Относящиеся к этому заявки
Данная заявка связана с патентной заявкой США с серийным номером 09/524006, зарегистрированной 13 марта 2000 г., которая включена здесь в качестве ссылки, как здесь полностью объяснено.
Область техники, к которой относится изобретение
В настоящем изобретении предлагаются усовершенствованные векторы с зависимой от условий репликацией, способы их получения, способы их размножения, селективной упаковки, модификации и применение векторов с зависимой от условий репликацией и лентивирусных векторов. Важно, что векторы обладают пониженной способностью становиться компетентными по репликации и при применении обладают, таким образом, повышенной безопасностью. Предлагаются также специфические последовательности нуклеотидов и аминокислот, относящиеся к векторам, фармацевтические композиции и клетки-хозяева, включающие векторы, применение таких клеток-хозяев для скрининга лекарств и способы применения векторов для определения функции гена. Способы также включают профилактическое и терапевтическое лечение заболевания, особенно вирусной инфекции и, в частности, HIV инфекции. Изобретение также направлено на прямые и непрямые способы создания новых вакцин для лечения инфекционных заболеваний, рака и других заболеваний, имеющих отношение к генетике, и на способы диагностики. Другие способы и композиции изобретения включают применение векторов с зависимой от условий репликацией и лентивирусных векторов для генно-инженерной терапии и для других целей.
Предпосылки изобретения
Открытие того, что вирус иммунодефицита человека (HIV), лентивирус, является причиной синдрома приобретенного иммунодефицита (СПИДа), способствовало всплеску исследований механизмов, лежащих в основе инфекционного цикла вируса и патогенеза вирусного заболевания. Исследование данных механизмов обеспечило исследователей все возрастающим количеством мишеней для разработки противовирусных агентов, эффективных не только против HIV, но и против продуктов HIV, его генома, а также других вирусов. Данные противовирусные агенты, особенно те, которые направлены против HIV, могут быть разделены на группы в зависимости от способа их действия. Такие группы включают ингибиторы обратной транскриптазы, соединения, конкурирующие с вирусом за вход в клетки, вакцины, ингибиторы протеаз, а также недавно появившуюся группу, обозначаемую здесь как "генетические противовирусные агенты".
Обычно каждый тип противовирусного агента имеет свои собственные преимущества и ограничения, и они должны быть оценены исходя из необходимости лечения при конкретной ситуации. Противовирусные агенты, такие как зидовудин (3'-азидо-3'-дезокситимидин, также известный как AZT), ингибиторы протеаз и тому подобное могут быть доставлены в клетки организма больного с относительной легкостью, и они интенсивно изучаются. Имея мишенью один конкретный фактор в инфекционном цикле вируса, такие агенты, как доказано, относительно малоэффективны против HIV. Это в первую очередь обусловлено тем фактом, что штаммы HIV быстро изменяются и становятся устойчивыми к агентам, имеющим единственный локус воздействия (Richman, AIDS Res. and Hum. Retrovir., 8, 1065-1071 (1992)). Соответственно, проблемы генетической вариабельности и быстрой мутации в геномах HIV вынуждают разрабатывать новые стратегии противовирусной терапии для лечения HIV инфекций. Среди данных стратегий привлекательными являются генетические противовирусные агенты, так как они работают на множестве различных внутриклеточных уровней.
Генетические противовирусные агенты отличаются от других терапевтических агентов тем, что они переносятся в клетку-мишень как молекулярные элементы, где они защищают клетку от вирусной инфекции (Baltimore, Nature, 325, 395-396 (1988); и Dropulic' et al., Hum. Gene Ther., 5, 927-939 (1994)). Генетические противовирусные агенты могут представлять собой любую генетическую последовательность и включают, но не ограничиваются этим, антисмысловые молекулы, ловушки РНК, трансдоминантные мутанты, интерфероны, токсины, иммуногены и рибозимы. В частности, рибозимы являются генетическими противовирусными агентами, подобными антисмысловым, которые расщепляют РНК-мишени, включая РНК HIV, специфически зависимым от последовательности образом. Специфичность опосредованного рибозимами расщепления РНК-мишени предполагает возможность применения рибозимов в качестве терапевтических ингибиторов репликации вирусов, включая репликацию HIV. Различные типы рибозимов, такие как рибозимы в форме головки молотка и шпильки для волос, применяли в анти-HIV стратегии (см., например, патенты США № 5144019, 5180818 и 5272262 и патентные заявки PCT № WO 94/01549 и WO 93/23569). Как рибозимы в форме головки молотка, так и рибозимы в форме шпильки для волос могут быть сконструированы для разрушения любой РНК-мишени, которая содержит последовательность GUC (Haseloff et al., Nature, 334, 585-591 (1988); Uhlenbeck, Nature, 334, 585 (1987); Hampel et al., Nuc. Acids Res., 18, 299-304 (1990); и Symons, Ann. Rev. Biochem., 61, 641-671 (1992)). В общем виде рибозимы в форме головки молотка имеют два типа функциональных доменов, консервативный каталитический домен с двумя примыкающими гибридизационными доменами. Гибридизационные домены связываются с последовательностями, окружающими последовательность GUC, и каталитический домен расщепляет РНК-мишень с 3' конца от последовательности GUC (Uhlenbeck (1987), выше; Haseloff et al. (1988), выше; и Symons (1992), выше).
Ряд исследований подтвердил, что рибозимы могут быть, по меньшей мере, частично эффективными при ингибировании размножения HIV в клетках тканевых культур (смотри, например, Sarver et al., Science, 247, 1222-1225 (1990); Sarver et al., NIH Res., 5, 63-67 (1993a); Dropulic' et al., J. Virol., 66, 1432-1441 (1992); Dropulic' et al., Methods: Comp.Meth. Enzymol., 5, 43-49 (1993); Ojwang et al., PNAS, 89, 10802-10806 (1992); Yu et al., PNAS, 90, 6340-6344 (1993); и Weerasinghe et al., J. Virol., 65, 5531-5534 (1991)). В частности Sarver et al. ((1990), выше) показали, что рибозимы в форме головки молотка, предназначенные для расщепления в транскрибируемой области гена gag HIV, т.е. рибозимы против gag, могут специфически расщеплять РНК gag HIV in vitro. Более того, когда клеточные линии, экспрессирующие рибозимы против gag, инфицировали HIV-1, наблюдали от 50- до 100-кратное ингибирование репликации HIV. Сходно Weerasinghe et al. ((1991), выше) показали, что ретровирусные векторы, кодирующие рибозимы, предназначенные для расщепления в последовательности U5 РНК HIV-1, придают трансдуцированным клеткам устойчивость к HIV при последующем инфицировании HIV. Хотя различные клоны трансдуцированных клеток проявляли разные уровни устойчивости к инфицированию, что определялось с помощью промоторной системы, применяемой для управления экспрессией рибозимов, большинство клеточных линий, экспрессирующих рибозимы, становятся способными экспрессировать HIV после ограниченного времени культивирования.
Трансдукция клеток тканевых культур провирусом в ген nef (который не является необходимым для репликации вирусов в тканевой культуре), в который был введен рибозим, гибридизационные домены которого были специфичными для области U5 HIV, как показано, ингибирует репликацию вирусов в трансдуцированных клетках в 100 раз по сравнению с клетками, трансдуцированными провирусами дикого типа (смотри, например, Dropulic' et al. (1992) и (1993), выше). Сходно, рибозимы в форме шпильки для волос, как показано, ингибируют репликацию HIV в Т-клетках, трансдуцированных векторами, содержащими рибозимы в форме шпильки для волос U5, и инфицированных HIV (Ojwang et al. (1992), выше). Другие исследования показали, что векторы, содержащие рибозимы, экспрессирующиеся с промотора тРНК, также ингибируют различные штаммы HIV (Yu et al. (1993), выше).
Доставка рибозимов или других генетических противовирусных агентов к клеточным мишеням HIV инфекции (например, к T-клеткам CD4+ и моноцитам/макрофагам) являлась существенным препятствием для эффективного генетического терапевтического лечения СПИДа. Современные подходы к направленной доставке к клеткам гемопоэтической системы (т.е. к первичным мишеням для HIV инфекции) требуют введение терапевтических генов в предшественник мультипотентных стволовых клеток, который в процессе дифференцировки дает ряд зрелых Т-клеток или, в противоположном варианте, превращается в сами зрелые Т-лимфоциты CD4+. Направленная доставка в стволовые клетки является, однако, проблематичной, так как клетки трудно культивировать и трансдуцировать in vitro. Направленная доставка в циркулирующие Т-лимфоциты также является проблематичной, так как данные клетки настолько широко распространены, что трудно охватить все клетки-мишени, применяя имеющиеся в настоящее время векторные системы доставки. Более того, макрофаги необходимо рассматривать как клеточные мишени, поскольку они представляют собой главный источник распространения вирусов к другим органам. Однако, так как макрофаги находятся на конечной стадии дифференцировки и, следовательно, не способны к клеточному делению, их нелегко трансдуцировать обычно применяемыми векторами.
Соответственно, преобладающий в настоящее время подход к лечению HIV основывается на применении вирусных векторов с дефектом репликации и упаковывающих (т.е. "хелперных") клеточных линий (смотри, например, Buchschacher, JAMA, 269(22), 2880-2886 (1993); Anderson, Science, 256, 808-813 (1992); Miller, Nature, 357, 455-460 (1992); Mulligan, Science, 260, 926-931 (1993); Friedmann, Science, 244, 1275-1281 (1989); и Cournoyer et al., Ann. Rev. Immunol., 11, 297-329 (1993)) для введения в клетки, восприимчивые к вирусной инфекции (такой как HIV инфекция), чужеродного гена, который специфически вмешивается в репликацию вирусов или который вызывает гибель инфицированной клетки (обобщено Buchschacher (1993), выше). Такие вирусные векторы с дефектом репликации содержат в дополнение к интересующему чужеродному гену цис-действующие последовательности, необходимые для репликации вирусов, но не содержат последовательности, которые кодируют важнейшие вирусные белки. Следовательно, такой вектор не способен завершить цикл репликации вируса и для его размножения применяется хелперная клеточная линия, которая содержит и конститутивно экспрессирует вирусные гены в своем геноме. После введения вирусного вектора с дефектом репликации в хелперную клеточную линию белки, требуемые для образования вирусной частицы, предоставляются в ней вектору и продуцируются векторные вирусные частицы, способные инфицировать клетки-мишени и экспрессировать в них ген, который вмешивается в репликацию вирусов или вызывает гибель инфицированной вирусом клетки.
Такие ретровирусные векторы с дефектом репликации включают аденовирусы и вирусы, ассоциированные с аденовирусами, а также те ретровирусные векторы, которые применяют в клинических испытаниях при генной терапии HIV, и, в частности, мышиный амфотропный ретровирусный вектор, известный как вирус мышиного лейкоза Молони (MuLV). Данные дефектные вирусные векторы применяли для трансдукции клеток CD4+ генетическими противовирусными агентами, такими как рибозимы против HIV, с различной степенью успешности (Sarver et al. (1990), выше; Weerasinghe et al. (1991), выше; Dropulic' et al. (1993), выше; Ojwang et al. (1992), выше; и Yu et al. (1993), выше). Однако применение данных векторов для генной терапии HIV существенно ограничено. Например, высокая частота трансдукции особенно важна для лечения HIV, когда вектор должен трансдуцировать либо редкие гемопоэтические стволовые клетки-предшественники CD34+, либо широко рассеянные клетки-мишени CD4+, большинство из которых в течение клинически "латентной" стадии заболевания уже инфицированы HIV. Векторы MuLV, однако, трудно получить с высоким титром, следовательно, результатом является редкая трансдукция. Более того, не получена длительная экспрессия трансдуцированной ДНК в стволовых клетках-предшественниках CD34+, в частности, после их дифференцировки в зрелые Т-лимфоциты. Кроме того, применение дефектных вирусных векторов требует стратегий переноса генов ex vivo (смотри, например, патент США No. 5399346), что может быть дорогостоящим и недоступным по цене для общей популяции.
Данные недостатки, связанные с применением доступных в настоящее время векторов для генного терапевтического лечения СПИДа, привели ученых к поиску новых вирусных векторов. Одним таким вектором является сам HIV. Векторы HIV применяли для исследований инфекционности (Page et al., J. Virol., 64, 5270-5276 (1990)) и для введения генов (таких как гены-самоубийцы) в клетки CD4+, особенно в клетки CD4+, инфицированные HIV (смотри, например, Buchschacher et al., Hum. Gener. Ther., 3, 391-397 (1992); Richardson et al., J. Virol., 67, 3997-4005 (1993); Carroll et al., J. Virol., 68, 6047-6051 (1994); и Parolin et al., J. Virol., 68, 3888-3895 (1994)). Стратегия данных исследований заключается в применении HIV-векторов для введения генов в Т-клетки CD4+ и моноциты.
В настоящий момент, однако, данные векторы являются крайне сложными. Более того, применение данных векторов сопровождается риском выработки HIV дикого типа в результате внутриклеточной рекомбинации. Совместная трансфекция/совместное инфицирование последовательностями дефектного вектора и хелперным вирусом, как обнаружено, ведет к рекомбинации между гомологичными областями вирусных геномов (Inoue et al., PNAS, 88, 2278-282 (1991)). Выявленная комплементация in vitro указывает на то, что сходный вектор с дефектной репликацией HIV может рекомбинировать in vivo, обостряя таким образом уже существующую HIV-инфекцию. Тот факт, что в ретровирусах упаковано два генома РНК в одном вирионе, привел исследователей к предположению о том, что ретровирусы несут две вирусных РНК для обхода любых генетических дефектов, вызываемых комплементацией и/или рекомбинацией (Inoue et al., (1991), выше).
В дополнение к риску внутриклеточной рекомбинации, в результате чего возникает HIV дикого типа, HIV-векторы имеют ассоциированный риск мутации in vivo, что увеличивает патогенность вирусного вектора. Это привело Sarver et al. (AIDS Res. and Hum. Retrovir., 9, 483-487 (1993b)) к предположению, касающемуся разработки второго поколения рекомбинантных HIV-векторов, которые компетентны в отношении репликации, но еще не патогенны. Такие векторы по сравнению с преимущественно используемыми нереплицирующимися векторами (т.е. векторами с дефицитом репликации) продолжают реплицироваться у больного, создавая таким образом постоянную конкуренцию HIV дикого типа. Однако до настоящего времени такие векторы не доступны.
В идеале наилучшая возможность лечения инфицированного индивидуума имеется в момент инокуляции, до того даже как вирус инфицирует хозяина. Однако это трудно достичь ввиду того, что многие индивидуумы не понимают, что они стали инфицированными HIV до клинической латентной фазы заболевания. Основываясь на этом, стадия, на которой противовирусное вмешательство максимально необходимо, представляет собой стадию клинически латентного периода. Терапия на данной стадии требует противостояния заражению, представленному большим числом уже инфицированных лимфоцитов CD4+, которые несут вирусные геномы. Это не обычное заражение, о чем свидетельствует тот факт, что к настоящему времени HIV остается неизлечимым и только в малой степени поддающимся лечению с помощью доступных к настоящему времени способов терапии. Эффективная вакцина не появляется и, хотя ингибиторы обратной транскриптазы и протеазы, как показано, предотвращают репликацию HIV в тканевой культуре, развитие устойчивости вируса in vivo ведет к несостоятельности лечения. Таким образом, терапия геном HIV может приносить небольшую пользу подавляющему большинству индивидуумов, инфицироанных HIV, которых к настоящему времени более 30 миллионов.
В свете представленного выше становится все более важным получение длительных и устойчивых иммунологических ответов на определенные патогены, особенно вирусы, в частности в связи, например, со СПИДом и раком. Рассматривались живые аттенуированные (LA) вакцины с применением компетентных в отношении репликации, но непатогенных вирусов (Daniel et al., Science, 258, 1938-1941 (1992); и Desrosiers, AIDS Res. & Human Retrovir., 10, 331-332 (1994)). Однако такие непатогенные вирусы, которые отличаются от соответствующих вирусов дикого типа делецией в одном или более генов, либо (i) не могут вызывать защитного иммунного ответа, потому что антиген не сохраняется (из-за того, что LA-вирус эффективно не реплицируется); или (ii) LA-вирус реплицируется, но имеет другой патогенный потенциал, что оценивается по способности LA-вируса вызывать заболевание в моделях молодых животных (Baba et al., Science, 267, 1823-1825 (1995)).
По указанным выше причинам остается необходимость в альтернативных профилактических и терапевтических модальностях лечения вирусной инфекции, особенно в плане СПИДа и рака. В настоящем изобретении предлагаются такие альтернативные способы в результате предложения вектора с зависимой от условий репликацией. В изобретении также предлагаются такие дополнительные способы, в которых может применяться такой вектор. В изобретении дополнительно предлагаются хелперные векторные конструкты, которые дополняют вектор с зависимой от условий репликацией, для обеспечения его репликации и упаковки в виде вирусных частиц и вирионов. Такие хелперные векторы модифицированы так, что рекомбинация с вектором с зависимой от условий репликацией сведена к минимуму. Описаны различные осуществления конструктов таких хелперных векторов. Данные и другие цели и преимущества настоящего изобретения, также как дополнительные черты изобретения, будут ясны из представленного здесь описания изобретения.
Краткое изложение существа изобретения
В настоящем изобретении предлагаются усовершенствованные векторы с зависимой от условий репликацией, а также усовершенствованные композиции и способы получения и применения указанных векторов. Вектор с зависимой от условий репликацией характеризуется способностью к репликации только в клетке-хозяине, которая является пермиссивной для репликации вектора. Предлагаемые здесь усовершенствованные векторы характеризуются повышенной безопасностью, обладая пониженным риском восстановления репликативной компетенции. Векторы как таковые могут также обозначаться векторами с "пониженной рекомбинацией".
В одном аспекте изобретения усовершенствованный вектор с зависимой от условий репликацией включает, по меньшей мере, одну последовательность нуклеиновой кислоты, присутствие, транскрипция или трансляция которой придает вектору в клетке-хозяине, разрешающей репликацию, селективное преимущество над штаммом вируса дикого типа, соответствующим вирусу, от которого произошел вектор. Предпочтительно усовершенствованный вектор с зависимой от условий репликацией включает, по меньшей мере, одну последовательность нуклеиновой кислоты, которая придает вектору селективное преимущество в отношении репликации над любым другим конкурирующим геномом или элементом генома. Применяемый здесь термин элемент генома определяется как последовательность нуклеиновой кислоты в клетке-хозяине, которая не происходит ни от вектора, ни от любого вируса дикого типа и которая может конкурировать, вмешиваться или влиять на репликацию вектора в клетке-хозяине. Элемент генома не является также полным геномом, таким как геном клетки-хозяина или другого вектора или вируса. Селективное преимущество для репликации придается благодаря присутствию, транскрипции или трансляции указанной последовательности нуклеиновой кислоты.
В другом предпочтительном осуществлении вектор с зависимой от условий репликацией представляет собой ретровирус, особенно лентивирус, и включает, по меньшей мере, одну последовательность нуклеиновой кислоты, присутствие, транскрипция или трансляция которой придает клетке-хозяину, которая инфицирована вектором, селективное преимущество над клеткой, инфицированной штаммом вируса дикого типа, соответствующим вирусу, от которого произошел вектор. В противоположном варианте вектор не является полностью производным штамма дикого типа, а является химерным, содержащим компоненты, которые происходят более чем от одного вируса дикого типа. Более чем один вирус дикого типа может представлять собой различные (или гетерологичные) вирусы или различные штаммы или изоляты одного вируса. Предпочтительно указанная последовательность нуклеиновой кислоты может быть экспрессирована для обеспечения профилактического эффекта в указанной клетке-хозяине. Это ведет к клетке, которой придано преимущество в жизнеспособности, так как последовательность нуклеиновой кислоты не дает любому инфицирующему вирусу реплицироваться до уровней, которые вызывают гибель клетки.
Другим предпочтительным осуществлением является усовершенствованный плазмидный вектор с зависимой от условий репликацией, включающий, по меньшей мере, одну последовательность нуклеиновой кислоты, которая придает вектору селективное репликативное преимущество над любой другой конкурирующей векторной или плазмидной молекулой. Например, такие векторы могут включать последовательность (такую как, например, рибозим или антисмысловую последовательность), способную расщеплять или разрушать или приводить к расщеплению или разрушению, что ведет к расщеплению конкурирующего вектора или плазмиды, когда они локализованы совместно с вектором. Если конкурирующий вектор и хелпер должны быть разрушены, то вектор изобретения сконструирован так, что содержит другие последовательности для придания ему полного селективного преимущества в репликации. Например, вектор изобретения может содержать антисмысловую последовательность, которая разрушает как конкурирующий вектор, так и хелпер, а вектор все еще будет иметь селективное преимущество в репликации, потому что он дополнительно содержит вторую первичную нуклеотидную последовательность (например, промотор, который продуцирует больше векторной РНК по сравнению с РНК конкурирующего вектора, число копий вектора в трансдуцированной клетке, большее, чем в конкурирующем геноме, или сигнал упаковки, который присутствует в векторе, но не в хелпере), что придает вектору с зависимой от условий репликацией полное селективное преимущество в репликации. Следовательно, по меньшей мере, первая нуклеотидная последовательность дополнительно обеспечивает полное селективное преимущество в репликации векторов изобретения в данном осуществлении, потому что две первичные нуклеотидные последовательности (т.е. первая первичная нуклеотидная последовательность, являющаяся мишенью конкурирующего вектора и/или хелпера, и вторая первичная нуклеотидная последовательность обеспечивает селективное преимущество в репликации) работают совместно для достижения эффекта полного селективного преимущества. Следовательно, синергические эффекты множественных первичных нуклеотидных последовательностей могут вести к полностью селективным условиям для вектора с зависимой от условий репликацией, где любая индивидуальная первичная нуклеотидная последовательность обеспечивает дискриминацию селекции по отношению к конкурирующему геному.
Расщепление или деструкция конкурирующего вектора или плазмиды снижает также возможность полноразмерной рекомбинации с вектором, который, таким образом, обладает свойством "пониженной рекомбинации". Вектор может быть защищен от расщепления путем индукции вырождения его последовательности для того, чтобы она не была мишенью рибозима или антисмысловой последовательности. Вектор может быть дополнительно защищен от расщепления путем конструирования вектора или геномной версии вектора для дифференциального отслеживания РНК, так что РНК геномного вектора существенно не расщепляется конкурирующим вектором или хелпером. В противоположном варианте может быть сходно и исключительно сконструирован хелпер путем, например, вставки элементов сплайсинга для внедрения хелпера в сплайсосому, при конструировании векторной геномной РНК для включения в нее первых последовательностей нуклеиновой кислоты, которые присутствуют только в не подвергнутой сплайсингу, но не в подвергнутой сплайсингу векторной РНК. Такие неограничивающие примеры усовершенствованных векторов особенно выгодны для применения при получении вирусного вектора или плазмидного вектора и клонирования или субклонирования последовательностей нуклеиновой кислоты в такие векторы, а также при их применении для мутагенеза, экспрессии индуцибельных генов и тому подобное.
В настоящем изобретении предлагается также фармацевтическая композиция, включающая вектор изобретения с зависимой от условий репликацией и фармацевтически приемлемый носитель. Дополнительно предлагается клетка-хозяин, включающая вирусный вектор с зависимой от условий репликацией. Предлагается также вектор, где указанный вектор, если это ДНК, включает нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NOS: 2, 3, 4, 5, 6, 14, в которых, по меньшей мере, один нуклеотид мутирован, 15 и 16, и где указанный вектор, если это РНК, включает нуклеотидную последовательность, кодируемую нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NOS: 2, 3, 4, 5, 6, 14, в которых, по меньшей мере, один нуклеотид мутирован, 15 и 16, в виде выделенных и очищенных молекул нуклеиновой кислоты, как здесь указано. Сходно предлагается способ конструирования вектора с рибозимом, способ модификации вектора и способ размножения и селективной упаковки вектора с зависимой от условий репликацией без использования упаковывающей клеточной линии.
В еще одном аспекте настоящего изобретения предлагается способ терапевтического и профилактического лечения клетки-хозяина от вирусной инфекции. В особенно предпочтительных осуществлениях такое лечение эффективно у указанного хозяина в результате придания доминантного фенотипа, который ингибирует вирусную инфекцию, или в результате придания фенотипа, который ингибирует инфицирование другими вирусами. Предпочтительно другие ингибируемые вирусы включают широкий спектр вирусных штаммов. Такие способы могут дополнительно включать применение хелперных экспрессионных векторов, также обозначаемых как "хелперный вектор" или "хелперный векторный конструкт", цитотоксического лекарства, белков/факторов или ингибитора протеазы/обратной транскриптазы, что подходит. Способ может быть применен, например, для ингибирования репликации вируса, для лечения заболеваний (включая рак, генетические, инфекционные, сосудистые и другие заболевания), для проведения эффективного переноса генов ex vivo и in vivo, для возможности получения безопасных векторов, для определения функции гена или для экспрессии интересующего гена в клетке-хозяине.
В еще одном аспекте изобретения предлагается способ применения клетки-хозяина, включающей вектор изобретения с зависимой от условий репликацией, для определения взаимодействия между лекарством/фактором и белком. Такой способ дает возможность характеризовать белок и осуществлять скрининг лекарств, факторов или других белков на активность или физические взаимодействия в отношении данного белка, кодируемого вектором. Способ дополнительно позволяет идентифицировать и охарактеризовать белки, которые являются функционально близкими с данным белком, кодируемым вектором. Например, если кодируемый белок является транскрипционным фактором, его экспрессия вектором изобретения позволяет идентифицировать гены, регулируемые транскрипционным фактором.
В изобретении предлагаются также композиции и условия для хранения векторов при различных условиях перед их применением. Примеры таких условий включают хранение при -80°С, -20°С и 4°С в присутствии различных носителей.
Дополнительные осуществления настоящего изобретения включают общие лентивирусные векторы, модификации конструктов хелперных векторов, которые служат для уменьшения, сведения к минимуму или исключения рекомбинации вектора-хелпера с вектором с зависимой от условий репликацией для создания репликационно компетентного вектора или вируса (RCV). Следовательно, изобретение включает хелперные векторы, которые служат для получения векторов со "сниженной рекомбинацией". Хелперный вектор изобретения также предпочтительно увеличивает титр получаемого вектора с зависимой от условий репликацией до уровней, больших чем 107 трансдуцирующих единиц на миллилитр. Другие осуществления включают концентрирование вектора с применением высокоскоростного (но не ультра) центрифугирования и хроматографических, ультрафильтрационных и диафильтрационных способов для концентрирования и очистки.
Модификации хелперного вектора изобретения включают вставку рибозима, такого как анти-U5 рибозим, или антисмысловой последовательности, которая расщепляет или ведет к разрушению или инактивации вектора с зависимой от условий репликацией в случае, когда хелперный вектор и вектор с зависимой от условий репликацией совместно локализованы или совместно упакованы. В одном осуществлении хелперные компоненты полностью интегрированы в одном плазмидном конструкте для создания двухплазмидной функциональной векторной хелперной системы, которая эффективно продуцирует неконцентрированный супернатант векторов с титрами, большими чем 107 трансдуцирующих единиц на мл (смотри фиг.3). В другом осуществлении нуклеотидная последовательность хелперного вектора является вырожденной для сведения к минимуму рекомбинации с вектором с зависимой от условий репликацией. В еще одном осуществлении хелперный вектор включает гетерологичные транс-активирующие элементы для применения при упаковке вектора с зависимой от условий репликацией. Примеры таких элементов включают, но не ограничиваются этим, белок оболочки вируса везикулярного стоматита VSV-G, белок оболочки RD114, белок оболочки вируса бешенства, белок оболочки вируса лейкоза человекообразных гиббонов (GALV) и химерные белки оболочки. Дополнительные модификации также включают гетерологичные элементы, чувствительные к rev (RREs), посттранскрипционные регуляторные элементы (PRE) или элементы конститутивного транспорта (CTEs).
В еще одном осуществлении конструкт хелперного вектора дополнительно включает донор сплайсинга, акцепторные сайты сплайсинга или сайты для вырожденных или гуманизированных нуклеотидных последовательностей. Например, сайты сплайсинга могут быть локализованы так, что сигнал упаковки и/или RRE может быть удален из транскрибируемых РНК в результате сплайсинга. Более того, сайты для вставки интронов в вектор или в хелперные конструкты предлагаются так, что димеризация, совместная локализация или рекомбинация между вектором с зависимой от условий репликацией и хелпером, или другим конкурирующим геномом или геномным элементом сводятся к минимуму. Вставка интронов может быть направлена на получение транспорта хелперной РНК в сплайсосомы и обратно из векторной РНК. Неожиданно оказалось, что некоторые конструкты хелперных векторов увеличивают титр вектора с зависимой от условий репликацией.
Краткое описание фигур
На фиг.1A-1K представлены схематические изображения конкретных векторов с зависимой от условий репликацией, охватываемых настоящим изобретением: pN1(cPT), pN1(cPTc)ASenvGFP(464), pN1(cPT)ASenvGFP(452), pN1(cPT2)ASenvGFP, pN1(cPT)cGFP, pN1GFP(cPT)T, pN1(cPT)GFPTAR, pN1GFP(cPT)VT, pN2GFP, pN2ASenvGFP(418) и pN2(spe)ASenvGFP. Разумеется, маркерный ген зеленого флуоресцентного белка (GFP) может быть удален перед использованием векторов для описанных здесь применений. Обозначения: N1, минимальный, ведущий свое начало от HIV-1 вектор без последовательностей gag/pol, но с упаковывающей последовательностью из gag, обозначаемой как gag' или gag", и кодоном терминации, помещенным на расстоянии приблизительно 40 пар оснований от ATG и последовательности gag; N2, вектор, берущий начало от HIV-1, способный экспрессировать последовательности gag/pol; AS, антисмысловая; ASenv, последовательность env, присутствующая в антисмысловой ориентации; gag, pol и env, кодирующая последовательность для белков, которые образуют сердцевину вируса, обратную транскриптазу и оболочку соответственно; tat, rev, rre и nef, дополнительные вирусные гены; cPTc, последовательность минимального центрального полипиримидинового тракта; cPT, центральный полипиримидиновый тракт, содержащий большую вставку из приблизительно 548 пар нуклеотидов; cPT2, центральный полипиримидиновый тракт, содержащий вставку из приблизительно 438 пар нуклеотидов (не включает последовательности gag в такой же степени, что и cPT); spe, нетранслируемые последовательности gag/pol; GFP, последовательность, кодирующая зеленый флуоресцентный белок.
На фиг.2 показаны последовательности ДНК U5-РНК дикого типа HIV SEQ ID NO:l (A) и модифицированной U5-РНК crHIV SEQ ID NO:2 (B). Числа относятся к номерам оснований ниже старта транскрипции.
Фиг.3A-3E иллюстрирует влияния отношения хелперного вектора к вектору с зависимой от условий репликацией (cr) на титр образующихся cr-векторов. На фиг.3A показано, что молярное отношение 1:0,5 обеспечивает наибольший титр pNl(cPTc)GFP. На фиг.3B и 3C показано, что молярное отношение 1:0,75 обеспечивает наибольший титр pNl(cPT)GFP и pNl(cPT2)ASenvGFP соответственно; а фиг.3D и 3E показывают, что молярное отношение 1:1,5 обеспечивает наибольший титр pNlcGFP и pN2cGFP соответственно. Векторы показаны на фиг.1, за исключением pNl(cPT)GFP, в котором нет антисмысловой последовательности env, и pNlcGFP и pN2cGFP, в которых имеется вставка промотора цитомегаловируса (CMV) для экспрессии GFP. На фигурах показано, что для двухплазмидной системы были получены условия для получения титров, составляющих, по меньшей мере, 1,5×107 трансдуцирующих единиц на мл.
На фиг.4A и 4B показаны карты векторов с зависимой от условий репликацией на основе HIV-2: pS1cGFP и pS2cGFP. Обозначения pS1 и pS2 относятся к отсутствию или присутствию последовательностей gag/pol, как описано выше, для pNl и pN2. Обозначение с указывает на наличие промотора CMV для управления экспрессией GFP.
На фиг.5A и 5B проиллюстрировано влияние различных конструктов хелперного вектора на упаковку pSlcGFP и pS2cGFP. Влияние различных молярных отношений хелперного вектора и вектора с зависимой от условий репликацией проверяли наряду с влиянием различных RRE на хелперный вектор. Как показано, применение отношения 1:1 хелперного вектора к вектору с зависимой от условий репликацией было более эффективным в образовании функциональных векторных частиц по сравнению с другими тестированными отношениями.
На фиг.6A-6G изображены структуры различных конструктов хелперного вектора: pVIRPAC-1, pVIRPAC-2, pVIRPAC-1.1Rz, pVIRPAC-1.2, VirPaс1.2Rz, pVIRPAC-l.2Rz2 и pVIRPAC 1.2RzIn, охватываемых настоящим изобретением. Rz относится к присутствию анти-U5 рибозима, а 1.1 и 1.2 относятся к хелперу, содержащему RRE, берущему начало от HIV-1 и HIV-2 соответственно.
На фиг.7 иллюстрируется влияние одного или более рибозимов на титры вирусного вектора. PN1(cPT)GFP упаковывали в клетках HeLa-tat в присутствии pvirPac1.2 (не содержащего рибозимы), pVirPac1.2RzIn (содержащего один рибозим и интрон), pVP1.2Rz (содержащего один рибозим) или pVP1.2Rz2 (содержащего два рибозима). Как показано на графике, присутствие одного рибозима (pVirPac1.2Rz) или рибозима и интрона, разработанного для влияния на клеточное перемещение хелперной РНК (pVirPac1.2RzIn), не оказывало существенного влияния на титр вектора. Анализ с помощью ПЦР образцов для титрования на совместную упаковку хелперных конструктов показал низкую совместную упаковку в присутствии рибозима по сравнению с высокой совместной упаковкой в отсутствие рибозима. Указатель "VirPac" может также обозначаться "VIRPAC" или "VP".
На фиг.8 иллюстрируется ингибирующее действие векторов из серий pNl и pN2 на репликацию HIV дикого типа в T-клетках. T-клетки сначала трансдуцировали вектором, а затем подвергали контрольному заражению вирусом дикого типа при кратности инфицирования 0,1 при 100% трансдуцированных клеток. Репликацию вируса оценивали с помощью теста ИФА поглощения антигена p24. pN2ASenvGFP показал сильное ингибирование репликации вируса дикого типа.
На фиг.9A и 9B показано сильное ингибирование репликации HIV дикого типа векторами на основе pNl и pN2. На фиг.9A показано сильное ингибирование HIV дикого типа в T-клетках человека под действием pNlGFP(cPT)VT и pN2ASenvGFP по сравнению с контрольными опухолевыми клетками после инфицирования HIV дикого типа. На фиг.9B показаны сходные результаты на T-клетках с pN1(cPT)ASenvGFP и pN2ASenvGFP.
На фиг.10A и 10B показаны векторы и их применение для отбора трансдуцированных клеток. На фиг.10A показана организация применяемых векторов, где pN1CMIG и pN1MCG содержат внутренний промотор CMV, в то время как pN1MIG и pN1MIG-W экспрессируют ген MGMT посредством промотора HIV-LTR. На фиг.10B показан график разрастания клеток SupT1, трансдуцированных указанными выше векторами и подвергнутыми селекции с помощью BG и BCNU, как описано ниже в примере 5.
На фиг.11, панели A-F, показана селекция трансдуцированных первичных клеток CD4+ с помощью BG и BCNU. Клетки CD4+, трансдуцированные pN2MIG, культивировали в присутствии 0, 0,5, 2, 5, 10 и 10 мкМ BG (показано на панелях A-F, соответственно), с указанными концентрациями BCNU. На панели F дополнительно показаны результаты разведения трансдуцированных клеток 1:5 с последующей обработкой 10 мкМ BG и указанными концентрациями BCNU. Исходный уровень 3% клеток GFP+ на панели F возрастал, по меньшей мере, в 32 раза до 97%, уровня, который ранее не наблюдали in vitro.
На фиг.12 показаны эффекты рибозимов в предотвращении совместной упаковки хелперной РНК в препараты вектора. Содержащие вирус супернатанты экстрагировали с помощью «шумной экстракции» и подвергали перевариванию ДНКазой I с последующим количественным определением области gal-pol, найденной в хелперном конструкте, с помощью обратной транскрипции-ПЦР (ОТ-ПЦР). Как показано, присутствие одного или двух рибозимов в примененном хелперном конструкте (pVP1.2Rz или pVP1.2Rz2) значительно снижало количество совместно упакованного хелперного вектора по сравнению с хелпером без рибозима (pVP1.2).
На фиг.13A и 13B показана возможность очистки опухолевых клеток от стволовых клеток CD34+. На фиг.13A показано, что при MOI 10 pN1(cPT)ASenvGFP трансдуцировал 98.52% опухолевых клеток SupTl после одного раунда трансдукции. На фиг.13B показано, что при трансдукции клеток CD34+не наблюдалось значительной трансдукции после одного раунда трансдукции. Лишь после трех раундов наблюдалась значительная трансдукция. Обозначения: 1/2/A, например, относится к раунду 1 трансдукции при MOI 2 и присутствию вирусных вспомогательных белков в трансдуцируемом векторе, тогда как 3/50, например, относится к трем раундам трансдукции и MOI 50.
На фиг.14A показаны структуры VSV-G дикого типа, RD114 дикого типа и гибридных белков оболочки с указанными экстраклеточными, трансмембранными и цитоплазматическими доменами. На фиг.14B показаны титры векторов HIV-1, псевдотипированных в HT1080 различными белками оболочки (VSV-G дикого типа, вируса бешенства G, RD114 дикого типа и RD114E, гибридным конструктом VSV-G и RD114).
На фиг.15A-15E представлены схематические изображения связанных с упаковкой конструктов, охватываемых настоящим изобретением: p(CGCRSRRE), p(TREtTApuro), p(BI-RevTat), p(EH-GP) и p(CMV-GP). На фиг.15F показана организация зависимых от rev конструктов VSV-G.
На фиг.16 показан выход векторов pN1(cPT)GFP на клетку до и после концентрирования в клетках HeLa-tat. Можно видеть, что выход остается на уровне приблизительно 1010 при обоих условиях.
На фиг.17 показаны результаты применения системы Rev/RRE/CRS для контроля экспрессии VSV-G.
На фиг.18 показано влияние буфера для хранения на извлечение вектора после хранения в течение 3-5 недель при различной температуре. Можно видеть, что присутствие 10% трегалозы или 10% глюкозы в D-PBS или HBS обеспечивает хороший выход вектора после хранения при -20 или при -80°С.
Описание предпочтительных осуществлений
В настоящем изобретении предлагаются усовершенствованные векторы с зависимой от условий репликацией, лентивирусные векторы и их применение. В дополнение к способности к селективной репликации векторы содержат конкретные модификации для снижения вероятности рекомбинации, что ведет к тому, что векторы становятся репликационно компетентными. Включены способы ингибирования репликации штамма вируса дикого типа и способы генной доставки в клетки, включающие такие усовершенствованные векторы. Способ включает контактирование хозяина, который может быть инфицирован, имеет риск быть инфицированным или предпочтительно действительно инфицирован таким штаммом вируса дикого типа с усовершенствованным вектором, который размножается только в хозяине, который является пермиссивным для репликации вектора (т.е. непатогенного или не вызывающего болезнь вектора с зависимой от условий репликацией (cr)).
Как здесь дополнительно описывается, конкретная цель способа состоит в создании конкурентной инфекции у хозяина с помощью такого непатогенного вектора с зависимой от условий репликацией. Обычно вектор с зависимой от условий репликацией в соответствии с изобретением включает, по меньшей мере, одну последовательность нуклеиновой кислоты, которая придает селективное преимущество репликации и распространения вектору с зависимой от условий репликацией по сравнению с вирусом дикого типа, и/или, по меньшей мере, одну последовательность нуклеиновой кислоты, которая придает селективное преимущество размножения вирусных частиц в клетке-хозяине, содержащей вектор с зависимой от условий репликацией, по сравнению с клеткой-хозяином, содержащей вирус дикого типа.
