Изобретение относится к препаратам и способам для введения генетического материала в клетки индивидуума. Препараты и способы изобретения могут применяться для доставки защитных и/или лечебных агентов, в том числе и генетического материала, кодирующего белковые мишени для вакцинации и лечебных белков.
В качестве средства замены дефектной генетической информации рассматривалось непосредственное введение в живой организм нормально функционирующего гена. В отдельных исследованиях ДНК вводят непосредственно в клетки живого организма без применения вирусной частицы или иного инфекционного вектора. В работе Nabel E.G. и др. (1990) Science 249, 1285-1288 раскрыта cайт-специфичная генная экспрессия in vivo гена бета-галактозидазы, перенесенного непосредственно в стенку артерии мыши. В работе Wolfe J.A. и др. (1990) Science 247, 1465-1468 раскрыта экспрессия разнообразных генов-репортеров, перенесенных непосредственно в мышцы мыши in vivo. В работе Acsadi G. и др. (1991) Nature 352, 815-818, раскрыта экспрессия гена дистрофина человека у мыши после внутримышечной инъекции ДНК конструкции. Работа Wolfe J. A. и др. (1991) Biotechniques 11(4), 474-485 которая вводится здесь в качестве ссылки, относится к условиям, влияющим на прямой перенос гена в мышцы грызуна in vivo. В работе Felgner P.L. и G. Rhodes (1991) Nature 349, 351-352 раскрыта непосредственная доставка в качестве лекарственных средств выделенных генов без применения ретровирусов.
В качестве альтернативы способу противопатогенной вакцинации предложено применение прямого переноса гена. В качестве возможной стратегии вакцинации против ВИЧ предложено применение прямого переноса гена путем единственной инъекции. Имеются сообщения о наблюдаемой клеточной иммунной реакции на ВИЧ-gp120, возникающей при введении в клетку плазмидной ДНК, кодирующей указанный гликопротеин. РСТ Международная заявка под номером PCT/US 90/01515, опубликованная 4 октября 1990, раскрывает способы вакцинации индивидуума против патогенной инфекции прямой инъекцией одних только полинуклеотидов в клетки индивидуума применением одностадийной методики. Применение трансфецирующих агентов, отличных от липофектинов, специально исключено из раскрываемых способов. Стимулирование инокулированных клеток не раскрывается и не предполагается. Раскрыта ВИЧ вакцина, состоящая из введенных полинуклеотидов, кодирующих вирусный белок gp120. Свидетельств о действенности вакцины не приводится.
В работе Thomason D. B. и др. (1990) Cell Physiol, 27, 578-581 и РСТ заявке на патент N WO 91/12329 раскрыто введение в мышечные клетки бупивакаина (bupivacaina) с целью вызвать пролиферацию сателитных клеток в виде части методики поставки гена при участии ретровируса.
Настоящее изобретение относится к способам введения в клетки индивидуума генетического материала. Способы включают стадии контактирования клеток индивидуума с усиливающим функцию полинуклеотида агентом, в качестве которого рекомендуется агент, облегчающий поглощение ДНК клеткой или усиливающий воспалительную реакцию, и введения в клетки молекул нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая либо кодирует целевой пептид или белок, либо служит матрицей для молекул функциональной нуклеиновой кислоты. Вводимые молекулы нуклеиновой кислоты не содержат ретровирусных частиц. Целевой белок может быть представлен либо белком, функционирующим в организме индивидуума, либо белком, служащим мишенью для иммунной реакции.
Настоящее изобретение относится к способу вакцинации индивидуума против патогена. Способ включает стадии контактирования клеток индивидуума с усиливающим функцию полинуклеотида агентом, в качестве которого рекомендуется агент, облегчающий поглощение ДНК клеткой или усиливающий иммунный ответ, и введения в клетки молекул нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую пептид, имеющий по меньшей мере один эпитоп, который идентичен или по существу аналогичен эпитопу, имеющемуся на антигене патогена и операбельно связанному с регуляторными последовательностями. Молекула нуклеиновой кислоты способна экспрессироваться в клетках индивидуума.
Настоящее изобретение относится к способу вакцинации человека против ВИЧ. Способ состоит во введении человеку молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую по меньшей мере один пептид, в состав которого входит по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопу, имеющемуся на ВИЧ белке, операбельно связанном с регуляторными последовательностями.
Настоящее изобретение относится к способу вакцинации человека против ВИЧ. Способ состоит во введении молекул двух различных нуклеиновых кислот в различные клетки человека. Молекула каждой нуклеиновой кислоты содержит нуклеотидную последовательность, кодирующую по меньшей мере один пептид, в состав которого входит по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопу, имеющемуся на ВИЧ белке, операбельно связанном с регуляторными последовательностями. Каждая из молекул различных нуклеиновых кислот содержит различные нуклеотидные последовательности, кодирующие по меньшей мере один отличный от другого пептид, и каждая нуклеиновая кислота способна экспрессироваться в клетках человека.
Настоящее изобретение относится к способу вакцинации индивидуума против гиперпролиферативного заболевания или аутоиммунных заболеваний. Способ состоит во введении в клетки индивидуума молекул нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую пептид, в состав которого входит по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопу, находящемуся в белке, связанном с гиперпролиферативным заболеванием, или белке, связанном с аутоиммунным заболеванием соответственно и операбельно связанном с регуляторными последовательностями, причем молекула нуклеиновой кислоты способна экспрессироваться в клетках.
Настоящее изобретение относится к способам лечения страдающего заболеванием индивидуума, включающим стадии контактирования клеток индивидуума с усиливающим функцию полинуклеотида агентом, в качестве которого рекомендуется агент, облегчающий поглощение ДНК клетками или усиливающий воспалительную реакцию, и введения в клетки индивидуума молекул нуклеиновой кислоты, содержащей нуклеотидную последовательность, функционирующую вместо дефектного гена или кодирующую молекулу, создающую лечебный эффект в организме индивидуума и операбельно связанную с регуляторными последовательностями, причем молекулы нуклеиновой кислоты способны экспрессироваться в клетках.
Настоящее изобретение относится к фармацевтическим препаратам, содержащим нуклеиновую кислоту и усилитель функции полинуклеотида. Настоящее изобретение относится к фармацевтическому набору, включающему контейнер с нуклеиновой кислотой и контейнер с усилителем функции полинуклеотида.
Настоящее изобретение относится к профилактическим и лечебным ВИЧ вакцинам, содержащим фармацевтически приемлемый носитель или разбавитель и нуклеиновую кислоту, кодирующую один или несклько пептидов, каждый из которых имеет по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопу, находящемуся в по меньшей мере одном ВИЧ белке, операбельно связанном с регуляторными последовательностями, причем нуклеиновая кислота способна экспрессироваться в клетках человека.
Настоящее изобретение относится к профилактическим и лечебным ВИЧ вакцинам, содержащим два инокулята. Первый инокулят состоит из фармацевтически приемлемого носителя или разбавителя и первой нуклеиновой кислоты. Молекула первой нуклеиновой кислоты включает нуклеотидную последовательность, кодирующую один или несколько пептидов, каждый из которых содержит по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопу, имеющемуся в по меньшей мере одном ВИЧ белке, операбельно связанном с регуляторными последовательностями, причем молекула нуклеиновой кислоты способна экспрессироваться в клетках человека. Второй инокулят состоит из фармацевтически приемлемого носителя или разбавителя и второй нуклеиновой кислоты. Молекула второй нуклеиновой кислоты включает нуклеотидную последовательность, кодирующую один или несколько пептидов, каждый из которых содержит по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопу, имеющемуся в по меньшей мере одном ВИЧ белке, операбельно связанном с регуляторными последовательностями, причем молекула нуклеиновой кислоты способна экспрессироваться в клетках человека. Молекулы первой и второй нуклеиновых кислот различны и кодируют различные пептиды.
На фиг. 1A приведена диаграмма, иллюстрирующая конструирование плазмиды pM160, созданной путем вставки образованного в ПЦР (полимеразной цепной реакции) фрагмента, кодирующего гликопротеин gp160 ВИЧ-НХВ2 в плазмиду pMAMneoBlue (Клонтех).
На фиг. 1B приведена фотография авторадиограммы вестерн блотинга полного клеточного лизата клеток, трансфецированных плазмидой pM160 (3G7 клетки), в сравнении с трансфецированными только вектором клетками (ТЕ671 клетки). Видно продуцирование gp120 и gp41 в 3G7 клетках, но не в ТЕ 671 клетках.
На фиг. 2 показана фотография авторадиограммы результатов иммуноосаждения при связывании антител сыворотки с 125J-gp160.
На фиг. 3A-3E приведены графики, показывающие результаты ELISA (ФИСА) при связывании различных сывороток с различными белками, иммобилизованными на титрационных михропланшетах.
На фиг. 4A и 4B показаны фотографии МТ-2 клеток, инфицированных ТСID50ВИЧ-1/III8 бесклеточным вирусом, предварительно инкубированным в последовательных разбавлениях антисыворотки.
На фиг. 4C приведены графики, показывающие изменение величин нейтрализации (Vn/Vo) в зависимости от факторов разбавления и построенные на основании результатов с применением контрольной сыворотки (x = мыши, иммунизируемые pMAMneoBlue вектором) и испытуемой сыворотки (о = мыши, иммунизируемые pM160).
На фиг. 4D-4G показаны фотографии H9/III8 клеток, применяемых в опытах с целью изучения синцитиального ингибирова- ния применением сыворотки от иммунизированных и контрольных животных.
На фиг. 5 приведена диаграмма, отражающая выживание иммунизированных и неиммунизированных мышей, контрольно зараженных gp160-мечеными и немечеными клетками опухоли. Мышей иммунизируют рекомбинантным gp160 белком только векторной ДНК или рекомбинантным вектором, включающим ДНК, кодирующую gp160. Мышам вводят SP2/0 клетки опухоли или SP2/0-gp160 клетки опухоли (SP2/0 клетки, трансфектированные ДНК, кодирующей gp160, и экспрессирующие gp160).
На фиг. 6 приведена карта плазмиды pGAGPOL.rev.
На фиг. 7 приведена карта плазмиды pENV.
На фиг. 8 показаны четыре макромолекулярных скелета (А, В, С и D), применяемых для построения генетического конструкта.
На фиг. 9 показаны четыре вставки (1, 2, 3 и 4), вставляемые в скелеты с получением генетических конструктов.
Настоящее изобретение относится к способу введения в клетки животного молекул нуклеиновой кислоты, чем обеспечивается высокий уровень поглощения и функционирования молекул нуклеиновой кислоты. Способ настоящего изобретения включает стадии введения в клетки индивидуума молекул нуклеиновой кислоты, не содержащих вирусных частиц, в частности ретровирусных частиц, в сочетании с введением ко-агента, усиливающего воспалительную реакцию и/или усиливающего экспрессию молекул нуклеиновой кислоты в тканях, и/или облегчающих поглощения клетками молекул нуклеиновой кислоты. Рекомендуемыми вариантами настоящего изобретения даются способы поставки в клетки индивидуума молекул нуклеиновой кислоты без применения инфекционных агентов.
Молекулы нуклеиновых кислот, поставляемые по изобретению в клетки, могут служить: 1) генетическими матрицами для белков, действующих в качестве профилактических и/или лечебных иммунизирующих средств; 2) копиями, заменяющими дефектные, отсутствующие или нефункционирующие гены; 3) генетическими матрицами для лечебных белков; 4) генетическими матрицами для антисмысловых молекул или 5) генетическими матрицами для рибозимов. В том случае, если молекула нуклеиновой кислоты кодирует белок, рекомендуется, чтобы молекула нуклеиновой кислоты включала последовательности, необходимые для транскрипции и трансляции в клетках животного. В случае молекул нуклеиновой кислоты, служащей матрицей для антисмысловых молекул и рибозимов, рекомендуется, чтобы такие нуклеиновые кислоты были связаны с регуляторными элементами, необходимыми для продукции достаточного числа копий кодируемых ими антисмысловых молекул и рибозимов соответственно. Молекулы нуклеиновой кислоты не содержат ретровирусных частиц и предпочтительно даются как ДНК, имеющей вид плазмиды.
Ко-агент также называется "усилителем функции полинуклеотида" или "УФП". УФП является соединением или смесью соединений, усиливающих воспалительную реакцию и/или усиливающих экспрессию молекул нуклеиновой кислоты в тканях, и/или облегчающих поглощение клетками молекул нуклеиновой кислоты, предпочтительно обладающих несколькими из указанных свойств. Усилители функции полинуклеотида, облегчающие поглощение клетками ДНК и РНК и стимулирующие деление и репликацию клеток, называются также стимулирующими клетку агентами. Рекомендуемые для настоящего изобретения ко-агенты выбирают из группы, включающей эфиры и анилиды бензойной кислоты. В рекомендуемых вариантах УФП представлен бупивакаином.
В соответствии с некоторыми аспектами настоящего изобретения даются препараты и способы, которыми индивидуум профилактически и/или терапевтически вакцинируют против патогена или ненормальной, связанной с заболеванием клеткой. Генетический материал кодирует пептид или белок, разделяющий по меньшей мере один эпитоп с иммуногенным белком, обнаруживаемым в являющихся мишенями патогене или клетках. Генетический материал экспрессируется клетками индивидуума, и служит иммуногенной мишенью, против которой возникает иммунная реакция. Возникшая иммунная реакция имеет широкую основу: помимо гуморальной иммунной реакции вызываются также оба вида клеточной иммунной реакции. Способы настоящего изобретения применимы для придания профилактического или терапевтического иммунитета. Таким образом, способ включает как методы защиты индивидуума от заражения патогеном или возникновения, или пролиферации специфичных клеток, так и методы лечения индивидуума, страдающего от инфицирования патогеном, гиперпролиферативного заболевания или аутоиммунного заболевания.
Настоящее изобретение позволяет вызвать широкий спектр иммунных реакций против целевых белков, т.е. белков, специфично ассоциированных с патогенами или собственными "ненормальными" клетками индивидуума. Настоящее изобретение применимо для вакцинации индивидуумов против патогенных агентов или микроорганизмов таким образом, что иммунная реакция против патогена обеспечивает защитный иммунитет против патогена. Настоящее изобретение применимо для борьбы с гиперпролиферативными заболеваниями и нарушениями, например раком, путем создания иммунной реакции против целевого белка, специфично ассоциируемого с гиперпролиферативными клетками. Настоящее изобретение позволяет бороться с аутоиммунными заболеваниями и нарушениями путем создания иммунной реакции против целевого белка, специфично ассоциируемого с клетками, ответственными за аутоиммунное состояние.
Некоторые аспекты настоящего изобретения относятся к генной терапии, т. е. к препаратам нуклеиновой кислоты и способам их введения в клетки индивидуума в виде экзогенных ионий гена, который либо соответствует дефектному, отсутствующему, нефункционирующему или частично функционирующему гену индивидуума, либо кодирует лечебные белки, т.е. белки, присутствие которых в организме индивидуума компенсирует их дефицит в организме и/или присутствие которых создает лечебное действие на организм индивидуума. Таким образом, настоящим изобретением предусмотрены средства доставки белка альтернативным путем, отличным от простого введения белка.
Подразумевается, что в применяемом здесь значении термин "целевой белок" относится к пептидам и белкам, кодируемым генными конструкциями настоящего изобретения, которые либо служат белковыми мишенями для иммунной реакции, либо действуют в качестве терапевтических или компенсирующих белков в режимах генной терапии.
Согласно настоящему изобретению ДНК и РНК, кодирующую целевой белок, вводят в клетки индивидуума, где происходит ее экспрессия с продуцированием в результате целевого белка. ДНК или РНК, кодирующая целевой белок, связана с регуляторными элементами, необходимыми для экспрессии в клетках индивидуума. Регуляторные элементы для экспрессии ДНК включают промотор и сигнал полиаденилирования. В генетический конструкт могут быть также включены и другие элементы, например область Козака.
В применяемом здесь значении термин "генетический конструкт" относится к молекуле ДНК или РНК, содержащей нуклеотидную последовательность, кодирующую целевой белок, и которая включает сигналы инициирования и терминирования, операбельно связанные с регуляторными элементами, в том числе промотором и сигналом полиаденилирования, способными направлять экспрессию в клетках вакцинированного индивидуума.
В применяемом здесь значении термин "экспрессируемая форма" относится к генному конструкту, содержащему необходимые регуляторные элементы, операбельно связанные с кодирующей последовательностью таким образом, что при их наличии в клетках индивидуума происходит экспрессия кодирующей последовательности. Кодирующая последовательность кодирует целевой белок.
В применяемом здесь значении термин "генетическая вакцина" относится к фармацевтическому препарату, содержащему генетический конструкт. Такой конструкт включает нуклеотидную последовательность, кодирующую целевой белок. В число таких препаратов входят и фармацевтические препараты, применимые для создания терапевтической иммунной реакции.
В применяемом здесь значении термин "генетический терапевтический (лечебный) препарат" относится к фармацевтическому препарату, содержащему генетический конструкт. Конструкт включает нуклеотидную последовательность, кодирующую терапевтический или компенсирующий белок.
В применяемом здесь значении термин "целевой белок" относится к белку, относительно которого может быть вызвана иммунная реакция. Целевой белок является иммуногенным белком, разделяющим по меньшей мере один эпитоп с белком из патогена или клетки нежелательного типа, например раковой клетки или клетки, участвующей в аутоиммунном заболевании, относительно которого требуется вакцинация. Иммунная реакция, направленная против целевого белка, способна защитить индивидуума от специфичных инфекции или заболевания, с которыми ассоциируется целевой белок, или вылечить от них.
В применяемом здесь значении выражение "разделять эпитоп" относится к белкам, содержащим по меньшей мере один эпитоп, который идентичен или по существу аналогичен эпитопу другого белка.
Имеется в виду, что в применяемом здесь значении выражение "по существу аналогичный эпитоп" относится к эпитопу, строение которого не идентично строению эпитопа белка, но тем не менее способному вызвать перекрестную клеточную или гуморальную иммунную реакцию.
В применяемом здесь значении термин "терапевтический (лечебный) белок" относится к белкам, присутствие которых оказывает лечебное действие на индивидуума.
Считается, что в применяемом здесь значении термин "компенсирующий белок" будет относится к белкам, присутствие которых компенсирует отсутствие в полной мере функционального, эндогенно продуцируемого белка, связанное с отсутствием, дефектом, нефункциональностью или частичной функциональностью эндогенного гена.
Генетические конструкты включают нуклеотидную последовательность, кодирующую целевой белок, операбельно связанный с необходимыми для генной экспрессии регуляторными элементами. Соответственно, введение в живую клетку молекулы ДНК или РНК приводит к экспрессии ДНК или РНК, кодирующей целевой белок, с продуцированием тем самым целевого белка.
При усвоении клеткой генетический констуркт, включающий нуклеотидную последовательность, кодирующую целевой белок, операбельно связанный с регуляторынми элементами, может остаться в клетке в виде функционирующей внехромосомной молекулы или может быть интегрирован в хромосомную ДНК клетки. ДНК может быть введена в клетку, где и будет оставаться в виде отдельного генетического материала в форме плазмиды. Или же в клетку может быть введена линейная ДНК, способная интегрироваться с хромосомой. При введении ДНК в клетку могут быть добавлены реагенты, способствующие интеграции ДНК с хромосомами. В молекулу ДНК могут быть также включены ДНК последовательности, способные промотировать интеграцию. Или же в клетку может быть введена РНК. Не исключено также создание генетического конструкта в виде линейной минихромосомы, включающей центромер, теломеры и ориджин репликации.
Молекула, кодирующая целевой белок, может представлять собой ДНК или РНК, содержащую нуклеотидную последовательность, кодирующую целевой белок. Такие молекулы могут быть представлены кДНК, геномной ДНК, синтетической ДНК или их гибридом, или молекулой РНК, например мРНК. Соответственно, считается, что в применяемом здесь значении термины "ДНК конструкт", "генетический конструкт" и "нуклеотидная последовательность" относятся к молекулам как ДНК, так и РНК.
Регуляторные элементы необходимые для генной экспрессии молекулы ДНК, включают: промотор, инициирующий кодон, терминирующий кодон и сигнал полиаденилирования. Кроме того, для генной экспрессии часто требуются усилители. Необходимо, чтобы такие элементы были операбельно связаны с кодирующей целевые белки последовательностью и чтобы регуляторные элементы были операбельны в организме индивидуума, которому они вводятся
Обычно считают, что инициирующий кодон и терминирующий кодон являются частью нуклеотидной последовательности, кодирующей целевой белок. Однако необходимо, чтобы указанные элементы были функциональны в организме индивидуума, которому вводят генный конструкт. Инициирующий и терминирующий кодоны должны находиться в одной рамке с кодирующей последовательностью.
Промоторы и сигналы полиаденилирования, которые применяют, должны быть функциональны в пределах клетки индивидуума.
Примеры промоторов, применимых в практике настоящего изобретения, особенно при получении генетической вакцины для человека, включают (но без ограничения только ими) промоторы из обезьяньего вируса 40 (ОВ40), вируса опухоли молочной железы мыши (ВОМЖМ) промотор, вируса иммунодефицита человека (ВИЧ), например ВИЧ длинного концевого повтора (ДКП) промотор, вирус Молони, ALV, цитомегаловируса (ЦМВ), например ЦМВ непосредственно ранний промотор, вируса Эпштейна-Барра (ВЭБ), вируса саркомы Рауса (ВСР), а также промоторы из генов человека, например актина человека, миозина человека, гемоглобина человека, мышечного креатина человека и металлотионеина.
Примеры сигналов полиаденилирования, применимых в практике настоящего изобретения, особенно при получении генной вакцины для человека, включают (во без ограничения только ими) ОВ40 сигналы полиаденилировавия и ДКП сигналы полиаденилирования. В частности, применяют ОВ40 сигнал полиаденилирования, являющийся рСЕР4 плазмидой (Инвитроген, Сан Диего, КА), называемый ОВ40 сигналом полиаденилирования.
Помимо регуляторных элементов, необходимых для экспрессии ДНК, в молекулу ДНК могут быть также включены и другие элементы. Также дополнительные элементы включают усилители. Усилитель может быть выбран из группы, включающей (но без ограничения только ими) актин человека, миозин человека, гемоглобин человека, мышечный креатин человека и вирусные усилители, например усилители из ЦМВ, ВСР и ВЭБ.
Генетические конструкты могут быть обеспечены ориджином репликации млекопитающего, цель которого удержание конструкта вне хромосомы и получение множества копий конструкта в клетке. Плазмиды рСЕР4 и pREP4 фирмы Инвитроген (Сан Диего, КА) содержат ориджин репликации вируса Эпштейна-Барра и область кодирования ядерного антигена EBNA-1, что обеспечивает высокопийную эписомальную репликацию без интеграции.
В некоторых рекомендуемых воплощениях изобретения применяемый вектор выбирают из векторов, описанных в Примере 46. В тех аспектах изобретения, которые относятся к генной терапии, рекомендуются конструкты с ориджином репликации, включающим необходимый для активации антиген.
В некоторых рекомендуемых воплощениях изобретения, относящихся к вакцинации, генетический конструкт содержит нуклеотидные последовательности, кодирующие целевой белок, и, кроме того, включает гены для белков, усиливающих иммунную реакцию против таких целевых белков. Примеры подобных генов включают гены, кодирующие цитокины и лимфокины, например альфа-интерферон, гамма-интерферон, происходящий из тромбоцитов фактор роста (ПТФР), GC-SF, GM-CSF, ФНО, эпидермальный фактор роста (ЭФР), ИЛ-1, ИЛ-2, ИЛ-4, ИЛ-6, ИЛ-8, ИЛ-10 и ИЛ-12. В некоторых вариантах рекомендуется, чтобы в применяемых в вакцинирующих препаратах генетических конструктах был включен ген для GM-CSF.
Может быть добавлен дополнительный элемент, служащий мишенью для разрушения клетки, если по какой-либо причине желательно исключить попадания в клетку генетического конструкта. В генетический конструкт может быть включен ген тимидин-киназы герпеса (tk). Индивидууму может быть введено лекарственное средство гангцикловир, ведущее к селективному уничтожению любых продуцирующих tk клеток, с обеспечением тем самым средств для селективного разрушения клеток генетическим конструктом.
Для максимального увеличения продуцирования белка могут быть подобраны регуляторные последовательности, хорошо соответствующие генной экспрессии в клетках, в которые был введен конструктор. Кроме того, могут быть подобраны кодоны, наиболее эффективно транскрибируемые в клетках. Любой специалист способен создать функционирующие в клетках ДНК конструкты.
Для выявления экспрессии генетические конструкты могут быть проанализированы in vltro на уровень экспрессии с применением тканевых культур клеток того же типа, что и клетки, в которые будет вводиться конструкт. К примеру, если генетическая вакцина должна быть введена в мышечные клетки человека, для определения уровня экспрессии в качестве модели in vitro могут быть использованы мышечные клетки, выращенные в культуре, например, клетки твердой мышечной опухоли рабдомиосаркомы.
Применяемые в настоящем изобретении генетические конструкты не вводятся в ретровирусные частицы. Генетические конструкты усвиваются клеткой без участия передаваемой ретровирусной частицей вставки, например такой, которая возникает, когда ретровирусными частицами с введенными в них РНК инфицируют клетку. В применяемом здесь значении термин "без ретровирусных частиц" относится к генетическим конструктам, которые не вводятся в ретровирусные частицы. В применяемом здесь значении выражение "отделенный от инфекционного агента" относится к генетическому материалу, не являющемуся частью вирусного, бактериального или эукариотного вектора (активного, инактивированного, живого или мертвого), способного инфицировать клетку.
В некоторых воплощениях изобретения генетические конструкты представляют собой недостаточно полные способные реплицироваться вирусные геномы, в результате чего при введении в клетку генетический конструкт обладает генетической информацией, недостаточной для обеспечения продукции инфекционных вирусных частиц. В применяемом здесь значении термин "неполный вирусный геном" относится к генетическому конструкту, содержащему недостаточно полный геном, в силу чего при введении такого генетического конструкта в клетку не происходит ввода генетической информации, достаточной для продуцирования инфекционного вируса.
В некоторых вариантах ослабленная вирусная вакцина может быть поставлена в виде генетического конструкта, содержащего достаточно генетического материала, чтобы позволить продуцирование вирусных частиц. Поставка ослабленной вакцины в виде генетического конструкта представляется более легким путем для продуцирования больших количеств безопасного, чистого активного вакцинирующего продукта.
Генетический конструкт может быть введен с применением или без применения "микроснарядов". Преимуществом является то, что генетический конструкт настоящего изобретения может быть доставлен в клетки индивидуума без твердых частиц. Имеется в виду, что выражение "без твердых частиц" в применяемом здесь значении относится к жидкости, которая не содержит каких-либо твердых "микроснарядов", применяемых в качестве средства для перфорации, прокалывания или каким-то иным путем протыкания клеточной мембраны с целью создания входного отверстия для поступления в клетку генетического материала.
Настоящее изобретение может быть применено для вакцинации индивидуума против всех патогенов, например вирусов, прокариотных и патогенных эукариотных микроорганизмов, таких как одноклеточные патогенные микроорганизмы и многоклеточные паразиты. Настоящее изобретение особенно пригодно для вакцинации индивидуума против тех патогенов, которые инфицируют клетку и которые не капсулируются, например вирусов и прокариотов, таких как гоноррея, листерез и шигеллез. Кроме того, настоящее изобретение применимо также для вакцинации индивидуума против патогенов из класса простейших, в жизненном цикле которых имеется стадия, когда они являются внутриклеточными патогенами. Имеется в виду, что в применяемом здесь значении термин "внутриклеточный патоген" к вирусу и патогенному микроорганизму, которые по меньшей мере часть своего репродуктивного или жизненного цикла проводят в клетке-хозяине, в которой продуцируют или принуждаются к продуцированию патогенных белков. В Таблице 1 приведен перечень некоторых вирусных семейств и родов, для которых могут быть изготовлены вакцины по настоящему изобретению. В вакцинах могут быть использованы ДНК конструкты, включающие ДНК последовательности, кодирующие пептиды, в состав которых входит по меньшей один эпитоп, идентичный или по существу аналогичный эпитопу, имеющемуся на антигене патогена, например тех антигенов, перечень которых приведен в таблицах. Более того, настоящее изобретение применимо также для вакцинации индивидуума против других патогенов, в том числе прокариотных и эукариотных патогенов из класса простейших, а также многоклеточных паразитов, например тех, перечень которых приведен в Таблице 2.
Для создания генетической вакцины, предназначенной для защиты от патогенной инфекции, в генетический конструкт должен быть включен генетический материал, кодирующий иммуногенные белки, против которых может быть создана защитная иммунная реакция. Будет ли инфекция патогеном внутриклеточной, что особенно приемлемо для настоящего изобретения, или внеклеточной, маловероятно, чтобы все антигены патогена создавали защитную реакцию. Поскольку и ДНК, и РНК сравнительно малы и могут быть получены сравнительно легко, настоящее изобретение обладает дополнительным преимуществом, заключающимся в том, что допускает вакцинацию рядом антигенов патогена. Применяемый в генетической вакцине генетический конструкт может включать генетический материал, кодирующий целый ряд патогенов антигена. К примеру, в один конструкт может быть включено несколько вирусных генов с обеспечением тем самым множества мишений. Кроме того, может быть приготовлен целый ряд инокулятов, которые могут быть поставлены в различные клетки, и совместно включающих в некоторых случаях полный или, что более предпочтительно, неполный, например почти полный, набор генов в вакцине. К примеру, полный набор вирусных генов может быть введен применением двух конструктов, каждый из которых содержит разные половины генома, вводимые в разных участках. Таким образом, может быть создана иммунная реакция против каждого антигена без риска вовлечения инфекционного вируса. Это позволяет вводить более одной мишени антигена, а также позволяет исключить требование идентифицировать защитные антигены.
