Настоящее изобретение обеспечивает выделенный мутантный белок atpE и начинающуюся с указанного мутантного белка atpE идентификацию домена связывания ATP-азы. Настоящее изобретение также обеспечивает связанные с этим нуклеиновые кислоты, векторы, клетки-хозяева, фармацевтические композиции и изделия. Кроме того, настоящее изобретение предоставляет способы определения того, взаимодействует ли исследуемое соединение с белком atpE, то есть с доменом связывания ATP-азы по настоящему изобретению, а также композиции фармацевтических препаратов, содержащие указанное исследуемое соединение, в частности, в качестве антимикробных агентов, более конкретно в качестве антимикобактериального агента, еще более конкретно агента для лечения туберкулеза у субъекта.
Уровень техники
После СПИДа туберкулез (TB) представляет собой ведущую причину смертности у взрослых (2-3 миллиона смертей в год) в мире и представляет собой критический сдерживающий фактор в облегчении глобальной бедности и страданий (1). Факторы, вносящие вклад в возобновление заболевания, включают сложности осуществления антитуберкулезных программ во многих странах, резкое увеличение количества индивидуумов с ослабленным иммунитетом, в основном, в связи с ВИЧ-инфекцией - и перемещение людей через области, где TB является эндемическим, и из них. Эпидемии TB и ВИЧ подпитывают друг друга в людях, инфицированных совместно - в настоящее время 11 миллионов взрослых, - увеличивая как заболеваемость, так и смертность (2, 3). В дополнение к этому, TB представляет собой ведущую причину смерти у людей, инфицированных ВИЧ (4).
Хотя режимы введения противотуберкулезных лекарственных средств первой линии могут достигать более чем 90% долей эффективности, их сложность может приводить к плохой совместимости, когда адекватная медицинская поддержка и программы лечения TB являются недоступными и, кроме того, к появлению резистентности (5). Устойчивые к множеству лекарственных средств (MDR) штаммы TB существенно осложняют лечение (6). The Global Alliance for TB Drug Development рекомендует, чтобы любое новое лечение предлагало, по меньшей мере, одно из трех следующих преимуществ по сравнению с существующими видами терапии: сокращение или упрощение эффективного лечения TB; увеличение эффективности против MDR-TB; и улучшение лечения латентной формы инфекции TB. Такое новое лекарственное средство сильно улучшило бы податливость пациента, при этом понижая стоимость программ лечения TB, подобных стратегии World Health Organization (WHO)'s Directly Observed Treatment Short-course (DOT) (7).
Более новые кандидаты в противотуберкулезные агенты, находящиеся в настоящее время в преклинической и клинической разработке, имеют тенденцию к тому, чтобы либо принадлежать к существующим семействам лекарственных средств (такие как моксифлоксацин), либо к аналогам лекарственных средств первой линии, таких как MJH-98-1-81 (из изониазида), оксазолидиноны и рифапентин (близкий аналог рифампина) (8). Хотя эти новые лекарственные средства могут быть сильнодействующими, аналоговые соединения обеспечивают только временные решения проблем резистентности (9), поскольку они основываются на таком же механизме действия, как и существующие семейства лекарственных средств.
Антибиотики, как правило, обычно ингибируют бактериальную репликацию посредством ингибирования бактериального метаболизма через специфический механизм. Например, изониазид вмешивается в работу ферментативного аппарата, который синтезирует миколиновые кислоты, необходимые компоненты клеточной стенки, в то время как рифампицин вмешивается в деятельность бактериального механизма транскрипции РНК из ДНК. Соответственно, представляет интерес обнаружение новых способов идентификации противотуберкулезных соединений, которые нацелены на другие конкретные микобактериальные аспекты роста клеток и репликации, по сравнению с известными агентами.
Сущность изобретения
Настоящее изобретение предоставляет выделенные мутантные белки atpE, в частности, кодируемые аминокислотными последовательностями, выбранными из (SEQ ID No.1), (SEQ ID No.2), (SEQ ID No.3), (SEQ ID No.4) и (SEQ ID No.5), выделенную нуклеиновую кислоту, кодирующую указанные мутантные белки atpE, в частности выбранную из группы, состоящей из (SEQ ID No.6), (SEQ ID No.7), (SEQ ID No.8), (SEQ ID No.9) и (SEQ ID No.10), и вектор, содержащий рассматриваемую нуклеиновую кислоту. В конкретном варианте осуществления мутантный белок atpE кодируется SEQ ID No.2, и выделенная последовательность нуклеиновой кислоты, кодирующая указанный белок, состоит из SEQ ID No.7.
Кроме того, настоящее изобретение обеспечивает систему хозяин-вектор, включающую клетку-хозяин, содержащую в себе рассматриваемый вектор экспрессии.
Кроме того, настоящее изобретение обеспечивает выделенную клетку, содержащую мутантный белок atpE, где указанный белок индуцирует в клетке антимикробную устойчивость.
Кроме того, настоящее изобретение обеспечивает способ идентификации антимикробного соединения, включающий стадии
(a) контактирования клетки, экспрессирующей белок atpE, с исследуемым соединением в физиологических условиях;
(b) определения того, взаимодействует ли исследуемое соединение с белком atpE.
Кроме того, настоящее изобретение предоставляет способ оценки потенциала исследуемого соединения к взаимодействию с белком atpE, включающий:
(a) использование методов молекулярного моделирования для генерирования трехмерной структуры белка atpE;
(b) использование компьютерных средств для осуществления операции совмещения исследуемого соединения и трехмерной структуры белка atpE; и
(c) анализ результатов указанной операции совмещения для количественного определения ассоциации исследуемого соединения с трехмерной структурой белка atpE.
Целью настоящего изобретения также является создание сайта связывания части F0 ATP-азы, содержащего, по меньшей мере, аминокислоты Ala24, Gly27, Phe53, Val57, Gly58, Glu61, Tyr64 и Phe65 одной субъединицы C; аминокислоты Ser182, Leu183, Ser184, Leu185 и Arg186 одной субъединицы A, и указанные аминокислоты имеют атомные координаты любой из таблиц 3, 4 или 5.
Еще в одном аспекте настоящее изобретение предусматривает использование указанного выше домена связывания в способе идентификации соединений, которые взаимодействуют с частью F0 ATP-азы, и их потенциала в качестве антимикробных соединений, в частности, в способе идентификации антимикобактериальных соединений.
Соответственно, целью настоящего изобретения является создание способа лечения субъекта с микробной инфекцией, включающего введение субъекту соединения, которое взаимодействует с частью F0 ATP-азы, в частности с белком atpE, на сайтах мутации, придающей устойчивость, или с сайтом связывания по настоящему изобретению. Кроме того, настоящее изобретение предусматривает способ лечения субъекта, страдающего туберкулезом, включающий введение субъекту агента, который взаимодействует с белком atpE, используя любой из указанных выше способов скрининга. В способах лечения, включающих использование соединения, которое взаимодействует с частью F0 ATP-азы, в частности с белком atpE, соединения, о которых ранее было известно, что они взаимодействуют с частью F0 ATP-азы, и, в частности с белком atpE, должны быть исключены. Более конкретно, должно быть исключено использование соединений DARQ J, описанных в (11), в любом из описанных способов лечения.
Кроме того, настоящее изобретение обеспечивает фармацевтическую композицию, содержащую агент, который взаимодействует с белком atpE в клетке, и фармацевтически приемлемый носитель. Наконец, настоящее изобретение предусматривает изделие, содержащее упаковку и фармацевтический агент, где (a) фармацевтический агент взаимодействует с белком atpE в клетке и (b) упаковка содержит этикетку, указывающую на использование агента для лечения бактериальной инфекции у субъекта. В конкретном варианте осуществления настоящее изобретение предусматривает использование DARQ J при производстве антимикробного лекарственного средства.
Этот и другие аспекты настоящего изобретения будут обсуждаться более подробно далее.
Краткое описание таблиц и чертежей
Таблица 1. Минимальные ингибирующие концентрации (MIC) ведущего соединения DARQ (J), которые ингибируют 90% роста различных видов микобактерий. Количество исследуемых штаммов было n=1, если не указано иначе.
Таблица 2. Аминокислоты, окружающие сайт связывания для соединения DARQ J.
Таблица 3. Атомные координаты аминокислот, окружающих сайт связывания для соединения DARQ J, полученного как из дикого типа, так и из мутанта DARQ J штамма M.tuberculosis.
Таблица 4. Атомные координаты сайта связывания для соединения DARQ J в диком типе M.tuberculosis.
Таблица 5. Атомные координаты сайта связывания для соединения DARQ J мутантного штамма DARQ J M.tuberculosis.
Таблица 6. Атомные координаты мутантного белка atpE (SEQ ID No.2) M.tuberculosis.
Таблица 7. Атомные координаты белка atpE дикого типа (SEQ ID No.l) M.tuberculosis.
Фиг.1. Абсолютная конфигурация R207910, далее также упоминаемого как J или DARQ J.
Фиг.2. Сравнения последовательностей белка atpE для мутантов M.tuberculosis и M.smegmatis. Mtb_S: чувствительный к лекарственным средствам штамм M.tuberculosis H37Rv, atpE (1-81). Номер доступа: Swiss-Prot Q10598 (SEQ ID No.1). Mtb_R: устойчивый к лекарственным средствам штамм M.tuberculosis BK12, atpE (1-81) (SEQ ID No.2). Msm_S: чувствительный к лекарственным средствам штамм M.smegmatis, atpE (1-86). Последовательность, полученная Institute for Genome Research (SEQ ID No.3). Msm_R09 (SEQ ID No.4) и R10 (SEQ ID No.5): устойчивые к лекарственным средствам штаммы M.smegmatis atpE (1-86). Последовательности, полученные в лаборатории. Для человека: Homo sapiens, ATP5G3 (66-142). Номер доступа: Ensembl ENSP00000284727. Верхняя нумерация: M.tuberculosis и M.smegmatis atpE. Нижняя нумерация: H sapiens ATP5G3 (66-142). Черный цвет указывает сходство аминокислот с использованием матрицы BLOSUM62 (черный = высокое, серый = среднее). Стрелки указывают положения точечных мутаций, наблюдаемых у стойких штаммов.
Фиг.3. Измерение общего клеточного ATP M. Tuberculosis в присутствии DARQ J, изониазида и DCCD. Относительные единицы люминесценции оксилюциферина, измеренной при 526 нм, как у дикого типа M.tuberculosis, так и у мутанта DARQ J M.tuberculosis.
Фиг.4. Ленточное представление трех C-субъединиц (цепь A, цепь K и цепь L) и субъединицы A (цепь M), которые вместе образуют сайт связывания для соединения DARQ J.
Подробное описание
Определения
Как используется в настоящей заявке, если только здесь в явном виде не предусматривается иначе, каждый из следующих далее терминов будет иметь значение, приведенное ниже.
"Белок atpE" должен обозначать цепь C субъединицы F0 комплекса ATP-азы, как представлено SwissProt, код доступа Q10598 для M.tuberculosis, или белок, имеющий, по меньшей мере, 70, 80, 90, 95, 97 или 99% идентичность последовательности с указанной последовательностью M.tuberculosis.
"F1F0 ATP-аза", упоминаемая также как ATP-аза, ATP-синтаза или F0F1 ATP-аза, должна обозначать большой комплекс из множества субъединиц, который катализирует синтез или гидролиз ATP. F0F1 ATP-азы состоят из двух доменов: части F1, которая является внешней по отношению к мембране и содержит каталитические сайты, и части F0, которая охватывает бислой и содержит протонный канал. ATP-азы находятся в плазматической мембране бактерий, тилакоидной мембране хлоропластов и во внутренней мембране митохондрии, где они используют энергию электрохимического градиента протонов для осуществления синтеза ATP.
"Введение" должно обозначать доставку способом, который осуществляется или производится с использованием любого из различных способов и систем доставки, известных специалистам в данной области. Введение может осуществляться, например, местно, внутривенно, перикардиально, перорально, посредством импланта, трансмукозально, трансдермально, внутримышечно, подкожно, внутрибрюшинно, интратекально, интралимфатически, в поврежденную ткань или эпидурально. Введение может также осуществляться, например, один раз, много раз и/или в течение одного или нескольких продолжительных периодов.
"Клетки-хозяева" включают, но не ограничиваются этим, бактериальные клетки, дрожжевые клетки, грибковые клетки, клетки насекомых и клетки млекопитающих. Бактериальные клетки могут быть трансфицированы способами, хорошо известными в данной области, такими как кальцийфосфатное осаждение, электропорация и микроинъекция.
"Выделенный", по отношению к белку atpE, должен обозначать препарат фрагмента мембраны, содержащий белок atpE, или другой подходящий для использования препарат, где atpE сохраняет свою природную функцию и является свободным от некоторых или всех иных белков в его нативной окружающей среде. Подразумевается включение препаратов мембран, содержащих часть F0 F0F1 ATP-азы, в частности часть F0, содержащую мутантные белки atpE по настоящему изобретению.
"Бактериальная клетка" должна означать любую бактериальную клетку. Бактериальные клетки включают, без ограничения, клетки, которые являются нормальными, аномальными и трансформированными, и иллюстрируются микобактериями, в частности, Mycobacterium tuberculosis и Mycobacterium smegmatis, corynebacteria, nocardia, грамположительными бактериями, такими как, например, стрептококк, стафилококк и энтерококк или грамотрицательными бактериями, такими как, например, Escherichia coli, Heamophilus influenzae и Helicobacter pylori.
Термины "нуклеиновая кислота" и "полинуклеотид" здесь используются взаимозаменяемо и каждый из них относится к полимеру дезоксирибонуклеотидов и/или рибонуклеотидов. Дезоксирибонуклеотиды и рибонуклеотиды могут быть встречающимися в природе соединениями или их синтетическими аналогами.
Термин "физиологические условия" должен означать по отношению к данной клетке такие условия, которые обычно составляли бы биохимическую окружающую среду клетки. Биохимическая окружающая среда клетки включает, без ограничения, некоторые или все протеазы, для которых клетка обычно экспонируется. Такие условия включают, но не ограничиваются этим, условия in vivo.
Термины "полипептид", "пептид" и "белок" используются здесь взаимозаменяемо и каждый из них означает полимер аминокислотных остатков. Аминокислотные остатки могут быть встречающимися в природе соединениями или их химическими аналогами. Полипептиды, пептиды и белки могут также включать модификации, такие как гликозилирование, присоединение липидов, сульфатирование, гидроксилирование и ADP-рибозилирование.
"Субъект" должен означать любое животное, такое как млекопитающее или птица, включая, без ограничения, корову, лошадь, овцу, свинью, собаку, кошку, грызуна, такого как мышь или крыса, индейку, курицу и примата. В предпочтительном варианте осуществления субъектом является человек.
"Лечение" должно включать, без ограничения, устранение, обращение течения, замедление развития, ослабление симптомов или облегчение другим образом заболевания субъекта.
"Вектор" должен означать любой вектор нуклеиновой кислоты, известный в данной области. Такие векторы включают, но не ограничиваются этим, плазмидные векторы, космидные векторы и векторы бактериофагов.
Термины "вещество-кандидат" и "исследуемое соединение" используются взаимозаменяемо и относятся к веществу, которое, как предполагается, взаимодействует с другим остатком, то есть с белком atpE, в качестве модификатора биологической реакции. Например, репрезентативное вещество-кандидат, как предполагается, взаимодействует с белком atpE и, как предполагается, модифицирует активность ATP-азы. Примеры веществ-кандидатов, которые могут быть исследованы с использованием способов по настоящему изобретению, включают, но не ограничиваются ими, пептиды, ферменты, субстраты ферментов, кофакторы, сахара, олигонуклеотиды, малые молекулы химических соединений и моноклональные антитела.
"Модулировать" должно означать увеличение, уменьшение или другое изменение каких-либо или всех химических и биологических активностей или свойств белка atpE дикого типа или мутантного.
"Взаимодействовать" должно означать детектируемые взаимодействия между молекулами, включая "связывающие" взаимодействия между молекулами. Взаимодействия могут представлять собой, например, взаимодействие белок-белок или белок-нуклеиновая кислота в природе. Такие взаимодействия могут быть выявлены с использованием методик, известных в данной области, например, двухгибридным анализом на дрожжах, иммунопреципитацией, анализом SPA или анализом связывания на фильтре.
Как здесь используется, термин "атомные координаты" или "структурные координаты" относится к математическим координатам, которые описывают положения атомов в формате Protein Data Bank (PDB), включая X, Y, Z и B, для каждого атома. Специалисты в данной области понимают, что набор структурных координат, определенных с помощью рентгеновской кристаллографии, не обходится без стандартной ошибки. Для целей настоящего изобретения любой набор структурных координат для ATP-синтазы из любого источника, имеющий среднеквадратичное отклонение для атомов, отличных от водорода, менее чем 1,5 Å, когда он накладывается на положение соответствующих атомных координат для атомов, отличных от водорода, из таблиц 3, 4, 5, 6 или 7, рассматривается как по существу идентичный или гомологичный. В более предпочтительном варианте осуществления, любой набор структурных координат для ATP-синтазы из любого источника, имеющий среднее квадратичное отклонение для атомов, отличных от водорода, менее чем 0,75 Å, когда он накладывается на положение соответствующих атомных координат атомов, отличных от водорода, из таблиц 3, 4, 5, 6 или 7, рассматривается как по существу идентичный.
Варианты осуществления изобретения
Мутантные белки atpE
Настоящее изобретение обеспечивает выделенные мутантные белки atpE, в частности бактериальные белки atpE, более конкретно, микобактериальные белки atpE, еще более конкретно, белки atpE M.tuberculosis или M.smegmatis. Мутация выбирается из единственной точечной мутации, инсерции или делеции. В одном из вариантов осуществления настоящего изобретения мутация состоит, по меньшей мере, из одной точечной мутации, расположенной в любой из аминокислот 20-40, в частности 30-40, предпочтительно, в аминокислоте 34 или в аминокислотах 60-75, в частности 62-73, предпочтительно, в аминокислоте 69, как показано в сравнении последовательностей на фиг.2. В дополнительном варианте осуществления выделенные мутантные белки atpE выбираются из Mtb_R (SEQ ID No.2), Msm_R09 (SEQ ID No.4) и Msm_R10 (SEQ ID No.5), как показано на фиг.2, или из аминокислотной последовательности, имеющей, по меньшей мере, 70, 80, 90, 95, 97 или 98% идентичность последовательности с любой из указанных выше аминокислотных последовательностей.
Кроме того, настоящее изобретение обеспечивает выделенную нуклеиновую кислоту, кодирующую указанные мутантные белки atpE. В одном из вариантов осуществления указанная последовательность нуклеиновой кислоты состоит из всех генов, которые кодируют часть F0, как описано, например, в J. Biol. Chem., 1994, Vol. 269(10), p.7285-7289, где указанные гены транскрибированы из одного промотора и содержат последовательность нуклеиновой кислоты, кодирующую мутантный белок atpE по настоящему изобретению. Нуклеиновая кислота может представлять собой ДНК или РНК, предпочтительно ДНК, и в дополнительном варианте осуществления они выбираются из последовательностей нуклеиновых кислот, кодирующих Mtb_R (SEQ ID No.7), Msm_R09 (SEQ ID No.9), Msm_R10 (SEQ ID No.10), или последовательности нуклеиновой кислоты, имеющей, по меньшей мере, 70, 80, 90, 95, 97 или 98% идентичность последовательности с любой из указанных выше последовательностей нуклеиновых кислот.
Процент идентичности последовательностей нуклеиновых кислот и полипептидов может быть вычислена с использованием коммерчески доступных алгоритмов, которые сравнивают эталонную последовательность с запрашиваемой последовательностью. Следующие далее программы (поставляемые National Center for Biotechnology Information) могут использоваться для определения гомологий/идентичностей: BLAST, BLAST с пропусками, BLASTN и PSI-BLAST, которые могут использоваться с параметрами по умолчанию.
Алгоритм GAP (Genetics Computer Group, Madison, WI) использует алгоритм Needleman and Wunsch для сравнения двух полных последовательностей, который доводит до максимума количество совпадений и сводит к минимуму количество пропусков. Как правило, используются параметры по умолчанию со штрафом при образовании пропуска = 12 и штрафом при расширении пропуска = 4.
Другой способ для определения наилучшего общего совпадения между последовательностью нуклеиновой кислоты или ее частью и исследуемой последовательностью представляет собой использование компьютерной программы FASTDB, основанной на алгоритме Brutlag et al (Comp. App. Biosci., 6; 237-245 (1990)). Программа обеспечивает общее сравнение последовательностей. Результат указанного общего сравнения последовательностей выражается в проценте идентичности. Подходящие параметры, используемые при поиске FASTDB последовательности ДНК, для вычисления процента идентичности представляют собой: Matrix (Матрица)=унитарная, k-tuple=4 (Кортеж из k=4 элементов), Mismatch Penalty (Штраф за несовпадение)=1, Joining Penalty (Штраф за сращивание)=30, Randomization Group Length (Длина группы рандомизации)=0, Cutoff Score (Балл отбрасывания)=1, Gap Penalty (Штраф за пропуск)=5, Gap Size Penalty (Штраф за размер пропуска)=0,05 и Window Size (Размер окна)=500 или длине исследуемой последовательности в нуклеотидных основаниях, то, что короче. Подходящие параметры для вычисления процента идентичности и сходства сравнения аминокислот представляют собой: Matrix=PAM 150, k-tuple=2, Mismatch Penalty=1, Joining Penalty=20, Randomization Group Length=0, Cutoff Score=1, Gap Penalty=5, Gap Size Penalty=0,05 и Window Size=500 или длине исследуемой последовательности в нуклеотидных основаниях, то, что короче.
Кроме того, настоящее изобретение предоставляет вектор, содержащий рассматриваемую нуклеиновую кислоту. В одном из вариантов осуществления вектор представляет собой плазмидный вектор.
Кроме того, настоящее изобретение предусматривает систему хозяин-вектор, содержащую клетку-хозяин, имеющую рассматриваемый плазмидный вектор. Клетка может быть прокариотической или эукариотической, в одном из вариантов осуществления клетка-хозяин представляет собой бактериальную клетку, в частности микобактериальную клетку, такую как, например, M.tuberculosis или M.smegmatis.
Кроме того, настоящее изобретение обеспечивает выделенную клетку, содержащую мутантный белок atpE, который индуцирует антимикробную устойчивость в клетке. В одном из вариантов осуществления выделенная клетка состоит из клетки M.smegmatis, трансформированной с помощью мутантного микобактериального белка atpE, в частности трансформированную с помощью мутантного микобактериального белка atpE, где мутация состоит, по меньшей мере, из одной точечной мутации, расположенной в любой из аминокислот 20-40, в частности 30-40, предпочтительно, в аминокислоте 34 или в аминокислотах 60-75, в частности 62-73, предпочтительно, в аминокислоте 69, как показано в сравнении последовательностей на фиг.2.
Способы скрининга
Кроме того, настоящее изобретение обеспечивает способ идентификации антимикробного соединения, включающий стадии
(a) контактирования клетки, экспрессирующей белок atpE, с исследуемым соединением в физиологических условиях;
(b) определения того, взаимодействует ли исследуемое соединение с белком atpE.
В одном из вариантов осуществления белок atpE, используемый в указанном выше способе, состоит из бактериального белка atpE, в частности микобактериального белка, и, как предполагается, включает как белки atpE дикого типа, так и также мутантные белки atpE, как описано здесь выше. В дополнительном варианте осуществления настоящего изобретения, микобактериальный белок atpE, используемый в указанном выше способе, состоит из мутантного микобактериального белка atpE в соответствии с настоящим изобретением. В конкретном варианте осуществления указанного выше анализа используется клетка-хозяин, трансформированная с помощью мутантного белка atpE по настоящему изобретению, и взаимодействие исследуемого соединения с указанным белком atpE оценивается путем определения возможного ингибирования ферментативной активности F1F0-ATP-азы, содержащей указанный мутантный белок atpE. Ингибирование активности F1F0-ATP-азы определяется с использованием методик, известных в данной области, например добавлением вещества в систему, содержащую F1F0-ATP-азу и ATP, в качестве субстрата, с детектированием ферментативной активности путем объединения продуцирования ADP с окислением NADH посредством реакций пируваткиназы и лактатгидрогеназы.
В одном из вариантов анализа, белок atpE может использоваться в анализе связывания. Анализы связывания могут быть конкурентными или неконкурентными. Такой анализ может быть приспособлен для быстрого скрининга большого количества соединений, для определения того, какие соединения способны связываться с полипептидами, если они есть.
В этом контексте, настоящее изобретение предусматривает способ идентификации того, связывается ли исследуемое соединение с выделенным белком atpE по настоящему изобретению, и таким образом, является ли оно потенциальным антимикробным соединением, причем указанный способ включает:
a) контактирование клеток, экспрессирующих белок atpE, где такие клетки обычно не экспрессируют указанный белок atpE, с исследуемым соединением, в присутствии и в отсутствие соединения, о котором известно, что оно связывает белок atpE,
b) определение связывания исследуемого соединения с белком atpE, с использованием соединения, о котором известно, что оно связывает белок atpE, в качестве эталона.
Связывание исследуемого соединения или соединения, о котором известно, что оно связывает белок atpE, упоминаемого также ниже как эталонное соединение, оценивается с использованием известных в данной области способов исследования взаимодействий белок-лиганд. Например, такое связывание может быть измерено посредством использования меченого вещества или эталонного соединения. Исследуемое соединение или эталонное соединение, в частности соединение J (фиг.1), можно метить любым удобным способом, известным в данной области, например радиоактивно, флуоресцентно или ферментативно. В конкретном варианте осуществления указанного выше способа, соединение, о котором известно, что оно связывает белок atpE, также известное как эталонное соединение, метят для детектирования и указанная метка используется для определения связывания исследуемого соединения с белком atpE. Указанное эталонное соединение метят, используя радиоактивную метку, флуоресцентную метку или ферментативную метку, более предпочтительно, радиоактивную метку.
