Область техники
Изобретение относится к медицинской и молекулярной генетике, а именно к генетическим конструкциям, экспрессирующим РНК-последовательности и/или гены, кодирующие белки, обладающие антивирусной активностью в отношении вируса иммунодефицита человека, и могут быть использованы в научных исследованиях.
Уровень техники
ВИЧ инфекция остается огромной проблемой для здравоохранения и общества в целом. Число ВИЧ инфицированных продолжает неуклонно расти. Современная высокоактивная антиретровирусная терапия (ВААРТ) позволяет эффективно сдерживать прогрессирование заболевания и значительно повышает уровень жизни ВИЧ-инфицированного пациента. Однако ВААРТ является дорогостоящим пожизненным лечением и приводит к накоплению побочных эффектов, связанных с токсичностью используемых химических препаратов. Кроме того, проблемой ВААРТ является возникновение лекарственно устойчивых мутантных штаммов, что приводит к необходимости в постоянной разработке и использовании все большего количества новых химических препаратов.
Генная терапия ВИЧ подразумевает введение в клетки генов, подавляющих развитие инфекции. На данный момент на клеточных культурах опробовано множество генетических конструкций, в той или иной мере усиливающих устойчивость клеток к ВИЧ. Усовершенствование клинических методов изоляции и трансплантации клеток позволяет рассматривать генотерапию как перспективное направление антиВИЧ терапии.
Патент RU 2426788 («Генетические конструкции для антиВИЧ терапии», 20.08.2011; МПК C12N 15/86) раскрывает генетические конструкции на основе модифицированного лентивирусного вектора, содержащие последовательность, кодирующую модифицированный белок TRIM5α (tripartite motif 5 а) человека, где модификация включает изменение аминокислотной последовательности TRIM5α в области SPRY домена, которое обеспечивает распознавание белком TRIM5α вирусного капсида ВИЧ. TRIM5α - белковый фактор врожденного иммунитета, обуславливающий устойчивость к ретровирусам. Различие между приматами в аминокислотном составе домена SPRY белка TRIM5 определяет различие в наборе ретровирусов, к которым устойчив данный вид. Гомолог TRIM5α у резус-макак обеспечивает устойчивость клеток этих животных к заражению ВИЧ. TRIM5α человека не способен распознавать ВИЧ, однако небольшое изменение аминокислотной последовательности позволяет модифицировать белок таким образом, что он приобретает противовирусную антиВИЧ активность.
Задачей изобретения согласно патенту RU 2426788 является расширение арсенала средств нового поколения, предназначенных для лечения ВИЧ-инфекции. Оно направлено на создание более эффективных антиВИЧ препаратов на основе РНК и белков, продуцируемых в клетках, с помощью введенных генетических конструкций, предложенных по изобретению. В RU 2426788 также продемонстрировано, что комбинирование двух и более антивирусных агентов, перечисленных ниже, в одной генетической конструкции обеспечивает синергетический эффект: каждый агент по отдельности в различной степени подавляет репродукцию ВИЧ (от 50% до 90%); одновременное использование нескольких противовирусных агентов замедляет образование мутантных штаммов, а комбинация противовирусных агентов с агентами, придающими резистентность клеткам к ВИЧ, позволяет получить близкую к 100% антивирусную активность препарата и предотвратить появление устойчивых мутантов. В качестве одного из агентов, придающих клетке устойчивость к ВИЧ, авторами были выбраны короткие интерферирующие РНК, направленные против гена хемокинового рецептора CCR5, задействованного в процессе проникновения ВИЧ в клетку. Техническим результатом является создание предложенных конструкций и реализация ими указанного назначения. Частные варианты выполнения изобретения включают генетические конструкции для антиВИЧ терапии на основе лентивирусного вектора, в том числе и самоинактивирующегося лентивирусного вектора, где лентивирусный вектор создан на основе одного или нескольких из лентивирусов: ВИЧ-1, ВИЧ-2, ВИО (вирус иммунодефицита обезьян, SIV), вирус иммунодефицита крупного рогатого скота (BIV), лентивирус артрита-энцефалита коз (CAEV), вирус инфекционной анемии лошадей (EIAV), вирус иммунодефицита кошек (FIV) и проч.
Заявка на патент США US2012076763 ("Combination anti-HIV vectors, targeting vectors, and methods of use"; 29.03.2012; МПК A61K 48/00, C12N 5/071, C12N 15/49, A61P 31/18, C12N 5/10, C12Q 1/70, C12N 15/63, C12N 7/01) раскрывает рекомбинантные лентивирусные векторы, включающие следующие элементы: лентивирусную основу, содержащую последовательности для интеграции в геном клетки-мишени; нуклеиновые кислоты, кодирующие интерферирующую РНК, направленную против гена рецептора человека CCR5; а также элемент контроля экспрессии нуклеиновой кислоты, кодирующей RNAi CCR5. Векторы также могут содержать полинуклеотиды, кодирующие последовательности TRIM5α и TAR-ловушки ВИЧ вместе с элементами регуляции экспрессии генов, и сочетаться с упаковочными плазмидами и сопряженными специфическими антителами против клеток-мишеней. В описании указанной заявки приводится полинуклеотид, кодирующий последовательность химерный TRIM5α, где последовательность 13 аминокислот резус-макаки вставлена в последовательность белка TRIM5α человека (SEQ ID NO 17, 994-1032). Полученный химерный TRIM5α эффективно ингибирует вирус ВИЧ-1 на стадии после проникновения вируса в клетку и перед его интеграцией. В заявке также приведены диагностические и терапевтические методы использования указанных конструкций.
В заявке на патент США US2012270773 ("TRIM5alpha mutants and uses thereof; 25.10.2012; МПК С07Н 21/04, A61K 31/7088, C12N 5/02, C12Q 1/70, C07K 16/18, C07K 14/435, A61K 38/00, C12N 15/63, C12N 5/10) описывается мутантный полипептид TRIM5α, придающий более высокую устойчивость лентивирусам по сравнению с диким типом TRIM5α человека. Указанный мутантный полипептид TRIM5α содержит мутацию в аминокислоте (замена или делеция), соответствующую аминокислоте 324, 328, 330, 333, 335, 336 или 337 дикого типа TRIM5α человека. В указанной заявке также описано выделение нуклеиновой кислоты, содержащей последовательности, кодирующие вышеупомянутые полипептиды TRIM5α, или их гомологи. Заявка раскрывает композиции, содержащие мутантные полипептиды TRIM5α, кодирующие их нуклеиновые кислоты, векторы для переноса генов, а также способы определения (in vivo или in vitro) устойчивости клеток к лентивирусной инфекции, включающие определение в биологическом образце наличия или отсутствия мутантного TRIM5α и/или кодирующих их нуклеиновые кислоты. Показано, что клетки, содержащие мутантные полипептиды TRIM5α, имеют повышенную устойчивость к лентивирусной инфекции.
В работе Андерсона (J.S. Anderson, J. Javien, J.A. Nolta, and G. Bauer "Preintegration HIV-1 Inhibition by a Combination Lentiviral Vector Containing a Chimeric TRIM5α Protein, а CCR5 shRNA, and a TAR Decoy", Molecular Therapy, 2009, vol. 17(12), pp.2103-2114) описан комбинированный лентивирусный вектор, кодирующий три высокоэффективных антиВИЧ гена и функционирующий на отдельных этапах жизненного цикла вируса:
- CCR5 короткие шпилечные РНК (shRNA) (перед проникновением вируса),
- химерный TRIM5α человека/ резус-макаки (на стадии после проникновения вируса ВИЧ в клетку и перед его интеграцией),
- TAR-ловушки (после интеграции).
Основные усилия ученых при дизайне указанного антиВИЧ вектора были сосредоточены на блокировке продуктивного заражения ВИЧ-1 и ингибировании любого образования провируса, который будет поддерживать вирусный резервуар.
Э. Бативелли и коллеги (Е. Battivelli, J. Migraine, D. Lecossier et al. "Modulation of TRIM5α Activity in Human Cells by Alternatively Spliced TRIM5 Isoforms", J. Virol., 2011, vol. 85(15), pp.7828-7835) установили, что активность TRIM5α в конкретном типе клеток может зависеть от соотношения экспрессированных изоформ TRIM5. Кроме TRIM5α могут присутствовать транскрипты TRIM5ι, TRIM5γ, TRIM5δ и TRIM5κ. Они не ингибируют репликацию ВИЧ-1, но обладают доминантно-негативной активностью в отношении TRIM5α. Авторами показано, что экспрессия TRIM5i уменьшает противовирусную активность TRIM5α в человеческих клетках U373-X4, что указывает на то, что физиологические уровни экспрессии усеченных изоформ TRIM5 в клетках человека могут снижать активность TRIM5α.
Задачей настоящего изобретения является усовершенствование генетических конструкций, содержащих последовательность, кодирующую модифицированный белок TRIM5α человека, с целью повышения уровня экспрессии гена, и, следовательно, количества белка, что приведет к повышению противовирусной эффективности данных конструкций в отношении вируса иммунодефицита человека. Технический результат - создание усовершенствованных генетических конструкций и реализация ими указанного назначения. Предлагаемые в объекте исследования генетические конструкции, кодирующие модифицированный белок TRIM5α, отличаются от известных генетических конструкций тем, что в последовательности модифицированного гена TRIM5α размером 1488 п.н. (496 кодонов) были произведены синонимичные замены нуклеотидов по всей длине таким образом, что процентный состав суммы всех гуанинов (G) и цитозинов (С) по отношению к общему числу нуклеотидов (GC-состав) последовательности увеличился.
Согласно результатам исследования, антиВИЧ активность конструкции, включающей последовательность (SEQ ID N1), кодирующую модифицированный белок TRIM5α человека, почти в 40 раз выше по сравнению с активностью аналогичной конструкции, включающей последовательность (SEQ ID N2), кодирующую модифицированный белок TRIM5α человека, описанной в патенте RU 2426788. Кроме того, увеличилась жизнеспособность клеток в присутствии ВИЧ.
Раскрытие изобретения
В настоящем изобретении "молекула нуклеиновой кислоты" и "полинуклеотид" означают полимерную форму нуклеотидов любой длины, либо дезоксирибонуклеотиды, либо рибонуклеотиды, либо их аналоги. Полинуклеотиды могут иметь трехмерную структуру и могут осуществлять любую функцию. Неограничивающими примерами полинуклеотидов являются ген, генный фрагмент, экзоны, интроны, открытая рамка считывания, матричная РНК (мРНК), транспортная РНК, рибосомная РНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, космиды, векторы, выделенная ДНК с любой последовательностью, выделенная РНК с любой последовательностью, нуклеиновокислотные зонды и праймеры.
Полинуклеотид обычно состоит из специфической последовательности из четырех нуклеотидных оснований: аденина (А), цитозина (С), гуанина (G) и тимина (Т) (а в случае, если указанным полинуклеотидом является РНК, то вместо тимина присутствует урацил (U)). Таким образом, термин "полинуклеотидная последовательность" означает буквенное представление полинуклеотидной молекулы. Это буквенное представление может быть введено в базу данных компьютера, имеющего центральный процессор, и используется в биоинформатике, например в функциональной геномике и в поиске гомологии.
Термин "конструкция" означает любую молекулу, способную переносить последовательности нуклеиновой кислоты (например, невирусные векторы, частицы-носители, липосомы и вирусные векторы) в клетки-мишени. Термин "плазмидная конструкция" означает внехромосомный генетический элемент, способный к саморепликации в клетке-хозяине. Обычно термины "вектор", "конструкция", "экспрессирующий вектор" и "вектор для переноса генов" означают любую конструкцию нуклеиновой кислоты, способную регулировать экспрессию нужного гена и переносить генные последовательности в клетки-мишени. Таким образом, этот термин включает в себя клонирующие и экспрессирующие носители, а также вирусные векторы.
Вектор может содержать либо ДНК, либо РНК. Например, для получения вектора может быть использован либо ДНК-, либо РНК-вирус. На основе геномной РНК-вируса может быть получена копия кДНК. И наоборот, фрагмент кДНК (или вирусной геномной ДНК) может быть транскрибирован in vitro с образованием РНК. Эти методики хорошо известны специалистам.
