Область, к которой относится изобретение
Настоящее изобретение относится к нуклеиновокислотным векторам, содержащим гены, необходимые для продуцирования ретровируса, и к их применению. Настоящее изобретение также относится к способам продуцирования дефектных по репликации частиц ретровирусного вектора, содержащих описанные здесь нуклеиновокислотные векторы.
Предшествующий уровень техники
В генотерапии, генетический материал доставляют в эндогенные клетки индивидуума, нуждающегося в лечении. Этот генетический материал позволяет вводить новые гены индивидууму или дополнительные копии уже существующих генов или различные аллели или варианты генов, присутствующих у индивидуума. Системы вирусных векторов были предложены как средства для эффективной доставки генов в методах генотерапии (Verma and Somia (1997) Nature 389: 239-242).
В частности, эти вирусные векторы были получены на основе членов семейства ретровирусов, что обусловлено их способностью интегрировать полезную генетическую нагрузку в геном хозяина. Ретровирусные векторы были сконструированы так, чтобы они сохраняли основные белки, необходимые для упаковки и доставки ретровирусного генома, но при этом удаляли любые неосновные вспомогательные белки, включая белки, ответственные за развитие заболевания. Примерами ретровирусных векторов являются лентивирусные векторы, такие как векторы, полученные на основе вируса иммунодефицита человека типа 1 (ВИЧ-1), которые широко применяются благодаря их способности интегрироваться в непролиферирующиеся клетки.
В настоящее время большинство вирусных векторов продуцируют путем временной ко-трансфекции вирусных генов в клеточную линию хозяина. Вирусные гены вводят с использованием бактериальных плазмид, которые присутствуют в клетках-хозяевах в течение лишь ограниченного периода времени, поскольку вирусные гены сохраняются на плазмидах и не интегрируются в геном. Так, например, временно трансфецированный генетический материал не проходит стадию образования последующих генераций в процессе деления клеток.
Однако применение современных методов временной трансфекции связано с рядом проблем, таких как вариабельность между партиями, высокая стоимость реагентов для трансфекции и трудности проведения контроля качества (см. Segura et al. (2013) Expert Opin. Biol. Ther. 13(7): 987-1011). Сам процесс трансфекции также является очень трудоемким и трудно масштабируемым. При этом также возникают проблемы с удалением плазмидных примесей, которые появляются в процессе получения вектора (см. Pichlmair et al. (2007) J. Virol. 81(2): 539-47).
Следовательно, целью настоящего изобретения является разработка усовершенствованного способа временной трансфекции, который позволял бы устранить один или более недостатков, ассоциированных с применением уже существующих методов.
Сущность изобретения
Авторами настоящего изобретения был разработан новый способ продуцирования ретровирусных векторов, включающий использование нуклеиновокислотных векторов, содержащих ориджин репликации животных, не являющихся млекопитающими, и способных сохранять по меньшей мере 25 тысяч пар оснований (т.п.о.) ДНК, таких как бактериальные искусственные хромосомы, включающие все ретровирусные гены, необходимые для продуцирования ретровирусного вектора. Для проведения современных методов временной трансфекции необходимо использовать 3-4 отдельных плазмиды, несущих различные компоненты, необходимые для введения полученного ретровируса в клетку-хозяина, но эти методы требуют много времени, и при их осуществлении возникают проблемы, связанные с давлением отбора. Способ, предложенный авторами настоящего изобретения, включает получение всех основных ретровирусных генов на одной конструкции, которая затем может быть введена в клетку-хозяина, что позволяет уменьшить количество материала, необходимого для переноса вирусного вектора в клетку-хозяина. Следовательно, это позволяет снизить стоимость продуктов, поскольку в данном случае используется один вектор, в отличие от известных методов, в которых используется множество плазмидных векторов.
Использование нуклеиновокислотного вектора, содержащего ориджин репликации животных, не являющихся млекопитающими, и способного сохранять по меньшей мере 25 т.п.о. ДНК (то есть, крупной ДНК-конструкции), имеет несколько преимуществ. В первом случае, векторы могут быть сначала модифицированы в клетках животных, не являющихся млекопитающими (например, в микробных клетках, таких как бактериальные клетки), но не в клетках-хозяевах млекопитающих, что позволяет существенно облегчить применение этих векторов (например, бактериальные искусственные хромосомы могут быть сначала модифицированы в E. coli). После получения нуклеиновокислотного вектора, он может быть введен в клетку-хозяина, такую как клетка-хозяин млекопитающего, для продуцирования ретровирусного вектора.
Применение нуклеиновокислотных векторов согласно изобретению дает преимущества при получении ретровирусных векторов.
Следовательно, в своем первом аспекте, настоящее изобретение относится к нуклеиновокислотному вектору, содержащему ориджин репликации животных, не являющихся млекопитающими, и способному сохранять по меньшей мере 25 тысяч пар оснований (т.п.о.) ДНК, где указанный нуклеиновокислотный вектор отличается тем, что он содержит последовательности нуклеиновой кислоты ретровируса, кодирующие:
белки gag и pol, и
белок env или его функциональный аналог.
В соответствии с другим своим аспектом, настоящее изобретение относится к определенному здесь нуклеиновокислотному вектору для применения в способе продуцирования частиц ретровирусного вектора.
В соответствии с другим своим аспектом, настоящее изобретение относится к способу продуцирования дефектной по репликации частицы ретровирусного вектора, где указанный способ включает:
(a) введение определенного здесь нуклеиновокислотного вектора в культуру клеток-хозяев млекопитающих; и
(b) культивирование клеток-хозяев млекопитающих в условиях, способствующих продуцированию дефектной по репликации частицы ретровирусного вектора.
В соответствии с другим своим аспектом, настоящее изобретение относится к дефектной по репликации частице ретровирусного вектора, полученной описанным здесь способом.
Краткое описание чертежей
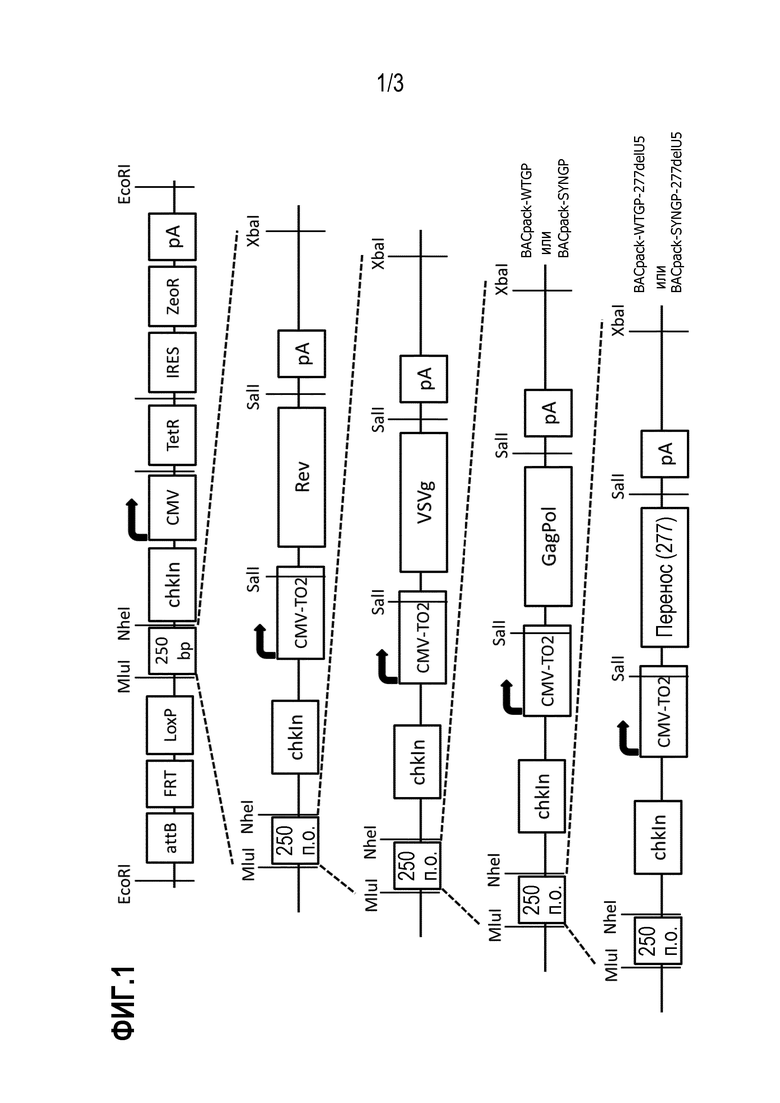
Фигура 1: Постадийный способ конструирования BACpack-WTGP-277delU5 и BACpack-SYNGP-277delU5.
