Предпосылки изобретения
Настоящее изобретение относится к лечению расстройств центральной нервной системы. Более конкретно, изобретение относится к лечению неврологических и психических расстройств, например психоза и расстройств с симптомом когнитивного дефицита. Кроме того, настоящее изобретение относится к лечению нейродегенеративных расстройств и состояний. Изобретение также относится к ингибированию PDE10. Изобретение также относится к способам идентификации химических соединений, обладающих селективной ингибиторной активностью в отношении PDE10.
Циклические нуклеотиды, циклический аденозинмонофосфат (цАМФ) и циклический гуанозинмонофосфат (цГМФ) действуют как внутриклеточные вторичные посредники, регулирующие большое количество внутриклеточных процессов, в частности, в нейронах центральной нервной системы. В нейронах их действие заключается в активации цАМФ- и цГМФ-зависимых киназ и последующем фосфорилировании белков, участвующих в регуляции быстрой синаптической передачи, а также в дифференцировке и выживании нейронов. О сложности сигнального пути с участием циклических нуклеотидов говорит большое молекулярное разнообразие ферментов, участвующих в синтезе и деградации цАМФ и цГМФ. Существует десять семейств аденилилциклазы, два семейства гуанилилциклазы и одиннадцать семейств фосфодиэстеразы (PDE). Более того, известно, что в различных типах нейронов экспрессируются многочисленные изозимы каждого фермента этих классов, что является убедительным подтверждением компартментализации и функциональной специфичности различных изозимов в указанных нейронах.
цАМФ синтезируется семейством мембранных ферментов, аденилилциклазами, о которых упоминалось выше. Активность этих ферментов регулируется разнообразными рецепторами семейства серпина посредством механизма присоединения, опосредованного гетеротримерным G-белком. Увеличение концентрации внутриклеточного цАМФ приводит к активации цАМФ-зависимых протеинкиназ, которые регулируют активность других сигнальных киназ, факторов транскрипции и ферментов посредством их фосфорилирования. Кроме того, циклический АМФ может непосредственно влиять на активность ионных каналов, регулируемых циклическими нуклеотидами, активность фосфодиэстераз и факторов обмена гуанин - нуклеотид. Последние исследования также показали, что внутриклеточный цАМФ может действовать как предшественник нейромодулятора аденозина, контролируя его выход из клетки.
Гуанилилциклаза, которая синтезирует цГМФ, находится как в мембранно-связанной, так и в цитоплазматической форме. Мембранно-связанная форма представляет собой рецепторы, связанные с G-белком, как, например, рецептор атриального натрийуретического пептида (ANP), тогда как растворимая форма гуанилилициклазы активируется окисью азота (Wang, X. and Robinson, P.J. Journal of Neurochemistry 68(2):443-456, 1997). Как и в случае с цАМФ, медиаторы, участвующие в сигнальном пути цГМФ центральной нервной системы, включают в себя ионные каналы, чувствительные к цГМФ, цГМФ-специфические фосфодиэстеразы и цГМФ-зависимые протеинкиназы. Так как циклические нуклеотиды играют важную роль в сигнальной трансдукции в центральной нервной системе, применение соединений, которые влияют на регуляцию сигнального пути с участием циклических нуклеотидов, может иметь важное терапевтическое значение.
Основным механизмом регуляции сигнального пути с участием циклических нуклеотидов является катаболизм циклических нуклеотидов, катализируемый фосфодиэстразами. Известны одиннадцать семейств фосфодиэстераз (PDE), кодируемых 21 разными генами. Каждый ген экспрессирует различные сплайс-варианты, из которых затем образуется большое количество изозимов. Семейства PDE различаются по функциональной специфичности в отношении субстрата, то есть циклических нуклеотидов, по механизму(ами) регуляции и чувствительности к ингибиторам. Более того, PDE не одинаково экспрессируются в организме, в том числе в центральной нервной системе. В результате такого различия ферментной активности и локализации, разные изозимы PDE могут сильно отличаться по физиологическим свойствам. Кроме того, соединения, которые могут селективно ингибировать разные семейства PDE или изозимы, могут иметь особые терапевтические эффекты, меньше побочных эффектов или и то, и другое.