В альтернативном варианте осуществления настоящего изобретения, указанные выше анализы связывания осуществляют на клеточной композиции, то есть на клеточном экстракте, клеточной фракции или органеллах клеток, содержащих белок atpE, как определено выше. Более конкретно, указанные выше анализы связывания осуществляют на клеточной композиции, то есть на мембранном препарате, содержащем белок atpE, как определено здесь, где указанная клеточная композиция, то есть мембранный препарат, получен из клеток M.smegmatis, трансформированных с помощью мутантного микобактериального белка atpE, в частности трансформированных с помощью мутантного микобактериального белка atpE, где мутация состоит, по меньшей мере, из одной точечной мутации, расположенной в одной из аминокислот 20-40, в частности 30-40, предпочтительно, в аминокислоте 34 или в аминокислотах 60-75 в частности, 62-73, предпочтительно, в аминокислоте 69, как показано в сравнении последовательностей на фиг.2. Принимая нумерацию Mtb_S (SEQ ID No.1) или Mtb_R (SEQ ID No.2) в качестве эталона, указанные выше области соответствуют аминокислотам 14-34, в частности 24-34, предпочтительно, в аминокислоте 28 или аминокислотах 54-69, в частности 56-67, предпочтительно, в аминокислоте 63.
В одном из вариантов осуществления анализы связывания осуществляют с использованием мембранных препаратов. Эти мембранные препараты могут использоваться в обычных анализах связывания на фильтре (например, с использованием оборудования для анализа на фильтре Brandel) или при высокопроизводительных анализах связывания типа метода близости сцинтилляций (технология SPA и Cytostar-T flashplate; Amersham Pharmacia Biotech) для детектирования связывания радиоактивно-меченных лигандов atpE (включая 3H-меченый DARQ) и замещения таких радиоактивно-меченных лигандов конкурентами за сайт связывания. Радиоактивность может быть измерена с помощью Packard Topcount или сходного инструмента, способного к осуществлению быстрых измерений в форматах 96-, 384-, 1536-луночных микротитровальных планшетов. Технология SPA/Cytostar-T является особенно пригодной для высокопроизводительного скрининга, и по этой причине эта технология является подходящей для использования при скрининге соединений, способных к замещению стандартных лигандов.
Другой подход к исследованию связывания лигандов с белком atpE в окружающей среде, приблизительно воспроизводящей нативную ситуацию, использует эффект поверхностного плазменного резонанса, применяемый инструментом Biacore (Biacore). Белок atpE в мембранных препаратах или в цельных клетках может присоединяться к биосенсорному чипу Biacore, и связывание лигандов исследуется в присутствии и в отсутствие соединений для идентификации конкурентов за сайт связывания.
Молекулярное моделирование
Кроме того, настоящее изобретение предусматривает способ оценки потенциала исследуемого соединения для взаимодействия с белком atpE, включающий:
(a) использование методов молекулярного моделирования для составления трехмерной структуры белка atpE;
(b) использование компьютерных средств для осуществления операции совмещения исследуемого соединения и трехмерной структуры белка atpE; и
(c) анализ результатов указанной операции совмещения для количественного определения ассоциации исследуемого соединения с трехмерной структурой белка atpE.
Методы молекулярного моделирования известны в данной области и включают как аппаратное обеспечение, так и программное обеспечение, пригодное для создания и использования моделей рецепторов и конформаций ферментов.
Многочисленные компьютерные программы являются доступными и подходящими для способов компьютерного моделирования, построения моделей и компьютерной идентификации, выбора и оценки потенциальных соединений, взаимодействующих с atpE, в способах, описанных здесь. Они включают, например, GRID (доступную от Oxford University, UK), MCSS (доступную от Accelrys, Inc., San Diego, CA), AUTODOCK (доступную от Oxford Molecular Group), FLEX X (доступную от Tripos, St Louis. MO), DOCK (доступную от University of California, San Francisco, CA), CAVEAT (доступную от University of California, Berkeley), HOOK (доступную от Accelrys, Inc., San Diego, CA) и системы 3-мерных баз данных, такие как MACCS-3D (доступную от MDL Information Systems, San Leandro, CA), UNITY (доступную от Tripos, St. Louis. MO) и CATALYST (доступную от Accelrys, Inc., San Diego, CA). Потенциальные вещества-кандидаты могут быть также сконструированы с помощью компьютера "de novo" с использованием пакетов программного обеспечения, таких как LUDI (доступного от Biosym Technologies, San Diego, CA), LEGEND (доступного от Accelrys, Inc, San Diego, CA) и LEAPFROG (доступного от Tripos, St. Louis. MO). Энергия деформации соединения и электростатическое отталкивание могут анализироваться с использованием таких программ, как GAUSSIAN 92, AMBER, QUANTA/CHARMM и INSIGHT H/DISCOVER. Эти методы компьютерной оценки и моделирования могут осуществляться на любом соответствующем аппаратном обеспечении, включая, например, рабочие станции, доступные от Silicon Graphics, Sun Microsystems, и другие. Эти методы моделирования, методы, аппаратное обеспечение и пакеты программного обеспечения являются репрезентативными и не предназначены в качестве полного их перечисления. Другие методы моделирования, известные в данной области, также могут использоваться в соответствии с настоящим изобретением. См., например, N.C. Cohen, Molecular Modeling in Drug Design, Academic Press (1996).
В одном из вариантов осуществления настоящего изобретения трехмерная структура белка atpE генерируется с использованием атомных координат Ile28, Glu61 и Ile63 E. coli (Protein Database 1Q01) +/- среднеквадратичное отклонение атомов основной цепи указанных аминокислот не более чем 10 Å, предпочтительно не более чем 5 Å.
Как приведено далее в примерах, целью настоящего изобретения является создание трехмерной структуры белка atpE. В таблицах 6 и 7 приводятся атомные координаты для белка atpE мутантного и дикого типа с SEQ ID No.2 и SEQ ID No.1. Таким образом, в одном из вариантов осуществления трехмерная структура белка atpE генерируется с использованием атомных координат из таблиц 6 или 7. В конкретном варианте осуществления трехмерная структура белка atpE генерируется с использованием атомных координат из таблицы 7. Соединение DARQ J ингибирует взаимодействие Arg186 субъединицы A с Glu61 субъединицы C в его депротонированной форме. Соответственно целью настоящего изобретения является обеспечение использования атомных координат из таблиц 6 или 7 в способе оценки потенциала исследуемого соединения к взаимодействию с белком atpE.
Сайт связывания
В другом варианте осуществления настоящего изобретения предусматривается характеризация сайта связывания в части F0 ATP-азы. Этот сайт связывания, идентифицируемый как способный к связыванию с соединением DARQ J, как обнаружено, совпадает с областями, идентифицированными до этого, как сайты придающих устойчивость мутаций в белках atpE M.tuberculosis и M.smegmatis (17). Следовательно, настоящее изобретение предоставляет сайт связывания в части F0 ATP-азы, отличающийся тем, что он содержит сайты придающих устойчивость мутаций белка atpE. Сайты придающих устойчивость мутаций, как здесь используется, относятся к аминокислотам 14-34, в частности 24-34, и к аминокислотам 53-69, в частности 56-67 белка atpE, принимая нумерацию Mtb_S (SEQ ID No.1) или Mtb_R (SEQ ID No.2) в качестве эталона.
В дополнительном варианте осуществления сайт связывания содержит, по меньшей мере, аминокислоты Ala24, Gly27, Phe53, Val57, Gly58, Glu61, Tyr64 и Phe65 одной субъединицы C и аминокислоты Ser182, Leu183, Leu185 и Arg186 одной субъединицы A (имеющие для субъединицы A, коды Ser 206 - Leu 207-Leu 209 и Arg 210 в таблицах 3, 4 и 5), где указанные аминокислоты имеют атомные координаты одной из таблиц 3, 4 или 5, или гомологичные структурные координаты, содержащие среднеквадратичное отклонение атомов, отличных от водорода, меньше, чем примерно 1,5 Å, предпочтительно, не более чем 0,75 Å, когда накладываются на положения соответствующих атомных координат атомов, отличных от водорода, из таблиц 3, 4 или 5. В конкретном варианте осуществления сайт связывания содержит аминокислоты Ala21, Gly25 первой субъединицы C; аминокислоты Ala24, Gly27, Phe53, Phe54, Val57, Gly58, Glu61, Tyr64, Phe65 второй субъединицы C; аминокислоты Met17, Gly19, Gly20, Ala21, Ile22, Gly23, Ala24, Gly25, Ile26, Gly27, Asp28, Gly29, Ala31, Phe53, Thr56, Val57, Gly58, Leu59, Val60, Glu61, Ala62, Ala63/Pro63, Tyr54, Phe65 третьей субъединицы C и аминокислоты Leu183, Leu185 и Arg186 субъединицы A; где указанные аминокислоты имеют атомные координаты любой из таблиц 3, 4 или 5, или гомологичные структурные координаты, содержащие среднеквадратичное отклонение для атомов, отличных от водорода, меньшее, чем примерно 1,5 Å, предпочтительно не более чем 0,75 Å, когда накладываются на положение соответствующих атомных координат для атомов, отличных от водорода, из таблиц 3, 4 или 5. В еще более конкретном варианте осуществления сайт связывания состоит из аминокислот Ala21, Gly25 первой субъединицы C; аминокислот Ala24, Gly27, Phe53, Phe54, Val57, Gly58, Glu61, Tyr64, Phe65 второй субъединицы C; аминокислот Met17, Gly19, Gly20, Ala21, Ile22, Gly23, Ala24, Gly25, Ile26, Gly27, Asp28, Gly29, Ala31, Phe53, Thr56, Val57, Gly58, Leu59, Val60, Glu61, Ala62, Ala63/Pro63, Tyr64, Phe65 третьей субъединицы C и аминокислот Leu183, Leu185 и Arg186 субъединицы A; где указанные аминокислоты имеют атомные координаты любой из таблиц 3, 4 или 5. В наиболее конкретном варианте осуществления сайт связывания состоит из аминокислот Ala21, Gly25 первой субъединицы C; аминокислот Ala24, Gly27, Phe53, Phe54, Val57, Gly58, Glu61, Tyr64, Phe65 второй субъединицы C; аминокислот Met17, Gly19, Gly20, Ala21, Ile22, Gly23, Ala24, Gly25, Ile26, Gly27, Asp28, Gly29, Ala31, Phe53, Thr56, Val57, Gly58, Leu59, Val60, Glu61, Ala62, Ala63/Pro63, Tyr64, Phe65 третьей субъединицы C и аминокислот Leu183, Leu185 и Arg186 субъединицы A; где указанные аминокислоты имеют атомные координаты из таблицы 3.
Соответственно, целью настоящего изобретения является оценка потенциала исследуемого соединения относительно взаимодействия с белком atpE с использованием атомных координат, как описывается выше, в компьютерных программах скрининга. В одном из вариантов осуществления настоящее изобретение обеспечивает способ оценки потенциала исследуемого соединения относительно взаимодействия с белком atpE, включающий методы молекулярного моделирования для генерирования трехмерной структуры сайта связывания части F0 ATP-азы использование компьютерных средств для осуществления операций совмещения исследуемого соединения и трехмерной структуры сайта связывания и анализ результатов указанной операции совмещения для количественного определения ассоциации исследуемого соединения с трехмерной структурой сайта связывания. В дополнительном варианте осуществления настоящего изобретения трехмерная структура сайта связывания генерируется с использованием атомных координат из таблиц 3, 4 или 5, или гомологичных структурных координат, содержащих среднеквадратичное отклонение для атомов, отличных от водорода, меньше, чем примерно 1,5 Å, предпочтительно не более чем 0,75 Å, когда накладываются на положение соответствующих атомных координат для атомов, отличных от водорода, из таблиц 3, 4 или 5. В конкретном варианте осуществления трехмерная структура генерируется с использованием атомных координат аминокислот Ala21, Gly25 цепи A любой из таблиц 3, 4 или 5; аминокислот Ala24, Gly27, Phe53, Phe54, Val57, Gly58, Glu61, Tyr64, Phe65 цепи K любой из таблиц 3, 4 или 5; аминокислот Met17, Gly19, Gly20, Ala21, Ile22, Gly23, Ala24, Gly25, Ile26, Gly27, Asp28, Gly29, Ala31, Phe53, Thr56, Val57, Gly58, Leu59, Val60, Glu61, Ala62, Ala63/Pro63, Tyr64, Phe65 цепи L любой из таблиц 3, 4 или 5; и аминокислот Ser206, Leu207, Leu207 и Arg210 цепи M любой из таблиц 3, 4 или 5.
При этом скрининге качество совмещения таких соединений с сайтом связывания может оцениваться либо по комплементарности формы, либо по оцениваемой энергии взаимодействия (Meng, E.C. et al., J. Coma. Chem 13:505-524 (1992)).
Использование сайта связывания
Конструирование соединений, которые связываются с atpE, усиливают или ингибируют его функциональную активность в соответствии с настоящим изобретением, как правило, включает рассмотрение двух факторов. Во-первых, соединение должно быть способным к физическому и структурному ассоциированию с atpE. Нековалентные молекулярные взаимодействия, важные при ассоциации atpE с соединением, включают водородную связь, Ван-дер-ваальсовы и гидрофобные взаимодействия. Во-вторых, соединение должно быть способным, предположительно, иметь конформацию, которая даст ему возможность ассоциации с atpE. Хотя определенные части соединения могут непосредственно не участвовать в ассоциации с atpE, эти части могут по-прежнему влиять на общую конформацию молекулы. Это, в свою очередь, может оказывать значительное воздействие на сродство при связывании, терапевтическую эффективность, качества, сходные с лекарственными средствами, и активность. Такие конформационные требования включают общую трехмерную структуру и ориентацию химического остатка или соединения по отношению к активному сайту в целом или к его части, или к другой области atpE, или расстояние между функциональными группами соединения, содержащего несколько химических остатков, которые непосредственно взаимодействуют с atpE.
Потенциал, предсказанный эффект ингибиторного агониста, антагониста или связывания лиганда или другого соединения на atpE, может анализироваться до его реального синтеза и исследования посредством использования методов компьютерного моделирования. Если теоретическая структура данного соединения говорит о недостаточном взаимодействии и ассоциации между ним и atpE, синтез и исследование соединения могут быть отменены. Однако если компьютерное моделирование указывает на сильное взаимодействие, молекула может быть затем синтезирована и исследована на ее способность к взаимодействию с atpE. Таким образом, синтез неработающих соединений может быть исключен. В некоторых случаях, синтезируются неактивные соединения, которые могут быть предсказаны при моделировании, а затем исследованы для развития SAR (соотношения структура-активность) для соединений, взаимодействующих с конкретной областью atpE. Специалист в данной области может использовать один из нескольких способов скрининга химических остатков, фрагментов, соединений или агентов на их способность к ассоциации с atpE, а более конкретно, с индивидуальными карманами связывания или активными сайтами atpE. Этот способ может начинаться, например, визуальным просмотром активного сайта на экране компьютера, на основе атомных координат atpE или комплекса atpE и лиганда. Выбранные химические остатки, соединения или агенты могут затем позиционироваться во множестве ориентаций или устанавливаться внутри индивидуального кармана связывания atpE. Установка может осуществляться с использованием такого программного обеспечения, как Quanta и Sybyl, с последующей минимизацией энергии и молекулярной динамикой со стандартными силовыми полями молекулярной механики, такой как CHARMM и AMBER.
Специализированные компьютерные программы могут также помочь в процессе выбора химических остатков. Они включают, но не ограничиваются этим: GRID (Goodford, P. J., "A Computational Procedure for Determining Energetically Favorable Binding Sites on Biologically Important Macromolecules," J. Med. Chem. 28:849-857 (1985), доступный от Oxford University, Oxford, UK); MCSS (Miranker, A. and M. Karplus, "Functionality Maps of Binding Sites: A Multiple Copy Simultaneous Search Methods." Proteins: Structure, Function and Genetics 11: 29-34 (1991), доступный от Molecular Simulations, Burlington, Mass.); AUTODOCK (Goodsell, D. S. and A. J. Olsen, "Automated Docking of Substrates to Proteins by Simulated Annealing" Proteins: Structure, Function, and Genetics 8:195-202 (1990), доступный от Scripps Research Institute, La Jolla, Calif.); и DOCK (Kuntz, I. D. et al., "A Geometric Approach to Macromolecule-Ligand Interactions," J.-Mol. Biol. 161:269-288 (1982), доступный от University of California, San Francisco, Calif.).
Использование такого программного обеспечения как GRID, программы, которая определяет вероятные сайты взаимодействия между зондами с различными характеристиками функциональных групп и макромолекулярной поверхностью, применяется для анализа поверхностных сайтов с целью определения структур сходных ингибирующих белков или соединений. Вычисления GRID с соответствующими ингибирующими группами на молекулах (например, протонированными первичными аминами), в качестве зонда, используются для идентификации потенциальных активных центров вокруг доступных положений на соответствующих контурных уровнях энергии. Программа DOCK может использоваться для анализа активного сайта или сайта связывания лиганда и предложить лиганды с комплементарными стерическими свойствами.
После выбора соответствующих химических остатков, соединений, или агентов они могут быть собраны в единый лиганд или соединение, или ингибитор, или активатор. Сборка может осуществляться путем визуальной проверки соотношения фрагментов друг с другом на трехмерном изображении. За этим может следовать построение модели вручную с использованием такого программного обеспечения, как Quanta или Sybyl.
Используемые программы для помощи при соединении индивидуальных химических остатков, соединений или агентов включают, но не ограничиваются этим: CAVEAT (Bartlett, P.A. et al., "CAVEAT: A Program to Facilitate Structure-Derived Design of Biologically Active Molecules." In Molecular Recognition in Chemical and Biological Problems, Special Pub., Royal Chem. Soc., 78, pp. 82-196 (1989)); системы 3D Database, такие как MACCS-3D (MDL Information Systems, San Leandro, Calif. and Martin, Y.C., "3D Database Searching in Drug Design", J. Med. Chem. 35: 2145-2154 (1992); и HOOK (доступные от Molecular Simulations, Burlington, Mass.).
Доступными являются несколько методологий для поиска трехмерных баз данных для исследования гипотетических фармакофоров и выбора соединений для скрининга. Они включают программу CAVEAT (Bacon et al., J. Mol. Biol. 225:849-858 (1992)). Например, CAVEAT использует базы данных циклических соединений, которые могут действовать в качестве "спейсеров" для соединения любого количества химических фрагментов, уже расположенных в активном сайте. Это позволяет специалисту в данной области быстро генерировать сотни возможных путей для соединения фрагментов, которые уже известны или предположительно рассматриваются как необходимые для прочного связывания. Вместо осуществления конструирования ингибитора активатора, агониста или антагониста atpE в виде последовательности шагов, по одному химическому остатку за раз, как описано выше, такие соединения могут быть сконструированы в целом или "de novo" с использованием либо пустого активного сайта, либо с необязательным включением некоторой части (частей) известных молекул. Эти способы включают LUDI (Bohm, H.-J., "The Computer Program LUDI: A New Method for the De Novo Design of Enzyme Inhibitors", J. ComR. Aid. Molec. Design, 6, pp. 61-78 (1992), доступный от Biosym Technologies, San Diego, Calif.); LEGEND (Nishibata, Y. and A. Itai, Tetrahedron 47:8985 (1991), доступный от Molecular Simulations, Burlington, Mass.); и LeapFrog (доступный от Tripos Associates, St. Louis, Mo.). Например, программа LUDI может определить список сайтов взаимодействия, в который нужно включать как водородные связи, так и гидрофобные фрагменты. Затем LUDI использует библиотеку линкеров для соединения до четырех различных сайтов взаимодействия во фрагментах. Затем "мостиковые" группы меньшего размера, такие как -CH2- и -COO-, используются для соединения этих фрагментов. Например, для фермента DHFR размещение ключевых функциональных групп в хорошо известном ингибиторе метотрексате воспроизводится посредством LUDI. См. также Rotstein and Murcko, J. Med. Chem. 36: 1700-1710 (1992).
Другие методы молекулярного моделирования также могут использоваться в соответствии с настоящим изобретением. См., например, Cohen, N.C. et al., "Molecular Modeling Software and Methods for Medicinal Chemistry, J. Med. Chem. 33:883-894 (1990). См. также Navia, M.A. and M. A. Murcko, "The Use of Structural Information in Drug Design," Current Opinions in Structural Biology, 2, pp. 202-210 (1992).
После того как соединение конструируется или выбирается указанными выше способами, сродство, с которым это соединение может связываться или ассоциироваться с atpE, может быть исследовано и оптимизировано путем компьютерной оценки и/или путем исследования биологической активности после синтеза соединения. Ингибиторы или соединения могут взаимодействовать с atpE в нескольких конформациях, которые являются сходными по общей энергии связывания. В этих случаях энергия деформации связи принимается как разность между энергией свободного соединения и средней энергией конформации, наблюдаемых, когда соединение связывается с atpE.
Соединение, сконструированное или выбранное в качестве связывающегося или ассоциирующегося с atpE, может быть дополнительно оптимизировано с помощью компьютера, так что в своем связанном состоянии оно должно бы предпочтительно не иметь отталкивающего электростатического взаимодействия с atpE. Такие некомплементарные (например, электростатические) взаимодействия включают отталкивающие взаимодействия заряд-заряд, диполь-диполь и заряд-диполь. Конкретно, сумма всех электростатических взаимодействий между ингибитором и atpE, когда ингибитор является связанным, предпочтительно делает нейтральным или благоприятным вклад в энтальпию связывания. Слабо связывающиеся соединения также будут конструироваться этими способами с тем, чтобы определить SAR. См., например, заявки на патент США №№ 60/275629; 60/331235; 60/379617 и 10/097249.
Специальное компьютерное программное обеспечение является доступным в данной области для оценки энергии деформации соединения и электростатического взаимодействия. Примеры программ, созданных для таких применений, включают Gaussian 92, revision C (M. J. Frisch, Gaussian, Inc., Pittsburgh, Pa., COPYRGT 1992); AMBER, version 4.0 (P. A. Kollman, University of California at San Francisco, COPYRGT 1994); QUANTA/CHARMM (Molecular Simulations, Inc., Burlington, Mass. COPYRGT 1994); и Insight II/Discover (Biosysm Technologies Inc., San Diego, Calif. COPYRGT 1994). Другие системы аппаратного обеспечения и пакеты программного обеспечения будут известны специалистам в данной области.
После того как соединение, которое ассоциируется с atpE, выбирается или конструируется оптимальным образом, как описано выше, затем могут быть сделаны замены в некоторых из его атомов или боковых групп для улучшения или модификации его свойств связывания. Как правило, начальные замещения являются консервативными, то есть группа-заместитель будет иметь приблизительно такой же размер, форму, гидрофобность и заряд, как и исходная группа. Разумеется, необходимо понять, что компоненты, известные в данной области как изменяющие конформацию, могут быть исключены. Такие замещенные химические соединения затем можно анализировать на эффективность совмещения с atpE такими же компьютерными способами, подробно описанными выше.
Кроме того, настоящее изобретение предоставляет системы, в частности системы на основе компьютера, которые содержат последовательность и/или структурные координаты, описанные здесь. Такие системы конструируются для определения структуры и для рациональной разработки лекарственных средств для atpE или для сайта связывания в части F0 ATP-азы. Системы на основе компьютера относятся к средствам аппаратного обеспечения, средствам программного обеспечения и средствам хранения данных, используемых для анализа последовательности и/или структурных координат по настоящему изобретению в любом из компьютерных способов, подробно описанных выше. Минимальные средства аппаратного обеспечения системы на основе компьютера по настоящему изобретению включают узел центрального процессора (CPU), средства ввода, средства вывода и средства хранения данных. Специалист в данной области легко определит, какие именно из доступных в настоящее время систем на основе компьютера являются пригодными для использования в настоящем изобретении.
Соответственно, целью настоящего изобретения является создание считываемой компьютером среды для хранения данных, содержащей структурные координаты, описанные здесь. Как здесь используется, "считываемая компьютером среда для хранения данных" относится к любой среде, которая может считываться компьютером или может быть доступной непосредственно с его помощью. Такие среды включают, но не ограничиваются этим: магнитные среды для хранения данных, такие как гибкие диски, среды для хранения данных на основе твердых дисков и магнитную ленту; оптические среды для хранения данных, такие как оптические диски или CD-ROM; электрические среды для хранения данных, такие как RAM и ROM; и гибриды этих сред, такие как магнитные/оптические среды для хранения данных.
Способы лечения
Как уже рассматривалось выше, целью настоящего изобретения является также обеспечение применения соединений, идентифицированных с использованием любого из указанных выше способов скрининга, в способе лечения субъекта с инфекцией, вызванной микробами. Как правило, бактериальные патогены могут классифицироваться либо как грамположительные, либо как грамотрицательные патогены. Антимикробные соединения с активностью, как против грамположительных, так и против грамотрицательных патогенов, как правило, рассматриваются как имеющие широкий спектр активности. Соединения по настоящему изобретению рассматриваются как активные против грамположительных и/или грамотрицательных бактериальных патогенов. В частности, настоящие соединения являются активными, по меньшей мере, против одной грамположительной бактерии, предпочтительно, против нескольких грамположительных бактерий, более предпочтительно, против одной или нескольких грамположительных бактерий и/или одной или нескольких грамотрицательных бактерий.
Примеры грамположительных и грамотрицательных аэробных и анаэробных бактерий включают Staphylococci, например S.aureus; Enterococci, например E.faecalis; Streptococci, например S.pneumoniae, S.mutans, S.pyogens; Bacilli, например Bacillus subtilis; Listeria, например Listeria monocytogenes; Haemophilus, например H.influenza; Moraxella, например M.catarrhalis; Pseudomonas, например Pseudomonas aeruginosa; и Escherichia, например E.coli. Грамположительные патогены, например Staphylococci, Enterococci и Streptococci, являются особенно важными из-за развития стойких штаммов, которые как сложно лечить, так и сложно ликвидировать, например, в окружающей среде больницы, после распространения. Примеры таких штаммов представляют собой устойчивый к метициллину Staphylococcus aureus (MRSA), устойчивый к метициллину коагулазо-отрицательный стафилококк (MRCNS), устойчивый к пенициллину Streptococcus pneumoniae и устойчивый к множеству лекарственных средств Enterococcus faecium.
