - Кристаллизация каталитического домена PDE5 дикого типа с силденафилом
Фракции PDE5 из последней гель-фильтрационной колонки собирали (общий объем составлял 25 мл) и определяли концентрацию белка (0,2 мг/мл). В раствор белка добавляли 10 мкМ Е-64 и 1 мг/мл лейпептина (Sigma, Dorset, UK). Раствор концентрировали до 3 мл/мл с использованием центрифужного концентратора Centriprep с отсечкой молекулярной массы 10 кДа (Amicon Bioseparations, Maine, USA) при скорости 3000 об/мин, 4°С. В раствор белка добавляли 3 х молярных эквивалента силденафила (10 мг/мл водного маточного раствора), а затем его концентрировали до 8 мг/мл. Затем к этому раствору добавляли еще один молярный эквивалент силденафила и концентрировали до 10 мг/мл. Перед кристаллизацией раствор белка центрифугировали в течение 5 минут при 14000 об/мин в центрифуге Эппендорфа.
Эксперименты по кристаллизации осуществляли путем диффузии из паровой фазы при 20°С методом "висячей" капли. Капли, состоящие из 2 мкл резервуарного буфера, смешанного с 2 мкл раствора белка, суспендировали на силиконизированных покровных стеклах в 900 мкл резервуарных растворах, содержащих 0,1 М трис, рН 8,0, 50 мМ фосфата аммония, рН 7,0, 16-26% PEG2КММЕ (все они были получены от Sigma, Dorset, UK). Через 2-5 дней, из осадка вырастали кристаллы прямоугольной формы, которые в своем наибольшем измерении достигали 300 мкм. Кристаллы переносили в раствор, содержащий 0,1 М трис, рН 8,0, 250 мМ NaCl, 10% глицерин и 12-22% PEG2КММЕ, используемый в качестве криопротектанта. Затем образцы быстро замораживали, а затем проводили сбор данных для рентгеноструктурного анализа.
Пример 10 - Кристаллизация PDE5*
Фракции PDE5* из последней гель-фильтрационной колонки собирали (общий объем составлял 25 мл) и определяли концентрацию белка (0,2 мг/мл). В раствор белка добавляли 10 мкМ Е-64 и 1 мг/мл лейпептина (Sigma, Dorset, UK). Раствор концентрировали до 10 мл/мл с использованием центрифужного концентратора Centriprep с отсечкой молекулярной массы 10 кДа (Amicon Bioseparations, Maine, USA) при скорости 3000 об/мин, 4°С. Перед кристаллизацией раствор белка центрифугировали в течение 5 минут при 14000 об/мин в центрифуге Эппендорфа.
Эксперименты по кристаллизации осуществляли путем диффузии из паровой фазы при 4°С методом "висячей" капли. Капли, состоящие из 2 мкл резервуарного буфера, смешанного с 2 мкл раствора белка, суспендировали на силиконизированных покровных стеклах в 900 мкл резервуарных растворов, содержащих 0,2 М ацетата натрия, 0,1 трис-гидрохлорид, рН 8,5, 30% мас./об. полиэтиленглиголя 8000 (раствор представляет собой компонент номер 22 в наборе Crystal Screen® от Hampton Research, California, USA). Через 1-2 дня из осадка вырастали кристаллы в форме пластинок, которые в своем наибольшем измерении достигали 700 мкм. Кристаллы переносили в раствор, содержащий 0,16 М ацетата натрия, 80 мМ трис-гидрохлорида, рН 8,5, 24% мас./об. полиэтиленглиголя 8000 и 10% глицерина, а затем замораживали во время сбора данных для рентгеноструктурного анализа.
Пример 11 - Кристаллизация PDE5* с силденафилом
К очищенному белку PDE5* добавляли 10 мкМ Е-64 и 1 мг/мл лейпептина (Sigma, Dorset, UK).
Методом, проводимым точно так же, как было описано выше для каталитического домена PDE5 дикого типа, получали комплекс PDE5* с силденафилом, и концентрировали до конечной концентрации белка 10 мг/мл.
Эксперименты по кристаллизации осуществляли путем диффузии из паровой фазы при 4°С методом "висячей" капли. Капли, состоящие из 2 мкл резервуарного буфера, смешанного с 2 мкл раствора белка, суспендировали на силиконизированных покровных стеклах в 900 мкл резервуарных растворов, содержащих 0,1 М трис, рН 7,4, 50 мМ фосфата аммония, рН 7,5, 30-24% PEG2КММЕ (Sigma, Dorset, UK). Через 2-5 дней, при добавлении 28% PEG2KMME, вырастали кристаллы в форме тонких пластинок, которые в своем наибольшем измерении достигали 600 мкм.
Кристаллы переносили в раствор, содержащий 0,1 М трис, рН 7,4, 250 мМ NaCl, 10% глицерина и 26-20% PEG2KMME, используемый в качестве криопротектанта. Затем образцы быстро замораживали, после чего проводили сбор данных для рентгеноструктурного анализа.