Термин "кодирующая последовательность" или последовательность, которая "кодирует" выбранный полипептид, означает молекулу нуклеиновой кислоты, которая транскрибируется (в случае ДНК) и транслируется (в случае мРНК) в полипептид in vivo при ее нахождении под контролем соответствующих регуляторных последовательностей (или "регуляторных элементов"). Границы кодирующей последовательности определены старт-кодоном у 5'(амино)-конца и кодоном терминации трансляции у 3'(карбокси)-конца. Кодирующей последовательностью может быть, не ограничиваясь ими, кДНК, происходящая от вирусной, прокариотической или эукариотической мРНК, геномные ДНК-последовательности, происходящие от вирусной или прокариотической ДНК, и даже синтезированные ДНК-последовательности. Последовательность терминации транскрипции может быть локализована со стороны 3'-конца по отношению к кодирующей последовательности. Транскрипция и трансляция кодирующих последовательностей обычно регулируются "регуляторными элементами", включая, но не ограничиваясь ими, промоторы транскрипции, энхансерные элементы транскрипции, последовательности Шайна-Дальгарно, сигналы терминации транскрипции, последовательности полиаденилирования (локализованные со стороны 3'-конца по отношению к кодону терминации трансляции), последовательности для оптимизации инициации трансляции (локализованные со стороны 5'-конца по отношению к кодирующей последовательности) и последовательности терминации трансляции. Термин "элемент регуляции экспрессии генов" означает нуклеотидную последовательность, содержащую, как минимум, промотор. Такой элемент может, кроме того, содержать другие последовательности, необходимые для экспрессии кодирующих последовательностей, функционально присоединенных к промотору, или влияющие на такую экспрессию. Компоненты этих элементов необязательно должны быть смежными, то есть они могут быть разделены промежуточными последовательностями. Компоненты этих элементов могут влиять на экспрессию кодирующих последовательностей на уровне транскрипции, стабильности РНК, процессинга и/или трансляции РНК. Обычно такой элемент не содержит кодирующих последовательностей, с которыми они функционально связаны. В некоторых случаях, элемент регуляции экспрессии гена может содержать природный ген либо, в основном, состоять из такого гена, кроме кодирующей последовательности данного гена.
Векторы для генной терапии ВИЧ
Главным требованием генной терапии является устойчивая экспрессия терапевтических генов, не вызывающая негативных клинических эффектов. Крайне желательно иметь вектор (последовательность, обеспечивающую встраивание ДНК последовательностей в геном клетки человека) с высоким титром, который способен стабильно интегрировать в клетки-мишени (включая делящиеся и неделящиеся клетки) и который не является патогенным и не вызывает иммунной реакции.
Ретровирусные векторные системы
Одной из первых систем для генотерапии был ретровирусный вектор, сконструированный на основе вируса мышиного лейкоза Moloney (MoMLV). Этот вектор обладает способностью инфицировать делящиеся клетки. ДНК-провирус MoMLV способен интегрироваться в геном делящихся клеток и передаваться по наследству вместе с клеточным геномом. В целом, клинические испытания с помощью векторных систем на основе вируса MoMLV составляют более 23% от клинических испытаний всех систем генотерапии различных заболеваний человека.
Векторы на основе спумовируса (foamy virus)
В настоящее время для генной терапии разработаны векторы на основе спумавируса. Спумавирусы - не патогенные, интегрирующие ретровирусы обладающие свойствами отличающими их от лентивирусов и гамма-ретровирусов. Такие векторы позволяют упаковывать и переносить большие фрагменты ДНК и эффективно трансдуцируют гемопоэтические клетки. Кроме того преимуществом данного вектора для генной терапии ВИЧ инфекции является низкая гомология нуклеотидной последовательности с вирусом иммунодефицита человека и, благодаря которой, резко понижается риск мобилизации вектора, обеспечивая безопасность терапии. Способность эффективно переносить антиВИЧ трансгены, которые не могут быть перенесены с помощью векторов на основе ВИЧ-1, является ключевым преимуществом этого вектора. Показано, что на титр вектора на основе спумавируса не влияют кассеты трансгенов, которые значительно уменьшают титр лентивирусного вектора. Сайты интеграции данного вектора значительно отличаются от сайтов интеграции лентивирусов и гамма-ретровирусов, что может уменьшить риск возникновения лейкемии при использовании в генной терапии.
Невирусные системы доставки генов - вектор на основе транспозопов
Невирусные системы доставки генов (с помощью ДНК плазмид) являются наиболее простыми и безопасными, однако эффективность таких способов долгое время оставалась очень низкой. На данный момент существуют несколько экспрессионных векторов на основе транспозонов, некоторые из которых обладают эффективностью переноса ДНК, сравнимой с ретровирусными системами. Примером такого вектора может служить транспозонная система Спящей красавицы (Sleeping Beauty). Эта система состоит из искусственного транспозона и соответствующей транспозазы, фермента, который обладает функцией разрезания и встраивания фрагментов ДНК. Процесс переноса гена состоит из двух ступеней: на первой транспозаза распознает короткие инвертированные или прямые повторы в составе транспозона, а на второй вырезает и встраивает транспозон в геномную ДНК, в области ТА-повторов. В системе транспозонов Спящей красавицы могут быть использованы две независимые плазмиды, кодирующие транспозон и транспозазу или оба компонента могут быть объединены на одной плазмиде. Усовершенствование фермента транспозазы привело к увеличению эффективности доставки генов с помощью этой системы. Показана возможность переноса генов в различные клетки млекопитающих, включая эмбриональные клетки мышей. Использование векторов транспозон/транспозаза представляет собой альтернативный способ доставки трансгена для генной терапии ВИЧ.
Лентивирусные векторы
На сегодняшний день лентивирусные векторы на основе ВИЧ-1 наиболее часто используются в генной терапии и молекулярно-биологических исследованиях, так как строение генома этого вируса детально изучено, что значительно облегчает проведение генноинженерных манипуляций. Однако также сконструированы векторы на основе других лентивирусов: ВИЧ-2, ВИО, BIV, CAEV, EIAV, FIV, вирус болезни Джембрана (JDV). Такие векторы потенциально могут обладать определенными преимуществами, такими как повышенная безопасность, так как они не могут инфицировать человека, а значит понижается вероятность возникновения репликационно-компетентных вирусных частиц. Кроме того многие антиВИЧ агенты действующие против вируса на этапе упаковки значительно снижают титр вирусных частиц при использовании векторов на основе ВИЧ-1, но не влияют на титр вирусных частиц на основе других лентивирусов.
Вирусы подсемейства лентивирусов обладают уникальной способностью стабильно переносить генетический материал в неделящиеся клетки. В отличие от MuLV и других γ-ретровирусов лентивирусные векторы обычно встраиваются после точки начала транскрипции, и, таким образом, не могут активировать онкогены. Широкомасштабный скрининг сайтов интеграции лентивирусов показал отсутствие злокачественных перерождений клеток. Разработка новых векторов на основе лентивирусов (в частности, ВИЧ) открывает широкие возможности в генной терапии ВИЧ. К РНК-вирусам подсемейства Lentivirus относят вирусы иммунодефицита человека типов 1 и 2 (т.е. ВИЧ-1 или ВИЧ-2, при этом ВИЧ-1 раньше называли с лимфаденопатией вирусом 3 (HTLV-III) и вирусы, ассоциированные с синдромом приобретенного иммунодефицита (СПИД) (ARV)), или другие вирусы, близкие ВИЧ-1 или ВИЧ-2, которые были идентифицированы и ассоциированы со СПИДом или СПИД-подобным заболеванием. Кроме того, к РНК-вирусам подсемейства Lentivirus относят вирус Висны/Маэди (например, инфицирующий овец), вирус иммунодефицита кошек (FIV), бычий лентивирус, вирус иммунодефицита обезьян (SIV), вирус инфекционной анемии лошадей (EIAV) и вирус артрита-энцефалита коз (CAEV).
Требования к вирусному вектору включают наличие промоторных элементов, размер генома, позволяющий упаковывать инородный генетический материал, и отсутствие вирулентных детерминант. Для уменьшения патогенности при производстве рекомбинантных вирусных векторов поступление структурных генов (кодирующих белки, образующие оболочку вируса, нуклеокапсид, и т.д.) обеспечивается in trans путем интеграции в геном упаковочных клеток, либо на плазмиде, вводимой в клетки совместно с вектором.
Репликация, интеграция и упаковка вирусных частиц требует наличия РНК (ДНК) областей, которые не кодируют белков. Большинство из таких цис-элементов необходимо включать в лентивирусный вектор (табл.1).
Структурные гены, которые поставляются in trans, включают три группы белков gag, pol и env (табл.2). Эти гены входят в состав нескольких отдельных упаковочных плазмид. Удаление таких генов из состава вектора не дает ему возможность самостоятельно реплицироваться и производить вирусные частицы, тем самым увеличивая безопасность применения таких векторов.
Векторы по настоящему изобретению также могут представлять собой самоинактивирующиеся лентивирусные векторы, которые придают конструкциям свойство самоинактивироваться и обеспечивают дополнительную безопасность при проведении терапии. При конструировании самоинактивирующихся векторов в 3'-LTR векторной ДНК (провирус) вносят делецию, затрагивающую энхансерно-промоторную область (она находится в U3-районе LTR), т.е. конструируют 3'LTR с делецией в области U3. После заражения клетки мишени из-за особенностей обратной транскрипции образуется провирус, у которого оба LTR лишены промоторно-энхансерной области. Таким образом, после интеграции в геном клетки-мишени самоинактивирующийся вектор не способен к репликации, поэтому вероятность возникновения репликационно-компетентного вируса (RCV) в культуре трансдуцированных клеток ничтожно мала. Кроме того исключается возможность инсерционного мутагенеза, так как отсутствие сильных энхансерно-промоторных участков на 3' конце вектора (которые находятся в делегированном участке LTR) предотвращает активацию собственных генов клетки, в том числе онкогенов.
Для получения векторных транскриптов в упаковочных клетках вместо вирусного промотора-энхансера в U5-pauone LTR используется RSV/5'-LTR гибридный промотор. Это позволяет не вводить дополнительно плазмиду, кодирующую вирусный белок tat, и делает систему еще более безопасной.
Согласно настоящему изобретению в предложенных генетических конструкциях могут быть использованы любые известные векторы с последовательностью, обеспечивающей встраивание ДНК последовательностей в геном клетки человека, с высоким титром, которые способны стабильно интегрировать в клетки-мишени (включая делящиеся и неделящиеся клетки) и которые не является патогенным и не вызывают иммунной реакции.
Доставка генов в заданные локусы генома с помощью эндонуклеаз
В настоящее время созданы искусственные эндонуклеазы, способные специфически распознавать и связываться с протяженными участками ДНК. Такие эндонуклеазы могут вносить разрыв в уникальный участок генома, что приводит к активации процессов репарации хромосомы, которые могут идти либо по пути негомологичного соединения концов ДНК, либо по пути гомологичной рекомбинации. Негомологичное соединение концов в большинстве случаев сопровождается делецией нуклеотидов или встраиванием дополнительных нуклеотидов, что приводит к сдвигу рамки считывания и нарушению функции соответствующего гена. Гомологичная рекомбинация может иметь место, если в ядре присутствует донорская двухцепочечная ДНК, имеющая гомологию с последовательностями, находящимися в непосредственной близости от места разрыва. В результате происходит встраивание донорской последовательности в область разрыва. Таким образом, эндонуклеазы, вносящие разрыв в уникальный участок генома, могут быть использованы для внесения мутаций в определенные гены, исправлению мутаций в генах за счет гомологичной рекомбинации или доставки целевых генов в заданные области генома. Преимуществом такой доставки целевых генов является известная локализация встраиваемой последовательности, что очень важно для понимания дальнейшей регуляции целевого гена и позволяет избежать генотоксичности, вероятность которой существует при случайном встраивании ДНК последовательности в геном.
До недавнего времени использовали два класса нуклеаз - мегануклеазы и нуклеазы с цинковыми пальцами. Существенным ограничением для использования данного метода доставки являлась трудоемкость создания этих нуклеаз, вносящих разрыв в выбранный участок генома. Открытие белков, подобных транскрипционным активаторам (TALE), которые были выделены из бактерии Xanthomonas, легло в основу создания нового класса нуклеаз TALE-нуклеаз или TALEN. Такие нуклеазы имеют модульную структуру. Они состоят из повторяющихся доменов в 33-35 аминокислот, причем две аминокислоты в середине повтора являются вариабельными и определяют нуклеотид, с которым связывается домен. Последовательное соединение доменов, однозначно определяет ДНК последовательность, с которой связывается нуклеаза. Таким образом, использование описанных нуклеаз позволяет осуществлять доставку генетических конструкций в заданный участок генома.
Промоторы
Последовательности, кодирующие генетические антивирусные агенты, могут быть включены в векторную частицу под контролем соответствующих промоторов, известных для специалиста в данной области.
Термин "промотор" означает нуклеотидную последовательность, которая контролирует инициацию и скорость транскрипции полинуклеотида. Промотор содержит элементы, с которыми могут связываться регуляторные белки и молекулы, такие как РНК-полимераза и другие факторы транскрипции, для инициации специфической транскрипции последовательности нуклеиновой кислоты. Выражения «функционально расположенный», «функционально связанный», «под контролем» и «под транскрипционным контролем» означают, что промотор находится в корректном функциональном расположении и/или ориентации по отношению к последовательности нуклеиновой кислоты для контроля инициации транскрипции и/или экспрессии такой последовательности.