Простота в обращении и недорогая природа ДНК и РНК позволяют, кроме того, более эффективно отбирать защитные антигены. Гены могут быть отсортированы и подвергнуты системному анализу более легко, чем белки. Отбирают патогенные агенты микроорганизмы, для защиты от которых создается вакцина, и затем идентифицируют иммуногенный белок. Таблицы 1 и 2 включают перечни некоторых патогенных агентов и микроорганизмов, для защиты от инфицирования которыми могут быть созданы генетические вакцины. В некоторых рекомендуемых вариантах способы вакцинации индивидуума от патогена направлены против ВИЧ, HTLV или ВЭБ.
Другим аспектом настоящего изобретения являются способ придания на широкой основе защитной иммунной реакции против гиперпролиферирующих клеток, характерных для гиперпролиферирующих заболеваний, и способ лечения страдающего гиперпролиферативным заболеванием индивидуума. Подразумевается, что в применяемом здесь значении термин "гиперпролиферативное заболевание" относится к тем заболеваниям и нарушениям, которые характеризуются гиперпролиферацией клеток. Примеры гиперпролиферативных заболеваний включают рак всех видов и псориаз.
Обнаружено, что введение генетического конструкта, включающего нуклеотидную последовательность, кодирующую иммуногенный, ассоциированный с "гиперпролиферирующей клеткой" белок, в клетку индивидуума приводит к продуцированию указанных белков в вакцинированных клетках индивидуума. Имеется в виду, что в применяемом здесь значении термин "гиперпролиферативно ассоциируемый белок" относится к белкам, ассоциируемым с гиперпролиферативным заболеванием. Для вакцинации от гиперпролиферативных заболеваний индивидууму вводят генетический конструкт, включающий нуклеотидную последовательность, кодирующую ассоциируемый с гиперпролиферирующим заболеванием белок.
Для того чтобы гиперпролиферативно ассоциируемый белок был эффективной иммуногенной мишенью, это должен быть белок, продуцируемый исключительно или на более высоком, чем в нормальных клетках, уровне в гиперпролиферативных клетках. Целевые антигены включают те белки, их фрагменты и пептиды, которые содержат по меньшей мере один эпитоп, обнаруживаемый в указанных белках. В отдельных случаях гиперпролиферативно ассоциируемый белок является продуктом мутации кодирующего белок гена. Мутировавший ген кодирует белок, почти идентичный нормальному белку за исключением некоторых отличий в аминокислотной последовательности, что приводит к отличному, не обнаруживаемому в нормальном белке эпитопу. Подобные целевые белки включают те белки, которые кодируются онкогенами, например: myb, myc, fyn, а также транслокационным геном bcr/ab1, ras, src, Р53, neu, trk и ECRF. Помимо онкогенных продуктов в качестве целевых антигенов целевые белки для противоракового лечения и защитных режимов включают переменные области антител, создаваемых лимфомами В-клеток, и переменные области рецепторов Т-клеток из Т-клеточных лимфом, а в некоторых вариантах применяют также целевые антигены для аутоиммунных заболеваний. В качестве целевых белков могут быть использованы и другие ассоциируемые с опухолью белки, например белки, обнаруживаемые на самом высоком уровне в клетках опухоли, в том числе белки, распознаваемые моноклональным антителом 17-1А, и связывающие фолат белки.
Хотя настоящее изобретение может быть использовано для вакцинации индивидуума от одной или нескольких форм рака, тем не менее настоящее изобретение особенно пригодно для профилактической вакцинации индивидуума, предрасположенного к развитию определенного вида рака или болевшего раком, вследствие чего склонного к рецидиву. Достижения генетики и технологии, а также эпидемиологии позволяют определять вероятность и риск развития рака у индивидуума. Применением генетического отбора и/или историй болезни семьи можно предсказать вероятность развития у конкретного индивидуума одного из нескольких типов рака.
Аналогично, те индивидуумы, которые уже перенесли рак и которые подвергались лечению с удалением опухоли или по каким-то иным причинам находящиеся в состоянии ремиссии, особенно склонны к рецидиву и повторным проявлениям рака. В виде части лечебного процесса такие индивидуумы могут быть вакцинированы от рака, который был у них диагностирован, с целью борьбы с повторными его проявлениями. Таким образом, после того как стало известно, что индивидуум был болен раком какого-то типа и имеется риск рецидива, этот индивидуум может быть вакцинирован с целью подготовки его иммунной системы для борьбы с любыми будущими проявлениями рака.
Настоящим изобретением дается способ лечения индивидуума, страдающего гиперпролиферативным заболеванием. В таких способах введение генетических конструктов служит иммунотерапевтическим, направляющим и промотирующим иммунную систему целям с целью борьбы с продуцирующими гиперпролиферативными клетками.
Настоящим изобретением дается способ лечения индивидуумов, страдающих аутоиммунными заболеваниями и нарушениями приданием на широкой основе иммунной реакции против целей, ассоциируемых с аутоиммунитетом, в том числе клеточных рецепторов и клеток, продуцирующих "само"-направленные антитела.
Передаваемые через Т-клетки аутоиммунные заболевания включают: ревматоидный артрит (РА), множественный склероз (МС), синдром Сьегрена, саркоидоз, инсулинозависимый сахарный диабет (ИЗСД), аутоиммунный тиреоидит, реактивный артрит, анкилозирующий спондилоартрит, склеродермия, полимиозит, дерматомиозит, болезнь Крона и язвенный колит. Каждое из указанных заболеваний характеризуется участием Т-клеточных рецепторов, связывающихся с эндогенными антигенами и инициирующих воспалительный каскад, ассоциируемый с аутоиммунными заболеваниями. Вакцинация от переменной области Т-клеток способна вызвать иммунную реакцию, в том числе ЦТЛ с удалением этих Т-клеток.
В случае РА охарактеризовано несколько специфичных переменных областей Т-клеточных рецепторов (ТКР), участвующих в заболевании. Подобные ТКР включают: Vβ-3,Vβ-14,Vβ-17 и Vα-17. Таким образом, вакцинация ДНК конструктом, кодирующим по меньшей мере один из указанных белков, вызовет иммунную реакцию, направленную на участвующие в RA Т-клетки. См.: Howell M.D. и др. , 1991, Proc. Natl. Acad. Sci. США 88, 10821-10925; Palliard X. и др., 1991, Science 253, 325-329; Williams W.V. и др., 1992, J.CIin.Invest. 90, 326-333; каждая из этих работ вводится здесь в качестве ссылки.
В случае МС охарактеризовано несколько специфичных переменных областей ТКР, участвующих в заболевании. Такие ТКР включают: Vβ-7 и Vα-10. Таким образом, вакцинация ДНК конструктом, кодирующим по меньшей мере один из указанных белков, создает иммунную реакцию, направленную на участвующие в МС Т-клетки. См.: Wucherphennig K.W. и др., 1990, Science 248, 1016-1019; Oksenberg J.R. и др., 1990, Nature 345, 344-346; каждая из этих работ вводится в качестве ссылки.
В случае склеродермии охарактеризовано несколько специфичных переменных областей ТКР, участвующих в заболевании. Эти ТКР включают:
Таким образом, вакцинации ДНК конструктом, кодирующим по меньшей мере один из указанных белков, создаст иммунную реакцию, направленную на участвующие в склеродермии Т-клетки.
При лечении больных, страдающих передаваемым через Т-клетки аутоиммунным заболеванием, в частности больных, для которых переменные области ТКР еще не охарактеризованы, может быть проведена синовиальная биопсия. Могут быть отобраны образцы присутствующих Т-клеток, в которых с помощью стандартных методик идентифицируют переменные области этих ТКР. На основе полученной информации могут быть приготовлены генетические вакцины.
Передаваемые через В-клетки аутоиммунные заболевания включают: волчанку (СКВ), тяжкую миастению, аутоиммунную гемолитическую анемию, аутоиммунную тромбоцитопению, астму, криоглобулинемию, первичный билиарный склероз и пернициозную анемию. Каждое из указанных заболеваний характеризуется антителами, связывающимися с эндогенными антигенами и инициирующими воспалительный каскад, ассоциируемый с аутоиммунными заболеваниями. Вакцинация от переменных областей антител способна вызвать иммунную реакцию, в том числе ЦТЛ с удалением продуцирующих антитело В-клеток.
Для лечения больного, страдающего передаваемым через В-клетки аутоиммунным заболеванием, необходимо идентифицировать переменную область антител, проявляющих аутоиммунную активность. Может быть проведена биопсия с отбором образцов антител, присутствующих в месте воспаления. Переменная область этих антител может быть идентифицирована с помощью стандартных методик. На основании полученной информации могут быть приготовлены генетические вакцины.
В случае СКВ один из антигенов, как полагают, представлен ДНК. Таким образом, у больных, подлежащих вакцинации от СКВ, их сыворотка может быть проанализирована на анти-ДНК антитела и может быть приготовлена вакцина, включающая ДНК конструкты, кодирующие переменную область таких, обнаруженных в сыворотке анти-ДНК антител.
Общие структурные признаки для переменных областей как ТКР, так и антител хорошо известны. ДНК последовательность, кодирующая конкретные ТКР и антитело, как правило, может быть выявлена с помощью хорошо известных методов, описанных, например, в издании: Kabat и др., 1987, "Последовательности представляющих иммунологический интерес белков", США, Департамент здоровья и службы человека, Бетесда, МД, которое вводится здесь в качестве ссылки. Кроме того, общий способ клонирования функциональных переменных областей из антител можно найти в работе: Chaudhery V.К. и др., 1990, Proc. Natl.Acad. Sci. США 87, 1066, которая вводится здесь в качестве ссылки.
В некоторых воплощениях изобретения, относящихся к генной терапии, генные конструкты содержат либо компенсирующие гены, либо гены, кодирующие лечебные белки. Примеры компенсирующих генов включают ген, кодирующий дистрофин или его функциональный фрагмент, ген, предназначенный компенсировать дефектный ген у больных, страдающих муковисцидозом, инсулин, ген, предназначенный компенсировать дефектный ген у больных, страдающих ADA, и ген, кодирующий фактор VIII. Примеры генов, кодирующих лечебные белки, включают гены, кодирующие эритропоиэтин, интерферон, LDL рецептор, GM-CSF, ИЛ-2, ИЛ-4 и ФНО. Кроме того, могут быть введены генетические конструкты, кодирующие одноцепочечные компоненты антитела, специфично связывающиеся с токсичными веществами.
В некоторых рекомендуемых вариантах ген дистрофина дается в виде минигена, который применяют для лечения индивидуумов, страдающих мышечной дистрофией. В отдельных рекомендуемых вариантах дается миниген, содержащий последовательность, кодирующую часть белка дистрофина. Нарушения в гене дистрофина ведут как к более мягкой мышечной дистрофии Бекера
(МДБ), так и к более тяжкой мышечной дистрофии Дученне (МДД) В случае МДБ дистрофин образуется, но имеет отклонения от нормы по размеру и/или по количеству. Больной находится в состоянии от слабого до умеренно слабого. При МДД белок не образуется, и больной прикован к креслу в возрасте 13 лет и обычно умирает в возрасте 20. У некоторых больных, в частности больных МДБ, частичный белок дистрофин, полученный экспрессией минигена, поставляемого способом изобретения, может привести к улучшению мышечной функции.
В некоторых рекомендуемых вариантах гены, кодирующие ИЛ-2, ИЛ-4, интерферон или ФНО, поставляются в клетки опухоли, которые либо присутствуют, либо удалены и затем возвращены в организм индивидуума. В отдельных вариантах индивидууму, страдающему множественным склерозом, вводят ген, кодирующий гамма-интерферон.
Антисмысловые молекулы и рибозимы также могут быть поставлены в клетки индивидуума путем введения генетического материала, действующего в качестве матрицы для копий таких активных агентов. Такие агенты дезактивируют или каким-то иным путем влияют на экспрессию генов, кодирующих белки, присутствие которых нежелательно. Конструкты, содержащие последовательности, кодирующие антисмысловые молекулы, могут применяться для подавления или предотвращения продуцирования белков в клетке. Так, продуцирование таких белков, как онкогеновые продукты, может быть исключено или снижено. Аналогично, рибозимы способны прервать генную экспрессию селективным разрушением матричной РНК до ее трансляции в белок. В некоторых вариантах клетки обрабатывают способом изобретения с применением конструктов, кодирующих антисмысловые молекулы или рибозимы в виде части лечебного процесса, включающего введение других лекарственных средств и другие процедуры. В генных конструктах, кодирующих антисмысловые молекулы и рибозимы, применяют вектора, аналогичные тем, которые применяют, когда желательно продуцирование белка, за исключением того, что кодирующая последовательность не имеет инициирующего кодона, инициирующего трансляцию РНК в белок. В некоторых вариантах рекомендуется, чтобы вектора, описанные, в частности, в Примере 46, содержали ориджин репликации и экспрессируемую форму соответствующего ядерного антигена.
Рибозимы являются каталитическими РНК, способными саморасщепляться или расщеплять молекулы других РНК. Специалистам известны рибозимы нескольких различных типов, например типа молота, шпильки, интрон группы I Tetrahymena, топора и РНКазы Р (S. Edgington, Biotechnology, 1992, 10, 256-262). Рибозимы типа молота обладают каталитическим участком, который был картирован до ядра в менее чем 40 нуклеотидов. Некоторые рибозимы в растительных вироидах и сателитных РНК делят общую вторичную структуру и определенные консервативные нуклеотиды. Хотя указанные рибозимы в природе выступают в роли собственного субстрата, домен субстрата может быть направлен на другой РНК субстрат путем спаривания оснований в последовательностях, фланкирующих консервированный участок расщепления. Такая способность рибозимов обычного строения позволяет применять их для последовательность-специфичного расщепления РНК (G. Peolella и др., ЕМВО 1992, 1913-1919). Таким образом, в компетенции специалиста применение различных каталитических последовательностей из рибозимов разных типов, например каталитической последовательности типа молота, и конструирование их вышеприведенным путем. Могут быть созданы рибозимы, направленные на разнообразные цели, в том числе патогенные нуклеотидные последовательности и онкогенные последовательности. Определенные рекомендуемые варианты изобретения включают достаточную комплементарность к специфично нацеленному abl-bcr слитому транскрипту при сохранении эффективности реакции расщепления.
В соответствии с некоторыми воплощениями настоящего изобретения клетки обрабатывают соединениями, облегчающими поглощение клеткой генетического конструкта. Согласно определенным воплощениям изобретения клетки обрабатывают соединениями, облегчающими клеточное деление и способствующими поглощению генетического конструкта. Введение соединений, облегчающих поглощение клеткой генетического конструкта, в том числе стимулирующих клетку соединений, приводит к более эффективной иммунной реакции против кодируемого генетическим конструктом целевого белка.
Согласно некоторым воплощениям настоящего изобретения генетический конструкт вводят в организм индивидуума с помощью безыгольного устройства для инъекций. В соответствии с некоторыми воплощениями настоящего изобретения генетический конструкт вводят одновременно в организм индивидуума внутрикожно, подкожно и внутримышечно с помощью безыгольного устройства для инъекций. Безыгольные устройства для инъекций хорошо известны и широко доступны. Обычный специалист, руководствуясь данным описанием, для поставки генетического материала в клетки индивидуума применяет безыгольное устройство для инъекций. Безыгольные устройства для инъекций хорошо подходят для поставки генетического материала во все ткани. Устройства особенно пригодны для поставки генетического материала в клетки кожи и мышц. В некоторых вариантах безыгольное устройство для инъекций может быть использовано для продвижения содержащей молекулы ДНК жидкости к поверхности кожи индивидуума. Жидкость продвигается с достаточной скоростью так, что при соударении с кожей жидкость проникает через поверхность кожи и проходит в нижерасположенные ткани кожи и мышц. Таким образом, генетический материал вводится одновременно чрескожно, подкожно и внутримышечно. В некоторых вариантах безыгольное устройство для инъекций может быть использовано для поставки генетического материала в ткани другого органа с целью введения молекулы нуклеиновой кислоты в клетки этого органа.
Согласно изобретению генетическая вакцина может быть введена непосредственно в организм подлежащего вакцинации индивидуума или может быть введена ex vivo в удаленные клетки индивидуума с последующей их реимплантацией после введения. При любом пути введения генетический материал вводится в клетки, присутствующие в организме индивидуума. Пути введения включают (но без ограничения только ими): внутримышечный, внутрибрюшинный, внутрикожный, подкожный, внутривенный, внутриартериальный, внутриглазной и пероральный, а также чрезкожный, в виде ингаляции или супозитория. Рекомендуемые пути введения включают внутримышечные, внутрибрюшинные, внутрикожные и подкожные инъекции. Поставка кодирующих целевые белки генных конструктов может придать защитные свойства слизистым оболочкам индивидуума, если введение осуществляют путем, в котором материал присутствует в тканях, ассоциируемых с защитными свойствами слизистых оболочек. Так, в некоторых примерах генный конструкт поставляется путем введения в щечный карман, находящийся во рту индивидуума.
Генетические конструкты могут быть введены применением (но не только их) традиционных шприцев, безыгольных устройств для инъецирования или "генных пистолетов для микробомбардировки". Или же генетическая вакцина может быть введена разнообразными способами в клетки, удаленные из организма индивидуума. Такие способы включают, например, ex vivo трансфекцию, электропорацию, микроинъецирование и микробомбардировку. После поглощения клетками генного конструкта их вновь имплантируют в организм индивидуума. Считается, что обычно неиммуногенные клетки после введения в них генных конструктов могут быть имплантированы в организм индивидуума, даже если вакцинированные клетки изначально были отобраны у другого индивидуума.
Генетические вакцины изобретения содержат ДНК в интервале от 1 нг до 1000 мкг. В некоторых рекомендуемых вариантах вакцина содержит ДНК в пределах от 10 нг до 800 мкг. В отдельных рекомендуемых вариантах вакцина содержит 0.1-500 мкг ДНК. В других рекомендуемых вариантах вакцины содержат 1-350 мгк ДНК. И в еще одних рекомендуемых вариантах вакцины содержат 25-250 мкг ДНК. В других рекомендуемых вариантах вакцины содержат около 100 мкг ДНК.
Генетические вакцины настоящего изобретения готовят в зависимости от намеченного к применению пути введения. Обычный специалист без труда приготовит генетическую вакцину, содержащую генетический конструкт. В случае, если в качестве пути введения выбраны внутримышечные инъекции, рекомендуют применять изотонические препараты. Применяемые для придания изотоничности добавки, как правило, включают: хлорид натрия, декстрозу, маннит, сорбит и лактозу. В некоторых случаях рекомендуются такие изотонические растворы, как фосфатные буферные солевые растворы. В некоторых вариантах к препарату добавляют суживающее сосуды средство. Стабилизаторы включают желатин и альбумин. Фармацевтические препараты настоящего изобретения даются стерильными и не содержащими пирогенов.
Генетические конструкты изобретения готовят в виде препарата или вводят в сочетании с усилителем функции полинуклеотида. Рекомендуемые соагенты настоящего изобретения выбирают из группы, включающей: эфиры, анилиды, амидины, уретаны бензойной кислоты и их гидрохлориды, принадлежащие, например, к семейству местных анестетиков.
УФП может быть соединением, представленным одной из следующих формул:
Ar-R1-O-R2-R3,
или
Ar-N-R1-R2-R3,
или
R4-N-R5-R6,
или
R4-O-R1-N-R7,
где Ar представляет бензол, п-аминобензол, м-аминобензол, о- аминобензол, замещенный бензол, замещенный п-аминобензол, замещенный м-аминобензол, замещенный о-аминобензол, где аминогруппа в аминобензольных производных может быть представлена амино-, С1-С5-алкиламино-, ди(С1-C5-алкил) аминогруппой, и заместители в замещенных производных представлены галогеном, С1-C5-алкилом и C1-C5-алкоксигруппой;
R1 представляет C=O;
R2 представляет C1-C10-алкил, в том числе разветвленные алкилы;
R3 представляет водород, амин, C1-C5-алкиламин, ди(C1-C5-алкил) амин,
R2 + R3 могут образовывать циклический алкил, C1-C5- алкилзамещенный циклический алкил, циклический алифатический амин, C1-C10-алкилзамещенный циклический алифатический амин, гетероцикл, C1-C10-алкилзамещенный гетероцикл, в том числе C1-C10)-алкил-N-замещенный гетероцикл;
R4 представляет Ar, R2 или C1-C5-алкоксигруппу, циклический алкил, C1-C10-алкилзамещенный циклический алкил, циклический алифатический амин, C1-C10-алкилзамещенный циклический алифатический амин, гетероцикл, C5-C10-алкилзамещенный гетероцикл и C1-C10-алкоксизамещенный гетероцикл, в том числе C1-C10-алкил-N-замещенный гетероцикл;
R5 представляет C=NH;
R6 представляет Ar, R2 или C1-C10-алкоксигруппу, циклический алкил, C1-C10-алкилзамещенный циклический алкил, циклический алифатический амин, C1-C10-алкилзамещенный циклический алифатический амин, гетероцикл, C1-C10-алкилзамещенный гетероцикл и C1-C10-алкоксизамещенный гетероцикл, в том числе C1-C10-aлкил-N-зaмeщeнный гетероцикл и
R7 представляет Ar, R2 или C1-C5-алкоксигруппу, циклический алкил, C1-C10-алкилзамещенный циклический алкил, циклический алифатический амин, C1-C10-алкилзамещенный циклический алифатический амин, гетероцикл, C1-C10-алкилзамещенный гетероцикл и C1-C10-алкоксизамещенный гетероцикл, в том числе C1-C10-алкил-N-замещенный гетероцикл.
Примеры эфиров включают эфиры бензойной кислоты, например пиперокаин, меприлкаин и изобукаин, эфиры пара-аминобензойной кислоты, например прокаин, тетракаин, бутетамин, пропоксикаин и хлорпрокаин; эфиры мета-аминобензойной кислоты, в их числе метабутамин и примакаин; эфиры пара-этоксибензойной кислоты, например парэтоксикаин. Примеры анилидов включают: лидокаин, этидокаин, мепивакаин, бупивакаин, пиррокаин и прилокаин. Другие примеры подобных соединений включают: дибукаин, бензокаин, диклонин, прамоксин, пропаракаин, бутакаин, беноксинат, карбокаин, метилбупивакаин, пикрат бутазина, фенакаин, диотан, луккаин, интракаин, нуперкаин, метабутоксикаин, пиридокаин, бифенамин, а также бициклические соединения растительного происхождения, например кокаин, цинамоилкокаин, труксиллин и кокаэтилен и все перечисленные соединения в виде гидрохлоридов.
В рекомендуемых вариантах УФП представлен бупивакаином. Разница между бипивакаином и мепивакаином заключается в том, что бупивакаин имеет N-бутильную группу вместо N-метильной группы мепивакаина. В этом случае речь идет о N-С1-С10- алкилпроизводных. Соединения могут быть замещены галогеном, как в случае с прокаином и хлорпрокаином. Анилиды более предпочтительны.
Бупивакаин вводят до, одновременно или после генетического конструкта. Бупивакаин и генетический конструкт могут быть приготовлены в виде совместного препарата. Бупивакаин особенно полезен в качестве стимулирующего клетку агента, что объясняется проявлением его активности и его многообразными свойствами при введении в ткани. Бупивакаин способствует и облегчает поглощение клеткой генетического материала. Сам по себе бупивакаин является трансфектирующим средством. Введение генетических конструктов совместно с бупивакаином облегчает вхождение генетического конструкта в клетку. Полагают, что бупивакаин разрывает клетку или каким-то иным образом делает клетку более проницаемой. Бупивакаин стимулирует деление и репликацию клеток. Следовательно, бупивакаин действует, как реплицирующее средство. Введение бупивакаина, кроме того, раздражает и повреждает ткани. Как таковой бупивакаин действует как воспалительное средство, вызывающее миграцию и хемотаксис иммунных клеток в месте введения. Помимо клеток, обычно присутствующих в месте введения, клетки иммунной системы, мигрировавшие в виде иммунного ответа на воспалительное средство, могут вступить в контакт с введенными генетическим материалом и бупивакаином. Бупивакаин, действующий в качестве трансфецирующего средства способен также промотировать поглощение генетического материала и такими клетками иммунной системы.
Химически и фармакологически бупивакаин относится к аминоацильным местным анестетикам. Бупивакаин является гомологом мепивакаина и родственен лидокаину. Бупивакаин придает мышечной ткани чувствительность к электрическому напряжению при введении натрия и влияет на концентрацию ионов в клетке. Полное описание фармакологической активности бупивакаина можно найти в издании: Ritchie J. M. и N.M. Greene, "Фармакологические основы терапии", ред. Gilman A.G. и др., 8-ое издание, глава 15, стр. 3111, которое вводится здесь в качестве ссылки. В способе настоящего изобретения рекомендуется применение бупивакаина и функционально аналогичных бупивакаину соединений.
Бупивакаин-HCl имеет следующее химическое название: 2-пиперидинкарбоксамид, 1-бутил-N-(2,6-диметилфенил)- моногидрохлорид, моногидрат, и широко выпускается для применения в фармацевтике многими фирмами, в том числе Астра Фармасьютикл Продуктс Инк. (Уэстборо, МА), Санофи Уинтроп Фармасьютиклз (Нью-Йорк, Н-Й), Истман Кодак (Рочестер, Н-Й). В продажу бупивакаин поступает с добавкой или без метилпарабена, а также с добавкой или без эпинефрина. Могут быть использованы любые такие препараты. Для применения в фармацевтике бупивакаин поступает в продажу в концентрации 0.25%, 0.5% и 0.75% и в таком виде может быть использован в изобретении. Или же при желании могут быть использованы альтернативные, создающие необходимый эффект концентрации, в частности 0.05-1%. Согласно настоящему изобретению бупивакаин вводится в количестве от 250 мкг до 10 мг. В некоторых вариантах вводят от 250 мкг до 7.5 мг бупивакаина. В отдельных вариантах вводят 0.05-5 мг бупивакаина. В других вариантах бупивакаин вводят в количестве 0.5-3 мг. И еще в одних вариантах вводят 5-50 мкг бупивакаина. К примеру, в некоторых вариантах в том же участке, что и вакцина вводят от 50 мкл до 2 мл, предпочтительно 50-1500 мкл, более предпочтительно около 1 мл 0.5% бупивакаин-HCl и 0.1% метилпарабена в изотоническом фармацевтическом носителе до, одновременно или после введения вакцины. Аналогично, в отдельных вариантах в том же участке, что и вакцина, вводят от 50 мкл до 2 мл, предпочтительно 50-1500 мкл и более предпочтительно около 1 мл 0.5% бупивакаин-HCl в изотоническом фармацевтическом носителе до, одновременно или после введения вакцины. Бупивакаин и любые другие, аналогично действующие соединения, в частности соединения, относящиеся к семейству местных анестетиков, могут быть введены в концентрациях, обеспечивающих необходимое облегчение поглощения клеткой генетических конструктов.
В некоторых воплощениях изобретения индивидууму вначале вводят инъекцию бупивакаина перед генетической вакцинацией внутримышечной инъекцией, т.е. в пределах от недели до десяти дней, например, индивидууму вначале вводят инъекцию бупивакаина. В некоторых вариантах перед вакцинацией индивидууму за 1-5 дней перед введением генетического конструкта вводят инъекцию бупивакаина. В отдельных вариантах перед вакцинацией, примерно за 24 часа до введения генетического конструкта индивидууму вводят инъекцию бупивакаина. Или же бупивакаин может быть инъецирован одновременно, за несколько минут до или после вакцинации. Соответственно бупивакаин и генетический конструкт могут быть объединены и инъецированы одновременно в виде смеси. В некоторых вариантах бупивакаин вводят после введения генетического конструкта. К примеру, индивидууму вводят инъекцию бупивакаина в пределах от недели до десяти дней после введения генетического конструкта. В некоторых вариантах индивидууму вводят инъекцию бупивакаина примерно через 24 часа после вакцинации. В отдельных вариантах индивидууму вводят инъекцию бупивакаина через 1-5 дней после вакцинации. В других вариантах индивидууму вводят бупивакаин в пределах от недели до десяти дней после вакцинации.
Другие агенты, способные выполнять роль трансфектирующих средств и/или реплицирующих средств, и/или воспалительных средств, и которые могут быть введены совместно с бупивакаином и аналогично действующими соединениями, включают: пектины, факторы роста, цитокины и лимфокины, например: α-интерферон, гамма-интерферон, происходящий из тромбоцитов фактор роста (ПТФР), GC-SF, GM-CSF, ФНО, эпидермальный фактор роста (ЭФР), ИЛ-1, ИЛ-2, ИЛ-4, ИЛ-6, ИЛ-8, ИЛ-10 и ИЛ-12, а также коллагеназу, фактор роста фибробласта, эстроген, дексаметазон, сапонины, поверхностно-активные вещества, например иммуностимулирующие комплексы (ИСКОМ), неполный адъювант Фрейнда, LPS аналог, включая монофосфорильный липид А (МФЛ), мурамиловые пептиды, аналоги хинона и пузырьки, например, сквалана и свалена, гиалуроновую кислоту и гиалуронидазу. Все эти вещества могут быть введены в сочетании с бупивакаином и генетическим конструктом. К примеру, совместно с генетическим конструктом вводят бупивакаин и либо гиалуроновую кислоту, либо гиалуронидазу.
Генетический конструкт может быть смешан с коллагеном в виде эмульсии и введен парентерально. Эмульсия коллагена дает средство для постепенного выделения ДНК. Коллаген применяют в количестве от 50 мкл до 2 мл. В рекомендуемом варианте с применением указанного состава смешивают около 100 мкг ДНК с 2 мл коллагена. Могут применяться и другие препараты продленного действия, описанные, например, в справочнике Ремингтона для фармацевтов (A. Osol), являющимся стандартным пособием в данной области, который вводится здесь в качестве ссылки. Такие препараты включают: водные суспензии, растворы и суспензии в масле, эмульсии и имплантанты, а также контейнеры и чрезкожные устройства. В некоторых вариантах рекомендуются выделяемые во времени препараты генетического конструкта. В некоторых вариантах рекомендуется, чтобы генетический конструкт выделялся во времени в пределах 6-144 часов, предпочтительно 12-96 часов, более предпочтительно 18-72 часов.
В некоторых воплощениях настоящего изобретения генетический конструкт инъецируют с помощью безыгольного устройства для инъекций. Безыгольные устройства для инъекций особенно предпочтительны для одновременного введения вещества внутримышечно, внутрикожно и подкожно.