Пример 12 - Сбор данных, определение структуры и уточнение параметров PDE5 дикого типа
Структуру рекомбинантного человеческого PDE5 определяли методом многоволнового аномального рассеяния (MAD) с использованием четырех длин волн у края LIII цинка.
Данные природной рентгеновской дифракции собирали с помощью MAR CCD на станции ВМ 14 ESRF, Grenoble, France. Все данные обрабатывали с использованием пакета программ HKL (Otwinkowski & Minor, 1997). Статистические данные систематизированы в таблице 1а.
Кристаллы принадлежали к пространственной группе Р62 с размерами элементарной ячейки а=94,921 Å, b=94,921 Å, c=81,850 Å, α=β=90°, γ=120°. Они содержали 1 молекулу на асимметричный элемент (Mw=39654,71 Да) и их вычисленное содержание растворителя составляло 43,23% (Vм=2,18; Matthews, 1968).
Место расположения аномальных тяжелых атомов определяли с использованием программы SOLVE (Terwilliger & Berendzen, 1997). Уточнение параметров тяжелых атомов и вычисление фаз осуществляли с использованием программы SHARP (de La Fortelle & Bricogne, 1997). Фазы уточняли за 100 циклов путем "уплощения" растворителя с использованием программы SOLOMON (Abrahams & Leslie, 1996). Полученная карта имела хорошее качество и была использована для выявления примерно 70% структуры с использованием QUANTA (Quanta 98, 1998, version 98.1111; Molecular Simulations Inc., San Diego, CA 92121-3752, USA).
Модель уточняли путем сравнения факторов нативной структуры (Fр-calc), полученных с помощью SHARP исходя из комбинации экспериментальных нативного (Fр) и выведенного (FРН) структурных факторов. Уточнение параметров проводили в диапазоне разрешения 30-2,5 Å с использованием программы XPLOR (Brünger et al., 1998). В процессе уточнения модели проводили коррекцию парциальных структурных факторов, исходя из плоской модели объема растворителя, и анизотропного В-фактора. R-фактор для текущей модели был равен 0,260 (фактор R-free, 5% данных, = 0,319) для всех данных в диапазоне разрешения 30-2,5 Å. Статистические данные уточнения систематизированы в таблице 2а.
Текущая модель содержит 296 из 342 аминокислотных остатков, вычисленных исходя из данной конструкции, и точно определенных в большинстве областей полипептидной цепи. Не поддающаяся определению электронная плотность наблюдалась для остатков: 534, 657-673, 790-804 и 863-875.
Анализ структуры с использованием программы PROCHECK (Laskowski et al., 1993) показал, что 12 остатков для четырех молекул в асимметричном элементе находятся в запрещенных областях.
Пример 13 - Сбор данных, определение структуры и уточнение параметров PDE5 дикого типа с силденафилом
Структуру рекомбинантного человеческого PDE5 определяли методом многоволнового аномального рассеяния (MAD) с использованием четырех длин волн у края LIII цинка.
Данные природной рентгеновской дифракции собирали с помощью MAR CCD на станции ВМ14 на ESRF, Grenoble, France. Все данные обрабатывали с использованием пакета программ HKL (Otwinkowski & Minor, 1997). Статистические данные систематизированы в таблице 1b.
Кристаллы принадлежали к пространственной группе Р212121 с размерами элементарной ячейки а=94,179 Å, b=103,6451 Å, с=141,942 Å, α=β=γ=90°. Они содержали 4 молекулы на асимметричный элемент (Mw=39654,71 Да), а вычисленное содержание растворителя составляло 43,23% (VМ=2,18; Matthews, 1968).
Место расположения аномальных тяжелых атомов определяли с использованием программы SOLVE (Terwilliger & Berendzen, 1997) и подтверждали с помощью SnB (Smith et al.,1998). Уточнение параметров тяжелого атома и вычисление фаз осуществляли с использованием программы SHARP (de La Fortelle & Bricogne, 1997). Фазы уточняли за 100 циклов путем "уплощения" растворителя с использованием программы SOLOMON (Abrahams & Leslie, 1996). Полученная карта имела хорошее качество и была использована для выявления примерно 80% структуры с использованием QUANTA (Quanta 98, 1998, version 98.1111; Molecular Simulations Inc., San Diego, CA 92121-3752, USA).