Промотор РНК-полимеразы II человека
Транскрипция всех кодирующих последовательностей эукариот находится под контролем промотора РНК-полимеразы II и приводит к образованию полностью зрелой мРНК, имеющей кэп на 5' конце и полиА на 3' конце последовательности мРНК. Промотор, в общем, содержит последовательность, которая определяет положение начала синтеза РНК. Лучшим известным примером такой последовательности является ТАТА-бокс, но в некоторых промоторах, лишенных ТАТА-бокса, таких как, например, промотор гена терминальной дезоксинуклеотидилтрансферазы млекопитающего и промотор поздних генов SV40, другой элемент, прилегающий к участку старта, способствует точному определению места инициации. Дополнительные промоторные элементы регулируют частоту инициации транскрипции. Обычно они локализованы в области на 30-110 н.п. выше участка старта, хотя некоторые промоторы, как было показано, содержат также функциональные элементы ниже участка старта. Для помещения кодирующей последовательности «под контроль промотора» располагают 5'-конец участка инициации транскрипции в транскрипционной рамке считывания «ниже» (т.е. в 3'-направлении) выбранного промотора.
Действие промотора может быть усилено «энхансером». Под энхансером подразумевают специфическую цис-действующую последовательность нуклеотидов, многократно усиливающую транскрипцию генов РНК-полимеразой II; например, энхансер вируса SV40 (размер - 72 пары нуклеотидов) может усиливать транскрипцию бета-глобинового гена в 200 раз, даже находясь на значительном удалении от него и в любой ориентации по отношению к промотору. Способность ряда энхансеров взаимодействовать со специфическими белками в дифференцированных клетках обеспечивает тканеспецифичный характер экспрессии соответствующих генов.
Естественно, важно использовать промотор и/или энхансер, который эффективно направляет экспрессию сегмента ДНК в органелле, клеточном типе, ткани, органе или организме, выбранном для экспрессии. Специалистам в области молекулярной биологии, в основном, известно применение промоторов, энхансеров и комбинаций клеточных типов для экспрессии белка (см., например, Sambrook et al., 1989, включенный сюда в качестве ссылки). Используемые промоторы могут быть конститутивными, тканеспецифичными, индуцируемыми и/или могут использоваться в условиях, подходящих для направления высокоуровневой экспрессии введенного сегмента ДНК. Промотор может быть гетерологичным или эндогенным.
Конститутивные промоторы
К конститутивным промоторам, т.е. промоторам работающим во всех типах клеток не зависимо от условий, относятся промоторы генов «домашнего хозяйства» (housekeeping genes). К генам «домашнего хозяйства» относятся гены, обеспечивающие жизнедеятельность эукариотической клетки и функционирующие повсеместно, на всех стадиях жизненного цикла организма. Они обеспечивают процесс гликолиза, биосинтез аминокислот и нуклеотидов, катаболизм белков и т.п. Неограничивающими примерами таких промоторов являются промотор гена фактора элонгации EF1α, гена фосфоглицераткипазы (PGK), гена β-актина, убиквитиновый промотор UbC, гистоновый промотор Н2 В (ТН2 В) и др.
Многие вирусные промоторы выполняют конститутивную функцию в эукариотических клетках. Эти промоторы включают:
- ранние и поздние промоторы SV40 (см. Bemoist and Chambon, Nature, 290:304 (1981));
- длинные концевые повторы (LTR) вируса лейкоза Молонея и других ретровирусов (см. R.Weisset et al., eds., Molecular Biology of Tumor Viruses, 2nd ed., RNA Tumor Viruses. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1985);
- промотор тимидинкиназы вируса простого герпеса (HSV) (см. Wagner et al., Proc. Natl. Acad. Sci. USA, 1981, vol.78, p.1441);
- самый ранний промотор цитомегаловируса (IE1) (см. Karasuyama et al., J. Ехр. Med., 1989, vol. 169 р.13;
- промотор вируса саркомы Рауса (RSV) (см. Yamamoto et al., Cell, 1980, vol.22, P.787);
- главный поздний промотор аденовируса (см. Yamada et al., Proc. Natl. Acad. Sci. USA, 1985, vol.82, p.3567) и многие другие.
Поэтому любые вышеуказанные конститутивные промоторы можно использовать для контроля транскрипции генной вставки.
Тканеспецифичиые промоторы
Тканеспецифичными называются промоторы, которые инициируют транскрипцию гена только в клетках определенной ткани. Тканеспецифический характер экспрессии генов обеспечивается различными механизмами взаимодействия белковых факторов транскрипции с регуляторными последовательностями нуклеиновых кислот. Отличительные черты тканеспецифических промоторов или элементов, а также анализы для характеристики их активности хорошо известны специалистам в данной области. Неограничивающие примеры генов под контролем таких промоторов охватывают человеческий ген LIMK2 (Nomoto et al., 1999), ген соматостатинового рецептора 2 (Kraus et al., 1998), ген связывающего ретиноевую кислоту белка придатков яичка мыши (Lareyre et al., 1999), человеческий CD4 (Zhao-Emonet et al., 1998), мышиный альфа-2-(XI)-коллаген (Tsumaki, et al., 1998), ген дофаминового рецептора D1A (Lee, et al., 1997), инсулиноподобный фактор роста II (Wu et al., 1997) и молекулу-1 адгезии тромбоцитов-эндотелиальных клеток человека (Almendro et al., 1996).
Ниже перечислены неограничивающие примеры тканеспецифичных элементов/промоторов РНК-полимеразы II, которые могут использоваться в контексте настоящего изобретения для регуляции экспрессии РНК: промотор гена тяжелой цепи иммуноглобулина человека, гена Т-клеточного рецептора, гена β-интерферона, гена IL2. гена рецептора IL2, генов МНС класса II, гена а-фетопротеина, гена γ-глобина, гена β-глобина, генов кодирующих α- и β-субъединицы гемоглобина человека, гена альбумина, гена коллагеназы, гена металлотионеина (MTII), гена эластазы I, гена c-HA-ras, гена инсулина, и др.
Регулируемые промоторы
Регулируемые или индуцибельные промоторы позволяют включать и/или выключать транскрипцию гена. Активность таких промоторов зависит от введения дополнительных факторов, в норме отсутствующих в экспрессирующих клетках. Существует несколько систем обратимого регулирования экспрессии, которые позволяют включать и выключать работу гена. Наиболее используемой и изученной является тетрациклин зависимая система переключения, которая состоит из трех основных частей: молекулы тетрациклина Tet или его аналога, молекулы белка тетрациклинового репрессора TetR или активатора tTA и участка промотора (т.н. тетрациклинового оператора tetO) связывающего репрессор или активатор. Существуют два варианта: а) в системе Tet-On используется репрессор TetR, связывающийся с оператором и блокирующий транскрипцию; добавление тетрациклина приводит к удалению репрессора и включение экспрессии гена б) в системе Tet-OFF, используется активатор tTA, способствующий экспрессии в отсутствие тетрациклина и выключающий работу гена при добавлении тетрациклина. Аналог тетрациклина доксициклин лучше проникает через клеточную мембрану и обладает большим сродством с репрессором и активатором, поэтому чаще используется в качестве переключателя. Другими примерами регулируемых промоторов могут служить LacZ регулируемая система или экдизон регулируемая система. Единственным примером необратимой индукции (или репрессии) является система, базирующаяся на Cre-Lox рекомбинации бактериофага Р1. Использование Cre-Lox рекомбинации позволяет необратимо включить или выключить экспрессию интересующей последовательности в генной конструкции после введения ее в клетку.
Промоторы полимеразы III человека
Для экспрессии коротких РНК-последовательностей (в нашем случае рибозим, TAR, shRNA) удобно использовать промоторы для РНК-полимеразы III. Промоторы для РНК-полимеразы III делятся на три типа, основываясь на композиции промоторных элементов и их положения относительно старта транскрипции (PAULE MR, WHITE RJ. Survey and summary: transcription by RNA polymerases I and III. Nucleic Acids Res 2000; 28(6): 1283-98). Промоторы 3-го типа для РНК-полимеразы III наиболее подходят для транскрипции коротких РНК-последовательностей, поскольку в природе они используются клеткой для наработки малых ядерных РНК-молекул. В отличие от промоторов 1-го и 2-го типа, промоторы третьего типа полностью расположены выше транскрипционного старта и содержат 3 специфических промоторных элемента, необходимые для их функционирования: 1. ТАТА-последовательность в позиции -20 п.о., 2. PSE последовательность проксимального элемента в позиции -50 п.н., 3. DSE последовательность дистального элемента в позиции -240 п.н. (где +1 - первый транскрибируемый нуклеотид). Оба элемента DSE и PSE содержат энхансер и последовательность связывающую транскрипционный фактор, которые абсолютно необходимы и делеция которых полностью элиминирует транскрипцию. Расстояние между этими элементами строго определено и критично для достижения максимальной скорости транскрипции. Характерной особенностью данных промоторов является четко определенный сайт инициации транскрипции и простой сигнал терминации состоящий из 4-х или 5-ти последовательных тимидинов (SCHRAMM L et al. Recruitment of RNA polymerase III to its target promoters. Genes Dev 2002; 16(20): 2593-620). Относительная простота и высокий уровень транскрипции (4*105 транскриптов на клетку) делает их максимально удобными для использования в генетических конструкциях.
К промоторам 3-го типа для РНК-полимеразы III относятся промоторы генов HI, U6 и 7SK, которые обычно используются в экспериментах для экспрессии shRNA. Ген HI кодирует РНК, которая является компонентом ядерной РНКазы Р и вовлечена в созревание тРНК. HI промотор человека является необыкновенно компактным - все элементы необходимые для транскрипции находятся в пределах 100 п.о. выше старта. РНК, кодируемая геном U6 играет центральную роль в процессинге РНК и является неотьемлимой частью сплайсосомы (BERGET SM, ROBBERSON BL. U1, U2, and U4/U6 small nuclear ribonucleoproteins are required for in vitro splicing but not polyadenylation. Cell 1986; 46(5): 691-6). Эксперименты in vivo и in vitro показывают, что U6 промотор намного активнее, чем HI. Однако по-аналогии с U1 РНК для U6 РНК предполагают наличие регуляторного механизма, который предотвращает избыточное накопление U6 РНК в клетке Noonberg SB, Scott GK, Benz CC. Evidence of posttranscriptional regulation ofU6 small nuclear RNA. J Biol Chem 1996; 271(18): 10477-81). Поэтому сверхэкспрессия shRNA контролируемая U6 промотором может приводить к уменьшению количества эндогенной U6 РНК и изменению фенотипа клетки. Все более популярен становится промотор 7SK, как более сильный по сравнению с U6, но не обладающий механизмом саморегуляции. 7SK РНК эволюционно консервативна и присутствует в клетке в большом количестве. Существуют искусственно созданные регулируемые промоторы РНК-полимеразы III, основанные на описанных выше тетрациклиновой и Cre-Lox системе.
В общем промоторы и энхансеры, которые контролируют транскрипцию в эукариотических клетках, составлены из множественных генетических элементов. Клеточный аппарат способен объединить и интегрировать регуляторную информацию, которую несет каждый элемент, что позволяет образовывать комплексные профили транскрипционной регуляции. Активация или репрессия промоторных и энхансерных элементов может осуществляться путем контакта данных элементов с подходящими белковыми активаторами или репрессорами транскрипции.
В контексте данного изобретения может использоваться любая комбинация элементов промотор/энхансер (согласно базе данных эукариотических промоторов EPDB) приводящая к эффективной инициации и поддержанию транскрипции последовательности в гемопоэтических клетках. Для экспрессии трансгенов могут быть использованы регулируемые промоторы, позволяющие включать/выключать транскрипцию в процессе производства вирусных частиц и/или в процессе терапии ВИЧ инфекции. Специалист на основании данного описания и уровня техники способен подобрать соответствующий промотор для каждого из перечисленных ниже антивирусных агентов или их комбинации.
Генетические антивирусные агенты
Генетические антивирусные конструкции, которые используются в генной терапии, можно разделить на 2 типа: агенты, взаимодействующие с вирусными элементами после проникновения в клетку и начала репликации, и агенты, предотвращающие проникновение вируса в клетку и/или интеграцию в геном, и таким образом придающие клетке-хозяину резистентность (устойчивость) к инфицированию.
Согласно настоящему изобретению предлагаются генетические конструкции, включающие один или несколько нижеописанных антивирусных агентов, одним из которых должна быть нуклеотидная последовательность, кодирующая модифицированный белок TRIM5α человека (SEQ ID N3), и отличающаяся от исходной нуклеотидной последовательности SEQ ID N2 множественными синонимичными заменами, обеспечивающими увеличение процента GC пар в составе ДНК последовательности.