Соединения по настоящему изобретению также проявляют активность против устойчивых бактериальных штаммов.
Соединения по настоящему изобретению являются особенно активными против тех бактерий, у которых выживаемость зависит от правильного функционирования F1F0 ATP-синтазы. Не ограничиваясь какой-либо теорией, считается, что активность настоящих соединений заключается в ингибировании F1F0 ATP-синтазы, в частности в ингибировании комплекса F0 F1F0 ATP-синтазы, более конкретно, в ингибировании переноса протона от Arg186 субъединицы A к Glu61 субъединицы C комплекса F0 F1F0 ATP-синтазы, что приводит к уничтожению бактерий посредством истощения клеточных уровней ATP бактерий. Соединения, идентифицированные с использованием любого из указанных выше способов скрининга, являются особенно активными против грамположительных бактерий, более конкретно, микобактерий, и наиболее конкретно, против инфекций, вызываемых M.africanum, M.avium, M.bovis, M.bovis-BCG, M.chelonae, M.fortuitum, M.gordonae, M.intracellulare, M.kansasii, M.microti, M.scrofulaceum, M.paratuberculosis, M.leprea, M.tuberculosis, M.ulcerans и M.ranae.
Используется ли выше или ниже то, что соединения могут излечивать бактериальную инфекцию, при этом подразумевается, что соединения могут излечивать инфекцию, вызванную одним или несколькими бактериальными штаммами. Когда это используется, однако, в отношении использования DARQ J в качестве антимикробного соединения, антимикробный агент, как подразумевается, представляет собой соединение, которое может лечить инфекцию, вызванную одним или несколькими бактериальными штаммами, при условии, что указанные бактериальные штаммы являются иными, чем микобактерия.
Бактериальные инфекции, которые могут лечиться с помощью настоящих соединений, включают, например, инфекции центральной нервной системы, инфекции наружного уха, инфекции среднего уха, такие как острый отит среднего уха, инфекции краниальных синусов, инфекции глаз, инфекции ротовой полости, такие как инфекции зубов, десен и слизистой, инфекции верхних дыхательных путей, инфекции нижних дыхательных путей, инфекции мочеполовой системы, инфекции желудочно-кишечного тракта, гинекологические инфекции, септицемию, инфекции костей и суставов, инфекции кожи и структуры кожи, бактериальный эндокардит, ожоги, антибактериальную профилактику при хирургическом вмешательстве и антибактериальную профилактику у пациентов с ослабленным иммунитетом, таких как пациенты, принимающие раковую химиотерапию, или пациенты с трансплантированными органами.
Кроме того, настоящее изобретение предусматривает способ лечения субъекта, страдающего туберкулезом, включающий введение субъекту агента, который взаимодействует с белком atpE.
Фармацевтические композиции
Кроме того, настоящее изобретение обеспечивает фармацевтическую композицию, содержащую агент, который взаимодействует с белком atpE в клетке, и фармацевтически приемлемый носитель.
Такие агенты могут быть составлены в виде композиций, содержащих агент вместе с фармацевтически приемлемым носителем или разбавителем. Агент может находиться в форме физиологически функционального производного, такого как сложный эфир или соль, такая как кислотно-аддитивная соль или соль неблагородного металла, или N или S оксид. Композиции могут быть составлены для любого подходящего пути и средств введения. Фармацевтически приемлемые носители или разбавители включают те, которые используются в препаратах, подходящих для перорального, ректального, назального, ингаляционного, местного (включая буккальное и сублингвальное), вагинального или парентерального (включая подкожное, внутримышечное, внутривенное, чрескожное, интратекальное и эпидуральное) введения. Выбор носителя или разбавителя будет, разумеется, зависеть от предлагаемого пути введения, который может зависеть от агента и его терапевтической цели. Удобно, чтобы препараты были представлены в виде стандартной дозированной формы и могли быть приготовлены любым из способов, хорошо известных в области фармации. Такие способы включают стадию приведения в ассоциацию активного ингредиента с носителем, который состоит из одного или нескольких вспомогательных ингредиентов. Как правило, препараты получают однородным и тесным приведением в ассоциацию активного ингредиента с жидкими носителями или мелкодисперсными твердыми носителями, или как с теми, так и с этими, а затем, если это необходимо, формованием продукта.
Для твердых композиций могут использоваться обычные нетоксичные твердые носители, включая, например, фармацевтические сорта маннита, лактозы, целлюлозы, производных целлюлозы, крахмала, стеарата магния, сахарина натрия, талька, глюкозы, сахарозы, карбоната магния и тому подобное. Активное соединение, как определено выше, может быть составлено в виде суппозиториев, с использованием, например, полиалкиленгликолей, ацетилированных триглицеридов и тому подобное, в качестве носителя. Жидкие фармацевтически вводимые композиции могут быть получены, например, растворением, диспергированием и тому подобное активного соединения, как определено выше, и необязательных фармацевтических вспомогательных веществ в носителе, таком как, например, вода, водный солевой раствор с декстрозой, глицерин, этанол и тому подобное, чтобы тем самым образовать раствор или суспензию. Если это желательно, фармацевтическая композиция, которая должна вводиться, может также содержать минорные количества нетоксичных вспомогательных веществ, таких как смачивающие или эмульгирующие агенты, буферизуюшие pH агенты и тому подобное, например ацетат натрия, монолаурат сорбитана, триэтаноламин натрий ацетат, монолаурат сорбитана, олеат триэтаноламина и тому подобное. Реальные способы получения таких дозированных форм являются известными или будут очевидны специалистам в данной области; см., например, Gennaro et al., Remington's Pharmaceutical Sciences, Mack Publishing Company, Easton, Pennsylvania, 18th Edition, 1990.
Композиция или препарат, которые должны вводиться, будут в каждом случае содержать некоторое количество активного соединения (соединений) в количестве, эффективном для ослабления симптомов у субъекта, подвергаемого лечению.
Точная дозировка и частота введения настоящих соединений зависит от конкретного используемого соединения, конкретного состояния, которое лечится, тяжести заболевания, которое лечится, возраста, массы, пола, диеты, времени введения и общего физического состояния конкретного пациента, способа введения, а также другого медицинского препарата, который индивидуум может принимать, как хорошо известно специалистам в данной области. Кроме того, ясно, что эффективное ежедневное количество может быть снижено или повышено в зависимости от реакции излечиваемого субъекта и/или в зависимости от оценки врача, предписывающего соединения по настоящему изобретению.
Могут быть приготовлены дозированные формы или композиции, содержащие активный ингредиент в пределах от 0,25 до 95%, при этом остаток состоит из нетоксичного носителя. В зависимости от способа введения, фармацевтическая композиция предпочтительно будет состоять из 0,05-99% мас., более предпочтительно от 0,1 до 70% мас. активных ингредиентов, и от 1 до 99,95% мас., более предпочтительно от 30 до 99,9% мас. фармацевтически приемлемого носителя, все проценты представлены в расчете на общую композицию.
Для перорального введения фармацевтически приемлемая нетоксичная композиция формируется посредством введения любого из обычно используемых эксципиентов, таких как, например, фармацевтические сорта маннита, лактозы, целлюлозы, производных целлюлозы, кроскармеллозы натрия, крахмала, стеарата магния, сахарина натрия, талька, глюкозы, сахарозы, карбоната магния и тому подобное. Такие композиции принимают форму растворов, суспензий, таблеток, пилюль, капсул, порошков, препаратов с замедленным высвобождением и тому подобное. Такие композиции могут содержать 1%-95% активного ингредиента, более предпочтительно 2-50%, наиболее предпочтительно 5-8%.
Парентеральное введение, как правило, характеризуется инъекцией, либо подкожно, внутримышечно, либо внутривенно. Препараты для инъекций могут быть получены в обычных формах, либо в виде жидких растворов или суспензий, твердых форм, подходящих для растворения или суспендирования в жидкости перед инъекцией, либо в виде эмульсий. Подходящие эксципиенты представляют собой, например, воду, солевой раствор, декстрозу, глицерин, этанол или тому подобное. В дополнение к этому, если это желательно, фармацевтические композиции, которые должны вводиться, могут также содержать минорные количества нетоксичных вспомогательных веществ, таких как смачивающие или эмульгирующие агенты, буферизующие pH агенты и тому подобное, такие как, например, ацетат натрия, монолаурат сорбитана, олеат триэтаноламина, триэтаноламин натрий ацетат и тому подобное.
Процент активного соединения, содержащегося в таких парентеральных композициях, сильно зависит от их конкретной природы, а также активности соединения и потребностей субъекта. Однако подходящими для использования являются проценты активного ингредиента от 0,1% до 10% в растворе, и они могут быть выше, если композиция представляет собой твердый продукт, который будет впоследствии разбавлен до указанных выше процентов. Предпочтительно, композиция будет содержать 0,2-2% активного агента в растворе.
Наконец, настоящее изобретение предоставляет готовое изделие, содержащее упаковку и фармацевтический агент, где (a) фармацевтический агент взаимодействует с белком atpE в клетке, и (b) упаковка содержит этикетку, указывающую на использование агента для лечения бактериальной инфекции у субъекта, в частности, в качестве антимикобактериального лекарственного средства.
В настоящем описании термины "стандартные способы", "стандартные протоколы" и "стандартные методики", когда используются в контексте методов молекулярной биологии, должны пониматься как протоколы и методики, находящиеся в обычном лабораторном справочнике, таком как Current Protocols in Molecular Biology, editors F. Ausubel et al., John Wiley and Sons, Inc. 1994, или Sambrook, J., Fritsch, E.F. and Maniatis, T., Molecular Cloning: A laboratory manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY 1989.
Настоящее изобретение будет лучше понято со ссылками на экспериментальные подробности, которые следуют далее, но специалисты в данной области легко заметят, что они представляют собой только иллюстрацию настоящего изобретения, как описано более подробно в формуле изобретения, которая следует ниже. В дополнение к этому, в настоящем описании цитируются различные публикации. Описание этих публикаций тем самым включается в качестве ссылок в настоящую заявку для более полного описания состояния области, к которой относится настоящее изобретение.
Экспериментальная часть
Используя Mycobacterium smegmatis в качестве примера, авторы обнаружили ряд DARQ с сильной активностью in vitro против нескольких микобактерий (11). К настоящему моменту 20 молекул ряда DARQ имеют минимальную ингибирующую концентрацию (MIC) ниже 0,5 мкг/мл против Mycobacterium tuberculosis H37Rv и для трех из этих антимикобактериальных агентов активность подтверждается на модели мышей in vivo.
Структурно и механистически DARQ сильно отличаются как от фторхинолонов (включая метоксихинолоны), так и от других классов хинолинов, включая мефлохин и его аналоги, 4-метилхинолины и 4-хинолилгидразоны (12-16). Одним из главных структурных различий между DARQ и другими классами хинолонов или хинолинов является специфичность функционализированной латеральной (3') цепи, порождаемой классом DARQ. В дополнение к этому, отсутствие микобактериальной перекрестной устойчивости с существующими химическими классами указывает на другой механизм действия.
Ведущее соединение DARQ, ниже упоминаемое как J или DARQ J (фиг.1), как обнаружено, имеет уникальный спектр сильной и селективной антимикобактериальной активности in vitro (таблица 1). Медианная MIC, полученная для лабораторного штамма H37Rv и шести полностью восприимчивых изолятов, составляет 0,060 мкг/мл, по сравнению с 1,00 мкг/мл для рифампина. J демонстрирует сходную эффективность in vitro против клинических изолятов M.tuberculosis, устойчивых к TB агентам первой линии рифампину, стрептомицину, этамбутолу и пиразинамиду; и TB агенту второй линии моксифлоксацину. Для восьми клинических изолятов, устойчивых к изониазиду, медианная MIC составляет 0,010 мкг/мл. Отсутствие перекрестной устойчивости с используемыми в настоящее время анти-TB агентами говорит о том, что J может сохранять активность против штаммов MDR-TB. На самом деле, при использовании системы культуры BACTEC™, четкое концентрационно-зависимое ингибирование бактериального роста видно, когда MDR-TB штаммы экспонируются для фиксированных концентраций J. Из 30 изолятов MDR-TB, 13 (43%), как обнаружено, являются восприимчивыми к 0,100 мкг/мл J и 17 (57%) являются восприимчивыми к 0,010 мкг/мл J. Схожий высокий уровень восприимчивости (MIC ниже 0,010 мкг/мл) виден только для одного из 10 дополнительных полностью восприимчивых к лекарственным средствам штаммов, когда они исследуются с использованием системы BACTEC™, в то время как все штаммы являются восприимчивыми к 0,100 мкг/мл J.
Сильная активность демонстрируется также против других микобактериальных видов, включая Mycobacterium bovis и Mycobacterium kansasii, а также видов, природно-устойчивых ко многим другим анти-TB агентам и вовлеченным в условно-патогенные инфекции, таких как комплекс Mycobacterium avium (MAC), Mycobacterium abcessus, Mycobacterium fortuitum и Mycobacterium marinum (таблица 1).
Неожиданно активность J, вероятно, является специфичной к микобактериям. J является слабо активным против видов, близких к микобактериям, таких как Corynebacterium (MIC 4,00 мкг/мл) и Nocardia (MIC>4,00 мкг/мл) и неактивным против других организмов, включая грамположительные Streptococcus pneumoniae, Staphylococcus aureus - включая устойчивые к метициллину штаммы (MIC>32 мкг/мл) - и Enterococcus faecalis, или грамотрицательные Escherichia coli, Haemophilus influenzae и Helicobacter pylori. Экспонирование M.tuberculosis в фазе логарифмического роста для концентраций J при 100 × MIC приводит к 103 логарифмическому уменьшению бактериальных отсчетов через 12 дней, указывая на то, что J имеет бактерицидную активность in vitro. Воздействие J на tubercle bacilli в стационарной фазе еще не исследовано.
Выделение мутантов, перекрестная устойчивость и постулированная мишень для лекарственных средств
При исследовании микобактериальной устойчивости авторы имели целью идентификацию молекулярной мишени лекарственного средства и установление вероятного механизма действия. Устойчивые мутанты of M.tuberculosis и M.smegmatis получают селекцией in vitro при ингибирующих концентрациях J для того, чтобы:
- количественно определить долю устойчивых мутантов в микобактериях (с рифампином в качестве контроля);
- оценить характер устойчивости для устойчивых мутантов (включая перекрестную/неперекрестную устойчивость к хинолонам);
- исследовать механизм действия.
Согласно селекционным экспериментам доля мутантов с пониженной чувствительностью к J составляет 5×10-7 и 2×10-8 при MIC×4, и 5×10-8, и 1×10-8 при MIC×8 для M.tuberculosis и M.smegmatis соответственно (поддерживая данный текст). В случае M.tuberculosis, эти доли сравнимы с долями мутантов, устойчивых к рифампину (от 10-7 до 10-8), и указывают на то, что природная устойчивость к J является редкой. В дополнение к этому, чувствительность штаммов M.tuberculosis, устойчивых к J, остается неизменной по отношению к анти-TB агентам изониазиду, рифампину, стрептомицину, амикацину, этамбутолу и моксифлоксацину. Дополнительный анализ мутантов M.tuberculosis и M.smegmatis с пониженной чувствительностью к J показывает, что нет мутаций в областях ДНК гиразы gyrA и gyrB, последовательностей, в которых обычно развивается устойчивость к хинолону. Это подтверждает, что молекулярная мишень для J отличается от мишени фторхинолонов.
Один из подходов к определению молекулярной мишени для J и вмешательству в механизм действия заключается в идентификации и сравнении вызывающих устойчивость мутаций в чувствительных и устойчивых штаммах M.tuberculosis и M.smegmatis. Геномы устойчивого штамма M.tuberculosis BK12 и двух устойчивых штаммов M.smegmatis R09 и R10, а также исходного M.smegmatis, секвенируются почти полностью. Авторы идентифицировали придающие устойчивость мутации путем сравнительного анализа геномных последовательностей чувствительных и устойчивых штаммов M.tuberculosis и M.smegmatis (фиг.2). Авторы показали, что только ген, затронутый во всех трех независимых мутантах, по сравнению с соответствующим исходным диким типом, кодирует для atpE часть субъединицы F0 ATP-синтазы. Это говорит о том, что atpE является ответственным за устойчивость к J в мутантных штаммах, указывая на то, что J ингибирует новую мишень M.tuberculosis, протонный насос ATP-синтазы.
Осуществляли исследование комплементарности, чтобы показать, что мутантный ген atpE является ответственным за устойчивость к J и, путем прямого вмешательства, что генный продукт atpE представляет собой мишень J в микобактериях. Принимая во внимание тот факт, что известно о том, что все гены оперона ATP-синтазы должны экспрессироваться координированным образом, то есть все гены, которые кодируют часть F0, должны экспрессироваться из одного и того же положения, авторы амплифицировали часть F0 оперона из устойчивого штамма M.smegmatis (D32V) и выбрали клоны, которые не приобретают дополнительных мутаций в процессе ПЦР. M.smegmatis дикого типа трансформируется с помощью плазмиды, содержащей выбранный таким образом фрагмент мутанта F0. Это делает клетки устойчивыми к J при MIC, практически идентичной MIC устойчивого штамма M.smegmatis R09 (D32V). В дополнение к этому, когда плазмида повторно выделяется из этих трансформантов и ген atpE секвенируется, он, как показано, имеет сохраняющуюся мутантную аллель (D32V).
Реальное воздействие DARQ J на продуцирование ATP в M.tuberculosis дополнительно демонстрируется посредством измерения воздействия J на общий клеточный уровень ATP, присутствующей в микобактерии, с использованием ATP Bioluminescence Luciferase Assay Kit HSII, Roche. Этот анализ основан на осуществляемом с помощью ATP преобразовании D-люциферина в оксилюциферин, которое может быть измерено при 526 нм.
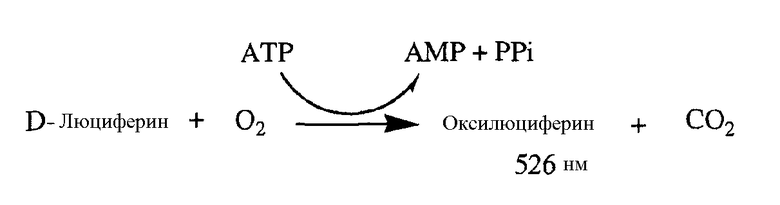
Вкратце, воздействие DARQ J на общий уровень ATP исследуют как на M.tuberculosis дикого типа, так и на мутантном штамме. DCCD, который представляет собой хорошо известный ингибитор ATP-синтазы, используют в качестве положительного контроля, и изониазид, который представляет собой ингибитор биосинтеза определенных компонентов клеточных стенок, но не влияет на продуцирование ATP, используют в качестве отрицательного контроля.
Как можно увидеть на фиг.3, обработка M.tuberculosis дикого типа DARQ J приводит к зависимому от дозы уменьшению продуцирования ATP в этих бактериях. В противоположность этому, изониазид не влияет на продуцирование ATP. Как уже описано выше, экспонирование этих бактерий для высокой концентрации DARQ J увеличивает количество устойчивых к диарилхинолинам мутантов M.tuberculosis. Когда эти устойчивые M.tuberculosis обрабатывают DARQ J, эти бактерии не показывают никакого уменьшения продуцирования ATP, даже при концентрации этого соединения, превышающей в 100 раз минимальную ингибирующую концентрацию (MIC). В противоположность этому, DCCD способен блокировать продуцирование ATP в этих бациллах, что говорит о том, что DARQ J и DCCD имеют различные карманы связывания в ATP-синтазах.
Компьютерное моделирование и идентификация области связывания DARQ J
Для дальнейшего исследования различных способов действия DCCD и DARQ J генерируют компьютерную 3-мерную модель ATP-синтазы как для дикого типа, так и для мутанта DARQ J M.tuberculosis. Атомные координаты, приведенные в таблицах 4 и 5, вычисляют путем построения модели 3-мерной структуры опубликованных аминокислотных последовательностей P63691 и AJ865377. Реальный сайт связывания DARQ J, как обнаружено, находится в контактной области субъединиц A и C, более конкретно, вблизи аминокислот 'Arg 210' субъединицы A и 'Glu 61' субъединицы C, как упоминается в таблицах 3, 4 или 5. Это прекрасно согласуется с результатами, наблюдаемыми для скрининга придающих устойчивость мутаций в чувствительных и устойчивых штаммах M.tuberculosis и M.smegmatis, описанных выше.
Модель основана на оптимизации относительного расположения A- и C-спиралей структуры ATP-азы и ориентации основной цепи и боковой цепи аминокислот по отношению к минимальной вычисленной внутренней деформации. Геометрию получают посредством ряда циклов моделирования с помощью молекулярной динамики и релаксаций молекулярной механики, начиная от опубликованной ранее геометрии спирали другого организма [E-Coli PDB код доступа 1C17 - V.K. Rastogi and M.E. Girvin, Nature, 402, 263-268 (1999)]. Как молекулярная динамика, так и релаксация геометрии осуществляется с помощью параметризации силового поля на основе MMFF94s [Halgren, T.A. (1996), J. Comput. Chem., 17, 490-519], но может использоваться любое программное обеспечение для молекулярной динамики, известное в данной области [Berendsen, H.J.C., van der Spoel, D. and van Drunen, R., Comp. Phys. Comm. 91 (1995), 43-56; Lindahl, E., Hess, B. and van der Spoel, D., J. Mol. Mod. 7(2001) 306-317.] с последующей соответствующей оптимизацией геометрии [J. W. Ponder and F. M. Richards, J. Comput. Chem., 8, 1016-1024 (1987)].
Вычисленные координаты в таблицах 3, 4 и 5 содержат часть (с объемом радиусом 30 Å) предсказанной структуры области, рассматриваемой в качестве релевантной для ингибирования активности MTB ATP-азы в этих ферментах, на основе предложенного способа ингибирования и появления точечных мутаций, вызывающих устойчивость, в биологических анализах.
Обсуждение
DARQ J является членом нового химического класса противотуберкулезных агентов с MIC, равной или более низкой, чем у эталонных соединений. Его спектр является уникальным по его специфичности к микобактериям, включая атипичные виды, важные для людей: MAC, M.kansasii и быстро растущие M.fortuitum и M.abscessus. Этот спектр антимикобактериальной специфичности отличается от спектра изониазида, который не обладает активностью против MAC. Клиническое использование J будет узко направленным на лечение TB и микобактериальных инфекций. Неспособность J к ингибированию не-микобактерий должна трансформироваться в меньшее селективное давление и более низкий риск развития устойчивости у других видов бактерий по сравнению с антибиотиками более широкого спектра (9).
Мишень и механизм действия J отличаются от параметров других противотуберкулезных агентов. Сравнение последовательностей ATP-синтаз различных бактерий и эукариотической ATP-синтазы и, в частности, цепи C субъединицы F0 комплекса ATP-азы, совместно с 3-мерным моделированием ATP-синтаз дикого типа и мутанта M.tuberculosis, дает объяснение специфичности антибактериального спектра и до меньшей степени профиля безопасности.
Исследование динамики, осуществляемое на сконструированных моделях ATP-азы Mycobacterium, показывает, что в этих структурах существует полость (сайт связывания в соответствии с атомными координатами таблицы 3) в области контакта субъединиц A и C (рядом с аминокислотой 'Arg 210' субъединицы A и 'Glu 61' субъединицы C). В таблицах 4 и 5 приводятся координаты двух изученных вариантов атомов, окружающих этот сайт, и их средние положения. Ингибитор DARQ J способен вмешиваться в стадию нормального переноса протона, в которую вовлечены эти две аминокислоты, посредством облегчения взаимодействия этих двух аминокислот. Стереоспецифичность DARQ J может быть понята из асимметрии предсказанного сайта связывания; активный хиральный энантиомер соответствует этой полости оптимально, другие формы соединения и варианты ATP-азы согласуются не так хорошо.
Сайт связывания, который авторы получили здесь, находится на совершенно другой части системы ATP-азы, чем DCCD (DARQ находятся в мембранной части фермента, связывание DCCD происходит приблизительно за 90 Å внутри клетки; на основе PDB структуры кристалла бычьей ATP-азы "1E79" - опубликовано в C.Gibbons, M.G. Montgomery, A.G.W. Leslie, J.E. Walker, Nat. Struct. Biol., 7, 1055 (2000). По этой причине атомы, которые могли бы потенциально быть вовлечены в ингибирование MTB ATP-азы типа DCCD, не находятся в какой-либо области из перечисленных в таблицах координат сайта связывания (который распространяется как раз только на мембранную часть фермента), и, наоборот, эта часть фермента, вовлеченная в способ ингибирования DARQ J, не присутствует в опубликованной структуре "1E79" (которая показывает только внутриклеточную часть). Это различие в сайтах связывания может объяснить различную реакцию M.Tuberculosis, наблюдаемую при анализе in vitro продуцирования ATP.
Не противореча указанному выше, другие исследования с помощью DCCD на митохондриальных ATP-азах предлагают другой сайт связывания, расположенный вблизи кислой аминокислоты в липофильной окружающей среде области F0 фермента, например Sebald W, Machleidt W, Wachter E., Proc Natl Acad Sci USA. 1980 Feb; 77(2):785-789. Это положение связывания на митохондриальных ATP-азах может считаться аналогичным положению, описанному здесь для видов Mycobacterium.
В то же время, нацеливание нового механизма обеспечивает то, что циркулирующие в настоящее время штаммы TB с мутациями, резистентными к доступным видам лечения, не являются перекрестно устойчивыми с J. Из исследований авторов in vitro очевидно, что J имеет, по меньшей мере, такое же высокое антибактериальное воздействие против изолятов MDR-TB, даже против изолятов с широкой устойчивостью к четырем лекарственным средствам, как и против абсолютно восприимчивых штаммов M.tuberculosis дикого типа. Это наблюдение является важным, поскольку оно четко демонстрирует, что нет перекрестной устойчивости с существующими противотуберкулезными лекарственными средствами. При условии дополнительной идентификации кармана связывания в мембранной части ATP-азы, результаты настоящего исследования сделают возможной дальнейшую разработку новых антибактериальных соединений, в частности антимикобактериальных соединений, нацеленных на синтез ATP в этих организмах.