Модель уточняли путем сравнения факторов нативной структуры (Fр-calc), полученный с помощью SHARP исходя из комбинации экспериментальных нативного (Fр) и выведенного (FРН) структурных факторов. Уточнение параметров проводили в диапазоне разрешения 30-2,2 Å с использованием CNX (Brünger et al., 1998) c целевой функцией максимального правдоподобия "mlhl". В процессе уточнения модели проводили коррекцию парциальных структурных факторов, исходя из плоской модели объема растворителя, и анизотропного В-фактора. R-фактор для текущей модели был равен 0,235 (фактор R-free, 5% данных, = 0,28) для всех данных в диапазоне разрешения 30-2,5 Å. Статистические данные уточнения систематизированы в таблице 2b.
Текущая модель содержала 1261 из 1364 аминокислотных остатков, вычисленных исходя из данной конструкции и точно определенных в большинстве областей полипептидной цепи. Не поддающаяся определению электронная плотность наблюдалась для остатков: молекула А: 534-536, 665-681 и 863-875; молекула D: 534, 667-681 и 865-875; молекула В: 534-536, 667 и 865-875; и молекула С: 534-536, 663-678 и 863-875.
Анализ структуры с использованием программы PROCHECK (Laskowski et al., 1993) показал, что только четыре остатка для четырех молекул в ассиметричном элементе находятся в запрещенных областях.
Пример 14 - Сбор данных, определение структуры и уточнение параметров PDE5*
Структуру генетически сконструированного на основе бакуловируса PDE5* определяли методом молекулярного замещения (MR) с использованием координат PDE5*, полученных для комплексов с силденафилом (см. Пример 15).
Данные дифракции рентгеновских лучей регистрировали на лабораторном визуализирующем плоскостном детекторе RaxisIV с вращающимся анодом RU200НВ (Rigaki) и c осмиевыми зеркалами (Blue Osmic mirror)(MSC). Все данные обрабатывали с использованием пакета программ HKL (Otwinkowski & Minor, 1997). Статистические данные систематизированы в таблице 1а.
Кристаллы принадлежали к пространственной группе Р21 с размерами элементарной ячейки а=54,983 Å, b=77,153 Å, с=80,660 Å, α=γ=90° β=101,311°. Они содержали 2 молекулы на асимметричный элемент (Mw=37562 Да), а вычисленное содержание растворителя составляло 45,1% (VМ=2,26; Matthews, 1968).
Молекулярное замещение осуществляли с использованием программы AMORE (CCP4). Полученная карта имела хорошее качество, и структуру корректировали с использованием программы QUANTA. Уточнение проводили в диапазоне разрешения 30-1,6 Å с использованием CNX (Brünger et al., 1998) c целевой функцией максимального правдоподобия "mlf". В процессе уточнения модели проводили коррекцию парциальных структурных факторов, исходя из плоской модели объема растворителя, и анизотропного В-фактора. R-фактор для текущей модели был равен 0,301 (фактор R-free, 5% данных, = 0,326) для всех данных в диапазоне разрешения 30-1,6 Е. Статистические данные уточнения систематизированы в таблице 2а.
Текущая модель содержит 323 остатка на молекулу, 537-858 (остаток Glu 681А был пронумерован с сохранением схемы нумерации PDE5). Не поддающаяся определению электронная плотность наблюдалась для остатков: 534, 535 и 536 в молекулах А или В. Анализ структуры с использованием программы PROCHECK (Laskowski et al., 1993) показал, что только два остатка в двух молекулах в асимметричном элементе находятся в запрещенных областях.
Пример 15 - Сбор данных, определение структуры и уточнение параметров PDE5* с силденафилом
Структуру сконструированного на основе бакуловируса PDE5* определяли методом молекулярного замещения (MR) с использованием объединенной модели PDE5 дикого типа со структурой второго субдомена от PDE4 в качестве поисковой модели.
Данные дифракции рентгеновских лучей регистрировали на лабораторном визуализирующем плоскостном детекторе RaxisIV с вращающимся анодом RU200НВ (Rigaku) и c осмиевыми зеркалами (Blue Osmic mirror) (MSC). Все данные обрабатывали с использованием пакета программ HKL (Otwinkowski & Minor, 1997). Статистические данные систематизированы в таблице 1b.