На основании первичной аминокислотной последовательности и особенной ферментативной активности семейство PDE10 идентифицировано как уникальное. Скрининг гомологов базы данных EST показал, что PDE10A мыши является первым членом семейства фосфодиэстрезы PDE10 (Fujishige et al., J. Biol. Chem. 274:18438-18445, 1999; Loughney, K. et al., Gene 234:109-117, 1999). Также были клонированы гомологи PDE мыши (Soderling, S. et al., Proc. Natl. Acad. Sci. USA 96:7071-7076, 1999) и идентифицированы N-концевые сплайс-варианты генов крыс и человека (Kotera, J. et al., Biochem. Biophys. Res. Comm. 261:551-557, 1999; Fujishige, K. et al., Eur. J. Biochem. 266:1118-1127, 1999). Степень межвидовой гомологичности очень велика. Мышиный PDE10A1 представляет собой белок из 779 аминокислот, который гидролизует как цАМФ, так и цГМФ, до АМФ и ГМФ, соответственно. Аффинность PDE10 в отношении цАМФ (Кm = 0,05 мкМ) выше аффинности в отношении цГМФ (Кm = 3 мкМ). Однако на основании того, что Vmax цГМФ приблизительно в 5 раз больше Vmax цАМФ, было сделано предположение, что PDE10 является уникальной цГМФазой, ингибируемой цАМФ (Fujishige et al., J. Biol. Chem. 274:18438-18445, 1999).
У млекопитающих PDE10 также имеет уникальную локализацию по сравнению с другими представителями семейства PDE. мРНК фермента PDE10 имеет высокую степень экспрессии только в яичках и мозге (Fujishige, K. et al., Eur J. Biochem. 266:1118-1127, 1999; Soderling, S. et al., Proc. Natl. Acad. Sci. USA 96:7071-7076, 1999; Loughney, K. et al., Gene 234:109-117, 1999). Эти первоначальные исследования показали, что в мозге экспрессия PDE10 выше в полосатом теле (хвостатом ядре и скорлупе), прилежащем ядре (n. accumbens) и обонятельных бугорках. Недавно был осуществлен подробный анализ характера экспрессии мРНК фермента PDE10 в мозге грызунов (Seeger, T.F. et al., Abst. Soc. Neurosci. 26:345.10, 2000) и PDE10 белка (Menniti, F.S., Stick, C.A., Seeger, T.F. и Ryan, A.M., Immunohistochemical localization of PDE10 in the rat brain. William Harvey Research Conference 'Phosphodiesterase in Health and Disease', Porto, Portugal, Dec. 5-7, 2001).
Краткое описание изобретения
Настоящее изобретение относится к способу лечения тревожного или психического расстройства у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для лечения указанного тревожного или психического расстройства.
Изобретение также относится к способу лечения тревожного или психического расстройства у млекопитающего, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для ингибирования PDE10.
Примеры психических расстройств, лечение которых может осуществляться в соответствии с настоящим изобретением, включают в себя, но ими не ограничиваются, шизофрению, например, параноидальную, гебефреническую, кататоническую, недифференцированную или остаточную шизофрению; шизофреноподобное расстройство; шизоаффективное расстройство, например, параноидного типа или депрессивного типа; бредовое расстройство; психические расстройство, вызванное приемом веществ, например, психоз, вызванный приемом алкоголя, амфетамина, марихуаны, кокаина, галлюциногенов, форм для ингаляции, опиоидов или фенциклидина; расстройство личности параноидального типа и расстройство личности шизоидного типа.
Примеры тревожных расстройств, лечение которых может осуществляться в соответствии с настоящим изобретением, включают в себя, но ими не ограничиваются, паническое расстройство; агрофобию; специфические фобии; социальную фобию; обсессивно/компульсивное расстройство; посттравматическое стрессовое расстройство; острую реакцию на стресс и генерализованное тревожное расстройство.
Настоящее изобретение также относится к способу лечения двигательных расстройств, выбранных из болезни Гентингтона и дискинезии, связанной с терапевтическим применением агонистов допамина, у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для лечения указанного расстройства.
Настоящее изобретение также относится к способу лечения двигательных расстройств, выбранных из болезни Гентингтона и дискинезии, связанной с терапевтическим применением агонистов допамина, у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для ингибирования PDE10.
Настоящее изобретение, кроме того, относится к способу лечения двигательного расстройства, выбранного из болезни Паркинсона, синдрома «беспокойных ног» и эссенциального тремора у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для лечения указанного расстройства.
Настоящее изобретение также относится к способу лечения двигательного расстройства, выбранного из болезни Паркинсона, синдрома «беспокойных ног» и эссенциального тремора у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для ингибирования PDE10.
Настоящее изобретение также относится к способу лечения расстройства, выбранного из обсессивно/компульсивных расстройств, синдрома Туретта и других тиков у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для лечения указанного расстройства.
Настоящее изобретение также относится к способу лечения обсессивно/компульсивных расстройств, синдрома Туретта и других тиков у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для ингибирования PDE10.