Модифицированная последовательность гена TRIM5α человека
TRIM5α (tripartite motif 5α) - белковый фактор, обуславливающий устойчивость резус-макак к ВИЧ, содержащий 3 различных домена (RING, B-box и RBCC домены). Кроме этих доменов в состав TRIM5α входит С-концевой SPRY домен, связывающийся с капсидом ВИЧ и отвечающий за устойчивость клетки к заражению ВИЧ. Различие в аминокислотном составе этого домена между приматами определяет различие в наборе ретровирусов, к которым устойчив данный вид. Белковые тримеры образуют множественные контакты с поверхностью вирусного капсида и распознают, по-видимому, пространственную структуру. Таким образом, TRIM5α можно назвать распознающим структуру рецептором (PRR) врожденной иммунной системы, направленным на распознавание мультимерной белковой поверхности капсида вируса. Механизмы действия белка пока мало изучены, известно, что он предотвращает интеграцию вирусной ДНК в хромосому, т.е. блокирует ВИЧ на ранней стадии. Предполагается, что блокирование вируса происходит на двух ступенях вирусного цикла: блокирование образования кДНК (до или после обратной транскрипции) и блокирование входа в ядро прединтеграционного комплекса.
Человеческий гомолог белка TRIM5α неэффективен по отношению к ВИЧ, вероятно из-за изменений вирусного капсида в процессе эволюции (Luban J., "Cyclophilin A, TRIM5, and resistance to human immunodeficiency virus type 1 infection", J. Virol., 2007, vol.81(3), pp.1054-1061). Изменения всего одной аминокислоты в последовательности человеческого гомолога TRIM5α делает клетки устойчивыми к ВИЧ инфекции. Таким образом, модифицированный TRIM5α является перспективным для использования в генной терапии - необходимое минорное изменение делает его не иммуногенным и в то же время введение такой модификации в клетки-мишени придает им устойчивость к инфицированию ВИЧ. На основе TRIM5α изоформы человека создан химерный белок, в котором 11 а.о. заменены на 13 а.о. из TRIM5α резуса-макаки (Sawyer S.L. et al., "Positive selection of primate TRIM5α identifies a critical species-specific retroviral restriction domain", Proc. Natl. Acad. Sci. USA, 2005, vol. 102(8), pp.2832-2837).
В заявке W02005081911 ("Methods and compositions for the treatment and prevention ofHIV infection using TRIM5α"; 09.09.2005; МПК С07Н 21/02) раскрываются TRIM5α и генетические конструкции их содержащие.
В патенте RU2426788 описаны генетические конструкции на основе модифицированного лентивирусного вектора, содержащие последовательность, кодирующую модифицированный белок TRIM5α человека, где модификация включает изменение аминокислотной последовательности TRIM5α в области SPRY домена (см. SEQ ID N3), которое обеспечивает распознавание белком TRIM5α вирусного капсида ВИЧ.
Задачей настоящего изобретения является усовершенствование генетических конструкций, содержащих последовательность, кодирующую модифицированный белок TRIM5α человека, с целью повышения уровня экспрессии гена, и, следовательно, количества белка, что приведет к повышению противовирусной эффективности данных конструкций в отношении ВИЧ.
Количество белка, продуцируемого в результате экспрессии гена TRIM5α, напрямую влияет на эффективность подавления репликации ВИЧ. Поэтому очень важно добиться максимально высокого уровня экспрессии гена. В работах (Kudia G., Lipinski L., Caffin F., Helwak A., Zylicz M. "High Guanine and Cytosine Content Increases mRNA Levels in Mammalian Cells", PLoS Biol., 2006, vol. 4(6), 180; Bauer A.P., Leikam D., Krinner S., et al. "The impact of intragenic CpG content on gene expression", Nucleic Acids Res., 2010, vol.38(12), pp.3891-3908; Al-Saif M., Khabar K.S.A., "UU/UA Dinucleotide Frequency Reduction in Coding Regions Results in Increased mRNA Stability and Protein Expression", Mol. Ther., 2012, vol.20(5), pp.954-959) было показано, что уровень экспрессии мРНК зависит от GC состава нуклеотидной последовательности гена: увеличение числа GC за счет синонимичных замен приводило к увеличению экспрессии гена на порядки.
Таким образом, главной задачей настоящего изобретения является создание генетических конструкций, содержащих GC богатую нуклеотидную последовательность, кодирующую модифицированный белок TRIM5α человека.
Модифицированный ген TRIM5α человека может находиться под контролем промотора РНК полимеразы II, который обеспечивает транскрипцию данного трансгена в клетке человека. В предпочтительном осуществлении промотор активен в конкретных клеточных типах или нисходящих ростках предшественников кроветворных клеток. Действие промотора может активироваться или подавляться факторами контроля транскрипции, активаторами или репрессорами.
В качестве промотора может быть использован промотор гена фактора элонгации EF1α, который является конститутивным промотором. Активность данного промотора в лимфоцитах человека значительно выше, чем активность некоторых других конститутивных промоторов, в частности промоторов генов UbC и Pgk. Это свойство промотора EF1α позволяет эффективно использовать его для направления экспрессии антиВИЧ генов, так как именно лимфоциты являются клетками, чувствительными к вирусу иммунодефицита человека и в этих клетках должен быть достигнут высокий уровень экспрессии антивирусных генов. Однако настоящее изобретение не ограничивается этим промотором, и специалисту в данной области известны другие промоторы, которые могут быть использованы без ограничения, например, промоторы генов PGK, EF1α, CD4, CD2, IL-2, промотор гена бета-глобина, промоторы генов МНС. Могут быть использованы гибридные промоторы, в составе которых дополнительно к природной последовательности введены последовательности энхансера или сайленсера, а также последовательности позволяющие регулировать активность промотора в зависимости от связывания с цитокинами/хемокинами или другими регулирующими молекулами.
Другой задачей настоящего изобретения является создание генетических конструкций, продуцирующих антивирусный агент на основе модифицированного гена ТММ5а с GC богатым нуклеотидным составом, обладающего повышенной экспрессией, в комбинации с одним или несколькими нижеперечисленными антивирусными агентами.
Комбинирование двух и более антивирусных генов в одной генетической конструкции представляет преимущество перед использованием одного агента, в частности, проявляется синергетический эффект. Разные противовирусные агенты предотвращают инфицирование клетки на разных этапах, блокируя проникновение вируса (CCR5 siRNA), предотвращая интеграцию вируса в геном (TRIM5α), замедляя репликацию вируса (TAR) и приводя к образованию неинфекционного вирусного потомства (env антисенс). Каждый агент в отдельности в различной степени подавляет репродукцию ВИЧ (от 50% до 90%). Одновременное использование нескольких противовирусных агентов замедляет образование мутантных штаммов, а комбинация противовирусных агентов с агентами, придающими резистентность клеткам к ВИЧ, позволяет получить близкую к 100% антивирусную активность препарата и предотвратить появление устойчивых мутантов.
Согласно настоящему изобретению предлагаются генетические конструкции, которые помимо вышеописанного агента на основе гена TRIM5a включают, по меньшей мере, один из перечисленных ниже антивирусных агентов.
Короткие интерферирующие РНК, направленные против гена рецептора человека CCR5 (CCR5si)
В качестве одного из агентов, придающих клетке устойчивость к ВИЧ, авторами были выбраны короткие интерферирующие РНК, направленные против гена хемокинового рецептора CCR5, задействованного в процессе проникновения ВИЧ в клетку.
РНК-интерференция - регуляторный механизм, присутствующий в большинстве эукариотических клеток который использует двухцепочечные РНК-молекулы как сигнал для регуляции активности генов, гомологичных этим РНК. В клетках млекопитающих эндогенный механизм РНК-интерференции запускается так называемыми молекулами микроРНК. Последовательности генов микроРНК образуют шпилечную структуру (первичная микроРНК), которая распознается и процессируется клеточным ферментом Drosha в ядре, после чего образовавшиеся короткие РНК-шпильки (предшественники микроРНК) экспортируются в цитоплазму. В цитоплазме специальный белковый комплекс (RISC) удаляет петлю шпильки, образуя короткие РНК-дуплексы. Эти дуплексы известные как короткие интерферирующие РНК - киРНК (short interfering RNAs, siRNAs) представляют собой двухцепочечные РНК размером 21-22 пар нуклеотидов; на 3' концах обеих цепей которых присутствуют два неспаренных нуклеотида. киРНК широко используются для ингибирования экспрессии генов при работе с клеточными культурами in vitro. Дальнейшее процессирование РНК-дуплекса приводит к выбору эффекторной короткой (~21 п.н.) цепи РНК, которая направляется к гомологичной мРНК и гибридизуется с комплементарным участком мРНК-мишени. В случае 100% гомологии связывание приводит к расщеплению мРНК и последующей полной деградации мРНК.
Запустить внутриклеточный механизм РНК-интерференции, направленный на подавление вирусных или клеточных генов, можно введя в клетку трансгены экспрессирующие короткие шпилечные РНК (short hairpin RNA, shRNA), которые образуют во вторичной структуре плотные шпильки и являются аналогами предшественников микроРНК. Также как и микроРНК предшественники они экспортируются в цитоплазму и далее процессируются, используя механизм РНК-интерференции. Если такая шпилечная РНК содержит последовательность, комплементарную последовательности гена CCR5, это приведет к деградации мРНК и отсутствию экспрессии рецептора. Короткие РНК, образующие шпильки, вводят в клетки при помощи вектора, использующего промотор РНК-полимеразы III для обеспечения конститутивной или индуцибельной экспрессии.
Последовательность, комплементарную последовательности гена CCR5, также можно встроить в тело эндогенной микроРНК; такая искусственно модифицированная первичная микроРНК будет процессирована и приведет к расщеплению мРНК соответствующего гена. Экспрессия микроРНК может регулироваться как промотором РНК-полимеразы II, так и промотором РНК-полимеразы III.
Интегрированный вектор, содержащий shRNA или микроРНК, передается дочерним клеткам и обеспечивает наследуемое выключения гена (Paddison P. et al., "Short hairpin RNAs (shRNAs) induce sequence-specific silencing in mammalian cells", Genes Dev., 2002, vol.16(8), pp.948-958).
РНК-интерференция является очень эффективным механизмом подавления экспрессии генов, с ее помощью можно достигнуть 99% ингибирования, то есть практически выключить работу гена.
Основная проблема, возникающая при использовании РНК-интерферирующих молекул направленных против генов ВИЧ, кроется в самой природе вируса, приводящей к возникновению устойчивых мутантных штаммов. Для успешного применения РНК-интерференции необходима строгая комплементарность siRNA и мРНК-мишени. Возникновение единственной замены нуклеотида в РНК ВИЧ, соответствующего 9-11 нуклеотиду siRNA, приводит к полной несостоятельности механизма РНК-интерференции и отсутствию подавления репликации. В клеточной культуре возникновение устойчивых вирусных мутантов, несущих замену единственного нуклеотида, как правило, возникает на 25 день. siRNA, направленные против высоко консервативных участков, лишь отчасти улучшают ситуацию, требуя более длительного времени для появления мутантов. Наиболее перспективный подход состоит в использовании siRNA, направленных против клеточных генов, белковые продукты которых взаимодействуют с ВИЧ, и которые отличаются высокой консервативностью.
В заявках WO 2004065549 ("Small interference RNA gene therapy"; 15.01.2004; МПК C12N 15/11, A61K 48/00) и WO 2003022052 ("Method for expression of small RNA molecules within a cell"; 13.09.2002; МПК C12N 15/11, C12N 15/867, A61K 48/00) раскрываются лентивирусные системы доставки siRNA в клетки для лечения рака.
В соответствие с настоящим изобретением предложены конструкции, содержащие последовательность, кодирующую siRNA, направленную против клеточного гена - гена хемокинового рецептора CCR5. CCR5-корецептор, необходимый для проникновения в клетку R5 тропных вирусов. Естественные мутации корецепторов широко встречаются в популяции как ВИЧ-инфицированных, так и неинфицированных людей. У некоторых людей такие мутации приводят к полному отсутствию рецепторной молекулы, что делает таких людей устойчивыми к инфицированию ВИЧ. Примечательно, что отсутствие CCR5 рецептора у таких людей не приводит к нарушениям фенотипа.
Последовательность, кодирующая данную siRNA, может быть клонирована в лентивирусный вектор в виде искусственной микроРНК (miCCR5), на основе микроРНК человека, транскрипция которой инициируется Pol II промотором.
Преимуществом данных siRNA (в виде микроРНК) является высокая эффективность подавления экспрессии гена CCR5 и в то же время отсутствие клеточной токсичности по сравнению с shRNA. Поскольку присутствие даже нескольких молекул на поверхности клетки делает ее чувствительной к заражению вирусом, то только полное подавление синтеза рецептора, за счет высокой эффективности siRNA, защищает клетку от проникновения вируса. Предложенные в соответствии с настоящим изобретением конструкции, включающие, в частности, miCCR5, обеспечивают превосходную антивирусную активность в сочетании с полным отсутствием токсичности.
РНК-шпилька, аналогичная TAR-последователъности
Еще одной группой антивирусных агентов являются РНК-аптамеры. Аптамеры-молекулы полинуклеотидов связывающие с высокой эффективностью белковые лиганды вируса. Аптамеры, синтезированные in vitro, демонстрируют высокую ингибирующую активность, однако при экспрессии в клетке эффективность заметно снижается. Проблемой является формирование правильной пространственной структуры в клеточном контексте. Исключение составляет использование двух последовательностей присутствующих в РНК ВИЧ - области TAR (trans-activating response region) и RRE (Rev response element), так называемые TAR и RRE ловушки, которые являются естественными аптамерами для tat и rev вирусных белков.