Таблица 1
Медианные значения
Ala24, Gly27, Phe53, Phe54, Val57, Gly38, Glu61, Tyr64, Phe65
Met17, Gly19, Gly20, Ala21, Ile22, Gly23, Ala24, Gly25, Ile26, Gly27, Asp28, Gly29, Ala31, Phe53, Thr56, Val57, Gly58, Leu59, Val60, Glu61, Ala62, Ala63/Pro63, Tyr64, Phe65.
Ser182, Leu183, Leu185, Arg186 (имеющие коды Ser 206-Leu 207-Leu 209 и Arg 210 в таблице 3)
Таблица 3
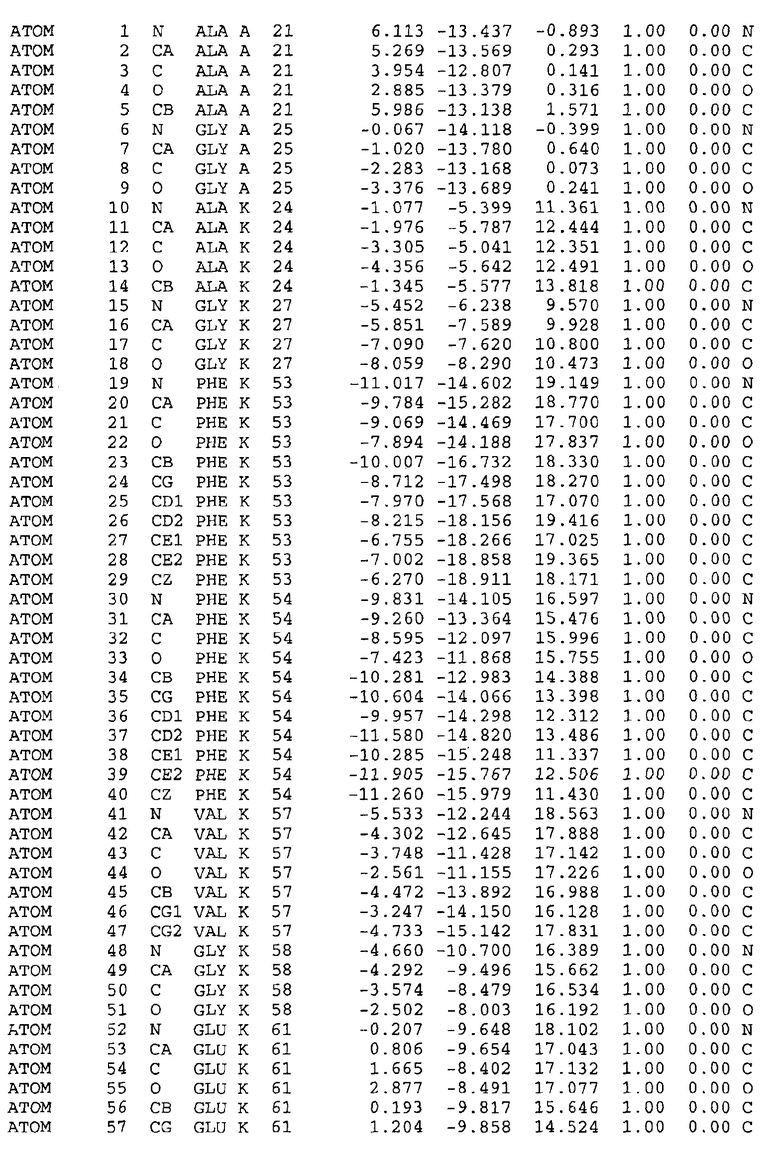
Таблица 3 -продолжение
Таблица 3 -продолжение
Таблица 3 -продолжение
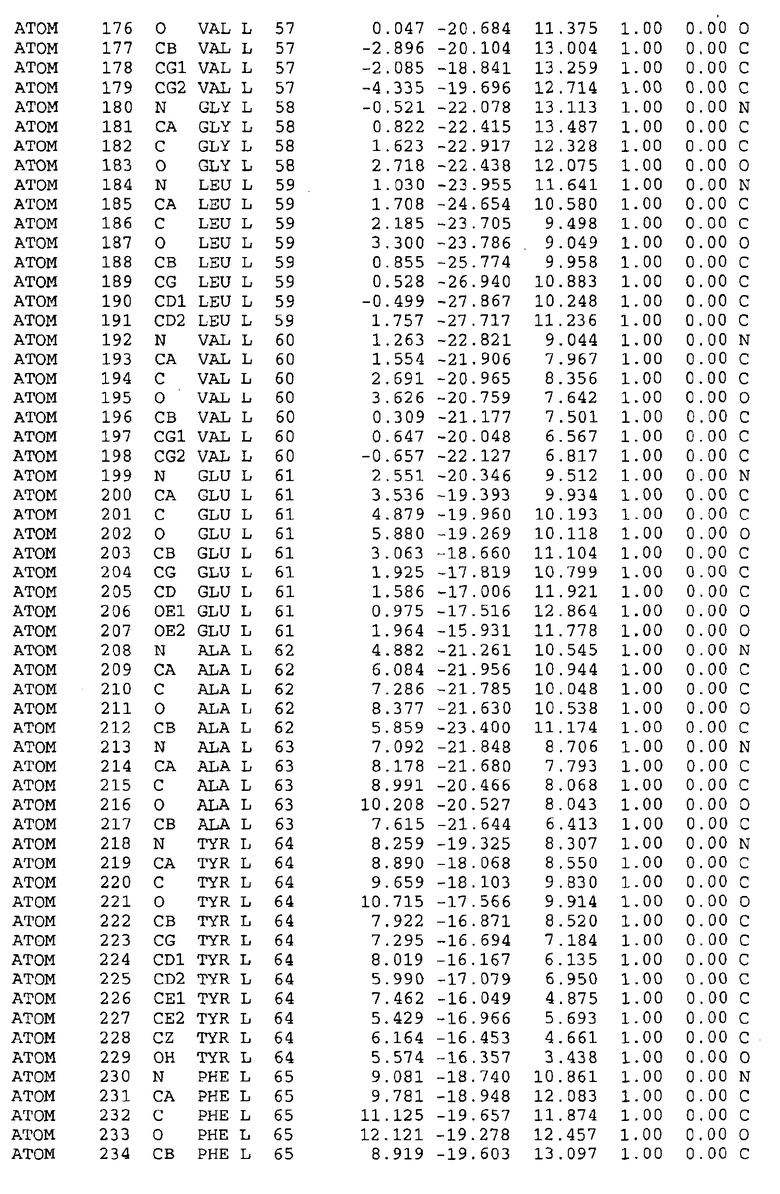
Таблица 3 -продолжение
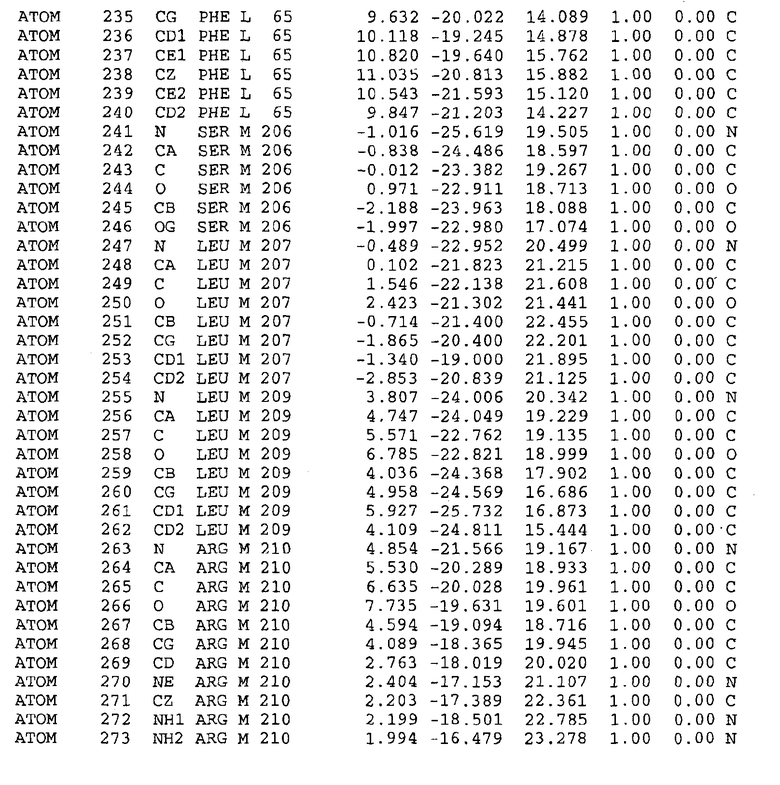
ТАБЛИЦА 4
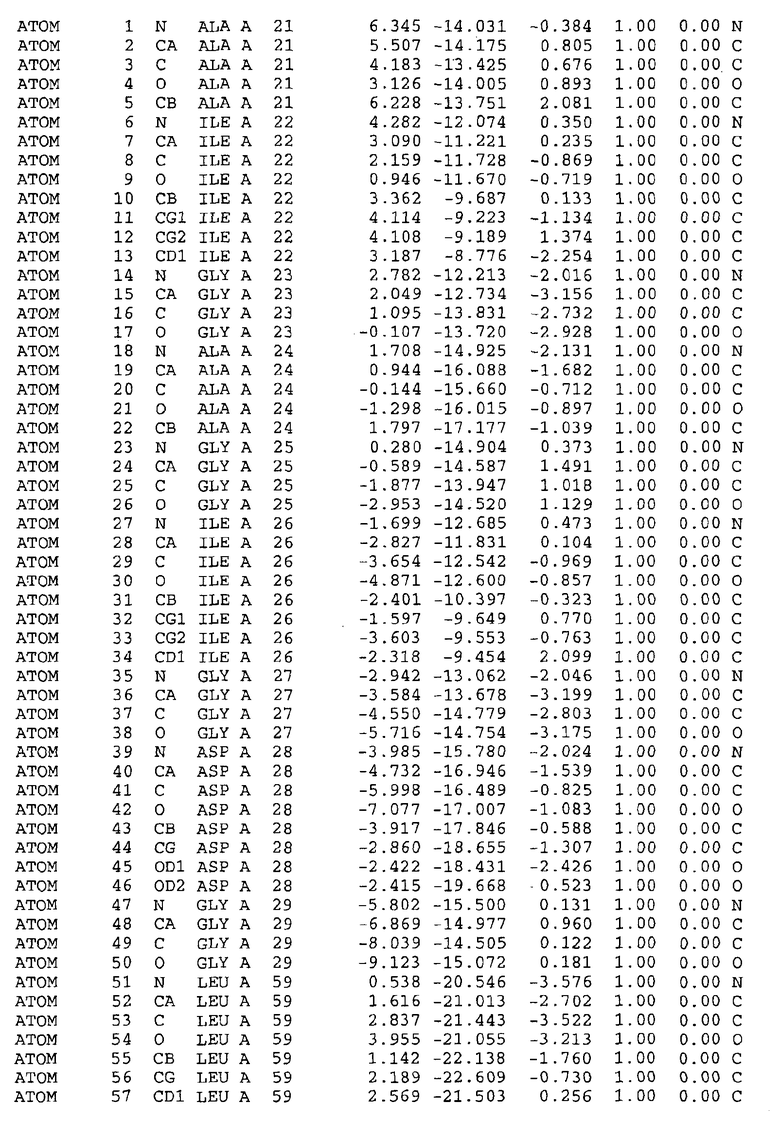
Таблица 4 -продолжение
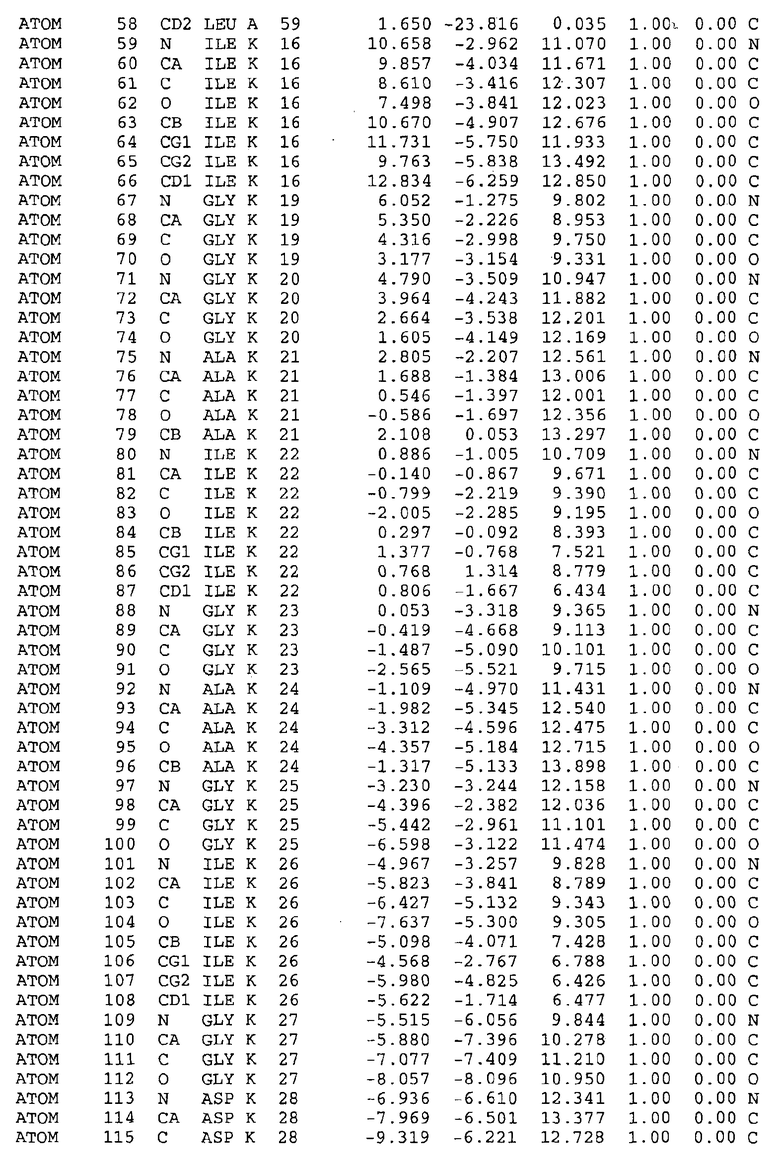
Таблица 4 -продолжение
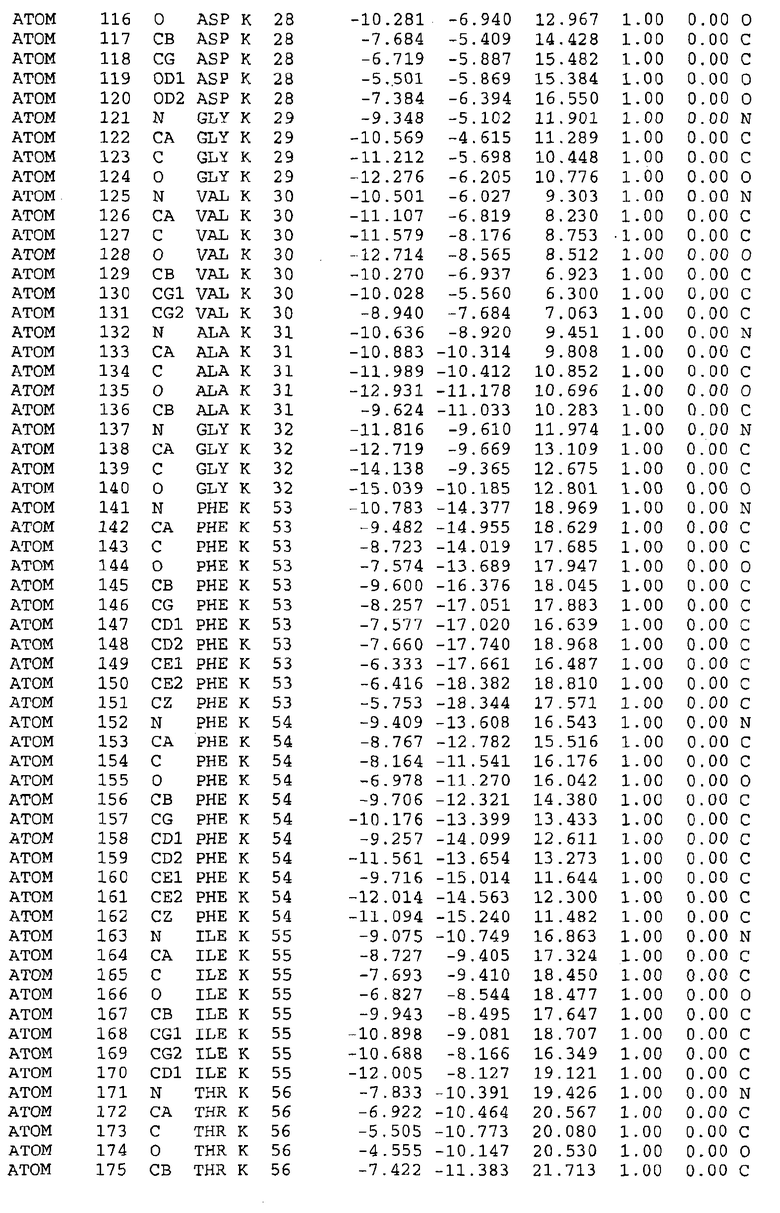
Таблица 4 -продолжение
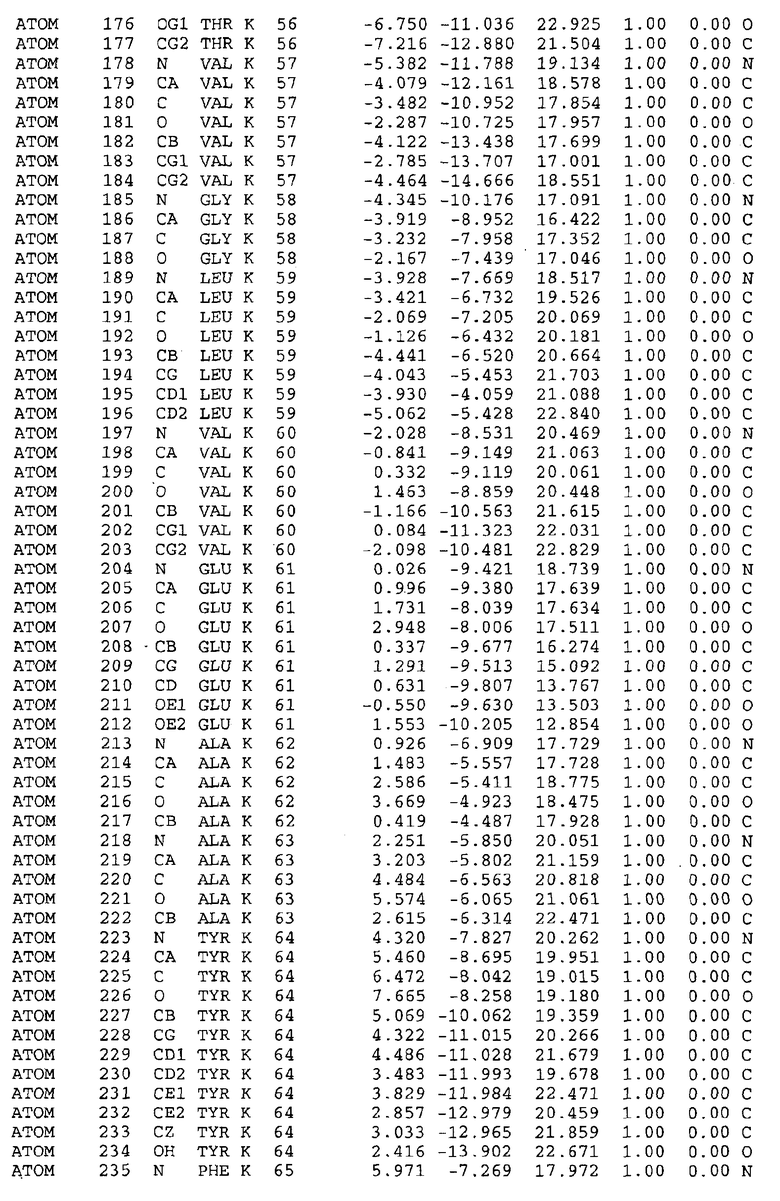
Таблица 4 -продолжение
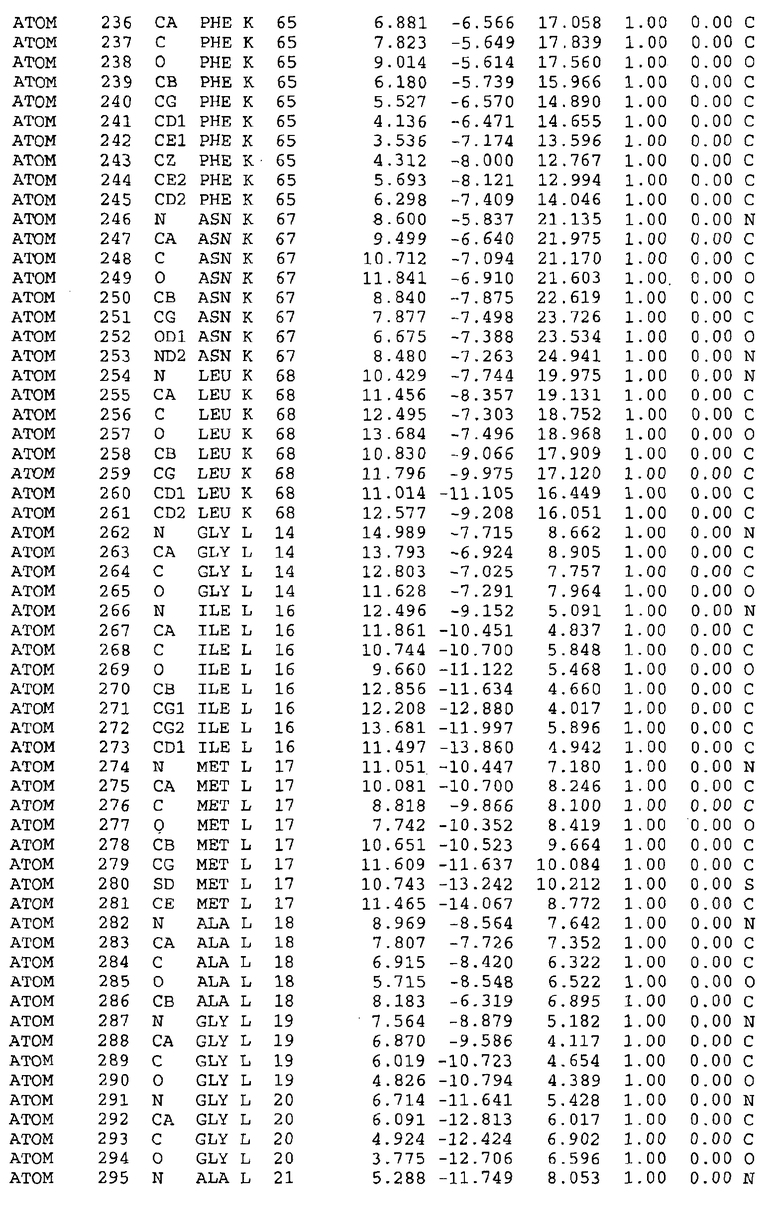
ТАБЛИЦА 4 -ПРОДОЛЖЕНИЕ
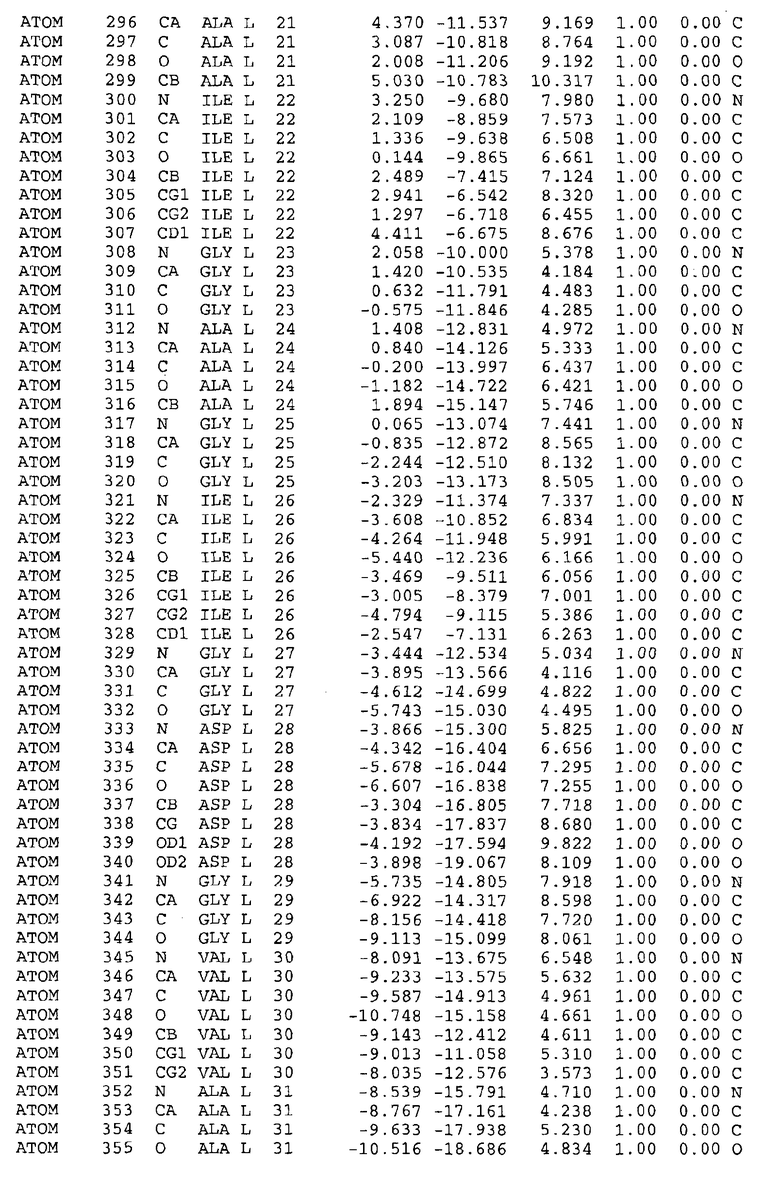
ТАБЛИЦА 4 -ПРОДОЛЖЕНИЕ
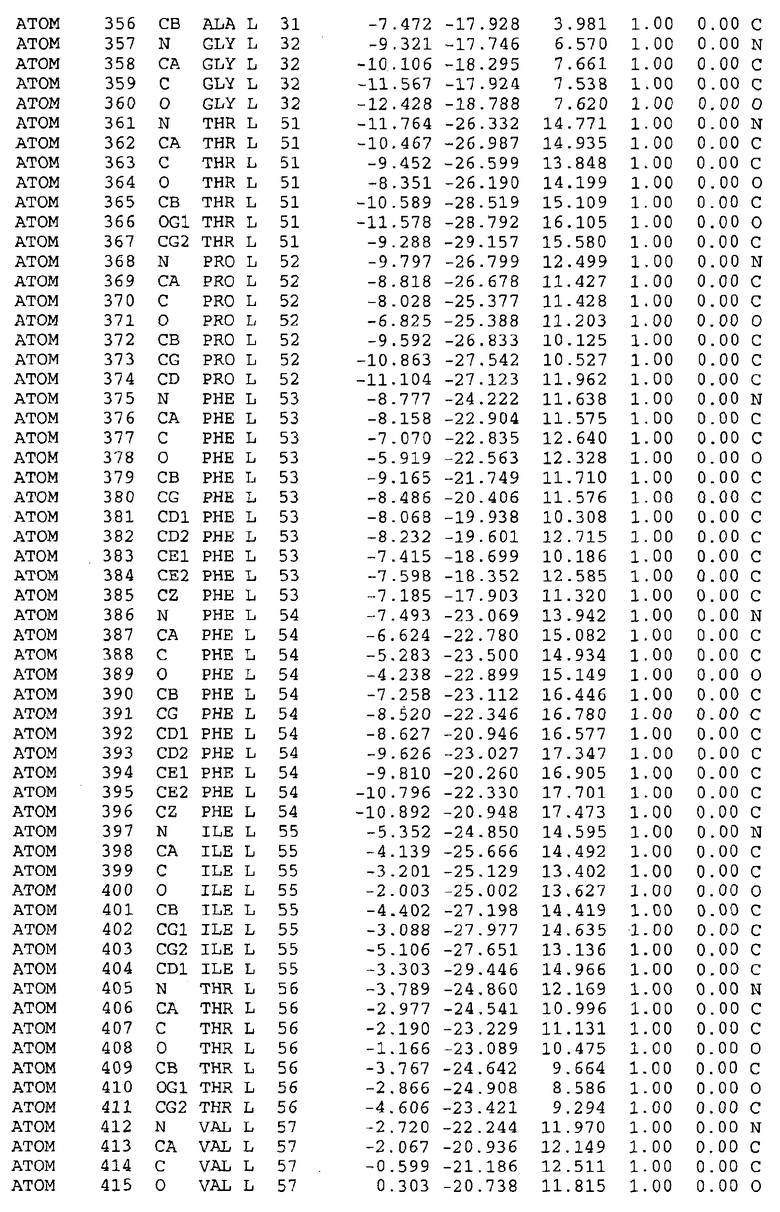
ТАБЛИЦА 4 -ПРОДОЛЖЕНИЕ
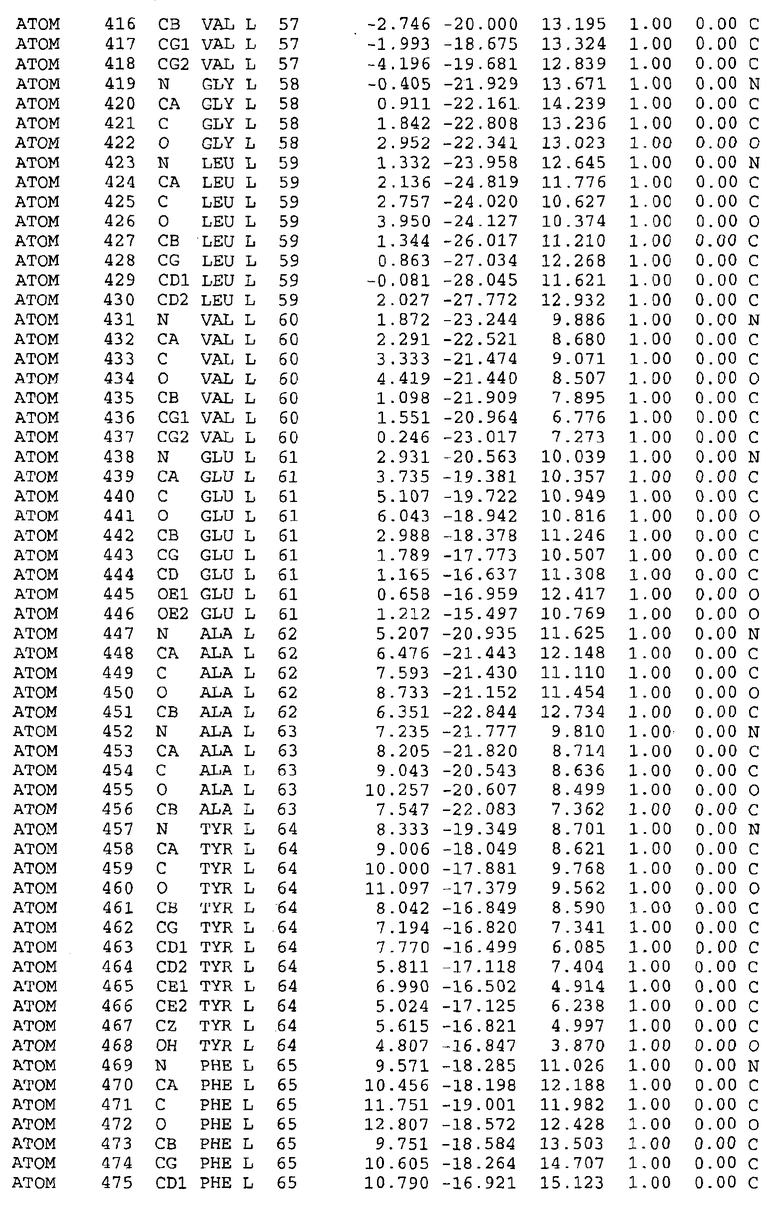
ТАБЛИЦА 4 -ПРОДОЛЖЕНИЕ
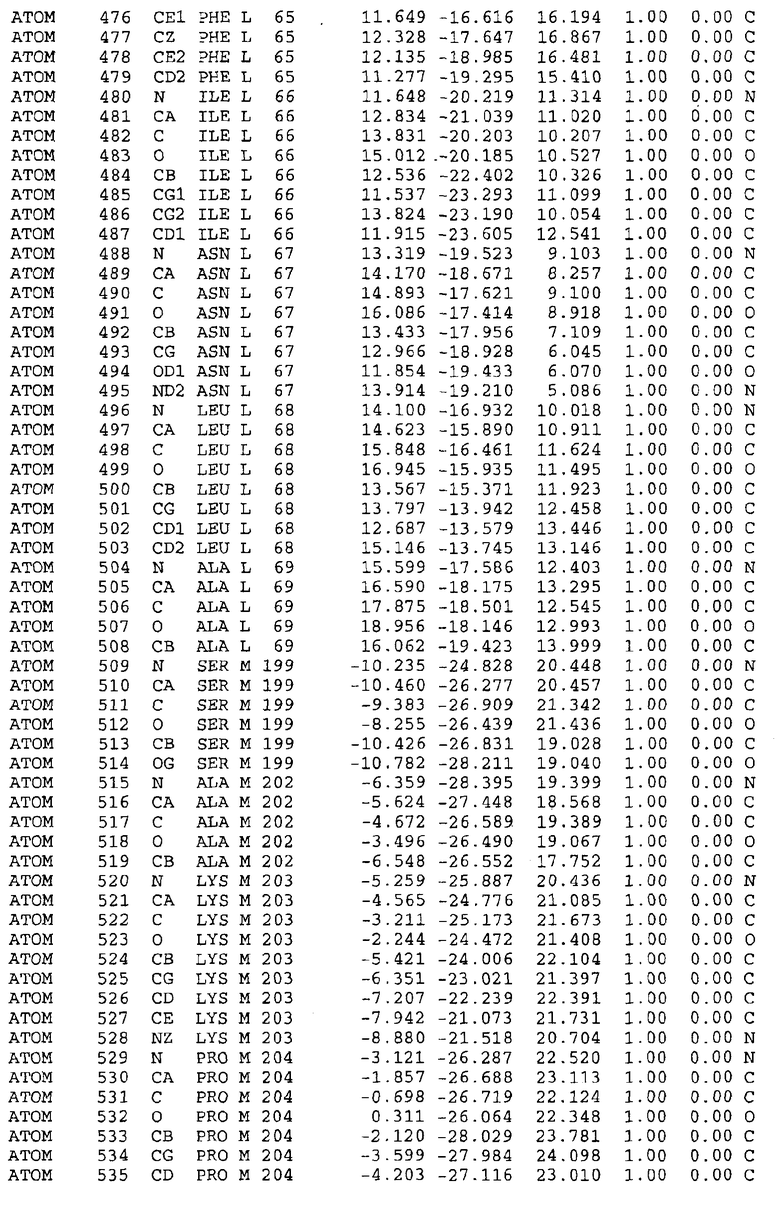
ТАБЛИЦА 4 -ПРОДОЛЖЕНИЕ
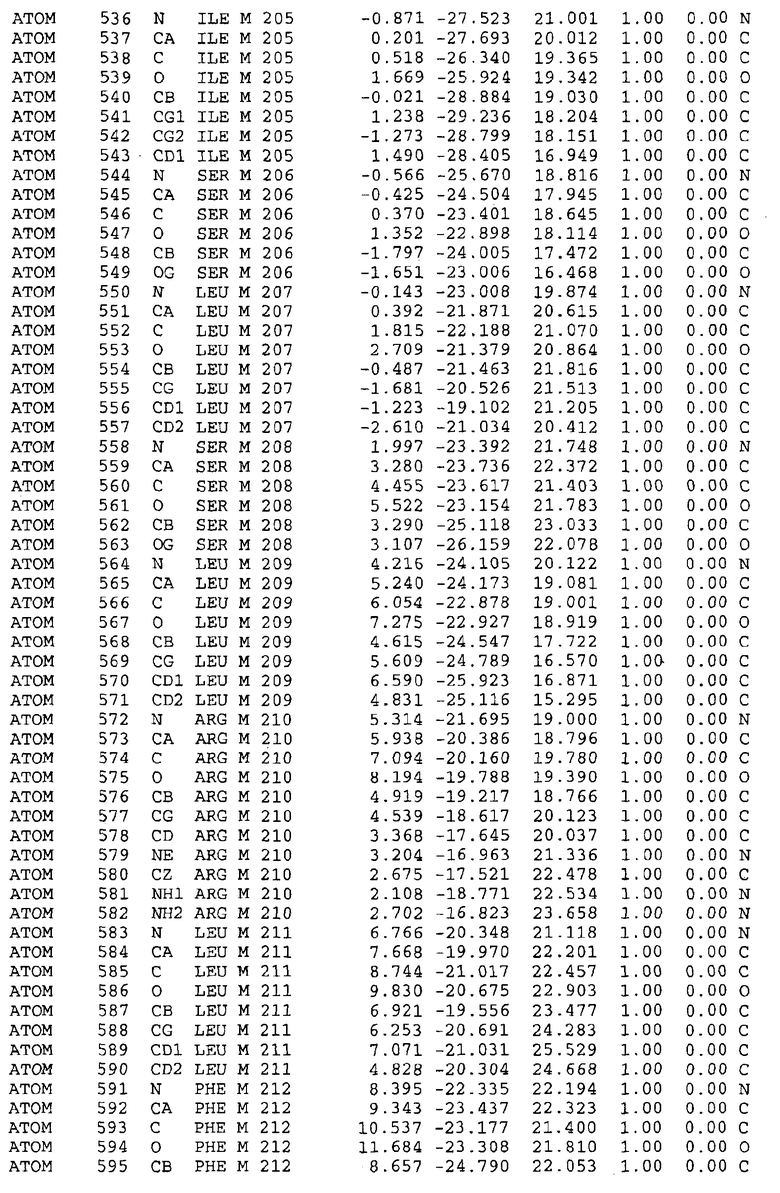
ТАБЛИЦА 4 -ПРОДОЛЖЕНИЕ
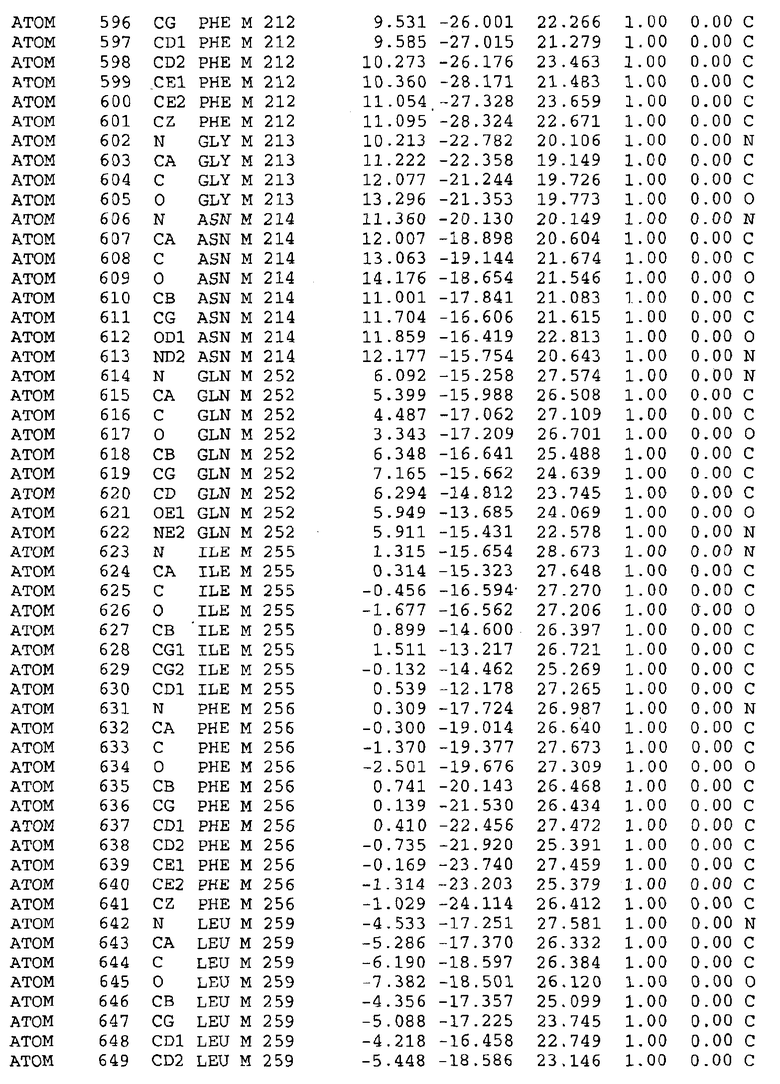
ТАБЛИЦА 5
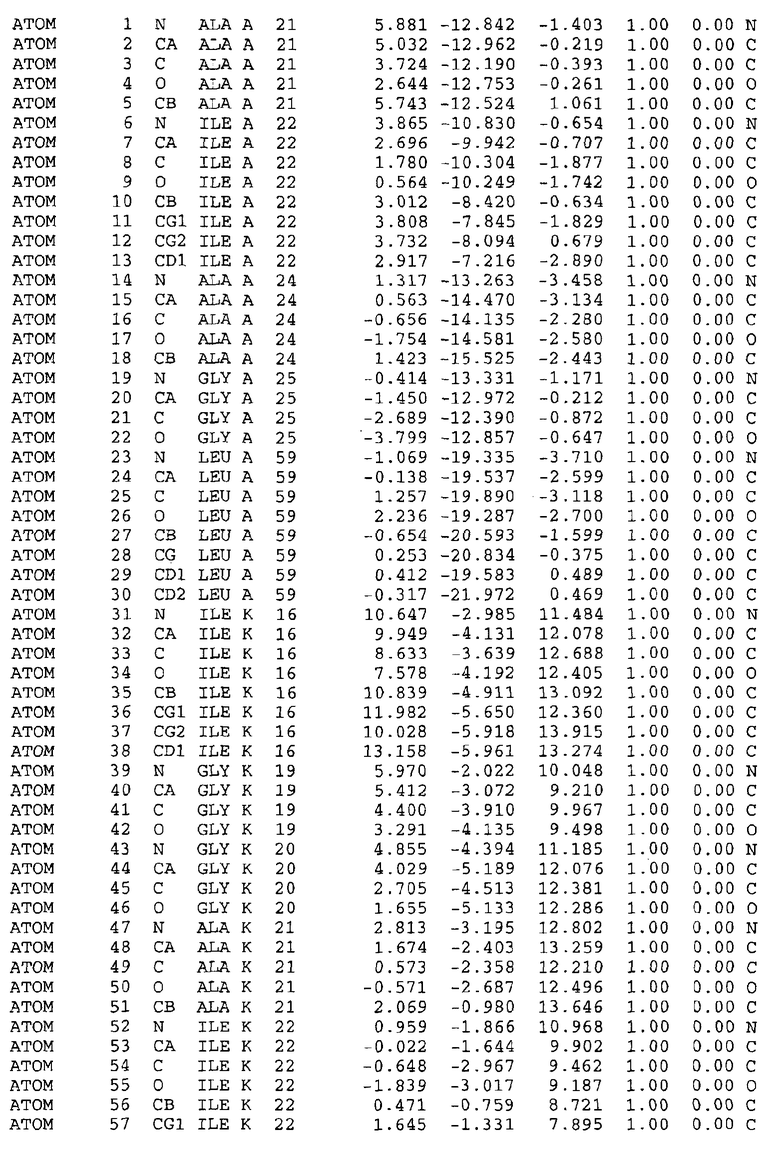
ТАБЛИЦА 5 -ПРОДОЛЖЕНИЕ
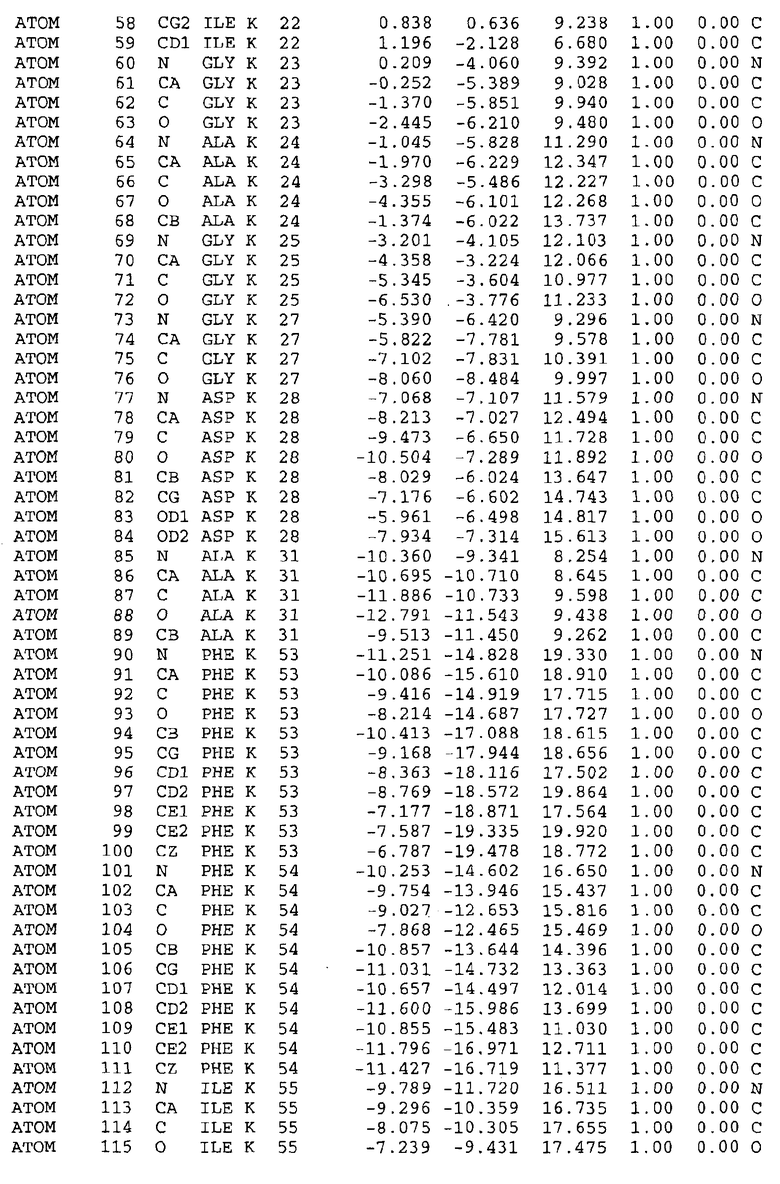
ТАБЛИЦА 5 -ПРОДОЛЖЕНИЕ
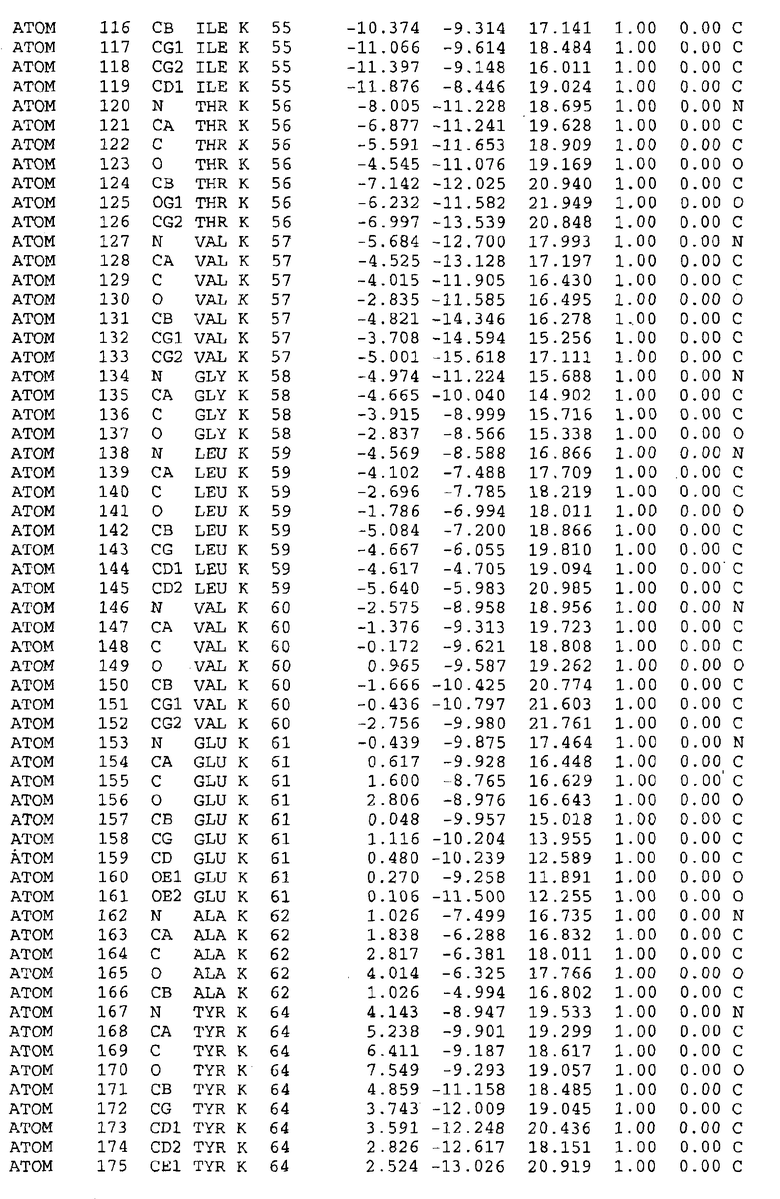
ТАБЛИЦА 5 -ПРОДОЛЖЕНИЕ
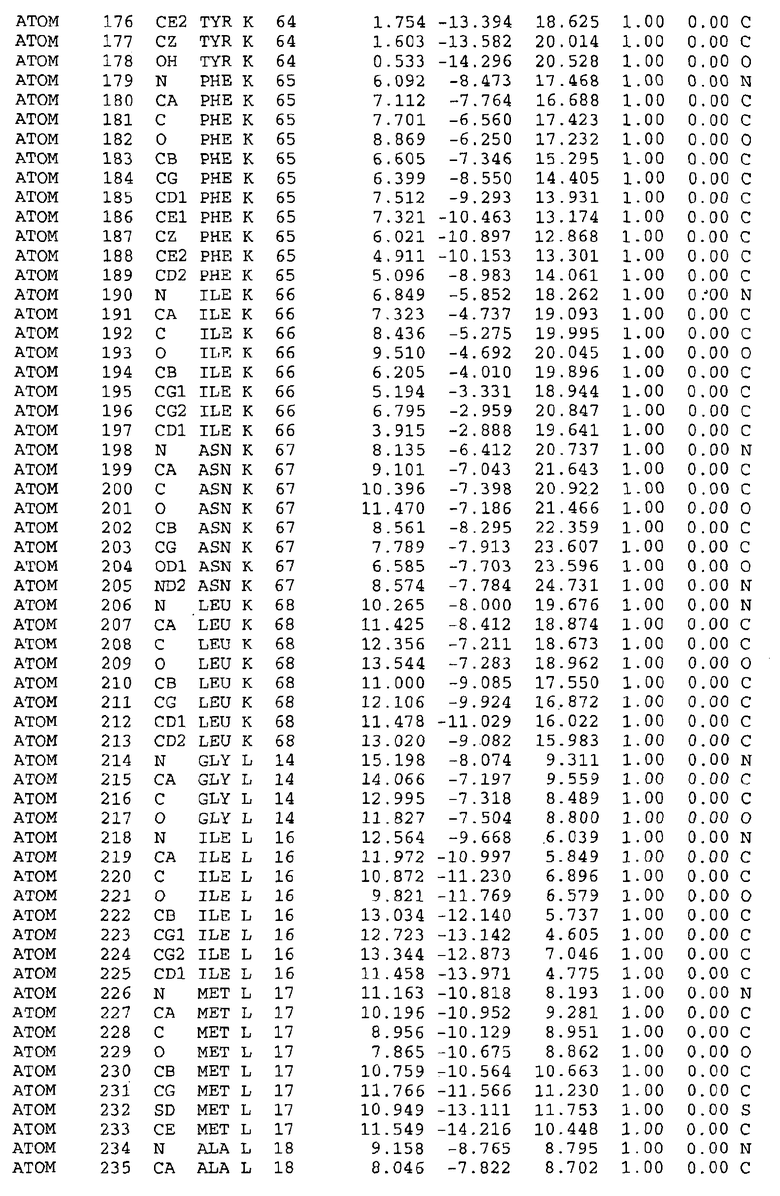
ТАБЛИЦА 5 -ПРОДОЛЖЕНИЕ
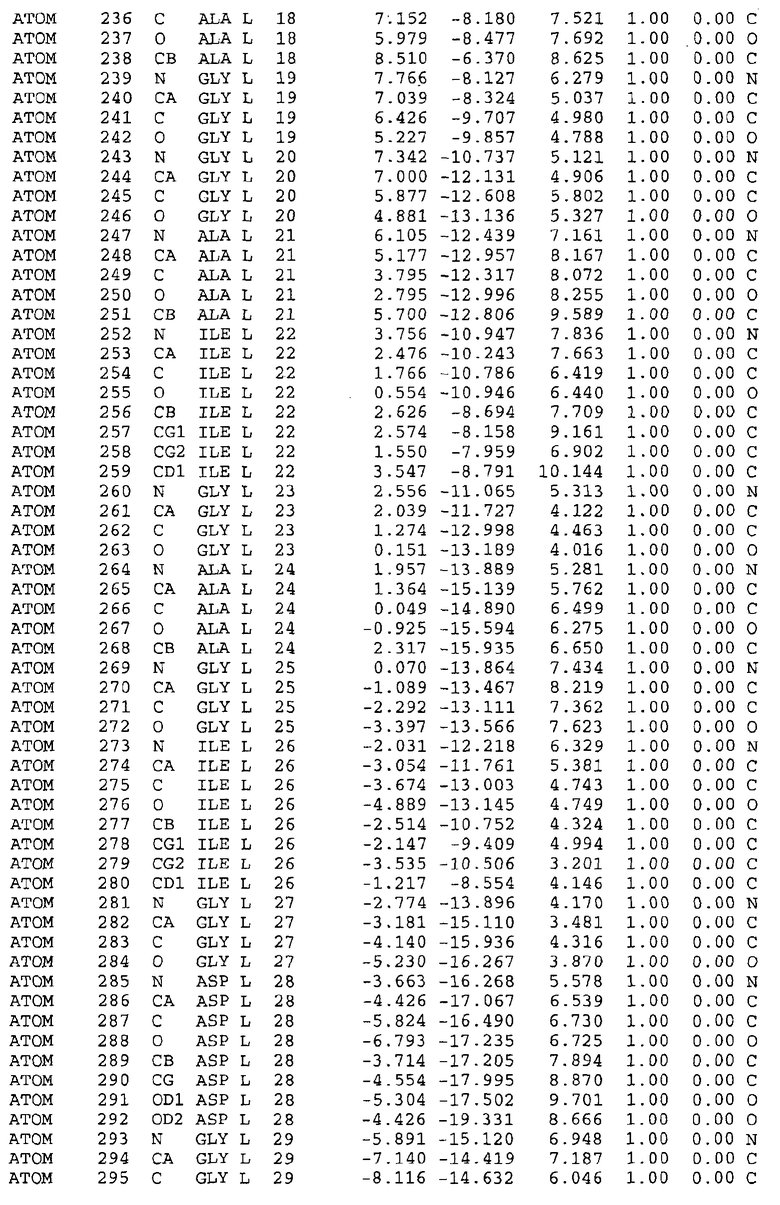