Настоящее изобретение также относится к способу лечения наркотической зависимости, например алкогольной, амфетаминовой, кокаиновой или опиоидной зависимости, у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для лечения указанной лекарственной зависимости.
Настоящее изобретение также относится к способу лечения наркотической зависимости, например алкогольной, амфетаминовой, кокаиновой или опиоидной зависимости, у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для ингибирования PDE10.
Под «наркотической зависимостью» понимают патологическую потребность в приеме наркотика, которая обычно характеризуется нарушением мотивационной сферы, таким как компульсивность принятия желаемого наркотика и эпизоды непреодолимого влечения к нему.
Настоящее изобретение также относится к способу лечения расстройства, включающего в себя симптом нарушения внимания и/или когнитивной функции, у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для лечения нарушения внимания и/или когнитивной функции.
Настоящее изобретение также относится к способу лечения расстройства, включающего в себя симптом нарушения внимания и/или когнитивных функций, у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для ингибирования PDE10.
Под использующейся здесь фразой «нарушение внимания и/или когнитивной функции» во фразе «расстройство, включающее в себя симптом нарушения внимания и/или когнитивной функции» понимают патологическую активность одной или нескольких когнитивных функций, таких как память, интеллект или способность к обучению и логическое мышление, у конкретного лица, по сравнению с лицами той же возрастной категории. «Нарушение внимания и/или когнитивной функции» также относится к снижению у любого конкретного лица активности одной или нескольких когнитивных функций, например происходящих при возрастном снижении когнитивных функций.
Примеры расстройств, включающих в себя в качестве симптома нарушение внимания и/или когнитивной функции, лечение которых может осуществляться в соответствии с настоящим изобретением, включают в себя деменцию, например деменцию при болезни Альцгеймера, мультиинфарктную деменцию, алкогольную деменцию или другую деменцию, вызванную приемом наркотических средств, деменцию, связанную с внутричерепной опухолью или травмой головного мозга, деменцию при болезни Гентингтона или болезни Паркинсона, деменцию при болезни, вызванной вирусом иммунодефицита человека (ВИЧ); делирий; амнестический синдром; посттравматическое стрессовое расстройство; умственную отсталость; расстройство развития учебных навыков, например расстройство чтения, расстройство арифметических навыков или расстройство развития экспрессивного письма; нарушение активности и внимания; возрастное снижение когнитивных функций.
Настоящее изобретение также относится к способу лечения расстройства настроения или депрессивного эпизода у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для лечения указанного расстройства или эпизода.
Настоящее изобретение также относится к способу лечения расстройства настроения или депрессивного эпизода у млекопитающих, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для ингибирования PDE10.
Примеры расстройств настроения и депрессивных эпизодов, лечение которых может осуществляться в соответствии с настоящим изобретением, включают в себя, но ими не ограничиваются, большой депрессивный эпизод легкой, умеренной или тяжелой степени, маниакальный или смешанный аффективный эпизод, гипоманию; депрессивный эпизод, атипический тип; депрессивный эпизод, меланхолический тип; депрессивный эпизод с кататоническими нарушениями; аффективный послеродовой эпизод; посттравматическое стрессовое расстройство; большую депрессию; дистимическое расстройство; малое депрессивное расстройство; предменструальное дисфорическое расстройство; постпсихотическую депрессию при шизофрении; депрессивный эпизод с психотическим расстройством, такой как бредовое расстройство или шизофрения; биполярное расстройство, например биполярное расстройство I типа, биполярное расстройство II типа и циклотимия.
Настоящее изобретение также относится к способу лечения нейродегенеративного расстройства или состояния у млекопитающего, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для лечения указанного расстройства или состояния.
Настоящее изобретение также относится к способу лечения нейродегенеративного расстройства или состояния у млекопитающего, включая человека, который заключается во введении указанному млекопитающему количества селективного ингибитора PDE10, эффективного для ингибирования PDE10.
В рамках изобретения, если не указано иное, к «нейродегенеративному расстройству или состоянию» относится расстройство или состояние, которое вызвано дисфункцией и/или гибелью нейронов центральной нервной системы. Эффективность лечения этих расстройств и состояний может быть повышена введением агента, который предупреждает дисфункцию или гибель нейронов, подверженных риску при этих расстройствах или состояниях, и/или усиливает функцию поврежденных или здоровых нейронов с тем, чтобы компенсировать потерю функции, вызванной дисфункцией или гибелью подверженных риску нейронов. Под используемым здесь термином «нейротрофический агент» понимают вещество или агент, который обладает всеми или частью этих свойств.