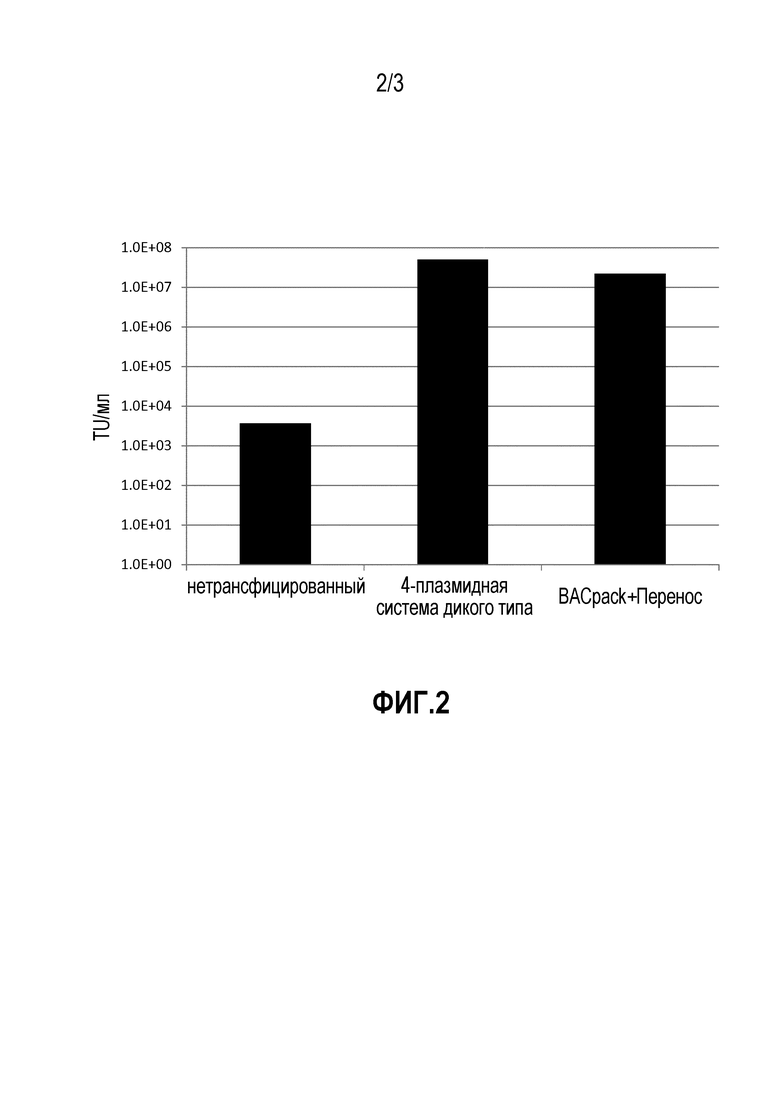
Фигура 2: Сравнение титров вирусов, полученных как описано в Примере 2. 106 клеток HEK293T высевали в 6-луночный планшет. На следующий день, прилипшие клетки трансфецировали с использованием ПЭИ в соответствии с инструкциями производителей. Клетки трансфецировали всего 4 мкг лентивирусных упаковывающих конструкций дикого типа (WT), состоящих из pMDL.gp (GagPol), pMD.G (VSVg), pK-Rev (Rev) и pCCL.277 (вектора для переноса GFP) или 2 мкг BACpack (одной BAC-конструкции, содержащей GagPol, VSVg и Rev) плюс 2 мкг вектора для переноса eGFP на отдельной плазмиде.
Через 48 часов после трансфекции, супернатант собирали, фильтровали через 0,22 мкм-фильтр и хранили при -80°C минимум 4 часа. Клетки HEK293T высевали для трансдукции при плотности 105 клеток на лунку в 24-луночном планшете. На следующий день, вирусный супернатант вводили в клетки в виде серийных разведений полибреном в конечной концентрации 8 мкг/мл. Через 3 дня после трансдукции, клетки собирали путем обработки трипсином и анализировали на GFP с помощью FACS-анализа. Титр вируса вычисляли как единицы трансдукции (ЕТ)/мл по следующему уравнению:
(GFP-позитивные клетки/100) × коэффициент разведения × число трансдукцированных клеток.
Титры вирусов сравнивали на гистограмме. Все инкубирования проводили при 37°C и в 5% CO2. Используемой средой является среда DMEM, в которую были добавлены 10% FBS и 1 мкг/мл доксициклина в BACpack+образец для переноса.
Фигура 3: Временная трансфекция BACpack в прилипших клетках HEK293T. Клетки HEK293T временно трансфецировали BACpackWTGP-277delU5, BACpackSYN-277delU5 или стандартной 4-плазмидной системой с применением кальций-фосфатного метода. Через 16 часов после трансфекции, среду+Dox индуцировали в течение 24 часов путем добавления 1 мкг/мл доксициклина. Вирусный супернатант собирали через 48 часов после трансфекции, фильтровали через 0,22 мкм-фильтр и титровали путем трансдукции клеток HEK293T. GFP-позитивные трансдуцированные клетки использовали для вычисления единиц трансдукции (ЕТ/мл).
Подробное описание изобретения
Определения
Если это не оговорено особо, то все используемые здесь технические и научные термины используются в своем общепринятом смысле, полятном специалисту в области, к которой относится изобретение. Все описанные здесь патенты и публикации полностью включены в настоящее описание посредством ссылки.
Термин «включающий» охватывает понятия «содержащий» или «состоящий из», например, композиция, «включающая» Х, может состоять исключительно из Х или может включать какие-либо другие компоненты, например, X+Y.
Термин «по существу, состоящий из» ограничивает объем признаков, относящихся к конкретным материалам или стадиям, и к тем материалам или стадиям, которые не оказывают значительного влияния на основное(ые) свойство(а) заявленного признака.
Термин «состоящий из» исключает присутствие любого(ых) дополнительного(ых) компонента(ов).
Термин «приблизительно», если он относится к численной величине Х, означает, например, Х ± 10%, 5%, 2% или 1%.
Термин «вектор» или «нуклеиновокислотный вектор» означает носитель, способный искусственно переносить чужеродный (то есть, экзогенный) генетический материал в другую клетку, где он может реплицироваться и/или экспрессироваться. Примерами векторов являются нуклеиновокислотные векторы животных, не являющихся млекопитающими, такие как бактериальные искусственные хромосомы (BAC), дрожжевые искусственные хромосомы (YAC), искусственные хромосомы, происходящие от P1 (PAC), космиды или фосмиды.
Другими примерами векторов являются вирусные векторы, такие как ретровирусные векторы и лентивирусные векторы, которые представляют особый интерес для авторов настоящего изобретения. Лентивирусные векторы, такие как векторы, полученные на основе вируса иммунодефицита человека типа 1 (ВИЧ-1), широко применяются благодаря их способности интегрироваться в непролиферирующиеся клетки. Вирусные векторы могут быть сделаны дефицитными по репликации путем расщепления вирусного генома на отдельные части, например, путем их помещения на отдельные плазмиды. Так, например, так называемая первая генерация лентивирусных векторов, разработанная в Институте биологических исследований Солка, была сконструирована как трехплазмидная экспрессионная система, состоящая из упаковывающего экспрессионного кластера, оболочечного экспрессионного кластера и векторного экспрессионного кластера. «Упаковывающая плазмида» содержит полноразмерные последовательности gag-pol, регуляторные последовательности (tat и rev) и вспомогательные последовательности (vif, vpr, vpu, nef). «Оболочечная плазмида» вместо нативного оболочечного белка ВИЧ-1 содержит гликопротеин вируса везикулярного стоматита (VSVg) под контролем промотора цитомегаловируса (CMV). Третья плазмида («плазмида для переноса») содержит длинные концевые повторы (LTR), инкапсулирующую последовательность (ψ), последовательность Rev-отвечающего элемента (RRE) и промотор CMV для экспрессии трансгена в клетке-хозяине.
Вторая генерация лентивирусного вектора была охарактеризована путем делеции вирулентных последовательностей vpr, vif, vpu и nef. Упаковывающий вектор был укорочен до генов gag, pol, tat и rev, что повышало безопасность системы.
Для усовершенствования лентивирусной системы были сконструированы векторы третьей генерации путем удаления гена tat из упаковывающей конструкции и инактивации LTR в вирусном кластере, что позволяет решить проблемы, связанные с эффектами инсерционного мутагенеза.
Различные лентивирусные генерации были описаны в следующих работах: Первая генерация: Naldini et al. (1996) Science 272(5259): 263-7; Вторая генерация: Zufferey et al. (1997) Nat. Biotechnol. 15(9): 871-5; Третья генерация: Dull et al. (1998) J. Virol. 72(11): 8463-7, где все указанные работы во всей своей полноте вводятся в настоящее описание посредством ссылки. Описание конструирования лентивирусных векторов можно найти в публикациях Sakuma et al. (2012) Biochem. J. 443(3): 603-18 и Picanço-Castro et al. (2008) Exp. Opin. Therap. Patents 18(5):525-539.
Термин «ориджин репликации животных, не являющихся млекопитающими» означает последовательность нуклеиновой кислоты, в которой инициируется репликация, и которая происходит от животного, не являющегося млекопитающим. Это позволяет нуклеиновокислотным векторам согласно изобретению стабильно реплицироваться и сегрегироваться во всех эндогенных хромосомах в подходящей клетке-хозяине (например, в микробной клетке, такой как бактериальная или дрожжевая клетка), что будет способствовать переносу этой последовательности в потомство клетки-хозяина, за исключением случая, когда клеткой-хозяином является клетка-хозяин млекопитающего. В клетках-хозяевах млекопитающих, нуклеиновокислотные векторы, содержащие ориджины репликации животных, не являющиеся млекопитающими, либо будут интегрироваться в эндогенные хромосомы клеток-хозяев млекопитающих, либо они будут исчезать после репликации в клетках-хозяевах млекопитающих. Так, например, нуклеиновокислотные векторы, содержащие ориджины репликации животных, не являющихся млекопитающими, такие как бактериальные искусственные хромосомы (BAC), искусственные хромосомы, происходящие от P1 (PAC), космиды или фосмиды, будут стабильно реплицироваться и сегрегироваться во всех эндогенных хромосомах в бактериальных клетках (таких как E. coli), однако, при их введении в клетки-хозяева млекопитающих, BAC, PAC, космида или фосмида либо будут интегрироваться в клетку-хозяина млекопитающего, либо будут исчезать после репликации в клетках-хозяевах млекопитающих. Дрожжевые искусственные хромосомы (YAC) будут стабильно реплицироваться и сегрегироваться во всех эндогенных хромосомах в дрожжевых клетках, однако, при их введении в клетки-хозяева млекопитающих, YAC либо будет интегрироваться в клетку-хозяина млекопитающего, либо будет исчезать после репликации в клетках-хозяевах млекопитающих. Следовательно, в этом контексте изобретения, нуклеиновокислотные векторы согласно изобретению действуют как резервуары ДНК (то есть, генов, играющих важную роль в продуцировании ретровируса), которые могут быть легко перенесены в клетки млекопитающих для получения частиц ретровирусного вектора. Примерами ориджинов репликации животных, не являющихся млекопитающими, являются бактериальные ориджины репликации, такие как oriC, oriV или oriS; вирусные ориджины репликации, такие как ориджин репликации SV40; или дрожжевые ориджины репликации, также известные как автономно реплицирующиеся последовательности (элементы ARS).