Кристаллы принадлежали к пространственной группе Р21 с размерами элементарной ячейки а=54,93 Å, b=77,77 Å, с=82,05 Å, α=γ=90° β=100,955°. Они содержали 2 молекулы на асимметричный элемент (Mw=37562,41 Да), и вычисленное содержание растворителя составляло 45,87% (VМ=2,29; Matthews, 1968).
Молекулярное замещение осуществляли с использованием программы AMORE (CCP4). Полученная карта имела хорошее качество, и структуру корректировали с использованием программы QUANTA. Уточнение проводили в диапазоне разрешения 30-1,6 Å с использованием CNX (Brünger et al., 1998) c целевой функцией максимального правдоподобия "mlf". В процессе уточнения модели проводили коррекцию парциальных структурных факторов, исходя из плоской модели объема растворителя, и анизотропного В-фактора. R-фактор для текущей модели был равен 0,286 (фактор R-free, 5% данных, = 0,307) для всех данных в диапазоне разрешения 30-1,6 Å. Статистические данные уточнения систематизированы в таблице 2b.
Текущая модель содержит 323 остатка на молекулу, 537-858 (остаток Glu 681А был пронумерован с сохранением схемы нумерации PDE5). Не поддающаяся определению электронная плотность наблюдалась для остатков: 534, 535 и 536 в молекулах А или В. Анализ структуры с использованием программы PROCHECK (Laskowski et al., 1993) показал, что только два остатка в двух молекулах в асимметричном элементе находятся в запрещенных областях.
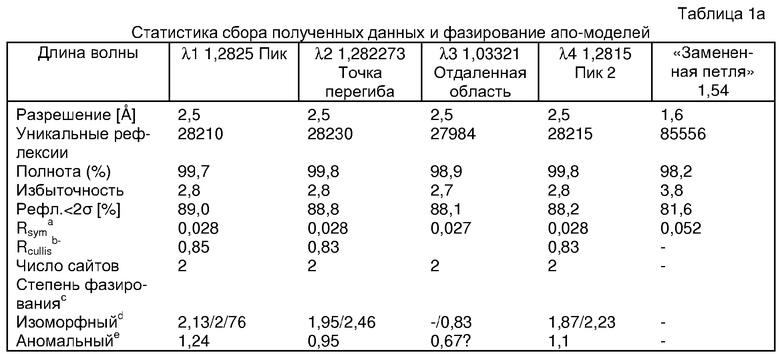
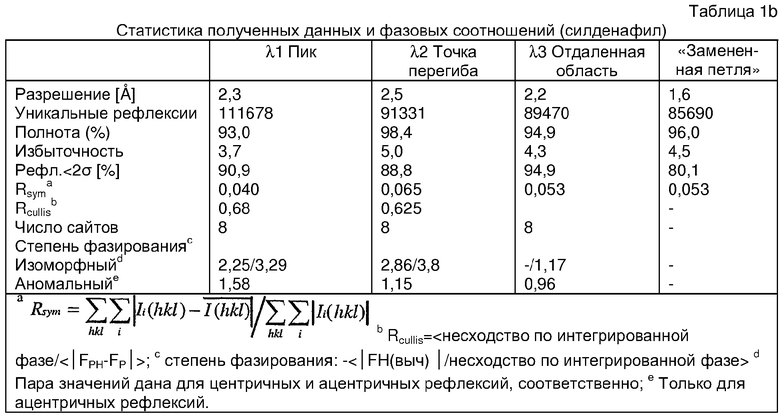
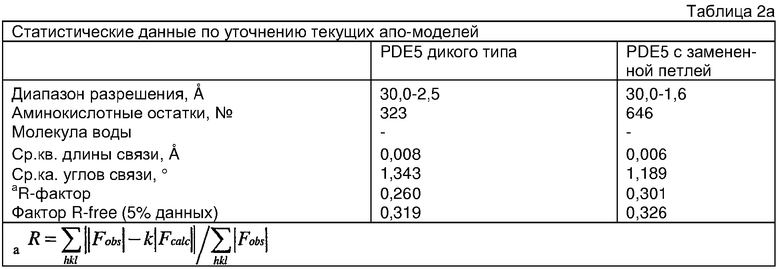
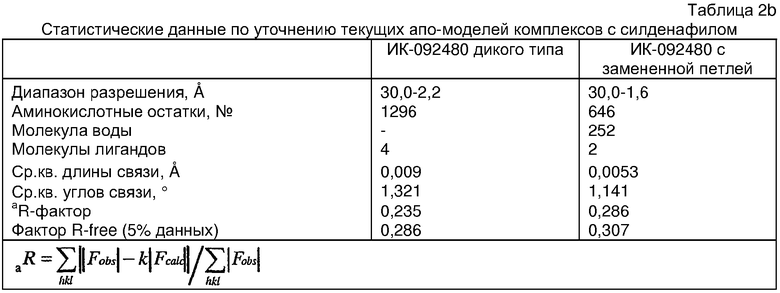
Библиография
- Abrahams, J.P. & Leslie, A. (1996) Acta Crystallogr. D 52, 30-42.
- Aravind, L. & Ponting, C. P. (1997) TIBS. 22, 458-459.
- Briinger, A. et al. (1998), Acta Crystallogr. D 54, 905-921.
- Butcher, R.W. & Sutherland, E.W. (1962) J. biol. Chem. 237, 1244-1250.
- Charbonneau, H, (1990) in Cyclic Nucleotide Phosphodiesterases: Structure, Regulation and Drug Action (Beavo, J., and Houslay, M.D., eds) pp. 267-296, John. Wiley & Sons, Inc. New York.
- Charbonneau. H. et al., (1986) Proc. Natl. Acad. Sci. U. S. A.. 83, 9308-9312.
- Collaborative Computational Project Number 4 (1994) Acta Crystallogr. D 50, 760-763.
- Corbin, J.D. & Francis, S.H. (1999) J. Biol. Chein. 274, 13729-13732.
- de La Portelle, E. & Bricogne, G. (1997) Methods Enzyvwi. 276, 472-494.
- Francis, S.H. et al. (1994) J. Biol. Chem. 269, 22477-22480.
- Goldberg, N.D. et al (1980) J. Biol. Chem. 255, 10344-10347.
- Hendrickson, W. A. et al. (1989) Basic Life Sci. 51 (Synchrotron Radiat. Struct. Biol.), 317-324.
- Jiang, H. et al. (1992) J. Biol. Chem. 267, 1015-1019.
- Laskowski, R. A. et al. (1993) J. Appl. Crystallogr. 26, 283-291.
- Lohman, S.M. et al. (1997) Trends in Biochem. Sci. 22, 307-312.
- Mardnez, S. et al. (2001) Poster presented at the American Crystallography Association annual meeting, Los Angeles, USA.
- Matthews, B.W. (1968) J. Mol. Biol. 33, 491-497.
- Nicholls, A. et al. (1993) J. Biophys, A166.
- Otwinowski, Z. & Minor, W. (1997) Methods Enzymol. 276, 307-326.
- Quanta 98, 1998, version 98.1111; Molecular Simulations Inc., San Diego, CA 92121-3752.
- Rail, T.W. & Sutherland, E.W. (1958) J. Biol. Chem. 232, 1065-1076.
- Soderling, S.H. & Beavo, J.A., (2000) Curr. Opin. Cell Biol. 12. 174-179.
- Smith, G.D. et al. (1998) Acta Crystallographica Section D-Biological Crystallography. 54 (Pt 5):799-804
- Terwilliger, T.C. & Berendzen, J. (1997) Acta Crystallogr. D 53, 571-579
Crystallogr. D 54, 799-804.
- Thomas et al. (1990) J. Biol. diem. 265, 14971-14978.
- Xu, R. X. et al. (2000) Science 288, 1822-1825.
Сокращения
| название | год | авторы | номер документа |
|---|---|---|---|
| ДОМЕН СВЯЗЫВАНИЯ БАКТЕРИАЛЬНОЙ АТР-СИНТАЗЫ | 2005 |
|
RU2418001C2 |
| ПОЛИПЕПТИД, ИМЕЮЩИЙ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ ФОСФОДИЭСТЕРАЗЫ | 1998 |
|