Tat - один из ранних вирусных белков, продуцируемый во время ВИЧ инфекции, играет ключевую роль в регуляции ВИЧ репликации. Связываясь с последовательностью TAR, Tat активизирует экспрессию вирусных генов более чем в 100 раз. TAR элемент локализован на 5 конце всех ВИЧ транскриптов, его структура представляет собой шпильку длиной 59 нуклеотидов. Исследования показали, что за связывание tat белка отвечает выступ из трех нуклеотидов, а петля на конце шпильки необходима для кооперативного взаимодействия с клеточным белком циклин Т1 (cyclinT1) Длинный двухцепочечный участок - ствол шпильки - играет лишь структурную роль.
Среди нескольких стратегий генной терапии, направленных на блокирование активности гена tat, одна из самых успешных - использование коротких РНК-молекул, работающих как ловушки. Такие РНК имитируют tat связывающую РНК-последовательность вируса, приводя к уменьшению свободного tat белка (Bohjanen P.R. et al., "A small circular TAR RNA decoy specifically inhibits Tat-activated HIV-1 transcription", Nucleic Acids Res., 1996, vol.24(19), pp.3733-3738).
Поскольку конструкция на основе последовательности (SEQ ID N4), кодирующей РНК-шпильку, аналогичную TAR-последовательности, замедляет репликацию ВИЧ, использование ее в комбинации с другими анти-ВИЧ агентами позволит усилить антивирусный эффект и избежать появления устойчивых мутантов.
Антисмысловая последовательность к участку гена env ВИЧ
Антисмысловые (антисенс) РНК - короткие или длинные РНК-последовательности, комплиментарные последовательности РНК ВИЧ, образуют нефункциональные дуплексы с вирусной РНК и способны блокировать репликацию вируса в клетках. Хотя точный механизм ингибирования не ясен, он может включать запуск деаминации цитозина ВИЧ:антисенс дуплексной РНК, приводя к задержке транскриптов в ядре и последующей их деградации или к образованию многочисленных нежизнеспособных вирусных мутантов. Другими возможными механизмами (которые зависят от структуры антисмысловой РНК) являются ингибирование экспрессии на уровне сплайсинга, ингибирование трансляции, упаковка химерного дуплекса с образованием дефектных вирусных частиц.
Множество антисенс-конструкций, исследованных in vitro, обладали способностью подавлять размножение вируса, в том числе антисмысловые РНК, блокирующие регуляторные цис элементы, такие как сигнал упаковки (Ψ), последовательность связывания tat белка (TAR) и сайт связывания праймера (PBS) (Cohli H., Fan В., Joshi R.L., et al. "Inhibition of HIV-1 multiplication in a human CD4+ lymphoid cell line expressing antisense and sense RNA molecules containing HIV-1 packaging signal and RREs", Antisense Res. Dev., 1994, vol.4(1), pp.19-26; Ding S.F., Noronha J., and Joshi S., "Co-packaging of sense and antisense RNAs: a novel strategy for blocking HIV-1 replication", Nucleic Acids Res., 1998, vol. 26, pp.3270-3278; Chadwick D.R., and Lever A.M., "Antisense RNA sequences targeting the 5 leader packaging signal region of human immunodeficiency virus type-1 inhibits viral replication at post-transcriptional stages of the life cycle". Gene Ther., 2000, vol.7, pp.1362-1368) или антисенс РНК, соответствующие кодирующим областям вирусных генов, таким как rev, env, vif и т.д. Известно, что антивирусный эффект в большой степени зависит от промотора и вектора использованного для доставки.
В статье Левайна (Levine B.L. et al., "Gene transfer in humans using a conditionally replicating lentiviral vector", Proc. Natl. Acad. Sci. USA, 2006, vol.103(46) pp.17372-17377) раскрываются конструкции на основе лентивирусных векторов, экспрессирующие антисмысловые гены против оболочки ВИЧ.
В предложенных в соответствии с настоящим изобретением конструкциях в качестве антисмысловой последовательности к участку гена env ВИЧ используют последовательность (SEQ ID N5), полученную из кДНК изолята вируса иммунодефицита человека субтипа А. Данный изолят был получен из клинического материала от Российского ВИЧ-инфицированного пациента. Поскольку в России в подавляющем большинстве ВИЧ-позитивные пациенты инфицированы ВИЧ субтипа А (более 90%), можно говорить о том, что противовирусный агент, полученный на основе данного изолята, будет более предпочтителен при лечении российских пациентов.
Специалист в данной области на основании информации, раскрытой в настоящем описании, легко может получить генетические конструкции, включающие любые комбинации перечисленных выше генетических антивирусных агентов.
Векторные конструкции в соответствии с настоящим изобретением могут быть созданы, например, на основе вектора pLVX-Puro (Clontech) (фиг.1), содержащего лентивирусные LTR и упаковочный сигнал (т). Также вектор содержит последовательность сРРТ размером 124 п.о., которая повышает эффективность транспорта вирусного генома в ядро. Это приводит к более эффективной интеграции генома, а, следовательно, к более эффективной трансдукции. Наличие RRE (Rev-responsive element) в векторе способствует увеличению титра за счет повышения экспорта из ядра несплайсированной вирусной РНК в присутствии белка rev. Наличие в векторе WPRE (посттранскрипционного регуляторного элемента вируса гепатита североамериканского лесного сурка) позволяет повысить эффективность событий, связанных с процессингом РНК и экспорта из ядра. WPRE обеспечивает двойной эффект. Во-первых, находясь в составе вирусного транскрипта, данный элемент повышает эффективность упаковки вектора, что способствует повышению титра вируса, продуцируемого упаковочными клетками 293Т. Во-вторых, он усиливает экспрессию гена (gene of interest) за счет облегчения образования зрелой матричной мРНК из транскрипта, инициированного с внутреннего промотора.
В соответствии с другим вариантом выполнения изобретения, генетические конструкции могут быть выполнены на основе самоинактивирующегося лентивирусного вектора, например, на основе вектора pLV-neo (фиг.2). Указанный вектор содержит те же основные элементы, что и вектор pLVX-Puro, за исключением последовательностей LTR. Вместо 5'LTR последовательности используется RSV/5'-LTR гибридный промотор, что позволяет не вводить дополнительно плазмиду кодирующую вирусный белок tat. 3'LTR содержит делецию в области U3, затрагивающую энхансерно-промоторную область. Так как, из-за особенностей обратной транскрипции, интегрирующий в геном провирус имеет U3 области LTR, идентичные длинному концевому повтору, расположенному на 3' конце, в данном случае он будет лишен промоторно-энхансерной области.
Неожиданно было дополнительно обнаружено, что модификация лентивирусного вектора, заключающаяся в клонировании последовательности SEQ ID N10 непосредственно перед областью сРРТ, позволяет еще более стабилизировать генную экспрессию, способствует повышению инфекционности вирусных частиц, улучшает эффективность введения генетического материала в клетки-мишени.
При необходимости, в вектор вместо гена для селекции пуромицина в качестве молекулярного маркера может быть введена другая последовательность для селекции и/или мониторинга модифицированных клеток. Маркерный ген представляет собой последовательность, которая присоединена к элементам гетерологичного промотора или энхансера, кодирующую продукт, который может быть легко и количественно проанализирован в том случае, когда эта конструкция введена в ткани или в клетки. Специалистам в данной области известны различные последовательности для мониторинга и/или селекции, например ген зеленого флюоресцентного белка (EGFP). Маркерный ген/последовательность может быть удален при использовании вектора в терапии ВИЧ.
Краткое описание чертежей
На фиг.1 представлена карта плазмиды pLVX-puro, характеризующейся:
5'LTR: 1-635
PBS: 636-653
Ψ: 685-822
RRE: 1303-1536
cPPT: 2028-2151
PCMV IE: 2185-2788
MCS: 2816-2880
PPGK: 2882-3390
Puror: 3411-4010
WPRE: 4024-4615
3'LTR: 4819-5455
pUC: 5925-6595
Ampr: 6740-7736
На фиг.2 представлена карта плазмиды pLV-neo, характеризующейся:
RSV/5'-LTR гибридный промотор: 1-416
PBS: 418-435
Ψ: 467-604
RRE: 1081-1314
cPPT: 1810-1928
PCMV IE: 1940-2525
Гистоновый промотор: 2576-2933
Ген устойчивости к неомицину: 2942-3736
WPRE: 3750-4291
3'LTR, с делецией в области U3: 4428-4662
SV40 polyA: 4734-4864
Ampr: 5828-6688
На фиг.3 приведены последовательности, кодирующие модифицированный белок TRIM5α человека, с GC богатым нуклеотидным составом (trim enh, SEQ ID N1) и исходная последовательность, описанная в патенте RU 2426788 (trim, SEQ ID N2).
На фиг.4 представлено сравнение антиВИЧ активностей конструкции, включающей последовательность, кодирующую модифицированный белок TRIM5α человека, с GC богатым нуклеотидным составом (trim enhanced) и конструкции, включающей исходную последовательность (trim).
Далее приводятся примеры осуществления изобретения со ссылками на прилагаемые фигуры.
Примеры осуществления изобретения и реализации назначения
Пример 1. Получение векторов для генетических конструкций
1а. Лентивирусный вектор
Вектор pLVX-puro доступен от Clontech, длиной 8102 пар нуклеотидов (фиг.1) модифицировали с помощью последовательности SEQ ID N10. Используя праймеры сРРТ 1g Hpa for (TTTGGTTAACCCAGTAAAAACAGTACATACAG) и сРРТ 1g Cla Barn rev (TTTTGGATCCATCGATAAATTTTGAATTTTTGTAATTTG), плазмиду pLP1 (Invitrogen) в качестве матрицы, наработали ПЦР-продукт длиной 370 п.о., который клонировали в вектор pGEM-T vector (Promega). После трансформации и последующей селекции данный фрагмент переклонировали в плазмиду pLV-neo, предварительно гидролизованную по сайтам HpaI и BamHI. Полученную плазмиду нарабатывали в препаративных количествах и резали с использованием эндонуклеаз MfeI и ClaI, a полученный фрагмент длиной 1532 п.о. лигировали в вектор pLVX-Puro, предварительно гидролизованный по этим же сайтам. Полученный после лигирования вектор pLVX-Puro сРРТ 1g нарабатывали в препаративных количествах и выделяли из Е. coli с помощью набора Axygen. Для последующего удобства клонирования, в вектор pLVX-Puro сРРТ 1g, вместо CMV промотора, по сайтам ClaI и EcoRI, был введен линкер
Link For (TTGATATCGATCTGAGTGAGACTCTGACGAATTCTTCT)
Link Rev (AGAAGAATTCGTCAGAGTCTCACTCAGATCGATATCAA).
Полученный вектор pLVX Puro сРРТ lg ACMV использовали для последующих процедур клонирования перечисленных последовательностей.
1б. Самоинактивирующийся лснтивирусный вектор
Модификацию вектора pLV-neo длиной 7907 пар нуклеотидов (фиг.2) с помощью последовательности SEQ ID N10 проводили аналогично предыдущему. С помощью праймеров сРРТ 1g Hpa for (TTTGGTTAACCCAGTAAAAACAGTACATACAG) и сРРТ 1g Cla Barn rev (TTTTGGATCCATCGATAAATTTTGAATTnTGTAATTTG), используя в качестве матрицы плазмиду pLP1 (Invitrogen), наработали ПЦР-продукт длиной 370 п.о., который клонировали в вектор pGEM-T vector (Promega). После трансформации и последующей селекции данный фрагмент переклонировали в плазмиду pLV-neo, предварительно гидролизованную по сайтам HpaI и BamHI. Полученная плазмида pLV-neo сРРТ 1g содержит последовательность SEQ ID N10 перед областью сРРТ. В плазмиду pLV-neo сРРТ 1g вместо CMV промотора ввели линкер также как в примере 1а и получили pLV-neo сРРТ 1g ACMV.
Пример 2. Модификация лентивирусного вектора - получение конструкции без гена PURO с геном-маркером EGFP
ПЦР фрагмент, содержащий ген EGFP и внутренний сайт связывания рибосомы IRES, получен при амплификации плазмиды pIRES-EGFP (Clontech #6064-1) с праймерами RI-ires for GACGGAATTCAGTGGATCCACTAGTAACGGC Sal-egfp rev TTTGTCGACCTTTACTTGTACAGCTCGTCCATGCC. Данный фрагмент встроен в pLVneo вектор по сайтам EcoRI-SalI. Кассета, содержащая сайт IRES и ген EGFP, переклонирована в вектор pLVX-Puro сРРТ 1g вместо гена устойчивости к пуромицину по сайтам рестрикции EcoRI-KpnI. Данный вектор pLVX Egfp cPPT1g вместо селективного гена устойчивости к пуромицину содержит ген зеленого флуоресцентного белка в качестве маркера, позволяющего сортировать клетки по наличию флуоресценции.