Нуклеиновокислотные векторы согласно изобретению содержат ориджин репликации животных, не являющихся млекопитающими и способны сохранять по меньшей мере 25 тысяч пар оснований (т.п.о.) ДНК. В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор способен сохранять по меньшей мере 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 210, 220, 230, 240, 250, 260, 270, 280, 290, 300, 310, 320, 330, 340 или 350 тысяч пар оснований ДНК. Следует отметить, что словосочетание «способность сохранять» употребляется в своем общепринятом смысле и означает, что верхний предел размеров вставки нуклеиновокислотного вектора должен составлять не ниже заявленного размера (то есть, не менее, чем 25 т.п.о. ДНК).
Целью настоящего изобретения является включение генов, играющих важную роль в упаковке ретровирусов в одной конструкции (то есть, в нуклеиновокислотном векторе). Следовательно, нуклеиновокислотные векторы согласно изобретению должны обладать способностью сохранять крупные вставки ДНК. Во избежание неясностей следует отметить, что термины «нуклеиновокислотные векторы» или «искусственные хромосомы» не означают природные бактериальные плазмиды (например, такие как плазмиды, используемые в настоящее время в методах временной трансфекции), поскольку они не способны сохранять по меньшей мере 25 т.п.о. ДНК. Максимальный размер вставки, которую может содержать плазмида, составляет приблизительно 15 т.п.о. Такие нуклеиновокислотные векторы также не являются бактериофагами, которые обычно сохраняют лишь вставки размером максимум 5-11 т.п.о. Поэтому, в одном из вариантов осуществления изобретения, нуклеиновокислотный вектор согласно изобретению не является плазмидой, бактериофагом или эписомой.
Термин «эндогенные хромосомы» означает геномные хромосомы, присутствующие в клетке-хозяине до генерирования или введения экзогенного нуклеиновокислотного вектора, такие как бактериальная искусственная хромосома.
Используемые здесь термины «трансфекция», «трансформация» и «трансдукция» могут употребляться для описания встраивания вектора животного, не являющегося млекопитающим, или вирусного вектора в клетку-мишень. Встраивание вектора обычно называется трансформацией бактериальных клеток и трансфекцией эукариотических клеток, хотя встраивание вирусного вектора может также называться трансдукцией. Специалисту в данной области известны различные часто применяемые методы не-вирусной трансфекции, которые включают, но не ограничиваются ими, физические методы (например, электропорацию, сжатие клеток, обработку ультразвуком, оптическую трансфекцию, слияние протопластов, конъюгирование, обработку магнитными частицами, «выстреливание» генов или бомбардировку частицами), химические методы (например, использование фосфата кальция, в высокой степени разветвленных органических соединений или катионных полимеров) или методы с использованием катионных липидов (например, липофекцию). Для проведения множества методов трансфекции необходимо контактирование растворов плазмидной ДНК с клетками, которые затем культивируют.
Термин «промотор» означает последовательность, которая запускает экспрессию гена. Для запуска экспрессии на высоком уровне, предпочтительно, использовать высокоэффективный промотор, такой как не-ретровирусный высокоэффективный промотор. Примерами подходящих промоторов могут быть такие промоторы, как предранный промотор человеческого цитомегаловируса (CMV), промотор фокус-образующего вируса селезенки (SFFV), промотор вируса саркомы Рауса (RSV) или промотор человеческого фактора элонгации 1-альфа (pEF).
Термин «polyA-сигнал» означает последовательность сигнала полиаденилирования, расположенную, например, у 3'-конца трансгена, что позволяет факторам хозяина присоединять полиаденозиновый (polyA) хвост к концу растущей мРНК в процессе транскрипции. polyA-хвост представляет собой фрагмент, содержащий до 300 аденозиновых рибонуклеотидов, которые защищают мРНК от ферментативного разложения, а также облегчают трансляцию. В соответствии с этим, нуклеиновокислотные векторы согласно изобретению могут включать последовательность polyA-сигнала, такую как последовательность polyA-сигналов человеческого бета-глобина или кроличьего бета-глобина, ранние или поздные polyA-сигналы обезьяньего вируса 40 (SV40), polyA-сигнал человеческого инсулина или polyA-сигнал бычьего гормона роста. В одном из вариантов осуществления изобретения, последовательность polyA-сигнала представляет собой polyA-сигнал человеческого бета-глобина.
Термин «интронная последовательность» означает нуклеотидную последовательность, которая удаляется из конечного генного продукта посредством РНК-сплайсинга. Было показано, что использование интрона, расположенного ниже энхансерной/промоторной области и выше кДНК-вставки, повышает уровень экспрессии гена. Повышение уровня экспрессии зависит от конкретной кДНК-вставки. В соответствии с этим, нуклеиновокислотный вектор согласно изобретению может включать такие интроны, как интрон человеческого бета-глобина, интрон кроличьего бета-глобина II или химерный интрон человеческого бета-глобина-иммуноглобулина. В одном из вариантов осуществления изобретения, интрон представляет собой интрон человеческого бета-глобина и/или интрон кроличьего бета-глобина II.
Термин «упаковывающая клеточная линия» означает клеточную линию, способную экспрессировать белок gag и pol и оболочечные гены гликопротеина. Альтернативно, термин «клеточная линия-продуцент» означает упаковывающую клеточную линию, которая также способна экспрессировать вектор для переноса, содержащий представляющий интерес трансген.
Термин «временно трансфецированный» относится к трансфецированным клеткам, в которых нуклеиновые кислоты-мишени (то есть, ретровирусные гены) перманетно не включаются в клеточный геном. Следовательно, эффекты нуклеиновых кислот в клетках происходят только в течение короткого периода времени.
Нуклеиновокислотные векторы
В своем первом аспекте, настоящее изобретение относится к нуклеиновокислотному вектору, содержащему ориджин репликации животных, не являющихся млекопитающими, и способному сохранять по меньшей мере 25 тысяч пар оснований (т.п.о.) ДНК, где указанный нуклеиновокислотный вектор отличается тем, что он содержит последовательности нуклеиновой кислоты ретровируса, кодирующие:
белки gag и pol, и
белок env или его функциональный аналог.
В частности, каждая последовательность нуклеиновой кислоты ретровируса может быть расположена в нуклеиновокислотном векторе в виде отдельных экспрессионных конструкций.
Авторами настоящего изобретения было обнаружено, что описанные здесь нуклеиновокислотные векторы могут быть использованы для получения частиц ретровирусного вектора, что позволяет обойти возникавшие ранее трудности, связанные с применением методов получения ретровирусного вектора. Так, например, при включении всех основных ретровирусных генов в нуклеиновокислотный вектор, эти ретровирусные гены могут быть затем введены в клетку-хозяина млекопитающего за одну стадию. Следовательно, применение нуклеиновокислотного вектора, предложенного авторами настоящей заявки, позволяет быстрее получить вектор и уменьшить количество материала, необходимого для получения ретровирусного вектора.
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно содержит последовательности нуклеиновой кислоты, кодирующие РНК-геном, состоящий из частиц ретровирусного вектора. Следует отметить, что РНК-геном, состоящий из частиц ретровирусного вектора, обычно вводят в «вектор для переноса», используемый в способах временной трансфекции. Плазмидный вектор для переноса обычно содержит промотор (такой как CMV), 3'-LTR (который может быть, а может и не быть, самоинактивирующимся (то есть, SIN) 3'-LTR), 5' LTR (который может содержать, а может и не содержать, область U5), инкапсидирующую последовательность (ψ) и может содержать трансген, присоединенный к промотору.
В одном из вариантов осуществления изобретения, в нуклеиновокислотный вектор включают множество копий РНК-генома, состоящего из частиц ретровирусного вектора (то есть, вектора для переноса). Предполагается, что множество копий вектора для переноса будет повышать титр вирусного вектора. Так, например, нуклеиновокислотный вектор может включать две или более, например, три, четыре, пять, шесть, семь, восемь, девять, десять или более копий РНК-генома, состоящего из частиц ретровирусного вектора (то есть, вектора для переноса).
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор содержит один или множество сайтов рекомбинации. Фермент рекомбиназа катализирует реакцию рекомбинации между двумя сайтами рекомбинации.
Специалистам известно множество типов систем сайт-специфической рекомбинации, и в настоящем изобретении может быть использована любая подходящая система рекомбинации. Так, например, в одном из вариантов осуществления изобретения, сайт(ы) рекомбинации выбирают или получают из системы int/att фага лямбда, системы Cre/lox бактериофага P1, дрожжевой системы FLP/FRT, рекомбиназной системы Gin/gix фага Mu, рекомбиназной системы Cin, рекомбиназной системы Pin E. coli и системы R/RS плазмиды pSR1 или любой их комбинации. В другом варианте осуществления изобретения, сайтом рекомбинации является сайт att (например, от фага лямбда), где сайт att позволяет осуществлять сайт-направленную интеграцию в присутствии интегразы лямбда. Следует отметить, что термин «интеграза лямбда» включает мутантные интегразы, которые являются совместимыми с системой int/att, например, модифицированные интегразы лямбда, описанные в WO 2002/097059.
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор выбирают из бактериальной искусственной хромосомы (BAC), дрожжевой искусственной хромосомы (YAC), искусственной хромосомы, происходящей от P1 (PAC), фосмиды или космиды. В другом варианте осуществления изобретения, нуклеиновокислотным вектором является бактериальная искусственная хромосома (BAC).
Бактериальные искусственные хромосомы
Термин «бактериальная искусственная хромосома» или «BAC» означает ДНК-конструкцию, происходящую от бактериальных плазмид, способных сохранять крупную вставку экзогенной ДНК. Эти плазмиды могут сохранять ДНК-вставку размером приблизительно максимум 350 т.п.о. BAC были получены из хорошо охарактеризованной бактериальной функциональной плазмиды, ответственной за фертильность (F-плазмиды), которая содержит расщеплающие гены, стимулирующие однородное распределение плазмид после деления бактериальных клеток. Это позволяет BAC стабильно реплицироваться и сегрегироваться во всех геномах эндогенных бактерий (таких как E. coli). BAC обычно содержит по меньшей мере одну копию ориджина репликации (такого как ген oriS или ген oriV), ген repE (ответственный за репликацию плазмиды и регуляцию числа копий) и расщепляющие гены (такие как sopA, sopB, parA, parB и/или parC), что обеспечивает стабильное сохранение BAC в бактериальных клетках. По своей природе, BAC имеют кольцевую и суперспирализованную структуру, что облегчает их выделение, в отличие от линейных искусственных хромосом, таких как YAC. Они могут быть также относительно легко введены в бактериальные клетки-хозяева простыми методами, такими как электропорация.