В предпочтительном осуществлении изобретения вектор включает последовательность HIV и применяется для лечения HIV инфекции. Таким образом, вектор или клетка-хозяин, содержащая вектор, включает, по меньшей мере, одну последовательность нуклеиновой кислоты, которая (1) обеспечивает геном crHIV селективным преимуществом над геномом HIV дикого типа в отношении упаковки в вирионы-потомки (т.е. в клетках, в которых оба они являются резидентами), и/или (2) обеспечивает клетку-хозяин, продуцирующую вектор (вирус) с зависимой от условий репликацией, селективным преимуществом в отношении продукции вириона crHIV по сравнению с клеткой-хозяином, продуцирующей вирус дикого типа. Одним способом (которым изобретение не ограничивается) является придание геномам crHIV селективного преимущества в упаковке путем предложения их с одним или более рибозимов, способных расщеплять геном HIV дикого типа.
В другом аспекте изобретения предлагаются векторы с пониженной способностью к рекомбинации с диким типом и хелперными конструктами, что ведет к получению репликационно компетентных векторов. Кроме того, предлагаются хелперные векторы, клетки и упаковывающие системы, которые также вносят вклад в снижение рекомбинации, для дальнейшего снижения количества событий продуктивной рекомбинации, чтобы не давать векторам больше условий для репликации.
Вирус дикого типа
В соответствии с изобретением "вирус" представляет собой инфекционный агент, который состоит из белка и нуклеиновой кислоты и который использует генетический аппарат клетки-хозяина для продукции вирусных продуктов, определяемых вирусной нуклеиновой кислотой. "Нуклеиновая кислота" обозначает полимер ДНК или РНК, который является одноцепочечным или двухцепочечным, линейным или кольцевым и необязательно содержит синтетические, неприродные или модифицированные нуклеотиды, которые способны включаться в полимеры ДНК или РНК. Полинуклеотидная ДНК предпочтительно состоит из последовательностей геномной или кДНК.
"Штамм вируса дикого типа" представляет собой штамм, который не включает какие-либо индуцированные человеком мутации, как здесь описано, т.е. любой вирус, который может быть выделен из природных источников. В противоположном варианте штаммом дикого типа является любой вирус, который культивировался в лаборатории, но до сих пор в отсутствие любого другого вируса способен продуцировать геномы или вирионы потомков, подобные тем, которые выделены из природных источников. Например, молекулярный клон pNL4-3 HIV-1, описанный в последующих примерах, представляет собой штамм дикого типа, который можно получить по AIDS Research and Reference Reagent Program Catalog через National Institutes of Health (смотри также Adachi et al., J. Virol., 59, 284-291 (1986)). pPSXB представляет собой молекулярный клон HIV-2, который был любезно предоставлен Dr Suresh Arya из National Institutes of Health, Bethesda, Maryland и описан в Arya et al. Human immunodeficiency virus type 2 lentivirus vectors for gene transfer: expression and potential for helper virus-free packaging. Hum Gene Ther. 1998 Jun 10;9(9):1371-80.
В целом, способ настоящего изобретения предпочтительно применяют для лечения вирусных заболеваний, которые являются результатом вирусной инфекции. Желательно, чтобы вирус (так же как вектор, как обсуждается ниже) представлял собой РНК-вирус, но также он может представлять собой ДНК-вирус. РНК-вирусы представляют собой разнообразную группу, которая инфицирует прокариот (например, бактериофаги), так же как многих эукариот, включая млекопитающих и, в частности, человека. Большинство РНК-вирусов обладает одноцепочечной РНК в качестве своего генетического материала, хотя, по меньшей мере, одно семейство имеет двухцепочечную РНК в качестве своего генетического материала. РНК-вирусы подразделены на три большие группы: вирусы с плюс-цепью (т.е. те, геном которых, переносимый вирусом, транслируется в белок и чья депротеинизированная нуклеиновая кислота существенна для инициации инфекции), вирусы с минус-цепью (т.е. те, геном которых, переносимый вирусом, комплементарен последовательности мессенджера и может наступать транскрипция, которая должна происходить с помощью ферментов, ассоциированных с вирионом, до трансляции) и двухцепочечные РНК-вирусы. Способ настоящего изобретения предпочтительно применяют для борьбы с вирусами с плюс-цепью, вирусами с минус-цепью и двухцепочечными РНК-вирусами.
Применяемый здесь РНК-вирус охватывает Sindbis-подобные вирусы (например, Togaviridae, Bromovirus, Cucumovirus, Tobamovirus, Ilarvirus, Tobravirus и Potexvirus), Picornavirus-подобные вирусы (например, Picornaviridae, Caliciviridae, Comovirus, Nepovirus и Potyvirus), вирусы с минус-цепью (например, Paramyxoviridae, Rhabdoviridae, Orthomyxoviridae, Bunyaviridae и Arenaviridae), двухцепочечные вирусы (например, Reoviridae и Birnaviridae), Flavivirus-подобные вирусы (например, Flaviviridae и Pestivirus), Retrovirus-подобные вирусы (например, Retroviridae), Coronaviridae и другие группы вирусов, включая, но не ограничиваясь этим, Nodaviridae.
Предпочтительный РНК-вирус в соответствии с изобретением представляет собой вирус семейства Flaviviridae, предпочтительно вирус рода Filovirus и особенно вирус Марбурга или Эбола. Предпочтительно вирус семейства Flaviviridae представляет собой вирус рода Flavivirus, такой как вирус желтой лихорадки, вирус Денге, вирус Западного Нила, вирус энцефалита Сант-Луиса, вирус японского энцефалита, вирус энцефалита долины Мюррей, вирус Роцио, вирус клещевого энцефалита и тому подобное.
Также предпочтительным является вирус семейства Picornaviridae, предпочтительно вирус гепатита А (HAV), вирус гепатита B (HBV) или вирус гепатита не-А или не-В.
Другой предпочтительный РНК-вирус представляет собой вирус семейства Retroviridae (т.е. ретровирус), особенно вирус рода или подсемейства Oncovirinae, Spumavirinae, Spumavirus, Lentivirinae и Lentivirus. РНК-вирус подсемейства Oncovirinae является желательно Т-лимфотропным вирусом человека типа 1 или 2 (т.е. HTLV-1 или HTLV-2) или вирусом лейкоза коров (BLV), вирусом лейкосаркомы птиц (например, вирусом саркомы Рауса (RSV), вирусом миелобластоза птиц (AMV), вирусом эритробластоза птиц (AEV) и вирусом, ассоциированным с саркомой Рауса (RAV; от RAV-0 до RAV-50), вирусом типа С млекопитающих (например, вирусом мышиного лейкоза Молони (MuLV), вирусом мышиной саркомы Харвея (HaMSV), вирусом мышиного лейкоза Абельсона (A-MuLV), AKR-MuLV, вирусом лейкоза кошачьих (FeLV), вирусом саркомы обезьян, вирусом ретикулоэндотелиоза (REV), вирусом некроза селезенки (SNV)), вирусом типа В (например, вирусом опухоли молочной железы мышей (MMTV)) и вирусом типа D (например, вирусом обезьян Мейсона-Пфазера (MPMV) и вирусами "SAIDS"). РНК-вирус подсемейства Lentivirus представляет собой желательно вирус типа 1 или 2 иммунодефицита человека (т.е. HIV-1 или HIV-2, где HIV-1 ранее был назван как вирус 3, ассоциированный с лимфоаденопатией (HTLV-III), и вирус, связанный с синдромом приобретенного иммунодефицита (СПИДом) (ARV)), или другой вирус, относящийся к HIV-1 или HIV-2, который идентифицирован и связан со СПИДом и заболеванием, подобным СПИДу. Акроним "HIV (ВИЧ)" или термины "вирус СПИДа" или "вирус иммунодефицита человека" применяются здесь для обозначения таких вирусов HIV и вирусов, относящихся к или ассоциированных с HIV, в целом. Более того, РНК-вирус подсемейства Lentivirus предпочтительно представляет собой вирус Visna/maedi (например, такой, который инфицирует овец), вирус иммунодефицита кошачьих (FIV), лентивирус коров, вирус иммунодефицита обезьян (SIV), вирус инфекционной анемии лошадей (EIAV) и вирус артрита-энцефалита козлов (CAEV).
Вирус в соответствии с изобретением также желательно является ДНК-вирусом. Предпочтительно ДНК-вирус представляет собой вирус Эпштейна-Барр, аденовирус, вирус простого герпеса, вирус папилломы, вирус коровьей оспы и тому подобное.
Многие из данных вирусов классифицированы как патогены "уровня 4 биологической безопасности" (т.е. "группы риска 4" по классификации Всемирной организации здравоохранения (ВОЗ)), для которых при всех лабораторных работах требуется максимум защитных приспособлений. Специалист в данной области техники, однако, близко знаком с этим и способен придерживаться мер предосторожности и безопасности, необходимых для данных вирусов.
"Клеткой-хозяином" может быть любая клетка и предпочтительно клетка эукариот. Желательно, чтобы клеткой-хозяином был лимфоцит (такой как Т-лимфоцит), или макрофаг (такой как моноцит/макрофаг), или предшественник любой из данных клеток, такой как гемопоэтическая стволовая клетка. Предпочтительно, чтобы клетки имели на клеточной поверхности гликопротеин CD4+, т.е. являлись CD4+. Желательно, однако, чтобы Т-лимфоцит CD4+, который инфицирован вирусом СПИДа, не был до этого активирован (т.е. предпочтительно, чтобы экспрессия nef еще не наступала, и даже более предпочтительно, чтобы экспрессия гена CD4 не регулировалась негативно, как будет далее обсуждаться ниже). Более того, клетка-хозяин предпочтительно представляет собой клетку, в которой отсутствует маркер CD4 и которая еще способна инфицироваться вирусом в соответствии с настоящим изобретением. Такая клетка включает, но не ограничивается этим, астроцит, фибробласт кожи, эпителиальную клетку кишечника, эндотелиальную клетку, эпителиальную клетку, дендритную клетку, клетки Лангерганса, моноцит, гематопоэтическую стволовую клетку, эмбриональную стволовую клетку, клетку, которая дает сперматогенный ряд или ооцит, стромальную клетку, мукозную клетку и тому подобное. Предпочтительно, чтобы клетка-хозяин представляла собой эукариотные, многоклеточные виды (например, в отличие от клетки одноклеточных дрожжей) и даже более предпочтительно, чтобы она представляла собой клетку млекопитающих, например, человека.
Клетка может быть представлена как один организм, или может быть частью более крупного скопления клеток. Такое "более крупное скопление клеток" может включать, например, клеточную культуру (либо смешанную, либо чистую), ткань (например, эндотелиальную, эпителиальную, мукозную или другую ткань, включая ткани, содержащие указанные выше клетки с отсутствием CD 4), орган (например, сердце, легкое, печень, мышцу, желчный пузырь, мочевой пузырь, гонады, глаз и другие органы), систему органов (например, систему циркуляции, респираторную систему, систему желудочно-кишечного тракта, мочевую систему, нервную систему, систему наружных покровов или другую систему органов) или организм (например, птицы, млекопитающего или тому подобное). Предпочтительно, чтобы органы/ткани/клетки, являющиеся мишенями, представляли циркуляторную систему (например, включая, но не ограничиваясь этим, сердце, кровеносные сосуды и кровь, включая лейкоциты и эритроциты), респираторную систему (например, нос, глотку, гортань, трахею, бронхи, бронхиолы, легкие и тому подобное), систему желудочно-кишечного тракта (например, включая рот, глотку, пищевод, желудок, кишечник, слюнные железы, поджелудочную железу, печень, желчный пузырь и другие), мочевую систему (например, такую как почки, мочеточники, мочевой пузырь, уретру и тому подобное), нервную систему (например, включая, но не ограничиваясь этим, мозг и спинной мозг, специфические сенсорные органы, такие как глаз) и систему кожных покровов (например, кожу, эпидермис и клетки подкожной или кожной ткани). Даже более предпочтительно, чтобы клетки, являющиеся мишенями, были выбраны из группы, состоящей из сердца, кровеносного сосуда, легкого, печени, желчного пузыря, мочевого пузыря и клеток глаза. Клетки-мишени необязательно должны быть нормальными клетками и могут представлять собой клетки, пораженные болезнью. Такие клетки, пораженные болезнью, могут представлять собой, но не ограничиваться этим, опухолевые клетки, инфицированные клетки, генетически измененные клетки или клетки, находящиеся вблизи или в контакте с поврежденной тканью, такие как эндотелиальные клетки сосудов опухоли.
Вектор
"Вектор" представляет собой молекулу нуклеиновой кислоты (обычно ДНК или РНК), которая служит для переноса переносимой последовательности нуклеиновой кислоты (т.е. ДНК или РНК) в клетку-хозяина. Три обычных типа векторов включают плазмиды, фаги и вирусы. Предпочтительно, чтобы вектор представлял собой вирус, который включает капсидные формы векторных нуклеиновых кислот, и вирусные частицы, в которые упакован вектор нуклеиновых кислот.
Желательно, чтобы вектор не представлял собой штамм вируса дикого типа, в виду того, что он включает индуцированные человеком мутации или модификации. Таким образом, вектор обычно получают от штамма вируса дикого типа путем генетических манипуляций (т.е. путем делеции) для охвата вируса с зависящей от условий репликацией, как будет далее здесь описано. Оптимально, чтобы вирусный вектор включал штамм вируса, который является тем же типом, что и вирус дикого типа, вызывающий инфекцию, подвергаемую лечению, который предпочтительно представляет собой один из указанных выше вирусов дикого типа. Соответственно, предпочтительно, чтобы вектор происходил от РНК-вируса, даже более предпочтительно, чтобы вектор происходил от ретровируса, и оптимально, чтобы вектор происходил от вируса иммунодефицита человека. Такой вектор, происходящий от вируса иммунодефицита человека, обозначается здесь обычно как вектор "crHIV".
Вектор также предпочтительно является "химерным вектором", например сочетанием вирусного вектора с другими последовательностями, таким как, например, сочетание последовательностей HIV с одним или более другими вирусами (которые желательно происходят от штамма вирусов дикого типа для вхождения в состав вектора с зависимой от условий репликацией). В частности, последовательности HIV желательно могут быть соединены с последовательностями модифицированного (т.е. не дикого типа) штамма аденовируса, вируса, ассоциированного с аденовирусом, вируса из Alphaviridae, вируса из Flaviviridae, вируса из Hepadnaviridae, вируса из Papovaviridae, вируса из Parvoviridae, вируса из Herpesviridae, вируса из Poxviridae, вируса из Paramyxoviridae, вируса из Rhabdoviridae, вируса из Retroviridae, включая онко-ретровирусы, спума-ретровирусы и ленти-ретровирусы. Включаются также вирусы или вирусоподобные геномы, происходящие из или связанные с данными семействами вирусов.
Предпочтительным химерным вектором является такой, в котором векторные последовательности (т.е. либо последовательности, кодирующие белки, фрагменты, либо некодирующие последовательности), происходящие не от лентивируса, вставлены в лентивирусный вектор. Предпочтительно последовательности не-HIV вставляют в вектор, происходящий от HIV.
Как здесь предусмотрено, вектор может включать либо ДНК, либо РНК. Например, либо ДНК, либо РНК-вектор может применяться для получения вируса. Сходно копия кДНК может быть создана из геномной РНК вируса. В противоположном варианте часть кДНК (или вирусной геномной ДНК) может быть транскрибирована in vitro для получения РНК. Данные способы хорошо известны специалистам в данной области техники и описаны также в последующих примерах.
"Вирус с зависимой от условий репликацией" представляет собой вирус с дефектом репликации, который обладает данным дефектом только при определенных условиях. В частности, вирус может завершать свой цикл репликации в пермиссирующей клетке-хозяине и не может завершать свой цикл репликации в запрещающей клетке-хозяине. "Клетка-хозяин" представляет собой клетку, способную инфицироваться или действительно инфицированную штаммом вируса дикого типа или псевдотипированным вектором. Такое инфицирование вирусом дикого типа может происходить либо перед, либо после инфицирования вирусом с зависимой от условий репликацией в соответствии с изобретением. В противоположном варианте "клеткой-хозяином" является такая клетка, которая кодирует генные продукты вируса дикого типа или хелпера, необходимые для репликации вируса. Таким образом, вектор с зависимой от условий репликацией в соответствии с изобретением представляет собой вирус (который предпочтительно является вирусом того же типа, что и вызывающий инфекцию, подвергаемую лечению), который реплицируется только при комплементации штаммом вируса дикого типа (или хелпером) или когда вирус дикого типа инфицирует клетки, содержащие геномы вектора с зависимой от условий репликацией.
В предпочтительном осуществлении вектор включает РНК-вирус (например, вирус HIV с зависимой от условий репликацией), который вводится в форме ДНК. В данном предпочтительном осуществлении предлагается стратегия вектора HIV-1 с зависимой от условий репликацией, которая дает непатогенные векторные геномы crHIV-1 с селективным преимуществом над патогенными геномами HIV дикого типа. Конкретно, в клетках, содержащих геномы как HIV дикого типа, так и crHIV, РНК crHIV имеют селективное преимущество в отношении упаковки в вирионы, потому что они содержат, например, рибозимы, которые расщепляют РНК дикого типа, но не РНК crHIV. Такие непатогенные crHIVs способны переходить в неинфицированные клетки, которые восприимчивы к HIV-инфекции (например, клетки CD4+), в присутствии хелперного вируса дикого типа. Таким способом селективно упаковывающийся и распространяющийся crHIV вмешивается в репликацию HIV дикого типа.
Дополнительное предпочтительное осуществление представляет собой непатогенный вектор crHIV, который является непатогенным, потому что он не содержит никакого сочетания последовательностей вирусных вспомогательных белков (таких как, но не ограничиваясь этим, Vif, Vpu, Vpr или Nef или их сочетания, или фрагменты), которое может сделать вектор патогенным. В противоположном варианте последовательности могут присутствовать, но являться транскрипционно молчащими или нетранслируемыми. Необязательно вектор не содержит какого-либо сочетания последовательностей регуляторных белков (таких как, но не ограничиваясь этим, Tat или Rev или их фрагменты), которые должны сделать вектор патогенным. В противоположном варианте данные последовательности могут присутствовать, но являться транскрипционно молчащими или нетранслируемыми.
Векторы, однако, предпочтительно содержат какое-либо сочетание последовательностей структурных белков (таких как, gag или его фрагмент), последовательностей ферментативных белков (таких как, pol или его фрагмент) и/или последовательностей белков оболочки (таких как, env или его фрагмент) в форме, способной либо активно транслироваться, либо в молчащей форме. Следовательно, вектор с зависимой от условий репликацией может быть способным к репликации, но не реплицироваться до уровней, которые являются патогенными для данного хозяина. Вместо этого вектор требует комплементации компонентом хелпера (таким как хелперный вектор), содержащим необходимые последовательности, происходящие от вируса дикого типа, для того чтобы реплицироваться до уровней, достаточных для индукции требуемого терапевтического, профилактического или биологического эффекта. Определение точного сочетания последовательностей белков или нуклеотидов, присутствующих в векторе и в хелпере, для обеспечения оптимального биологического эффекта включает только прямое применение простых процессов скрининга, включающих добавление в векторные или хелперные конструкты или изъятие из них различного сочетания последовательностей нуклеотидов, причем данные процессы являются рутинными для специалистов в данной области техники.
Более того, указанные выше последовательности нуклеотидов могут быть модифицированы или подвергнуты мутагенезу для того, чтобы модифицировать биологический эффект или, например, снизить возможность рекомбинации с хелперным конструктом. Хорошо известны в данной области техники многочисленные протоколы для модификации или мутагенеза векторных или хелперных конструктов для получения более оптимизированных конструктов (например, Current Protocols in Molecular Biology, Harcourt Brace and Jovanovich, 2000; Molecular Cloning, Sambrook et al, Cold Spring Harbor Press, 1989; и Soong et al Nature Genetics 25: 436-439, 2000, все включены здесь полностью).
Подход, возможный с указанными выше векторами, отличается от применения живых аттенуированных (LA) вакцин, в которых используются компетентные в отношении репликации вирусы, которые не имеют вспомогательных белков (смотри Daniel et al., Science, 258, 1938-1941 (1992); и Desrosiers, AIDS Res. & Human Retrovir., 10, 331-332 (1994)), потому что с LA-вакцинами не делалось попыток восполнить недостатки, которые, например, препятствуют эффективной продукции эффективного иммунного ответа и при все еще сохраняющейся безопасности. Например, известно, что вакцины LA SIV с множественными делециями не могут вызывать эффективного иммунного ответа, в то время как вакцины LA SIV с одной делецией (Nef негативные) являются патогенными у неполовозрелых макак (Baba et al, 1995).
Альтернативный подход к вакцине LA SIV, предлагаемый в настоящем изобретении, как описано выше, должен заключаться в использовании, по меньшей мере, двух векторов, где, по меньшей мере, один является вектором HIV с множественными делециями, а второй (хелперный) вектор экспрессирует все вспомогательные белки, за исключением Nef. Следовательно, многократно аттенуированный вектор HIV может реплицироваться зависимым от условий образом без провоцирования заболевания при комплементации Vif, Vpr и Vpu из хелперного вектора. Первый вектор может быть сконструирован как содержащий генетические противовирусные части, в результате чего может вмешиваться в репликацию и распространение HIV дикого типа. Альтернативно, такой первый вектор может быть использован для индукции эффективного иммунного ответа против вируса дикого типа. Специалистам в данной области техники должно быть ясно, что простой процесс скрининга должен позволить оценить какая(ие) фактическая(ие) комбинация(и) последовательностей, удаленных из первого вектора, но присутствующих в хелперном векторе, должны позволить первому вектору оптимально обеспечить терапевтический или профилактический ответ и все еще сохранять безопасность, будучи непатогенным.
При дополнительном предпочтительном осуществлении векторно-хелперной системы с зависимой от условий репликацией для экспрессии gag, pol, env, tat и rev будет применяться вектор HIV-1, а хелперный вектор, происходящий от HIV-2, будет применяться, например, для экспрессии Vif, Vpu, Vpr и необязательно Nef генов. Изобретение не ограничивается данным примером, потому что равно возможно любое сочетание указанных выше генов, помещенных в любое совместимое сочетание векторов, включая обратные HIV-1 и HIV-2, или химерные форматы HIV. Экспрессия Tat остовом HIV-1 должна трансактивировать как HIV-1, так и HIV-2 LTRs для комплементации одного другим для получения векторных частиц, которые содержат либо векторные геномы HIV-1, либо векторные геномы HIV-2. Однако две геномных РНК не должны эффективно димеризоваться, предотвращая таким образом совместную локализацию как векторного, так и хелперного геномов, которые иначе оба будут упакованы в одну вирионную частицу. Если они совместно упакованы в одну и ту же вирионную частицу, может наступить рекомбинация между векторным и хелперным геномом с получением репликационно компетентного вектора (RCV) в процессе обратной транскрипции после того, как частица инфицирует последующую клетку-мишень.
Для дополнительного управления возможной продукцией совместно упакованных конструктов векторы могут дополнительно содержать одну или более последовательностей нуклеиновой кислоты, которые снижают риск любой рекомбинации. Например, первый вектор может содержать рибозимную (или антисмысловую/рибозимную) последовательность, специфичную в отношении расщепления хелперного вектора. Совместная локализация таких векторов в том же самом клеточном, субклеточном или внеклеточном месте должна приводить к расщеплению хелперного вектора для увеличения безопасности путем уничтожения димеризации или совместной локализации векторов, что ведет к возникновению неактивного генома после рекомбинации или по-другому предотвращает возникновение получения RCV. Данный подход может быть изменен путем включения рибозима в хелперный вектор вместо первого вектора или дополнительно усовершенствован путем включения рибозимов в оба вектора для обеспечения дополнительной безопасности.
В противоположном варианте антисмысловые молекулы могут быть применены для замещения указанных выше рибозимов, так что образование гибридной двухцепочечной РНК будет быстро разрушаться клеточными эндонуклеазами. Дополнительное предпочтительное осуществление заключается в том, что антисмысловые последовательности должны превышать в длину РНК рибозима, используемую как мишень, на приблизительно 16 оснований.
В модифицированном подходе хелперный вирус указанного выше примера может происходить от гетерологичного вируса (такого как любой из вирусов и семейств вирусов, описанных выше, но предпочтительно происходить от аденовируса, вируса, ассоциированного с аденовирусом, мышиного онкоретровируса или лентивируса, не относящегося к HIV), в результате чего белки могут экспрессироваться конститутивно. В противоположном варианте гетерологичный хелперный вектор может быть сконструирован для получения HIV-LTR для индуцируемой или ауторегулируемой экспрессии Tat с тем, чтобы трансактивировать экспрессию вектора HIV-1. Преимущество, обеспечиваемое гетерологичным вирусным хелперным вектором, состоит в ограниченной ассоциации между геномами первого и второго векторов, таким образом дополнительно снижая вероятность рекомбинации с возможным образованием RCV.
В другом осуществлении вектор содержит антисмысловую последовательность, которая присутствует в хелперном геноме или включена в хелперный геном. Неограничивающий пример можно рассмотреть с вектором pN1cptASgag, который аналогичен вектору pN1cptASenv за исключением того, что антисмысловая последовательность направлена на последовательность gag, присутствующую в хелперном векторе в дополнение к или вместо последовательности env, присутствующей в HIV дикого типа. Анти-gag антисмысловая последовательность расположена выше акцепторного сайта сплайсинга, который находится ниже последовательности RRE, присутствующей в векторе. Следовательно, последовательность gag будет упакована только в геномных, а не в подгеномных или сплайсингованных видах векторных РНК. Таким образом, хелперные геномы интронов, содержащих хелперы, такие как относящиеся к системе VIRPAC, должны быть предпочтительно направлены к сплайсосоме клетки, в то время как геномные векторные РНК, которые содержат анти-gag антисмысловую последовательность, должны предпочтительно обходить систему сплайсинга.
Различающиеся пути в клетке векторного и хелперного геномов должны приводить к минимальному влиянию на титр вектора, даже если векторный и хелперный геномы будут по «счастливой случайности» совместно локализованы, то гибридизация пар оснований векторного и хелперного геномов будет приводить к инактивации или разрушению таких частиц, содержащих векторный и хелперный геномы, и предотвращать рекомбинацию вектор-хелпер с образованием RCV. В противоположном осуществлении хелпер может быть сконструирован с наличием анти-U5 векторных антисмысловых последовательностей, направленных на U5 последовательности, находящиеся в векторе с зависимой от условий репликацией. Не выходя за пределы сущности данного аспекта изобретения, анти-U5 антисмысловые последовательности могут быть вставлены дистальнее хелперных кодирующих последовательностей, но перед сайтом терминации транскрипции. Однако если векторная и хелперная РНК будут по «счастливой случайности» совместно локализованы и будут подвергаться совместной упаковке и возможной рекомбинации, то антисмысловые последовательности будут разрушать или инактивировать такие совместно упакованные вирусные частицы и предотвращать рекомбинацию.
В еще одном осуществлении хелпер может содержать последовательности-мишени для первой нуклеотидной последовательности, присутствующей в векторе. Нелимитирующим примером является вставка смыслового фрагмента env в хелпер из двух пар плазмид, содержащих вектор pN1cptSenv. Следовательно, если векторная и хелперная РНК будут совместно локализованы, смысловая последовательность env в хелпере будет образовывать пары оснований с антисмысловой последовательностью env в векторе с предотвращением рекомбинации. Представленные выше примеры не предназначены для ограничения изобретения только одним или двумя типами генетической противовирусной последовательности. Специалисту в данной области техники должно быть ясно, что множество генетических противовирусных последовательностей, также как родственных им последовательностей-мишеней, может быть вставлено в векторный и/или хелперный конструкты. В качестве дополнительного неограничивающего примера вектор pN1cptASgagASenv и хелпер, который содержит смысловую последовательность gag и последовательность фрагмента env, могут быть применены в сочетании друг с другом для увеличения безопасности путем снижения вероятности совместной упаковки и рекомбинации с образованием RCV.
В другом предпочтительном осуществлении при экспрессии векторного или хелперного компонентов компоненты экспрессируются временно. Одним способом создания вектора или хелпера, экспрессирующего свой геном или компоненты генома временно, является конструирование векторного или хелперного вектора с применением, например, негативного по интегразе гена Pol. Такие лентивирусные интегразные мутанты известны в данной области техники и, как показано, не являются инфекционными (Hirsch et al 1989 Nature 341: 573-574). Следовательно, векторный и/или хелперный геномы, которые получают из негативных по интегразе клеток-продуцентов, не будут интегрироваться, но могут экспрессироваться транзиторным образом с тем, чтобы регулировать уровень репликации вектора или хелпера. Еще одним способом транзиторной экспрессии либо вектора, либо хелпера является разрыв сайтов AAT в векторе LTRs. Данные сайты отвечают за активную интеграцию обратно транскрибируемой геномной векторной ДНК в хромосому клетки-хозяина.
Дополнительным средством для достижения указанного выше является применение гибридного белка для упаковки функциональной молекулы интегразы в вирусные частицы. В данном осуществлении ни вектор с зависящей от условий репликацией, такой как лентивектор, ни хелперный вектор, не должны кодировать интегразу, но гибридный белок функциональной интегразы мог бы быть доступным за счет экспрессии другим плазмидным конструктом, таким как хелперный вектор, или копией, интегрированной в применяемую упаковывающую клетку, в процессе продукции или упаковки хелперного конструкта, обеспечивая таким образом функциональную активность интегразы при инфекции. В противоположном варианте белок-интеграза предлагается отдельно путем экспрессии хелперным конструктом, который его кодирует. В данном осуществлении изобретения вектор с зависящей от условий репликацией или лентивектор не должен кодировать интегразу. Гибридный белок может представлять собой, но не ограничиваться этим, гибридный белок vpr-интегразы, содержащий сайт расщепления протеазами в месте соединения между аминокислотными последовательностями vpr и интегразы.
Указанное выше описание двойной системы вектор-хелпер не должно ни в коей мере ограничивать изобретение двумя векторными или хелперными конструктами. Может быть использовано любое сочетание двух, трех или более векторов и/или хелперов для обеспечения компонентами, необходимыми для получения вектора. Разделение необходимых геномных элементов на большее количество векторных или хелперных компонентов будет создавать эффект повышенной безопасности, так как для множественных векторных и хелперных геномов будет труднее создавать RCV путем рекомбинации. Безопасность может быть дополнительно увеличена путем конструирования вектора(ов) и хелпера(ов), содержащих мало или не содержащих участков гомологии между ними, что является дополнительным предпочтительным осуществлением. Разделение необходимых геномных элементов на множественные компоненты может также дополнительно ограничить репликацию вектора с зависящей от условий репликацией, так как менее вероятно, чтобы более двух геномов по сравнению с только двумя геномами будет одновременно присутствовать в клетке-хозяине. Оптимальное количество требуемых вектора(ов) и хелпера(ов) может быть легко определено с помощью простого скрининга различных сочетаний и может варьироваться в зависимости от конкретной применяемой вирусной векторной системы.
Одно простое и неограничивающее средство ограничения или удаления участков гомологии между вектором(ами) и хелпером(ами) заключается в простой вырожденности нуклеотидной последовательности при поддержании последовательности кодируемых аминокислот. Способы создания вырожденных последовательностей хорошо известны в данной области техники и являются рутинными. Один предпочтительный способ заключается в приближении последовательностей к человеческим, если их терапевтически применяют у человека. Применение кодонов у приматов сведено в таблицу и описывается в Wada et al. (Nucleic Acids Research vol. 18 Supplement: 2367-2411, 1990), которая включена здесь в полном изложении.
Вырожденность может также быть использована для защиты хелперного конструкта от действия агента, предназначенного для направления вектора в сторону упаковки с хелпером. Это может быть достигнуто просто с помощью вырожденности родственной последовательности-мишени, если какая-то найдена в хелперном конструкте. Дополнительно вырожденность как векторного, так и хелперного конструктов может быть предназначена для снижения рекомбинации с другими вирусными последовательностями, включая те из других последовательностей дикого типа, которые могут быть случайно найдены в векторной или хелперной последовательности в клетке. Например, применение векторной и хелперной пары в упаковывающей системе, основанной на HIV-1 и HIV-2, можно модифицировать так, что векторная и хелперная пара становится вырожденной также для эндогенного ретроэлемента. Таким образом, вырожденная нуклеотидная последовательность либо вектора, либо хелпера должна снижать совместную локализацию предполагаемых рекомбинантов и таким образом снижать риск создания репликационно компетентного вируса между векторным и хелперным геномами или между векторным или хелперным геномами и конкурирующим геномом, таким как эндогенный ретроэлемент.
В частности, геномы crHIV вводятся в инфицированные вирусом клетки или в неинфицированные клетки. Инфицированные клетки снабжают геном crHIV белками, требуемыми для инкапсидации и продукции потомства вирионов. Геномы crHIV вводят в неинфицированные клетки предпочтительно либо прямо путем трансдукции (например, это может быть сделано, например, с помощью опосредованной липосомами трансдукции ДНК crHIV или путем применения химерного вирусного вектора), либо путем инфицирования частицами crHIV, что является результатом трансфекции клеток, инфицированных HIV дикого типа. Неинфицированные клетки, содержащие усовершенствованный вектор crHIV изобретения, не продуцируют такое количество частиц crHIV, которое является патогенным для хозяина. Некоторые осуществления клеток с вектором crHIV, не суперинфицированных HIV дикого типа, будут продуцировать некоторое количество частиц crHIV, но на уровне, который не является патогенным для хозяина. Клетки, содержащие любой вектор изобретения, могут оставаться восприимчивыми к суперинфицированию вирусом дикого типа, который будет поставлять белки, требуемые для дополнительной продукции частиц crHIV. В этом смысле вектор с зависящей от условий репликацией в соответствии с изобретением в присутствии дополнительного суперинфицирования диким типом функционирует также как тип "вирусного вектора для доставки", например, могут происходить множественные циклы инфицирования crHIV (т.е. в присутствии сопутствующей инфекции HIV дикого типа). Такой вектор является источником вируса в течение более чем одного цикла репликации вируса и таким образом инфицирует другие клетки, или в течение множественных циклов репликации количество вирусов доводится до уровня, который должен вызывать биологический ответ, такой как иммунный ответ. Данное усовершенствование контрастирует с другими векторами, такими как те, которые применяют со стандартными упаковывающими клеточными линиями и которые дают только один цикл репликации или множественные циклы репликации, которые либо не достаточны для индукции соответствующего биологического ответа, либо являются патогенными для хозяина.
Если это желательно (например, для облегчения применения вектора in vitro), генные продукты вируса дикого типа могут параллельно поставляться клетке, инфицированной вектором с зависящей от условий репликацией. Генные продукты вируса дикого типа могут поставляться не только с помощью совместного инфицирования штаммом вируса дикого типа (или кДНК, или провирусом, или РНК-вирусом), но также путем предоставления их клетке в форме их генов, субклонированных в экспрессионном векторе, например в хелперном экспрессионном векторе ("хелпер" или "хелперный вектор"), который способен передавать клетке-хозяину транскрипцию или трансляцию последовательностей (регуляторных или структурных), или в альтернативном варианте генные продукты могут поставляться экзогенно, т.е. путем добавления белковых продуктов к клетке.
Например, вектор crHIV может быть сконструирован так, чтобы он содержал все белки HIV дикого типа за исключением, например, последовательности, кодирующей Tat. Вместо применения хелперного экспрессионного конструкта, который экспрессирует Tat, в качестве хелпера может быть применен сам белок Tat. Преимущество использования белка вместо хелперного конструкта заключается в том, что не существует теоретической возможности создания вируса дикого типа, так как никаких последовательностей нуклеиновых кислот не присутствует для рекомбинации. Таким образом, crHIV может размножаться, используя белок Tat, а не хелперный конструкт нуклеиновой кислоты per se. Tat 1 или Tat 2 (соответствующие первому или первому и второму экзонам Tat соответственно) могут быть использованы в качестве хелпера. Способы получения и очистки таких белков хорошо известны в данной области техники.
В одном предпочтительном осуществлении применяют химерный белок Tat. Tat вставляли в гибридный белок (например, Tat-VP16) и, как показано, он сохранял свою трансактивирующую активность на промоторе HIV-LTR. Для увеличения желаемого биологического эффекта могут быть созданы другие химерные белки Tat. Например, если желаемый биологический эффект состоит в увеличении иммунного ответа, то может быть сконструирован гибридный белок Tat-GM-CSF. Данный подход не ограничивается одним типом химерного белка и может быть желательным добавление, по меньшей мере, второго химерного (или нехимерного) белка Tat (например, Tat-IFN-alpha, Tat-TNF-alpha, Tat-IL-4, Tat-G-CSF, TNF-alpha, GM-CSF, IL-4, G-CSF, IFN-alpha в качестве примера, но охватывая только некоторые возможности). Специалист в данной области техники может, несомненно, с легкостью сконструировать и протестировать другие эквивалентные химерные белки и провести их скрининг на способность увеличивать иммунный ответ, предоставляя в то же время хелперную функцию для вектора. Другое предпочтительное осуществление заключается в конструировании гибридного белка Tat-Rev или гибридного белка Tat-Rev природного HIV, например Trev. Следовательно, представленное выше описание химерных белков Tat не ограничивается включением только цитокинов или клеточных белков, но в изобретении могут быть использованы также вирусные белки (продукция гомологичных или гетерологичных вирусных гибридных белков) в зависимости от желаемого биологического эффекта.
В отношении "хелперного вектора" его экспрессия может быть специфичной или неспецифичной для клетки, и он может быть введен в клетку-хозяин вместе с вирусным вектором с зависящей от условий репликацией, как здесь указано, давая возможность постоянной репликации вирусному вектору с зависящей от условий репликацией.
"Хелперные векторы" изобретения могут поставлять генные продукты, необходимые для репликации, с помощью либо единичного вектора, либо множественных векторов. Такие векторы предпочтительно применяют во временной трансфекционной системе. Альтернативно, генные продукты могут также быть интегрированы в геном клетки-хозяина или клеточной линии. В соответствии с изобретением единичный вектор или плазмида (которая может быть моно-, би- или мультицистронной в клетке-хозяине) являются предпочтительными из-за увеличенных векторных титров, возникающих благодаря увеличенной вероятности совместной трансфекции хелперного вектора и вектора с зависящей от условий репликацией в одной и той же клетке. Пониженная вероятность одновременной трансфекции более двух различных векторов в одной и той же клетке делает применение единичного хелперного вектора более желательным. Снижение количества векторов или плазмид, включенных в трансфекционную систему, является также целесообразным в плане цены, так как меньшее количество плазмид по сравнению с общепринятой трехплазмидной трансфекционной системой должно продуцироваться для упаковки ретровирусных конструктов.
Применяемый здесь термин "комплементация" обозначает негенное взаимодействие вирусных генных продуктов из различных источников в клетках. Конкретно, в случае смешанной инфекции комплементация включает увеличение выхода вирусов одного или обоих родительских геномов, в то время как генотипы родительских геномов остаются неизмененными. Комплементация может быть неаллельной (т.е. межгенной, при которой мутанты, дефектные в отношении различных функций, помогают друг другу при вирусной репликации путем снабжения функцией, которая является дефектной у другого вируса) или аллельной (т.е. внутригенной, при которой два родителя имеют дефекты в различных доменах мультимерного белка).
Желательно, чтобы типы клеток, которые могут быть трансфицированы (трансдуцированы) ДНК crHIV (т.е. с помощью липосом или применения гетерологичного вектора для получения химерного вектора, как описано выше), могли быть либо клетками, инфицированными HIV, либо неинфицированными клетками. Клетки, инфицированные HIV, могут быть активированными или неактивированными. Если они активированы, они будут немедленно транскрибировать РНК HIV дикого типа и РНК crHIV, приводя к селективной упаковке РНК crHIV в вирионы потомков. Если клетки, инфицированные HIV, неактивированы, ДНК crHIV будет находиться в них до тех пор, пока они не станут активированными (например, путем стимуляции митогенами, антигенами и тому подобное), опять приводя к селективной упаковке РНК crHIV в вирионы потомков. Как активированные, так и неактивированные неинфицированные клетки, которые трансфицированы ДНК crHIV, не будут продуцировать вирионы до тех пор, пока они не станут суперинфицированными HIV дикого типа и активированными путем стимуляции, опять приводящей к селективной упаковке РНК crHIV в вирионы потомков.