В некоторых воплощениях настоящего изобретения генетический конструкт вводят вместе с (УФП) применением методики микрозарядной бомбардировки частицами, раскрытой Sanford и др. в патенте США 4945050, выдан 31 июля 1990 г., который вводится здесь в качестве ссылки.
В некоторых воплощениях настоящего изобретения генетический конструкт вводят в виде части липосомного комплекса с усиливающим функцию полинуклеотида агентом.
В некоторых воплощениях изобретения индивидуум подвергают единственной вакцинации с созданием полной, широкой иммунной реакции. В отдельных воплощениях изобретения индивидуума подвергают ряду вакцинаций и создают полную, широкую иммунную реакцию. Согласно некоторым воплощениям изобретения в течение какого-то времени проводят по меньшей мере две, предпочтительно четыре-пять инъекций. Промежуток времени между инъекциями может составлять от 24 часов до двух недель и более, предпочтительно одна неделя. Или же по меньшей мере две и вплоть до четырех инъекций проводят одновременно в различных участках.
В некоторых воплощениях изобретения полная вакцинация включает инъецирование единственного инокулята, содержащего генетический конструкт, включающий последовательности, кодирующие один или несколько целевых эпитопов.
В некоторых воплощениях изобретения исчерпывающая вакцинация включает инъецирование двух или более разных инокулятов в разных участках. К примеру, ВИЧ вакцина по изобретению содержит два инокулята, каждый из которых включает генетический материал, кодирующий разные вирусные белки. Такой способ вакцинации позволяет вводить возможно более полный набор вирусных генов без риска введения инфекционной вирусной частицы. Так, у вакцинированного индивидуума может быть создана иммунная реакция к большинству или всем вирусам. Инъецирование каждого инокулянта осуществляют в разных участках, предпочтительно на некотором расстоянии, чтобы избежать получение клетками обоих генетических конструктов. Для большей безопасности некоторые гены могут быть изъяты или изменены с дополнительной гарантией предотвращения возможности появления инфекционного вирусного ансамбля. В применяемом здесь значении термин "фармацевтический набор" относится собирательно к множественному инокуляту, применяемому в настоящем изобретении. Такие наборы включают отдельные контейнеры с разными инокулятами и/или стимулирующими клетку агентами. Предполагается, что эти наборы включают ряд инокулятов, применяемых в способе вакцинации.
Способы настоящего изобретения применимы в области медицины человека и ветеринарии. Соответственно настоящее изобретение относится к генетической вакцинации млекопитающих, птиц и рыб. Способы настоящего изобретения могут быть особенно полезными в применении к млекопитающим, в том числе человеку, коровам, овцам, свиньям, козам, собакам и кошкам.
Нижеприведенные Примеры включают характерные примеры аспектов настоящего изобретения. Примеры не предназначены для ограничения объема изобретения, но скорее служат целям иллюстрации. Кроме того, разнообразные аспекты изобретения суммированы в следующем описании. Однако данное описание не предназначено для ограничения объема изобретения, но скорее дается для освещения разнообразных аспектов изобретения. Обычный специалист способен легко выявить дополнительные аспекты изобретения и его варианты.
Пример 1
Настоящим изобретением дается ВИЧ вакцина, применимая для прямой генетической вакцинации. Даются генетические конструкты, которые при поступлении в клетки индивидуума экспрессируются с продуцированием ВИЧ белков. Согласно некоторым вариантам продуцирование всех вирусных структурных белков в клетках индивидуума создает защитную иммунную реакцию, защищающую от ВИЧ инфекции. ВИЧ вакцина настоящего изобретения может быть использована для вакцинации неинфицированвых индивидуумов от ВИЧ инфекции или может служить в качестве иммунотерапевтического средства для уже инфицированных индивидуумов. ВИЧ вакцина настоящего изобретения создает иммунную реакцию, в том числе ЦТЛ, позволяющую распознавать и атаковать ВИЧ инфицированные клетки и распознавать широкий спектр ВИЧ белков. Таким образом, неинфицированные индивидуумы оказываются защищенными от ВИЧ инфекции.
В некоторых своих воплощениях настоящее изобретение относится к способу вакцинации индивидуума от ВИЧ путем введения двух инокулятов. Такие два инокулята включают по меньшей мере два и предпочтительно более двух наборов генов или все гены ВИЧ вируса. Однако инокуляты не поставляются совместно. Соответственно в инокулированную клетку не поступает полный набор генов. Вакцинированный индивидуум получит по меньшей мере два разных и предпочтительно более двух, более предпочтительно целый ряд или все вирусные гены.
Такая стратегия повышает вероятность того, что генетический материал, кодирующий наиболее эффективные целевые белки, будет включен в вакцину, и снижает вероятность того, что вирусной частице удастся избежать обнаружения при иммунной реакции, несмотря на структурные изменения в одном или нескольких вирусных белках, возникающих при мутации вируса. Соответственно желательна вакцинация индивидуума целым рядом и предпочтительно почти полным или полным набором генов, кодирующих вирусные белки.
Если единичная клетка обеспечивается полным набором вирусных генов, возникает вероятность того, что в пределах клетки появится полный инфекционный вирус. Соответственно, генетический конструкт изобретения не содержит полного набора таких генов. Более того, два или более инокулята, каждый из которых имеет неполный набор генов, а все вместе имеют вплоть до полного набора вирусных генов, вводят в разные клетки, предпочтительно расположенные на некотором расстоянии друг от друга с целью избежать возможности действия на вакцинированную клетку полного набора генов. К примеру, часть ВИЧ генома может быть вставлена в первый конструкт, а оставшаяся ВИЧ генома вставлена во второй конструкт. Первый конструкт вводят индивидууму в виде генетической вакцины в мышечную ткань одной руки, а второй конструкт вводят индивидууму в виде генетической вакцины в другую руку. Индивидуум подвергается воздействию полного набора вирусных генов с вакцинацией тем самым по существу от полного вируса, но без риска возникновения инфекционной вирусной частицы.
В целях дополнительной безопасности, даже если генетический материал поставляется двумя или несколькими инокулятами в расположенные на расстоянии части тела индивидуума, один или несколько существенных генов может быть изъято или преднамеренно изменено с еще большей гарантией невозможности образования инфекционной вирусной частицы. В подобных вариантах индивидууму вводят неполный функциональный набор вирусных генов.
Дополнительными мерами безопасности даются неперекрывающиеся части вирусного генома в отдельных генетических конструктах, образующих соответственно отдельные инокуляты. Тем самым предотвращается рекомбинация между двумя генетическими конструктами.
В некоторых воплощениях настоящего изобретения дается полный набор структурных генов. Структурные гены ВИЧ включают gag, pol и env. Эти три гена даются в двух разных ДНК или РНК конструктах. Соответственно в одном рекомендуемом варианте gag и pol находятся в одном ДНК или РНК конструкте, а env - в другом. В другом рекомендуемом варианте gag находится в одном ДНК или РНК конструкте, а pol и env - в другом. В еще одном рекомендуемом варианте gag и env находятся в одном ДНК или РНК конструкте, а pol - в другом. В некоторых рекомендуемых вариантах конструкты, содержащие rev, имеют сплайс-акцептор в восходящем направлении от инициирующего кодона для rev. В отдельных рекомендуемых вариантах конструкты, содержащие gag, имеют сплайс-донор в восходящем направлении от gag трансляционного инициирующего кодона. Возможно, что в любой из приведенных комбинаций могут также присутствовать ВИЧ регуляторные гены. К ВИЧ регуляторным генам относятся: vpr, vif, vpu, tat и rev.
ДНК конструкты в рекомендуемом варианте включают промотор, усилитель и сигнал полиаденилирования. Промотор может быть выбран из группы, включающей ВИЧ ДКП, актин человека, миозин человека, ЦМВ, ВСР, Молони, ВОМЖМ, гемоглобин человека, мышечный креатин человека и ВЭБ. Усилитель может быть выбран из группы, включающей актин человека, миозин человека, ЦМВ, ВСР, гемоглобин человека, мышечный креатин человека и ВЭБ. Сигнал полиаденилирования может быть выбран из группы, включающей ДКП сигнал полиаденилирования и ОВ40 сигнал полиаденилирования, в частности среди прочих меньший ОВ40 сигнал полиаденилирования.
В некоторых вариантах две инокулятные вакцины вводят внутримышечно в пространственно разделенные ткани индивидуума, предпочтительно в разные части тела индивидуума, такие как, например, его правая и левая руки. Каждый инокулят настоящего изобретения может содержать 0.1-10000 мкг ДНК. Каждый инокулят предпочтительно содержит 1-500 мкг ДНК. Более предпочтительно, если каждый инокулят содержит 25-250 мкг ДНК. Наиболее предпочтительно, если каждый инокулят содержит около 100 мкг ДНК.
Инокулят в некоторых вариантах представляет собой стерильный изотонический носитель, предпочтительно фосфатный буферный солевой раствор или простой солевой раствор.
В некоторых вариантах перед введением вакцины в подлежащие вакцинации ткани инъецируют способствующее клеточной пролиферации средство, предпочтительно бупивакаин. Инъекции бупивакаином могут быть проведены вплоть до примерно 24 часов перед вакцинацией. Инъекцию бупивакаина целесообразно проводить непосредственно перед вакцинацией. В место введения вакцины вводят от 50 мкл до 2 мл 0.5% бупивакаин-HCl и 0.1% метилпарабена в изотоническом растворе NaCl, предпочтительно 50-1500 мкл, более предпочтительно около 1 мл.
В других вариантах способствующее пролиферации клеток средство, предпочтительно бупивакаин, включают в препарат совместно с генетическим конструктом. В место, куда должна быть введена вакцина, вводят от 50 мкл до 2 мл 0.5% бупивакаин-HCl и 0.1% метилпарабена в изотоническом растворе NaCl, предпочтительно 50-1500 мкл, более предпочтительно около 1 мл.
Соответственно, некоторыми вариантами предлагаются две инокулятные вакцины: один инокулят включает ДНК или РНК конструкт, содержащий два ВИЧ структурных гена, второй инокулят включает ДНК или РНК конструкт, содержащий третий, оставшийся структурный ген, так что вместе инокуляты содержат полный набор ВИЧ структурных генов. Структурные гены в каждом ДНК конструкте операбельно связаны с промотором, усилителем и сигналом полиаденилирования. Экспрессию вирусных генов могут регулировать одинаковые или разные регуляторные элементы. При вакцинации индивидуума два инокулята вводят внутримышечно в различных местах, предпочтительно в разные руки индивидуума. В некоторых воплощениях изобретения в место введения вакцины вначале вводят бупивакаин. В отдельных воплощениях изобретения бупивакаин включен в общий с генетическим конструктом препарат.
В некоторых вариантах процедуру вакцинации повторяют по меньшей мере еще раз, предпочтительно два или три раза. Каждую вакцинацию разделяет промежуток времени от 24 часов до двух месяцев.
В некоторых вариантах вакцину вводят с помощью безыгольного устройства для инъекций. В отдельных вариантах вакцину вводят гиподермально с помощью безыгольного устройства для инъекций с обеспечением тем самым внутримышечного, внутрикожного и подкожного введения одновременно и осуществлением также промежуточного введения вещества.
Рекомендуемые генетические конструкты включают следующее: плазмиды и конструкты, полученные клонированием.
Сконструированы две плазмиды: одна содержит ВИЧ gag/pol, а другая содержит ВИЧ env.
ВИЧ-1 геномный клон pNL43 получен от Др. Malcolm Martin, Программа НИЗ СПИД, исследования и ссылочные реагенты (ARRRR), отдел СПИД, NIAID, NIH. Клон может быть использован в качестве исходного продукта для ВИЧ-1 вирусных генов, предназначенных для генетических конструктов. Или же любой ВИЧ молекулярный клон инфицированной клетки может быть с помощью полимеразной цепной технологии модифицирован в достаточной для конструирования степени, например: НХВ2 клон, MN клон, а также SP или BAL-1 клон. Клон pNL43 является конструктом, состоящим из ВИЧ-1 провирусной ДНК плюс 3 к.о. хозяйской последовательности из участка интеграции клонированием в плазмиду pUC18.
Конструирование pNL-puro-env- плазмиды
Данная плазмида сконструирована для экспрессии gag/pol. StuI сайт в пределах не- ВИЧ 5'-фланкирующей человеческой ДНК из pNL43 разрушают частичным гидролизом в присутствии StuI с последующим гидролизом свободных концов в присутствии Е. coli полимеразы 1. Линейную плазмиду наполняют и затем самолигируют, оставляя в пределах ВИЧ генома единственный StuI сайт. Данную плазмиду (pNLDstu) затем гидролизуют в присутствии затупляющих ферментов StuI и BsaBI, удаляющих фрагмент кодирующей gp120 последовательности. В присутствии EcoRI и ClaI из pBABE-puro (Morgenstern, Land, 1990, Nucl.Acids Res. 18(12), 3587-3596, вводится здесь в качестве ссылки, плазмида предоставлена Др. Hartmut Land, Имперский раковый исследовательский центр) выделяют ОВ40 промотор и область кодирования устойчивости к пуромицину (пуромицин-ацетилтрансфераза (ПАТ)). Данный фрагмент затупляют и затем клонируют в StuI/BsaBI-гидролизованную pNLDstu. Отбирают клон с OB40-puro-фрагментом в необходимой ориентации с тем, чтобы 3'-ДКП из ВИЧ мог обеспечить поли-А функции для трансляции ПАТ. Полученная плазмида обозначена pNLpuro.
Стратегия клонирования для делеции vpr регуляторного гена из ВИЧ gag pol вектора
Область, находящуюся сразу же в восходящем направлении от единственного PflMI сайта до сразу же после vif терминирующего кодона, амплифицируют методом ПЦР с применением праймеров, которыми вводятся неконсервативная замена аминокислоты (glu ---> val) у аминокислоты 22 в vpr, терминирующий кодон в vpr рамке считывания после аминокислоты 22 и EcoRI сайт, расположенный сразу же после нового терминирующего кодона. Данный ПЦР фрагмент замещают PflMI-EcoRI фрагментом из pNLpuro или pNL43. Такое замещение приводит к делеции 122 нуклеотидов в открытой рамке считывания vpr с исключением тем самым возможности обращения, характерного для стратегии точетной мутации. Полученная плазмида (pNLpuro vpr) кодирует первые 21 природные аминокислоты из vpr плюс валин плюс все остальные оставшиеся ВИЧ-1 гены и сплайс-сочленения в их нативной форме. Подобная стратегия делеции применима также к nef, vif и vpu и допускает экспрессию структурного гена с одновременной защитой от образования живого рекомбинантного вируса.
Конструирование плазмиды, предназначенной для экспрессии оболочки
ДНК сегмент, кодирующий ген оболочки ВИЧ-1 НХВ2, клонируют амплификацией методом полимеразной цепной реакции (ПЦР) с применением лямда-клонированной ДНК, полученной от Программы СПИД, исследования и ссылочные реагенты. Последовательности 5'- и 3'-праймеров имеют следующий вид: 5'-AGGCGTCTCGAGACAGAGGAGAGCAAGAAATG-3' (ПОСЛЕД. N 1) с введением XhoI сайта и 5'-TTTCCCTCTAGATAAGCCATCCAATCACAC-3' (ПОСЛЕД. N 2) с введением XbaI сайта, направленных на gp160, tat и rev кодирующую область. Ген-специфичную амплификацию осуществляют применением Tag ДНК полимеразы согласно инструкциям изготовителя (Перкин-Эльмер Цетус Корп.). Продукт ПЦР обрабатывают тридцать минут при 37oC 0.5 мкг/мл протеиназы К с последующим экстрагированием смесью фенол-хлороформ и осаждением этанолом. Выделенную ДНК затем гидролизуют два часа при 37oC в присутствии XhoI и XbaI и подвергают электрофорезу на агарозном геле. Выделенный и очищенный XhoI-XbaI фрагмент клонируют в Bluescript плазмиду (Стратеген Инк., Ля Джолла, Ка) и затем субклонируют в эукариотный экспрессионный вектор pMAMneoVlue (Клонтех Лаборэториз, Инк., Рало Альто, КА). Полученный конструкт обозначен как рМ160 (фиг. 1А). Плазмидную ДНК очищают ультрацентрифугированием с градиентом CsCl. ДНК конструкт рМ160 кодирует ВИЧ-1/НХВ2 (Fisher A.G. и др., (1985) Nature 316, 262-265) связанный с мембраной гликопротеин gp160 под контролем ВСР усиливающего элемента и ВОМЖМ ЦДК в качестве промотора.
Конструирование альтернативной плазмиды экспрессии оболочки, названной ВИЧ-1 env-rev плазмидой
Область, кодирующую два экзона из rev и vpu, и оболочечные открытые рамки считывания из ВИЧ-1 НХВ2 амплифицируют методом ПЦР и клонируют в экспрессионный вектор pCNDA/neo (Инвитроген). Данная плазмида направляет продуцирование оболочки при участии ЦМВ промотора.
Получение и очистка
Плазмиду в E.coli (DH5 альфа) выращивают следующим образом. LB плюс ампициллин агаровые пластинки засевают штрихованием культурой целевой плазмиды из замороженной общей партии. Пластинки выдерживают при 37oC в течение ночи (14-15 часов). С пластинки отбирают единственную колонию, которой инокулируют 15 мл LB среды с препаратом пептона и 50 мкг/мл ампициллина. Полученную культуру выращивают при 37oC со встряхиванием (прим. 175 об/мин) в течение 8-10 часов. Показания ОП600 должны быть равны по меньшей мере 1. LB среду с пептоном и 50 мкг/мл ампициллина (1 л) инокулируют культурой со значением ОП = 1. Приготовленную культуру (1-2 литра) выращивают в течение ночи при 37oC со встряхиванием (175 об/мин).
Плазмиды, выращенные в Е. coli (штамм DH5 альфа) собирают и очищают по следующей методике. Общие методики лизиса клеток и очистки плазмид можно найти в издании: "Молекулярное клонирование. Лабораторное руководство", 2-ое издание, J. Sambrook, E.F. Fritsch, Т. Maniatis, Коулд Спринг Харбор Пресс, 1989. Клетки концентрируют и промывают глюкоза-трис-ЭДТК буфером (pH 8). Концентрированные клетки лизируют действием лизозима и кратковременно обрабатывают 0.2 н. КОН, последующим добавлением калийацетат-уксусной кислоты буфера устанавливают pH 5.5. Нерастворимые вещества удаляют центрифугированием. Для осаждения плазмиды к надосадочной жидкости добавляют 2-пропанол. Плазмиду вновь растворяют в трис-ЭДТК буфере и дополнительно очищают экстрагированием фенол-хлороформенной смесью и повторным осаждением 2-пропанолом.
Возможно удаление эндотоксина применением самых разнообразных способов, в том числе следующих: специфичная адсорбция иммобилизованными веществами, например полимиксином (Tani и др., Biomater. Aptif. Cells Inimobilization Biotechnol. 20(2-4), 457-62 (1992); Issekutz, J. Immunol. Methods 61(3), 275-81 (1983); применение антиэндотоксиновых моноклональных антител, например: 8А1 и НА-1АТМ (Центокор, Мальверн, ПА, Bogard и др., J. Immunol. 150(10), 4438-4449 (1993); Rietschel и др. Infect. Immunity, стр. 3863 (1993)); применение положительно заряженных глубинных фильтров (Hou и др., J. Parenter. Sci. Technol. 44(4), 204-9 (июль-август 1990)); применение поли(гамма-метил-L-глютамата), Hirayama и др., Chem. Pharm. Vull. (Токио) 40(8), 2106-9 (1992)); применение гистидина (Matsumae и др., Biotechnol. Appl. Biochem. 12(2), 129-40 (1990)); применение колонок и мембран гидрофобного взаимодействия (Aida и др., J. Immunol. Methods 132(2), 191-5 (1990); Umeda и др., Biomater. Artif. Cells Artif. Organs 18(4), 491-7 (1990); Hou и др. , Biochem. Biophys. Acta 1073 (1), 149-54 (1991); Sawada и др., J. Hyg. Лондон) 91(1), 103-14 (1986)); применение специфичных смол, пригодных для удаления эндотоксина, в том числе полистирол-дивинилбензольных или дивинилбензольных смол, например Браунли Полипор смолы (Эпплайд Биосистемс, Пало Альто, КА); XUS 40323 (Дау Кемикл, Мидлэнд, МИ); НР20, УНР20Р (Мицубиси Касей, США); Гамильтон PRP-1, PRP-сродство (Гамильтон, Рено, НВ); Джорди с обращенными фазами ДВБ, Джорди гель ДВБ, Полимер Лабс ПЛ-гельТМ (Олтех, Дирфилд, ИЛ); Видак PLxТМ (Сепарейшн Групп, Гесперия, КА); другие удаляющие эндотоксин материалы и методы включают; Детокси-ГельТМ, удаляющий эндотоксин гель (Пирс Кемикл, Рокфорд, ИЛ); Примечание 206 к применению (Фармация Биотех, Инк., Пискэтэвей, Н-Й). См. также в целом: Sharma, Biotech. Арр. Biochem. 8, 5-22 (1986). Рекомендуемые антиэндотоксиновые моноклональные антитела связываются с консервативными доменами эндотоксина и предпочтительно являются антителами к липиду А - наиболее структурно сохраняемой части молекулы эндотоксина. Такие антилипид А моноклональные антитела включают: моноклональные антитела 8А1 с высоким сродством к мышиному IgG и человечьи антилипид A IgM(k) моноклональные антитела НА-1АТМ. Источником НА-1АТМ служит человечья В Е. coli 15 вакцина. Как сообщается НА-1АТМ широко перекрестно реакционноспособно с самыми различными бактериальными эндотоксинами (липополисахаридами).
Пример 2
В опытах, предназначенных для сравнения иммуногенного ответа, возникающего после генетической вакцинации и вакцинации белками, созданы модели на животных с применением клеток опухоли, специфично экспрессирующих чужеродный долевой белок. Созданы три модели на иммунокомпетентных мышах, экспрессирующих чужеродные антигены. Применяют три линии клональных клеток опухоли, источником которых служат мыши линии Balb/c. Клеточные линии представлены: 1) лимфоидной клеточной линией, не дающей существенных метастазов в другие ткани, но образующей пальпируемую опухоль, которая, видимо, и убивает животное в пределах 8-12 недель; 2) клеточной линией мышиной меланомы с той же способностью образовывать метастазы, по большей части в легкое, и которая после инокуляции 1 миллионом клеток приводит к развитию у мыши больших пальпируемых опухолей, аналогичным образом убивающих животное в пределах 10-12 недель; и 3) клеточной линией мышиной легочной аденокарциномы, которая дает метастазы в множество тканей и убивает животное в течение 12 или более недель. Отбирают субклоны, в которых обнаруживаются чужеродные антигены в нераспознаваемом виде. После имплантации трансфектированной опухоли мыши родоначальной линии, в отличие от большинства
аналогичных линий мышиных опухолей, у животных не возникает защитной реакции на имеющиеся чужеродные антигены, и опухоль приживается. Эта опухоль впоследствии убивает животное с тем же фенотипом и в те же отрезки времени, что и первоначальная нетрансфецированная опухоль. Применением таких моделей можно определить иммунную реакцию, создаваемую генетической вакцинацией против антигена.
Сделано наблюдение, что мыши, вакцинированные генетическими вакцинами, включающими генетический конструкт, в результате продуцируют в своих клетках целевой белок, создающий иммунную реакцию, в том числе и сильную цитотоксичную реакцию, полностью уничтожающую опухоли, в которых обнаруживается целевой белок, но безо всякого влияния на опухоли без целевого белка. У мышей, инокулированных самим целевым белком, создаваемая в результате иммунная реакция была гораздо менее эффективной. Опухоль уменьшалась в размерах, но из-за отсутствия цитотоксичной реакции ее уничтожения не происходило. В качестве контроля в опытах по сравнению иммунной реакции животных, вакцинированных генетической вакциной, субъединичной вакциной, и невакцинированных животных применялись нетрансфецированные опухоли. В этих опытах четко показано, что генетическая вакцина создает более эффективную иммунную реакцию с более широким спектром действия, способную при участии ЦТЛ полностью уничтожить опухоль. И напротив, иммунизация применением интактного целевого белка создает более ограниченную, менее эффективную иммунную реакцию.
Пример 3
В другом воплощении изобретения создана генетическая вакцина против ВИЧ. Вирусный белок gp160, превращенный в gp120 и gp41, является целевым белком, против которого получена генетическая вакцина. Генетическая вакцина содержит ДНК конструкт, включающий ДНК последовательность, кодирующую gp160 операбельно связанные регуляторные элементы. При введении индивидууму ДНК конструкт генетической вакцины внедряется в клетки индивидуума, после чего продуцируется gp160. Иммунная реакция, создаваемая белком, имеет широкую основу и включает гуморальную и оба вида клеточной иммунной реакции. Широкая биологическая реакция обеспечивает более высокую степень защиты по сравнению с той, которая достигается при введении самого белка.
Мышам внутримышечно инъецируют рМ160 (см. Пример 1), после чего проводят анализ на анти-ВИЧ иммунную реакцию. В антисыворотке иммунизированных таким путем животных создается иммунная реакция на ВИЧ оболочечный белок, определяемая ферментативным иммуносорбентным анализом (ФИСА) и анализом иммуноосаждением. Антисыворотка нейтрализует ВИЧ-1 инфекцию и подавляет индуцируемое ВИЧ-1 образование синцитий.
Наблюдаемая нейтрализация и противосинцитиальная активность могут быть результатом реакционноспособности созданных антител к белку оболочки ВИЧ-1, например: V3 петле gp120, CD4 участку связывания и N-концевой "иммунодоминантной" области gp41 среди прочих.
В приведенной здесь методике генетической иммунизации в квадрицепсные мышцы мышей линии BALB/c инъецируют 100 мкл 0.5% бупивакаина-HCl и 0.1% метилпарабена в изотоническом растворе NaCl с применением иглы 27-го размера. Цель инъекции - стимулировать регенерацию мышечных клеток и облегчить поглощение генетического конструкта. Спустя двадцать четыре часа в те же места инъекции инъецируют либо 100 мкг рМ160, либо 100 мкг pMAMneoBlue в качестве контрольной плазмиды (фиг. 1А). Мышей ревакцинируют инъецированием того же количества ДНК конструкта трижды с интервалом в две недели указанным путем, но без предварительной обработки бупивакаин-HCl.
При иммунизации рекомбинантным gp160 мышей линии BALB/c первоначально иммунизируют 1 мкг гликозилированного рекомбинантного (ВИЧ-1/III8) gp160 (МикроГенСис Инк.) в полном адъюванте Фрейнда с последующим трехкратным ревакцинированием 1 мкг gp160 каждый раз в неполном адъюванте Фрейнда с интервалом в две недели. Продуцирование антител против ВИЧ-1 gp160 определяют анализом сыворотки мышей на способность иммуноосаждать gp160. Иммуноосаждение проводят использованием 1 • 106 cpm 125J-меченого rgp160 сыворотки мышей и агарозных шариков протеина-G (ГИБКО, Инк.) по ранее описанной методике Osther К. и др., Hybridoma 10, 673-683 (1991), которая вводится здесь в качестве ссылки. Результаты специфичного осаждения анализируют на 10% НДС-ПАГЭ. Полоса 1 соответствует 1 мкл сыворотки предсенсибилизированной мыши в реакции с 125J-gpl60. Полоса 2 соответствует 1 мкл сыворотки мыши, иммунизированной рМ160. Полоса 3 соответствует 1 мкл 1:100 разбавления моноклональных анти-gp120 антител ID6 (Ugen К.E. и др. (1992) "Создание моноклональных антител против аминообласти gp120, вызывающей зависимую от антител клеточную цитотоксичность", Коулд Спринг Харбор Лаборэтори) в качестве положительного контроля. Стрелкой указан специфично иммуноосажденный гликопротеин оболочки 125J-gpl60.
125J-меченый gpl60 специфично иммуноосаждается антисывороткой от рМ160-иммунизированных животных (фиг. 2, полоса 2), а также применяемыми в качестве положительного контроля анти-gp160 моноклональными антителами ID6 (фиг. 2, полоса 3). Напротив, сыворотка предсенсибизированных мышей (фиг. 2, полоса 1) показывает минимальную активность в том же анализе.
Восемь из десяти мышей, иммунизированных рМ160 конструктом, оказались положительными с точки зрения реакционности к gpl60 при определении методом ФИСА, и иммунная реакция животных с наиболее высоким анти-gp160 титром подвергнута подробному анализу. Все четыре мыши, иммунизированные контрольным вектором, показали одинаковую отрицательную реакционность к gp160 при определении методом ФИСА, и сыворотка одной из этих мышей использована в качестве контрольной в последующих опытах.
Показано, что нейтрализующие ВИЧ антитела специфично направлены на некоторые эпитопы gp120 и gp41, в том числе V3 петлю в gp120 (Goudsmit J. и др., (1988) AIDS 2, 157-164; Putney S.D. и др., (1989) "Создание ВИЧ субъединичной вакцины", Монреаль), CD4 участок связывания вблизи карбоксиокончания gp120 (Lasky L.A. и др. (1987) Cell 50, 975-985), a также иммунодоминантную петлю gp41 непосредственно в нисходящем направлении от N-концевой области слияния (Schrier R.D. и др. (1988) J. Virol. 62, 2531-2536).