В одном из вариантов осуществления изобретения, бактериальная искусственная хромосома содержит ген oriS. В одном из вариантов осуществления изобретения, бактериальная искусственная хромосома содержит ген repE. В одном из вариантов осуществления изобретения, бактериальная искусственная хромосома содержит расщепляющие гены. В другом варианте осуществления изобретения, расщепляющие гены выбраны из sopA, sopB, parA, parB и/или parC. В другом варианте осуществления изобретения, бактериальная искусственная хромосома содержит ген sopA и sopB.
BAC, используемая в настоящем изобретении, может быть получена из коммерчески доступных источников, например, pSMART BAC от LUCIGEN™ (см. полноразмерную последовательность остова в геноме регистрационный No. EU101022.1). Эта BAC содержит «многокопийную» систему L-арабинозы, которая также содержит ориджин репликации oriV со средним числом копий, который является активным только в присутствии белка репликации TrfA. Ген TrfA может быть введен в геном бактериальных клеток-хозяев под контролем индуцибельного промотора L-арабинозы araC-PBAD (см. Wild et al. (2002) Genome Res. 12(9): 1434-1444). Присоединение L-арабинозы индуцирует экспрессию TrfA, который активирует oriV, что приводит к репликации плазмиды до 50 копий на клетку.
Дрожжевые искусственные хромосомы
Термин «дрожжевая искусственная хромосома» или «YAC» означает хромосомы, в бактериальные плазмиды которых была введена дрожжевая ДНК. Эти хромосомы содержат автономную реплицирующуюся последовательность (ARS) (то есть, ориджин репликации), центромеру и теломеры. В отличие от BAC, YAC является линейной, а поэтому содержит дрожжевые теломеры у каждого конца хромосомы, что позволяет защищать эти концы от разложения при передаче потомству клеток-хозяев. YAC могут сохранять ДНК-вставку размером в пределах от 100 до 2000 т.п.о.
Искусственные хромосомы, происходящие от P1
Термин «искусственная хромосома, происходящая от Р1» или «PAC» означает ДНК-конструкции, происходящие от ДНК P1-бактериофага и бактериальной F-плазмиды. Они обычно сохраняют ДНК-вставку размером приблизительно максимум 100-300 т.п.о. и используются в качестве клонирующих векторов в E. coli. PAC имеют такие же преимущества, как и BAC, а именно, они облегчают очистку и введение в бактериальные клетки-хозяева.
Космиды и фосмиды
Термин «космида» означает ДНК-конструкции, происходящие от бактериальных плазмид, которые дополнительно содержат сайты cos, происходящие от бактериофага лямбда. Космиды обычно содержат бактериальный ориджин репликации (такой как oriV), маркер отбора, сайт клонирования и по меньшей мере один сайт cos. Космиды обычно имеют ДНК-вставку размером максимум 40-45 т.п.о. Было показано, что космиды являются более эффективными для инфицирования клеток E. coli, чем стандартные бактериальные плазмиды. Термин «фосмиды» означает нуклеиновокислотные векторы животных, не являющихся млекопитающими, и такие векторы аналогичны космидам, за исключением того, что они были сконструированы на основе бактериальной F-плазмиды. В частности, эти фосмиды используют F-плазмидный ориджин репликации и действуют по механизмам расщепления, что позволяет клонировать крупные фрагменты ДНК. Фосмиды обычно имеют ДНК-вставку размером максимум 40 т.п.о.
Ретровирусы
Ретровирусы принадлежат к семейству вирусов, содержащих псевдодиплоидный одноцепочечный РНК-геном. Они кодируют обратную транскриптазу, продуцирующую ДНК из РНК-генома, который может быть встроен в ДНК клетки-хозяина. Описанное здесь изобретение может быть применено для продуцирования дефектных по репликации частиц ретровирусного вектора. Частица ретровирусного вектора согласно изобретению может быть выбрана из любого подходящего ретровируса или может происходить от него.
В одном из вариантов осуществления изобретения, частица ретровирусного вектора происходит от, или выбрана из них, лентивируса, альфа-ретровируса, гамма-ретровируса или пенистого ретровируса, такого как лентивирус или гамма-ретровирус, а в частности, лентивирус. В другом варианте осуществления изобретения, частицей ретровирусного вектора является лентивирус, выбранный из группы, состоящей из ВИЧ-1, ВИЧ-2, SIV, FIV, EIAV и висны. Лентивирусы способны инфицировать неделящиеся (то есть, «покоящиеся») клетки, что делает ретровирусные векторы привлекательным средством для генотерапии. В другом варианте осуществления изобретения, частицей ретровирусного вектора является ВИЧ-1 или вирус, происходящий от ВИЧ 1. Геномная структура некторых ретровирусов описана в литературе. Так, например, подробное описание ВИЧ-1 можно найти в базе данных Genbank NCBI (геном под регистрационным No. AF033819). ВИЧ-1 является одним из самых хорошо охарактеризованных ретровирусов, а поэтому он часто используется как ретровирусный вектор.
Гены ретровирусов
Последовательности нуклеиновых кислот, которые являются общими для всех ретровирусов, более подробно описаны ниже.
Длинные концевые повторы (LTR): Основная структура ретровирусного генома содержит 5'-LTR и 3'-LTR, между которыми находятся гены, необходимые для продуцирования ретровируса. LTR необходимы для интерграции и транскрипции ретровируса. Они могут также действовать как промоторные последовательности, регулирующие экспрессию ретровирусных генов (то есть, они представляют собой цис-действующие гены). LTR состоят из трех подобластей, обозначенных U3, R, U5, где U3 происходит от уникальной последовательности, расположенной у 3'-конца РНК; R происходит от последовательности, повторяющейся у обоих концов РНК, а U5 происходит от уникальной последовательности, расположенной у 5'-конца РНК. Поэтому, в одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно содержит 5'- и 3'-LTR. В другом варианте осуществления изобретения, область U5 5'-LTR может быть делетирована и заменена polyA-хвостом вируса, не являющегося ВИЧ-1 (см. Hanawa et al. (2002) Mol. Ther. 5(3): 242-51).
Для решения проблем безопасности, связанных с получением компетентного по репликации вируса, был сконструирован самоинактивирующийся (SIN) вектор путем делеции части в области U3 3′-LTR, которая включает TATA-бокс и сайты связывания с факторами транскрипции Sp1 и NF-κB (см. Miyoshi et al. (1998) J. Virol. 72(10):8150-7). Делецию переносят в 5′-LTR после обратной транскрипции и интеграции в инфицированные клетки, что приводит к инактивации транскрипции LTR. Этот вектор известен как самоинактивирующаяся векторная система на основе лентивирусов, которая может быть использована в настоящем изобретении.
ψ: Инкапсидирование ретровирусных РНК достигается посредством последовательности ψ (пси), локализованной у 5'-конца ретровирусного генома. Специалистам также хорошо известно, что последовательности, расположенные ниже последовательности пси и простирающиеся до gag-кодирующей области, используются для эффективного продуцирования ретровирусного вектора (см. Cui et al. (1999) J. Virol. 73(7): 6171-6176). В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно содержит последовательность ψ (пси).
Сайт связывания с праймером (PBS): Ретровирусный геном содержит PBS, который находится за областью U5 5'-LTR. Этот сайт связывается с тРНК-праймером, необходимым для инициации обратной транскрипции. В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно содержит последовательность PBS.
PPT: Ретровирусные геномы содержат короткие пуриновые фрагменты, называемые полипуриновым трактом (PPT), расположенным возле 3'-конца ретровирусного генома. Эти PPT функционируют как РНК-праймеры для синтеза плюс-цепи ДНК во время обратной транскрипции. Сложные ретровирусы (такие как ВИЧ-1) содержат второй PPT, расположенный ближе к центру (то есть, центральный полипуриновый тракт (cPPT)), и этот РРТ представляет собой второй сайт для инициации синтеза ДНК. Было показано, что ретровирусные векторы, кодирующие cPPT, усиливают трансдукцию и повышают уровень экспрессии трансгенов (см. Barry et al. (2001) Hum. Gene Ther. 12(9):1103-8). В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно содержит последовательность 3'-PPT и/или последовательность cPPT.
Геномная структура вышеописанных некодирующих областей хорошо известна специалистам. Так, например, подробное описание геномной структуры некодирующих областей в ВИЧ-1 можно найти в базе данных Genbank NCBI для генома под регистрационным номером No. AF033819 или в ВИЧ-1 HXB2 (хорошо известном эталонном штамме ВИЧ-1) для генома под регистрационным номером No. K03455. В одном из вариантов осуществления изобретения, некодирующие области происходят от последовательностей, имеющихся в базе данных для генома под регистрационным номером No. K03455, например, от пар оснований 454-1126 (для R-U5-PBS-Gag), 7622-8479 (для RRE) или 7769-8146 (для RRE), 4781-4898 (для cPPT), 9015-9120 и 9521-9719 (для dNEF-PPT-sinU3-R-U5).
Gag/pol: Экспрессия генов gag и pol зависит от сдвига трансляционной рамки считывания между gag и gagpol. Оба они кодируют полипротеины, которые расщепляются в процессе созревания. Основная структурная матрица, капсид и нуклеокапсидные белки ретровирусного вектора кодируются gag. Ген pol кодирует ретровирусные ферменты: i) обратную транскриптазу, играющую главную роль в реакции обратной транскрипции ретровирусного РНК-генома с образованием двухцепочечной ДНК; ii) интрегразу, ответственную за интеграцию ретровирусного ДНК-генома в хромосоме клетки-хозяина; и iii) протеазу, которая расщепляет синтезированный полипротеин с образованием зрелых и функциональных белков ретровируса. В одном из вариантов осуществления изобретения, ретровирусная последовательность нуклеиновой кислоты, кодирующая белки gag и pol, происходит от последовательности ВИЧ-1 HXB2, которая имеется в базе данных для генома под регистрационным номером No. K03455, например, от пар оснований 790-5105.