RU2272841C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, КОДИРУЮЩАЯ ПОЛИПЕПТИД, КОТОРЫЙ СВЯЗЫВАЕТСЯ С MORT-1 (ВАРИАНТЫ), ПОЛИПЕПТИД (ВАРИАНТЫ) И СПОСОБ ЕГО ПОЛУЧЕНИЯ, ВЕКТОР, СПОСОБ МОДУЛИРОВАНИЯ ДЕЙСТВИЯ ЛИГАНДА FAS-R ИЛИ TNF НА КЛЕТКИ (ВАРИАНТЫ), СПОСОБ ОБРАБОТКИ КЛЕТОК, СПОСОБ ВЫДЕЛЕНИЯ И ИДЕНТИФИКАЦИИ БЕЛКОВ, СПОСОБ МОДУЛИРОВАНИЯ ИНДУЦИРУЕМОГО MORT-1 ДЕЙСТВИЯ ИЛИ ИНДУЦИРУЕМОГО MORT-1 СВЯЗЫВАЮЩИМ БЕЛКОМ ДЕЙСТВИЯ НА КЛЕТКИ | 1996 |
|
RU2235779C2 |
| НОВЫЙ ПЕПТИД-ИНГИБИТОР PI3Kγ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ОРГАНОВ ДЫХАНИЯ | 2015 |
|
RU2704826C2 |
| БЕЛКИ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2549678C2 |
| БЕЛКИ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2673724C2 |
| ДНК-ВАКЦИНЫ ПРОТИВ ОПУХОЛЕВОГО РОСТА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2343195C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, СОСТОЯНИЙ ИЛИ ПРОЦЕССОВ, ХАРАКТЕРИЗУЕМЫХ АБЕРРАНТНОЙ ПРОЛИФЕРАЦИЕЙ ФИБРОБЛАСТОВ И ОТЛОЖЕНИЕМ ВНЕКЛЕТОЧНОГО МАТРИКСА | 2012 |
|
RU2620066C2 |
| НЕПРИРОДНЫЕ СЕМАФОРИНЫ КЛАССА 3 И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2722832C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ЛЕГОЧНОЙ ГИПЕРТЕНЗИИ | 2017 |
|
RU2748278C2 |
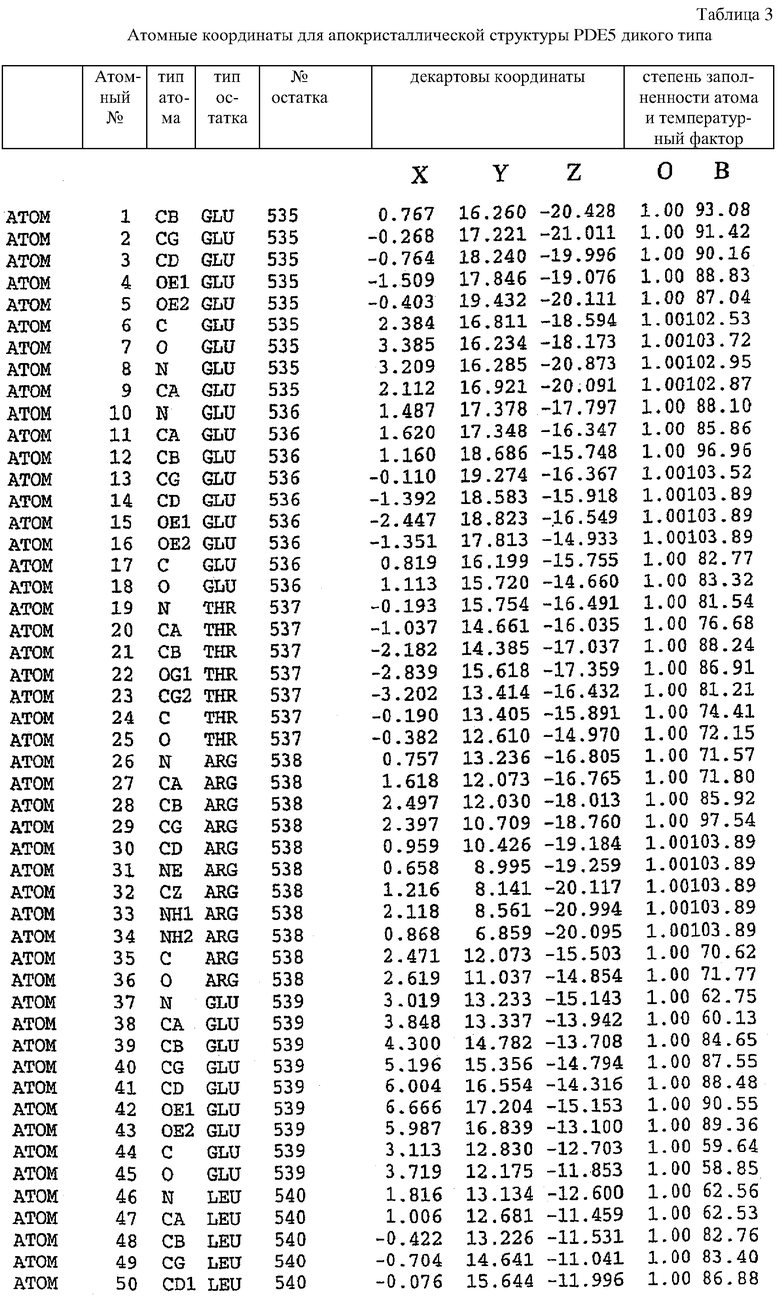
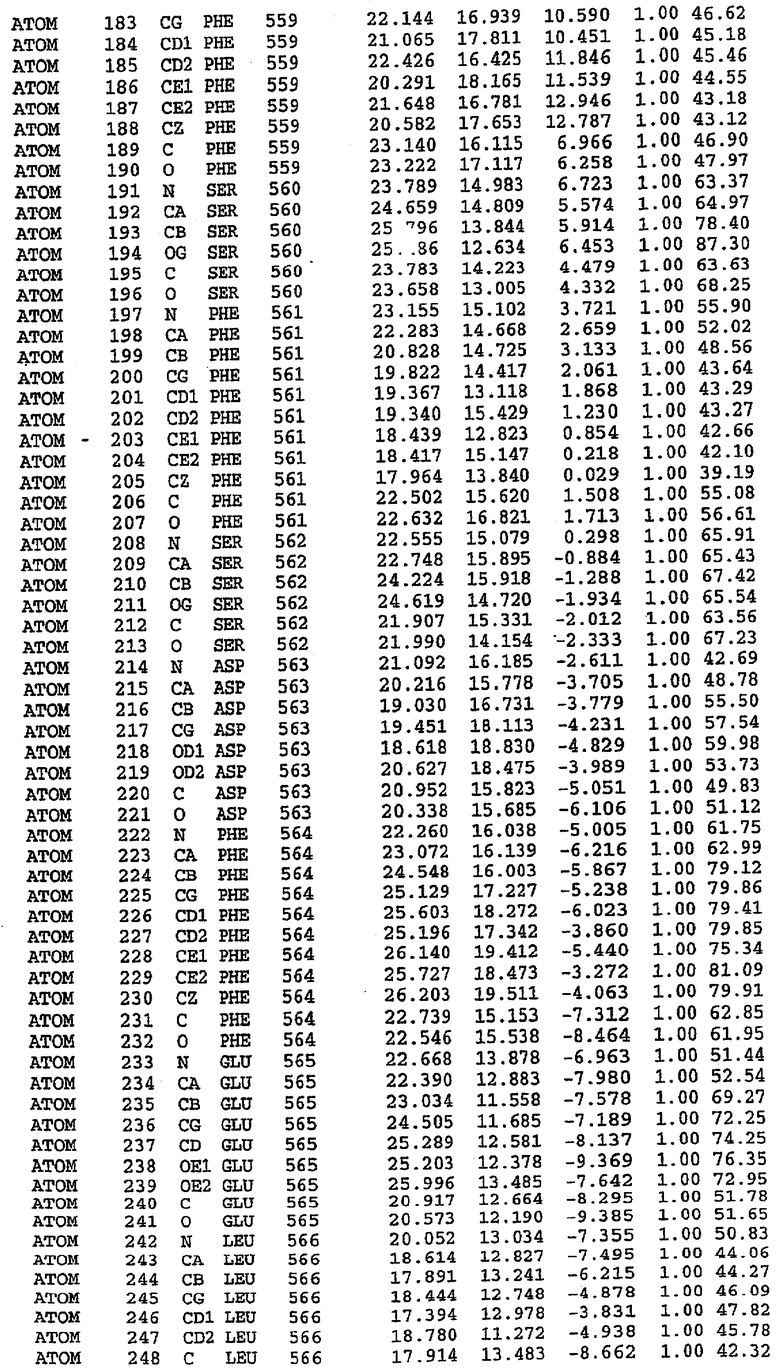
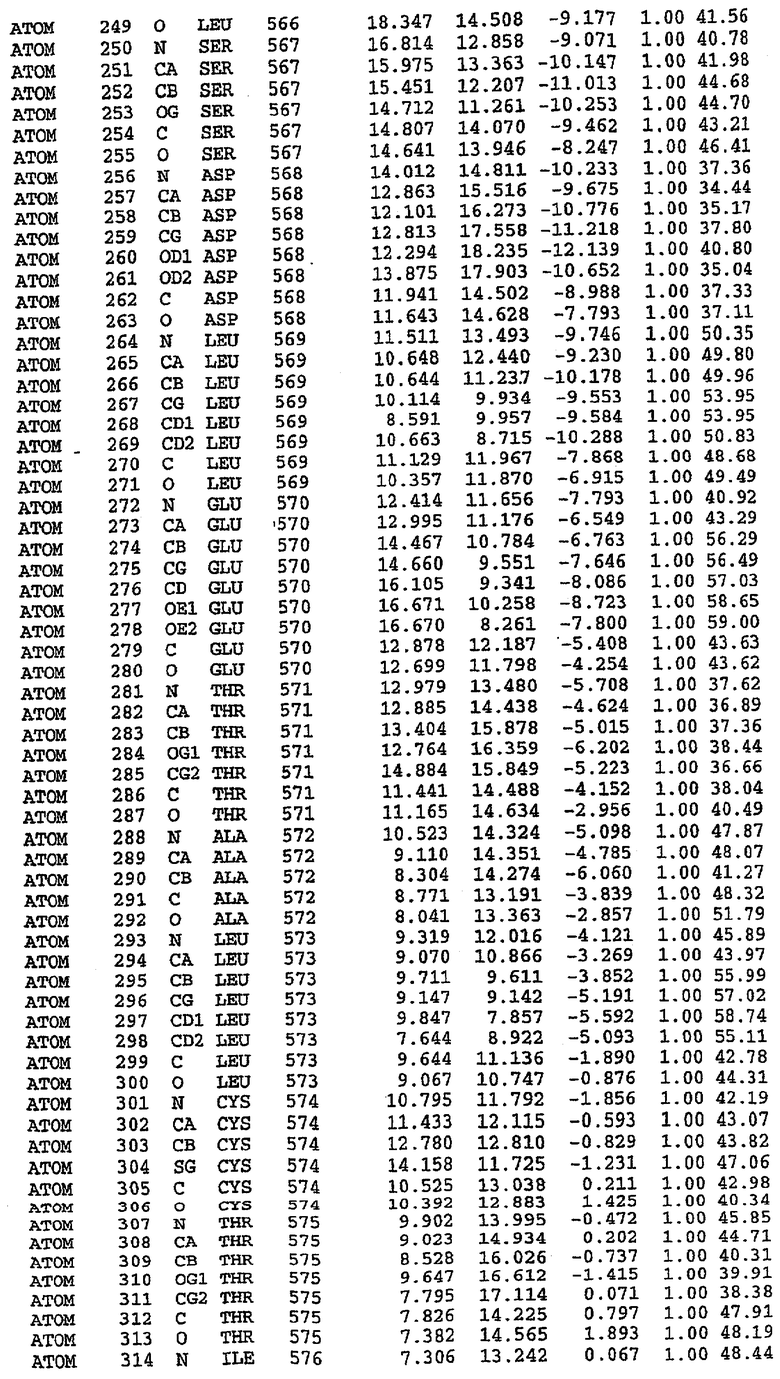
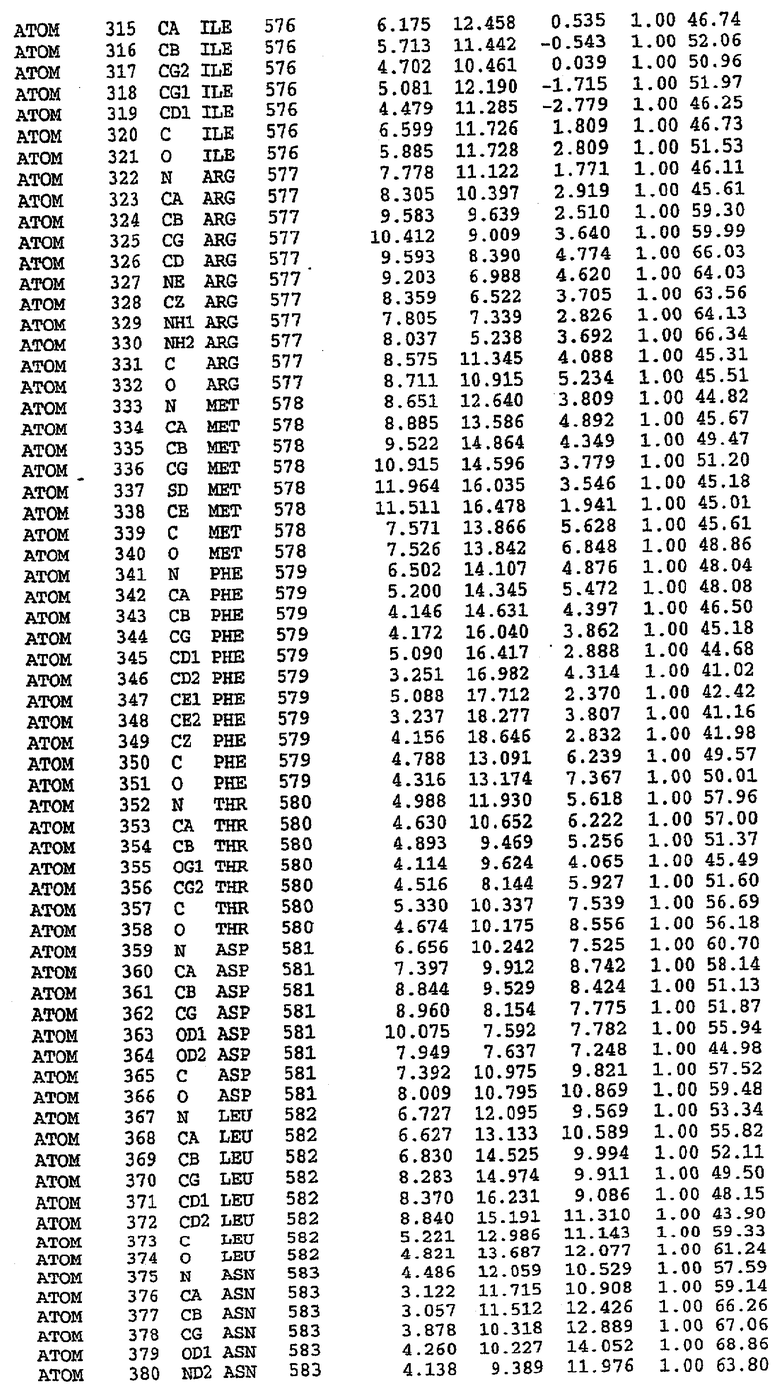
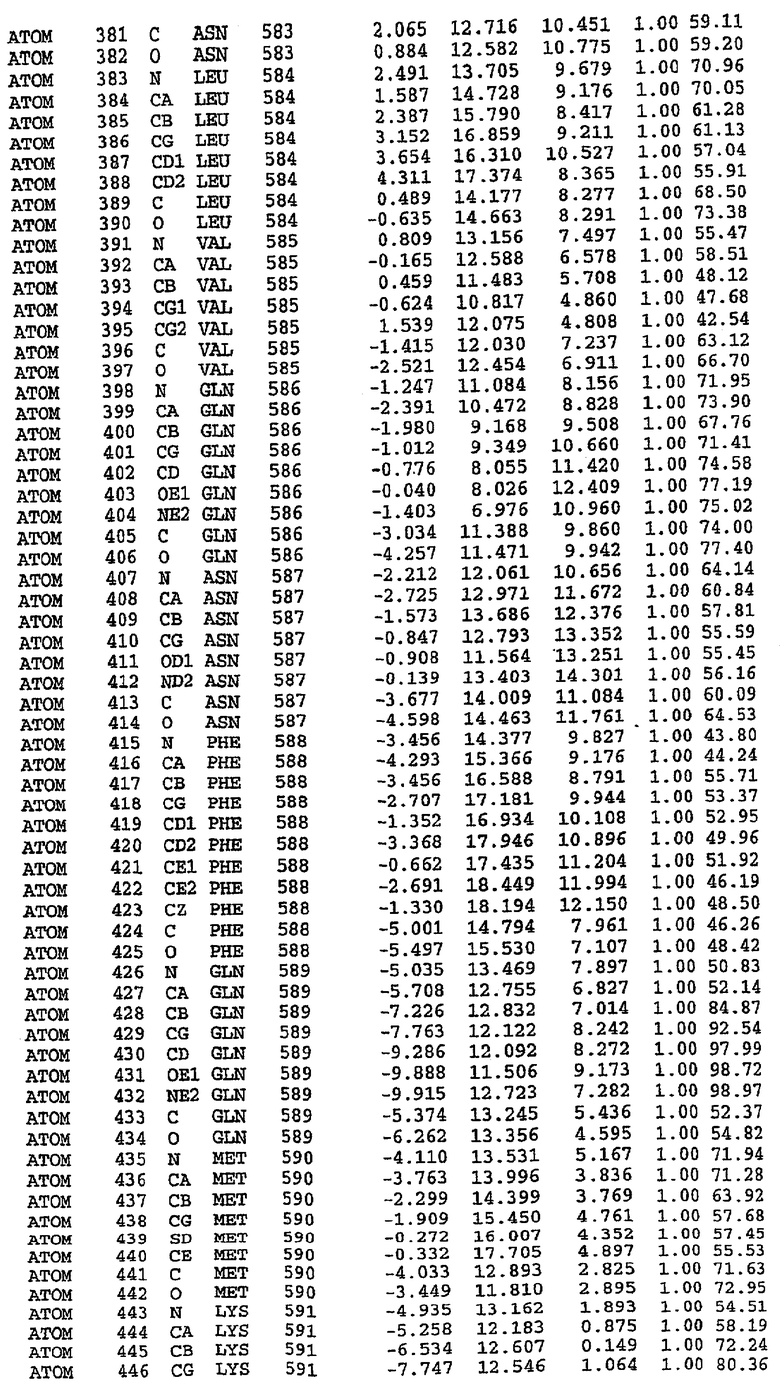
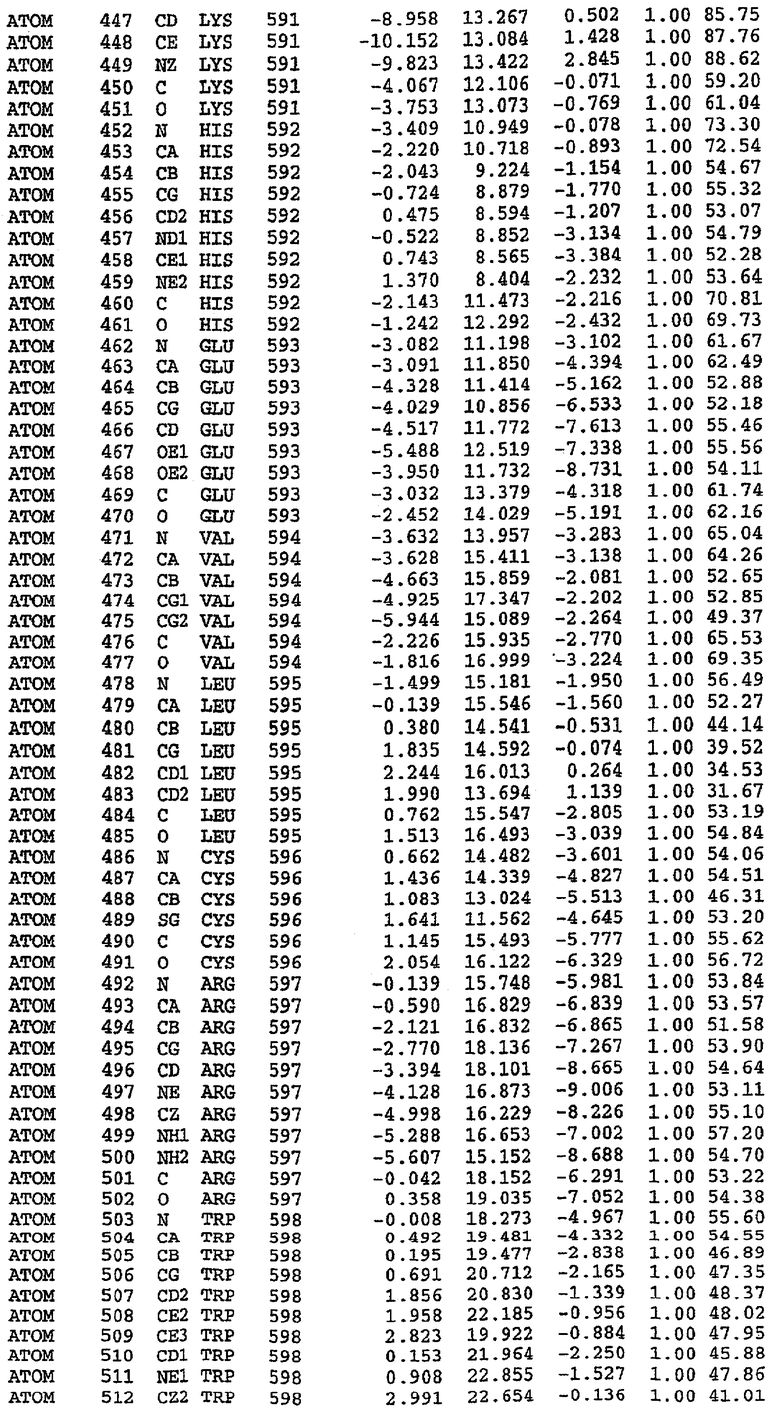
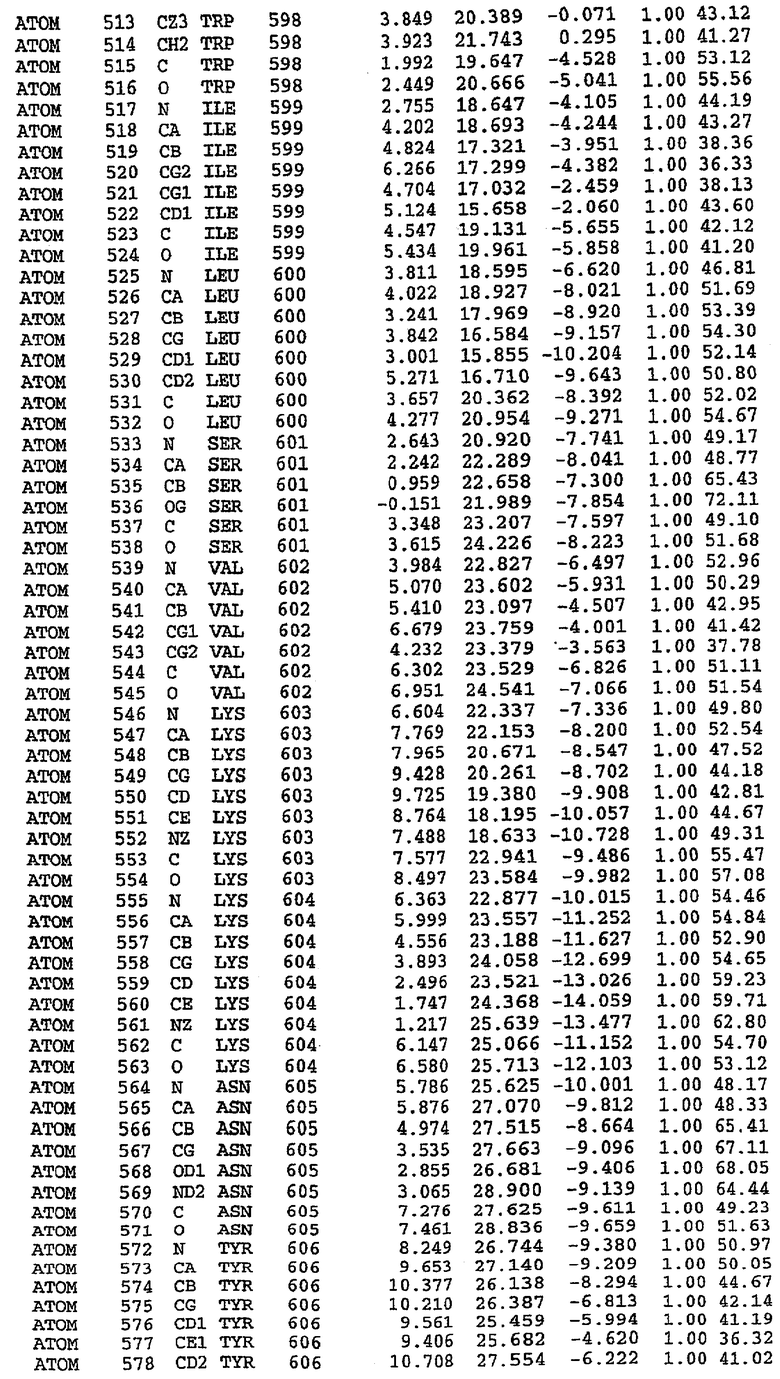
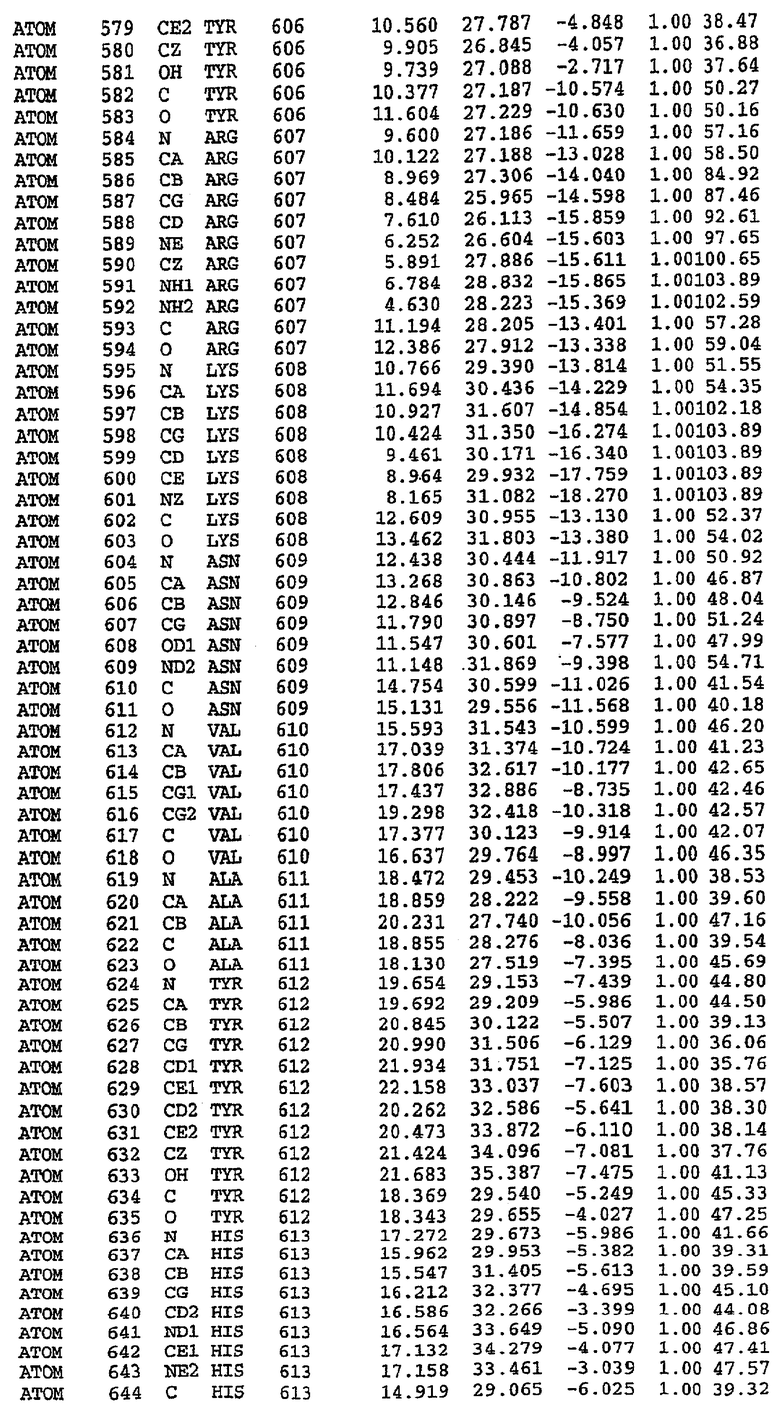
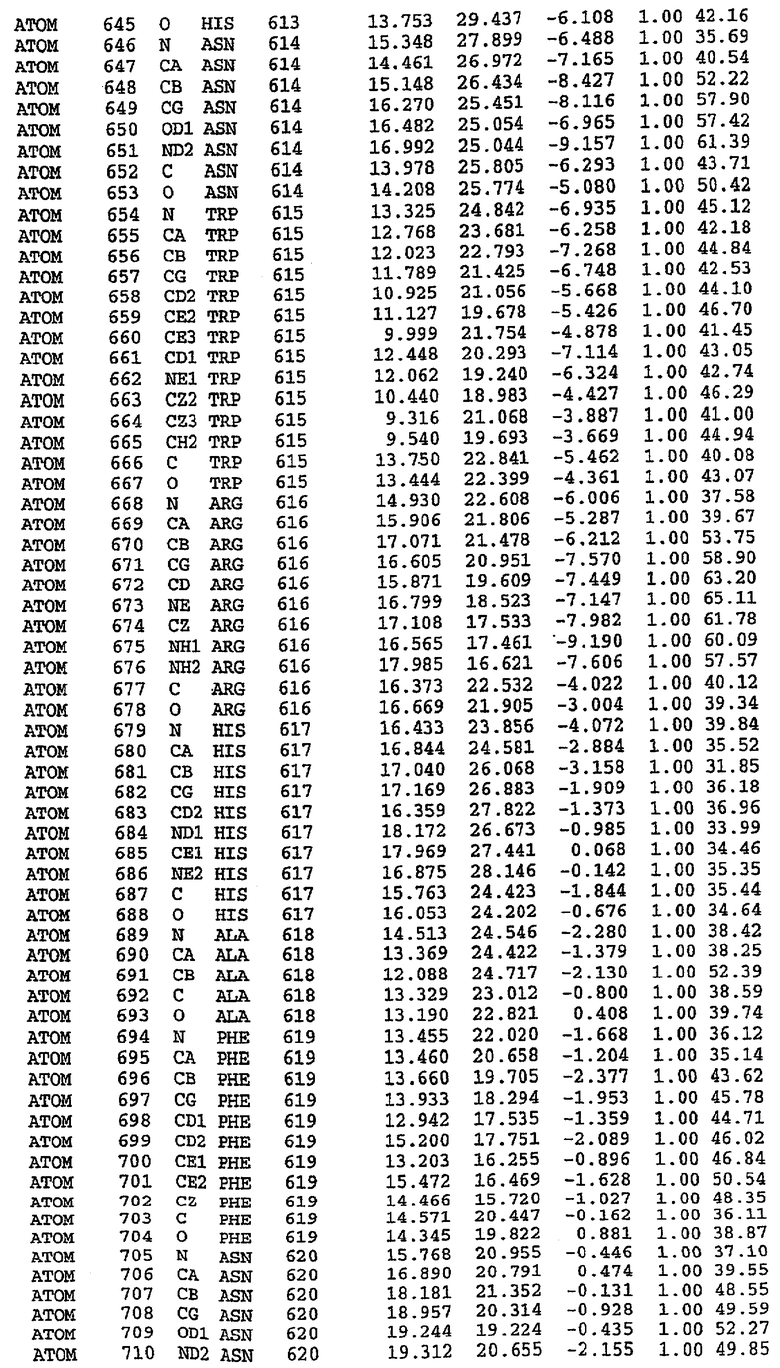
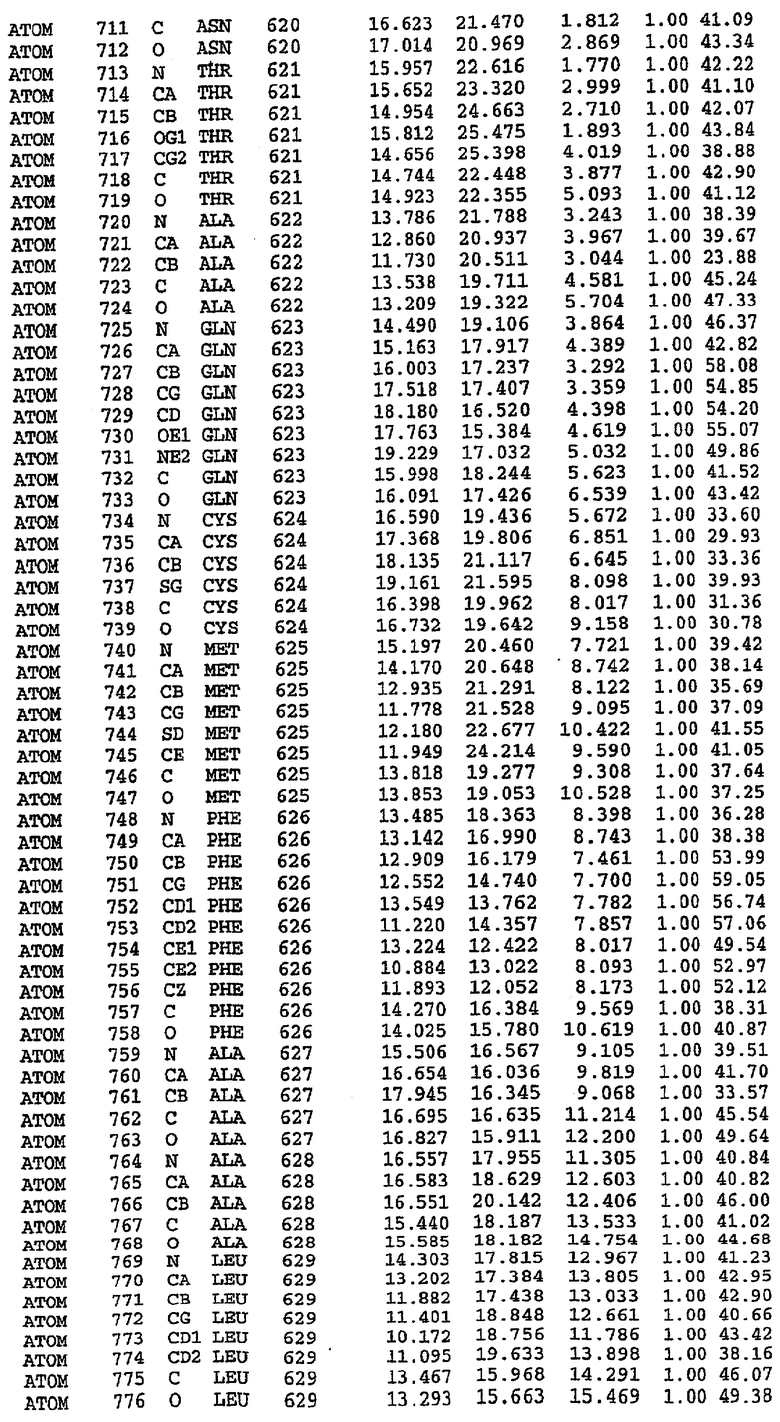
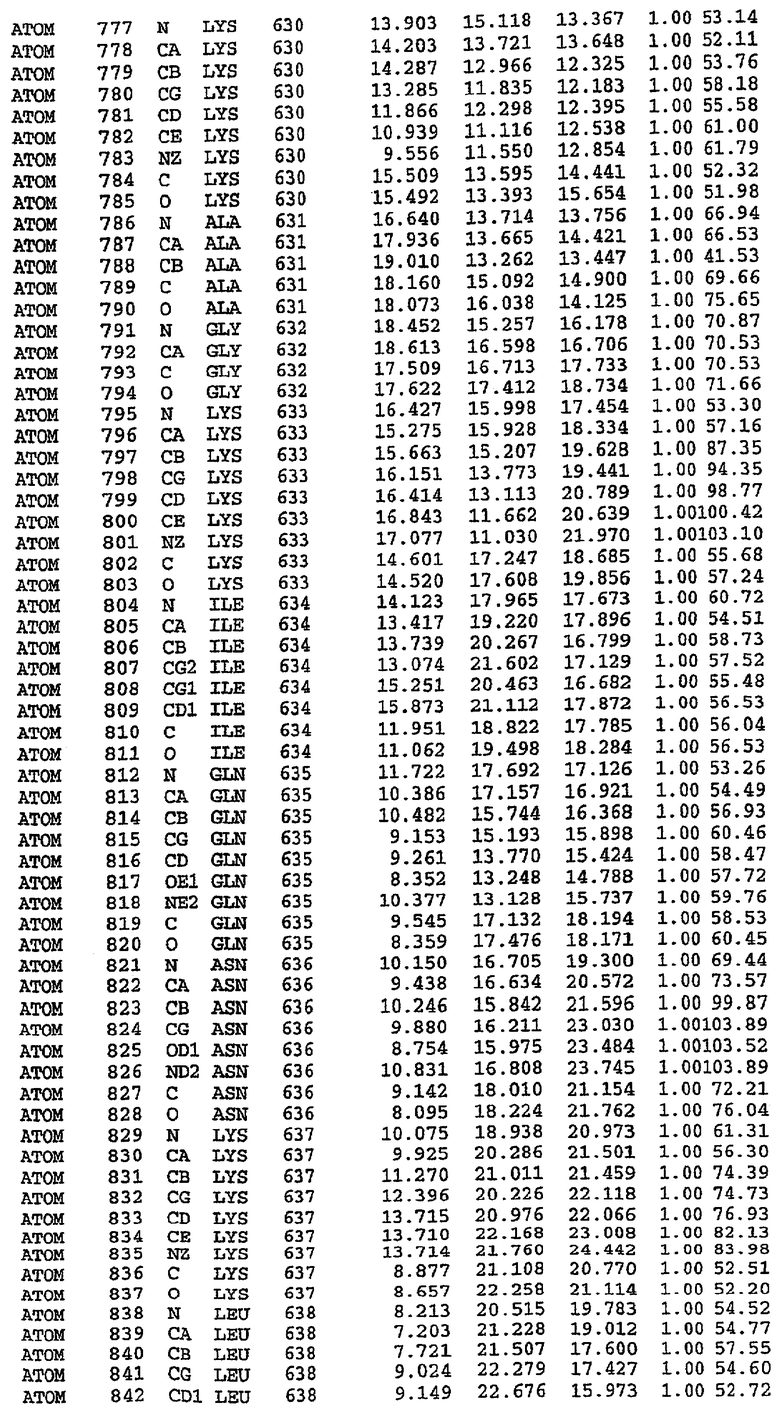
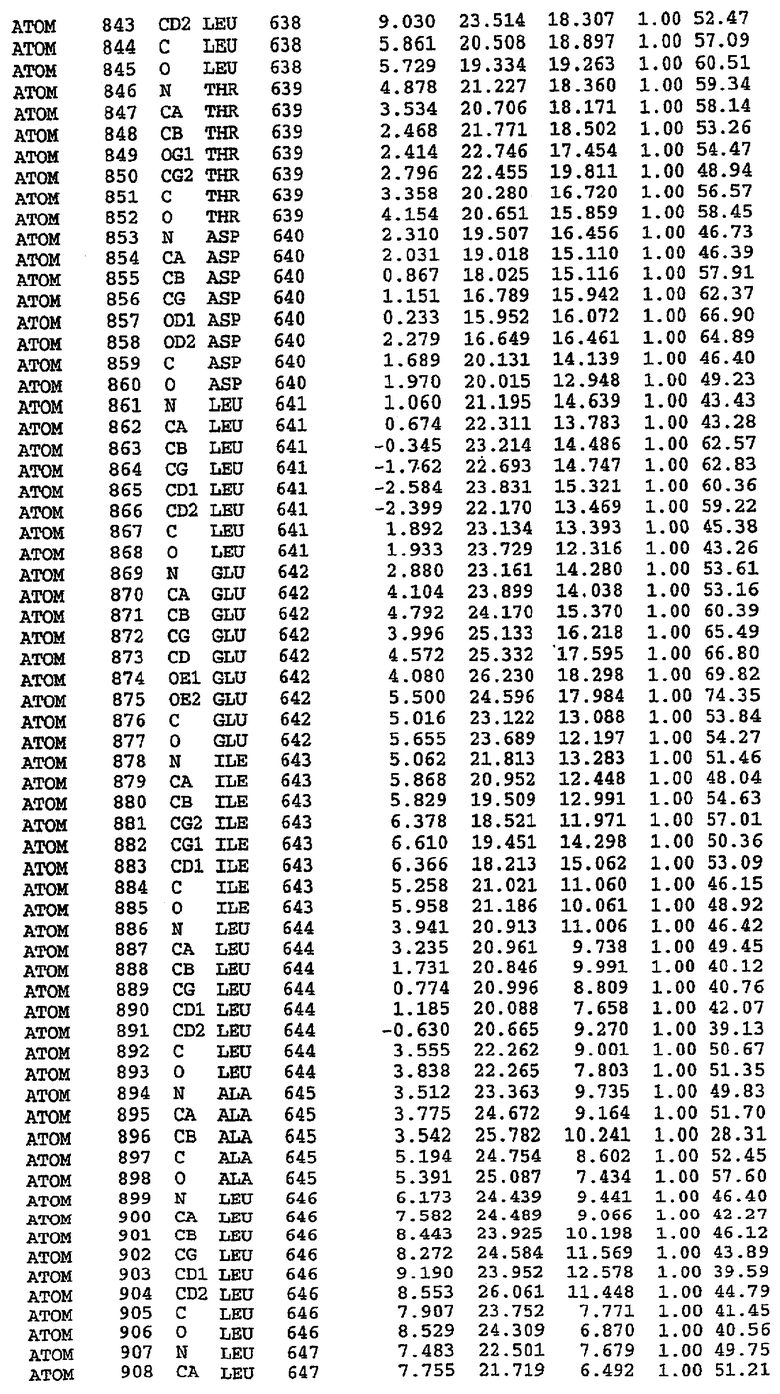
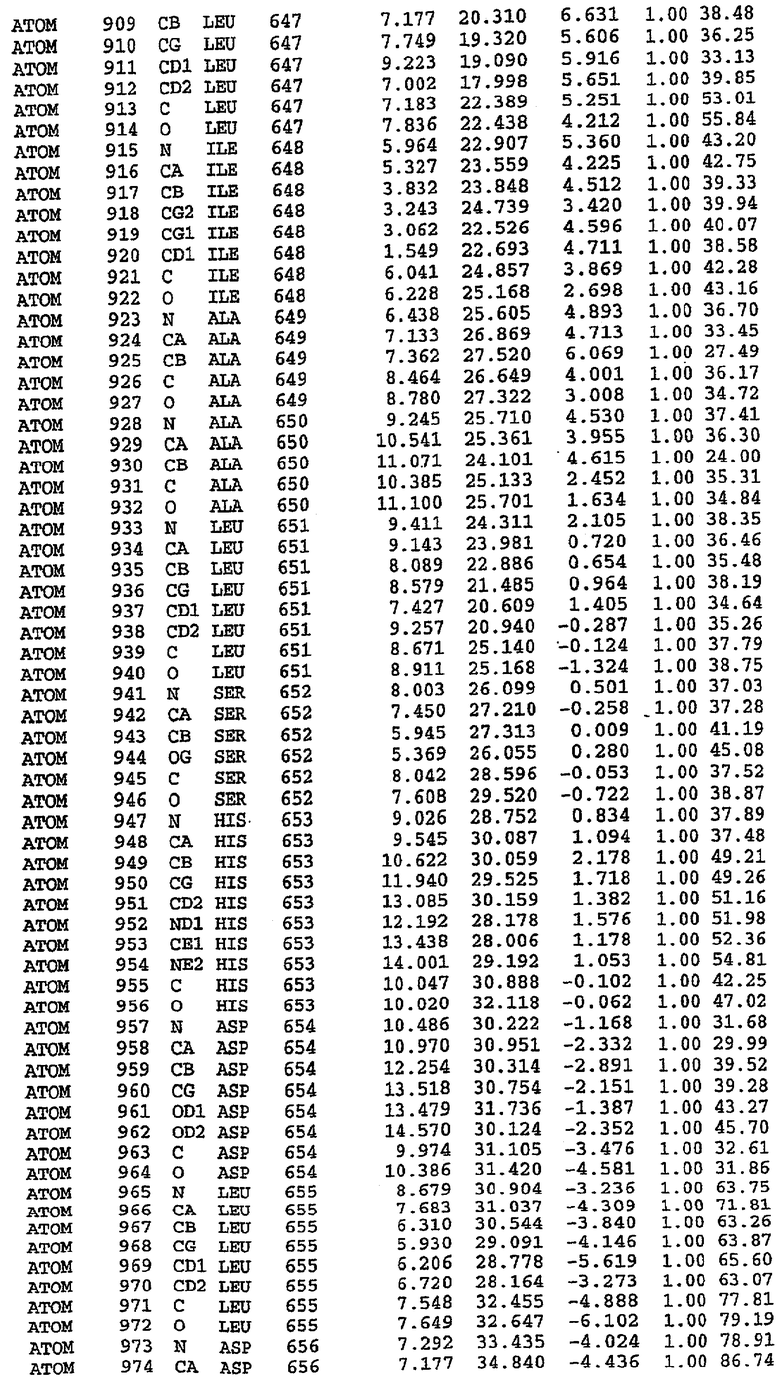
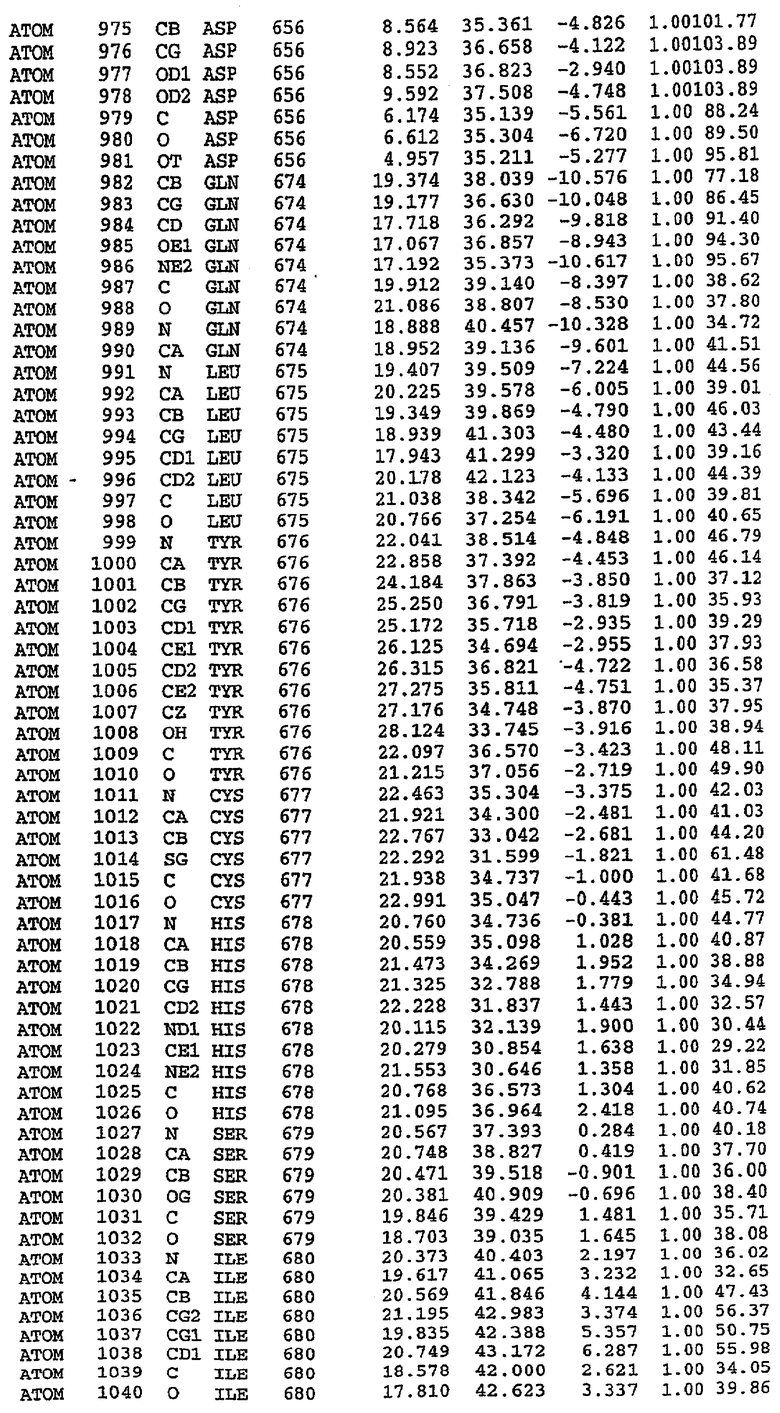
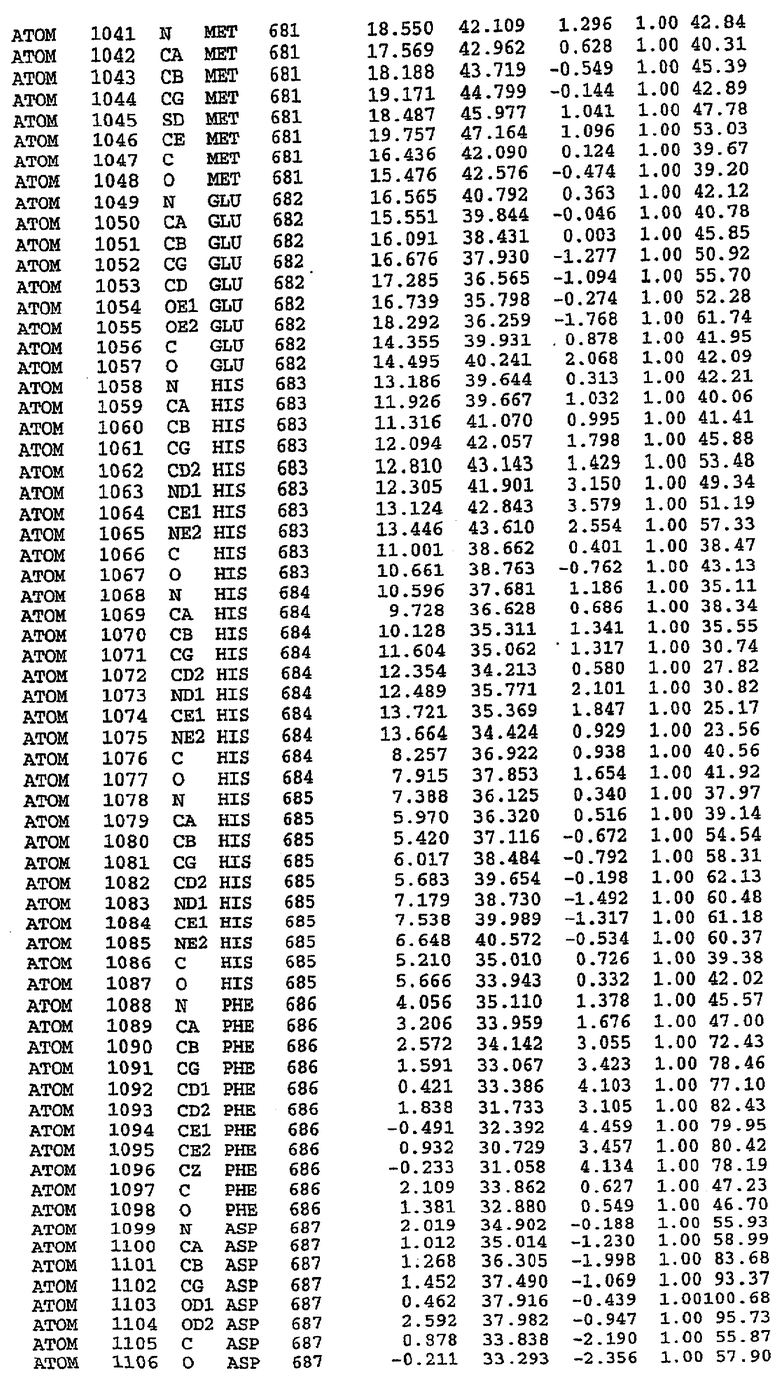
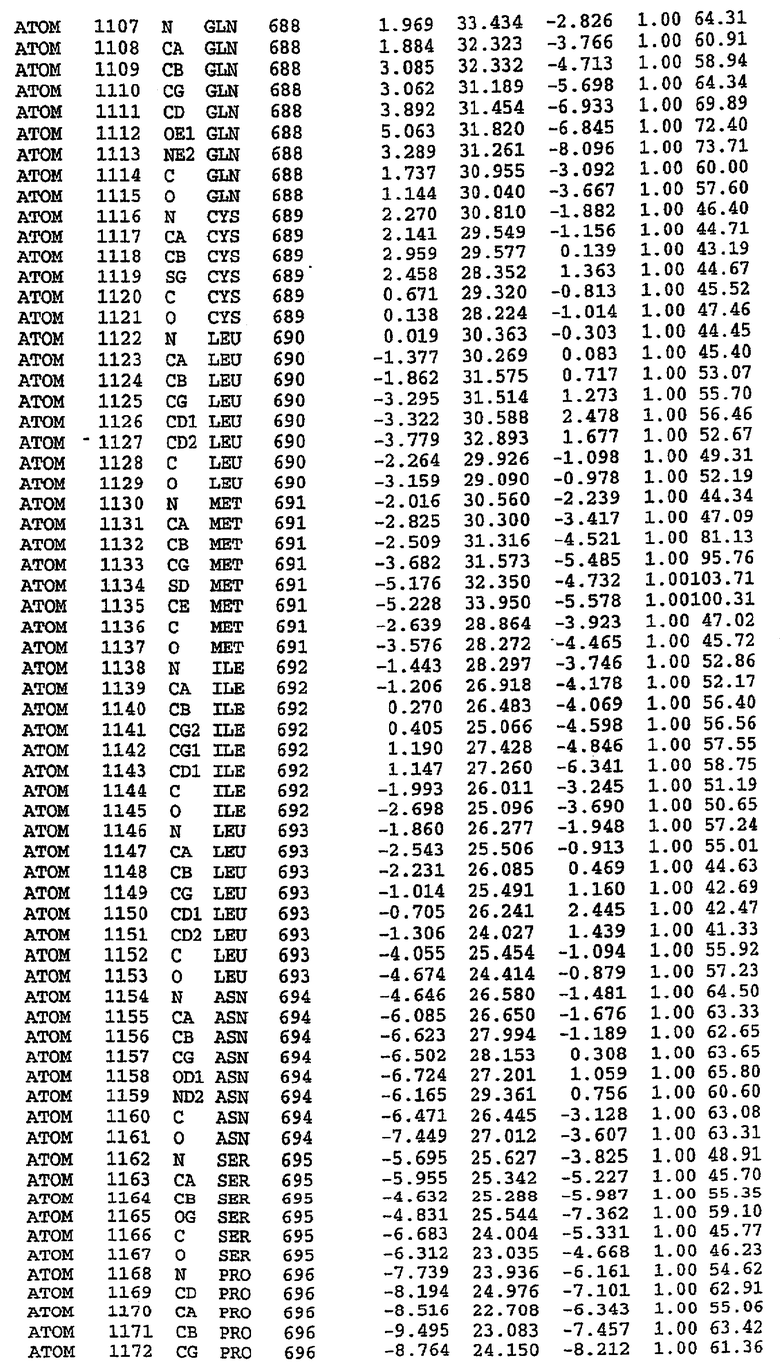
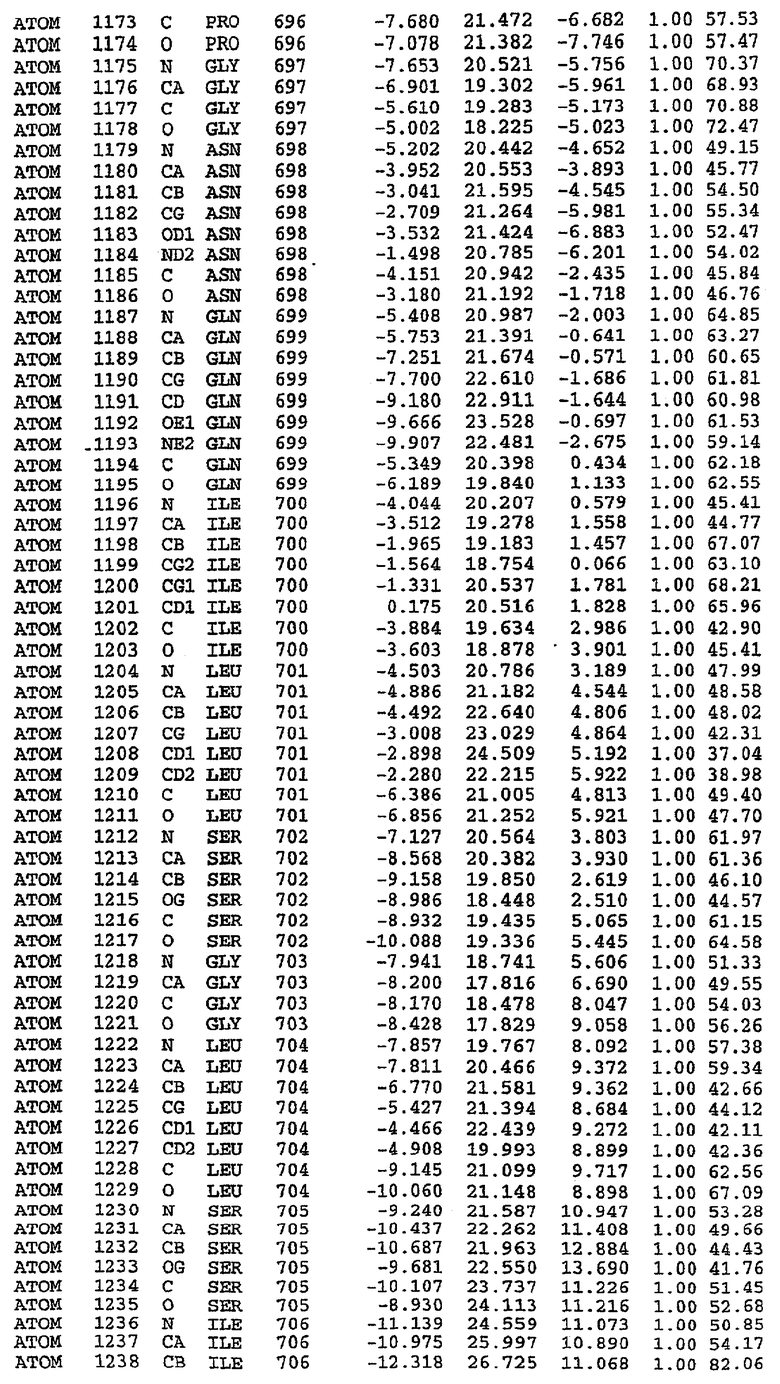

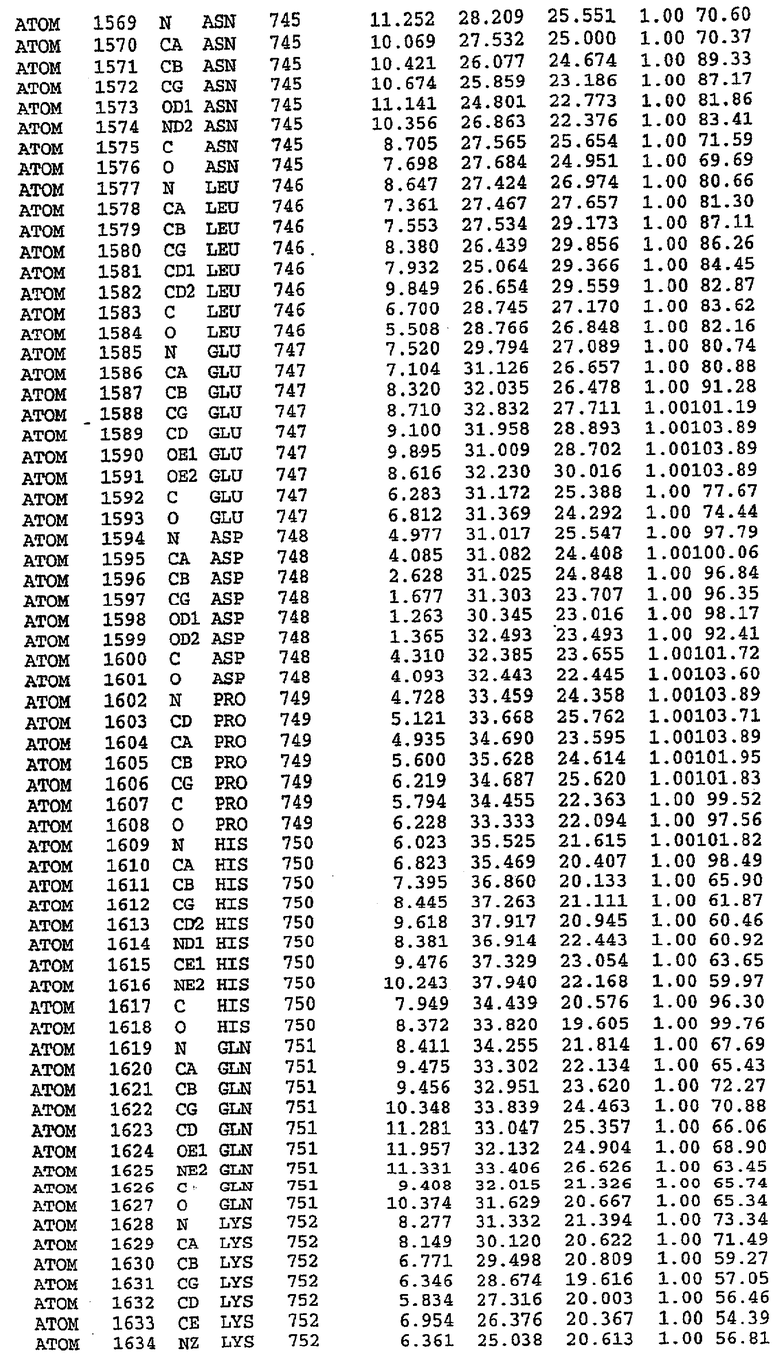
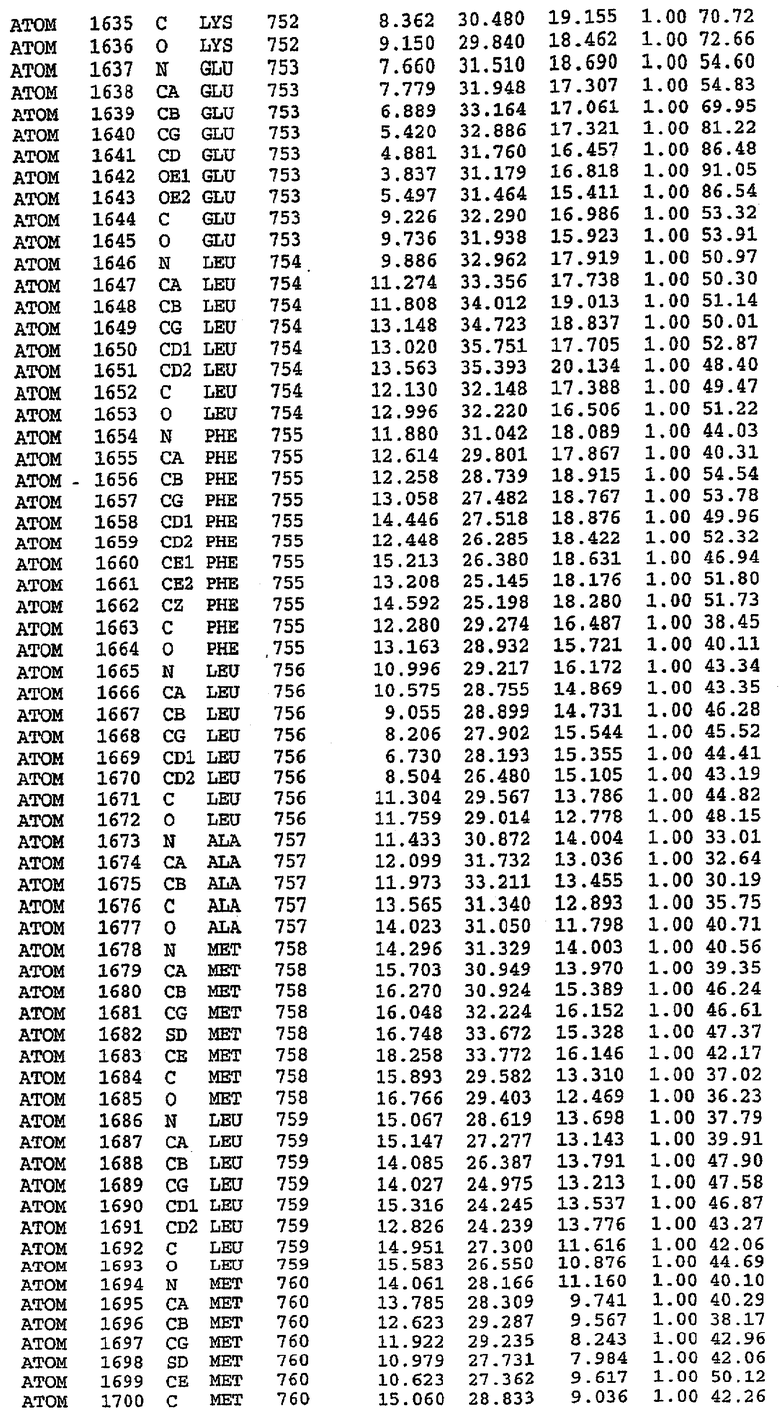
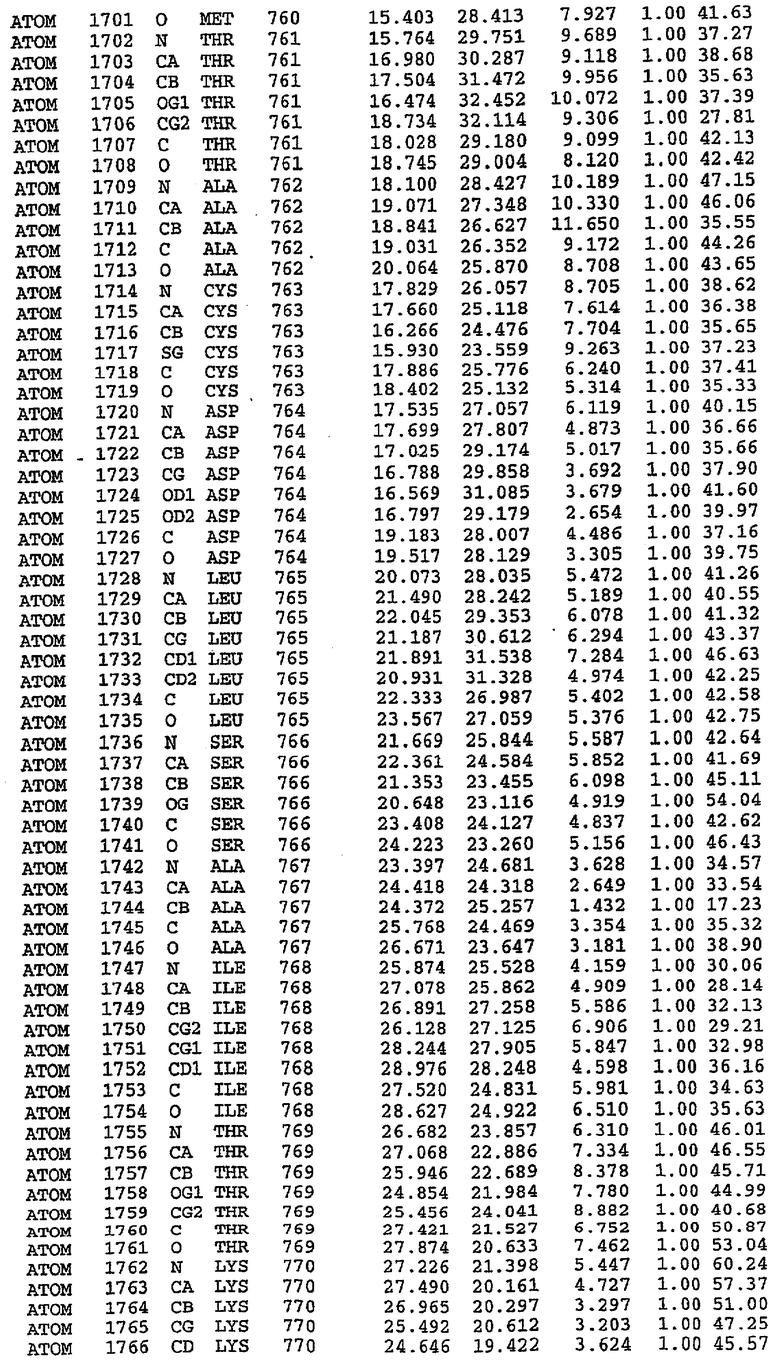
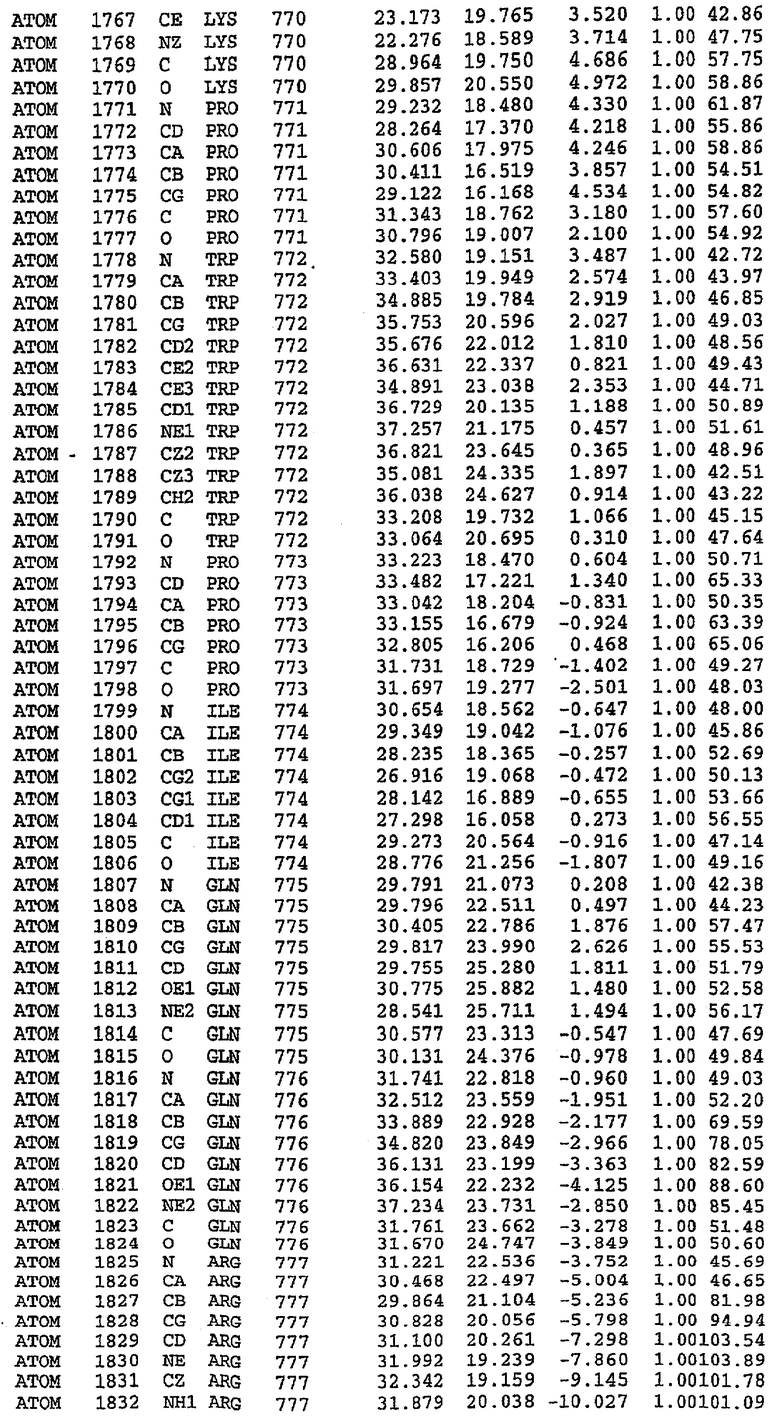
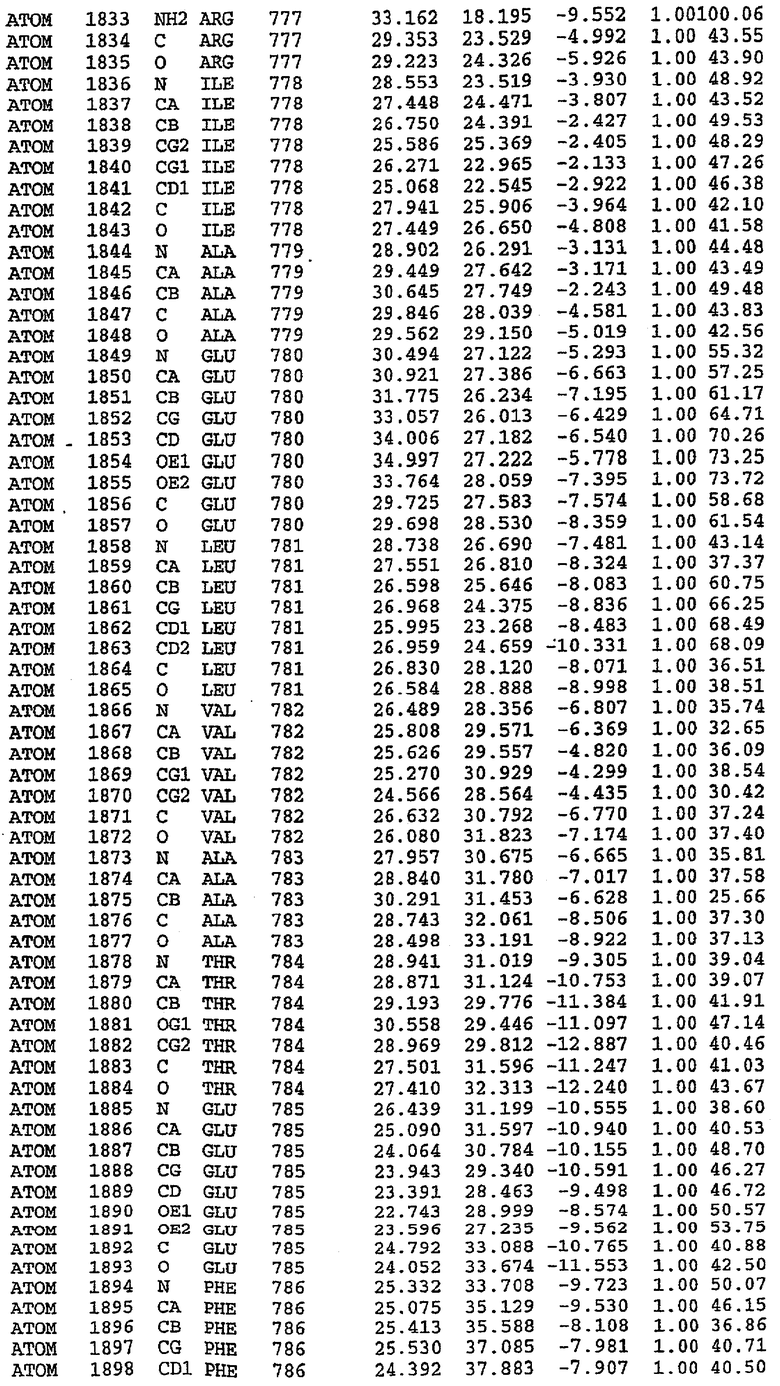
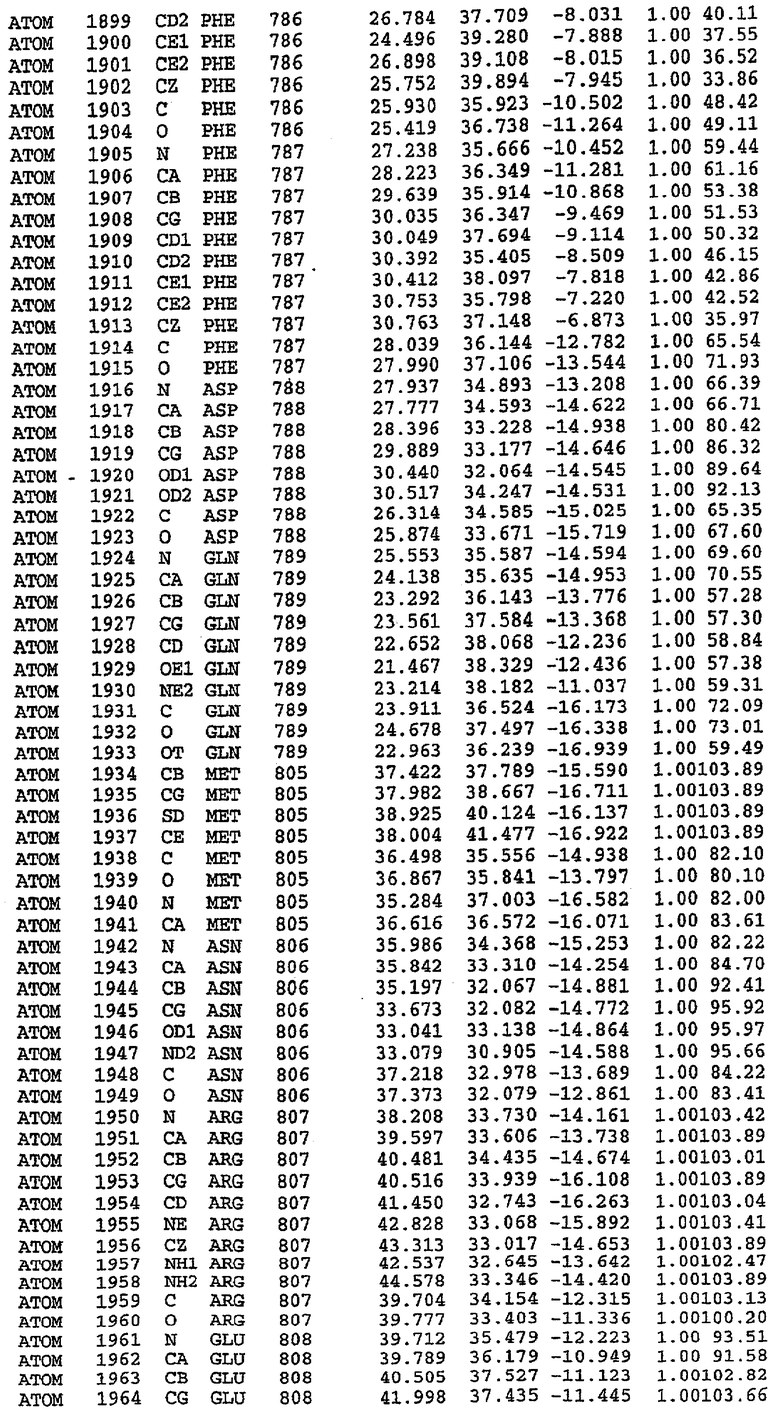
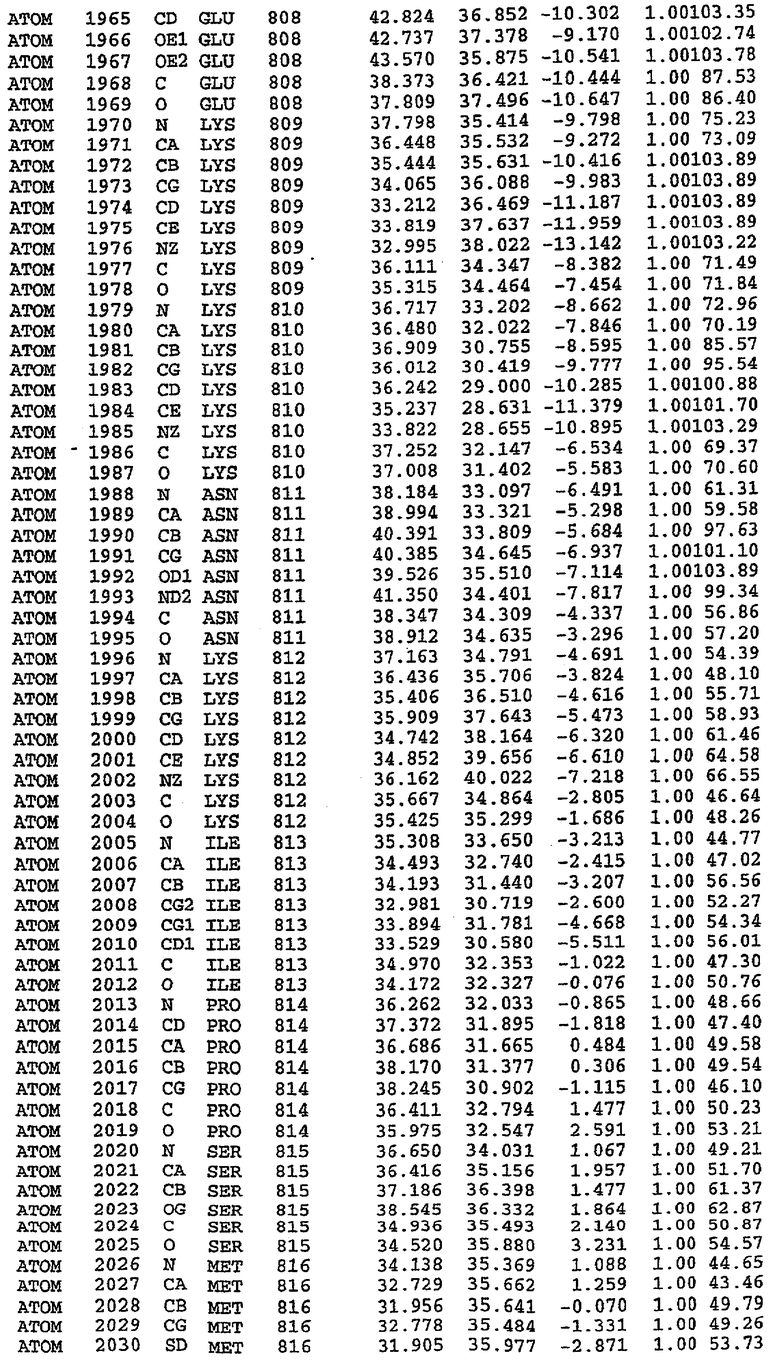
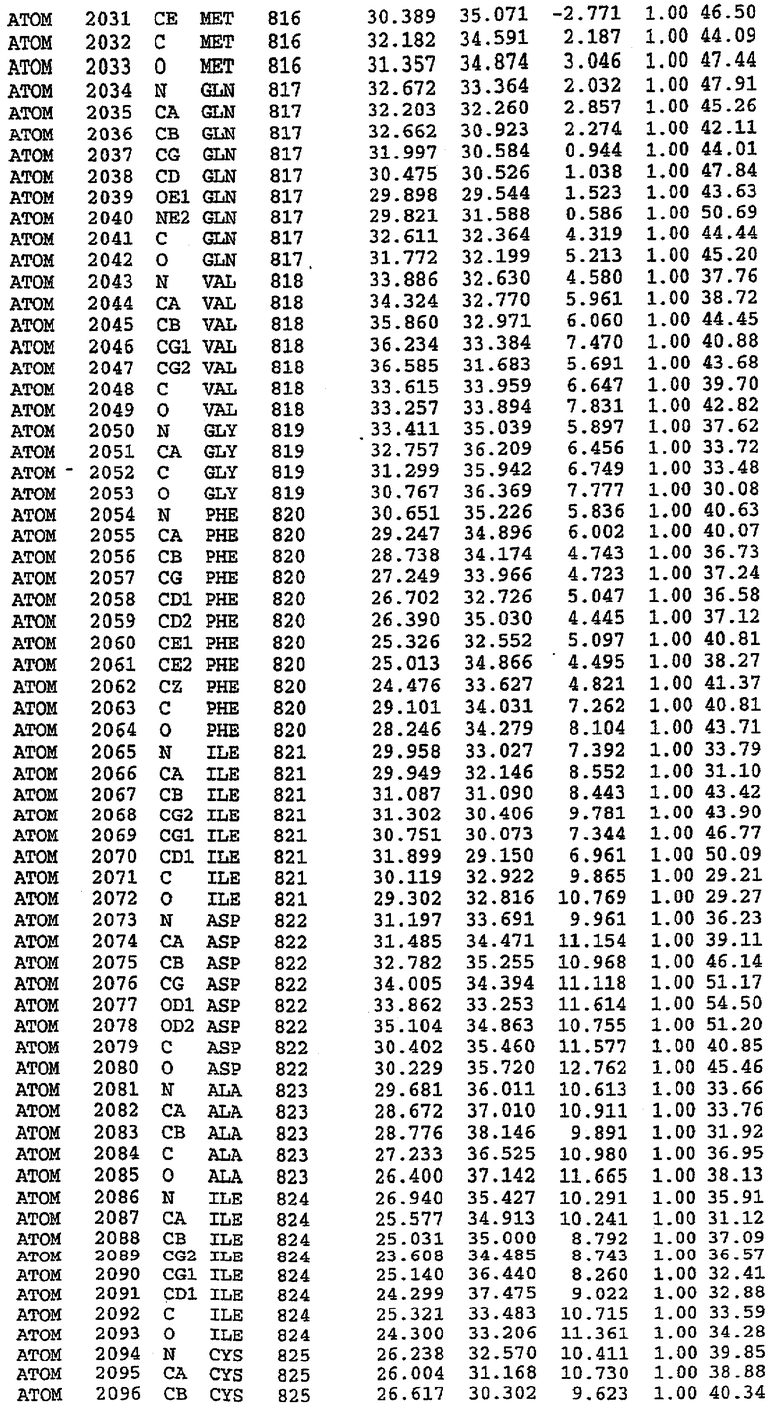
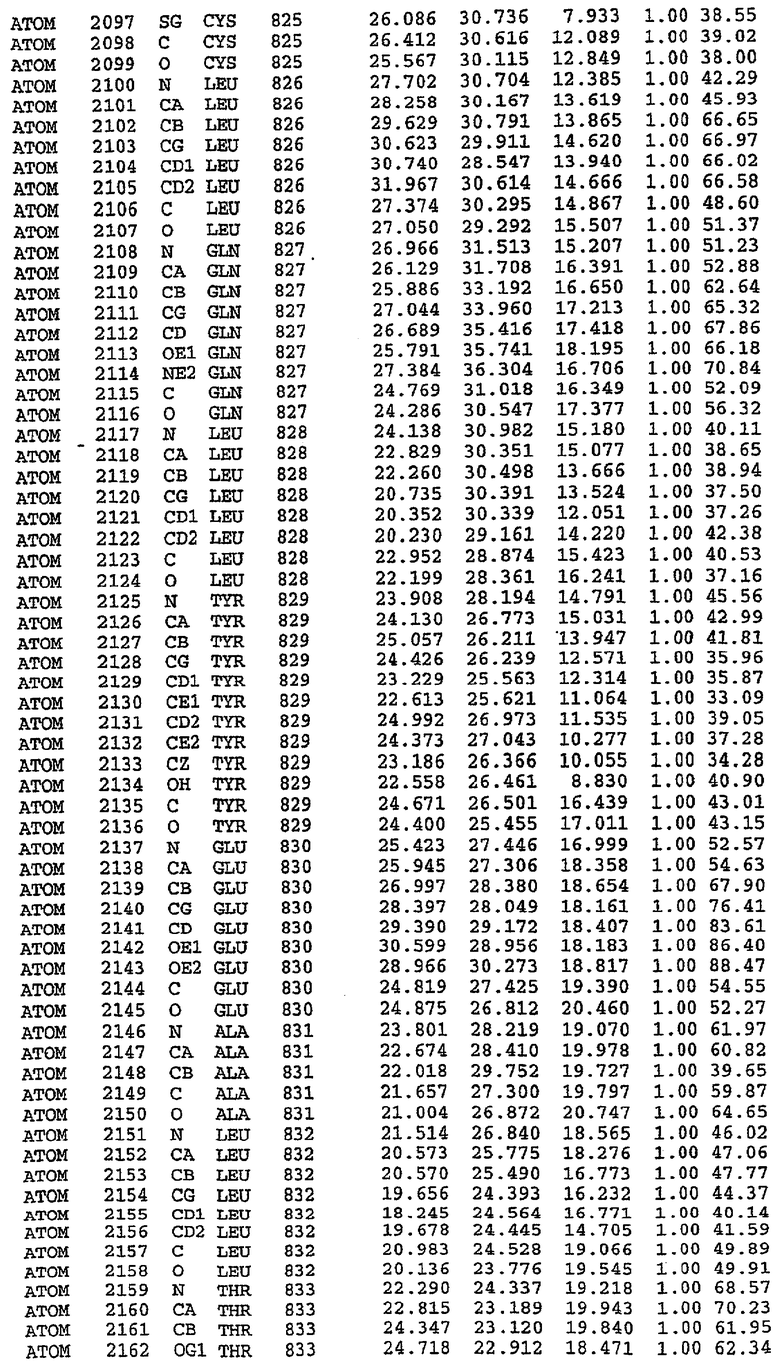
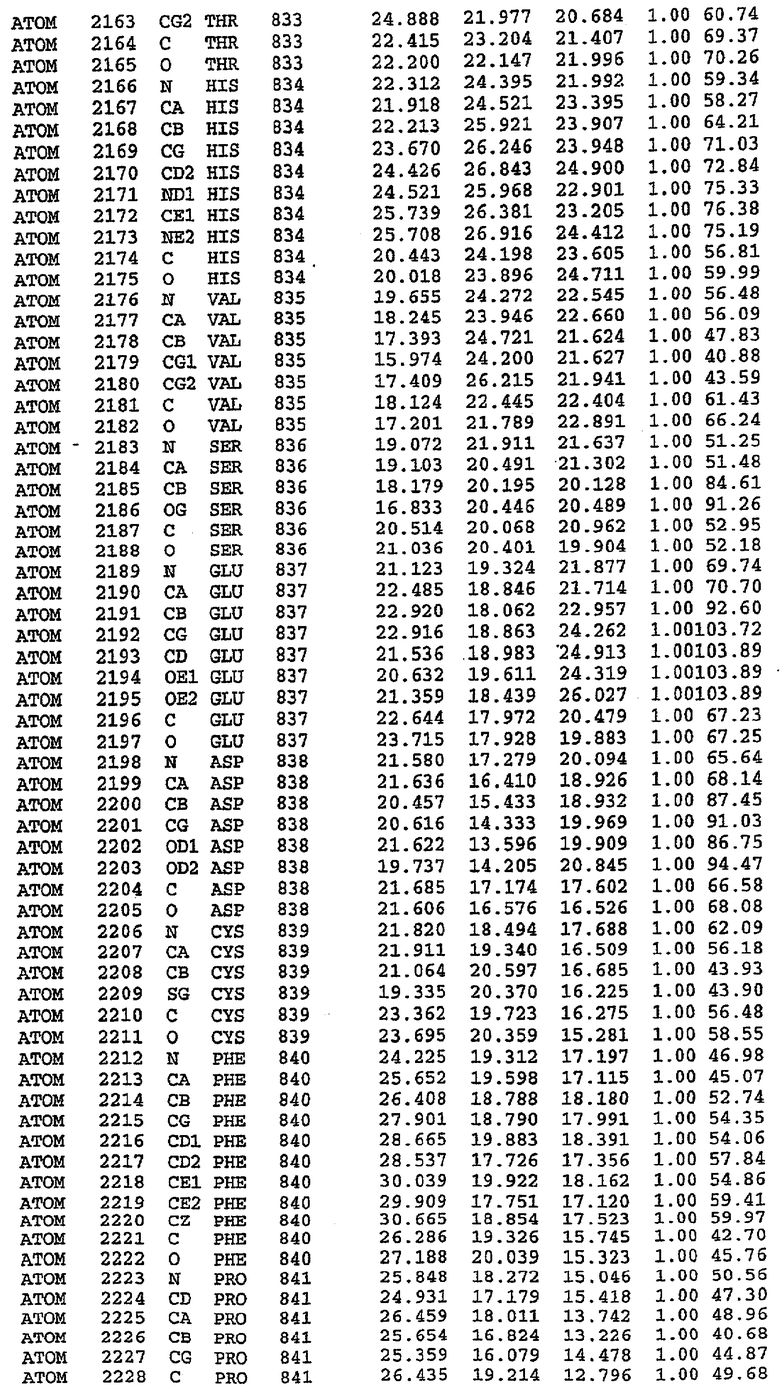
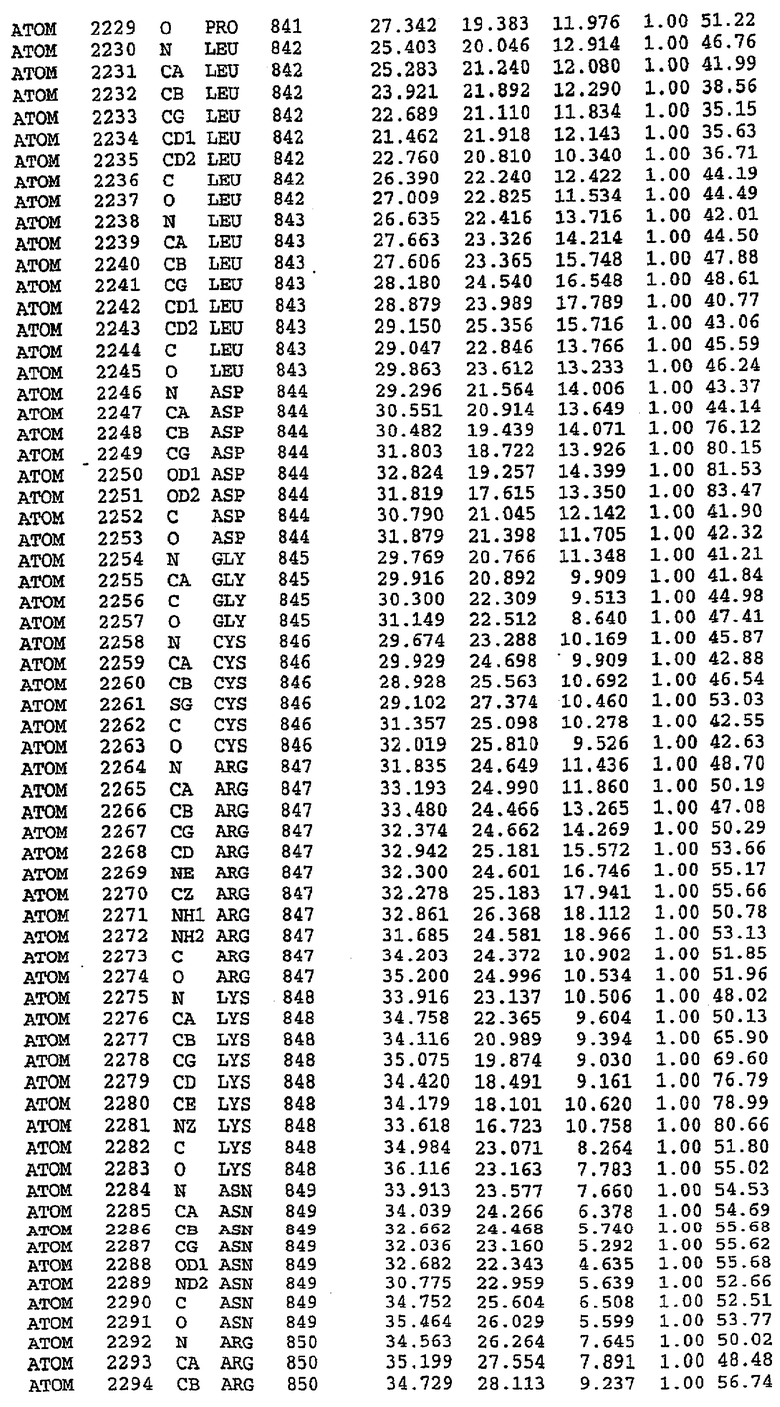
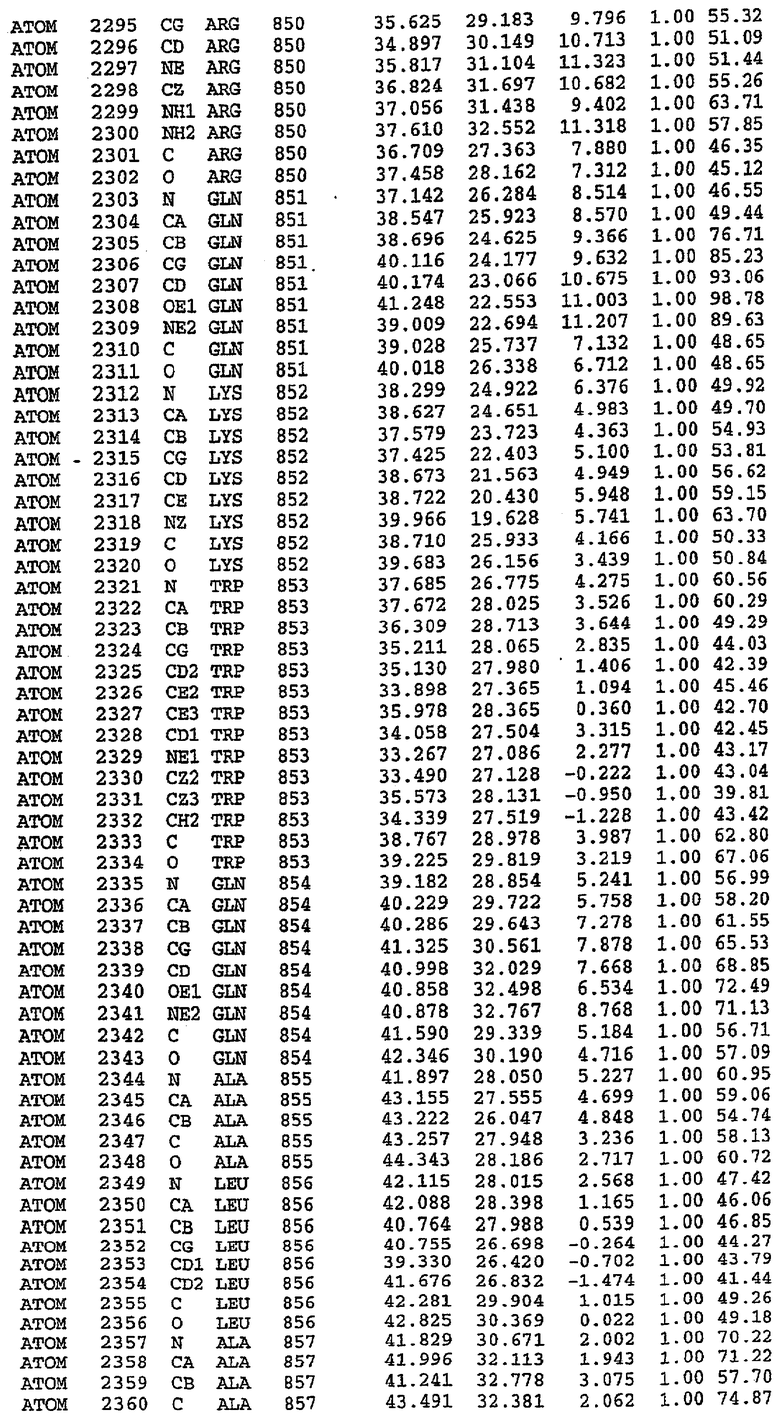
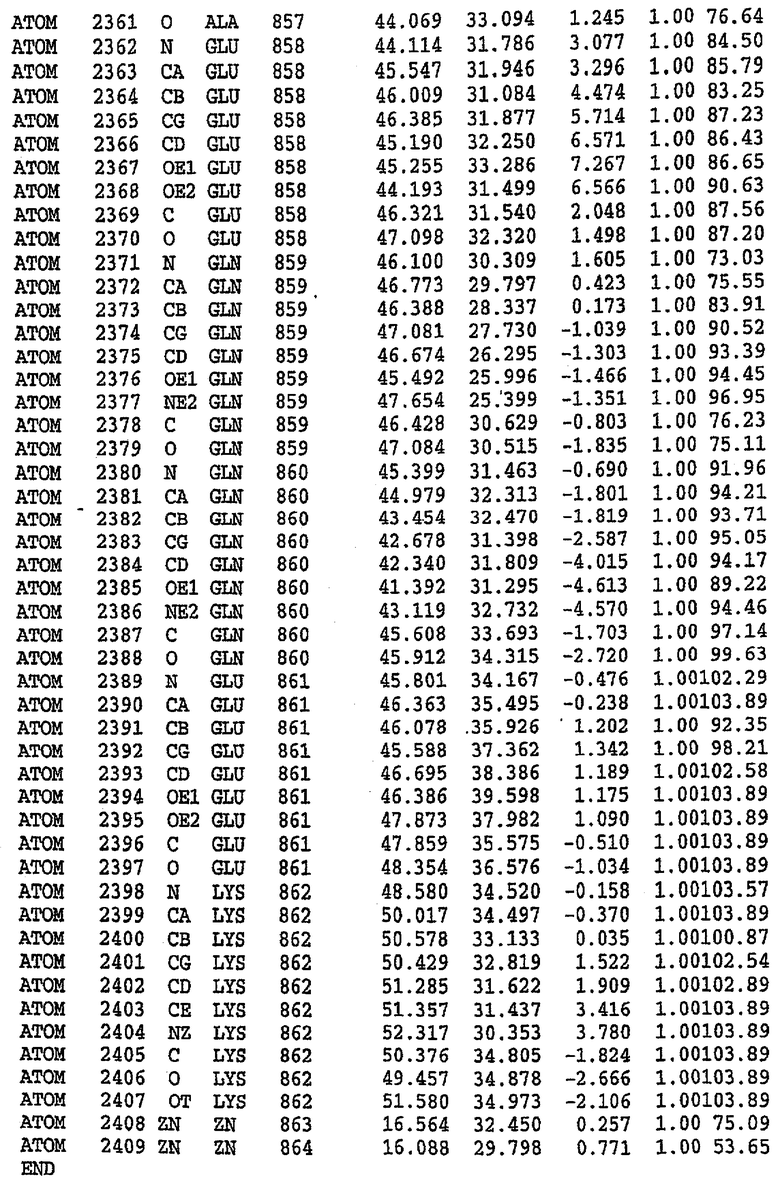

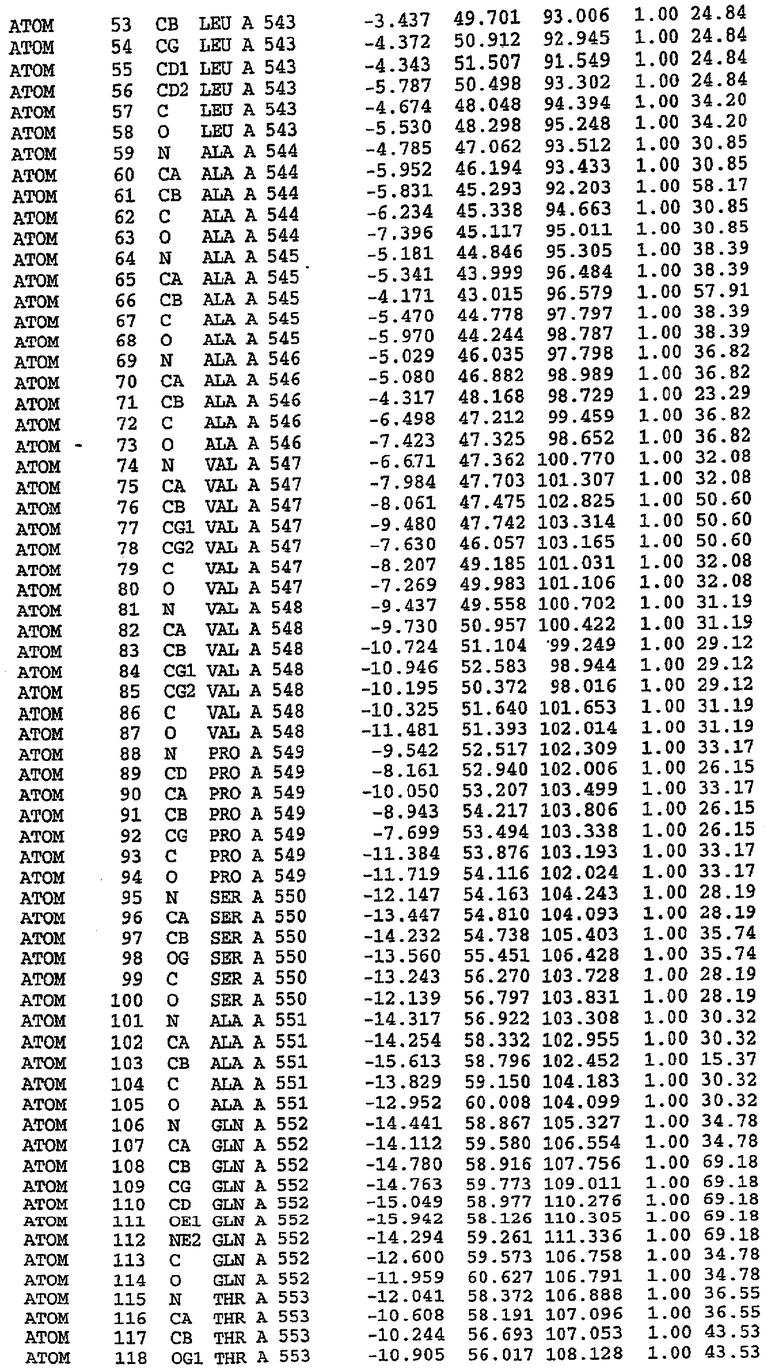
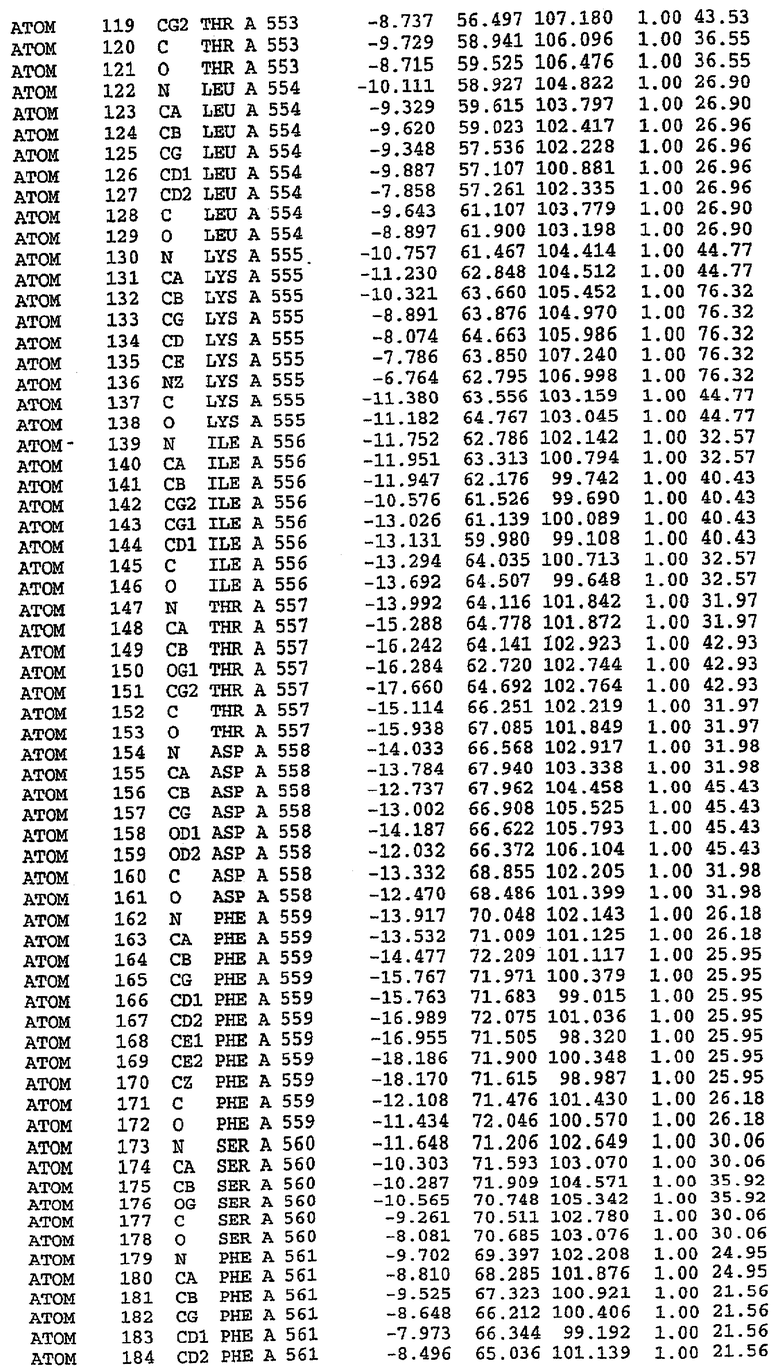
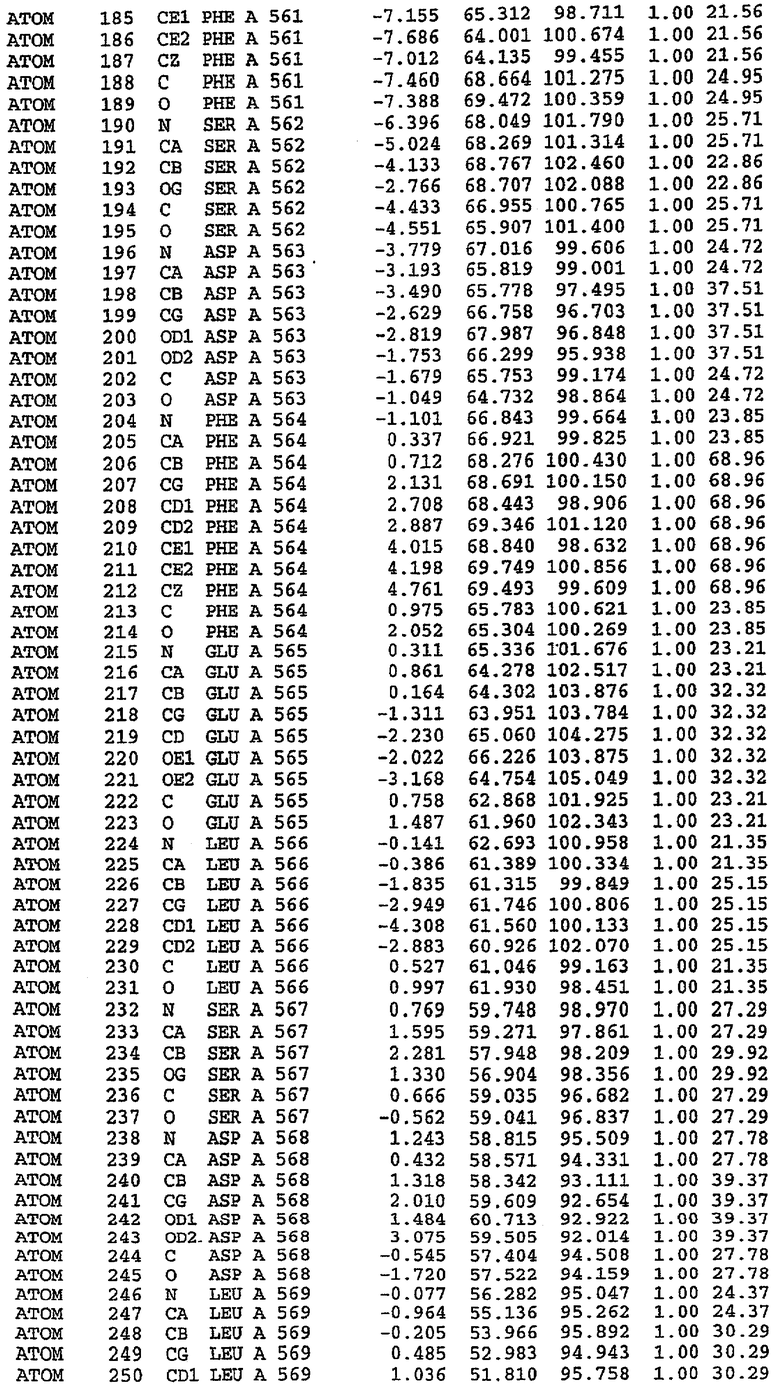

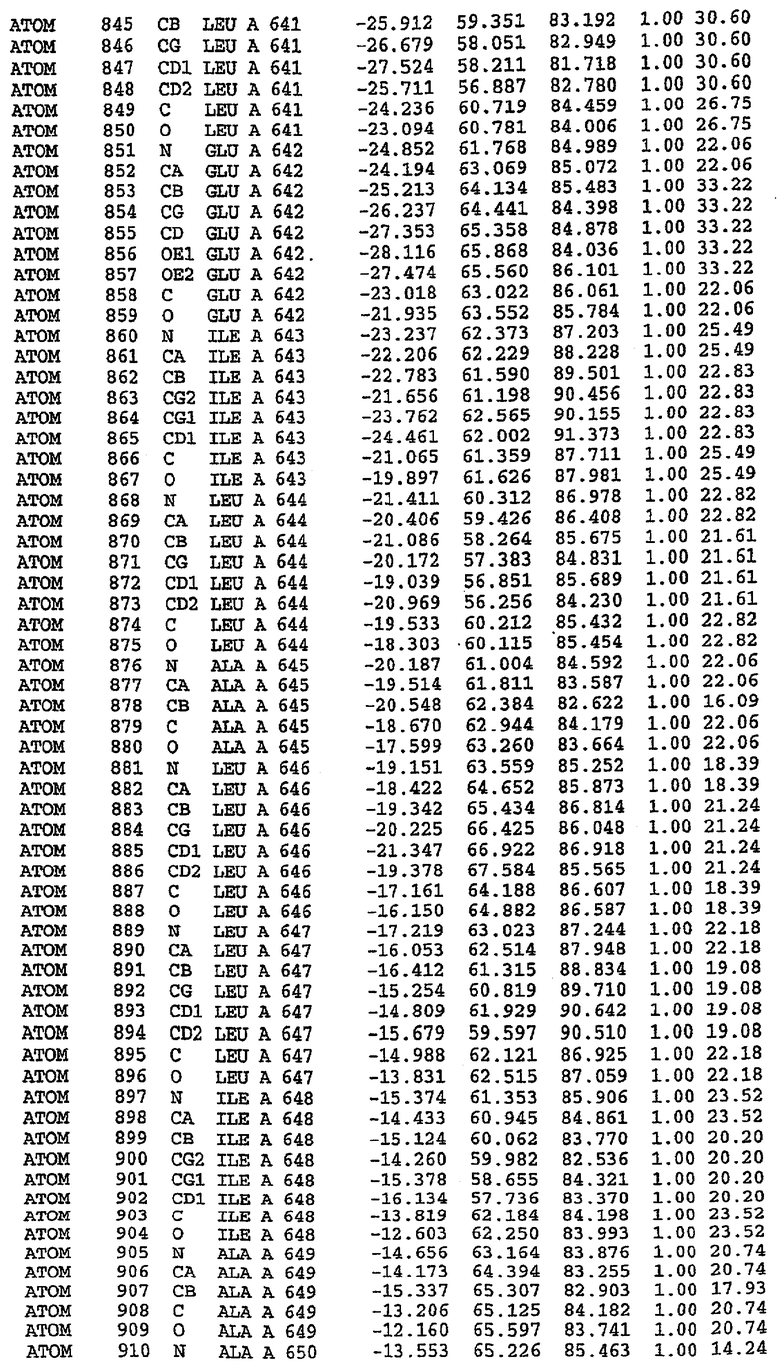
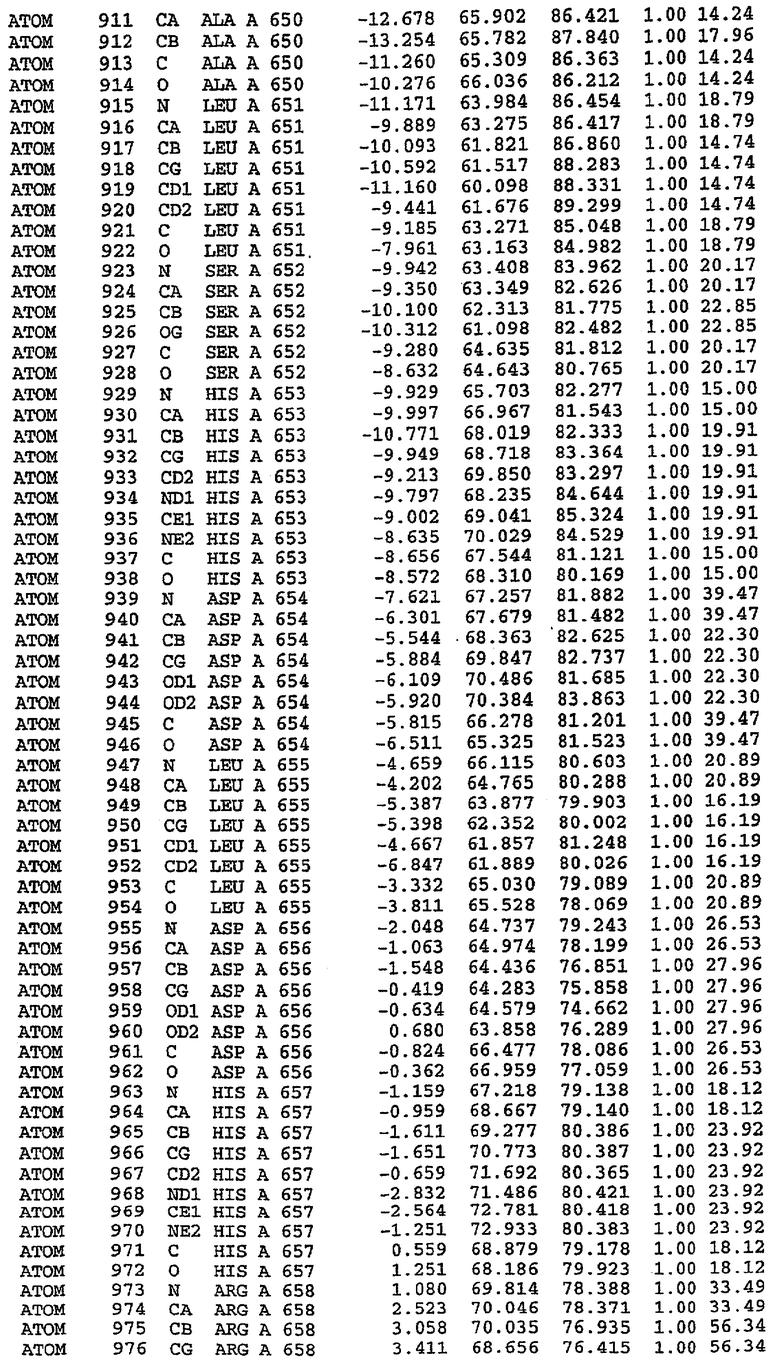
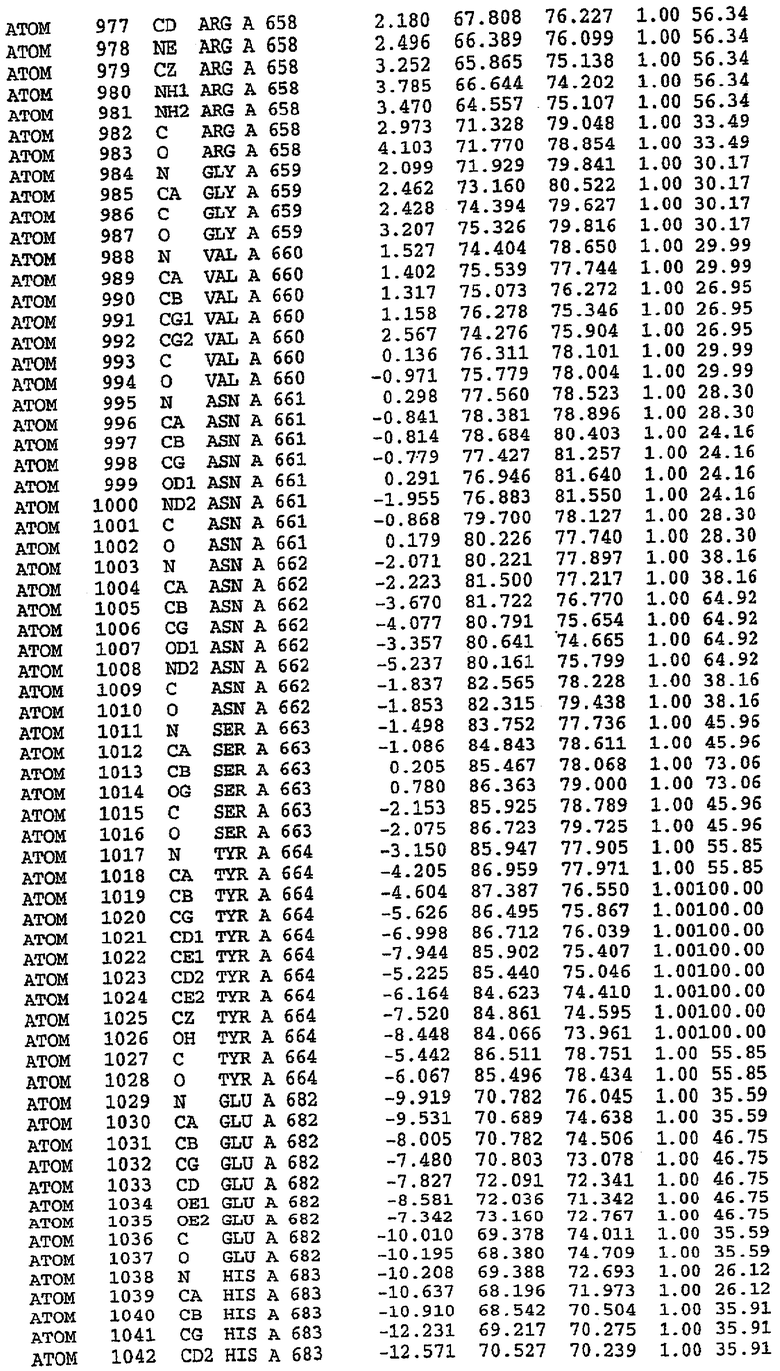
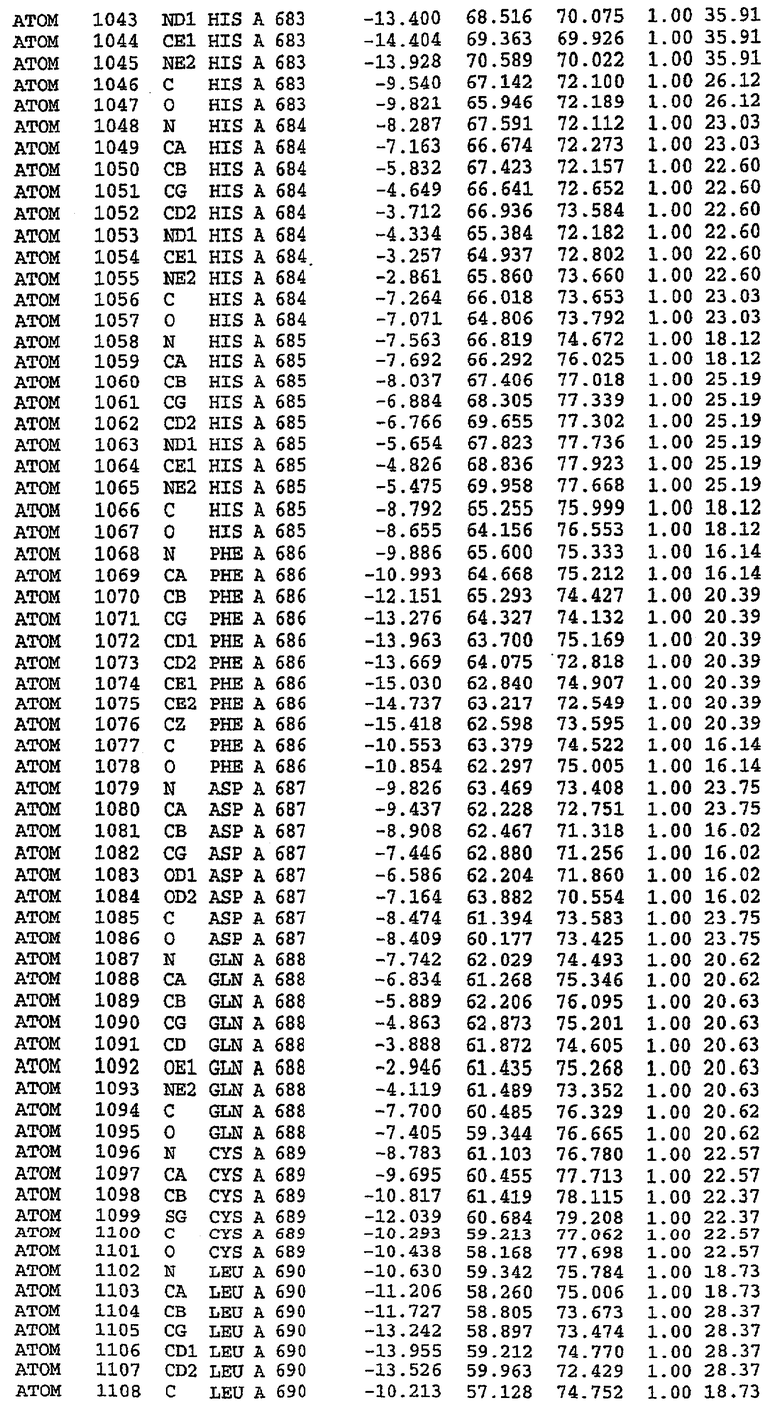
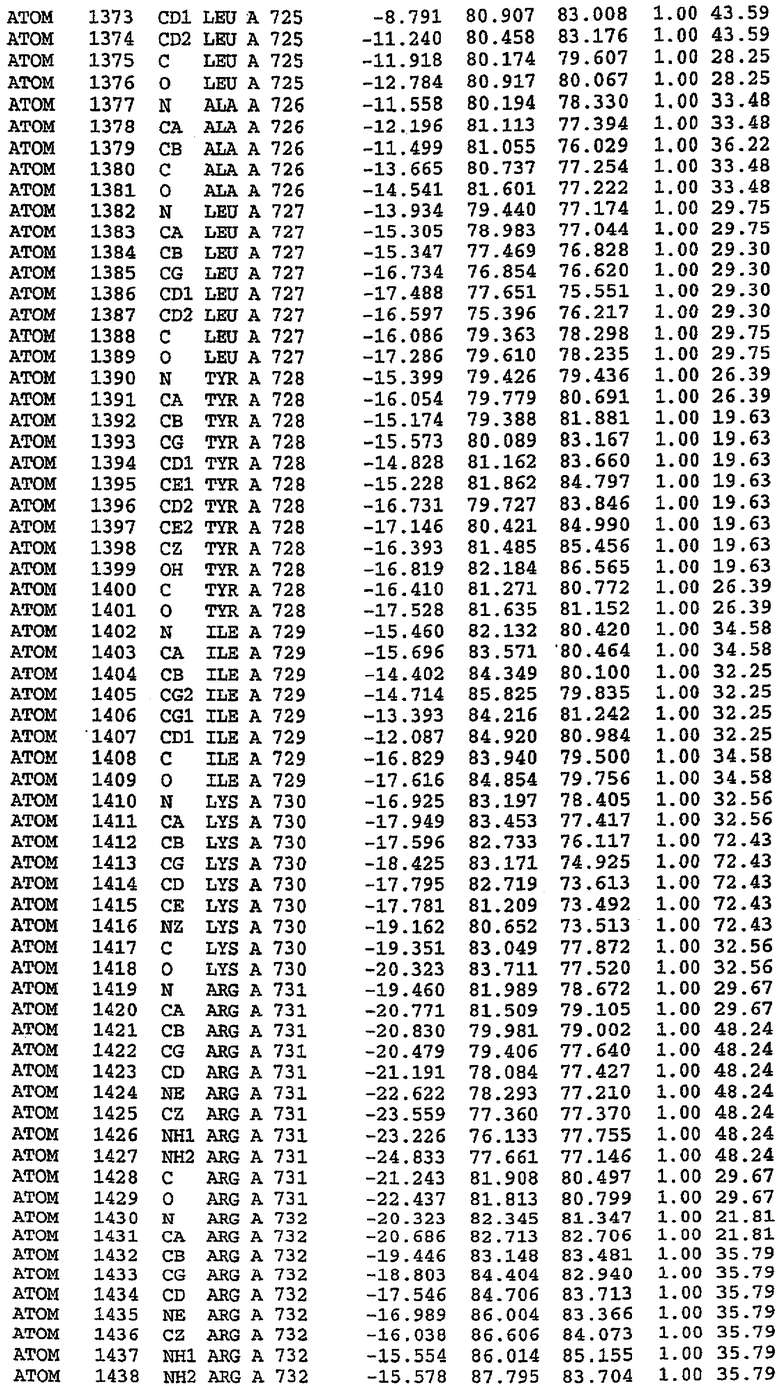
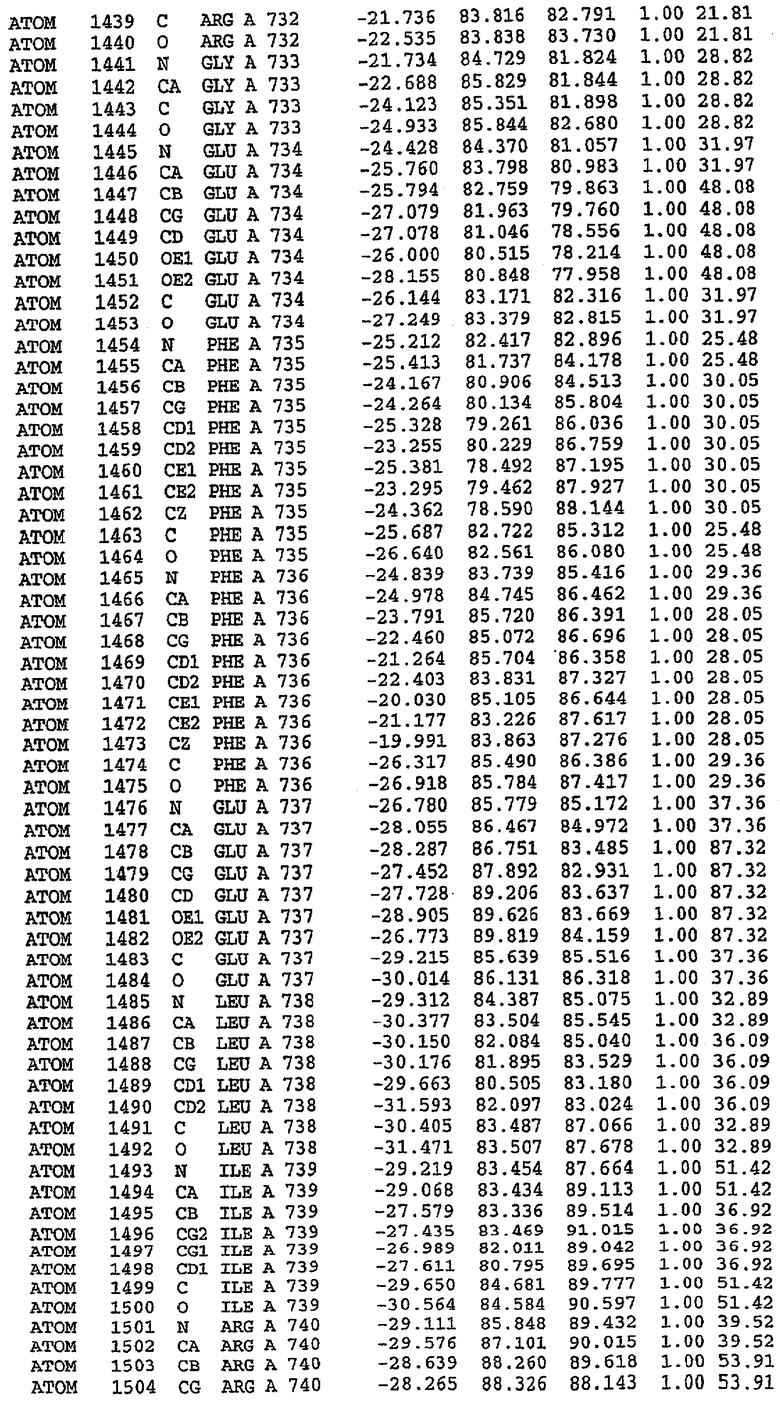
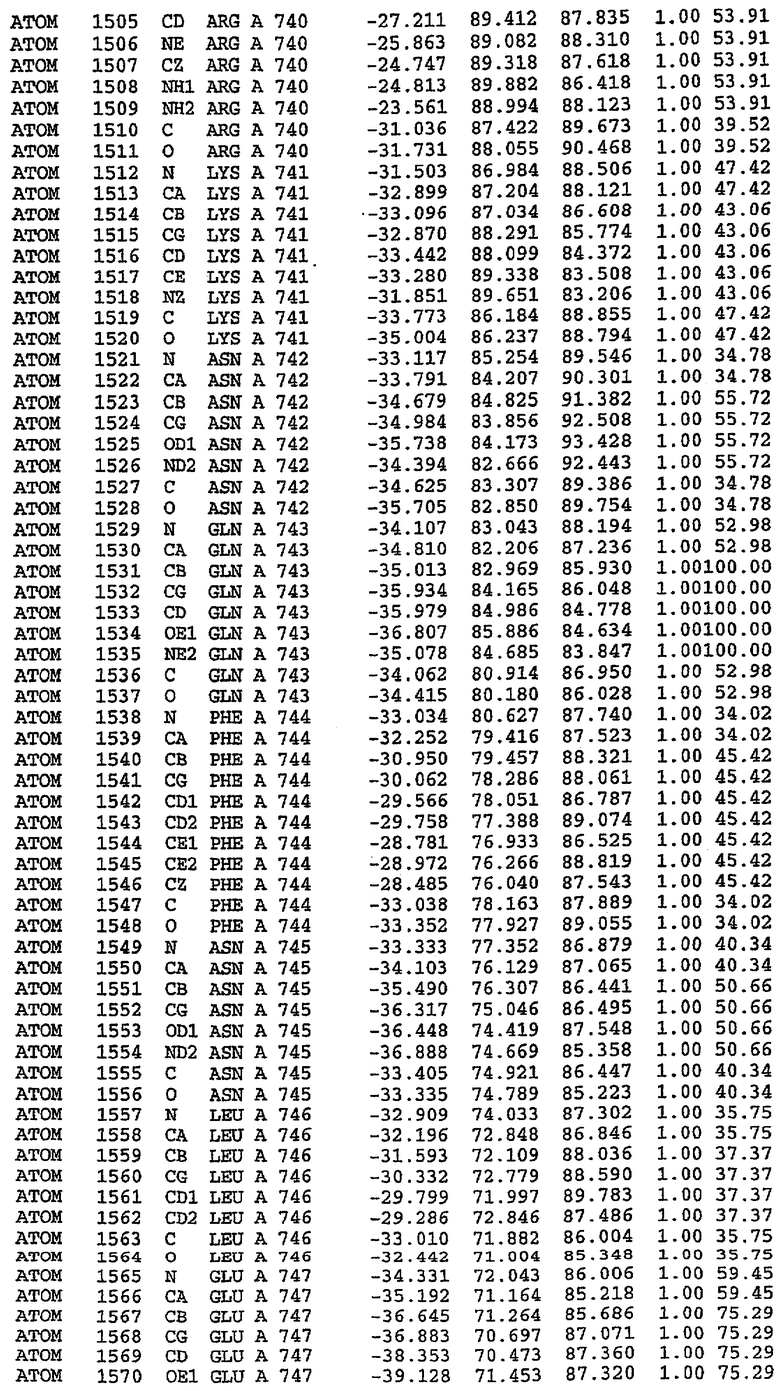
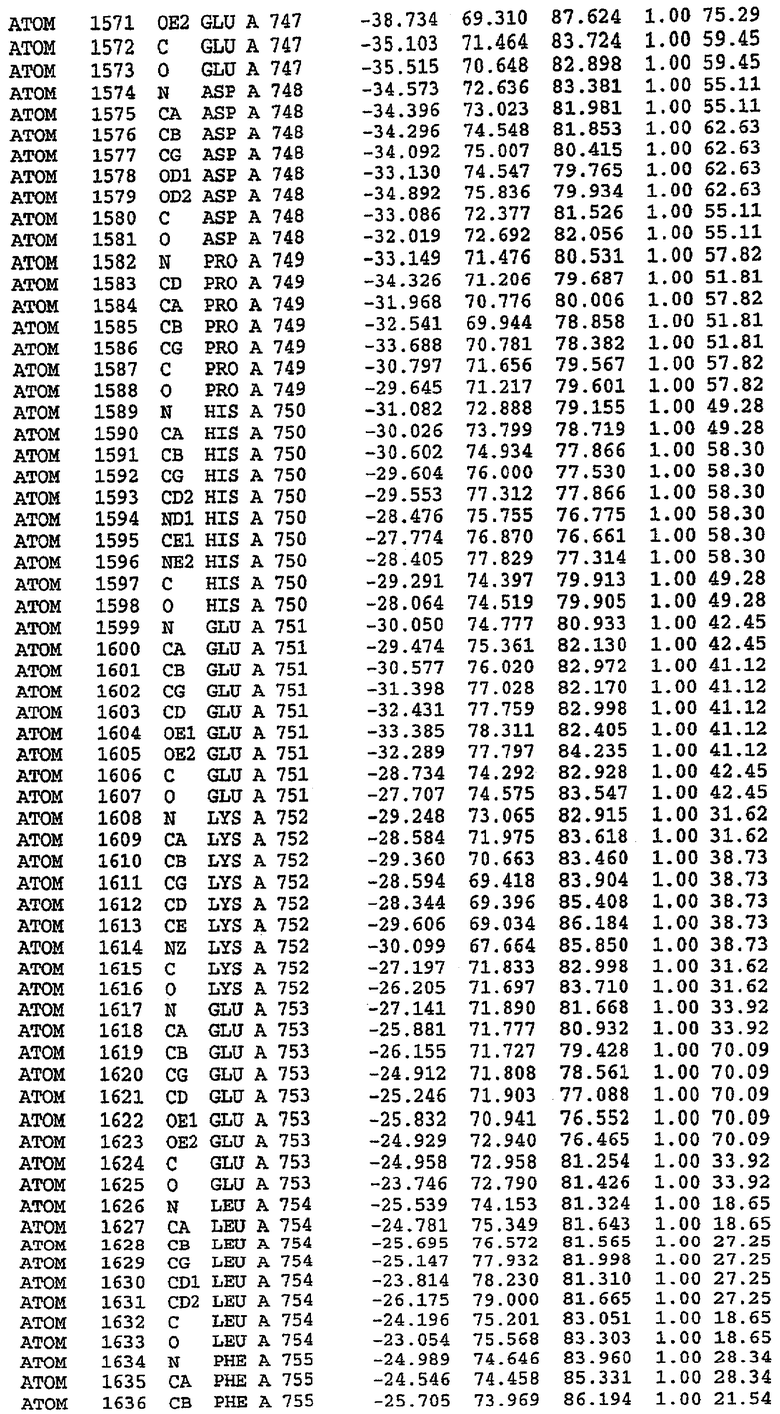
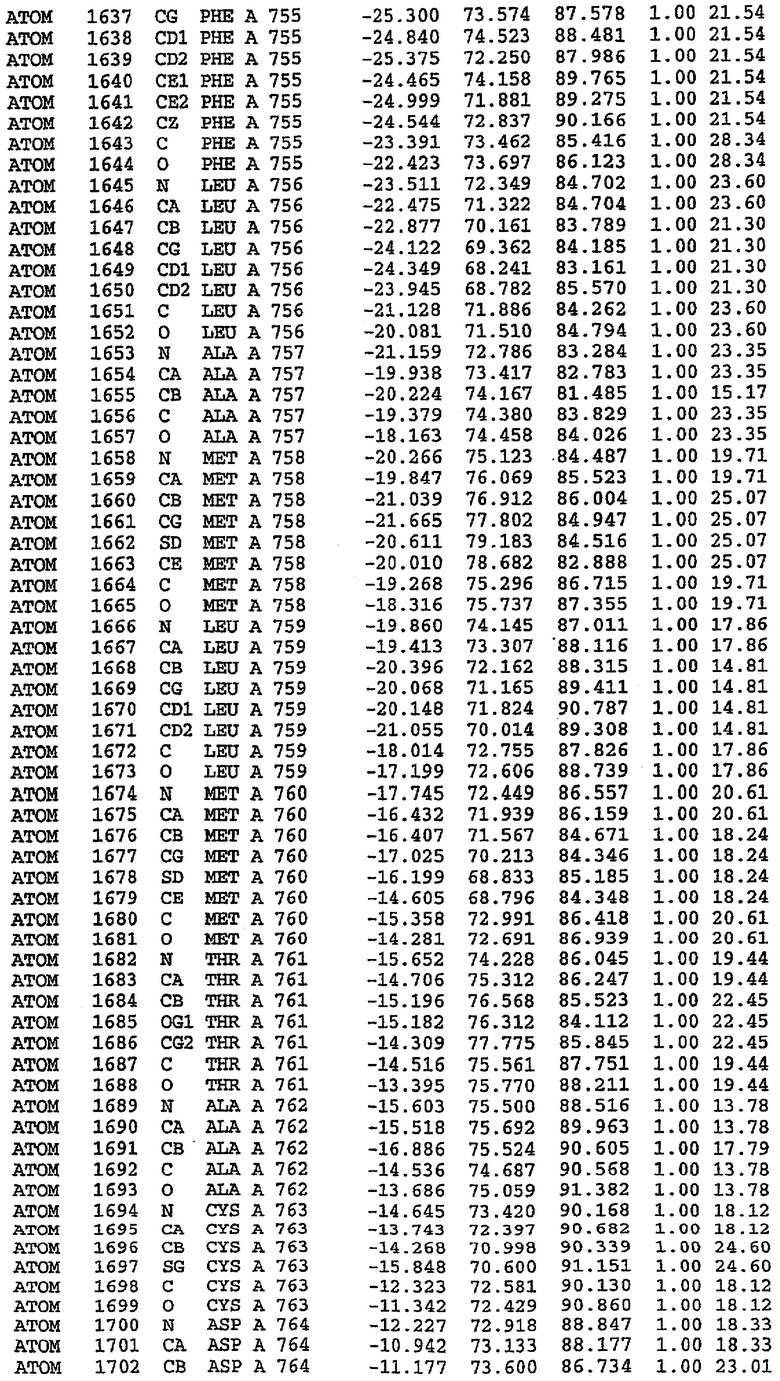
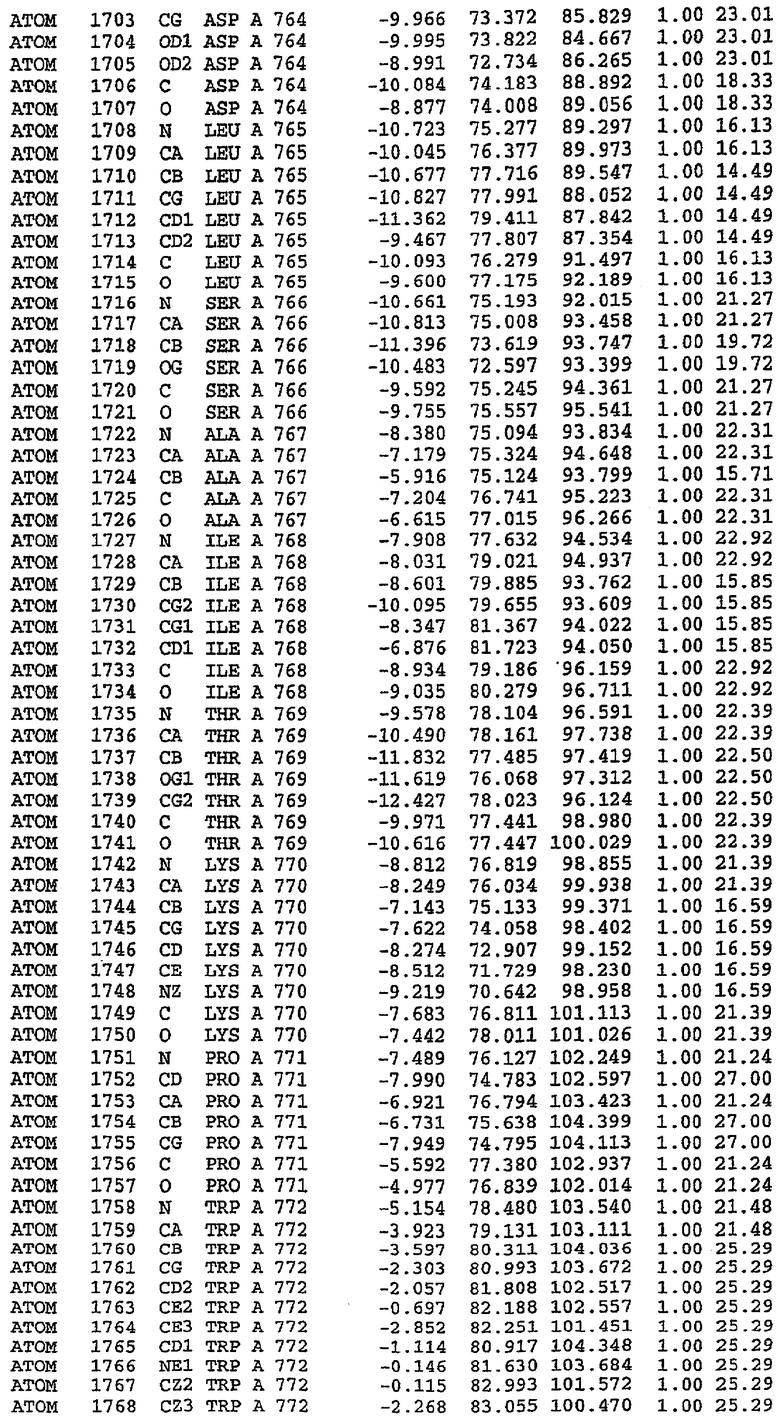
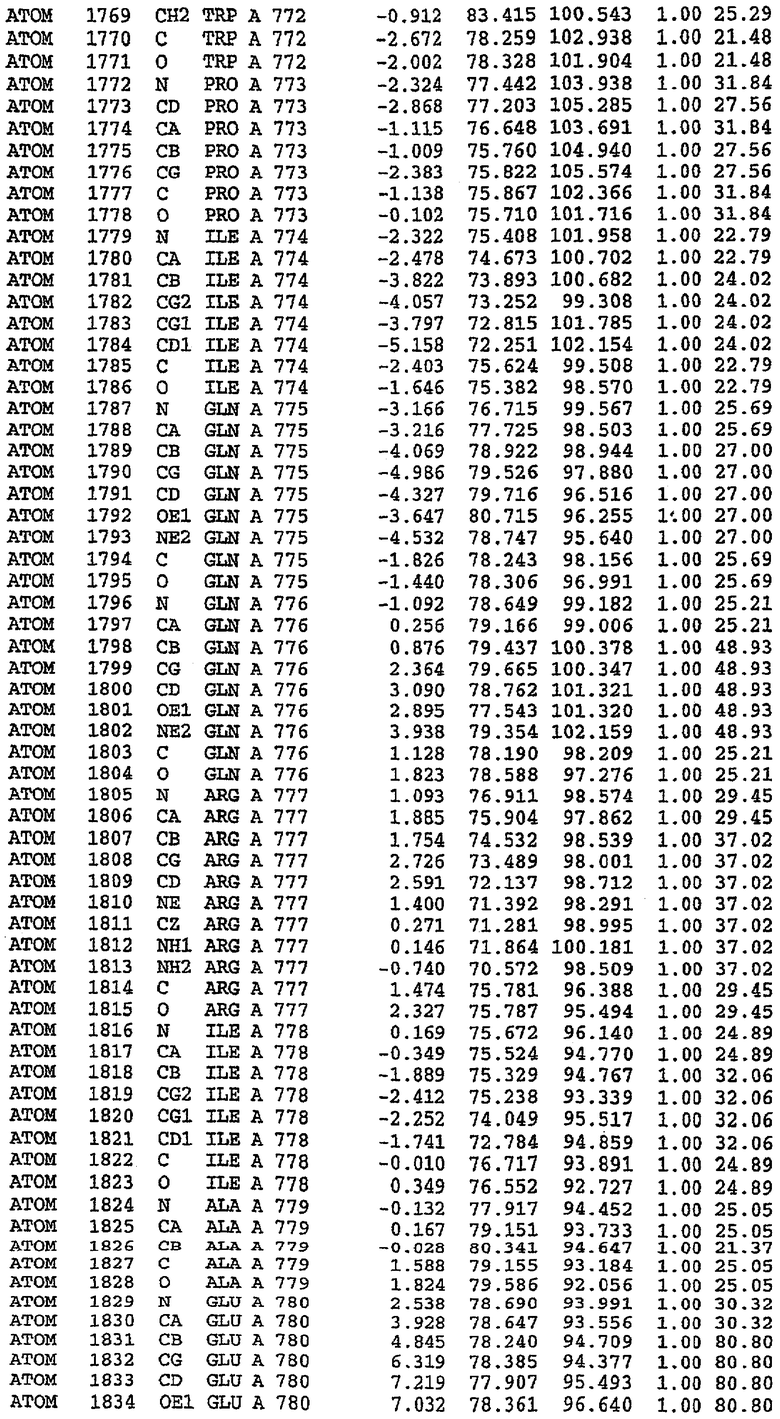
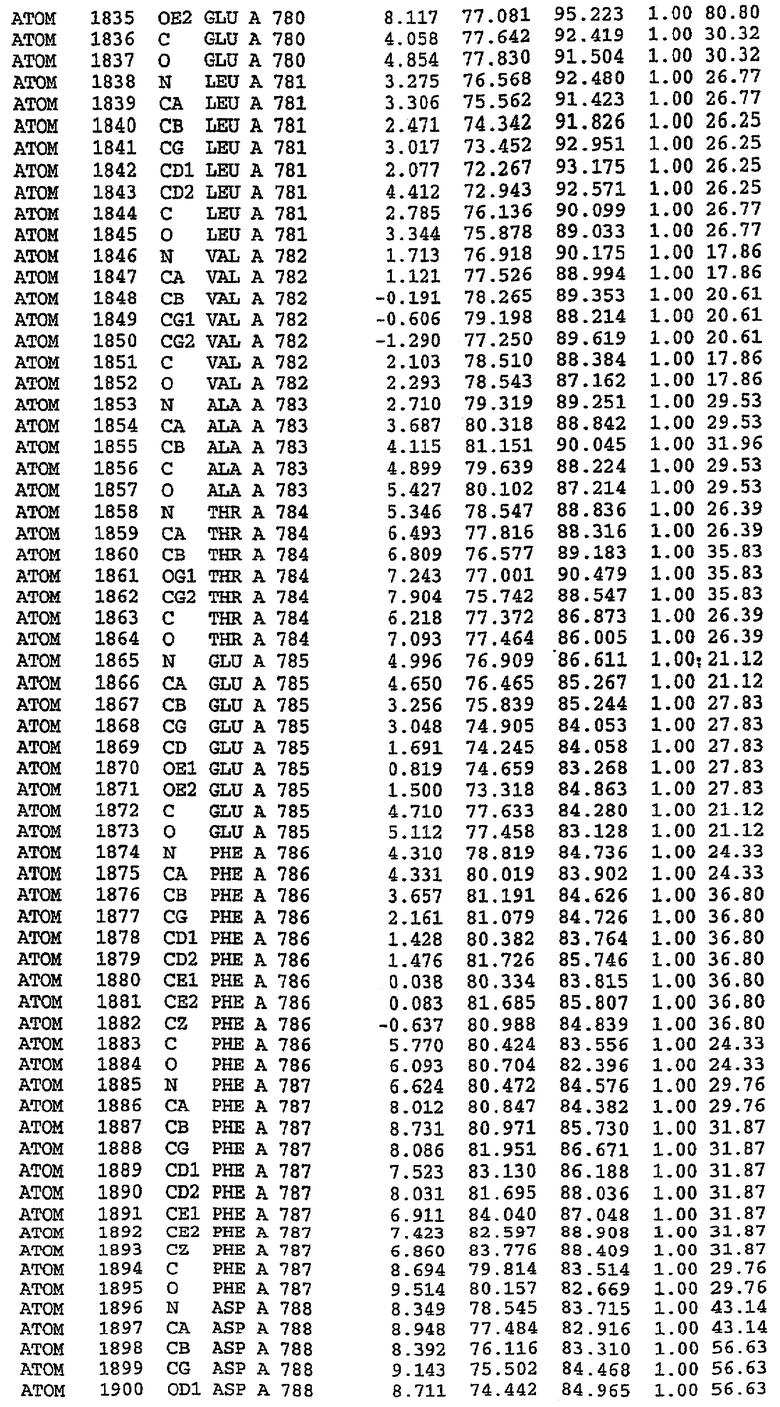
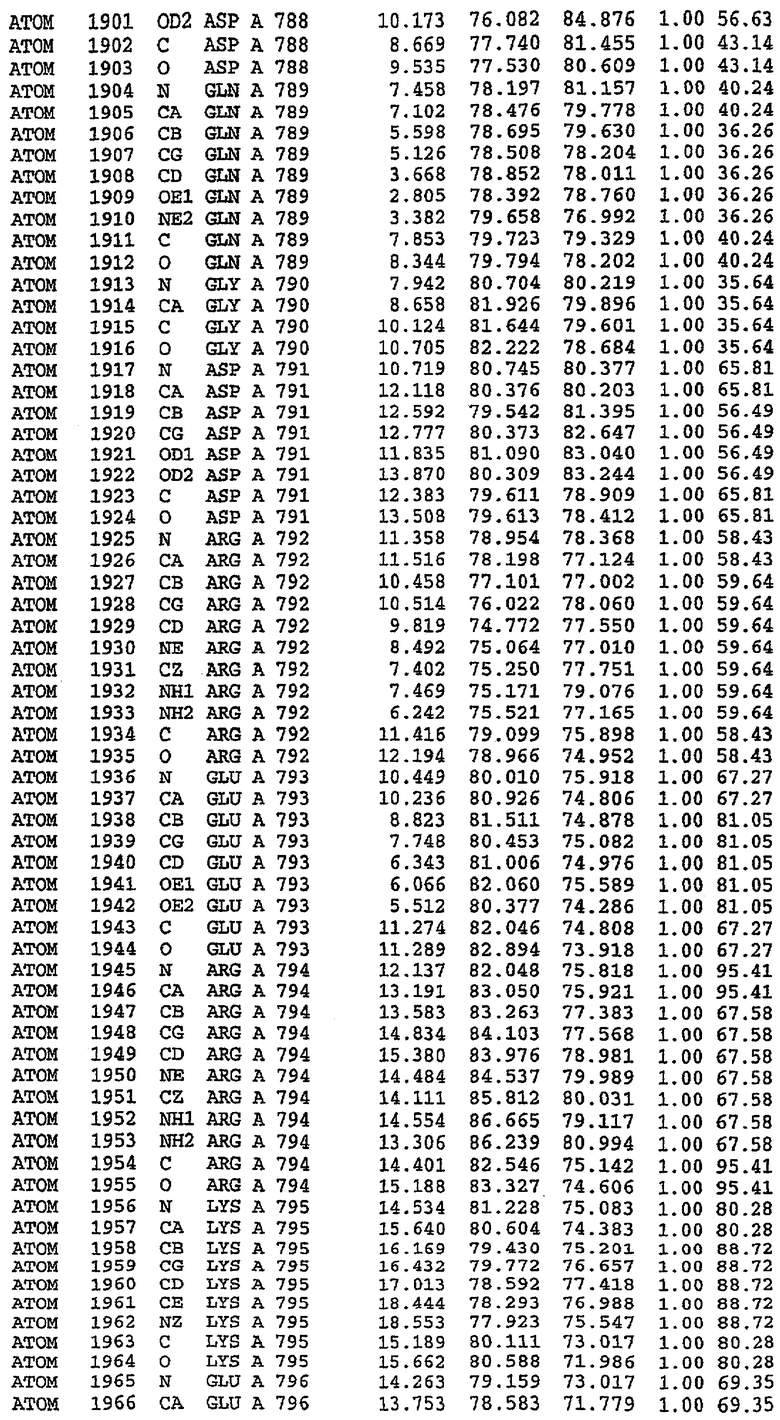
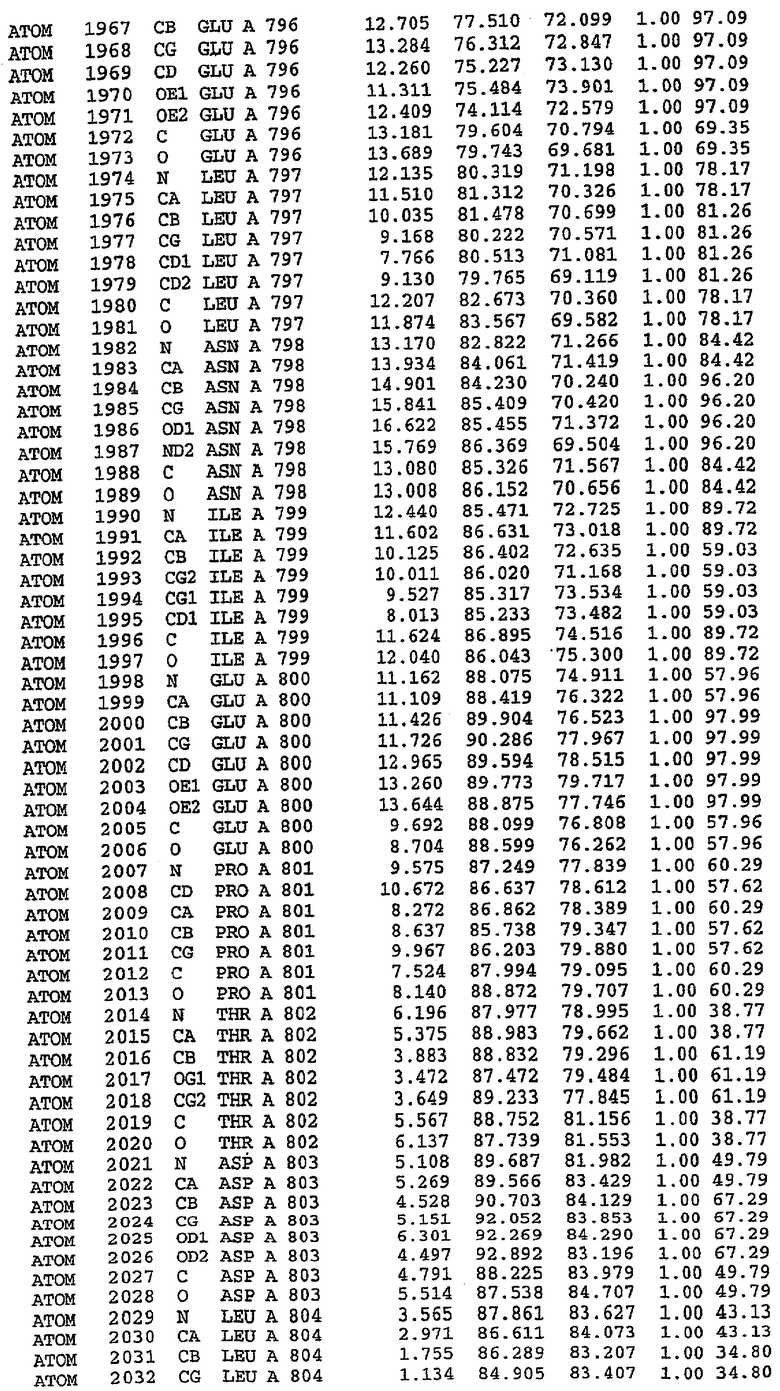
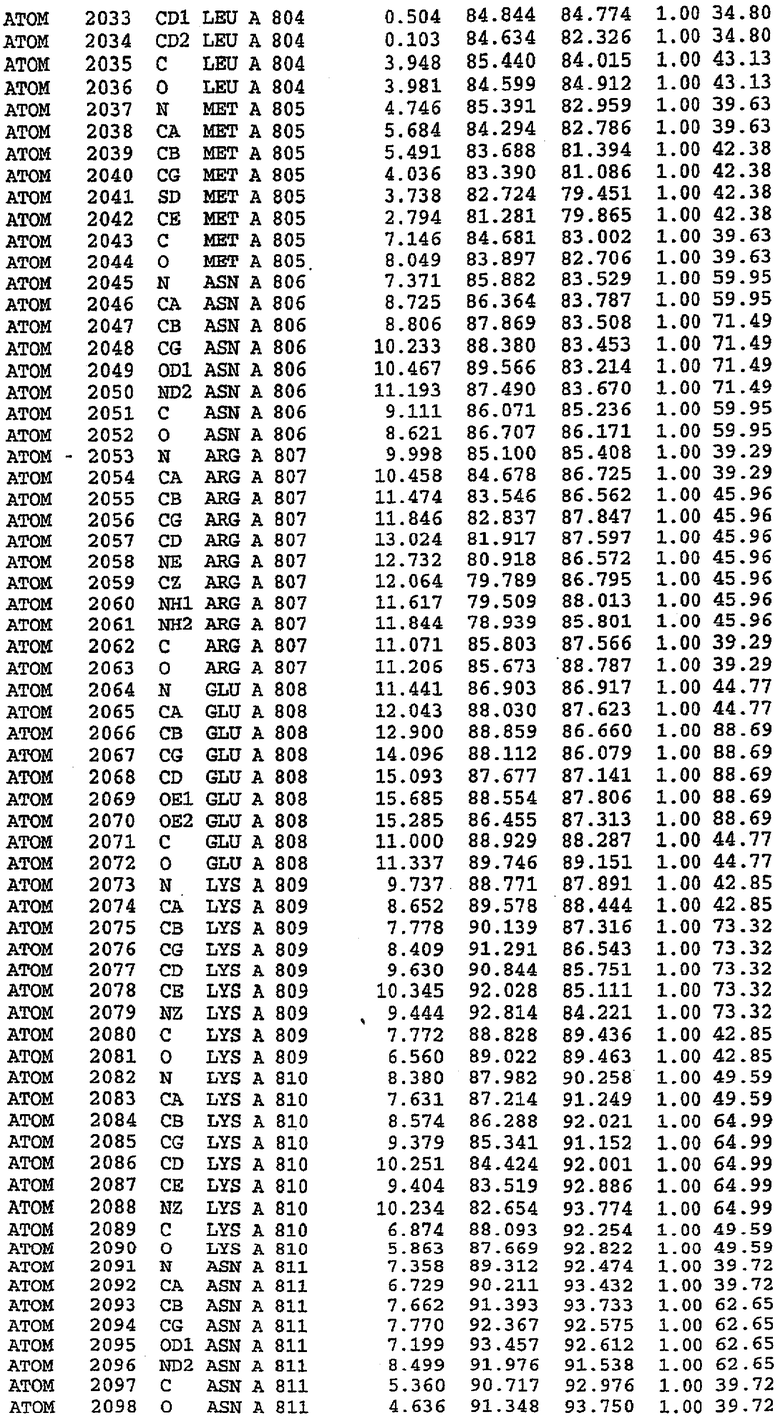
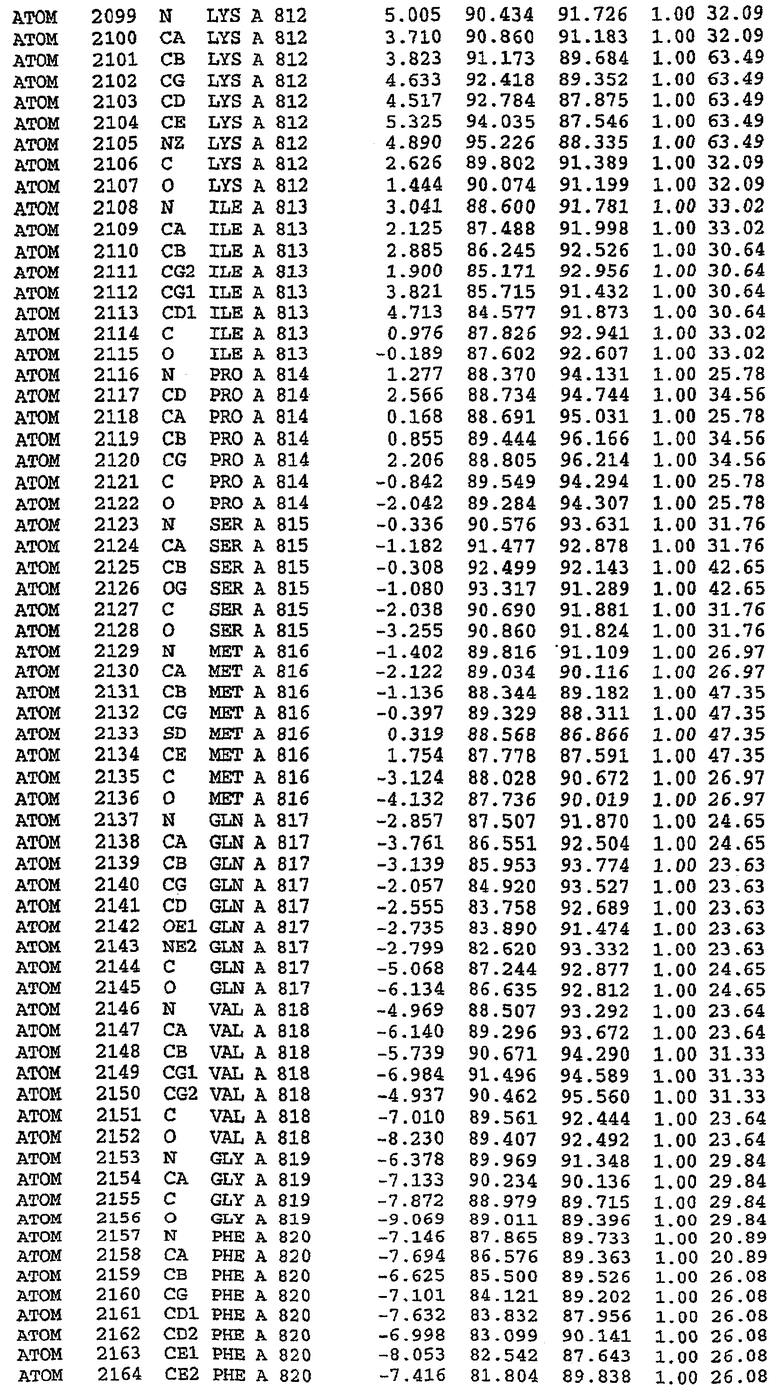
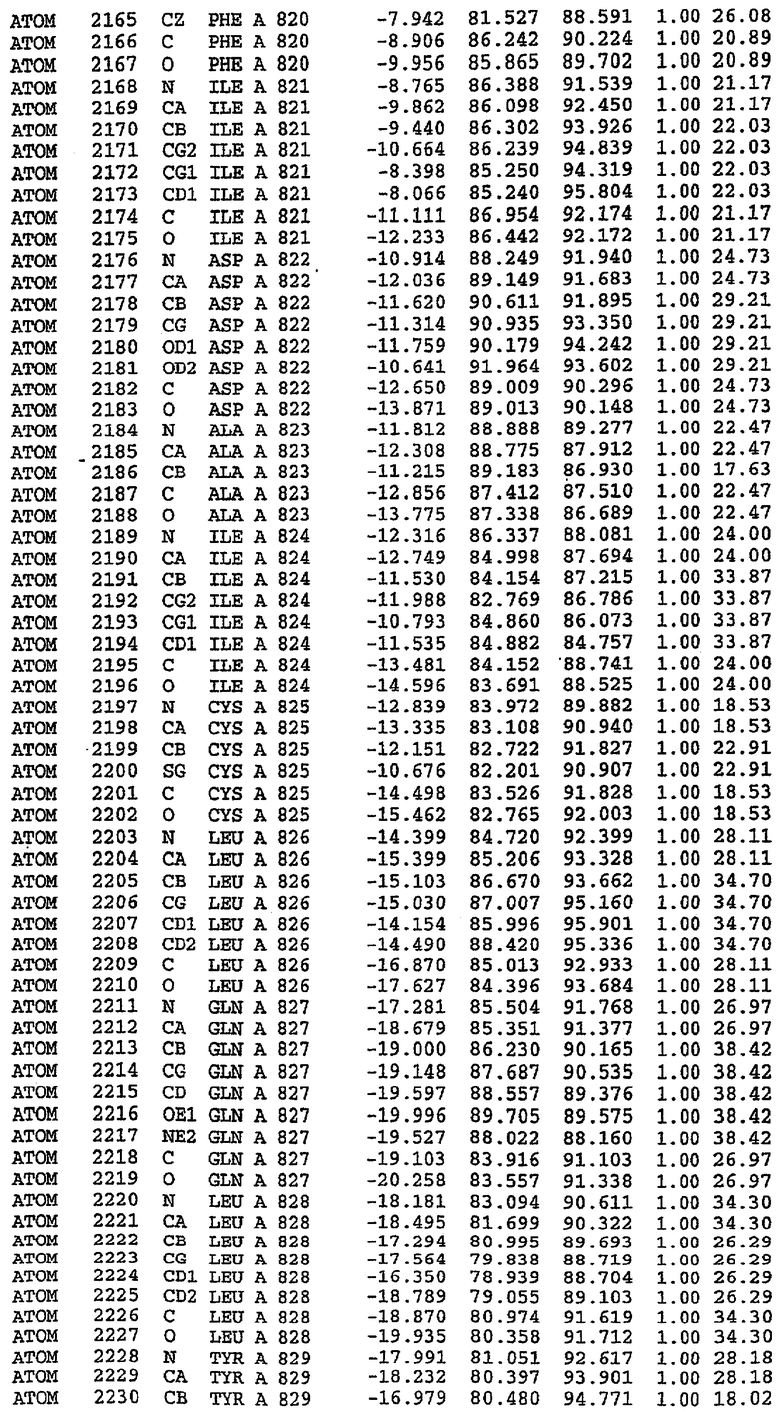
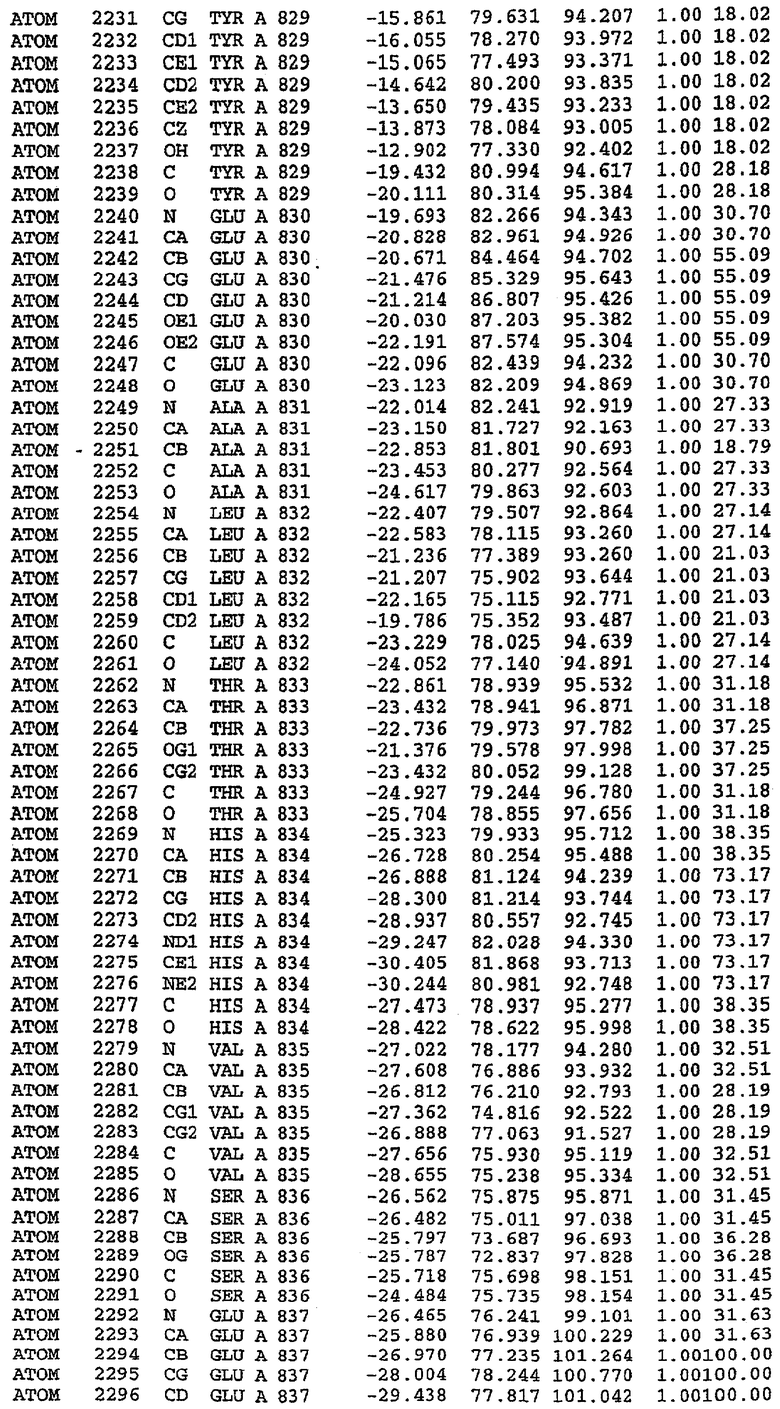
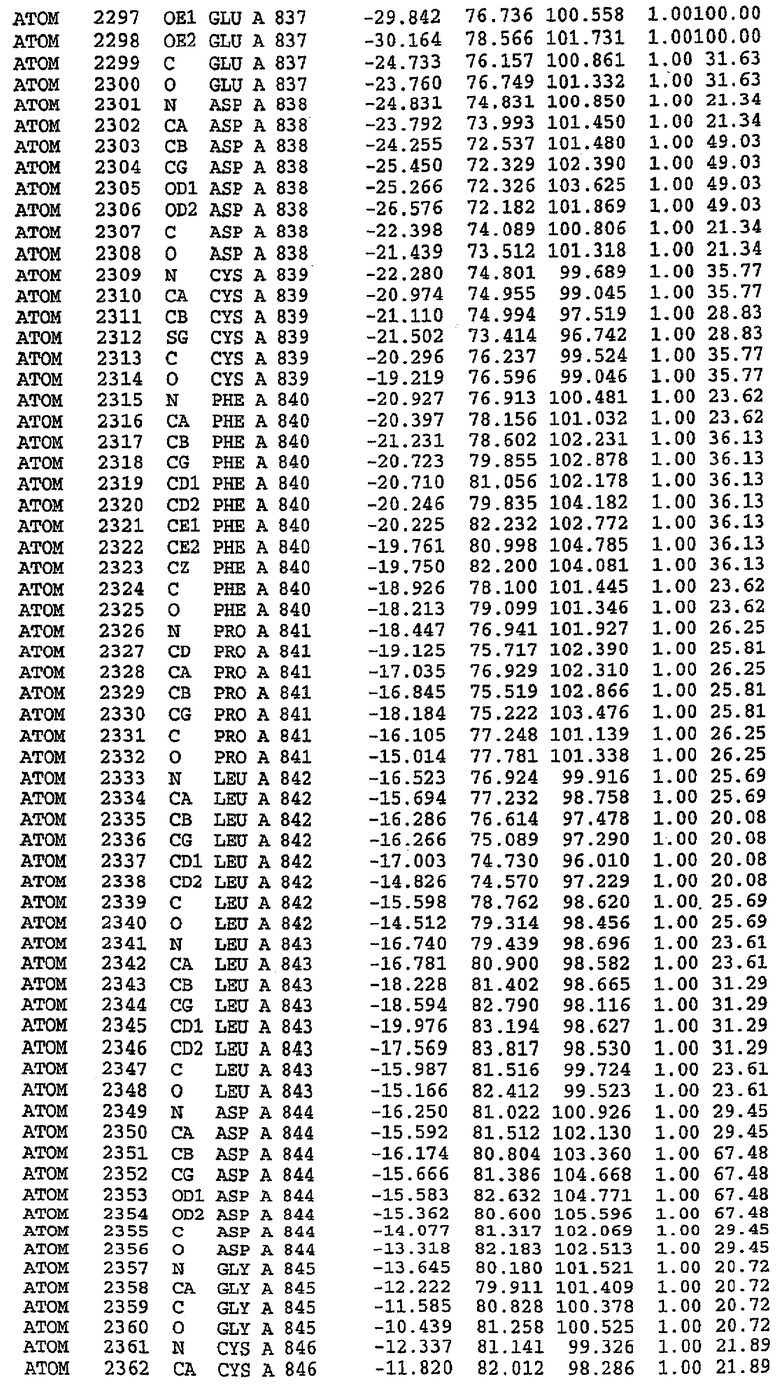
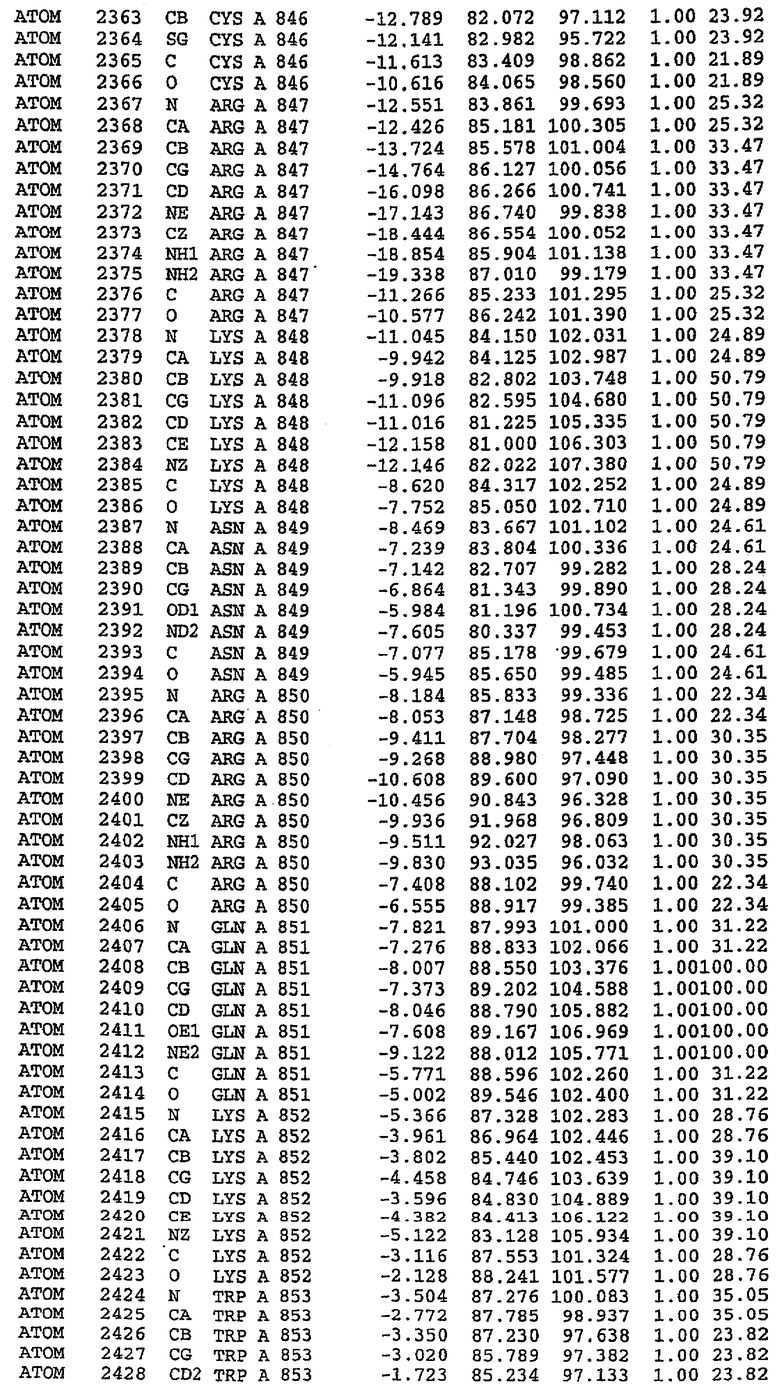
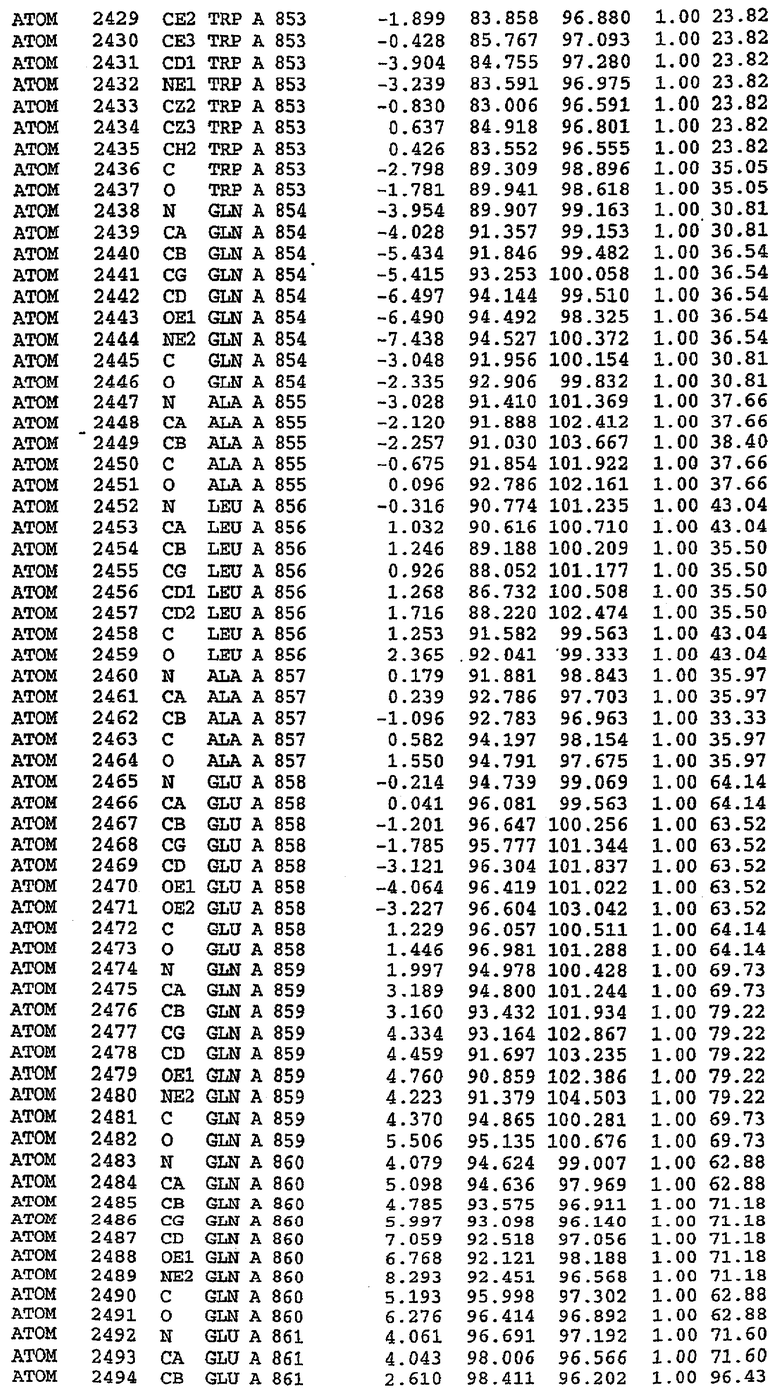
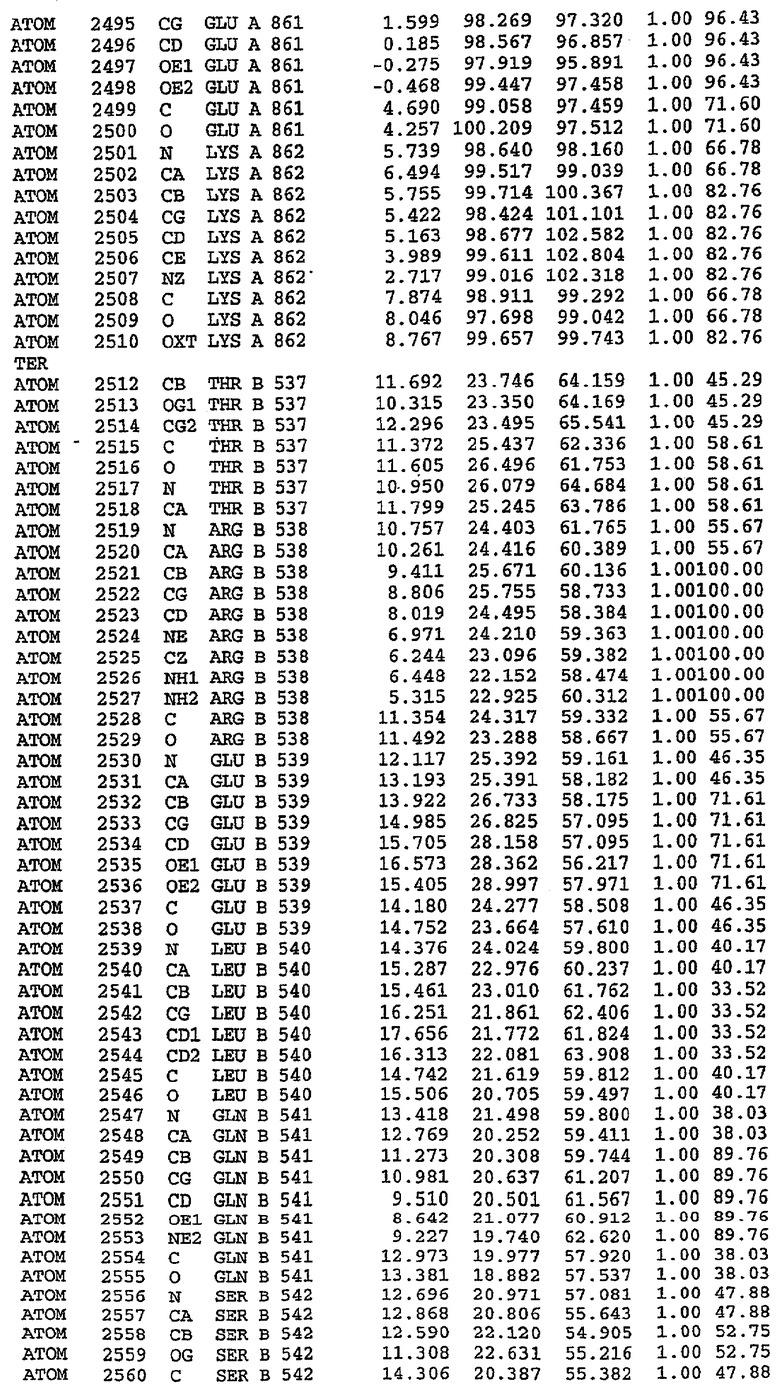
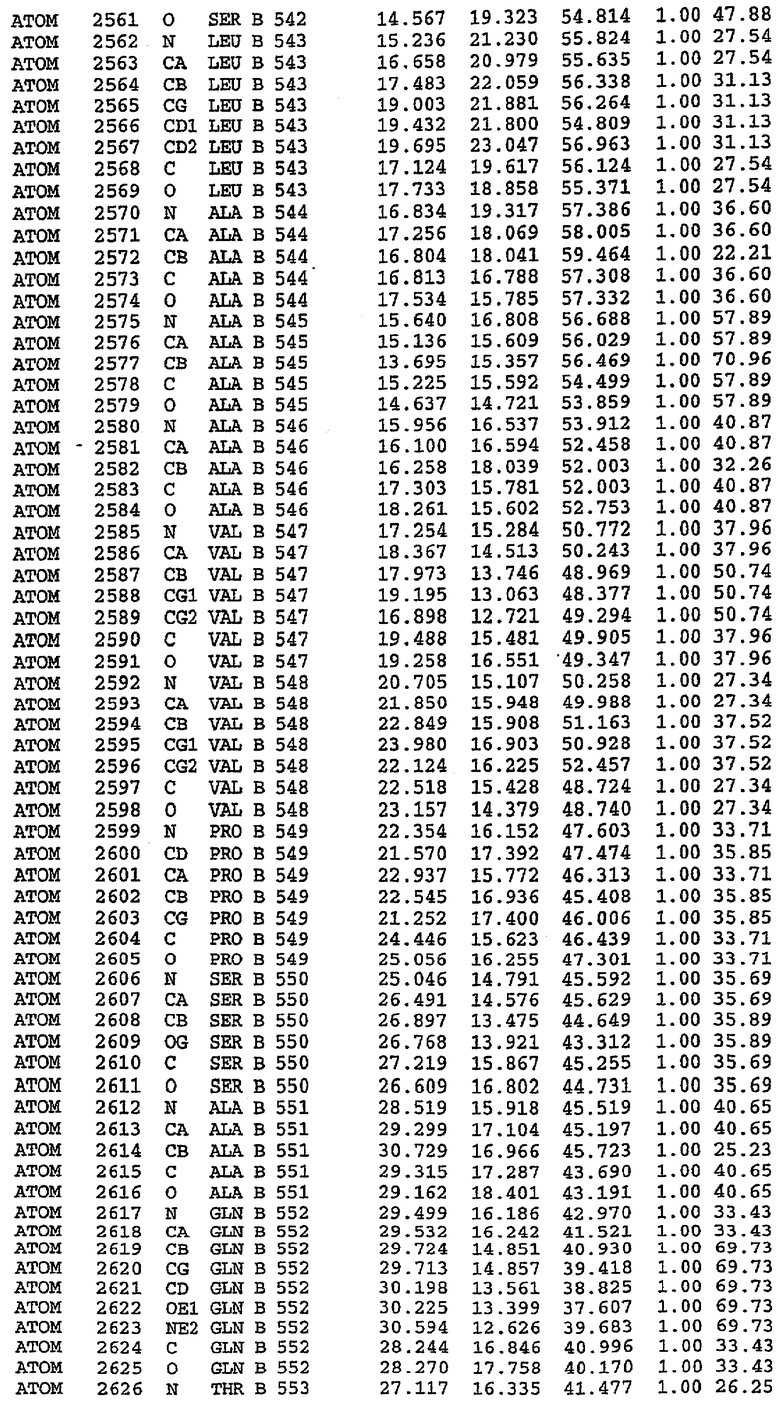
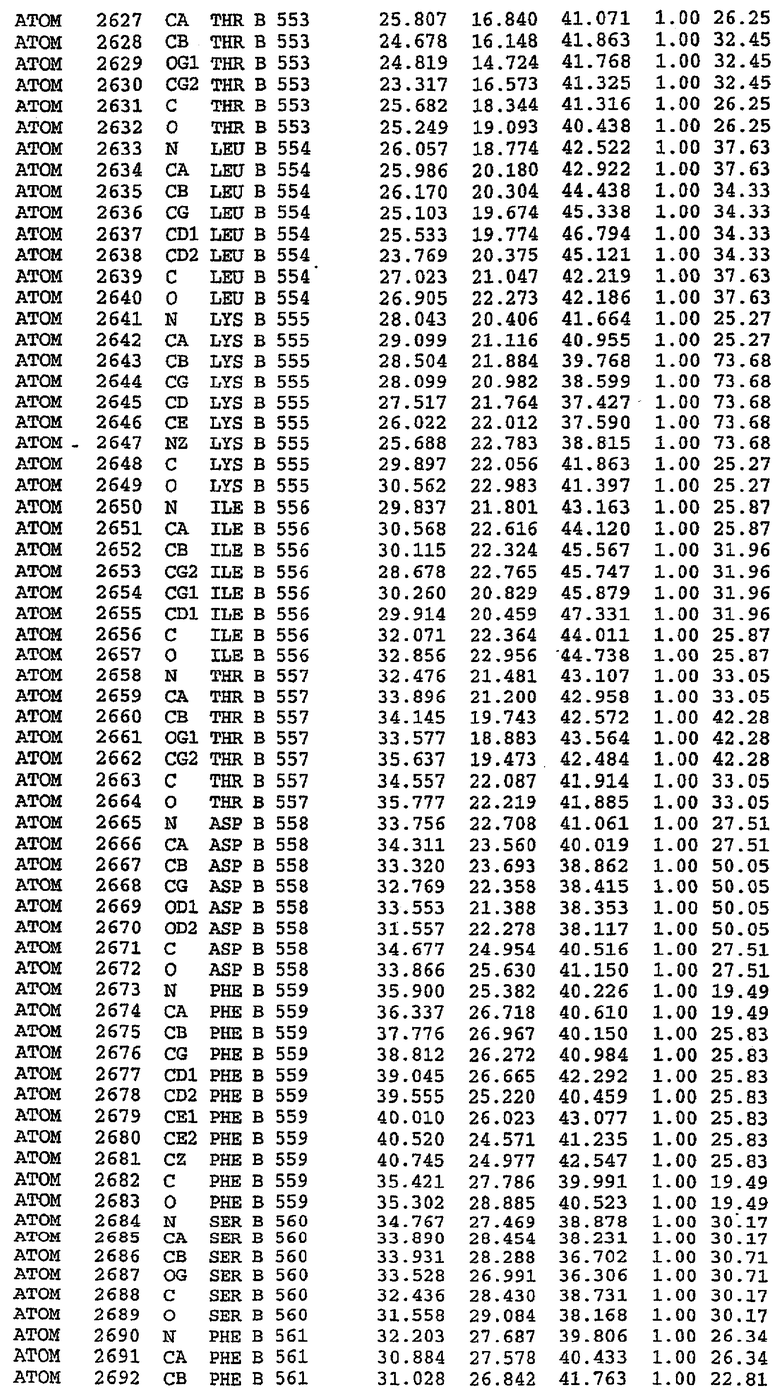
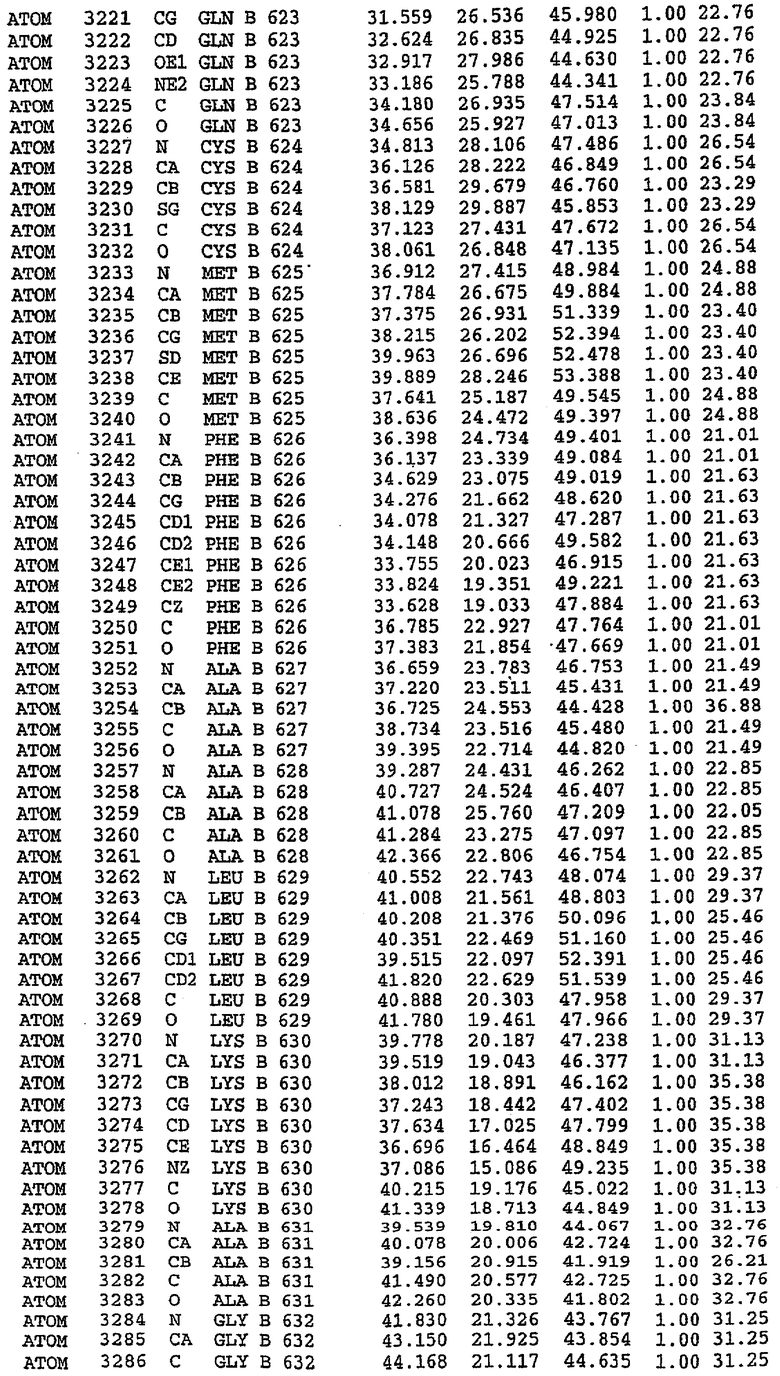
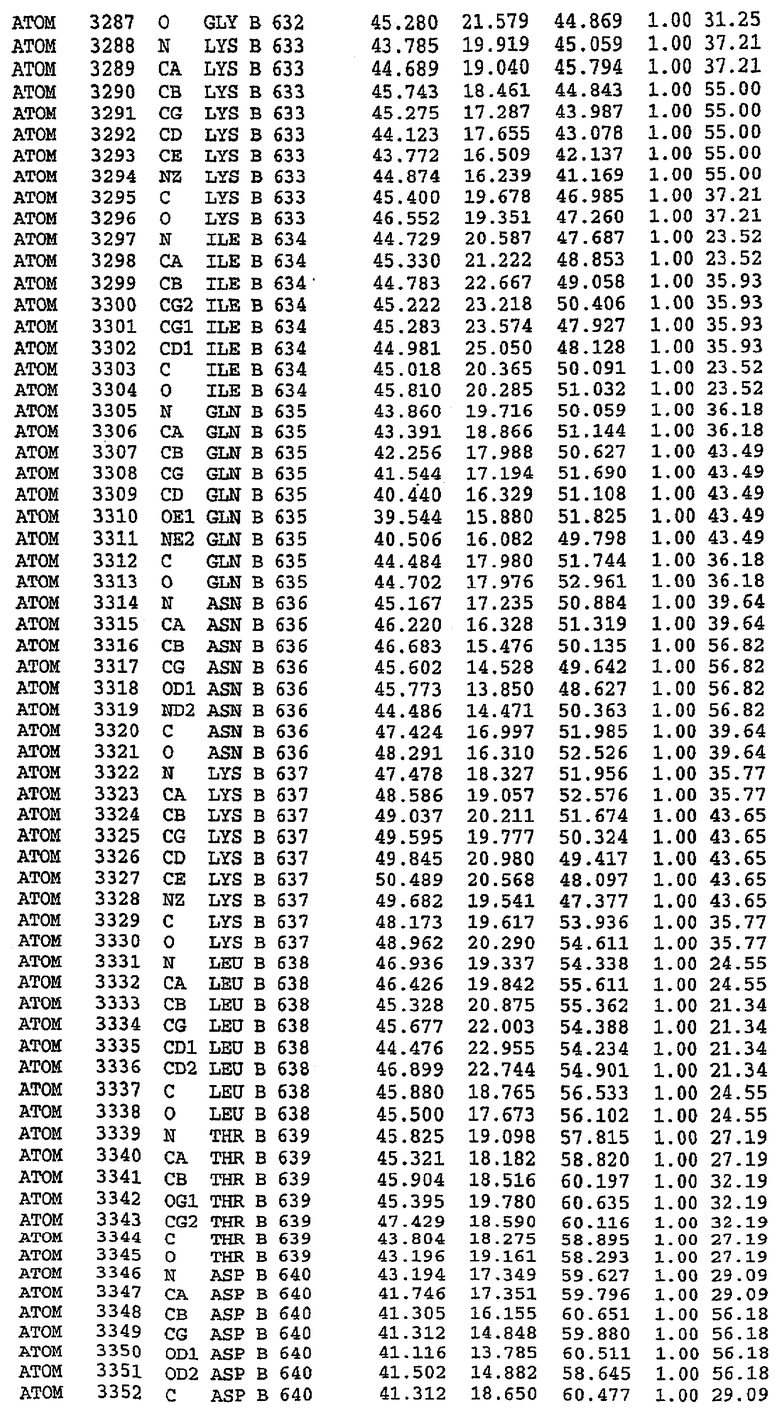
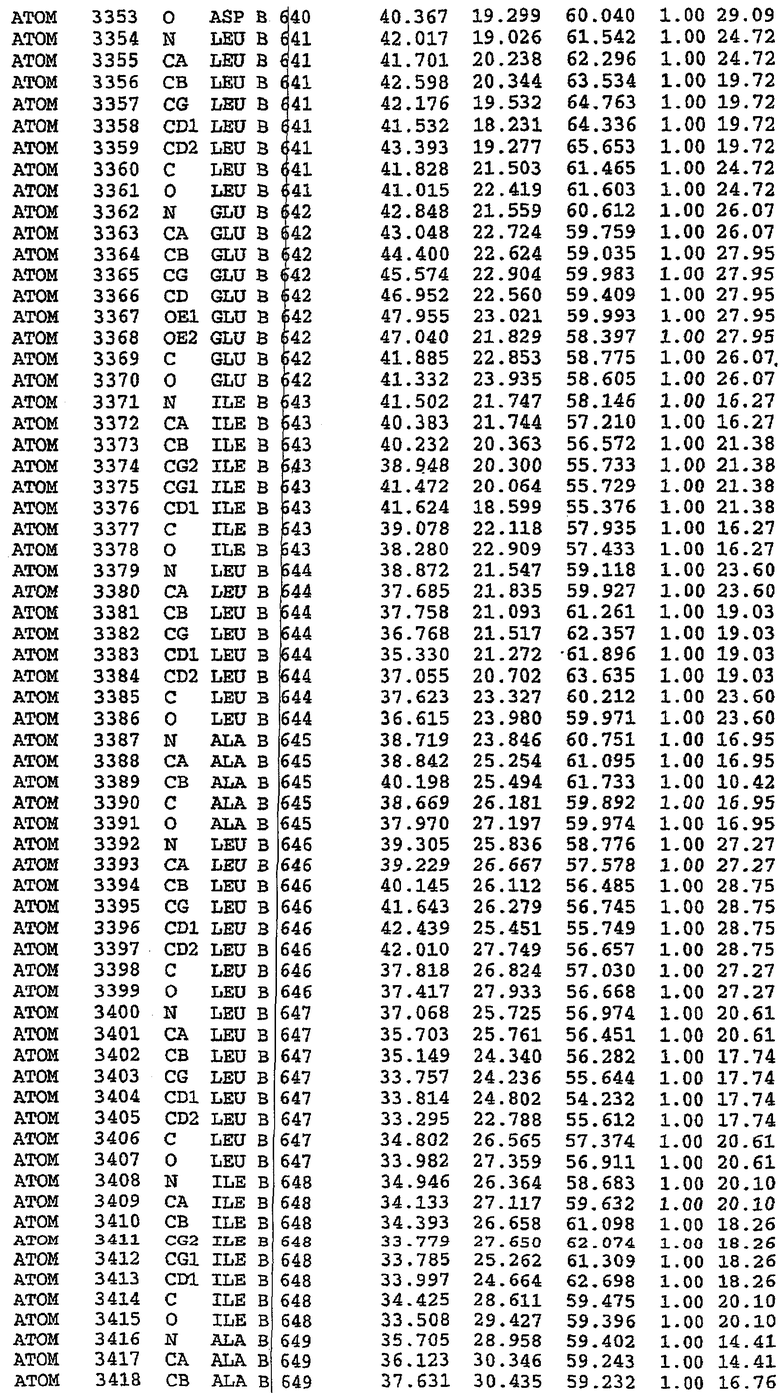
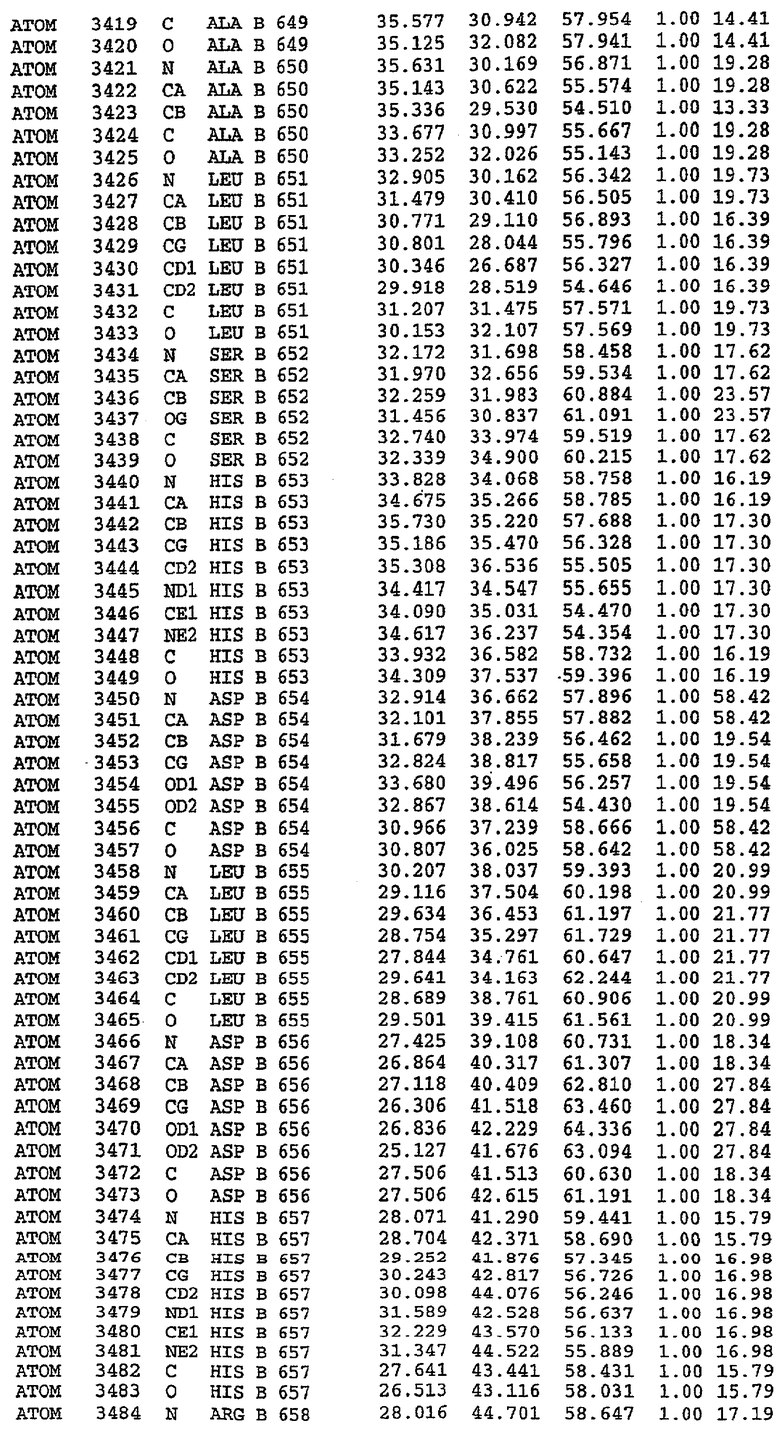
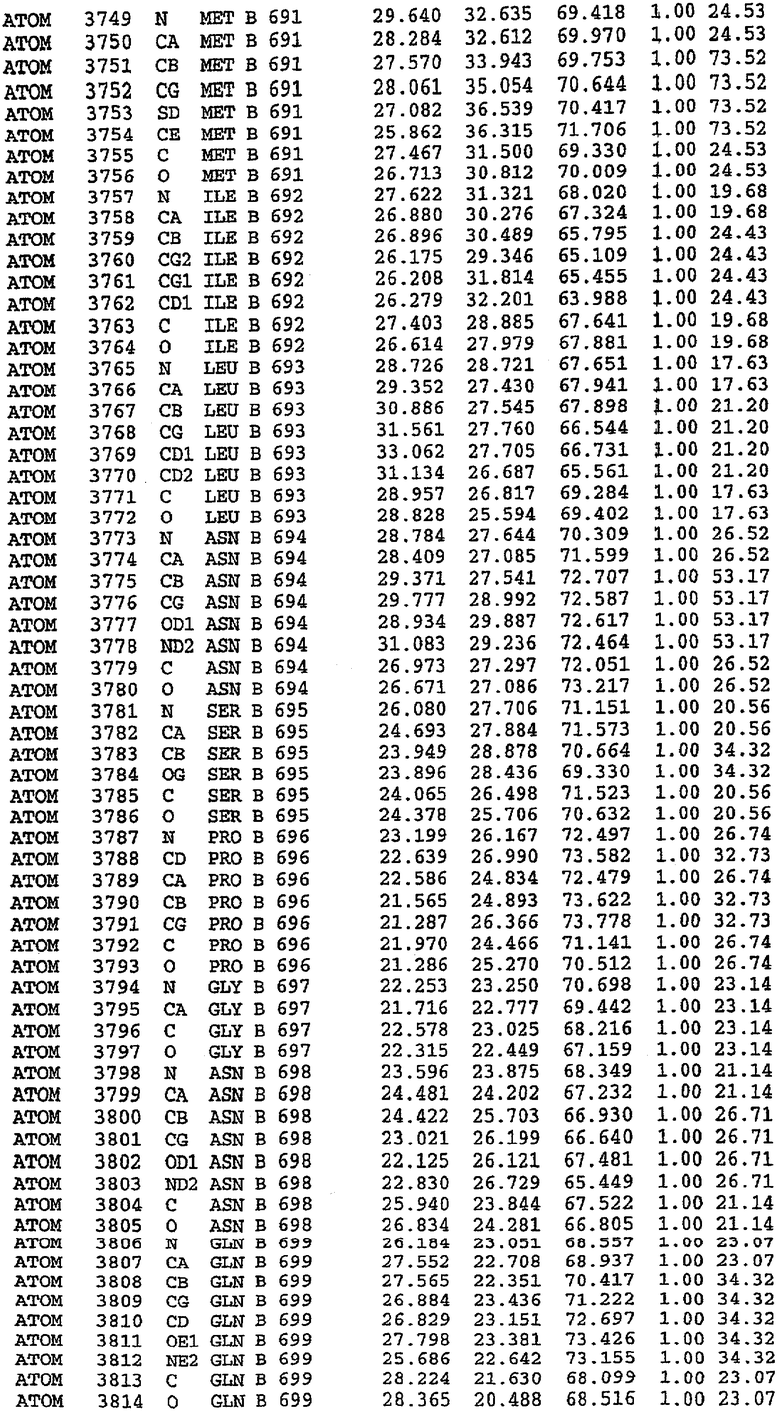
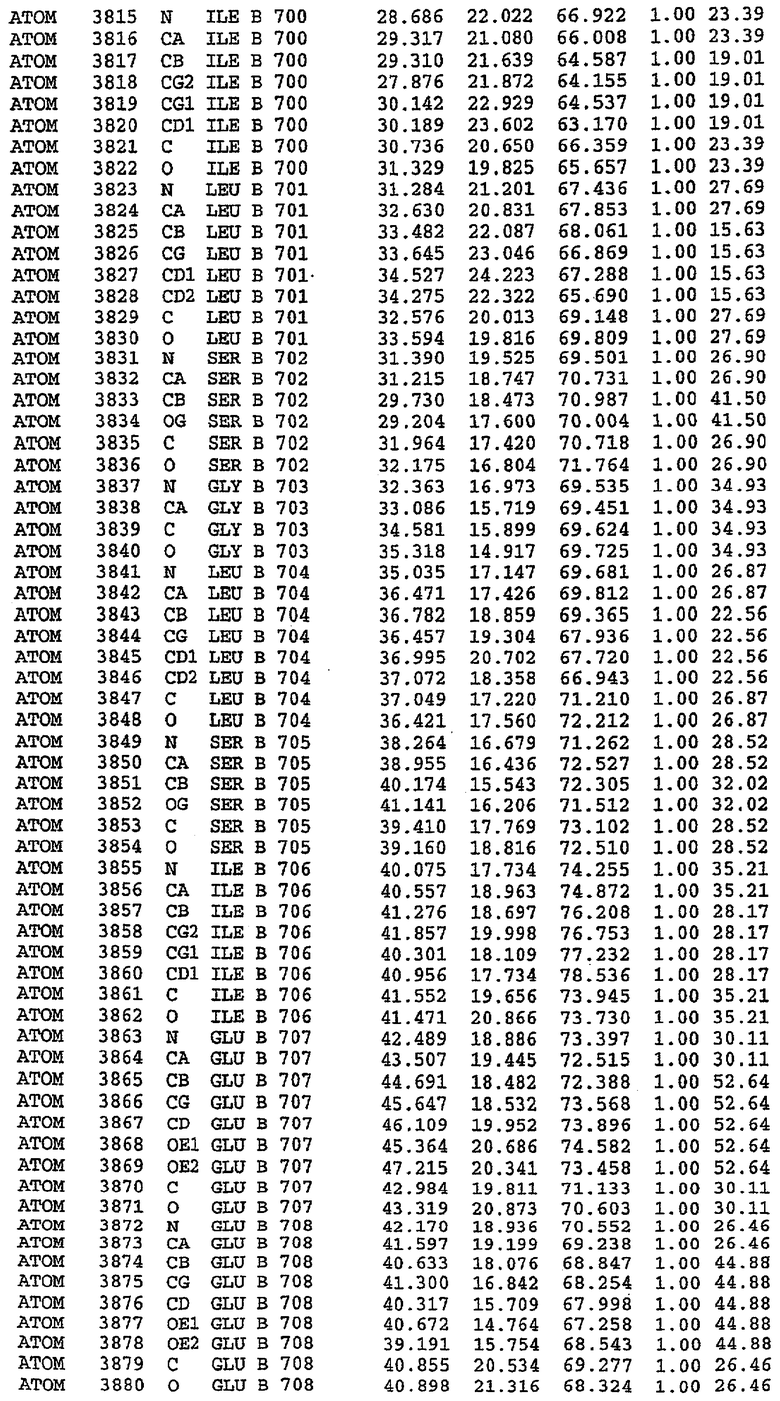
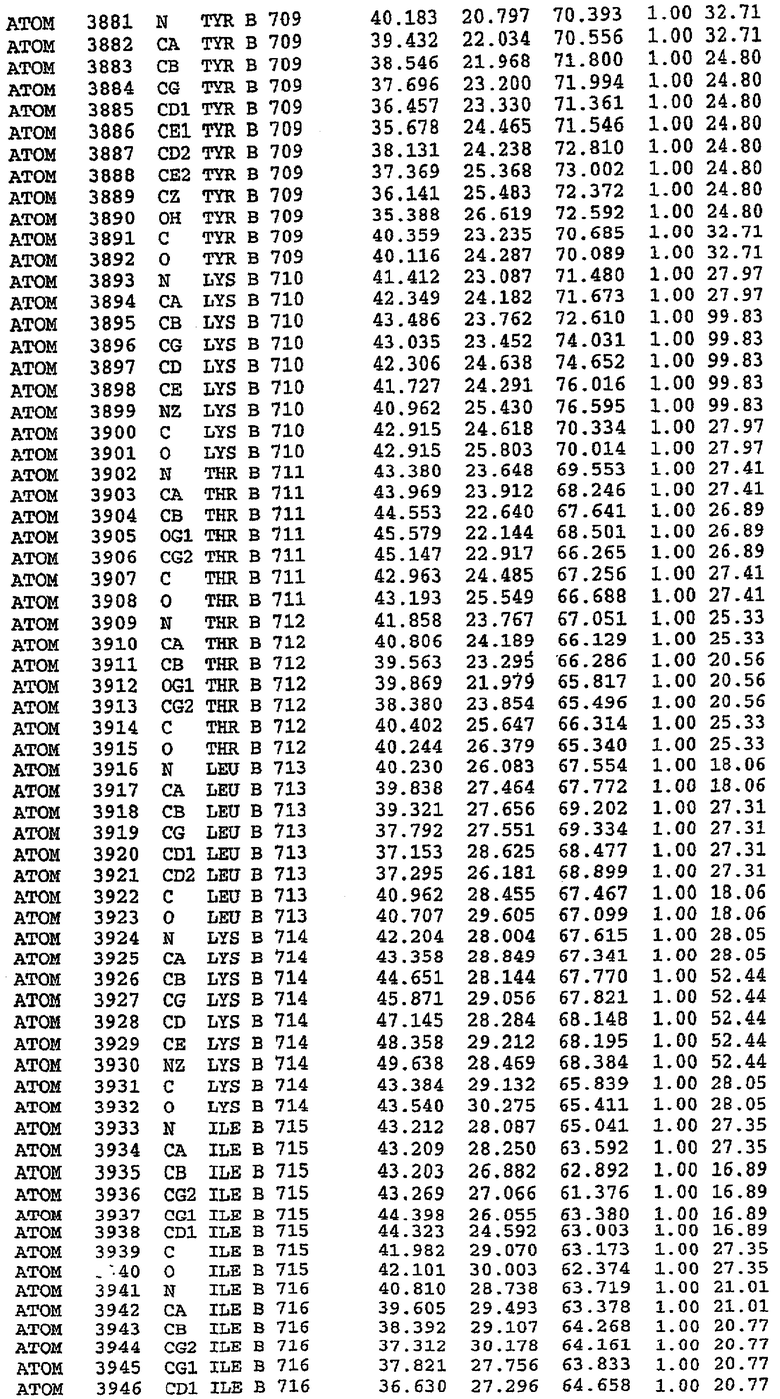
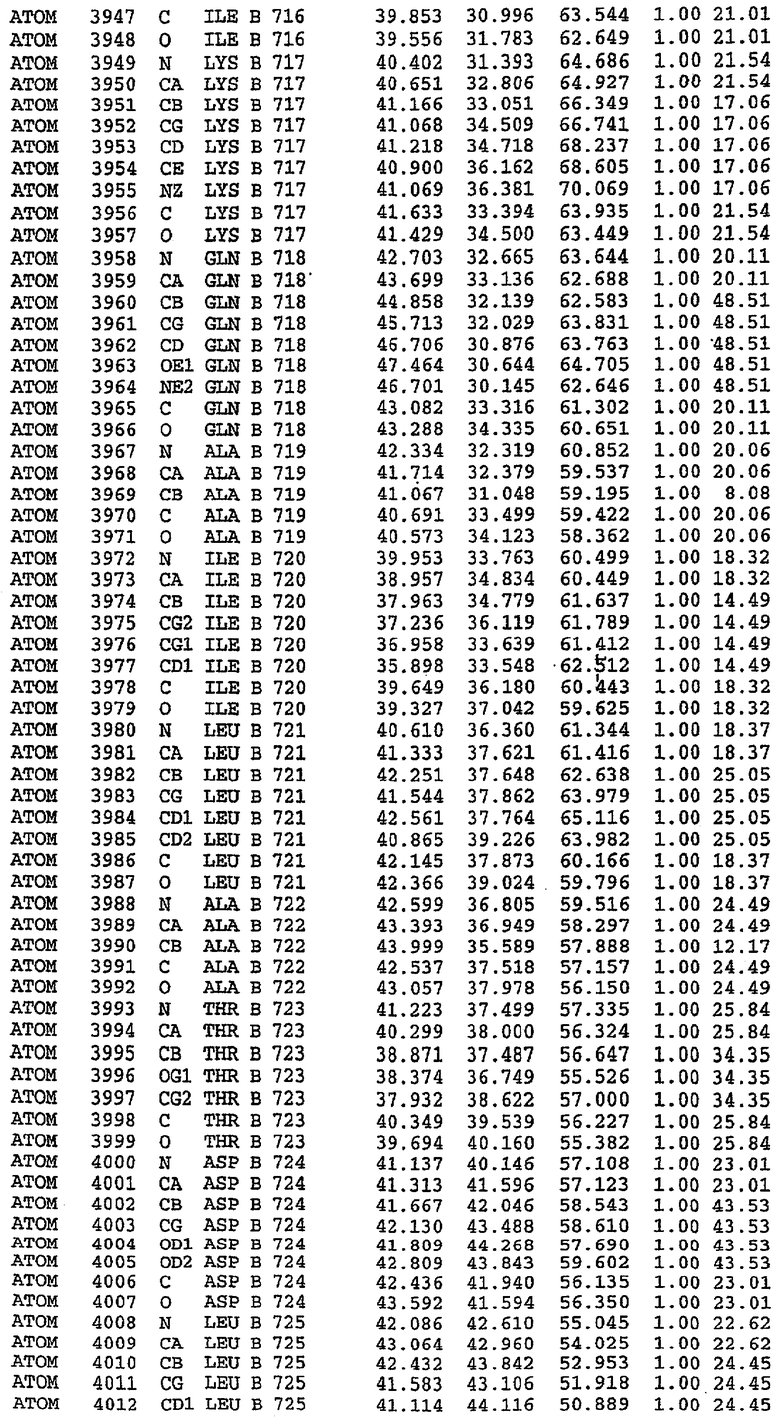
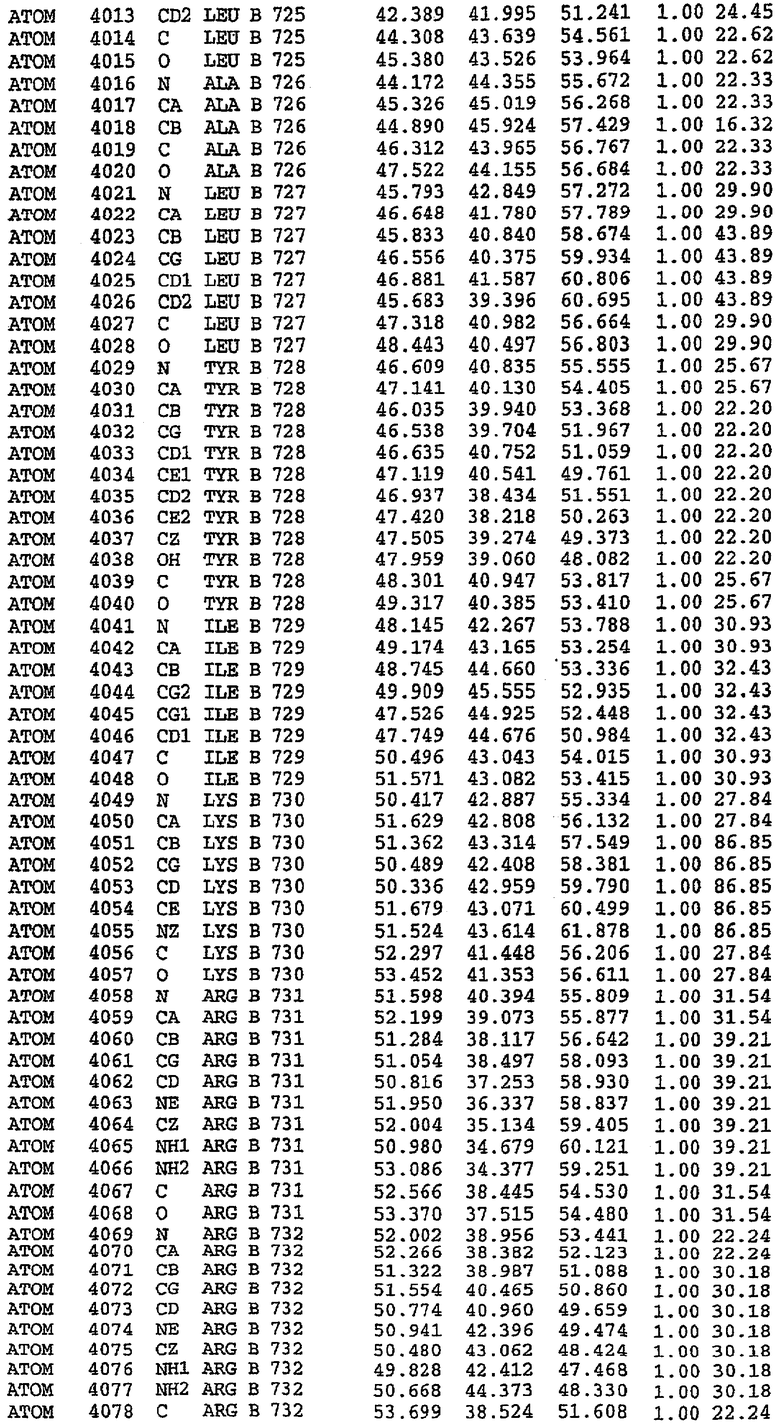
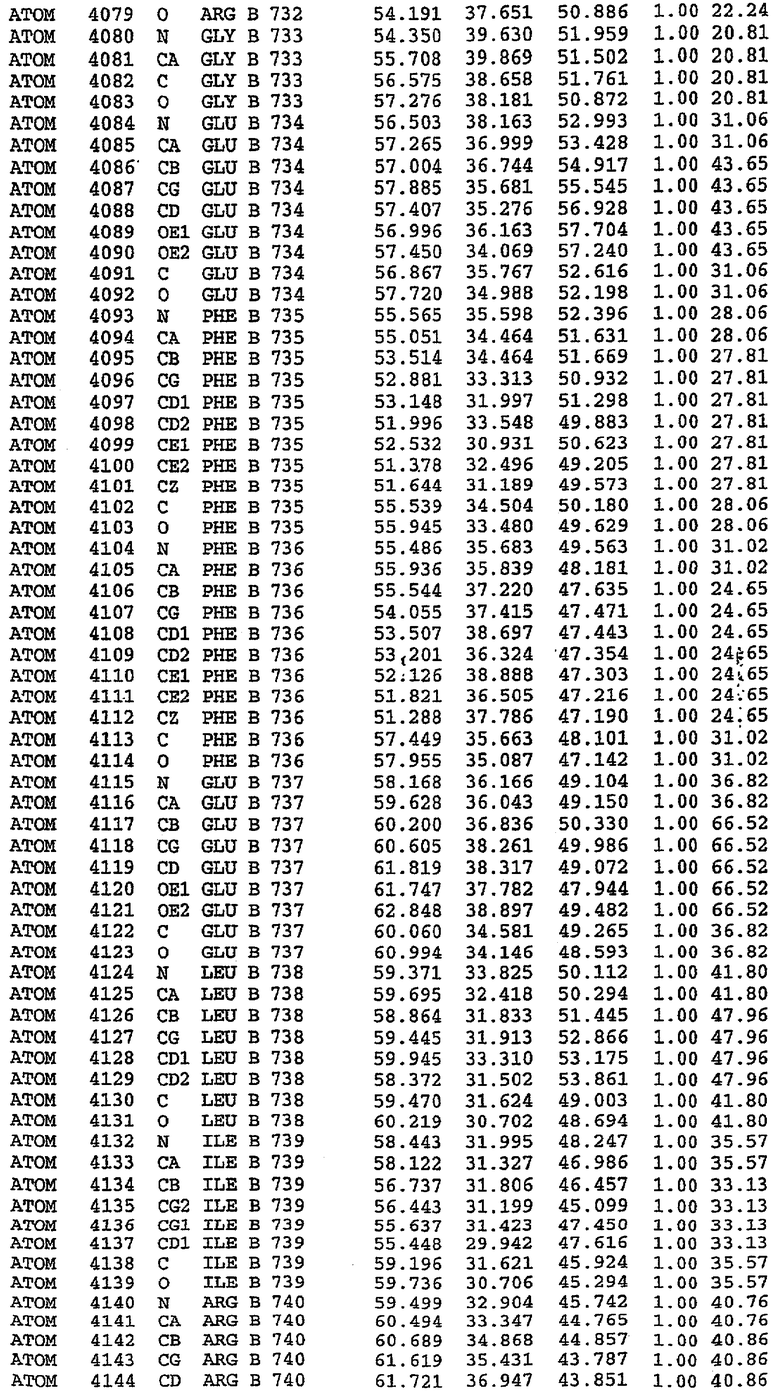
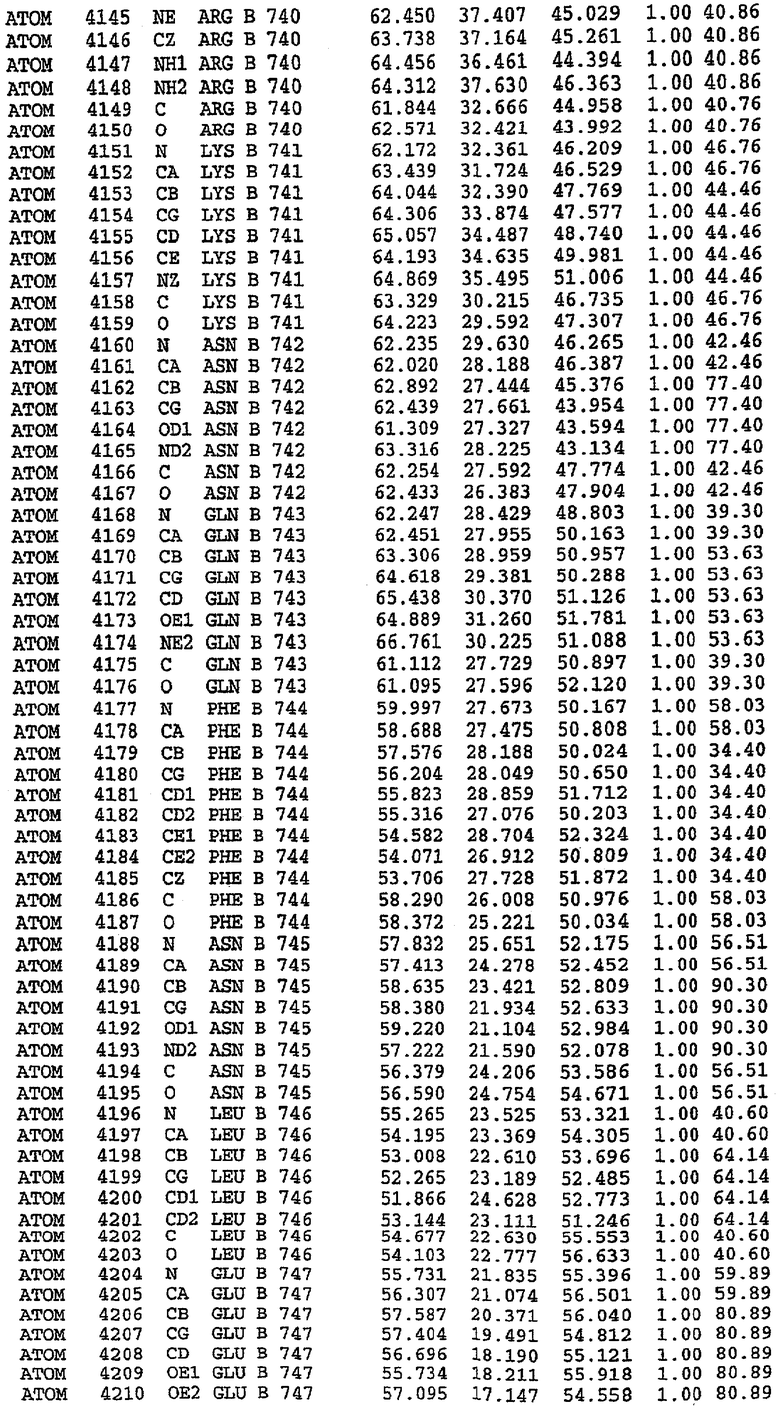
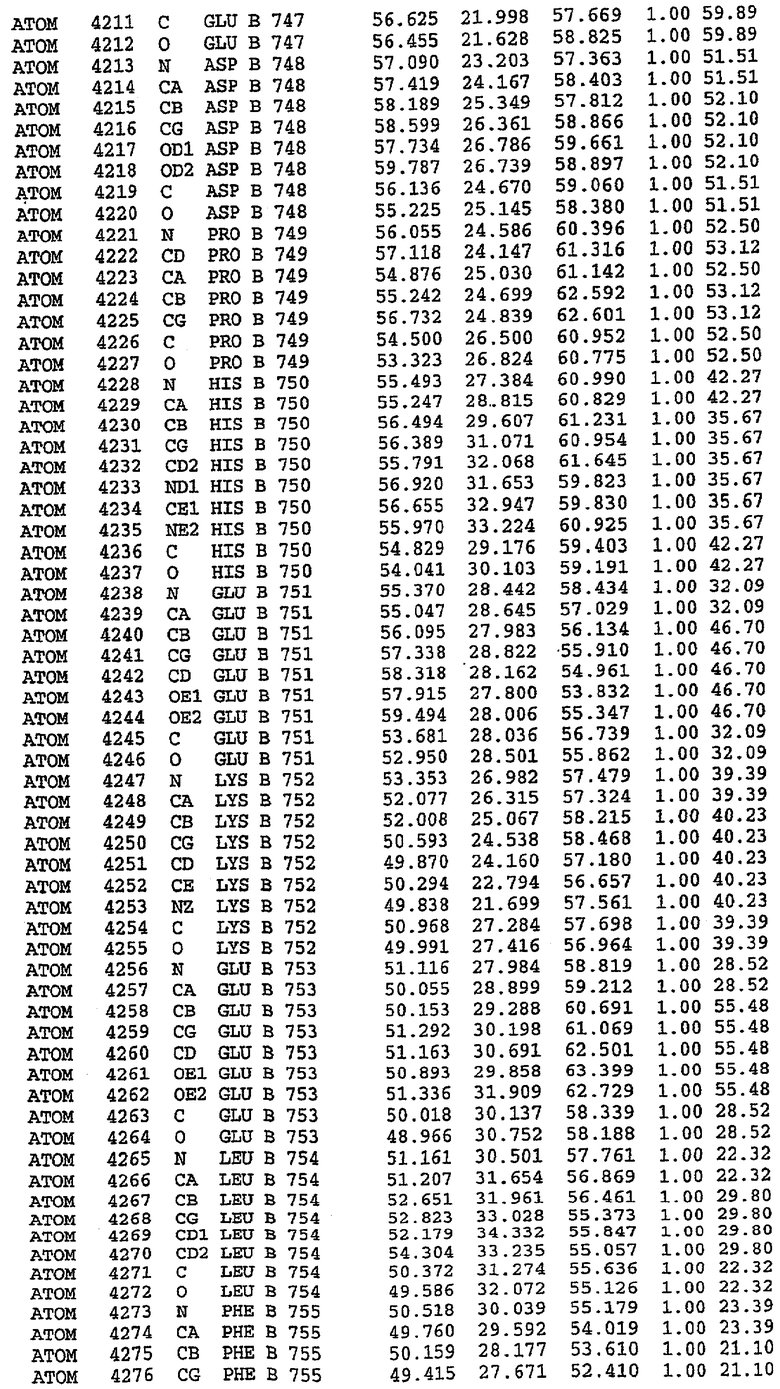
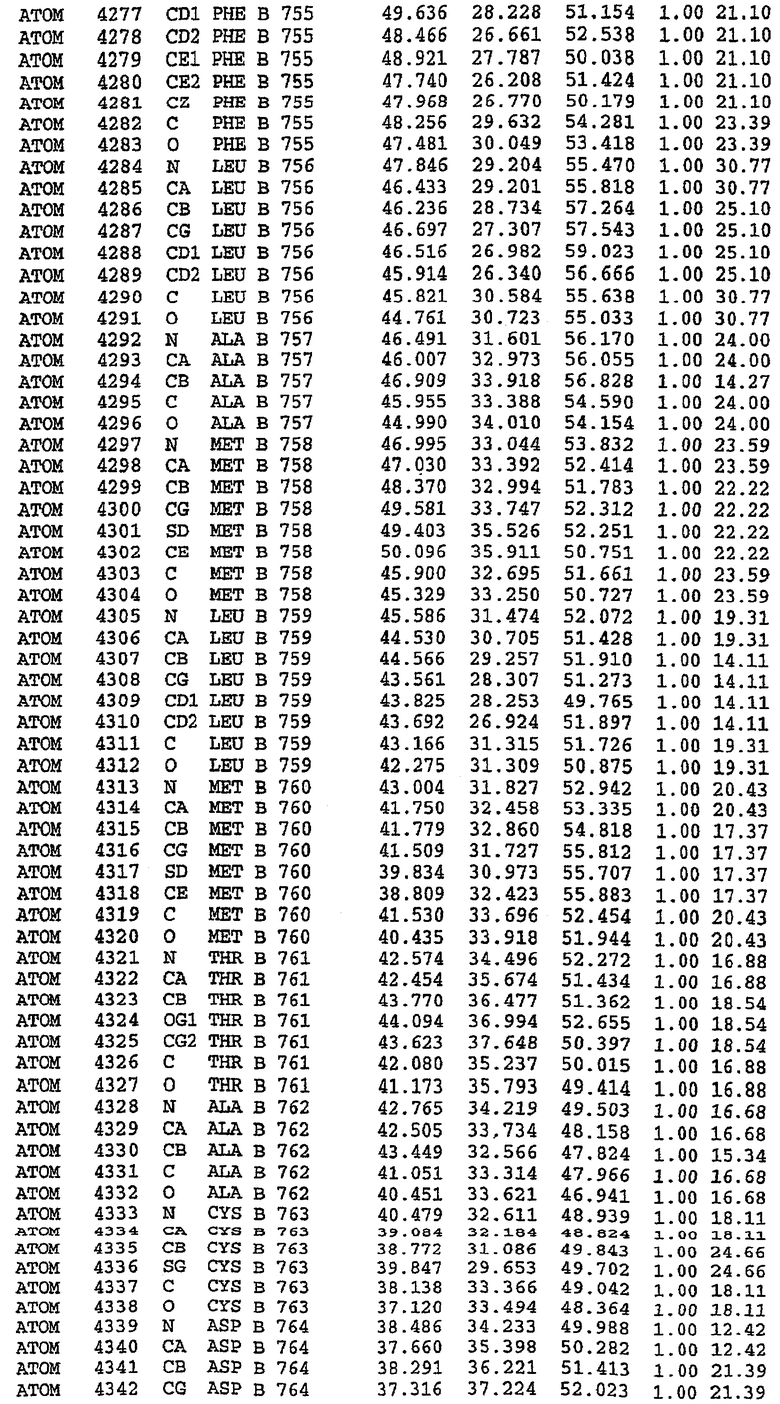
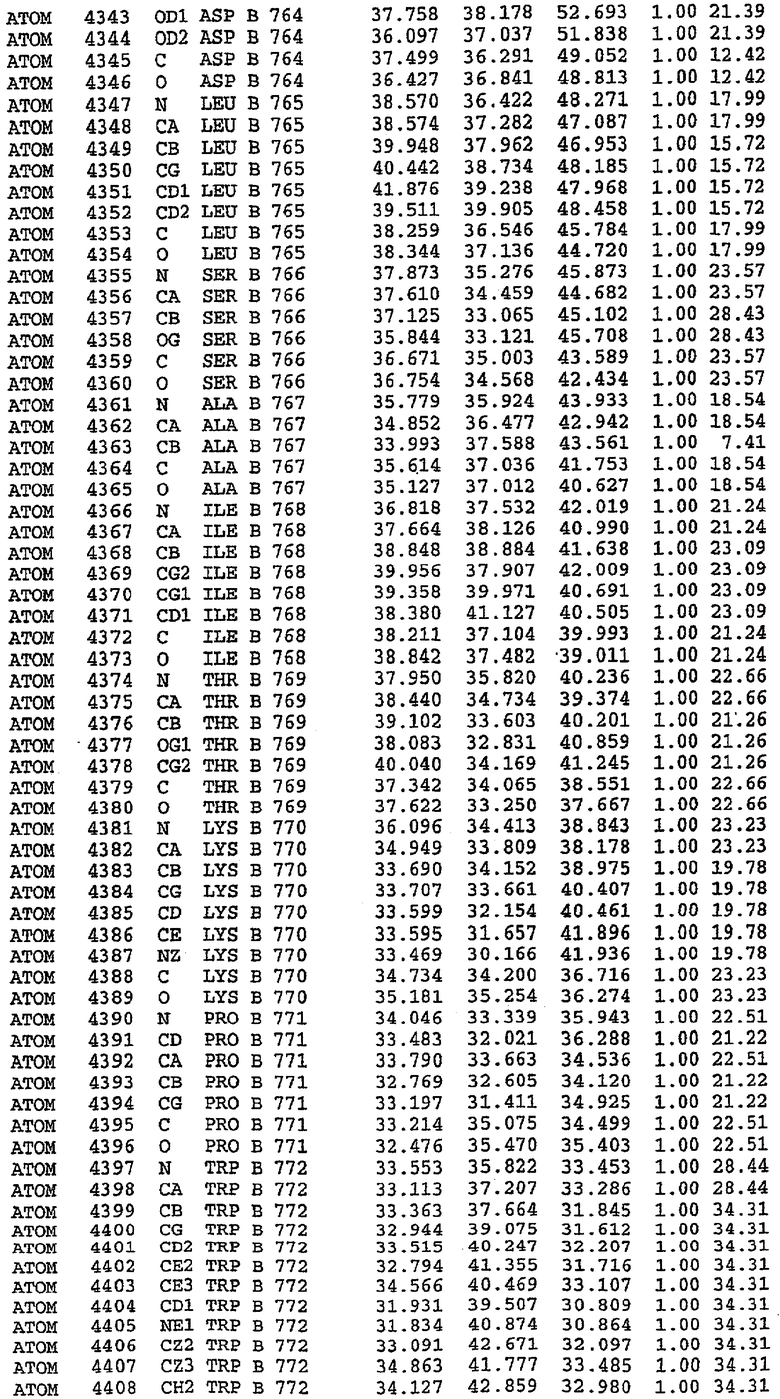