Одним вариантом клетки-хозяина, трансдуцированной вектором изобретения, является такой, при котором клетка содержит максимальное количество векторных копий, которые не являются существенно токсичными ни для клетки-хозяина, ни для организма-хозяина, содержащего клетку. Другим вариантом клетки-хозяина, трансдуцированной вектором, является такой, при котором клетка содержит, по меньшей мере, одну копию вектора, но предпочтительно содержит минимальное количество векторных копий, требуемых для индукции биологического эффекта. Количество копий в клетке может быть определено с помощью, например, TaqMan ПЦР, и приемлемое количество копий или диапазон количества копий, подходящие как для желаемого биологического эффекта, так и в плане отсутствия токсичности, может быть легко определено специалистом в данной области техники. Приемлемое количество копий будет варьироваться в зависимости от факторов, включая тип трансдуцируемой клетки, природу вектора (например, вирусное происхождение) и желаемый биологический эффект, но будет оставаться легко определимым путем обычного скрининга специалистом в данной области техники.
Суперинфицирование клеток, содержащих геномы crHIV (например, в качестве результата трансфекции или инфекции), наступает, потому что геномы crHIV не кодируют вирусные белки, которые блокируют суперинфицирование (такие, как env и nef). Полученные вирионы crHIV могут инфицировать неинфицированные клетки, так как вирусные частицы содержат молекулу обратной транскриптазы, которую несут все частицы HIV, так что они могут создавать ДНК-провирус из их геномной РНК. Данный процесс называют обратной транскрипцией. Как только вирионы crHIV инфицируют неинфицированные клетки, они могут подвергаться обратной транскрипции и продуцировать провирус из своей геномной РНК. Таким образом, данные клетки являются эквивалентными тем неинфицированным клеткам, которые прямо трансдуцируются ДНК crHIV. Они не могут продуцировать частицы crHIV до тех пор, пока данные клетки не станут суперинфицированными HIV дикого типа и не будут активированными, тогда еще раз произойдет селективная упаковка РНК crHIV в вирионы потомков. Возможно, что частицы crHIV смогли также инфицировать некоторые клетки, которые уже инфицированы HIV (смотри, например, Yunoki et al., Arch. Virol., 116, 143-158 (1991); Winslow et al., Virol., 196, 849-854 (1993); Chen et al., Nuc. Acids Res., 20, 4581-4589 (1992); и Kim et al., AIDS Res. & Hum. Retrovir., 9, 875-882 (1993)). Однако для того, чтобы это наступило, данные инфицированные HIV клетки не должны экспрессировать белки, которые негативно регулируют экспрессию CD4, потому что это будет препятствовать вирионам crHIV инфицировать данные клетки. Активированные клетки, инфицированные HIV, обычно негативно регулируют экспрессию CD4. Соответственно, клетки, инфицированные HIV, которые являются неактивированными, потенциально восприимчивы к суперинфицированию crHIV и, таким образом, могут быть другим источником продукции частиц crHIV. В отношении предпочтительного вектора crHIV изобретения вектор включает последовательности, требуемые для транскрипции РНК, связывания праймеров тРНК, димеризации и упаковки, и либо не имеет последовательностей, кодирующих белки, которые блокируют суперинфицирование HIV дикого типа (например, белки nef или env), или включают такие последовательности, но они либо не транскрибируются, либо не транслируются в функционирующий белок, так что их экспрессия считается "молчащей". Даже более предпочтительно, чтобы вектор не имел области или последовательностей, кодирующих область HIV дикого типа, начиная от последовательности, кодирующей gag, до и включая ген nef. Оптимально, однако, чтобы вектор действительно включал rev-чувствительный элемент (RRE), который клонирован в вектор в область делеции или какую-либо другую подходящую область. Такой предпочтительный вектор HIV, как говорят, "лишен области или последовательностей, кодирующих область", в виду того, что данный вектор может быть введен в своем РНК варианте или, альтернативно, в виде ДНК, как описано ранее.
В соответствии с изобретением для того, чтобы достичь успешной комплементации в системе, которая использует хелперный вектор в сочетании с вектором с зависящей от условий репликацией, любой вирусный компонент, необходимый для вирусной упаковки, но не экспрессируемый вектором с зависящей от условий репликацией, должен быть поставлен хелперным вектором. Таким образом, настолько долго, насколько два или более различных вектора вместе обладают полным дополнением необходимых вирусных компонентов, композиция индивидуальных векторов является предметом для многих перестановок. Каждая из данных перестановок охватывается настоящим изобретением. В качестве примера гены gag и pol, необходимые для успешной упаковки и репликации, могут оба быть включенными либо в хелперный вектор, либо в вектор с зависящей от условий репликацией, или два гена могут индивидуально присутствовать в одном из каждых векторов. Более того, один хелперный векторный конструкт может включать последовательности как gag/pol, так и env, включая гетерологичные последовательности env, под контролем одного ряда транскрипционных регуляторных элементов, включая промотор, или под контролем отдельных промоторных элементов. Например, LTR может быть использован для экспрессии генных продуктов в векторах изобретения. Применение различных промоторов, включая индуцибельные промоторы, дает возможность дифференциальной регуляции, например, последовательностей gag/pol и env. Таким образом, последовательность env, особенно гетерологичная последовательность env, такая как последовательность оболочки VSV-G, может экспрессироваться на более высоком уровне.
Векторное конструирование хорошо известно специалистам в данной области техники. Данные способы могут быть использованы для конструирования как вектора с зависящей от условий репликацией, так и хелперного вектора. Таким образом, применяемый здесь термин "вектор" может относиться к обоим типам векторов. Дополнительно термин вектор охватывает любой лентивирусный вектор, не обязательно являющийся вектором с зависящей от условий репликацией. Такие векторы могут также относиться к общим лентивирусным векторам. В предпочтительных осуществлениях изобретения, однако, вектор представляет собой лентивирусный вектор с зависящей от условий репликацией. Например, и как описано в примере 1, манифестированная ДНК РНК-вируса, такого как HIV, расщепляется при применении рестриктазных ферментов для удаления последовательностей, кодирующих HIV, начиная от области, кодирующей gag, до U3 области, следующей за геном nef, включительно. Клонирующую кассету, включающую полилинкер, содержащий множественные сайты рестрикции, вставляют в область делеции перед лигированием для обеспечения удобных сайтов рестрикции для клонирования в векторе. Фрагмент ДНК, содержащий RRE, субклонируют в один из данных сайтов. Полученный вектор продуцирует укороченный транскрипт gag и не продуцирует белок Gag дикого типа или любые другие белки HIV дикого типа. Более того, не является необходимым, чтобы вектор экспрессировал даже укороченный белок gag, в виду того что последовательность инициации трансляции gag может быть подвергнута мутации для предотвращения его трансляции.
С применением того же самого подхода последовательности crHIV могут быть соединены с другими последовательностями, такими как последовательности вируса или другого вектора, с получением химерного вектора, как описано выше. Например, последовательности crHIV могут быть соединены с таковыми вируса лихорадки Синдбиса, AAV, аденовируса или амфотропного ретровируса, упоминая некоторые из них. Дополнительные вирусные последовательности, которые могут быть применены, включают последовательности вируса герпеса, поксвируса и других описанных здесь вирусов, включая любой вирус, который может быть использован для обеспечения доставки последовательностей crHIV. Такой химерный вектор может быть введен в клетку либо с применением механизма соединения вирусов для входа в клетку (например, путем опосредуемого рецепторами эндоцитоза для аденовируса), либо с помощью других способов, например с помощью липосом.
Предпочтительно в соответствии с изобретением вектор (т.е. вирус с зависящей от условий репликацией, который предпочтительно является вектором crHIV) включает, по меньшей мере, одну последовательность нуклеиновой кислоты, наличие (т.е. присутствие, транскрипция или трансляция) которой придает селективное преимущество. Существует два типа таких последовательностей нуклеиновой кислоты, предназначенных для включения в вектор: (1) последовательность нуклеиновой кислоты, наличие которой дает оптимальное селективное преимущество для вирусной репликации и распространения вектору, включающему такую последовательность, над штаммом вируса дикого типа (т.е. предпочтительно штаммом дикого типа, из которого происходит вектор и который не включает последовательность), и (2) последовательность нуклеиновой кислоты, наличие которой дает оптимальное селективное преимущество клеткам, инфицированным вектором, включающим последовательность, по сравнению с клетками, инфицированными или неинфицированными штаммом вируса дикого типа (т.е. предпочтительно штаммом дикого типа, из которого происходит вектор (и также, например, хелперный экспрессионный вектор, который стимулирует репликацию и/или функцию вектора в неинфицированной клетке-хозяине), который не включает последовательность, с помощью, например, стимуляции выживания клеток, содействия продукции и/или размножению векторных частиц, стимуляции получения crHIV-векторных вирионов из клеток, продуцирующих вектор crHIV, индукции апоптоза, облегчения продукции белков или стимуляции иммунологической функции или ее направленности, так чтобы достичь желаемого профилактического, терапевтического или биологического выхода. Каждая из данных последовательностей или множественность из каждой из данных последовательностей, т.е. последовательность, которая одна или в сочетании с другим(ими) фактором(ами) стимулирует размножение вектора и/или стимулирует конкретную функцию клетки-хозяина, так чтобы возникал благоприятный профилактический, терапевтический и/или биологический эффект, может быть включена в вектор либо в отсутствие, либо в присутствии другой последовательности, т.е. вектор может включать, "по меньшей мере, одну последовательность нуклеиновой кислоты" и, "по меньшей мере, одну дополнительную последовательность нуклеиновой кислоты".
Третий тип последовательности нуклеиновой кислоты представляет собой такой, который придает клетке-хозяину фенотип, который снижает, сводит к минимуму или предотвращает вирусную инфекцию. Такие последовательности нуклеиновой кислоты включают те, которые кодируют генный продукт, который, когда экспрессируется, защищает клетку-хозяин от вирусной инфекции. Например, индивидуумы, гомозиготные по аллелю Δ32 белка CCR5, были защищены от инфицирования HIV-1 из-за отсутствия главного со-рецептора для входа HIV-1 в клетки, экспрессирующие на своей поверхности усеченный белок CCR5. Исследования гетерозиготных индивидуумов показали, что аллель Δ32 имеет трансдоминантный эффект на экспрессию СCR5 дикого типа на клеточной поверхности. Это может быть обусловлено способностью аллеля Δ32 димеризоваться с белком дикого типа и предотвращать его корректную экспрессию на клеточной поверхности.
Аллель Δ32 имеет делецию 32 нуклеотидов, что вызывает мутацию со сдвигом рамки считывания, ведущую к синтезу укороченного белка CCR5. Так как укорочение вызывает появление 31 аминокислот на С-конце аллеля Δ32, которые не присутствуют в белке дикого типа и могут вызвать нежелательный иммунный ответ при присутствии на клеточной поверхности, настоящее изобретение включает экспрессию укороченного мутантного белка CCR5Δ32 (CCR5Δ32T), у которого отсутствует потенциально антигенная последовательность из 31 аминокислоты. Данный белок должен сохранять свою способность создавать трансдоминантный эффект для обеспечения клеткам защиты против инфицирования HIV. Последовательность, кодирующая CCR5Δ32T, легко создается путем введения стоп-кодона непосредственно после последней аминокислоты, общей как для CCR5 дикого типа, так и для CCR5Δ32 (после тирозина в положении 184).
В дополнительном осуществлении другой третий тип последовательности нуклеиновой кислоты представляет собой такой, который дополнительно усиливает приданный фенотип, который снижает, сводит к минимуму или предотвращает вирусную инфекцию. Такая последовательность нуклеиновой кислоты может кодировать генетический противовирусный агент, который, например, направлен на клеточный ген. Например, вектор может экспрессировать указанный выше мутантный белок CCR5Δ32T и дополнительно экспрессировать антисмысловые молекулы или молекулы рибозима с промотора U1 и содержащиеся в U1 snРНК (описано в патенте США 5814500), например, направленные на молекулу CCR5 дикого типа. Антисмысловая область, которая направлена против CCR5 дикого типа, должна быть вырожденной в ограниченной последовательности вектора CCR5Δ32T, таким образом, чтобы создать трансдоминантную мутантную РНК, устойчивую к действию антисмысловой последовательности против CCR5 или рибозимным молекулам, но все еще транслируемой в правильную аминокислотную последовательность для получения функционального белка CCR5Δ32T.
Представленный выше способ одновременной экспрессии рибозима или антисмысловой последовательности, направленных против генного продукта, и вырождения гена в сайтах-мишенях для удержания рибозима или связывания или расщепления антисмысловой последовательности не ограничен CCR5 или даже молекулой, которая может свести к минимуму репликацию вируса. Другой неограничивающий пример для терапевтического применения заключается в направленности против онкогенного белка, такого как гибридный белок Bcr-Abl, который является этиологическим агентом хронического миелогенного лейкоза. Векторный конструкт, содержащий ген Abl или Bcr или оба, может экспрессировать данные гены одновременно с экспрессией антисмысловой последовательности или рибозима, которые направлены на любой сайт по всей длине транскрипта Bcr-Abl. Следовательно, ненормальный гибридный транскрипт Bcr-Abl должен быть разрушен, в то время как природный Bcr или Abl или оба должны экспрессироваться для уменьшения ненормального дефекта. Таким образом, гены Bcr или Abl должны иметь селективное преимущество для амплификации над генами дикого типа или гибридным геном Bcr-Abl. Предпочтительный вектор должен экспрессировать конструкт Bcr и/или Abl с их природных промоторов, в то время как рибозимы или антисмысловая последовательность против Bcr-Abl должны экспрессироваться с промотора U1 и содержащихся в U1 последовательностей snРНК.
Представленный выше пример Bcr-Abl не ограничивает изобретение направленностью против онкогенов, и специалисту в данной области техники должно быть очевидно, что данный подход может быть использован для терапевтического или профилактического замещения любого экспрессируемого ненормального гена, нежелательного гена или даже интересующего(их) желательного(ых) гена(ов). Ген-мишень может быть гомологичным или гетерологичным по отношению к модифицированному гену, который устойчив к эффектам генной противовирусной молекулы. Другие заболевания, на которые может быть направлен данный способ, должны быть ясны для специалиста в данной области медицинской и клинической техники. Например, молекулярные мишени для лечения заболеваний крови с помощью раскрытых здесь способов изобретения описаны в "The molecular basis of blood diseases"(Stamatoyannopoulos, Majerus, Perlmutter & Varmus, 3rd Ed., Philadelphia: WB Sauders Co., авторские права 1987, 1994 и 2001, все из которых включены здесь в качестве ссылки).
Изобретение не должно быть ограничено клиническим применением. Может быть использован подход для определения функции интересующей генной последовательности. Например, интересующая генная последовательность, модифицированная в интересующем домене, может быть одновременно экспрессирована с антисмысловой последовательностью или рибозимом, которые ингибируют или нарушают экспрессию любой немодифицированной генной последовательности, будучи направленными против соответствующей немодифицированной области. Таким образом, интересующая модифицированная генная последовательность должна иметь селективное преимущество для амплифицированной экспрессии над немодифицированной генной последовательностью, позволяя определить функцию модифицированной генной последовательности или кодируемого ею генного продукта с помощью способов тестирования, известных в данной области техники.
"Нуклеиновая кислота" представляет собой описанное ранее. "Последовательность нуклеиновой кислоты", в частности, включает любой ген или кодирующую последовательность (т.е. ДНК или РНК) потенциально любого размера (т.е., конечно, ограниченного любыми упаковывающими ограничениями, навязанными вектором), наличие которой придает селективное преимущество, как здесь определяется далее. "Ген" представляет собой любую последовательность нуклеиновой кислоты, кодирующую белок, или возникающую молекулу мРНК (не взирая на то, будет ли последовательность транскрибироваться и/или транслироваться). В то время как ген включает кодирующие последовательности, а также некодирующие последовательности (например, регуляторные последовательности), "кодирующая последовательность" не включает никакую некодирующую ДНК.
1. Последовательность нуклеиновой кислоты, наличие которой придает вектору, включающему такую последовательность, в клетке-хозяине селективное преимущество над штаммом вируса дикого типа, или конкурирующая геномная последовательность, которая должна вмешиваться или влиять на амплификацию вектора.
Последовательность нуклеиновой кислоты, которая придает селективное преимущество вектору в клетке-хозяине над штаммом вируса дикого типа, предпочтительно является любой последовательностью, которая позволяет вирусным частицам размножаться из вектора, будучи селективно продуцируемыми или упакованными по сравнению с вирусными частицами, полученными размножением вируса дикого типа. Такие последовательности включают, но не ограничиваются этим, последовательность, которая ведет к увеличению количества векторных геномов, продуцируемых внутриклеточно, по сравнению с геномами дикого типа, и противовирусную последовательность нуклеиновой кислоты. Такие последовательности также включают, но не ограничиваются этим, последовательность, которая ведет к увеличению количества, свойства или состояния векторных геномов, продуцируемых внутриклеточно по сравнению с геномами дикого типа, которые не происходят от родственного вируса дикого типа.
Первая категория последовательностей нуклеиновой кислоты, которая придает селективное преимущество в клетке-хозяине вектору, содержащему последовательность, над штаммом вируса дикого типа, представляет собой последовательности, такие как промоторные. "Промотор" представляет собой последовательность, которая направляет связывание РНК-полимеразы и в результате этого стимулирует синтез РНК и которая может включать один или более энхансеров. "Энхансеры" представляют собой цис-действующие элементы, которые стимулируют или ингибируют транскрипцию примыкающих генов. Энхансер, который тормозит транскрипцию, также обозначают как "сайленсер". Сайленсер может также быть "изоляционным" элементом, как это найдено в сайтах гиперчувствительности к ДНКазе эритроидного изоляционного элемента цыплят. Энхансеры отличаются от сайтов ДНК, взаимодействующих с ДНК-связывающими белками, специфически узнающими последовательность ДНК, найденными только в промоторе (которые также называют "промоторными элементами"), тем, что энхансеры могут функционировать в любой ориентации и на расстоянии до нескольких тысяч пар нуклеотидов (т.п.н.) от положения даже ниже транскрибируемой области.
Соответственно и предпочтительно промотор (например, длинный концевой повтор (LTR)) вектора HIV с зависящей от условий репликацией модифицирован так, что вектор становится более чувствительным к определенным цитокинам, чем штамм HIV дикого типа. Например, имеется модифицированный промотор HIV, который проявляет увеличенную транскрипционную активность в присутствии интерлейкина-2. Включение данного промотора в вектор и включение вектора в клетки, инфицированные HIV дикого типа, предпочтительно ведет к увеличенной продукции и упаковке вирионов-потомков векторного генома по сравнению с геномом HIV дикого типа. Сходным образом для стимуляции селективной упаковки вирионов, кодируемых вектором, могут быть применены другие цитокины и/или хемокины (например, включая, но не ограничиваясь этим, интерферон-альфа, фактор некроза опухолей альфа, RANTES и тому подобное). Положение элементов, которые делают HIV-LTR чувствительным к цитокинам, не в коей мере не ограничивают объем изобретения такими осуществлениями. Любая нуклеотидная последовательность может быть включена в HIV-LTR для модификации чувствительности данного промотора. Перечень факторов, которые связываются с сайтами ДНК, которые могут быть вставлены в HIV-LTR, можно найти на сайте http://transfac.gb.de/TRANSFAC/lists/browse.html доступном на 8 сентября 2000 г., который включен здесь в качестве ссылки в полном изложении. Перечень ДНК-связывающих факторов для Homo sapiens (http://transfac.gbf.de/TRANSFAC/lists/factor/species/humanHomosapiens.html) доступен на 8 сентября 2000 г., который сходно включен здесь в качестве ссылки.
Сходно сайленсеры или изоляторы могут быть также включены в промоторную или энхансерную области природного или модифицированного ретровирусного LTR, такого как HIV-LTR, так, чтобы четко регулировать экспрессию с данного промотора. В определенных случаях вставка элементов в HIV-LTR может придать вектору селективно невыгодное положение в отношении инфицирования вирусом дикого типа. Однако применение данного типа вектора может не создавать конкуренции с вирусом дикого типа за упаковку вирионов-потомков, но лучше экспрессировать последовательность интересующей нуклеиновой кислоты (или "гена полезной нагрузки"), которая, например, ингибирует репликацию HIV без конкуренции. В данном случае первая последовательность нуклеиновой кислоты не дает вектору селективного преимущества, но должна в минимальном количестве, например, ингибировать рекомбинацию между вектором и хелпером в процессе продукции вектора.
Вторая категория последовательности нуклеиновой кислоты, которая придает селективное преимущество вектору, содержащему последовательность, над штаммом вируса дикого типа, включает в качестве предпочтительной последовательности нуклеиновой кислоты противовирусную последовательность нуклеиновой кислоты. "Противовирусные агенты" классифицируются по типу своего действия, например ингибиторы обратной транскриптазы, конкуренты с вирусом за вход в клетки, вакцины, ингибиторы протеаз и генетические противовирусные агенты. "Генетические противовирусные агенты" представляют собой молекулы ДНК или РНК, которые переносятся в клетки и влияют на свои внутриклеточные мишени либо прямо (т.е. в том виде, в котором введены внутрь клетки), либо после своего превращения либо в РНК, либо в белок (обобщено Dropulic et al. (1994), выше). Генетическая противовирусная последовательность также является предпочтительной последовательностью нуклеиновой кислоты. Генетические противовирусные агенты включают, но не ограничиваются этим, антисмысловые молекулы, ловушки РНК, трансдоминантные мутанты, токсины, агенты, модифицирующие и модулирующие сплайсинг РНК и белка, EGS (сами или в прямой связи с элементами M1, U1, PRE или CTE), иммуногены, РНКi (смотри Zamore et al. Cell 101:25-33, 2000), нуклеотидные последовательности или молекулы, которые модифицируют или модулируют сплайсинг, конститутивные элементы транспорта (CTE), факторы ядерного импорта и экспорта, факторы интеграции ДНК, элементы, стабилизирующие или дестабилизирующие РНК, посттранскрипционные регуляторные элементы (PRE), белки, которые вмешиваются в репликацию вирусов, интерфероны, токсины, иммуногены (определяемые любым геном, связанным с иммунной системой), антитела (цельные антитела, одноцепочечные антитела или молекулы, представляющие лиганды), рибозимы или любой другой элемент, который любым образом влияет на репликацию вектора или вируса дикого типа, и рибозимы. Желательно, чтобы генетический противовирусный агент представлял собой антисмысловую молекулу, трансдоминантный мутант, иммуноген и рибозим. Соответственно, предпочтительная последовательность нуклеиновой кислоты, которая придает селективное преимущество вектору над штаммом вируса дикого типа, является таковой генетического противовирусного агента, выбранного из группы, состоящей из антисмысловой молекулы, иммуногена и рибозима.
Генетические противовирусные агенты, применяемые в настоящем изобретении, ограничиваются только нацеливанием на вирусную последовательность, особенно когда они не помещены в вирусные векторы с зависящей от условий репликацией. В качестве неограничивающего примера генетическая противовирусная последовательность может быть помещена в лентивирусный вектор так, что агент направлен на вирусные или невирусные мишени, например клеточные, бактериальные или паразитарные мишени. В данном случае лентивирусный вектор содержит интересующий ген, оперативно связанный с модифицированным или немодифицированным HIV-LTR (для экспрессии неподвергнутой или подвергнутой сплайсингу РНК), и дополнительно содержит генетический противовирусный агент или последовательность, которая присоединена к промоторной последовательности, которая оперативно не связана с HIV-LTR. Предпочтительная последовательность представляет собой антисмысловую последовательность или рибозимы, которые представляют собой химеры с snРНК, предпочтительно U1, U2, U3, U4, U5 или U6 snРНК.
Также генетический противовирусный агент не ограничивается одной или однотипной последовательностью, присутствующей либо в векторной, либо в хелперной последовательности. Два или более агентов могут одновременно присутствовать в векторном или хелперном конструкте, либо локализованных дистально, либо предпочтительно соединенных в виде тандема. Предпочтительно два или более различных генетических противовирусных агентов соединяют в виде тандема. Дополнительное осуществление генетических противовирусных агентов для применения в изобретении представляет собой любое, при котором соединяют два различных типа генетических противовирусных агентов.
"Антисмысловая молекула" представляет собой молекулу, которая является "зеркальным отражением" на основе правил пар нуклеотидов и наличия "неоднозначности" пар нуклеотидов, короткого участка гена, экспрессию которого надо блокировать. Антисмысловая молекула, направленная против HIV, гибридизуется с РНК HIV дикого типа, создавая возможность ее предпочтительной деградации под действием клеточных нуклеаз. Антисмысловые молекулы предпочтительно являются олигонуклеотидами ДНК, желательно от приблизительно 20 до приблизительно 200 пар нуклеотидов в длину, предпочтительно от приблизительно 20 до приблизительно 50 пар нуклеотидов в длину и оптимально менее 25 пар нуклеотидов в длину. С РНК crHIV может быть экспрессирована антисмысловая молекула, которая предпочтительно связывается с геномной РНК дикого типа, придавая в результате этого РНК crHIV селективное преимущество для упаковки вирионов-потомков.
Антисмысловые молекулы, экспрессируемые в векторах в виде РНК, предпочтительно составляют, по меньшей мере, от 20 нуклеотидов до любого размера в длину, но предпочтительно до 2000 нуклеотидов в длину, более предпочтительно от приблизительно 50 до 500 нуклеотидов в длину. Такие антисмысловые молекулы предпочтительно связываются с геномной РНК дикого типа, а не с векторной РНК, придавая вектору селективное преимущество над вирусом дикого типа. Предпочтительной областью для создания антисмысловой молекулы в векторе, происходящем от HIV, например, является область из области оболочки (env), областей белков Tat или Rev, областей вспомогательных генов (Vif, Nef, Vpr, Vpu), областей Gag, которые находятся 3'("правее") по отношению к сайту рестрикции Nsi I на pNL4-3, и областей Pol, которые не содержат область 545 нуклеотидов в Pol, как описано ниже.
"Иммуноген" относится к любому относящемуся к иммунной системе гену и кодируемому им генному продукту. Хотя исторически термин "иммуноген" применялся в отношении адъюванта, применяемого для вакцинации, термины "иммуно-" и "-ген" применяются здесь для обозначения относящегося к иммунной системе гена и кодируемого им генного продукта. Например, иммуноген может кодировать одноцепочечное антитело (scAb), направленное против структурного белка вируса, или он может кодировать антитело, которое секретируется клеткой во внеклеточную среду. Иммуноген переносится в виде нуклеиновой кислоты и экспрессируется внутриклеточно. Сходно иммуноген также может кодировать любой антиген, белок клеточной поверхности (включая ограниченные определенным классом) или индикаторно-подобное антитело, который облегчает селекцию вектора и/или клетки-хозяина. В предпочтительном векторе последовательность нуклеиновой кислоты включает последовательность, кодирующую scAb, которое связывается с белком Rev HIV дикого типа. Это предпочтительно предотвращает созревание белка Rev в результате его удержания в эндоплазматическом ретикулуме. Конкретно, белки Rev, сами или в сочетании с другими клеточными факторами, экспортируют несплайсированную и раздельно сплайсированную РНК HIV из ядра в цитоплазму с помощью связывания с RRE (высоко структурированной цис-действующей последовательностью РНК-мишени для Rev, обозначаемой Rev-чувствительным элементом) и затем олигомеризуются вокруг РНК HIV. РНК HIV, которые комплексируются с Rev, экспортируются в цитоплазму и обходят клеточный механизм сплайсинга. Таким образом, если Rev дикого типа не связывается с RRE дикого типа, то РНК HIV дикого типа не экспортируются в цитоплазму и не инкапсидируются в вирионы-потомки.
Оптимально, чтобы вектор, содержащий последовательность нуклеиновой кислоты scAb, дополнительно содержал модифицированную последовательность RRE и кодировал мутантный белок Rev, который узнает модифицированный RRE, но не RRE дикого типа. Соответственно, в клетках, содержащих HIV дикого типа и вектор, включающий последовательность нуклеиновой кислоты scAb, вектор предпочтительно упаковывается в вирионы. Сходная стратегия предпочтительно применяется, когда белки матрикса или нуклеокапсида HIV дикого типа (т.е. или любой белок, вовлеченный во взаимодействия белок/РНК, который влияет на инкапсидацию вирусной РНК) являются мишенями scAb.
"Рибозим" представляет собой антисмысловую молекулу с каталитической активностью, т.е. вместо связывания РНК и ингибирования трансляции рибозимы связывают РНК и вызывают сайт-специфическое расщепление связанной молекулы РНК. В целом существуют четыре группы рибозимов: вмешивающаяся последовательность группы I Tetrahymena, EGS (внешняя руководящая последовательность) и рибозимы в форме головки молотка и шпильки для волос. Однако существуют также дополнительные каталитические мотивы в других молекулах РНК, например РНК вируса гепатита дельта и рибосомальных РНК в митохондриях грибов.
Предпочтительным рибозимом является рибозим, в котором каталитический домен расщепляет 3'-нуклеотидную последовательность NUH, где N может быть любым нуклеотидом (т.е. G, A, U или C) и H может быть либо A, C или U. Однако в виду того, что последовательность, которая расщепляется наиболее эффективно такими рибозимами, представляет собой сайт GUC, предпочтительно, чтобы последовательность NUH включала сайт GUC.
Желательно, чтобы такой рибозим расщеплял в области штамма вируса дикого типа или его транскриптов, но не расщеплял в области вектора или его транскриптов. Рибозим расщепляет вирус или его транскрипты, в том смысле, что такой вирус или вектор может представлять собой либо РНК, либо ДНК, как описано ранее. Под расщеплением "в области" подразумевается расщепление в области-мишени, т.е. предпочтительно в области вируса, которая необходима для размножения вируса. Желательно, чтобы вектор был модифицирован так, чтобы данная конкретная область, являющаяся мишенью (т.е. если она вообще присутствует в векторе), не расщеплялась рибозимом. Необязательно рибозим может расщеплять вектор, если расщепление не происходит в области, требуемой для размножения вирусных, например crHIV, частиц.
Оптимально, чтобы рибозим кодировался последовательностью, выбранной из группы, состоящей из SEQ ID NO:3 (т.е. CACACAACACTGATGAGGCCGAAAGGCCGAAACGGGCACA) и SEQ ID NO:4 (т.е. ATCTCTAGTCTGATGAGGCCGAAAGGCCGAAACCAGAGTC). В то время как SEQ ID NO:3 включает рибозим, который направляют к сайту +115 (т.е. в терминах количества нуклеотидов ниже старта транскрипции) области U5 HIV дикого типа, SEQ ID NO:4 включает рибозим, который направляют к сайту +133 области U5 HIV дикого типа.
Такой рибозим способен расщеплять геном HIV дикого типа (или его транскрипты), но не векторный геном (или его транскрипты), в виду того, что последовательности вектора U5 модифицированы с помощью сайт-специфического мутагенеза in vitro, как это известно в данной области техники и описано в примере 1. В частности, последовательности вектора предпочтительно модифицируют так, что вектор включает последовательность, выбранную из группы, состоящей из SEQ ID NO:2 (т.е. GTGTGCCCACCTGTTGTGTGACTCTGGCAGCTAGAGAAC), SEQ ID NO:5 (т.е. GTGTGCCCGCCTGTTGTGTGACTCTGGTAACTAGAGATC), SEQ ID NO:6 (т.е., GTGTGCCCGTCTGTTGTGTGACTCTGGCAAC TAGAGATC), SEQ ID NO:14, в которой, по меньшей мере, один N мутирован, SEQ ID NO:15 и SEQ ID NO:16. В форме РНК вектор предпочтительно включает последовательность, кодируемую последовательностью, выбранной из группы, состоящей из SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:14, в которой, по меньшей мере, один N мутирован, SEQ ID NO: 15 и SEQ ID NO: 16. В отличие от этого HIV дикого типа включает последовательность US, кодируемую последовательностью SEQ ID NO:1 (т.е. GTGTGCCCGTCTGTTGTGTGACTCTGGTAACTAGAGATC). Модификации в целом и сравнение с последовательностью U5 дикого типа (в форме ДНК) представлены на Фиг.2.
Более того, могут быть применены другие рибозимы, направленные на другие области вирусного генома, и особенно генома HIV, либо по одиночке, либо в сочетании. Например, рибозим может расщеплять другие последовательности РНК, необходимые для репликации вирусов, например в последовательностях обратной транскриптазы, протеазы, или трансактиваторного белка, в последовательности Rev или в других необходимых последовательностях, таких как описанные. Предпочтительно, чтобы вектор включал множество рибозимов, например, направленных на множественные сайты. В таких случаях аналогичные последовательности в векторе модифицируют с помощью сайт-направленного мутагенеза или некоторых других способов, таких как известные в данной области техники, для получения вектора, который устойчив к такому расщеплению рибозимами.
Когда вектор представляет собой вирус иммунодефицита человека, предпочтительно, чтобы вектор был лишен гена tat и его 5' сайта сплайсинга и на его месте содержал тройную кассету анти-Tat рибозимов, где каталитический домен каждого рибозима тройной кассеты рибозимов расщепляет молекулу нуклеиновой кислоты вируса дикого типа иммунодефицита человека по различным сайтам, в частности по различным сайтам внутри tat. Предпочтительно, чтобы каталитический домен каждого рибозима расщеплял нуклеотидную последовательность в области молекулы нуклеиновой кислоты вируса дикого типа иммунодефицита человека, для которой не существует рибозим-чувствительной копии в самом векторе.
Дополнительное осуществление рибозима в качестве генетического противовирусного агента доступно в рибозимах, которые направлены на последовательности не только со спариванием нуклеотидов по Уотсону-Крику, но и с образованием неоднозначного спаривания нуклеотидов G-U. Предшествующие исследования показали, что неоднозначное спаривание нуклеотидов G-U в двуспиральной РНК имеет сходные черты с образованием пар нуклеотидов по Уотсону-Крику. Принимая во внимание высокую скорость мутаций даже в высококонсервативных областях HIV, приводящую к образованию различных штаммов HIV, возможность создания рибозима, который направлен на многие штаммы, является существенным преимуществом настоящего изобретения. Например, высококонсервативная область-мишень в гене tat, которая может быть использована в качестве последовательности-мишени для рибозимов настоящего изобретения, содержит либо "G", либо "A" в 40 хорошо изученных штаммах HIV-1. Таким образом, рибозим, направленный на данную область, может быть создан как содержащий "U" вместо "C" в подходящей направляющей последовательности для обеспечения образования пар с остатком либо "G", либо "A". Это можно сравнить с применением "С" в направляющей последовательности, которое должно снижать способность рибозима поражать штаммы HIV-1, содержащие в соответствующем положении "А". Данный подход с применением "U" везде, где имеется вариация G, A в положении, узнаваемом рибозимом, существенно увеличивает применимость рибозимов изобретения.
Дополнительное осуществление для применения раскрытого изобретения заключается в использовании кассеты рибозимов (которая содержит более одного рибозима, связанных в тандем), вырожденной так, что она не представляет собой прямо повторенную последовательность в векторе. Присутствие вырожденных последовательностей предотвращает делецию повторов последовательностей, как наблюдается в ретровирусных векторах и других нуклеиновых кислотах. Следовательно, во время множественных циклов репликации может произойти делеция кассеты рибозимов, состоящей из тандемно-организованных прямых повторов последовательностей. Вырожденные последовательности рибозимов могут быть использованы для преодоления данной проблемы, потому что последовательности в каталитическом домене молекулы рибозимов в форме головки молотка, например, могут быть замещены другими нуклеотидами на основе принципа образования неоднозначных пар нуклеотидов без существенной потери активности. Ранее был осуществлен мутагенез рибозимов в форме головки молотка и определены сайты, которые могут быть вырожденными (Ruffner DE, Stormo GD, Uhlenbeck OC. Sequence requirements of the hammerhead RNA self-cleavage reaction. Biochemistry. 1990, Nov 27; 29(47):10695-702). Другой подход к снижению вероятности делеций заключается в направлении индивидуальных рибозимов кассеты к различным сайтам-мишеням РНК-мишени. Таким образом, кассета не будет иметь в наличии прямых повторов последовательностей.
"Трансдоминантная" или "доминантная негативная" последовательность нуклеиновой кислоты дает свой кодируемый фенотип даже в присутствии последовательности дикого типа. Одним примером является обсуждение укороченного белка CCR5 (смотри выше). Другие примеры включают любую мутантную ретровирусную или лентивирусную последовательность, которая дает трансдоминантный фенотип. Примеры включают гены rev и gag.
Одним примером применения для обеспечения селективного преимущества, а также ингибирования инфицирующего вируса дикого типа является применение трансдоминантных последовательностей gag, которые, как показано, ингибируют репликацию HIV-1, возможно, путем создания препятствия для сборки капсида. Таким образом, такую последовательность предпочтительно применяют в сочетании с ретровирусным вектором не на основе HIV-1 для обеспечения его репликации и упаковки без торможения трансдоминантной последовательности. Например, ретровирусный вектор, содержащий гены gag HIV-2 и pol, может включать трансдоминантную последовательность gag HIV-1. Такой вектор может быть реплицирован и упакован в результате комплементации подходящим хелперным векторным конструктом HIV-2. Но после развертывания в клетке, подвергнутой инфицированию HIV-1, трансдоминантная последовательность gag способна экспрессироваться при инфицировании HIV-1, блокируя мобилизацию вируса HIV-1. Ретровирусный вектор, кодирующий гены gag и pol HIV-2, является, однако, все еще доступным для инкапсидации и мобилизации вектора.
2. Последовательность нуклеиновой кислоты, наличие которой придает селективное преимущество клеткам, инфицированным или неинфицированным вектором, включающим последовательность, по сравнению с клетками, инфицированными штаммом вируса дикого типа.
Последовательность нуклеиновой кислоты, которая придает селективное преимущество клетке, содержащей вектор, включающий последовательность, по сравнению с клеткой, содержащей или не содержащей штамм вируса дикого типа (т.е. у которой отсутствует последовательность), предпочтительно представляет собой любую последовательность, которая позволяет клетке, содержащей вектор, способствовать выживанию и размножению вирусных частиц (т.е. вирусных частиц crHIV) по сравнению с клеткой, содержащей вирус дикого типа, или с клеткой, у которой отсутствует последовательность нуклеиновой кислоты. Такие последовательности включают, но не ограничиваются этим, любую последовательность, которая позволяет клетке или вектору, содержащемуся в клетке, избежать разрушения, последовательности, которые стимулируют выживание клетки, последовательности, которые индуцируют апоптоз, последовательности, которые облегчают продукцию белков, или последовательности, которые стимулируют иммунную функцию или ее направленность.
Например, предпочтительно, чтобы такая последовательность нуклеиновой кислоты, содержащаяся в векторе, кодировала гены множественной устойчивости к лекарствам (смотри, например, Ueda et al., Biochem. Biophys. Res. Commun. 141, 956-962 (1986); Ueda et al., J. Biol. Chem., 262, 505-508 (1987); и Ueda et al., PNAS, 84, 3004-3008 (1987)). В присутствии добавленного цитотоксического лекарства (например, как применяется при химиотерапии рака) это позволяет клетке, содержащей вектор, выжить, в то время как клетка, которая содержит вирус дикого типа, такой как HIV, не способна выжить. Такие цитотоксические лекарства включают, но не ограничиваются этим, актиномицин D, сульфат винбластина, сульфат винкристина, BCNU (с или без улучшения с помощью BG), дауномицин, адриамицин, VP-16 и AMSA.
Другим примером такой последовательности нуклеиновой кислоты является любой вариант последовательности O6-метилгуанин-ДНК-метилтрансферазы (MGMT). MGMT может использоваться для защиты гематопоэтических предшественников от токсичности алкилирующих агентов. MGMT дикого типа ингибируется O6-бензилгуанином (BG), который усиливает токсичность алкилирующего агента. Варианты MGMT, которые устойчивы к опосредуемой BG инактивации и способны защищать против алкилирующих агентов, могут быть предоставлены любому типу клеток с помощью настоящего изобретения. Примером такого варианта является G156A MGMT.
Например, гематопоэтические клетки-предшественники, трансдуцированные таким вариантом настоящего изобретения, могут быть отобраны при введении BG и агента, метилирующего или хлорэтилирующего O6-гуанин. Примеры таких метилирующих агентов для применения в изобретении включают, но не ограничиваются этим, клинически пригодные темозоломид, нитрозомочевину, тетразины, триазины, дакабазин, темозоломид, стрептозотоцин, прокарбазин. Примеры хлорэтилирующих агентов включают, но не ограничиваются этим, BCNU (1,3-бис(2-хлорэтил)-1-нитрозомочевину), CCNU (3-циклогексил-1-хлорэтил-нитрозомочевину), ACNU (1-(4-амино-2-метил-5-пиримидинил)метил-3-(2-хлор)-3-нитрозомочевину) и MeCCNU (1-(2-хлорэтил)-3-(4-метилциклогексил)-1-нитрозомочевину. Предпочтительным алкилирующим агентом является BCNU.