Для определения того, будут ли анти-gp160 антитела, созданные у этих мышей, вступать в реакцию с указанными важными участками гликопротеинами оболочки, пептиды для BRU/V3 петли, пептиды для MN/V3 петли, пептиды для БХВ2/gp41 N-окончания или пептиды для HXB2/CD4 участка связывания адсорбируют на титрационных микропланшетах, после чего методом ФИСА определяют удельную реакционность мышиной антисыворотки. Один мкг/мл gp160 или 10 мкг/мл каждого пептида наносят в виде покрытия на титрационные микропланшеты в 0.1 М бикарбонатного буфера (pH 9.5), выдерживают в течение ночи при 4oC, блокируют 2% альбумина бычьей сыворотки в ФБС, проводят в течение одного часа при 37oC реакцию с козьим антимышиным IgG, конъюгированным с ПОХ (Фишер) и проявляют 10-30 минут при комнатной температуре в темноте ТМВ субстратом (Сигма). Полученные результаты приведены на фиг. 3. Показаны следующие антисыворотки: (-+-) соответствует сыворотке предсенсибилизированной мыши, (-х-) соответствует сыворотке мыши, иммунизированной pMAMneoBlue вектором, (-o-) соответствует сыворотке мыши, иммунизированной рМ160, (-Δ-) соответствует сыворотке мыши, иммунизированной белком rgp160. На фиг. 3А приведены результаты, полученные применением планшетов, покрытых rgp160 белком. На фиг. 3В показаны результаты, полученные применением планшетов, покрытых BRU/V3 петли пептидов (CNTRKR- IRIQRGPGRAFVTIGK (ПОСЛЕД. N 11)). На фиг. 3С показаны результаты, полученные применением планшетов, покрытых MN/V пептидами (YNKRKRIHIQRGPGRAFYTTKNIIC (ПОСЛЕД. N 12)) с QR последовательностью из ВИЧ-1/III8 подчеркнутого типа. На фиг. 3D приведены результаты, полученные применением планшетов, покрытых HXB2/CD4 пептидов участка связывания (CRIKQFINMWQEVGKAMYAPPISGIRC (ПОСЛЕД. N 13)). На фиг. 3Е приведены результаты, полученные применением планшетов, покрытых пептидами BRU/gp41 иммунодоминантной области (RILAVERYTKDQQLLGIWGCSGKLIC (ПОСЛЕД. N 14)).
Из фиг. 3 видно, что антисыворотка от мыши, иммунизированной рМ160 конструктом, показывает значительно более высокую реакционоспособность к пептидам BRU и MN/V3 петли, пептиду CD4 участка связывания и иммунодоминантному пептиду gp41, чем сыворотка при иммунизации рекомбинантным gp160 белком (rgp160). Антисыворотка от иммунизированной rgp160 мыши имела гораздо более высокий титр относительно rgp160 по сравнению с антисывороткой при иммунизации рМ160, но более низкую активность относительно трех специфичных нейтрализующих эпитопов gp160, подвергнутых анализу.
Для определения того, будет антисыворотка, созданная при иммунизации ДНК, обладать антивирусной активностью, исследована способность антисыворотки нейтрализовать инфекцию ВИЧ-1. Бесконечный ВИЧ-1/III8 вирус при 100 TCID50 инкубируют с последовательными разбавлениями антисыворотки перед использованием для инфицирования МТ-2 целевых клеток (Montefiori D.C. (1988) J. Clin. Microbiol. 26, 231-235).
Сто (100) TCID50 ВИЧ/III8 бесклеточного вируса выдерживают один час при 37oC с последовательными разбавлениями антисыворотки. После выдерживания подвергнутые предварительной обработке вирусы наносят на пластинки с целевой МТ-2 клеточной линией (4 • 106), выдерживают один час при 37oC, после инфицирования МТ-2 клетки промывают трижды и затем инкубируют при 37oC и 5% СО2. Спустя три дня определяют количественно слияние путем визуального подсчета числа синцитий на лунку в опытах с тремя повторами под фазовоконтрастным микроскопом.
Полученные результаты показаны на фиг. 4. На фиг. 4А показаны результаты, полученные использованием сыворотки от иммунизированной вектором мыши в сравнении с сывороткой при иммунизации рМ160. Графики зависимости величин нейтрализации (Vn/Vo) от фактора разбавления (Nara P. (1989) "Методы исследования ВИЧ", ред. Aldovini A., Walkter B.D., 77-86, M. Стоктон Пресс) показаны на фиг. 4С. Контрольная сыворотка (-х-) получена от иммунизированной pMAMneoBlue вектором мыши. Испытуемая сыворотка (-о-) получена от иммунизированных рМ160 мышей.
Подавление синцитий осуществляют по методике Osther К. и др. (1991) Hybridoma 10, 673-683. Клеточную линию Н9/III8 предварительно инкубируют тридцать минут при 37oC и 5% СО2 с последовательными разбавлениями (1:100, 1:200 и 1:400) антисыворотки, приготовленными в планшетах на 96 лунок в общем объеме 50 мкл. Спустя три дня определяют количественно слияние путем визуального подсчета числа синцитий на лунку под фазовоконтрастным микроскопом. Фиг. 4D иллюстрирует совместное культивирование целевых клеток с ВИЧ-1/III8 клеточной линией, обработанной сывороткой от предиммунизации. Фиг. 4Е иллюстрирует то же самое, что и фиг. 4D, но обработка ведется сывороткой от контрольной иммунизации вектором. Фиг. 4F иллюстрирует то же самое, что и фиг. 4D, но обработка ведется сывороткой от иммунизации rgp160. Фиг. 4G иллюстрирует то же самое, что и фиг. 4D, но обработка ведется сывороткой от иммунизации рМ160. Фиг. 4D-4G показывают, что в данном анализе подавление синцитий становится очевидным при разбавлении 1:200. МТ-2 клетки инфицируют бесклеточным ВИЧ-1/III8 вирусом, предварительно инкубированным синцитиями, легко образующимися в антисыворотке от иммунизации вектором (фиг. 4А).
Для сравнения: предварительное инкубирование с сывороткой от иммунизированной рМ160 мыши предотвращает образование синцитий (фиг. 4В). Кинетику нейтрализации определяют построением графика зависимости Vn/Vo от последовательных разбавлений (Nara P. (1989) "Методы исследования ВИЧ", ред. Aldovini А., Walker B.D., 77-86, М. Стоктон Пресс) (фиг. 4С). Сыворотка от рМ160 иммунизированной мыши проявляет биологическую нейтрализующую активность при разбавлениях вплоть до 1:320, в то время как контрольная антисыворотка не показывает аналогичной активности.
Для определения того, будет ли антисыворотка от рМ160 иммунизированной мыши способна подавлять передаваемое через оболочку распространение вируса путем слияния непосредственно клетка с клеткой, проведены опыты с подавлением синцитий. Антисыворотка от рМ160 иммунизированной мыши подавляют ВИЧ-1 индуцируемое образование синцитий при разбавлении 1:200 (фиг. 4G). И напротив, предиммунная сыворотка, антисыворотка от rgp160 иммунизированной мыши (фиг. 4F) и антисыворотка от иммунизированных контрольных вектором животных (фиг. 4Е) оказались не способны подавлять образование синцитий при том же разбавлении.
Наблюдения, сделанные в опытах по нейтрализации (фиг. 4А-С) и подавлению образования синцитий (фиг. 4D-G) для указанных сывороток, коррелируются с наблюдениями реакционности, сделанные в ФИСА (фиг. 3). Антисыворотка от рМ160 иммунизированной мыши, показавшая высокий уровень связывания с нейтрализующими эпитопами, проявила также и высокий уровень антивирусной активности. И напротив, с низким уровнем связывания с теми же эпитопами, в том числе антисыворотка от rgp160 иммунизированной мыши, обладает низкой антивирусной активностью.
Для выявления того, будет ли антисыворотка от рМ160 иммунизированной мыши способна подавлять связывание gp120 с СD4- содержащими Т-клетками, проведен анализ прямого подавления с применением флюроцитометрии (Chen Y.H. и др. (1992) AIDS 6, 533-539). Сделано наблюдение о том, что сыворотка от иммунизированный рМ160 конструктом мыши способна блокировать связывание gp120 с СD4-содержащими Т-клетки. Иммунная сыворотка в разбавлении 1:15 подавляет связывание FITC-gp120 с CD4+ SupT1 клетками на 22±2% при определении проточной цитометрией в дублированных опытах. Это указывает на то, что указанная область проникновения ВИЧ в целевые клетки может быть также функционально подавлена данной антисывороткой. Полученные данные коррелируются с наблюдаемой в ФИСА реакционностью антисыворотки к пептидам CD4 участка связывания (фиг. 3C).
Применением продажного набора мышиных моноклональных антител для определения изотипов (Сигма) проведены исследования изотипа иммуноглобулина. Из анти-gp160 специфичных антител, создаваемых рМ160 иммунизацией, 19% относятся к IgG1, 51% - в IgG2, 16% - к IgG3, 10% - к IgM и 5% относятся к IgA.
Преобладание IgC изотипов указывает на протекание вторичной иммунной реакции, что еще больше усиливает предположение о возможности возникновения при генетической вакцинации хелперных Т-клеток.
Титрационные микропланшеты покрывают рМ160 и PMAMneoBlue ДНК и методом ФИСА с применением сыворотки всех иммунизированных животных определяют специфичное связывание. Никакого значительного связывания с плазмидной ДНК не отмечено. Таким образом, маловероятно, что применением генетического материала для инокуляции мышечной ткани можно создать реакцию против плазмидной ДНК.
Введение ДНК конструкта в мышцы мыши инъекций с помощью иглы может привести к непредсказуемым результатам, поскольку такая методика не обеспечивает средств регулирования поглощения ДНК клетками мышц. Проведено сравнение инициирования одного только ДНК конструкта (n ≈ 4) с животными (n ≈ 4), предварительно обработанных бупивакаином. Наблюдаемые в двух группах иммунные реакции не совпадают, и методом ФИСА для этих групп получены величины соответственно в 25% и 75% для респондирующих животных. Повышенная эффективность может быть достигнута применением системы прямой доставки ДНК, например, бомбардировкой частицами (Klein Т.М. и др. (1992) Biotechnology 10, 286-291).
Факты нейтрализации, подавления синцитий, подавления CD4-gp120 связывания и специфичного связывания с некоторыми важными областями в gp160 показывают, что введение ДНК конструкта, кодирующего ВИЧ gp160 связанный с мембраной гликопротеин, непосредственно в мышцы животного вызывает специфичные гуморальные реакции и ведет к образованию биологически уместных антивирусных антител.
Для выявления того, будет ли вакцина способна создавать защитную иммунную реакцию, использована вышеописанная модель на животных. Клетки опухоли трансфецируют ДНК, кодирующей gp160, затем подтверждают экспрессию белка и имплантируют животному. Контроль включает нетрансфецированные клеточные линии.
Генетически иммунизированных животных вакцинируют плазмидой рМ160. Контроль включает невакцинированных животных, животных, вакцинированных только векторной ДНК, и животных, которым введен gp160 белок.
Полученные результаты показывают, что иммунная реакция вакцинированной мыши достаточна для полного удаления трансфецированных опухолей без оказания какого-либо воздействия на нетрансфецированные опухоли. Вакцинация gp160 белком приводит к некоторому уменьшению размера трансфецированных опухолей по сравнению с нетрансфецированными опухолями, но не оказывает никакого влияния на смертность. Невакцинированные животные показали одинаковую смертность как для трансфецированных, так и нетрансфецированных опухолей.
Пример 4
Ниже приводится перечень конструктов, которые могут быть использованы в способах настоящего изобретения. Впервые сообщение о создании вектора pBabe. puro, применяемого в качестве исходного вещества при создании многих из нижеперечисленных конструктов, появилось в работе Morgenstenn J.P., Land H. (1990) Nucl. Acids Res. 18(12), 3587-3596, которая вводится здесь в качестве ссылки. Плазмида pBabe. puro особенно приемлема для экспрессии экзогенных генов в клетках млекопитающих. Подлежащие экспрессии ДНК последовательности вставляют в участки клонирования под контролем повтора длинного конца (ПДК) вируса Молони мышиного лейкоза (Мо MuLV) промотора. Плазмида содержит отбираемый маркер для устойчивости к пуромицину.
Пример 5
Плазмида pBa.Vα3 является плазмидой в 7.8 к.о., содержащей 2.7 к. о. EcoRI геномного фрагмента, кодирующего Vα3 область Т-клеточного рецептора, содержащую L, V и J сегменты, клонированные в EcoRI сайте плазмиды pBabe.puro. Происходящий из Т-клеточного рецептора целевой белок применим для вакцинации против и лечении передаваемого через Т-клетки аутоиммунного заболевания, а также клонотипичной Т-клеточной лимфомы и лейкоза.
Пример 6
Плазмида pBa. gagpol-vpr является плазмидой в 9.88 к.о., содержащей gag/pol и vlf гены из ВИЧ/MN, клонированные в pBabe.puro. Ген vpr изъят. Плазмида, содержащая указанные вирусные гены, кодирующие ВИЧ целевые белки, применима для иммунизации от ВИЧ инфекции и СПИД и для их лечения. ВИЧ ДНК последовательность опубликована в работе: Reiz M.S. (1992) AIDS Res. Human Retro. B, 1549, которая вводится здесь в качестве ссылки. Последовательность может быть получена в Генбанке под N М17449, который указан здесь в виде ссылки.
Пример 7
Плазмида рМ160 является плазмидой в 11 к.о., содержащей 2.3 к. о. ПЦР фрагмента, кодирующего ВИЧ-1/3В оболочечный белок и rev/tat гены, клонированные в pMAMneoBlue. Область nef изъята. Плазмида, содержащая указанные вирусные гены, кодирующие целевые белки, применима для вакцинации от ВИЧ инфекции и СПИД и для их лечения. ДНК последовательность ВИЧ-1/3В опубликована в работе; Fisher А. (1995) Nature 316, 262, которая вводится здесь в качестве ссылки. Последовательность может быть получена в Генбанке под N К03455, который указан здесь в виде ссылки.
Пример 8
Плазмида pBa.VL является плазмидой в 5.4 к.о., содержащей ПЦР фрагмент, кодирующий VL область анти-ДНК антитела, клонированную в pBabe.puro в XbaI и EcoRI сайтах. Происходящий из антитела целевой белок служит примером целевого белка, применимого для вакцинации от передаваемого через В-клетки аутоиммунного заболевания и клонотипичной В-клеточной лимфомы и лейкоза. Общую методику клонирования функциональных переменных областей из антител можно найти в работе: Chaudhary V.К. и др. (1990) Proc. Natl. Acad. Sci. США 87, 1066, которая вводится здесь в качестве ссылки.
Пример 9
Плазмида pOspa.B является плазмидой в 6.84 к.о., содержащей области кодирования, кодирующие OspA и OspB антигены Borrelia burgdorferi - спирохеты, ответственной за болезнь Лима. Антигены клонированы BamHI и SalI сайтах плазмиды pBabe. puro. ПЦР праймеры, применяемые для создания OspA и OspB фрагментов, имеют следующие последовательности: 5'-GAAGGATCCATGAAAAAATATTTATTGGG-3' (ПОСЛЕД. N 3) и 5'-ACTGTCGACTTATTTTAAAGCGTTTTTAAG-3' (ПОСЛЕД. N 4). Cм.: Williams W.V. и др. (1992) DNA and Cell. Viol. 11(3), 207, которая вводится здесь в качестве ссылки. Плазмида, содержащая указанные патогенные гены, кодирующие целевые белки, применима для вакцинации от болезни Лима.
Пример 10
Плазмида pBa. RB-G является плазмидой в 7.1 к.о., содержащей созданный методом ПЦР фрагмент, кодирующей белок С бешенства, клонированный в BamHI сайт плазмиды pBabe.puro. Содержащая этот патогенный ген плазмида, кодирующая белок С бешенства, применима для вакцинации от бешенства. ДНК последовательность находится в Генбанке под N М32751, который указан здесь в виде ссылки. См. также: Aniliopis А. и др. (1981) Nature 294, 275, которая вводится здесь в качестве ссылки.
Пример 11
Плазмида pBa.HPV-L1 является плазмидой в 6.8 к.о., содержащей созданный методом ПЦР фрагмент, кодирующий L1 капсидный белок вируса папилломы человека (ВПЧ), в том числе его штаммов 16, 18, 31 и 33. Фрагмент клонирован в BamHI и EcoRI сайтах плазмиды pBabe.puro. ДНК последовательность помещена в Генбанк под N М15781, который указан здесь в виде ссылки. См. также: Howley P. (1990) "Разделы вирусологии", том 2, ред. Channock R.M. и др., глава 58, 1625 и Shah К., Howley P., "Разделы вирусологии" (1990), том 2, ред. Channock R.M. и др., глава 59, каждое из которых вводится здесь в качестве ссылки. Плазмида применима для вакцинации от ВПЧ инфекции и в терапии раковых заболеваний.
Пример 12
Плазмида pBa.HRV-L2 является плазмидой в 6.8 к. о., содержащей созданный методом ПЦР фрагмент, кодирующий L2 капсидный белок вируса папилломы человека (ВПЧ), в том числе штаммов 16, 18, 31 и 33 ВПЧ. Фрагмент клонирован в BamHI и EcoRI сайтах плазмиды pBabe.puro. Плазмида применима для вакцинации от ВПЧ инфекции и в терапии раковых заболеваний. ДНК последовательность помещена в Генбанк под N М15781, который указан здесь в виде ссылки. См. также издания: Howley P. (1990) "Разделы вирусологии", том. 2, ред. Channock и др., глава 58, 1625; Shah К., P. Howley (1990) "Разделы вирусологии", том 2, ред. Channock и др., глава 59, каждое из которых вводится здесь в качестве ссылки.
Пример 13
Плазмида pBa. MNp7 является плазмидой в 5.24 к.о., содержащей созданный методом ПЦР фрагмент, кодирующий область кодирования р7, включающую ВИЧ MN gag (белок ядра) последовательность, клонированную в BamHI сайте плазмиды pBabe. puro. Содержащая указанные ВИЧ вирусные гены, кодирующие ВИЧ целевые белки, плазмида применима для вакцинации от ВИЧ инфекции и СПИД и для их лечения. См. работу: Reiz M.S. (1992) AIDS Res. Human Retro. 8, 1549, которая вводится здесь в качестве ссылки. Последовательность может быть получена в Генбанке под N М17449, который указан здесь в виде ссылки.
Пример 14
Плазмида pGA733-2 является плазмидой в 6.3 к.о., содержащей поверхностный антиген GA-733-2 опухоли, клонированный из клеточной линии SW948 колоректальной карциномы в pCUM8 вектор (Seed В., A. Aruffo (1987) Proc. Natl. Acad. Sci. США 84, 3365, вводится здесь в качестве ссылки) в BstXI сайте. Ассоциируемый с опухолью целевой белок является примером целевого белка, пригодного для вакцинации от и лечения гиперпролиферативного заболевания, например рака. GA733-2 антиген является целевым антигеном против рака ободочной кишки. Сообщения об GA733 антигене приводятся в работе: Szala S. и др. (1990) Proc. Natl. Acad. Sci. США 87, 3542- 3546, которая вводится здесь в качестве ссылки.
Пример 15
Плазмида pT4-pMV7 является плазмидой в 11.15 к.о., содержащей кДНК, кодирующую CD4 рецептор человека, клонированный в pMV7 вектор по EcoRI сайту. CD4 целевой белок применим для вакцинации от Т-клеточной лимфомы и ее лечения. Плазмида pT4-pMV7 может быть получена в СПИД Хранилище, каталожный номер 158.
Пример 16
Плазмида pDJCA733 является плазмидой в 5.1 к.о., содержащей GA733 поверхностный антиген опухоли, клонированный в pBabe.puro по BamHI сайту. Ассоциируемый с опухолью целевой белок является примером целевого белка, применимого для вакцинации от и для лечения гиперпролиферативного заболевания, например рака. GA733 антиген является целевым антигеном против рака ободочной кишки.
Пример 17
Плазмида pBa.RAS является плазмидой в 6.8 к. о., содержащей ras область кодирования, которую вначале субклонируют из pZIPneoRAS и клонируют в pBabe. puro по BamHI сайту. Целевой белок ras служит примером цитоплазмический сигнальной молекулы. Способ клонирования ras приводится в работе: Weinberg (1984) Mol. Cell. Biol. 4, 1577, которая вводится здесь в качестве ссылки. Кодирующая ras плазмида применима для вакцинации от и для лечения гиперпролиферативного заболевания, например рака, в частности, рака мочевого пузыря, мышц, легких, мозга и костей.
Пример 18
Плазмида pBa.MNp55 является плазмидой в 6.38 к.о., содержащей созданный методом ПЦР фрагмент, кодирующий область кодирования р55, включающую последовательность ВИЧ MN gag предшественника (белок ядра), клонированную в pBabe. puro по BamHI сайту. Плазмида, содержащая указанные вирусные гены ВИЧ, кодирующие ВИЧ целевые белки, применима для вакцинации от ВИЧ инфекции и СПИД и для их лечения. См. работу: Reiz M.S. (1992) AIDS Res. Human Retro. 8, которая вводится здесь в качестве ссылки. Последовательность может быть получена в Генбанке под N М17449, который указан здесь в виде ссылки.
Пример 19
Плазмида pBa.MNp24 является плазмидой в 5.78 к.о., содержащей созданный методом ПЦР фрагмент из pMN-SFl матрицы, кодирующей область кодирования р24, включающей полную ВИЧ MN gag область кодирования, клонированную в pBabe.puro по BamHI и EcoRI сайтам. Плазмида, содержащая указанные ВИЧ вирусные гены, кодирующие ВИЧ целевые белки, применима для вакцинации от ВИЧ инфекции и СПИД и для их лечения. См. работу: Reiz M.S. (1992) AIDS Res. Human Retro. 8, 1549, которая вводится здесь в качестве ссылки. Последовательность может быть получена в Генбанке под N М17449, который указан здесь в виде ссылки.
Пример 20
Плазмида pBa.MNpl7 является плазмидой в 5.5 к. о., содержащей созданный методом ПЦР фрагмент, кодирующий область кодирования р17, включающую ВИЧ MN gag (белок ядра) последовательность, клонированную в pBabe.puro по BamHI и EcoRI сайтам. Плазмида, содержащая указанные ВИЧ вирусные гены, кодирующие ВИЧ целевые белки, применима для вакцинации от ВИЧ инфекции и СПИД и для их лечения. См. работу: Reiz M.S. (1992) AIDS Res. Human Retro. 8, 1549, которая вводится здесь в качестве ссылки. Последовательность может быть получена в Генбанке под N М17449, который указан здесь в виде ссылки.
Пример 21
Плазмида pBa.SIVenv является плазмидой в 7.8 к. о., содержащей созданный методом ПЦР фрагмент в 2.71 к.о., амплифицированный из конструкта, содержащего SIV 239 в рВР322 и клонированный в pBabe.puro по BamHI и EcoRI сайтам. Применяют следующие праймеры: 5'-GCCAGTTTTGGATCCTTAAAAAAGGCTTGG-3' (ПОСЛЕД. N 5) и 5'-TTGTGAGGGACAGAATTCCAATCAGGG-3' (ПОСЛЕД. N 6). Последовательность может быть получена от Программы исследования СПИД и ссылочных реагентов, каталожный N 210.
Пример 22
Плазмида pcTSP/ATK.env является плазмидой в 8.92 к.о., содержащей созданный методом ПЦР фрагмент, кодирующий полную область кодирования HTLV оболочки из HTLV-1/TSP и /АТК изолятов, субклонированных pcDNA1/neo вектор. Применяют следующие праймеры: 5'-CAGTGATATCCCGGGAGACTCCTC-3' (ПОСЛЕД. N 7) и 5'- GAATAGAAGAACTCCTCTAGAATTC-3' (ПОСЛЕД. N 8). Сообщения о плазмиде pcTSP/ATK. env даются в работе: Proc. Natl. Acad. Sci. США 85, 3599 (1988), которая вводится здесь в качестве ссылки, HTLV env целевой белок применим для вакцинации от HTLV и Т-клеточной лимфомы и для их лечения.
Пример 23
Плазмида pBa.MNgpl60 является плазмидой в 7.9 к.о., содержащей созданный методом ПЦР фрагмент в 2.8 к. о., амплифицированный из содержащего MNenv в pSR72 конструкта и клонированного в pBabe.puro по BamHI и EcoRI сайтам. Применяют следующие праймеры: 5'-GCCTTAGGCGGATCCTATGGCAGGAAG-3' (ПОСЛЕД. N 9) и 5'-TAAGATGGGTGGCCATGGTGAAT-3' (ПОСЛЕД. N 10). См. работу: Reiz M.S. (1992) AIDS Res. Himan Retro. 8, 1459, которая вводится здесь в качестве ссылки. Последовательность может быть получена в Генбанке под N М17449, который указан здесь в виде ссылки. Плазмида, содержащая указанные ВИЧ вирусные гены, кодирующие ВИЧ целевые белки, применима для вакцинации от инфекции ВИЧ и СПИД и для их лечения.
Пример 24
Плазмида pC. MNp55 является плазмидой в 11.8 к.о., содержащей созданный методом ПЦР фрагмент в 1.4 к.о., амплифицированный из gag области MN изолята и клонированный в рСЕР4 вектор. Плазмида, содержащая указанные ВИЧ вирусные гены, кодирующие ВИЧ целевые белки, применимы для вакцинации от ВИЧ инфекции и СПИД и для их лечения. См. работу: Reiz M.S. (1992) AIDS Res. Human Retro. 8, 1549, которая вводится здесь в качестве ссылки. Последовательность может быть получена в Генбанке под N М17449, который указан здесь в виде ссылки.
Пример 25
Плазмида pC.Neu является плазмидой в 14.2 к.о., содержащей ДНК фрагмент в 3.8 к. о., в состав которого входит область кодирования neu онкогена человека, вырезанного из LTR-2/erbB-2 конструкта и субклонированного в рСЕР4 вектор. Сообщение о pC.Neu плазмиде приводится в работе: DiFiore (1987) Science 237, 178, которая вводится здесь в качестве ссылки. Целевой белок - peu онкоген служит примером рецептора фактора роста, применимого в качестве целевого белка для вакцинации от и для лечения гиперпролиферативного заболевания, например рака, в частности рака ободочной кишки, молочной железы, легкого и мозга.
Пример 26
Плазмида pC.RAS является плазмидой в 11.7 к.о., содержащей ДНК фрагмент в 1.4 к. о. , включающей область кодирования gas онкогена, которую сначала субклонируют из pZIPneoRAS и субклонируют в рСЕР4 по BamHI сайту. Сообщение о pC. RAS плазмиде приводится в работе: Weinberg (1984) Mol. Cell. Biol. 4, 1577, которая вводится здесь в качестве ссылки. Целевой белок ras служит примером цитоплазмической сигнальной молекулы. Кодирующая ras плазмида применима для вакцинации от и для лечения гиперпролиферативного заболевания, например рака, в частности рака мочевого пузыря, мышц, легкого, мозга и костей.
Пример 27
Плазмида pNLpuro является плазмидой в 15 к.о., содержащей gag/pol и SV40-puro вставки. Плазмида, содержащая указанные ВИЧ вирусные гены, кодирующие ВИЧ целевые белки, применима для вакцинации от ВИЧ инфекции и СПИД и для их лечения.
Пример 28
Для проверки эффективности генетической вакцины против человеческого CD4 на мыши создан ДНК конструкт. Данные опыты были созданы для выявления способности вакцины защищать от антигена Т-клеточной лимфомы. При Т-клеточной лимфоме CD4 является специфичным для опухоли антигеном. Соответственно, данная модель показывает способность генетической вакцины защищать от Т-клеточной лимфомы. Кроме того, в данных опытах выявлялась эффективность против одного из членов сверхсемейства иммуноглобулиновых молекул. CD4 характеризуется высокой консервацией между человеческим и мышиным видами.
Применяемая модель на животных приведена выше. Клетки опухоли трансфецируют ДНК, кодирующей CD4, подтверждают экспрессию клетками белка, после чего их имплантируют животному. Контроль включает нетрансфецированные клеточные линии. Хотя животные и стали иммунокомпетентными, иммунная реакция не была направлена против имплантированной, CD4-меченой опухоли у невакцинированных животных.
Генетически иммунизированных животных вакцинируют плазмидой pT4-pMV7, описанной в Примере 15. Контроль включает невакцинированных животных и животных, которым вводился CD4 белок.
В описываемой здесь процедуре генетической иммунизации в квадрицепсные мышцы мышей линии BALB/c инъецируют 100 мкл 0.5% бупивакаина-HCl и 0.1% метилпарабена в изотоническом растворе NaCl применением иглы номер 27 с целью стимулирования регенерации мышечных клеток и облегчения поглощения генетического конструкта. Спустя двадцать четыре часа в тот же участок инъецирования инъецируют либо 100 мкг pT4-pMV7, либо 100 мкг pMV7 в качестве контрольной плазмиды. Мышей ревакцинируют инъецированием того количества ДНК трижды с интервалом в две недели и тем же способом, но без предварительного введения бупивакаина-HCl.
Животным вводят 1000000 СD4-меченых клеток опухоли. У невакцинированных животных образуются большие опухоли и смерть наступает в пределах 7-10 недель. У вакцинированных животных аналогичные смертельные опухоли не развивались.
Полученные результаты показывают, что иммунная реакция у генетически вакцинированных мышей достаточна для полного удаления трансфецированных опухолей, и в то же время не влияет на нетрансфецированные опухоли. Вакцинация CD4 белком приводит к некоторому уменьшению размера опухоли в случае трансфецированных опухолей по сравнению с нетрансфецированными опухолями, но на смертность не влияет. Невакцинированные животные показали одинаковую смертность как в случае трансфецированных, так и нетрансфецированных опухолей.
Пример 29
Для выявления эффективности генетической вакцины против GA733 человека на мышах создан ДНК конструкт. Данные опыты были проведены для определения способности вакцины защищать от ассоциируемого с GA733 раком, например раком ободочной кишки. Применяемая модель на животных приведена выше. Клетки опухоли трансфецируют ДНК, кодирующей GA733, подтверждают способность клеток экспрессировать белок, после чего их имплантируют животному. Контроль включает нетрансфецированные клеточные линии.
Вышеприведенным способом иммунизированных животных вакцинируют плазмидой pGA733-2, описанной в Примере 14. Контроль включает невакцинированных животных и животных, которым вводился GA733 белок.
Полученные результаты показывают, что иммунная реакция генетически вакцинированных мышей достаточна для полного удаления трансфецированных, и в то же время никак не влияет на нетрансфецировавные опухоли. Вакцинация GA733 белок приводит к некоторому уменьшению размера трансфецированной опухоли по сравнению с нетрансфецированной опухолью, но не влияет на смертность. Невакцинированные животные показали одинаковую смертность как в случае трансфецированных, так и нетрансфецированных опухолей.