ТАБЛИЦА 5 -ПРОДОЛЖЕНИЕ
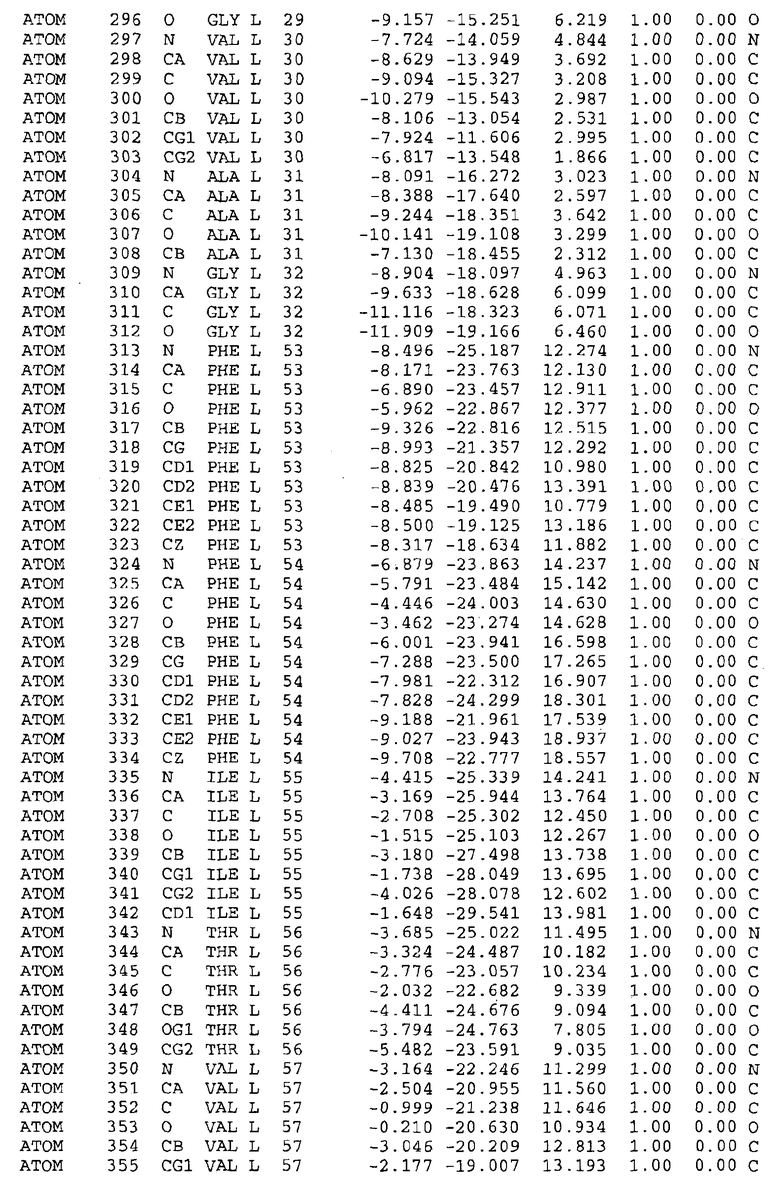
ТАБЛИЦА 5 -ПРОДОЛЖЕНИЕ
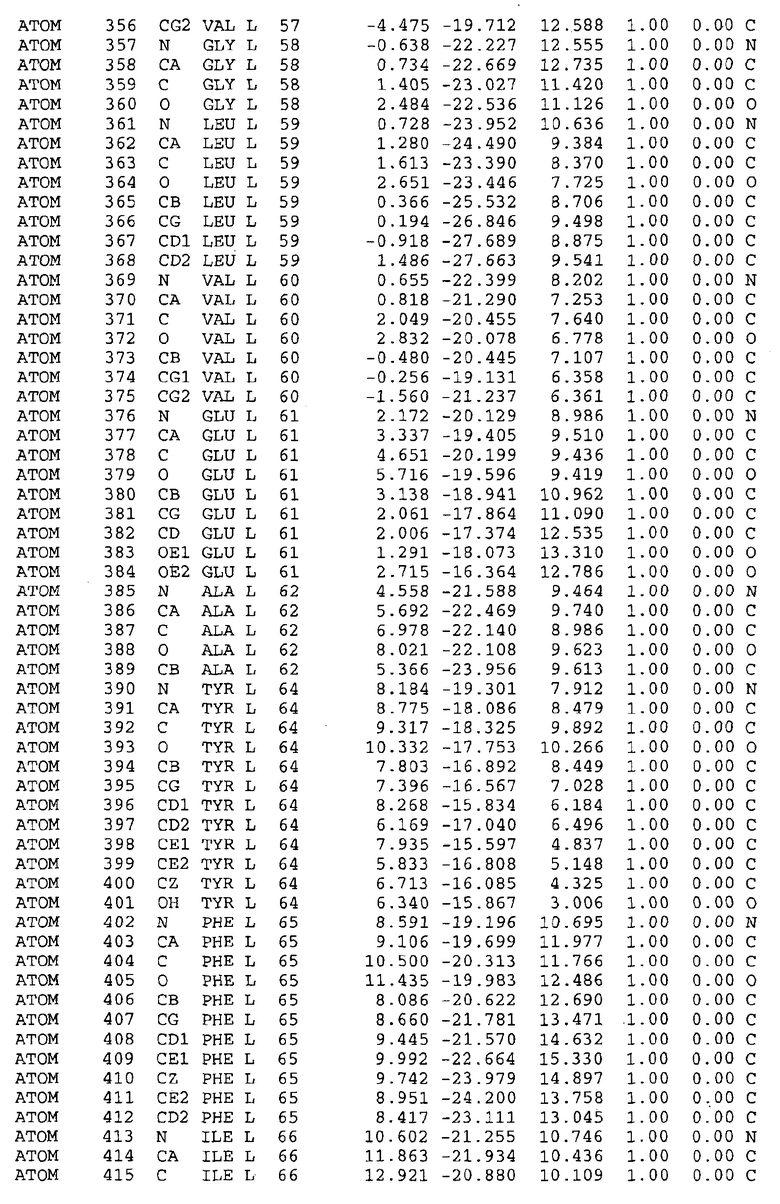
ТАБЛИЦА 5 -ПРОДОЛЖЕНИЕ
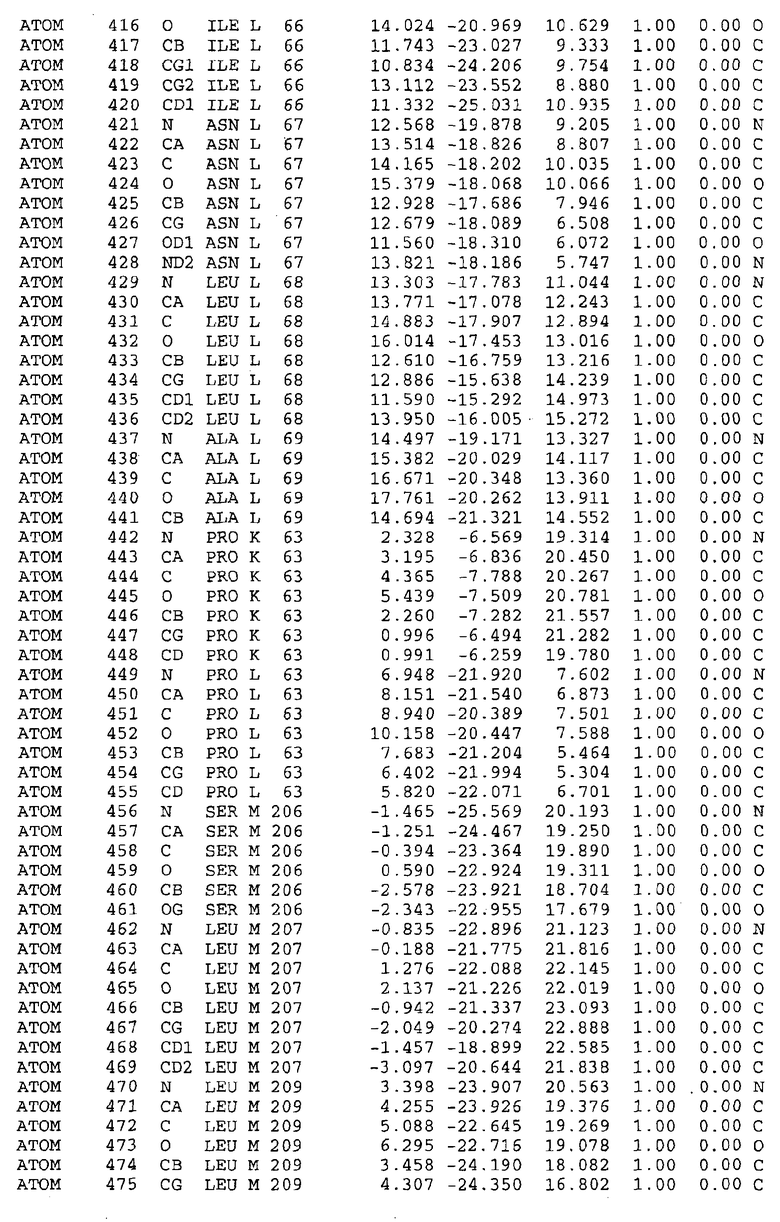
ТАБЛИЦА 5 -ПРОДОЛЖЕНИЕ
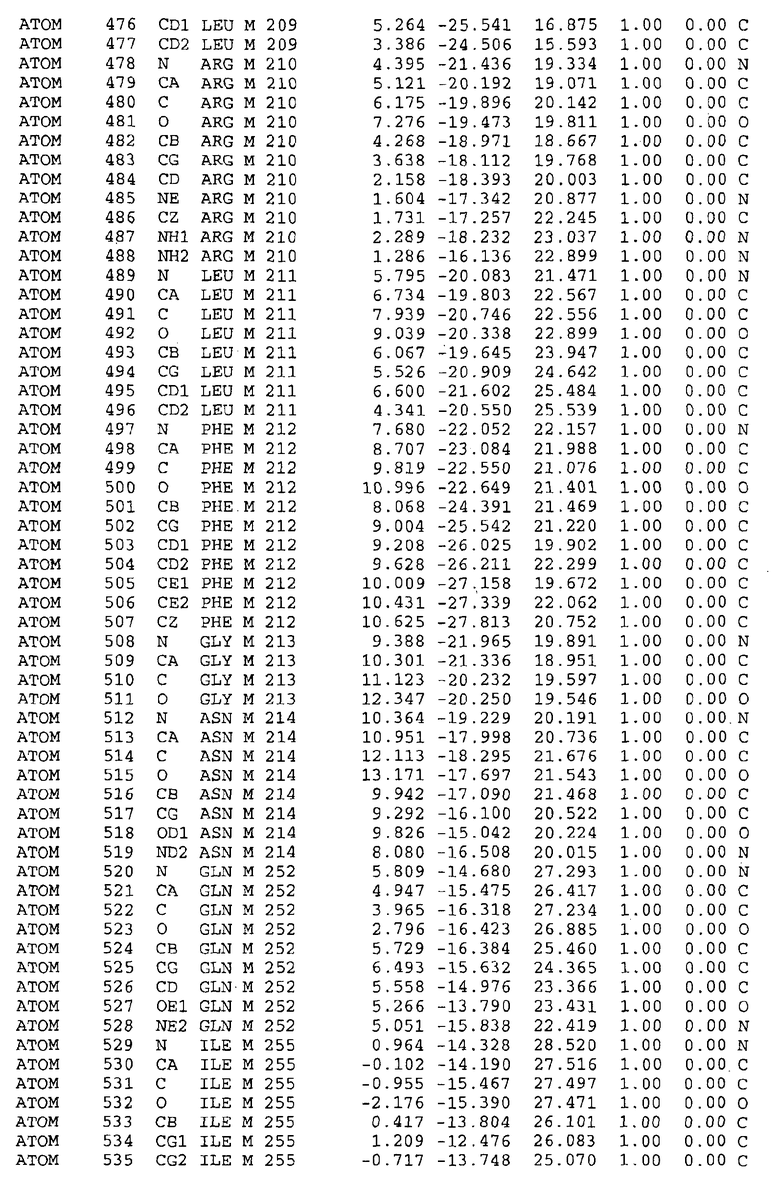
ТАБЛИЦА 5 -ПРОДОЛЖЕНИЕ
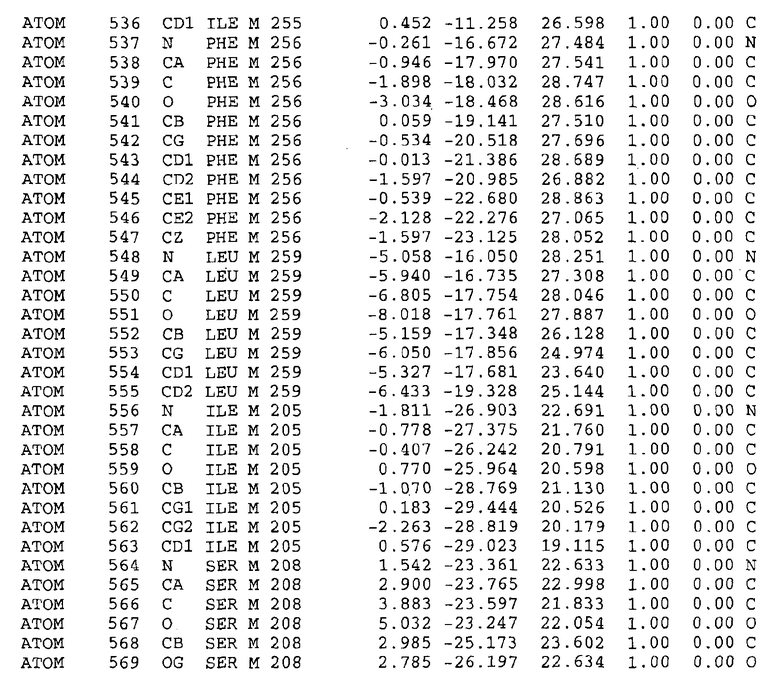
ТАБЛИЦА 6
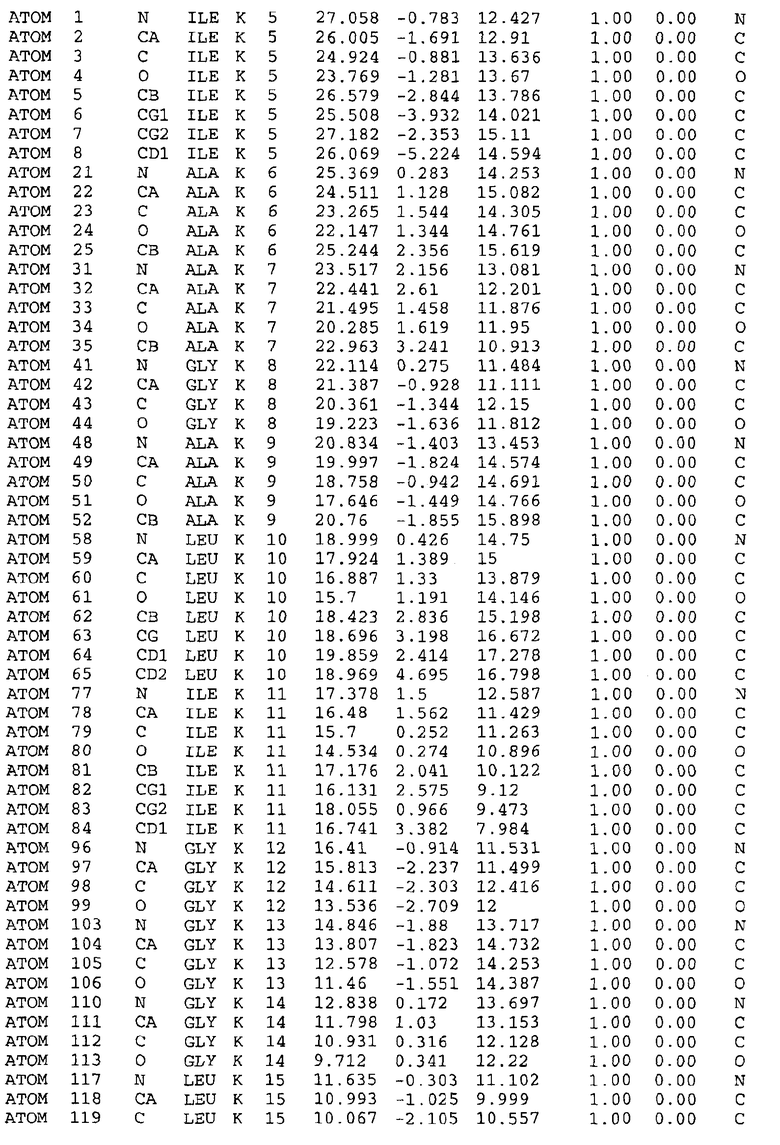
ТАБЛИЦА 6 -ПРОДОЛЖЕНИЕ
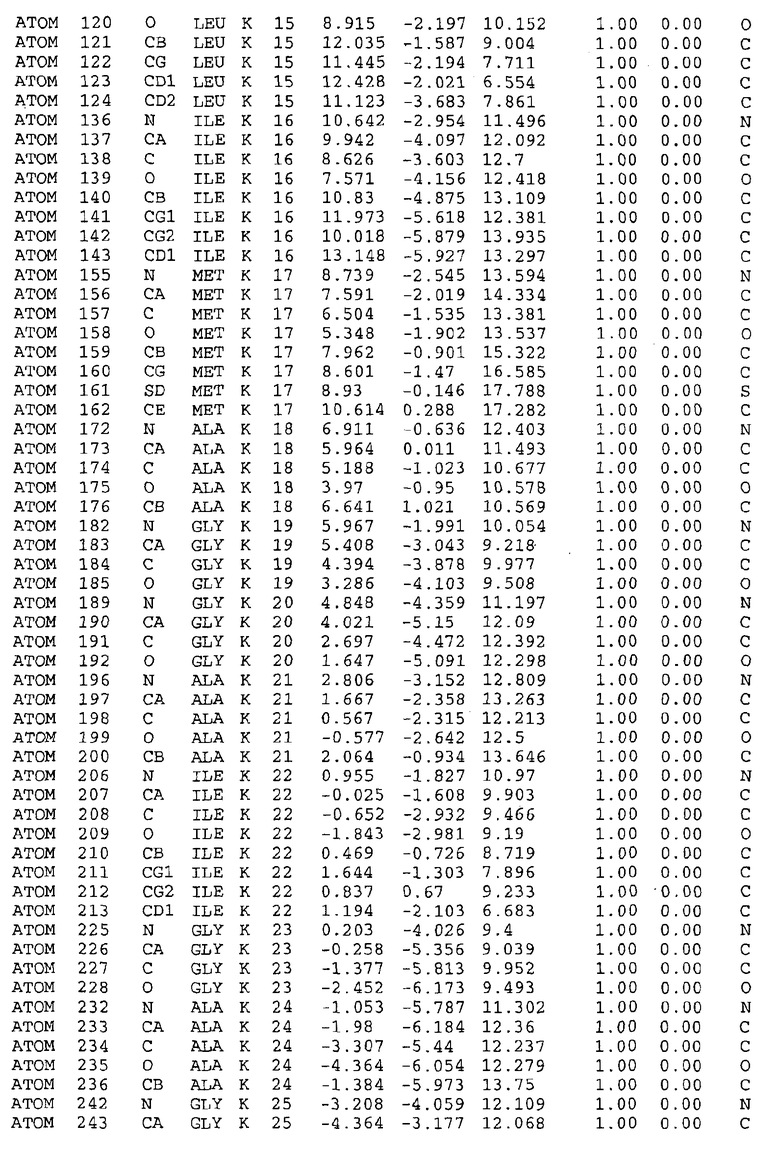
ТАБЛИЦА 6 -ПРОДОЛЖЕНИЕ
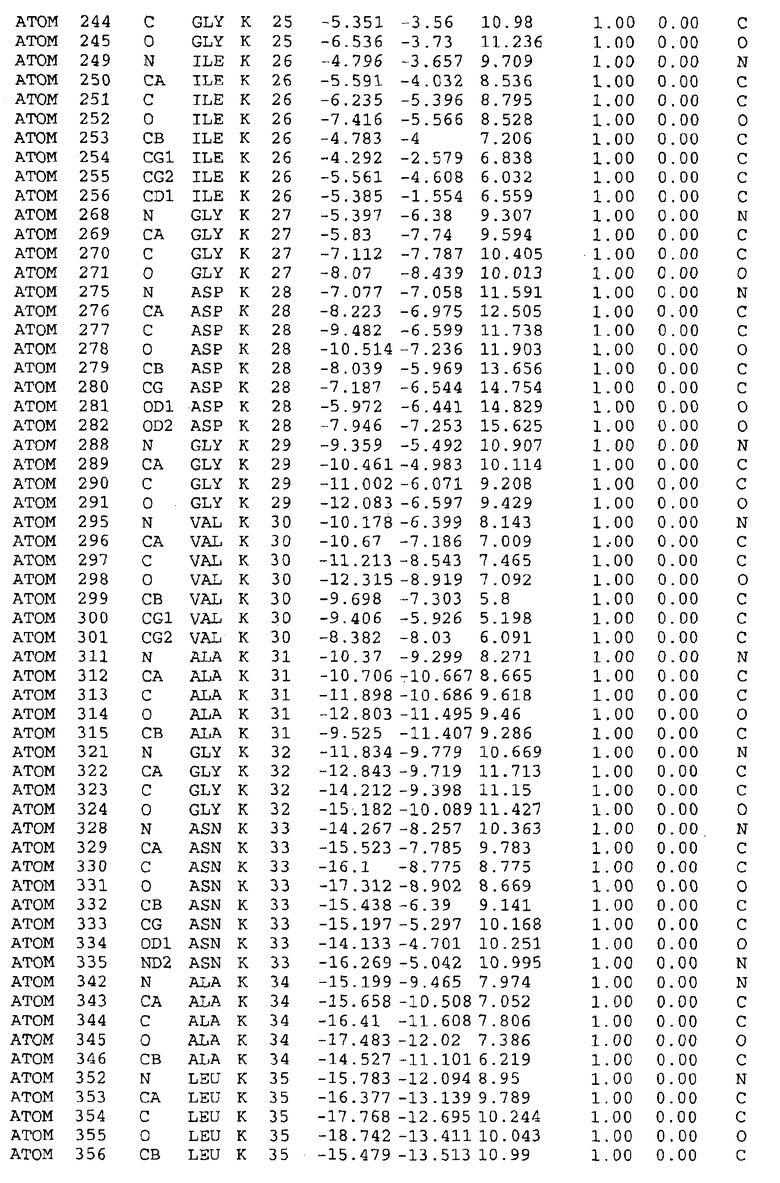
ТАБЛИЦА 6 -ПРОДОЛЖЕНИЕ
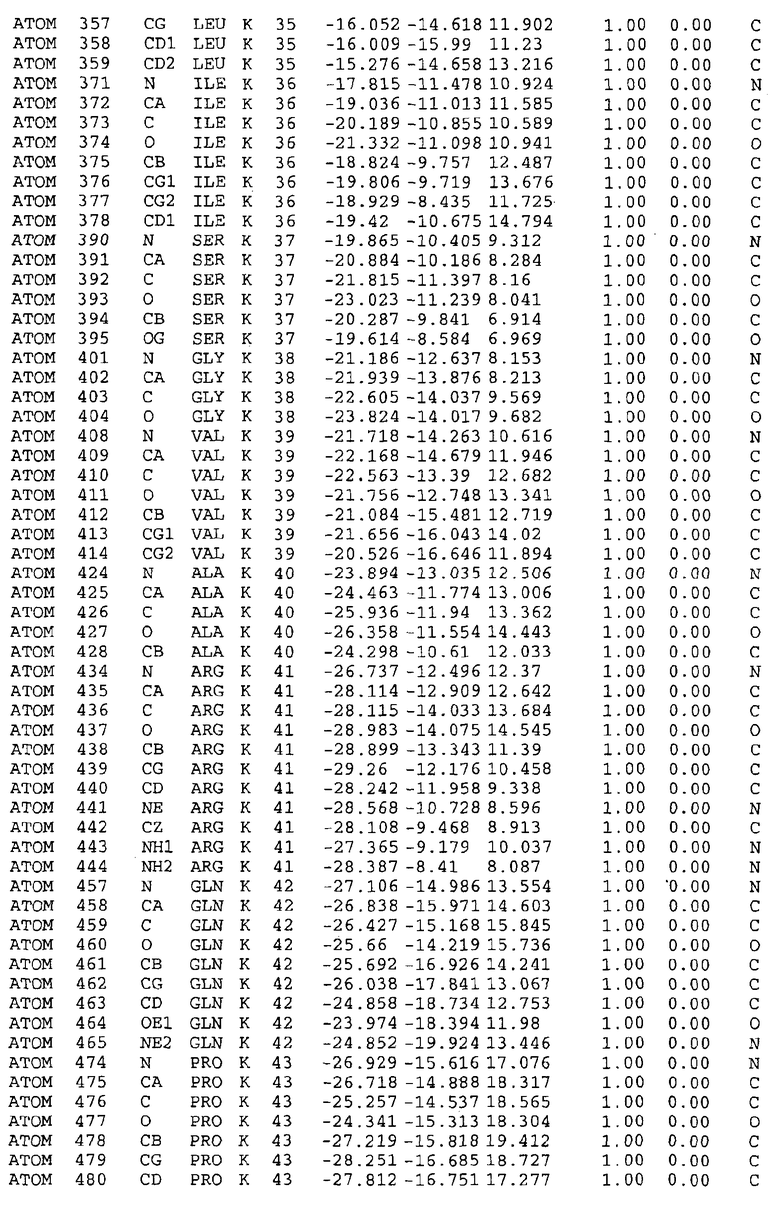
ТАБЛИЦА 6 -ПРОДОЛЖЕНИЕ
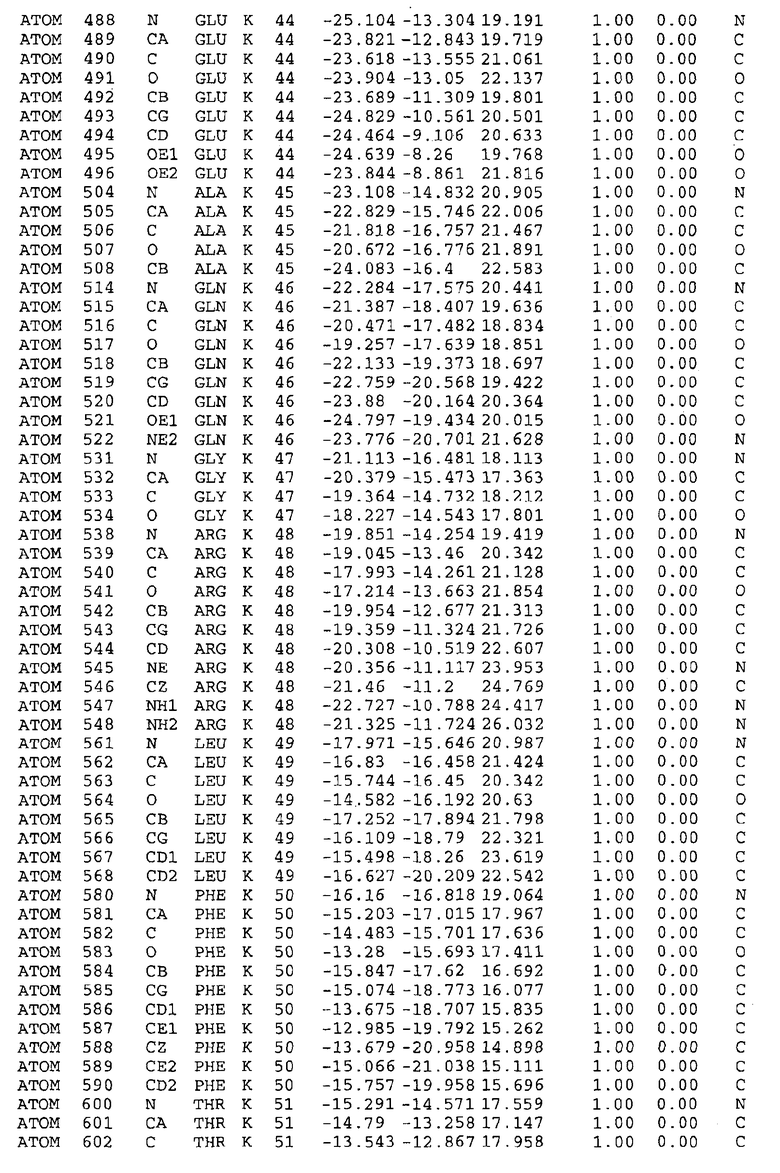
ТАБЛИЦА 6 -ПРОДОЛЖЕНИЕ
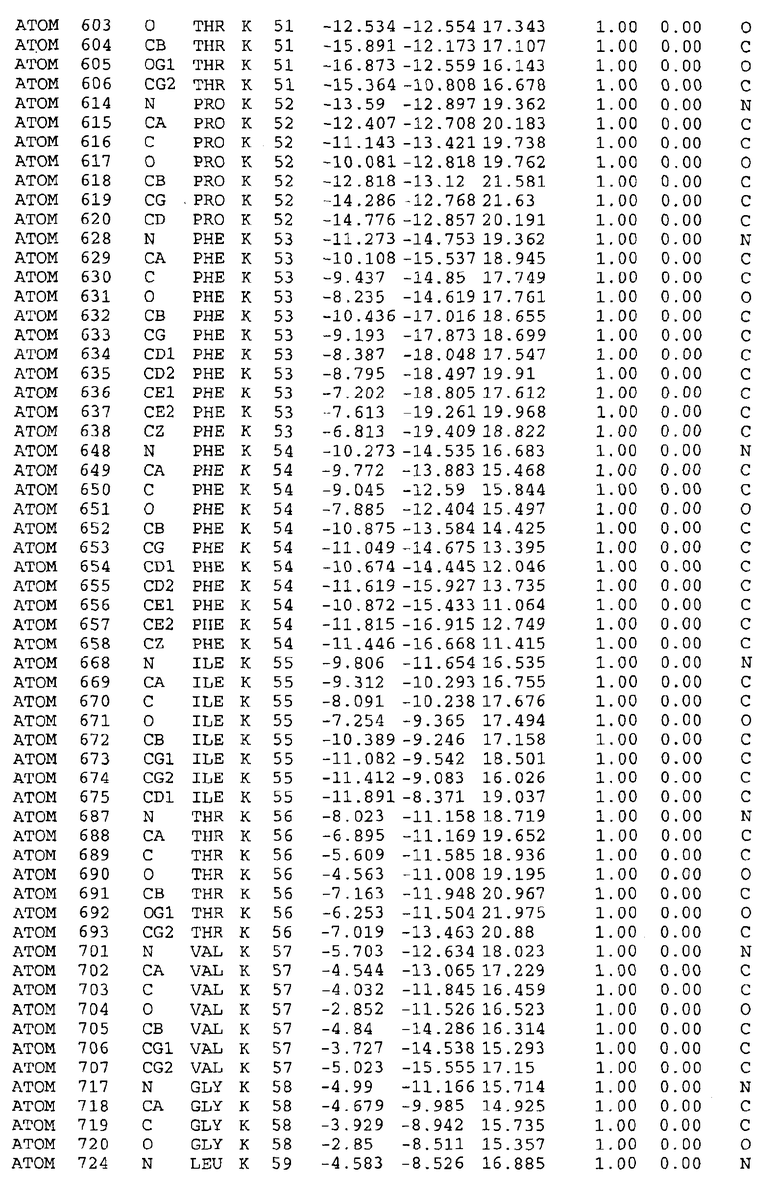
ТАБЛИЦА 6 -ПРОДОЛЖЕНИЕ
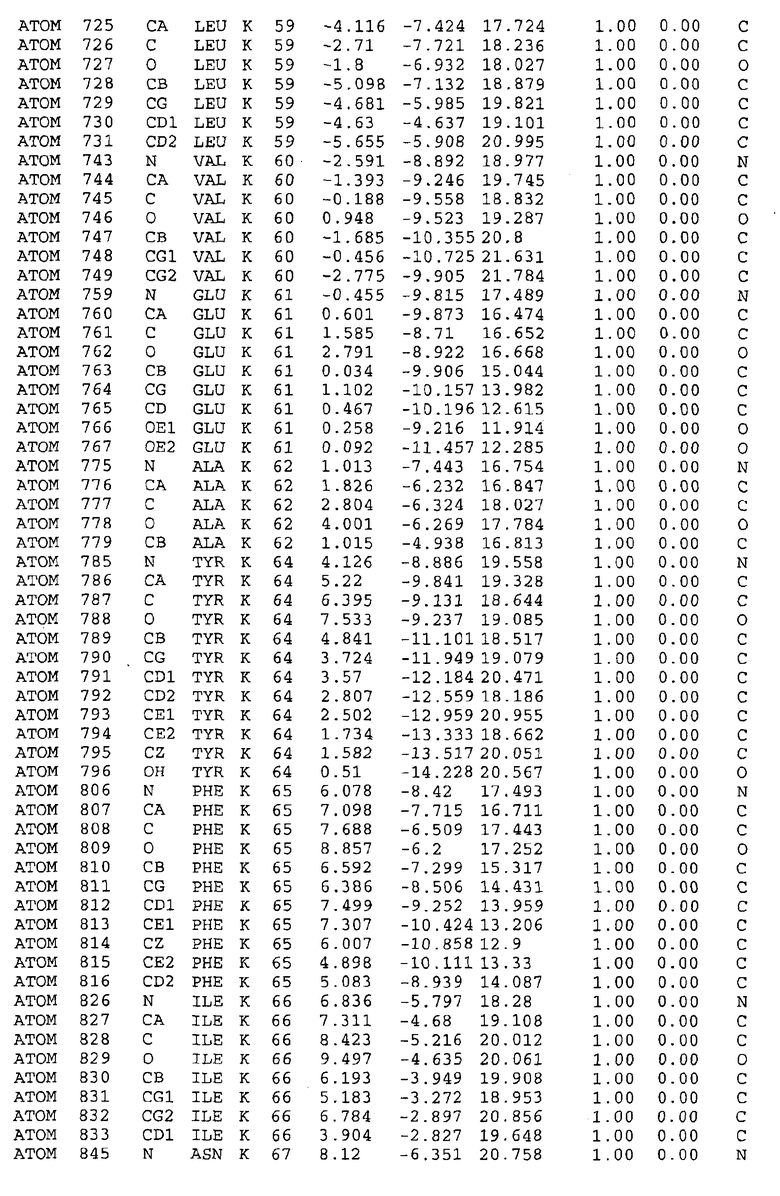
ТАБЛИЦА 6 -ПРОДОЛЖЕНИЕ
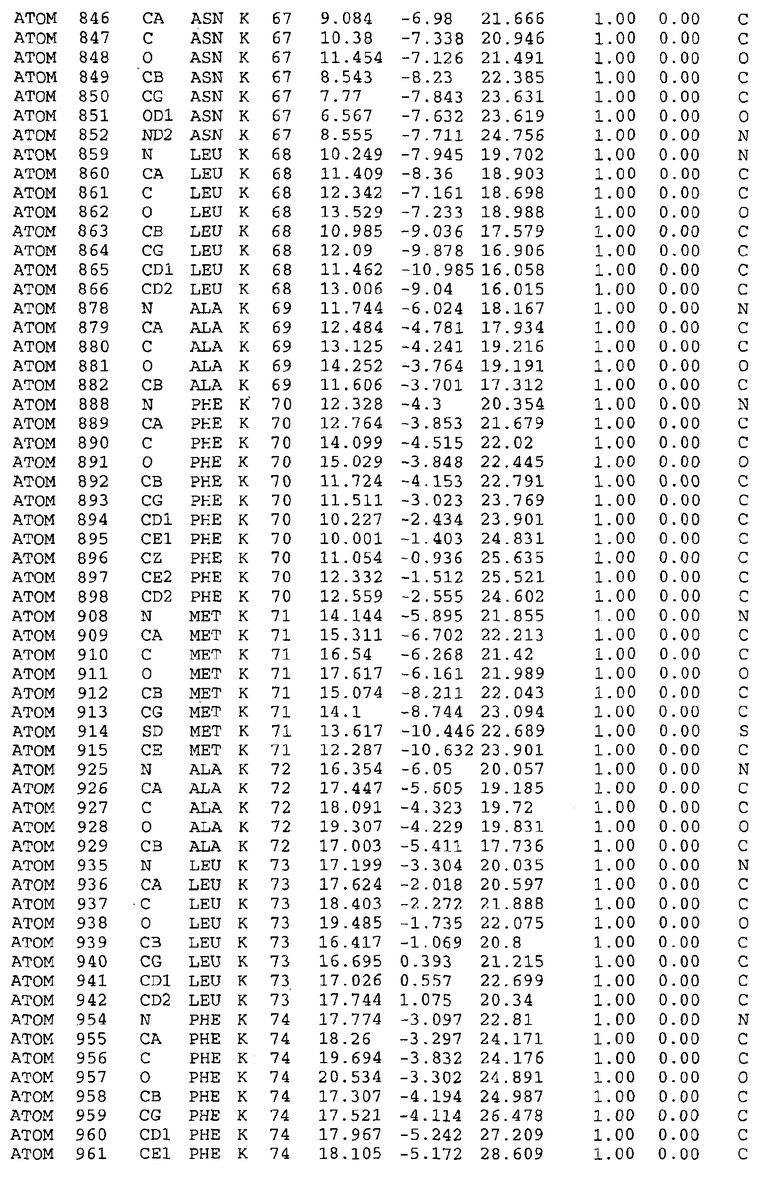
ТАБЛИЦА 6 -ПРОДОЛЖЕНИЕ
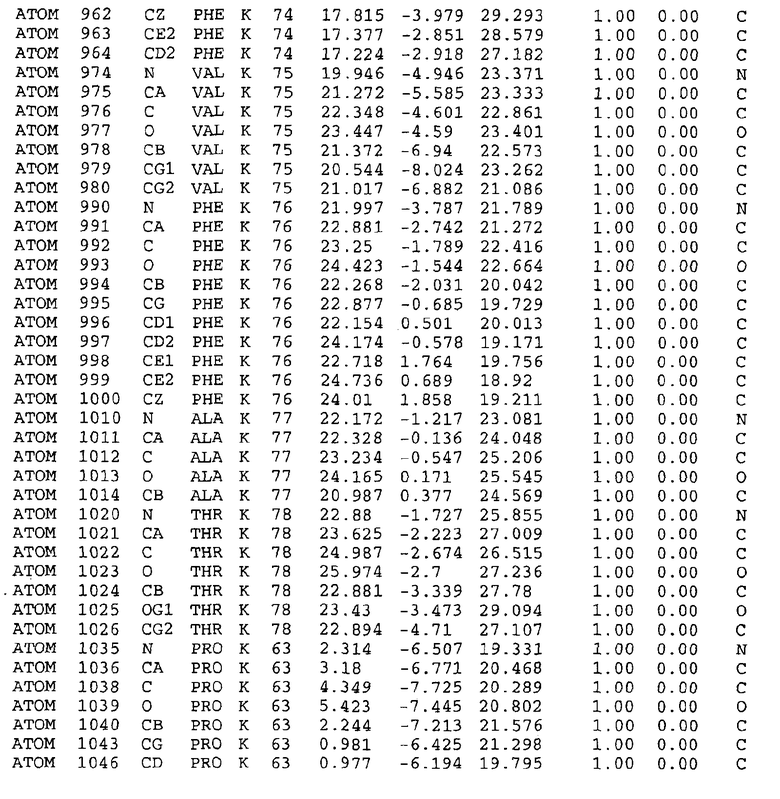
ТАБЛИЦА 7
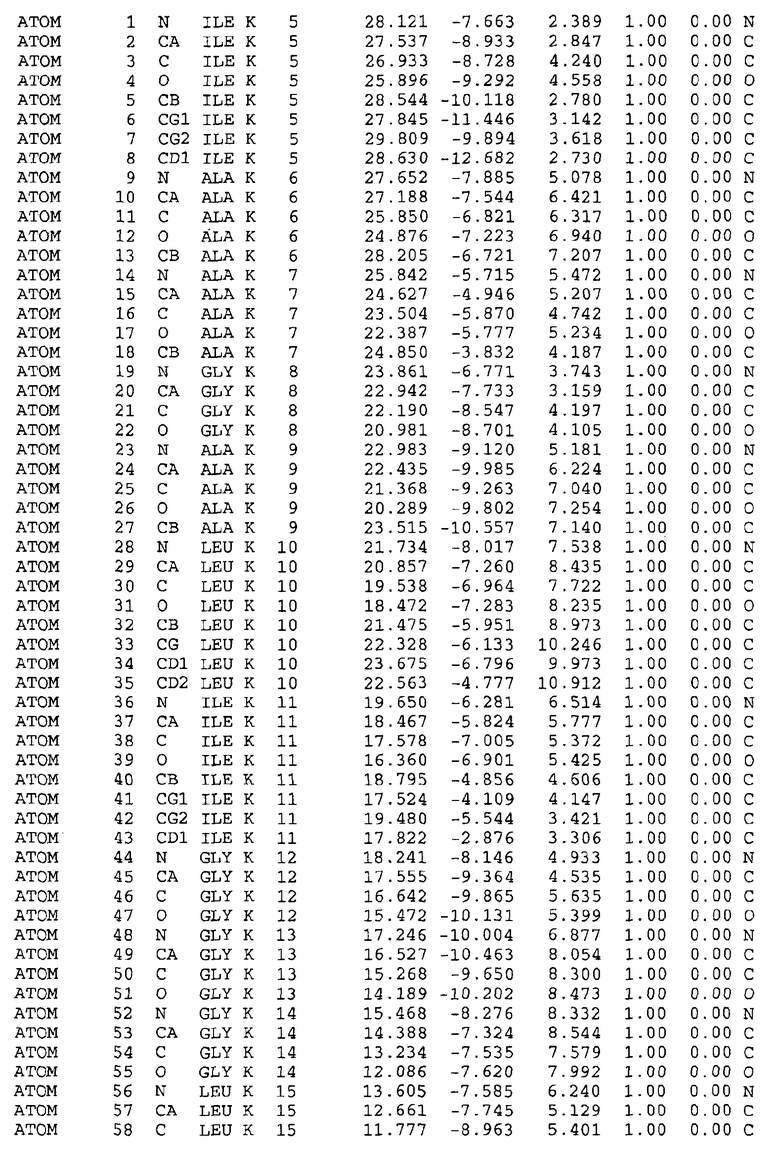
ТАБЛИЦА 7 -ПРОДОЛЖЕНИЕ
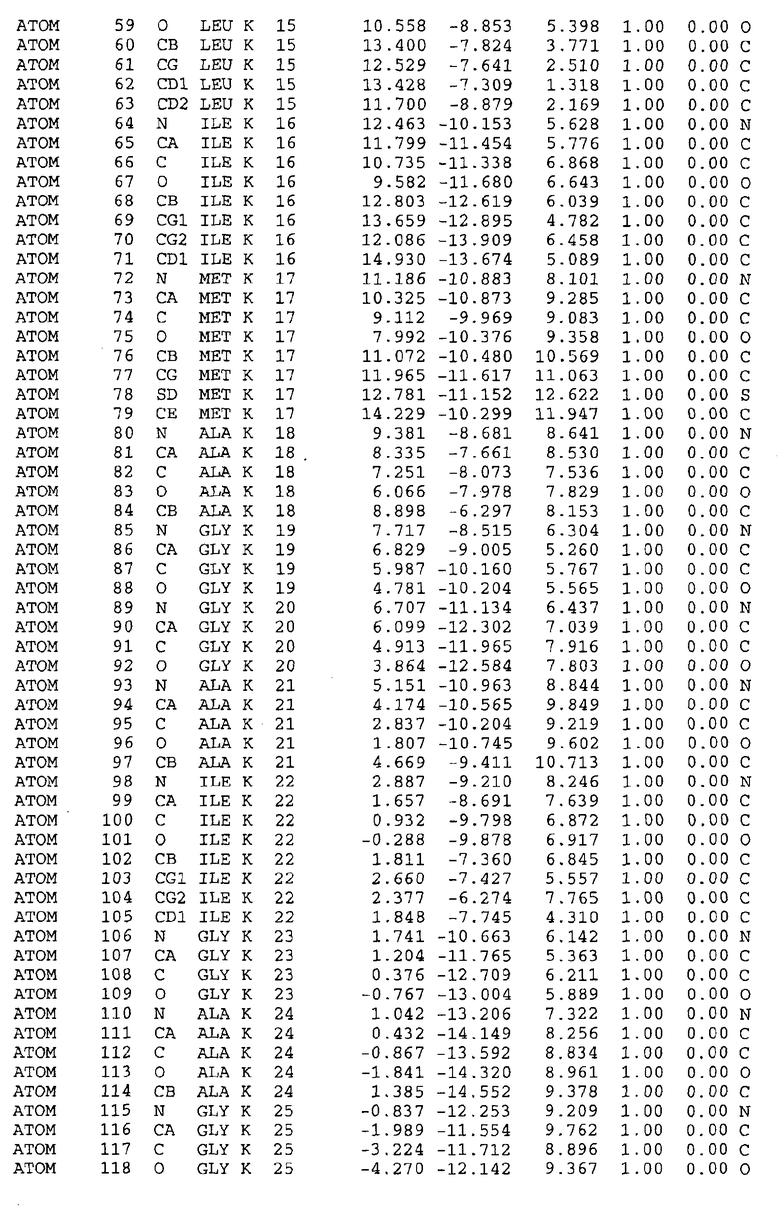
ТАБЛИЦА 7 -ПРОДОЛЖЕНИЕ
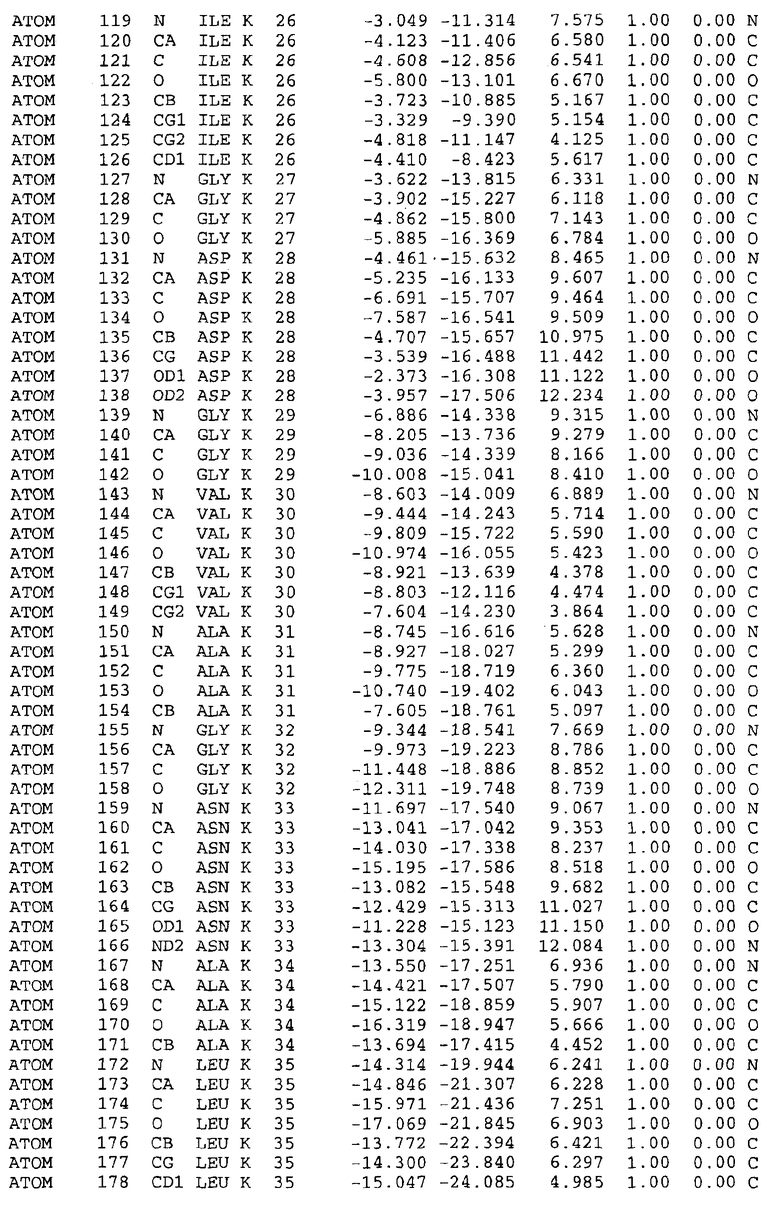
ТАБЛИЦА 7 -ПРОДОЛЖЕНИЕ
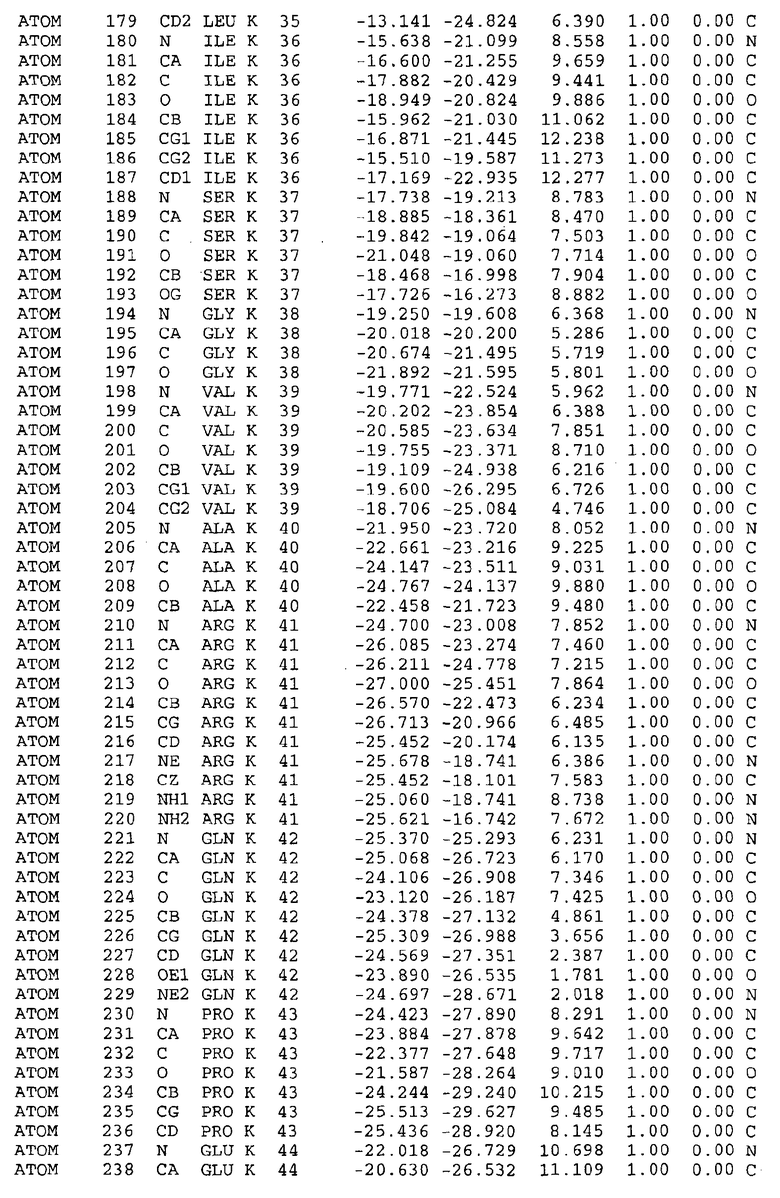
ТАБЛИЦА 7 -ПРОДОЛЖЕНИЕ
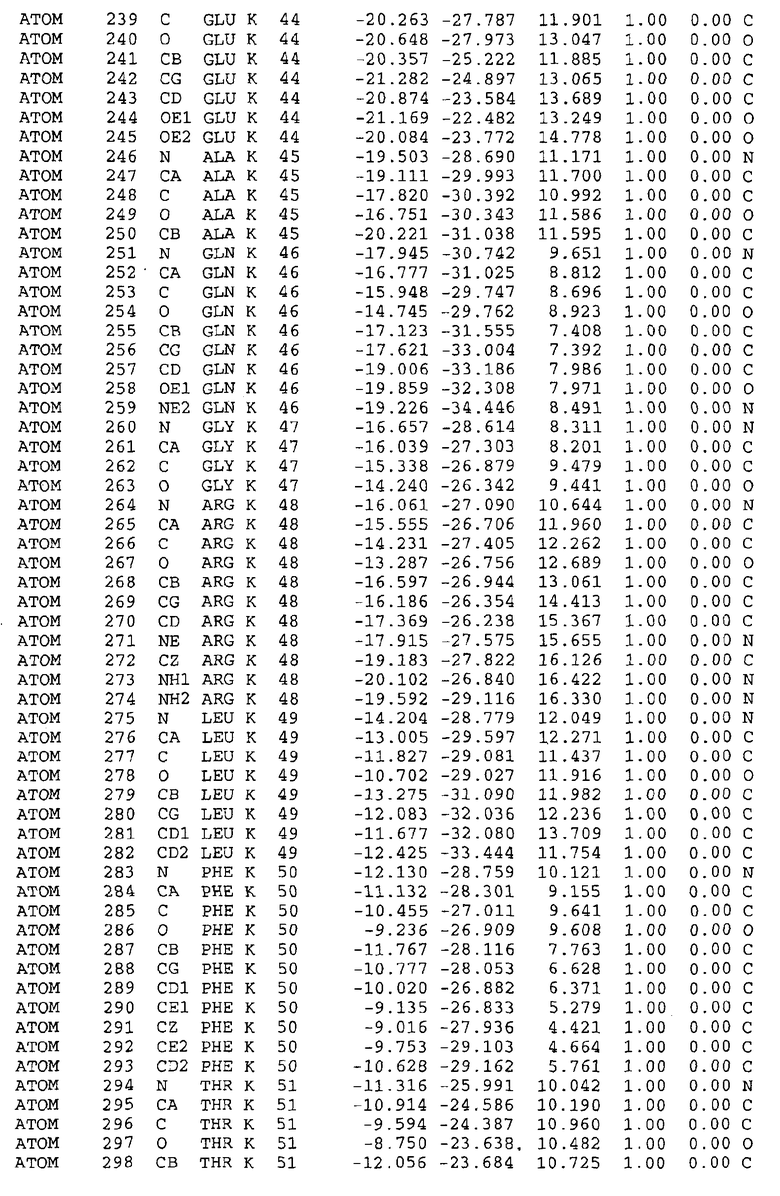
ТАБЛИЦА 7 -ПРОДОЛЖЕНИЕ
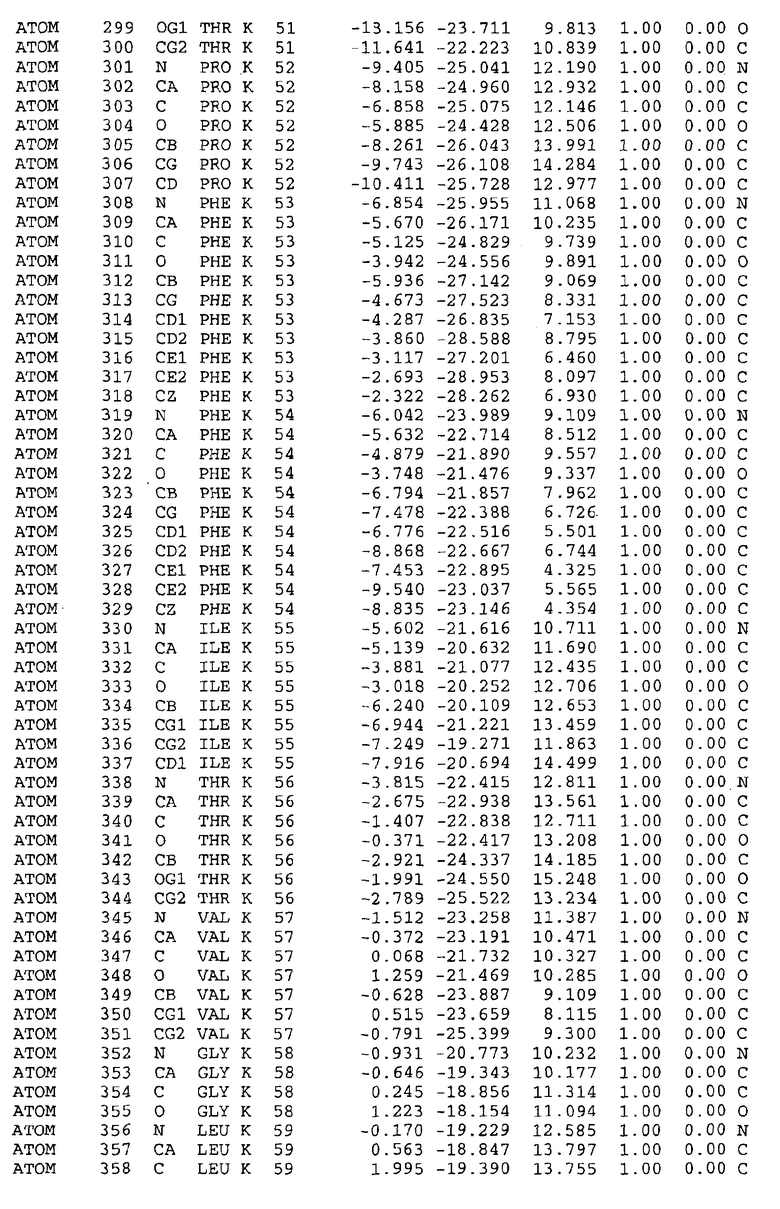
ТАБЛИЦА 7 -ПРОДОЛЖЕНИЕ
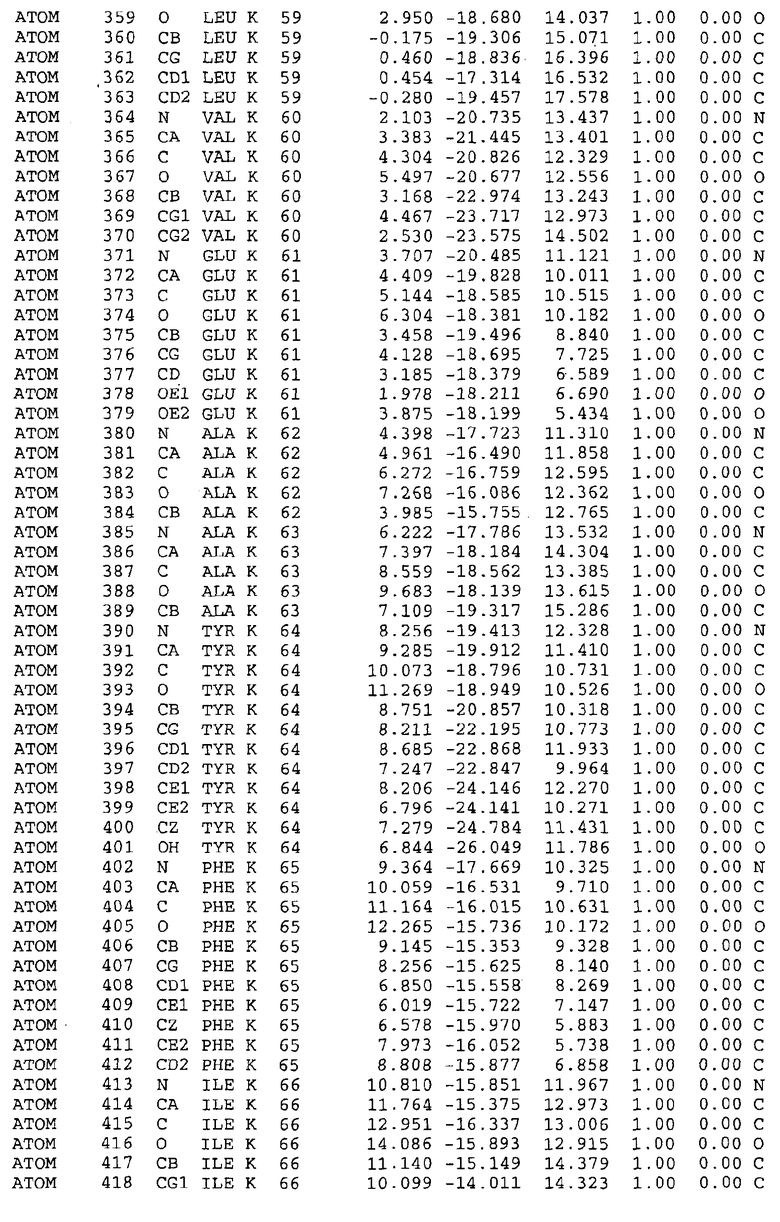
ТАБЛИЦА 7 -ПРОДОЛЖЕНИЕ
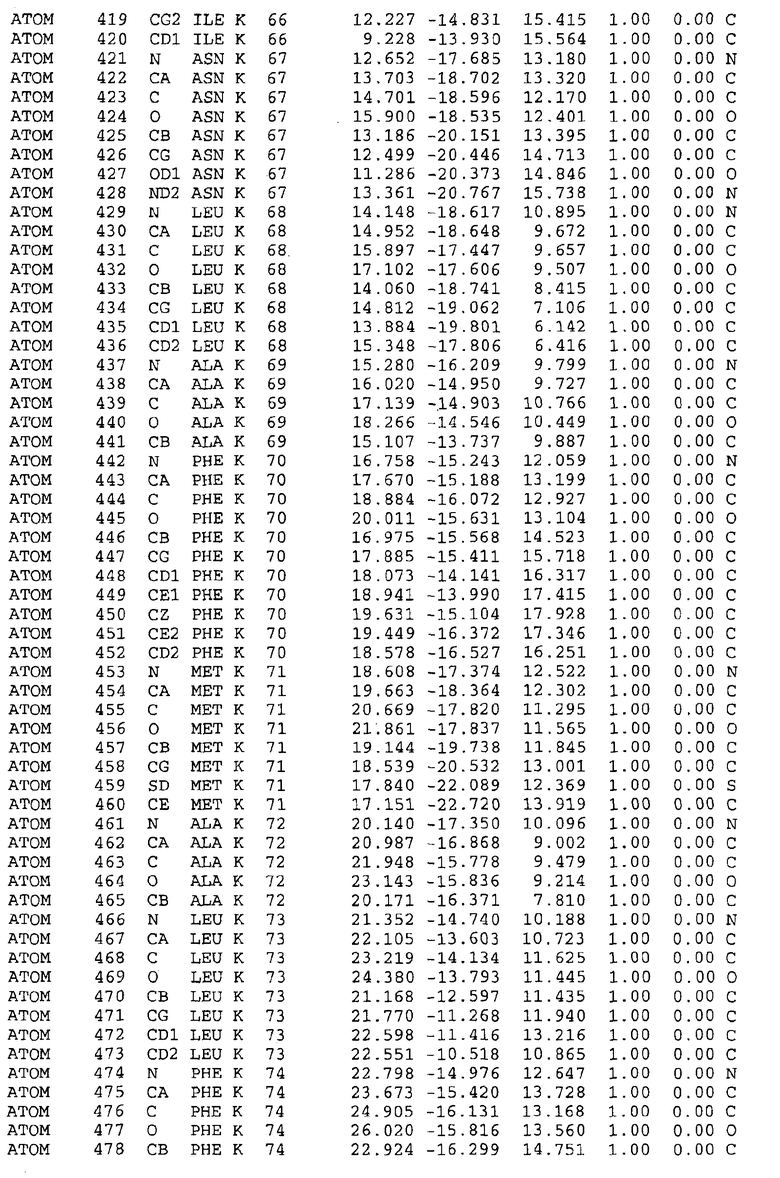
ТАБЛИЦА 7 -ПРОДОЛЖЕНИЕ
Источники информации
1. Global Alliance for TB Drug Development, Developing a faster TB cure (2004; http://www.tballiance.org).
2. E. L. Corbett et al, Arch. Intern. Med. 163, 1009 (2003).