BCNU, как показано, формирует постоянное повреждение путем ковалентной межцепочечной сшивки ДНК в клетках, как в стадии покоя, так и в цикле пролиферации. Данные повреждения являются цитотоксическими для клеток в процессе репликации ДНК. MGMT является первичным механизмом репарации ДНК после повреждений под действием BCNU. В результате передачи его токсичности клеткам в стадии покоя отбор вариантов MGMT, доставляемых лентивирусными векторами изобретения, может также позволить отбирать тотипотентные гемопоэтические стволовые клетки (HSCs), которые находятся преимущественно в стадии G0 клеточного цикла, но которые циклируют периодически для получения большего количества стволовых клеток и клеток-предшественников крови. Стволовые клетки обычно устойчивы к ретровирусной трансдукции, но могут быть трансдуцированы ретровирусными векторами, если они находятся не только в стадии G0 цикла. Подобно онкоретровирусам, лентивирусные векторы изобретения могут быть использованы для трансдукции гематопоэтических стволовых клеток, включая пригодные для тестирования гемопоэтические ELTC-IC и NOD-SCID или SCID-hu клетки с воспроизводимой популяцией. Однако в отличие от онкоретровирусов состав цитокинов, требуемый для трансдукции стволовых клеток без их дифференцировки, более сходен с лентивирусными векторами, так как лентивирусы (например, HIV) могут инфицировать клетки, которые активно не делятся в момент инфицирования. Селекция HSC невозможна с помощью других стратегий, которые заключаются в отборе циклирующих гематопоэтических предшественников. Таким образом, селекция, опосредуемая вариантами MGMT и лентивирусными векторами изобретения, может увеличить присутствие трансдуцированных тотипотентных HSCs в отсутствие необходимости длительного введения лекарства для селекции.
Представленное выше обсуждение может быть обобщено в контексте гематопоэтических клеток in vitro и ex vivo, а также in vivo. Таким образом, в дополнение к трансдукции вариантами MGMT клеток in vivo гематопоэтические клетки субъекта могут быть трансдуцированы ex vivo с последующим лекарственным лечением либо ex vivo, либо in vivo. Такие клетки, обработанные ex vivo, могут быть затем инфузированы субъекту, необязательно в качестве частичной замены популяции клеток костного мозга трансдуцированными клетками.
Перед осуществлением описанного выше может быть также использован алкилирующий агент BCNU для снижения в целом инфицированных HIV клеток CD4+ у HIV-позитивных субъектов. Как установлено выше, BCNU (после истощения эндогенной MGMT) дает цитотоксические повреждения как в клетках в стадии покоя, так и в пролиферирующих клетках. Такие повреждения образуются даже без принудительного истощения MGMT, если доза лекарства превышает уровень функционального белка AGT. Повторное системное введение BCNU вызывает кумулятивную миелосупрессию и панцитопению. Среди лимфоидных клеток клетки CD4+, очевидно, являются особенно чувствительными к BCNU. Таким образом, у субъектов, инфицированных ретровирусом, таких как субъекты, инфицированные HIV, введение BCNU, включая введение в малых дозах, для избежания миелосупрессии, если это желательно, будет снижать суммарную популяцию клеток CD4+ и таким образом снижать нагрузку вирусом. За данным подходом может следовать инфузия гематопоэтических клеток-предшественников, как обсуждалось выше.
Так как BCNU может действовать "скрытым" способом путем образования цитотоксических повреждений даже в клетках в стадии покоя, данный подход имеет дополнительное преимущество в снижении потребности повторных обработок. Конечно, данный подход можно сочетать с применением BG для потенцирования эффектов BCNU. В противоположном варианте подход может быть применен в сочетании с введением вариантов MGMT с векторами изобретения для обеспечения защиты трансдуцированных клеток CD4+. Данное последнее средство может быть либо независимым, либо применяться в сочетании с обработкой гематопоэтических клеток, как обсуждалось выше. Следовательно, предпочтительное осуществление изобретения заключается в экспрессии мутантной MGMT (второй последовательности нуклеиновой кислоты) в векторе, который содержит анти-HIV антисмысловую последовательность или рибозим (первую последовательность нуклеиновой кислоты), так, чтобы вектор обеспечивал селективное преимущество над клетками, инфицированными HIV дикого типа, и селективное преимущество над вирусом HIV дикого типа, соответственно. Дополнительное предпочтительное осуществление изобретения заключается в том, что вектор экспрессирует ген MGMT (или вторую последовательность нуклеиновой кислоты) вне промотора HIV-LTR в виде сплайсированной мРНК.
Наконец, хотя векторы экспрессируют анти-HIV рибозимы или антисмысловые последовательности, изобретение этим не ограничивается, так как специалист в данной области техники может легко вставить любую ингибиторную нуклеотидную последовательность в вектор для конкретного терапевтического, профилактического или биологического эффекта. Предпочтительный способ экспрессии ингибиторной последовательности заключается во включении ее в U1 snРНК/промоторную кассету, как описано Dietz (патент США 5814500).
В противоположном варианте такая последовательность нуклеиновой кислоты желательно включает последовательность, выбранную из группы, состоящей из последовательности (или последовательности, которая кодирует) мутантной (т.е. мутанта) протеазы и последовательности (или последовательности, которая кодирует) мутантной (т.е. мутанта) обратной транскриптазы. Предпочтительно, чтобы мутантную обратную транскриптазу конструировали так, чтобы она была устойчива к нуклеозидным и ненуклеозидным ингибиторам обратной транскриптазы, а мутантную протеазу конструировали так, чтобы она была устойчива к обычно применяемым ингибиторам протеаз.
Введение данных ингибиторов протеазы или обратной транскриптазы хозяину в сочетании с вектором применяется для отбора на клетки, продуцирующие вектор в противоположность клеткам, продуцирующим вирус дикого типа. Сходно данный подход модифицируется для применения с любым лекарством, которое ингибирует репликацию вируса так, что вирус может быть подвергнут мутации для избежания ингибирования. Соответственно, для лечения HIV селективная последовательность нуклеиновой кислоты, включенная в вектор, предпочтительно включает мутантные последовательности HIV. Оптимально, однако, чтобы данные последовательности не предотвращали суперинфицирование HIV дикого типа.
Предпочтительно, чтобы вектор представлял собой один из указанных выше и, в частности, усовершенствованные векторы с зависящей от условий репликацией, изображенные на фиг.1А-1К. Конечно, при применении в способах изобретения последовательности, кодирующие GFP, могут быть удалены или замещены другими последовательностями. Последовательности, кодирующие GFP, присутствуют в данных примерах осуществлений векторов в качестве маркера или индикатора присутствия вектора или экспрессии гена из вектора.
Предпочтительные векторы изобретения могут содержать различные элементы, описанные в настоящем раскрытии. В частности, в лентивирусных векторах может отсутствовать ген tat и содержаться, по меньшей мере, одна антисмысловая последовательность. Дополнительно векторы могут содержать последовательность, такую как центральный полипиримидиновый тракт HIV или содержащий его более крупный фрагмент нуклеиновой кислоты.
Оптимально, чтобы вектор был совместим с клеткой, в которую его вводят, например, был способен осуществлять экспрессию в клетке последовательностей нуклеиновой кислоты, кодируемых вектором. Желательно, чтобы вектор включал ориджин репликации, функционирующий в клетке. Когда последовательность нуклеиновой кислоты переносится в форме своей кодирующей последовательности ДНК (например, в отличие от формы полного гена, включающего свой собственный промотор), оптимально, чтобы вектор также содержал промотор, который способен индуцировать экспрессию кодирующей последовательности и который оперативно связан с кодирующей последовательностью. Кодирующая последовательность является "оперативно связанной" с промотором (например, когда как кодирующая последовательность, так и промотор совместно составляют природный или рекомбинантный ген), когда промотор способен направлять транскрипцию кодирующей последовательности.
В рекомбинантном векторе настоящего изобретения предпочтительно все присущие сигналы транскрипции (например, сигналы инициации и терминации), трансляции (например, вход рибосом или сайт связывания или тому подобное), сигналы процессинга (например, донорские и акцепторные сайты сплайсинга, если это необходимо, и сигналы полиаденилирования), транслокации, сборки, сайтов интеграции, входа рибонуклеиновых комплексов, стабильности и элементов транслокации в цис- или транс-положениях организованы в векторе корректно, так что любой ген или кодирующая последовательность транскрибируется присущим ей образом (и/или транслируется, если это желательно) в клетках, в которые введен вектор. Манипуляции с такими сигналами для уверенности в их соответствующей экспрессии в клетках-хозяевах хорошо известны и находятся в компетенции специалистов в данной области техники.
Предпочтительно, чтобы вектор содержал последовательность Pol, которая увеличивает эффективность стабильной трансдукции, определяемой как интегрированные копии вектора в геноме клетки-хозяина. Последовательность, ранее идентифицированная Zennou et al. (Cell 101:173-185 (2000)) как центральное крыло ДНК (фрагмент из 178 пар нуклеотидов в положениях от 4793 до 4971 в pLAI3, соответствующий положениям от 4757 до 4935 в pNL4-3), как указывалось, увеличивает эффективность трансдукции, но было найдено, что она неэффективна в отношении значительного увеличения эффективности стабильной трансдукции. Настоящее изобретение включает раскрытие того, что в то время как данный небольшой фрагмент не достаточен для увеличения эффективности стабильной трансдукции, более крупный фрагмент из 545 пар нуклеотидов (положения от 4551 до 5096 в pNL4-3) или содержащие его еще более крупные фрагменты, как описано в патенте США 5885806, способны увеличивать стабильную трансдукцию, являясь частью настоящего изобретения. Увеличение эффективности стабильной трансдукции было определено с помощью экспрессии GFP и анализа FACS, а также Taqman-анализа интегрированных копий векторного генома.
Дополнительные средства для увеличения эффективности трансдукции описаны в параллельно поданной патентной заявке США с серийным номером (еще будет присвоен), зарегистрированной 31 августа 2000 г., под номером выданного патентным поверенным документа № 397272000400, которая включена здесь в качестве ссылки в полном изложении.
Вирусные векторы, применяемые в настоящем изобретении, могут также приводить к образованию "псевдотипа", когда совместное инфицирование клетки различными вирусами дает вирионы-потомки, содержащие геном одного вируса и имеющие внешний слой капсида, содержащий один или более белков оболочки от другого вируса. Данный феномен был применен для упаковки интересующих вирусных векторов в "псевдотипные" вирионы с помощью совместной трансфекции или совместного инфицирования упаковывающей клетки, как интересующим вирусным вектором, так и генетическим материалом, кодирующим, по меньшей мере, один белок оболочки другого вируса или молекулу клеточной поверхности. Смотри патент США 5512421. Такие смешанные вирусы могут быть нейтрализованы с помощью антисывороток против одного или более используемых гетерологичных белков оболочки. Одним вирусом, обычно применяемым при образовании псевдотипа, является вирус везикулярного стоматита (VSV), который представляет собой рабдовирус. Применение псевдотипирования расширяет диапазон клеток-хозяев для вируса путем включения элементов механизма вирусного входа от применяемого гетерологичного вируса. Псевдотипирование вирусных векторов и VSV для применения в настоящем изобретении дает вирусные частицы, содержащие нуклеиновую кислоту вирусного вектора, инкапсулированную в нуклеокапсид, который окружен мембраной, содержащей белок G VSV. Нуклеокапсид предпочтительно содержит белки, обычно ассоциированные с вирусным вектором. Окружающая мембрана, содержащая белок G VSV, образует часть вирусной частицы при выходе ее из клетки, применяемую для упаковки вирусного вектора. Примеры упаковывающих клеток описываются в патенте США 5739018. В предпочтительном осуществлении изобретения вирусная частица происходит от HIV и псевдотипирована с белком G VSV. Псевдотипированные вирусные частицы, содержащие белок G VSV, могут инфицировать разнообразный набор клеточных типов с большей эффективностью, чем амфотропные вирусные векторы. Диапазон клеток-хозяев включает виды как млекопитающих, так и немлекопитающих, такие как человек, грызуны, рыбы, амфибии и насекомые.
Предпочтительно, чтобы вектор также включал некое средство, с помощью которого вектор или содержащую его субклонированную последовательность идентифицировали и отбирали. Идентификация и/или отбор вектора осуществляется с применением множества подходов известных специалистам в данной области техники. Например, векторы, содержащие конкретные гены или кодирующие последовательности, предпочтительно идентифицируют с помощью гибридизации, в присутствии или в отсутствие так называемых "маркеров" функций генов, кодируемых маркерными генами, присутствующими в векторах, и/или при экспрессии конкретных последовательностей. При первом подходе присутствие конкретной последовательности в векторе определяется с помощью гибридизации (например, с помощью гибридизации ДНК-ДНК) с применением зондов, включающих последовательности, которые являются гомологичными соответствующей последовательности. При втором подходе рекомбинантную систему вектор/хозяин идентифицируют и отбирают на основе присутствия или отсутствия определенных функций маркерных генов, таких как устойчивость к антибиотикам, активность тимидинкиназы и тому подобное, обусловленных определенными генами, кодирующими данные функции, присутствующими в векторе. При третьем подходе векторы идентифицируют с помощью тестирования конкретного продукта гена, кодируемого вектором. Такие тесты основаны на физических, иммунологических или функциональных свойствах продукта гена.
Соответственно, в настоящем изобретении также предлагается вектор, который, если это ДНК, включает нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NOS: 2, 4, 5, 6, 15 и 16, и который, если это РНК, включает нуклеотидную последовательность, кодируемую нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NOS: 4, 5, 6.
В настоящем изобретении дополнительно предлагается способ конструирования вектора, который происходит от вируса дикого типа иммунодефицита человека и который способен реплицироваться только в клетке-хозяине, которая является пермиссивной для репликации указанного вектора, с рибозимом. Рибозим, который включают в вектор или который кодируется вектором, расщепляет нуклеиновую кислоту вируса дикого типа иммунодефицита человека, но не самого вектора и его транскриптов, если они есть. Способ включает получение вектора, который происходит от вируса дикого типа иммунодефицита человека и который способен реплицироваться только в клетке-хозяине, которая является пермиссивной для репликации указанного вектора, и включение в вектор последовательности нуклеиновой кислоты, которая включает или кодирует рибозим, каталитический домен которого расщепляет нуклеиновую кислоту вируса дикого типа иммунодефицита человека, но не самого вектора и его транскриптов, если они есть. В таком способе нуклеотидная последовательность, включающая или кодирующая последовательность U5 вируса дикого типа иммунодефицита человека, может быть удалена из вектора и замещена нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NOS: 2, 5, 6, 14, в которой, по меньшей мере, один N мутирован, 15 и 16, если вектор представляет собой ДНК, и нуклеотидной последовательностью, кодируемой нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NOS: 2, 5, 6, 14, в которой, по меньшей мере, один N мутирован, 15 и 16, если вектор представляет собой РНК. Предпочтительно, чтобы вектор реплицировался в клетке-хозяине, пермиссивной для репликации указанного вектора, более одного раза.
В настоящем изобретении также предлагается способ модификации вектора. Способ включает получение вектора и введение в вектор нуклеотидной последовательности, выбранной из группы, состоящей из последовательностей ДНК SEQ ID NOS: 2, 3, 4, 5, 6, 14, в которой, по меньшей мере, один N мутирован, 15 и 16, если вектор представляет собой ДНК, и нуклеотидной последовательности, кодируемой нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NOS: 2, 4, 5, 6, 15 и 16, если вектор представляет собой РНК.
В настоящем изобретении дополнительно предлагается способ размножения и селективной упаковки вектора с зависящей от условий репликацией без применения упаковывающей клеточной линии. Способ включает контактирование вектора с зависящей от условий репликацией с клеткой, способной быть инфицированной другим вектором, который является вектором того же самого типа, что и вектор с зависящей от условий репликацией, и который отличается от вектора с зависящей от условий репликацией сохранением репликационной компетенции вектора дикого типа; последующее контактирование клетки с другим вектором и затем культивирование клетки в условиях, приводящих к размножению вектора с зависящей от условий репликацией. Хелперный вектор, обсуждаемый выше и более подробно описываемый ниже, является одним из таких дополнительных векторов.
Предлагается также молекула выделенной и очищенной нуклеиновой кислоты, выбранная из группы, состоящей из молекулы ДНК, включающей нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NOS: 2, 5, 6, 14, в которой, по меньшей мере, один N мутирован, 15 и 16, и молекулы РНК, включающей нуклеотидную последовательность, кодируемую нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NOS: 2, 6, 15 и 16.
Способ применения
Описанные выше векторы предпочтительно вводят в клетку-хозяин для профилактического или терапевтического лечения вирусной инфекции, для облегчения поддержания вектора, а также по другим причинам. Соответственно, в настоящем изобретении предлагается клетка-хозяин, включающая вектор согласно изобретению. Выделение клеток-хозяев и/или поддержание таких клеток или ведущих от них начало линий клеток в культуре стало обычным делом, в котором хорошо разбирается специалист в данной области техники.
В частности, вирусный вектор с зависимой от условий репликацией или предпочтительно лентивирусный вектор, как описано выше, предпочтительно применяют для профилактического или терапевтического лечения вирусной инфекции, предпочтительно такой, где инфекция обусловлена вирусом дикого типа, предпочтительно РНК-вирусом дикого типа, еще более предпочтительно ретровирусом дикого типа и оптимально ВИЧ дикого типа.
Способ включает контактирование клетки-хозяина, восприимчивой к инфицированию вирусом дикого типа, с вектором с зависимой от условий репликацией, способным реплицироваться только в клетке-хозяине, допускающей репликацию вектора, наличие, транскрипция или трансляция которого ингибирует репликацию штамма вируса дикого типа в клетке-хозяине. Желательно, чтобы вектор реплицировался более одного раза и включал, по меньшей мере, одну последовательность нуклеиновой кислоты, обладание (т.е. наличие, транскрипция или трансляция) которой придает селективное преимущество вектору в клетке-хозяине по сравнению со штаммом вируса дикого типа, который оптимально является штаммом, от которого происходит вектор.
Согласно данному способу последовательность нуклеиновой кислоты предпочтительно включает нуклеотидную последовательность, которая включает или кодирует генетический противовирусный агент, который неблагоприятно влияет на репликацию и/или экспрессию вируса, отличного от указанного вектора. Желательно, чтобы генетический противовирусный агент был выбран из группы, состоящей из антисмысловой молекулы, рибозима и иммуногена. Оптимально, чтобы генетический противовирусный агент был рибозимом, каталитический домен которого предпочтительно расщепляет 3' нуклеотидную последовательность NUH (т.е. в особенности последовательность GUC). Необязательно рибозим кодируется, по меньшей мере частично, последовательностью, выбранной из группы, состоящей из SEQ ID NO:3 и SEQ ID NO:4. Желательно, чтобы рибозим вызывал расщепление в области штамма вируса дикого типа или его транскриптов, но не вызывал расщепление в области вектора или его транскриптов. Предпочтительно, чтобы это происходило благодаря тому, что штамм вируса дикого типа включает последовательность, кодируемую SEQ ID NO:1, тогда как вектор, если он представлен ДНК, включает нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NOS:2, 5, 6, 14, в которых, по меньшей мере, один N мутирован, 15 и 16, и, если он представлен РНК, включает нуклеотидную последовательность, кодированную нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NOS:2, 5, 6, 14, в которых, по меньшей мере, один N мутирован, 15 и 16.
Способ также желательно осуществлять, когда вектор включает, по меньшей мере, одну последовательность нуклеиновой кислоты, обладание (т.е. наличие, транскрипция или трансляция) которой придает селективное преимущество инфицированной вектором клетке-хозяину по сравнению с клеткой, инфицированной штаммом вируса дикого типа, который оптимально является штаммом вируса, от которого берет начало вектор. В этом отношении вектор может включать, по меньшей мере, одну последовательность нуклеиновой кислоты, которая придает селективное преимущество клетке-хозяину, инфицированной вирусом, и, по меньшей мере, последовательность нуклеиновой кислоты, которая придает селективное преимущество вектору по сравнению со штаммом вируса дикого типа, соответствующего вирусу, от которого берет начало вектор.
Соответственно, предпочтительно реализуют способ, в котором последовательность нуклеиновой кислоты включает нуклеотидную последовательность, кодирующую ген устойчивости ко многим лекарствам. В противоположном варианте реализуют способ, в котором последовательность нуклеиновой кислоты включает нуклеотидную последовательность, кодирующую мутированную (мутантную) протеазу, и нуклеотидную последовательность, кодирующую мутированную (мутантную) обратную транскриптазу, в случае, когда подлежащая профилактическому или терапевтическому лечению вирусная инфекция обусловлена ретровирусом.
Способ предпочтительно дополнительно включает введение в клетку-хозяина агента, выбранного из группы, состоящей из цитотоксичного лекарства, ингибитора протеазы и ингибитора обратной транскриптазы (т.е. в дополнение к введению вектора).
Соответственно, вектор может быть применен в соответствии с описанным выше способом не только для терапевтического лечения вирусной инфекции, но и для защиты потенциальной клетки-хозяина от вирусной инфекции, т.е. способом профилактического лечения вирусной инфекции или «вакцинации» против интересующего вируса, такого как РНК-вирус, в особенности ретровирус, такой как HIV (ВИЧ). Способ в значительной мере ингибирует репликацию штамма вируса дикого типа перед тем, как клетка-хозяин входит в контакт с штаммом вируса дикого типа. В этом контексте вектор может включать или кодировать белки, которые блокируют суперинфекцию вирусом дикого типа. Способ включает контактирование клетки-хозяина с вектором с зависимой от условий репликацией, как описано выше, и «вектором, экспрессирующим хелпер», т.е. геномом вируса, стимулирующим репликацию «вектора» в неинфицированном хозяине. Вектор с зависимой от условий репликацией имеет селективное преимущество при упаковке и/или размножении. Более того, вектор, например, может содержать последовательность, которая повышает выживаемость клетки, стимулирует образование вируса, индуцирует апоптоз, облегчает образование белка и/или стимулирует иммунную функцию и/или определяет направленное воздействие. Конструкт «вектора, экспрессирующего хелпер» представляет собой любой экспрессионный вектор, который комплементирует неспособность «вектора» к репликации. Такие экспрессирующие хелпер векторы являются обычными и могут быть легко сконструированы специалистами в данной области техники. Экспрессирующий хелпер вектор может быть либо упакован в вирионы, подобно вектору, либо может быть экспрессирован без необходимости упаковки. Поскольку «вектор» обладает селективным преимуществом при упаковке и/или размножении, данная система обеспечивает средства безопасности для достижения значительной репликации вируса без возможных патогенных эффектов, которые мог бы потенциально вызвать живой аттенуированный вирус. Кроме того, вектор может быть смешан с неспецифическими адъювантами для повышения иммуногенности. Такие адъюванты известны специалистам в данной области техники и включают, но не ограничиваются этим, полный или неполный адъювант Фрейнда, эмульсии, составленные из компонентов стенки бактериальной или микобактериальной клетки, и тому подобное.
Векторы с зависимой от условий репликацией или предпочтительно лентивирусные векторы изобретения могут быть, например, также применены для иммунотерапии при лечении вирусной инфекции или для лечения онкогенных заболеваний. Дендритные клетки или их предшественники (например, гематопоэтические клетки CD34+ или моноциты крови, но не ограничиваясь данными клеточными типами), а также другие презентирующие антиген клетки могут быть трансдуцированы вектором, который экспрессирует белки ВИЧ или белковую последовательность, которая содержит главные эпитопы, благодаря которым хозяин формирует иммунный ответ против чужеродного агента, такого как вирус. Такие белковые эпитопы для ВИЧ описаны в "HIV molecular immunology database", 1998, National Institutes of Allergy and Infection Diseases, Maryland & Los Alamos National Laboratory, New Mexico, которая включена здесь во всей своей полноте. Предпочтительным осуществлением для эпитопа белка ВИЧ является интегрированная или композитная последовательность ЦТЛ (или ее фрагмент), представленная ниже в примере 14. Другие эпитопы, которые могли бы быть экспрессированы сходным образом, берут начало от других вирусов, бактерий грибков, паразитов, опухолевых клеток и генетически модифицированных клеток. В одной белковой последовательности может быть объединено более одного эпитопа таким образом, что неиммуногенные сайты исключаются для создания вектора с повышенной безопасностью, поскольку кодирующие белок последовательности являются строго прерывистыми. Предпочтительное осуществление прерывистой последовательности представляет собой вырожденную прерывистую последовательность (при наличии или в отсутствие подходящих линкерных аминокислотных последовательностей для стабилизации белковой структуры, если это необходимо), кодирующую иммунологически значимые эпитопы. Применение вырожденных последовательностей снижает риск рекомбинации. Другим предпочтительным осуществлением является экспрессия описанной белковой последовательности (или ее фрагмента) с помощью подвергнутого сплайсингу месенджера, управляемого ретровирусным LTR, таким как HIV-LTR.
В связи с применением ретровирусных векторов для лечения инфекции и заболевания человека вызывает озабоченность возможность возникновения способного к репликации вируса (RCV). Одним из основных путей возникновения RCV, по-видимому, является гомологичная рекомбинация между хелперным вектором и вектором с зависимой от условий репликацией. Таким образом, в настоящем изобретении предлагается модификация конструкта хелперного вектора для сведения к минимуму или исключения возможности гомологичной рекомбинации. Настоящим изобретением рассматривается любая модификация, которая служит для снижения вероятности димеризации, совместной упаковки и/или рекомбинации хелперного вектора и вектора с зависимой от условий репликацией.
Осуществлением настоящего изобретения является вставка одного или более антивекторных рибозимов или антисмысловой последовательности (не обязательно в форме кассеты, определяемой как, по меньшей мере, две такие последовательности, соединенные последовательно, в отношении рибозимов, и определенной как, по меньшей мере, две последовательности, соединенные последовательно и направленные против прерывистых областей векторной нуклеиновой кислоты-мишени, в отношении антисмысловых последовательностей) в 3'-конец кодирующих последовательностей хелперного гена, но со стороны 5' от сайта терминации транскрипции. Хотя неспецифическая упаковка хелперных геномов в вектор может происходить, такая упаковка, по-видимому, зависит от вектора. Следовательно, может быть применен ряд подходов для снижения упаковки хелперного вектора в векторные частицы, которая в противном варианте могла бы быть первой стадией возможного возникновения RCV.
Предпочтительный хелперный конструкт включает антивекторный рибозим или антисмысловую молекулу на 3' конце последовательности, кодирующей структурный белок, или последовательности, кодирующей (гомологичную или гетерологичную) оболочку. Более предпочтительный хелперный конструкт включает антивекторный рибозим или антисмысловую молекулу на 3' конце последовательности, кодирующей структурный белок, и последовательности, кодирующей оболочку. Если последовательность нуклеиновой кислоты является антисмысловой молекулой, то она может действовать внутриклеточно, разрушая совместно локализованные двухцепочечные вектор-хелперные молекулы посредством нуклеаз клетки. Если хелпер упакован с векторной РНК в вирусные частицы, то такие молекулы не способны подвергаться полной обратной транскрипции для образования RCV.
В другом предпочтительном осуществлении в хелперный конструкт помещают последовательности, которые стимулируют дифференцированное перемещение или локализацию хелперной нуклеиновой кислоты в стороне от вектора. Неограничивающим примером является включение в хелперные конструкты гетерологичных интронных и поли-A последовательностей так, как это показано на фиг.6. Данные последовательности должны облегчать дифференцированное перемещение векторной и хелперной нуклеиновых кислот в различные участки клетки или внутриклеточных структур. Предпочтительно, чтобы титр образованного вектора менялся не более чем в 100 раз, более предпочтительно - не более чем в 10 раз, и наиболее предпочтительно - не зависел от такой модификации хелпера.
На фиг.7 показано, что присутствие одного рибозима (pVirPac1.2Rz) или рибозима и интрона, созданного для оказания влияния на перемещение в клетке хелперной РНК (pVirPac1.2RzIn), не оказывало существенного влияния на титр вектора. На фиг.12 показано, что анализ с помощью ПЦР образцов для титрования совместной упаковки хелперных конструктов показал сниженную совместную упаковку в присутствии рибозима по сравнению с высокой совместной упаковкой в отсутствие рибозима. Наличие двух рибозимов полностью предотвращало совместную упаковку хелперного вектора в векторные вирусные частицы. Таким образом, изобретение включает эффективные средства применения системы упаковки двух плазмид как для получения высоких титров вектора, так и для усовершенствования безопасности таких векторов путем снижения или исключения совместной упаковки хелперного вектора.
Результаты позволяют предполагать, что упаковка хелпера в вирионы не является случайной и зависит от вектора. В альтернативном предпочтительном осуществлении для предотвращения совместной упаковки хелперных нуклеиновых кислот с таковыми вектора применяют вырождение хелперного конструкта в областях, важных для ассоциации хелперной и векторной последовательностей. Данный подход может быть дополнительно модифицирован полным вырождением хелперной последовательности для предотвращения совместной упаковки хелперной и векторной последовательностей.
Например, нуклеотидная последовательность хелперного вектора может быть вырожденной частично или полностью. В то время как такое вырождение значительно снижает или снимает вероятность гомологичного спаривания и рекомбинации хелперного вектора и вектора с зависимой от условий репликацией, оно не влияет на способность хелперного вектора кодировать белковые продукты, необходимые для репликации и упаковки вируса. Данное вырождение может быть направлено на любой возможный ген и открытую рамку считывания (ОРС), которая является гомологичной для хелпера и векторов с зависимой от условий репликацией, или более селективно направлено на конкретные гены или ОРС внутри хелперного вектора. Следует отметить, что полное вырождение не является необходимым для снижения вероятности гомологичной рекомбинации, поскольку отсутствие гомологии последовательности из более 18 нуклеотидов в длину должно быть достаточным. Таким образом, вырождение до уровней, которые не превышают 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7 или 6 нуклеотидов, идентичных в последовательностях, будет подавлять или предотвращать рекомбинацию. Последовательности могут быть вырождены с применением кодонов, которые обладают преимущественной экспрессией у человека или в конкретном типе клеток, если это желательно.
Например, нуклеотидная последовательность, кодирующая gag, rev и/или tat, может быть вырождена полностью или частично. Другие примеры включают дегенерацию первых 42 нуклеотидов последовательности gag, первых 208 нуклеотидов последовательности rev, последних 183 нуклеотидов последовательности tat и последних 545 нуклеотидов последовательности Pol. Примерами хелперных векторов с таким вырождением являются pVirPac1.2, pVirPac1.2Rz, pVirPac1.2Rz2 и pVirPac1.2RzIn или их производные. Разумеется, нуклеотидная последовательность, кодирующая любой другой продукт вирусного гена, также может быть вырождена согласно изобретению, что тем самым снижает, сводит к минимуму или исключает вероятность гомологичной рекомбинации.
Альтернативным осуществлением настоящего изобретения является включение в хелперный вектор элемента, который направлен на вектор с зависимой от условий репликацией и вызывает его разрушение в случае совместной локализации, совместной упаковки или иного совместного спаривания двух векторов. Рибозим является примером такого элемента. В изобретении рассматривается любой рибозим, который селективно поражает вектор с зависимой от условий репликацией. Могут быть также применены множественные рибозимы, такие как описанные здесь кассеты из двух или трех рибозимов. Например, рибозим может поражать область U5 (например, область U5 HIV-1, HIV-2 или другого ретровируса, применяемого в векторе с зависимой от условий репликацией) вектора с зависимой от условий репликацией. Расщепление вектора с зависимой от условий репликацией предотвращает событие рекомбинации с вектором с зависимой от условий репликацией с образованием RCV, которое может произойти, если два вектора были упакованы совместно. Примерами хелперных векторов, содержащих рибозим, являются pVIRPAC1.1Rz и pVIRPAC1.2Rz2, как показано на фигуре 6.
Еще одним осуществлением настоящего изобретения является замена гетерологичного RRE, включая любой ретровирусный RRE, но предпочтительно другой лентивирусный RRE, в хелперном векторе, такая как замена RRE HIV-1, CTE или PRE на RRE HIV-2. Данный подход вносит вклад в снижение, сведение к минимуму или снятию возможности гомологичной рекомбинации на основе того, что разные RRE имеют разные последовательности. Дополнительным преимуществом такой замены согласно изобретению является удивительное и неожиданное повышение образования вектора с зависимой от условий репликацией, равное пятикратному повышению. Вне связи с теорией возможно, что различные RRE в хелперном векторе и векторе с зависимой от условий репликацией связываются с одним или более различными факторами клетки, количество которых может быть ограниченным. В результате применение разных RRE может снижать конкуренцию между двумя векторами за ограниченные клеточные факторы. Примерами хелперных векторов, содержащих элемент RRE HIV-2 для упаковки ретровирусного вектора на основе HIV-1, являются pVIRPAC-1.2 и pVIRPAC-1.2Rz2, как показано на фиг.6.
Еще одним осуществлением является включение в хелперный вектор нуклеотидной последовательности, кодирующей гетерологичный белок env, такой как VSV-G из вируса везикулярного стоматита, белок G бешенства, GaLV, гликопротеин E1/2 альфавируса или RD114 - белок env эндогенного вируса кошачьих. Это обеспечивает упаковку вектора с зависимой от условий репликацией в частицы с гетерологичными белками env. Более того, в белок оболочки необязательно может быть вставлен лиганд для стимуляции клетки-мишени во время трансдукции, если это желательно. Поскольку модификация белков оболочки может нарушать их связываюшую способность, предпочтительным осуществлением является экспрессия модифицированного лиганда, содержащего гибридный белок оболочки, с немодифицированным псевдотипированным белком оболочки во время образования. Таким образом, оба типа белков оболочки должны находиться на поверхности клетки, один из которых предназначен для стимуляции клетки-мишени, а другой - для опосредования связывания и вхождения. Предпочтительным псевдотипированным лентивирусным вектором, который также экспрессирует гибридный рецептор для стимуляции клеток, является белок оболочки VSV-G и гибридный белок оболочки VSV-G. Более предпочтительным гибридным белком оболочки является гибридный белок оболочки VSV-G/RD114 (смотри фиг.14A и 14B). Другие предпочтительные белки (или их фрагменты) для создания гибридов с VSV-G включают notch, дельта, лиганд FLT-3, TPO, лиганд Kit, лиганд, который связывается с CD3, лиганд, который связывается с CD28, и лиганд, который связывается с GM-CSF.
Последовательность, кодирующая гетерологичный белок env, может быть необязательно оперативно связана со вторым индуцибельным промотором для обеспечения более обильной поставки белка env для упаковки вируса. Данный подход может дополнительно увеличивать продукцию вируса, в особенности в условиях, при которых продукция белка env является лимитирующим фактором. Последовательность, кодирующая гетерологичный белок env, может находиться либо в том же хелперном векторе, в котором находятся последовательности, кодирующие другие дополняющие вирусные белки, или в отдельном векторе. Она необязательно может быть также интегрирована в продуцирующую клетку-хозяин или клеточную линию.
Дополнительным осуществлением настоящего изобретения является селективное позиционирование донорных и акцепторных сайтов сплайсинга в хелперном векторе, в результате чего селективное удаление за счет сплайсинга некоторых компонентов вируса обеспечивает сведение к минимуму или исключение возможности гомологичной рекомбинации с вектором с зависимой от условий репликацией. Например, сайты сплайсинга могли бы быть расположены так, чтобы в результате событий сплайсинга обеспечивалась делеция RRE и/или сигналов упаковки или димеризации. Следовательно, контекст того, какие нуклеиновые кислоты будут экспрессироваться, может контролироваться помещением в вектор сайтов сплайсинга. Например, сайты сплайсинга можно было бы расположить дистально по отношению к рибозимной или антисмысловой последовательности так, чтобы, например, рибозимы или антисмысловая последовательность включались в не подвергнутые сплайсингу, но не в подвергнутые сплайсингу РНК. В противоположном варианте рибозимы или антисмысловую последовательность можно было бы поместить ниже сайта сплайсинга, если бы целью являлась экспрессия молекулы рибозимов или антисмысловой последовательности со всех молекул мРНК, а не их конкретного подтипа.
Когда вектор применяют согласно описанному выше способу для профилактического лечения вирусной инфекции, вектор может кодировать антиген белка, который не кодируется вирусом дикого типа, такой как мутантный вирусный белок или невирусный белок. Соответственно, кодируемый вектором антиген может иметь, например, бактериальное происхождение или может происходить от раковых клеток. Более того, вирусный вектор с зависимой от условий репликацией или предпочтительно лентивирусный вектор может также кодировать ген MHC для правильной презентации антигена иммунной системе хозяина. Таким образом, такие векторы могут быть применены для облегчения продолжительного иммунологического ответа против широкого спектра потенциальных патогенов и/или эндогенных белков (например, специфичного для опухоли антигена), которые селективно экспрессируются в ненормальных клетках.
Приведенный выше пример не ограничивает применение изобретения предоставлением антигена дендритным клеткам для иммунотерапии. Напротив, в изобретении предлагаются средства и способы для доставки и экспрессии любого интересующего гена в любом желаемом типе клеток. Более того, изобретение также обеспечивает доставку и экспрессию многих генов посредством одного вектора. Множество генов может быть экспрессировано при применении любой подходящей стратегии генной экспрессии. Неограничивающим примером является применение внутреннего сайта вхождения в рибосому (IRES), который может быть помещен дистально по отношению к первому гену, который подлежит экспрессии. Предпочтительным примером является экспрессия интересующего гена с применением последовательности IRES, расположенной дистально по отношению к варианту гена MGMT. В более предпочтительном осуществлении вектор экспрессирует интересующий ген и соединенный с IRES интересующий ген с немодифицированного или модифицированного промотора HIV-LTR, исключаемого из подвергнутого сплайсингу мессенджера. Следующий предпочтительный вектор экспрессирует два интересующих гена с немодифицированного или модифицированного промотора HIV-LTR, такого, что если он модифицирован, то модификация касается последовательностей, которые связываются с транскрипционными факторами. Разумеется, в дополнение к LTR HIV могут быть применены любые немодифицированные или модифицированные лентивирусные LTR.
Другим неограничивающим примером является экспрессия интересующего гена с HIV-LTR с применением сайтов сплайсинга, берущих начало от HIV. Например, один интересующий ген мог бы экспрессироваться с акцепторного сайта сплайсинга Nef, а второй интересующий ген мог бы экспрессироваться с акцепторного сайта сплайсинга Tat. В данном варианте два гена могли бы экспрессироваться без необходимости в наличии элемента IRES. Мог бы быть применен любой сайт сплайсинга, включая таковые генов Tat, Rev, Vif, Vpr, Vpu, Nef и Env. Белок Env экспрессируется с подвергнутого однократному сплайсингу мессенджера, и поэтому для экспрессии интересующего гена через данный сайт сплайсинга может иметься потребность в Rev, если необходима значительная экспрессия интересующего гена. Более того, если это желательно, экспрессия с HIV-LTR может быть сделана зависимой от Tat и Rev.
Более того, может быть создан экспрессионный вектор «хелпер-вирус» (также обозначаемый здесь как «хелпер») для экспрессии лишь в специфичных типах клеток (например, стволовых клетках, профессиональных антиген-презентирующих клетках и опухолевых клетках) путем добавления или удаления специфического генетического элемента/фактора (либо в векторе, либо в экспрессионном конструкте хелпер-вирус), что обеспечивает специфичную в отношении клетки репликацию и распространение вектора. Таким образом, вектор продолжает распространяться благодаря комплементации хелпер-вирусным конструктом, но это распространение является специфичным для клеток и зависит от того, добавлен ли в вектор или в хелпер-вирусный экспрессионный конструкт или удален из них определенный генетический элемент/фактор. Это можно применять отдельно или в сочетании с другими упомянутыми выше стратегиями.
Например, может быть создан вектор HIV с зависимой от условий репликацией для специфичной репликации в макрофагах, а не в T-клетках. Вектор, который создавал бы дефектный по Tat HIV (вектор кодирует другие белки HIV, но они не экспрессируются из-за отсутствия транскрипционного трансактиватора Tat), может кодировать рибозим, который расщепляет HIV дикого типа, но не РНК HIV с зависимой от условий репликацией. Экспрессионный вектор хелпера для данного вектора может кодировать ген tat, экспрессируемый со специфичного для макрофагов промотора. Таким образом, crHIV должен был бы реплицироваться в зависимости от условий только в клетках-макрофагах и в то же время был бы неспособен реплицироваться в T-клетках или клетках других типов.