Env: Ген env («оболочки») кодирует поверхностные и трансмембранные компоненты ретровирусной оболочки (например, гликопротеины gp120 и gp41 ВИЧ-1) и участвует в слиянии ретровируса и клеточной мембраны. Для расширения тропизма ретровирусных векторов к тканям, описанные здесь ретровирусные векторы могут быть псевдотипированы оболочечным белком, происходящим от другого вируса. Термин «псевдотипирование» означает процесс, посредством которого клетки-хозяева, содержащие ретровирусные векторы, включая лентивирусные векторы, могут быть размножены или модифицированы путем замены гликопротеинов (GP) на частицах ретровирусного вектора (например, с использованием GP, полученных или происходящих от других оболочечных вирусов или с использованием синтетических/искусственных GP). Гликопротеинами, наиболее часто используемыми для псевдотипирования ретровирусных векторов, являются GP вируса везикулярного стоматита (VSVg), что обусловлено его широким тропизмом и высокой стабильностью векторных частиц. Однако, совершенно очевидно, что для псевдотипирования могут быть использованы и другие гликопротеины (см., публикацию Cronin et al. (2005) Curr. Gene Ther. 5(4):387-398, которая во всей своей полноте вводится в настоящее описание посредством ссылки). Выбор вируса, используемого для псевдотипирования, может также зависеть от типа клетки и/или органа-мишени, поскольку было показано, что некоторые псевдотипы являются более специфичными для тканей конкретного типа.
В одном из вариантов осуществления изобретения, белок env или его функциональный аналог получают из вируса, или он происходит от вируса, выбранного из везикуловируса (например, вируса везикулярного стоматита), лиссавируса (например, вируса бешенства, вируса Мокола), аренавируса (например, вируса лимфоцитарного хориоменингита (LCMV)), альфавируса (например, вируса реки Росс (RRV), вируса Синдбис, вируса лесов Семлики (SFV), вируса Венесуэльского лошадиного энцефалита), филовируса (например, вируса Эбола штамма Рестон, вируса Эбола штамма Заир, лассавируса), альфа-ретровируса (например, вируса птичьего лейкоза (ALV)), бета-ретровируса (например, ретровируса овец породы Ягсигте (JSRV)), гамма-ретровируса (например, вируса мышиного лейкоза Молони (MLV), вируса лейкоза гиббонов (GALV), кошачьего эндогенного ретровируса (RD114)), дельта-ретровируса (например, человеческого T-лимфотропного вируса 1 (HTLV-1)), спумавируса (например, человеческого пенистого вируса), лентивируса (например, вируса Маеди-висны (MVV)), коронавируса (например, SARS-CoV), респировируса (например, вируса Сендай, респираторно-синцитиального вируса (RSV)), гепацивируса (например, вируса гепатита C (HCV)), вируса гриппа (например, вируса гриппа A) и нуклеополиэдровируса (например, множественного нуклеополиэдровируса Autographa californica (AcMNPV)). В другом варианте осуществления изобретения, белок env или его функциональный аналог получают из вируса, или он происходит от вируса везикулярного стоматита. В этом варианте осуществления изобретения может быть использован белок гликопротеина вируса везикулярного стоматита (VSVg), который позволяет ретровирусным частицам инфицировать более широкий круг клеток-хозяев и исключить возможность рекомбинации с образованием оболочечных белков дикого типа. В другом варианте осуществления изобретения, последовательность нуклеиновой кислоты ретровируса, кодирующая белок env или его функциональный аналог происходит от последовательности, имеющейся в базе данных для генома под регистрационным номером J02428.1, например, от пар оснований 3071-4720.
Описанные здесь структурные гены являются общими для всех ретровирусов. В ретровирусах различных типов могут присутствовать и другие вспомогательные гены. Так, например, лентивирусы, такие как ВИЧ-1, содержат шесть других вспомогательных генов, известных как rev, vif, vpu, vpr, nef и tat. Другие ретровирусы могут иметь вспомогательные гены, аналогичные описанным здесь генам, однако, они не всегда могут иметь такое же название, как в литературе. Описание различных ретровирусных вспомогательных генов можно найти в публикациях Tomonaga and Mikami (1996) J. Gen. Virol. 77(Pt 8):1611-1621.
Rev: Вспомогательный ген rev («регулятор вириона») кодирует вспомогательный белок, который связывается с Rev-отвечающим элементом (RRE) и облегчает экспорт ретровирусных транскриптов. Продукт белка, кодируемого геном, позволяет фрагментам ретровирусной мРНК, содержащей Rev-отвечающий элемент (RRE), экспортироваться из ядра в цитоплазму. Было предсказано, что последовательность RRE образует структуру, имеющую сложную укладку. Эта конкретная роль rev указывает на тесную взаимосвязь между стадиями сплайсинга и экспорта из ядра. В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор содержит последовательность RRE. В другом варианте осуществления изобретения, последовательность RRE происходит от последовательности ВИЧ-1 HXB2, которая имеется в базе данных для генома под регистрационным номером No. K03455, например, от пар оснований 7622-8479 или 7769-8146, а в частности, пар оснований 7622-8479.
Rev связывается с RRE и облегчает экспорт одиночных сплайсированных (env, vif, vpr и vpu) или не-сплайсированных (gag, pol и геномной РНК) вирусных транскриптов, что приводит к инициации последующих событий, таких как трансляция и упаковка генов (см. Suhasini and Reddy (2009) Curr. HIV Res. 7(1): 91-100). В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно содержит вспомогательный ген rev или аналогичный ему ген (то есть, он происходит от других ретровируов или функционально аналогичной системы). Включение гена rev обеспечивает эффективный экспорт РНК-транскриптов генома ретровирусного вектора из ядра в цитоплазму, а особенно в том случае, когда на транспортируемом транскрипте также присутствует элемент RRE. В другом варианте осуществления изобретения, ген rev содержит последовательность, которая по меньшей мере на 60%, например, по меньшей мере на 70% идентична парам оснований 970-1320 генома под регистрационным номером No. M11840 (то есть, кДНК клона 12 ВИЧ-1, локуса HIVPCV12). В альтернативном варианте осуществления изобретения, ген rev содержит последовательность, которая по меньшей мере на 60%, например, по меньшей мере на 70%, 80%, 90% или 100% идентична парам оснований 5970-6040 и 8379-8653 генома под регистрационным номером No. K03455.1 (то есть, вируса иммунодефицита человека типа 1, HXB2).
Считается, что вспомогательные гены играют определенную роль в репликации и патогенезе ретровирусов, а поэтому многие современные системы продуцирования вирусных векторов не включают некоторые из этих генов. Исключение составляет ген rev, который обычно присутствует, или система, аналогичная системе rev/RRE, которая также может быть использована. Поэтому, в одном из вариантов осуществления изобретения, последовательности нуклеиновой кислоты, кодирующие один или более вспомогательных генов vpr, vif, vpu, tat и nef, или аналогичные вспомогательные гены, подвергают дизрупции так, чтобы указанные вспомогательные гены были удалены из РНК-генома частиц ретровирусного вектора или были неспособны кодировать функциональные вспомогательные белки. В другом варианте осуществления изобретения, по меньшей мере два или более, три или более, четыре или более или все вспомогательные гены vpr, vif, vpu, tat и nef или аналогичные вспомогательные гены подвергают дизрупции так, чтобы указанные вспомогательные гены были удалены из РНК-генома частиц ретровирусного вектора или были неспособны кодировать функциональные вспомогательные белки. Для удаления функционального вспомогательного гена необязательно удаление всего гена, то есть, в данном случае, достаточно удалить часть гена или осуществить дизрупцию гена.
Следует отметить, что последовательности нуклеиновой кислоты, кодирующие дефектные по репликации частицы ретровирусного вектора, могут быть такими же, как последовательности дикого типа ретровируса или происходить от этих последовательностей, то есть, эти последовательности основаны на последовательностях частиц ретровирусного вектора, а именно, они могут представлять собой генетически или как-либо иначе модифицированные варианты последовательностей, содержащихся в вирусе дикого типа. Следовательно, ретровирусные гены, включенные в нуклеиновокислотные векторы или в геномы клеток-хозяев, могут также называться оптимизированными по кодонам вариантами генов дикого типа.
Дополнительные компоненты
Нуклеиновокислотные векторы согласно изобретению могут также включать дополнительные компоненты. Эти дополнительные признаки могут быть использованы, например, для повышения стабильности транскриптов для трансляции, повышения уровня экспрессии гена и включения/отключения транскрипции гена.
Частицы ретровирусного вектора, продуцируемые в соответствии с настоящим изобретением, могут быть использованы в способах генотерапии. Следовательно, в одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно включает один или более трансгенов. Этот трансген может представлять собой терапевтически активный ген, кодирующий генный продукт, которы может быть использован для лечения или для ослабления симптомов заболевания, подвергаемого лечению. Трансген может кодировать, например, антисмысловую РНК, рибозим, белок (например, белок-супрессор опухоли), токсин, антиген (кторый может быть использован для индуцирования антител или хелперных T-клеток или цитотоксических T-клеток) или антитело (такое как одноцепочечное антитело). В одном из вариантов осуществления изобретения, трансген кодирует бета-глобин.
Считается, что множество копий вектора для переноса, содержащего трансген, будут повышать титр ретровирусного вектора, а поэтому, в одном из вариантов осуществления изобретения, нуклеиновокислотный вектор содержит множество копий трансгена, например, две или более, а в частности, три или более копий трансгена. В некоторых случаях, для лечения заболевания требуется более, чем один генный продукт, а поэтому, в другом варианте осуществления изобретения, нуклеиновокислотный вектор дополнительно содержит два или более, а именно, три или более, или четыре или более различных трансгенов.