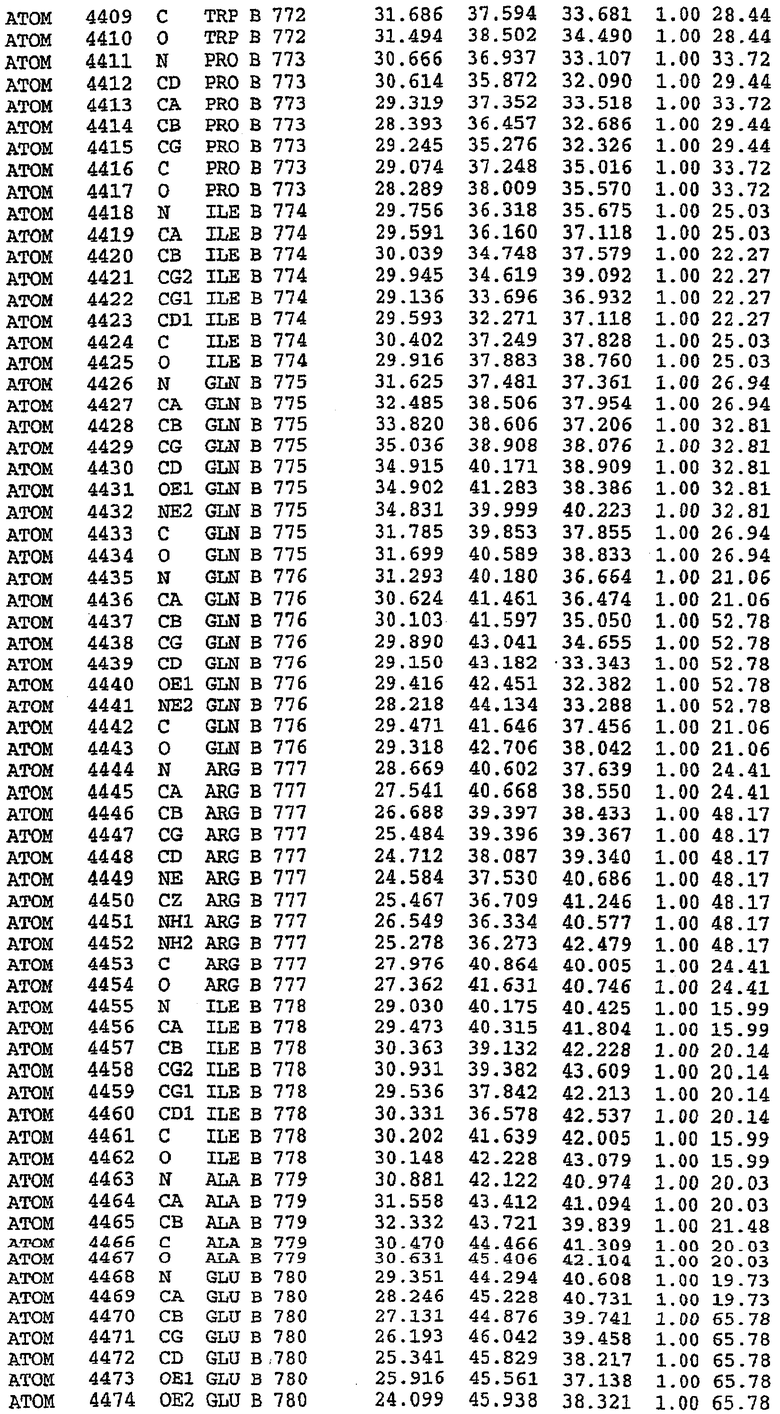
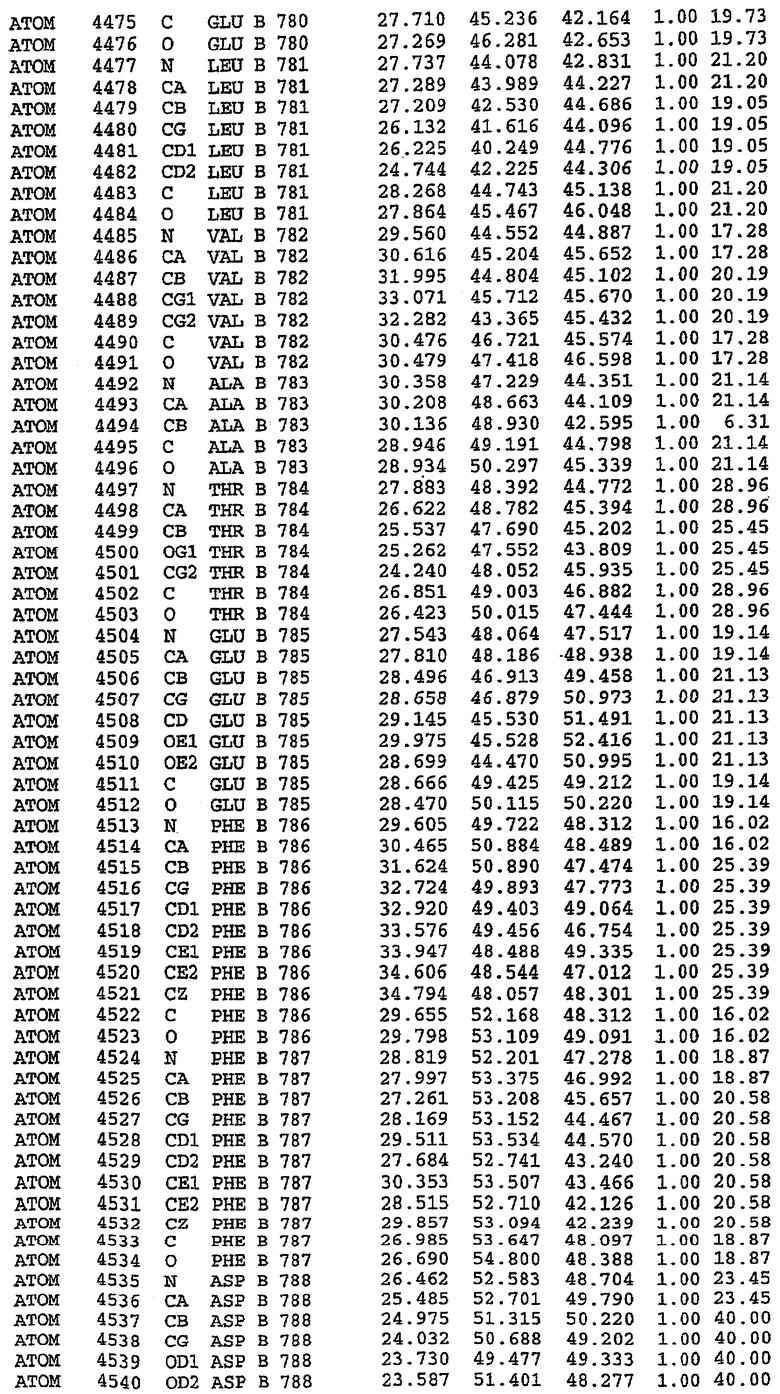
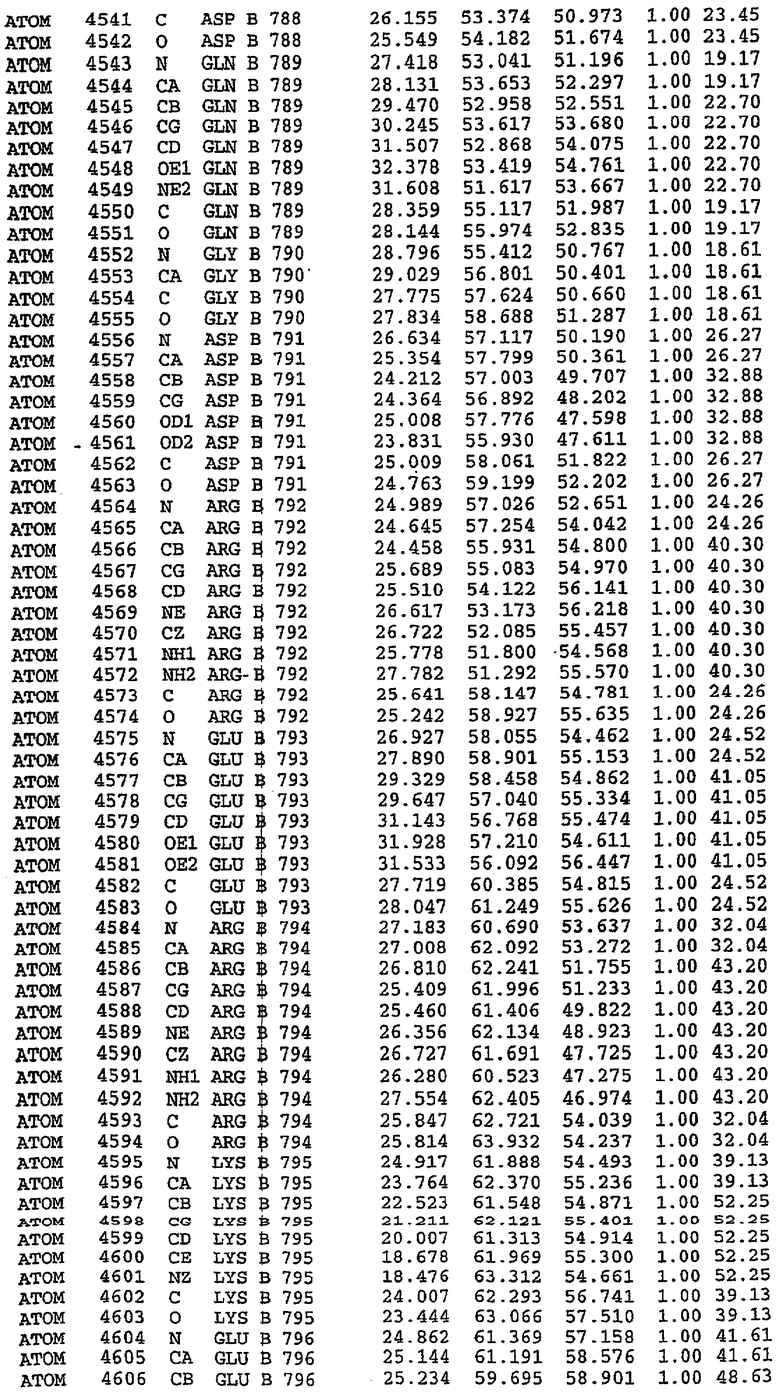
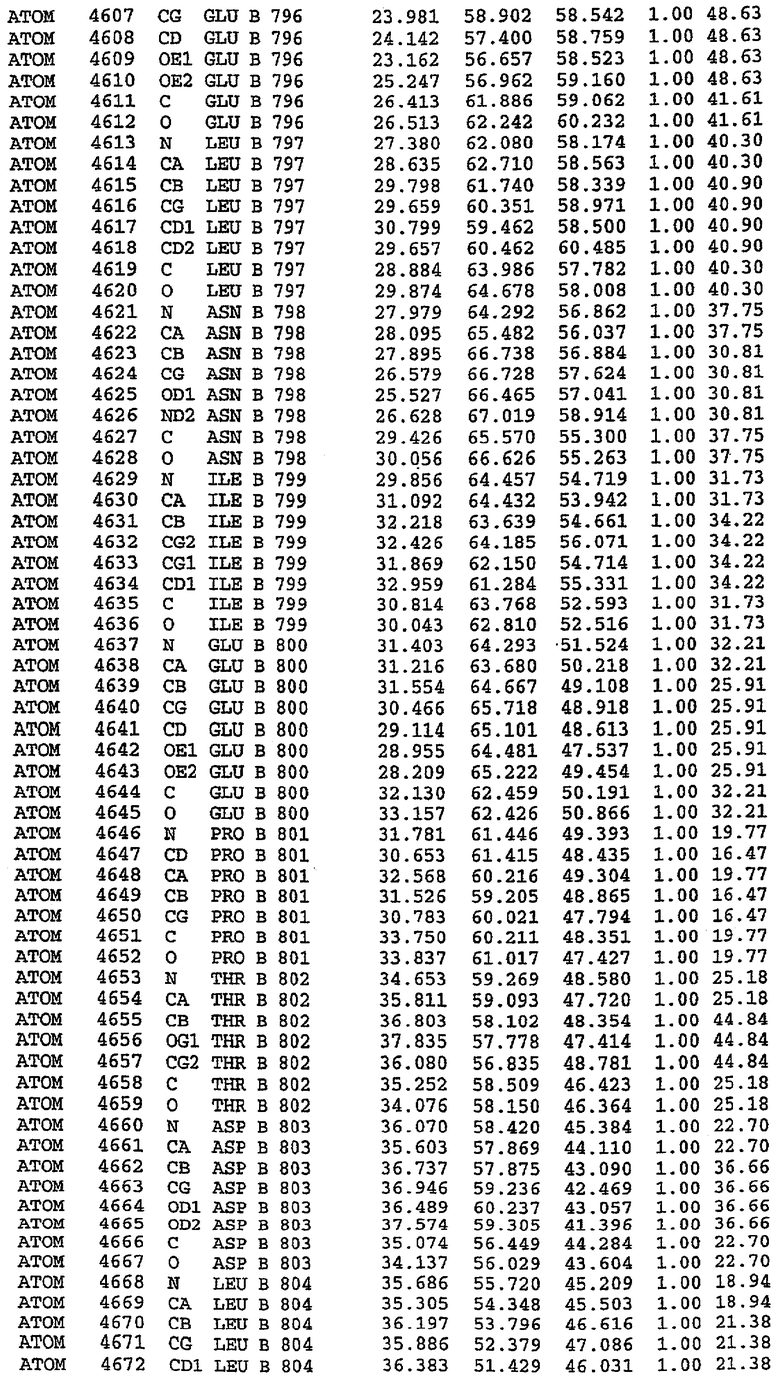
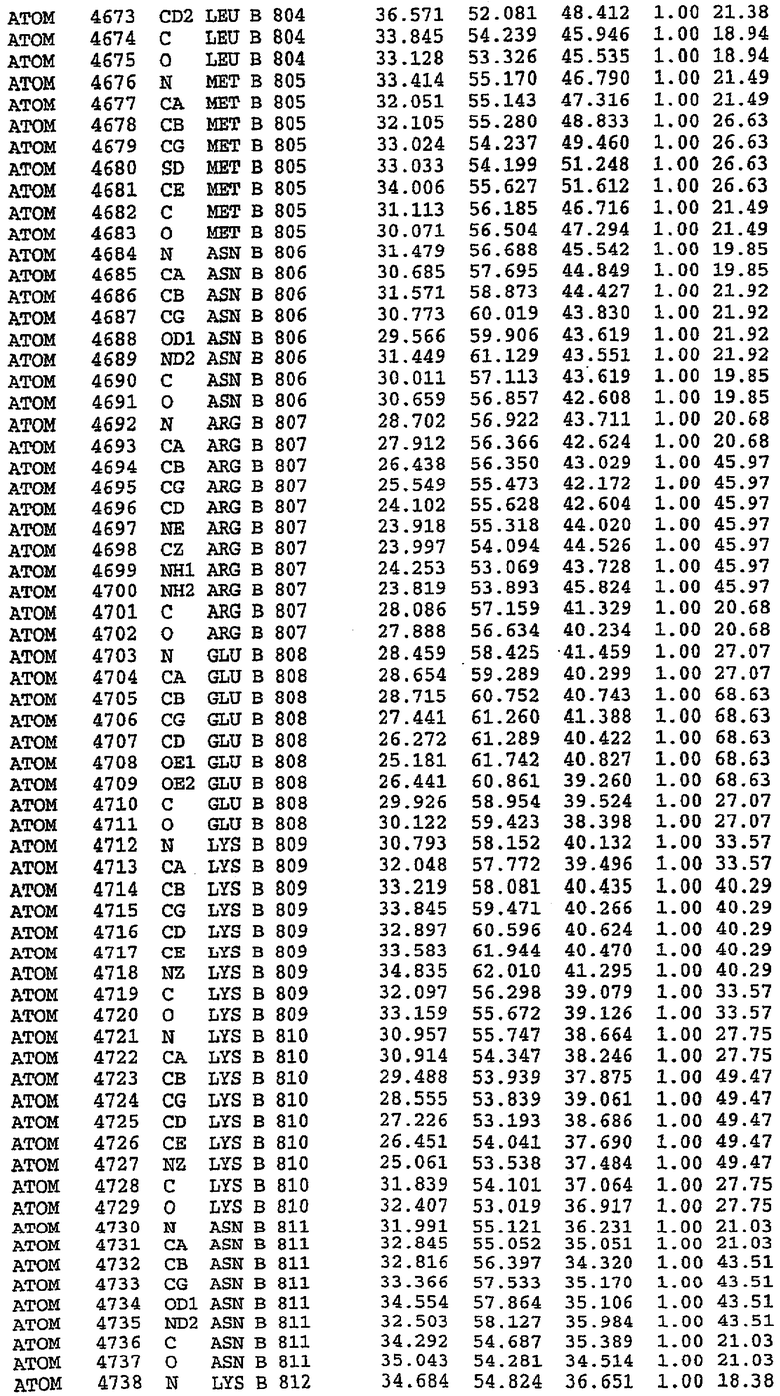
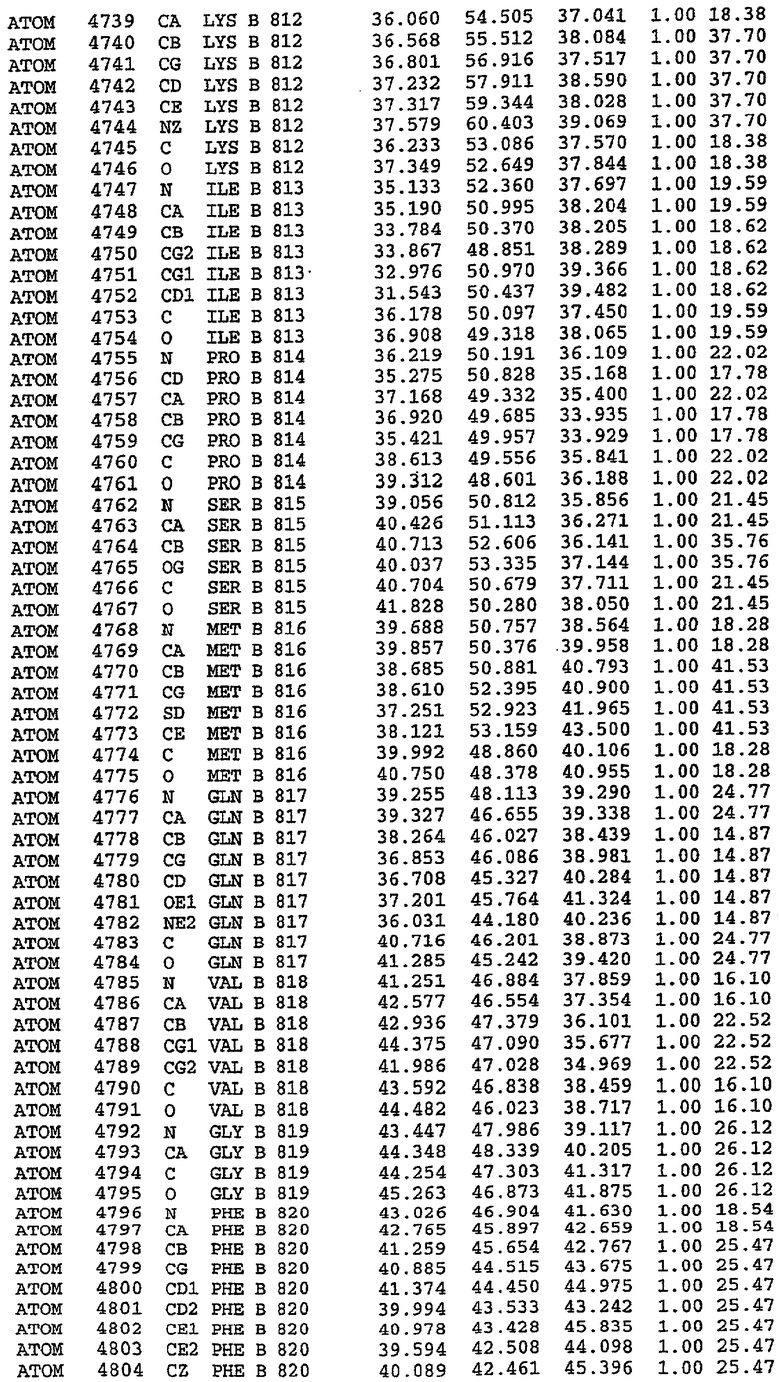


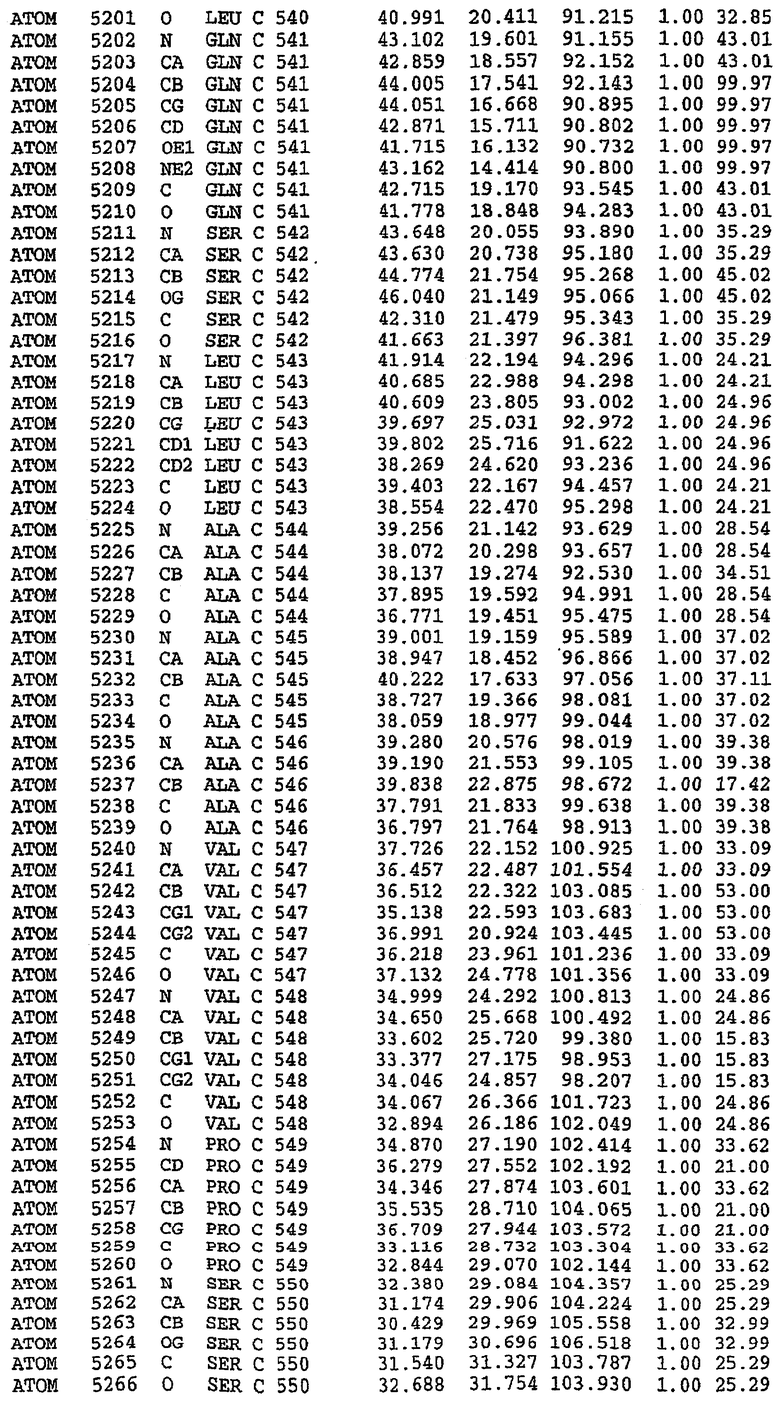
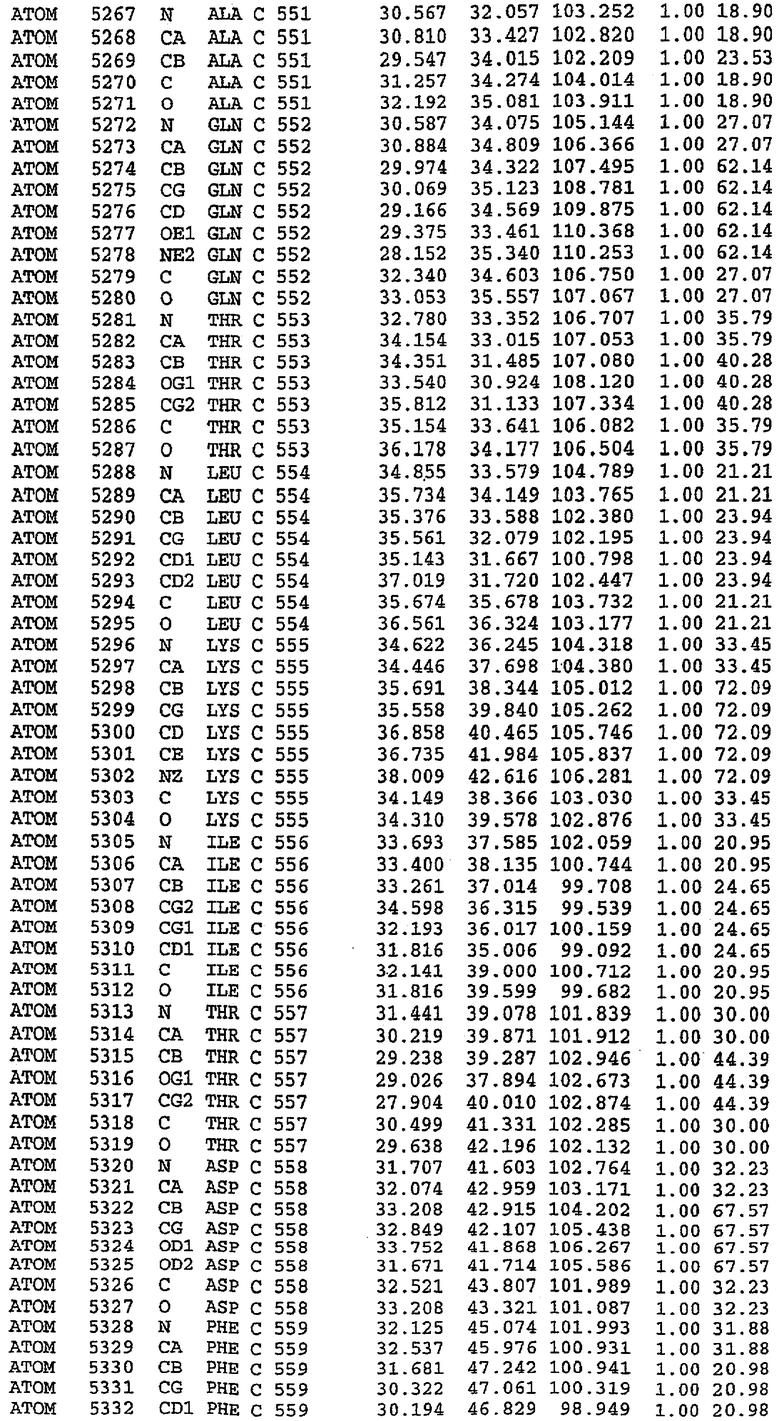
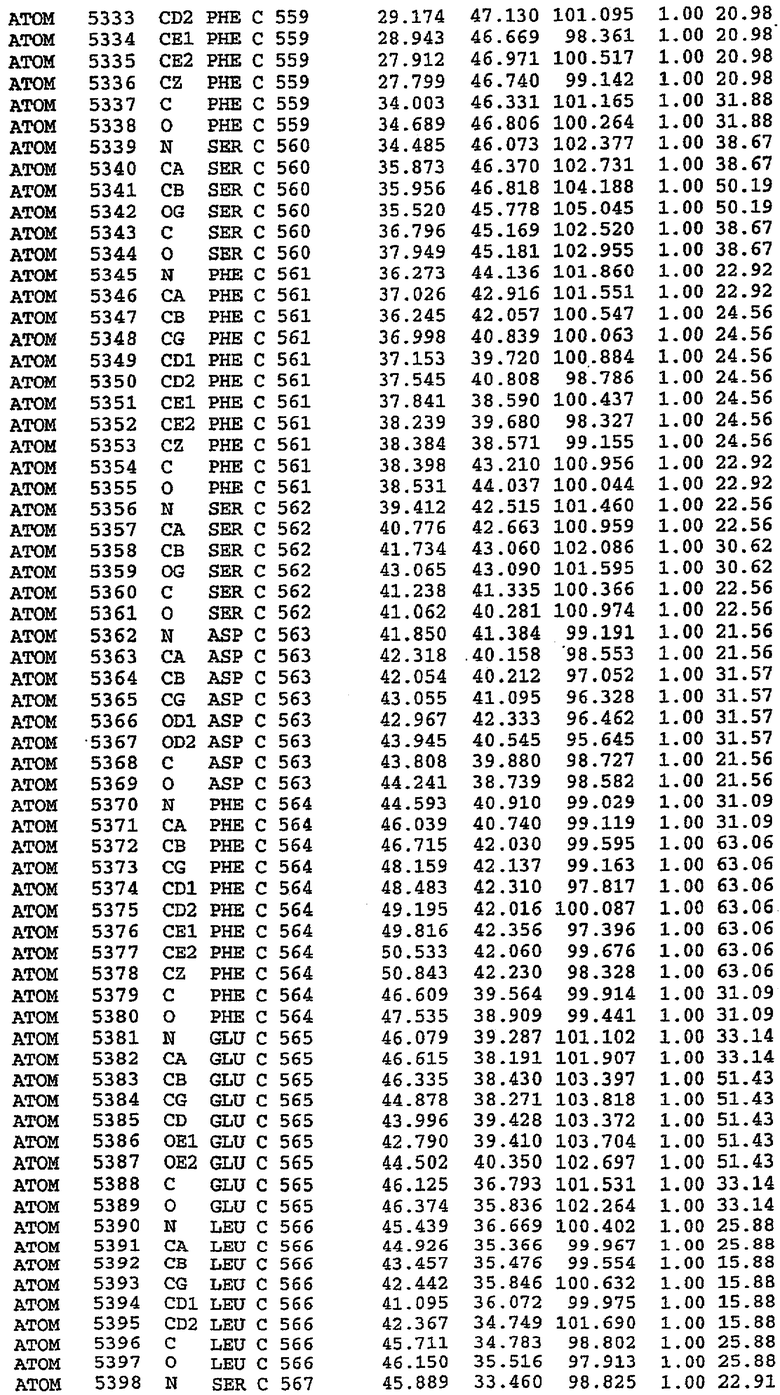
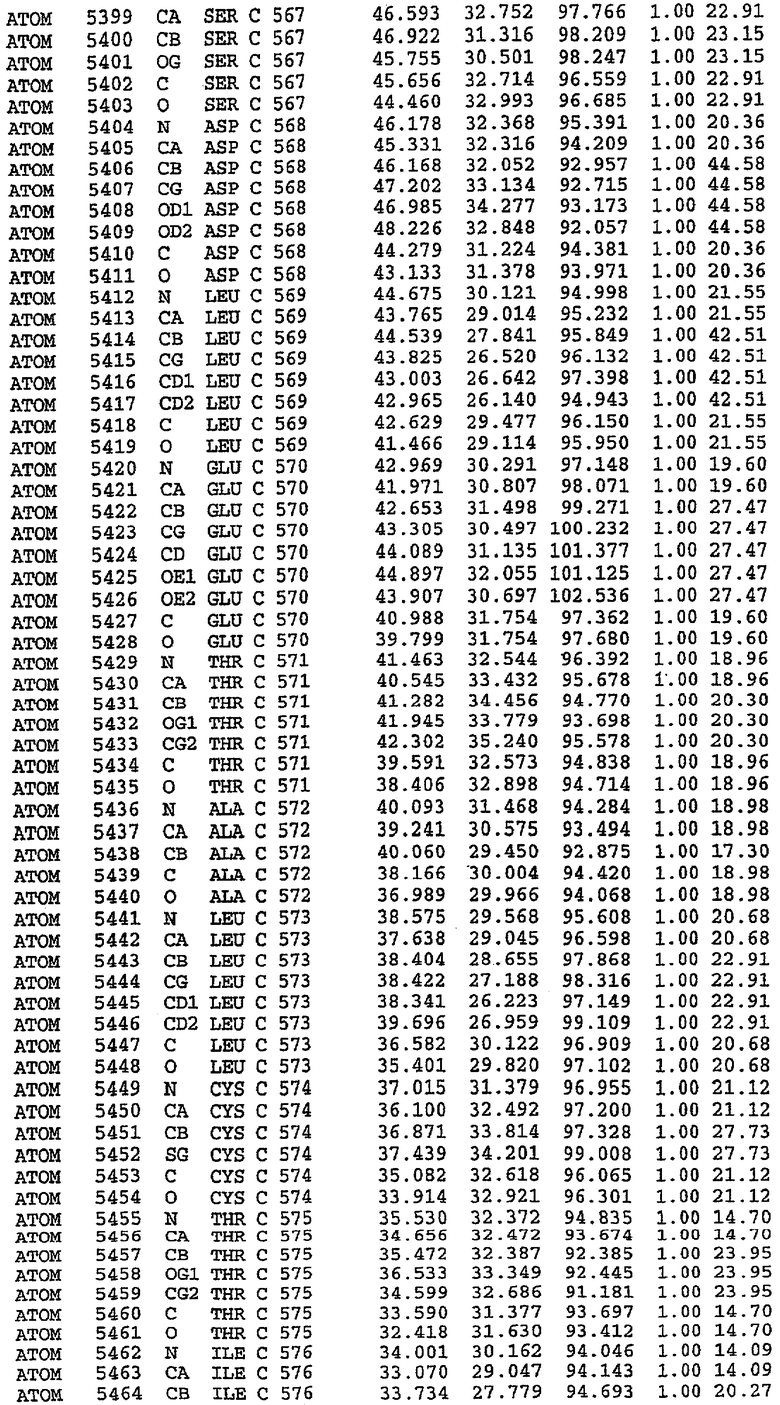

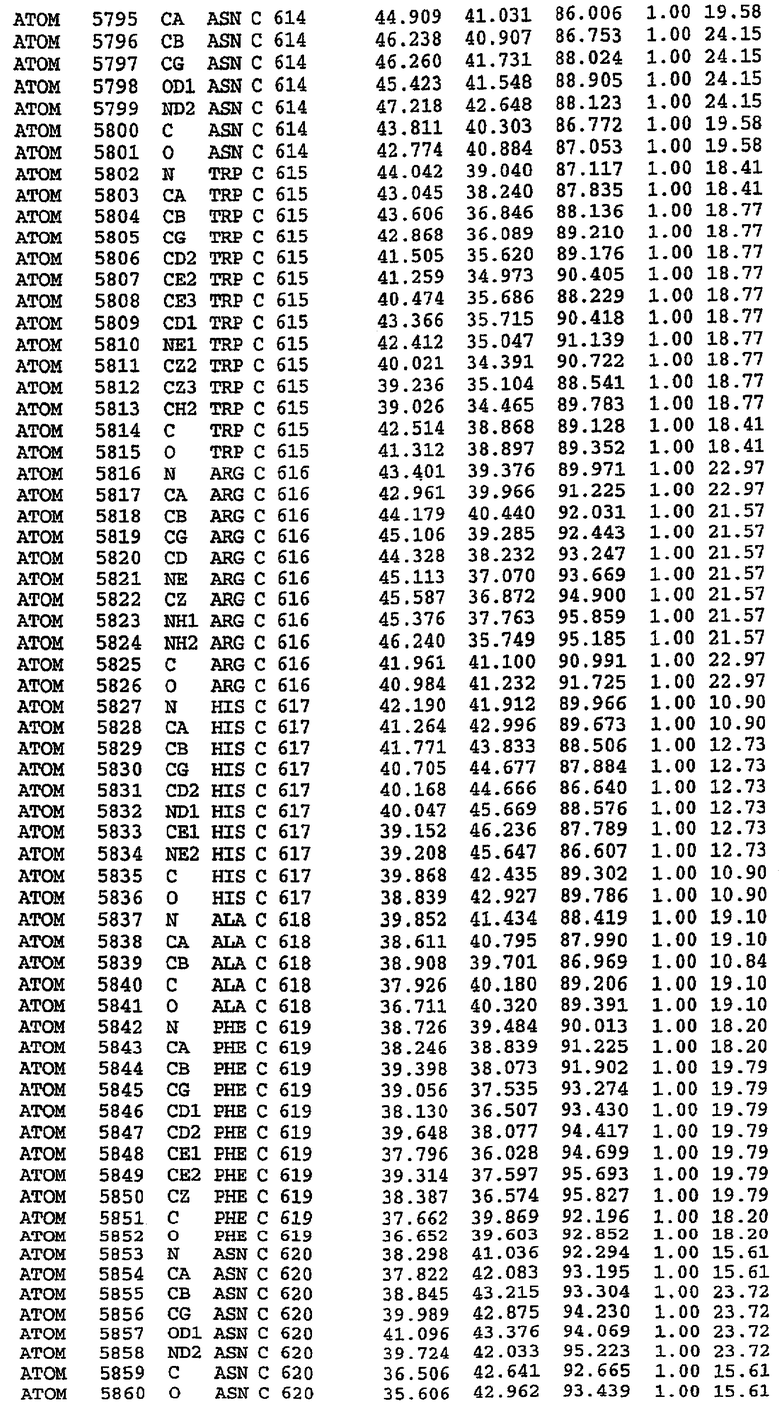
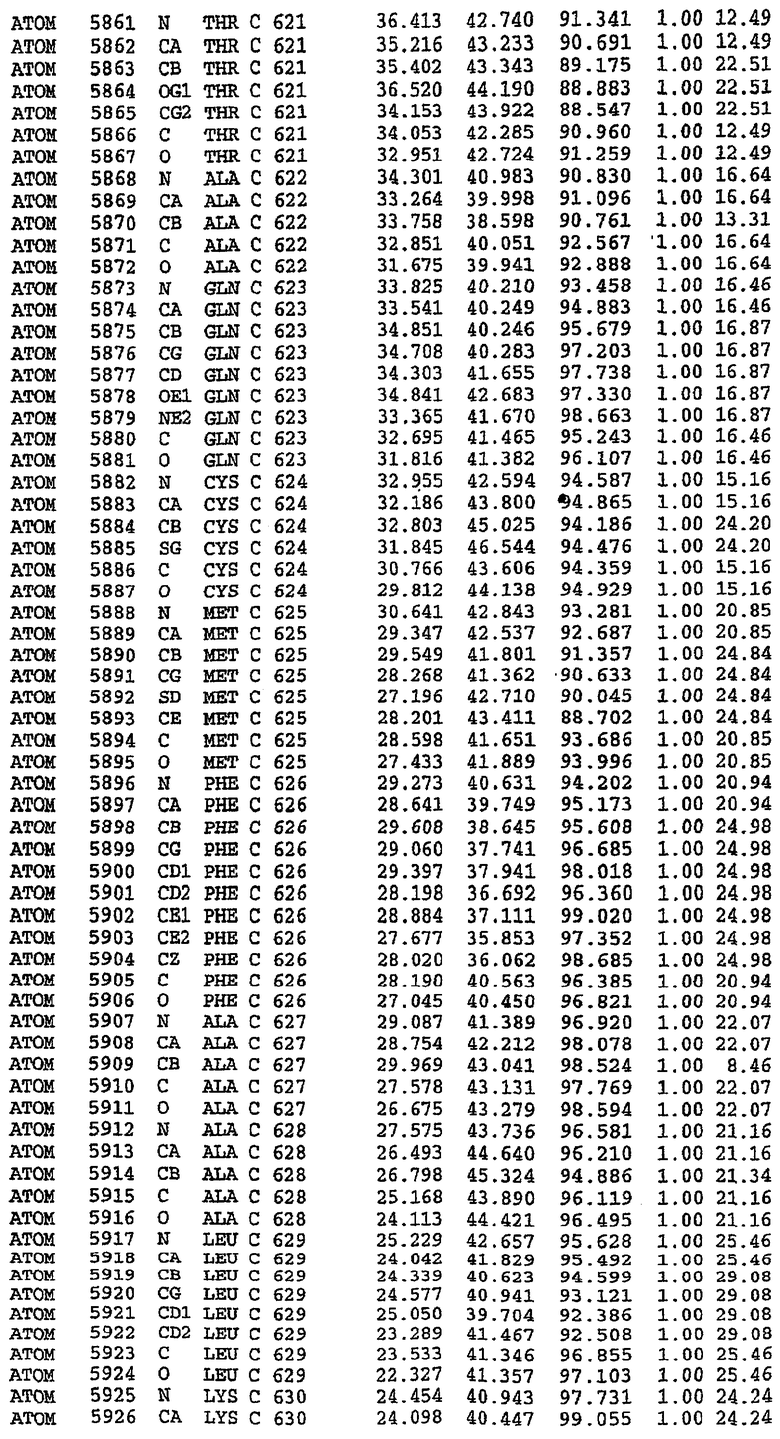
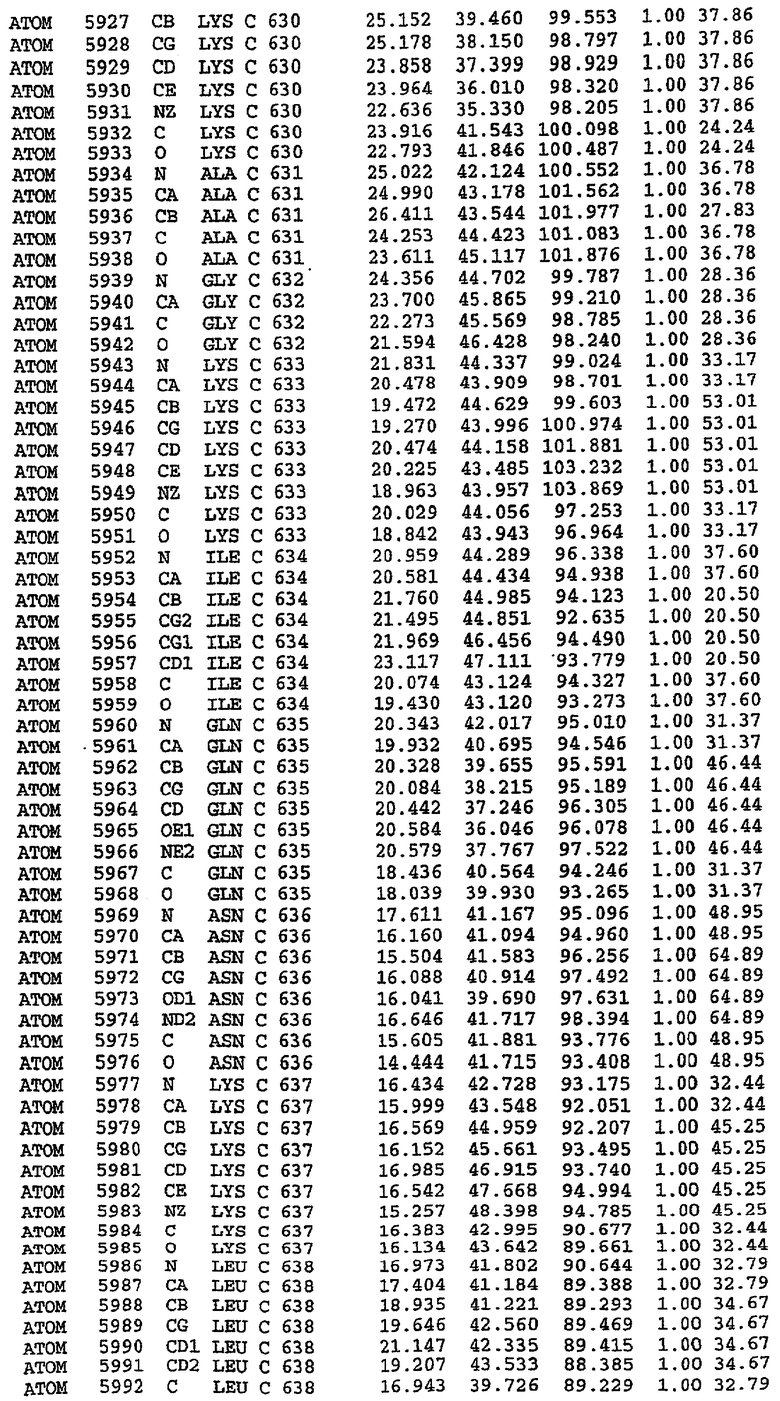
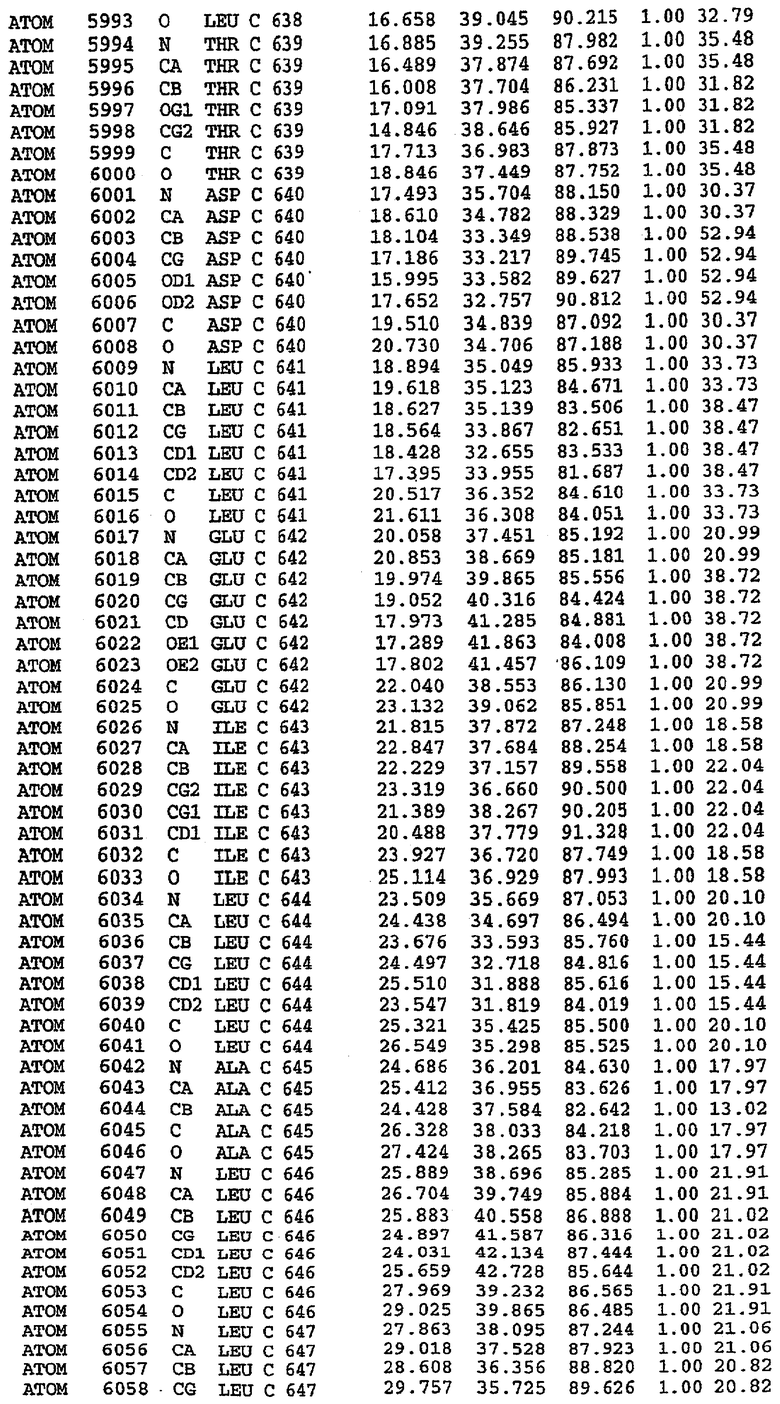
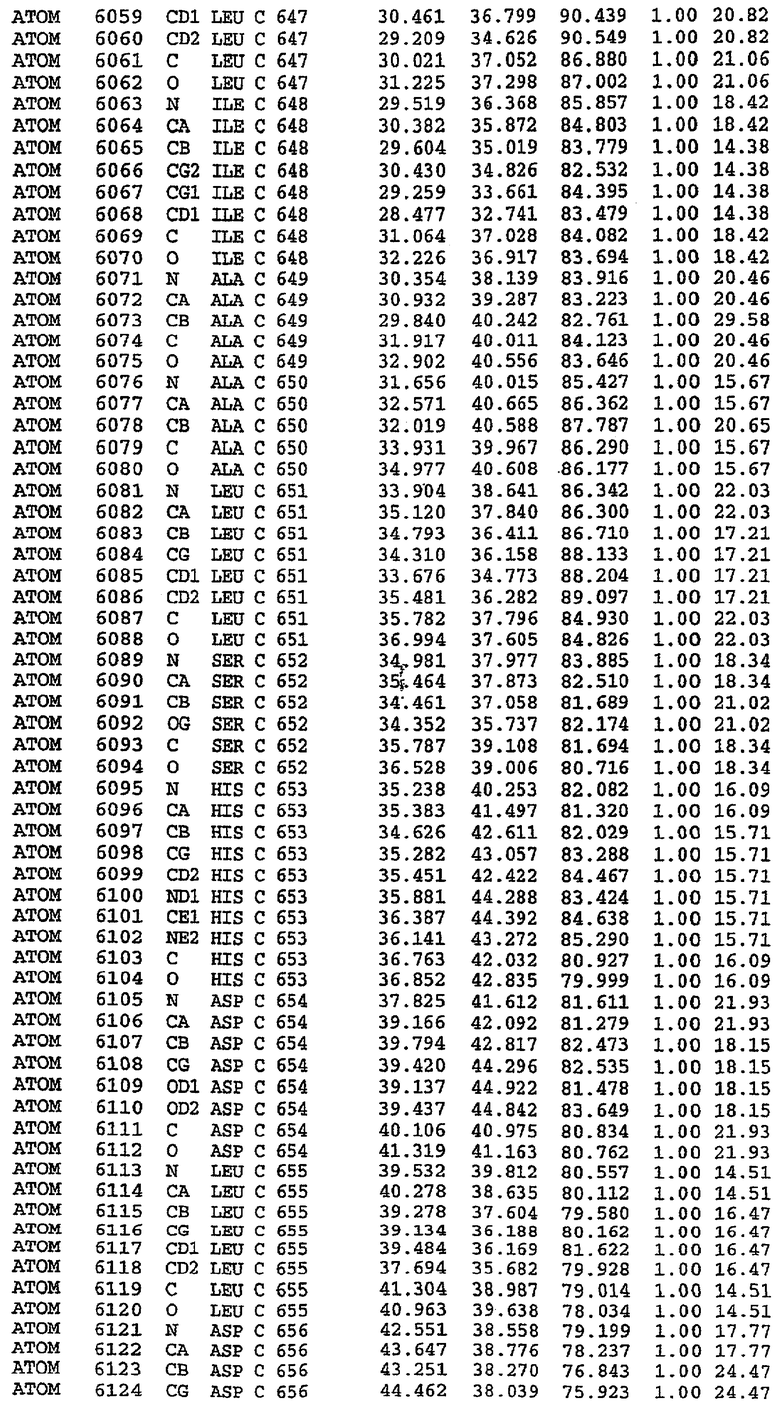
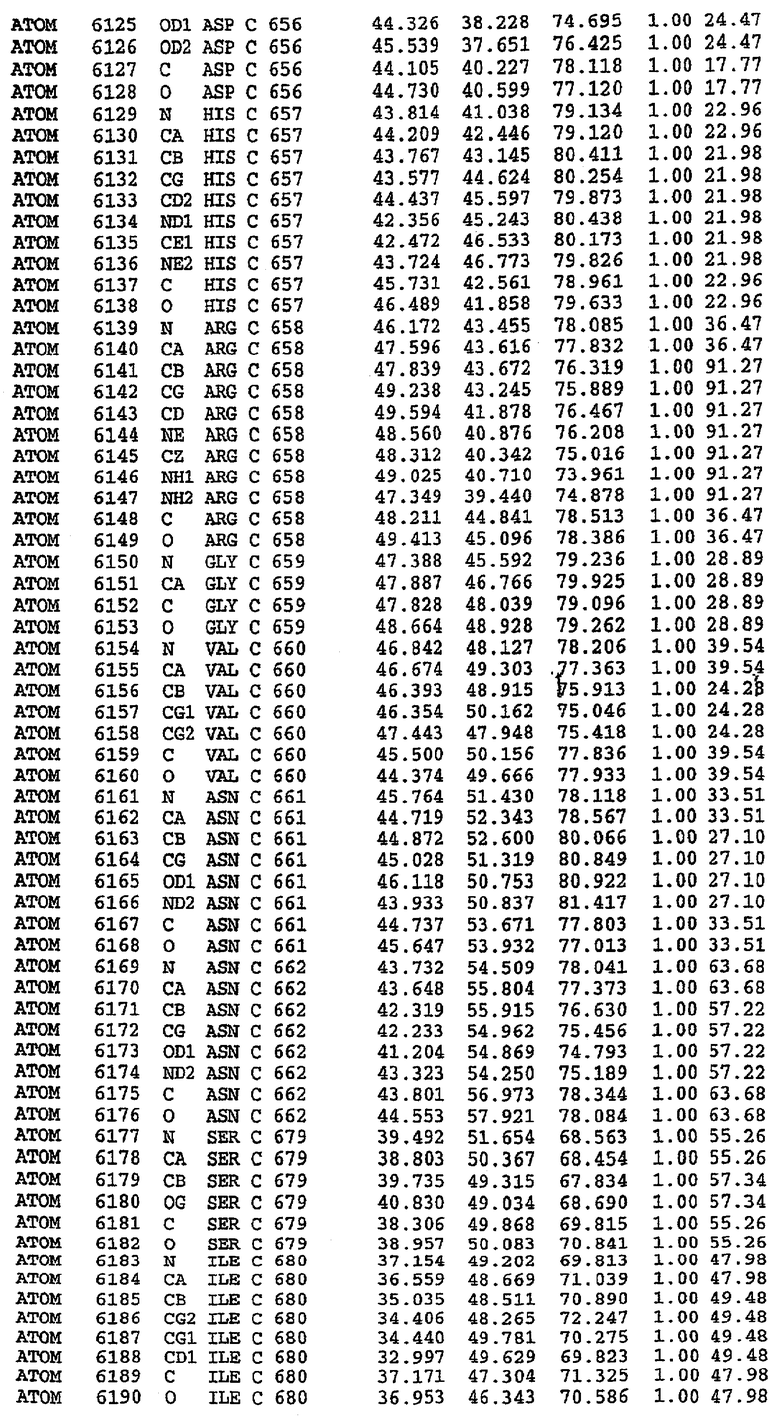
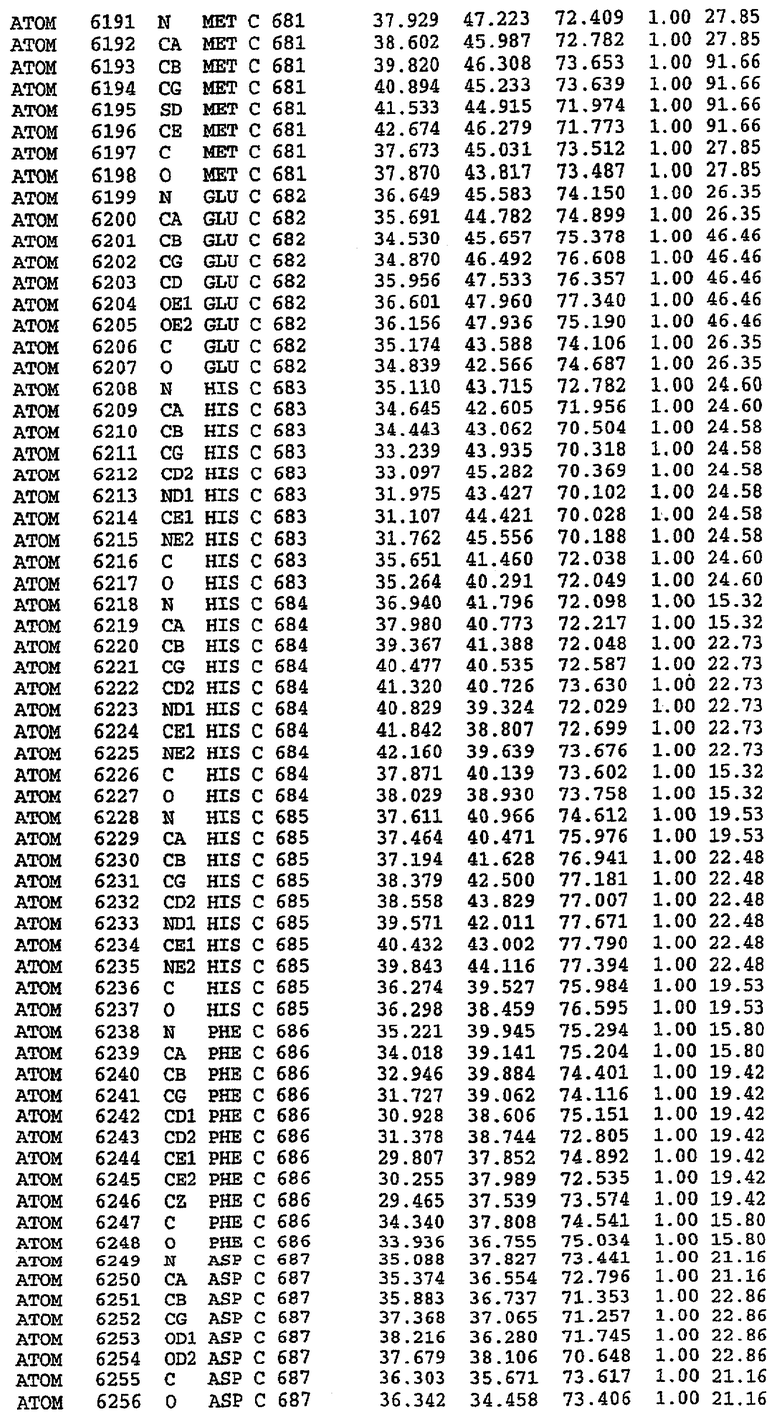
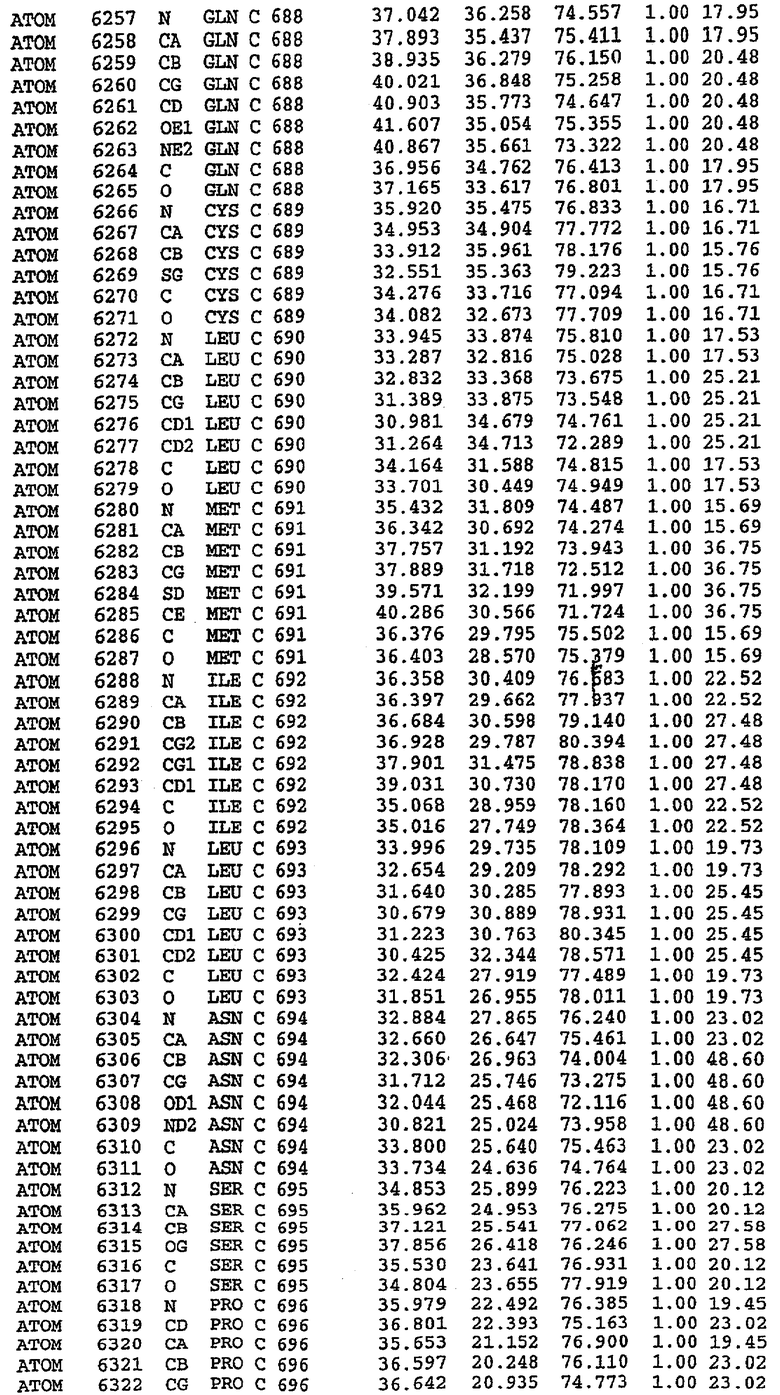
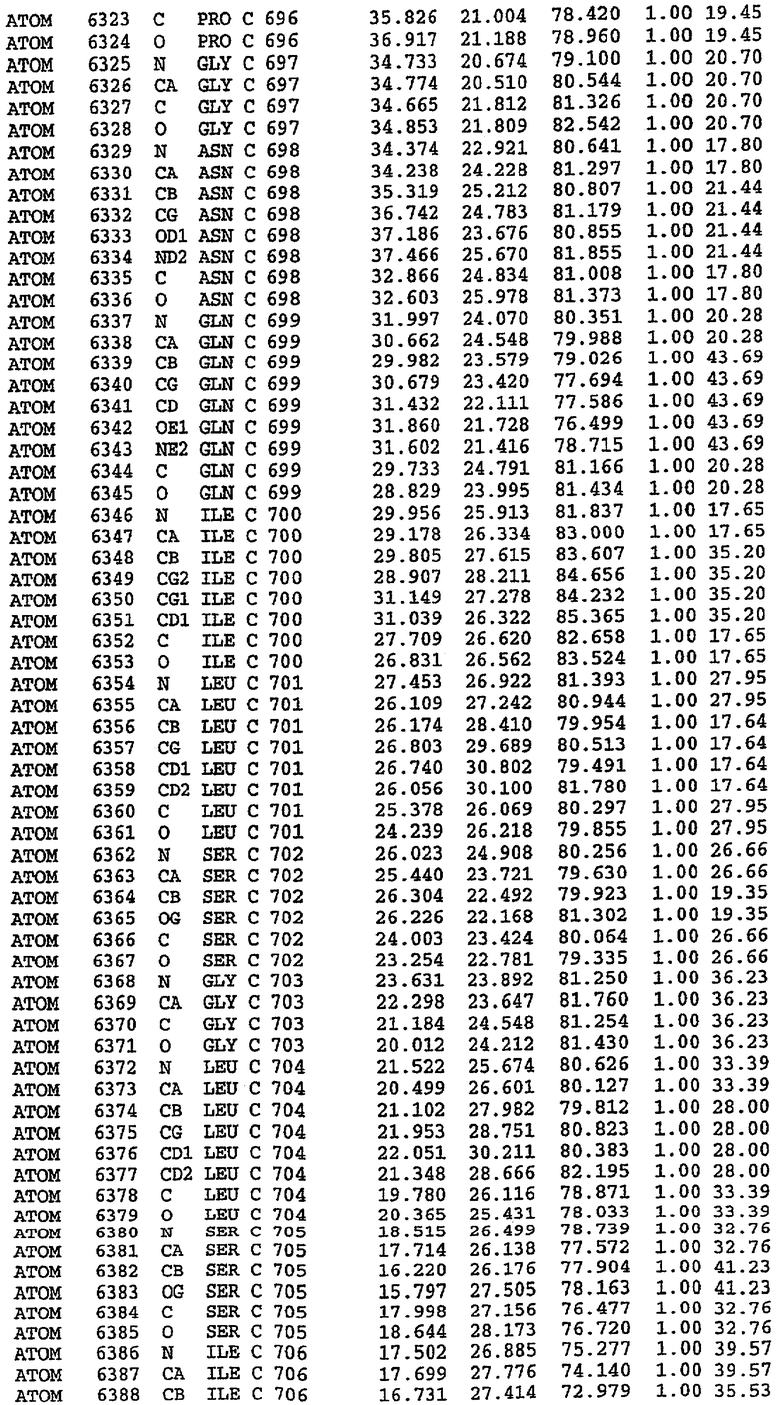
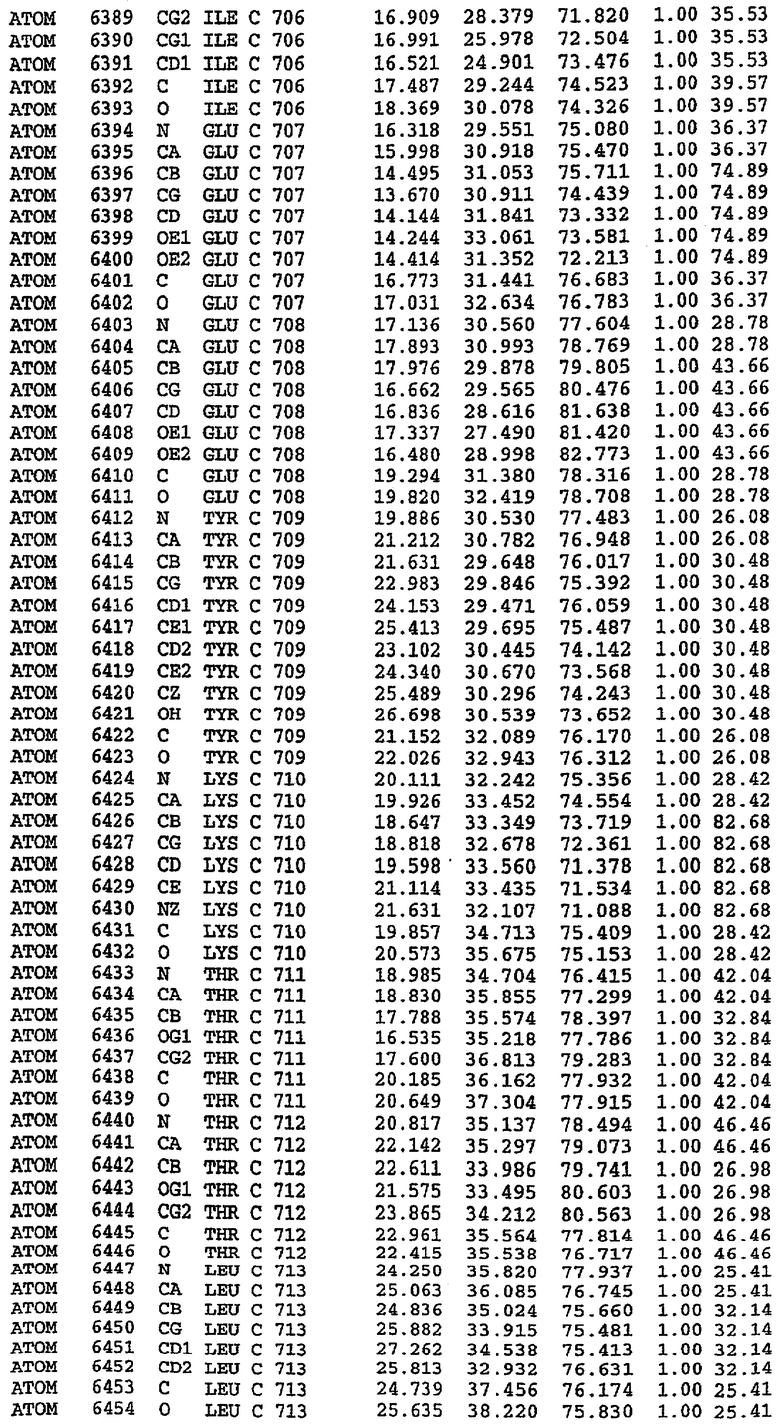
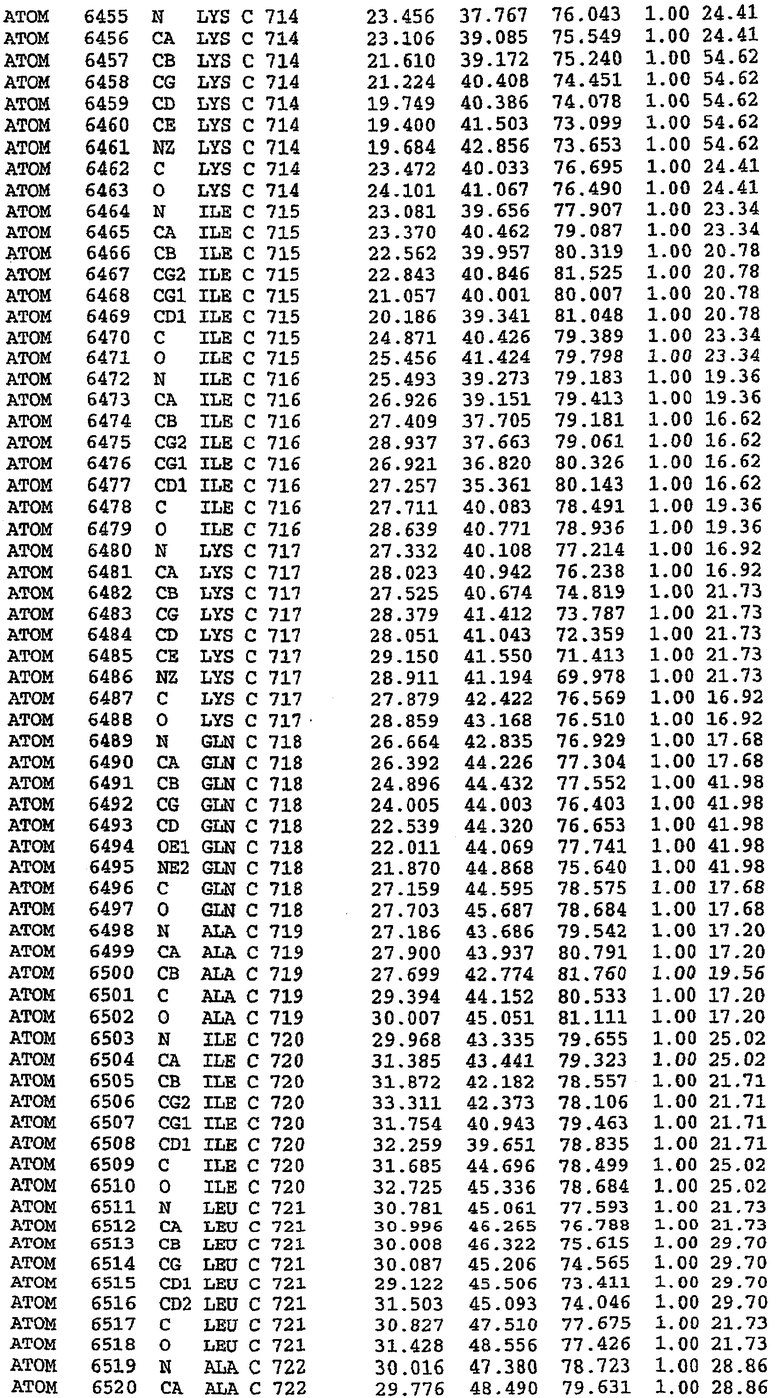
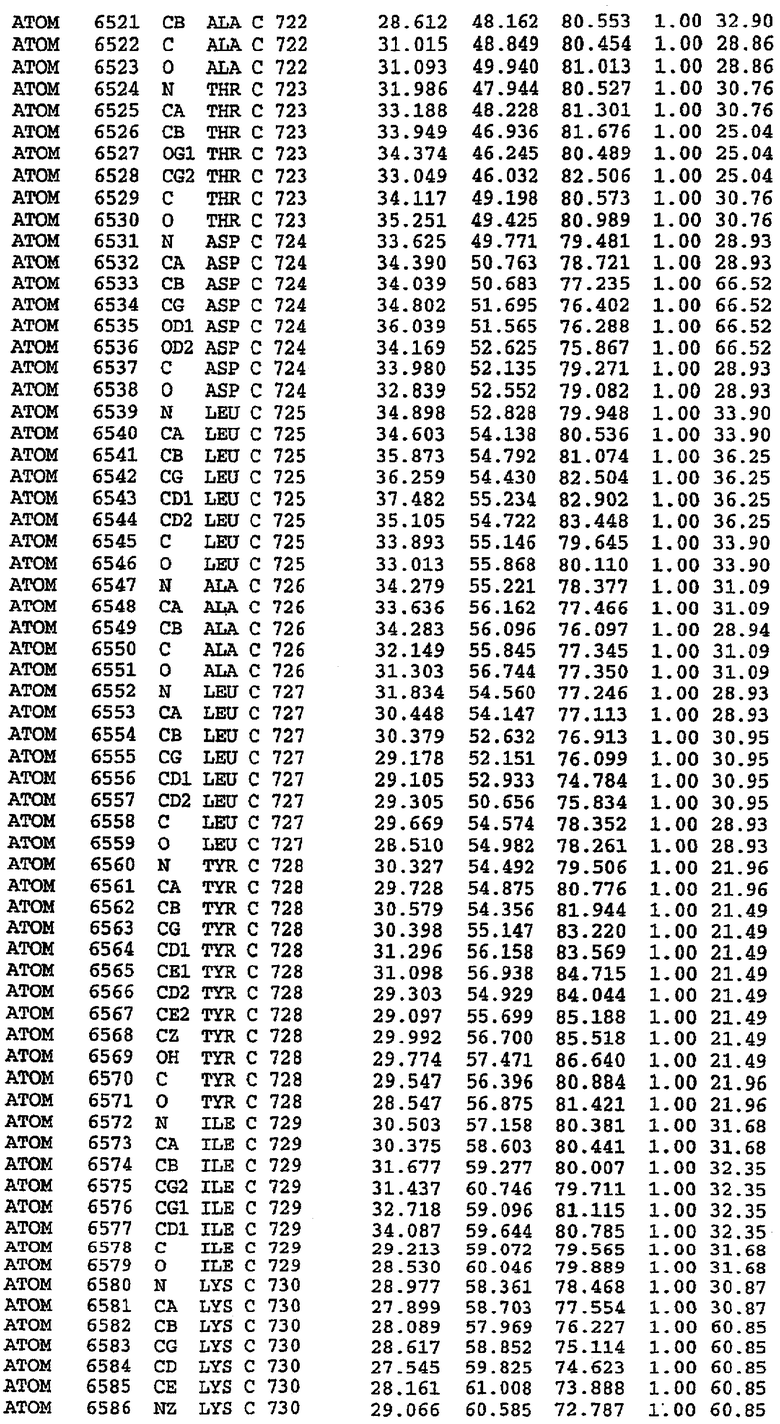
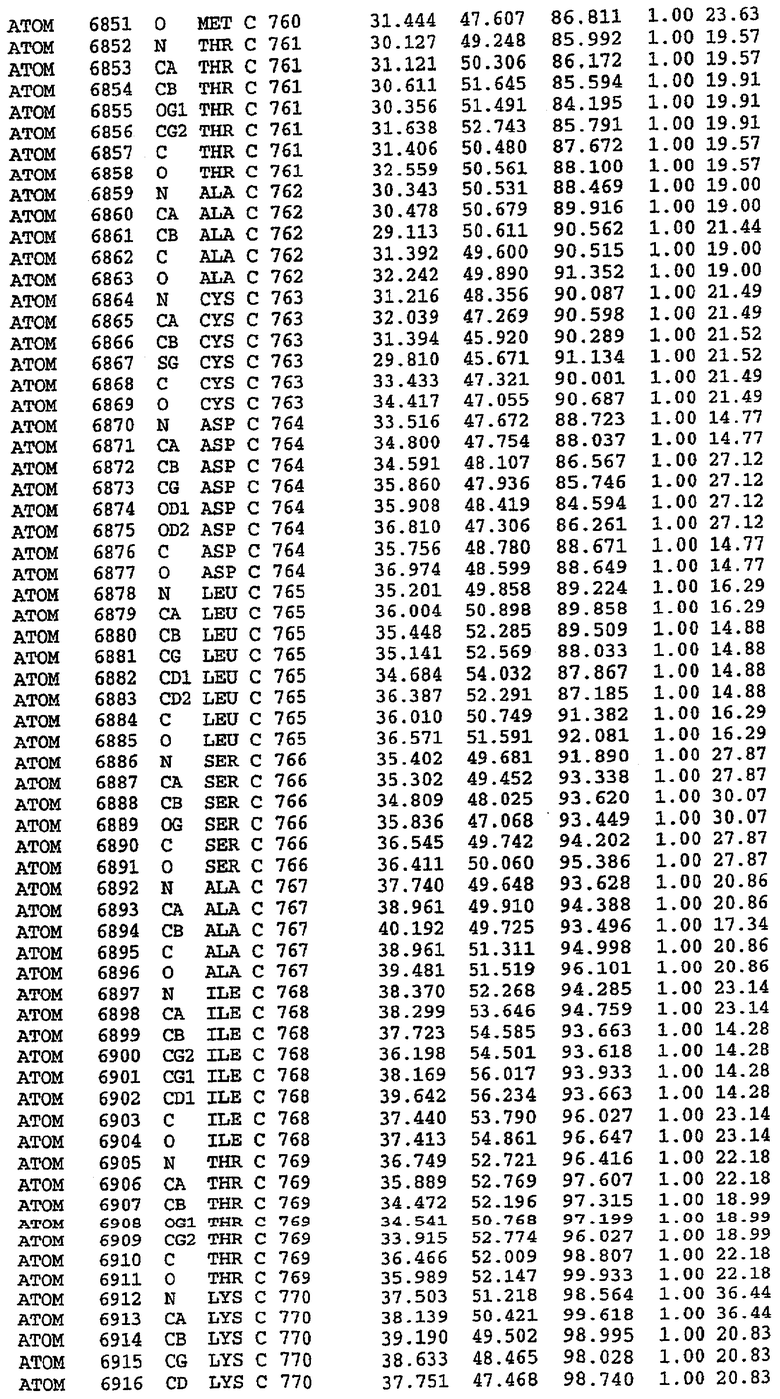
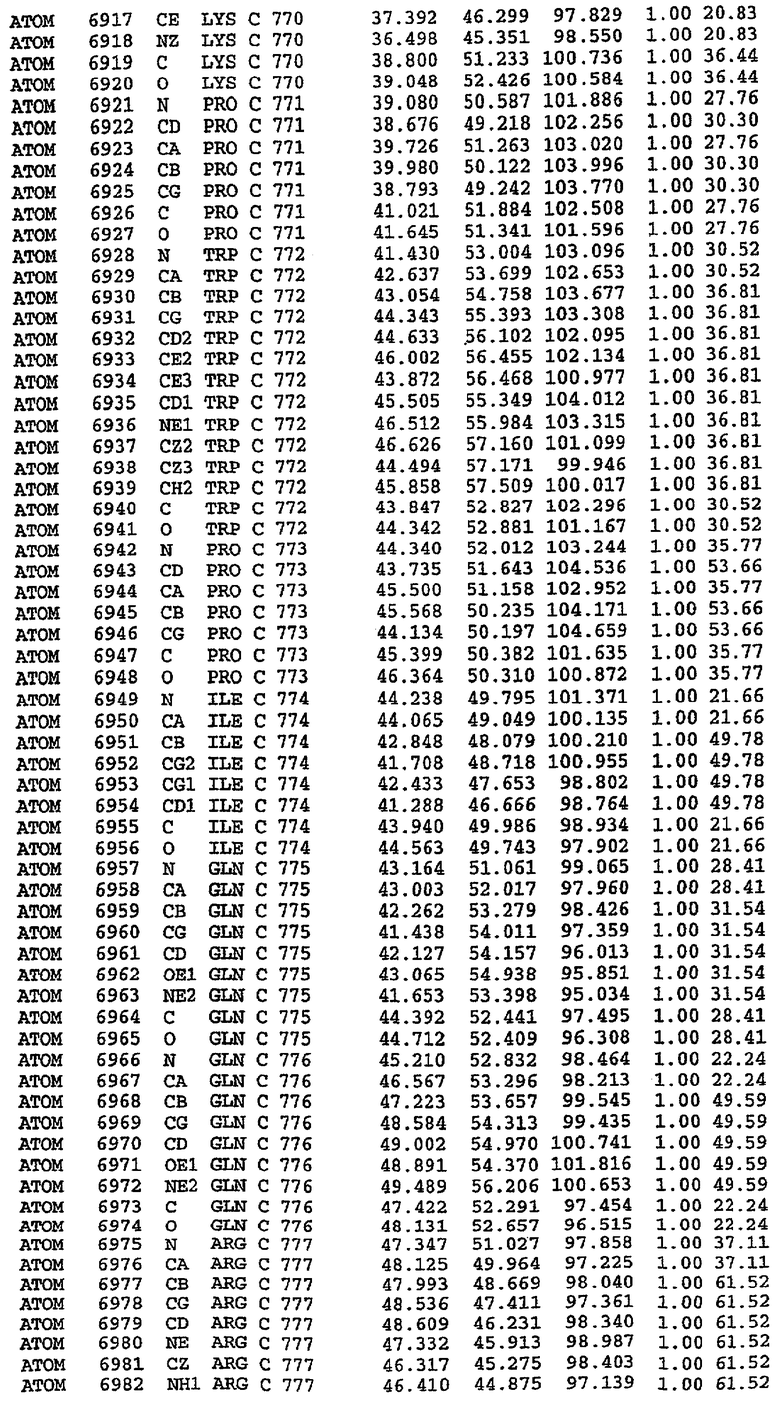
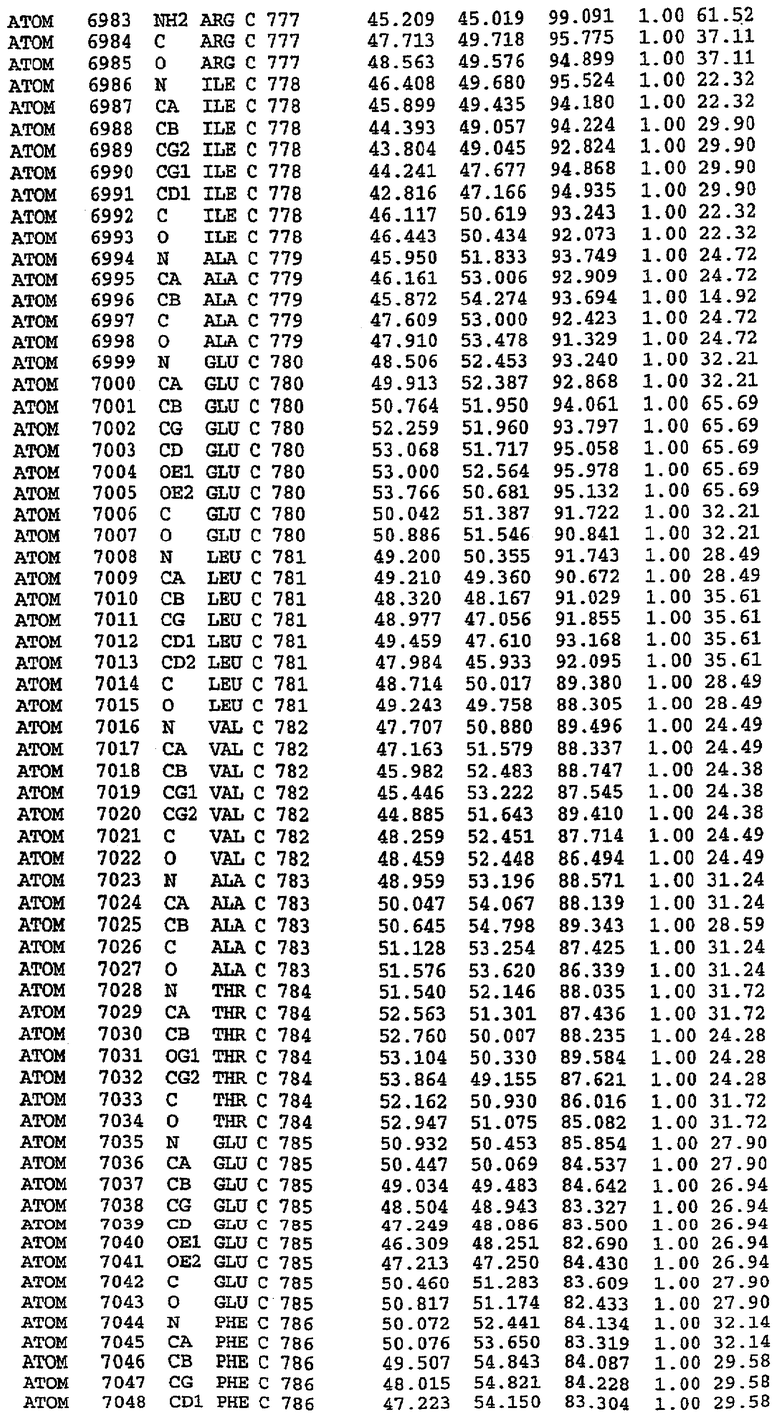
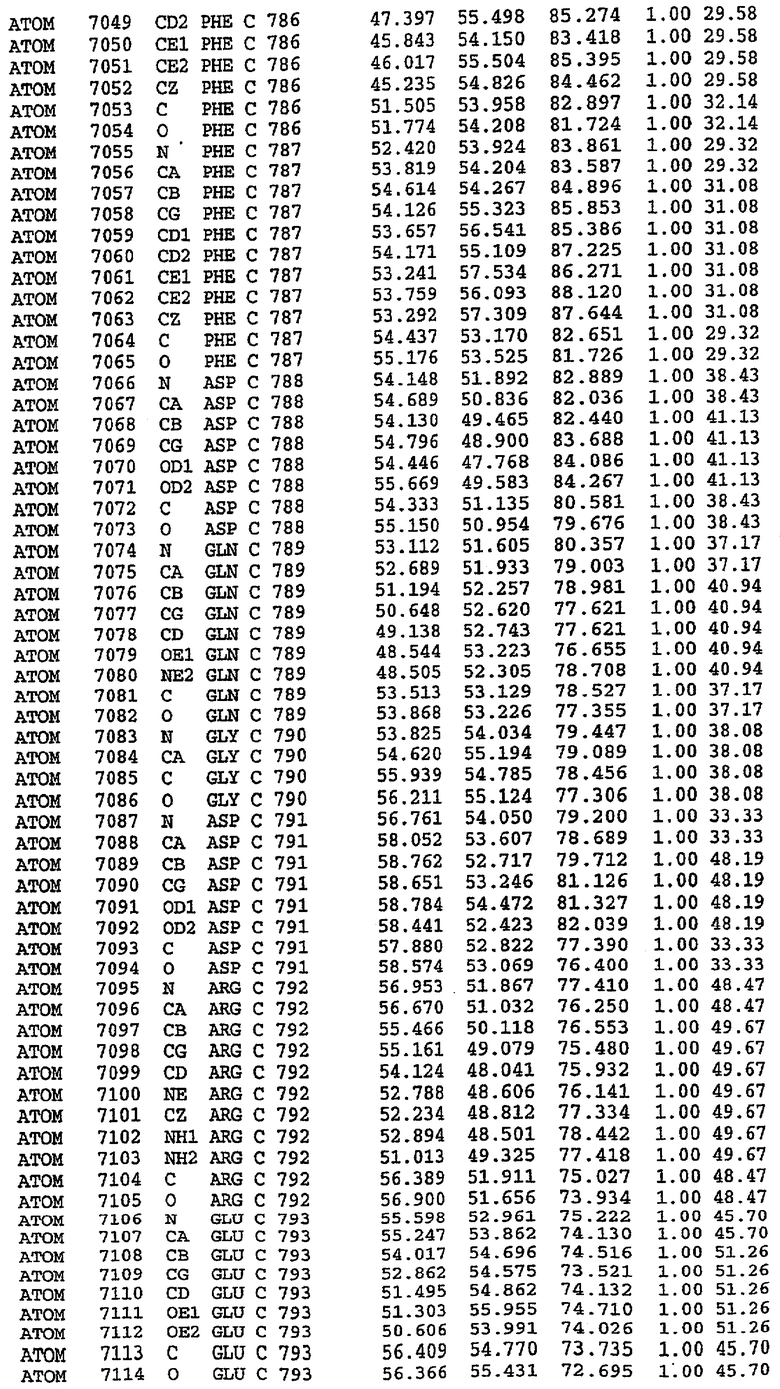
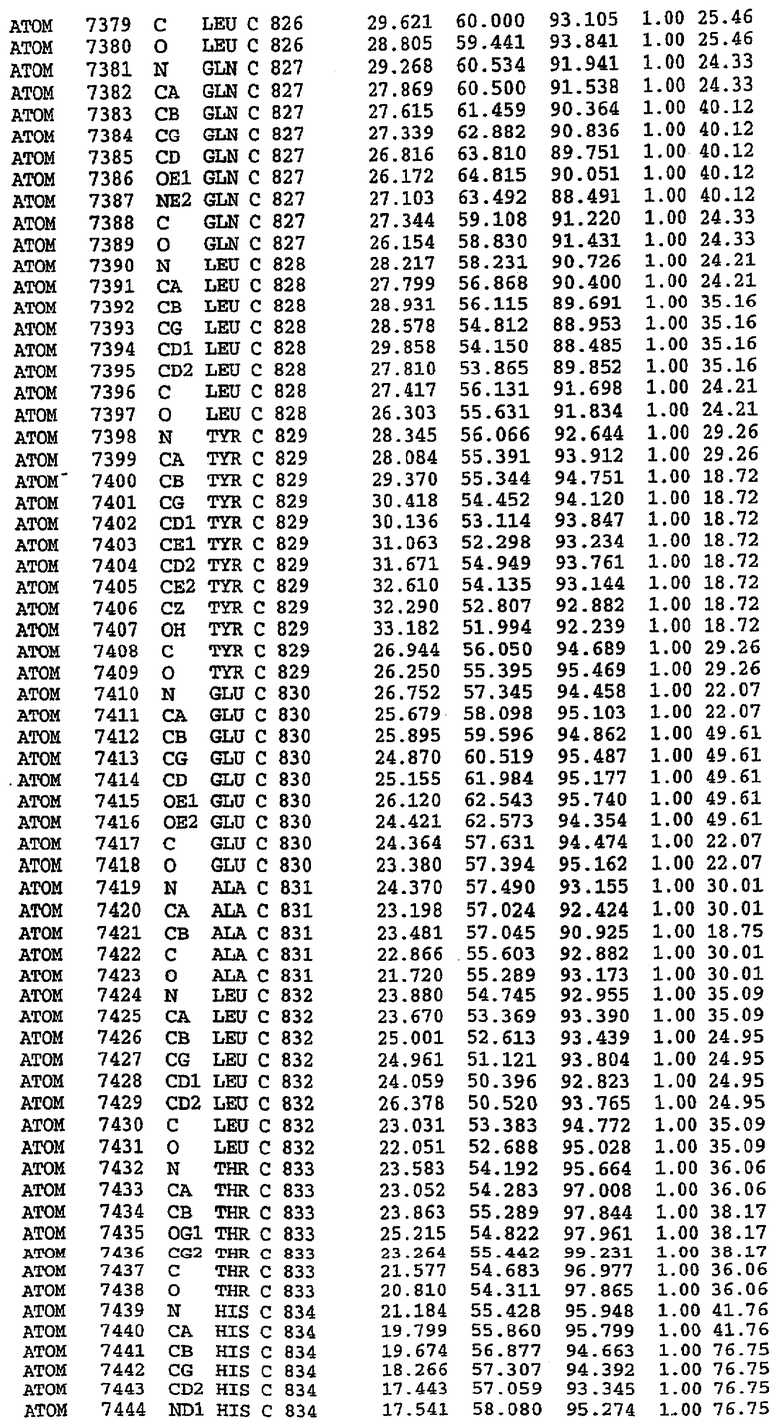
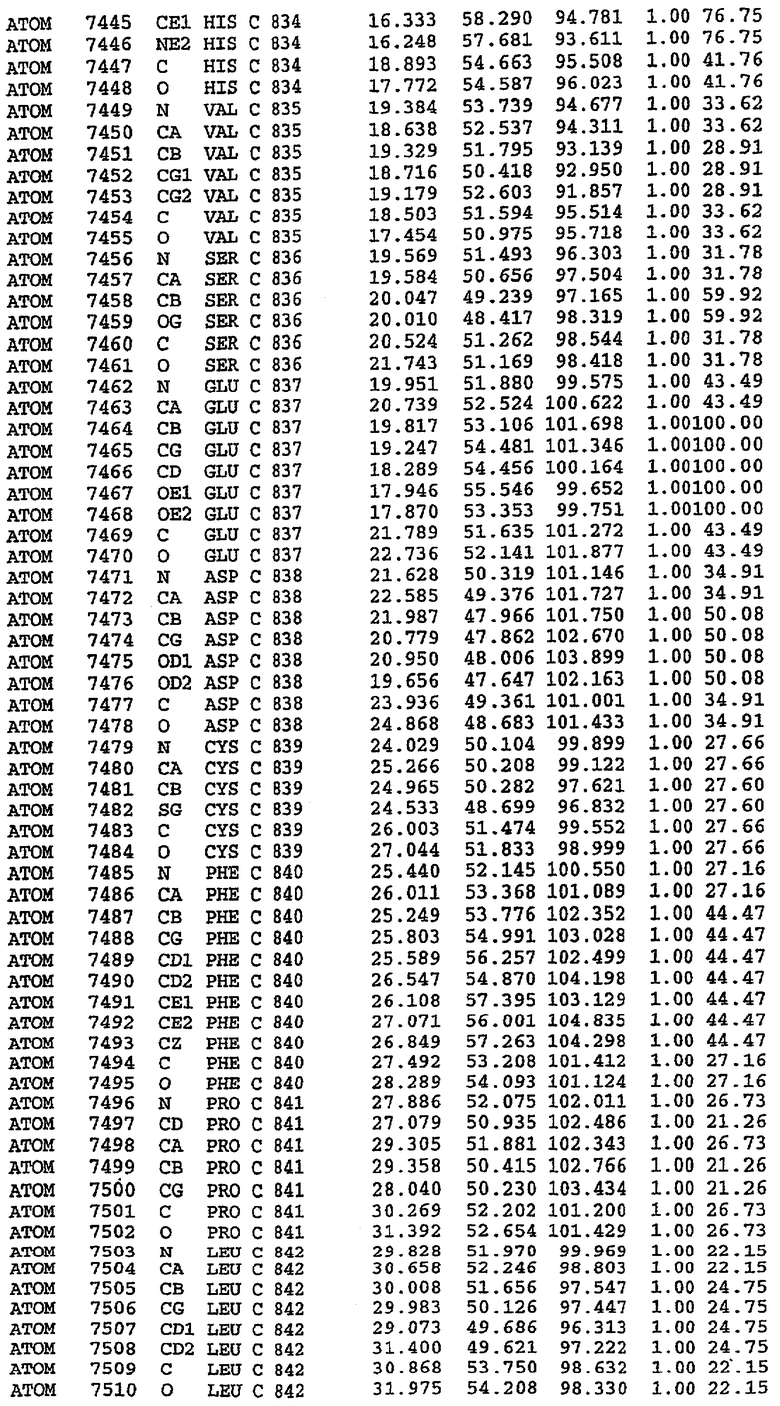
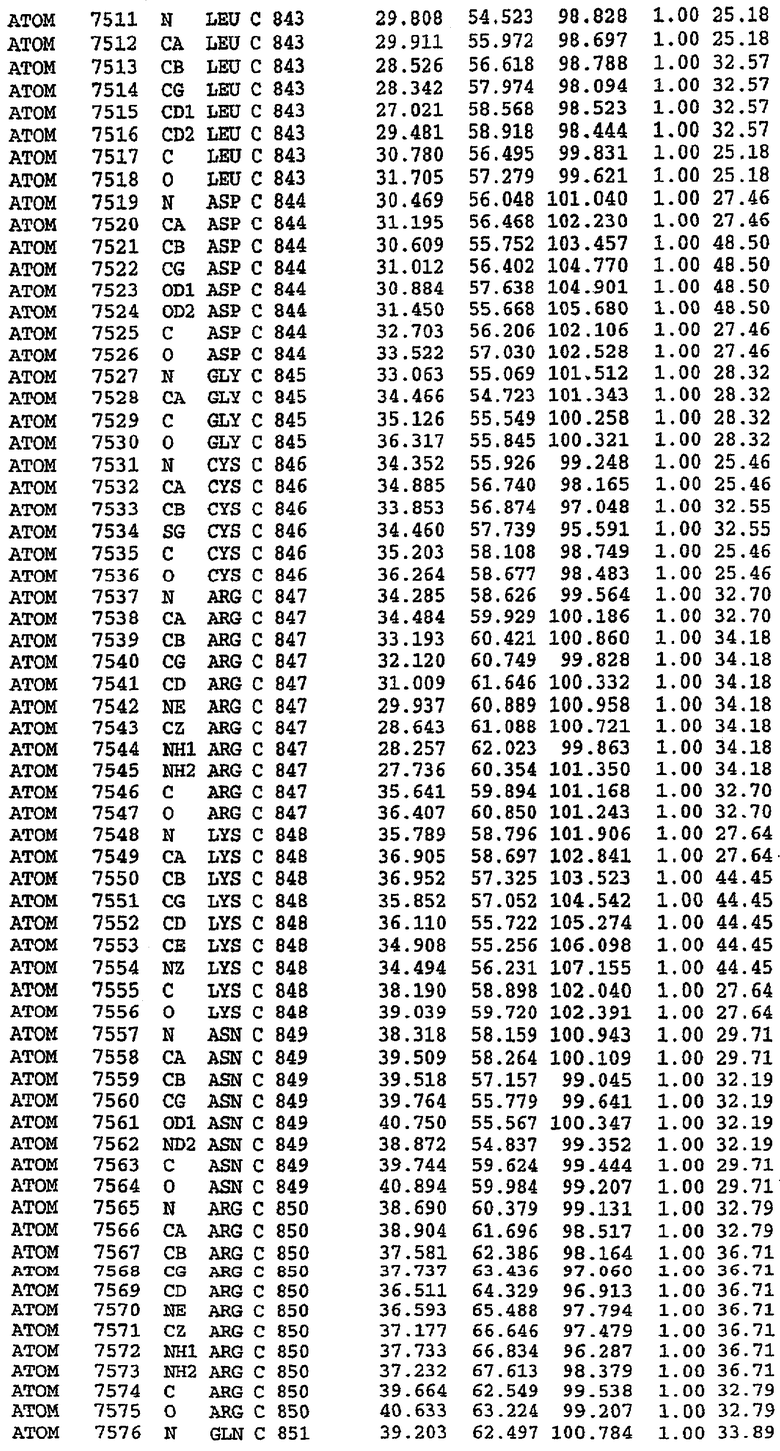
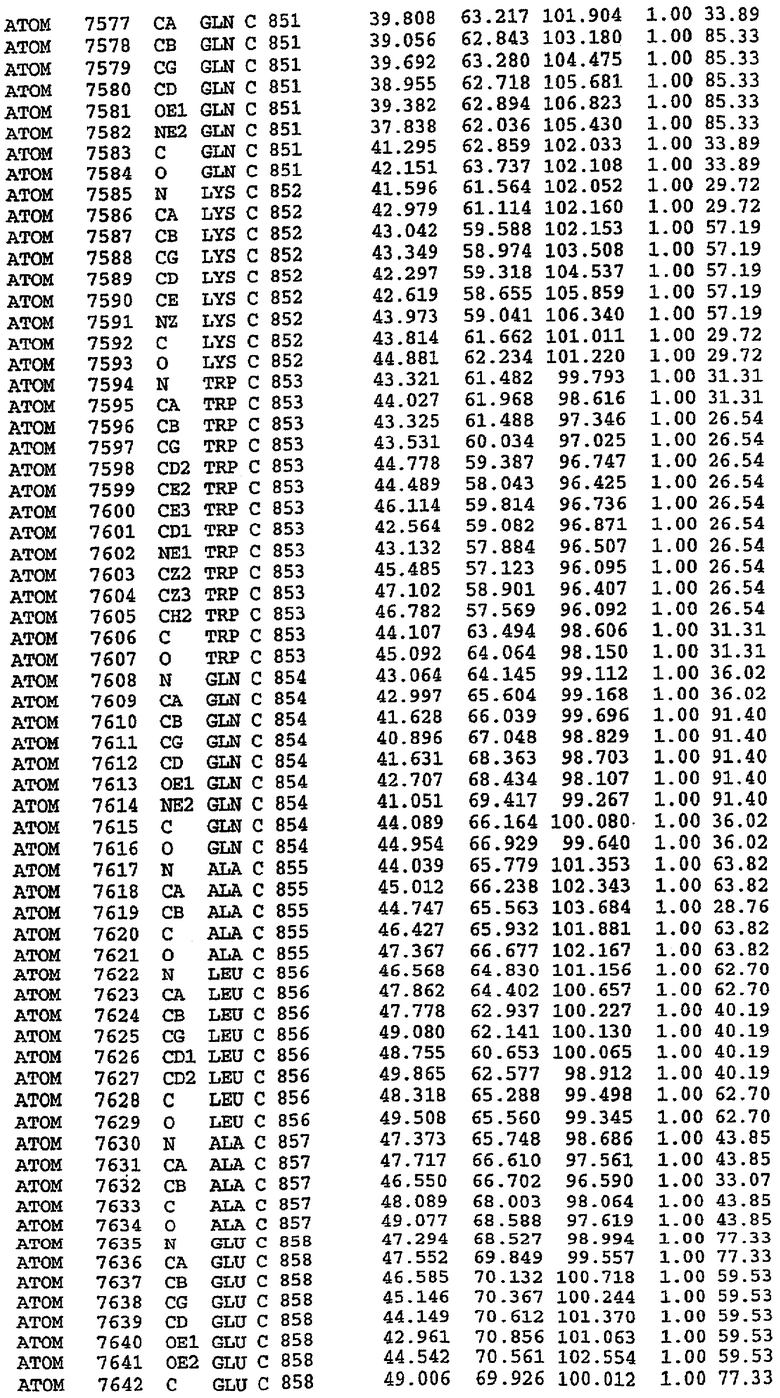
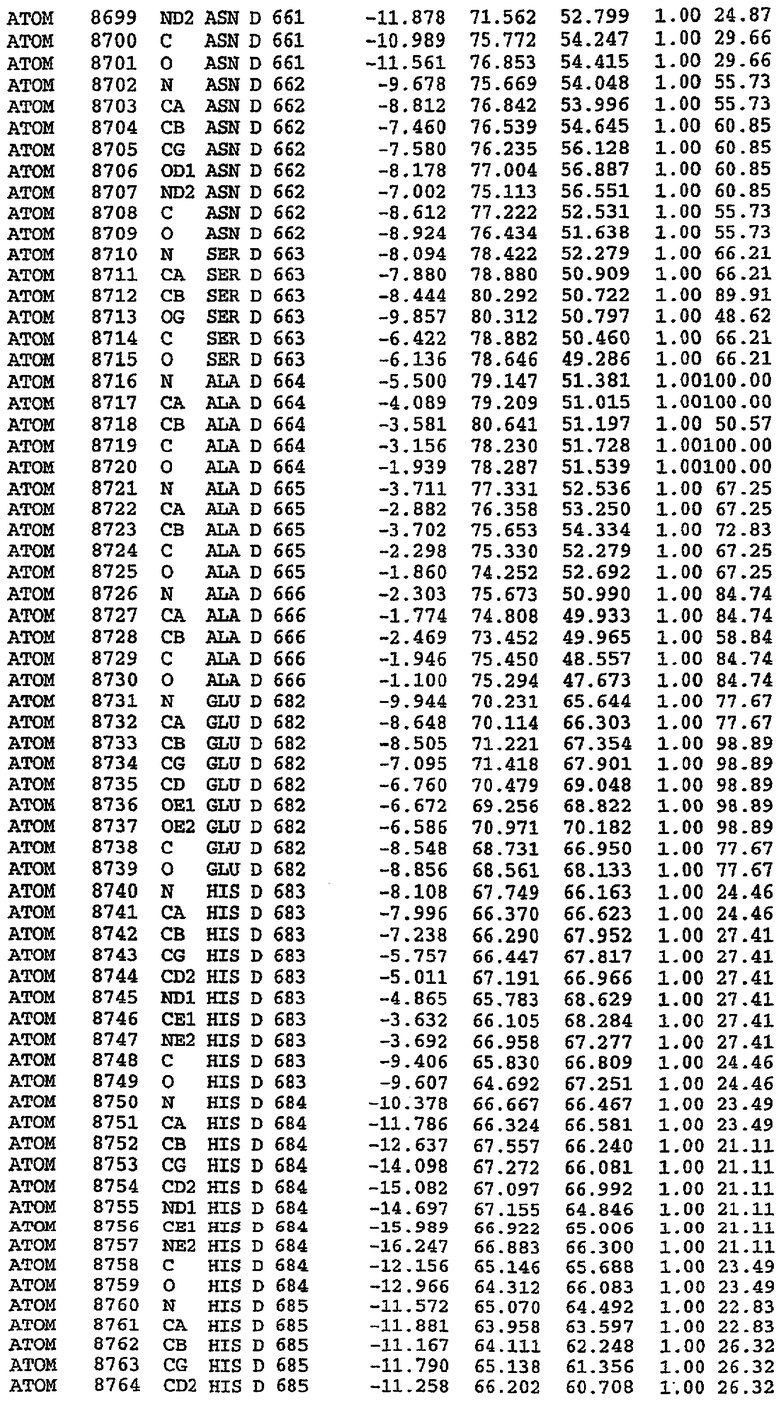
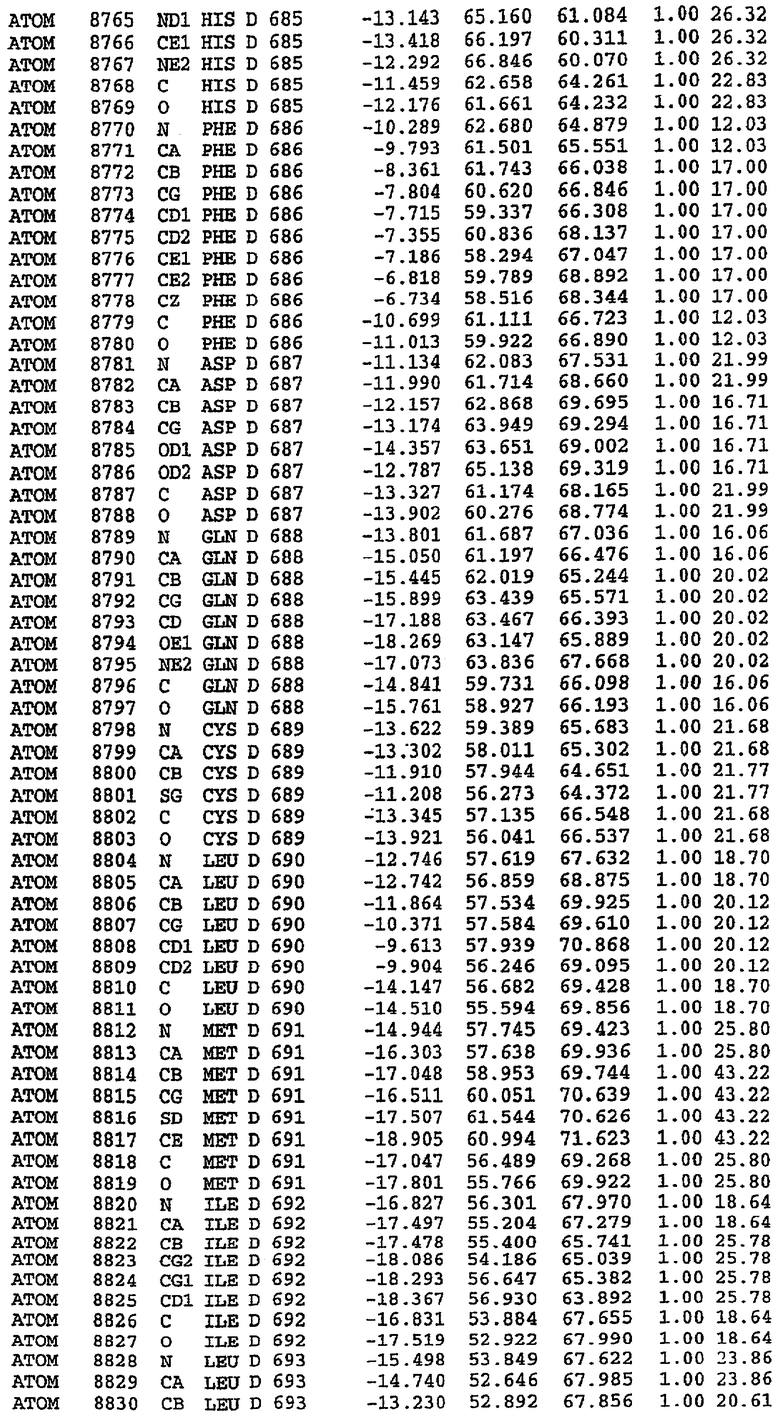
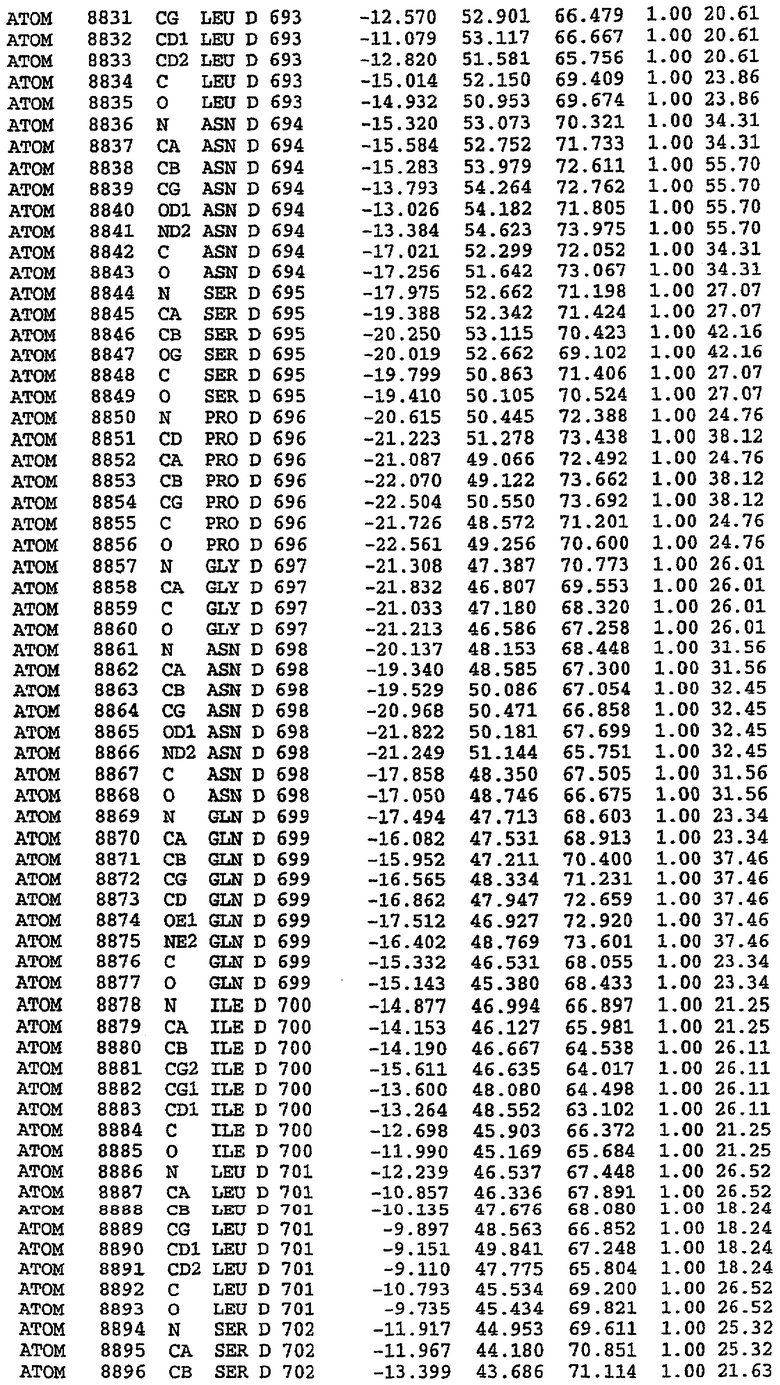
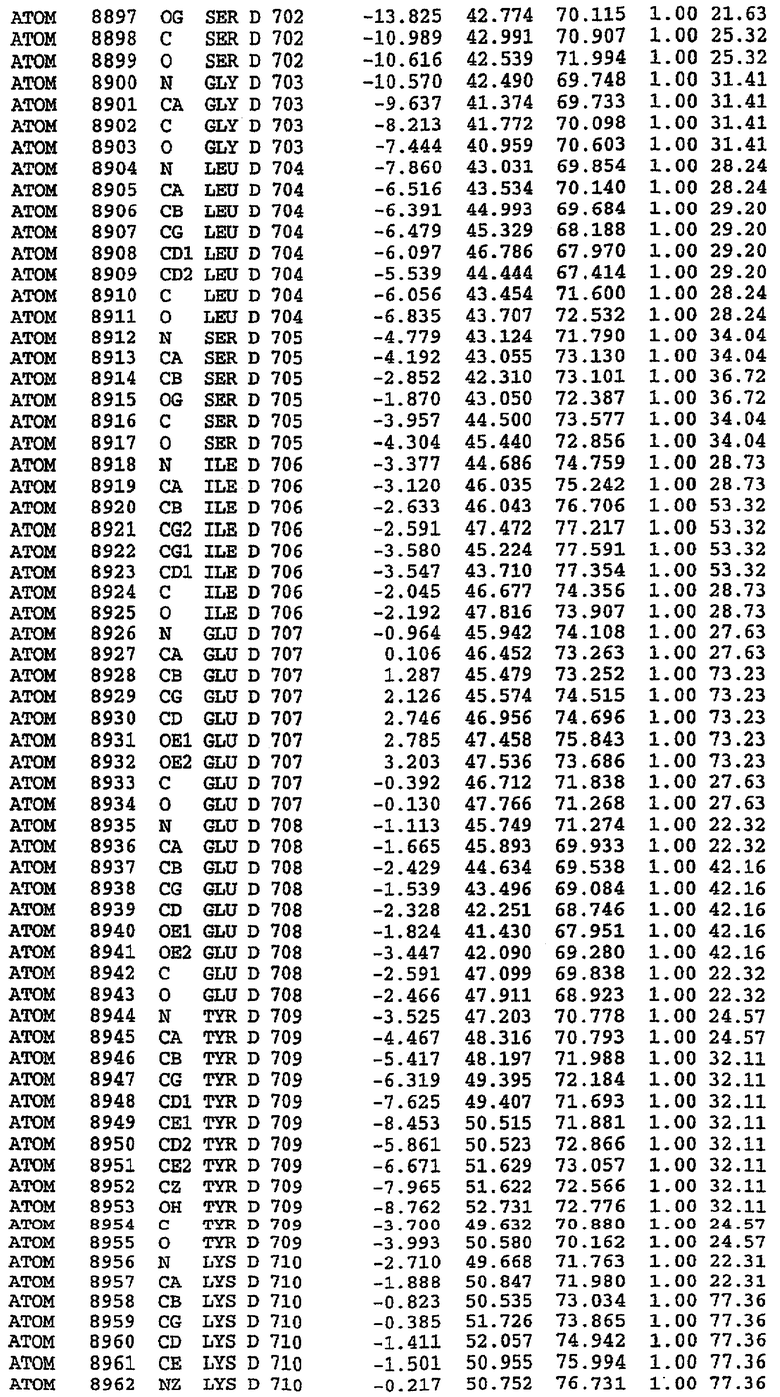
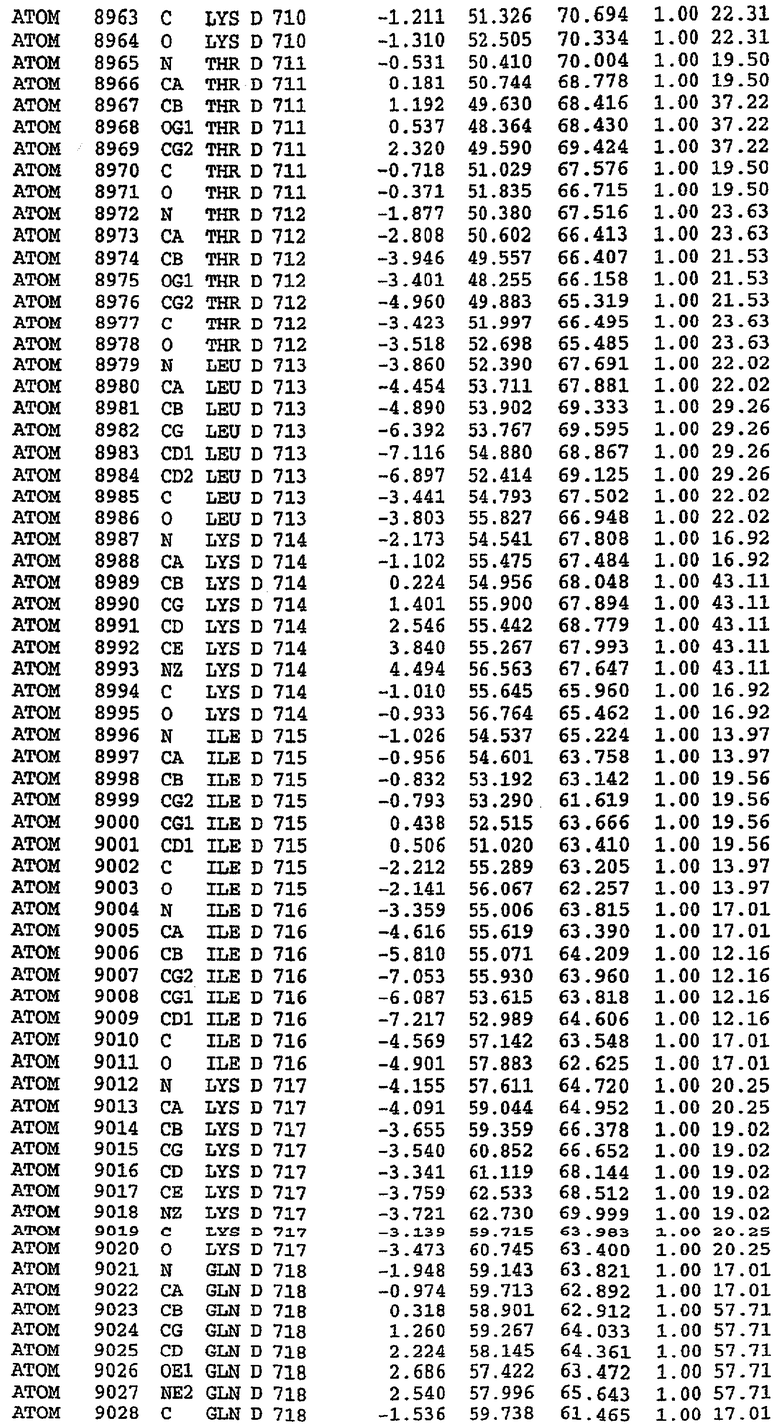
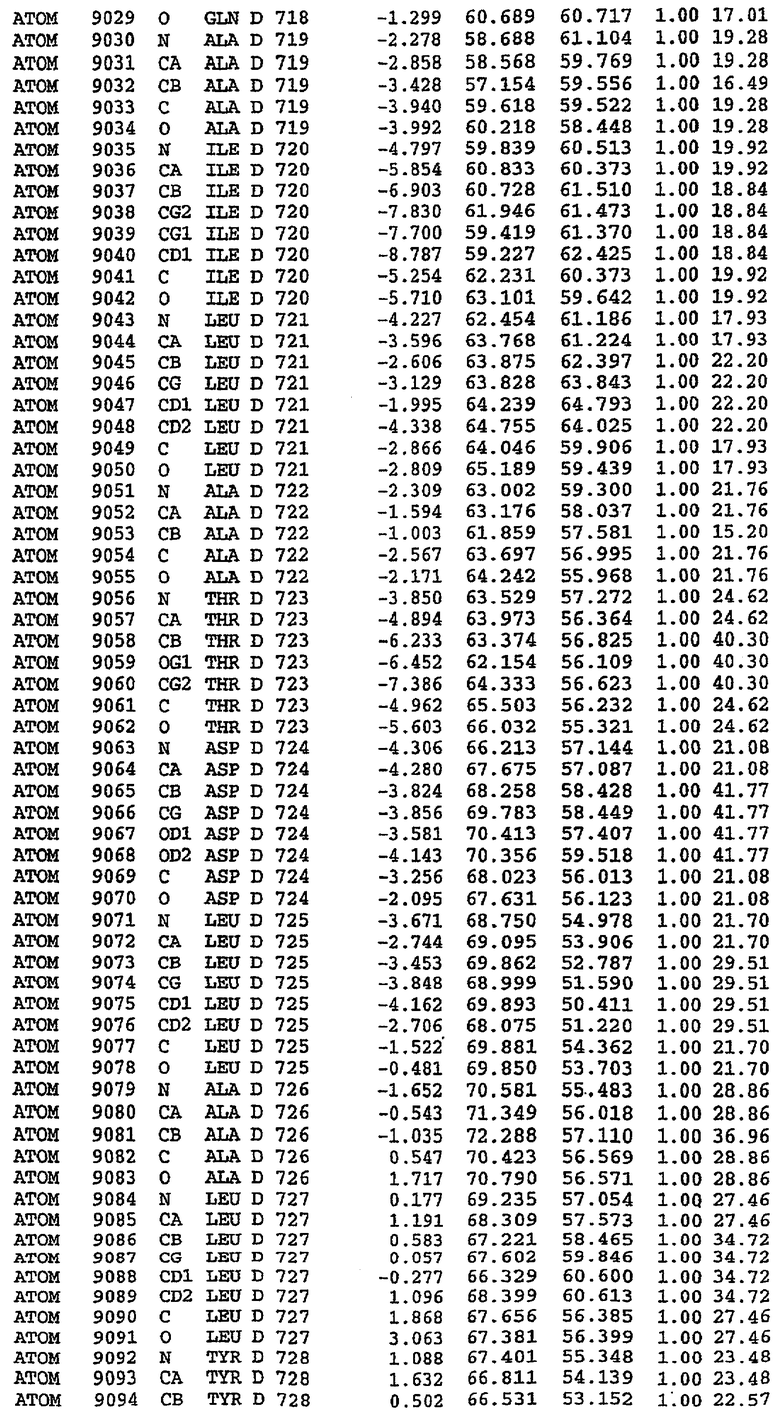
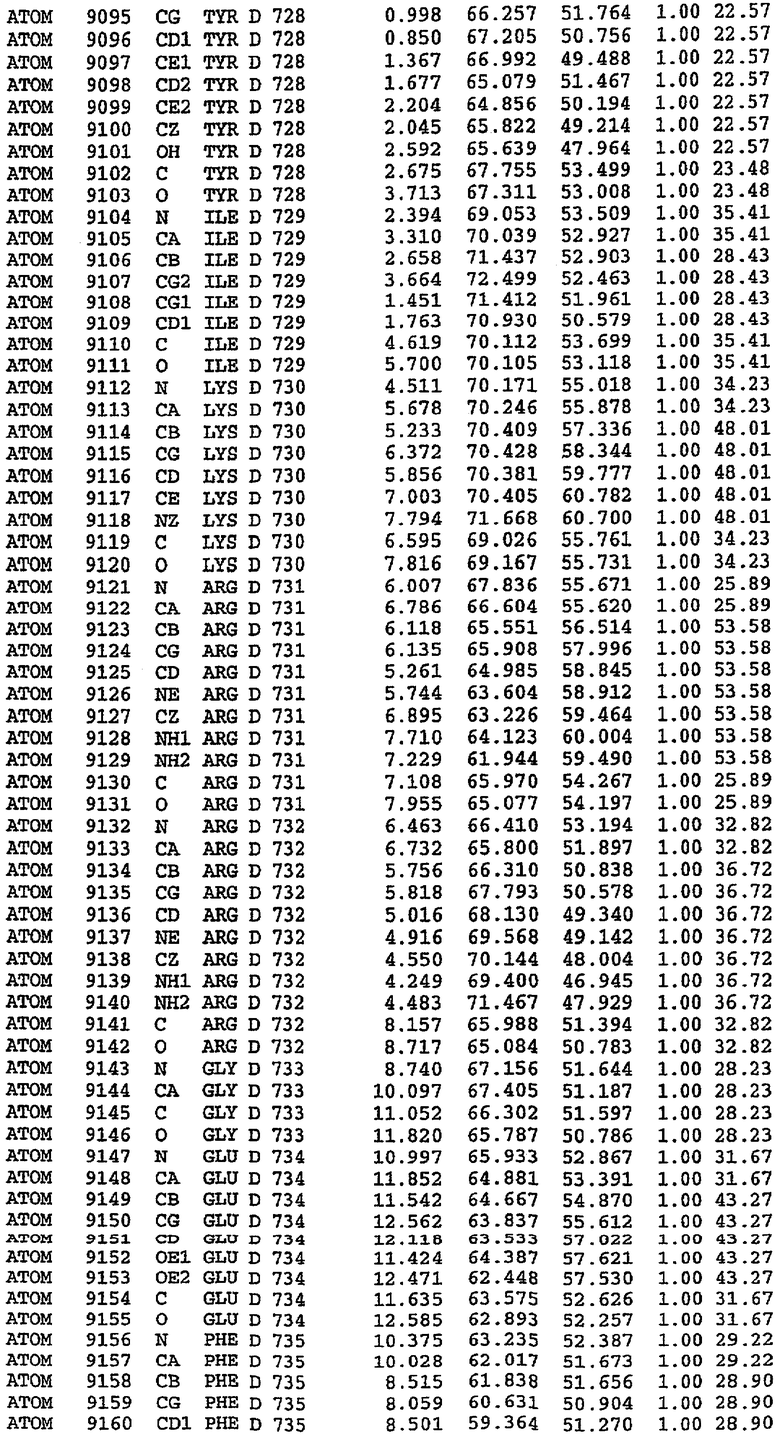
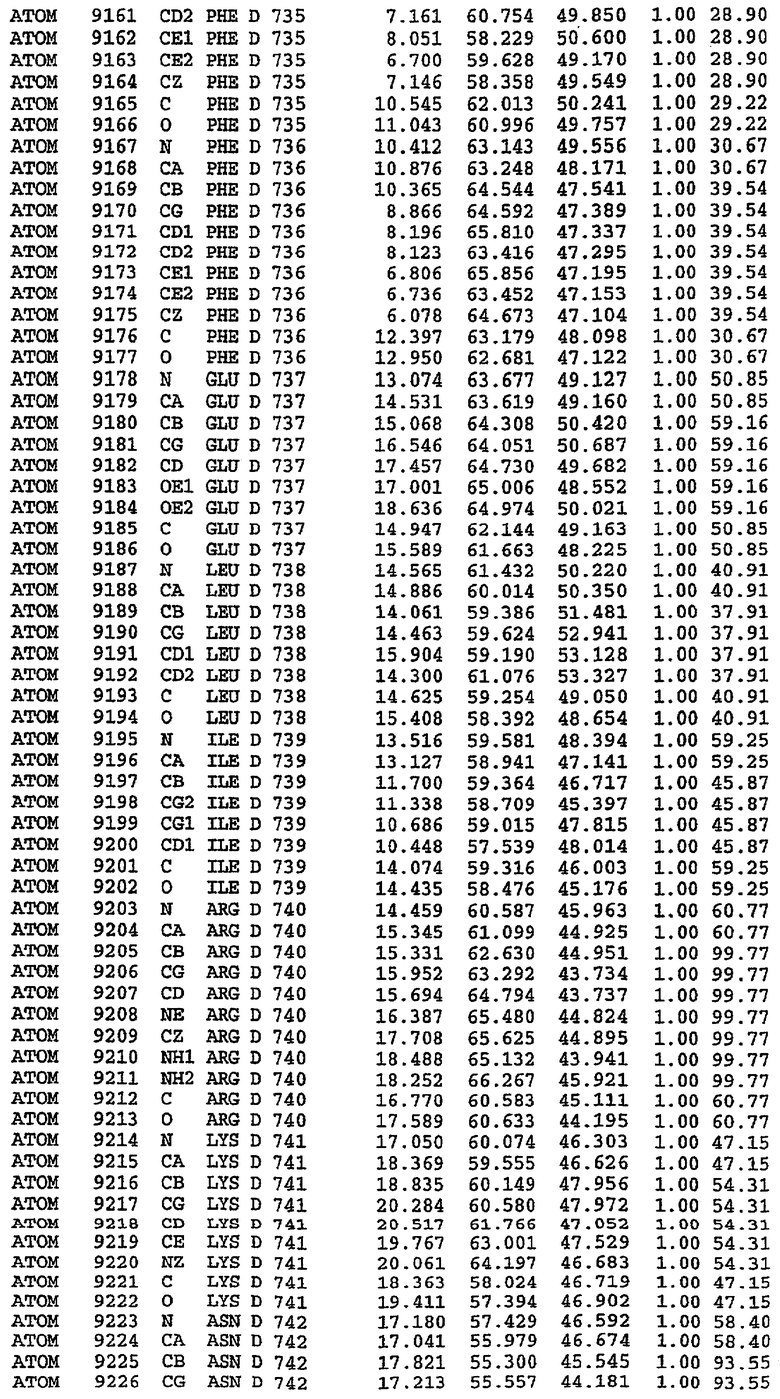
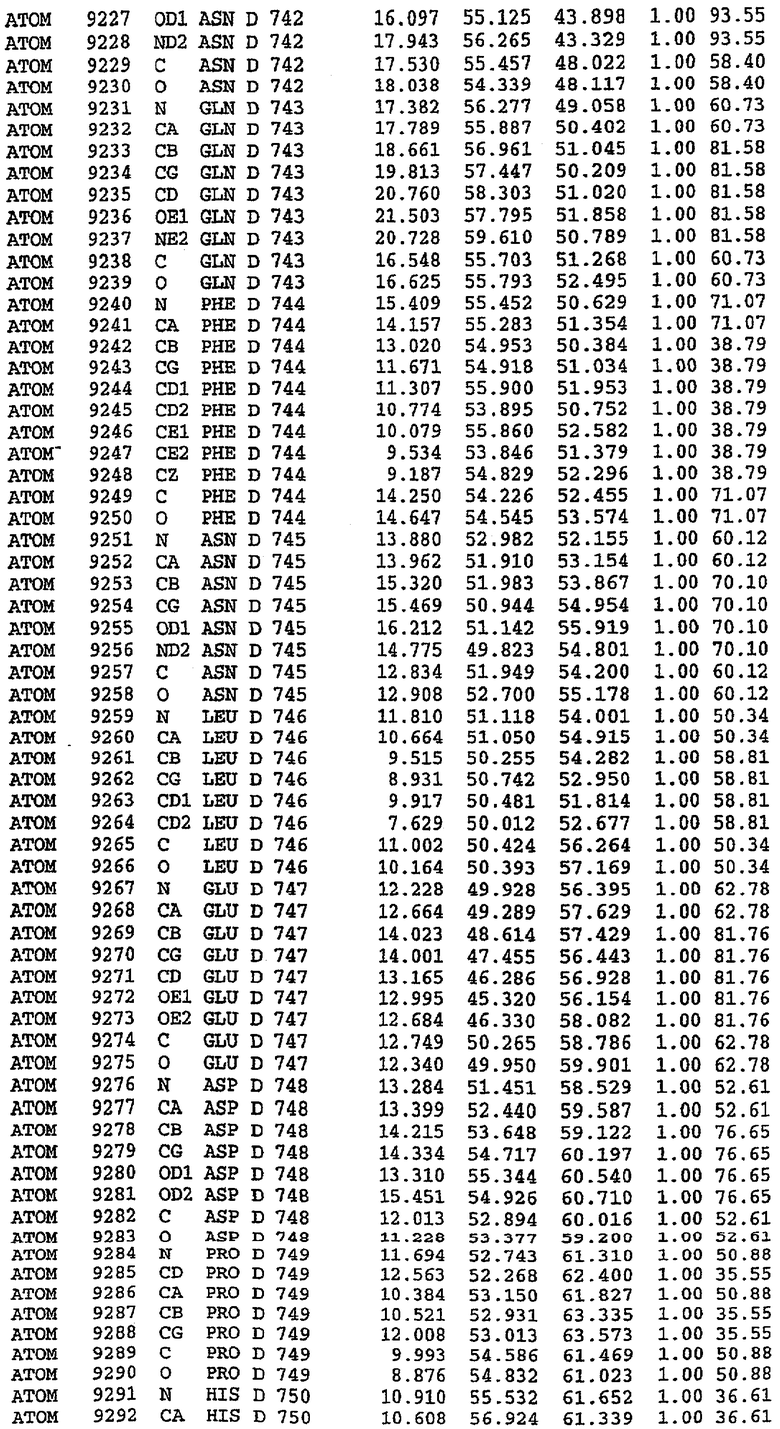

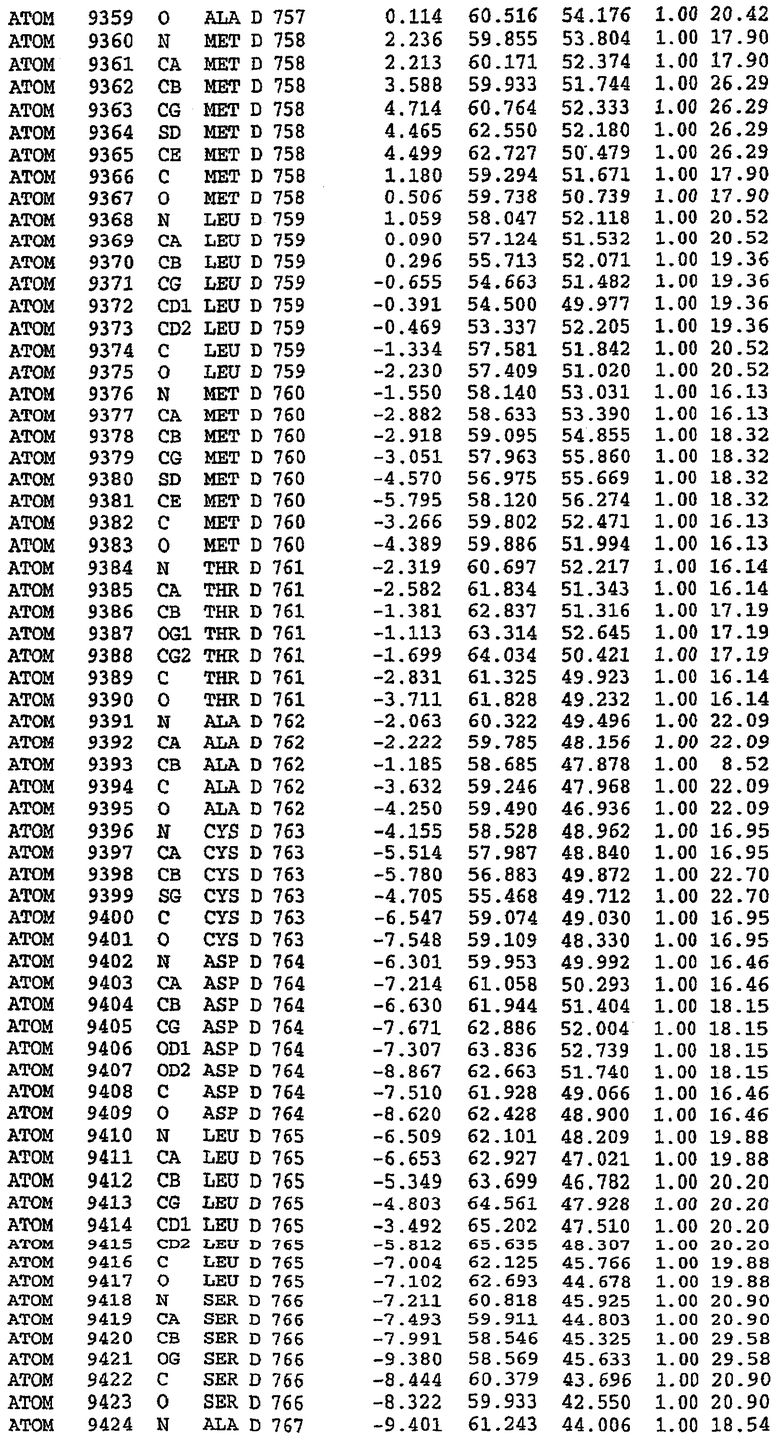
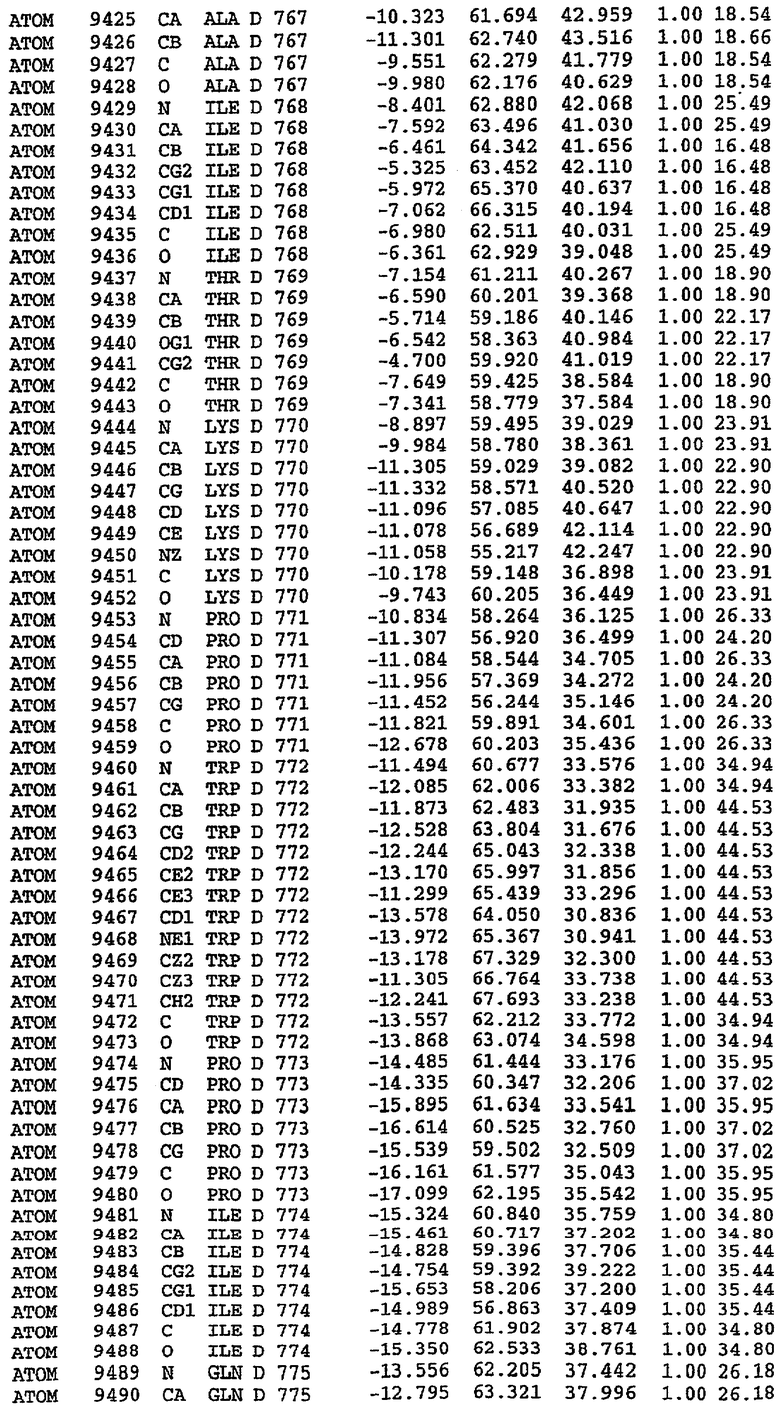
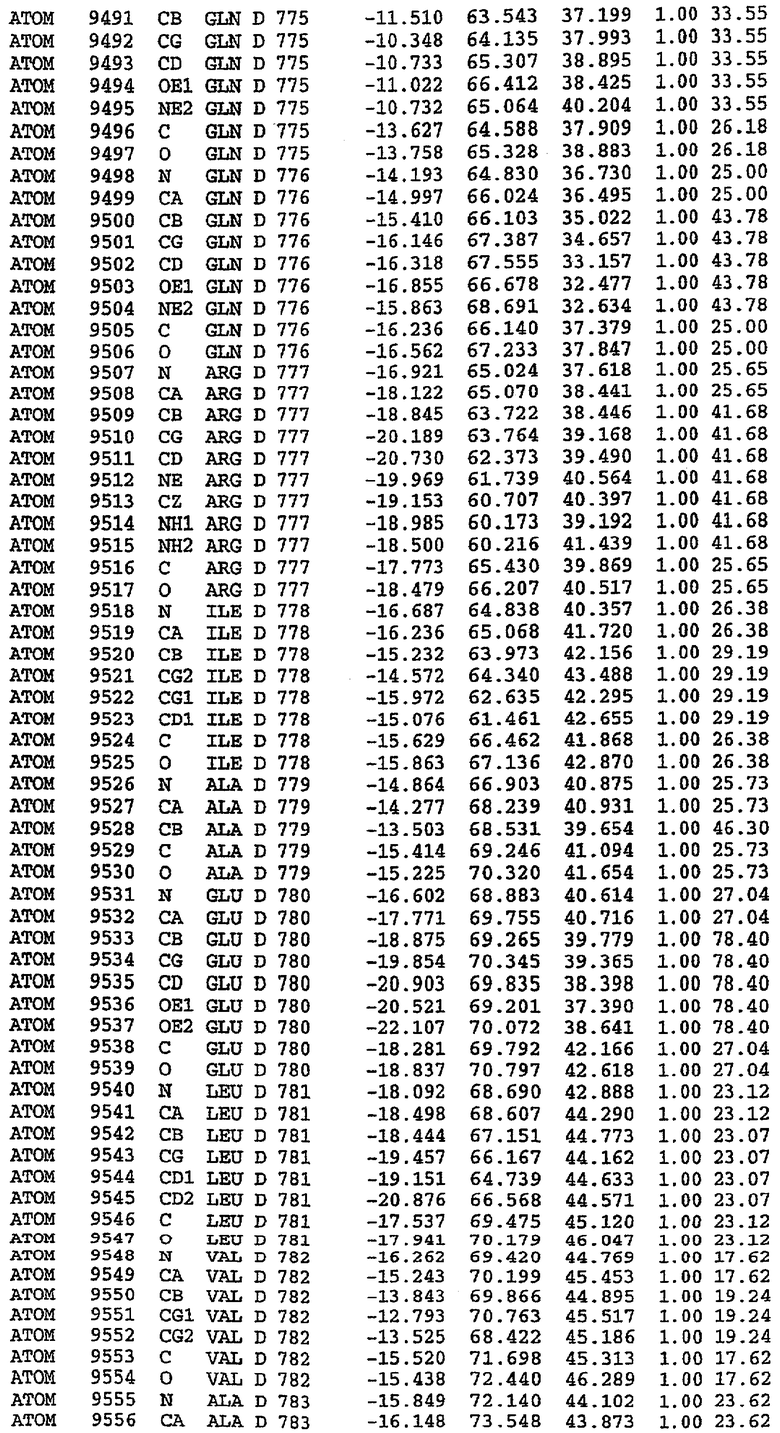

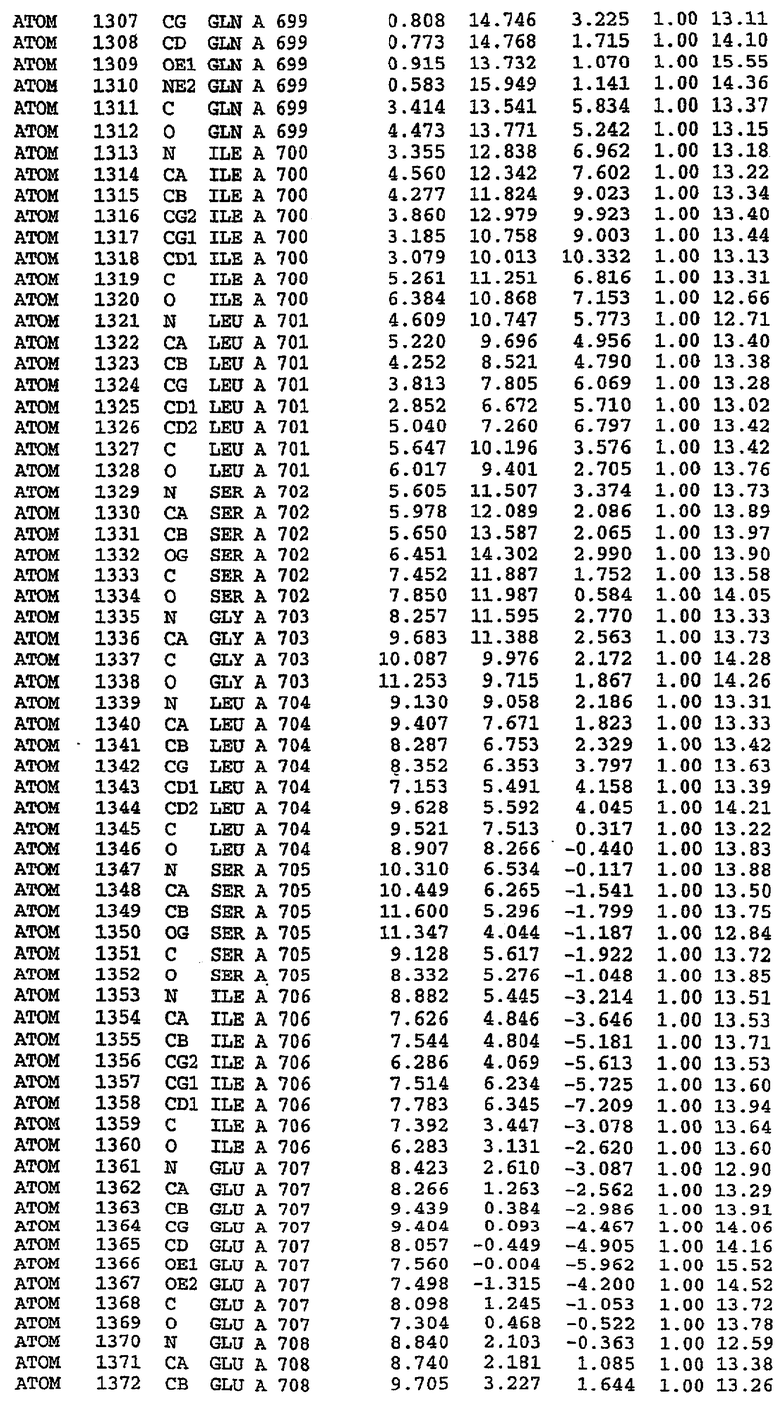
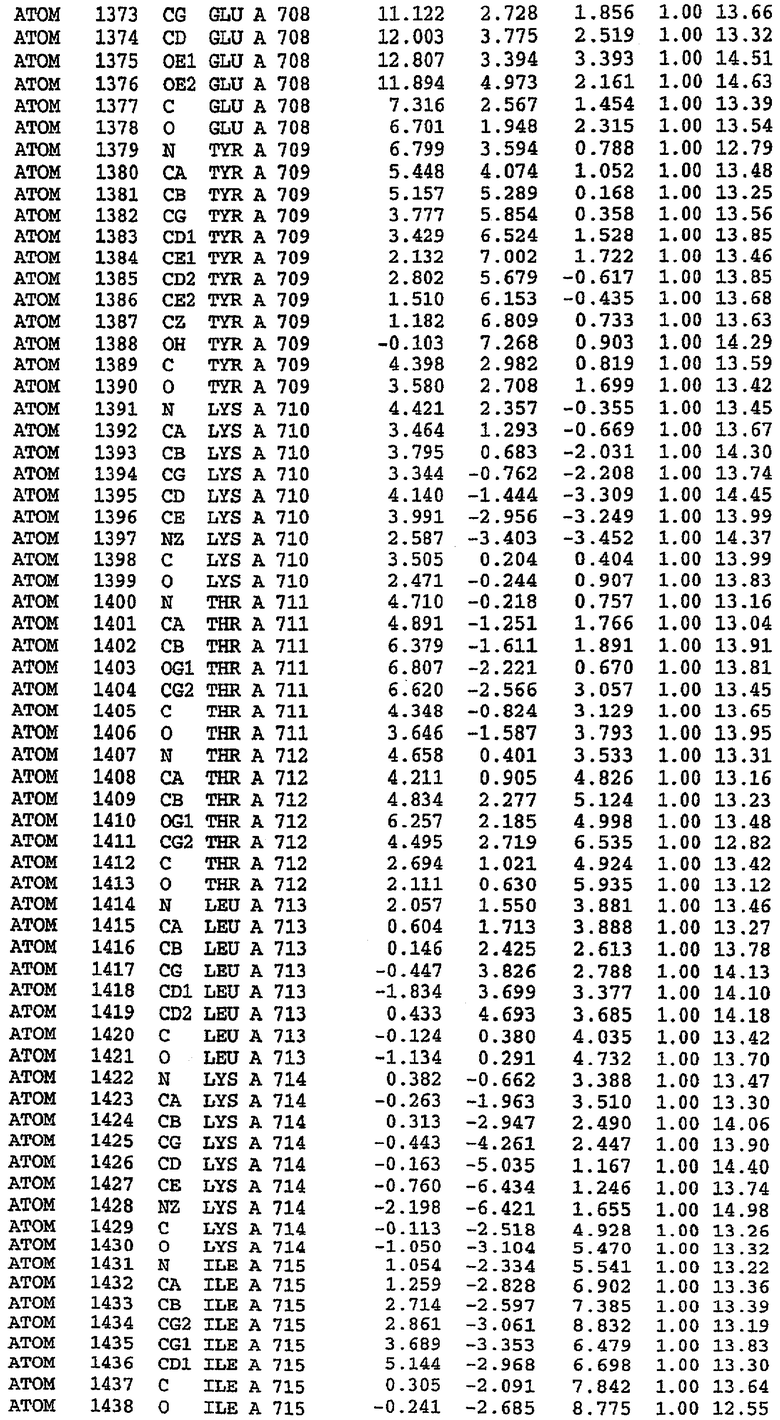
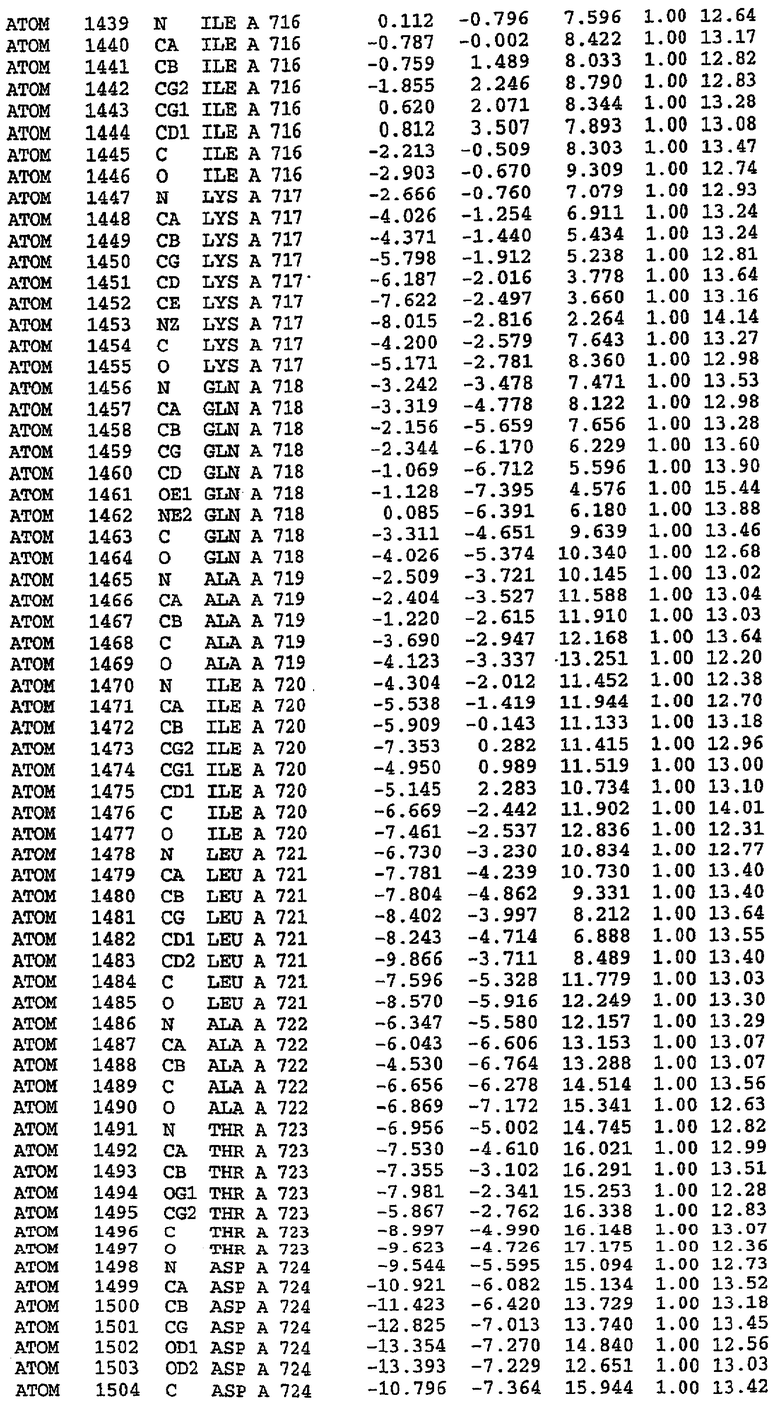
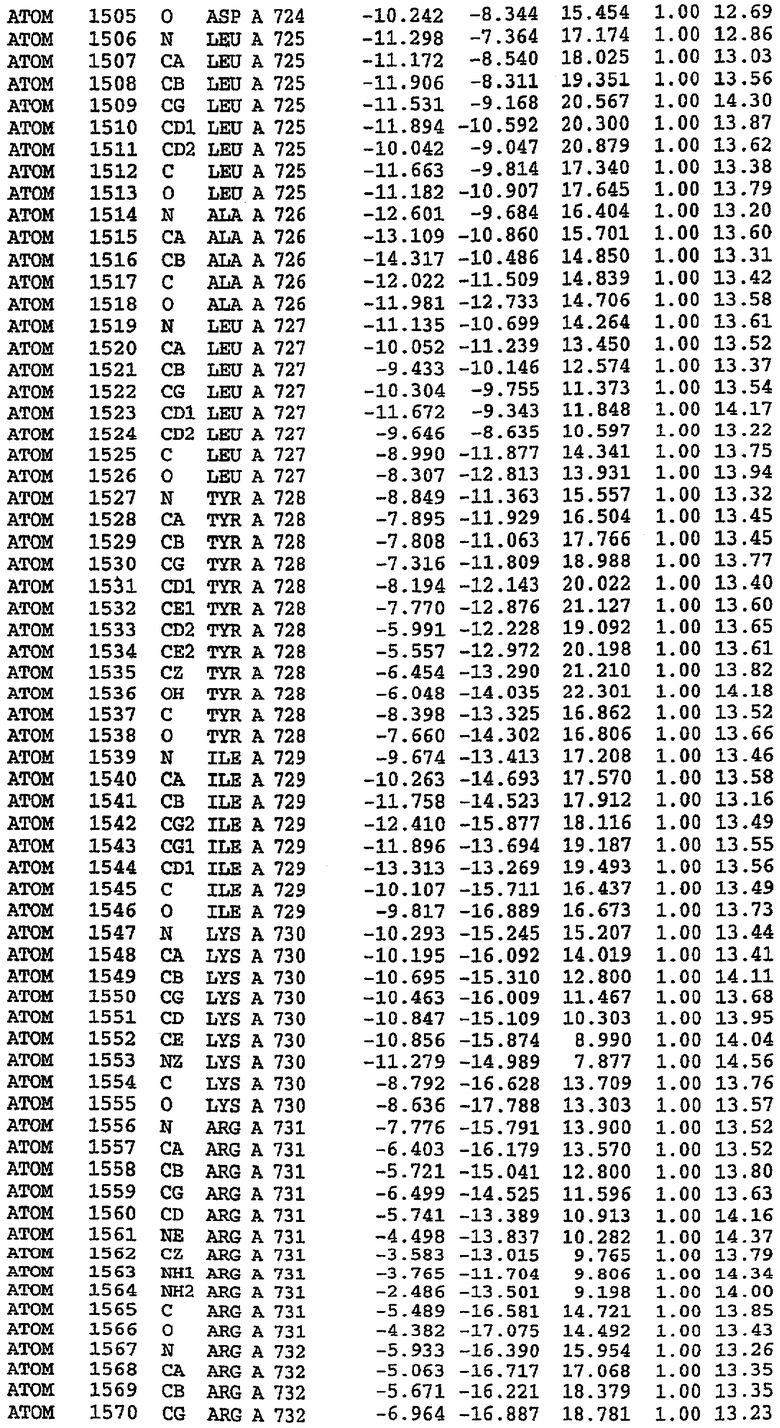
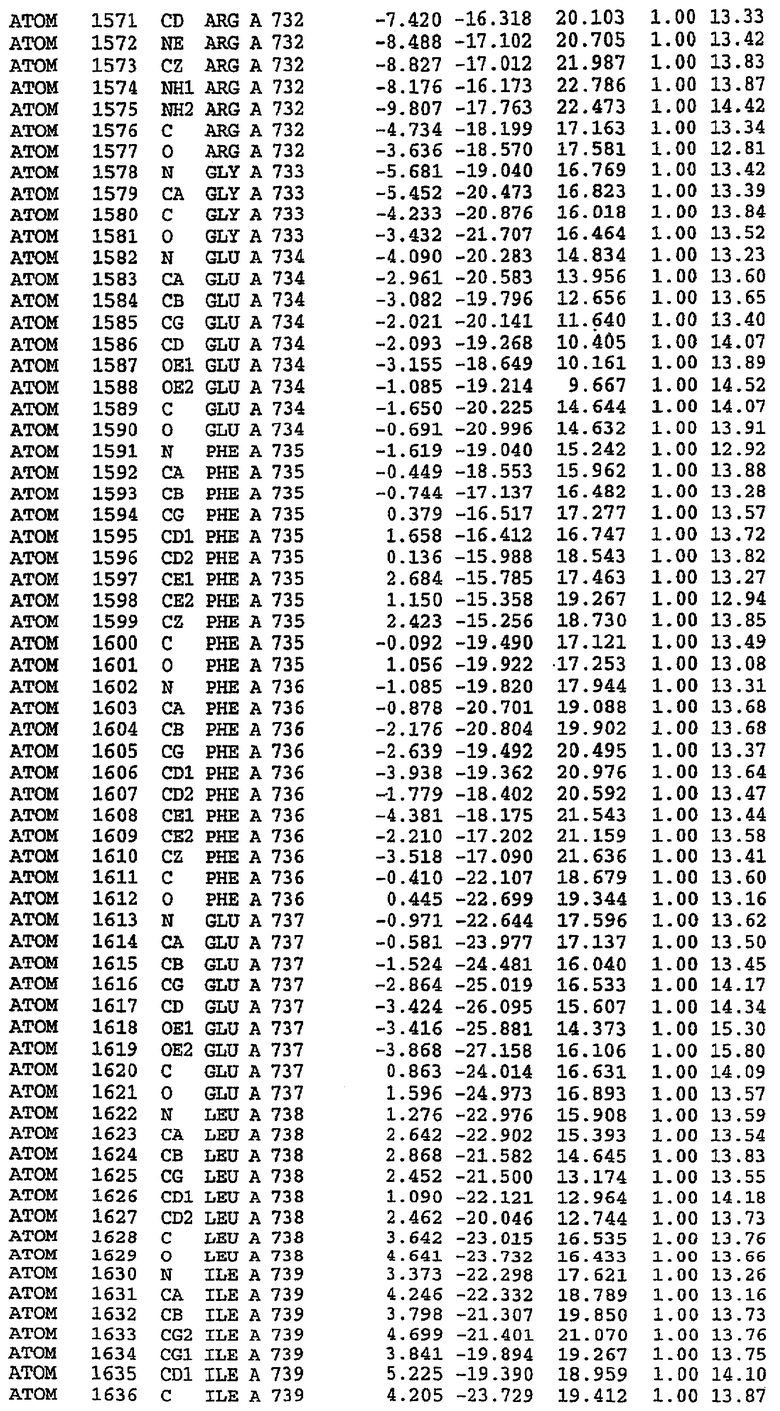
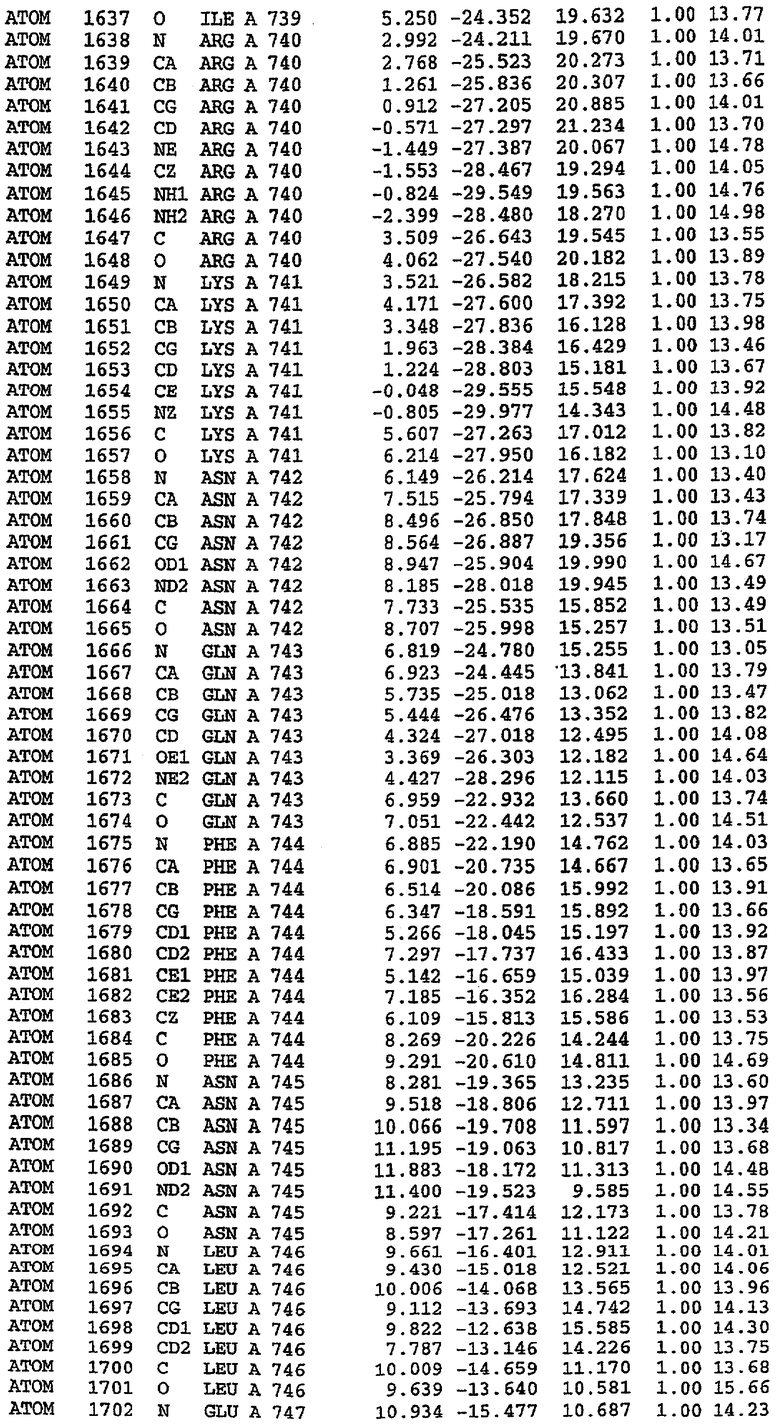
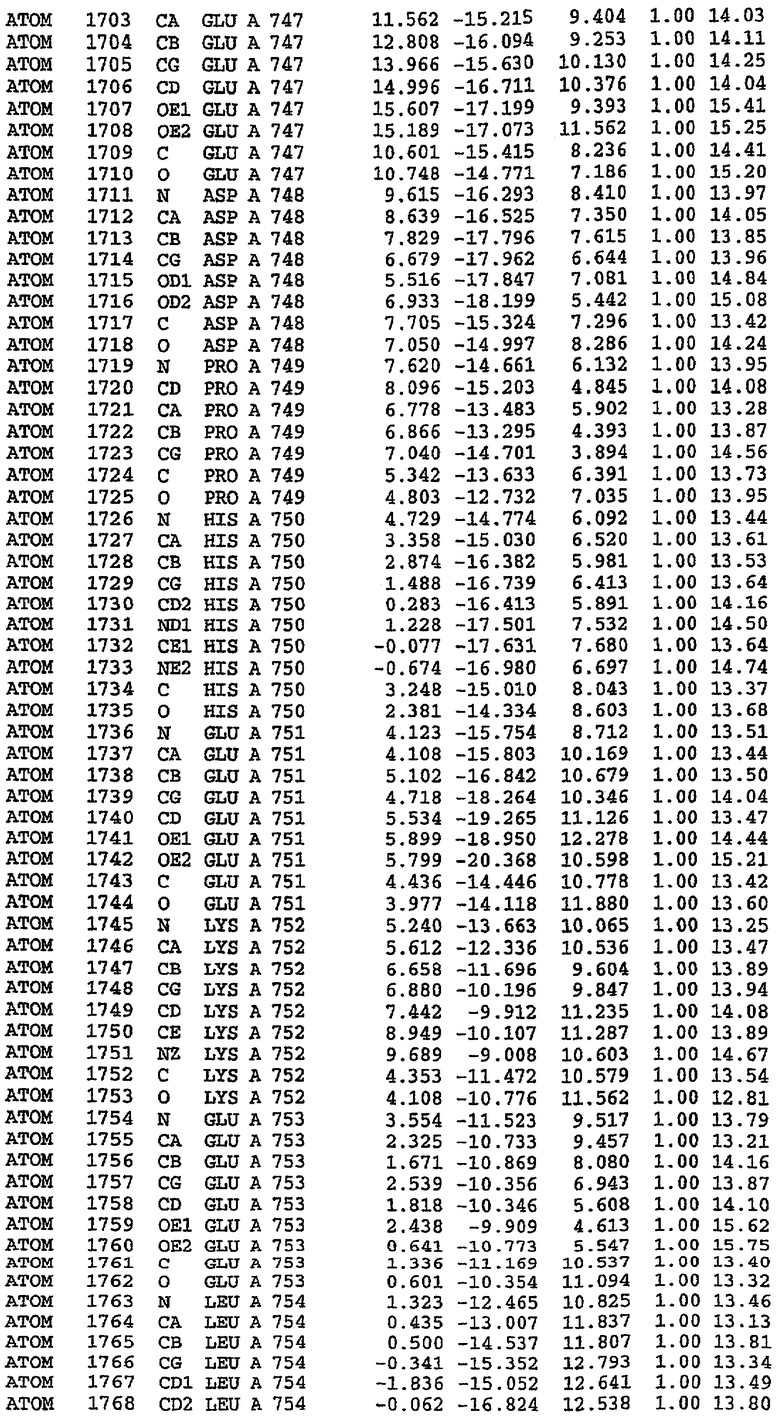
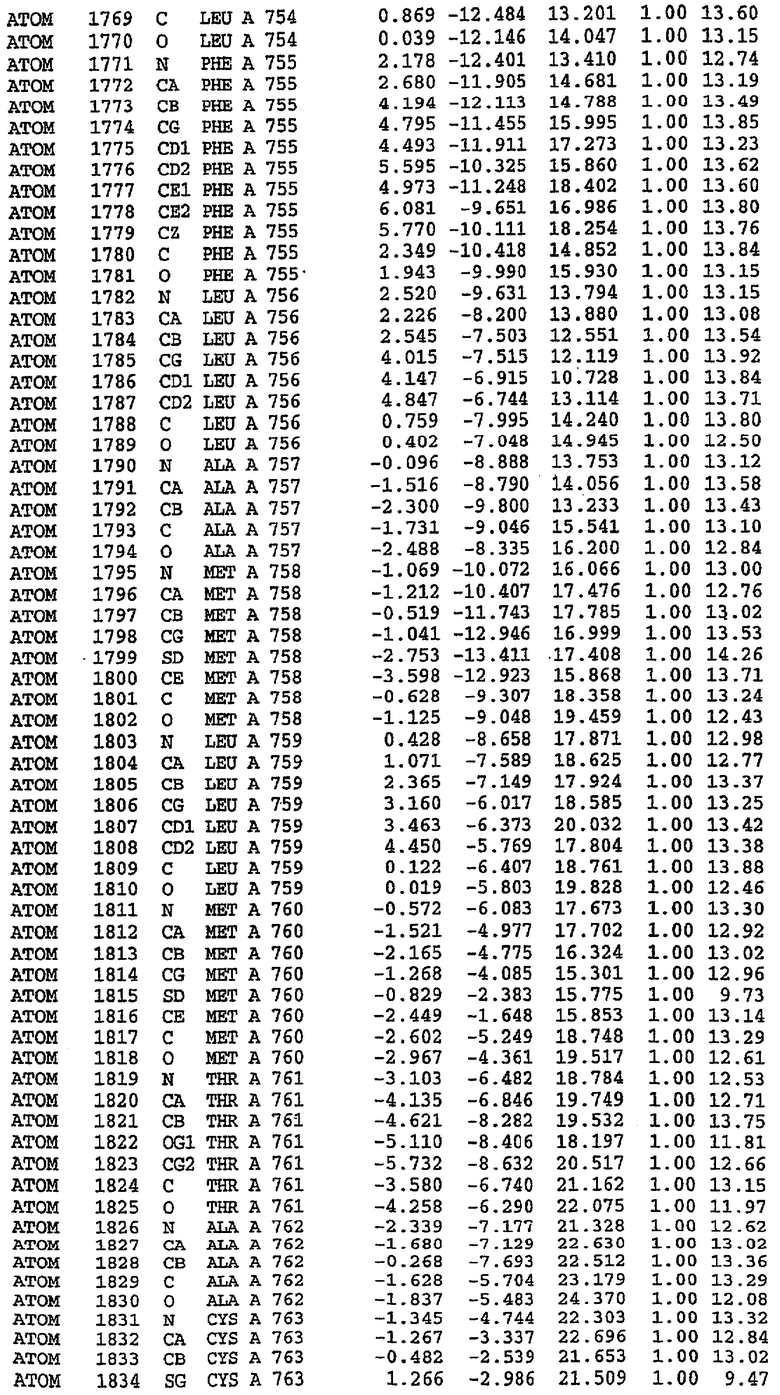
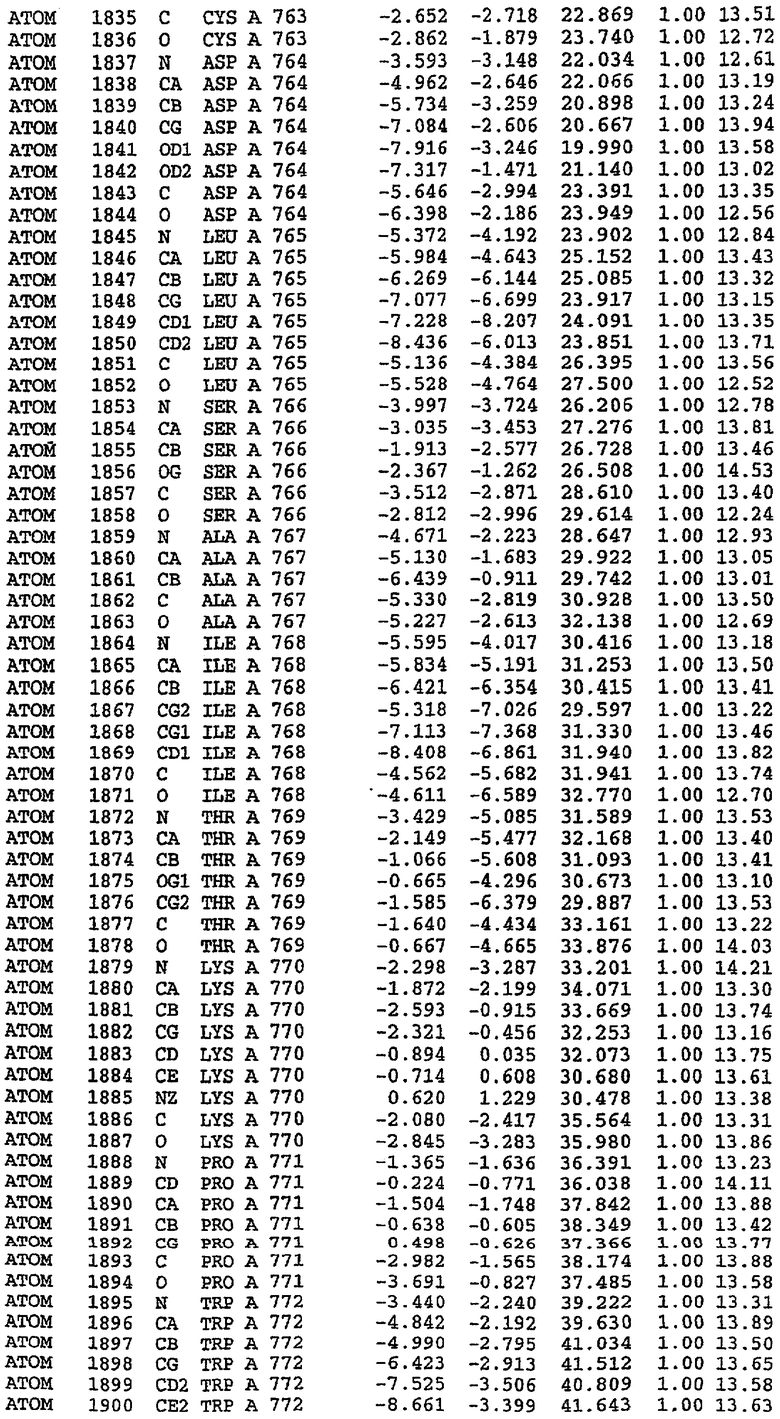
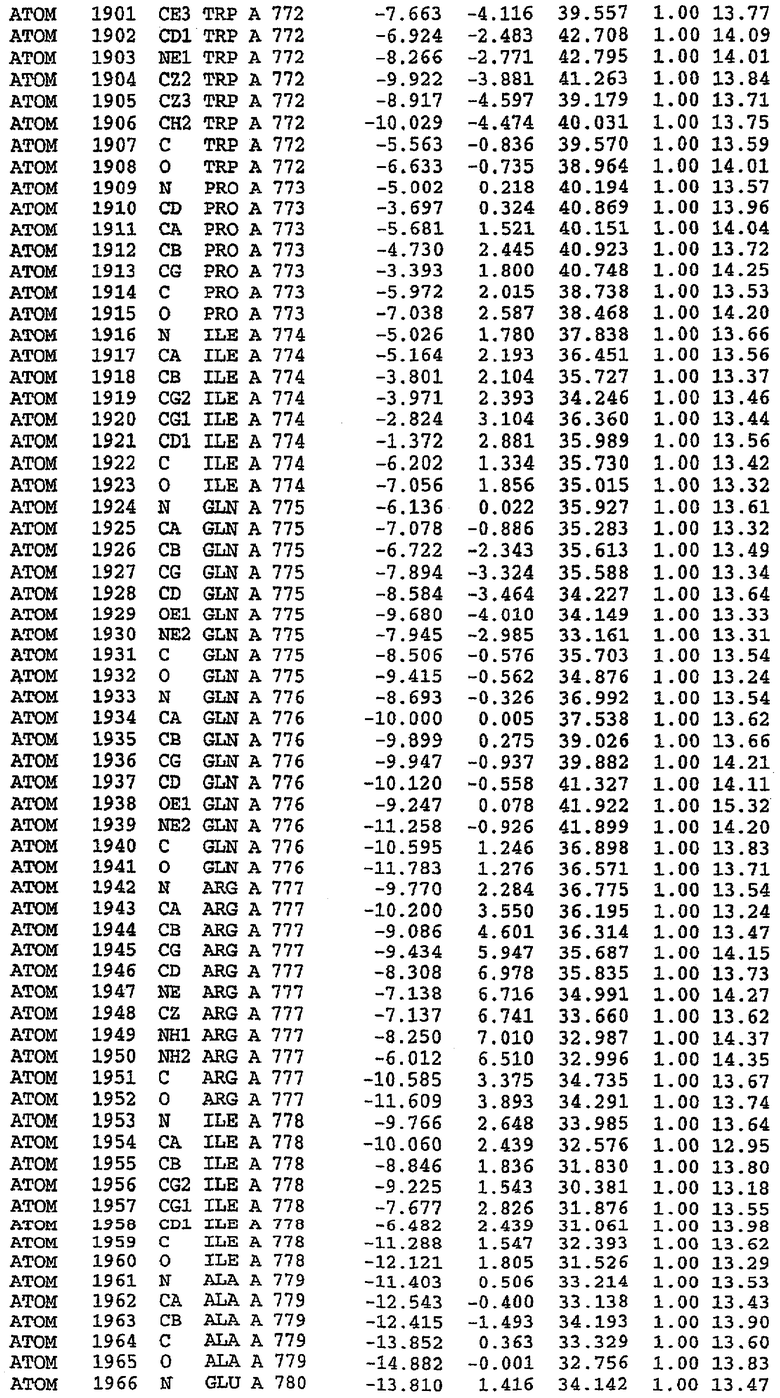
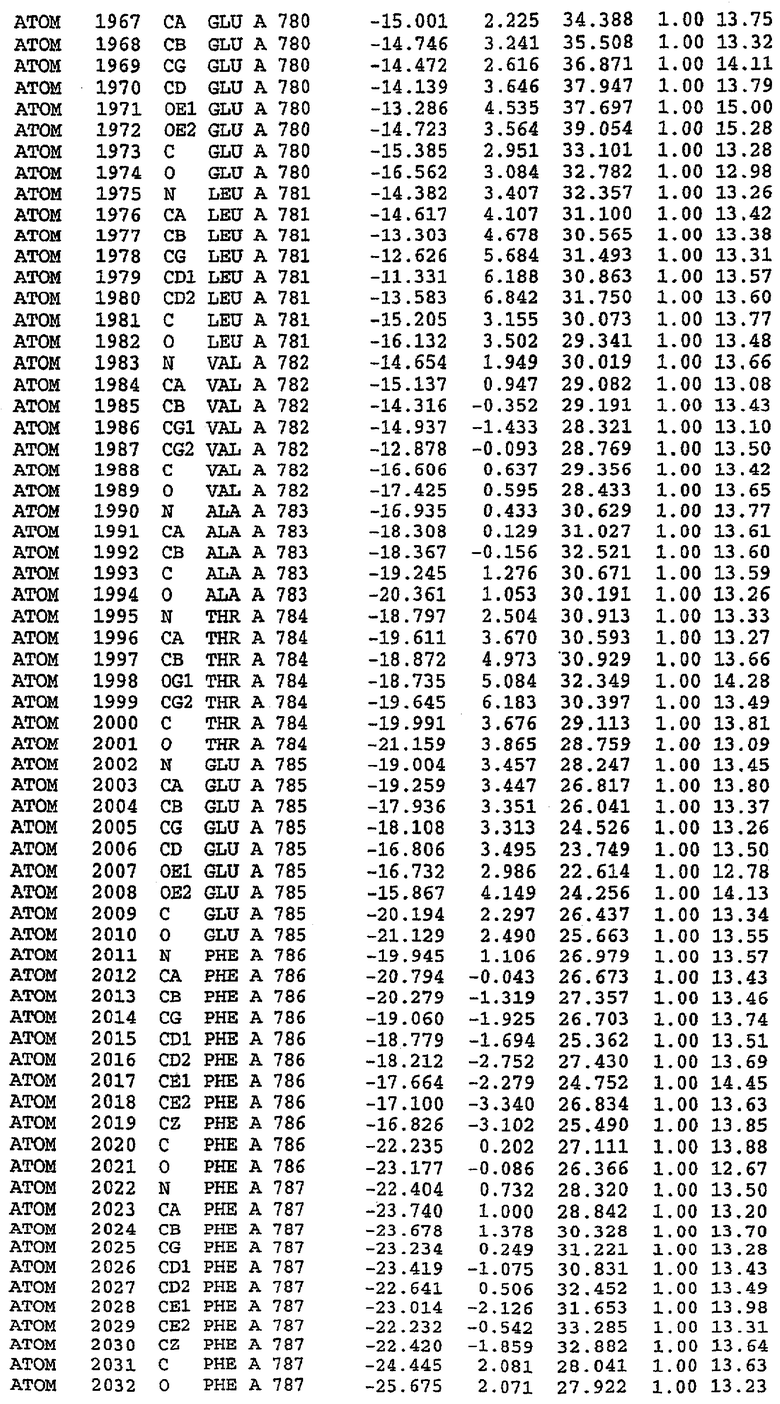
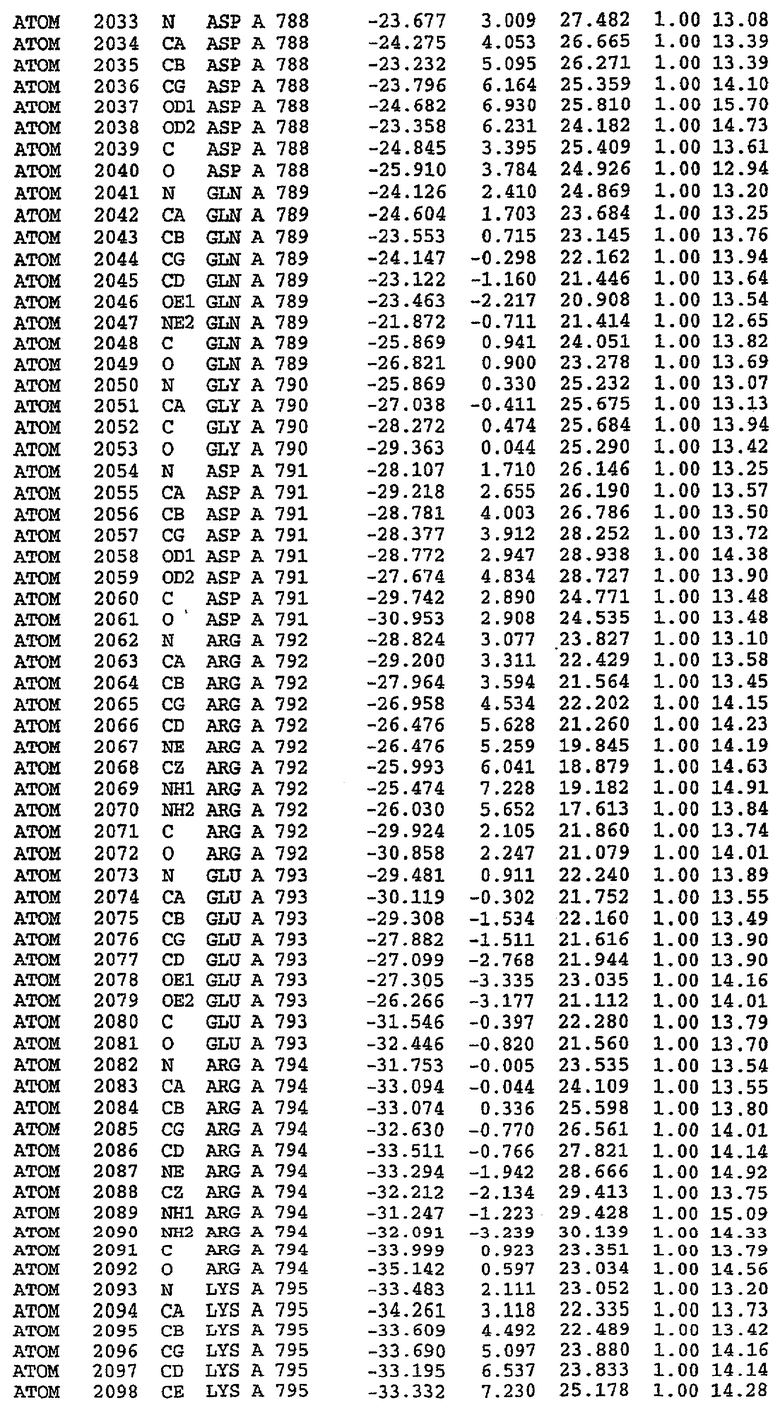
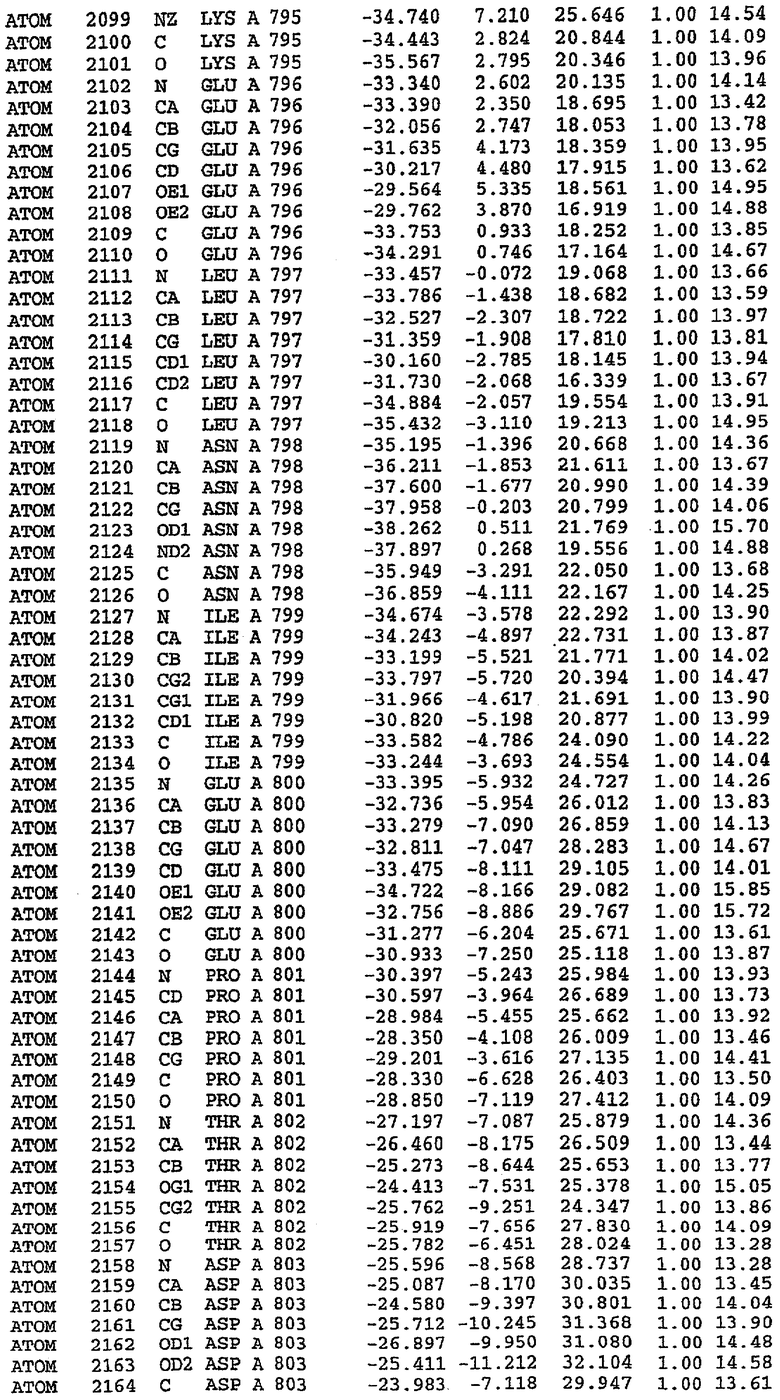
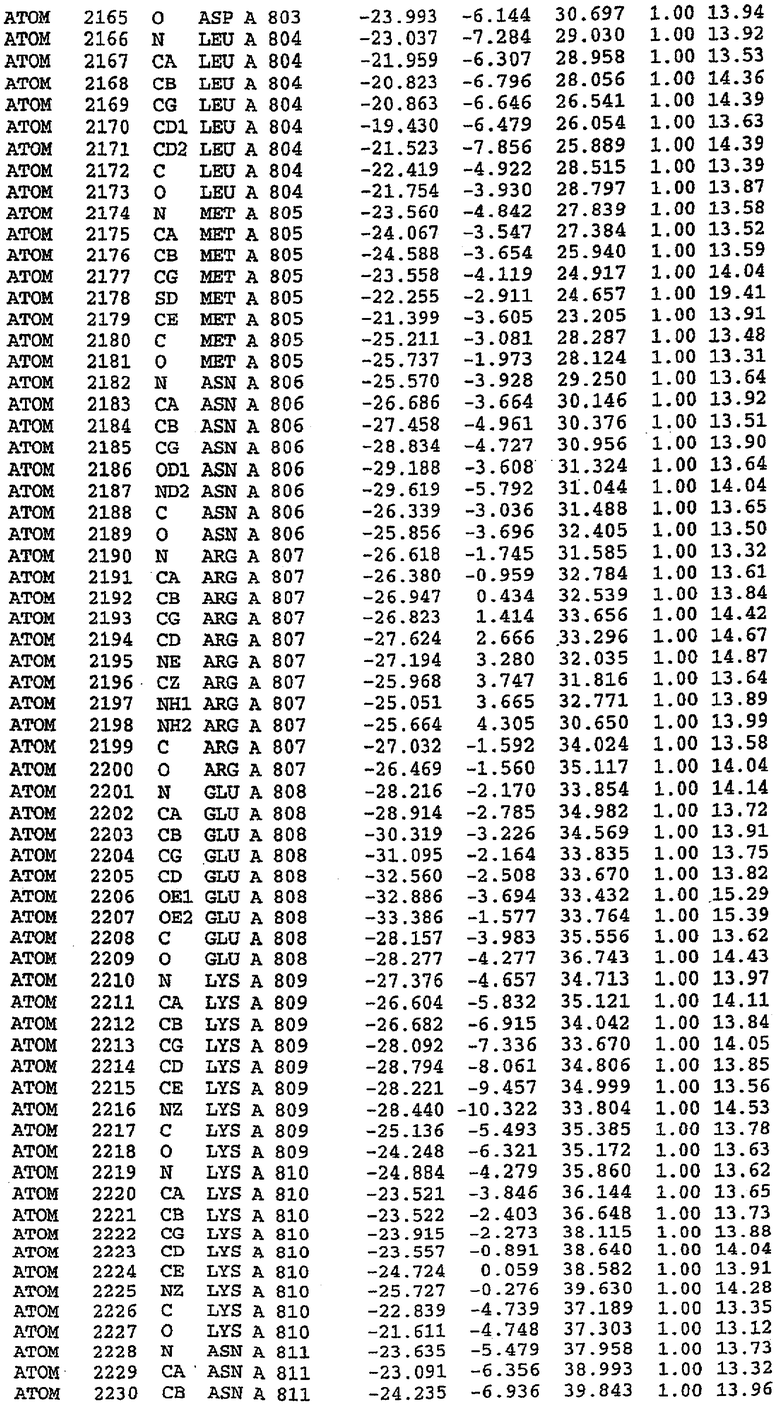
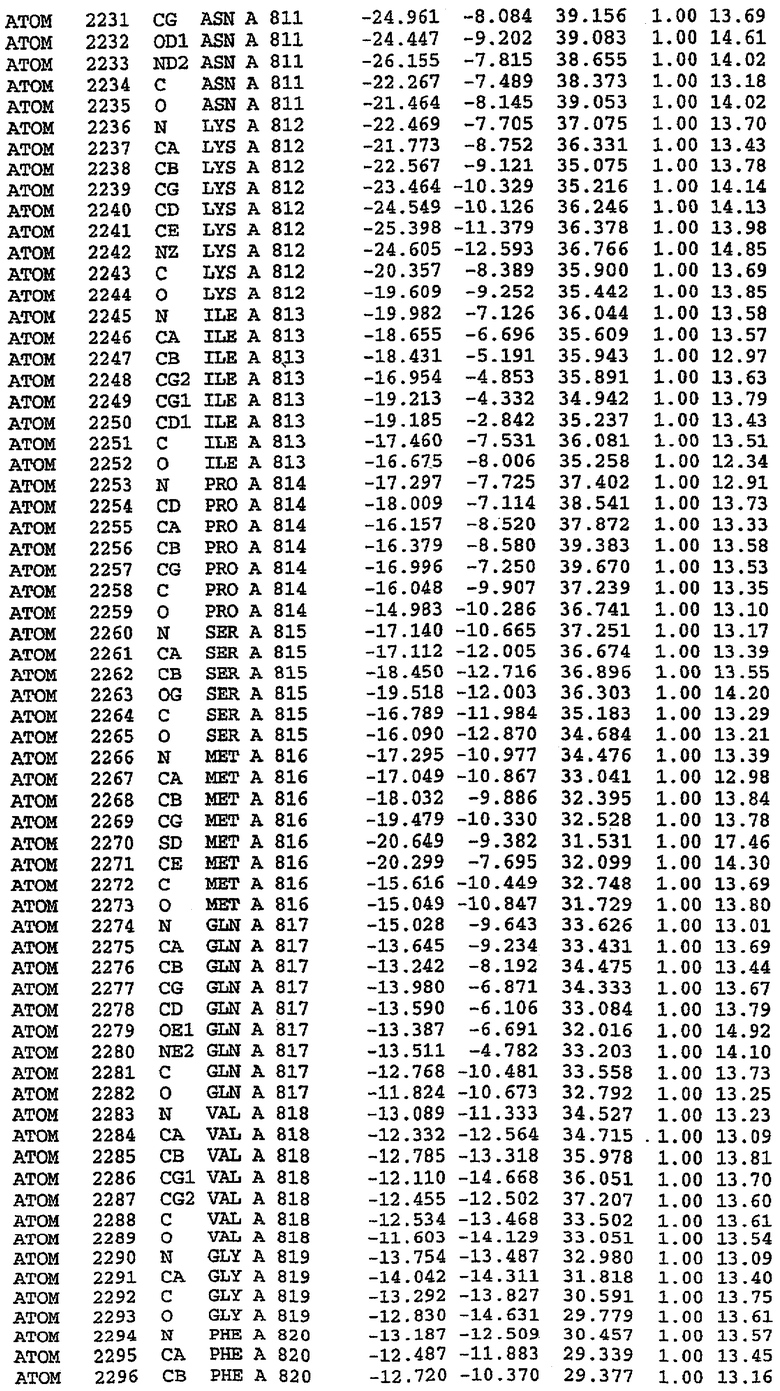
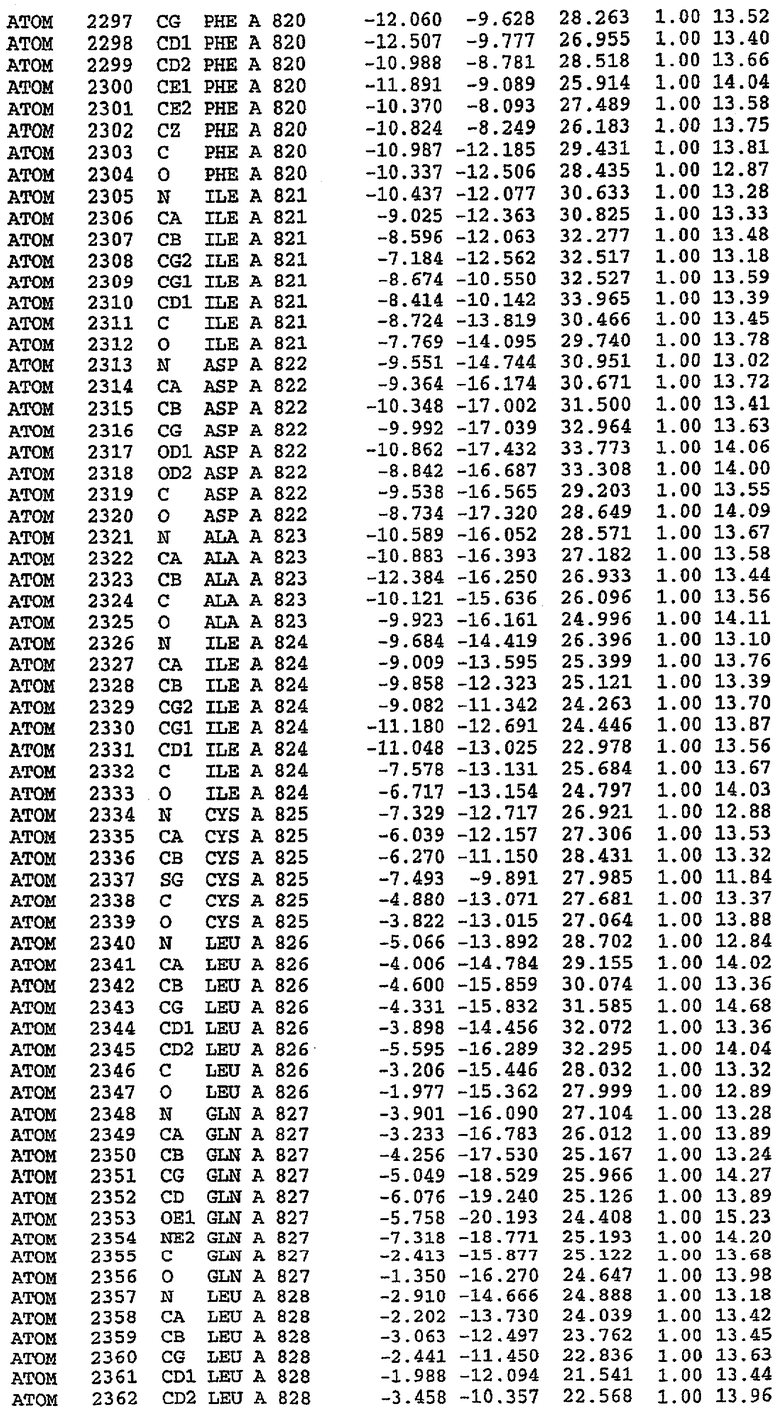

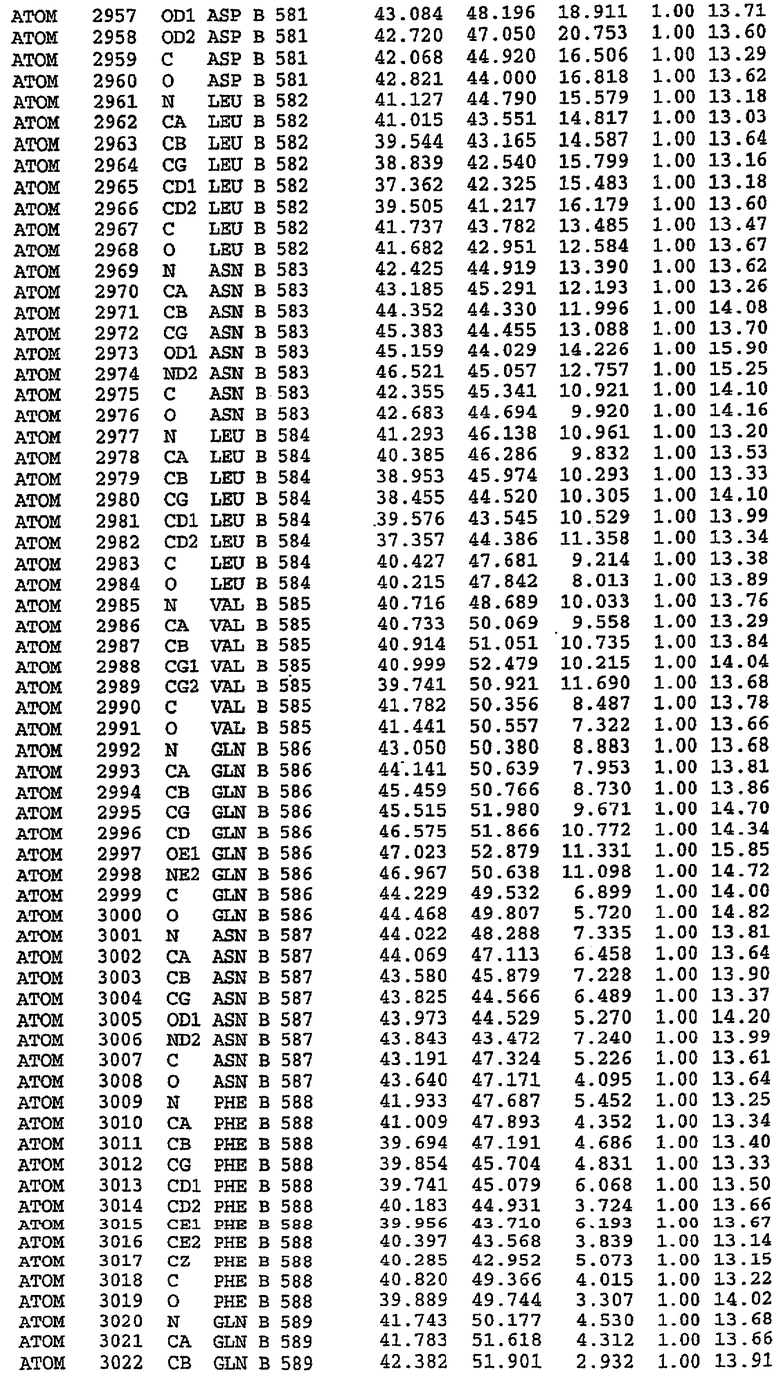
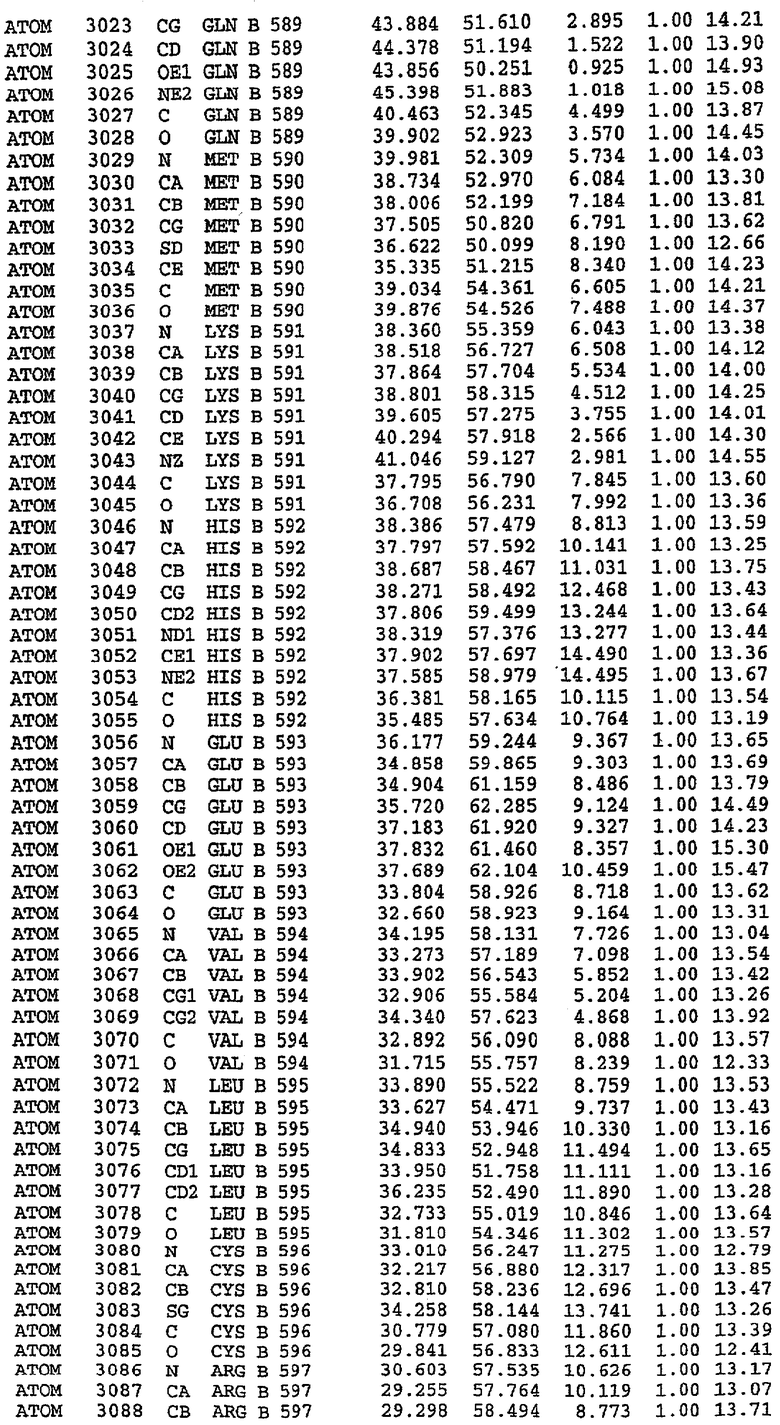
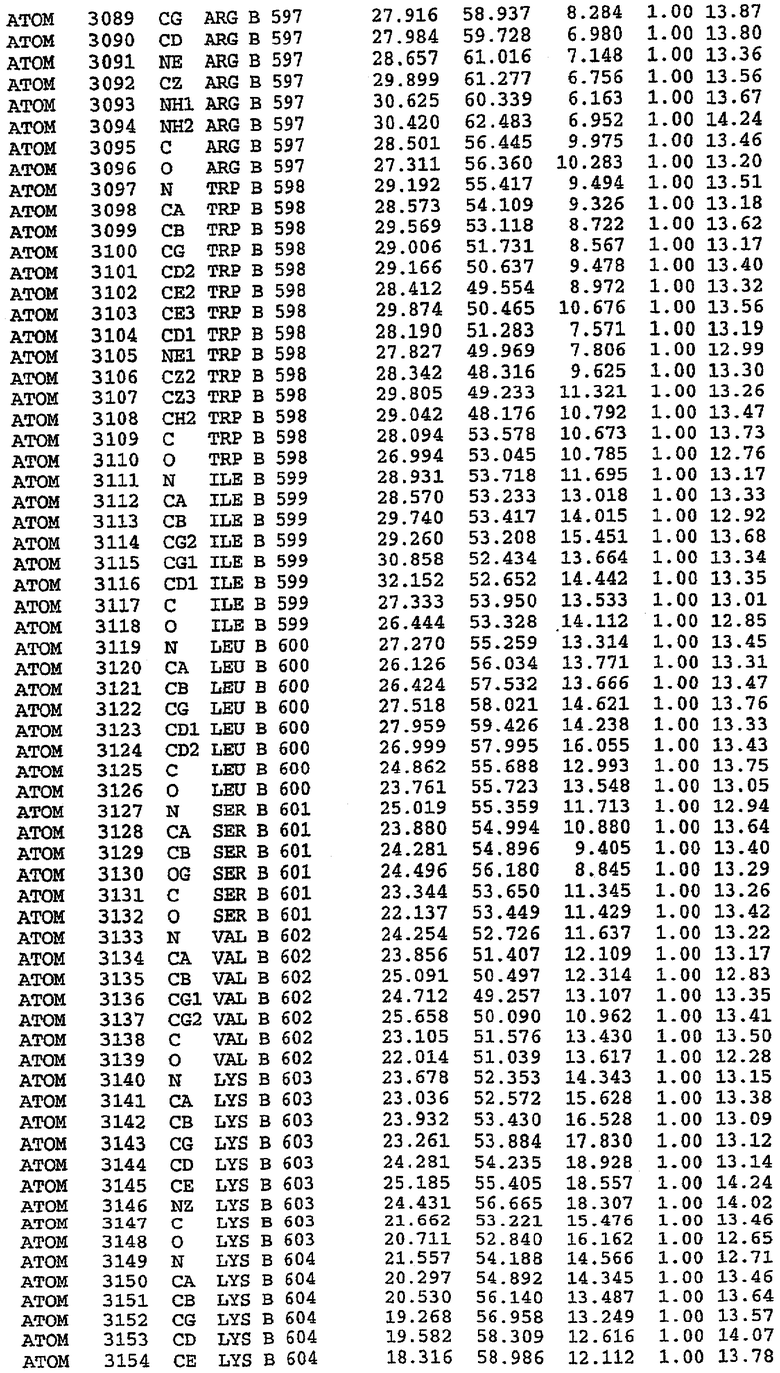
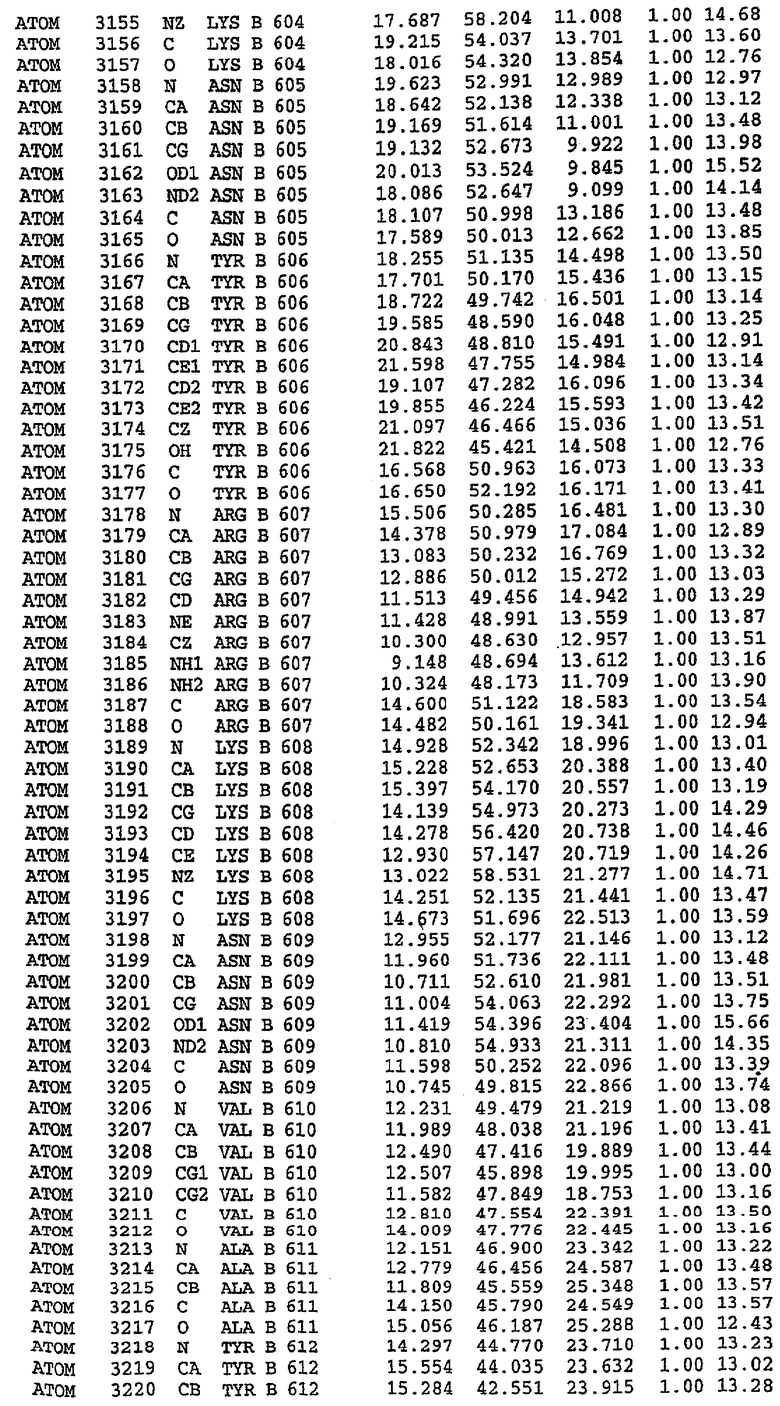
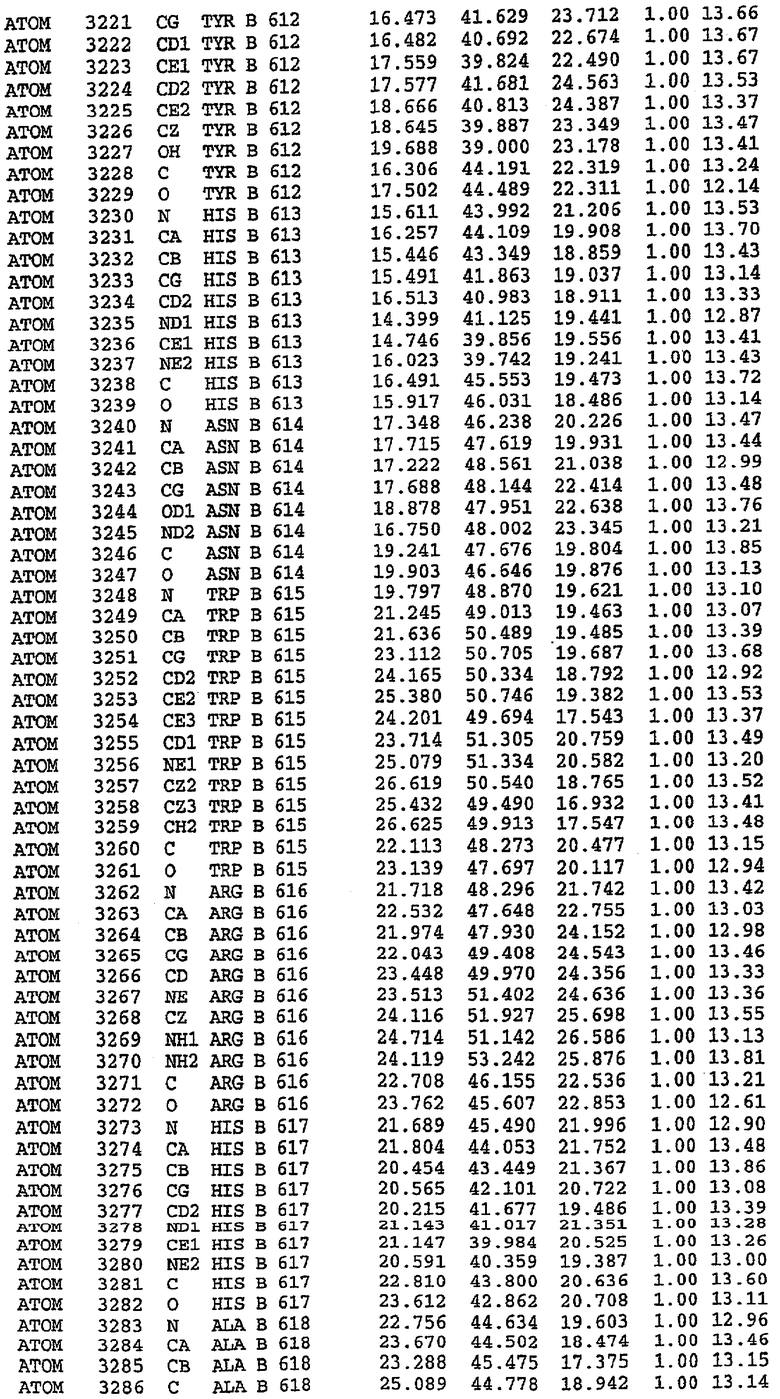
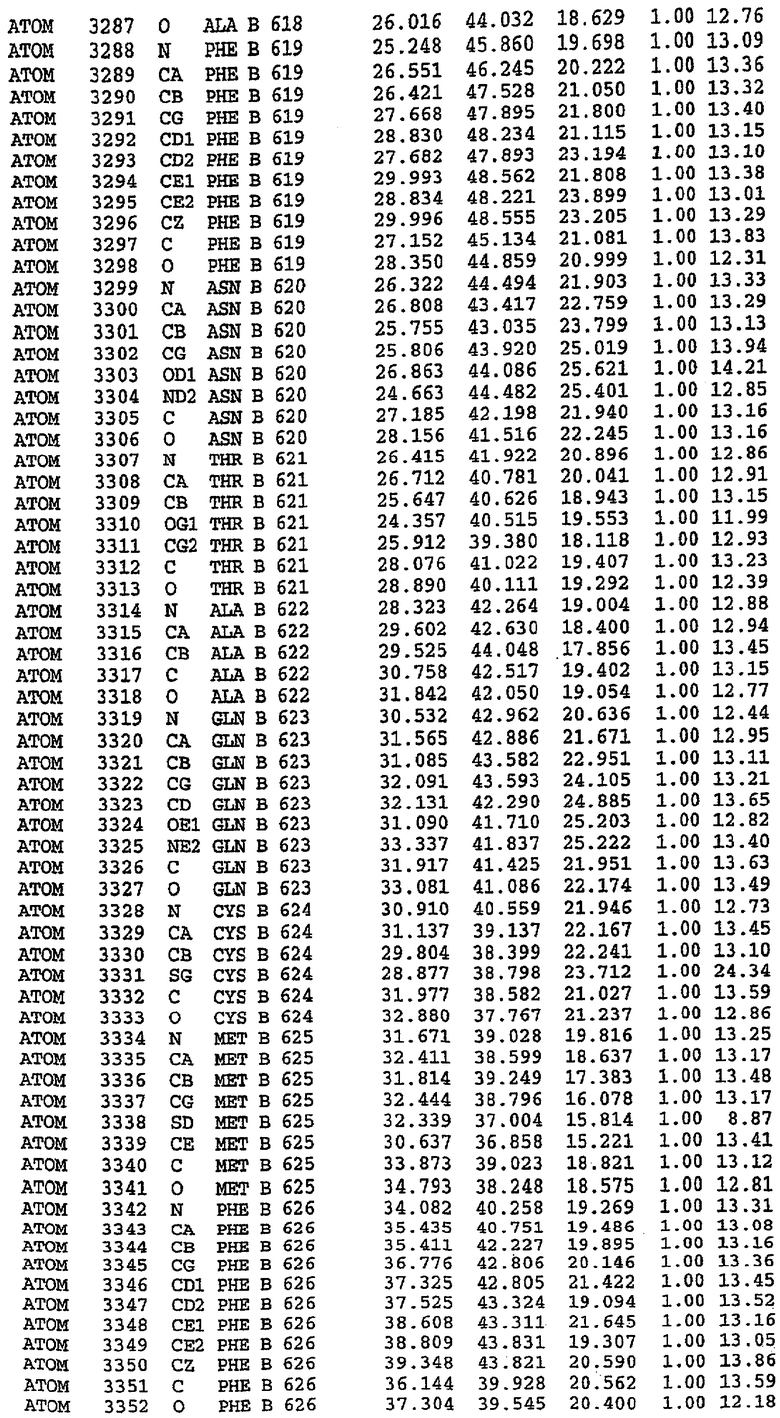
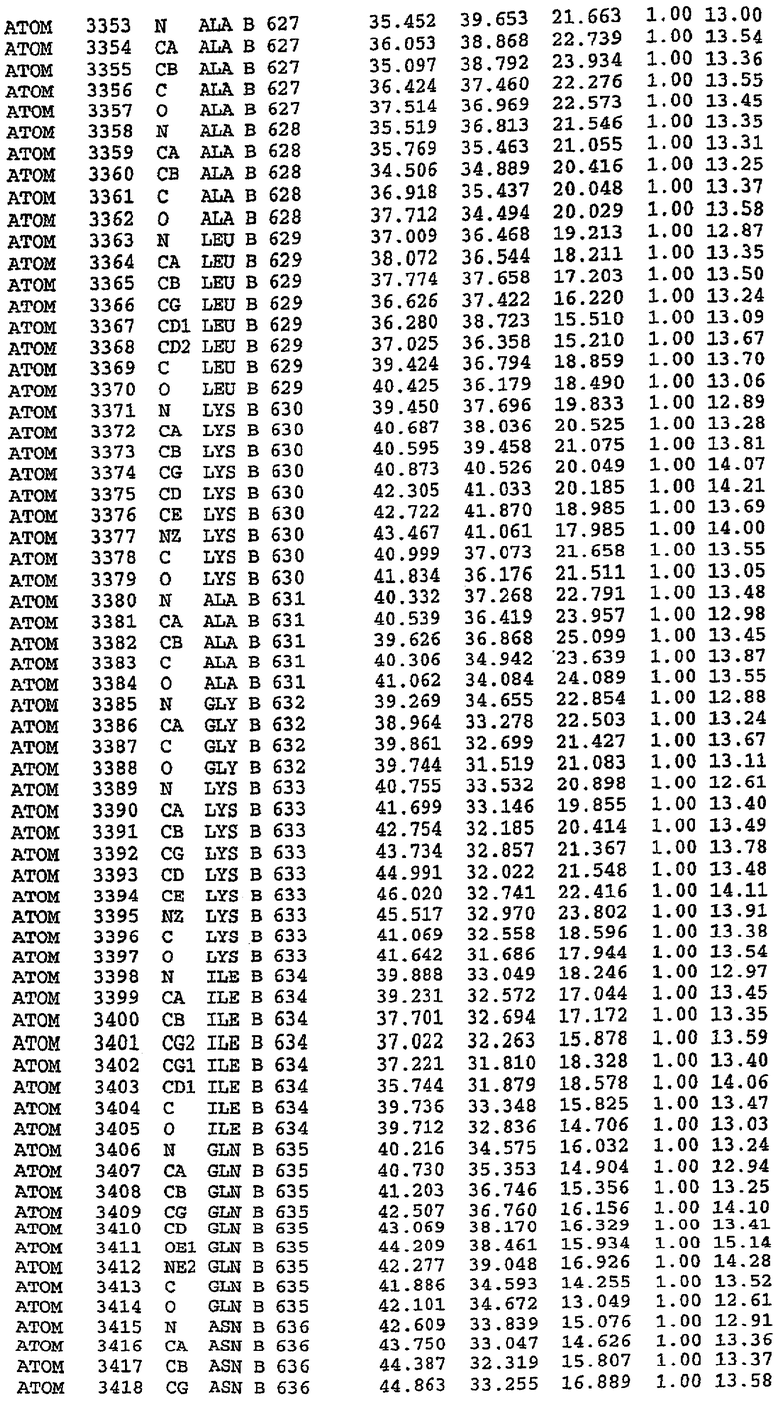
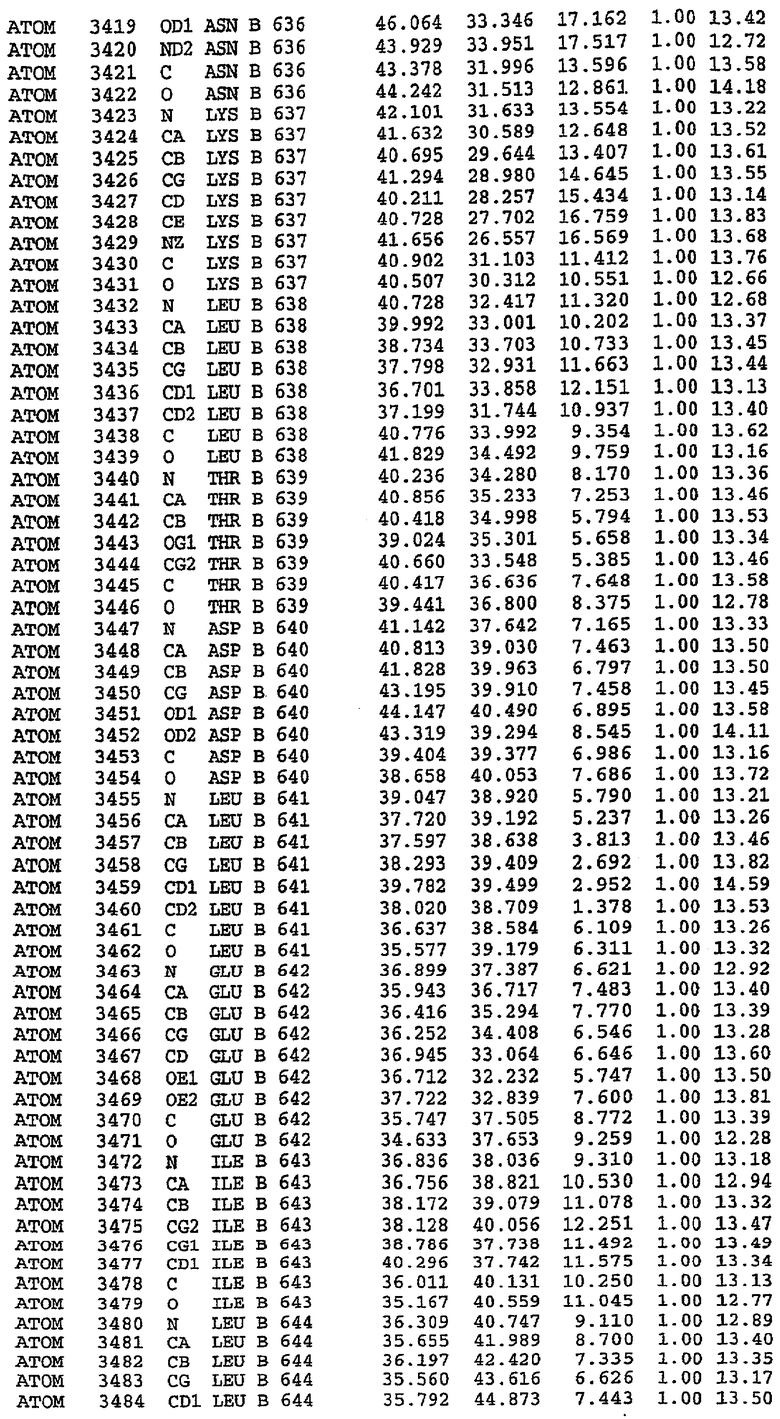
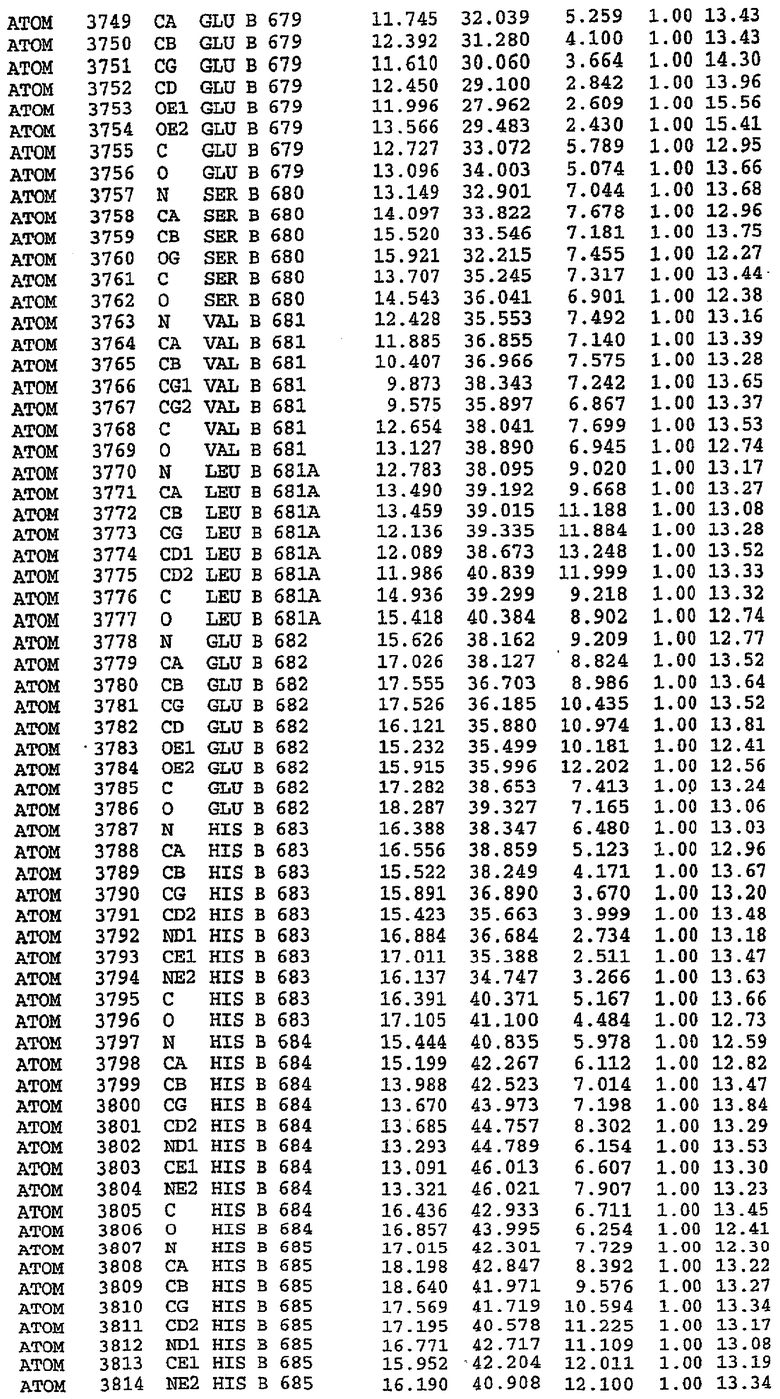
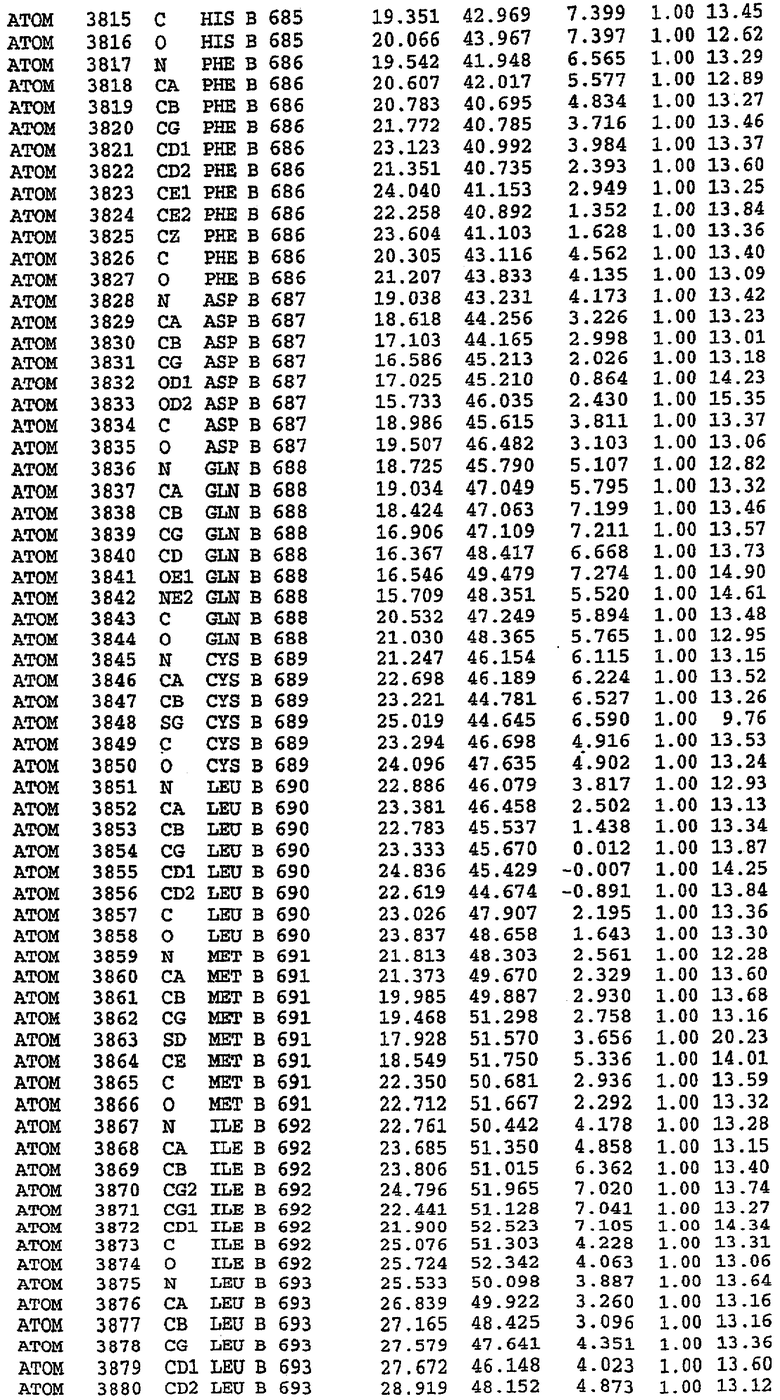
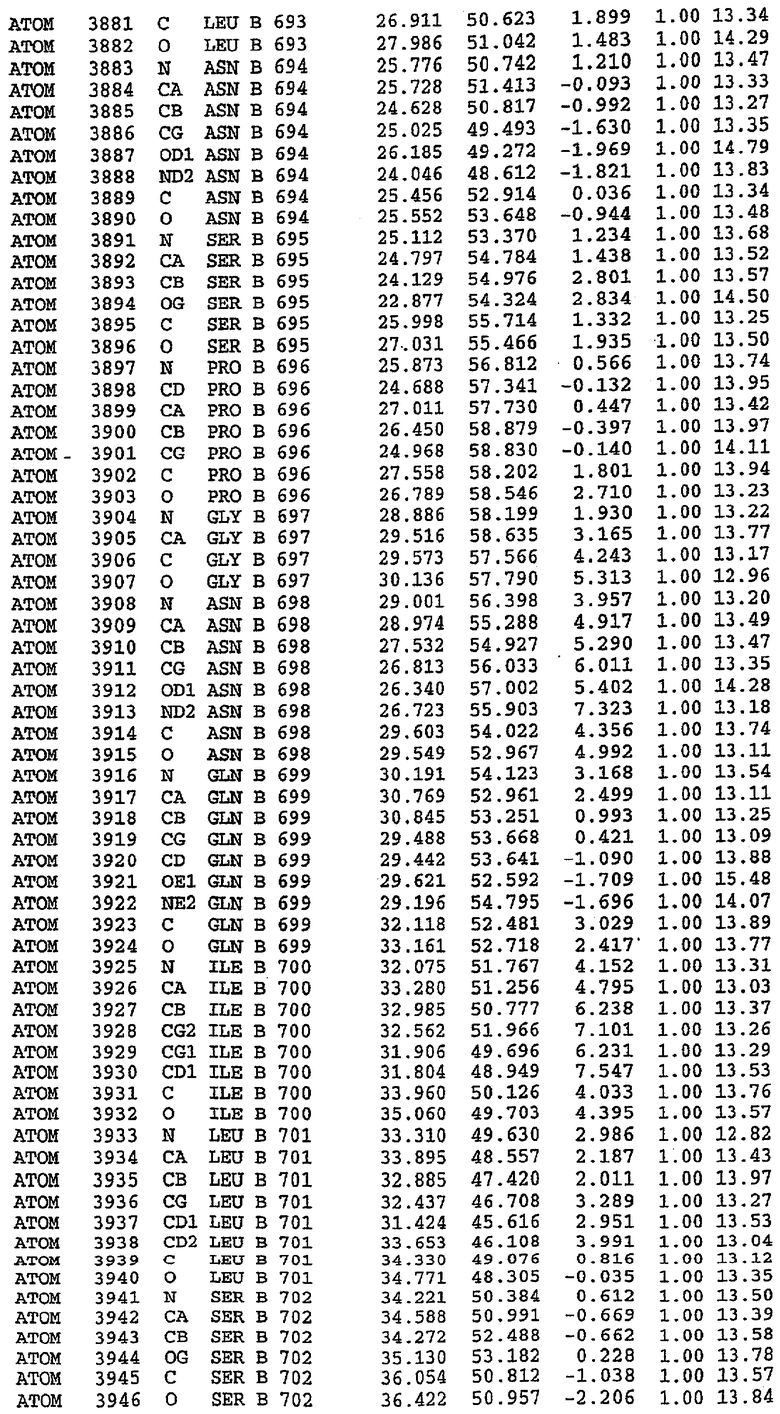
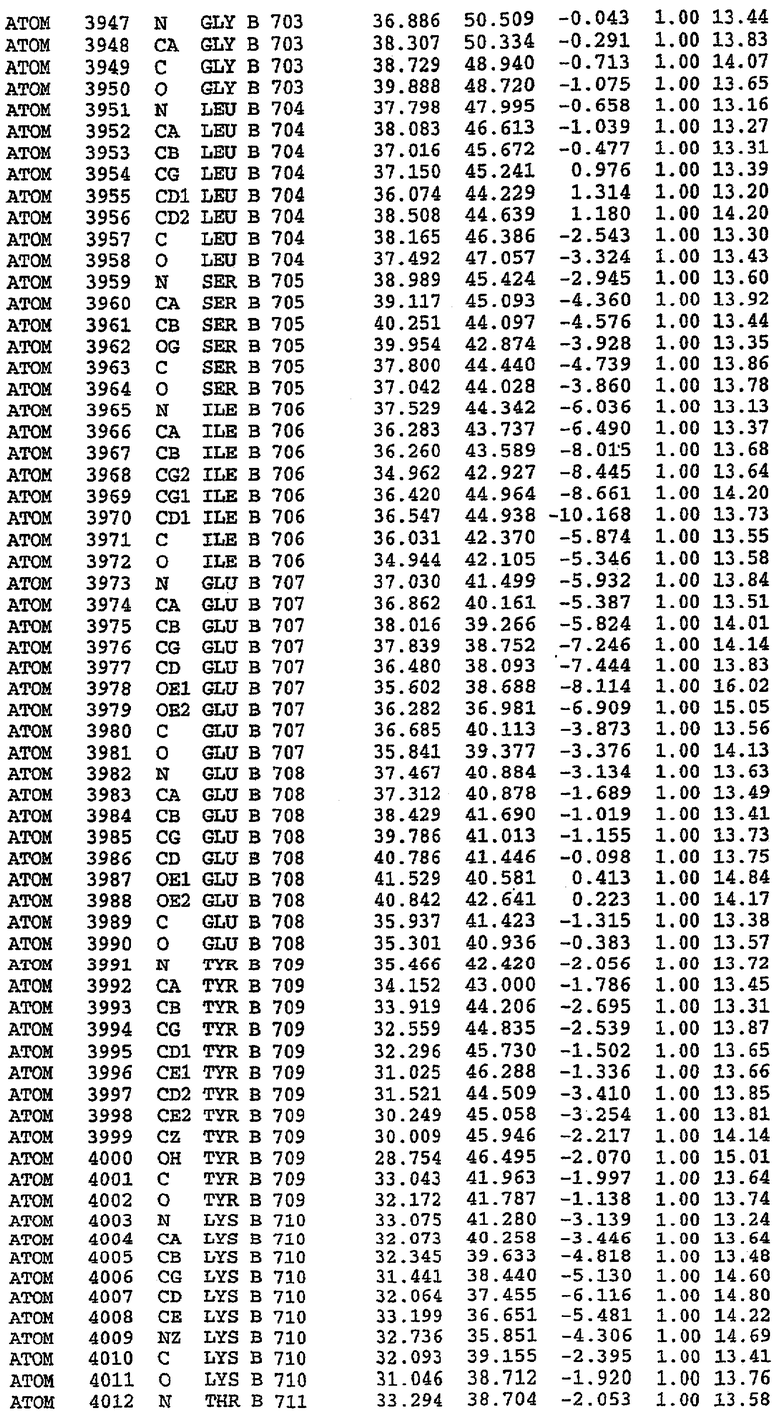



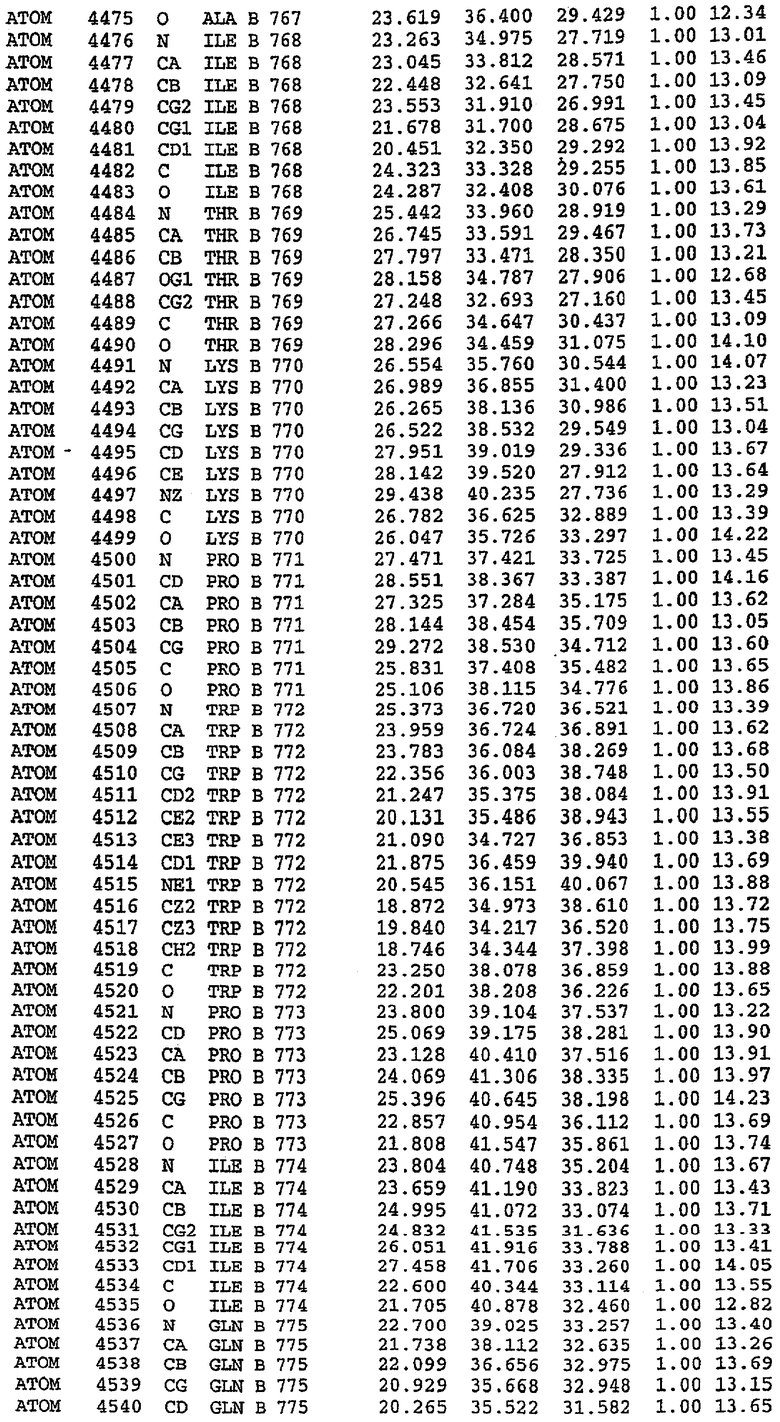
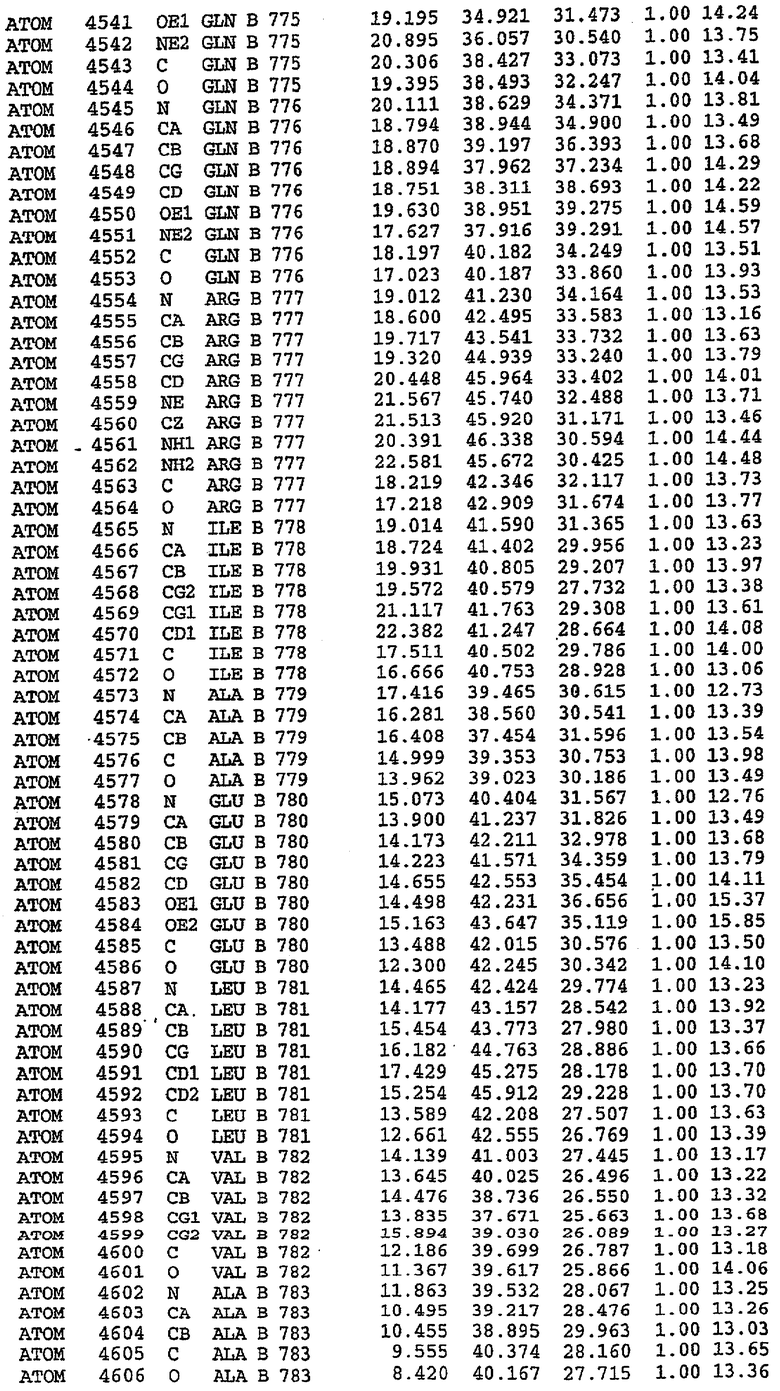
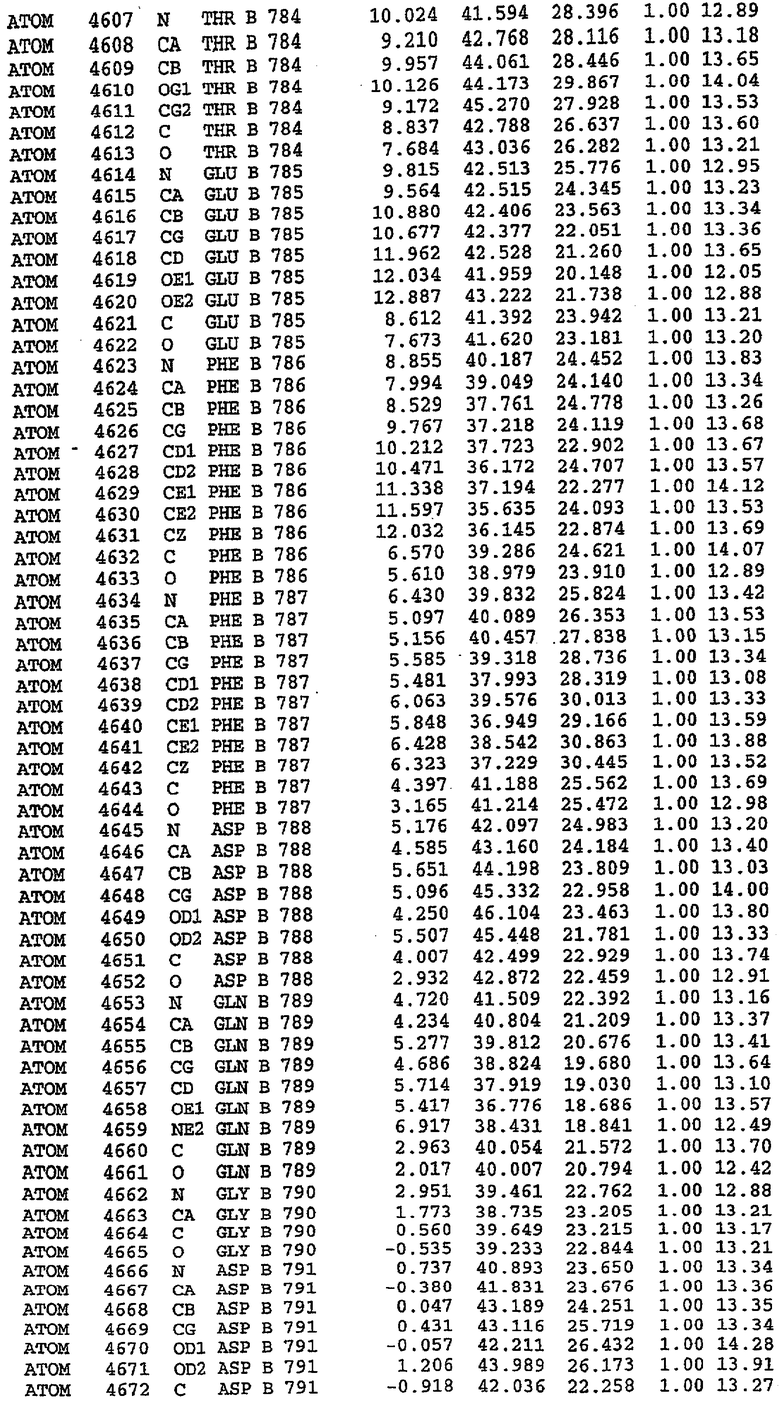
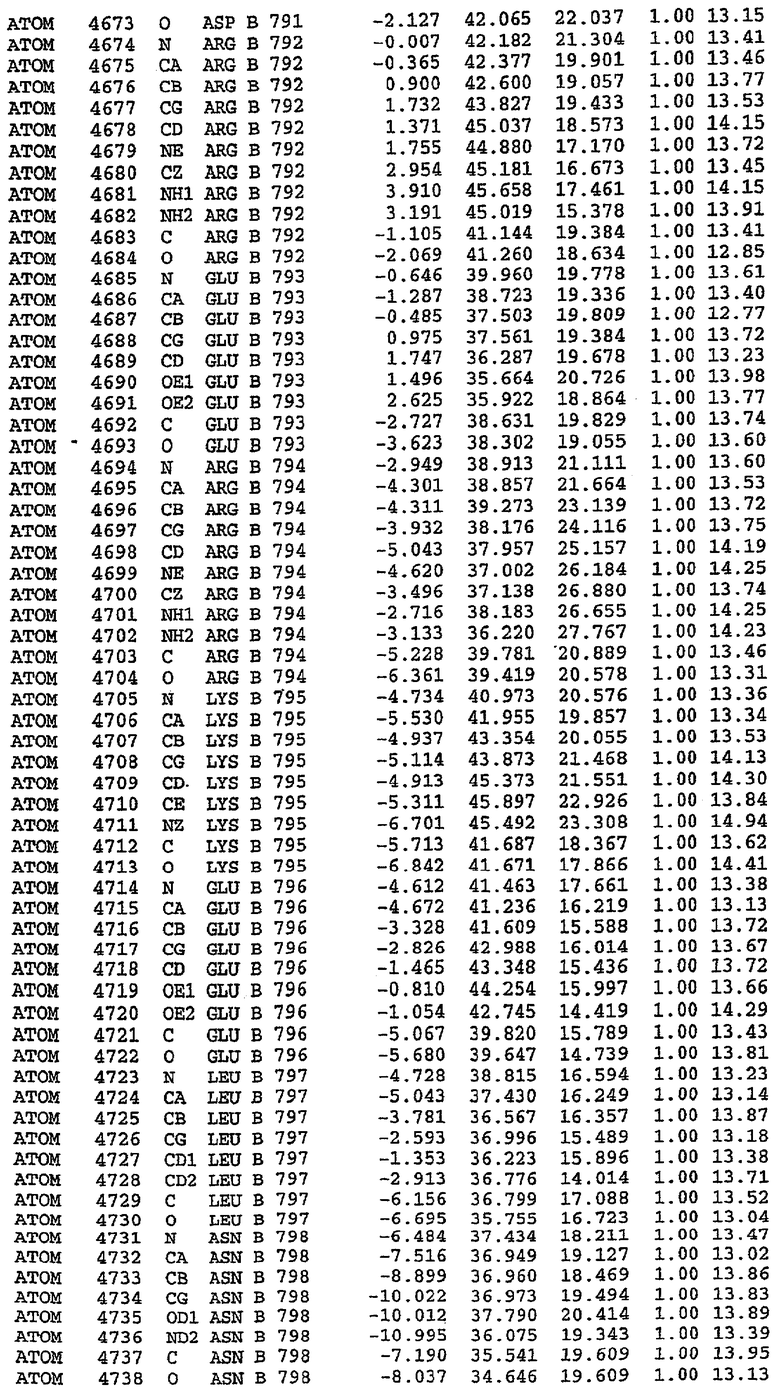
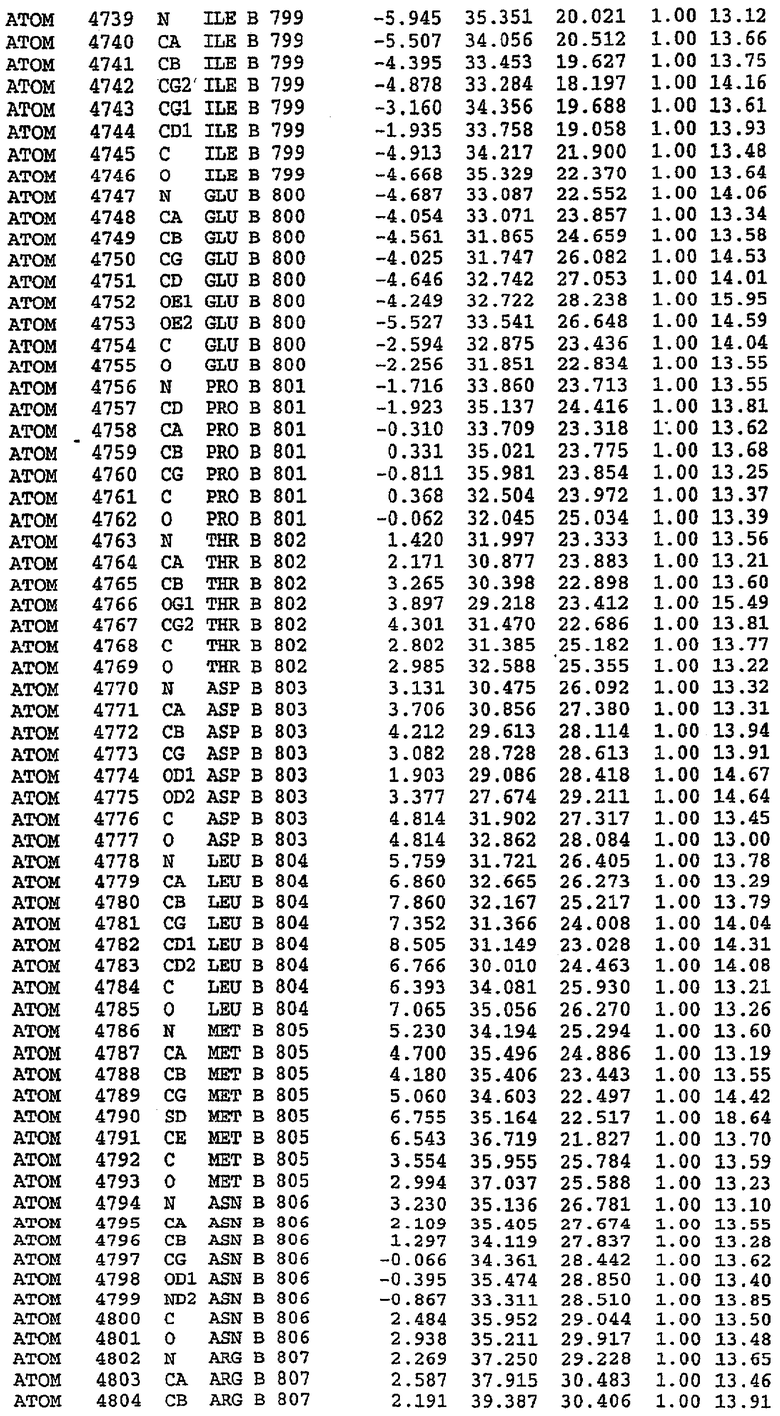
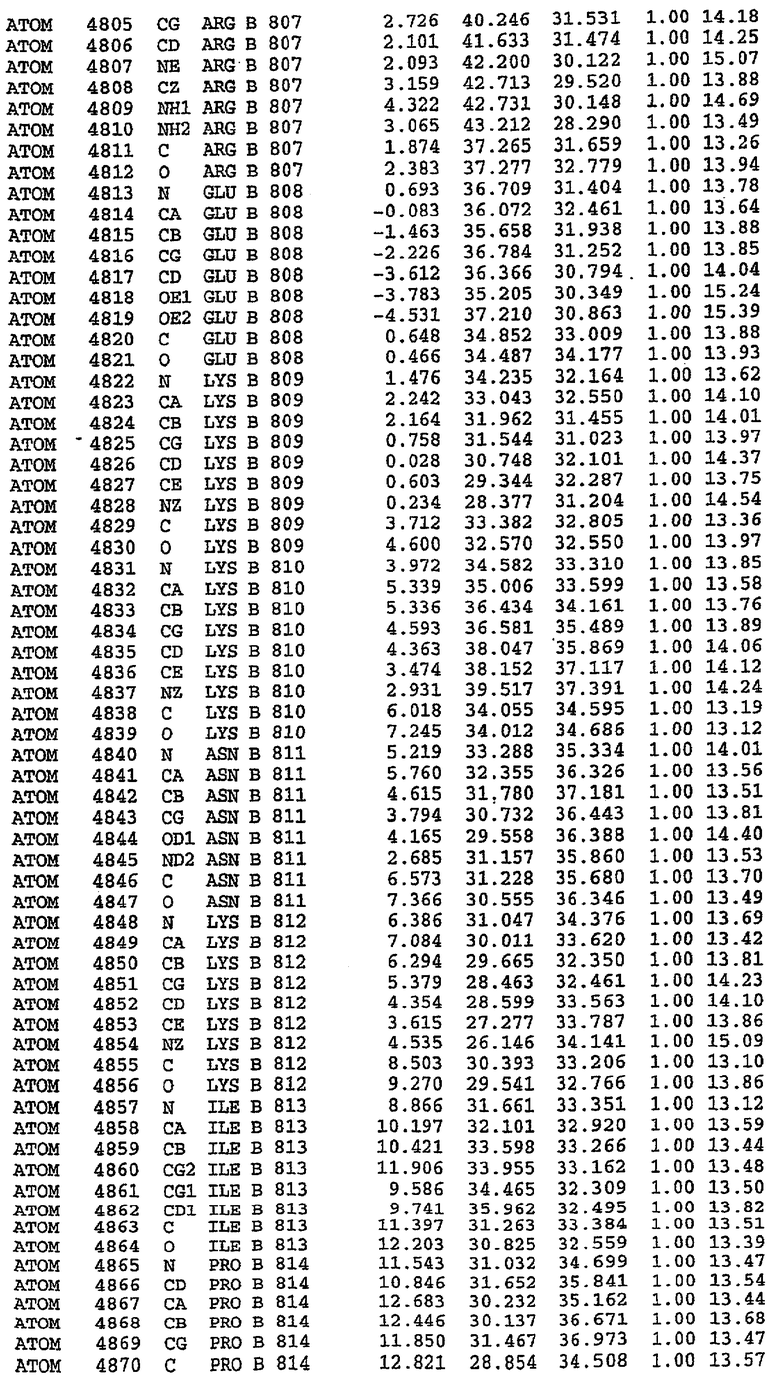
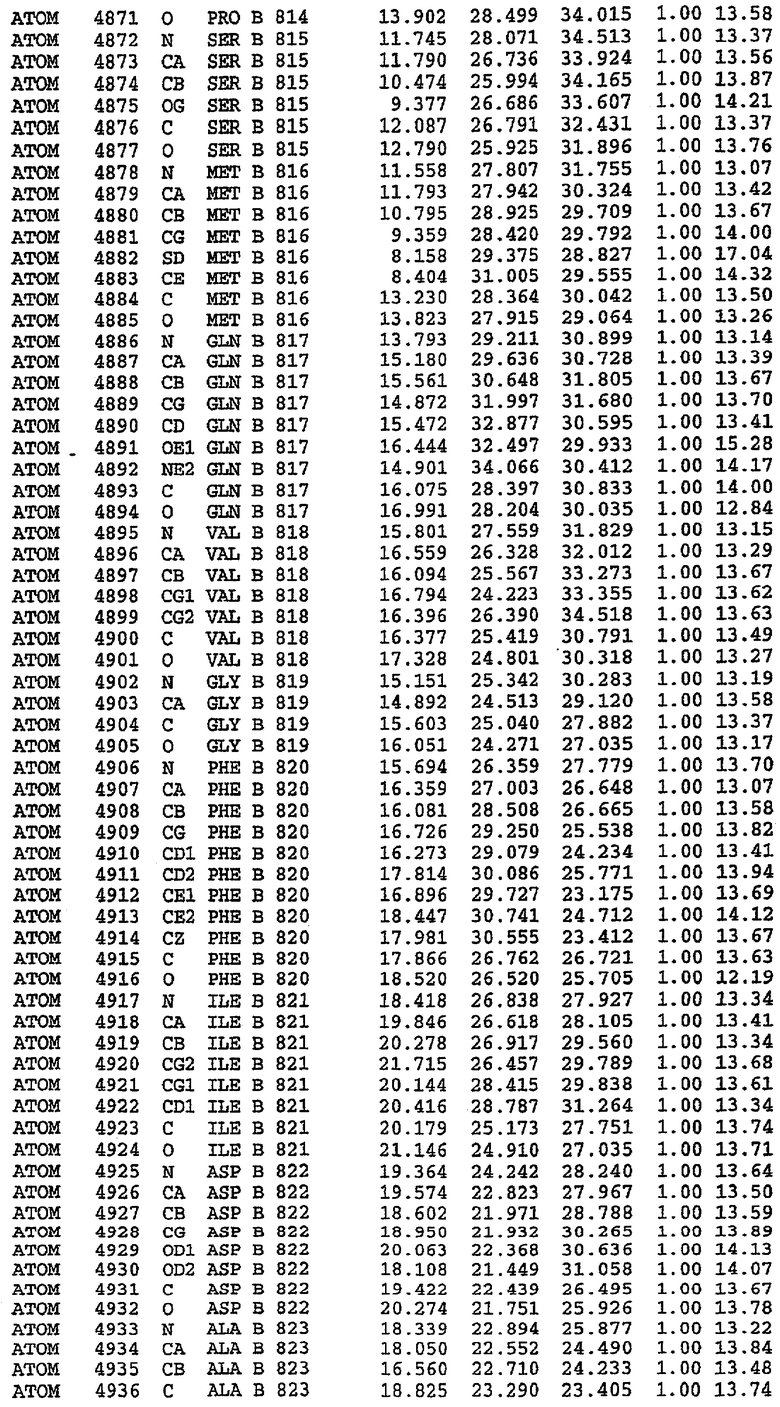
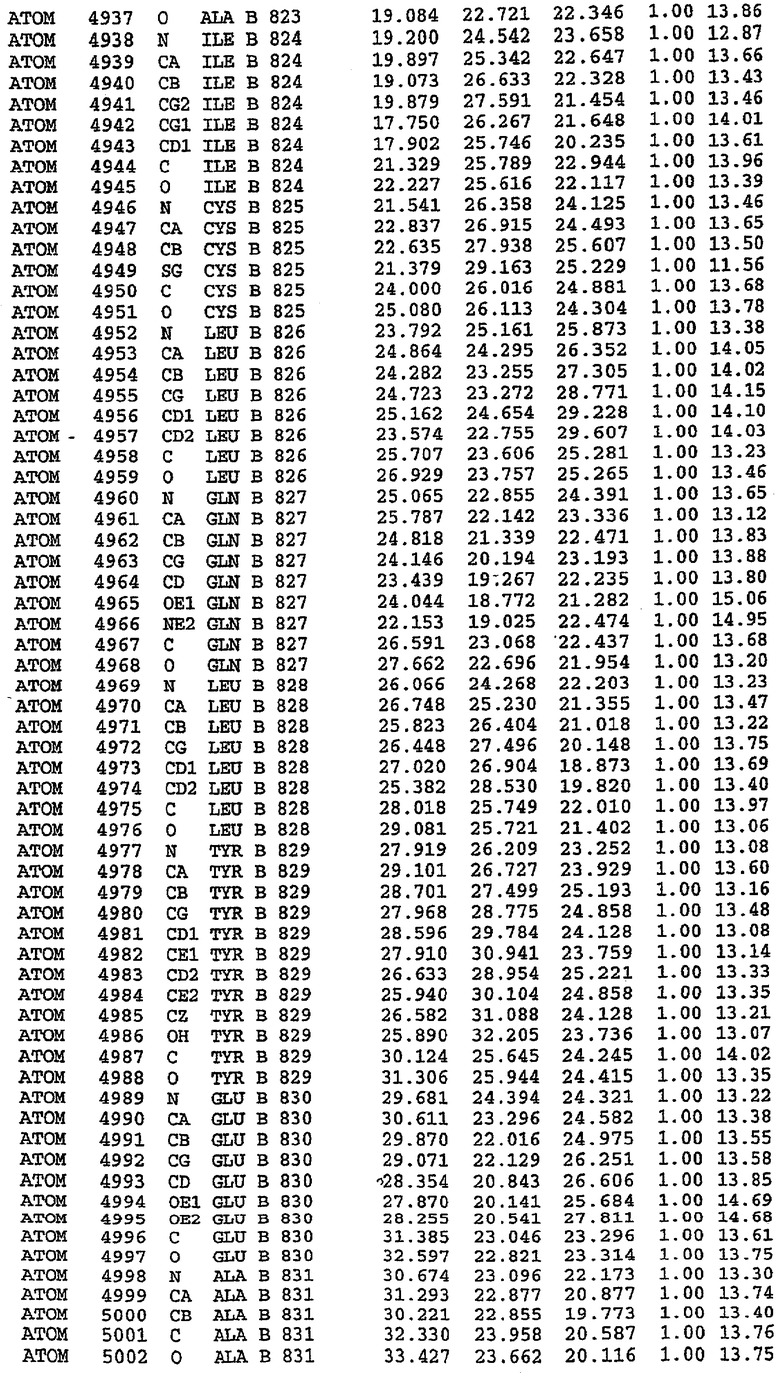
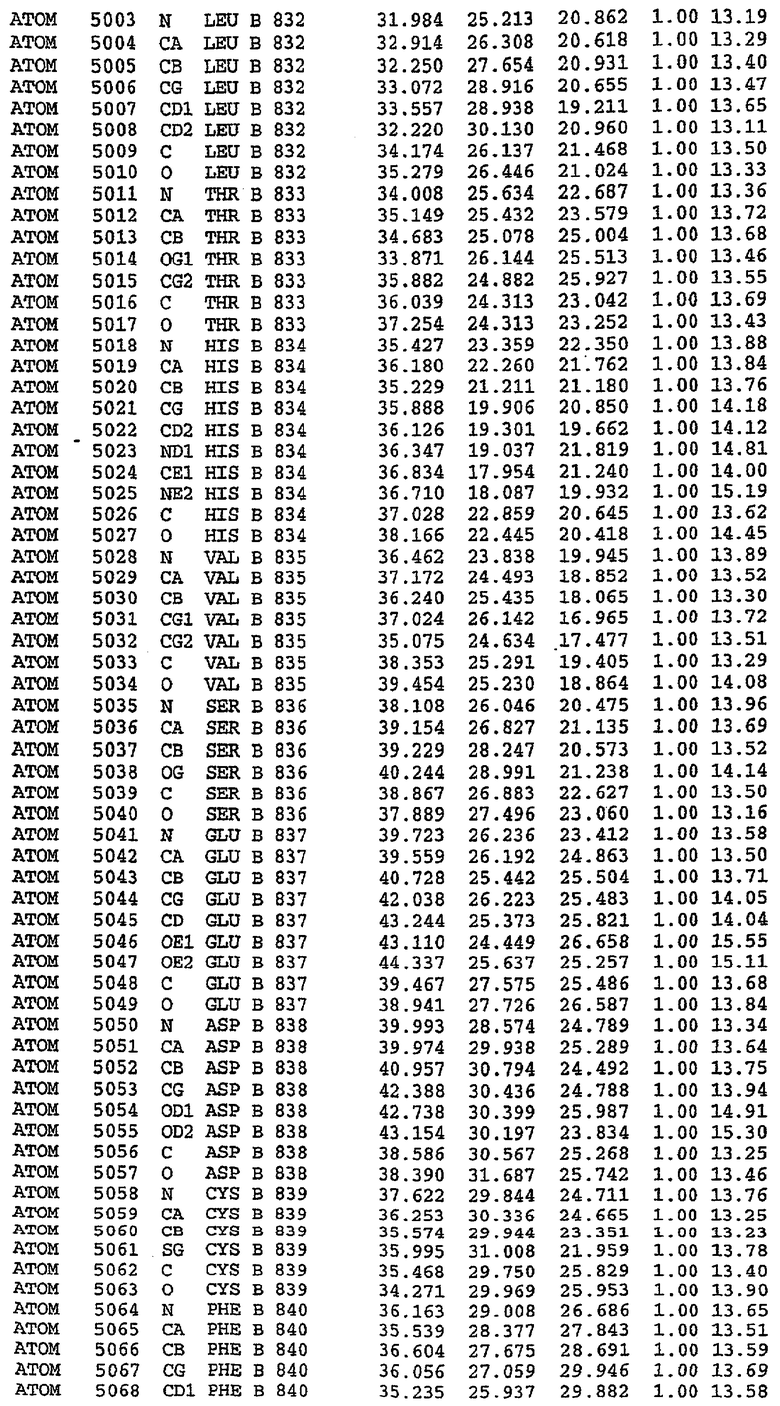
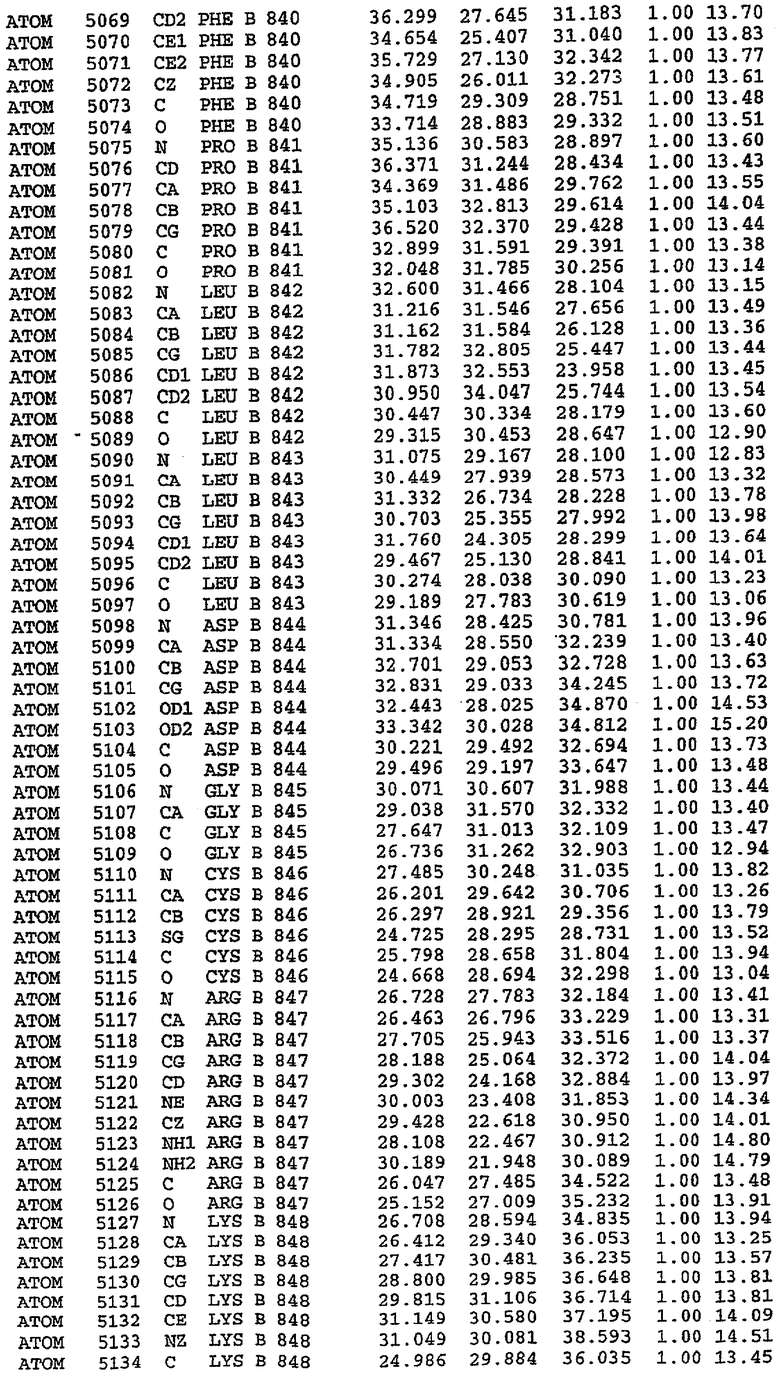
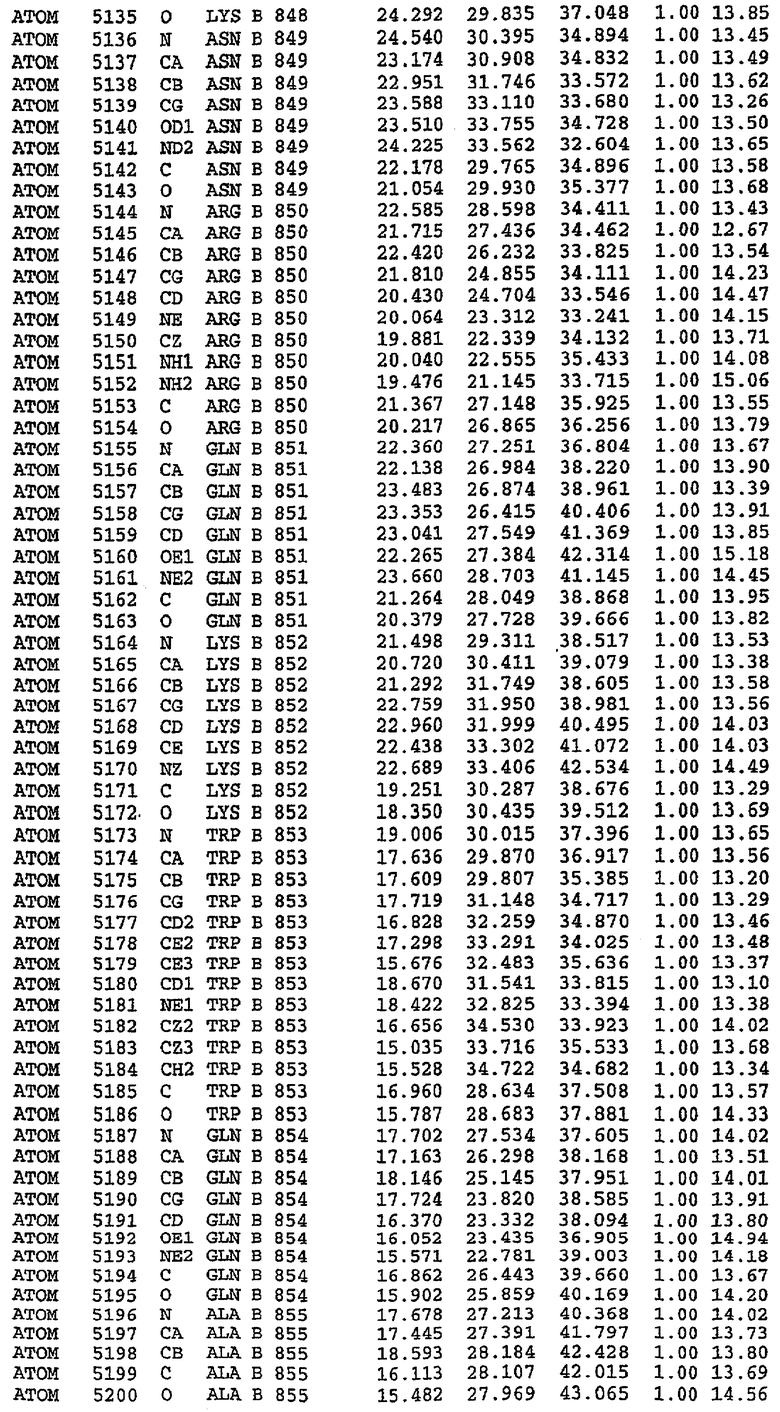

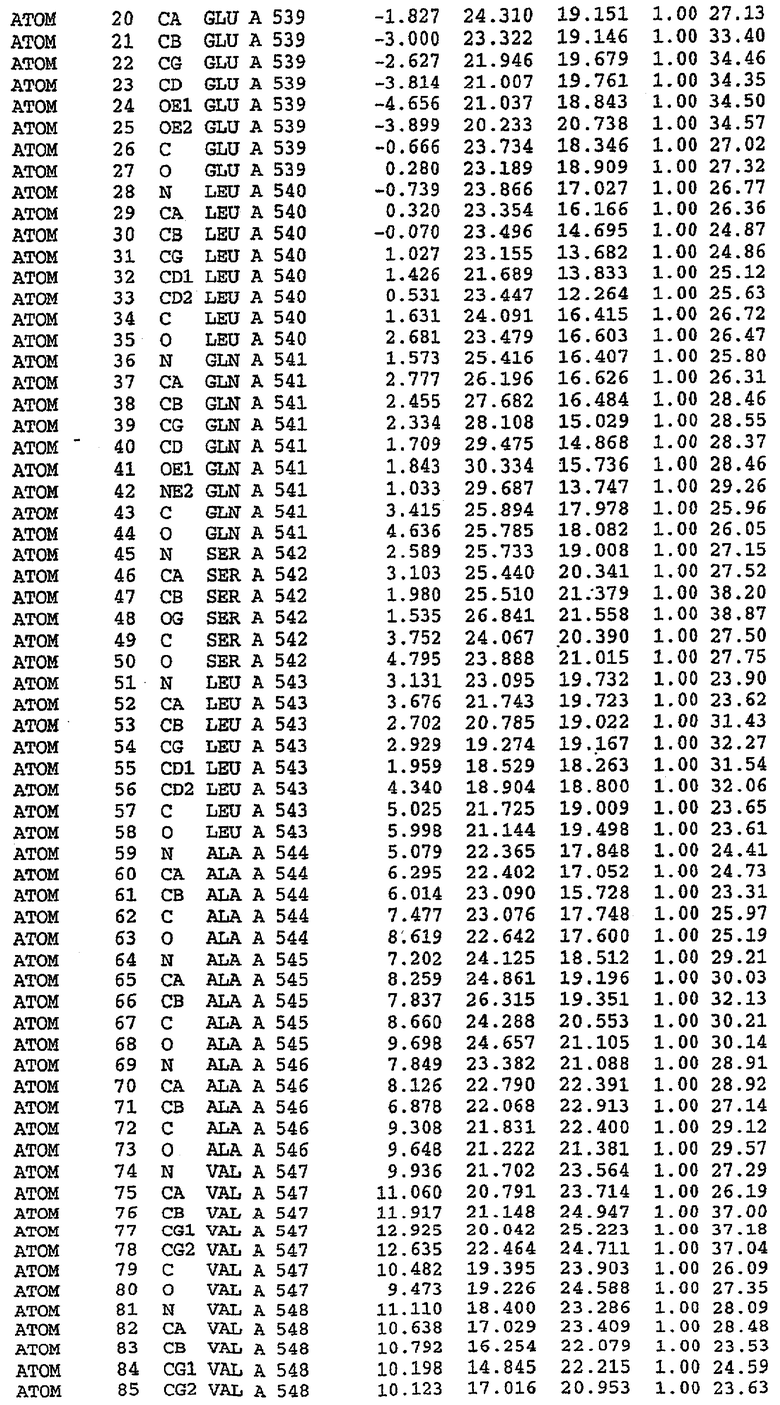
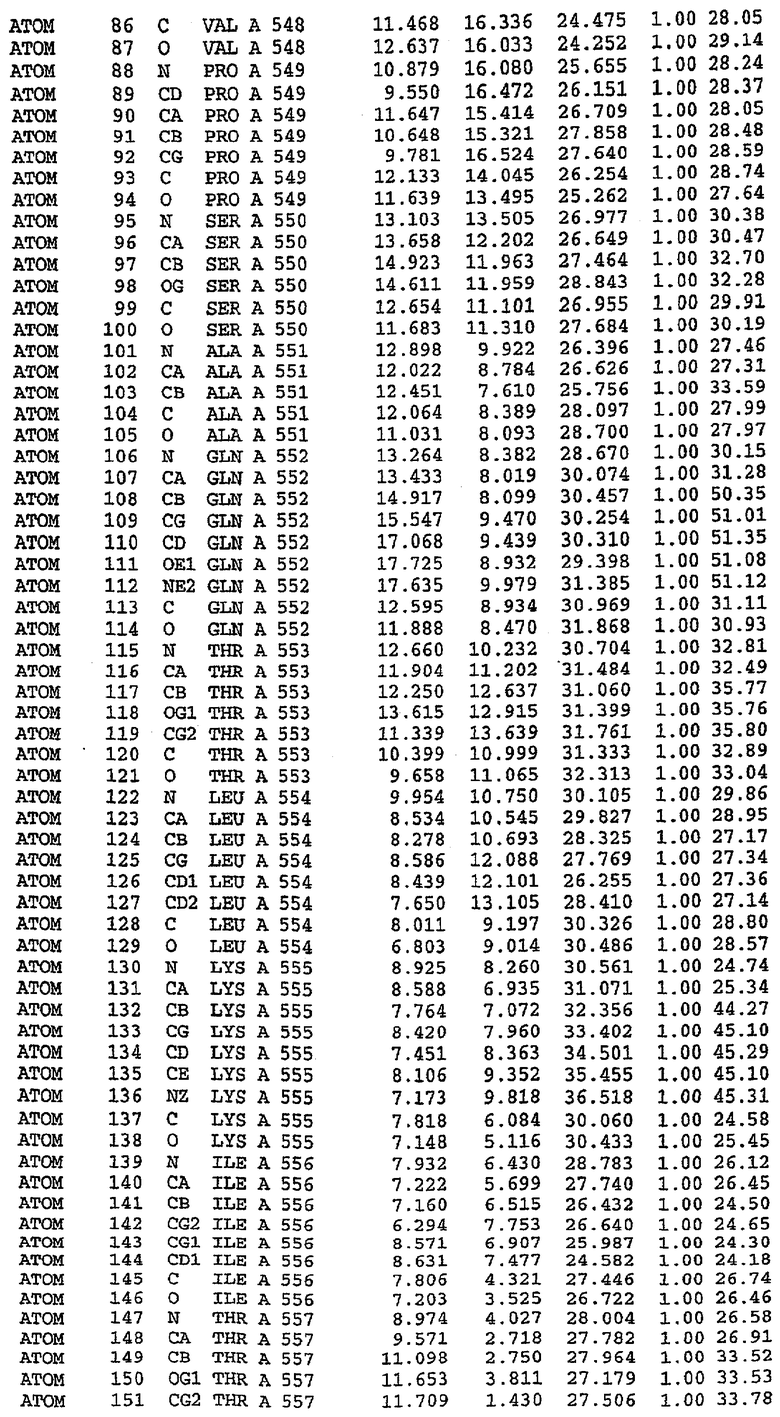
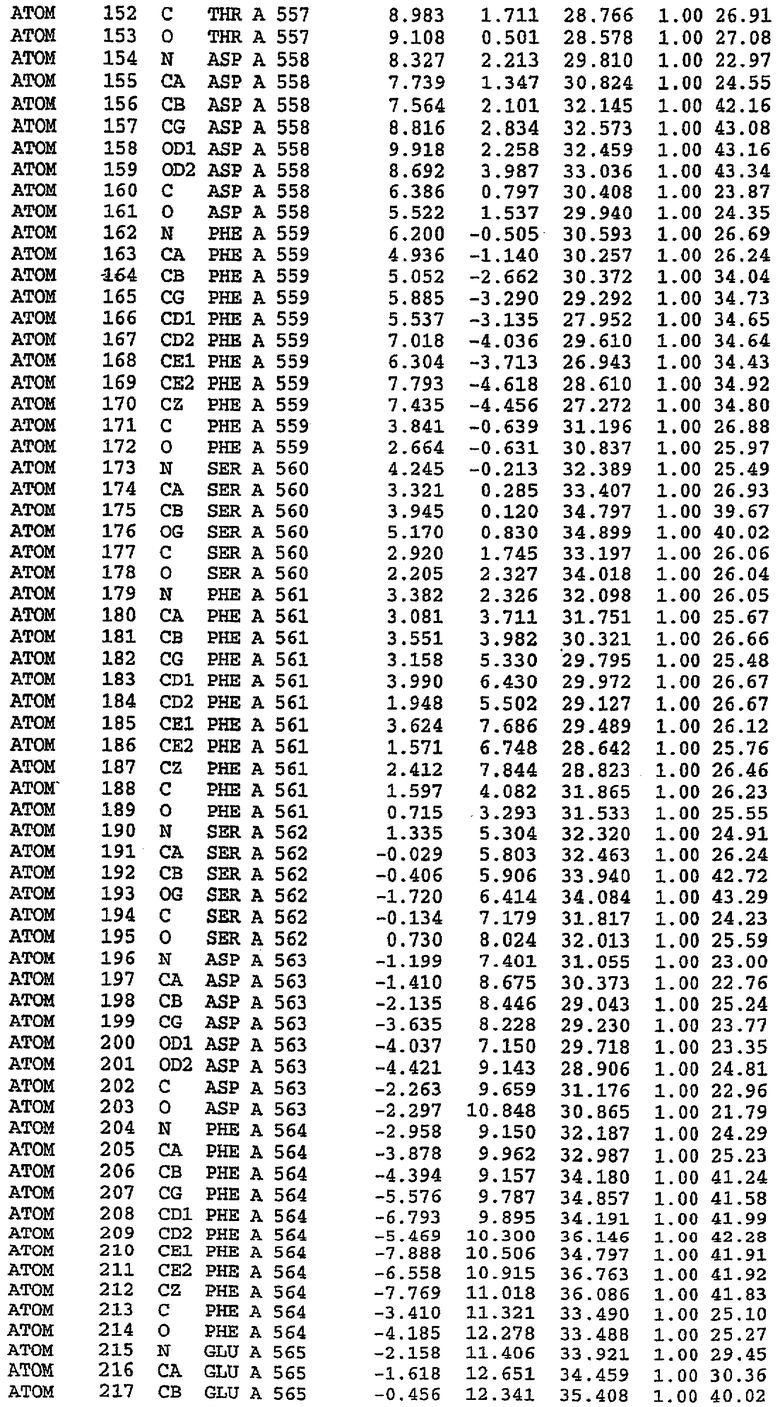
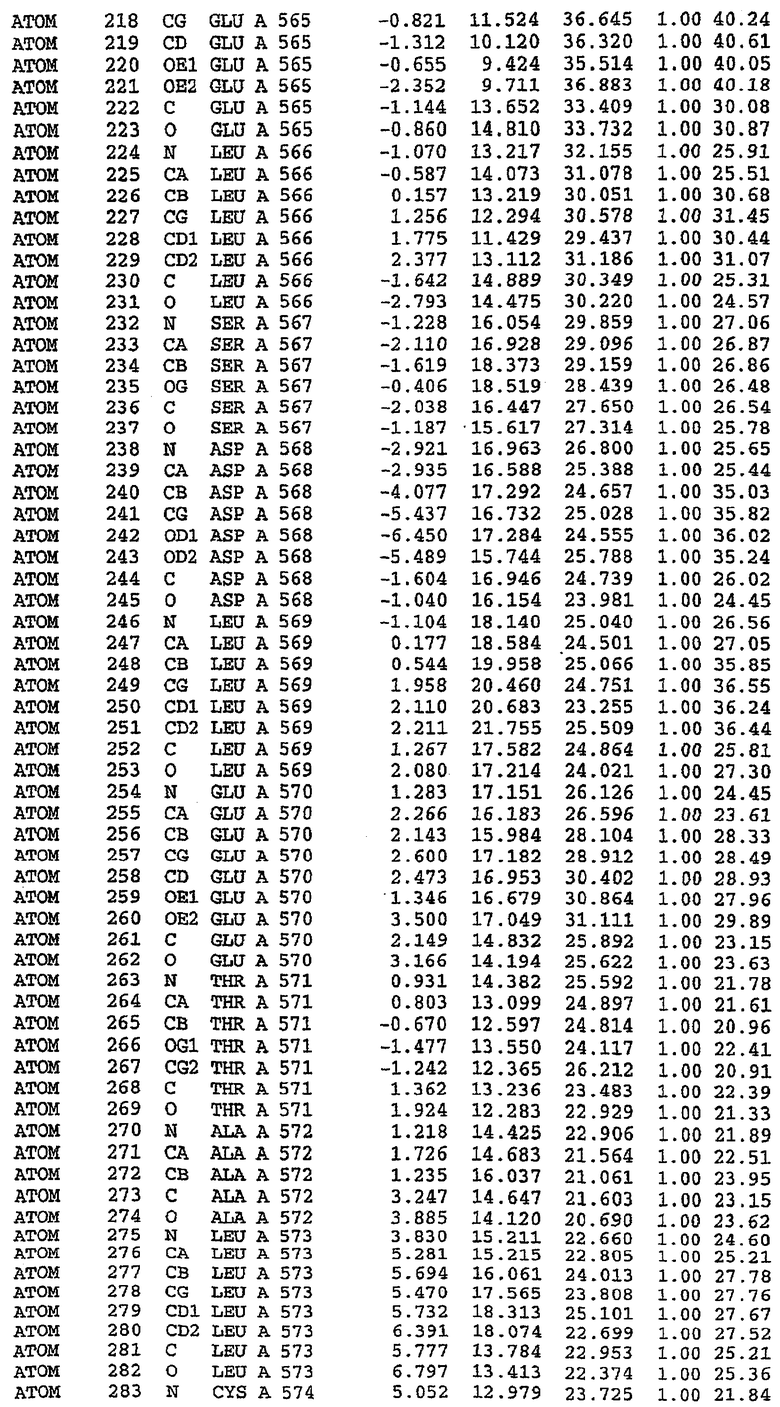
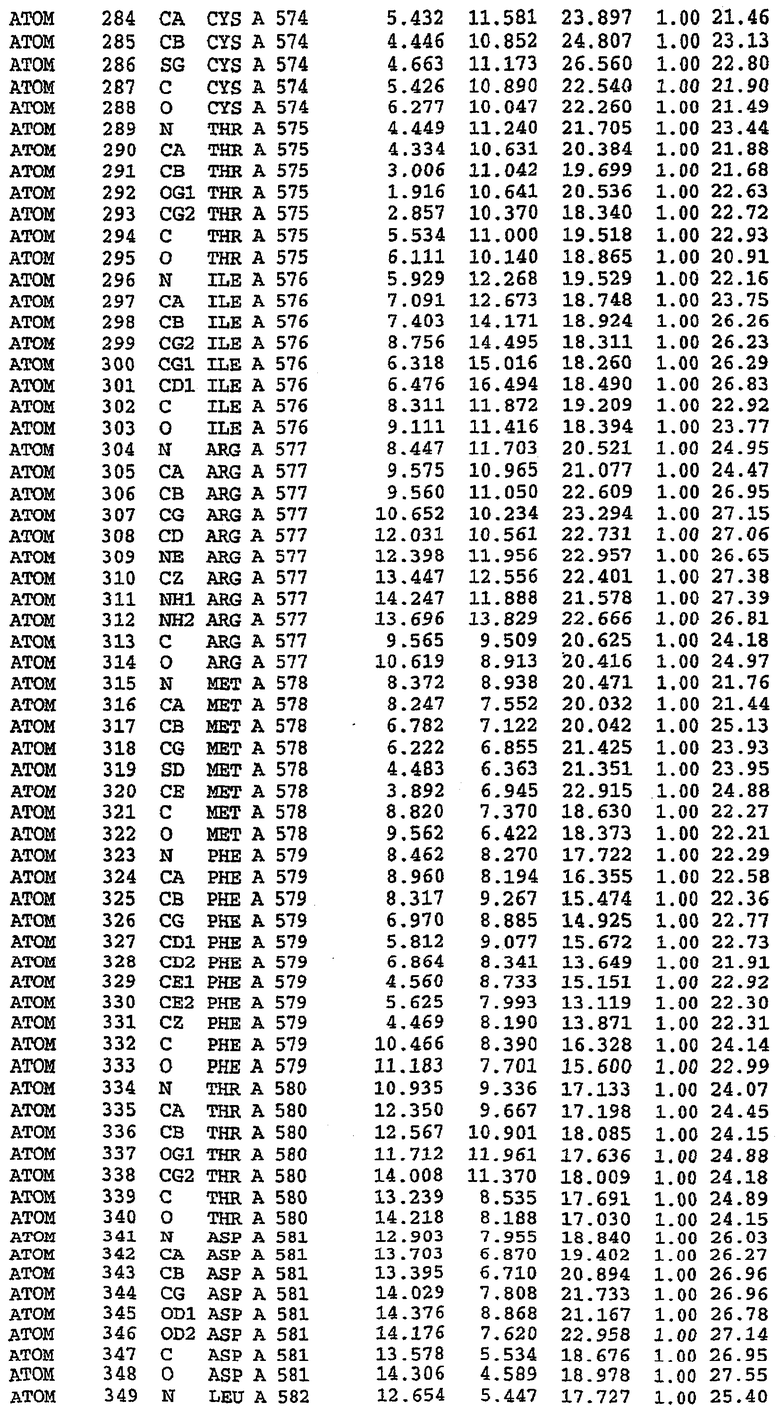


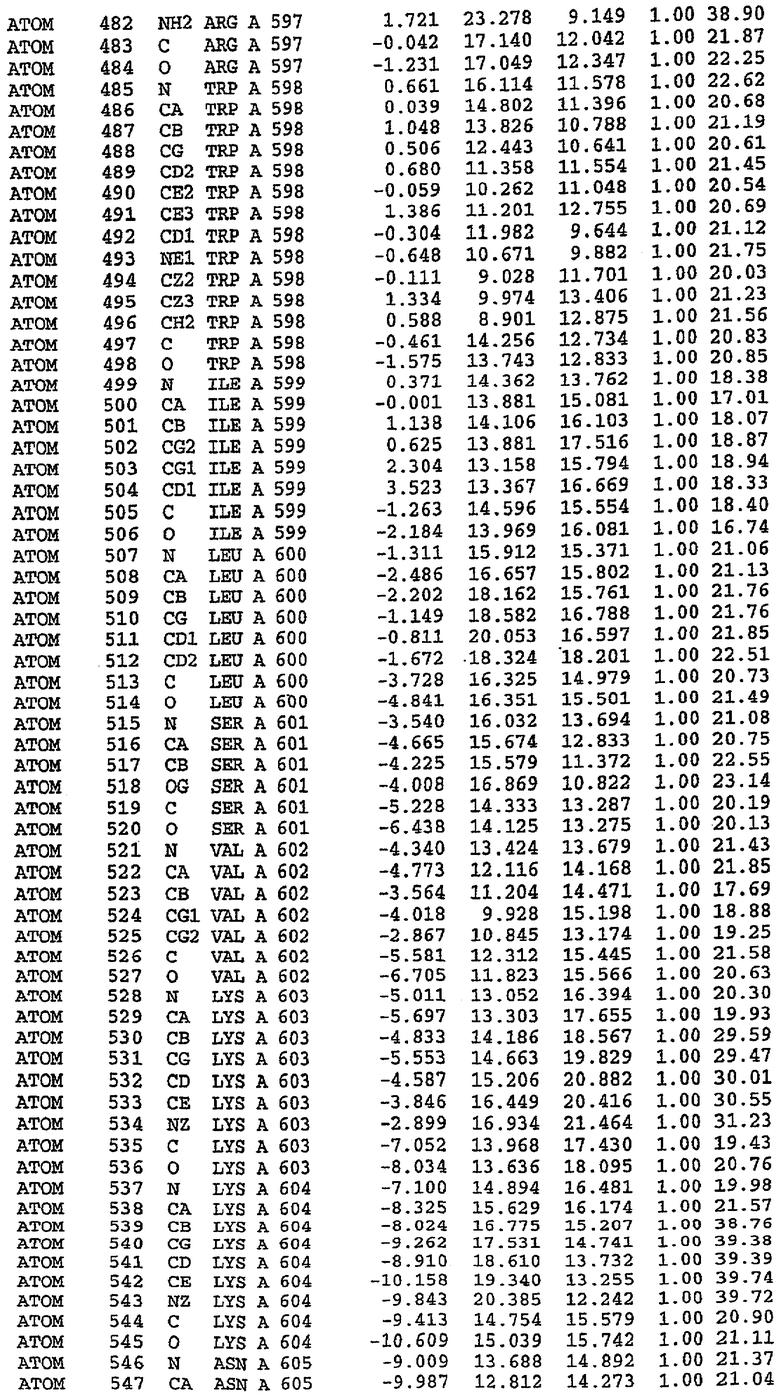
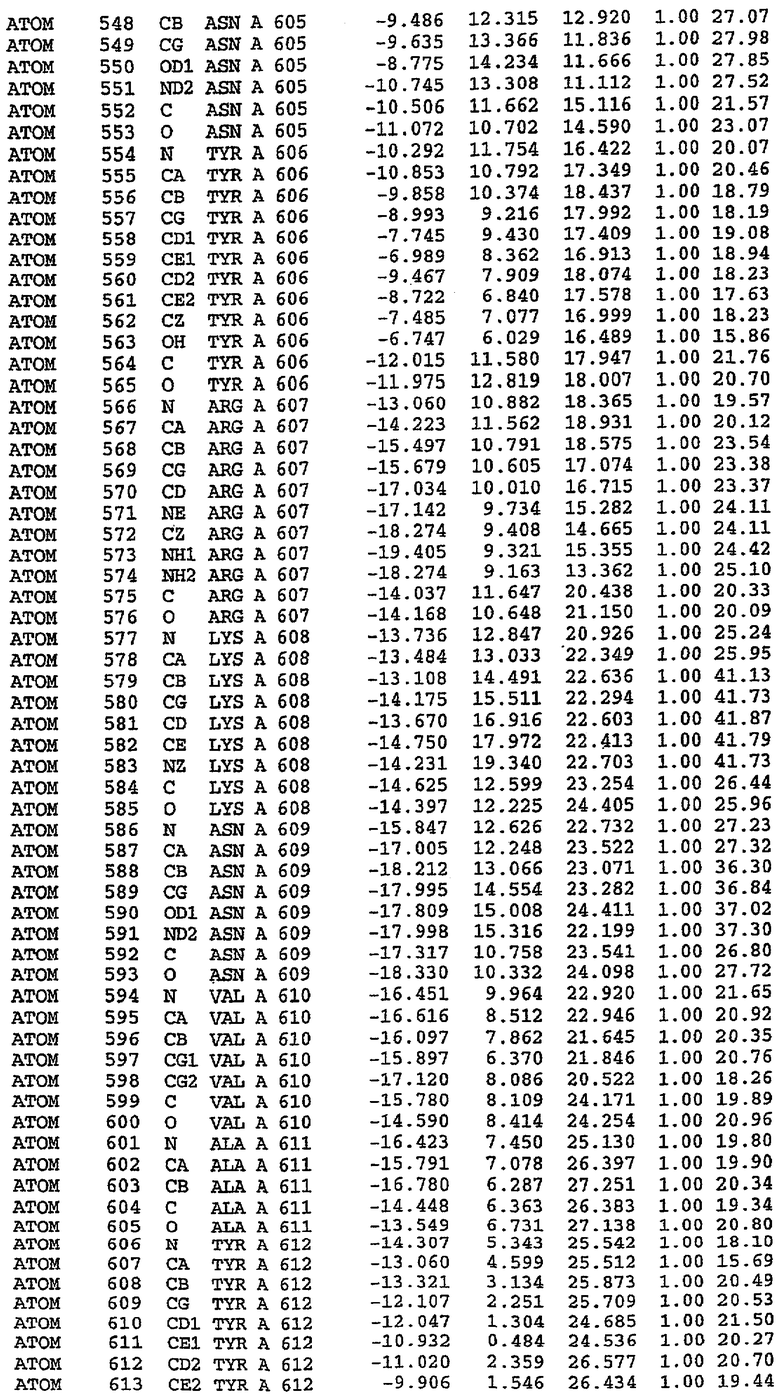
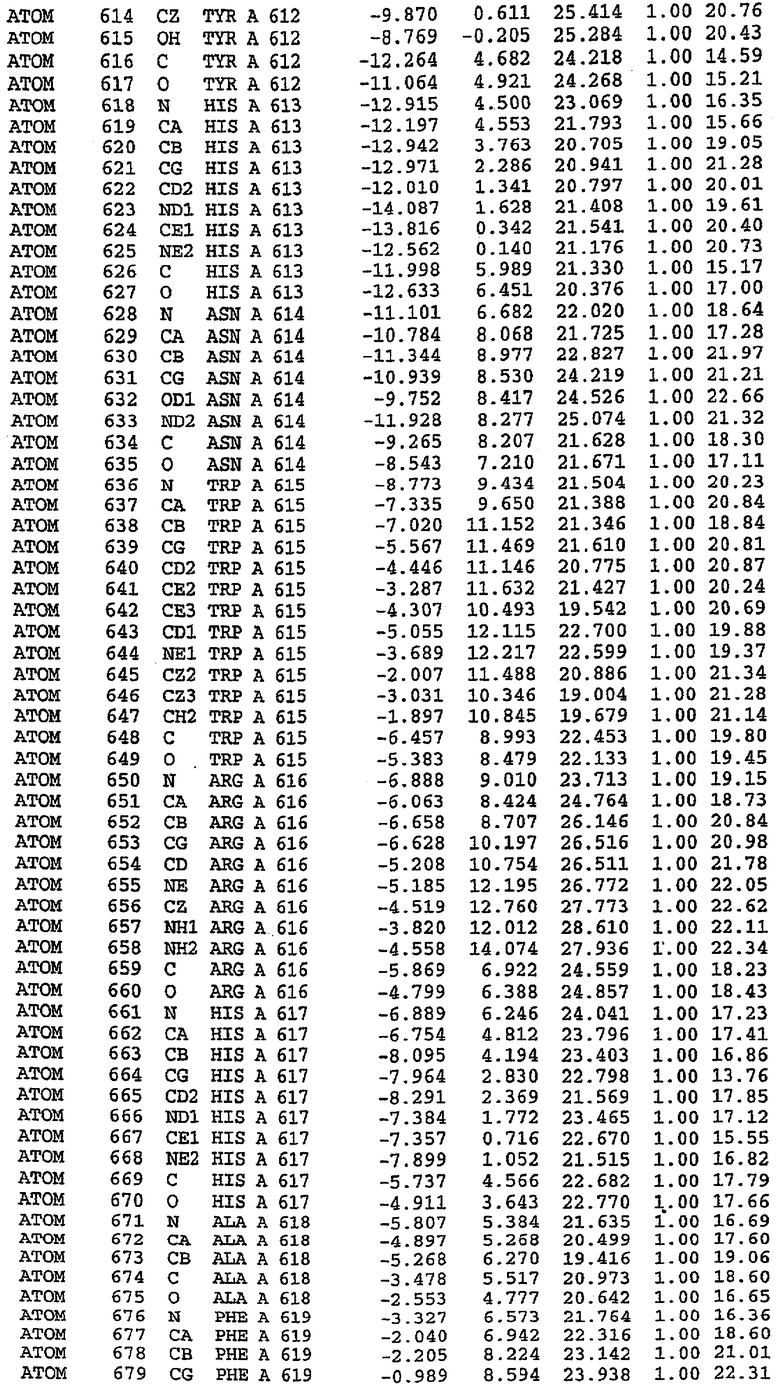
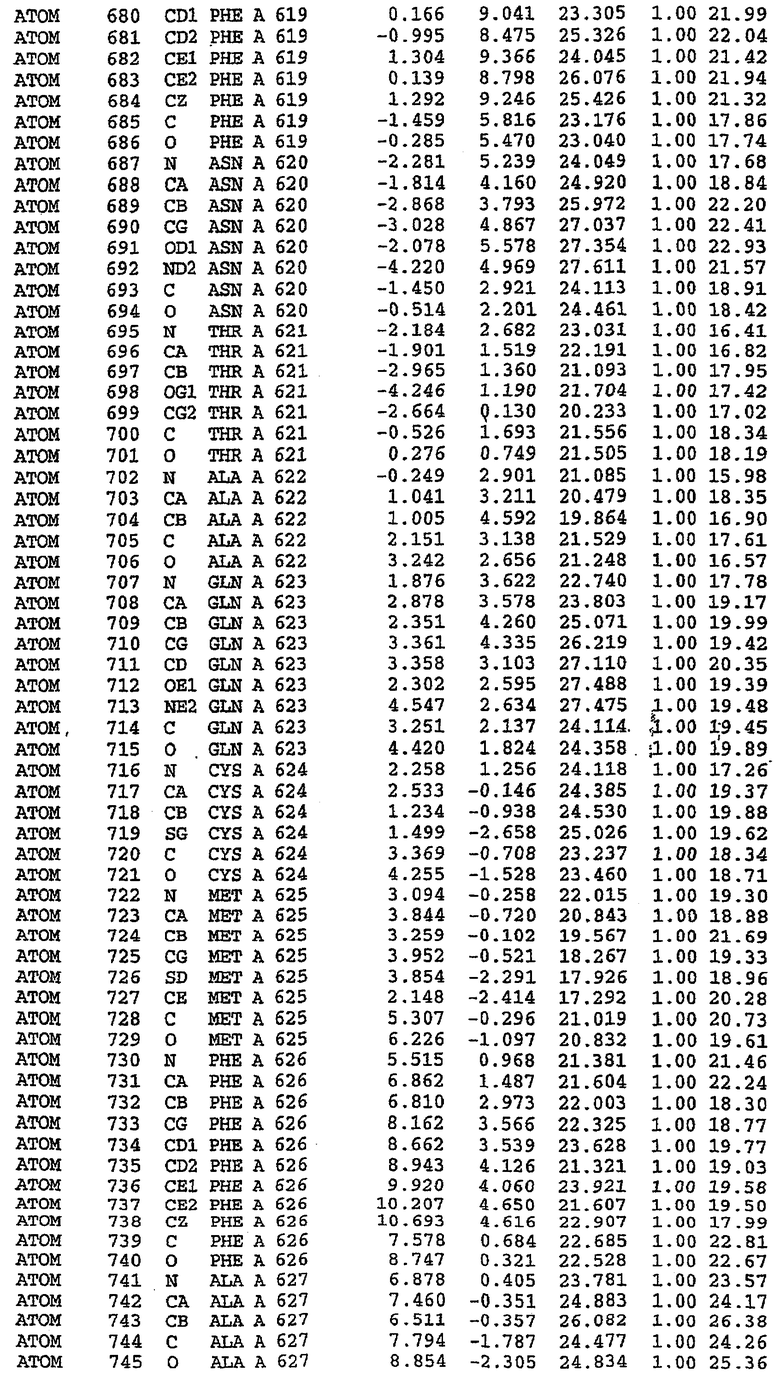
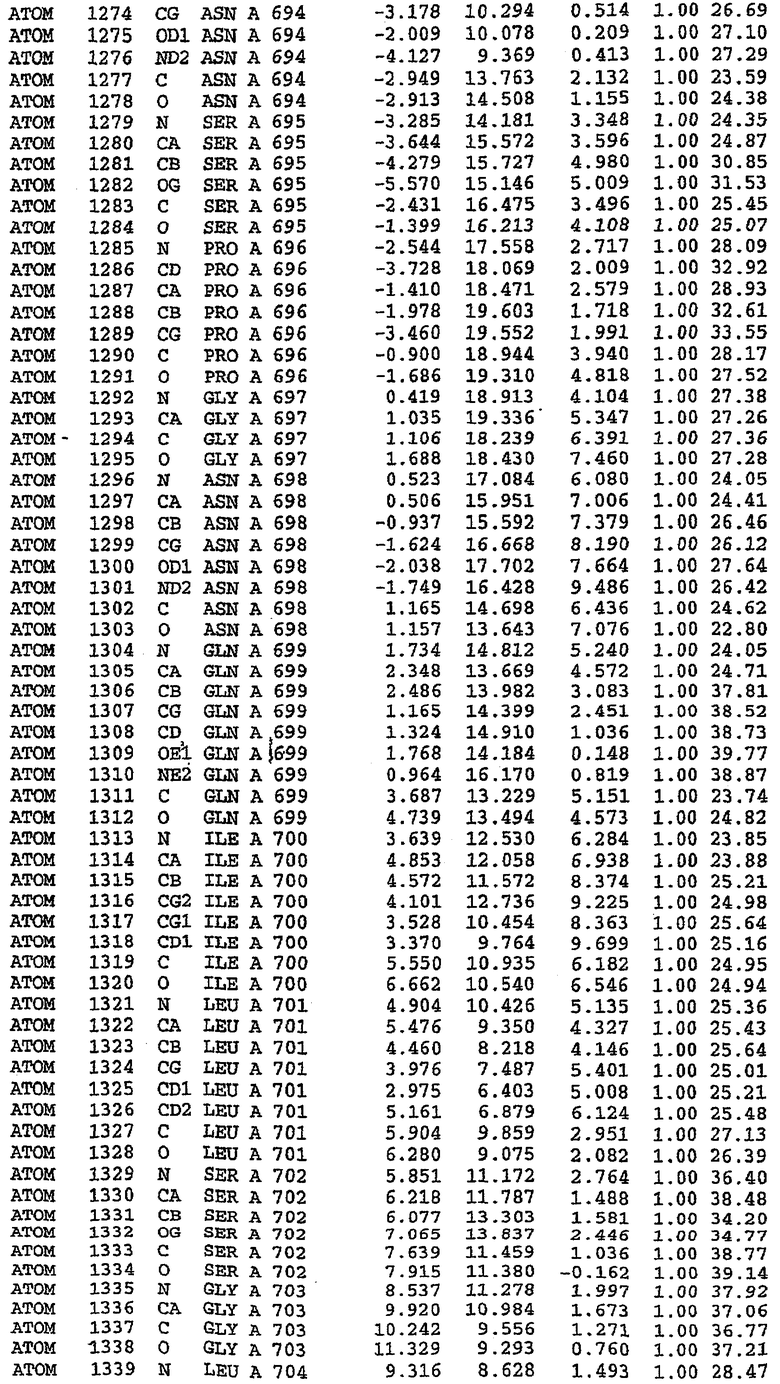
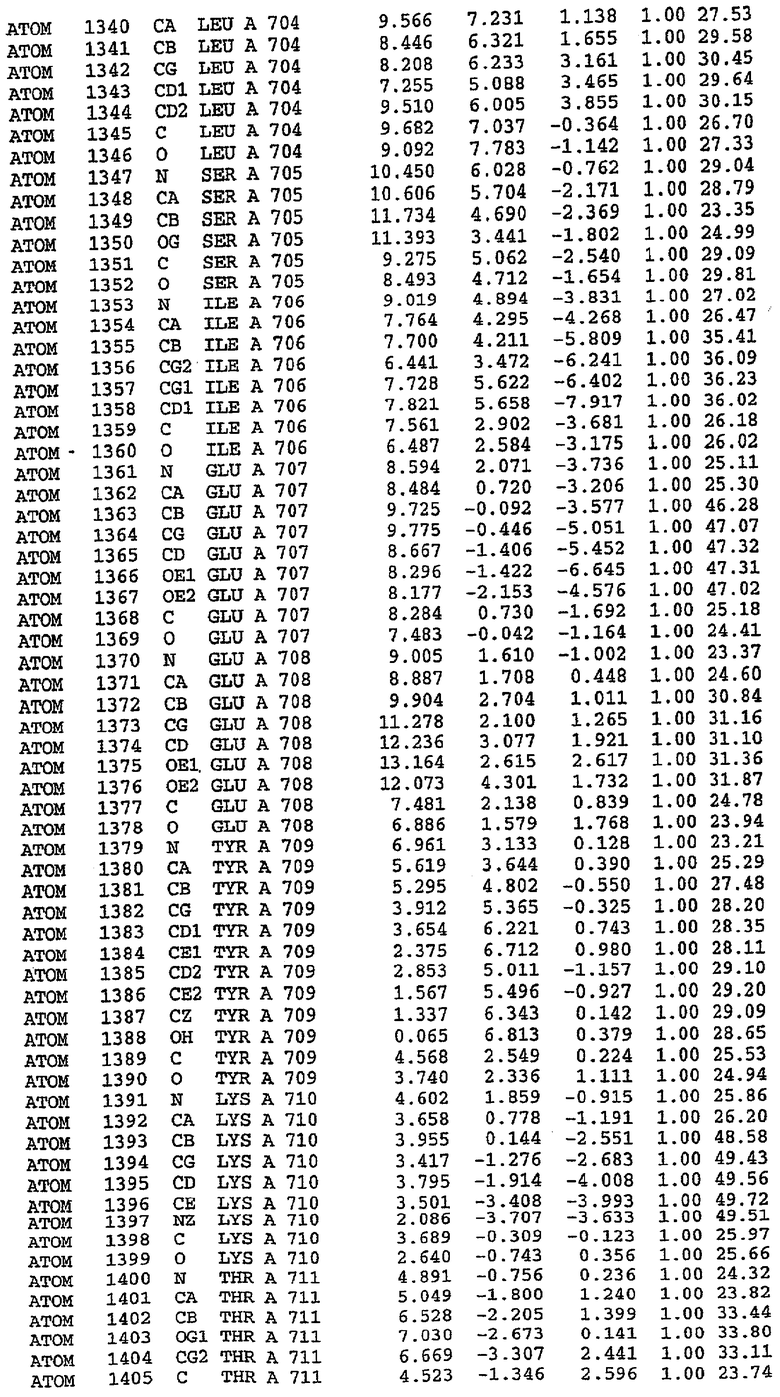
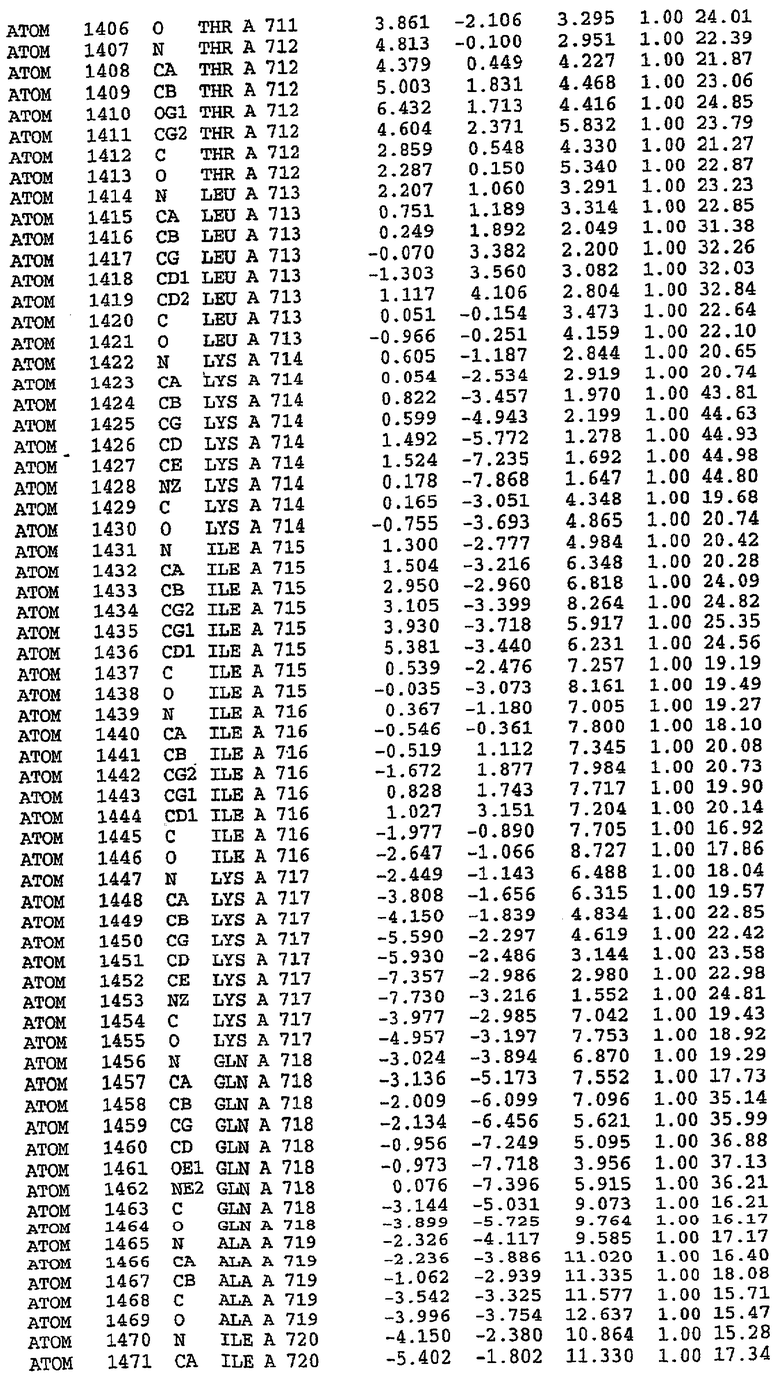
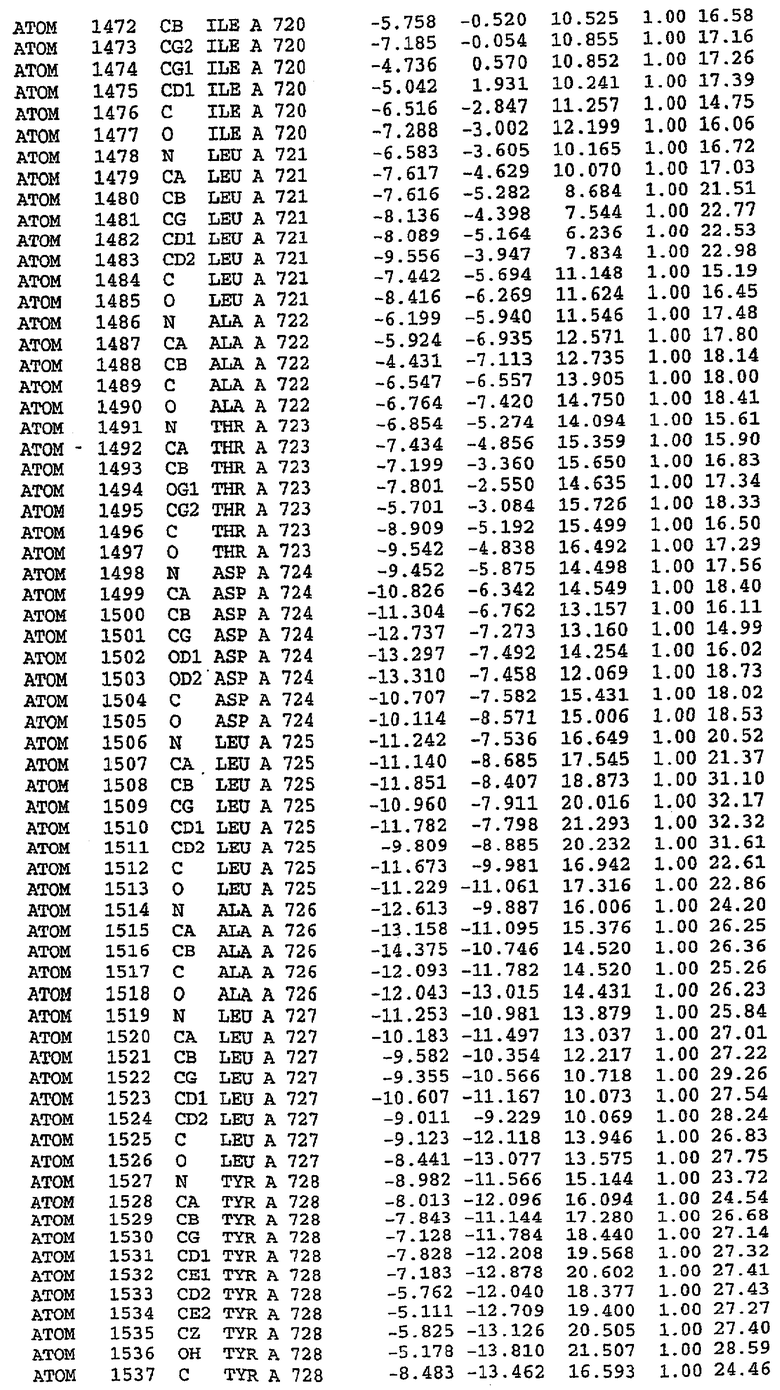
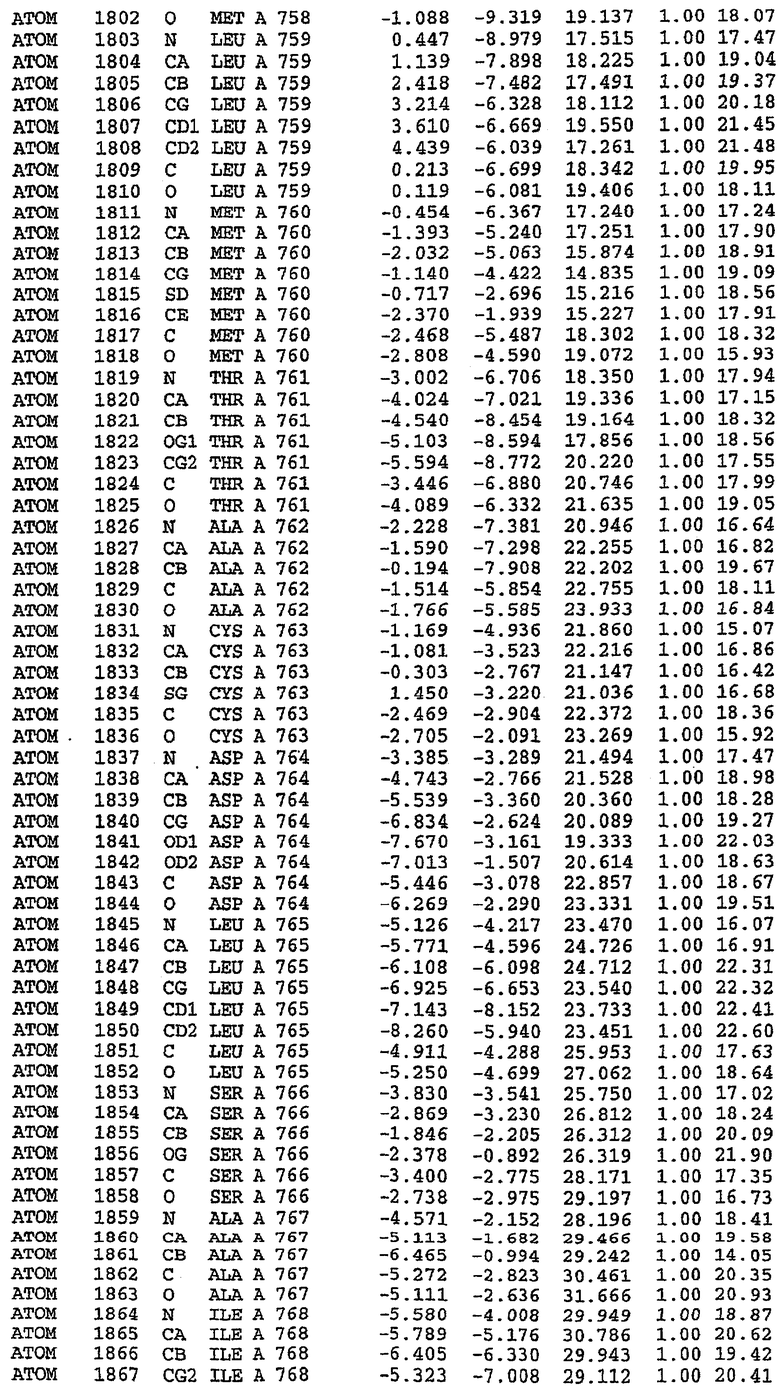
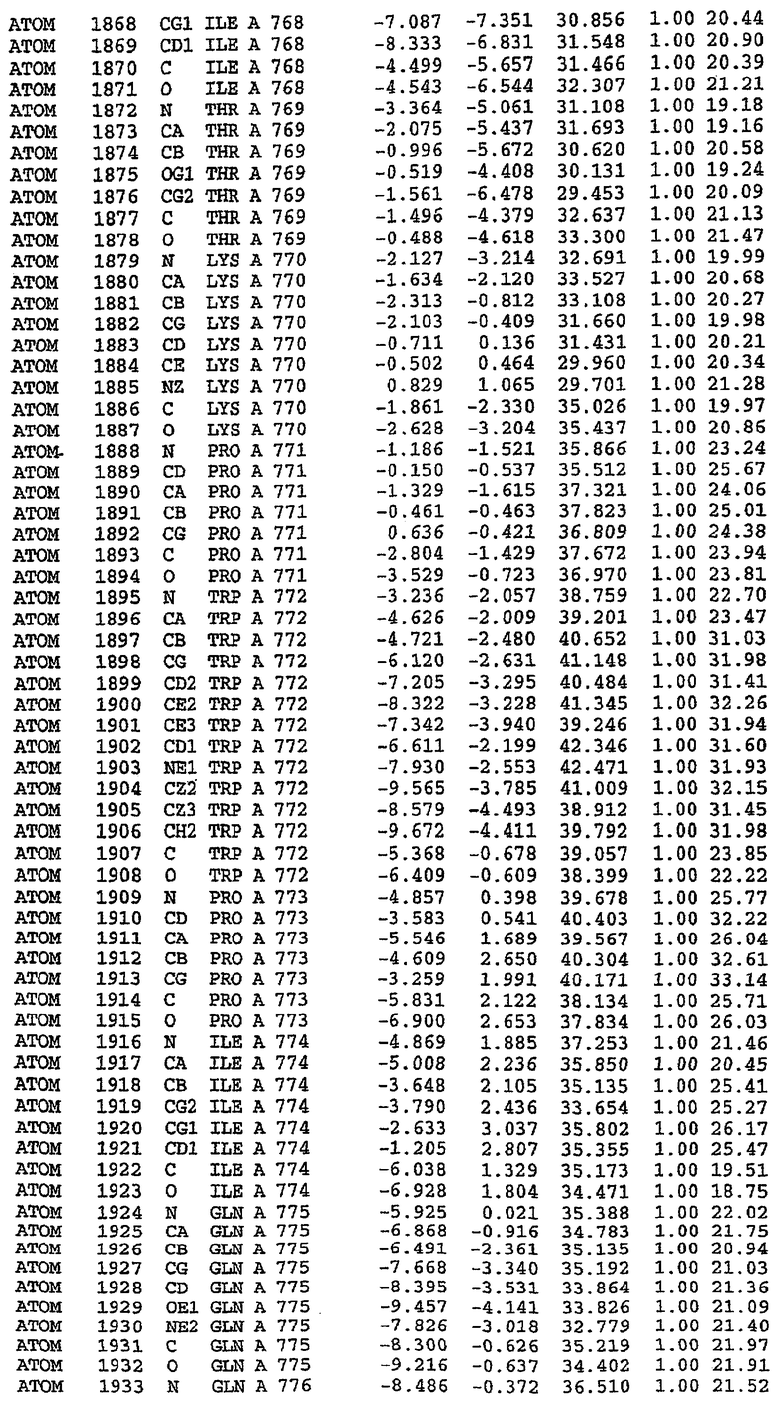
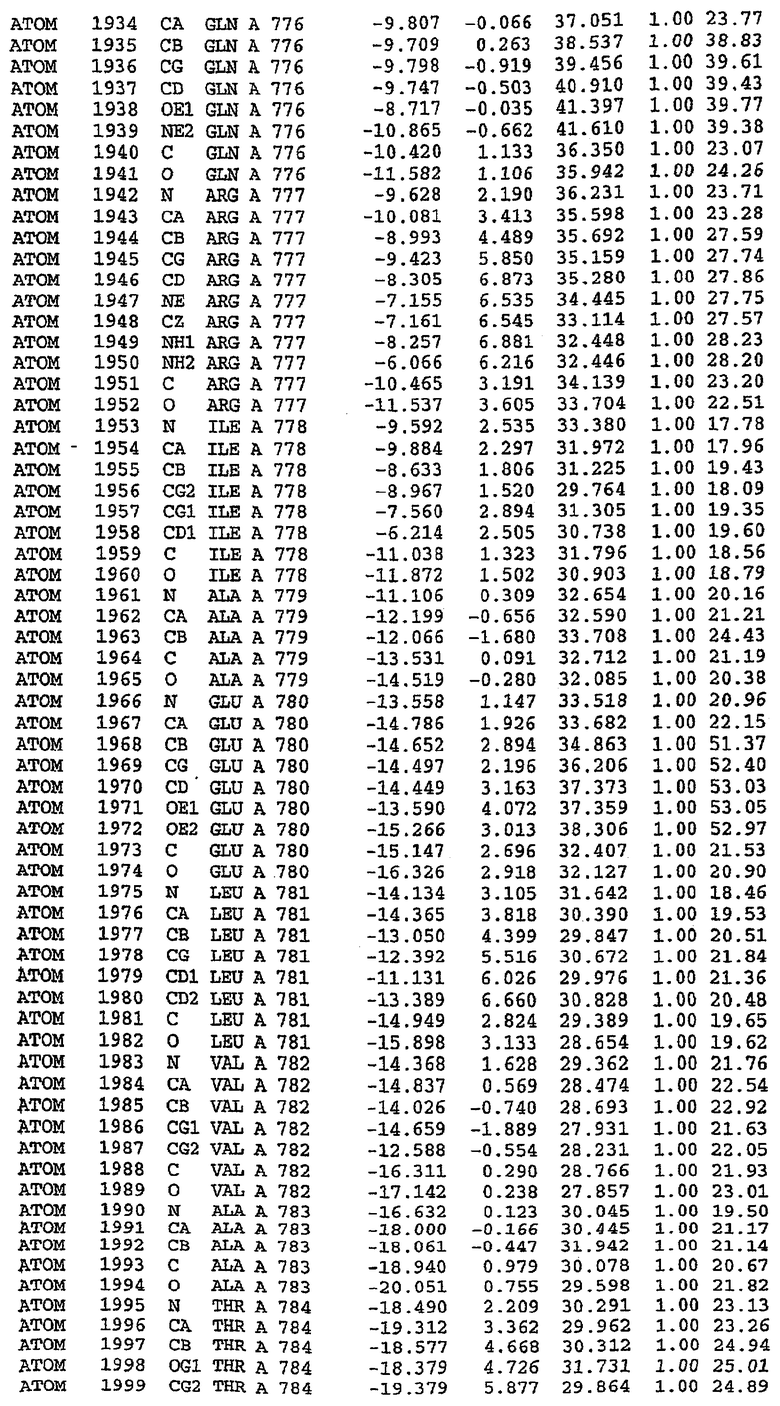
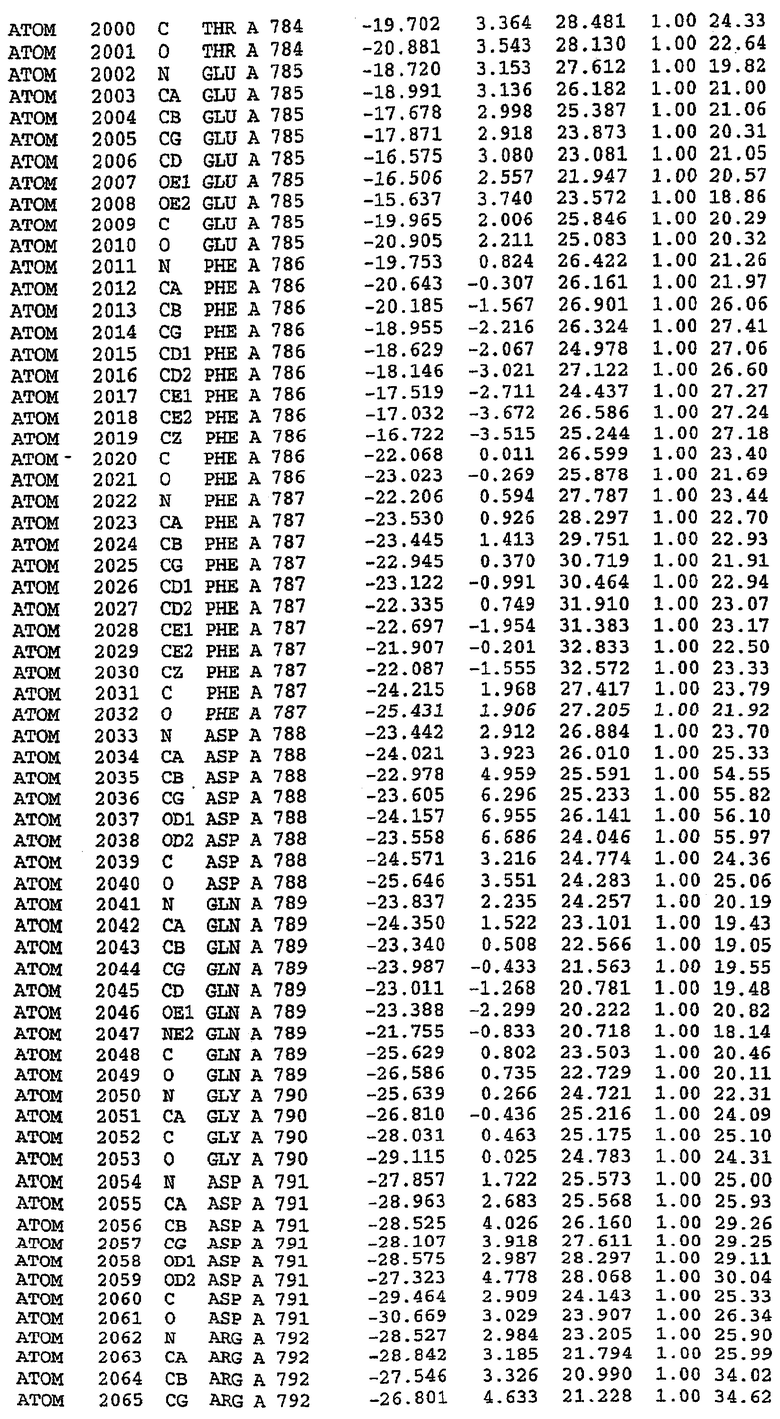