В противоположном варианте ген tat может быть оперативно соединен со специфичным для опухоли промотором; так что crHIV мог бы затем реплицироваться только в опухолевых клетках CD4, но не в нормальных клетках CD4. Генетический элемент/фактор может быть также модификацией промоторной последовательности вектора, так что он экспрессируется только в специфичных типах клеток, но не в других типах клеток вместе с «хелпер-вирусным» экспрессионным конструктом.
В другом осуществлении белки оболочки хелперного экспрессионного конструкта или векторного конструкта (если такие конструкты были разработаны для включения белков оболочки) могут быть модифицированы таким образом, что вектор-вирион будет специфично инфицировать определенные типы клеток (например, опухолевые клетки), не будучи способным инфицировать другие типы клеток (например, нормальные клетки). В еще одном осуществлении аденовирус, лишенный одного или нескольких ключевых для репликации факторов, мог бы быть дополнен применением хелперного конструкта, который обеспечивает снабжение такими факторами, связанными со специфичным для опухоли промотором. Таким образом, факторы, комплементирующие репликацию аденовируса, должны были бы экспрессироваться только в опухолевых клетках, тем самым обеспечивая вирусную репликацию в опухолевых клетках (с экспрессией белков, необходимых для гибели клеток), но не в нормальных клетках.
В другом осуществлении вектор мог бы экспрессировать ген негативной селекции для вызова гибели клеток с помощью агента, алкилирующего O6-гуанин, такого как, но не ограничиваясь этим, BCNU (или функциональный аналог BCNU) в качестве лекарства для негативной селекции. Лентивирусный вектор, экспрессирующий антисмысловую последовательность или рибозим, направленные на ген MGMT, может быть доставлен к нормальной клетке. Позже в желаемый момент клетки можно обработать BCNU или его аналогом ex vivo или in vivo для индукции гибели трансдуцированных вектором клеток. В предпочтительном осуществлении нормальные клетки профилактически трансдуцируют экспрессионным вектором с антисмысловой последовательностью MGMT и затем вызывают гибель, когда клетки становятся ненормальными. Примером данного последнего подхода является возможность индукции гибели клеток обработкой клеток BCNU или его функциональным аналогом после превращения клеток в раковые. Другим предпочтительным осуществлением является экспрессия антисмысловой последовательности против MGMT или рибозимной молекулы c промотора U1 и содержащейся в snRNA U1. Еще более предпочтительным осуществлением является трансдукция анти-MDMT лентивирусного вектора в популяцию гематопоэтических клеток, таких как лимфоциты, стволовые клетки и дендритные клетки.
В другом предпочтительном осуществлении нормальными клетками являются гематопоэтические клетки, предпочтительно T клетки, которые трансдуцируют вектором перед трансплантацией аллогенному хозяину. Гибель таких клеток может быть вызвана обработкой BCNU или его функциональным аналогом, если клетки становятся вредными для хозяина (например, происходит реакция трансплантат против хозяина). В еще одном осуществлении тот же самый лентивирусный вектор, экспрессирующий антисмысловую последовательность против MGMT или рибозим, мог бы быть применен для удаления нежелательных клеток из смешанных клеточных популяций. Неограничивающим примером является случай зараженного опухолевыми клетками костного мозга, готового к трансплантации. Было показано, что опухолевые клетки очень эффективно трансдуцируются при относительно низких MOI при одном раунде трансдукции (смотри фиг.13A). Напротив, нормальные клетки, в особенности, но не ограничиваясь этим, гематопоэтические стволовые клетки CD34+, труднее эффективно трансдуцировать, и для высокой эффективности требуется много раундов трансдукции (смотри фиг.13B). Следовательно, привлекательная стратегия очистки заключается в трансдукции зараженного костного мозга вектором, содержащим антисмысловую последовательность против MGMT, с применением MOI, который бы эффективно трансдуцировал загрязняющие опухолевые клетки, но не нормальные клетки. Данный пример не ограничивает объем изобретения применением ex vivo или связанными исключительно с раком применениями. Другие применения включают избирательную доставку гена in vivo, в особенности, но не ограничиваясь этим, для лечения рака мозга и любых других заболеваний, при которых дифференцированная между больными и нормальными клетками эффективность трансдукции может быть использована для направленного воздействия в соответствии с настоящим изобретением.
Таким образом, в настоящем изобретении также предлагается способ лечения рака и, в частности, Т-клеточного лейкоза. «Лечение рака» согласно изобретению включает введение хозяину дополнительно модифицированного вектора, как изложено здесь, с целью индукции терапевтического действия. Такое действие может быть оценено, например, с помощью отслеживания задержки опухолевого роста и/или регрессии опухоли. «Опухолевый рост» включает увеличение размера и/или количества опухолей. «Регрессия опухоли» включает снижение массы опухоли.
«Рак» согласно изобретению включает раки, которые характеризуются ненормальной клеточной пролиферацией и отсутствием контактного ингибирования, которые могут быть подтверждены образованием опухоли. Термин охватывает рак, локализованный в опухолях, а также рак, не локализованный в опухолях, такой как, например, раковые клетки, распространяющиеся из опухоли локально путем инвазии или системно путем метастазирования. Теоретически любой тип рака может быть объектом лечения согласно изобретению. Предпочтительно, однако, чтобы рак имел вирусное происхождение.
Наконец, описанные выше векторы могут быть непосредственно применены для генной терапии in vivo. Современные стратегии генной терапии ущербны из-за того, что они не могут опосредовать доставку гена к большому проценту клеток; лишь определенный процент клеток подвергается инфицированию. Это особенно важно в противоопухолевых стратегиях, когда трансдукция гена всей опухолевой популяции является решающей. Путем добавления «вектора» вместе с «хелпером» непосредственно трансдуцированные клетки будут продуцировать вирусные частицы, которые могут инфицировать соседние клетки и, таким образом, способны к высокой и, возможно, полной эффективности трансдукции. В одном осуществлении данного изобретения ретровирус человека (которым мог бы быть HIV или ретротранспозонный элемент) мог бы доставляться в ткань (или клетки in vitro) с помощью «хелперного» конструкта. Клетки, непосредственно содержащие вектор и хелпер, будут продуцировать вирус и будут зависимо от условий упаковывать вектор в вирионы. Данные вирионы будут способны опосредовать высокую эффективность трансдукции соседних клеток (поскольку межклеточные контакты являются наиболее эффективным путем трансдукции клеток). Непосредственно трансдуцированные клетки могут погибать или не погибать, в зависимости от того, приводит ли сочетание вектор/хелпер к цитолитической инфекции. В случае ретротранспозона может не иметься необходимости в содержании в хелпере структурных белков, поскольку нормальные или опухолевые клетки могут содержать белок/фактор, необходимый для инкапсуляции в вирионы. В данном случае хелпер может быть, но не ограничиваясь этим, лишь трансактиваторным белком, который активирует транскрипцию факторов, необходимых для инкапсулирования ретротранспозона. В случае HIV для инкапсулирования генома HIV в вирионы потомства для инфицирования/трансдукции клеток могут, но не обязательно, требоваться другие факторы.
Описанные выше векторы могут быть применены для биологической и химической защиты в военных целях. Например, вектор с зависимой от условий репликацией может быть введен индивидууму, недавно инфицированному высоко патогенным вирусом, или бактерией, или химическим агентом (например, токсином). Вектор должен препятствовать репликации патогенного вируса, как описано ранее. Однако вектор с зависимой от условий репликацией может быть также применен в целях бактериальной и химической защиты совместно с хелперным экспрессионным вектором («хелпером»).
Например, вектор с зависимой от условий репликацией может секретировать антибактериальные или противотоксинные антитела после обеспечения «хелпером» его экспрессии и размножения. «Хелпер» может, но не обязательно, управляться индуцибельным промотором, который обеспечивает его экспрессию при активации бактерией, цитокином (в ответ на бактериальную инфекцию), антибиотиком (как в случае индуцируемых тетрациклином промоторных систем (Paulus et al., J. Virol., 70, 62-67 (1996)) или химическим агентом (например, самим токсином). Таким образом, вектор с зависимой от условий репликацией будет не только селективно размножаться с помощью «хелпера» в ответ на вторжение патогена или токсина (в результате активации хелпера), но и секретировать противопатогенные или противотоксинные антитела для торможения патологических эффектов опухолевого антигена, патогена или химического агента (например, токсина). Таким образом, любой белок, фактор или генетический элемент, который может быть транскрибирован в мРНК и/или белок, может быть вставлен в вектор с зависимой от условий репликацией для торможения патогенного действия - совместно с «хелпером», который способствует его селективному размножению и экспрессии (селективному благодаря тому, что продукты хелпера экспрессируются в зависимости от условий (например, но не ограничиваясь этим, (a) индуцибельная промоторная система - фактор опухолевых клеток активирует продукцию хелперного фактора, чувствительной к токсину последовательности, которая экспрессирует хелперный фактор, или цитокинчувствительный промотор индуцирует продукцию хелперного фактора, (b) хелперные РНК/белок/фактор селективно стабилизируются в одних, но не в других клетках) и (c) непрямая индукция гена третьей стороны, который влияет на хелперный вирусный белок через его продукцию, сопровождение, направление, структуру или другую биологическую функцию). Такие стратегии могут быть применены у трансгенных растений и животных для их защиты от патогенов. Сходным образом такие стратегии могут быть применены в трансгенных системах для получения ценных гетерологичных белков/факторов.
В другом осуществлении способа согласно настоящему изобретению может быть создана клеточная линия для скрининга лекарства/фактора для определения, например, того, какая часть белка/фактора является важной для конкретной функции. Может быть создан вектор для экспрессии интересующего мутированного белка в данной клеточной линии. РНК, кодирующую мутированный белок, однако, делают устойчивой к рибозиму путем, например, вставки молчащих точечных мутаций. В то же время экспрессия белка дикого типа в клеточной линии ингибируется. Для экспрессии мутантного тестируемого белка могут быть также сконструированы векторы, которые экспрессируют рибозим против интересующего белка. Когда вектор трансдуцируют в клетки, большая часть природной РНК, кодирующей нормальный белок, расщепляется, тогда как мутантный тестируемый белок экспрессируется. Данный способ может быть применен с использованием недавно разработанных способов доставки и селекции в качестве быстрого и действенного способа определения того, как функционирует данный белок и как данный фактор/лекарство взаимодействует с белком.
В еще одном осуществлении для введения вектора для лечения врожденных заболеваний крови больного подвергают лейкаферезу для извлечения из крови достаточного количества лейкоцитов. После лейкафереза выделяют желаемые клетки, трансдуцируют вектором и размножают в культуре до желаемого количества клеток. Во время размножения желаемого типа клеток ex vivo больному вводят антитела, направленные против белка клеточной поверхности клеток желаемого типа, с целью разрушения данных клеток в период времени, совпадающий с размножением достаточного количества клеток желаемого типа. Трансдуцированные клетки затем обратно вводят больному для компенсации вызванной антителом in vivo утраты клеток.
Предпочтительным средством реализации указанного выше является выделение T-клеток CD4+ из лейкаферезного материала с последующей трансдукцией клеток вектором HIV, который содержит антисмысловую или рибозимную последовательность против HIV, для размножения ex vivo. Во время размножения клеток ex vivo эндогенные T-клетки CD4+ больного разрушают введением, например, антитимоцитного глобулина (ATG) (Pasteur Merieux Serums et Vaccins, Lyon, Франция, распространяемого SangStat Medical Corporation, Menlo Park, CA, США) или Atgam (Upjohn Company, Kalamazoo, MI, США), однако могло бы быть применено любое снижающее количество клеток антитело после подходящего скрининга, который должен быть очевиден для специалиста в данной области техники. После размножения трансдуцированные T-клетки CD4+ затем обратно вводят больному. В предпочтительном осуществлении CD4-негативные клетки, полученные во время выделения CD4-позитивных клеток, сохраняют, замораживают и размораживают для введения в момент введения назад больному трансдуцированных CD4-позитивных клеток.
Приведенный выше пример не ограничивает объем изобретения ни ATG, ни T-клетками CD4+. Любой тип клеток может быть направленно поражен путем применения антитела, которое связывается с белком клеточной поверхности указанной клетки и является цитоцидным. Антитело могло бы быть введено экзогенно больному, или в организм мог бы быть введен второй вектор, секретирующий антитело. Данный вектор мог бы продуцировать антитело конститутивно (на протяжении жизни клетки), временно (например, но не ограничиваясь этим, с вектором, лишенным интегразы) или индуцибельно (например, но не ограничиваясь этим, с индуцируемой тетрациклином промоторной системой).
Существует также множество применений способа и векторов настоящего изобретения in vitro. Например, векторы можно применять для выяснения некоторых нюансов репликации вирусов и рибозимной функции. Сходным образом, содержащие рибозим векторы можно применять в качестве средств диагностики, например, для оценки мутаций, имеющихся в больных клетках, или для проверки генетического дрейфа. Векторы могут быть также применены для определения или подтверждения функции гена в зависимости от того, была ли эта функция известна ранее, является известной или лишь предполагается или допускается. Данное вышеизложенное обсуждение никоим образом не является исчерпывающим в отношении применения настоящего изобретения.
Преимущества изобретения
Преимущества применения стратегии crHIV для генетического терапевтического лечения СПИД и других заболеваний являются значительными. Например, становится решенной проблема направления вектора к клеткам, инфицированным HIV. После трансфекции crHIVs in vivo в инфицированные клетки CD4+ crHIV упаковываются в вирионы потомства с использованием эндогенных белков оболочки инфекционного HIV. Таким образом, РНК crHIV переносится внутри вирионов потомства и инфицирует типы клеток, в норме инфицируемые данным конкретным штаммом HIV, с образованием непатогенных вирионов. Это включает затруднение с направленным лечением клеток, таких как микроглия мозга, которые являются главным резервуаром для инфекции HIV (ВИЧ) в центральной нервной системе. По-видимому, с векторами crHIV, которые инфицируют неинфицированные клетки CD4+, связана слабая токсичность, поскольку вирусные белки не кодируются векторами crHIV. Более того, за счет конкуренции вектора crHIV с HIV дикого типа образуются непатогенные частицы, что приводит к снижению вирусного заражения. Снижение заражения патогенным HIV-1 не только повышает время выживания инфицированных индивидуумов, но и снижает скорость заражения неинфицированных индивидуумов, поскольку частицы crHIV также могут распространяться системно (т.е. как и инфекционный HIV). Сниженное заражение патогенным HIV-1 крови может быть особенно важным для беременных инфицированных HIV (ВИЧ) индивидуумов, поскольку образование crHIV также может снижать передачу HIV-1 от инфицированных матерей их плодам in utero.
Смесь плазмидная ДНК/липид, которая может быть применена для введения вектора crHIV в клетку-хозяин, должна быть стабильной и дешевой для получения в обход дорогих стратегий ex vivo. Разумеется, способу изобретения присуща врожденная гибкость, поскольку он мог бы быть также применен для доставки гена ex vivo, если это было бы желательно. Независимо от этого доступность опосредуемого липосомами подхода открывает возможность лечения всей популяции - того, что нереально с современными стратегиями генной терапии. Могут быть также созданы векторы crHIV, которые содержат несколько рибозимов, которые могут быть изготовлены против разных мишеней в геноме HIV. Это снижает возможность мутации и ускользания инфекционного HIV из-под действия рибозимов против HIV. Более того, стратегия вирусов, способных к зависимой от условий репликации, может быть применена для лечения других вирусных инфекций, в особенности таких, при которых оборот вируса является высоким.
Особенно полезной особенностью векторов crHIV является то, что их можно применять для экспрессии генетических противовирусных агентов, например рибозима, действующих посттранскрипционно. Так, инфекция векторами crHIV неинфицированных клеток сопровождается низкой токсичностью из-за слабой экспрессии с промотора длинного концевого повтора (LTR) HIV в отсутствие белка Tat. Высокий уровень экспрессии crHIV и его последующей противовирусной активности имеют место только тогда, когда белок Tat обеспечивается комплементацией HIV дикого типа. Таким образом, векторы crHIV предназначены не для зашиты клеток от инфекции HIV, а для снижения общей нагрузки вирусом HIV дикого типа за счет селективной аккумуляции непатогенных частиц crHIV.
Не желая быть связанными какой-либо конкретной теорией, касающейся действия или функционирования изобретения, заявители полагают, что, как это подтверждено в следующих примерах, рибозимы могут быть применены для придания геномам crHIV селективного преимущества за счет двух полезных свойств: (1) они обладают высокой степенью специфичности, и (2) они обладают относительной эффективностью, которая зависит от их способности к совместной локализации с РНК-мишенями (Cech, Science, 236, 1532-1539 (1987)). Специфичность рибозимов подтверждается их специфичной гибридизацией с комплементарными последовательностями-мишенями, содержащими сайт XUY. Рибозимы относительно эффективны, поскольку они расщепляют РНК-мишени с высокой эффективностью лишь тогда, когда они эффективно локализуются совместно с РНК-мишенями. При смешанной инфекции HIV/crHIV должна иметь место совместная локализация содержащей рибозим РНК crHIV и РНК HIV дикого типа, поскольку геномы РНК HIV димеризуются перед упаковкой в вирионы потомства. Расщепление негеномных форм РНК HIV дикого типа, необходимое для продукции вирусных белков, по-видимому, происходит менее эффективно по сравнению с геномными РНК HIV дикого типа, поскольку негеномные РНК HIV не димеризуются. В описанных здесь экспериментах было обнаружено, что сообщаемое РНК crHIV селективное преимущество обусловлено селективной упаковкой crHIVs в вирусные частицы. Данные результаты позволяют предполагать, что наиболее эффективное расщепление происходит внутриклеточно во время димеризации, что приводит к селективному разрушению РНК HIV дикого типа нуклеазами хозяина. Это обеспечивает преимущественную упаковку РНК crHIV в вирусные частицы.
Применение векторов crHIV для терапии HIV (ВИЧ) может включать не только геномную селекцию crHIV, но и клеточную селекцию клеток, продуцирующих частицы crHIV. В противном случае клетки, продуцирующие HIV дикого типа, продуцировали бы частицы HIV дикого типа с селективным преимуществом по сравнению с клетками, продуцирующими частицы crHIV, и быстро становились бы преобладающими. Селективное преимущество может быть сообщено экспрессирующим crHIV клеткам путем вставки в геномы crHIV гена (например, гена устойчивости ко многим лекарствам), который придает экспрессирующим crHIV клеткам (в присутствии лекарства) преимущество в выживании по сравнению с клетками, экспрессирующими HIV дикого типа. В данных условиях клетки, экспрессирующие HIV дикого типа, постепенно гибнут, но все еще продуцируют некоторое количество HIV дикого типа, а клетки, экспрессирующие crHIV, которые селективно продуцируют crHIV, выживают. Инфицирование клеток, содержащих crHIV, остающимся HIV дикого типа должно приводить к дополнительной продукции вирусных частиц, содержащих crHIV. Таким образом, вирусный геномный сдвиг может приводить к кумулятивному инфицированию клеток CD4+геномами crHIV, что тем самым смещает вирусный баланс у хозяина от патогенного HIV дикого типа к непатогенным геномам crHIV. Такая стратегия может приводить к удалению HIV дикого типа, поскольку баланс между геномами HIV селективно смещается от HIV дикого типа в сторону crHIV. Вирусная репликация постепенно прекращается, поскольку crHIV может реплицироваться только в присутствии геномов хелперного HIV дикого типа. Следовательно, при таких взаимно ограничивающих условиях возможно создание векторов crHIV, которые не только снижают уровень вирусного заражения HIV дикого типа, но и удаляют вирус из инфицированного HIV хозяина.
Средства получения
Вектор может быть получен с помощью процесса комплементации либо путем применения временной трансфекции геномов вектора и хелпера в клеточную линию, предпочтительно, но не ограничиваясь этим, в хорошо известную клеточную линию 293T; или путем применения клеточной линии для упаковки. Предпочтительно клетку временно трансфицируют при высокой эффективности трансфекции с применением реагента трансфекции, предпочтительно фосфата кальция, электропорации или липидного реагента трансфекции. После того как трансфекция произошла, вектор собирают из супернатанта, предпочтительно, но не ранее чем через 12 часов после трансфекции и не более чем через 7 дней после трансфекции. Вектор может быть необязательно по желанию сконцентрирован высокоскоростным центрифугированием без осаждения или ультрацентрифугированием. Предпочтительными условиями центрифугирования являются приблизительно 5000-12000g, более предпочтительно приблизительно 10000g. Более предпочтительными условиями являются центрифугирование в течение ночи при 4°С.
Способы очистки вектора
Способ 1
1. Осветление вирусного супернатанта
2. Концентрирование ультрафильтрацией
3. Диафильтрация с обработкой или без обработки бензоназой
4. Ионообменная хроматография
5. Хроматография с исключением по размеру или диафильтрация
6. Необязательно концентрирование ультрафильтрацией
Способ 2
1. Осветление вирусного супернатанта
2. Ионообменная хроматография
3. Диафильтрация или хроматография с исключением по размеру
4. Необязательная обработка бензоназой
5. Хроматография с исключением по размеру или диафильтрация
6. Необязательно концентрирование ультрафильтрацией
Для указанной выше процедуры могут быть применены различные смолы, включая Poros 50HQ от Perseptive Biosystems. Для ультрафильтрации или диафильтрации могут быть применены картриджи из полых волокон, включая картриджи (серия UFP-750) от A/G Technologies.
Средства введения
Согласно изобретению вектор вводят в клетку-хозяин, нуждающуюся в генной терапии вирусной инфекции, как описано ранее. Средства введения включают контактирование хозяина, способного быть инфицированным, с вирусным вектором согласно изобретению. Предпочтительно, чтобы такое контактирование включало любые средства, с помощью которых вектор вводится в клетку-хозяин; способ не зависит от любого конкретного средства введения и не нуждается в таком толковании. Средства введения хорошо известны специалистам в данной области техники и также проиллюстрированы здесь.
Соответственно, введение может быть осуществлено, например, либо in vitro (например, в способе генной терапии типа ex vivo), либо in vivo, которое включает применение электропорации, трансформации, трансдукции, конъюгации или трехродительского спаривания, трансфекции, инфицирования, слияния мембран с катионными липидами, высокоскоростной бомбардировки покрытыми ДНК микроснарядами, инкубации с преципитатом фосфат кальция-ДНК, прямой микроинъекции в одиночную клетку и тому подобного. Другие способы также доступны и известны специалистам в данной области техники.
Предпочтительно, однако, чтобы векторы или рибозимы вводились с помощью катионных липидов, например липосом. Такие липосомы имеются в продаже (например, Lipofectin™, Lipofectamine™ и тому подобное, предоставляемые Life Technologies, Gibco BRL, Gaithersburg, Md). Более того, в настоящем изобретении могут быть использованы липосомы с повышенной переносящей способностью и/или сниженной токсичностью in vivo (например, как рассмотрено в обзоре в патентной заявке PCT no. WO 95/21259). Для введения липосом можно следовать рекомендациям, указанным в патентной заявке PCT no. WO 93/23569. В целом при таком введении состав захватывается большей частью лимфоцитов в течение 8 ч при 37°С с обнаружением более 50% введенной дозы в селезенке через час после внутривенного введения. Сходным образом другие средства доставки включают гидрогели и полимеры с контролируемым высвобождением.
Форма вектора, вводимого в клетку-хозяин, может варьировать в зависимости частично от того, подлежит ли вектор введению in vitro или in vivo. Например, нуклеиновая кислота может быть сомкнута в кольцо, разрезана или линеризована в зависимости от того, подлежит ли вектор внегеномному поддержанию (т.е. в виде автономно реплицируемого вектора), интеграции в виде провируса или профага, временной трансфекции, временной инфекции, как при применении вирусов с дефектной репликацией или с зависимой от условий репликацией, или стабильному включению в геном хозяина посредством событий двойной или одиночной рекомбинации кроссинговера.
Перед введением хозяину вектор настоящего изобретения может быть составлен в различные композиции для применения в способах терапевтического и профилактического лечения. В частности, вектор может быть выполнен в виде фармацевтической композиции путем сочетания с подходящими фармацевтически приемлемыми носителями или разбавителями и может быть составлен для того, чтобы быть приемлемым для применений у человека или в ветеринарии.
Так, композиция для применения в способе настоящего изобретения может включать один или более упомянутых выше векторов, предпочтительно в сочетании с фармацевтически приемлемым носителем. Фармацевтически приемлемые носители хорошо известны специалистам в данной области техники, так же как и подходящие способы введения. Выбор носителя должен определяться отчасти конкретным вектором, а также конкретным способом, применяемым для введения композиции. Специалист в данной области техники также должен понимать, что доступны разные пути введения композиции, и, хотя для введения могут быть применены более одного пути, определенный путь может обеспечить более быструю и более эффективную реакцию по сравнению с другим путем. Соответственно, существует множество подходящих составов композиции настоящего изобретения.
Композиция, составленная из вектора настоящего изобретения, отдельно или в сочетании с другими противовирусными соединениями, может быть выполнена в виде состава, пригодного для парентерального введения, предпочтительно внутрибрюшинного введения. Такая композиция может включать водные и неводные изотоничные стерильные растворы для инъекции, которые могут содержать антиоксиданты, буферы, бактериостатики и растворы, которые придают композиции изотоничность с кровью подлежащего лечению реципиента, и водные или неводные стерильные суспензии, которые могут включать суспендирующие агенты, солюбилизаторы, загустители, стабилизаторы и консерванты. Композиции могут быть представлены в форме единицы дозы или множества доз в запаянных контейнерах, таких как ампулы или флаконы, и могут храниться в лиофилизованной форме, для которой требуется лишь добавление стерильного жидкого носителя, например воды для инъекций, непосредственно перед применением. Приготовленные для немедленного применения растворы и суспензии для инъекций могут быть получены из стерильных порошков, гранул и таблеток, как описано здесь.
Вектор может храниться в любом подходящем растворе, буфере или в лиофилизованной форме, если это желательно. Предпочтительным буфером для хранения является забуференный фосфатом солевой раствор Дульбекко; забуференный фосфатом солевой раствор Дульбекко, смешанный с 1-50% раствором трегалозы в воде (1:1), предпочтительно 10% раствором трегалозы в воде (1:1), так что конечная концентрация трегалозы равна 5%; забуференный фосфатом солевой раствор Дульбекко, смешанный с 1-50% раствором глюкозы в воде (1:1), предпочтительно 10% раствором глюкозы в воде (1:1), так что конечная концентрация глюкозы равна 5%; солевой раствор, забуференный 20 мМ HEPES, смешанный с 1-50% раствором трегалозы в воде (1:1), предпочтительно 10% раствором трегалозы в воде (1:1), так что конечная концентрация трегалозы равна 5%; забуференный фосфатом солевой раствор Дульбекко, смешанный с 1-50% раствором маннита в воде (1:1), предпочтительно 5% раствором маннита в воде (1:1), так что конечная концентрация маннита равна 2,5%.
Композиция, пригодная для перорального введения, может состоять из жидких растворов, таких как эффективное количество соединения, растворенного в разбавителях, таких как вода, солевой раствор или фруктовый сок; капсул, пакетиков или таблеток, каждый из которых содержит заранее определенное количество активного ингредиента в виде твердого вещества или гранул; растворов или суспензий в водной жидкости и эмульсий масла в воде или эмульсий воды в масле. Таблетированные формы могут включать одно или более из лактозы, маннита, кукурузного крахмала, картофельного крахмала, микрокристаллической целлюлозы, камеди, желатина, коллоидного диоксида кремния, кроскармеллозы натрия, талька, стеарата магния, стеариновой кислоты и других наполнителей, красителей, разбавителей, забуферивающих агентов, увлажняющих агентов, консервантов, корригентов и фармакологически совместимых носителей.
Может быть также изготовлена аэрозольная композиция, пригодная для введения посредством ингаляции. Аэрозольная композиция может быть помещена в приемлемый сжатый движущий газ, такой как дихлордифторметан, пропан, азот и тому подобное.
Сходным образом композиция, пригодная для перорального введения, может включать форму лепешек, которые могут включать активный ингредиент в сластях, обычно из сахарозы и камеди или трагаканта; пастилок, включающих активный ингредиент в инертной основе, такой как желатин, или глицерин, или сахароза и камедь; и полосканий для рта, включающих активный ингредиент в подходящем жидком носителе; а также в виде кремов, эмульсий, гелей и тому подобного, содержащих в дополнение к активному ингредиенту такие носители, которые известны в данной области техники.
Композиции, пригодные для местного нанесения, могут находиться в форме кремов, мазей или лосьонов.
Композиция для ректального введения может быть представлена в форме суппозитория с подходящей основой, например маслом какао или салицилатом. Композиция, подходящая для влагалищного введения, может быть представлена в виде пессария, тампона, крема, геля, пасты, пены или спрея, содержащих в дополнение к активному ингредиенту такие носители, которые, как известно в данной области техники, являются подходящими. Сходным образом активный ингредиент может быть объединен со смазывающим агентом в качестве покрытия презерватива.
Доза, вводимая животному, в особенности человеку, в контексте настоящего изобретения должна быть достаточной для оказания терапевтического действия у инфицированного индивидуума в рамках разумного периода времени. Доза должна определяться эффективностью конкретного вектора, применяемого для лечения, тяжести заболевания, а также массой тела и возрастом инфицированного индивидуума. Размер дозы должен также определяться наличием каких-либо побочных эффектов, которые могут сопровождать применение конкретного используемого вектора. Всегда желательно при любой возможности сводить нежелательные побочные эффекты к минимуму.
Доза может находиться в виде единицы лекарственной формы, такой как таблетка или капсула. Применяемый здесь термин «единица лекарственной формы» относится к физически дискретным единицам, подходящим в качестве единиц доз для субъектов, являющихся человеком и животным, причем каждая единица содержит предварительно определенное количество вектора, одного или в сочетании с другими противовирусными агентами, рассчитанное в виде количества, достаточного для получения желаемого эффекта, в сочетании с фармакологически приемлемым разбавителем, носителем или наполнителем. Характерные особенности единиц лекарственных форм настоящего изобретения зависят от конкретного применяемого соединения или соединений и эффекта, который должен быть достигнут, а также от фармакодинамики каждого соединения у хозяина. Вводимая доза должна быть «эффективным против вируса количеством» или количеством, необходимым для достижения «эффективного уровня» у отдельного больного.
Так как «эффективный уровень» применяют в качестве предпочтительного предела дозирования, действительная доза и схема могут варьировать в зависимости от межиндивидуальных особенностей фармакокинетики, распределения лекарства и метаболизма. «Эффективный уровень» может быть определен, например, как желательный уровень в крови или ткани больного, который соответствует концентрации одного или более вектора(векторов) согласно изобретению, которая ингибирует вирус, такой как HIV, в тесте прогноза клинической противовирусной активности химических соединений. «Эффективный уровень» для соединений настоящего изобретения может также варьировать, когда композиции настоящего изобретения применяют в сочетании с зидовудином или другими известными противовирусными соединениями или их сочетаниями.
Специалист в данной области техники может легко определить подходящую дозу, схему и способ введения для применяемого конкретного состава композиции для достижения желаемого «эффективного уровня» у отдельного больного. Специалист в данной области техники также может легко определить и использовать подходящий индикатор «эффективного уровня» соединений настоящего изобретения с помощью прямого (например, аналитического химического анализа) или непрямого (например, с суррогатными индикаторами вирусной инфекции, такими как p24 или обратная транскриптаза для лечения СПИДа или подобного СПИДу заболевания) анализа подходящих образцов больного (например, крови и/или ткани).
Затем в отношении определения эффективного уровня у больного для лечения СПИДа или подобного СПИДу заболевания, в частности, существуют подходящие животные модели, и они широко применяются для оценки эффективности in vivo различных протоколов генной терапии против HIV (Sarver et al. (1993b), см. выше). Данные модели включают мышей, обезьян и кошек. Даже хотя данные животные по своей природе не восприимчивы к инфекции HIV, химерные мышиные модели (например, SCID, bg/nu/xid, NOD/SCID, SCID-hu, иммунокомпетентные SCID-hu, BALB/c с удаленным костным мозгом), воссозданные с мононуклеарными клетками периферической крови человека (PBMCs), лимфатическими узлами, фетальной печенью/тимусом или другими тканями, могут быть инфицированы вектором или HIV и применены в качестве моделей патогенеза HIV и генной терапии. Сходным образом может быть применена модель вирус иммунодефицита обезьян (SIV)/обезьяна, так же как и модель вирус иммунодефицита кошачьих (FIV)/кошка.
Данные модели могут быть также использованы для определения безопасности вектора с целью подтверждения возможности применения векторной системы в клинических испытаниях. Важным применением является использование данных животных моделей для исследований биологического распределения. Трансдуцированные клетки, предпочтительно человека, но не ограничиваясь этим, содержащие вектор, вводят животному-модели, не являющемуся человеком, и безопасность вектора определяют по отсутствию генетического материала вектора в ткани животного. Отсутствие генетического материала вектора в ткани животного должно означать, что вектор не реплицируется автономно в отсутствие хелпера или вируса дикого типа и, таким образом, должен рассматриваться как безопасный для клинического применения у человека. Присутствие вектора в отсутствие хелперного вируса или хелперного вектора может рассматриваться как угроза безопасности, если не ожидается, что вектор будет реплицироваться автономно. Однако в том случае, когда ожидается, что векторы будут реплицироваться автономно, необходимо установить другие критерии безопасности, например отсутствие репликации в определенных тканях или уровень репликации у животного. Присутствие или отсутствие вектора может быть определено с помощью ПЦР или FASC-анализом, если тестируемый вектор экспрессирует маркерный ген, который может быть визуализован с помощью FASC, но не ограничивается такими способами определения.
Обычно предпочтительно количество вектора, достаточное для достижения концентрации в ткани вводимого рибозима (или вектора), от приблизительно 5 мкг/кг до приблизительно 300 мг/кг массы тела в день, особенно от приблизительно 10 мкг/кг до приблизительно 200 мг/кг массы тела в день. В определенных применениях, например местном, глазном или вагинальном применениях, предпочтительны множественные дневные дозы. Более того, количество доз должно варьироваться в зависимости от средств доставки и конкретного вводимого вектора.
При лечении некоторых индивидуумов, инфицированных вирусом, может быть желательным применение режима «мега-дозировки», при которой вводится большая доза вектора, дается время, чтобы соединение подействовало, и затем индивидууму вводится подходящий реагент для инактивации активного(ных) соединения(ний). В способе настоящего изобретения лечение (т.е. репликацию вектора, конкурирующую с таковой вируса, от которого лечат) необходимо ограничить. Другими словами, как только уровень, например, HIV снижается, уровень вектора, зависимого от HIV с целью продукции вирионов, должен также снижаться.
Фармацевтическая композиция может содержать также другие фармацевтические компоненты в сочетании с вектором в соответствии с изобретением при применении для терапевтического лечения СПИДа. Данные другие фармацевтические компоненты могут быть применены в их традиционном виде (т.е. в качестве агентов для лечения HIV инфекции), так же как более предпочтительно в способе отбора вирусов crHIV in vivo. Такой отбор, как здесь описано, должен стимулировать распространение HIV с зависимой от условий репликацией и позволять HIV с зависимой от условий репликацией более эффективно конкурировать с HIV дикого типа, что будет неизбежно ограничивать патогенность HIV дикого типа. В частности, предполагается применение антиретровирусного агента, такого как предпочтительно зидовудин. Дополнительные репрезентативные примеры данных дополнительных фармацевтических агентов, которые могут быть применены в дополнение к описанным ранее, включают противовирусные соединения, иммуномодуляторы, иммуностимуляторы, антибиотики и другие агенты и режимы терапии (включающие те, которые известны как альтернативная медицина), которые могут применяться для лечения СПИДа. Противовирусные компоненты включают, но не ограничиваются этим, ddI, ddC, ганцикловир, фторированные дидезоксинуклеотиды, ненуклеозидные аналоги соединений, такие как невирапин (Shih et al., PNAS, 88, 9878-9882 (1991)), производные TIBO, такие как R82913 (White et al., Antiviral Research, 16, 257-266 (1991)) и BI-RJ-70 (Shih et al., Am. J. Med., 90 (Suppl. 4A), 8S-17S (1991)). Иммуномодуляторы и иммуностимуляторы включают, но не ограничиваются этим, различные интерлейкины, CD4, цитокины, препараты антител, трансфузии крови и клеточные трансфузии. Антибиотики включают, но не ограничиваются этим, противогрибковые агенты, антибактериальные агенты и агенты против Pneumocystis carinii.
Введение ингибирующего вирусы соединения с другими противоретровирусными агентами и особенно с известными ингибиторами ОТ, такими как ddC, зидовудин, ddI, ddA, или с другими ингибиторами, которые действуют против других белков HIV, такими как агенты против TAT, должно обычно ингибировать большинство или все репликативные стадии жизненного цикла вируса. Дозировки ddC и зидовудина, применяемые у больных СПИДом или заболеванием, ассоциированным со СПИДом (ARC), опубликованы. Статический для вируса диапазон ddC находится обычно между 0,05 мкМ и 1,0 мкМ. Диапазон в приблизительно 0,005-0,25 мг/кг массы тела является статическим для вирусов у большинства больных. Диапазоны доз для перорального введения являются несколько более широкими, например от 0,001 до 0,25 мг/кг, при приеме в виде одной или более доз с интервалами в 2, 4, 6, 8 и 12 и т.д. час. Предпочтительно, чтобы дозу ddC в 0,01 мг/кг массы тела давали каждые 8 ч. При приеме в виде комбинированной терапии другое противовирусное соединение, например, может быть дано в то же самое время, что и вектор в соответствии с изобретением, или дозирование может по желанию разнесено. Вектор также может сочетаться в композиции. Дозы каждого могут быть меньшими при применении в комбинации, чем когда каждый агент применяют по отдельности.
Примеры
Соединения и способы настоящего изобретения далее описываются в контексте следующих примеров. Данные примеры служат для дополнительной иллюстрации настоящего изобретения и не предназначены для ограничения объема изобретения.
Пример 1
В данном примере описывается конструкция векторов, способных к зависимой от условий репликации, в соответствии с изобретением. В частности, в данном примере описывается конструкция векторов с зависимой от условий репликацией на основе HIV (ВИЧ), т.е. векторы crHIV.
Одним из наиболее выраженных аспектов патогенеза ВИЧ-1 является образование генетических вариантов вируса. Быстрое образование вариантов ВИЧ in vivo указывает на то, что вирус можно рассматривать в рамках Дарвиновской генетической модели (смотри, например, Coffin, Curr. Top.Microbiol. Immunol., 176, 143-164 (1992); и Coffin, Science, 267, 483-489 (1995)). Варианты возникают в результате неточности работы молекулы обратной транскриптазы HIV-1, которая создает мутации в новых провирусах при транскрипции вирусной геномной РНК. Таким образом, в условиях in vivo при отсутствии существенных ограничений и многочисленных циклах репликации имеют место значительные генетические изменения с образованием многих вариантов вируса. При этом, однако, все еще преобладает HIV дикого типа, поскольку при таких неограничивающих условиях он обладает наибольшим селективным преимуществом. Однако в присутствии ингибитора, например зидовудина, будет отбираться вариант вируса, обладающий большим селективным преимуществом по сравнению со штаммом дикого типа, и вследствие этого он будет преобладать (Coffin (1992) и (1995), выше). На этой основе в настоящем изобретении предлагается стратегия вирусного вектора с зависимой от условий репликацией, которая дает непатогенные геномы HIV-1 с селективным преимуществом по сравнению с патогенным HIV-1 дикого типа.
Данные непатогенные векторы HIV (crHIV) с зависимой от условий репликацией представляют собой дефектные HIV, которые подвергаются репликации и упаковке только в клетках, инфицированных HIV дикого типа. Геномы crHIV конкурируют с патогенным HIV дикого типа и снижают вызываемое им заражение. Эффект снижения заражения HIV дикого типа у инфицированного хозяина должен вести к повышению вероятности сохранения жизни. Это должно также снижать способность инфицированных хозяев передавать HIV дикого типа неинфицированным индивидуумам. Для успешной конкуренции crHIV с HIV-1 дикого типа, по-видимому, важны два фактора: (1) селективное преимущество геномов crHIV по сравнению с геномами HIV дикого типа, и (2) селективное преимущество клеток, экспрессирующих crHIV, по сравнению с клетками, экспрессируюшими HIV дикого типа, (т.е. селективное преимущество в отношении продукции вирионов crHIV из клеток, экспрессирующих crHIV, по сравнению с клетками, экспрессируюшими HIV дикого типа).