Используемый здесь термин «трансген» означает гетерологичную или чужеродную ДНК, которая не присутствует в клетке-хозяине млекопитающего, в которую ее вводят, или экспрессируется в этой клетке на недостаточном уровне. Так, например, если ген-мишень не экспрессируется в клетке-хозяине млекопитающего на соответствующем уровне, то в эту клетку может быть включен скорректированный вариант гена-мишени, используемый в качестве трансгена. Следовательно, трансгеном может быть ген, представляющий интерес с терапевтической точки зрения. Трансген может быть получен из клеток другого типа или другого вида, либо он может быть получен путем синтеза. Альтернативно, трансген может быть получен из клетки-хозяина, но он может быть функционально присоединен к регуляторным областям, которые отличаются от областей, присутствующих в нативном гене. Альтернативно, трансгеном могут быть различные аллели или варианты гена, присутствующие в клетке-хозяине.
Целью генотерапии является модификация генетического материала живых клеток для проведения терапии, и такая модификация включает инсерцию функционального гена в клетку для достижения терапевтического эффекта. Ретровирусный вектор, продуцируемый с использованием нуклеиновокислотных векторов и клеток-хозяев, описанных в настоящей заявке, может быть использован для трансфекции клеток-мишеней и для индуцирования экспрессии гена, который может представлять интерес с терапевтической точки зрения. Следовательно, ретровирусный вектор может быть использован для лечения млекопитающего, такого как человек, страдающий заболеваниями, включая, но не ограничиваясь ими, наследственные расстройства, рак и некоторые вирусные инфекции.
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно содержит элемент регуляции транскрипции. Так, например, любой из описанных здесь элементов может быть функционально присоединен к промотору так, чтобы могла осуществляться регуляция экспрессии. Используемый здесь термин «промоторы» может включать известные промоторы, как целые, так и их части, которые могут быть конститутивно активными или индуцибельными, например, в присутствии регуляторного белка. В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно включает высокоэффективный промотор, такой как промотор CMV. Этот промотор имеет то преимущество, что он стимулирует высокий уровень экспрессии элементов, кодируемых нуклеиновокислотным вектором животных, не являющихся млекопитающими. В другом варианте осуществления изобретения, промотор CMV содержит последовательность, происходящую от штамма человеческого цитомегаловируса AD169. Эта последовательность имеется в базе данных для генома под регистрационным номером No. X17403, например, с парами оснований от 173731 до 174404.
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно содержит изолятор, такой как изолятор хроматина. Термин «изолятор» означает генную последовательность, которая блокирует взаимодействие между промоторами и энхансерами. В другом варианте осуществления изобретения, изолятор (такой как изолятор хроматина) находится между каждой последовательностью нуклеиновой кислоты ретровируса. Этот изолятор способствует предупреждению интерференции промотора (то есть, где промотор, происходящий от одной транскрипционной единицы, негативно влияет на экспрессию смежной транскрипционной единицы), присутствующего между смежными последовательностями нуклеиновой кислоты ретровируса. Следует отметить, что если изоляторы присутствуют в нуклеиновокислотном векторе, расположенном между каждой последовательностью нуклеиновой кислоты ретровируса, то эти изоляторы могут быть сконструированы в виде отдельных экспрессионных конструкций в нуклеиновокислотном векторе. Так, например, каждая последовательность, кодирующая последовательности нуклеиновой кислоты ретровируса, имеет свой собственный промотор и/или интрон и/или polyA-сигнал. В одном из вариантов осуществления изобретения, изолятор хроматина имеет последовательность, которая по меньшей мере на 90%, например, по меньшей мере на 95% идентична куриной (Gallus gallus) последовательности изолятора HS4 (см, например, геном под регистрационным номером No. U78775.2, пары оснований 1-1205).
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно включает polyA-сигнал. Использование polyA-сигнала имеет то преимущество, что этот сигнал защищает мРНК от ферментативного разложения и стимулирует трансляцию. В другом варианте осуществления изобретения, polyA-сигнал получают от SV40, бычьего гормона роста и/или человеческого бета-глобина, либо он происходит от них. В одном из вариантов осуществления изобретения, polyA-сигнал происходит от раннего polyA-сигнала SV40 (см., например, геном под регистрационным номером No. EF579804.1, пары оснований 2668-2538 от минус-цепи). В одном из вариантов осуществления изобретения, polyA-сигнал происходит от polyA-сигнала человеческого бета-глобина (см., например, геном под регистрационным номером No. GU324922.1, пары оснований 3394-4162).
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно включает интронную последовательность. Известно, что использование интрона, расположенного ниже энхансерной/промоторной области и выше кДНК-вставки (то есть, трансгена), повышает уровень экспрессии вставки. В другом варианте осуществления изобретения, интронной последовательностью является последовательность интрона человеческого бета-глобина или интрона II кроличьего бета-глобина. В одном из вариантов осуществления изобретения, интрон человеческого бета-глобина происходит от последовательности, имеющейся в базе данных для генома под регистрационным номером No. KM504957.1 (например, пары оснований 476-1393). В одном из вариантов осуществления изобретения, интрон II кроличьего бета-глобина происходит от последовательности, имеющейся в базе данных для генома под регистрационным номером No. V00882.1 (например, пары оснований 718-1290).
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно включает посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE). Было показано, что присутствие WPRE повышает уровень экспрессии и таким образом, вероятно, является предпочтительным для сохранения высоких уровней экспрессии. В другом варианте осуществления изобретения, WPRE происходит от последовательности, имеющейся в базе данных для генома под регистрационным номером No. J04514.1 (например, пары оснований 1093-1684).
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно включает внутренний сайт связывания с рибосомой (IRES). IRES представляет собой структурированный элемент РНК, который обычно присутствует в 5'-нетранслируемой области, расположенный ниже 5'-кэпа (и который необходим для сборки инициирующего комплекса). IRES распознается факторами инициации трансляции и позволяет осуществлять независимую от кэпа трансляцию. В другом варианте осуществления изобретения, IRES происходит от генома вируса энцефаломиокардита (EMCV) (см., например, геном под регистрационным номером No. KF836387.1, пары оснований 151-724).
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор дополнительно включает сайт множественного клонирования (MCS). MCS представляет собой короткий сегмент ДНК в нуклеиновокислотном векторе, который содержит множество рестрикционных сайтов (например, 10, 15 или 20 сайтов). Эти сайты обычно присутствуют только в одном положении нуклеиновокислотного вектора, а поэтому эндонуклеаза осуществляет рестриктирующее действие только в одном сайте. Это облегчает встраивание ретровирусных генов с использованием соответствующих эндонуклеаз (то есть, рестриктирующих ферментов).
Для специалистов в данной области очевидно, что конструкции могут быть расположены в нуклеиновокислотном векторе в любом порядке. В репрезентативном варианте осуществления изобретения, нуклеиновокислотный вектор содержит следующие вставки: последовательность нуклеиновой кислоты ретровируса, кодирующую белки gag и pol; последовательность нуклеиновой кислоты ретровируса, кодирующую белок env или его функциональный аналог (такой как VSVg) и последовательность нуклеиновой кислоты ретровируса, содержащую вспомогательный ген rev (такую как оптимизированную по кодонам последовательность rev) или аналогичный ему ген или функционально аналогичную систему (то есть, GagPol-Env-Rev-содержащую последовательность BAC («остов BAC»), такую как: GagPol-(дикого типа)VSVg-(оптимизированная по кодонам)Rev-pSMARTBAC). В другом варианте осуществления изобретения, изолятор (такой как изолятор хроматина) расположен между последовательностями gagpol, env и rev. В другом варианте осуществления изобретения, промотор расположен перед каждой из последовательностей gagpol, env и rev. В другом варианте осуществления изобретения, по меньшей мере одна копия последовательности вектора для переноса (то есть, содержащая последовательности нуклеиновой кислоты, кодирующие РНК-геном частицы ретровирусного вектора) расположена перед последовательностью gagpol.
В одном из вариантов осуществления изобретения, нуклеиновокислотный вектор включает следующие вставки: изолятор (такой как изолятор хроматина); промотор (такой как промотор CMV), интрон (такой как интрон человеческого бета-глобина); последовательность нуклеиновой кислоты ретровируса, кодирующую белки gag и pol; ретровирусную нуклеиновую кислоту, кодирующую RRE, polyA-сигнал (такой как polyA-сигнал человеческого бета-глобина), изолятор (такой как изолятор хроматина); промотор (такой как промотор CMV), интрон (такой как интрон человеческого бета-глобина); последовательность нуклеиновой кислоты ретровируса, кодирующую белок env или его функциональный аналог (такой как VSVg), polyA-сигнал (такой как polyA-сигнал человеческого бета-глобина), изолятор (такой как изолятор хроматина); промотор (такой как промотор CMV), последовательность нуклеиновой кислоты ретровируса, содержащую вспомогательный ген rev или аналогичный ему ген или функционально аналогичную систему; polyA-сигнал (такой как polyA-сигнал человеческого бета-глобина), изолятор (такой как изолятор хроматина); промотор (такой как промотор CMV), интрон (такой как интрон кроличьего бета-глобина); polyA-сигнал и сайт множественного клонирования. Следует отметить, что вместе с этой вставкой и/или внутри этой вставки могут присутствовать и другие последовательности.
Последовательности нуклеиновой кислоты могут быть последовательно введены в нуклеиновокислотный вектор. Это позволяет проводить отбор после каждой интеграции для гарантии того, что все необходимые последовательности нуклеиновой кислоты будут успешно интегрированы в нуклеиновокислотный вектор. Альтернативно, по меньшей мере две или более последовательностей нуклеиновой кислоты одновременно вводят в нуклеиновокислотный вектор.
Следует отметить, что описанные здесь дополнительные гены могут быть введены в нуклеиновокислотный вектор стандартными методами молекулярного клонирования, известными специалистам, например, с использованием рестриктирующих эндонуклеаз, и методами лигирования. Кроме того, нуклеиновокислотный вектор, а в частности, BAC, PAC, фосмиды и/или космиды, могут быть введены в бактериальные клетки-хозяева (такие как клетки E. coli, а в частности, штамм E. coli DH10B) стандартными методами, такими как электропорация.
Применения
В соответствии с другим своим аспектом, настоящее изобретение относится к описанному здесь нуклеиновокислотному вектору, используемому в способе продуцирования частиц ретровирусного вектора. В настоящей заявке описано множество преимуществ использования нуклеиновокислотных векторов в способах временной трансфекции, а главным образом, возможность введения только 4-плазмидной системы трансфекции в один нуклеиновокислотный вектор, и тем самым снижения количества используемого материала.