Пример 30
Для выявления эффективности генетической вакцины против человечьего p185neu создан ДНК конструкт. Данные опыты были проведены для определения способности вакцины защищать от ассоциируемого с р185 neu рака, например рака молочной железы, легких и мозга. Применяемая модель на животных приведена выше. Клетки опухоли трансфецируют ДНК, кодирующей neu, подтверждают экспрессию белка клетками, после чего имплантируют животному. Контроль включает линии нетрансфецированных опухолей.
Генетически иммунизированных животных вакцинируют плазмидой pLTR-2/egbB-2, т. е. плазмидой в 14.3 к.о., содержащей область кодирования человечьего neu онкогена, клонированную по вышеприведенной методике в LTP-2 вектор по XhoI сайту. 5'-ДКП и 3'-ДКП являются ДКП из Молони-MuLV. Контроль включает невакцинированных животных и животных, которым вводился pl85neu белок.
Полученные результаты показывают, что иммунная реакция генетически вакцинированных мышей достаточна для полного удаления трансфецированных опухолей, и в то же время никак не влияет на нетрансфецированные опухоли. Вакцинация р185 белком приводит к некоторому уменьшению размера опухоли в случае трансфецированной опухоли по сравнению с нетрансфецированной опухолью, но не влияет на смертность. Невакцинированные животные показали одинаковую смертность как в случае трансфецированыых, так и нетрансфецированных опухолей.
Пример 31
Для выявления эффективности генетической вакцины против человечьего Ras на мышах создан ДНК конструкт. Данные опыты предназначались для определения способности вакцины защищать от ассоциируемого в Ras рака, например рака мочевого пузыря, мышц, легких, мозга и костей. Применяемая модель на животных приведена выше. Клетки опухоли трансфецируют ДНК, кодирующей Ras, подтверждают экспрессию клетками белка, после чего их имплантируют животному. Контроль включает нетрансфецированные линии опухоли.
Вышеприведенным способом вакцинации генетически вакцинированных животных вакцинируют плазмидой pBa. RAS, описанной в примере 17. Целевой белок ras является примером цитоплазмической сигнальной молекулы. Способ клонирования ras приводится в работе: Weinberg (1984) Mol. Cell. Biol. 4, 1577, которая вводится здесь в качестве ссылки. Контроль включает невакцивированных животных и животных, которым вводился Ras белок.
Пример 32
Для выявления эффективности генетической вакцины на мышах против антигена G-белка вируса бешенства человека создан ДНК конструкт. Применяемая модель на животных приведена выше. Клетки опухоля трансфецируют ДНК, кодирующий белок G вируса бешенства, подтверждают экспрессию белка клетками, после чего их имплантируют животному.
Генетически иммунизированных животных вакцинируют плазмидой рВа.РЬ-G, описанной в Примере 10, применением вышеприведенного способа вакцинации. Целевой G-белок вируса бешенства служит примером патогенного антигена. ДНК последовательность находится в Генбанке под N 32751. Контроль включает невакцинированных животных и животных, которым вводился G-белок вируса бешенства.
Пример 33
Для выявления эффективности генетической вакцины на мышах против антигена болезни Лима создан ДНК конструкт. Применяемая модель на животных приведена выше. Клетки опухоли трансфецируют ДНК, кодирующей OspA и OspB, подтверждают экспрессию белка клетками, после чего их имплантируют животному.
Генетически иммунизированных животных вакцинируют плазмидой pOspA.B, описанной в Примере 9. Контроль включает невакцинированных животных и животных, которым вводились белки OspA и OspB.
Пример 34
Для выявления на мышах эффективности генетической вакцины против переменной области Т-клеточного рецептора человека создан ДНК конструкт. Данные опыты предназначались для определения способности вакцины защищать от происходящего из Т-клеточного рецептора белка, ассоциируемого с раком, например: Т-клеточной лимфомой и передаваемой через Т-клетки аутоиммунной болезнью. Применяемая модель на животных приведена выше. Клетки опухоли трансфецируют ДНК, кодирующей Ras, подтверждают экспрессию белка клетками, после чего их имплантируют животному.
Генетически иммунизированных животных вакцинируют плазмидой рВа.Vα3, описанной в Примере 5, применением вышеприведенного способа вакцинации.
Пример 35
Плазмида рМ160 может быть использована в качестве исходного продукта для получения ряда плазмид, применимых для экспрессии одного или нескольких генов из env части ВИЧ. Описание
конструирования рМ160 приведено выше. Плазмида направлена на gp160, tat и rev область кодирования. Ген nef отсутствует.
Промотором, контролирующим экспрессию gp160/rev/tat гена, является ВОМЖМ ДКП. Промотор может быть изъят и заменен актиновым промотором, миозиновым промотором, ВИЧ ДКП промотором и ЦМВ промотором.
Ген, придающий устойчивость к ампициллину, может быть изъят или каким-то иным путем дезактивирован. Ген, придающий устойчивость к неомицину, может быть помещен под контроль бактериального промотора.
Из плазмиды может быть удален усилитель вируса саркомы Рауса. ВСР усилитель может быть заменен усилителем мышечного креатина.
Гены gp160/rev/tat перекрываются и разделяют одинаковые нуклеотидные последовательности в разных рамках считывания. Ген rev может быть изъят путем замены его инициирующего кодона другим кодоном. Аналогичным путем может быть удален и tat ген. В каждой плазмиде за исключением тех, в которых для контроля gp160/rev/tat применяется ВИЧ ДКП, может быть удален один из rev или tat, или оба rev и tat. В плазмидах, использующих ВИЧ ДКП промотор, присутствие tat обязательно.
В Таблице 3 дается перечень рМ160-модифицированных плазмид. В каждой удален ВСР усилитель. В некоторых из них усилитель отсутствует (нет), в других присутствует усилитель мышечного креатина (УМК). В некоторых из них присутствует ВИЧ rev ген (да), а в других этот ген отсутствует (нет). Некоторые из них имеют ВИЧ tat ген (да), в то время как в других этот ген отсутствует (нет).
Конструкты RA-29 - RA-56 идентичны соответственно конструктам RA-1 - RА-32 за исключением того, что в каждом случае промотор, контролирующий ген неомицина, является бактериальным промотором.
Пример 36
Плазмида pNLpuro может быть использована в качестве исходного продукта в создании нескольких различных плазмид, экспрессирующих ВИЧ gag/pol гены. Как указано выше pNLpuro создана для экспрессии gag/pol. Описанная выше плазмида pNL- puroΔvpr сконструирована с изъятием vpr регуляторного гена из ВИЧ gag pol вектора с целью исключить присутствие необходимого регуляторного белка из ряда генов, вводимых при вакцинации. Помимо изъятия vpr обычным специалистам в данной области в плазмиду pNLpuro могут быть внесены и другие изменения применением стандартных методик молекулярной биологии и широкодоступных исходных продуктов.
Человечьи фланкирующие 5' и 3' последовательности из ВИЧ последовательностей могут быть удалены несколькими способами. К примеру, методом ПЦР могут быть амплифицированы и реконструированы только ВИЧ, OB40-puro и pUC18 последовательности.
Из основанных на pNLpuro плазмид может быть изъята psi область, имеющая важное значение для упаковки вируса. Для удаления psi области pNLpuro плазмиду обрезают в присутствии SacI и SpeI. В результате гидролиза удаляется psi область, а также 5'-ДКП, находящийся в нисходящем направлении, и часть gag/pol области в нисходящем направлении от psi. Для повторной вставки не относящихся к psi последовательностей методом ДКП осуществляют амплификацию с регенерацией этих последовательностей. Созданы праймеры, позволяющие регенерировать часть ВИЧ последовательности 5' и 3' до psi без регенерации psi. Праймеры реформируют SacI сайт в 5'-части из 5'-ДКП плазмиды. Праймеры идут в восходящем направлении от SacI сайта до участка, находящегося непосредственно у 3'-части 5'-конца psi области с образованием у 3'-конца AatI сайта. Праймеры, начинающиеся непосредственно в 5'-части psi области, кроме того, создают AatI сайт, а начинающиеся в 3'-части SpeI сайта, регенерируют этот сайт. Созданные методом ПЦР фрагменты гидролизуют в присутствии SacI, AatI и SpeI и лигируют совместно с гидролизованным в присутствии SacI/SpeI pNLpuro-psi фрагмента. ВИЧ 5'-ДКП промотор может быть удален в заменен промотором вируса Молони, ВОМЖМ ДКП, актиновым промотором, миозиновым промотором и ЦМВ промотором.
ВИЧ 3'-ДКП участок полиаденилирования может быть удален и заменен ОВ40 участком полиаденилирования.
Ген, придающий устойчивость к ампициллину, может быть удален или дезактивирован каким-то иным путем.
В Таблице 4 приводится перечень основанных на pNLpuro конструктов, в которых изъяты ВИЧ psi и vpr области и, кроме того, удалены фланкирующие 5'- и 3'-области ВИЧ последовательности человека.
Конструкты от LA-17 до LA-32 идентичны соответственно конструктам от LA-1 до LA-16, за исключением того, что в каждом случае остается по меньшей мере одна из человечьих фланкирующих последовательностей.
Пример 37
При создании других конструктов для экспрессии env гена эта область ВИЧ может быть вставлена в коммерчески доступную плазмиду рСЕР4 (Инвитроген). рСЕР4 плазмида особенно приемлема, поскольку содержит ориджин репликации вируса Эпштейна-Барра и область кодирования ядерного антигена EBNA-1, создающих высококопийную репликацию без интеграции. рСEР4 содержит также гигромициновый маркер под регуляторным контролем тимидинкиназного промотора и участок полиаденилирования. Область кодирования ВИЧ env помещают под регуляторный контроль промотора ЦМВ и участка полиаденилирования ОВ40. Область кодирования ВИЧ env в виде ПЦР фрагмента в 2.3 к.о. из ВИЧ/3В, последовательность в Генбанке под N К03455. Полученную на основе рСЕР4 плазмиду (pRA-100) внедряют вне хромосомы, и плазмида продуцирует gp160 белок.
Пример 38
Для получения других конструктов для экспрессии env гена эта область ВИЧ может быть вставлена в коммерчески доступную плазмиду pREP4 (Инвитроген). pREP4 плазмида особенно приемлема, поскольку содержит ориджин репликации вируса Эпштейна-Барра и область кодирования ядерного антигена EBNA-1, обеспечивающих высококопийную репликацию без интеграции. pREP4 содержит также гигромициновый маркер под регуляторным контролем тимидинкиназного промотора и участка полиаденилирования. Область кодирования ВИЧ env помещают под регуляторный контроль ВСР промотора и ОВ40 участка полиаденилирования. Область кодирования ВИЧ env получена в виде ПЦР фрагмента в 2.3 к.о. ВИЧ/3В формы, последовательность в Генбанке N К03455. Полученную на основе рRЕР4 плазмиду (pRA-101) внедряют внехромосомно, и плазмида продуцирует gp160 белок.
Пример 39
Для создания других конструктов, предназначенных для экспрессии gag/pol генов, эта область ВИЧ может быть вставлена в продажную плазмиду pСЕР4 (Инвитроген). рСЕР4 плазмида особенно приемлема, поскольку содержит ориджин репликации вируса Эпштейна-Барра и область кодирования ядерного антигена EBNA-1, обеспечивающий высококопийную эписомальную репликацию без интеграции. рСЕР4 содержит также гигромициновый маркер под регуляторным контролем тимидинкиназного промотора и участка полиаденилирования. Область кодирования ВИЧ gag/pol помещают под регуляторный контроль ЦМВ промотора и ОВ40 участка полиаденилирования Область кодирования ВИЧ gag/pol получена из ВИЧ MN (последовательность в Генбанке N М17449), и включает vif ген. Ген vpr не включен. Полученная на основе рСЕР4 плазмида (pLA-100), внедренная внехромосомно, продуцирует белки GAG55, обратной транскриптазы, протеазы и интегразы.
Пример 40
Для создания других конструктов, предназначенных для экспрессии gag/pol, эта область ВИЧ может быть вставлена в коммерческую плазмиду pREP4 (Инвитроген). pREP4 плазмида особенно приемлема, поскольку содержит ориджин репликации вируса Эпштейна-Барра и область кодирования ядерного антигена EBNA-1, обеспечивающих высококопийную эписомальную репликацию без интеграции. pREP4 содержит также гигромициновый маркер под регуляторным контролем тимидинкиназного маркера и участка полиаденилирования. Область кодирования ВИЧ gag/pol помещают под регуляторный контроль ЦMB промотора и ОВ40 участка полиаденилирования. Область кодирования ВИЧ gag/pol получена из ВИЧ MN (последовательность в Генбанке N М17449), и включает vif ген. Ген vpr не включен. Полученную на основе pREP4 плазмиду (pLA-101) внедряют внехромосомно, и плазмида продуцирует белки GAG55, обратной транскриптазы, протеазы и интегразы.
Пример 41
Следующий конструкт, называемый здесь pGACPOL.rev, применим для экспрессии ВИЧ gag/pol генов.
Плазмида включает ген устойчивости к канамицину и pBR322 ориджин ДНК репликации. Необходимые для регуляции транскрипции последовательности включают: цитомегаловирусный промотор, усилитель вируса саркомы Рауса и ОВ40 сигнал полиаденилирования. Содержащиеся в pGAGPOL.gev ВИЧ-1 последовательности включают: последовательность, кодирующую р17, р24 и р15 из gag открытой рамки считывания, последовательность, кодирующую протеазу, последовательность, кодирующую обратную транскриптазу и содержащую небольшую делецию, и последовательность, кодирующую неактивное аминоокончание интегразы из роl открытой рамки считывания, а также кодирующую rev последовательность. Каждая ВИЧ последовательность происходит из штамма НХВ2 ВИЧ-1.
В pGAGPOL.rev предусмотрено несколько уровней безопасности. Они включают применение ЦМВ промотора и неретровирусный поли (А) участок. Более того, делеция ψ последовательности ограничивает возможности упаковки вирусной РНК. Кроме того, многочисленные мутации в обратной транскриптазе приводят к форментативно неактивному продукту. Более того, большая делеция в интегразе приводит к неактивному продукту, а для стабилизации бактериальных трансформантов применяют маркер устойчивости к канамицину.
Плазмиду pGAGROL.rev конструируют следующим образом.
Стадия 1. Применяют субклон части ВИЧ-1 (НХВ2) генома, клонированный в Bluescript (Стратаген). Субклон ВИЧ-1 содержит полный 5'-ДКП и остальную часть ВИЧ-1 генома до нуклеотида 5795 (нумерация Генбанка). ВИЧ-1 последовательности получены из HXB2D (СПИД хранилище).
Стадия 2. Подвергают ПЦР часть gag из открытой рамки считывания HXB2D плазмиды (СПИД хранилище). ПЦР фрагмент обрезают в присутствии NotI и SpeI и лигируют с вышеописанным ВИЧ-1 субклоном, рестриктированным в присутствии NotI и SpeI.
Стадия 3. Подвергают ПЦР комплекс gag/pol и часть pol-кодирующих последовательностей из HXB2D плазмиды (СПИД хранилище) с праймерами ПОСЛЕД. N 15 и ПОСЛЕД. N 16. Обрезают ПЦР продукт в присутствии ClaI и лигируют друг с другом. Обрезают лигированные фрагменты в присутствии BclI и SalI и лигируют с плазмидой со стадии 2, гидролизованной в присутствии BclI и SalI.
Стадия 4. Обрезают плазмиду со стадии 3 в присутствии BspMI и EcoRI и вновь лигируют с адаптерами, образованными отжигом линкеров (ПОСЛЕД. N 17 и ПОСЛЕД. N 18).
Стадия 5. Плазмиду со стадии 4 обрезают в присутствии NotI и SalI и лигируют либо с плазмидой 4a или 4b из описания, приведенного для pENV (см. ниже). Обрезают также в присутствии NotI и SatI.
Стадия 6. Плазмиду со стадии 5 рестриктируют в присутствии SalI и MluI и лигируют с ПЦР продуктом, полученным ПЦР rev с праймерами ПОСЛЕД. N 19 и ПОСЛЕД. N 20.
Стадия 7. Плазмиду со стадии 6 обрезают в присутствии NotI и лигируют с продуктом, полученным методом ПЦР rev-ответственного элемента в HXB2D плазмиде (СПИД хранилище) с праймерами ПОСЛЕД. N 21 и ПОСЛЕД. N 22.
Стадии 6 и 7 необязательны.
Пример 42
Следующий конструкт, называемый здесь pENV, применим для экспрессии ВИЧ env генов.
Плазмида включает ген устойчивости к канамицину и pBR322 ориджин ДНК репликации. Необходимые для регуляции транскрипции последовательности включают: цитомегаловирусный промотор, усилитель вируса саркомы Рауса и ОВ40 сигнал полиаденилирования. Содержащиеся в pENV ВИЧ-1 последовательности включают последовательность, кодирующую vpu, последовательность, кодирующую rev, последовательность, кодирующую gp160, последовательность, кодирующую 50% nef, последовательность, кодирующую vif, и последовательность, кодирующую vpr с делецией 13 аминокислот карбокси-конца. Последовательности vpu, rev, gp160 и nef происходят ив ВИЧ-1 штамма MN. Источником vif и vpr последовательностей является ВИЧ-1 штамм НХВ2.
В pGAGROL.rev предусмотрено несколько уровней безопасности. Они включают применение ЦМВ промотора и неретровирусно поли (А) участка. Более того, удален tat, а 50-ая делеция nef приводит к "неактивному" nef продукта. Кроме того, vif и vpr выведены из нормальной последовательности, а частичная делеция vpr дополнительно гарантирует неактивный vpr продукт.
Плазмида pENV создана следующим образом.
Стадия 1. Исходную плазмиду pUC18 гидролизуют в присутствии HindIII и EcoRI. Полученный фрагмент, содержащий ColEl ориджин репликации и laci ген, необходимо лигировать с EcoRI/HindIII фрагментом из pMAMneoBlue, содержащий наш усилитель вируса саркомы. Полученная плазмида или pMAMneoBlue от фирмы Клонтех (Пало Альто, КА) может быть затем гидролизована в присутствии HindIII и BglI. С помощью стандартных методик лигируют с фрагментом, содержащим kan ген, полученным методом ПЦР из генблочной плазмиды (Фармация).
Стадия 2. Если в качестве исходной плазмиды применяют pMAMneoBlue, ее гидролизуют в присутствии MluI и EcoRI, наполняют концы фрагментом Кленова из полимеразы I и повторно лигируют.
Стадия 3. Затем плазмиду, полученную из pMAMneoBlue либо из pUC18, гидролизуют в присутствии HindIII и лигируют с OB40 поли (А) участком и областью раннего сплайсинга, полученную методом ПЦР из рCEP4 (Инвитроген, Сан-Диего, КА) с праймерами ПОСЛЕД N 23 и ПОСЛЕД. N 24.
Стадия 4a. Гидролизуют в присутствии BamHI и лигируют с ЦМВ промотором, полученным методом ПЦР из рСЕР4 (Инвитроген, Сан-Диего, КА) с праймерами ПОСЛЕД. N 25 и ПОСЛЕД. N 26.
Стадия 4b. Гидролизуют в присутствии BamHI и лигируют с MoMLV ДКП, полученным методом ПЦР с праймерами ПОСЛЕД. N 27 и ПОСЛЕД. N 28.
Стадия 5. Гидролизуют в присутствии NotI и MluI и лигируют с областью кодирования gp160, полученной из pMN-ST1 методом ПЦР с праймерами ПОСЛЕД. N 29 и ПОСЛЕД. N 30.
Стадия 6. Гидролизуют в присутствии MluI и лигируют с последовательностями, кодирующими vif во всей его полноте и vpr с делецией 13 а. к. у карбокси-конца, получены из HX2BD плазмиды (СПИД хранилище) методом ПЦР с праймерами ПОСЛЕД. N 31 и ПОСЛЕД. N 32.
Пример 43
Дается система вакцинации, включающая:
фармацевтический препарат, содержащий около 100 мкг pGAGPOL.rev в изотоническом фармацевтически приемлемом растворе и фармацевтический препарат, содержащий 100 мкг pENV в изотоническом фармацевтически приемлемом растворе. Кроме того, система вакцинации предпочтительно включает фармацевтический препарат, содержащий 1 мл 0.5% бупивакаин-HCl и 0.1% метилпарабена в изотоническом фармацевтическом носителе.
В случае применения такой рекомендуемой системы вакцинации вначале проводят первую серию введений, в которой бупивакаин и один из двух фармацевтических препаратов вводят индивидууму внутримышечно, предпочтительно в мышцы руки и ягодицы. Бупивакаин и второй из двух фармацевтических препаратов вводят индивидууму внутримышечно в другом месте, предпочтительно удаленном от места введения первого фармацевтического препарата, предпочтительно в другую руку или ягодицу. Последовательность серии введений может быть разделена во времени, предпочтительно от 48 часов до двух или более недель между двумя введениями.
Система вакцинации может быть использована для вакцинации индивидуума с целью его защиты от ВИЧ инфекции или для лечения ВИЧ-инфицированного индивидуума иммунотерапевтическим средством.
Пример 44
В некоторых своих воплощениях изобретение относится к способу вакцинации индивидуума от ВИЧ введением единственного инокулята. Такой инокулят включает генетический конструкт, содержащий по меньшей мере один, предпочтительно два, более предпочтительно более двух или ряд генов ВИЧ вируса или все структурные гены. Однако инокулят не содержит полный набор всех ВИЧ генов. При попадании в единичную клетку полного набора вирусных генов возникает вероятность сборки в пределах клетки целого инфекционного вируса. Соответственно, в генетическом конструкте настоящего изобретения отсутствует такой полный набор генов. В целях безопасности один или несколько существенных генов может быть удалено или преднамеренно изменено с дополнительной гарантией исключения образования инфекционной вирусной частицы.
В некоторых воплощениях настоящего изобретения дается по меньшей мере части одного, двух или всех ВИЧ структурных генов. Структурные гены ВИЧ включают; gag, pol и env. В генетическом конструкте даются части по меньшей мере одного из этих генов. Соответственно, в некоторых вариантах в генетическом конструкте даются по меньшей мере части каждого из gag и pol; в отдельных вариантах в генетическом конструкте дается по меньшей мере часть env; в других вариантах в генетическом конструкте дается по меньшей мере часть gag; в отдельных вариантах в генетическом конструкте дается по меньшей мере часть каждого из pol и env; в отдельных вариантах в генетическом конструкте дается по меньшей мере часть каждого из gag и env; в других вариантах в генетическом конструкте дается по меньшей мере часть pol. Возможно присутствие гена целиком. В любом из указанных конструктов возможно также присутствие ВИЧ регуляторных генов. ВИЧ регуляторные гены включают: vpr, vif, nef, vpu, tat и rev.
Пример 45
В применяемом здесь значении термин "экспрессионная единица" относится к последовательности нуклеиновой кислоты, содержащей промотор, операбельно связанный с кодирующей последовательностью, в свою очередь операбельно связанной с сигналом полиаденилирования. Кодирующая последовательность может кодировать один или несколько белков или их фрагментов. В рекомендуемых вариантах экспрессионная единица находится в пределах плазмиды.
В применяемом здесь значении термин "ВИЧ экспрессионная единица" относится к последовательности нуклеиновой кислоты, содержащей промотор, операбельно связанный с кодирующей последовательностью, в свою очередь операбельно связанной с сигналом полиаденилирования, причем кодирующая последовательность кодирует пептид, имеющий эпитоп, который идентичен или по существу аналогичен эпитопу, обнаруживаемому в ВИЧ белке. Выражение "по существу аналогичный эпитоп" относится к эпитопу, строение которого не идентично строению эпитопа ВИЧ белка, но который, тем не менее, создает клеточную или гуморальную иммунную реакцию, перекрестно реакционную к ВИЧ белку. В рекомендуемых вариантах ВИЧ экспрессионная единица содержит последовательность, кодирующую один или несколько ВИЧ белков или их фрагменты. В рекомендуемых вариантах ВИЧ экспрессионная единица находится в пределах плазмиды.
В некоторых воплощениях настоящего изобретения дается единичный генетический конструкт, содержащий единственную ВИЧ экспрессионную единицу, включающую ДНК последовательности, кодирующие один или несколько ВИЧ белков или их фрагменты. В применяемом здесь значении термин "конструкт с единственной ВИЧ экспрессионной единицей" относится к единичному генетическому конструкту, содержащему единственную ВИЧ экспрессионную единицу. В рекомендуемых вариантах конструкт с единственной ВИЧ экспрессионной единицей имеет вид плазмиды.
В некоторых воплощениях настоящего изобретения дается единичный генетический конструкт, включающий более одной ВИЧ экспрессионной единицы, каждая из которых содержит ДНК последовательности, кодирующие один или несколько ВИЧ белков или их фрагменты. В применяемом здесь значении термин "генетический конструкт с рядом ВИЧ экспрессионных единиц" относится к единичной плазмиде, содержащей более одной ВИЧ экспрессионной единицы. В рекомендуемых вариантах конструкт с рядом ВИЧ экспрессионных единиц имеет вид плазмиды.
В некоторых воплощениях настоящего изобретения дается единичный генетический конструкт, включающий две ВИЧ экспрессионных единицы, каждая из которых содержит ДНК последовательности, кодирующие один или более ВИЧ белков или их фрагменты. В применяемом здесь значении термин "генетический конструкт с двумя ВИЧ экспрессионными единицами" относится к единичной плазмиде, содержащей две ВИЧ экспрессионные единицы, т.е. к генетическому конструкту с рядом ВИЧ экспрессионных единиц, содержащему две ВИЧ экспрессионных единицы. В случае генетического конструкта с двумя ВИЧ экспрессионными единицами рекомендуется, чтобы одна из ВИЧ экспрессионных единиц функционировала в направлении, противоположном функционированию другой ВИЧ экспрессионной единицы. В рекомендуемых вариантах генетический конструкт с двумя ВИЧ экспрессионными единицами имеет вид плазмиды.
В некоторых воплощениях настоящего изобретения дается ВИЧ генетическая вакцина, содержащая единственный генетический конструкт. Единственный генетический конструкт может представлять собой генетический конструкт с единственной ВИЧ экспрессионной единицей, двумя ВИЧ экспрессионными единицами или генетический конструкт с рядом ВИЧ экспрессионных единиц, в котором содержится более двух ВИЧ экспрессионных единиц.
В некоторых воплощениях настоящего изобретения дается ВИЧ генетическая вакцина, содержащая более одного генетического конструкта в единственном инокуляте.
В некоторых воплощениях настоящего изобретения дается ВИЧ генетическая вакцина, содержащая более одного генетического конструкта в более чем одном инокуляте. В применяемом здесь значении термин "множественный инокулят" относится к генетической вакцине, включающей более одного генетического конструкта, каждый из которых вводят отдельно. В некоторых воплощениях настоящего изобретения дается ВИЧ генетическая вакцина, содержащая два генетических конструкта. Каждый генетический конструкт может представлять собой независимо от другого генетический конструкт с единственной ВИЧ экспрессиониой единицей, генетический конструкт с двумя ВИЧ экспрессионными единицами или генетический конструкт с рядом ВИЧ экспрессионных единиц, в котором содержится более двух ВИЧ экспрессионных единиц. В некоторых вариантах оба генетических конструкта представляют собой генетический конструкт с единственной ВИЧ экспрессионной единицей. В отдельных вариантах оба генетических конструкта представлены генетическими конструктами с двумя ВИЧ экспрессионными единицами. В других вариантах оба генетических конструкта представлены генетическими конструктами с рядом ВИЧ экспрессионных единиц. В некоторых вариантах один генетический конструкт является генетическим конструктом с единственной ВИЧ экспрессионной единицей, а другой - генетическим конструктом с двумя ВИЧ экспрессионными единицами. Для обычного специалиста совершенно очевидны многие вариации, зависящие от числа генетических конструктов, применяемых в генетической вакцине, и числа ВИЧ экспрессионных единиц, которые могут присутствовать в каждом генетическом конструкте.
Рекомендуется, чтобы генетические конструкты настоящего изобретения не содержали определенных ВИЧ последовательностей, в частности, тех, которые принимают участие в интеграции ВИЧ генома в хромосомное вещество клетки, в которую вводится конструкт. Рекомендуется, чтобы генетические конструкты настоящего изобретения не содержали ПДК из ВИЧ. Рекомендуется также, чтобы генетические конструкты настоящего изобретения не содержали psi сайта ВИЧ. Кроме того, рекомендуется, чтобы были подвергнуты делеции ген обратной транскриптазы и ген интегразы. Делеции включают делецию только некоторых кодонов или замену отдельных кодонов с целью существенного изменения гена. К примеру, инициирующий кодон может быть делетирован, изменен или смещен за рамку с образованием нуклеотидной последовательности, кодирующей неполный или нефункциональный продукт.
Рекомендуется также, чтобы генетические конструкты настоящего изобретения не содержали транскрибируемый tat из ВИЧ. Ген tat, перекрывающий rev ген, может быть полностью изъят путем замены кодонов, кодирующих rev, другими кодонами, кодирующими ту же аминокислоту для rev, но не кодирующими необходимую tat аминокислоту в рамке считывания, в которой кодируется tat. Или же отключаются только некоторые кодоны путем замены, а по существу делеции инициирующего кодона для tat и/или путем замены, а по существу делеции достаточных кодонов с образованием в результате нуклеотидной последовательности, кодирующей неполный или нефункциональный tat.
Рекомендуется, чтобы генетический конструкт включал кодирующие последовательности, кодирующие пептиды, имеющие по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопу из ВИЧ gag, pol, env или rev белков. Еще более предпочтительно, если генетический конструкт включает кодирующие последовательности, которые кодируют по меньшей мере один из ВИЧ gag, pol, env или rev белков или их фрагменты. Рекомендуется, чтобы генетический конструкт включал кодирующие последовательности, кодирующие пептиды, содержащие более одного эпитопа, идентичного или по существу аналогичного эпитопу из ВИЧ gag, pol, env или rev белков. Более предпочтительно, если генетический конструкт включает последовательности, кодирующие более одного ВИЧ gag, pol, env или rev белков или их фрагменты.