Примеры нейродегенеративных расстройств и состояний, лечение которых может осуществляться в соответствии с настоящим изобретением, включают в себя, но ими не ограничиваются, болезнь Паркинсона; болезнь Гентингтона; деменцию, например, при болезни Альцгеймера, мультиинфарктную деменцию, деменцию при СПИДе и фронто-темпоральную деменцию; нейродегенеративный процесс, связанный с травмой головного мозга; нейродегенеративный процесс, связанный с инсультом, нейродегенеративный процесс, связанный с инфарктом мозга; нейродегенеративный процесс, вызванный гипогликемией; нейродегенеративный процесс, связанный с эпилептическими припадками; нейродегенеративный процесс, связанный с нейротоксическим отравлением и множественную системную атрофию.
В одном из вариантов осуществления настоящего изобретения нейродегенеративное расстройство или состояние включает в себя нейродегенеративный процесс медиальных шиповатых нейронов полосатого тела у млекопитающих, включая человека.
В другом варианте осуществления настоящего изобретения нейродегенеративное расстройство или состояние представляет собой болезнь Гентингтона.
«Нейротоксическое отравление» относится к отравлению, вызванному нейротоксическим веществом. Нейротоксическое вещество представляет собой любое химическое вещество или вещество, которое может вызывать гибель нейронов и, таким образом, неврологическое повреждение. Примером нейротоксического вещества является алкоголь, злоупотребление которого беременными женщинами может привести к алкогольному отравлению и неврологическому повреждению, известному как синдром внутриутробного алкоголизма (FAS) новорожденных. Другие примеры нейротоксических веществ включают в себя, но ими не ограничиваются, каиновую кислоту, домоевую кислоту и акромеловую кислоту; некоторые пестициды, такие как DDT; некоторые инсектициды, такие как органофосфаты; летучие органические растворители, такие как гексакарбоны (например, толуол); тяжелые металлы (например, свинец, ртуть, мышьяк и фосфор); алюминий; некоторые химические соединения, используемые в качестве химического оружия, такие как реактив «эйджент орандж» и нервно-паралитический газ; и нейротоксические противоопухолевые агенты.
Под используемым здесь термином «селективный ингибитор PDE10» понимают вещество, например органическое вещество, которое более эффективно ингибирует ферменты семейства PDE10 по сравнению с ферментами семейств PDE 1-9 или семейства PDE11. В одном варианте осуществления селективный ингибитор PDE10 представляет собой соединение, например органическое вещество, имеющее Ki ингибирования PDE10, меньше или равное приблизительно одной десятой Ki ингибирования любого другого фермента PDE. Другими словами, это вещество ингибирует активность PDE10 в той же степени при концентрации, составляющей приблизительно одну десятую или меньше от концентрации, необходимой для ингибирования любого другого фермента PDE.
В общих чертах, считается, что вещество эффективно ингибирует активность PDE10, если его Ki меньше или приблизительно равно 10 мкМ, предпочтительно меньше или приблизительно равно 0,1 мкМ.
В одном из вариантов осуществления терапевтических способов, описанных в настоящем изобретении, селективным ингибитором PDE10 является папаверин.
«Селективный ингибитор PDE10» может быть идентифицирован, например, путем сравнения способности вещества ингибировать активность PDE10 с его способностью ингибировать ферменты PDE разных семейств PDE. Например, вещество может быть исследовано на способность ингибировать активность PDE10, а также PDE1, PDE2, PDE3A, PDE4A, PDE4B, PDE4C, PDE4D, PDE5, PDE6, PDE7, PDE8, PDE9 и PDE11.
В одном из вариантов осуществления терапевтических способов по изобретению селективным ингибитором PDE10 является папаверин.
Настоящее изобретение также относится к способу селективного ингибирования PDE10 у млекопитающих, включая человека, заключающемуся во введении указанному млекопитающему папаверина в количестве, эффективном для ингибирования PDE10.
Термин «лечение» в «способе лечения расстройства» относится к реверсированию, облегчению или ингибированию развития расстройства, к которому относится данный термин, или одного или нескольких симптомов этого расстройства. Используемый здесь термин также охватывает, в зависимости от состояния пациента, профилактику расстройства, включая профилактику возникновения расстройства или любого симптома с ним связанного, а также уменьшение тяжести расстройства или любого его симптома до его возникновения. «Лечение» в рамках настоящего изобретения также относится к профилактике рецидива расстройства.
Например, «лечение шизофрении, или шизофреноподобного, или шизоаффективного расстройства» в рамках изобретения включает в себя лечение одного или нескольких симптомов (положительных, отрицательных и других связанных симптомов) указанных расстройств, например лечение бреда и/или галлюцинации, с ними связанных. Другие примеры симптомов шизофрении и шизофреноподобных и шизоаффективных расстройств включают в себя спонтанную речь, эмоциональную тупость, алогичность, ангедонию, неуместную тревогу, дисфорическое настроение (в форме, например, депрессии, тревожности или раздражительности), и некоторые указания на когнитивную дисфункцию.