Способы
В соответствии с другим своим аспектом, настоящее изобретение относится к способу продуцирования дефектной по репликации частицы ретровирусного вектора, где указанный способ включает:
(a) введение определенного здесь нуклеиновокислотного вектора в культуру клеток-хозяев млекопитающих; и
(b) культивирование клеток-хозяев млекопитающих в условиях, способствующих продуцированию дефектной по репликации частицы ретровирусного вектора.
Преимущество включения всех ретровирусных генов в крупный нуклеиновокислотный вектор завключается в том, что эти гены могут быть сначала получены в микробных клетках (таких как бактериальные или дрожжевые клетки), которые гораздо легче транспортировать и модифицировать, а затем введены в клетки млекопитающих за одну стадию.
В одном из вариантов осуществления изобретения, клеткой-хозяином является клетка млекопитающего. В другом варианте осуществления изобретения, клетки млекопитающих выбраны из клеток HEK 293, клеток HEK 6E, клеток CHO, клеток Jurkat, клеток KS62, клеток PerC6, клеток HeLa, клеток HOS, клеток H9 или их производных или функциональных эквивалентов. В другом варианте осуществления изобретения, клетками-хозяевами млекопитающих являются клетки HEK 293 или клетки, происходящие от клеток HEK 293. Такие клетки могут представлять собой адгезивные клеточные линии (то есть, они могут быть культивированы в отдельном слое, связанном с поверхностью) или суспензионно адаптированые/неадгезивные клеточные линии (то есть, они могут быть культивированы в суспензионной культуральной среде). В еще одном варианте осуществления изобретения, клетками HEK 293 являются клетки HEK 293T или клетки HEK 6E. Термин «клетка HEK 293» означает клеточную линию почек человеческого эмбриона 293, которая чаще всего используется в биотехнологии. В частности, клетки HEK 293T обычно используются для продуцирования различных ретровирусных векторов. Другими примерами подходящих коммерчески доступных клеточных линий являются клеточные линии T REX™ (Life Technologies).
Клетки-хозяева, трансдуцированные описанными здесь способами, могут быть использованы для получения ретровирусного вектора с высоким титром.
Используемый здесь термин «высокий титр» означает эффективное количество ретровирусного вектора или частицы, способных трансдуцировать клетку-мишень, такую как клетка пациента. В одном из вариантов осуществления изобретения, высокий титр составляет более, чем 106 ЕТ/мл, без концентрирования [ЕТ (TU)=единицы трансдукции].
Специалисту в данной области известно, что введение нуклеиновокислотного вектора в клетку-хозяина может быть осуществлено подходящими стандартными методами, например, посредством опосредуемой липидом трансфекции (липофекции), микроинжекции, слияния клеток (таких как микроклетки), электропорации, химических методов трансфекции или бомбардировки микрочастицами. Следует отметить, что выбор способа введения нуклеиновокислотного вектора может зависеть от типа используемой клетки-хозяина млекопитающего. В одном из вариантов осуществления изобретения, стадию введения (a) осуществляют методом липофекции, электропорации или химической трансфекции. В другом варианте осуществления изобретения, нуклеиновокислотный вектор вводят в клетку-хозяина посредством липофекции. В альтернативном варианте осуществления изобретения, нуклеиновокислотный вектор вводят в клетку-хозяина химическим методом трансфекции, таким как обработка фосфатом кальция. Обработку фосфатом кальция проводят с использованием коммерчески доступных реагентов, поставляемых, например, Promega.
Для каждого специалиста очевидно, что условия проведения описанного здесь способа зависят от типа клетки-хозяина. Типичные условия, например, культуральная среда или температура хорошо известны специалистам (см., например, Kutner et al. (2009) Nature Protocols 4(4); 495-505). В одном из вариантов осуществления изобретения, стадию культивирования (b) осуществляют путем инкубирования клеток-хозяев млекопитающих в условиях повышенной влажности. В другом варианте осуществления изобретения, условия повышенной влажности включают инкубирование трансфецированных клеток при 37°C в 5% CO2. В одном из вариантов осуществления изобретения, стадию культивирования (b) осуществляют с использованием культуральной среды, выбранной из модифицированной по способу Дульбекко среды Игла (DMEM), содержащей 10% (об/об) фетальную бычью сыворотку (FBS), или бессывороточной среды UltraCULTURETM (Lonza, Cat. No. 12-725F), или экспрессионной среды FreeStyleTM (Thermo fisher, Cat. No. 12338 018).
В одном из вариантов осуществления изобретения, указанный способ дополнительно включает выделение дефектной по репликации частицы ретровирусного вектора. Так, например, в одном из вариантов осуществления изобретения, выделение осуществляют с использованием фильтра. В другом варианте осуществления изобретения, фильтром является мембрана, связывающаяся с низкомолекулярным белком (например, 0,22 мкм-мембрана, связывающаяся с низкомолекулярным белком, или 0,45 мкм-мембрана, связывающаяся с низкомолекулярным белком), такая как искусственная мембрана на основе поливинилиденфторида (PVDF) или полиэфирсульфона (PES).
В одном из вариантов осуществления изобретения, дефектные по репликации частицы ретровирусного вектора выделяют в течение не более, чем 72 часов после стадии введения (a). В другом варианте осуществления изобретения, дефектные по репликации частицы ретровирусного вектора выделяют в течение периода времени от 48 до 72 часов после стадии введения (a), например, в течение 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71 или 72 часов.
После выделения, частицы ретровирусного вектора могут быть подвергнуты концентрированию для их применения in vivo. Методы концентрирования включают, например, ультрацентрифугирование, преципитацию или анионообменную хроматографию. Ультрацентрифугирование представляет собой быстрый метод концентрирования ретровирусного вектора в лабораторном масштабе. Альтернативно, анионообменная хроматография (например, проводимая с использованием анионообменных мембранных кассет Mustang Q) или преципитация (например, с использованием ПЭГ 6000) являются особенно подходящими для обработки больших объемов супернатантов лентивирусных векторов.
В соответствии с другим своим аспектом, настоящее изобретение относится к дефектной по репликации частице ретровирусного вектора, полученной описанным здесь способом.
Настоящее изобретение более подробно проиллюстрировано на нижеследующих неограничивающих примерах.
Примеры
Пример 1: Схема конструирования
На фигуре 1 проиллюстрирована постадийная схема конструирования BACpack-WTGP-277delU5 и BACpack-SYNGP-277delU5. Благодаря совместимым концам XbaI- и NheI-гидролизата, лентивирусные упаковывающие гены постепенно загружали в вектор pSmart BAC. После присоединения GagPol были получены 2 конструкции, содержащие либо GagPol дикого типа (WTGP), либо оптимизированный по кодонам GagPol (SYNGP). В соответствии с номенклатурой, они были обозначены BACpack-WTGP и BACpack-SYNGP, соответственно. Затем кластер для переноса загружали на обе эти конструкции и получали BACpackWTGP-277delU5 и BACpackSYNGP-277delU5.
Пример 2: Эксперимент по проверке концепции с использованием конструкции BAC
106 клеток HEK293T высевали в 6-луночный планшет. На следующий день, прилипшие клетки трансфецировали с использованием ПЭИ в соответствии с инструкциями производителей. Клетки трансфецировали всего 4 микрограммами лентивирусных упаковывающих конструкций дикого типа (WT), состоящих из pMDL.gp (GagPol), pMD.G (VSVg), pK-Rev (Rev) и pCCL.277 (вектора для переноса GFP) или 2 мкг BACpack (одной BAC-конструкции, содержащей GagPol, VSVg и Rev) плюс 2 мкг вектора для переноса eGFP на отдельной плазмиде.
Через 48 часов после трансфекции, супернатант собирали, фильтровали через 0,22 мкм-фильтр и хранили при -80°C минимум 4 часа. Клетки HEK293T высевали для трансдукции при плотности 105 клеток на лунку в 24-луночном планшете. На следующий день, вирусный супернатант вводили в клетки в виде серийных разведений полибреном в конечной концентрации 8 мкг/мл. Через 3 дня после трансдукции, клетки собирали путем обработки трипсином и анализировали на GFP с помощью FACS-анализа. Титр вируса вычисляли как единицы трансдукции (ЕТ)/мл по следующему уравнению:
(GFP-позитивные клетки/100) × коэффициент разведения × число трансдукцированных клеток.
Титры вирусов сравнивали на гистограмме (фигура 2). Все инкубирования проводили при 37°C и в 5% CO2. Используемой средой является среда DMEM, в которую были добавлены 10% FBS и 1 мкг/мл доксициклина в BACpack+образец для переноса.
Наблюдения:
В этом примере, свойства конструкции BACpack, состоящей из экспрессионных кластеров GagPol, VSVg и Rev, сравнивали со свойствами стандартных 3-плазмидных упаковывающих систем, в которых GagPol, VSVg и Rev были доставлены по отдельности. В обоих случаях, вектор для переноса вводили в каждую отдельную плазмиду до достижения полного комплекта основных компонентов вирусного вектора.
В этом случае, BACpack плюс вектор для переноса давали титр вируса в неконцентрированном супернатанте 2,2 × 107 ЕТ/мл, по сравнению с титром 5 × 107 ЕТ/мл, полученным с использованием 4 отдельных плазмидных лентивирусных систем. Хотя при использовании BACpack наблюдался более низкий титр, однако, этот анализ был проведен до оптимизации, а после оптимизации мог быть достигнут более выскоий титр.
В этом анализе по проверке данной концепции было обнаружено, что конструкция BACpack способна упаковывать вектор для переноса при титре вируса, сравнимом с титром вируса для отдельной упаковывающей плазмидной системы, полученной посредством временной трансфекции.
Пример 3: Временная трансфекция BACpack в адгезивных клетках HEK293T
Для подтверждения способности двух конструкций BACpack-277delU5 продуцировать лентивирусный вектор в системе временной трансфекции, адгезивную клеточную линию, HEK293T, обычно используемую для продуцирования лентивирусного вектора посредством временной трансфекции, трансфецировали используемой здесь системы из 4 упаковывающих плазмид, BACpackWTGP-277delU5 или BACpackSYNGP-277delU5. Конструкции из двух плазмид, BACpack-277delU5, либо индуцировали для того, чтобы определить, может ли экспрессия генов способствовать продуцированию лентивирусного вектора, либо оставляли неиндуцированной для оценки эффективности системы Tet-репрессора.