Векторы crHIV реплицируются в зависимости от условий благодаря тому, что они содержат последовательности, необходимые для экспрессии, димеризации и упаковки РНК, но не экспрессируют функциональные (т.е. дикого типа) белки HIV-1. Селективное преимущество было придано векторам crHIV путем вставки рибозимной кассеты, которая вызывает расщепление в области U5 генома HIV дикого типа, но не U5 РНК crHIV.
Имеющиеся в векторах рибозимы не расщепляют РНК crHIV, поскольку область U5 РНК crHIV была модифицирована консервативной заменой оснований (замены оснований, имеющиеся в других штаммах HIV) для предотвращения эффективного связывания и расщепления рибозимами по данным сайтам. Более того, crHIVs являются непатогенными, поскольку они не кодируют белки, которые, как полагают, ответственны за гибель клеток CD4+. Когда инфицированные HIV клетки (предварительно трансфицированные вектором crHIV) активируются, клетки становятся способными комплементировать недостатки генома crHIV, что приводит к образованию вирионного потомства crHIV.
В целом, геномы crHIV конструировали из полноразмерного инфекционного клона HIV, pNL4-3 (Adachi et al. (1986), см. выше). Все реакции клонирования и манипуляции с ДНК, РНК и белками производили с применением способов, хорошо известных специалистам в данной области техники и которые были описаны, например, в Maniatis et al., Molecular cloning: A Laboratory Manual, 2nd ed. (Cold Spring Harbor Laboratory, NY (1982)). Ферменты и реагенты, применяемые в данных реакциях, были получены от торговых поставщиков (например, New England Biolabs, Inc., Beverly, Mass.; Clontech, Palo Alto, Calif. и Boehringer Mannheim, Inc., Indianapolis, Ind.) и применялись в соответствии с рекомендациями производителей. Кроме того, поддержание и размножение векторов осуществляли с применением широко известных способов, описанных ранее (например, Dropulic' et al. (1992), см. выше, и Dropulic' et al. (1993), см. выше).
PNL4-3 расщепляли с помощью ферментов Pst I (который разрезает в области gag, в положении приблизительно +1000 от старта транскрипции) и Xho I (который разрезает в области nef, в положении приблизительно +8400 от старта транскрипции) и вставляли полилинкер, содержащий подходящие сайты рестрикции. Фрагмент размером 0,86 т.п.н. от Bgl II до Bam HI, содержащий rev-чувствительный элемент (RRE), клонировали в имеющийся в полилинкере Bam HI-сайт. Данные манипуляции привели к делеции в геноме HIV дикого типа, включающей фрагмент от кодирующей gag области до области кодирующей U3 (т.е., таким образом, с делецией и гена nef). Хотя вектор способен продуцировать укороченный транскрипт gag, вектор не продуцирует полноразмерный функциональный белок Gag. Однако поскольку функции Gag дикого типа не являются необходимыми согласно изобретению, последовательности gag могут быть подвержены мутациям для предотвращения трансляции белка Gag.
Рибозимную кассету, содержащую либо один, либо множество рибозимов, как описано здесь, вставляли в Sal I-сайт ниже Bam HI сайта. Для этого синтезировали комплементарные дезоксиолигонуклеотиды, кодирующие последовательности рибозимов, отжигали и затем клонировали в Sal I-сайт. Рибозимы, примененные для конструирования векторов crHIV, являлись рибозимами в форме головки молотка. Данные рибозимы содержали каталитический домен, состоявший из 22 пар оснований, и два домена гибридизации, каждый из которых включал 9 пар оснований. Рибозимы были направлены либо к сайту +115, либо к сайту +133 (т.е. соответственно количеству пар оснований ниже старта транскрипции) последовательности U5 РНК. Домены гибридизации и каталитический домен (подчеркнут) рибозимов, направленных к сайту +115 и сайту +133, были следующими:
CACACAACACTGATGAGGCCGAAAGGCCGAAACGGGCACA ("рибозим +115") SEQ ID NO:3
ATCTCTAGTCTGATGAGGCCGAAAGGCCGAAACCAGAGTC ("рибозим +133") SEQ ID NO:4
Рибозимная кассета была составлена из одного, двух или трех рибозима(ов), расположенных в виде тандема. Векторы, содержащие либо один, либо три рибозима, могут быть легко сконструированы и направлены на один и тот же сайт РНК U5 HIV в положении +115. Векторы, содержащие сдвоенные рибозимы, могут быть легко сконструированы и направлены либо на тот же сайт в положении +115, либо на другие сайты в положениях +115 и +133 РНК U5 HIV.
Для завершения конструирования векторов векторам crHIV придавали устойчивость к расщеплению рибозимами (т.е. при их манифестации в форме РНК) путем мутации сайта, узнаваемого рибозимами в форме головки молотка, находящегося в области U5 генома crHIV. Для этого для введения модифицированных сайтов в вектор применяли двухцепочечный олигонуклеотид (т.е.
AAGCTTGCCTTGAGTGCTCAAAGTAGTGTGTGCCCACCTGTTGTGTGACTCTGGCAG
CTAGAGATCCCACAGACCCTTTTAGTCAGTGTGGAAAATCTCTAGCAGTGGCGCC SEQ ID NO:13), содержащий замены оснований, описанные на фиг.2 SEQ ID NO:2. В частности, были введены замены оснований в сайты гибридизации и расщепления рибозимов в пары оснований 115 и 133. Конкретно, как показано на фиг.2, были введены мутации в пары оснований 113, 114, 132, 134 и 142. Данные сайты могут быть модифицированы для введения любой мутации (т.е. GTGTGCCCNNCTGTTGTGTGACTCTGGNANCTAGAGANC, где N может быть любым мутантным нуклеотидом SEQ ID NO:14). Предпочтительно, однако, чтобы последовательности мутировали таким образом, чтобы A заменял G в положении +113 (т.е. как в последовательности GTGTGCCCATCTGTTGTGTGACTCTGGTAACTAGAGATC SEQ ID NO:15), U (т.е. T в контексте последовательности ДНК) был заменен на C в сайте +114 SEQ ID NO:5, U (т.е. T в контексте последовательности ДНК) был заменен на C в сайте +132 SEQ ID NO:6, A был заменен на G в сайте +134 (т.е. так, что последовательность включает GTGTGCCCGTCTGTTGTGTGACTCTGGTAGCTAGAGATC SEQ ID NO: 16) и U (т.е. T в контексте последовательности ДНК) был заменен на A в сайте +142, причем данные мутации могут быть осуществлены изолированно или в сочетании. В частности, в отсутствие других мутаций U5 замена U (т.е. T в контексте последовательности ДНК) на C в сайте +114 SEQ ID NO:5 и/или сайте +132 SEQ ID NO:6 в РНК US crHIV предотвращает ее расщепление рибозимами (Uhlenbeck (1987), см. выше). Введенные замены оснований имеются в различных других штаммах HIV (Myers et al., HIV Sequence Database, Los Alamos Nat. Lab. (1994)), что указывает на то, что данные замены не снижают репликативной способности генома HIV.
Представленный здесь способ может быть применен для конструирования других векторов с зависимой от условий репликацией, например, составленных из различных вирусных геномов (например, различных РНК-вирусов) или составленных из различных генетических противовирусных агентов. Более того, вектор с зависимой от условий репликацией может быть дополнительно модифицирован для придания клетке-хозяину, в которую введен вектор, селективного преимущества по сравнению с клеткой-хозяином, содержащей вирус дикого типа. Например, такой вектор может быть модифицирован для дополнительного кодирования устойчивости ко многим лекарствам или мутантной протеазы или обратной транскриптазы.
Данные способы, конечно, могут быть также применены для конструирования описанных здесь различных конструктов хелперных векторов.
Пример 2
В данном примере описана устойчивость к рибозимному расщеплению векторов с зависимой от условий репликацией и, в частности, векторов crHIV.
Для подтверждения устойчивости к рибозимному расщеплению векторов crHIV проводили транскрипцию in vitro. Для этого рибозимные последовательности клонировали в Xho I-сайт pBluescript KSII (Stratagene, La Jolla, Calif.). Bgl II-фрагмент размером 0,21 тысячи пар нуклеотидов (т.п.н.), содержащий мутированную область U5 crHIV, был сходным образом вырезан из вектора crHIV и вставлен в Bam I-сайт pBluescript KSII. Полученные модифицированные векторы pBluescript KSII линеризовали с помощью Bss HII перед транскрипцией in vitro. Сходную плазмиду, экспрессирующую РНК U5 HIV-дикого типа (описанную в Myers et al. (1994), см выше), применяли в качестве контроля. Ее линеризовали с помощью Eco RI перед транскрипцией in vitro.
Радиоактивно меченную РНК U5 HIV и рибозимную РНК получали с помощью транскрипции in vitro векторов, как описано ранее (Dropulic' et al. (1992), см. выше). Радиоактивно меченные транскрипты инкубировали совместно (при молярном отношении мишени к рибозиму 1:2) в буфере 1X для транскрипции, содержащем 40 мМ трис-HCl, pH 7,5, 6 мМ MgCl2, 2 мМ спермидин и 10 мМ NaCl. Образцы нагревали до 65°С и затем охлаждали до 37°С в течение 5 мин перед добавления останавливающего буфера, содержащего 95% формамид, 20 мМ ЭДТА, 0,05% бромфеноловый синий и 0,05% Xylene Cyanol FF. Затем продукты разделяли с помощью электрофореза в полиакриламидном геле в денатурирующих условиях (PAGE) и определяли авторадиографией.
Когда РНК U5 HIV дикого типа инкубировали с транскриптом, содержащим единственный рибозим к сайту +115, расщепление легко наблюдалось. Такое расщепление давало продукты P1 и P2. Расщепление также наблюдалось при инкубации РНК HIV дикого типа с РНК, содержащими сдвоенные рибозимы, направленные либо на тот же сайт, либо на другие сайты. Когда содержащий рибозим транскрипт, направленный на два различных сайта, инкубировали с РНК HIV дикого типа, образовывались продукты P1, P2 и P3. P3 возникал в результате расщепления по сайту +133.
Напротив, когда инкубировали модифицированную РНК, содержащую U5 crHIV, с либо одним рибозимом, направленным на сайт +115, либо сдвоенным рибозимом, направленным либо на сайт +115, либо на сайт +133, продукты расщепления не обнаруживались. Таким образом, данные результаты подтверждают, что РНК U5 crHIV устойчивы к расщеплению рибозимами, тогда как РНК U5 HIV дикого типа расщепляются анти-US рибозимами. Более того, результаты подтверждают, что подход настоящего изобретения может быть применен для придания векторам с зависимой от условий репликацией (включая векторы, отличные от векторов HIV) селективного преимущества в репликации при введении в клетку-хозяин по сравнению со штаммом вируса дикого типа.
Пример 3
В данном примере кратко описаны ключевые стадии получения конструктов хелперных векторов изобретения. Если это специально не указано, последовательности нуклеиновой кислоты, кодирующие различные ретровирусные элементы, получали из общедоступных источников. На всех описанных стадиях требовалось секвенирование полной кодирующей области для всех белков для выявления ошибок и при необходимости прямой коррекции.
Для вырождения последовательности tat и обеспечения ее атаки анти-tat рибозимами плазмиду pTAT подвергали мутагенезу с помощью набора QuickChange от Stratagene для включения сайтов-мишеней для кассеты рибозимов против tat. Затем мутированную последовательность tat клонировали в экспрессионный вектор на основе плазмиды pMG от InvitroGen. Последовательность tat соединяли с IRES по первому ATG с применением NcoI-сайта. Вектор дополнительно модифицировали для включения гена резистентности к ампициллину и старта репликации SV40.
Последовательности, содержащие элементы rev и RRE HIV-1, были получены вырезанием третьего экзона rev вместе с RRE из pNL4-3 и клонированием в плазмиду pLITMUS38. Последовательность Rev до нуклеотида 208, которая включает второй экзон и часть третьего экзона до BamHI-сайта, была собрана из 3 синтетических нуклеотидов с применением ETR и ПЦР. Продукт ПЦР клонировали в pUC18 и затем субклонировали в pLIT/RRErev3. Две части rev соединяли с помощью набора QuickChange. RRE из HIV-1 заменяли RRE из HIV-2 и оба конструкта клонировали в вектор gag/pol/Rz. Вырожденные последовательности rev также непосредственно применяли для субклонирования в упаковывающий конструкт для таких векторов, как pVIRPac-2.
Для индукции вырождения первых 42 нуклеотидов gag, которые необходимы для упаковки и должны быть частью векторной плазмиды, фрагмент BssHII/SpeI из pNL4-3 субклонировали в pLITMUS38. Вырождение последовательности gag вызывали с помощью набора QuickChange, так что все компоненты сигнала упаковки были ликвидированы, и главный донор сплайсинга HIV был вновь вставлен перед первым кодоном ATG gag. Вырожденный gag субклонировали в плазмиду pNgp, любезно предоставленную Dr. Conde, на место 5' LTR для образования длинной последовательности gag. Данный конструкт все еще содержал последовательности акцептора сплайсинга (SA) и донора сплайсинга (SD), используемые для экспрессии белка vif. Данный сплайсинг может происходить в присутствии рибозимной кассеты в подвергаемой сплайсингу мРНК, что нежелательно из-за возможного расщепления подвергнутой сплайсингу ретровирусной (с зависимой от условий репликацией) векторной РНК и возникающего в результате снижения титра упакованного трансдуцирующего вектора. Для мутирования последовательностей SA и SD применяли перекрывающую ПЦР. Продукт ПЦР вставляли и клонировали в конструкт gag/pol с последующей вставкой рибозимовой кассеты против U5 и элементов rev и RRE.
Белок G везикулярного стоматита (VSV-G) клонировали в первый MCS вектора pMG-tat для создания экспрессионной кассеты оболочки. Для упрощения последующих этапов клонирования VSV-G клонировали в виде фрагмента с тупыми концами в Bg1II-сайт, замещенный с помощью полимеразы Кленова. Была создана конструкция со стартом репликации (ori) SV40. Для завершения построения плазмид упаковки в качестве хелперных векторов последовательность rev субклонировали во второй MCS. Полученная плазмида была названа pVIRPAC-2 (смотри фиг.6).
Для создания хелперных векторов из некоторых упаковывающих плазмид кассету gag/pol/Rz/RRE/rev субклонировали в плазмиду pMG-tat-G (без SV40 ori). Были получены конструкты с RRE либо из HIV-1, либо из HIV-2 (pVIRPac-1.1 и 1.2Rz на фиг.6). В качестве контроля для тестирования рибозимной функции из конечного содержащего RRE HIV-2 конструкта рибозимная кассета была удалена, что дало pVIRPac-1.2. Наконец, для проверки гипотезы о том, что введение РНК gal/pol в механизм сплайсинга может предотвращать расщепление ретровирусной (с зависимой от условий репликацией) векторной геномной РНК перед сигналом поли-A SV40 был вставлен интрон из β-глобина (pVIRPac-1.2RzIn). Разумеется, в других осуществлениях изобретения вставленный интрон происходит от HIV, предпочтительно полный интрон HIV с подтвержденной способностью направлять РНК в механизм клеточного сплайсинга (например, сплайсосому).
Пример 4
В данном примере описывается эффективность различных хелперных векторных конструктов. В данных экспериментах применяли стандартные протоколы получения ретровирусных векторов путем преходящей трансфекции. Большие количества плазмидной ДНК получали с применением известных способов. Для получения вируса клетки 293T совместно трансфицировали ретровирусными векторными плазмидами и хелперными векторными плазмидами с помощью кальций-фосфатного способа. Супернатанты клеток собирали через приблизительно 40 ч после трансфекции, фильтровали через 0,45 мкм фильтры для шприца и титровали на клетках HT1080. Титр вектора рассчитывали на основе его трансдуцирующей эффективности, измеренной проточной цитометрией через приблизительно 72 ч после инфицирования. Для вычислений титров применяли условную формулу N*400000*, где N представляет долю трансдуцированных клеток; 400000 является приблизительным количеством клеток во время инфицирования; и F является фактором разведения.
Для повышения сохранности хелперного вектора путем снижения возможности возникновения RCV при рекомбинации с ретровирусным вектором на основе HIV-1 в некоторые конструкты хелперного вектора вставляли анти-U5 рибозимную кассету. Рибозим был бы должен разрезать и разрушать ретровирусную векторную РНК, если бы происходила совместная упаковка в вирион. Возможно, однако, что рибозим мог бы также расщеплять векторную РНК внутри продуцирующей клетки, что влияло бы на образование вируса. Для дополнительного повышения сохранности хелперные конструкты содержали элемент RRE-2 для дополнительного снижения вероятности совместной упаковки хелпера и вектора в вирусную частицу. Данные хелперы применяли для упаковки вектора pN1(cPT)GFP.
Как показано на фиг.7, присутствие рибозима на хелперном конструкте оказывало слабое влияние на титр вектора. Важно, что хелперный конструкт (pVirPac1.2RzIn), содержащий интрон для влияния на перемещение в клетке хелперной РНК от векторной РНК, также не оказывал заметного влияния на титр вектора. Титр вектора, индуцированный данной 2-плазмидной системой, сопоставим с титром, полученным с применением традиционных 3-плазмидных систем трансфекции. Сходные эксперименты с хелперным конструктом, содержащим RRE-1, pVirPac1.1, показали приблизительно в 5 раз более низкую эффективность упаковки. Таким образом, включение RRE-2 в конструкт хелперного вектора не только увеличивало его сохранность за счет снижения возможности гомологичной рекомбинации, но также неожиданно повышало эффективность упаковки.
Пример 5
В данном примере проверяли, является ли экспрессия гена MGMT из подвергнутой сплайсингу РНК HIV-LTR достаточной для эффективной селекции трансдуцированных MGMT клеток с BG и BCNU. На фиг.10A показаны примененные векторы. На фиг.10B показаны эффективное выживание и размножение клеток в присутствии гибельных для клеток концентраций BG/BCNU. Буквенные обозначения представляют следующее: M, MGMT; I IRES; G, GFP; W, посттранскрипционный элемент Woodchuck; и C, промотор CMV. Все указанные выше элементы клонировали в вектор pN1 ниже элемента RRE, так что любой ген, расположенный сразу с 3' конца от элемента RRE, должен был бы экспрессироваться с подвергнутой сплайсингу РНК, поскольку акцепторный сайт сплайсинга был помещен непосредственно перед сайтом вставки гена. На фигуре показано, что в то время как контрольные клетки, не трансдуцированные лентивирусным вектором ("CGFP"), не распространялись или выживали, все векторы, экспрессирующие MGMT, выживали и увеличивались в числе.
Клетки, трансдуцированные pN1CMIG, погибали через 14 недель, в то время как клетки, трансдуцированные pN1MCG, гибли через 21 неделю. Важно, что клетки, трансдуцированные pN1MIG и pN1MIG-W, оставались живыми после 29 недель, что свидетельствует о длительном выживании клеток, трансдуцированных вектором без вставленного внутреннего промотора.
Неожиданно оказалось, что векторы, экспрессирующие MGMT с подвергнутой сплайсингу мРНК HIV-LTR, отбираются очень эффективно. Экспрессия MGMT не ограничивает объем загружаемых генов MGMT или генами селекции. Данные результаты показывают, что любой ген может быть экспрессирован с промотора HIV-LTR в T-клетках. Сходные данные были получены в клетках, отличных от T-клеток, что показывает, что экспрессия генов с подвергнутой сплайсингу мРНК HIV-LTR могла бы быть применена для экспрессии генов во многих типах клеток.
Кроме того, бицистронная природа конструктов "MIG" показывает, что множество генов может быть экспрессировано после сращивания с помощью IRES. Примеры, выходящие за рамки описанного выше, включают MGMT или другой ген селекции с хемокином, интерфероном или другим генетическим противовирусным агентом.
Пример 6
T-клетки CD4+ человека тестировали на их чувствительность к BG и BCNU. Клетки CD4+ с недостаточностью CD14 очищали из периферической крови с или без мобилизации цитокином. Клетки культивировали с 3 мкг/мл PHA и 5 нг/мл ИЛ-2 в течение 4-7 дней перед обработкой BG и BCNU. В подходящих условиях в отсутствие BFG и BCNU клетки CD4+ могут разрастаться 100-кратно на протяжении 10 дней в культуре. Тестированные дозы равнялись 0, 0,2, 0,5, 2, 5, 10, 20, 30 и 40 мкМ BG и 0, 2, 5, 10, 20, 30 и 40 мкМ BCNU. Были проведены серии из трех экспериментов.
На фиг.11 показаны типичные результаты указанных выше экспериментов. Разрастание снижалось до 20-кратного после 10 мкМ BG и мкМ BCNU и до 10-кратного или ниже после 20 мкМ или более BCNU. Обработка клеток CD4+ BCNU не приводила к задержке роста. Таким образом, в отличие от клеток SupT1, примененных в указанном выше примере, клетки CD4+ не чувствительны к низкой дозе одного BCNU. Это предполагает большую экспрессию MGMT в первичных клетках CD4+, хотя на основании вариации среди доноров T-клеток можно ожидать вариации в экспрессии MGMT.
Присутствие и BG, и BCNU, каждый в концентрации 10 мкМ или выше, было наилучшим при сенсибилизации клеток CD4+, тогда как BG в концентрации 0,5 мкМ и BCNU в концентрации 15 мкМ или выше сходно повышали чувствительность клеток. Повышение дозы BG не увеличивало гибели клеток, хотя токсичность к клеткам CD4 возрастала с дозой BCNU.
Как показано на фиг.11, уровень селекции в целом возрастал 5-кратно от исходных 20% до 100%. В одном эксперименте селекция возрастала от 3% до >90%. 10-100-кратное разрастание наблюдалось после селекции и одной недели; 20-400-кратное разрастание наблюдалось при 2,5-кратной селекции при средней кратности, равной 80.
Пример 7
Предшествующая работа позволяет предполагать корреляцию между очисткой аутотрансплантатов костного мозга и улучшением безболезненного выживания после аутотрансплантации. Удаление нормальных T-клеток из аллотрансплантата было также применено для снижения риска заболевания трансплантат против хозяина. На фиг.13 показана эффективность трансдукции опухолевых клеток в одном раунде трансдукции с применением pN1(cPT)ASenvGFP. Фиг.13B подтверждает, что опухолевые клетки могут быть селективно и эффективно убиты по отношению к стволовым клеткам CD34+, поскольку последние не проявляют значительной трансдукции после одного раунда трансдукции.
Применение описанного выше для удаления опухолевых клеток из аутотрансплантата костного мозга начинают со сбора костного мозга или сбора стволовых клеток периферической крови. Клетки CD34+ очищают посредством колонки с антителом к CD34 и затем трансдуцируют вектором с зависимой от условий репликацией при MOI от 2 до 400 в течение от 2 до 16 часов. Клетки необязательно инкубируют в присутствии одного или более активирующих клетки факторов, таких как цитокины, либо для содействия трансдукции, либо для поддержания клеток. Клетки затем переносят в подлежащий лечению субъект.
Способность сходным образом поражать T-клетки в аллотрансплантате применяет векторы с зависимой от условий репликацией в сочетании с высокоэффективным протоколом стабильной трансдукции, таким как имеющийся в параллельно поданной патентной заявке США с порядковым номером (еще не присвоенным), зарегистрированной 31 августа 2000 в качестве заверенной описи 397272000400, для селективной трансдукции T-клеток.
Пример 8
На фиг.14, панель B, показано эффективное псевдотипирование векторов HIV-1 изобретения различными белками оболочки. Сходные исходные титры с лентивирусным вектором, псевдотипированным белком оболочки VSV-G, могут быть эффективно увеличены путем очистки с применением либо высокоскоростного центрифугирования, либо диафильтрации. Вектор pN1(cPT2)ASenvGFP упаковывали с pVirPac1.2Rz2. В таблице показаны (a) общий белок в образце, (b) титр вектора в образце и (c) кратность очистки вектора от чужеродного материала в образце.
Как видно из таблицы, общий белок в образце векторного супернатанта после осветления от обломков клеток равен 3072 мг, а титр вектора на этой стадии равен 5,9×106. Образец затем подвергают концентрированию ультрафильтрацией, и к этому моменту общий белок в этом образце составляет 722 мг, а титр вектора возрастает до 8,2×107 при 61,7-кратной очистке. После концентрирования ультрафильтрацией образец подвергают диафильтрации, и к этому моменту общий белок в образце дополнительно снизился до 224 мг, тогда как титр вектора увеличился до 6,8×108 на мл, что соответствует 1650-кратной очистке. Это показывает, что невекторный чужеродный белок может быть удален, и лентивирусный вектор, который псевдотипируется с VSV-G, может быть очищен с применением описанного выше протокола.
Описанное выше может быть также применено в сочетании с векторами, продуцированными в линии клеток HeLa-tat, дающей титры приблизительно 1010, как показано на фиг.16.
Пример 9
Альтернативные хелперные конструкты для псевдотипирования векторов с зависимой от условий репликацией были разработаны путем постановки экспрессии VSV-G под зависимый от Rev контроль. Поскольку содержащие RRE мРНК не экспрессируются в отсутствие Rev, было предположено, что они дефективны из-за наличия репрессорных цис-последовательностей (CRS), находящихся в генах gag/pol, pol и env. Сообщалось, что CRS улавливает мРНК в ядре, дестабилизирует мРНК или предотвращает ассоциацию мРНК с рибосомами. Таким образом, присутствие CRS и Rev, расположенных в транс-положении, в сочетании со сплайсингом мРНК может быть применено для получения регулируемой экспрессии гена.
Систему Rev/RRE/CRS использовали для контроля экспрессии VSV-G зависимым от Rev образом. Хелперные конструкты для данного подхода содержат 5'донорный сайт сплайсинга; RRE HIV-2 и 3'акцепторный сайт сплайсинга (предпочтительно для tat/rev); фрагмент CRS из области pol HIV-2 для снижения вероятности рекомбинации с основанными на HIV-1 векторами; и необязательно неиндуцибельный промотор, такой как немедленный ранний (IE) промотор CMV, вместо любых индуцибельных промоторов. РНК, кодирующие VSV-G, образующиеся в отсутствие индукции, продолжают оставаться регулируемыми благодаря присутствию Rev, поскольку лишь не подвергнутые сплайсингу РНК могут быть экспортированы и экспрессированы в цитоплазме без Rev. Экспрессия VSV-G остается опосредуемой Rev, поскольку Rev препятствует ядерной задержке транскриптов, контролируемых сплайсингом мРНК и CRS.
Предпочтительно, чтобы донорный сайт сплайсинга не был синтетическим 5' донорным сайтом сплайсинга β-глобина, а был нативным донорным сайтом сплайсинга HIV или аналога HIV. RRE HIV-2 представляет собой фрагмент из 280 п.н., содержащий и RRE HIV-2, и акцепторный сайт сплайсинга для Tat/Rev; а CRS является внутренним фрагментом размером 550 п.н. из последовательности pol HIV-2. Элементы CRS и RRE из HIV-2 могут быть получены с помощью ПЦР. Неограничивающие примеры таких хелперных конструктов включают pCGCRSRRE, pCGCRSRzRRE и другие, показанные на фиг.15A-15E. Конечные плазмидные конструкты были подтверждены расщеплением множеством ферментов рестрикции. Хотя pEH-GP и pCMV-GP являются конструктами, которые конститутивно экспрессируют полипротеин gag-pol, могут быть применены другие промоторы, включая лентивирусный промотор или промотор HIV, если это желательно.
На фиг.17 показаны результаты, полученные при применении различных комбинаций доноров сплайсинга для зависимой от Rev экспрессии VSV-G. Как показано, удаление тетрациклина приводит к зависимой от Rev экспрессии.
Дополнительные примеры сочетаний донорных сайтов сплайсинга, а также консенсусная последовательность приведены ниже. Хотя все они могут быть использованы, сочетания доноров сплайсинга главного HIV, HIV-1 env, главного HIV-2 и аналога являются предпочтительными.
Пример 10
Для выявления подходящих условий для хранения векторов перед применением было проверено множество буферов для хранения и условий. Результаты показаны на фиг.18. Возможность хранить векторы при -20°С в фармацевтически приемлемом буфере без значительного снижения выхода вектора должно быть огромным достижением в сохранении векторов перед применением. Разумеется, хранение векторов при приблизительно -80°С, приблизительно -20°С и приблизительно 4°С в других фармацевтически приемлемых композициях также может быть легко осуществлено на практике.
Пример 11
В данном примере описана аминокислотная последовательность гибридного эпитопа ЦТЛ ВИЧ для практического применения изобретения. Последовательность (SEQ ID NO: 18) содержит первый метионин (M) для инициации трансляции, сопровождаемый различными смежными последовательностями, соответствующими p17, p24, p15, Pol, Rev, gp120env, gp41env и nef соответственно.


Пример 12
В данном примере предлагается краткое изложение осуществлений изобретения, как описано выше. Изобретение включает вирусный вектор с зависимой от условий репликацией, включающий ген (или другую нуклеиновую кислоту), подлежащий экспрессии, и одну или более первых нуклеотидных последовательностей, где вектор (a) реплицируется в клетке-хозяине лишь при комплементации штаммом вируса дикого типа, хелперным вирусом или хелперным вектором, каждый из которых чувствителен к присутствию указанных одной или более первых нуклеотидных последовательностей; (b) устойчив к присутствию указанных одной или более первых нуклеотидных последовательностей; и (c) содержит одну или более замен, вставок или делеций, что снижает вероятность возникновения способного к репликации вектора.
Для снижения вероятности возникновения способного к репликации вектора в изобретении предлагаются векторы, содержащие вырожденные последовательности в отношении одной или более последовательностей хелперного конструкта, состава нуклеиновой кислоты клетки-хозяина или других нуклеиновых кислот. Кроме того, предлагаются хелперные конструкты для применения в сочетании с такими векторами для их упаковки. Такие хелперные конструкты могут также содержать вырожденные последовательности в отношении одной или более последовательностей вектора, состава нуклеиновой кислоты клетки-хозяина или других нуклеиновых кислот.
Векторы изобретения могут также содержать одну или более последовательностей, которые направлены на хелперный конструкт. Такие последовательности включают генетические противовирусные агенты и предпочтительно являются рибозимами или антисмысловыми нуклеиновыми кислотами (или кодируют их). Предпочтительные рибозимы и антисмысловые последовательности способны к формированию комплементарных пар оснований со своими соответствующими мишенями посредством спаривания оснований G-U в дополнение к стандартному спариванию оснований A-T, A-U и G-C. Сходным образом хелперные конструкты изобретения могут также содержать тот же тип последовательностей, направленных на вектор.
Хелперные векторы изобретения могут также содержать одну или более последовательностей, которые влияют на локализацию или перемещение в клетке экспрессируемых хелпером РНК по сравнению с вектором. Например, хелперные векторы могут содержать интрон, гетерологичный RRE по сравнению с вектором или гетерологичную последовательность gag по сравнению с вектором. Хелперные векторы могут также содержать вырожденную последовательность(последовательности) gag и/или pol по сравнению с вектором, в результате чего совместная локализация хелперных и векторных РНК является менее вероятной. Хелперные векторы могут также содержать донорные сайты сплайсинга, в результате чего сигналы (последовательности) упаковки и RRE удаляются из РНК для предотвращения их совместной локализации с векторными РНК. Такие донорные сайты сплайсинга могут также иметься в хелперных векторах для индукции дифференцированного перемещения вектора по сравнению с хелперным вектором или вирусами дикого типа.
Хелперные векторы изобретения могут быть также способны псевдотипировать векторы изобретения. Типичные примеры включают VSV-G, RD114, вирус бешенства, GALV и гибридные белки оболочки. Хелперные векторы изобретения также предпочтительно берут свое начало от гетерологичного вируса в отношении вектора. Примером являются хелперные векторы, происходящие от HIV-2. Способы изобретения включают применение таких хелперных векторов для репликации векторов изобретения.
Типичные векторы изобретения включают векторы серий pN1 и pN2, показанные на фиг.1A-1K, в особенности с удаленными или замененными последовательностями GFP. Предпочтительно, чтобы векторы сохраняли, по меньшей мере, антисмысловую последовательность и содержащую cPT последовательность. Типичные хелперные конструкты изобретения включают конструкты серии pVirPac, как показано на фиг.6A-6G, с рибозимами или без них.
Все векторы и хелперы изобретения могут быть вырожденными, как описано выше, или в других частях конструктов, как это необходимо. Предпочтительная вырожденность касается кодонов, предпочтительно используемых в клетках человека. Векторы и хелперы могут быть также гибридными по своей природе, такими что они происходят от различных естественно существующих, а также синтетических нуклеиновых кислот. Предпочтительные гибридные конструкты содержат последовательности и HIV-1, и HIV-2. Однако особенно предпочтительными векторами являются производные HIV-1 или кодирующие белок Tat HIV-1. В противоположном варианте векторы не кодируют белок Tat, который обеспечивается хелперным вектором, вирусом дикого типа или клеткой-хозяином.
Векторы изобретения также предпочтительно не кодируют или не экспрессируют вспомогательные вирусные белки. Векторы могут также включать нуклеиновую кислоту, которая снижает, минимизирует или предотвращает вирусную инфекцию при ее наличии в клетке-хозяине или придает указанной клетке устойчивость к токсичным агентам. Примеры таких нуклеиновых кислот включают белок клеточной поверхности или укороченный белок CCR5, предпочтительно белок CCR5Δ32T. Другие примеры включают нуклеиновые кислоты, кодирующие устойчивость к лекарствам или повреждающий ДНК агент, предпочтительно устойчивость ко многим лекарствам или вариант MGMT, в особенности вариант G156A. Дополнительные примеры включают нуклеиновые кислоты, кодирующие трансдоминантный фенотип. Типичным примером является последовательность трансдоминантного gag HIV-1.
Векторы изобретения также могут содержать последовательности для помощи в экспрессии гетерологичной последовательности. Типичным примером служит последовательность изолятора. Векторы также могут содержать последовательность, которая улучшает трансдукцию вектора в клетки. Типичным примером является последовательность из 545 пар нуклеотидов, содержащая описанный выше cPT, хотя могут быть применены также меньшие или большие фрагменты, содержащие cPT. Векторы могут также содержать LTR для экспрессии генов, а также нуклеиновые кислоты, включающие LTR, модифицированные по локализованным в них транскрипционным сайтам связывания. Предпочтительно, чтобы такие модифицированные LTR содержались в лентивирусных векторах. Способы изобретения включают применение таких векторов.
Векторы изобретения могут также экспрессировать, по меньшей мере, два гена с использованием подходящим образом расположенных сайтов сплайсинга или элементов IRES. Векторы могут быть также упакованы в вирусные частицы с одним или более гибридными белками оболочки, которые стимулируют мишень или клетку-хозяин для трансдукции. Кроме того, векторы могут быть упакованы в клетках, где хелпером является белок, или хелпер содержит систему Rev/RRE/CRS для регуляции генной экспрессии. Способы изобретения включают применение таких векторов.
Векторы изобретения могут быть также применены для индукции селективной гибели трансдуцированных клеток в сочетании с вводимым агентом, который в нормальных условиях не является токсичным для нетрансдуцированных клеток. Предпочтительно, чтобы такие векторы действовали путем образования молекулы нуклеиновой кислоты, а не белка. Типичным примером является образование антисмысловой последовательности против MGMT или молекула рибозимной нуклеиновой кислоты. Способы изобретения включают применение таких векторов.
В изобретении также предлагаются способы хранения векторов изобретения.
Все цитированные здесь ссылки, включая патенты, патентные заявки и публикации, включены здесь во всей своей полноте в качестве ссылки. Все примененные здесь термины, представленные в форме единственного числа, предназначены для включения форм как единственного, так и множественного числа.
Хотя данное изобретение было описано с подчеркиванием предпочтительных осуществлений, специалистам в данной области техники должно быть ясно, что могут быть осуществлены и применены вариации в предпочтительных осуществлениях и что изобретение может быть реализовано на практике иначе, чем это конкретно описано здесь. Настоящее изобретение предназначено для включения таких вариаций и альтернативных практических осуществлений. Соответственно, данное изобретение включает все модификации, охватываемые духом и объемом изобретения, как определено формулой изобретения.