Пример 3. Получение конструкции ТММ5а с GC богатым нуклеотидным составом.
В последовательности модифицированного гена Trim5a (SEQ ID N2) размером 1488 п.н. (496 кодонов) было спроектировано 200 синонимичных замен нуклеотидов по всей длине таким образом, что GC-состав последовательности увеличился с 48% до 61%. Сравнение исходной и GC-богатой последовательностей (SEQ ID N1) гена TRIM5α приведено на фиг.3. Для конструирования GC-богатой последовательности была проведена сборка гена de novo с помощью лигазной цепной реакции с последующей ПЦР в соответствие со стандартным протоколом (Au L.C., Yang F.Y., Yang W.J., Lo S.H., Kao C.F., "Gene synthesis by a LCR-based approach: high-level production of leptin-L54 using synthetic gene in Escherichia coli", Biochem. Biophys. Res. Соттип., 1998, vol.248(1), pp.200-203). Для сборки использовали Tth лигазу фирмы «Fermentas». Дизайн 5'-фосфорелированных олигонуклеотидов был проведен при помощи программы Gene201igo (Rouillard J.M., Lee W., Truan G., Gao X., Zhou X., Gulari E., "Gene201igo: oligonucleotide design for in vitro gene synthesis". Nucleic Acids Res., 2004, vol.32 (Web Server issue), W176-80). Полученный ампликон, содержащий новую модифицированную последовательность Trim5a, клонировали в вектор pGEM (Promega). Правильность сборки гена в отдельных клонах была проверена с помощью секвенирования. Созданная плазмида была обозначена pGEM-TRIM enh.
Пример 4. Клонирование GC богатой последовательности модифицированного гена TRIM5α в лентивирусный вектор.
Продукт ПЦР амплификации плазмиды pGEM-TRIM enh с использованием праймеров ACTTCTAGATTACGAGCTCGGCGAGCACAG и TTTTCTCGAGACTAGTGATGCCGCCACCATG резали по сайтам рестриктаз XhoI и XbaI и лигировали в вектор pLVX-Puro cPPT 1g, порезанный по тем же сайтам. Полученная плазмида обозначена pLVX-Puro cPPT 1g CMV-Trim enh.
Пример 5. Клонирование GC богатой последовательности модифицированного гена TRIM5α в самоинактивирующийся лентивирусный вектор.
Продукт ПЦР амплификации плазмиды pGEM-TRIM enh с использованием праймеров ACTTCTAGATTACGAGCTCGGCGAGCACAG и TTTTCTCGAGACTAGTGATGCCGCCACCATG резали по сайтам рестриктаз XhoI и XbaI и лигировали в вектор pLV-neo cPPT 1g, порезанный по тем же сайтам. Полученная плазмида обозначена SIV CMV-Trim enh.
Пример 6. Клонирование последовательности, кодирующей микроРНК, направленную против CCR5 гена в вектор.
Нуклеотидную последовательность микроРНК мыши miR155 SEQ ID N6 использовали для создания искусственных микроРНК: SEQ ID N7, SEQ ID N8 или SEQ ID N9, направленных против гена рецептора CCR5 (miCCR5).
Для получения микроРНК SEQ ID N7 были использованы праймеры O1-1 TGCTGTAAGAGGTAGTTTCTGAACGTTTTG и O3-1 GTGGCCAAAACGTTCAGAAACTACCTCTTAC, которые отжигали друг на друга с получением короткого двухцепочечного фрагмента ДНК olig1-3 с липкими концами. Аналогично отжигали праймеры O2-1 GCCACTGACTGACGTTCAGAATACCTCTTA и O4-1 CCTGTAAGAGGTATTCTGAACGTCAGTCA с получением двухцепочечного олига olig2-4. Смесь, состоящую из плазмиды pcDNA6.2-GW/miR (набор Block-iT PolII miR RNA expression Vector Kit, Invitrogen) и двух олигов olig1-3 и olig2-4, лигировали в течение 1 часа при комнатной температуре в соответствии с инструкцией из набора. Лигазную смесь трансформировали в клетки E. Coli, выросшие клоны отбирали по сиквенсу. Трансформанты, содержащие искомую плазмиду, наращивали и выделяли ДНК плазмиды miR-CCR5-1. Фрагмент, содержащий микроРНК (SEQ ID N7), вырезали из плазмиды по сайтам рестрикции SalI-BgIII. Вектор pLVX Puro cPPTIg рестрицировали ферментами XhoI-BamHI и лигировали с фрагментом, содержащим микроРНК. Полученную конструкцию Puro/miR-CCR5-1 проверяли рестрикционным анализом и секвенировали. Для трансфекции нарабатывали в препаративных количествах в E.coli и выделяли с использованием QIAGEN plasmid maxi kit.
Аналогичным образом получали конструкции, содержащие miRNA SEQ ID N8 или SEQ ID N9. Для получения микроРНК SEQ ID N8 использовали следующие праймеры:
TgCTgCATAgATTggACTTgACACgTTTTg
CCTgCATAgATTgCTTgACACgTCAgTCA
gCCACTgACTgACgTgTCAAgCAATCTATg
gTggCCAAAACgTgTCAAgTCCAATCTATgC
Для получения микроРНК SEQ ID N9 использовали следующие праймеры:
TgCTgAATTgATgTCATAgATTggACTTgACACgTTTTg
CCTgAATTgATgTCATAgATTgCTTgACACgTCAgTCA
gCCACTgACTgACgTgTCAAgCAATCTATgACATCAATT
gTggCCAAAACgTgTCAAgTCCAATCTATgACATCAATTC
Пример 7. Получение конструкции, содержащей одновременно GC-богатый модифицированный ген TRIM5α и микроРНК против CCR5.
Вектор pLVX Egfp cPPT1g рестрицировали по сайтам XbaI и ClaI. Фрагмент, содержащий микроРНК, вырезали из плазмиды Puro/miR-CCR5-l по сайтам ClaI и XbaI, а фрагмент, содержащий GC-богатый модифицированный ген TRIM 5a, вырезали из плазмиды pLVX-Puro cPPT 1g CMV-Trim enh по сайтам NheI и XbaI. Для получения комбинированной конструкции вектор лигировали с двумя фрагментами и трансформировали в клетки E. Coli. Отбирали клоны, содержащие оба фрагмента, и анализировали выделенные плазмиды методом секвенирования. Полученная конструкция была обозначена pLVX-Puro miR-CCR5-1-Trim enh.
Аналогично получали конструкции, содержащие Trim enh и miRNA SEQ ID 8 или SEQ ID 9.
Пример 8. Трансфекция упаковочных клеток и получение лентивирусных частиц.
Для получения лентивирусов использовали клеточную линию НЕК 293Т и набор упаковочных плазмид Lenti-X HT Packaging Mix (Clontech). Среда для роста НЕК 293Т представляет собой: Dulbecco's Modified Eagle's Medium (DMEM), содержащая 4 ммоль L-глутамина, 4,5 г/л глюкозы, 1 ммоль натрия пирувата, 1,5 г/л натрия бикарбоната и 10% эмбриональной бычьей сыворотки. Такой уровень натрия бикарбоната (NaHCO3; 1,5 г/л) используется в инкубаторах, содержащих 5% CO2.
За день до трансфекции 4-5*10 клеток вносили на чашку 100 мм в 10 мл полной среды, содержащей сыворотку, свободную от тетрациклина (Tet System Approved FBS Clontech Cat# 631106). В день трансфекции в 15 мл пробирке смешивали 15 мкл смеси Lenti-X HT Packaging Mix и векторную ДНК 3 мкг. Затем в пробирку добавляли 1 мл среды OptiMEM (Invitrogen) и 20 мкл реагента для трансфекции Turbofect (Fermentas). Смесь инкубировали 20 минут при комнатной температуре. Во время инкубации клеточную среду заменяли на 10 мл среды OptiMEM, содержащей 10% сыворотки, свободной от тетрациклина. После окончания инкубации смесь плазмид, OptiMEM и Turbofect по каплям добавляли в клеточную среду. Клетки инкубировали в течение ночи при 37°С в СО2 инкубаторе. На следующий день удаляли среду, содержащую комплекс ДНК-Turbofect, и заменяли на 10 мл культуральной среды, содержащей сыворотку, свободную от тетрациклина. Клетки инкубировали при 37°С в СО2 инкубаторе 48-72 часа. Отбирали супернатант, содержащий лентивирусные частицы. Супернатант центрифугировали при 3000 об/мин в течение 15 минут при +4°С. Фильтровали супернатант через 0,45 мкм. Готовый препарат хранили при -80°С.
Для получения самоинактивирующихся лентивирусных частиц использовали конструкции на основе самоинактивирующегося вектора и набор упаковочных плазмид ViraPower Packaging Mix (Invitrogen). Трансфекция клеток проводилась аналогично, сыворотку, свободную от тетрациклина, заменяли на сыворотку Invitrogen.
Пример 9. Определение титра вируса
За день до трансдукции (1 день) фибробласты линии НТ1080 (приблизительно 2×105 клеток на лунку) помещали в плашку и инкубировали в течение ночи при 37°С в CO2 инкубаторе. На следующий день (2 день) размораживали лентивирусный сток и выполняли последовательные 10-кратные разведения от 10-2 до 10-6. Для каждого разведения доводили объем до 1 мл культуральной средой. Удаляли из клеток среду и добавляли каждое разведение вируса в свою лунку (в объеме 1 мл). Инкубировали в течение ночи при 37°С в CO2 инкубаторе.
На следующий день (3 день) удаляли среду, содержащую вирус, и заменяли на 2 мл культуральной среды. Инкубировали в течение ночи при 37°С в СО2 инкубаторе.
4 день: обрабатывали клетки антибиотиком пуромицином. Удаляли среду, заменяли на среду, содержащую 2 мкг/мл пуромицина. Заменяли среду, содержащую антибиотик, каждые 3-4 дня.
Через 7-10 дней селекции удаляли среду и отмывали клетки PBS 2 раза. Добавляли раствор crystal violet и инкубировали 10 минут при комнатной температуре. Отмывали клетки PBS 2 раза. Подсчитывали синие колонии и определяли титр вирусного стока.
Пример 10. Трансдукция МТ-4 клеток
Наращивали клетки в культуральной среде RPMI+10% FBS до концентрации 0,5×106 клеток на мл. В первый день размораживали лентивирусный сток и развовдили в свежей культуральной среде до необходимого MOI. К 3 мл суспензии клеток с концентрацией 0,5×106 кл/мл добавляли 1,5×106 вирусных частиц в 2 мл культуральной среды+5 мкл полибрен 4 мг/мл.
Инкубировали в течение ночи при 37°С в CO2 инкубаторе.
На следующий день (день 2) центрифугировали клетки, удаляли среду, содержащую вирус, и заменяли ее на свежую культуральную среду. Инкубировали в течение ночи при 37°С в CO2 инкубаторе.
На 3 день в среду добавляли антибиотик, для селекции трансдуцированных клеток. Меняли среду на свежую с антибиотиком каждые 3-4 дня.
Клетки, устойчивые к пуромицину, наращивали до титра 0,9×106. Далее клетки были использованы для тестирования на устойчивость к заражению вирусом ВИЧ.
Пример 11. Оценка эффективности конструкции на основе модифицированного гена TRIM5a на клетках МТ-4.
Методика.
Суспензию модифицированных клеток МТ-4 делили на два культуральных сосуда и разводили до концентрации 5*105 клеток на миллилитр. Одну часть клеток инфицировали вирусом иммунодефицита человека (pNL4-3) для определения противовирусной активности, другая служила контролем цитотоксичности препарата. Клетки культивировали параллельно.
Клетки инфицировали вирусом с множественностью заражения 0,01 инфекционных единиц на клетку. Инкубировали культуры клеток при 37°С в течение 1 час и дважды отмывали. После этого к культуре клеток добавляли питательную среду RPMI-1640 (конечная концентрация клеток 400000 клеток/мл). Далее смена среды производилась на 2, 4, 8, 12, 15 и 19 день культивирования. При смене среды старая среда полностью удалялась, клетки отмывались свежей средой RPMI-1640 без сыворотки, после чего добавлялась свежая питательная среда (конечная концентрация клеток 400000 клеток/мл). Учет результатов противовирусной активности производили по жизнеспособности клеток, путем окрашивания клеток красителем трипановым синим. Методом иммуноферментного анализа, с использованием коммерческого иммуноферментного набора фирмы «Вектор-Бест» (Россия) и разведении стандартного препарата р24 антигена (Aalto, Ирландия), проводили определение количества р24 антигена (нг/мл) в культуральной жидкости, согласно инструкции изготовителя. Результаты учитывали с помощью фотометра "Multiscan" при длине волны 630 нм.
Результаты.