Результаты, представленные на фигуре 3, указаны как титры единиц трансдукции (ЕТ)/мл супернатантов лентивирусного вектора, собранных при каждом условии трансфецирования. Результаты титрования показали, что клетки, трансфецированные либо BACpackWTGP-277delU5, либо BACpackSYNGP-277delU5 и индуцированнные 1 мкг/мл доксициклина (+Dox), продуцировали лентивирусный вектор в концентрациях, сравнимых с концентрациями, полученными при использовании стандартной 4-плазмидной системы. Кроме того, в не-индуцированных условиях наблюдалась значительно меньшая способность продуцировать лентивирусный вектор, чем в индуцированных условиях, и хотя такое продуцирование достигалось на более высоком уровне, чем в случае нетрансфецированного контроля, однако, это не может рассматриваться как недостаток системы временной трансфекции.
Эти результаты позволяют предположить, что один вектор BAC, содержащий все упаковывающие гены, необходимые для продуцирования лентивируса, может быть использован вместо стандартной 4-плазмидной системы.
Следует отметить, что описанные здесь варианты осуществления изобретения могут быть применены ко всем аспектам изобретения. Кроме того, все публикации, включая, но не ограничиваясь ими, патенты и патентные заявки, цитируемые в описании настоящей заявки, во всей своей полноте вводятся в настоящее описание посредством ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| СТАБИЛЬНЫЕ КЛЕТОЧНЫЕ ЛИНИИ ДЛЯ ПРОДУЦИРОВАНИЯ РЕТРОВИРУСОВ | 2016 |
|
RU2752498C2 |
| НУКЛЕАЗА PaCas9 | 2018 |
|
RU2706298C1 |
| УСЛОВНО РЕПЛИЦИРУЮЩИЙСЯ РЕТРОВИРУСНЫЙ ВЕКТОР (ВАРИАНТЫ), СПОСОБ ЕГО ПОЛУЧЕНИЯ И ИСПОЛЬЗОВАНИЯ (ВАРИАНТЫ), ВЫДЕЛЕННАЯ И ОЧИЩЕННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ | 1996 |
|
RU2270250C2 |
| ВИРУСНЫЕ ВЕКТОРЫ С ЗАВИСИМОЙ ОТ УСЛОВИЙ РЕПЛИКАЦИЕЙ И ИХ ПРИМЕНЕНИЕ | 2001 |
|
RU2301260C2 |
| ВЕКТОРЫ ДЛЯ ПРОИЗВОДСТВА БЕЛКОВ | 2019 |
|
RU2820218C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДОСТАВКИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА | 1994 |
|
RU2174845C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ МУЛЬТИПОТЕНТНЫХ КЛЕТОЧНЫХ ПОПУЛЯЦИЙ | 2008 |
|
RU2535365C2 |
| УСОВЕРШЕНСТВОВАННЫЕ ВЕКТОРЫ ЭКСПРЕССИИ МЛЕКОПИТАЮЩИХ И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2502800C2 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНЕЙ МОЗГА | 2014 |
|
RU2664471C2 |

Изобретение относится к биотехнологии. Описан нуклеиновокислотный вектор для получения дефектной по репликации частицы ретровирусного вектора, содержащий ориджин репликации животных, не являющихся млекопитающими, и способный сохранять по меньшей мере 25 тысяч пар оснований (т.п.о.) ДНК, где вектор выбран из: бактериальной искусственной хромосомы; дрожжевой искусственной хромосомы; искусственной хромосомы, происходящей от P1; фосмиды; или космиды, отличающийся тем, что указанный нуклеиновокислотный вектор содержит последовательности нуклеиновой кислоты ретровируса, кодирующие: белки gag и pol и белок env, где каждая последовательность нуклеиновой кислоты ретровируса имеет собственный промотор в нуклеиновокислотном векторе. Также раскрыт способ продуцирования дефектной по репликации частицы ретровирусного вектора. Изобретение расширяет арсенал средств для генной терапии. 2 н. и 25 з.п. ф-лы, 3 ил., 3 пр.
1. Нуклеиновокислотный вектор для получения дефектной по репликации частицы ретровирусного вектора, содержащий ориджин репликации животных, не являющихся млекопитающими, и способный сохранять по меньшей мере 25 тысяч пар оснований (т.п.о.) ДНК, где вектор выбран из: бактериальной искусственной хромосомы; дрожжевой искусственной хромосомы; искусственной хромосомы, происходящей от P1; фосмиды; или космиды, отличающийся тем, что указанный нуклеиновокислотный вектор содержит последовательности нуклеиновой кислоты ретровируса, кодирующие:
белки gag и pol, и
белок env, где каждая последовательность нуклеиновой кислоты ретровируса имеет собственный промотор в нуклеиновокислотном векторе.
2. Нуклеиновокислотный вектор по п. 1, который дополнительно включает последовательности нуклеиновой кислоты, кодирующие РНК-геном частицы ретровирусного вектора.
3. Нуклеиновокислотный вектор по п. 1, который дополнительно включает вспомогательный ген rev или его аналог.
4. Нуклеиновокислотный вектор по п. 1, где вектор представляет собой бактериальную искусственную хромосому.
5. Нуклеиновокислотный вектор по п. 1, где последовательности нуклеиновой кислоты ретровируса происходят от ретровируса, выбранного из лентивируса, альфа-ретровируса, гамма-ретровируса или пенистого ретровируса.
6. Нуклеиновокислотный вектор по п. 5, где последовательности нуклеиновой кислоты ретровируса происходят от лентивируса, выбранного из группы, состоящей из ВИЧ-1, ВИЧ-2, SIV, FIV, EIAV и висны.
7. Нуклеиновокислотный вектор по п. 6, где последовательности нуклеиновой кислоты ретровируса происходят от ВИЧ-1.
8. Нуклеиновокислотный вектор по п. 1, где белок env происходит от вируса везикулярного стоматита.
9. Нуклеиновокислотный вектор по п. 1, который дополнительно включает элемент регуляции транскрипции.
10. Нуклеиновокислотный вектор по п. 9, где элементом регуляции транскрипции является промотор CMV.
11. Нуклеиновокислотный вектор по п. 1, который дополнительно включает изолятор.
12. Нуклеиновокислотный вектор по п. 11, где изолятор находится между каждой из последовательностей нуклеиновой кислоты ретровируса.
13. Нуклеиновокислотный вектор по п. 1, который дополнительно включает один или более трансгенов.
14. Нуклеиновокислотный вектор по п. 1, который дополнительно включает внутренний сайт связывания с рибосомой (IRES).
15. Нуклеиновокислотный вектор по п. 1, который дополнительно включает polyA-сигнал.
16. Нуклеиновокислотный вектор по п. 1, который дополнительно включает интронную последовательность.
17. Нуклеиновокислотный вектор по п. 1, который дополнительно включает сайт множественного клонирования (MCS).
18. Нуклеиновокислотный вектор по любому из пп. 1-17 для применения в способе продуцирования частиц ретровирусного вектора.
19. Способ продуцирования дефектной по репликации частицы ретровирусного вектора, включающий:
(a) введение нуклеиновокислотного вектора по любому из пп. 1-17 в культуру клеток-хозяев млекопитающих; и
(b) культивирование клеток-хозяев млекопитающих в условиях, способствующих продуцированию дефектной по репликации частицы ретровирусного вектора.
20. Способ по п. 19, где клеткой-хозяином млекопитающих является клетка HEK 293.
21. Способ по п. 19, где стадию введения (a) осуществляют методом липофекции, электропорации или химической трансфекции, такой как обработка фосфатом кальция.
22. Способ по п. 19, где стадию культивирования (b) осуществляют путем инкубирования клеток-хозяев млекопитающих в условиях повышенной влажности.
23. Способ по п. 19, который дополнительно включает выделение дефектной по репликации частицы ретровирусного вектора.
24. Способ по п. 23, где выделение осуществляют с использованием фильтра, такого как мембрана, связывающаяся с низкомолекулярным белком.
25. Способ по п. 23 или 24, где дефектные по репликации частицы ретровирусного вектора выделяют не более чем через 72 часа после стадии введения (a).
26. Способ по п. 25, где дефектные по репликации ретровирусные частицы выделяют в течение периода времени от 48 до 72 часов после стадии введения (a).
27. Способ по п. 19, в котором клетка-хозяин млекопитающих представляет собой суспензионно адаптированую/неадгезивную клетку.
| КОМПОЗИЦИИ И СПОСОБЫ ДОСТАВКИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА | 1994 |
|
RU2174845C2 |
| Замочный механизм висячего замка | 1929 |
|
SU18124A1 |
| ФОРМУЛА КОШАЧЬЕЙ ВАКЦИНЫ | 2002 |
|
RU2312676C2 |
| СПОСОБ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ ИНТЕРЕСУЮЩЕГО ЭНДОГЕННОГО ГЕНА ИЛИ ХРОМОСОМНОГО ЛОКУСА (ВАРИАНТЫ) И ЕГО ИСПОЛЬЗОВАНИЕ | 2001 |
|
RU2290441C2 |
| Способ нагрева прессующего полотна непрерывно работающего пресса и устройство для его осуществления | 1990 |
|
SU1829963A3 |
| Приспособление для записи прогибов железнодорожного пути | 1927 |
|
SU12723A1 |
| ПОЛИВАЛЕНТНЫЕ ВИРУСНЫЕ ВЕКТОРЫ И СИСТЕМА ДЛЯ ИХ ПОЛУЧЕНИЯ | 2005 |
|
RU2416646C2 |
| WO 2012028681 A1, 08.03.2012 | |||
| КАПСУЛА ДЛЯ КЛЕТОК, ПРОДУЦИРУЮЩИХ ВИРУСНЫЕ ЧАСТИЦЫ, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ИСПОЛЬЗОВАНИЯ | 1996 |
|
RU2187301C2 |