В некоторых вариантах генетический конструкт включает последовательности, кодирующие пептиды, имеющие по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопу из ВИЧ vif, vpr, vpu или nef белков. В отдельных вариантах генетический конструкт включает последовательности, кодирующие по меньшей мере один из ВИЧ vif, vpr, vpu или nef белков.
Генетический конструкт с единственной ВИЧ экспрессионной единицей может включать области кодирования одного или нескольких пептидов, разделяющих по меньшей мере один эпитоп с ВИЧ белком или его фрагментом в единственной экспрессионной единице под регуляторным контролем единственного промотора и сигнала полиаденилирования. Рекомендуется, чтобы генетические конструкты кодировали более одного ВИЧ белка или его фрагменты. Промотором может быть любой, функциональный в клетке человека промотор. В качестве промотора рекомендуется ОВ40 промотор или ЦМВ промотор, предпочтительно ЦМВ непосредственно ранний промотор. Сигналом полиаденилирования может служить любой функциональный в клетке человека сигнал полиаденилирования. В качестве сигнала полиаденилирования рекомендуется ОВ40 сигнал полиаденилирования, предпочтительно ОВ40 малый сигнал полиаденилирования. Если в единственной экспрессионной единице имеются более одной области кодирования, эти области могут непосредственно примыкать друг к другу или могут быть разделены некодирующими областями. Для правильной экспрессии область кодирования должна иметь инициирующий кодон и терминирующий кодон.
Генетический конструкт с двумя ВИЧ экспрессионными единицами может включать области кодирования одного или нескольких пептидов, разделяющих по меньшей мере один эпитоп с ВИЧ белком или его фрагментом в каждой из двух экспрессионных единиц. Каждая экспрессионная единица находится под регуляторным контролем единственного промотора и сигнала полиаденилирования. В некоторых вариантах рекомендуется, чтобы генетические конструкты кодировали более одного ВИЧ белка или его фрагмента. В некоторых вариантах рекомендуется, чтобы в одной экспрессионной единице присутствовали нуклеотидные последовательности, кодирующие gag и pol, а в другой присутствовали нуклеотидные последовательности, кодирующие env и rev. Промотор может быть представлен любым функциональным в клетке человека промотором. В качестве промотора рекомендуется ОВ40 промотор или ЦМВ промотор, предпочтительно непосредственно ранний ЦМВ промотор. Сигнал полиаденилирования может быть представлен любым функциональным в клетке человека сигналом полиаденилирования. В качестве сигнала полиаденилирования рекомендуется ОВ40 сигнал полиаденилирования, предпочтительно ОВ40 малый сигнал полиаденилирования. Если в экспрессионной единице имеется более одной области кодирования, эти области могут непосредственно примыкать друг к другу или могут быть разделены некодирующими областями. Для правильной экспрессии области кодирования должны иметь инициирующий кодон и терминирующий кодон.
Согласно некоторым воплощениям настоящего изобретения МНС Класса II перекрестно реакционный эпитоп в env изъят и заменен аналогичной областью из ВИЧ II.
Если генетический конструкт содержит gag и/или pol, как правило, важно, чтобы в этом случае присутствовал и rev. Для повышенной экспрессии gag и pol вместе с этими генами помимо rev может быть предусмотрен и rev-респондирующий элемент.
При получении генетических конструктов рекомендуется, чтобы применяемый в плазмиде 1 env ген происходил из MN или MN-подобных изолятов, включая клинические MN-подобные изоляты, предпочтительно несинцитиально индуцируемые клинические изоляты, предпочтительно те клинические изоляты, которые являются макрофаговыми тропиками из клинических изолятов ранней стадии.
Альтернативным сплайсингом из единственной экспрессионной единицы может быть получен целый ряд белков. Обеспечением сигналов сплайсинга можно достигнуть альтернативного сплайсинга, в котором создаются различные матрицы, кодирующие разные белки.
Пример 46
На фиг. 8 показаны четыре скелета: A, B, C и D. На фиг. 9 показаны вставки: 1, 2, 3 и 4. Вставка 1 поддерживает экспрессию gag и pol; rev-респондирующий элемент клонирован путем, при котором сохраняется ВИЧ сплайс-акцептор. Вставка 2 аналогична вставке 1 в том, что также поддерживает экспрессию gag и pol, но при этом rev-респондирующий элемент клонируют без сохранения ВИЧ сплайс-акцептора. Вставка 3 поддерживает экспрессию gag и pol, включает делецию гена интегразы и не включает цис-действующего rev-респондирующего элемента. Вставка 4 поддерживает экспрессию rev, vpu и env. Ген env может иметь МНС Класса II перекрестно реакционный эпитоп, измененный с удалением перекрестной реакционности; петля V3 может быть изменена с целью исключения возможности образования синцитий.
В некоторых вариантах скелет А используют со вставкой 1. Подобные конструкты возможно содержат ОВ40 ориджин репликации. Плазмида pAlori+ представляет собой скелет А со вставкой 1 и ОВ40 ориджином репликации. Плазмида pAlori- представляет собой скелет А со вставкой 1 и без ОВ40 ориджина-репликации. Кроме того, как pAlori+, так и pAlori- могут включать интегразу, что приводит к плазмидам pAlori+int+ и pAlori-int+ соответственно. Плазмиды pAlori+, pAlori-, pAlori+int+ и pAlori-int+ могут быть дополнительно модифицированы функциональной делецией гена обратной транскриптазы (RT) с образованием соответственно pAlori+RT-, pAlori-RT-, pAlori+int+RT-и pAlori-int+RT-.
В некоторых вариантах скелет А применяют со вставкой 2. Подобные конструкты возможно содержат ОВ40 ориджин репликации. Плазмида pA2ori+ состоит из скелета А со вставкой 2 и ОВ40 ориджина репликации. Плазмида pA2ori- состоит из скелета А со вставкой 2, но без ОВ40 ориджина репликации. Кроме того, как pA2ori+, так и pA2ori- могут включать интегразу, что приводит к плазмидам pA2ori+int+ и pA2ori-int+ соответственно. Плазмиды pA2ori+, pA2ori-, pA2ori+int+ и pA2ori-int+ могут быть дополнительно модифицированы функциональной делецией гена обратной транскриптазы (RT) с образованием соответственно pA2ori+RT-, pA2ori-RT-, pA2ori+int+RT- и pA2ori-int+RT-.
В некоторых вариантах скелет В применяют со вставкой 1. Подобные конструкты возможно содержат ОВ40 ориджин репликации. Плазмида pBlori+ состоит из скелета В со вставкой 1 и ОВ40 ориджином репликации. Плазмида pBlori- состоит из скелета В со вставкой 1, но без ОВ40 ориджина репликации. Кроме того, как плазмида pBlori+, так и pBlori- могут включать интегразу, что приводит к плазмидам pBlori+int+ и pBlori-int+ соответственно. Плазмиды pBlori+, pBlori-, pBlori+int+ и pBlori-int+ могут быть дополнительно модифицированы функциональной делецией гена обратной транскриптазы (RT) с образованием соответственно pBlori+RT-, pBlori-RT-, pBlori+int+RT- и pBlori-int+RT-.
В некоторых вариантах скелет В применяют со вставкой 2. Такие конструкты возможно содержат ОВ40 ориджин репликации Плазмида pB2ori+ состоит из скелета В со вставкой 2 и ОВ40 ориджина репликации. Плазмида pB2ori- состоит из скелета В со вставкой 2, но без ОВ40 ориджина репликации. Кроме того, как pB2ori+, так и pB2ori- могут включать интегразу, что приводит к плазмидам pB2ori+int+ и pB2ori-int+ соответственно. Плазмиды pB2ori+, pB2ori-, pB2ori+int+ и pB2ori-int+ могут быть дополнительно модифицированы с функциональной делецией гена обратной транскриптазы (RT) с образованием соответственно pB2ori+RT-, pB2ori-RT-, pB2ori+int+RT- и pB2ori-int+RT-.
В некоторых вариантах скелет А минус rev применяют со вставкой 3. Такие конструкты возможно содержат ОВ40 ориджин репликации. Плазмида pA/r-3ori+ имеет скелет А со вставкой 3 и ОВ40 ориджином репликации. Плазмида pA/r-3ori- имеет скелет А минус rev со вставкой 3 и без ОВ40 ориджина репликации. Кроме того, как pA/r-3ori+, так и pA/r-3ori- могут включать интегразу, что приводит к плазмидам pA/r-3ori+int+ и pA/r-3ori-int+ соответственно. Плазмиды pA/r-3ori+, pA/r-3ori-, pA/r-3ori+int+ и pA/r-3ori-int+ могут быть дополнительно модифицированы функциональной делецией гена обратной транскрип-тазы (RT) с образованием соответственно pA/r-3ori+RT-, pA/r-3ori-RT-, pA/r-3ori+int+RT- и pA/r-3ori-int+RT-.
В некоторых вариантах скелет С применяют со вставкой 1. Такие конструкты возможно содержат ОВ40 ориджин репликации. Плазмида pClori+ имеет скелет С со вставкой 1 и ОВ40 ориджином репликации. Плазмида pClori- имеет скелет С со вставкой 1, но без ОВ40 ориджина репликации. Кроме того, как pClori+ так и pClori- могут включать интегразу, что приводит к плазмидам pClori+int+ и pClori-int+ соответственно. Плазмиды pClori+, pClori-, pClori+int+ и pClori-int+ могут быть дополнительно модифицированы функциональной делецией гена обратной транскриптазы (RT) с образованием соответственно pClori+RT-, pClori-RT-, pClori+int+RT- и pClori-int+RT-.
В некоторых вариантах скелет С применяют со вставкой 2. Такие конструкты возможно содержат ОВ40 ориджин репликации. Плазмида pC2ori+ имеет скелет С со вставкой 2 и ОВ40 ориджином репликации. Плазмида pC2ori- имеет скелет С со вставкой 2, но без ОВ40 ориджнна репликации. Кроме того, как pC2ori+, так и pC2ori- могут включать интегразу, что приводит к плазмидам pC2ori+int+ и pC2ori-int+ соответственно. Плазмиды pC2ori+, pC2ori-, pC2ori+int+ и pC2ori-int+ могут быть дополнительно модифицированы функциональной делецией гена обратной транскриптазы (RT) с образованием соответственно pC2ori+RT-, pC2ori-RT-, pC2ori+int+RT- и pC2ori-int+RT-.
В некоторых вариантах скелет С применяют со вставкой 3. Такие конструкты возможно содержат ОВ40 ориджин репликации. Плазмида pC3ori+ имеет скелет С со вставкой 3 и ОВ40 ориджином репликации. Плазмида pC3ori- имеет скелет С со вставкой 3, но без ОВ40 ориджина репликации. Кроме того, как pC3ori+, так и pC3ori- могут включать интегразу, что приводит к плазмидам pC3ori+int+ и pC3ori-int+ соответственно. Плазмиды pC3ori+, pC3ori-, pC3ori+int+ и pC3ori-int+ могут быть дополнительно модифицированы функциональной делецией гена обратной транскриптазы (RT) с образованием соответственно pC3ori+RT-, pC3ori-RT-, pC3ori+int+RT- и pC3ori-int+RT-.
В некоторых вариантах скелет D применяют со вставкой 4. Такие конструкты возможно содержат ОВ40 ориджин репликации. Плазмида pD4ori+ имеет скелет D со вставкой 4 и ОВ40 ориджином репликации. Плазмида pD4ori- имеет скелет D со вставкой 4, но без ОВ40 ориджина репликации.
Пример 47
В некоторых вариантах в качестве генетической вакцины дается единственная экспрессионная единица/единственный инокулят. Вакцина представляет собой генетический конструкт, включающий последовательность, кодирующую пептид, в состав которого входит по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопам ВИЧ белков. Кодирующая последовательность находится под регуляторным контролем непосредственно раннего промотора ЦМВ и малого сигнала полиаденилирования ОВ40.
В некоторых вариантах в качестве генетической вакцины дается единственная экспрессионная единица/единственный инокулят. Вакцина представляет собой генетический конструкт, включающий последовательность, кодирующую по меньшей мере один ВИЧ белок или его фрагмент. Кодирующая последовательность находится под регуляторным контролем непосредственно раннего промотора ЦМВ и малого сигнала полиаденилирования ОВ40. ВИЧ белок выбирают из группы, включающей: gag, pol, env и rev. В некоторых рекомендуемых вариантах генетическая вакцина представляет собой генетический конструкт, включающий последовательность, кодирующую по меньшей мере два ВИЧ белка или их фрагменты, выбранных из группы, включающей: gag, pol, env и rev или их фрагменты. В отдельных рекомендуемых вариантах генетическая вакцина представляет собой генетический конструкт, включающий последовательность, кодирующую по меньшей мере три ВИЧ белка или их фрагменты, выбранных на группы, включающей: gag, pol, env и rev или их фрагменты. В некоторых рекомендуемых вариантах дается генетическая вакцина, представляющая собой генетический конструкт, включающий последовательность, кодирующая gag, pol, env и rev или их фрагменты.
В некоторых вариантах дается двойная экспрессионная единица/единственный инокулят в качестве генетической вакцины. Вакцина представляет собой генетический конструкт, включающий две экспрессионные единицы, каждая из которых содержит последовательности, кодирующие пептид, имеющий по меньшей мере один эпитоп, идентичный или по существу аналогичный эпитопам ВИЧ белков. Кодирующая последовательность находится под контролем ЦМВ непосредственно раннего промотора ЦМВ и ОВ40 малого сигнала полиаденилирования.
В некоторых вариантах в качестве генетической вакцины дается двойная экспрессионная единица/единственный инокулят. Вакцина представляет собой генетический конструкт, включающий две экспрессионные единицы, каждая из которых содержит последовательность, кодирующую по меньшей мере один ВИЧ белок или его фрагмент. Каждая экспрессионная единица содержит кодирующую последовательность, находящуюся под регуляторным контролем ЦМВ непосредственно раннего промотора и ОВ40 малого сигнала полиаденилирования. ВИЧ белки выбирают из группы, включающей: gag, pol, env и rev. В некоторых рекомендуемых вариантах дается генетическая вакцина представляющая собой генетический конструкт, включающий две экспрессионные единицы, по меньшей мере одна из которых содержит последовательность, кодирующую по меньшей мере два ВИЧ белка или их фрагменты, выбранные из группы, включающей: gag, rol, env и rev или их фрагменты, а другая содержит по меньшей мере один ВИЧ белок или его фрагменты, выбранные из группы, включающей gag, pol, env и rev или их фрагменты. В отдельных рекомендуемых вариантах дается генетическая вакцина, представляющая собой генетический конструкт, включающий две экспрессионные единицы, по меньшей мере одна из которых содержит последовательность, кодирующую по меньшей мере три ВИЧ белка или их фрагменты, выбранные из группы, включающей: gag, pol, env и rev или их фрагменты, а другая содержит по меньшей мере один ВИЧ белок или его фрагменты, выбранный из группы, включающей: gag, pol, env и rev или их фрагменты. В других рекомендуемых вариантах дается генетическая вакцина, представляющая собой генетический конструкт, состоящий из двух экспрессионных единиц и включающий последовательность, кодирующую gag, pol, env и rev или их фрагменты.
Пример 48
Для применения в анти-ВИЧ фармацевтическом наборе или фармацевтическом препарате создан генетический конструкт (плазмида pCMN160Δ16). pCMN160Δ16 конструируют следующим образом.
Стадия 1. Для получения из ВИЧ/MN геномной ДНК ПЦР фрагмента применяют праймеры ПОСЛЕД. N 35 и ПОСЛЕД. N 34.
Стадия 2. Для получения ПЦР фрагмента из ВИЧ/MN геномной ДНК применяют праймеры ПОСЛЕД. N 33 и ПОСЛЕД. N 36.
Стадия 3. Праймеры ПОСЛЕД. N 35 и ПОСЛЕД. N 36 смешивают с 2 мкл продукта реакции стадий 1 и 2.
Стадия 4. Продукт реакции стадии 3 разрезают с помощью NotI и MluI и вставляют в скелет А, описанный в Примере 46 и разрезанный NotI и Mlul.
В результате создают плазмиду pCMN160Δ16, содержащую в качестве вставки в скелет А область кодирования, кодирующую ENV белок MN штамма с rev областью и половиной nef, имеющий изменения в HLA-DB области ВИЧ-2.
Пример 49
Плазмиду pGAGPOL. rev2 получают следующим образом. Сначала готовят скелет. Затем образуют вставку с ВИЧ gag и pol генами, которую вставляют в скелет.
Скелет получают следующим образом.
Стадия 1. Плазмиду pMAMneo (Клонтех) гидролизуют в присутствии BglI. Наполняют фрагментом Кленова полимеразы I. Разрезают HindIII. Выделяют на геле фрагмент в 1763 п.о.
Стадия 2. Из плазмиды pJ4Ωkan+ амплифицируют KanR ген (ген устойчивости к канамицину, получен от фирмы Фармация Инк., клонирован в pJ4Ω, полученную в виде дара от Имперского Фонда раковых исследований, Великобритания; впервые сообщения о конструировании pJ4Ω появились в работе: Morgenstern J. P., Н. Land, Nucl. Acids Res. 18(4), 1068, которая вводится здесь в качестве ссылки, с олигонуклеотидами ПОСЛЕД. N 37 и ПОСЛЕД. N 38. Затупляют ПЦР продукт. Разрезают HindIII. ПЦР фрагмент очищают на геле.
Стадия 3. Лигируют векторный скелет, образованный из pMAMneo и описанный для стадии 1, с ПЦР продуктом, кодирующим KanR ген и описанным для стадии 2. Выделяют плазмиду, содержащую KanR ген и бактериальный ориджии репликации.
Стадия 4. Полученную плазмиду гидролизуют в присутствии MluI, наполняют фрагментом Кленова из ДНК полимеразы I. Лигируют с SacII линкером (Нью Ингланд Биолабз).
Стадия 5. Полученную на стадии 4 плазмиду гидролизуют в присутствии AseI и SspI.
Стадия 6. Подвергают ПЦР часть KanR гена из плазмиды, описанной для стадии 3, с применением праймеров ПОСЛЕД. N 39 и ПОСЛЕД. 40. Разрезают ПНР продукт SspI и AseI.
Стадия 7. Лигируют самый крупный фрагмент, полученный на стадии 5, с ПЦР продуктом, полученным на стадии 6.
Стадия 8. Разрезают продукт лигирования/плазмиду,
полученный на стадии 7, в присутствии HindIII. Наполняют фрагментом Кленова ДНК полимеразы I.
Стадия 9. В присутствии SelI разрезают рСЕР4 (Инвитроген) с выделением ДНК фрагмента, содержащего промотор ЦМВ, полилинкер и поли(А) участок ОВ40. Полученный фрагмент очищают и затупляют фрагментом Кленова ДНК полимеразы I.
Стадия 10. Лигируют полученную на стадии 8 плазмиду с полученным на стадии 9 фрагментом. Выделяют плазмиду, содержащую бактериальный ориджин репликации, KanR ген, ВСР усилитель, ЦМВ промотор, полилинкер и ОВ40 поли(А) участок.
Стадия 11. Полученную на стадии 10 плазмиду разрезают BamHI и NbeI.
Стадия 12. Отжигают олигонуклеотиды ПОСЛЕД. N 41 и ПОСЛЕД. N 42.
Стадия 13. Лигируют полученную на стадии 10 плазмиду с полученными на стадии 12 олигонуклеотидами. Выделяют плазмиду, содержащую адаптор со стадии 12.
Стадия 14. Полученную на стадии 13 плазмиду гидролизуют в присутствии SalI и MluI.
Стадия 15. Методом ПЦР амплифицируют rev открытую рамку считывания с применением BBG35 (РД Системс Инк. Миннеаполис, МН, содержит область кодирования rev из ВИЧ штамма НХ3В в pUC19) в качестве матрицы и праймеров ПОСЛЕД. N 43 и ПОСЛЕД. N 44. ПЦР продукт гидролизуют в присутствии SalI и MluI.
Стадия 16. Полученную на стадии 14 плазмиду лигируют с полученным на стадии 15 ПЦР продуктом. Выделяют плазмиду, содержащую область кодирования rev.
Получение gag/pol вставки.
Стадия 1. Субклонируют часть ВИЧ-1 (НХВ2) генома, клонированного в Bluescript (Стратаген). Субклон ВИЧ-1 содержит полный 5'-ДКП и остаток ВИЧ-1 генома до нуклеотида 5795 (Нумерация Генбанка), клонированные в XbaI и SalI сайтах Blue-script. Из HXB2D плазмиды (СПИД хранилище) получают ВИЧ-1 последовательности.
Стадия 2. Подвергают ПЦР часть области кодирования gag из открытой рамки считывания плазмиды, описанной на стадии 1 (субклон части ВИЧ-1 НХВ2 генома, клонированного в Blue-script), с применением праймеров ПОСЛЕД. N 45 и ПОСЛЕД. N 46.
Стадия 3. Описанную на стадии 1 плазмиду (субклон части ВИЧ-1 НХВ2 генома, клонированного в Bluescript) гидролизуют в присутствии EcoRI. Очищают плазмиду, содержащую скелет pBluescript, 5' ВИЧ-1 ДКП, область кодирования gag и часть области кодирования pol, и религируют.
Стадия 4. Обрезают полученную на стадии 3 плазмиду в присутствии NotI и SpeI и лигируют с описанным для стадии 2 ПЦР фрагментом после его гидролиза в присутствии NotI и SpeI. Выделяют плазмиду, содержащую ПЦР фрагмент вместо NotI/SpeI фрагмента, включающего 5' ВИЧ-1 ДКП.
Стадия 5. Полученную на стадии 4 плазмиду гидролизуют в присутствии EcoRI и SalI.
Стадия 6. Отжигают олигонуклеотиды ПОСЛЕД. N 47 и ПОСЛЕД. N 48.
Стадия 7. Лигируют полученную на стадии 5 плазмиду с полученным на стадии 6 адаптором. Выделяют плазмиду, содержащую адаптор, клонированный в EcoRI/SalI сайтах.
Стадия 8. Полученную на стадии 7 плазмиду гидролизуют в присутствии NdeI и EcoRI.
Стадия 9. Методом ПЦР амплифицируют Rev-респондирующий элемент (RRE) из плазмиды, содержащей RRE последовательности из ВИЧ-1 штамма НХВ2, с применением праймеров ПОСЛЕД. N 49 и ПОСЛЕД. N 50. ПЦР продукт гидролизуют в присутствии NdeI и EcoRI.
Стадия 10. Лигируют полученную на стадии 8 плазмиду с полученным на стадии 9 ПЦР продуктом. Выделяют плазмиду, содержащую вставку с RRE последовательностью.
Стадия 11. Полученную на стадии 10 плазмиду гидролизуют в присутствии NotI и SalI и выделяют фрагмент, содержащий область кодирования gag, область кодирования модифицированного pol и RRE последовательность.
Стадия 12. По вышеприведенной методике получения скелета в присутствии NotI и SalI гидролизуют полученную на стадии 16 плазмиду.
Стадия 13. Полученную на стадии 12 плазмиду лигируют со вставкой, полученной на стадии 11. Выделяют плазмиды, содержащие вставку, включающую область кодирования gag, область кодирования модифицированного pol и RRE последовательность.
Стадия 14. Полученную на стадии 13 плазмиду гидролизуют в присутствии XbaI и NheI, концы затупляют и повторно лигируют.
Стадия 15. Получение на стадии 14 плазмиду гидролизуют в присутствии KpnI и выделяют самый крупный фрагмент.
Стадия 16. Отжигают олигонуклеотиды ПОСЛЕД. N 51 и ПОСЛЕД. N 52.
Стадия 17. Очищенный плазмидный фрагмент, полученный на стадии 15, лигируют с полученным на стадии 16 адаптором. Выделяют плазмиду, содержащую адаптор, вставленный в KpnI сайт полученной на стадии 15 плазмиды.
Пример 50. Генетическая вакцинация генами регуляторных белков
Затруднения, возникающие при борьбе с ВИЧ, частично связаны с чрезвычайной изменчивостью вируса и его способностью быстро мутировать в новые формы. Существуют существенные изменения белковой последовательности не только в ВИЧ изолятов, обнаруженные среди населения земли в целом, но вирус мутирует настолько быстро, что каждый ВИЧ-инфицированный индивидуум на деле является носителем целого ряда микровариантов ВИЧ. Такие ВИЧ изоляты характеризуются отличиями в эффективности репликации, тропизме, склонности к нейтрализации и устойчивости к лекарственному средству. С появлением устойчивых к лекарству мутантов преимущества лекарственной терапии исчезают. В случае АЗТ устойчивость к лекарственному средству обычно возникает в течение первого года лечения. Такое постоянное возникновение "ускользнувших" мутантов может частично влиять на способность ВИЧ в конце концов подавлять защиту хозяина после длительного периода, в течение которого вирус, видимо, находится под контролем.
Имеются сообщения о таком мутационном дрейфе в различных участках gp120 оболочечного гликопротеина, включая принципиальные нейтрализующие домены V3 петли, а также в белках ядра ВИЧ. ВИЧ регуляторные белки гораздо более консервативны, чем структурные белки, и, кроме того, характеризуются меньшим мутационным дрейфом во времени. Таким образом, регуляторные белки представляются привлекательной целью для противовирусной атаки.
ВИЧ отличается заметной временной регуляцией экспрессии регуляторных белков по сравнению со структурными. На ранних этапах вирусной репликации преобладает мРНК, кодирующая регуляторные белки Tat, Rev и Nef в то время, как на поздних этапах наблюдается повышенная экспрессия мРНК, кодирующей структурные белки, в том числе Gag, Pol и Env предшественники и многие вспомогательные белки. Такое смещение от ранних к поздним этапам прекращается при достижении определенного уровня Rev белка. Преобладание Tat, Rev и Nef на ранних этапах вирусного цикла также делает эти белки благоприятными целями для противовирусной атаки. Это особенно справедливо для tat и rev, играющих абсолютно важную роль в транскрипционной и посттранскрипционной регуляции ВИЧ генной экспрессии, и которые преобладают на ранних этапах цикла вирусной репликации перед транскрипцией вирусных структурных белков и продуцированием инфекционных вирусных частиц.
В отличие от tat и rev, однозначно играющих существенную роль в ВИЧ репликации, другие белки, например: nef, vpr, vif и vpu иногда называют "вспомогательными" белками. Их функции менее хорошо изучены, и степень, в которой может быть ослаблена вирусная репликация при потере определенной функции, колеблется в широких пределах и может зависеть от инфицированной клетки-хозяина. Тем не менее, сильная степень консервативности таких функций в самых различных ВИЧ изолятах, а также в других приматных вирусах иммунодефицита предполагает важность таких "вспомогательных" функций в процессе естественного инфицирования (см. в целом: Terwilliger E.F. (1992) AIDS Research Peviews 2, 3-27, ред. W.C. Koff, F. Wong-Staal, R.C. Kennedy, Нью-Йорк, Марсель Деккер, Инк.). В самом деле, приматные рекомбинантные вирусы с делецией vpr, nef или vif непатогены in vivo, что еще больше подчеркивает важность этих вспомогательных генов в жизненном цикле вируса.
Существуют некоторые свидетельства того, что защитной иммунной реакции против ВИЧ на более высоком уровне можно достигнуть проведением отбора нескольких регуляторных и/или ферментативных белков, но не всего набора ВИЧ генов. Соответственно, стратегия сфокусированной вакцинации желательно включает генетическую вакцинацию с применением последовательностей, кодирующих один или несколько регуляторных, неструктурных ВИЧ белков, в том числе tat, rev, vpr, nef, vpu или vif. Только vpr, как найдено, ассоциируется с вирусными частицами, в то время как другие регуляторные белки, включая tat, rev, nef, vif и vpu не связаны с вирионом.
В некоторых вариантах генетической вакцинации от ВИЧ с применением регуляторных генов один из tat, rev, nef, vif или vpu генов или несколько таких генов вставляют в описанный в Примере 46 скелет А. Рекомендуется применение tat и/или rev. Е некоторых вариантах в скелет А, описанный в Примере 46, вставляют tat или rev. В некоторых вариантах применяют (далее следует перечень в порядке снижения приемлемости в качестве целей) nef, vpr, vif и vpu. Рекомендуется применение более одного регуляторного гена, в том числе: tat и rev; tat, rev и nef; tat, rev, nef и vpr; tat, rev, nef, vpr и vif; tat, rev, nef, vpr, vif и vpu, а также их комбинаций и возможно таких дополнительных регуляторных генов, как tev.
Tat белок является трансактиватором ДКП-направленной генной экспрессии. Белок абсолютно необходим для ВИЧ репликации. Tat. продуцируется на ранних этапах цикла вирусной репликации, и функциональный Tat необходим для экспрессии Gag, Pol, Env и Vpr. Преобладающей формой Tat является белок из 86 аминокислот, происходящий из двухэкзонной мРНК. Амино-концевых 58 аминокислот достаточно для трансактивации, хотя и с пониженной активностью. Tat влияет на цис-действующую последовательность, называемую tar, с обеспечением значительного повышения ДКП-ведомой генной экспрессии. Влияние Tat может осуществляться частично путем повышения синтеза РНК и частично путем увеличения количества белка, синтезируемого через РНК транскрипт. До недавнего времени считалось, что Tat влияет только на ВИЧ-1 ДКП. Однако появились также сообщения о Tat-активируемой экспрессии из позднего промотора JC вируса. В качестве экзогенного фактора Tat может также стимулировать клеточную пролиферацию и может вносить свой вклад в промотирование роста саркомы Капоши у ВИЧ-инфицированных индивидуумов. Вследствие такого потенциально опасного действия на ВИЧ-инфицированных и неинфицированных индивидуумов рекомендуемые tat конструкты, применяемые для генетической вакцинации, модифицированы с экспрессией только нефункционального Tat. Мутации, способные дезактивировать Tat или Rev, могут, кроме того, действовать, как трансдоминантные мутации с потенциальной дезактивацией в результате любого функционального Tat, продуцируемого в организме ВИЧ-инфицированных индивидуумов.