На фиг.4 показана антиВИЧ активность конструкции, включающей последовательность, кодирующую модифицированный белок TRIM5α человека с GC богатым нуклеотидным составом (trim enh), по сравнению с конструкцией, включающей последовательность, кодирующую модифицированный белок TRIM5α человека, описанной в патенте RU 2426788.
Активность конструкции, включающей последовательность, кодирующую модифицированный белок TRIM5α человека с GC богатым нуклеотидным составом, превышает почти в 40 раз (на 7-ой день культивирования) активность конструкции с исходным нуклеотидным составом. Кроме того, увеличилась жизнеспособность клеток в присутствие ВИЧ.
Пример 12. Трансдукция клеток U373-MAGI-CCR5
В первый день клетки U373-MAGI-CCR5 рассевали по 1*105 клеток в лунки 48-луночного планшета в 0,5 мл среды DMEM с 10% фетальной телячьей сывороткой и 8 мкг/мл полибрена. На второй день полностью удаляли среду и добавляли лентивирусные частицы, несущие противовирусные ген/гены или контрольную вставку, в количестве 1 инфекционная частица на каждую клетку в 100 мкл полной среды DMEM с полибреном. Через 2 часа добавляли еще 300 мкл DMEM и оставляли на ночь. На третий день проводили смену среды. На пятый день проводили оценку количества трансдуцированных клеток с помощью проточной цитометрии, измеряя процент клеток, содержащих зеленый флуоресцентный белок. Если процент трансдуцированных клеток был не менее 50, полученные линии были использованы для последующей оценки противовирусной активности.
Пример 13. Оценка эффективности комбинированных конструкций на клетках U373-MAGI-CCR5.
Методика.
По 1*106 клеток каждой линии засевали в 2 флакона (для заражения и контрольный) площадью 25 см2 в 10 мл полной ростовой среды DMEM и 4 мкг/мл полибрена. На следующий день проводили заражение клеток вирусом ВИЧ pNL(AD)8, обладающего тропностью к рецептору CCR5. Для этого удалили ростовую среду из флаконов и внесли по 1 мл вирусной суспензии, содержащей 1*104 инфекционных единиц в 1 из флаконов каждой линии. В контрольные флаконы внесли по 1 мл ростовой среды. Инкубировали 2 часа при 37°С, покачивая каждые 30 минут. Через 2 часа клетки отмыли один раз PBS и второй раз поддерживающей средой. Затем в каждый флакон внесли по 7 мл полной ростовой среды. Начиная со следующего дня, в течение 9 дней проводили ежедневный отбор проб. Для этого из каждого флакона отбирали 4 пробы по 500 мкл и взамен вносили по 2 мл свежей ростовой среды. Полученные образцы использовали для измерения вирусной нагрузки ВИЧ и уровня вирусного антигена р24.
Результаты.
В таблице 3 представлены результаты измерения уровня р24 антигена ВИЧ в течение 9 дней после инфицирования клеток U373-MAGI-CCR5, трансдуцированных с помощью комбинированных конструкций. Во всех случаях наблюдалось замедление развития инфекции по сравнению с контролем (вектор без вставки). Использование тройной комбинации оказывало более сильное противовирусное воздействие, чем двойной.
Таким образом, настоящими экспериментальными данными продемонстрировано, что антиВИЧ активность конструкции, включающей последовательность, кодирующую модифицированный белок TRIM5α человека с GC богатым нуклеотидным составом, почти в 40 раз выше по сравнению с активностью аналогичной конструкции, включающей последовательность, кодирующую модифицированный белок TRIM5α человека, описанной в патенте RU 2426788 (без синонимичных замен). Добавление к данной конструкции дополнительно одного или двух антиВИЧ агентов повышали противовирусный эффект.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ АНТИВИЧ-ТЕРАПИИ | 2010 |
|
RU2426788C1 |
| Генетические конструкции и их смеси для антиВИЧ терапии | 2017 |
|
RU2666991C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ УРОВНЯ ЭКСПРЕССИИ ХИМЕРНОГО ГЕНА Trim5a | 2015 |
|
RU2592675C1 |
| ДВОЙНОЙ ВЕКТОР ДЛЯ ПОДАВЛЕНИЯ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА | 2010 |
|
RU2562868C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ МУЛЬТИПОТЕНТНЫХ КЛЕТОЧНЫХ ПОПУЛЯЦИЙ | 2008 |
|
RU2535365C2 |
| ВСПОМОГАТЕЛЬНЫЙ ПЛАЗМИДНЫЙ ЛЕНТИВИРУСНЫЙ ЭКСПРЕССИОННЫЙ ВЕКТОР ДЛЯ ПОЛУЧЕНИЯ ВЫСОКИХ ТИТРОВ VPX-СОДЕРЖАЩИХ ЛЕНТИВИРУСНЫХ ЧАСТИЦ, ОБЕСПЕЧИВАЮЩИЙ ЭФФЕКТИВНОЕ ЗАРАЖЕНИЕ МОНОЦИТОВ И ДЕНДРИТНЫХ КЛЕТОК ЧЕЛОВЕКА | 2017 |
|
RU2697781C2 |
| РЕКОМБИНАНТНЫЙ ЛЕНТИВИРУСНЫЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН, ТРАНСДУЦИРОВАННАЯ ЛЕНТИВИРУСНЫМ ВЕКТОРОМ, СПОСОБ ЕЕ ТРАНСДУКЦИИ И ПРИМЕНЕНИЕ | 2002 |
|
RU2305708C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ ГЕМОГЛОБИНОПАТИЙ | 2015 |
|
RU2707540C2 |
| ВИРУСНЫЕ ВЕКТОРЫ С ЗАВИСИМОЙ ОТ УСЛОВИЙ РЕПЛИКАЦИЕЙ И ИХ ПРИМЕНЕНИЕ | 2001 |
|
RU2301260C2 |
| РЕПОРТЕРНАЯ СИСТЕМА НА ОСНОВЕ ЛЕНТИВИРУСНЫХ РЕПОРТЕРНЫХ КОНСТРУКЦИЙ ДЛЯ ИЗУЧЕНИЯ БЕЛОК-БЕЛКОВЫХ ВЗАИМОДЕЙСТВИЙ | 2015 |
|
RU2639539C2 |







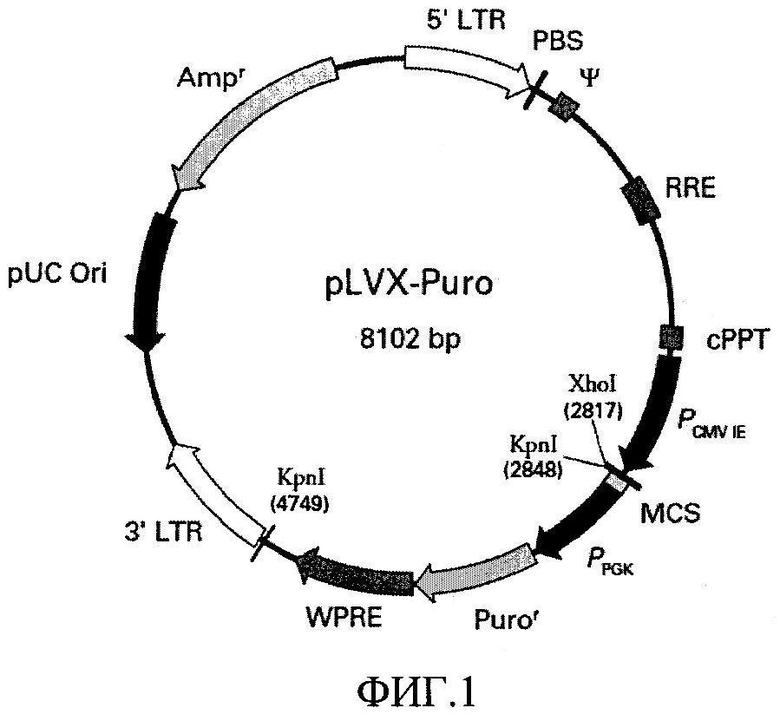
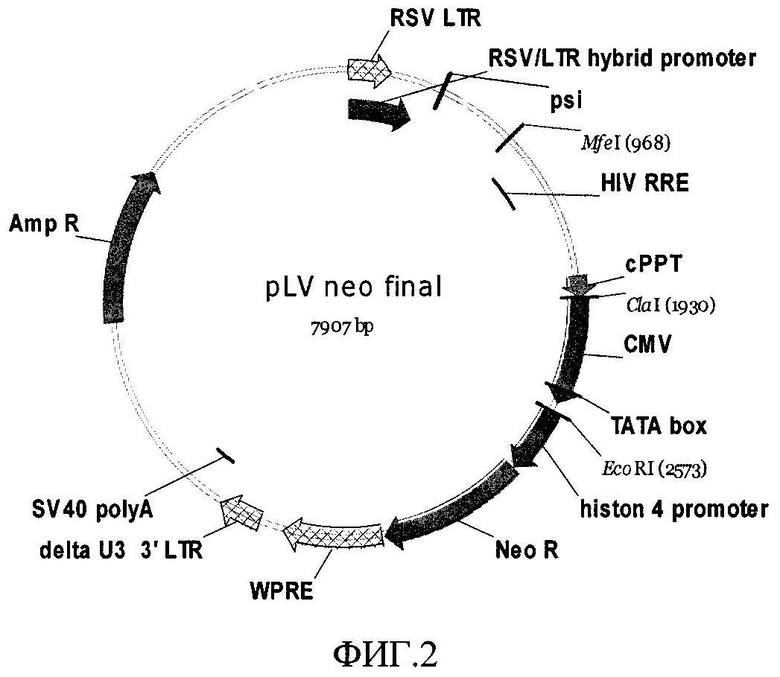
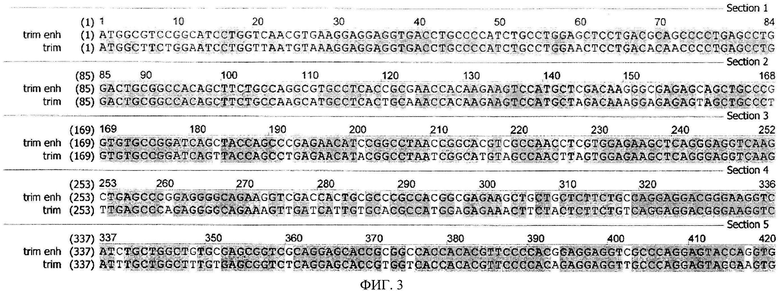
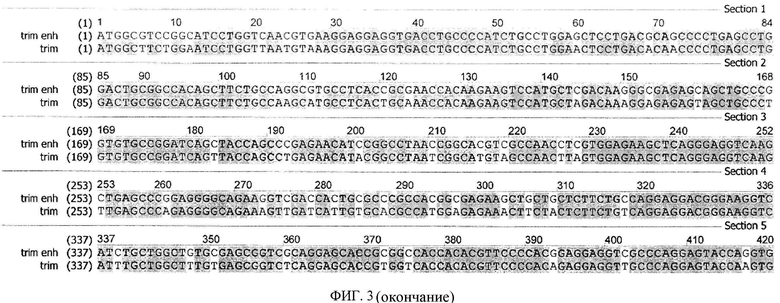
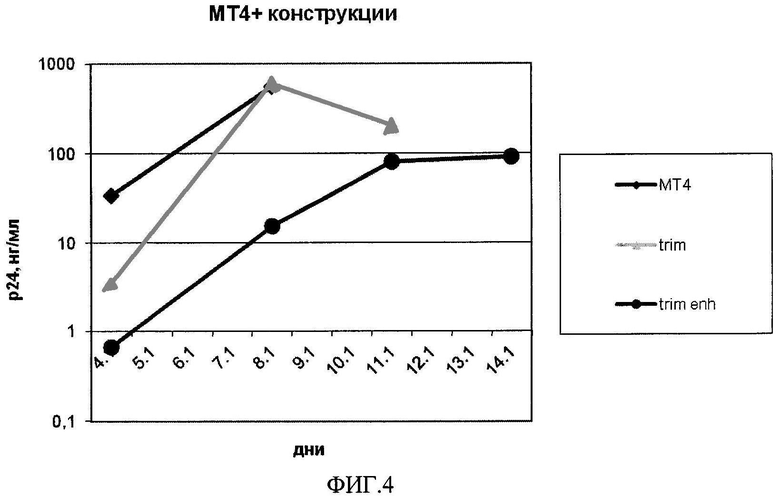
Изобретение относится к медицинской и молекулярной генетике. Описаны генетические конструкции, экспрессирующие РНК-последовательности и гены, кодирующие белки, обладающие антивирусной активностью в отношении вируса иммунодефицита человека. Генетические конструкции содержат последовательность, кодирующую модифицированный белок TRIM5a человека. Предложенные конструкции позволяют повысить уровень экспрессии модифицированного гена TRIM5a человека. Изобретение может быть использовано в научных исследованиях. 4 н. и 36 з.п. ф-лы, 4 ил., 3 табл., 13 пр.