Использующийся здесь термин «млекопитающие» относится к любому представителю класса «Mammalia», включая, но ими не ограничиваясь, человека, собак и кошек.
Настоящее изобретение также относится к новому способу скрининга соединений на наличие способности селективно ингибировать PDE10.
Например, изобретение также относится к способу определения селективной ингибиторной активности в отношении PDE10 у химического соединения, который включает в себя: a) введение химического соединения в культуру медиальных шиповатых нейронов и b) измерение повышения фосфорилирования CREB в культуре; повышение фосфорилирования CREB, тем самым, показывает, что соединение, использованное в стадии (a) обладает селективной ингибиторной активностью в отношении PDE10.
В другом примере изобретение относится к способу определения селективной ингибиторной активности в отношении PDE10 у химического соединения, который включает в себя: a) введение химического соединения в культуру медиальных шиповатых нейронов и b) измерение повышения количества продуцируемого GABA медиальными шиповатыми нейронами в указанной культуре; повышение продукции GABA указанными медиальными шиповатыми нейронами, тем самым, показывает, что соединение, использованное в стадии (a) обладает селективной ингибиторной активностью в отношении PDE10.
Культура медиальных шиповатых нейронов может быть получена специалистом в данной области с использованием известных способов, например, но ими не ограничиваясь, способов, подробно описанных ниже.
Химические соединения могут вводиться в культуру медиальных шиповатых клеток одним из вышеуказанных способов, с использованием известных способов. Введение химических соединений может осуществляться автоматизировано или вручную. Более того, группы химических соединений можно скринировать в соответствии с любым из способов путем высокопроизводительного скрининга. Необязательно, для одновременного и/или последовательного анализа различных соединений на селективную ингибиторную активность в отношении PDE10 может быть использовано более одной культуры медиальных шиповатых нейронов и/или могут быть использованы аликвоты одной культуры медиальных шиповатых нейронов. Каждое из этих исследований может включать в себя одну или несколько автоматизированных, например компьютеризированных, стадий.
Фосфорилирование CREB в культуре(ах) шиповатых нейронов среднего размера может быть измерено способами, хорошо известными специалистам в данной области. Например, фосфорилирование CREB может быть измерено Вестерн-блоттингом белковой смеси, полученной в результате гомогенизирования обрабатываемой культуры шиповатых нейронов среднего размера, с антителами, специфичными в отношении CREB. Комплекс антитело-CREB может быть измерен в соответствии с одним или несколькими известными способами, например с использованием вторичных флуоресцентно-меченных антител, антител, меченных радиактивной меткой или антител, меченных ферментом или фермент-субстратом.
GABA в культуре(ах) медиальных шиповатых нейронов может быть измерена способами, известными специалисту в данной области. Например, нейроны в культуре медиальных шиповатых нейронов сначала могут быть обнаружены с помощью одного из нескольких известных ядерных красителей и тубулина для идентификации клеток, в которых происходит процесс. Вслед за этим для определения нейронов, экспрессирующих GABA, могут быть использованы флуоресцентно-меченные антитела. Число нейронов, экспрессирующих GABA, может быть определено любой автоматизированной системой или визуально. Могут быть использованы системы обработки нефлуоресцентного изображения, включая, например, но ими не ограничиваясь, изображения, полученные с помощью радиомеченных GABA-специфических антител. В качестве другого способа, обрабатываемая культура медиальных шиповатых нейронов может быть гомогенизирована и GABA в ней может быть количественно определена любым известным способом, включая, но ими не ограничиваясь, ВЭЖХ, ELISA или ферментативную реакцию.