3. UNMDS, AIDS epidemic update 2003 (2003; www.unaids.org/Unaids/EN/Resources).
4. World Health Organization, Tuberculosis (2004; http://www.who.int/health topics/tuberculosis/en/).
5. R. J. O'Brien, P. P. Nunn, Am. J. Respir. Crit. Care Med. 163, 1055 (2001).
6. World Health Organization, Tuberculosis Fact Sheet No 104 (2004; http://www.who.int/mediacentre/factsheets/fs104/en/).
7. A. J. Claxton, J. Cramer, C. Pierce, Clin. Ther. 23, 1296 (2001).
8. N. Lounis et al., Antimicrob. Agents Chemother. 45, 3482 (2001).
9. A. S. Ginsburg, J. H. Grosset, W. R. Bishai, Lancet Infect. Dis. 3, 432 (2003).
10. C. K. Stover et al, Nature 405, 962 (2000).
11. Guillemont J, Emile G, Patent (International Publication Number. WO 2004/011436. International Publication Date 5 February, 2004).
12. Barbachyn MR, Brickner SJ, Patent (International Publication Number: WO 93/09103, International Publication Date: 13 May 1993, International Application Number: PCT/US92/08267, International Filing Date: 1992).
13. R. Jain, B. Vaitilingam, A. Nayyar, P. B. Palde, Bioorg. Med. Chem. Lett. 13, 1051 (2003).
14. C. M. Kunin, W. Y. Ellis, Antimicrob. Agents Chemother. 44, 848 (2000).
15. L. Savini, L. Chiasserini, A. Gaeta, C. Pellerano, Bioorg. Med Chem 10, 2193 (2002).
16. S. Vangapandu, M. Jain, R. Jain, S. Kaur, P. P. Singh, Bioorg. Med. Chem. 12, 2501 (2004).
17. K. Andries et al, Science 307, 223 (2005).
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИНА С УЛУЧШЕННОЙ ЭФФЕКТИВНОСТЬЮ | 2004 |
|
RU2342400C2 |
| НОВАЯ МУТАНТНАЯ КАРБАМОИЛФОСФАТСИНТЕТАЗА И СПОСОБ ПРОДУКЦИИ СОЕДИНЕНИЙ - ПРОИЗВОДНЫХ КАРБАМОИЛФОСФАТА | 2001 |