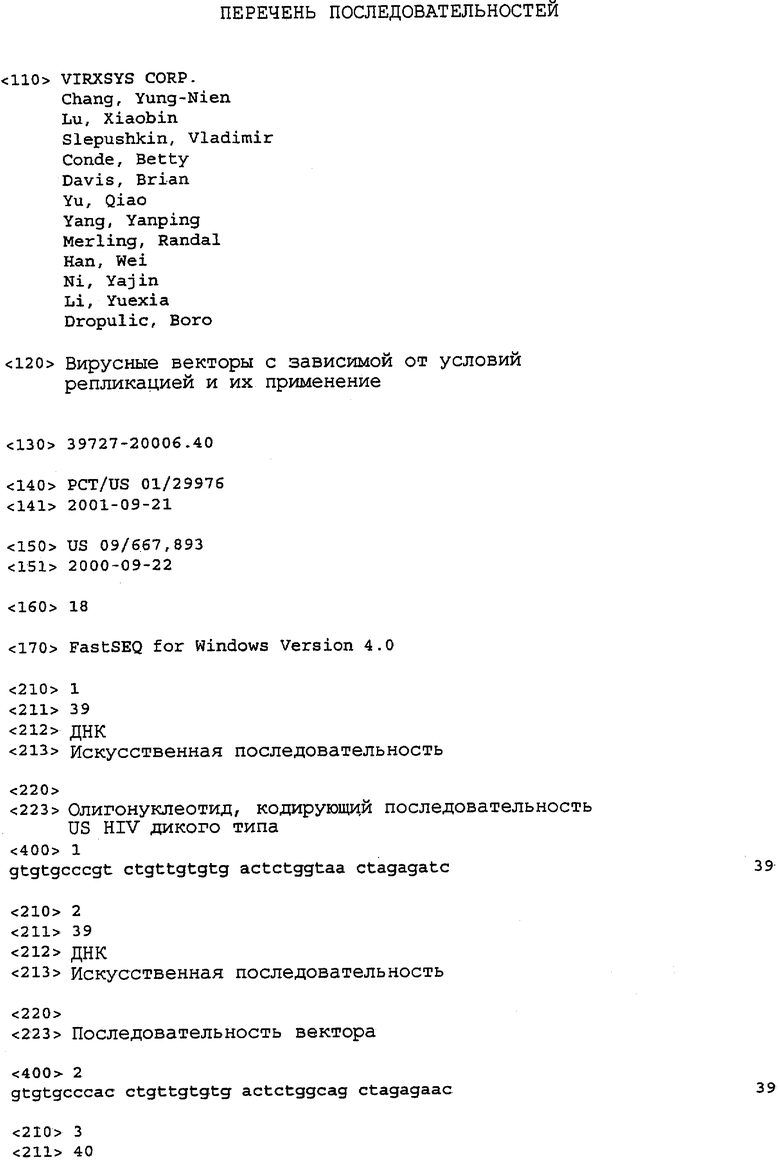








| название | год | авторы | номер документа |
|---|---|---|---|
| УСЛОВНО РЕПЛИЦИРУЮЩИЙСЯ РЕТРОВИРУСНЫЙ ВЕКТОР (ВАРИАНТЫ), СПОСОБ ЕГО ПОЛУЧЕНИЯ И ИСПОЛЬЗОВАНИЯ (ВАРИАНТЫ), ВЫДЕЛЕННАЯ И ОЧИЩЕННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ | 1996 |
|
RU2270250C2 |
| СПОСОБ ВРЕМЕННОЙ ТРАНСФЕКЦИИ ДЛЯ ПРОДУЦИРОВАНИЯ РЕТРОВИРУСА | 2016 |
|
RU2749717C2 |
| СТАБИЛЬНЫЕ КЛЕТОЧНЫЕ ЛИНИИ ДЛЯ ПРОДУЦИРОВАНИЯ РЕТРОВИРУСОВ | 2016 |
|
RU2752498C2 |
| ПОЛИВАЛЕНТНЫЕ ВИРУСНЫЕ ВЕКТОРЫ И СИСТЕМА ДЛЯ ИХ ПОЛУЧЕНИЯ | 2005 |
|
RU2416646C2 |
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ АНТИВИЧ-ТЕРАПИИ | 2010 |
|
RU2426788C1 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| СПОСОБЫ СТАБИЛЬНОЙ ТРАНСДУКЦИИ КЛЕТОК ВИРУСНЫМИ ВЕКТОРАМИ | 2001 |
|
RU2280074C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДОСТАВКИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА | 1994 |
|
RU2174845C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ С MND-ПРОМОТОРОМ | 2015 |
|
RU2708311C2 |
| Направляемое РНК уничтожение вируса JC человека и других полиомавирусов | 2015 |
|
RU2747722C2 |
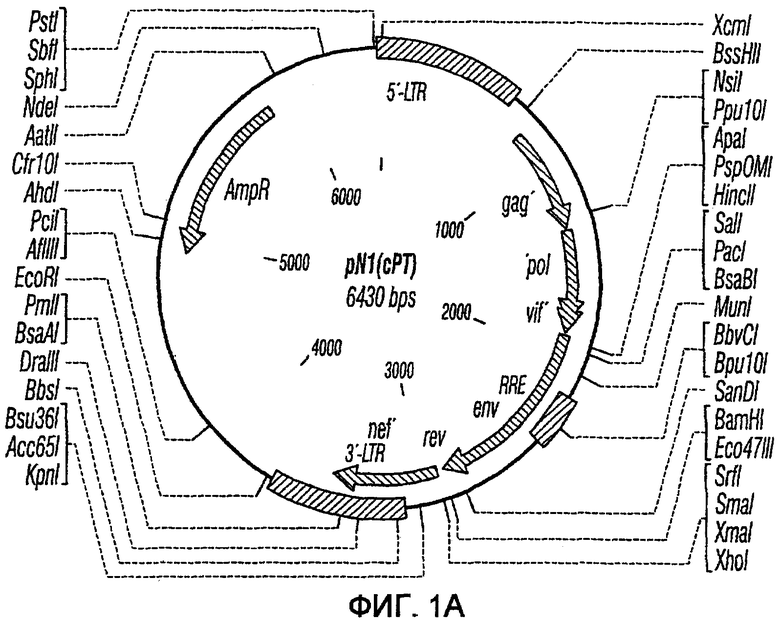
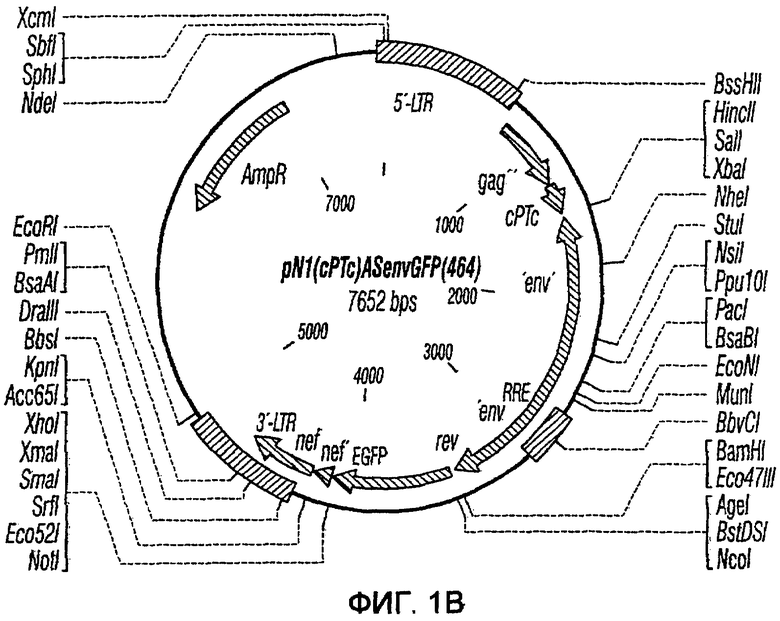

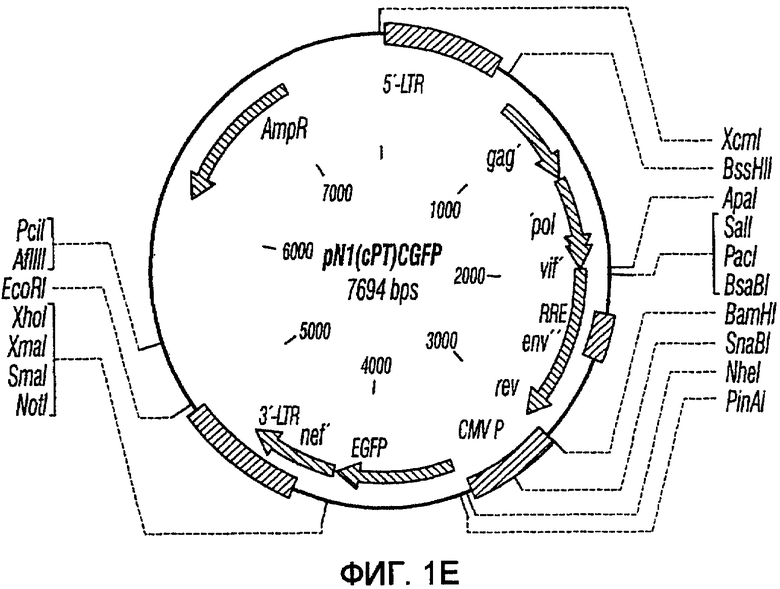
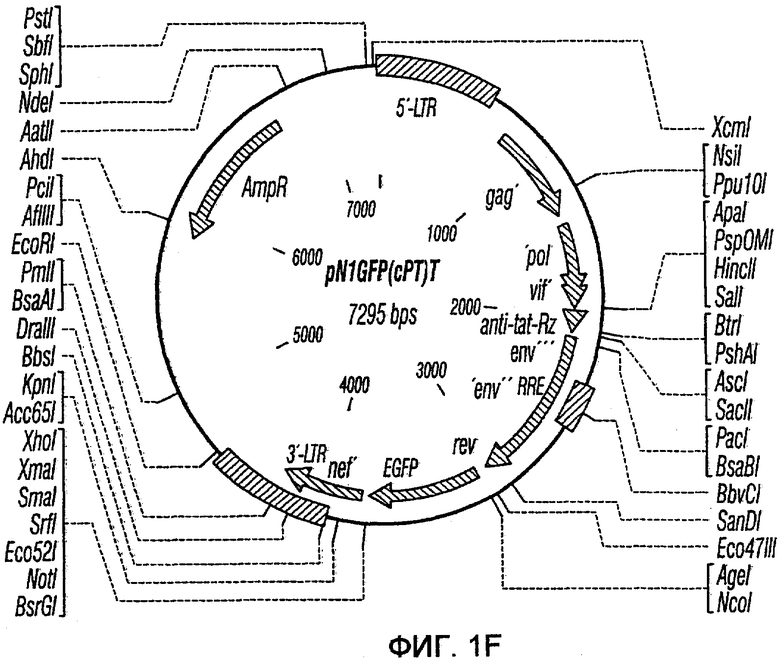
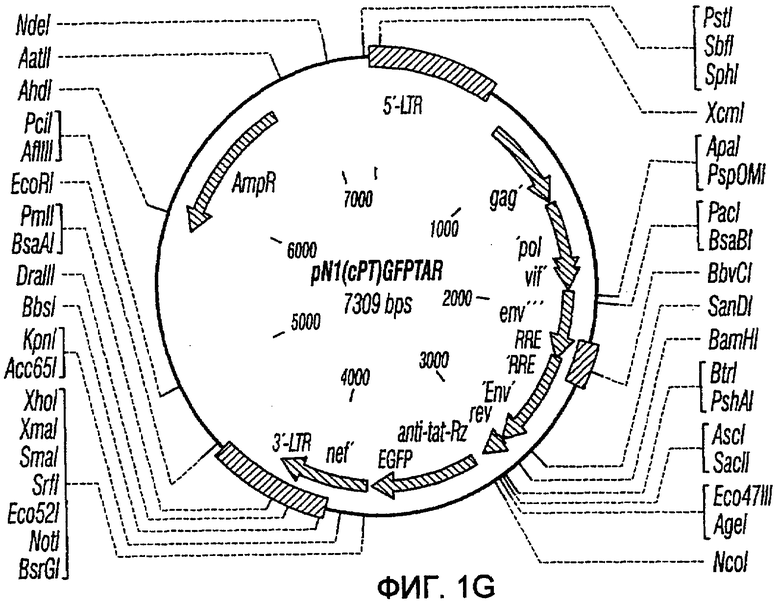
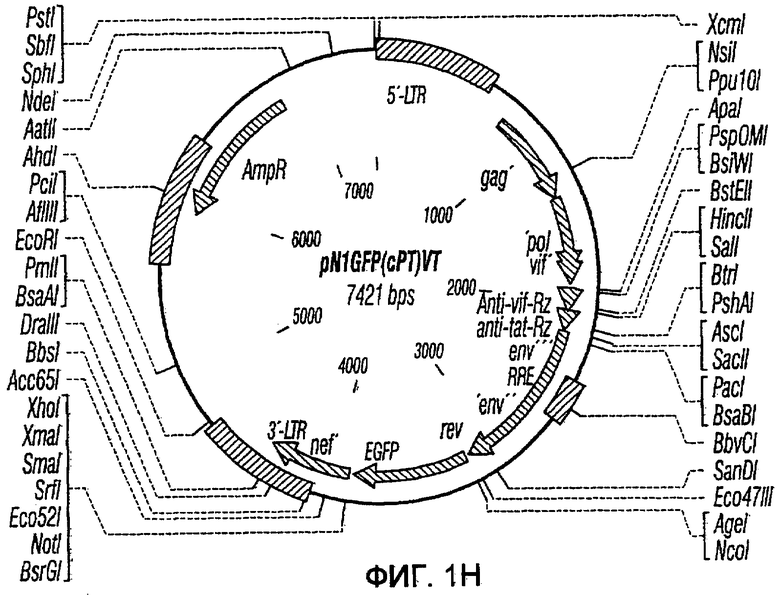
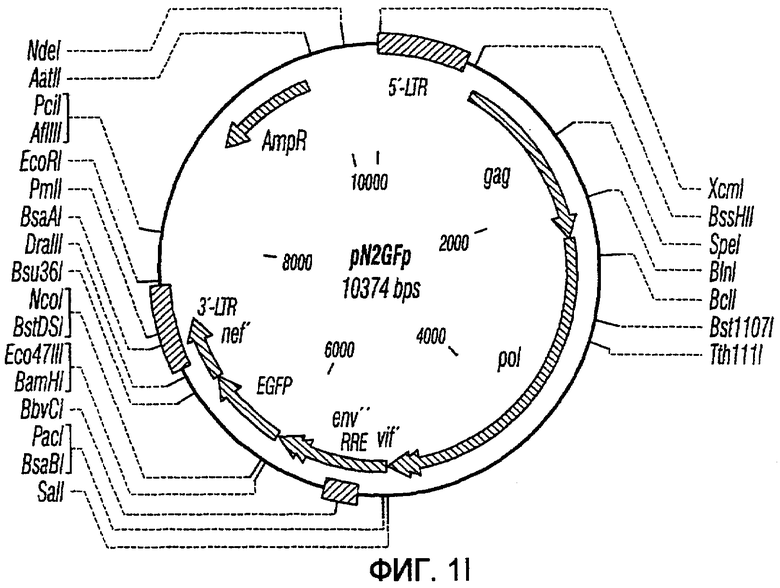
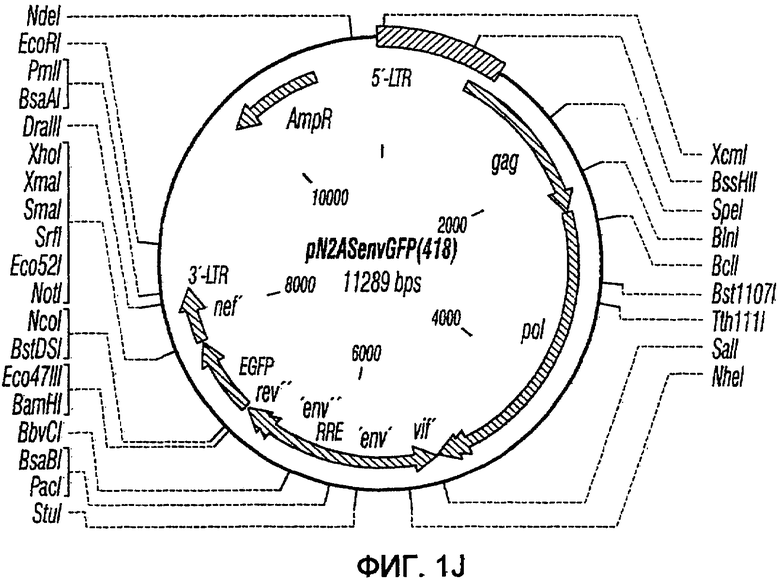
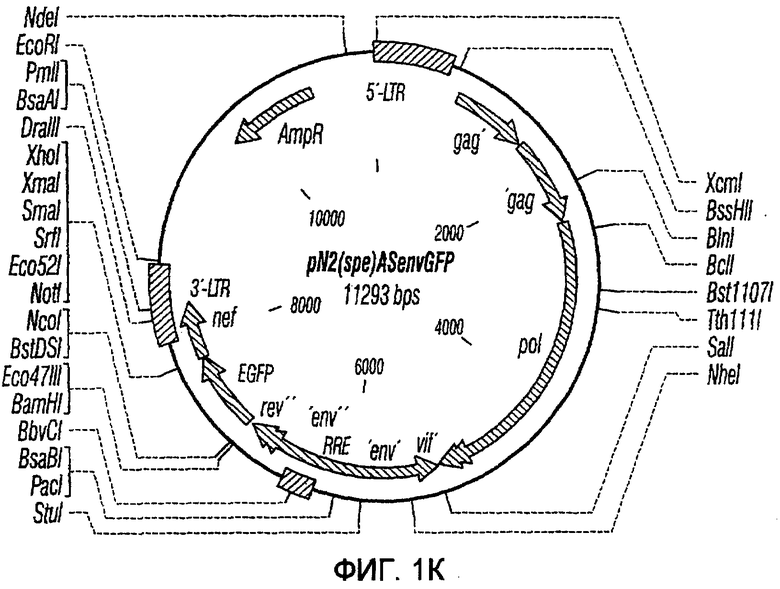
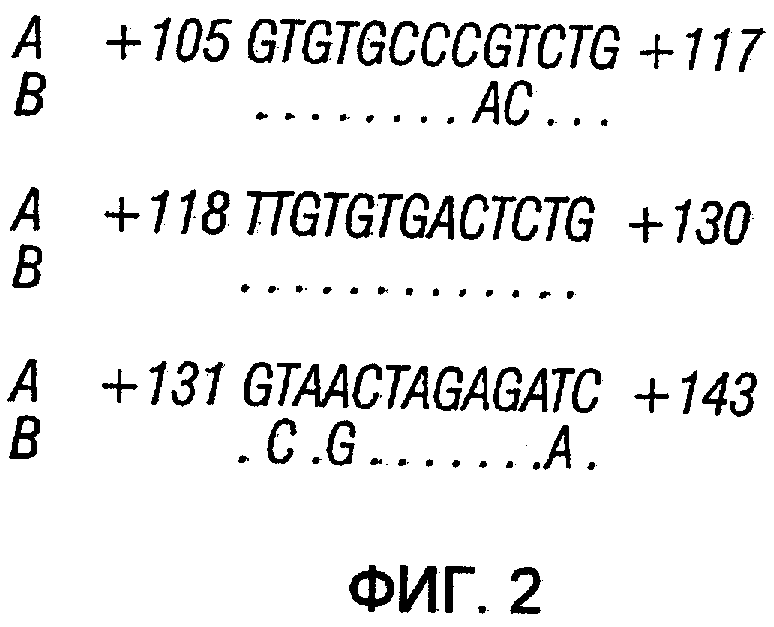
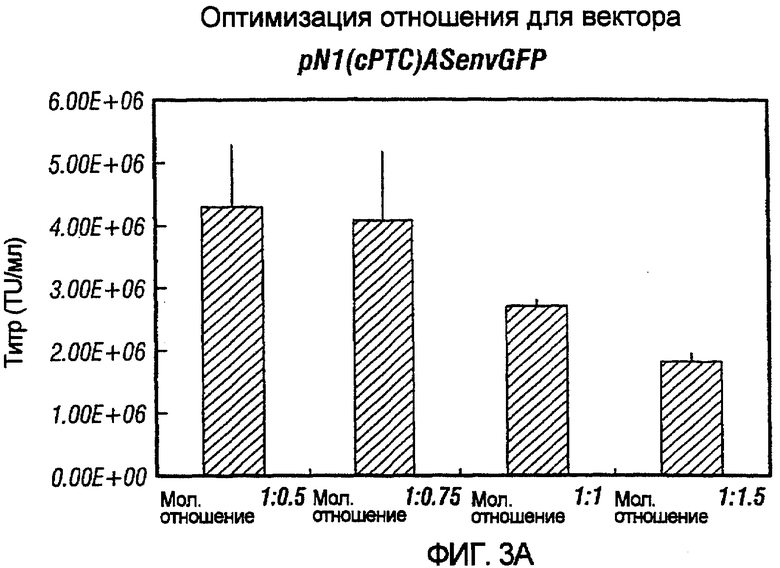
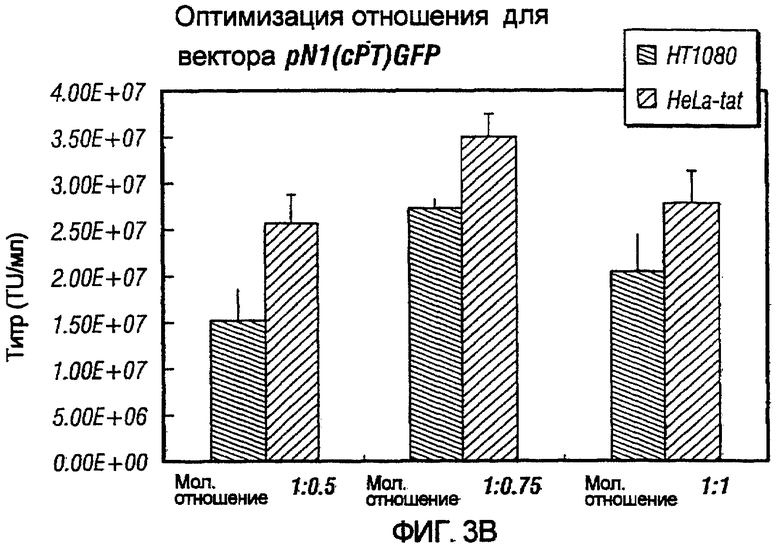
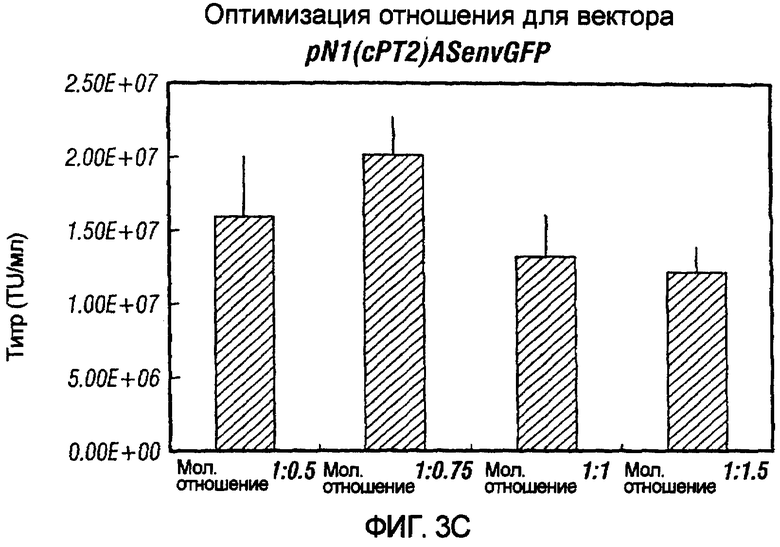
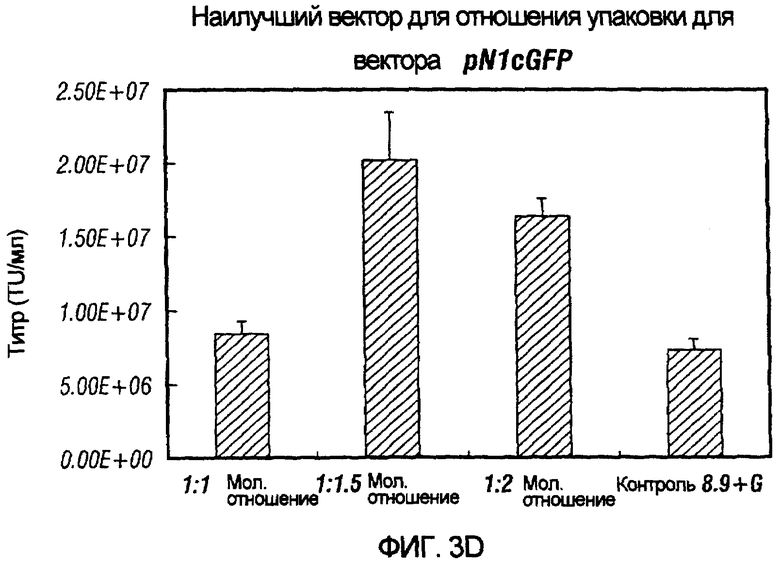
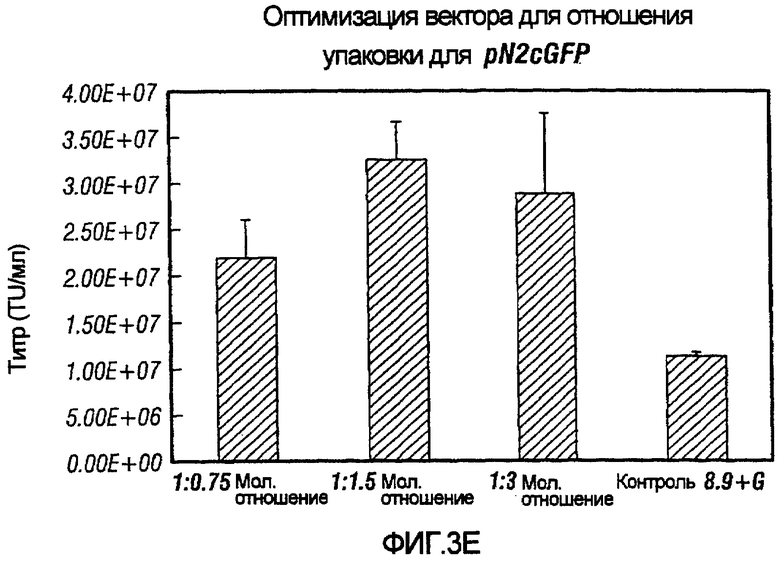
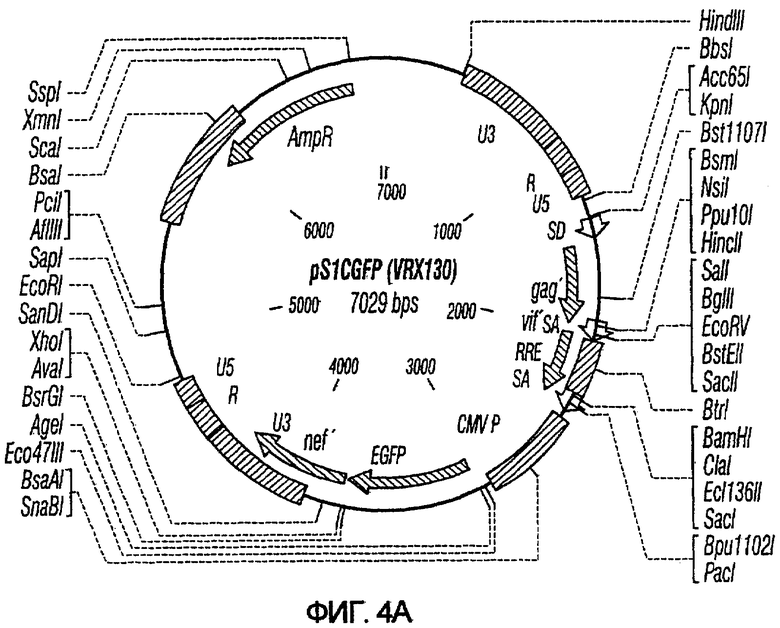
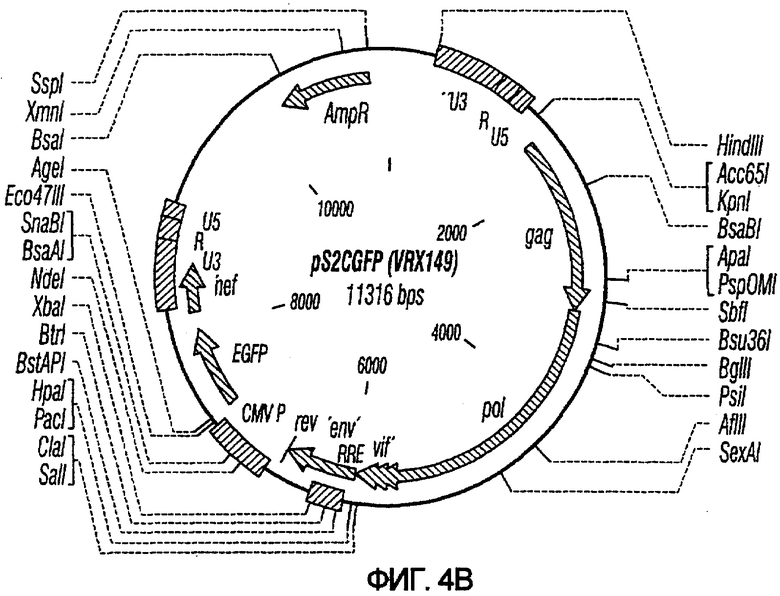
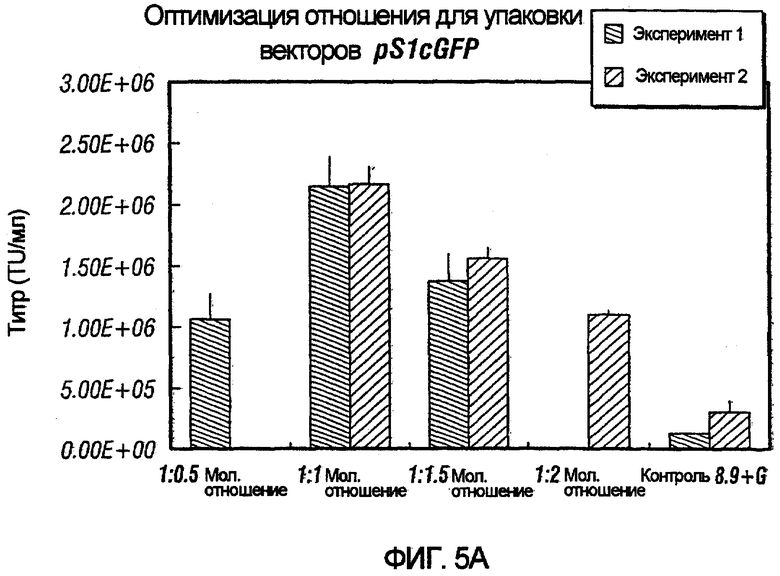
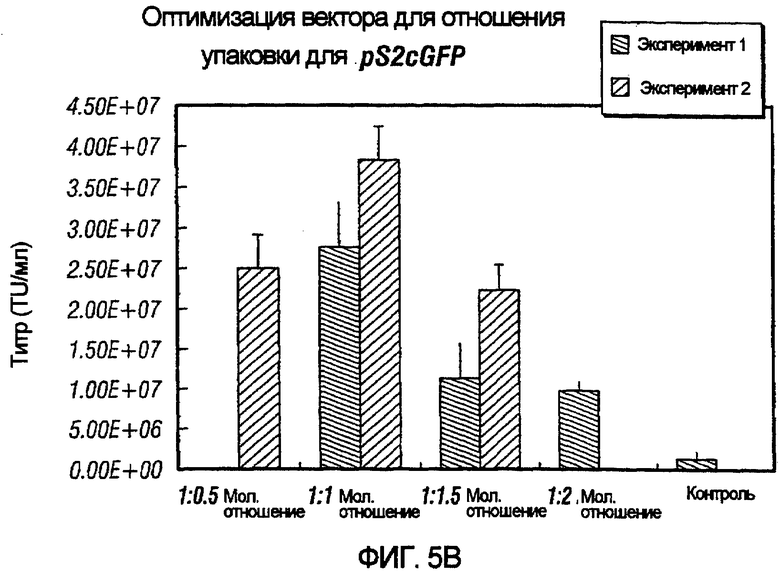
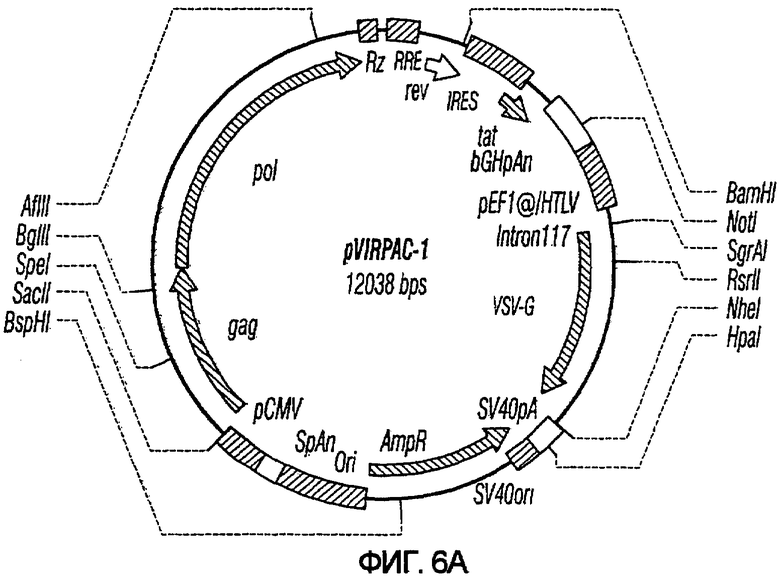
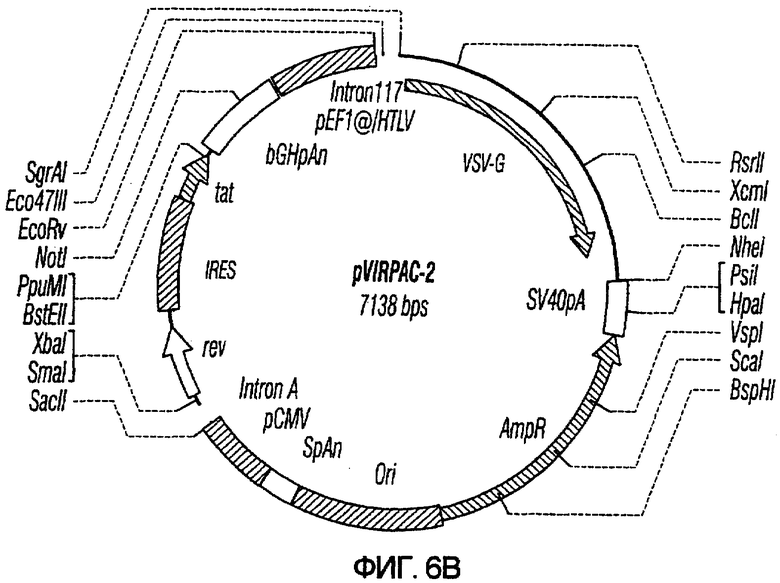
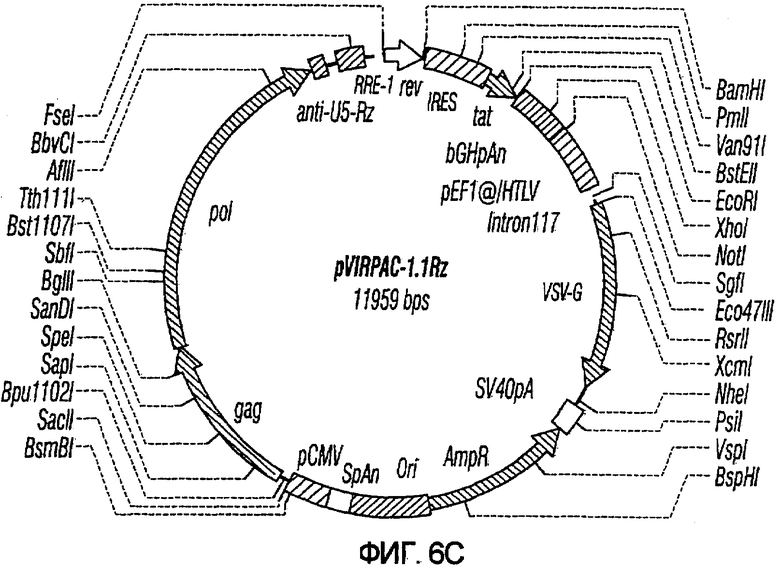
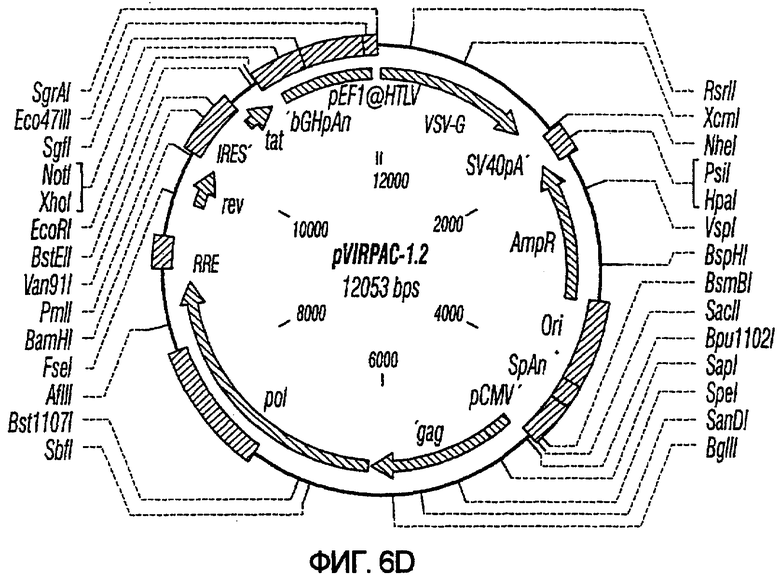
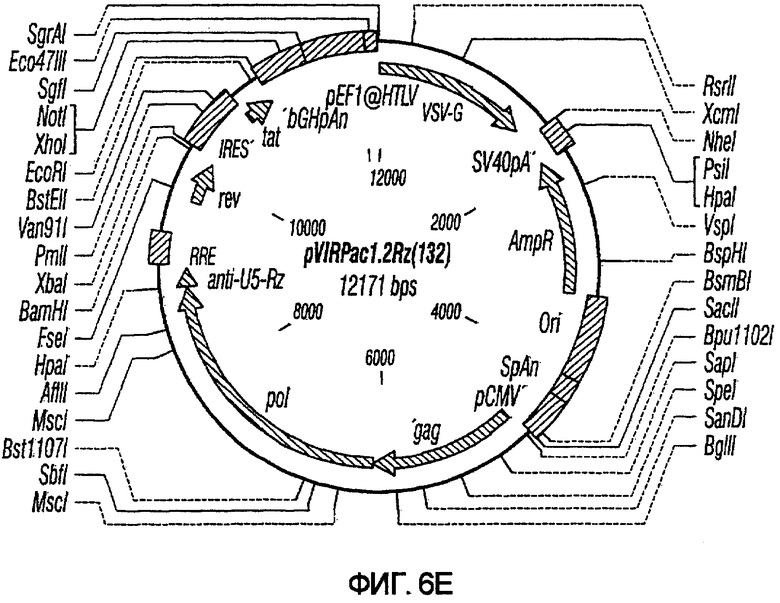
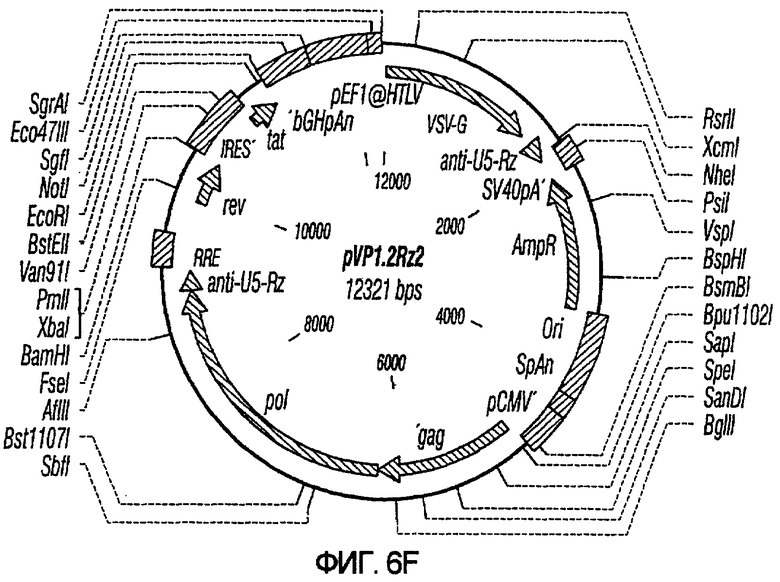
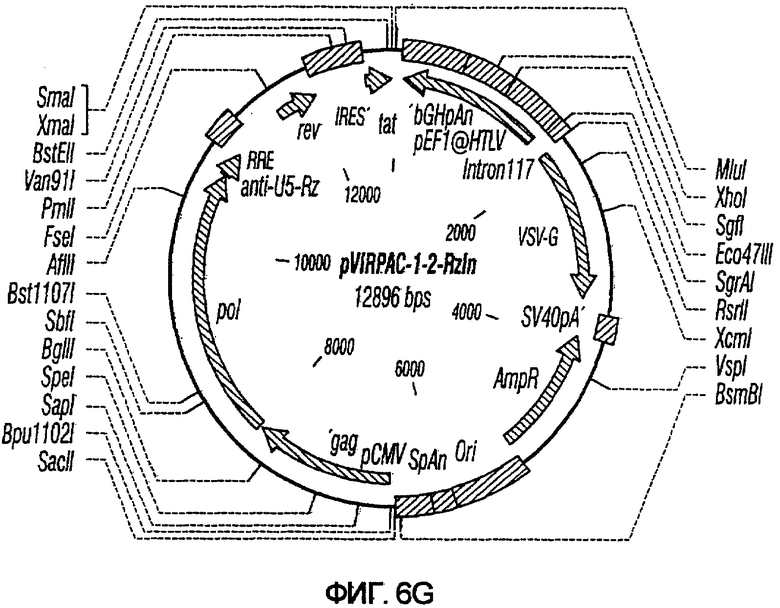
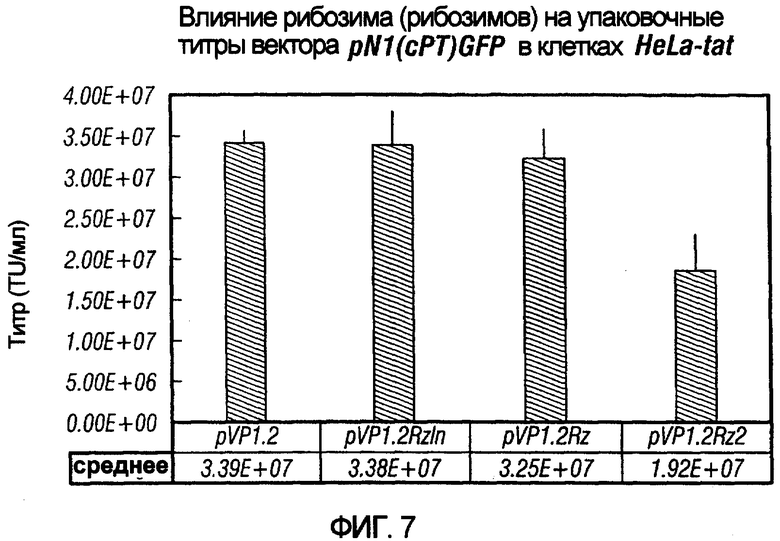
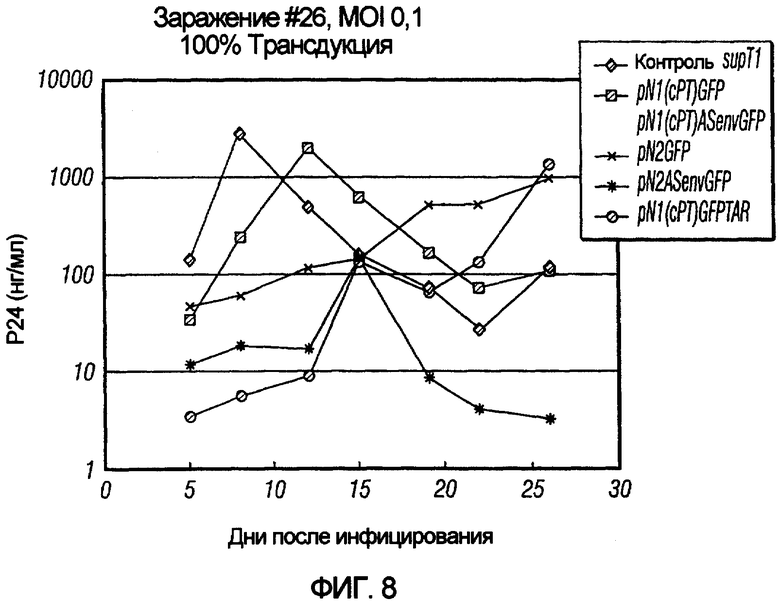
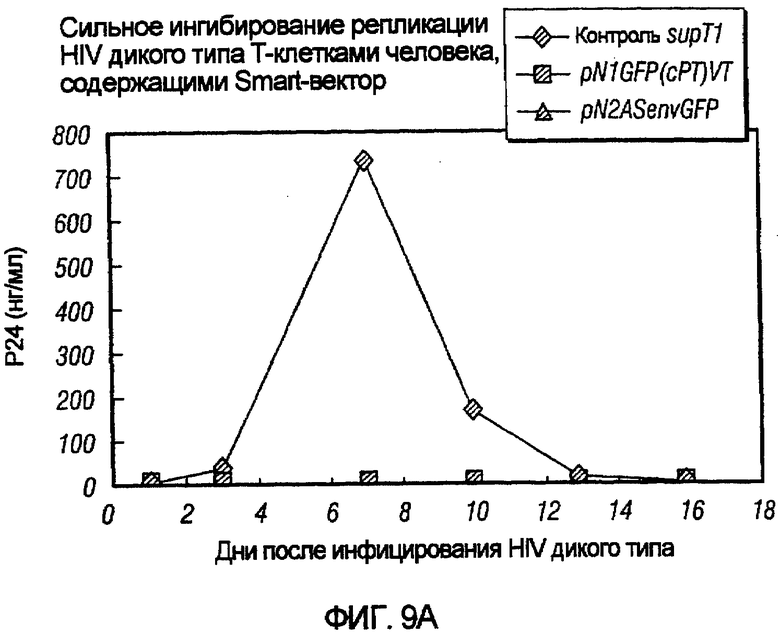
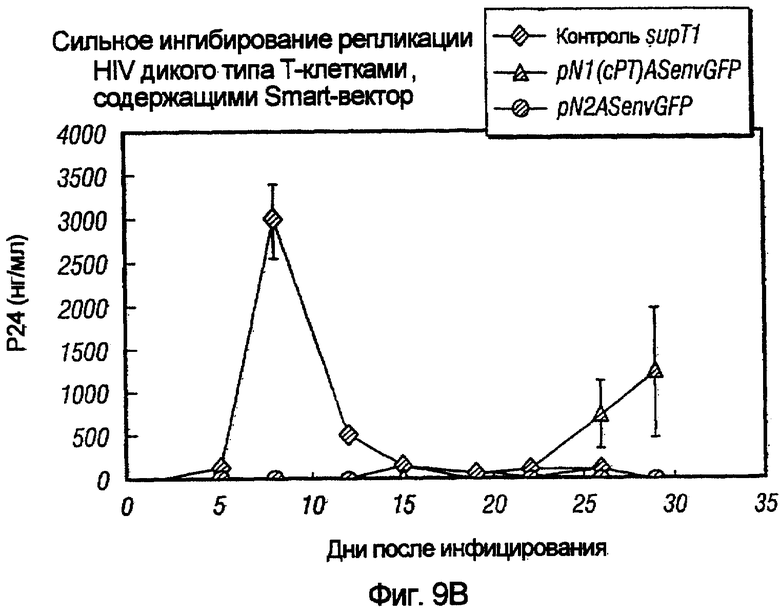
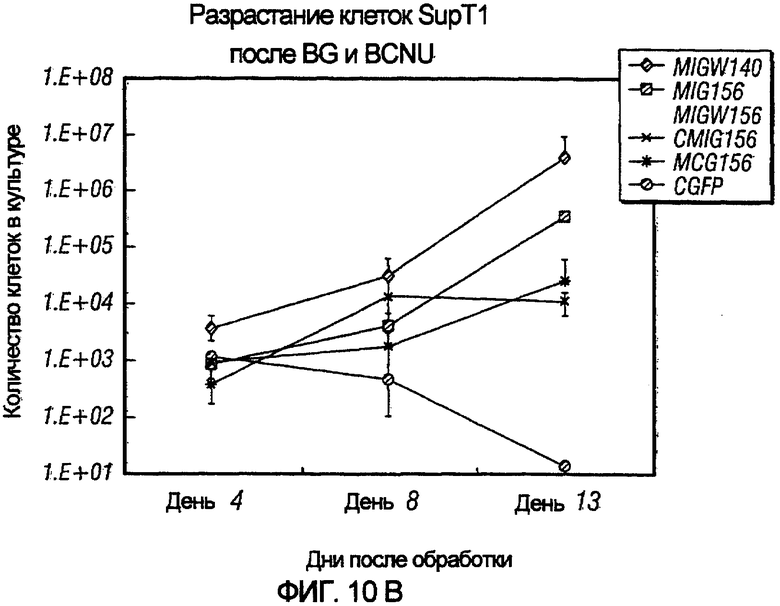
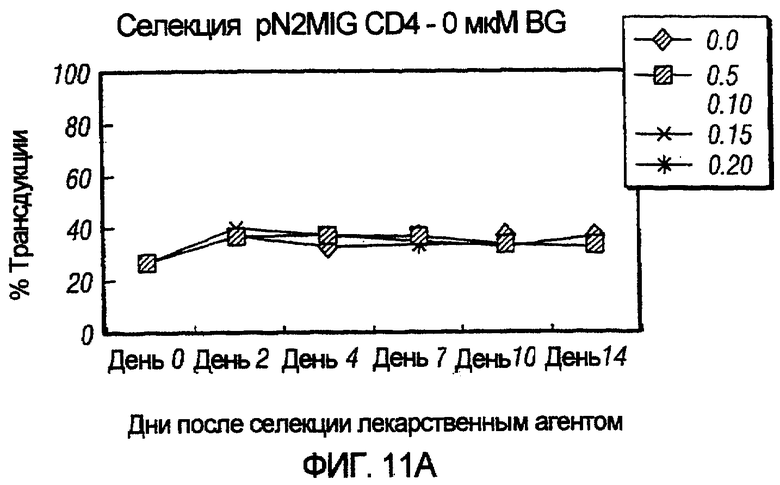
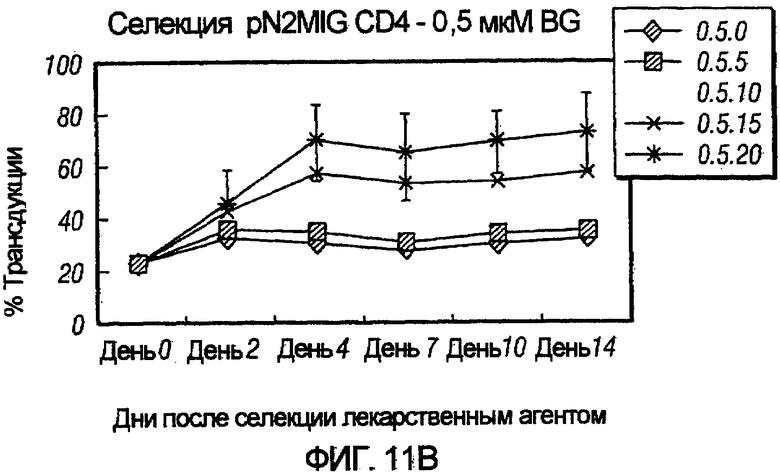
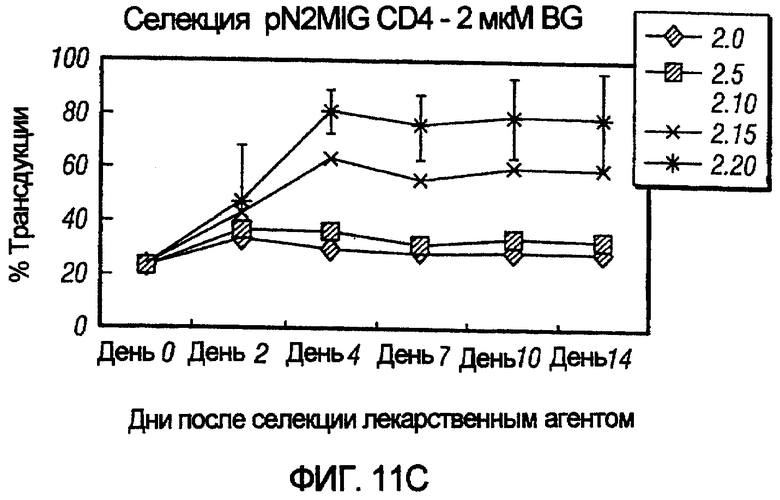
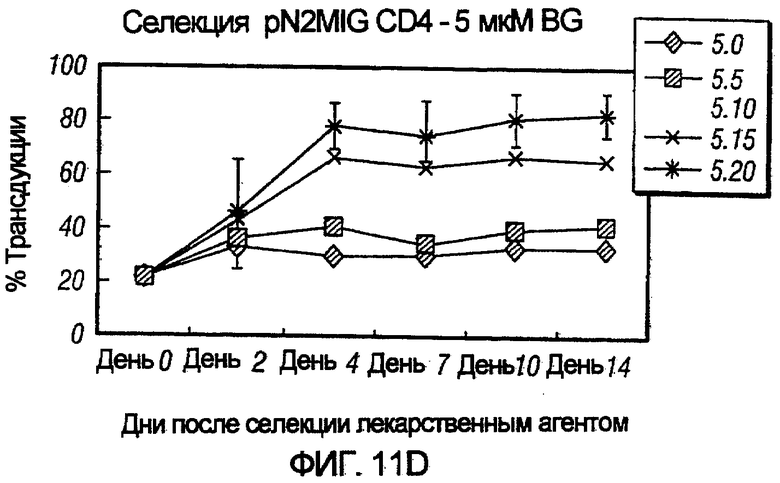
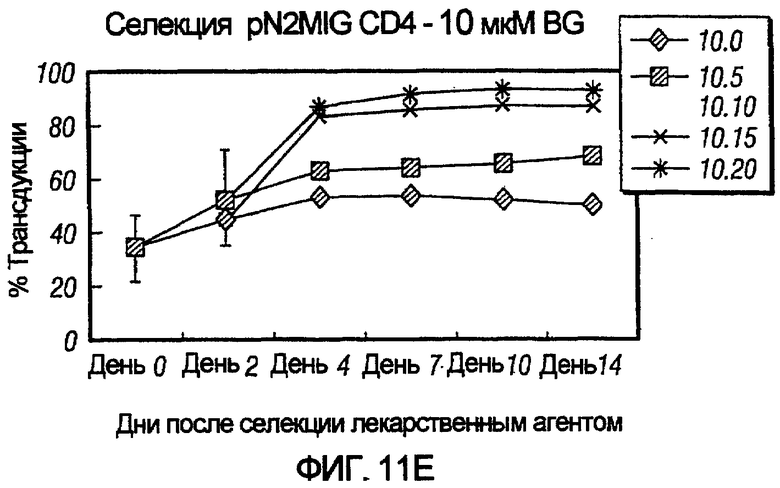
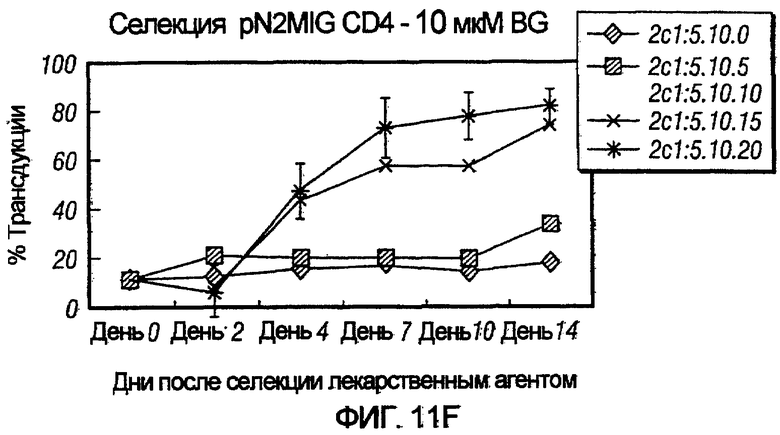
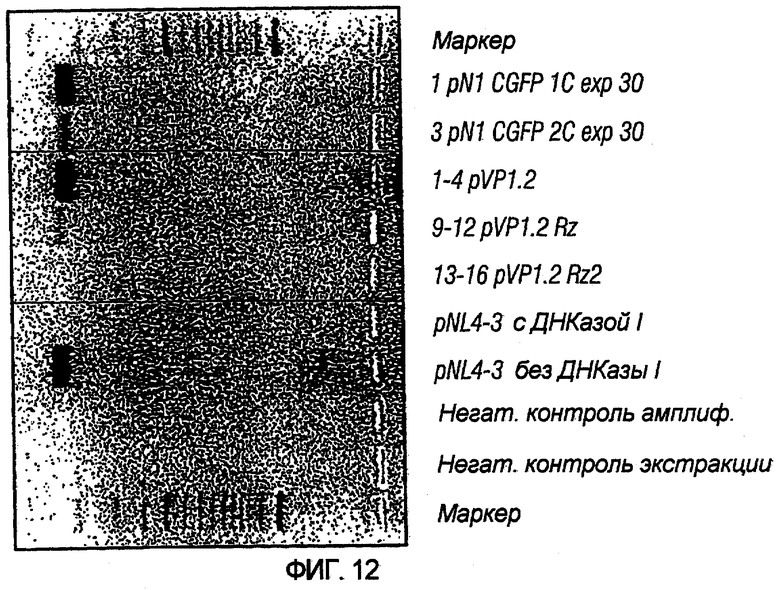
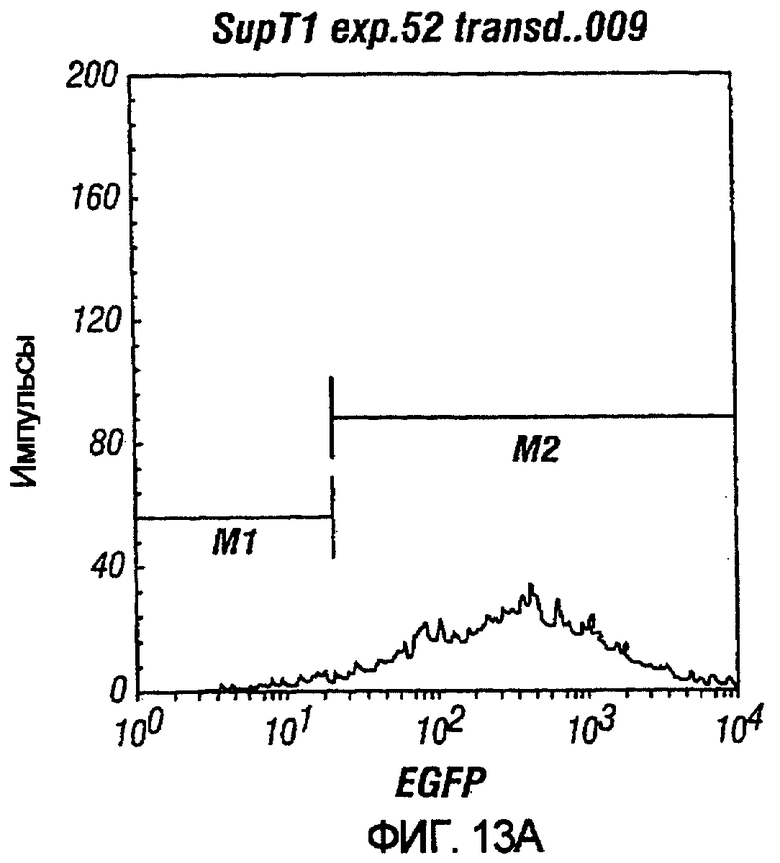
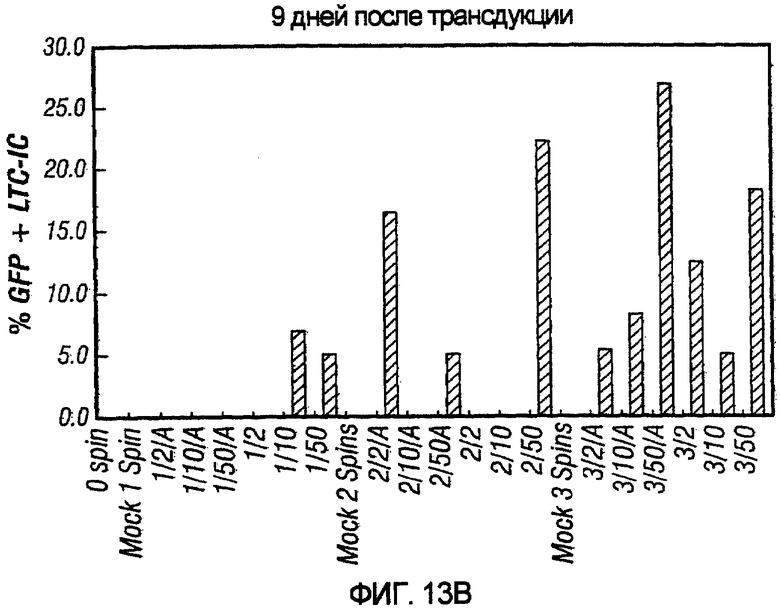
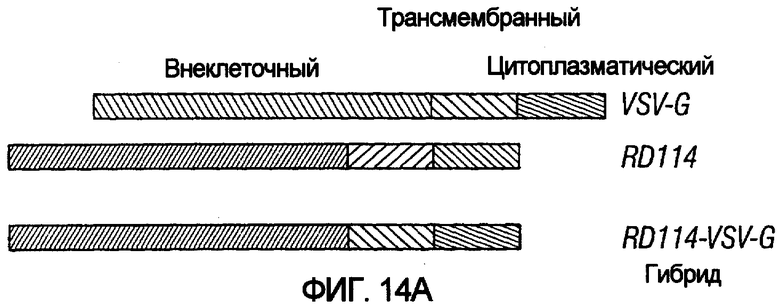

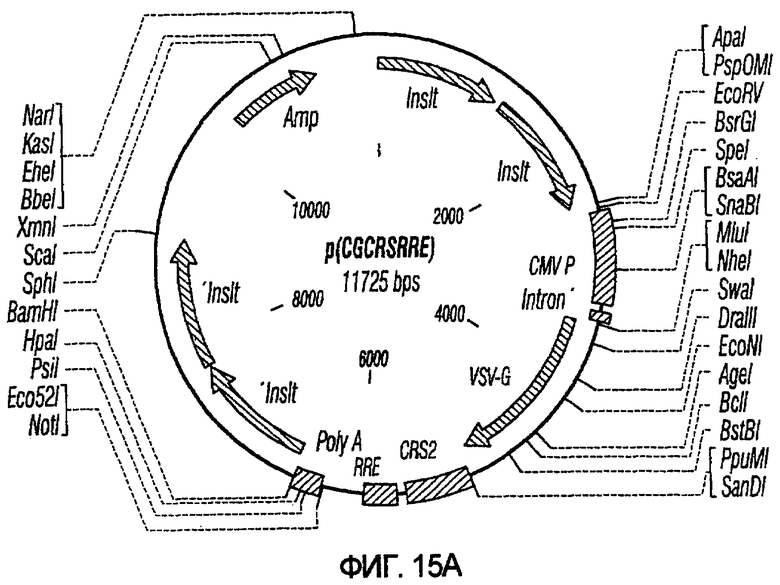
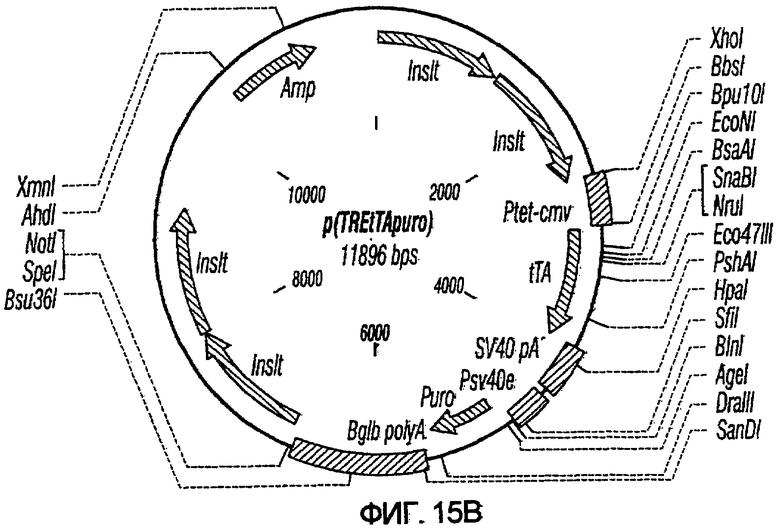
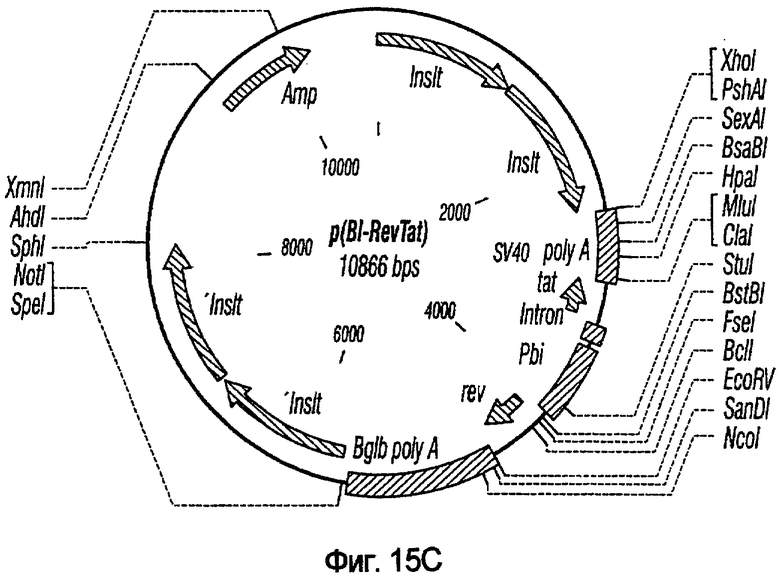
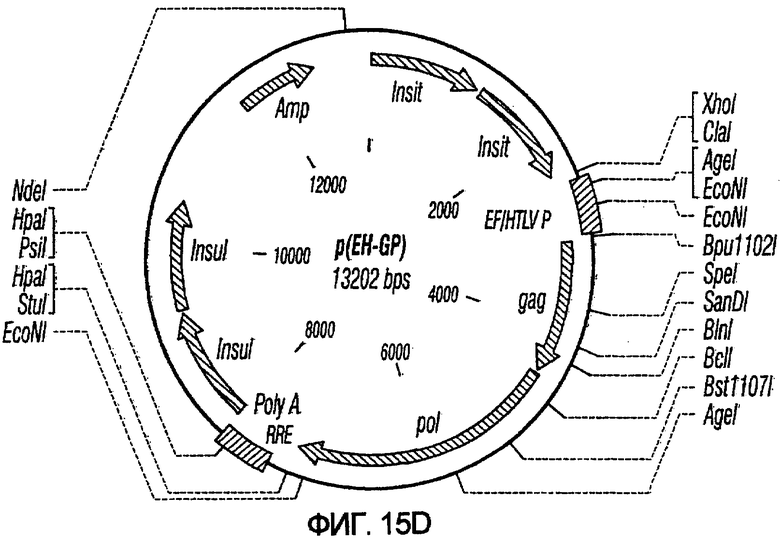
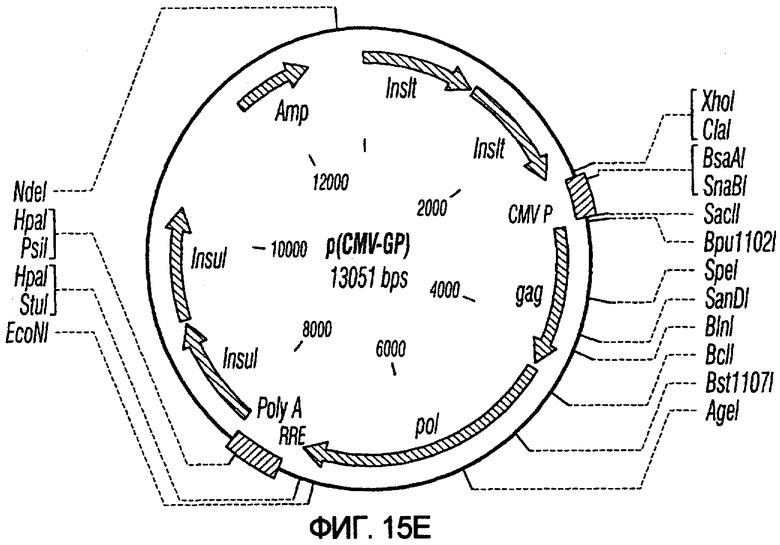
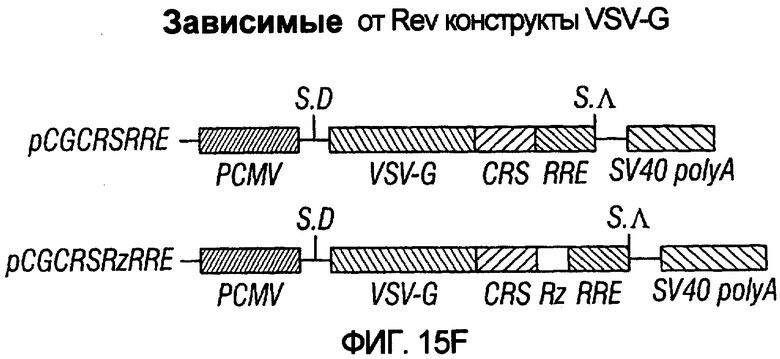
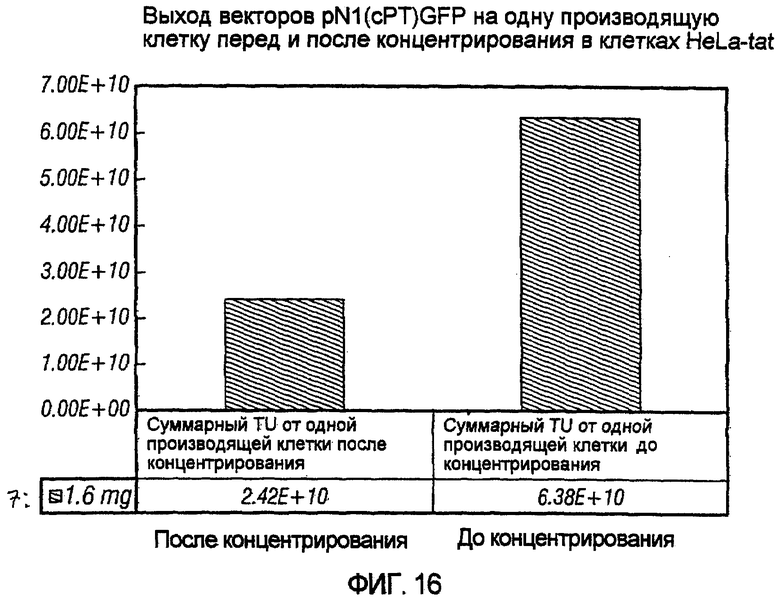

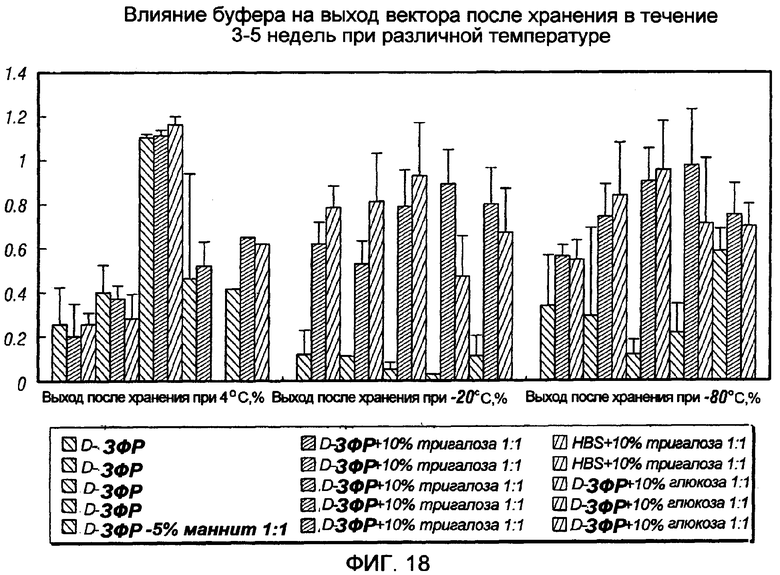
Изобретение относится к области генной инженерии. Лентивирусный вектор с зависимой от условий репликацией содержит две нуклеотидных последовательности. Первая нуклеотидная последовательность снижает вероятность образования лентивирусного вектора, способного к репликации. Вторая нуклеотидная последовательность ингибирует репликацию лентивируса дикого типа, хелперного вируса или хелперного вектора, но не ингибирует репликацию зависимых векторов, или кодирует белки, имеющие те же свойства. Также раскрыты сами нуклеотидные последовательности, способ получения таких векторов и их использование. Изобретение может быть использовано в медицине для профилактического и терапевтического лечения вирусных заболеваний, в частности HIV инфекции. 14 н. и 31 з.п. ф-лы, 18 ил.
(a) по меньшей мере первую нуклеотидную последовательность, содержащую последовательность, которая снижает вероятность образования лентивирусного вектора, способного к репликации, и
(b) одну или несколько вторых нуклеотидных последовательностей, содержащих
(i) нуклеотидную последовательность, которая ингибирует репликацию лентивируса штамма дикого типа, хелперного вируса или хелперного вектора, но не ингибирует репликацию лентивирусного вектора с зависимой от условий репликацией; или
(ii) нуклеотидную последовательность, которая кодирует белок, который ингибирует репликацию лентивируса штамма дикого типа, хелперный вектор или хелперный вирус, но не ингибирует репликацию лентивирусного вектора с зависимой от условий репликацией;
где первая нуклеотидная последовательность, которая снижает вероятность образования лентивирусного вектора, способного к репликации, содержит одну или несколько модификаций пар нуклеотидных оснований последовательности, включающих замену, инсерцию или делецию, что уменьшает вероятность событий гомологичной репликации между лентивирусной последовательностью в векторе и лентивирусной последовательностью в вирусе дикого типа, хелперным вирусом или хелперным вектором и что может привести к образованию лентивирусного вектора, способного к репликации, и замену, инсерцию или делецию вводят в любую часть лентивирусного вектора, которая, по существу, гомологична последовательности лентивируса дикого типа, с тем, чтобы происходили события гомологичной рекомбинации, если последовательность не подвергалась модификациям, и, необязательно, первая нуклеотидная последовательность, которая снижает вероятность события гомологичной рекомбинации, содержит по меньшей мере один вырожденный кодон вместо кодона, предпочтительно использующегося в клетке-хозяине, где указанная одна или несколько первых нуклеотидных последовательностей представляют собой один или несколько рибозимов или антисмысловых последовательностей или кодируют один или несколько рибозимов или антисмысловых последовательностей для профилактического и терапевтического лечения заболеваний, особенно вирусной инфекции и, в частности, HIV инфекции, для лечения инфекционных заболеваний, рака и других заболеваний, имеющих отношение к генетике, и для методов диагностики.
(a) является химерным вектором, содержащим последовательности ретровируса и одного или несколько других вирусов, где, необязательно, ретровирус содержит HIV и, необязательно, HIV представляет собой HIV-1 или HIV-2, и, необязательно, последовательности с связаны с последовательностями модифицированного штамма (неприродного типа) аденовируса, аденоассоциированного вируса, вируса рода Alphaviridae, вируса рода Flaviviridae, вируса рода Hepadnaviridae, вируса рода Papovaviridae, вируса рода Parvoviridae, вируса рода Herpesviridae, вируса рода Poxiviridae, вируса рода Paramyxoviridae, вируса рода Rhabdoviridae, вируса рода Retroviridae, онкоретровируса, спумаретровируса или лентиретровируса; или
(b) содержит последовательность ретровируса, где, необязательно, ретровифус содержит HIV, и, необязательно, HIV представляет собой HIV-1 или HIV-2, или их сочетание.
18 Лентивирусный вектор с пониженной, зависимой от условий репликацией, содержащий