Краткое описание чертежей
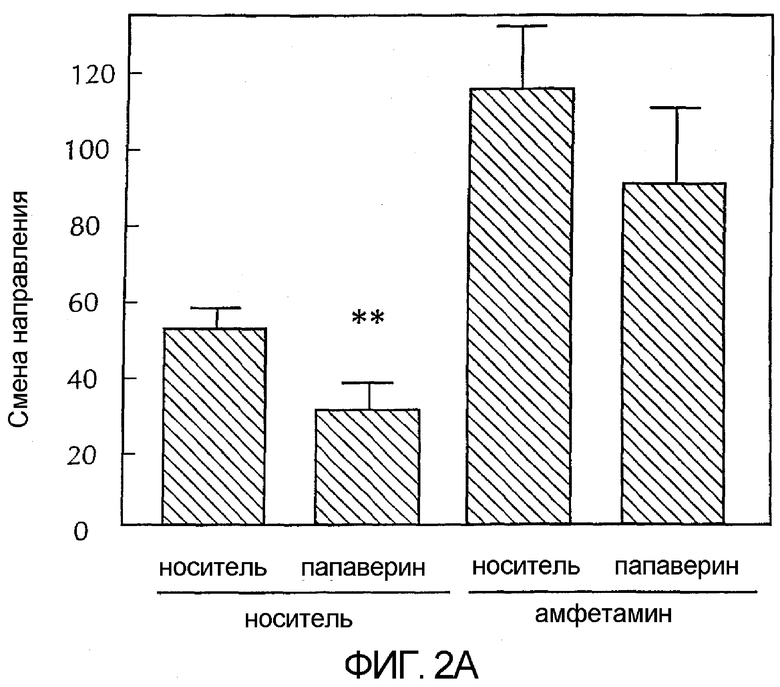
Фигура 1 - гистограмма, демонстрирующая каталепсию у животных в зависимости от возрастающих доз папаверина. Серые полосы представляют собой папаверин в сочетании с галоперидолом и показывают усиление папаверином вызванной галоперидолом каталепсии. Черные полосы представляют собой только папаверин. Эти черные полосы показывают, что папаверин самостоятельно не вызывает каталепсию в дозах до 32 мг/кг. Более конкретно, папаверин вводили в указанных дозах либо в чистом виде, либо с галоперидолом (0,32 мг/кг) за 30 минут до исследования. Каждая полоса обозначает средний латентный период до снятия передних лап с приподнятого барьера у шести сходно обрабатываемых животных. Для сравнения упорядоченных данных латентного периода папаверина в чистом виде против папаверина с галоперидолом использовали анализ изменения по Крускаллу-Уоллису. Post-hoc-анализ показал, что у животных, которым вводили 3,2, 10 и 32 мг/кг папаверина плюс галоперидол, длинный латентный период (**) был значительно больше, чем у животных, которым вводили только галоперидол.
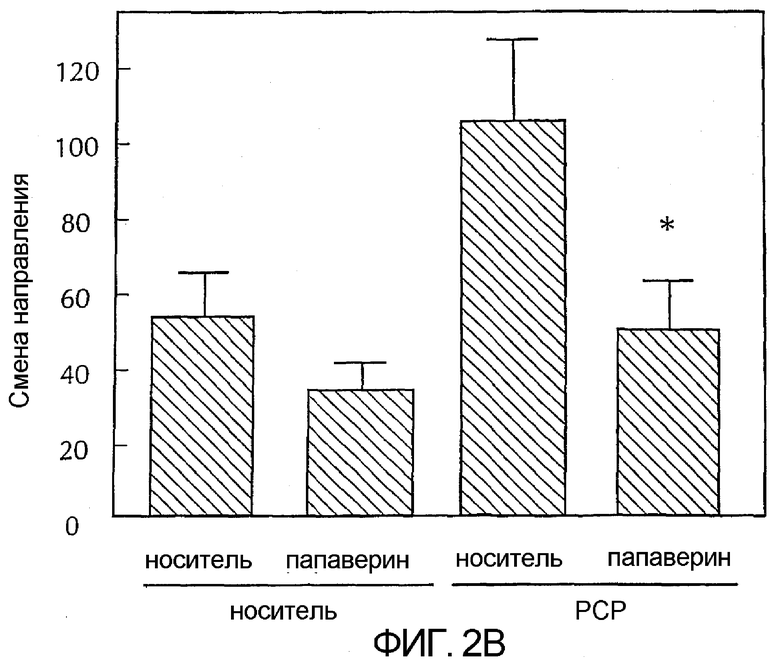
Фигура 2 - две графические полосы, каждая из которых показывает среднее + SEM число изменений направления у животных в эксперименте с челночной коробкой в течение первых 60 минут после введения препарата. На верхней диаграмме сравниваются эффекты папаверина, вводимого в чистом виде, на двигательную активность с эффектами папаверина на двигательную активность, вызванную амфетамином. На нижней диаграмме сравниваются эффекты папаверина, вводимого в чистом виде, на двигательную активность с эффектами папаверина на двигательную активность, вызванную PCP. Амфетамин вводили с дозой 1 мг/кг, внутрибрюшинно. PCP вводили с дозой 3,2 мг/кг, внутрибрюшинно. Папаверин вводили вместе с каждым препаратом с дозой 32 мг/кг, внутрибрюшинно. Данные показывают среднее + SEM число изменений направления в первые 60 минут после введения препарата у n = 8 крыс/группа.