|
RU2264459C2 |
| НОВЫЕ РЕГУЛИРУЮЩИЕ ЭКСПРЕССИЮ ПОСЛЕДОВАТЕЛЬНОСТИ И ПРОДУКТ ЭКСПРЕССИИ В ОБЛАСТИ ГИФОМИЦЕТОВ | 2001 |
|
RU2272835C2 |
| САМОИНДУЦИРУЕМАЯ ЭКСПРЕССИОННАЯ СИСТЕМА И ЕЕ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ ПОЛЕЗНЫХ МЕТАБОЛИТОВ С ПОМОЩЬЮ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae | 2013 |
|
RU2549708C2 |
| МУТАНТНАЯ АЦЕТОЛАКТАТСИНТАЗА И СПОСОБ ПРОДУКЦИИ РАЗВЕТВЛЕННЫХ L-АМИНОКИСЛОТ | 2006 |
|
RU2355763C2 |
| ПОВЫШЕНИЕ ВЫХОДА ВИРУСОПОДОБНЫХ ЧАСТИЦ В РАСТЕНИЯХ | 2012 |
|
RU2682752C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИНОЗИНА И 5'-ИНОЗИНОВОЙ КИСЛОТЫ МЕТОДОМ ФЕРМЕНТАЦИИ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ Escherichia | 2003 |
|
RU2271391C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE, В КОТОРОЙ ОСЛАБЛЕНА ЭКСПРЕССИЯ ГЕНОВ, КОДИРУЮЩИХ ТРАНСПОРТЕР ЛИЗИНА/АРГИНИНА/ОРНИТИНА | 2011 |
|
RU2468083C1 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2016 |
|
RU2745306C2 |
| Новый промотор и его применение | 2017 |
|
RU2733425C1 |
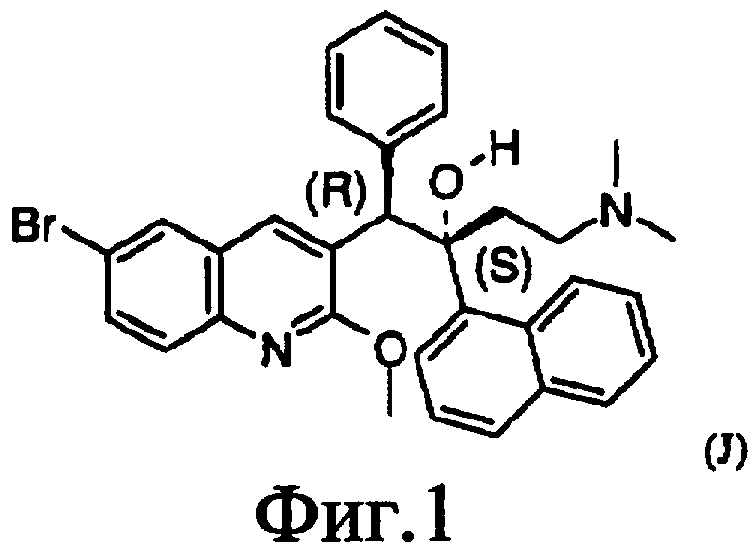
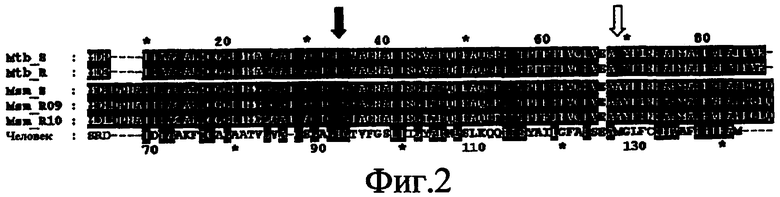
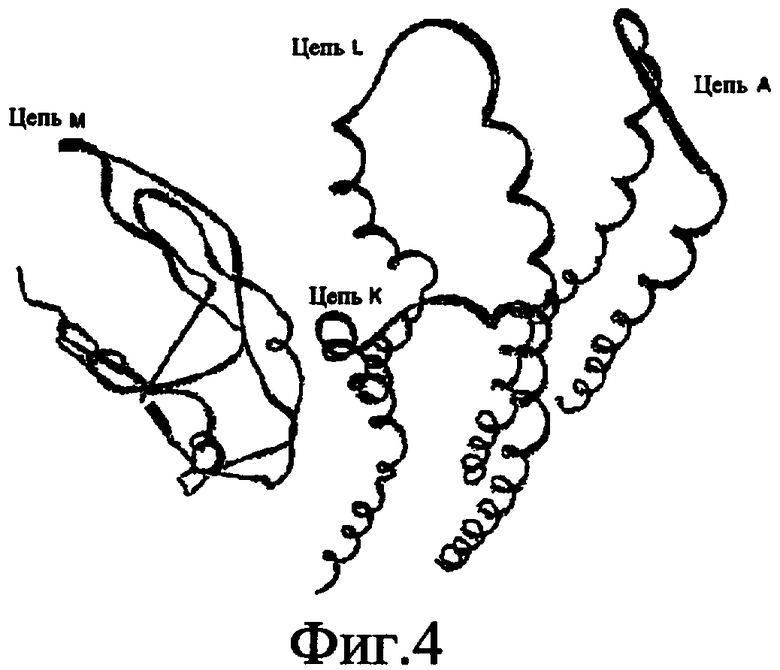
Настоящее изобретение относится к области генетической инженерии и может быть использовано в медико-биологической промышленности при поиске и/или разработке новых антимикобактериальных препаратов. Определены мутации аминокислотной последовательности белка atpE микобактерий (М.tuberculosis и М.smegmatis), определяющие их устойчивость к DARQ J. Получены кодирующие мутантные формы atpE, нуклеиновые