Rev является вторым регуляторным белком ВИЧ, необходимым для вирусной репликации. Rev является белком в 19 кД (116 аминокислот), экспрессируемым из двух кодирующих экзонов, в широком спектре сплайсированных мРНК. Идентифицированы два отличных друг от друга домена: основная область, участвующая в связывании с RRE (Rev-респондирующий элемент) содержащими транскриптами, и домен "активации", индуцирующий в результате связывания ядерный экспорт таких транскриптов. В ходе естественной вирусной инфекции Rev необходим для экспрессии ВИЧ структурных белков Gag, Pol и Env, а также Vpr.
Vpr, являющийся белком в 15 кД (96 аминокислот), находится в большинстве ВИЧ-1 штаммов, хотя Vpr открытая рамка считывания интенсивно усекается во многих вирусных штаммах при интенсивном прохождении в клеточной культуре. Vpr открытая рамка считывания присутствует также в ВИЧ-2 и большинстве изолятах. Vpr является первым ретровирусным регуляторным белком, для которого обнаружена связь с ВИЧ вирусными частицами. Его присутствие в ВИЧ вирионах предполагает возможность участия белка в некоторых ранних этапах цикла вирусной репликации. Vpr ускоряет репликацию ВИЧ, особенно на ранних этапах инфекции. Vpr примерно в три раза повышает уровень экспрессии генов-репортеров, связанных в ВИЧ ДКП. Более того, Vpr и Tat, видимо, действуют синергически на ДКП-связанные гены. Vpr может быть выделен из сыворотки ВИЧ-инфицированных индивидуумов, и, видимо, повышает способность вируса инфицировать новые клетки. Найдено также, что Vpr подавляет клеточную пролиферацию и индуцирует дифференцировку клеток (Levy D.M. и др. Cell (1993) 72, 1-20), открытие, которое может оказаться важным ввиду сообщений о том, что первичные моноциты/макрофаги оказываются инфекционными in vitro только претерпевая дифференцировку (Schuitemaker Н. и др. , J. Clin. Invest. 89, 1154-1160 (1992)). Одинаковые клетки, неспособные поддерживать репликацию ВИЧ, под влиянием Vpr могут быть разрегулированы. К примеру, Vpr может нести ответственность за потерю) мышечной ткани, часто наблюдаемую у больных СПИДом. Вследствие потенциально опасной активности Vpr генетическую вакцинацию следует предпочтительно проводить с модифицированным vpr конструктом, экспрессирующим нефункциональный Vpr белок.
Nef (называемый также в более старых источниках 3'orf) является белком в 25-27 кД. Предполагалось, что Nef может участвовать в разрегулировании CD4+ Т-лимфоцитов. Кроме того, Nef может участвовать в клеточных сигналах. Nef, видимо, имеет важное значение для установления ВИЧ инфекции in vivo. Полагают, что Nef-специфичные ЦТЛ имеют важное значение для регулирования ВИЧ инфекции in vivo.
Vif является цитоплазмическим белком в 23 кД, называемым "фактором инфекционности вируса". Хотя Vif-дефектные мутанты и не являются компромиссными в отношении трансмиссии типа клетка-к-клетке, такие мутанты характеризуются заметным снижением способности инфицировать многие CD4+ клеточные линии. В отсутствие Vif наблюдается снижение отмочковывания и снижение инфекционности. В опытах на приматах мутанты Vif с делецией проявляют жестко сниженную способность создавать инфекцию in vivo. Эти исследования подтверждают клиническую роль Vif в репликации вируса в хозяине.
Vpu является белком в 15-20 кД (81 аминокислота). Хотя Vpu(+) и Vpu(-) вирусы продуцируют одинаковое количество белка, последние отличаются повышенным внутриклеточным аккумулированием вирусных белков одновременно с уменьшением внеклеточного вируса. Это предполагает участие Vpu в сборке и/или выделении вирусных частиц.
Простые ретровирусы, например: мышиные к птичьи вирусы не имеют белков, аналогичных ВИЧ-1, ВИЧ-2 и SIV регуляторным белкам. У таких животных ретровирусная инфекция имеет тенденцию к самоограничению с клиренсом вируса и сниженной патогенностью. Аналогично, HTLV-1, включающий только Tax (действующий почти как Tat и, кроме того, проявляющий vpr-подобную активность) и Rex (действующий почти, как Rev), подвергается клиренсу в организме многих индивидуумов. Генетическая вакцинация регуляторными генами, как считают, уместна не только от ВИЧ, но также в случае ВЭБ (X генный продукт), HCV и HTLV-I (Tax) и (Rex). У всех указанных вирусов регуляторные гены, как полагают, играют решающую роль в жизненном цикле вируса и создании инфекции.
Пример 51. Конструирование ВИЧ-1 регуляторной плазмиды (рРЕС)
Плазмиду рРЕС конструируют постадийно, и каждое промежуточное вещество перед продолжением конструирования может быть проверено на экспрессию белка. Экспрессионный вектор, поддерживающий экспрессию tat и rev, конструируют в две стадии. Вначале продукт амплификации, содержащий 5' NheI сайт, ВИЧ-1 основной сплайс-донорный участок, основную часть области кодирования tat, область кодирования, кодирующей аминоконцевую область rev белка, и AvaII сайт, амплифицируют из синтетической матрицы. Такая синтетическая матрица создана применением опубликованных последовательностей НХВ2 штамма ВИЧ-1, полученных из базы данных GenBank, и последовательности изменены мутацией в белке tat цистеиновых остатков в положениях 22 и 30. Такие мутации, как показано, придают tat нефункциональность (Kuppuswany и др. (1989) Hucleic Acids Research 17(9), 3551-3561).
ПЦР продукт лигируют в вектор, гидролизованный в присутствии NheI и AvaII и содержащий ген устойчивости к канамицину и рВР322 ориджин репликации. Кроме того, эта плазмида содержит цитомегаловирусный промотор, усилитель вируса саркомы Рауса, область кодирования rev и ОВ40 сигнал полиаденилирования. Присутствующая в указанной плазмиде rev последовательность происходит из провирусного клона ВИЧ-1 III8. В результате создают экспрессионный вектор, содержащий полную, но мутированную область кодирования tat и полную область кодирования rev.
Проведением последующей стадии создают ПЦР продукт, содержащий AvaII сайт у своего 5'-конца, мутацию у аминокислоты в положении 81 rev, примерно 30% области кодирования rev, примерно 30% области кодирования nef и MluI сайт и 3'-конца. Замена аминокислоты в положении 81, как показано, приводит к удалению rev функции, вследствие чего полученная плазмида будет продуцировать нефункциональный rev белок (Bogard Н., Greene W.C. (1993) J. Virol. 67(5), 2496-2502). Считается, что основная делеция в nef области кодирования приведет к продуцированию нефункционального nef белка. 5' AvaII сайт и мутацию у аминокислоты в положении 81 rev белка вводят в 5' ПЦР праймер, комплементарный области кодирования rev, содержащей как AvaII сайт, так и нуклеотид, кодирующий аминокислоту 81. Применением 3' ПЦР праймера вводятся терминирующая кодон, вызывающий обрыв Nef у аминокислоты в положении 63, и 3' кодирующий клонирование MluI сайт. Матрицей для такой ПЦР амплификации служит плазмида или синтетическая матрица, содержащие области кодирования rev и nef из MN штамма ВИЧ-1. Полученный ПЦР продукт гидролизуют в присутствии AvaII и MluI и применяют для замены меньшего AvaII-MluI фрагмента, образующегося после гидролиза в присутствии AvaII и MluI описанной в предыдущем параграфе tat-rev плазмиды.
Возможно введение в один из двух сайтов этой плазмиды vpr. В одном из подходов vpr может быть амплифицирован применением 5' ПЦР праймера, содержащего MluI сайт в восходящем направлении в последовательности, охватывающей vpr трансляционный инициирующий кодон, и 3' ПЦР праймера, комплементарного vpr терминирующему кодон и последовательностям, фланкирующих кодон и содержащих также MluI сайт клонирования. Последовательности в восходящем направлении от инициирующего кодона содержат сплайс-акцептор. ПЦР продукт может быть гидролизован в присутствии Mlul и вставлен в tat rev net плазмиду, описанную выше, после ее гидролиза в присутствии Mlul.
Или же vpr амплификация может быть осуществлена аналогичным путем, однако ПЦР праймеры должны содержать рестрикционные сайты, совместимые с клонированием в другой вектор так, что тот экспрессируется под контролем второго эукариотного промотора. Происходящая из такой плазмиды кассета, содержащая второй промотор, за которым следует область кодирования vpr, затем поли(А), может быть получена гидролизом ферментами рестрикции, фланкирующими кассету, но не вклинивающимися в ее пределы. Полученный ДНК фрагмент необходимо клонировать в единственный сайт tat, rev, vpr плазмиды выпадающей из области, необходимой для экспрессии tat rev vpr. Таким путем образуют плазмиду с двумя экспрессионными единицами.
Пример 52. Конструирование HCV и HTLV-I плазмид
Аналогичный подход может быть использован для создания плазмиды, экспрессирующей HTLV-1 или HCV кодируемые белки, обладающей ферментативными свойствами, необходимыми для жизненного цикла вируса и/или для регуляторных белков указанных вирусов. В случае HTLV-1 применением плазмидного скелета и стратегии клонирования, аналогичных вышеприведенным, создают плазмиду, кодирующую регуляторный белок Tax. Подобные HCV гены, кодирующие ферментативные белки, включают РНК-зависимую РНК-полимеразу, белок с функциями геликазы/протеазы. Сведения о необходимых последовательностях опубликованы и они могут быть получены в Генбанке. Данные о строении HTLV-1 и HCV приводятся в издании: Cann A.J., Chen I.S.Y. "Вирусология", 2-ое издание, под редакцией B.N. Fiddr, Равен Пресс, Лтд., Нью-Йорк, 1990 и работе: Bradley V.W. Transfusion Medicine Peviews 1(2), 93-102, 1992, соответственно.
Пример 53. Генетическая вакцинация ферментативными генами
Генетическая вакцинация генами, кодирующими белки с функцией ферментов, например ВИЧ pol геном, может также являться важной стратегией борьбы с вирусом, поскольку такие ферменты, как Pol необходимы для продуцирования живого вируса. Без полимеразы или ее любого функционального компонента ВИЧ непатогенен и неинфекционен. Аналогично, ферментативные гены других вирусов, например HBV полимеразы, представляются привлекательными целями для генетической вакцинации. См., напр., Radziwill и др. "Мутационный анализ генного продукта вируса Р гепатита В. Строение домена и активность РНазы Н", J. Virol. 64(2), 613-620 (1990).
Одна из причин привлекательности вирусных ферментов в качестве иммунологических мишений состоит в ограниченной способности аминокислотных последовательностей таких ферментов к мутации с сохранением ими при этом своей ферментативной способности. К примеру, в случае ВИЧ-1 фермент Pol характеризуется ограниченным числом "ускользнувших" мутаций, ассоциируемых с устойчивостью к нуклеотидным аналогам, например АЗТ. Однако подавляющее большинство иммунологических мишеней в белке сохраняются даже в ускользнувших от лекарственного средства мутантах.
Пример 54. Конструирование HBV полимеразной плазмиды
Освещенные в литературе опыты указывают на то, что экспрессия HBV полимеразы достигается в тканевой культуре клеток в случае присутствия в молекуле мРНК как коровой, так и полимеразной открытых рамок считывания. Также показано, что в такой ситуации мутация в АТС ядра не влияет на экспрессию полимеразы.
HBV геном амплифицируют из плазмиды, содержащей димер типа голова-к-хвосту ADW HBV штамма. Поскольку желательна экспрессия только полимеразы, но не ядра, созданный 5' ПСР праймер приводит к мутации трансляционных инициирующих кодонов преядра и ядра. Кроме того, такой праймер вводит также мутантную DRI последовательность с целью исключить возможность образования репликационно-компетентной HBV геномной РНК. Этот ПЦР продукт помещают в плазмиду, содержащую ген устойчивости к канамицину и pBR322 ориджин репликации. Кроме того, такая плазмида содержит цитомегаловирусный промотор, усилитель вируса саркомы Payса и ОВ40 сигнал полиаденилирования. Для предотвращения экспрессии HBV и X генных продуктов трансляционные инициирующие кодоны для поверхностного антигена и продукта области кодирования X подвергают мутации.
Согласно другому подходу к достижению экспрессии HBV полимеразы ПЦР продукт, кодирующий область кодирования полной полимеразы, амплифицируют и клонируют в вектор, содержащий ген устойчивости к канамицину и pBR322 ориджин репликации. Кроме того, такая плазмида содержит цитомегаловирусный промотор, усилитель саркомы Рауса и ОВ40 сигнал полиаденилирования. 5' ПЦР праймер для такой амплификации содержит сайт клонирования и охватывает трансляционный инициирующий кодон гена полимеразы. 3' ПЦР продукт содержит сайт рестрикции для клонирования вставки в экспрессионный вектор и, кроме того, комплементарен традиционному терминирующему кодону гена HBV полимеразы и последовательностям, фланкирующим этот терминирующий кодон. После лигирования указанного ПЦР продукта в плазмиду, содержащую ген устойчивости к канамицину, pBR322 ориджин репликации, цитомегаловирусный промотор, усилитель вируса саркомы Рауса и ОВ40 сигнал полиаденилирования, трансляционные инициирующие кодоны для поверхностного антигена гепатита В и X генов подвергают мутации с целью предотвратить экспрессию этих генных продуктов. Применяемая альтернативная стратегия аналогична вышеприведенной, однако 3' ПЦР праймер в этом случае включает HBVполи(А) сигнал и фланкирующие этот сигнал последовательности. Такой 3' праймер применяют в том случае, если последовательности, включающие и/или окружающие HBVполи(А) сигнал, имеют важное для экспрессии значение. Мутационный анализ показал, что функционирование HBV полимеразного генного продукта можно исключить заменой конкретного нуклеотида (Radziwell G. и др. (1990) J. Virol. 64(2), 613-620). Перед употреблением плазмиды, сконструированной вышеприведенным образом, экспрессируемая полимераза может быть подвергнута мутации созданием одной из таких мутаций или других аналогичных мутаций.
Пример 55
Гранулоцит-макрофаг колониестимулирующий фактор (GM-CSF) оказывает стимулирующее действие на разнообразные клеточные линии, в том числе нейтрофилы, моноциты/макрофаги и эозинофилы. Действие GM-CSF делает его привлекательной терапевтической моделью. GM-CSF одобрен FDA для применения в аутологичной пересадке костного мозга, и начаты клинические испытания эффективности фактора в лечении различных нейтропений. В настоящее время GM-CSF вводят в виде белка, обычно требующего введения многими дозами. Необходимо получить и очистить такие белки.
Альтернативный подход к применению GM-CSF белка состоит в прямом введении генного конструкта, содержащего ген, кодирующий GM-CSF, в сочетании с введением бупивакаина. Генетически конструкт создают методом ПЦР из GM-CSF гена, включающего сигнальную последовательность. Генетический конструкт предпочтительно содержит ген устойчивости к канамицину (ген аминогликозид-3'фосфотрансферазы), бактериальный ориджин репликации, последовательности, поддерживающие экспрессию области кодирования GM-CSF в клетках, в которые вводят плазмиду, например, векторов, охарактеризованных в виде скелетов в Примере 46. Плазмида предпочтительно содержит ориджин репликации млекопитающего, индуцируемый клеточной репликацией, связанной с введением бупивакаина. В случае применения ВЭБ ориджина репликации включают также последовательность, кодирующую ядерный антиген EBNA-1, вместе с соответствующими регуляторными последовательностями. Праймеры для ПЦР амплификации вставки сайты рестрикции ферментами, позволяющими проводить клонирование в экспрессионный вектор, и комплементарными 5'- и 3'-концам GM-CSF кодирующей последовательности. ПЦР реакцию осуществляют с кДНК клоном по методике Lee и др., Proc. Natl. Acad. Sci. США 82, 4360- 4364.
Пример 56
Хронический миелогенный лейкоз (ХМЛ) является клональным миелопролиферативным нарушением кроветворных стволовых клеток, ассоциируемым с Филадельфийской хромосомой, т.е. хромосомой, возникающей в результате транслокации между хромосомами 9 и 22. Точечный разрыв в хромосоме 22 образует кластер в области в 6 к. о., называемой кластерной областью точечного разрыва (КОКР), в то время как в хромосоме 9 точечные разрывы рассеяны по всей области в 90 к. о. в восходящем направлении от c-abl экзона 2. Различные возникающие 9:22 транслокации могут быть подразделены на два типа: К28 транслокации и L6 транслокации. Транскрипция через bcr-abl приводит к образованию слитых мРНК. Антисмысловая молекула, нацеленная на bcr-abl сочленение мРНК, как показано, снижает способность кроветворных клеток, отобранных у больных ХМЛ, образовывать колонии.
Генетический конструкт, кодирующий антисмысловую молекулу, вводят вместе с бупивакаином в клетки индивидуума, страдающего ХМЛ, ex vivo. Обработанные клетки затем вновь вводят индивидууму.
Пример 57
Генные конструкты, применимые в фармацевтических наборах и препаратах для вакцинации от HBV и его лечения, конструируют с векторами, охарактеризованными в виде скелетов в Примере 46. Плазмиды содержат HBV структурные гены, в частности генов, кодирующих HBV поверхностный антиген и/или HBV антиген ядра, и/или HBV антиген преядра.
Пример 58
Генные конструкты, применимые в фармацевтических наборах и препаратах для вакцинации от HCV и его лечения, конструируют с векторами, охарактеризованными в виде скелета в Примере 46. Плазмиды содержат HCV структурные гены, в частности гены, кодирующие HCV белок ядра и/или HCV белок оболочки.
Пример 59
Создают генный конструкт pREV, содержащий нуклеотидную последовательность, кодирующую ВИЧ rev в качестве единственного целевого белка. Кодирующую rev последовательность клонируют в скелете В, описанный в Примере 46, из ВВС35 (RD Системе Инк., Миннеаполис, МН), содержащей область кодирования rev из ВИЧ штамма НХ3В в pUC19.
ТАБЛИЦА 1
Семейство пикорнавирусов.
Род
Риновирусы: (медицина) ответственны за ≈50% общих простудных состояний.
Этеровирусы: (медицина) включают полиовирусы, вирус коксаки, эховирусы и энтеровирусы человека, например вирус гепатита А.
Аптовирусы: (ветеринария) включают вирусы ящура.
Целевые антигены: VP1, VP2, VP3, VP4, VPG.
Семейство кальцивирусов.
Род
Вирусы Группы Norwalk: (медицина) вирусы являются важными возбудителями эпидемического гастроэнтерита.
Семейство тогавирусов.
Род
Альфавирусы: (медицина и ветеринария) примеры включают вирус преждевременного старения, вирус Росс-Ривер, вирус восточного и западного энцефалита лошадей.
Реовирус: (медицина) вирус краснухи.
Семейство флавирусов.
Примеры включают: (медицина) вирус денге, желтой лихорадки, японского энцефалита, энцефалита Сент-Луис и клещевого энцефалита.
Вирус гепатита С: (медицина) эти вирусы не объединены в семейство до настоящего времени, но, как полагают, они относятся либо к тогавирусам, либо флавивирусам. Наибольшее сходство с семейством тогавирусов.
Семейство коронавирусов:
(медицина и ветеринария) вирус инфекционного бронхита (домашняя птица), переносимый вирус гастроэнтерита свиней (свиньи), вирус гемагглютинирующего энцефаломиелита свиней (свиньи), вирус кошачьего инфекционного перитонита (кошки), кошачий брюшной коронавирус (кошки), собачий коронавирус (собаки). Респираторные коронавирусы человека вызывают ≈40% случаев общих простудных состояний, напр., 22КЕ, ОСК3.
Примечание: коронавирусы могут вызвать гепатит не-А, В или С.
Целевые антигены:
Е1, называемый также М или матриксным белком; Е2, называемый также S или белком шипов; Е3, называемый также НЕ или гемагглютинелтерозным гликопротеином (присутствует не во всех коронавирусах; N или нуклеокапсид.
Семейство рабдовирусов.
Род: везилиовирус, лиззавирус (медицина и ветеринария): бешенство.
Целевой антиген: G-протеин, N-протеин.
Семейство филовирусов.
Род
Парамиксовирус: (медицина и ветеринария) вирус эпидемического паратита, вирус ньюкастлской болезни (важный патоген у цыплят).
Морбилливирус: (медицина и ветеринария) корь, собачья чума.
Пневминвирус: (медицина и ветеринария) респираторный синцитиальный вирус.
Семейство ортомиксовирусов (медицина): вирус гриппа.
Семейство буньявирусов (Bungavirus Family).
Род
Буньявирус (медицина): калифорнийский энцефалит, ЛЯ Кросс.
Флебовирус (медицина): вирус лихорадки долины Рифт.
Хантавирус: пуремала, являющийся вирусом гемахагиновой (hemahagin) лихорадки.
Найрвирус (ветеринария): найробийская болезнь овец.
Кроме того, много неклассифицированных буньявирусов.
Семейство ареновирусов.
(медицина): LCM, вирус лихорадки Ласса.
Семейство Реовирусов.
Род
Реовирус (возможный патоген человека).
Ротавирус: острый гастроэнтерит у детей.
Орбивирусы (медицина и ветеринария): колорадская клещевая лихорадка, лемомбо (человек), лошадиный энцефалит, болезнь "синего языка".
Семейство ретровирусов.
Подсемейство: онковирусное (ветеринария и медицина): вирус кошачьего лейкоза, HTLV-1, HTLV-2.
Лентивирусно (медицина и ветеринария): ВИЧ, вирус иммунодефицита кошек, инфекции лошадей, вирус анемии.
Спумавирусное.
Семейство паповавирусов.
Подсемейства: полиомавирусы (медицина): BKU и JCU вирусы.
Папилломавирус (медицина): многие вирусные типы, ассоциируемые с раком или злокачественным развитием папилломы.
Аденовирус (медицина): напр. : AD7, ARD, А.В. - вызывают респираторные заболевания, некоторые аденовирусы, например, 275 вызывают энтерит.
Семейство парвовирусов (ветеринария): кошачий парвовирус, вызывает энтерит у кошек; кошачий вирус панлейкопении кошек; собачий парвовирус; парвовирус свиней.
Семейство герпесвирусов.
Подсемейство: альфагерпесвиридное.
Род: симплексвирус (медицина); HSVI, HSVII; варицелловирус (медицина и ветеринария): псевдобешенство - ветряная оспа.
Подсемейство: бетагерпесвиридное.
Род: цитомегаловирус; HCMV; мурамегаловирус.
Подсемейство: гаммагерпесвиридное.
Род: (медицина) лимфокриптовирус; ВЭБ (буркитт лимфо); радиновирус.
Семейство поксвируса.
Подсемейство: хордопоксвиридное (медицина и ветеринария)
Род вариола (натуральная оспа), вакциния (коровья оспа); парапоксвирус (ветеринария); ауипоксвирус (ветеринария); каприпоксвирус; лепорипоксвирус, суипоксвирус.
Подсемейство: энтемопоксвиридное.
Семейство гепадневируса: вирус гепатита В.
Неклассифицирован: вирус гепатита дельта.
ТАБЛИЦА 2
Бактериальные патогены
Патогенные грамположительные кокки включают: пнемококк, стафилококк, стрептококк. Патогенные грамотрицательные кокки включают: менингококк и гонококк.
Патогенные энтерические грамотрицательные бациллы включают: энтеробактерии, псевдомонады, ацинетобактерии и эйкенеллы, мелиоидоза, сальмонеллы, шигеллоза, гемофилии, шанкроида, бруцеллоза, тулеремии, йерсинии (пастеурелла), стрептобацилла монилиформис, спирилла, листериозные моноцитогены, эризипелотриксные рузиопаты, дифтерии, холеры, сибирской язвы, донованоза (паховой грануломы) и бартенеллоза.
Патогенные анаэробные бактерии включают: столбняка, ботулизма, другие клостридии, туберкулеза, проказы и другие микробактерии. Патогенные спирохетные заболевания включают: сифилис, трепонематоз, тропическую гранулему, пинту, эндемический сифилис и лептоспироз. Другие инфекции, возбудителями которых являются высшие патогенные бактерии и патогенные грибки, включают: актиномикоз, нокардиоз, криптококкоз, бластомикоз, гистоплазмоз, мукормикоз, споротрихоз, паракокцидиомукоз, петриеллидиоз, торулопсоз, мицетому, хромомикоз и дерматофитоз.
Риккетсиозные инфекции включают риккетсиоз.
Примеры микоплазмических и хламидиальных инфекций включают: микоплазмической пневмонии, венерической лимфогрануломы, пситакоз и перинатальные хламидиальные инфекции.