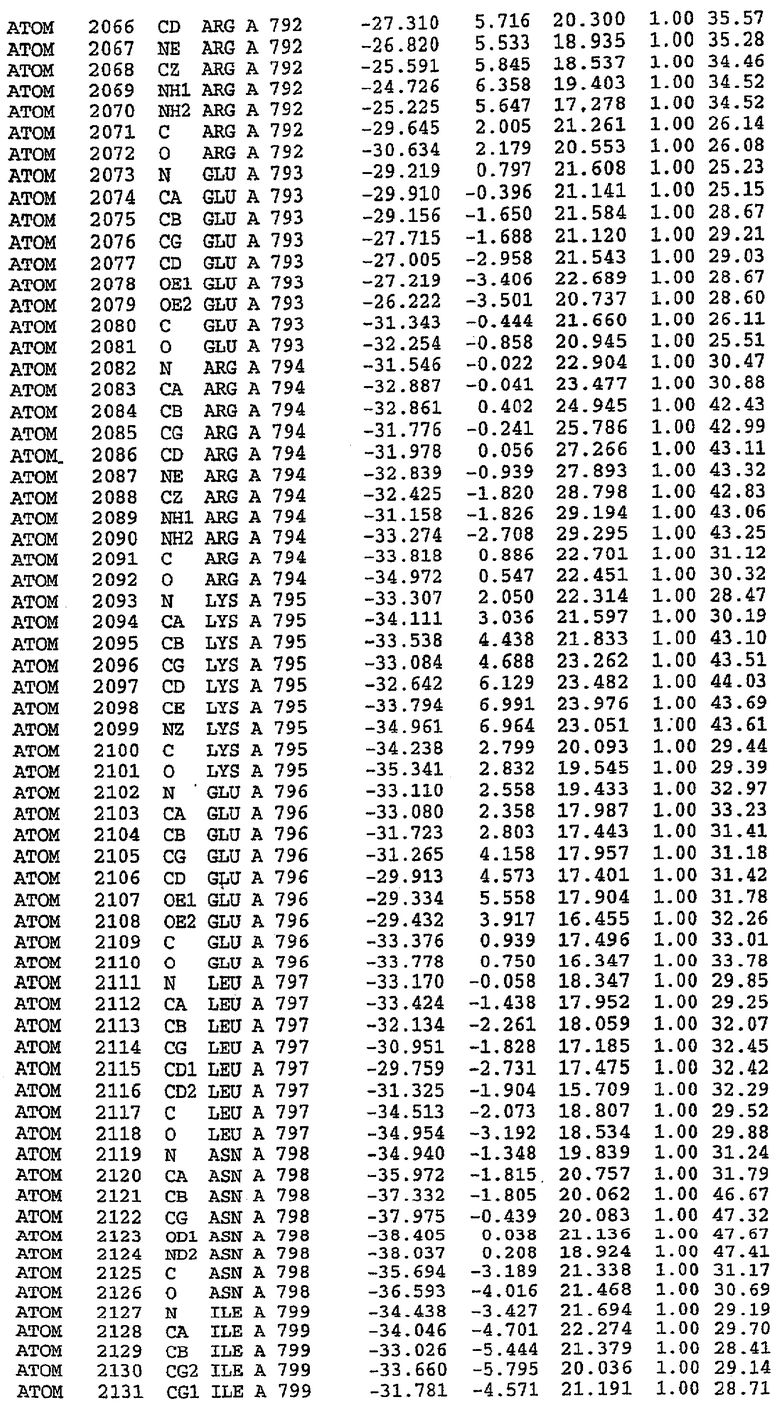
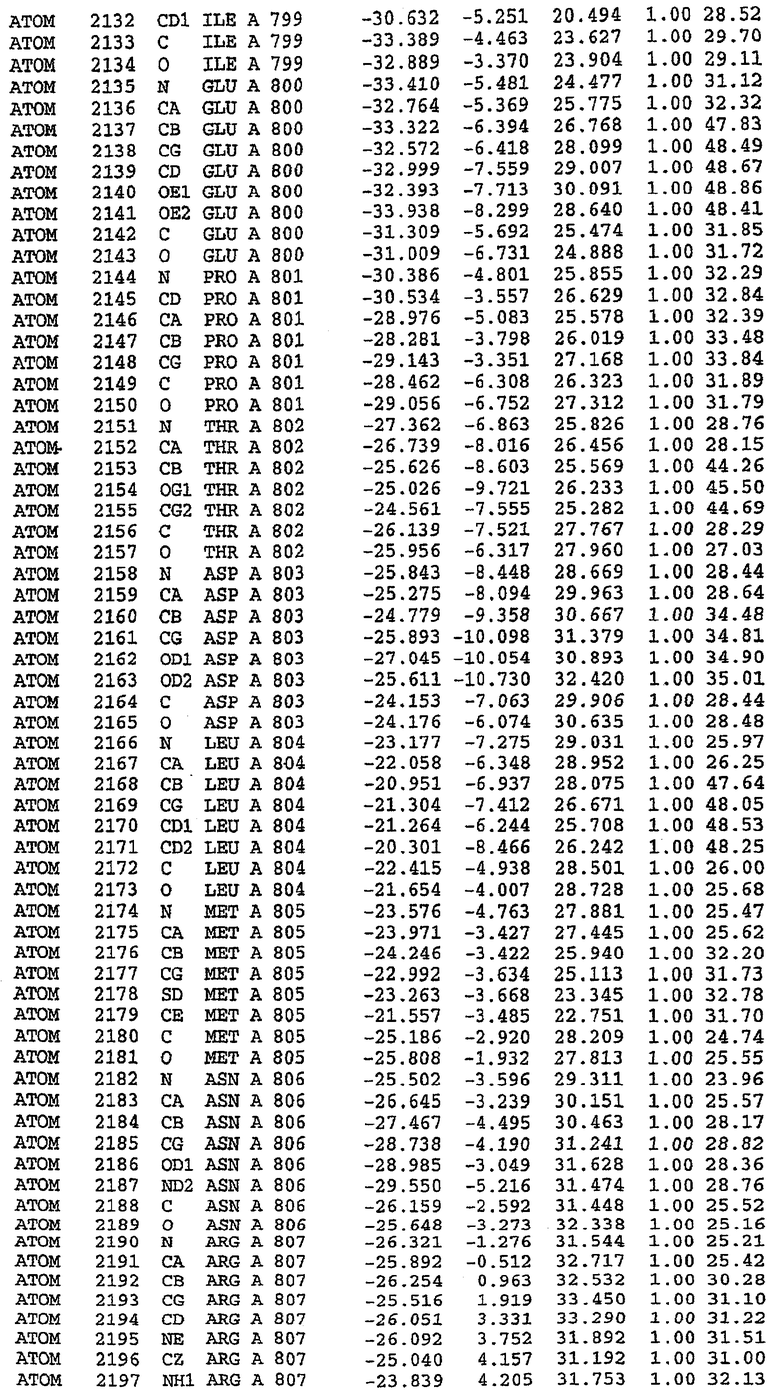
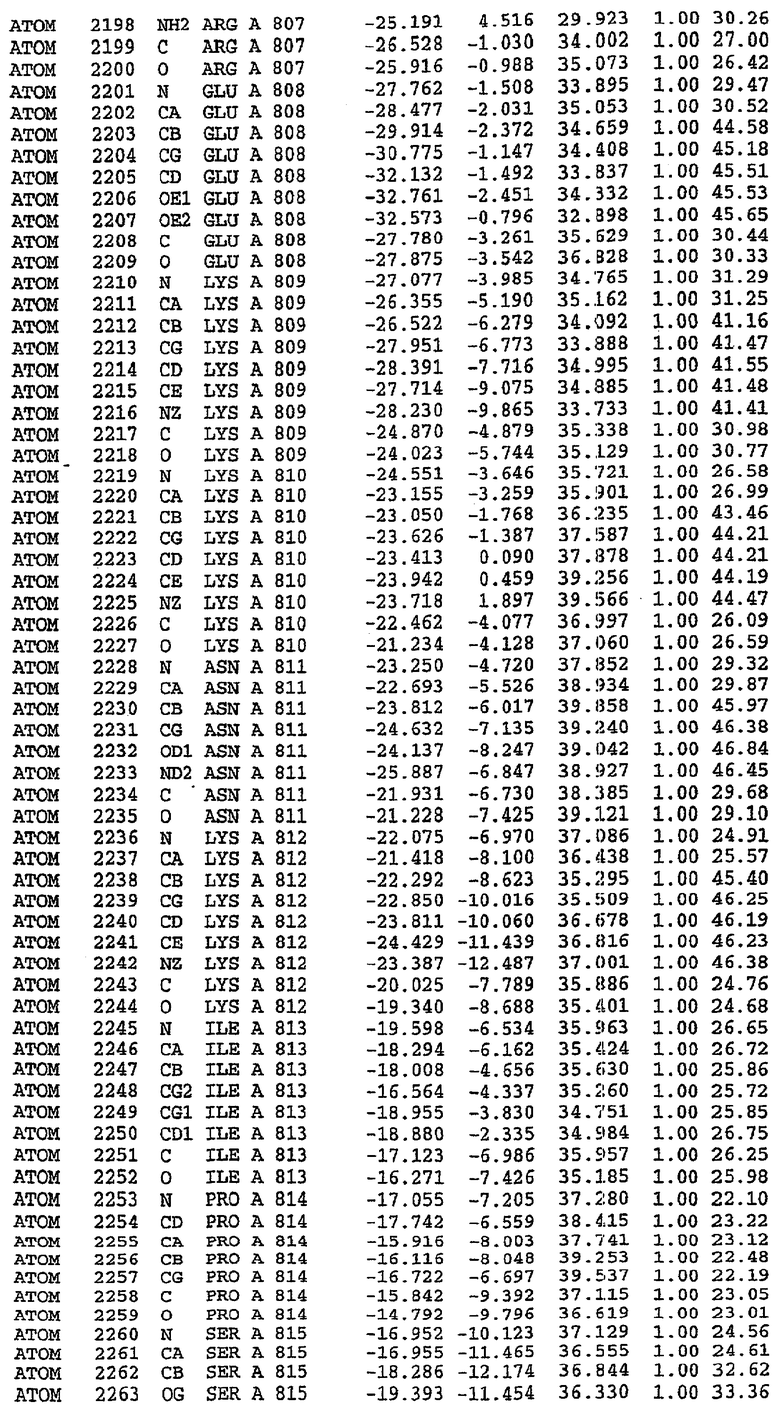
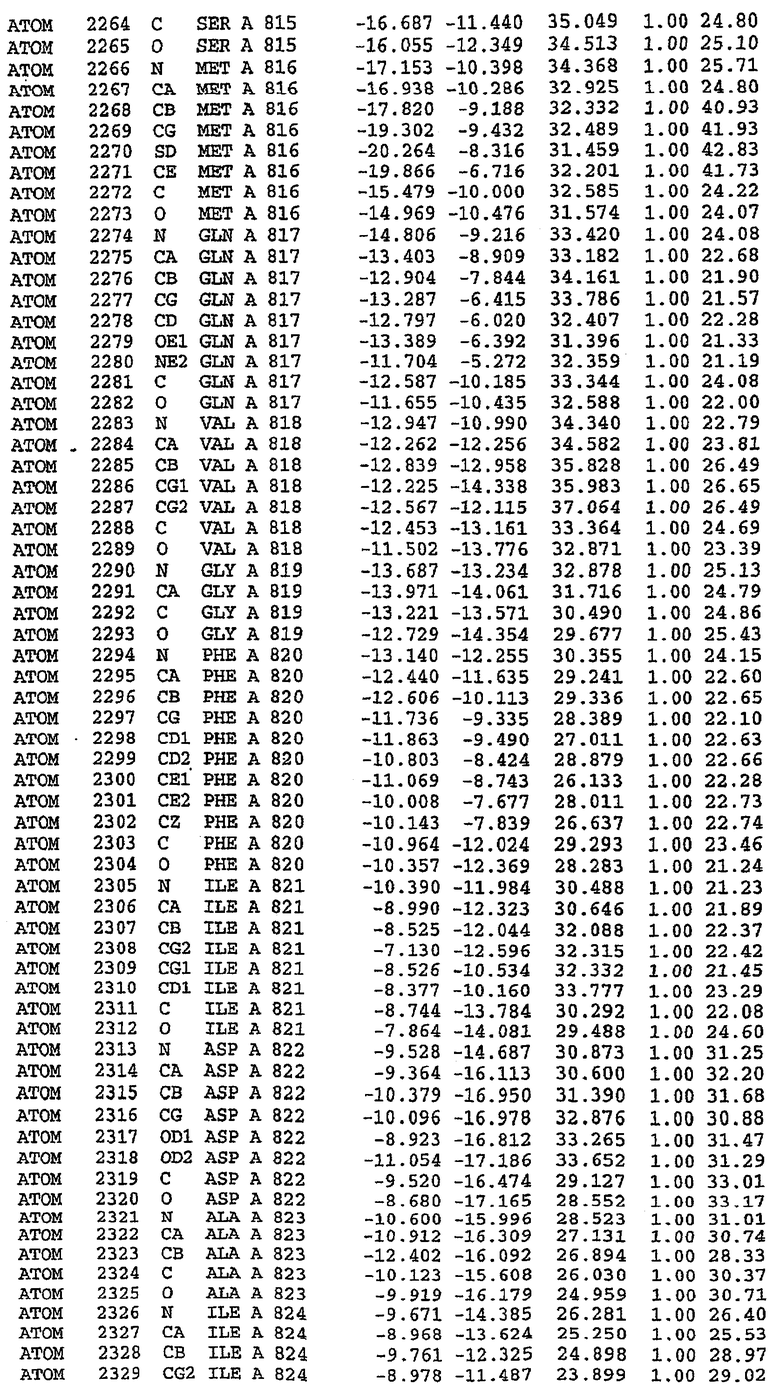
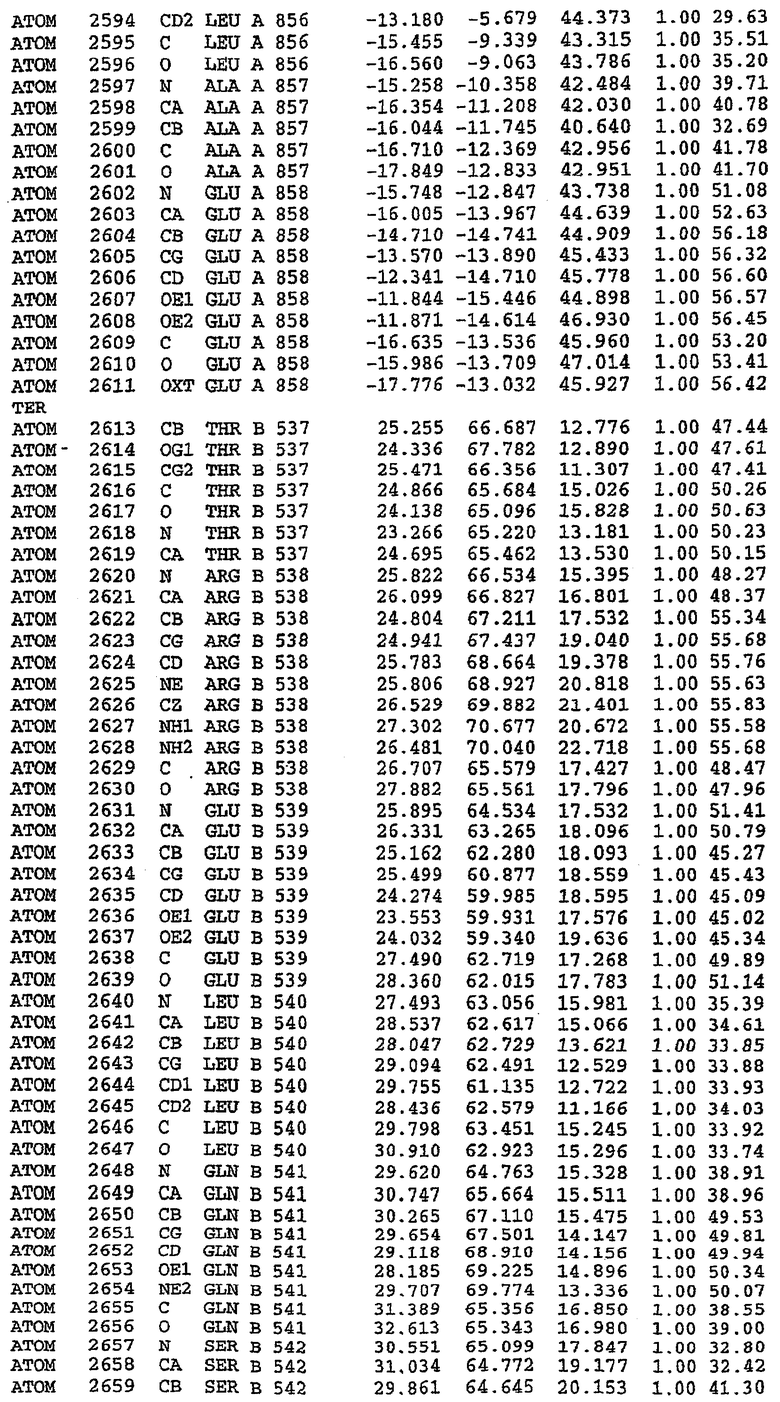
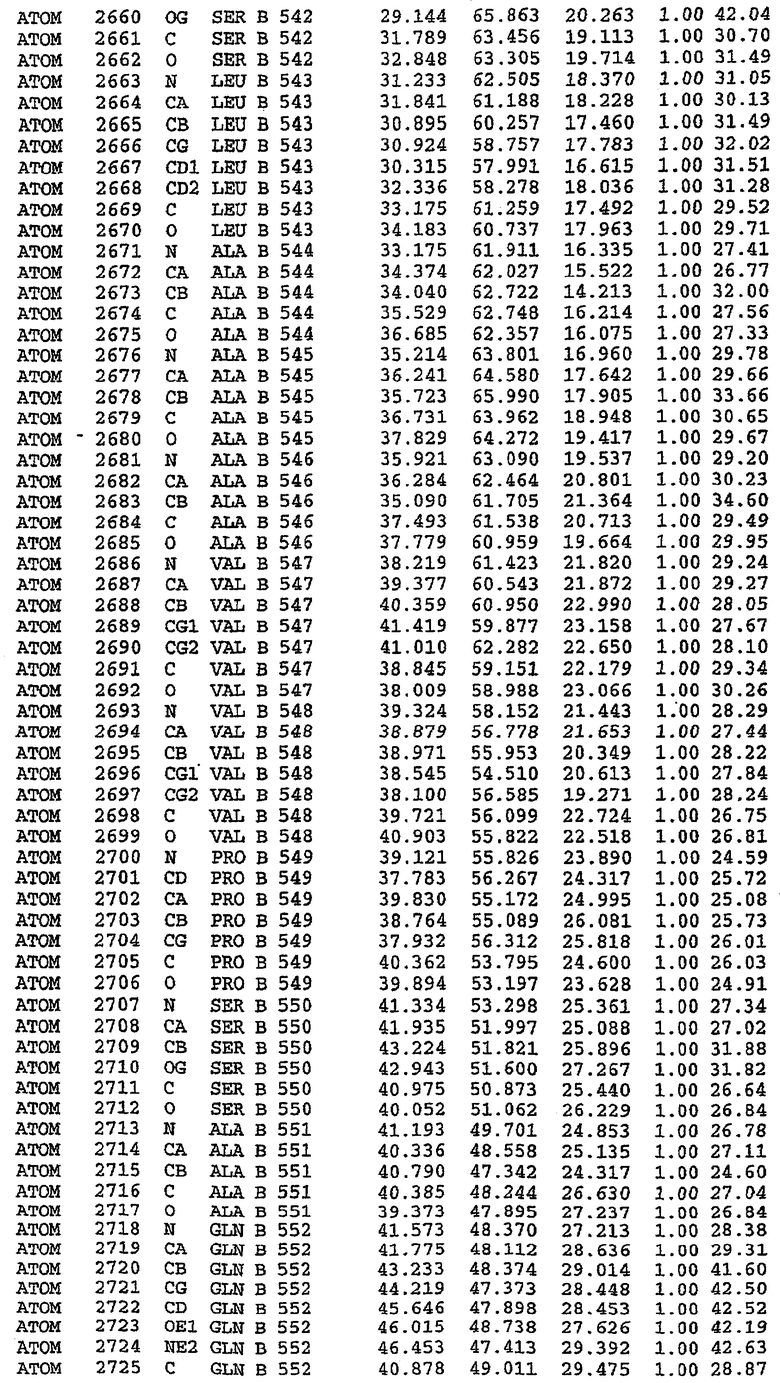
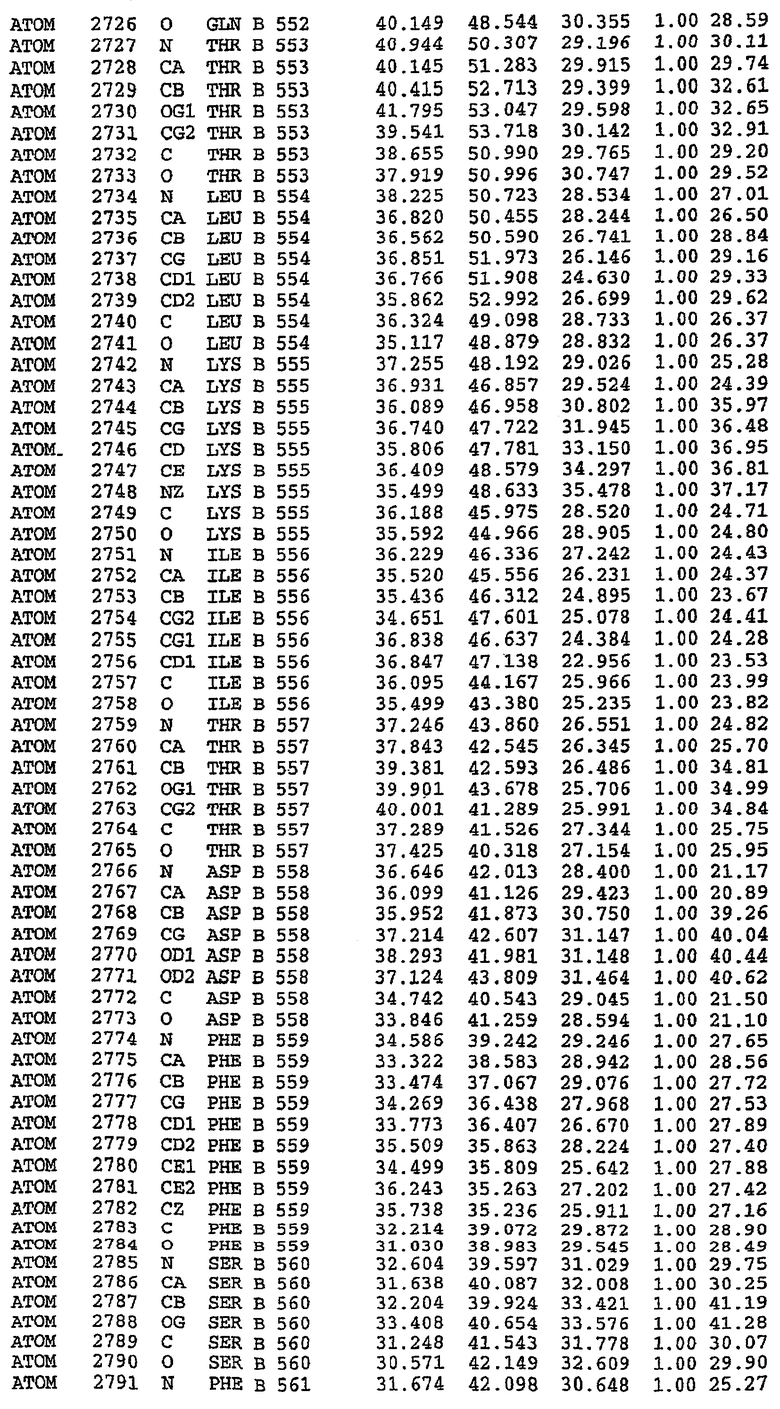
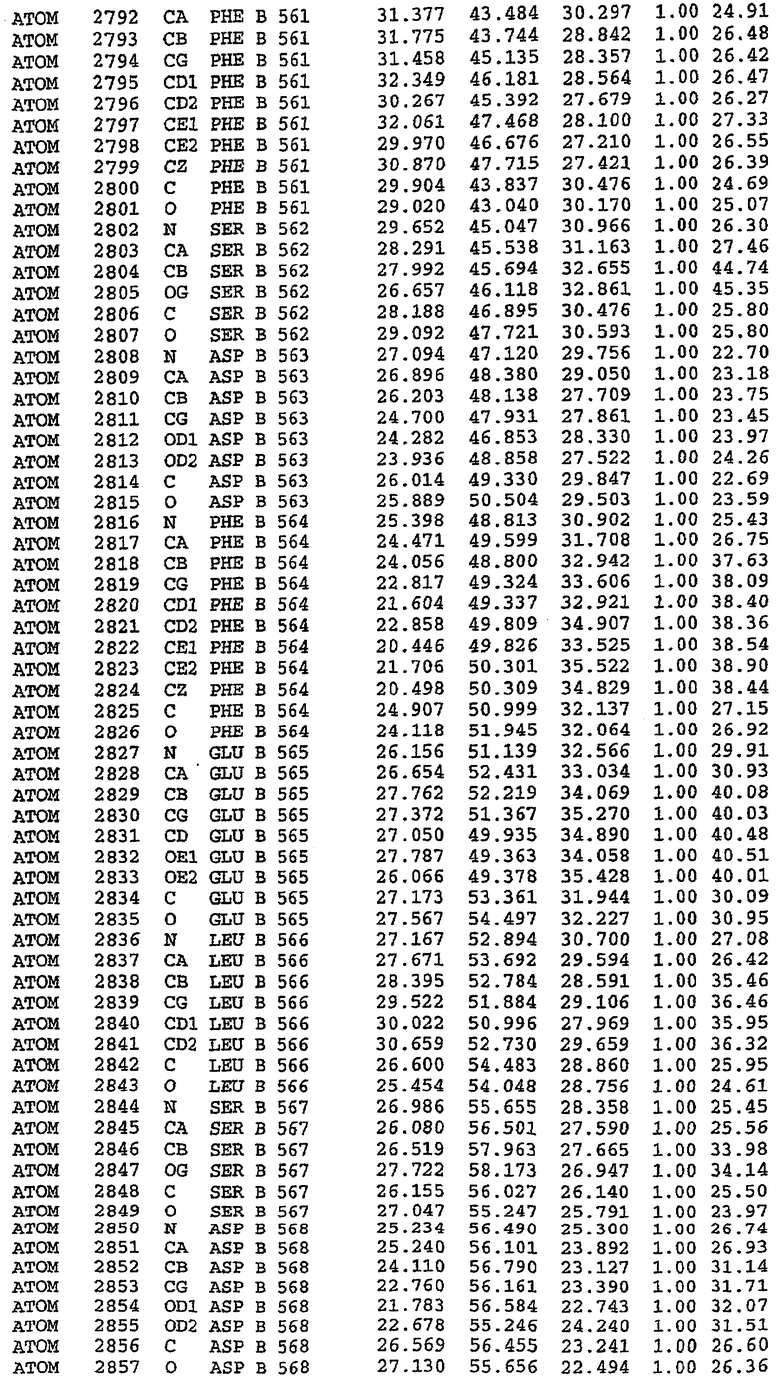
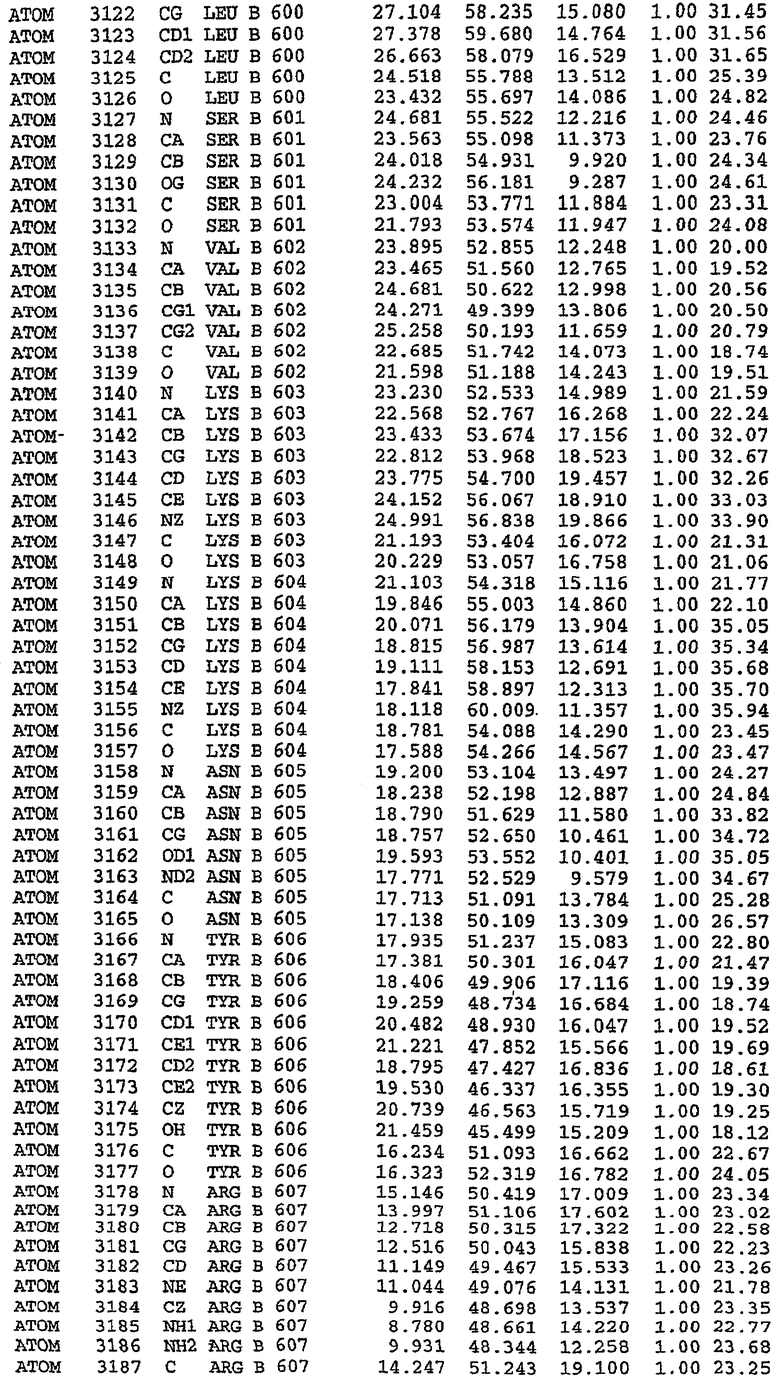
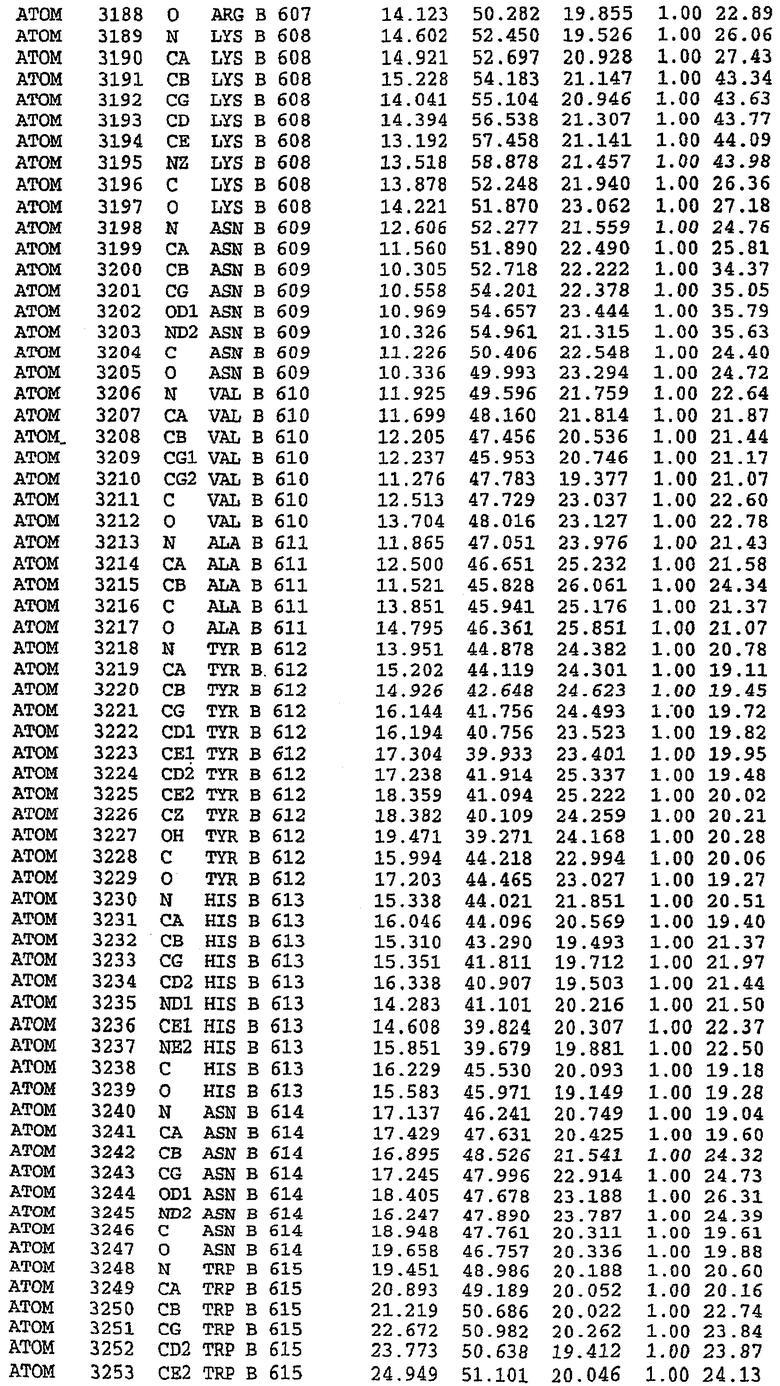
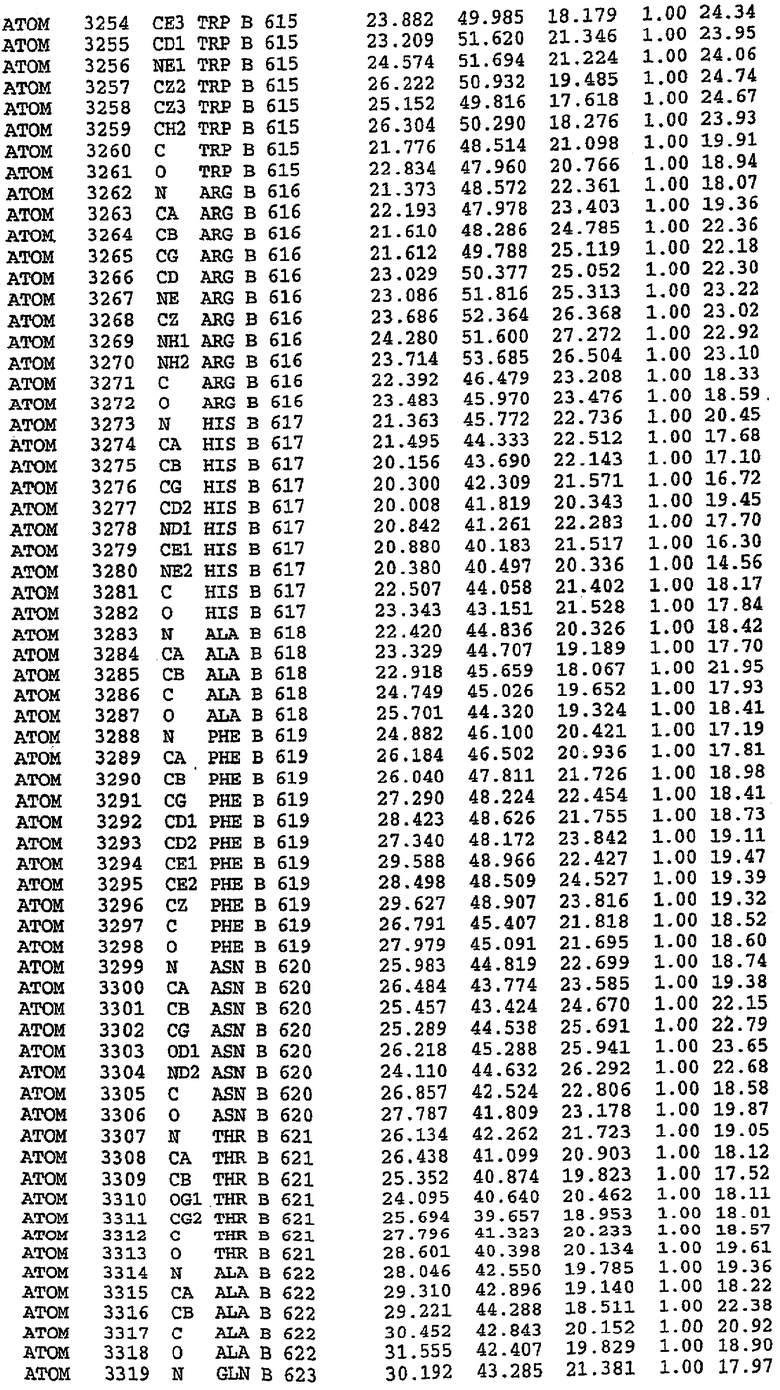
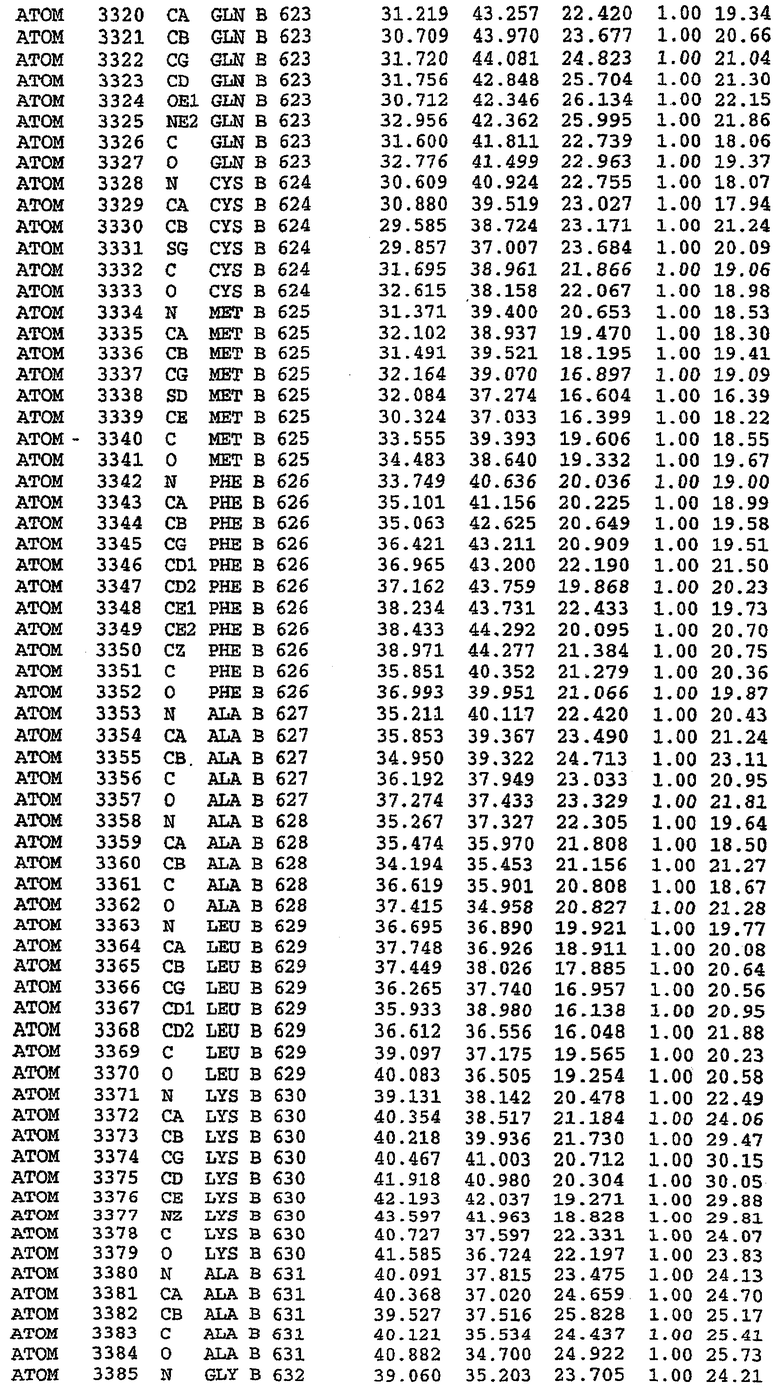
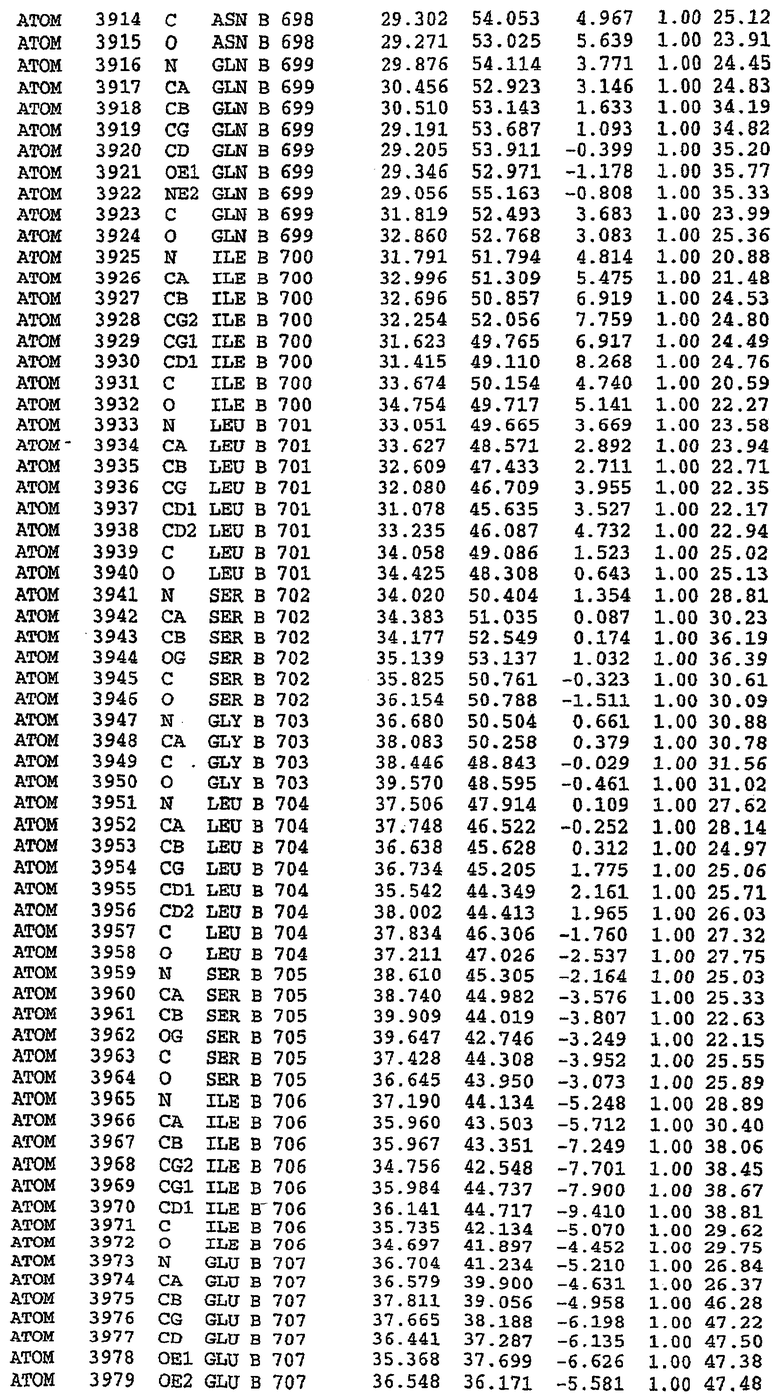
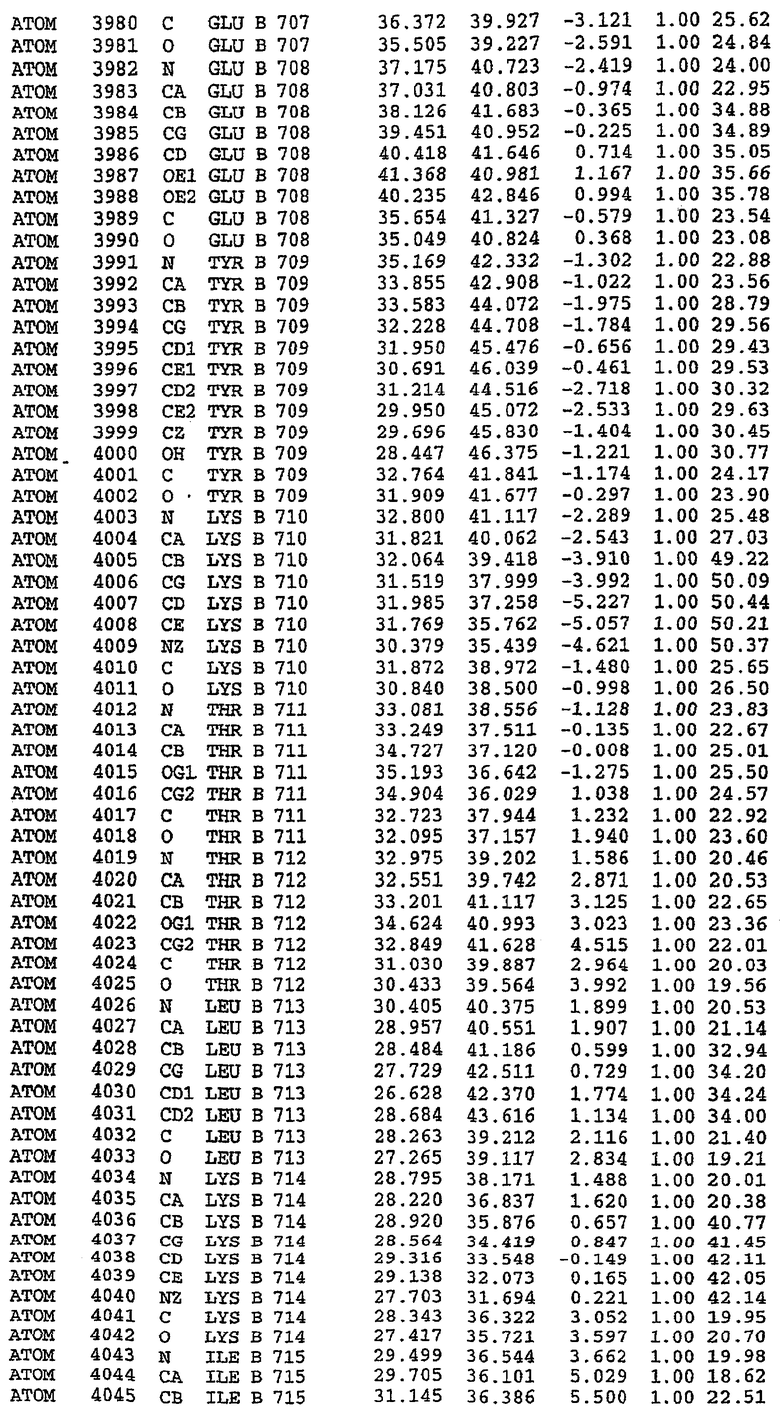
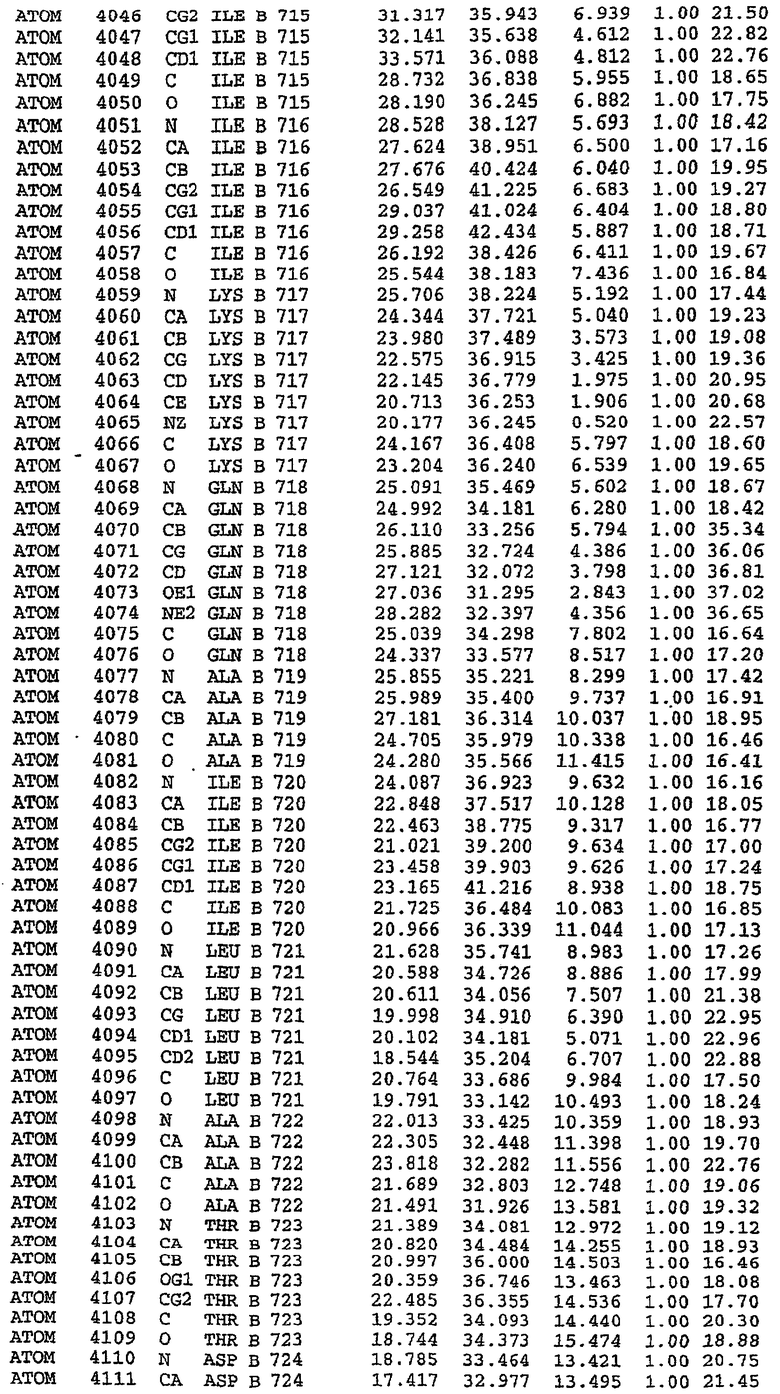
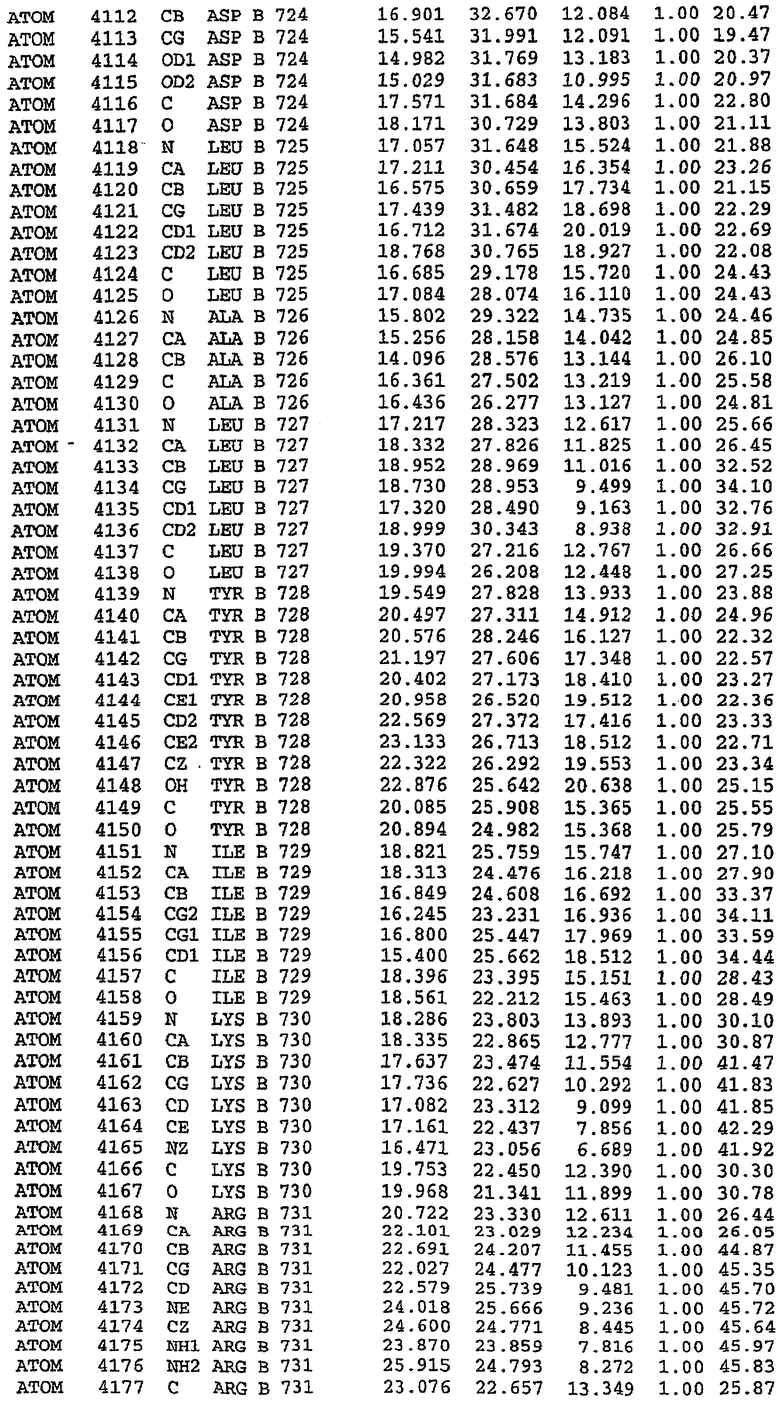
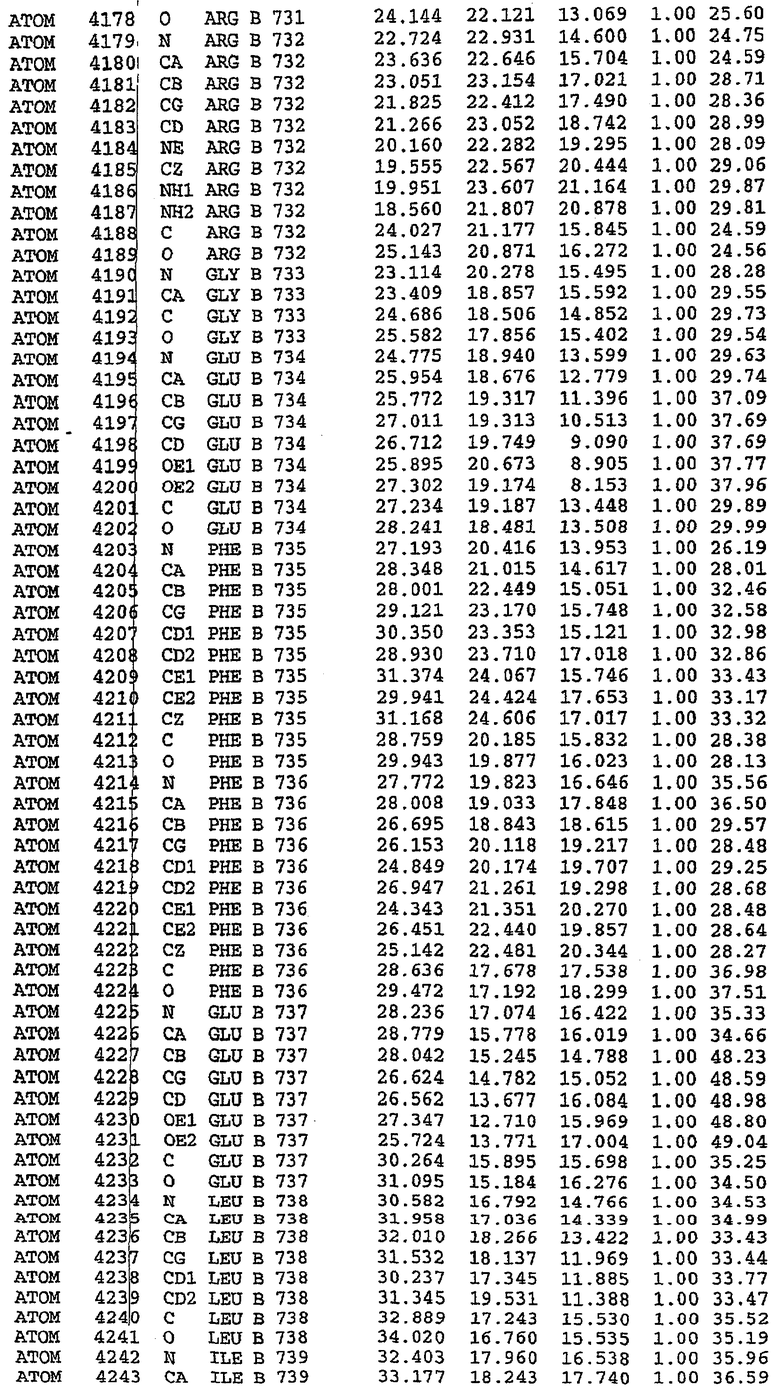
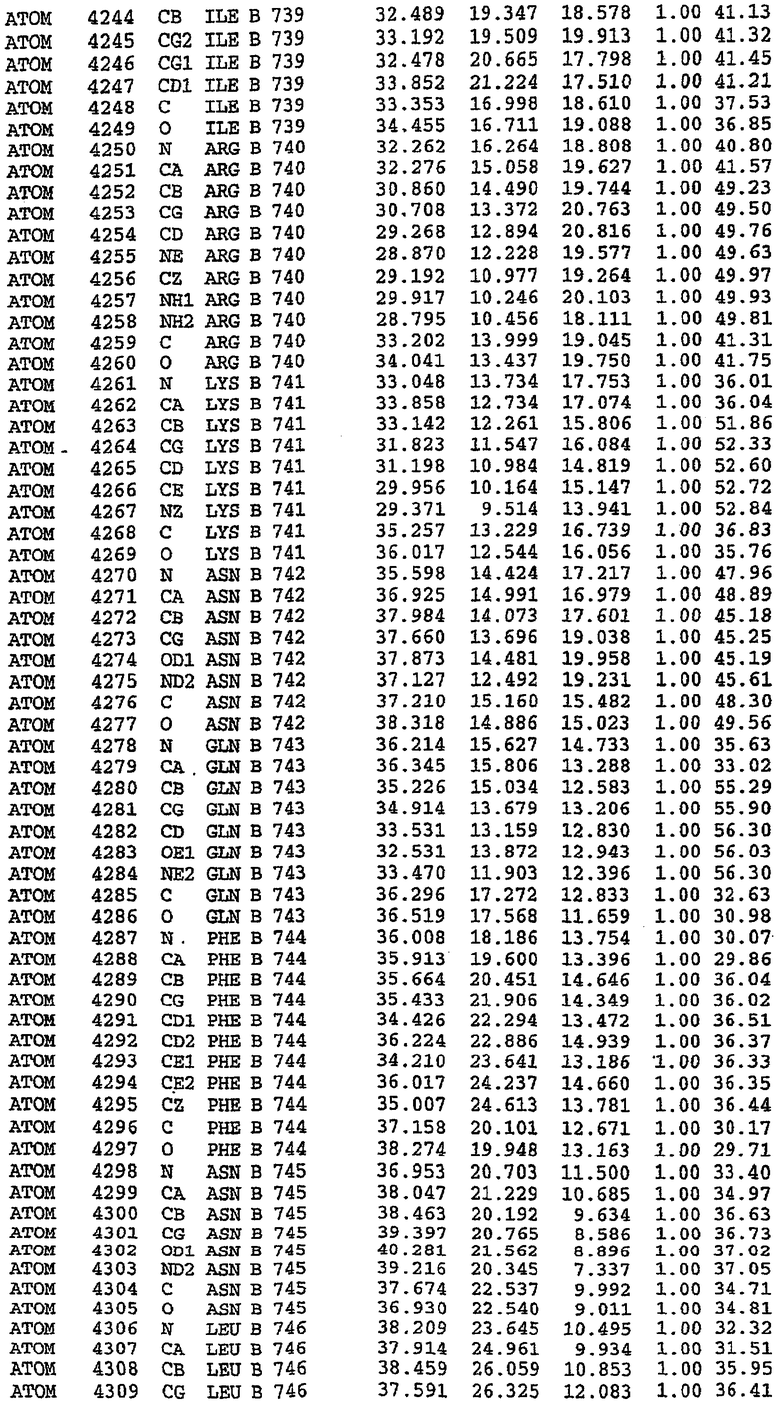
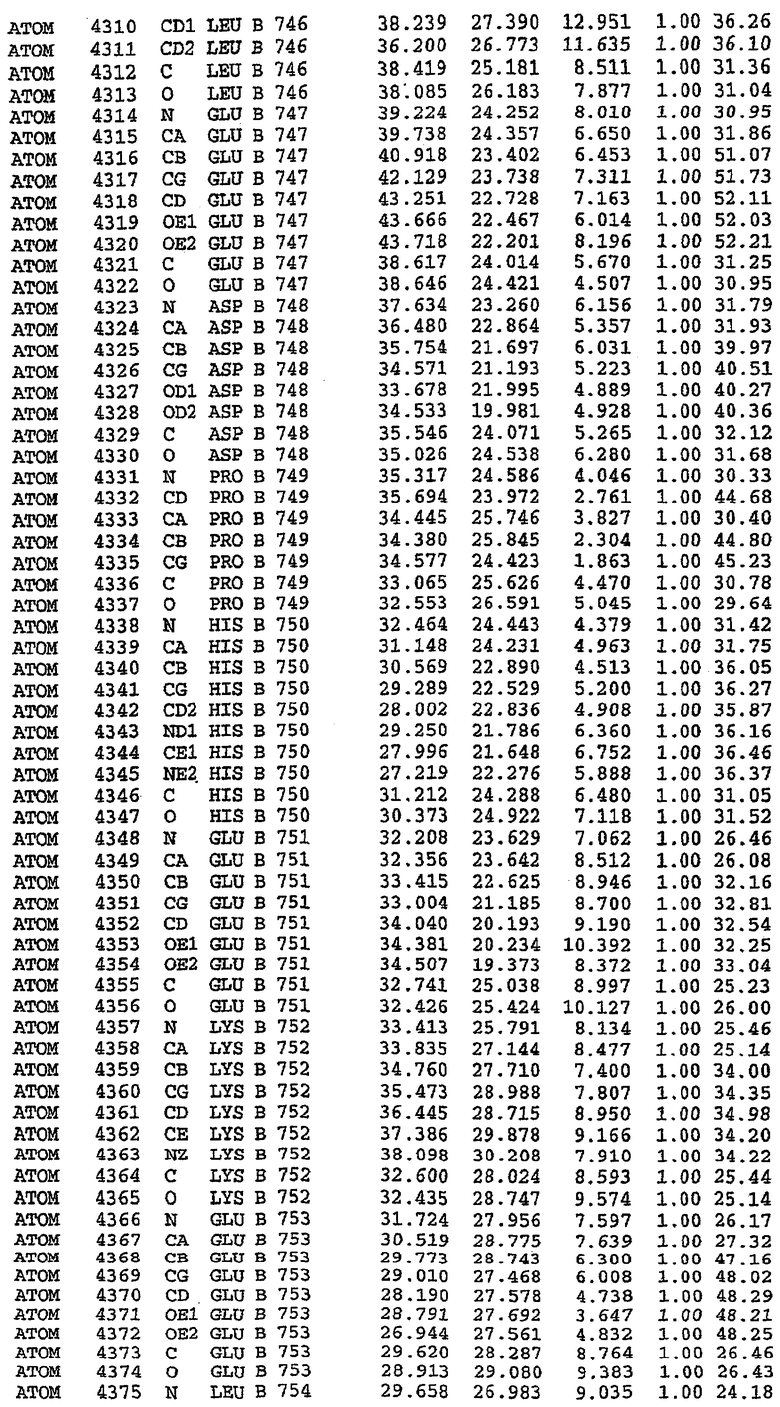
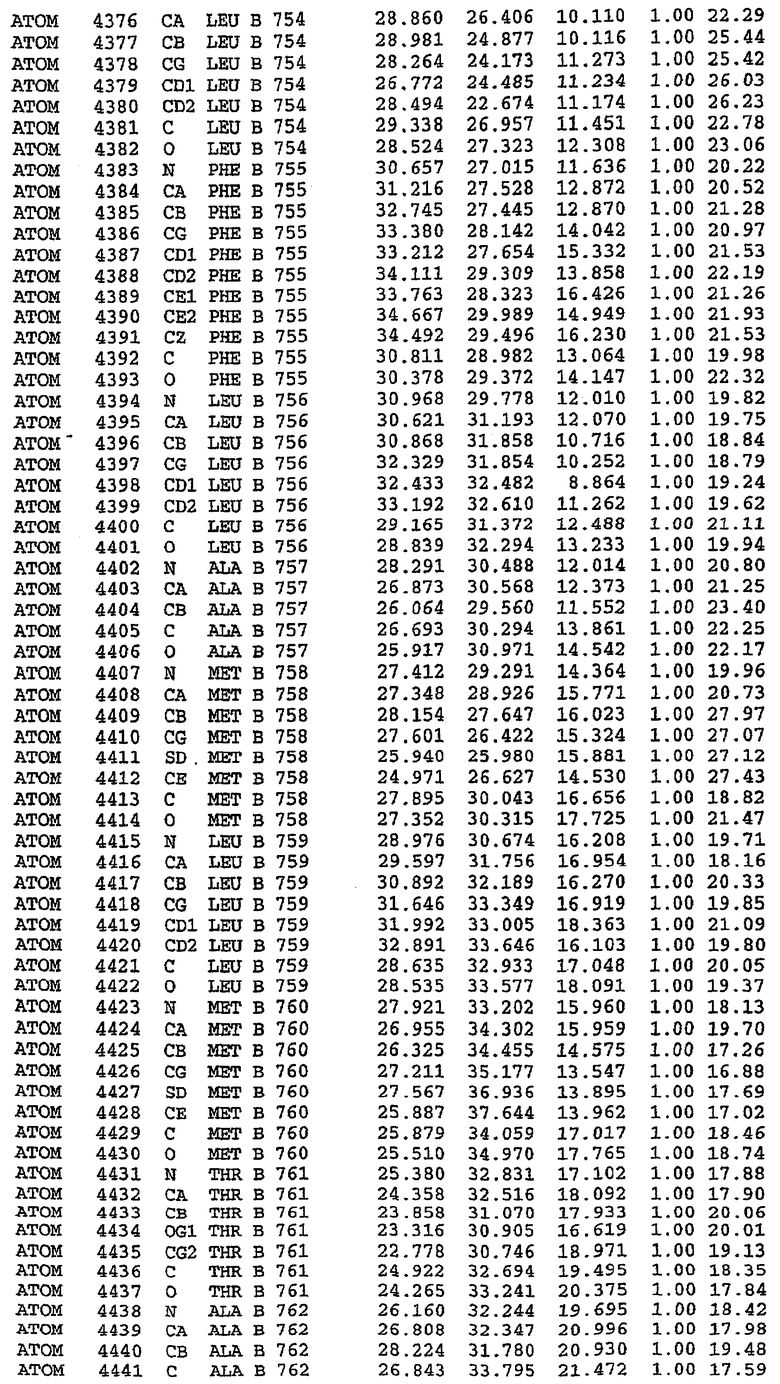

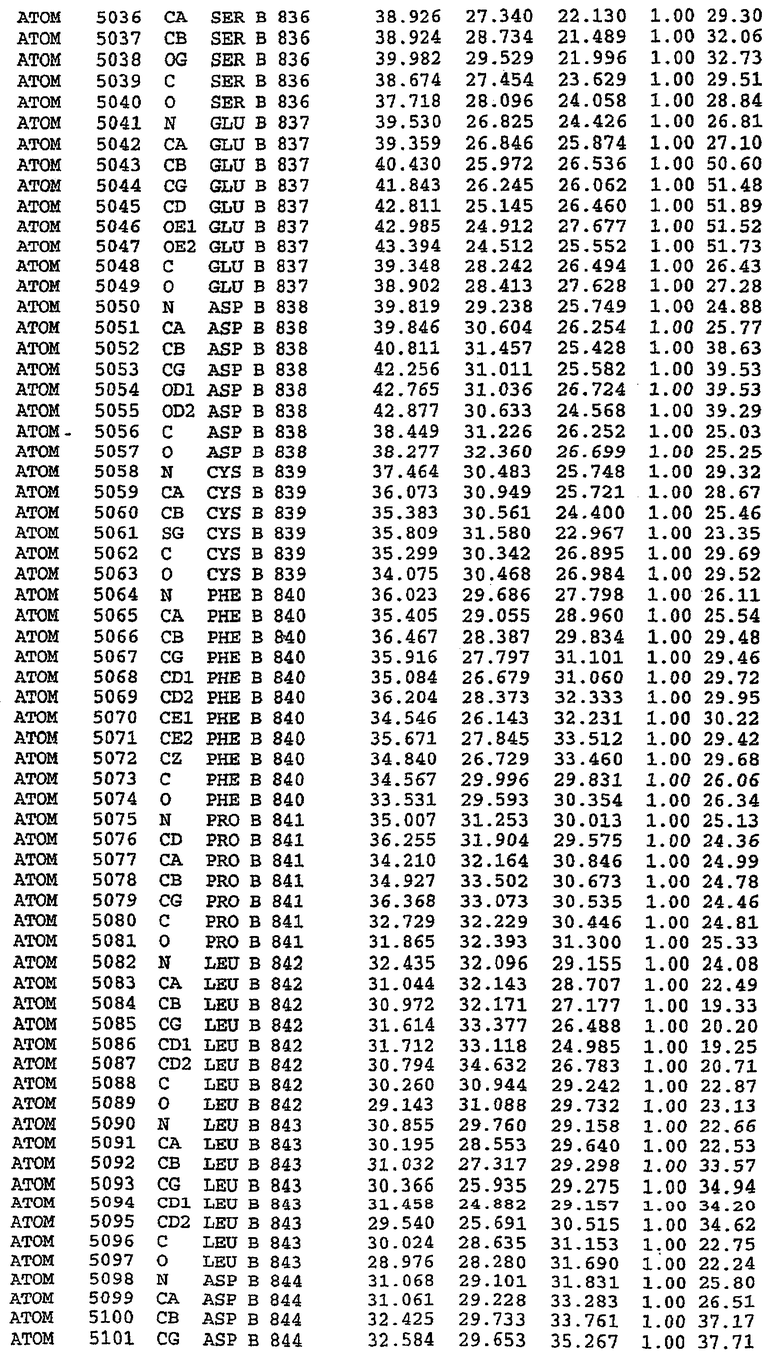
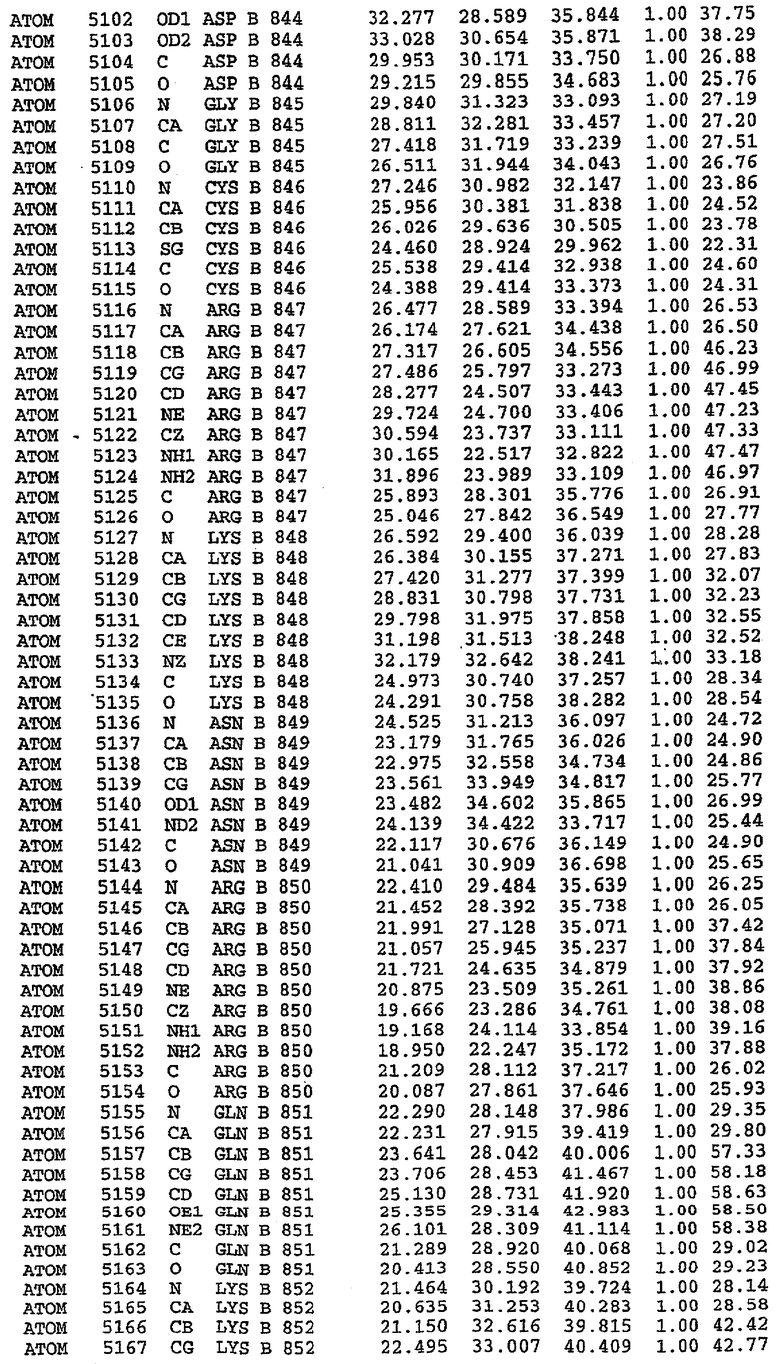












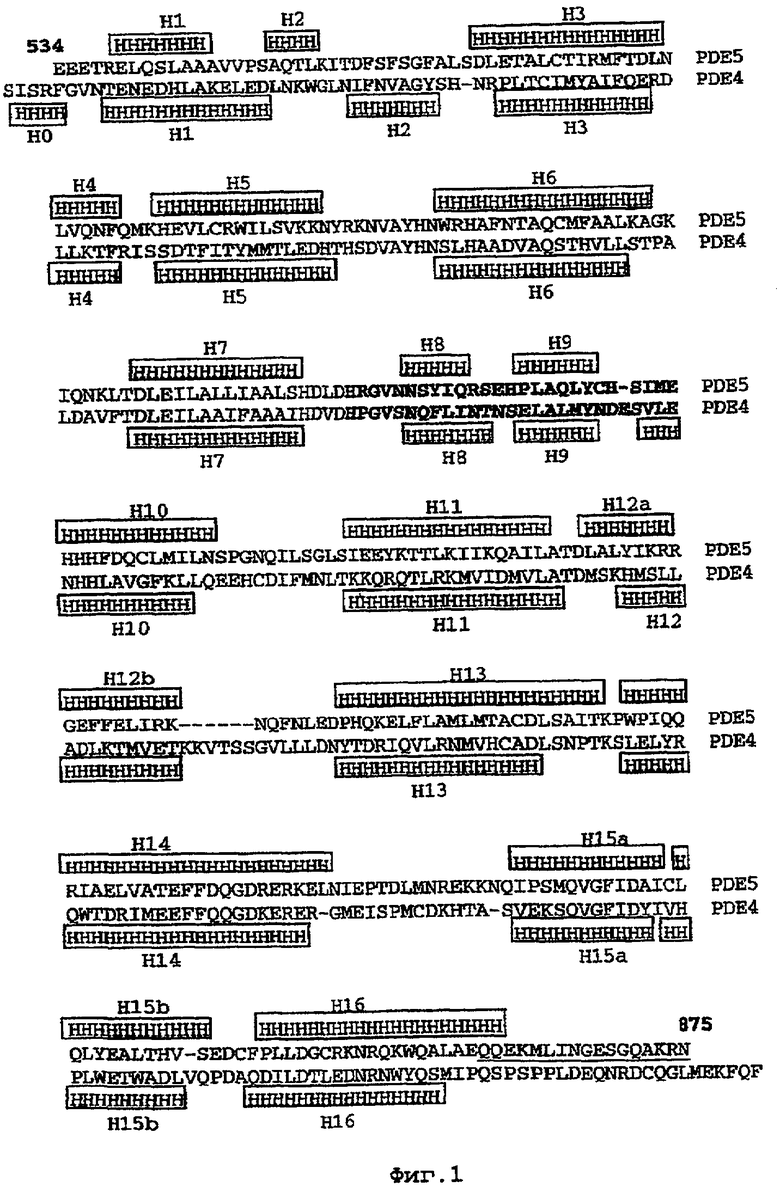
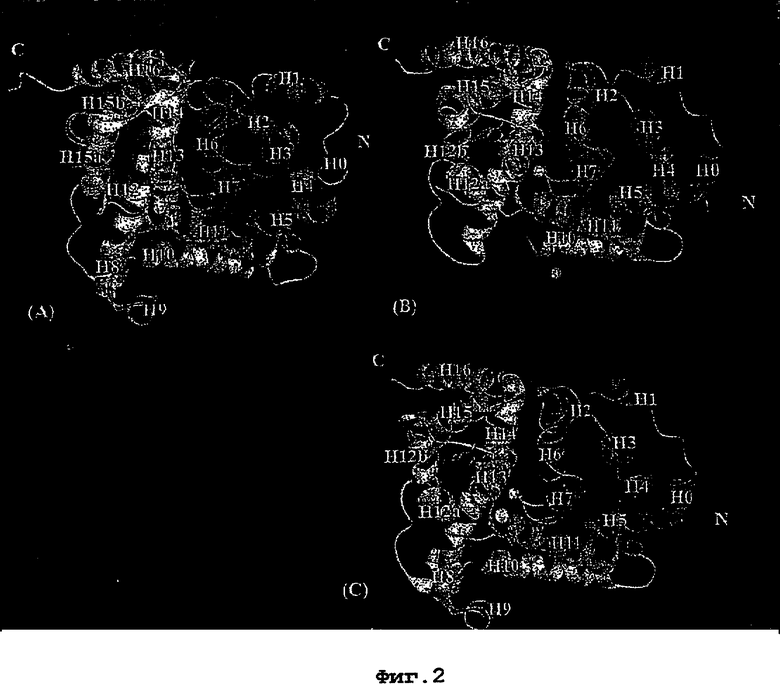
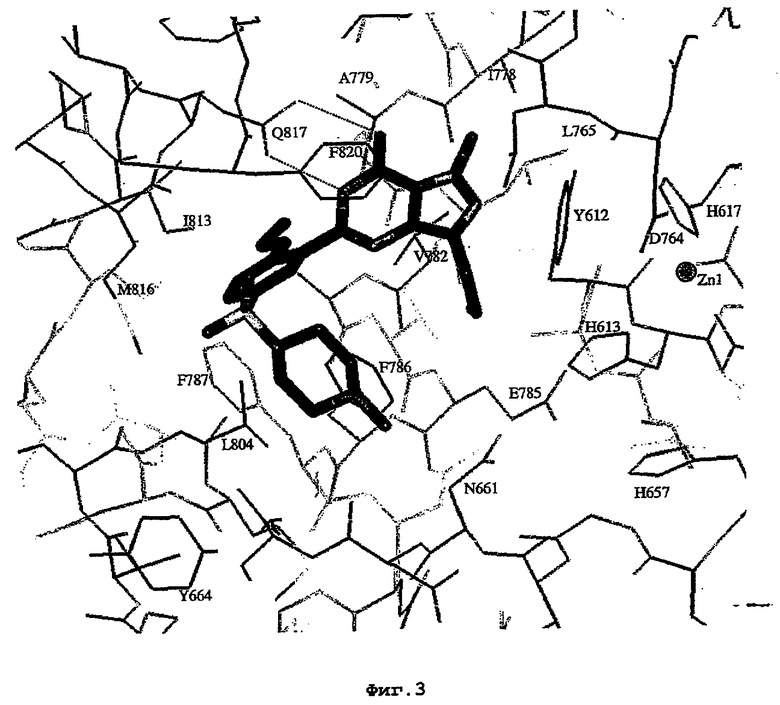
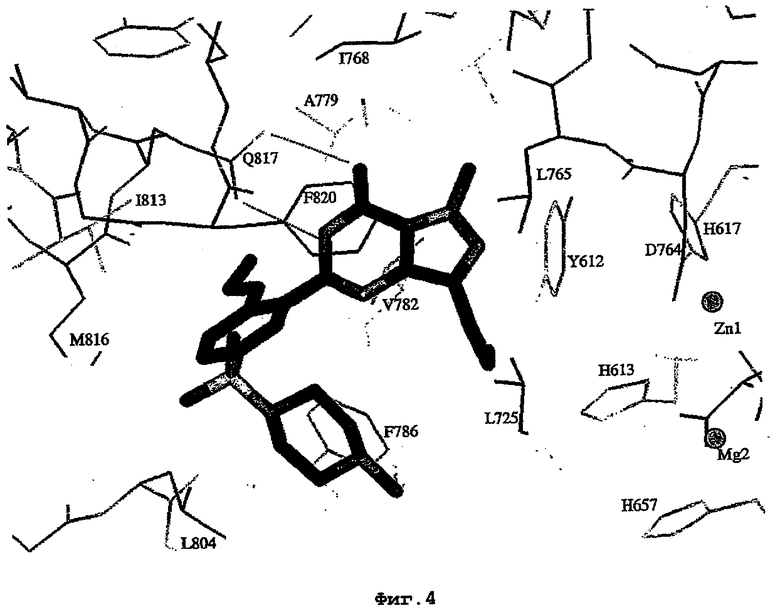
Изобретение относится к биотехнологии, в частности к кристаллическим структурам фосфодиэстеразы 5 (PDE5) и комплексов "РDЕ5/лиганд PDE5" и к их применениям для идентификации лигандов PDE5, включая соединения-ингибиторы PDE5. Настоящее изобретение также относится к способам идентификации указанных PDE5-ингибирующих соединений. Заявленное изобретение позволяет получить новый кристалл. 11 н. и 13 з.п. ф-лы, 5 ил., 8 табл.
(i) указанный каталитический домен PDE5 содержит последовательность SEQ ID NO: 2 или последовательность его гомолога, способного воспроизводить такие же характеристики кристалла при тех же условиях кристаллизации,
(ii) кристалл каталитического домена PDE5 имеет одну или несколько из следующих характеристик:
(a) спейсерную группу Р62;
(b) размеры элементарной ячейки a˜95 Å ± 1%, b˜95 Å ± 1%, c˜82 Å ± 1%, α=β=90°, γ=120°;
(c) 1 молекулу на один асимметричный элемент;
(d) содержит каталитический домен PDE5 с молекулярной массой приблизительно 40 кДа ± 2 кДа;
(e) расчетное содержание растворителя составляет приблизительно (43±5)%; и
(f) гексагональную кристаллическую систему;
(iii) каталитический домен PDE5 имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 3, или производным, выраженным в любой системе отсчета.
(i) указанный каталитический домен PDE5 содержит последовательность SEQ ID NO: 1 или последовательность его гомолога, способного воспроизводить такие же характеристики кристалла при тех же условиях кристаллизации,
(ii) кристалл каталитического домена PDE5 имеет одну или несколько из следующих характеристик:
(a) спейсерную группу Р62;
(b) размеры элементарной ячейки а˜95 Å ± 1%, b˜95 Å ± 1%, с˜82 Å ± 1%, α=β=90°, γ=120°;
(c) 1 молекулу на один асимметричный элемент;
(d) содержит каталитический домен PDE5 с молекулярной массой приблизительно 40 кДа ± 2 кДа;
(e) расчетное содержание растворителя составляет приблизительно (43±5)%; и
(f) гексагональную кристаллическую систему;
(iii) каталитический домен PDE5 имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 3, или производным, выраженным в любой системе отсчета.
(i) указанный каталитический домен PDE5 содержит последовательность SEQ ID NO: 5 или последовательность его гомолога, способного воспроизводить такие же характеристики кристалла при тех же условиях кристаллизации,
(ii) кристалл каталитического домена PDE5 имеет одну или несколько из следующих характеристик:
(a) спейсерную группу P21;
(b) размеры элементарной ячейки а˜55 Å ± 1%, b˜78 Å ± 1%, с˜82 Å ± 1%, α=γ=90°, β˜101°±2°;
(c) 2 молекулы на один асимметричный элемент;
(d) содержит каталитический домен PDE5 с молекулярной массой приблизительно 38 кДа ± 2 кДа;
(e) расчетное содержание растворителя составляет приблизительно (46±5)%; и
(f) моноклинную кристаллическую систему;
(iii) каталитический домен PDE5 имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 5, или производным, выраженным в любой системе отсчета.