кислоты, содержащие их векторы, клетки-хозяева, экспрессирующие мутантные белки. Предложено использование указанных клеток-хозяев в способе идентификации соединений, которые могут быть применены в качестве антимикробных агентов для резистентных к известным препаратам штаммов микобактерий. 5 н.п. ф-лы, 4 ил., 7 табл.
1. Выделенный мутантный белок atpE, определяющий резистентность клетки к DARQ J, с аминокислотной последовательностью, выбранной из SEQ ID No. 2 (Mtb_R), SEQ ID No. 4 (Мsm_R09) и SEQ ID No. 5 (Msm_R10).
2. Выделенная последовательность нуклеиновой кислоты, кодирующая выделенный мутантный белок atpE и характеризующаяся нуклеотидной последовательностью, определяющей аминокислотную последовательность белка по п.1.
3. Вектор экспрессии, содержащий последовательность нуклеиновой кислоты по п.2.
4. Клетка-хозяин, экспрессирующая мутантный белок atpE, несущая вектор по п.3.
5. Способ идентификации антимикобактериального агента, включающий:
(a) контактирование клеток, экспрессирующих белок atpE по п.1, с исследуемым соединением при физиологических условиях;
(b) определение того, взаимодействует ли исследуемое соединение с белком atpE;
(c) составление заключения о возможности использования тестируемого соединения в качестве антимикобактериального агента в случае установления указанного в (b) взаимодействия.
| WO 2004011436 A (JANSSEN PHARMACEUTICA N.V.), 05.02.2004 | |||
| GERUM ET AL | |||
| Antimicrobial Agents and Chemotherapy, 2002, 3362-3369 | |||
| HUMBERT R | |||
| ET AL | |||
| J.Bacteriol., 1989, 171 (3), 1435-1444 | |||
| RU 2002128741 A, 27.03.2004. |