** p < 0,01 по сравнению с контролем носитель/носитель;
* p < 0,05 по сравнению с носителем/PCP по критерию Стьюдента.
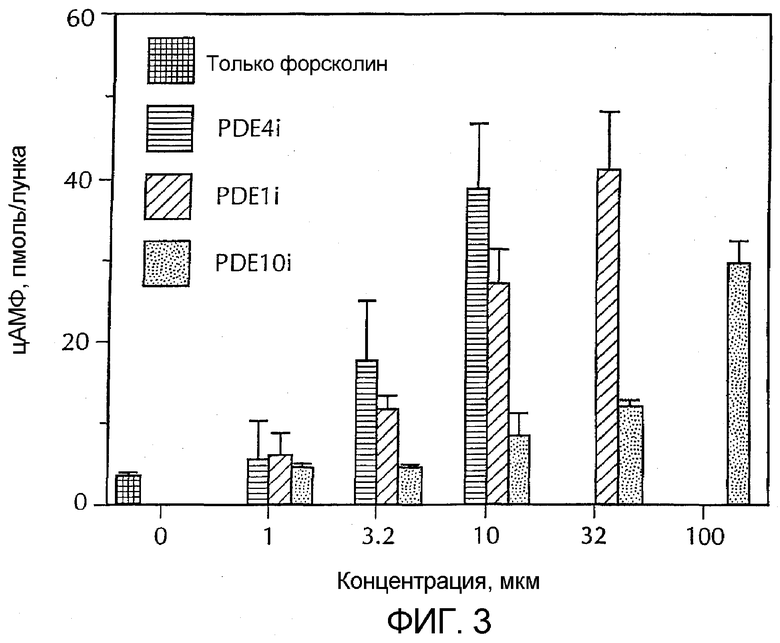
Фигура 3 - концентрация цАМФ в культуре медиальных шиповатых нейронов, стимулированных форсколином. Также показан эффект селективного ингибитора PDE 10, селективного ингибитора PDE 1B и селективного ингибитора PDE 4 на концентрацию цАМФ в стимулированных нейронах.
Фигура 4 - концентрация цГМФ в культуре медиальных шиповатых нейронов, стимулированных SNAP. Также показан эффект селективного ингибитора PDE 10, селективного ингибитора PDE 1B и селективного ингибитора PDE 4 на концентрацию цГМФ в стимулированных нейронах.
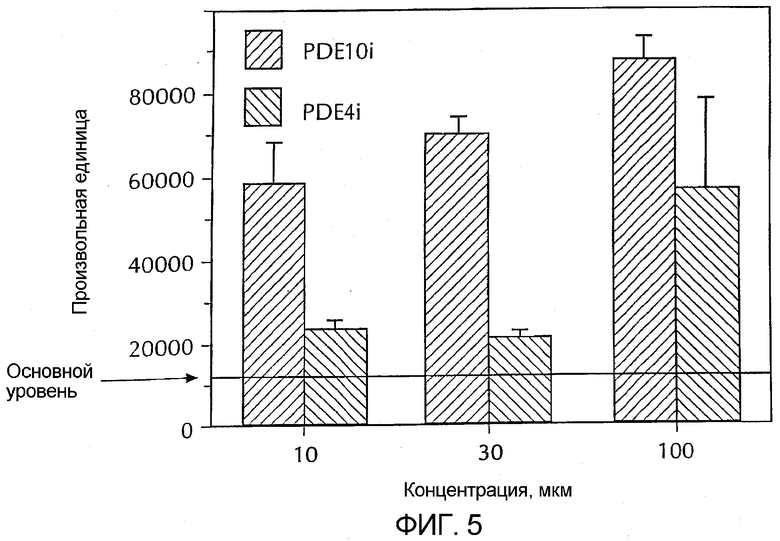
Фигура 5 - сравнение относительного эффекта селективного ингибитора PDE 10 и ролипрама (селективный ингибитор PDE 4) на фосфорилирование CREB (цАМФ-зависимый связывающий белок) в культуре медиальных шиповатых нейронов. Количество фосфорилированного CREB определяли с помощью Вестерн-блота.