Патогенные эукариоты
Патогенные простейшие и гельминты и создаваемые ими инфекции включают: амебиоз, малярию, лейшманиоз, трипаносомоз, токсоплазмоз, пневмоцистизный каринез, бебизиоз, гиардиаз, трихиноз, филариоз, шистосомоз, нематоды, трематоды или трематоды и цестодные (ленточные черви) инфекции.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
(2) Информация для ПОСЛЕД. N 1:
(I) Характеристики последовательности
(А) Длина:; 32 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(IX) Описание последовательности: ПОСЛЕД. N 1:
AGGCGTCTCG AGACAGAGGA GAGCAAGAAA TG
(2) Информация для ПОСЛЕД. N 2:
(I) Характеристики последовательности
(А) Длина: 30 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 2:
ТТТСССТСТА GATAAGCCAT ССААТСАСАС
(2) Информация для ПОСЛЕД. N 3:
(I) Характеристики последовательности:
(А) Длина: 29 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 3:
GAAGGATCCA TGAAAAAATA TTTATTGGG
(2) Описание для ПОСЛЕД. N 4:
(I) Характеристики последовательности:
(А) Длина: 30 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 4:
ACTGTCGACT TATTTTAAAG CGTTTTTAAG
(2) Информация для ПОСЛЕД. N 5:
(I) Характеристики последовательности:
(А) Длина: 30 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 5:
GCCAGTTTTG GATCCTTAAA AAAGGCTTGG
(2) Информация для ПОСЛЕД. N 6:
(I) Характеристики последовательности:
(А) Длина: 27 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 6:
TTGTGAGGGA CAGAATTCCA ATCAGGG
(2) Информация для ПОСЛЕД. N 7:
(I) Характеристики последовательности:
(А) Длина: 24 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 7:
CAGTGATATC CCGGGAGACT CCTC
(2) Информация для ПОСЛЕД. N 8:
(I) Характеристики последовательности:
(А) Длина: 25 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 8:
GAATAGAAGA ACTCCTCTAG ААТТС
(2) Информация для ПОСЛЕД. N 9:
(I) Характеристики последовательности:
(А) Длина: 27 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 9:
GCCTTAGGCG GATCCTATGG CAGGAAG
(2) Информация для ПОСЛЕД. N 10:
(I) Характеристики последовательности:
(А) Длина: 24 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 10:
TAAGATGGGT GGCCATGGTG AATT
(2) Информация для ПОСЛЕД. N 11:
(I) Характеристики последовательности:
(А) Длина: 22 аминокислоты
(В) Тип: аминокислота
(D) Топология: линейная
(II) Тип молекулы: пептид
(XI) Описание последовательности: ПОСЛЕД. N 11:
(2) Информация для ПОСЛЕД. N 12:
(I) Характеристики последовательности:
(А) Длина: 25 аминокислот
(В) Тип: аминокислота
(D) Топология: линейная
(II) Тип молекулы: пептид
(XI) Описание последовательности: ПОСЛЕД. N 12:
(2) Информация для ПОСЛЕД. N 13:
(I) Характеристики последовательности:
(А) Длина: 27 аминокислот
(В) Тип: аминокислота
(D) Топология: линейная
(II) Тип молекулы: пептид
(XI) Описание последовательности: ПОСЛЕД. N 13:
(2) Информация для ПОСЛЕД. N 14:
(I) Характеристики последовательности:
(А) Длина: 25 аминокислот
(В) Тип: аминокислота
(D) Топология: линейная
(II) Тип молекулы: пептид
(XI) Описание последовательности: ПОСЛЕД. N 14:
(2) Информация для ПОСЛЕД. N 15:
(I) Характеристики последовательности:
(А) Длина: 27 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 15:
TTGTTTAACT TTTGATCGAT ССАТТСС
(2) Информация для ПОСЛЕД. N 16:
(I) Характеристики последовательности:
(А) Длина: 21 пара оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 16:
GATTTGTATC GATGATCTGA С
(2) Информация для ПОСЛЕД. N 17:
(I) Характеристики последовательности:
(А) Длина: 25 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 17:
TGTAGTAGCA AAAGAAATAG TTAAG
(2) Информация для ПОСЛЕД. N 18:
(I) Характеристики последовательности:
(А) Длина: 25 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 18:
ААТТСТТААС ТАТТТСТТТТ GCTAC
(2) Информация для ПОСЛЕД. N 19:
(I) Характеристики последовательности:
(А) Длина: 40 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 19:
ATTTGTCGAC TGGTTTCAGC CTGCCATGGC AGGAAGAAGC
(2) Информация для ПОСЛЕД. N 20:
(I) Характеристики последовательности:
(А) Длина: 29 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 20:
ACGACGCGTA TTCTTTAGCT CCTGACTCC
(2) Информация для ПОСЛЕД. N 21:
(I) Характеристики последовательности:
(А) Длина: 24 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 21:
GCTGACGGTA GCGGCCGCAC ААТТ
(2) Информация для ПОСЛЕД. N 22:
(I) Характеристики последовательности:
(А) Длина: 22 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 22:
GTATTAAGCG GCCGCAATTG ТТ
(2) Информация для ПОСЛЕД. N 23:
(I) Характеристики последовательности:
(А) Длина: 78 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 23:
AAAAAGCTTC GCGGATCCGC GTTGCGGCCG CAACCGGTCA CCGGCGACGC GTCGGTCGAC CGGTCATGGC TGGGCCCC
(2) Информация для ПОСЛЕД. N 24:
(I) Характеристики последовательности:
(А) Длина: 29 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 24:
CCCAGCTTA GACATGATAA GATACATTG
(2) Информация для ПОСЛЕД. N 25:
(I) Характеристики последовательности:
(А) Длина: 22 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 25:
CTAGCAGCTG GATCCCAGCT TC
(2) Информация для ПОСЛЕД. N 26:
(I) Характеристики последовательности:
(А) Длина: 24 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 26:
GGATTTCTGG GGATCCAAGC TAGT
(2) Информация для ПОСЛЕД. N 27:
(I) Характеристики последовательности:
(А) Длина: 31 пара оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 27:
TATAGGATCC GCGCAATGAA AGACCCCACC Т
(2) Информация для ПОСЛЕД. N 28:
(I) Характеристики последовательности:
(А) Длина: 31 пара оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 28:
ATATGGATCC GCAATGAAAG ACCCCCGCTG А
(2) Информация для ПОСЛЕД. N 29:
(I) Характеристики последовательности:
(А) Длина: 30 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 29:
TAAAGCGGCC GCTCCTATGG CAGGAAGACG
(2) Информация для ПОСЛЕД. N 30:
(I) Характеристики последовательности:
(А) Длина: 34 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 30:
ATTACGCGTC TTATGCTTCT AGCCAGGCAC AATG
(2) Информация для ПОСЛЕД. N 31:
(I) Характеристики последовательности:
(А) Длина: 40 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 31:
ATTACGCGTT TATTACAGAA TGGAAAACAG ATGGCAGGTG
(2) Информация для ПОСЛЕД. N 32:
(I) Характеристики последовательности:
(А) Длина; 32 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 32:
ATTACGCGTT ATTGCAGAAT TCTTATTATG GC
(2) Информация для ПОСЛЕД. N 33:
(I) Характеристики последовательности:
(А) Длина: 38 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 33:
GAGGCTTGGA GAGGATTATA GAAGTACTGC AAGAGCTG
(2) Информация для ПОСЛЕД. N 34:
(I) Характеристики последовательности:
(А) Длина: 39 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 34:
GAATCCTCTC CAAGCCTCAG CTACTGCTAT AGCTGTGGC
(2) Информация для ПОСЛЕД. N 35:
(I) Характеристики последовательности:
(А) Длина: 39 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 35:
AAAAATAAAG CGGCCGCTCC TATGGCAGGA AGAGAAGCG
(2) Информация для ПОСЛЕД. N 36:
(I) Характеристики последовательности:
(А) Длина: 39 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 36:
ААААААТТАС GCGTCTTATG CTTCTAGCCA GGCACAATG
(2) Информация для ПОСЛЕД. N 37:
(I) Характеристики последовательности:
(А) Длина: 31 пара оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 37:
CCCAAGCTTG GGAATGCTCT GCCAGTGTTA С
(2) Информация для ПОСЛЕД. N 38:
(I) Характеристики последовательности:
(А) Длина: 36 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 38:
GGGGGCCGGA AGGGCACAAT AAAACTGTCT GCTTAC
(2) Информация для ПОСЛЕД. N 39:
(I) Характеристики последовательности:
(А) Длина: 33 пары оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 39:
CCTGATTCAG GTGAAAATAT TGTTGATGCG CTG
(2) Информация для ПОСЛЕД. N 40:
(I) Характеристики последовательности:
(А) Длина: 111 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 40:
ААСАТСААТА СААССТАТТА АТТТССССТС GTCAAAAATA AGGTTATCAA GTGAGAAATC ACCATCAGTG ACGACTGAAT CCGGTGAGAA TGGCAAAAGT TTATGCATTT С
(2) Информация для ПОСЛЕД. N 41:
(I) Характеристики последовательности:
(А) Длина: 55 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 41:
CTAGCTCGGG GATCCGCGTT GCGGCCGCAA AAAGTCGACG GGCGACGCGT AAAAA
(2) Информация для ПОСЛЕД. N 42:
(I) Характеристики последовательности:
(А) Длина: 55 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 42:
GATCTTTTTA CGCGTCGCCC GTCGACTTTT TGCGGCCGCA ACGCGGATCC CCGCG
(2) Информация для ПОСЛЕД. N 43:
(I) Характеристики последовательности:
(А) Длина: 48 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 43:
ATGTCGACTG GTTTCAGCCT GCCATGGCAG GAAGAAGC
(2) Информация для ПОСЛЕД. N 44:
(I) Характеристики последовательности:
(А) Длина: 35 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 44:
CCCCACGACG CGTCTATTCT TTAGCTCCTG АСТСС
(2) Информация для ПОСЛЕД. N 45:
(I) Характеристики последовательности:
(А) Длина: 35 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 45:
TTTGCGGCCG CGTAAGTGGA GAGAGATGGT GCGAG
(2) Информация для ПОСЛЕД. N 46:
(1) Характеристики последовательности
(А) Длина: 21 пара оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 46:
CTGGTGGGGC TGTTGGCTCT G
(2) Информация для ПОСЛЕД. N 47:
(I) Характеристики последовательности:
(А) Длина: 80 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 47:
ААТТТААТАА GTAAGTAAGT GTCATATGTT TGTTTGAATT CTGCAACAAC TGCTGTTTAT CCATTTTCAG AATTGGGTG
(2) Информация для ПОСЛЕД. N 48:
(I) Характеристики последовательности:
(А) Длина: 80 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 48:
TCGACACCCA ATTCTGAAAA TGGATAAACA GCACTTGTTG CAGAATTCAA АСАААСАТАТ GACACTTACT ТАСТТАТТА
(2) Информация для ПОСЛЕД. N 49:
(I) Характеристики последовательности:
(А) Длина: 41 пара оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 49:
GGGGTTTTTG GGCATATGTA TGAGGGACAA TTGGAGAAGT G
(2) Информация для ПОСЛЕД. N 50
(I) Характеристики последовательности:
(А) Длина: 70 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 50:
AAGCTTGTGG AATTCTTAAT TTCTCTGTCC GGGGTTTTTG GGCATATGTA TGAGGGACAT TGGAGAAGTG
(2) Информация для ПОСЛЕД. N 51:
(I) Характеристики последовательности:
(А) Длина: 29 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 51:
CAGTATCTGG CATGGGTAC
(2) Информация для ПОСЛЕД. N 52:
(I) Характеристики последовательности:
(А) Длина: 29 пар оснований
(В) Тип: нуклеиновая кислота
(С) Число нитей: одна
(D) Топология: линейная
(II) Тип молекулы: ДНК
(XI) Описание последовательности: ПОСЛЕД. N 52:
CCATGCCAGA TACTGGTACй


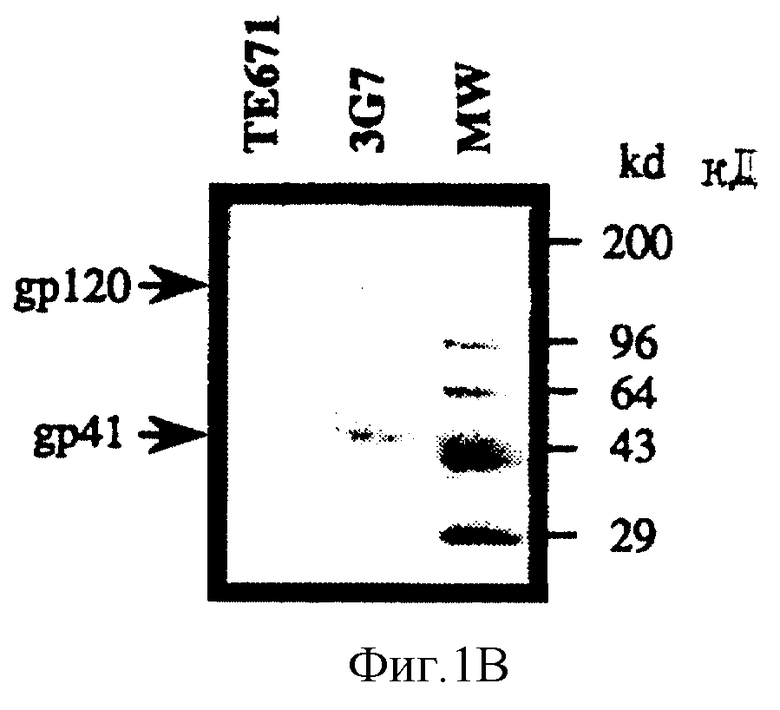
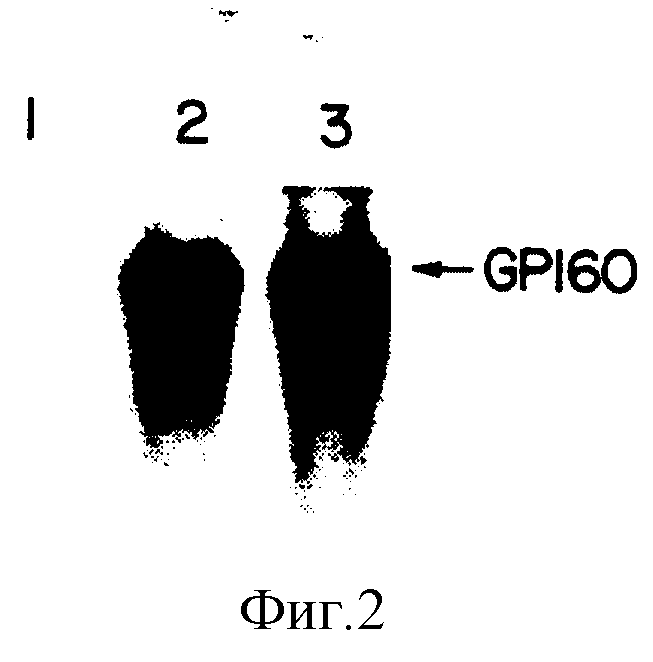
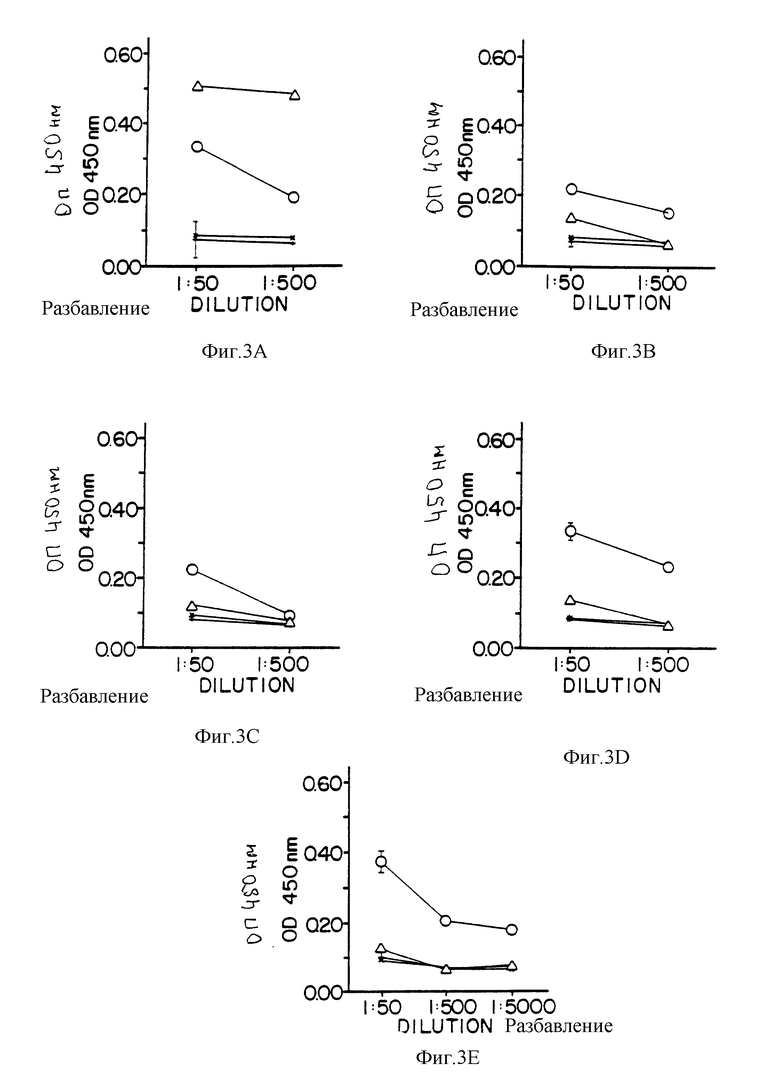
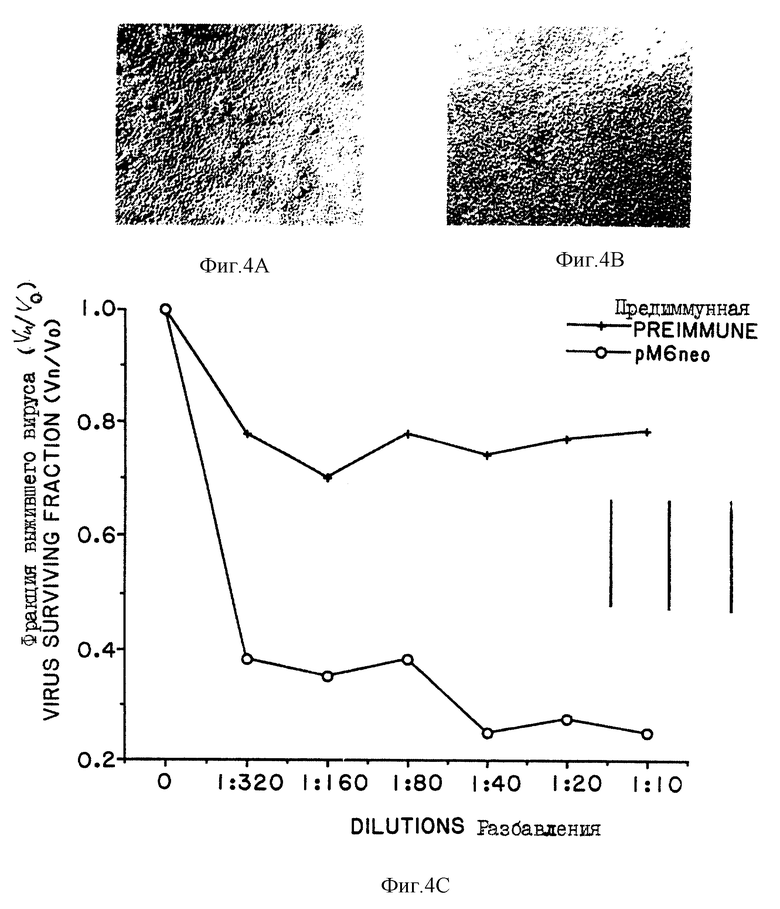
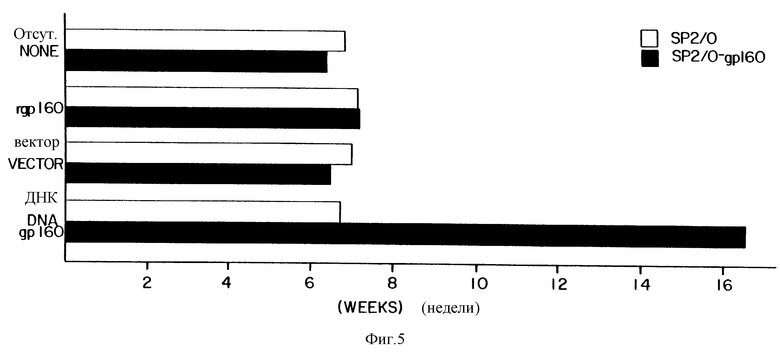
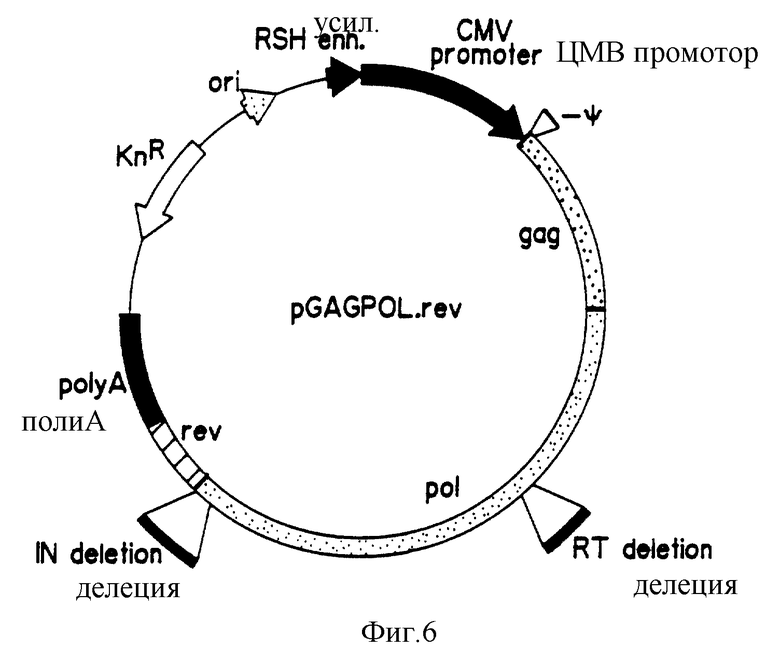
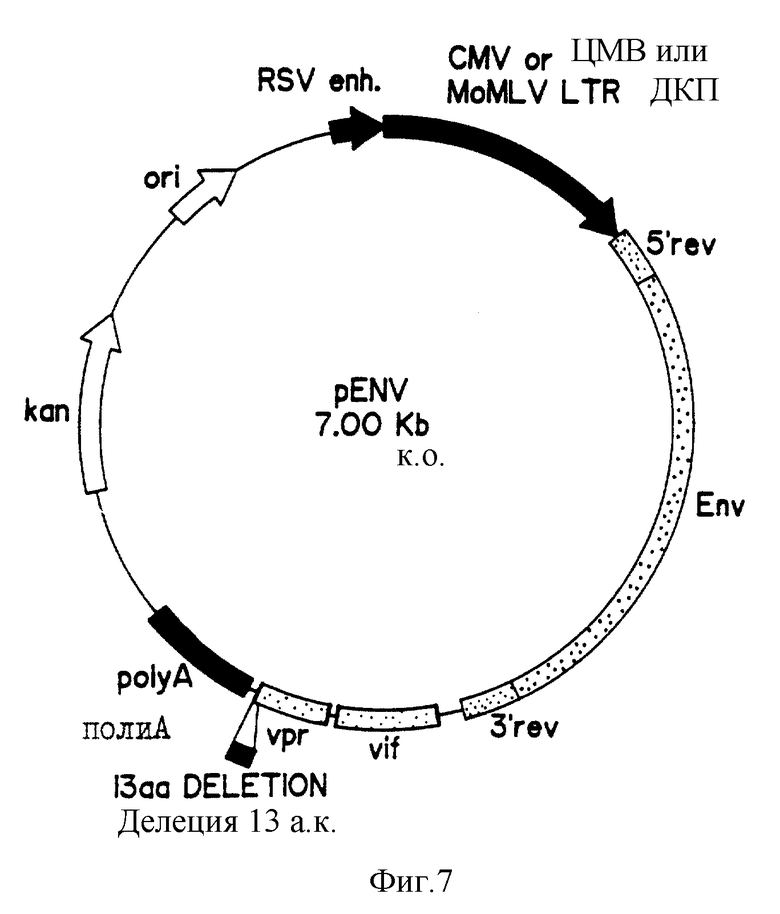
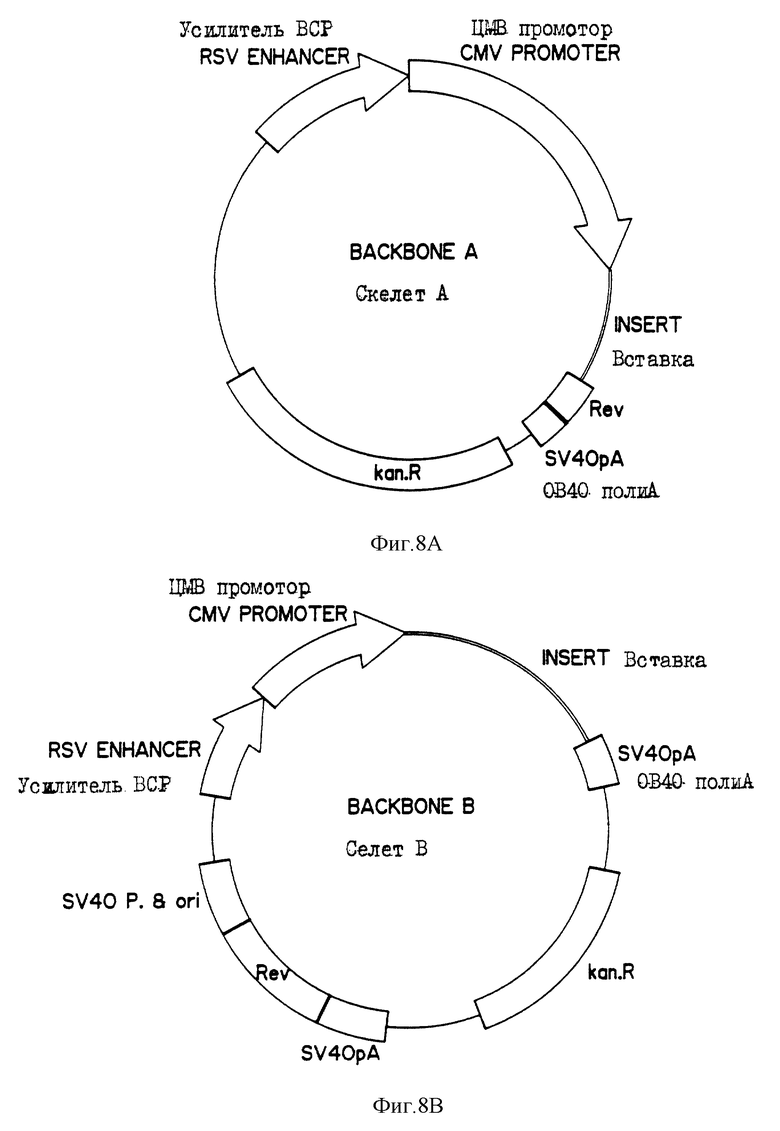
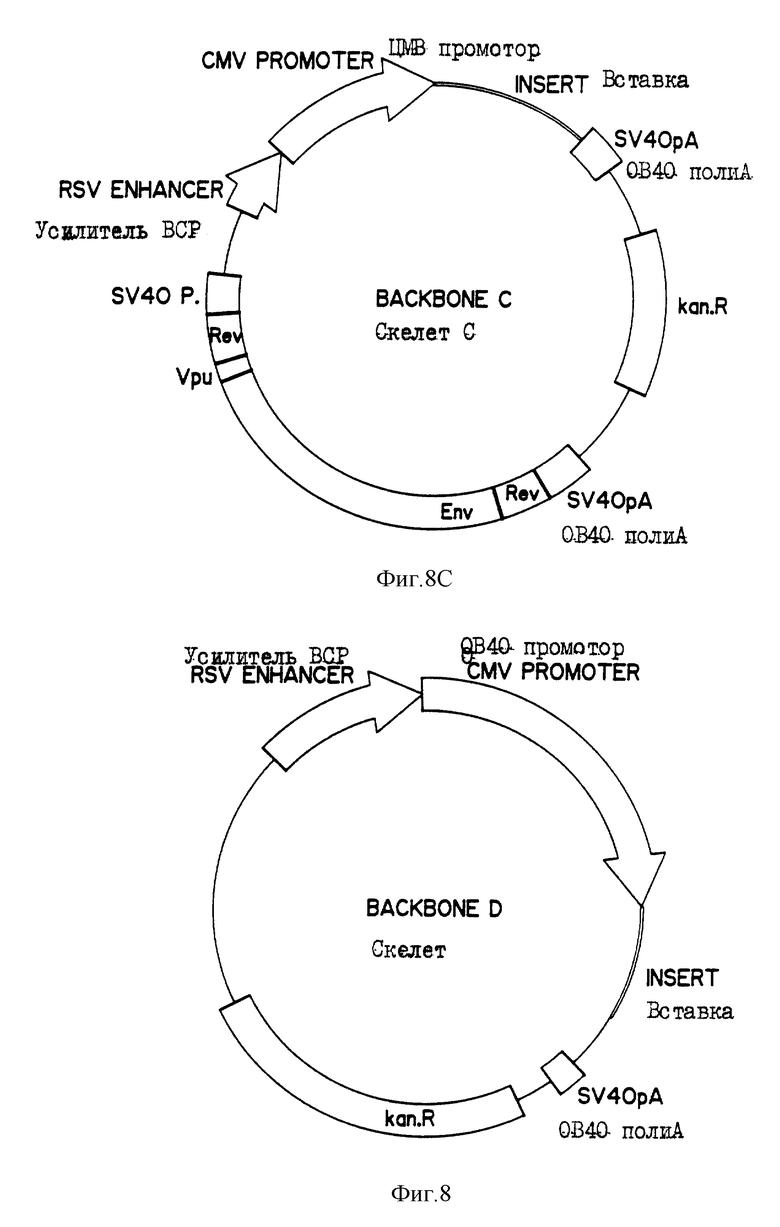
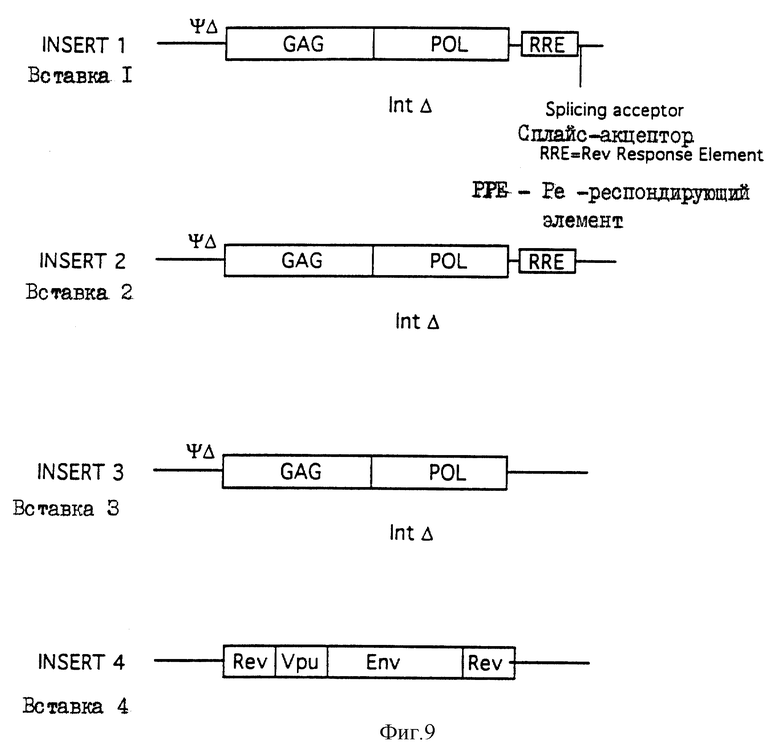
Изобретение относится к области медицины, а именно медицинской генетике. Сущность изобретения: в клетки индивидуума вводят генетический материал с усилителем функции полинуклеотида (например, бупивакаином), а также вводят молекулы нуклеиновой кислоты, не содержащей частиц ретровируса, молекула нуклеиновой кислоты включает такую последовательность, которая кодирует белок, в состав последнего входит по меньшей мере один эпитоп, идентичный или, по существу, аналогичный эпитопу антигена, против которого желателен иммунный ответ, причем нуклеотидная последовательность операбельно связана с регуляторными последовательностями. Техническим результатом изобретения является расширение арсенала средств против ВИЧ. Изобретение описывает также фармацевтическую композицию и набор для осуществления способа. 3 с. и 45 з. п. ф-лы, 4 табл., 9 ил.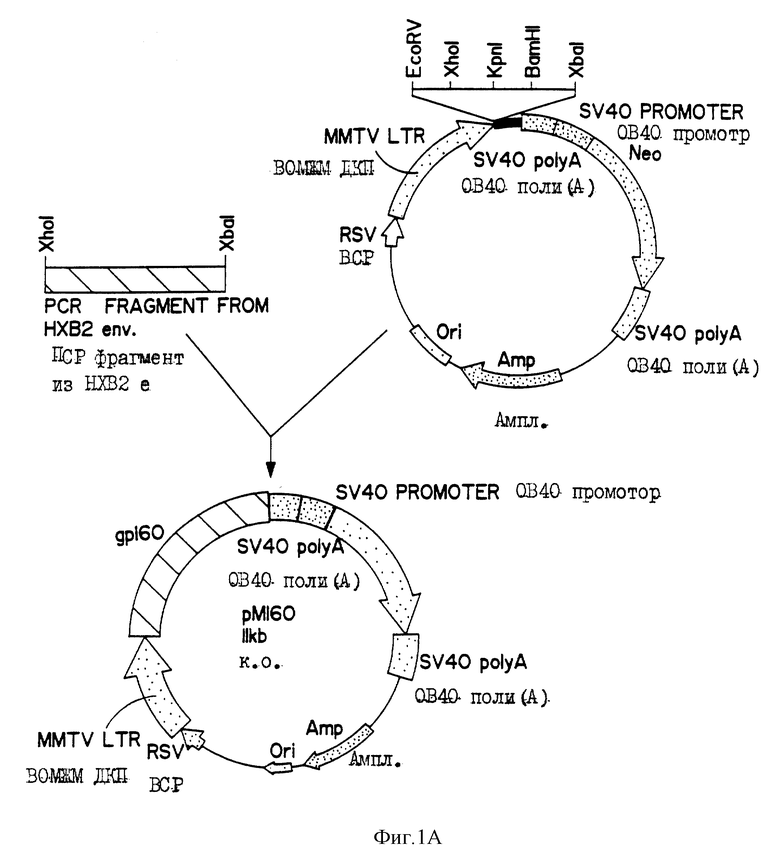
Приоритеты по пунктам:
26.01.1993 по пп.2, 17, 44, 45 и 48;
11.03.1993 по пп.6, 18 - 25 и 27.
| US 4636385 А, 13.01.1987 | |||
| Wolfe J.A | |||
| et al | |||
| "Biotechniges", 11(4), p.474-485. |