(a) первую нуклеотидную последовательность, функционально связанную с промотором, которая снижает вероятность образования вектора, способного к репликации, где последовательность нуклеиновой кислоты, которая снижает вероятность образования вектора, способного к репликации, снижает вероятность рекомбинации между одним или несколькими гомологичными областями лентивирусного вектора с зависимой от условий репликацией, и геномом лентивируса, способного к репликации, хелперным вектором или хелперным вирусом, следовательно, снижает вероятность рекомбинантного события в клетке-хозяине, что модифицирует лентивирусный вектор с зависимой от условий репликацией в форму, способную к репликации; и
(b) вторую неуклеотидную последовательность, которая ингибирует или предотвращает репликацию или упаковку генома лентивируса с зависимой от условий репликацией в клетке-хозяине, таким образом давая преимущество селективной репликации или упаковки лентивирусному вектору с зависимой от условий репликацией по сравнению с репликацией или упаковкой генома лентивируса, где указанная одна или несколько первых нуклеотидных последовательностей представляют собой один или несколько рибозимов или антисмысловых последовательностей или кодируют один или несколько рибозимов или антисмысловых последовательностей, для профилактического и терапевтического лечения заболеваний, особенно вирусной инфекции, и, в частности, HIV инфекции, для лечения инфекционных заболеваний, рака и других заболеваний, имеющих отношение к генетике, и для методов диагностики.
(a) стадии
(b) стадии
(a) получение клетки-хозяина, вектора с зависимой от условий репликацией по п.1 или 18, нуклеиновой кислоты вируса штамма дикого типа, хелперного вируса или хелперного вектора, способного дополнять вектор с зависимой от условий репликацией в клетке-хозяина;
(b) временную трансфекцию, инфицирование или введение последовательности вектора с зависимой репликацией и хелперной последовательности с высокой трансфецирующей или трансдуцирующей эффективностью, необязательно, используя трансфецирующий агент;
где, необязательно, трансфицирующим агентом является фосфат кальция, реагент липидной трансфекции, или трансфекцию осуществляют электропорацией; и
(с) выделение вектора из супернатанта;
где, необязательно, выделение проводят не ранее, чем через 12 ч после трансфекции, и не позднее, чем через 7 дней после трансфекции,
и, необязательно, выделяемый вектор концентрируют, необязательно, с помощью высокоскоростного центрифугирования без осаждения или ультрацентрифугирования,
и, необязательно, выделяемый вектор дополнительно очищают с помощью по крайней мере одной, нескольких или всех стадий способа по п.26.
где, необязательно, вирусом является лентивирус и, необязательно, лентивирус представляет собой HIV.
(а) трансдукцию HSC вектором по любому из пп.1-24,
где вектор кодирует последовательность варианта О6-метилгуанин-ДНК-метилтрансферазы (MGMT), устойчивого к инактивации, опосредованной O-бензилгуанином (BG), и, необязательно, вариантом является G 156A MGMT;
(b) контактирование клеток с O-бенизлгуанином (BG) в количестве, достаточном для инактивации MGMT дикого типа, при котором не происходит инактивации варианта MGMT, устойчивого к инактивации, опосредованной O-бензилгуанином (BG);
(c) контактирование клеток с цитотоксическим алкилирующим агентом после контакта этих клеток с количеством, достаточным для индукции гибели нетрансдуцированных клеток, при котором не происходит индукции гибели трансдуцированных клеток HSC;
и, необязательно, цитотоксический алкилирующий агент представляет собой 6-гуанинметилирующий или хлорэтилирующий агент или, необязательно, цитотоксический алкилирующий агент представляет собой темозоломид, нитрозомочевину, тетразины, триазины, дакабазин, темозоломид, стрептозотоцин, прокарбазин, BCNU (1,3-бис(2-хлорэтил)-1-нитрозомочевину, CCNU (3-циклогексил-1-хлорэтил-нитрозомочевину), ACNU (1-(4-амино-2-метил-5-пиримидинил)метил-3-(2-хлор)-3-нитрозомочевину) и MeCCNU (1-(2-хлорэтил)-3-(4-метилциклогексил)-1-нитрозомочевину).
где, необязательно, клетка-хозяин содержит любое сочетание двух, трех или более векторов с зависимой от условий репликацией или хелперных векторов,
для профилактического и терапевтического лечения заболеваний, особенно вирусной инфекции и, в частности, HIV инфекции, для лечения инфекционных заболеваний, рака и других заболеваний, имеющих отношение к генетике, и для методов диагностики.
где геном ретровируса с зависимой от условий репликацией включает длинный 5' концевой повтор (LTR) и длинный 3' концевой повтор (LTR) и транскрипционная активность последовательности, ингибирующей репликацию ретровируса, находится под контролем ретровирусного LTR,
и последовательность, ингибирующая репликацию вируса, или продукт последовательности, ингибирующей репликацию вируса, ингибирует репликацию ретровируса или дает некоторые преимущества репликации генома ретровируса с зависимой от условий репликацией по сравнению с репликацией генома ретровируса дикого типа.
где раствор, буфер или носитель включает фосфатно-буферный солевой раствор (PBS) или PBS включает PBS, который смешан с 1-50%-ным раствором трегалозы в воде (1:1) или с 10%-ным раствором трегалозы в воде (1:1) так, чтобы конечная концентрация составляла 5% трегалозы; или PBS смешан с 1-50%-ным раствором глюкозы в воде (1:1) или с 10% раствором глюкозы в воде (1:1), так, чтобы конечная концентрация глюкозы составляла 5%; 20 мМ HEPES-забуференный солевой раствор, который смешан с 1-50%-ным раствором трегалозы в воде (1:1) или с 10%-ным раствором трегалозы в воде (1:1) так, чтобы конечная концентрация трегалозы составляла 5%; или PBS смешан с 1-50%-ным раствором маннита в воде (1:1) или 5%-ным раствором маннита в воде (1:1) так, чтобы конечная концентрация маннита составляла 2,5%,
или композиция находится в лиофилизируемой форме.
(i) экспрессируемый ген или последовательность, где ген или последовательность: (а) могут излечивать или предотвращать развитие заболевания или состояния или (b) могут действовать как генетический противовирусный агент, где, необязательно, генетический противовирусный агент содержит антисмысловую молекулу, РНК-ловушку, трансдоминантный мутант, токсин, модификатор или модулятор РНК или белкового сплайсинга, внешнюю вспомогательную последовательность (EGS), иммунноген, мРНК, нуклеотидную последовательность или молекулу, которая модифицирует или модулирует спласинг, конститутивный транспортный элемент (СТЕ), нуклеарный фактор импорта и экспорта, факторы интеграции ДНК, элементы, стабилизирующие или дестабилизирующие РНК, посттранскрипционные регуляторные элементы (PRE), белок, который препятствует вирусной репликации, интерферон, антитело, молекулу-лиганд или рибозим, и
(ii) нуклеотидную последовательность, которая снижает вероятность образования лентивирусного вектора, способного к репликации, содержащего одну или несколько замен, инсерций или делеций пар нуклеотидных оснований, что уменьшает вероятность событий гомологичной рекомбинации между последовательностью лентивируса в векторе и последовательностью лентивируса дикого типа, хелперным вирусом или хелперным вектором, и что может привести к образованию лентивирусного вектора, способного к репликации.
| DROPULIC В | |||
| at al., «A conditionally replicating HIV-1 vector interferes with wild-type HIV-1 replication and spread»; Proc Nati Acad Sci USA | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| WO 9720060 (UNIV JOHNS HOPKINS MED), 05.06.1997. | |||