1. Генетическая конструкция для антиВИЧ терапии, которая включает нуклеотидную последовательность (SEQ ID N1), кодирующую модифицированный белок TRIM 5α человека (SEQ ID N3) и отличающуюся от исходной последовательности SEQ ID N2 множественными синонимичными заменами, обеспечивающими увеличение процента GC пар в составе ДНК последовательности.
2. Генетическая конструкция по п.1, где процент GC пар в последовательности, кодирующей модифицированный белок TRIM 5α человека (SEQ ID N3), составляет от 49 до 64%.
3. Генетическая конструкция по п.1, где в качестве последовательности, кодирующей модифицированный белок TRIM 5α человека (SEQ ID N3), использована последовательность SEQ ID N1.
4. Конструкция по пп.1, 2 или 3, дополнительно включающая молекулярный маркер, представляющий собой ген устойчивости к пуромицину, ген зеленого флуоресцентного белка или другую последовательность для селекции и/или мониторинга модифицированных клеток.
5. Генетическая конструкция по пп.1, 2 или 3, где последовательность, кодирующая модифицированный белок TRIM5a человека, находится под контролем конститутивного, тканеспецифичного или индуцибельного промотора.
6. Генетическая конструкция по пп.1, 2 или 3, где в качестве промотора используется один из следующих промоторов: промотор гена PGK, гена EF1α, гена UbC, промотор генов MHC класса II, промотор гена Т-клеточного рецептора, промотор гена β-интерферона, промотор гена IL2, промотор гена рецептора IL2 или тетрациклин зависимый промотор.
7. Генетическая конструкция для антиВИЧ терапии, которая включает:
векторную последовательность, обеспечивающую встраивание ДНК последовательностей в геном клетки человека, и
последовательность (SEQ ID N1), кодирующую модифицированный белок TRIM 5α человека (SEQ ID N3) и отличающуюся от исходной последовательности SEQ ID N2 множественными синонимичными заменами, обеспечивающими увеличение процента GC пар в составе ДНК последовательности.
8. Генетическая конструкция по п.7, где процент GC пар в последовательности, кодирующей модифицированный белок TRIM 5α человека (SEQ ID N3), составляет от 49 до 64%.
9. Генетическая конструкция по п.7, где в качестве последовательности, кодирующей модифицированный белок TRIM 5α человека (SEQ ID N3), использована последовательность SEQ ID N1.
10. Генетическая конструкция по пп.7, 8 или 9, где последовательность, кодирующая модифицированный белок TRIM5a человека, находится под контролем конститутивного, тканеспецифичного или индуцибельного промотора.
11. Генетическая конструкция по пп.7, 8 или 9 где в качестве промотора используется один из следующих промоторов: промотор гена PGK, гена EF1α, гена, UbC, промотор генов МНС класса II, промотор гена Т-клеточного рецептора, промотор гена β-интерферона, промотор гена IL2, промотор гена рецептора IL2 или тетрациклин зависимый промотор.
12. Генетическая конструкция по пп.7, 8 или 9, где последовательность, кодирующая модифицированный белок TRIM5a человека, может быть в прямом или обратном направлении по отношению к вектору.
13. Генетическая конструкция по п.п.7, 8 или 9, дополнительно включающая молекулярный маркер, представляющий собой ген устойчивости к пуромицину, ген зеленого флуоресцентного белка или другую последовательность для селекции и/или мониторинга модифицированных клеток.
14. Генетическая конструкция по пп.7, 8 или 9, где в качестве вектора используется лентивирусный вектор на основе ВИЧ-1, который включает по меньшей мере следующие области:
5′-LTR и 3′LTR,
участок связывания специфической тРНК (PBS),
упаковочный сигнал (ψ),
rev-отвечающий элемент (RRE),
посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE),
область сРРТ.
15. Генетическая конструкция по пп.7, 8 или 9, где в качестве вектора используется лентивирусный вектор на основе ВИЧ-1, который включает, по меньшей мере, следующие области:
5′-LTR и 3′LTR,
участок связывания специфической тРНК (PBS),
упаковочный сигнал (ψ),
rev-отвечающий элемент (RRE),
посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE),
область сРРТ,
и где последовательность вектора модифицирована с помощью последовательности SEQ ID N10 перед областью сРРТ.
16. Генетическая конструкция по пп.7, 8 или 9, где в качестве вектора используется самоинактивирующийся лентивирусный вектор, который включает, по меньшей мере, следующие области:
RSV/5′-LTR гибридный промотор,
3′LTR, с делецией в области U3,
участок связывания специфической тРНК (PBS),
упаковочный сигнал (ψ),
rev-отвечающий элемент (RRE),
посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE),
область сРРТ.
17. Генетическая конструкция по пп.7, 8 или 9, где в качестве вектора используется самоинактивирующийся лентивирусный вектор, который включает, по меньшей мере, следующие области:
RSV/5′-LTR гибридный промотор,
3′LTR, с делецией в области U3,
участок связывания специфической тРНК (PBS),
упаковочный сигнал (ψ),
rev-отвечающий элемент (RRE),
посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE),
область сРРТ
и где последовательность вектора модифицирована с помощью последовательности SEQ ID N10 перед областью сРРТ.
18. Генетическая конструкция по пп.7, 8 или 9, где в качестве вектора используется лентивирусный вектор на основе одного или нескольких из лентивирусов: ВИЧ-1, ВИЧ-2, ВИО, вирус иммунодефицита крупного рогатого скота, лентивирус артрита-энцефалита коз, вирус инфекционной анемии лошадей, вирус иммунодефицита кошек, и/или вирус болезни Джембрана, или вектор, созданный на основе спумавируса.
19. Генетическая конструкция по пп.7, 8 или 9, где в качестве вектора используется невирусный вектор, созданный на основе векторной системы транспозон/транспозаза.
20. Генетическая конструкция по пп.7, 8 или 9, где встраивание ДНК последовательности в определенное место хромосомы человека обеспечивается за счет внесения разрыва в заданный участок хромосомы с помощью специфической эндонуклеазы, и последующей гомологичной рекомбинации между векторными последовательностями, входящими в конструкции по пп.7, 8 или 9 и прилегающими к разрыву участками хромосомы.
21. Генетическая конструкция для антиВИЧ терапии, которая включает:
последовательность (SEQ ID N1), кодирующую модифицированный белок TRIM 5α человека (SEQ ID N3) и отличающуюся от исходной последовательности SEQ ID N2 множественными синонимичными заменами, обеспечивающими увеличение процента GC пар в составе ДНК последовательности,
и одну или более последовательностей, выбранных из SEQ ID N 7, 8, 9, кодирующих искусственную микроРНК, которая ингибирует экспрессию гена рецептора человека CCR5 (miRNA CCR5).
22. Генетическая конструкция по п.21, где процент GC пар в последовательности, кодирующей модифицированный белок TRIM 5α человека (SEQ ID N3), составляет от 49 до 64%.
23. Генетическая конструкция по п.21, где в качестве последовательности, кодирующей модифицированный белок TRIM 5α человека (SEQ ID N3), использована последовательность SEQ ID N1.
24. Генетическая конструкция по пп.21-23, дополнительно включающая последовательность SEQ ID N4, кодирующую РНК шпильку, аналогичную TAR-последовательности (TAR), и/или антисмысловую последовательность к участку гена env ВИЧ SEQ ID N5 (а/s env).
25. Генетическая конструкция по пп.21-23, где по меньшей мере одна из последовательностей: микроРНК CCR5 и/или последовательность, кодирующая модифицированный белок TRIM 5α человека, находится под контролем конститутивного, тканеспецифичного или индуцибельного промотора.
26. Генетическая конструкция по пп.21, 22 или 23 где в качестве промотора используется один из следующих промоторов: промотор гена PGK, гена EF1α, гена UbC, промотор генов МНС класса II, промотор гена Т-клеточного рецептора, промотор гена β-интерферона, промотор гена IL2, промотор гена рецептора IL2, тетрациклин зависимый промотор или промотор РНК полимеразы III.
27. Конструкция по пп.21-23, дополнительно включающая молекулярный маркер, представляющий собой ген устойчивости к пуромицину, ген зеленого флуоресцентного белка или другую последовательность для селекции и/или мониторинга модифицированных клеток.
28. Генетическая конструкция для антиВИЧ терапии, которая включает:
векторную последовательность, обеспечивающую встраивание ДНК последовательностей в геном клетки человека,
последовательность (SEQ ID N1), кодирующую модифицированный белок TRIM 5α человека (SEQ ID N3) и отличающуюся от исходной последовательности SEQ ID N2 множественными синонимичными заменами, обеспечивающими увеличение процента GC пар в составе ДНК последовательности,
и одну или более последовательностей, выбранных из SEQ ID N 7, 8, 9, кодирующих искусственную микроРНК, которая ингибирует экспрессию гена рецептора человека CCR5 (miRNA CCR5).
29. Генетическая конструкция по п.28, где процент GC пар в последовательности, кодирующей модифицированный белок TRIM 5α человека (SEQ ID N3), составляет от 49 до 64%.
30. Генетическая конструкция по п.28, где в качестве последовательности, кодирующей модифицированный белок TRIM 5α человека (SEQ ID N3), использована последовательность SEQ ID N1.
31. Генетическая конструкция по пп.28-30, дополнительно включающая последовательность SEQ ID N4, кодирующую РНК шпильку, аналогичную TAR-последовательности (TAR), и/или антисмысловую последовательность к участку гена env ВИЧ SEQ ID N5 (а/s env).
32. Генетическая конструкция по пп.28-30, где по меньшей мере одна из последовательностей: микроРНК CCR5 и/или последовательность, кодирующая модифицированный белок TRIM 5α человека, находится под контролем конститутивного, тканеспецифичного или индуцибельного промотора.
33. Генетическая конструкция по пп.28-30, где в качестве промотора используется один из следующих промоторов: промотор гена PGK, гена EF1α, гена UbC, промотор генов МНС класса II, промотор гена Т-клеточного рецептора, промотор гена β-интерферона, промотор гена IL2, промотор гена рецептора IL2, тетрациклин зависимый промотор или промотор РНК-полимеразы III.
34. Генетическая конструкция по пп.28-30, где в качестве вектора используется лентивирусный вектор на основе ВИЧ-1, который включает по меньшей мере следующие области:
5′-LTR и 3′LTR,
участок связывания специфической тРНК (PBS),
упаковочный сигнал (ψ),
rev-отвечающий элемент (RRE),
посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE),
область сРРТ.
35. Генетическая конструкция по пп.28-30, где в качестве вектора используется лентивирусный вектор на основе ВИЧ-1, который включает по меньшей мере следующие области:
5′-LTR и 3′LTR,
участок связывания специфической тРНК (PBS),
упаковочный сигнал (ψ),
rev-отвечающий элемент (RRE),
посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE),
область сРРТ
и где последовательность вектора модифицирована с помощью последовательности SEQ ID N1 перед областью сРРТ.
36. Генетическая конструкция по пп.28-30, где в качестве вектора используется самоинактивирующийся лентивирусный вектор, который включает, по меньшей мере, следующие области:
RSV/5′-LTR гибридный промотор,
3′LTR, с делецией в области U3,
участок связывания специфической тРНК (PBS),
упаковочный сигнал (ψ),
rev-отвечающий элемент (RRE),
посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE),
область сРРТ.
37. Генетическая конструкция по пп.28-30, где в качестве вектора используется самоинактивирующийся лентивирусный вектор, который включает, по меньшей мере, следующие области:
RSV/5′-LTR гибридный промотор,
3′LTR, с делецией в области U3,
участок связывания специфической тРНК (PBS),
упаковочный сигнал (ψ),
rev-отвечающий элемент (RRE),
посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE),
область сРРТ
и где последовательность вектора модифицирована с помощью последовательности SEQ ID N1 перед областью сРРТ.
38. Генетическая конструкция по пп.28-30, где в качестве вектора используется лентивирусный вектор на основе одного или нескольких из лентивирусов: ВИЧ-1, ВИЧ-2, ВИО, вирус иммунодефицита крупного рогатого скота, лентивирус артрита-энцефалита коз, вирус инфекционной анемии лошадей, вирус иммунодефицита кошек, и/или вирус болезни Джембрана, или вектор, созданный на основе спумавируса.
39. Генетическая конструкция по пп.28-30, где в качестве вектора используется невирусный вектор, созданный на основе векторной системы транспозон/транспозаза.
40. Генетическая конструкция по пп.28-30, где встраивание ДНК последовательности в определенное место хромосомы человека обеспечивается за счет внесения разрыва в заданный участок хромосомы с помощью специфической эндонуклеазы, и последующей гомологичной рекомбинации между векторными последовательностями, входящими в конструкции по пп.28, 29 или 30 и прилегающими к разрыву участками хромосомы.
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ АНТИВИЧ-ТЕРАПИИ | 2010 |
|
RU2426788C1 |
| US 2012076763 A1, 29.03.2012 | |||
| WO 2007104633 A1, 20.09.2007 | |||