(i) указанный каталитический домен PDE5 содержит последовательность SEQ ID NO: 4 или последовательность его гомолога, способного воспроизводить такие же характеристики кристалла при тех же условиях кристаллизации,
(ii) кристалл каталитического домена PDE5 имеет одну или несколько из следующих характеристик:
(a) спейсерную группу Р21;
(b) размеры элементарной ячейки а˜55 Å ± 1%, b˜78 Å ± 1%, с˜82 Å ± 1%, α=γ=90°, β˜101°±2°;
(c) 2 молекулы на один асимметричный элемент;
(d) содержит каталитический домен PDE5 с молекулярной массой приблизительно 38 кДа ± 2 кДа;
(e) расчетное содержание растворителя составляет приблизительно (46±5)%; и
(f) моноклинную кристаллическую систему;
(iii) каталитический домен PDE5 имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 5, или производным, выраженным в любой системе отсчета.
(i) кристалл комплекса каталитический домен PDE5/силденафил имеет одну или несколько их следующих характеристик:
(a) спейсерную группу P212121;
(b) размеры элементарной ячейки а˜94 Å ± 1%, b˜104 Å ± 1%, с˜142 Å ± 1%, α=β=γ=90°;
(c) 4 молекулы на один асимметричный элемент;
(d) содержит домен PDE5 с молекулярной массой приблизительно 40 кДа ± 2 кДа;
(e) расчетное содержание растворителя составляет приблизительно (43±5)%; и
(f) орторомбическую кристаллическую систему;
(ii) комплекс каталитический домен РDЕ5/силденафил имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 4, или производным, выраженным в любой системе отсчета.
(i) кристалл комплекса каталитический домен PDE5/силденафил имеет одну или несколько их следующих характеристик:
(a) спейсерную группу P21;
(b) размеры элементарной ячейки а˜55 Å ± 1%, b˜78 Å ± 1%, с˜82 Å ± 1%, α=γ=90°; β˜101°±2°;
(c) 2 молекулы на один асимметричный элемент;
(d) содержит PDE5 с молекулярной массой приблизительно 38 кДа ± 2 кДа;
(e) расчетное содержание растворителя составляет приблизительно (46±5)%; и
(f) моноклинную кристаллическую систему;
(ii) комплекс каталитический домен РDЕ5/силденафил имеет трехмерную структуру, характеризующуюся атомными координатами, представленными в таблице 6, или производным, выраженным в любой системе отсчета.
(a) компьютерное моделирование трехмерного представления структуры PDE5, полученной исходя из атомных координат, определенных в п.7, и трехмерного представления структуры потенциального лиганда PDE5;
(b) совместное отображение трехмерного представления потенциального лиганда PDE5 вместе с трехмерным представлением структуры PDE5 и
(c) установление факта соответствия трехмерного представления потенциального лиганда PDE5 трехмерному представлению активного сайта структуры PDE5.
(d) включение потенциального лиганда PDE5 в анализ на биологическую активность PDE5 и
(e) установление факта модуляции активности PDE5 потенциальным лигандом PDE5 в указанном анализе.
(a) выравнивание аминокислотной последовательности PDE5 белка с аминокислотной последовательностью (i) PDE4, (ii) каталитического домена PDE4, показанного на фигуре 1, или (iii) SEQ ID NO: 4;
(b) идентификацию структурно эквивалентного субдомена PDE5 белка, который соответствует последовательности SEQ ID NO: 4; и
(c) осуществление методами генной инженерии замены структурно эквивалентного субдомена PDE5 или его части последовательностью SEQ ID NO: 4 или последовательностью ее гомолога, способного воспроизводить такие же характеристики кристалла при тех же условиях кристаллизации.
(a) трис- или MES-буфер, фосфат аммония и/или PEG2KMME;
(b) трис-буфер, ацетат натрия и/или PEG4K; или
(с) 0,16 М ацетат натрия, 80 мМ трис-гидрохлорид, рН 8,5, 24% мас./об. PEG8KMME.
| Погрузчик | 1956 |
|
SU127101A1 |
| LA 9902793, 19.10.2001 | |||
| UKITA T | |||
| et al | |||
| Novel, potent, and selective phosphodiesterase 5 inhibitors: synthesis and biological activities of a series of 4-aryl-1-isoquinolinone derivatives, J Med Chem | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| ПРОИЗВОДНЫЕ 8-АРИЛ-1,7-НАФТИРИДИНА И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ОБЛАДАЮЩАЯ ПРОТИВОВОСПАЛИТЕЛЬНОЙ АКТИВНОСТЬЮ | 1997 |
|
RU2174123C2 |