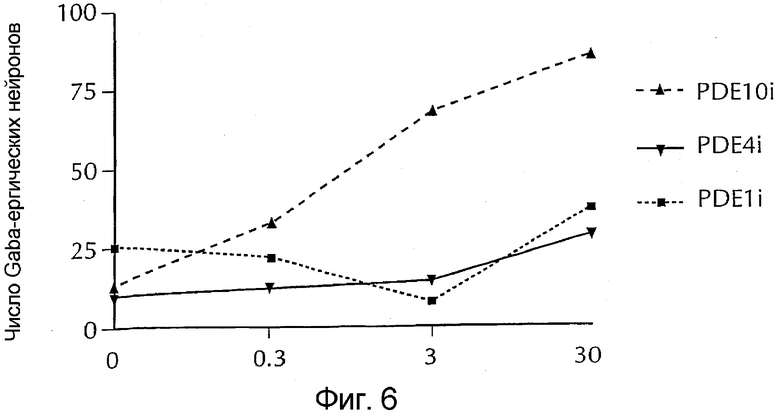
Фигура 6 - относительное число GABA-позитивных медиальных шиповатых нейронов в сравнении с нейронами, обработанными селективным ингибитором PDE 10, селективным ингибитором PDE 4 (ролипрам) и селективным ингибитором PDE 1B.
Подробное описание изобретения
В настоящем изобретении авторы идентифицируют селективный ингибитор PDE10, используют этот и сходные селективные ингибиторы PDE10 для определения ингибиторов PDE10, обладающих характеристиками и уникальным эффектом на метаболизм циклических нуклеотидов в популяции нейронов, высокоэкспрессирующих PDE10, медиальных шиповатых нейронов полосатого ядра. Эти ингибиторы также увеличивают фосфорилирование фактора регуляции транскрипции цАМФ-зависимого связывающего белка (CREB) в этих нейронах. Фосфорилирование CREB связано с изменением транскрипции различных генов, что, в свою очередь, имеет функциональные последствия, которые включают в себя, но ими не ограничиваются, эффекты на выживание нейронов и дифференцировку, и изменяет симпатическую организацию, что влияет на увеличение времени действия препарата. Заявители в настоящем изобретении описывают ингибиторы PDE10, которые обладают такими эффектами на медиальные шиповатые нейроны, то есть стимулируют дифференцировку этих нейронов до фенотипа GABA. Более того, авторы описывают ингибиторы PDE10, которые оказывают функциональное влияние на центральную нервную систему интактных млекопитающих. Конкретно, описывают ингибиторы PDE10, которые усиливают каталепсию у крыс, вызванную антагонистом рецептора допамина D2 галоперидолом, но не вызывают каталепсию, при самостоятельном введении с той же дозой. Ингибиторы PDE10 также ингибируют гиперлокомоцию, вызванную антагонистом рецептора NMDA фенциклидином. Эти данные говорят о том, что ингибиторы PDE10 воздействуют на центральную нервную систему и могут быть терапевтически использованы для лечения расстройств центральной нервной системы, перечисленных в формуле изобретения.
PDE 2, 3 и 5, изозимы, включая PDE человека, могут быть, например, получены из трабекул пещеристых тел; PDE1, изозимы, включая человеческий, из желудочка сердца; и PDE4, изозимы, включая человеческий, из скелетных мышц. PDE6 может быть получен, например, из сетчатки собаки. Способ получения ферментов из нативной ткани описан, например, Boolell, M. et al., Int. J. Impotence Research 8:7-52, 1996, что приведено здесь в качестве ссылки.
PDE 7-11 могут быть получены аналогичным образом из нативной ткани. Альтернативно, изозимы семейств PDE 7-9 и 11 могут быть получены из рекомбинантных клонов первичного транскрипта человека, трансфицированных, например, в клетки SF9, как описано Fisher, D.A., et al., Biochem. Biophys. Res. Comm. 246, 570-577, 1998; Soderling, S.H. et al., PNAS 96: 7071-7076, 1999; Fisher, D.A. et al., J. Biol. Chem. 273, 15559-15564, 1998b; и Fawcett, L., et al., PNAS 97: 3702-3707, 2000; соответственно. PDE10 может быть также получен из рекомбинатного клона крысы, трансфицированного в клетки SF9 (Fujishige et al., European Journal of Biochemistry, Vol. 266, 1118-1127 (1999)). Затем ферменты получали с помощью ЖХБР из растворимой фракции клеточных лизатов, как описано для PDE6. Вышеуказанные ссылки приведены здесь в качестве ссылки в полном объеме.
В одном из способов вещество скринировали на ингибирование гидролиза циклического нуклеотида с помощью PDE10 и PDE других семейств генов. В этом способе концентрация субстрата циклического нуклеотида каждого отдельного PDE составляла 1/3 от концентрации Km, что давало возможность сравнения значений IC50 различных ферментов. Активность PDE измеряли с помощью способа на основе сцинциляционного анализа (SPA), как описано ранее (Fawcett et al., 2000). Эффект ингибиторов PDE определяли, исследуя постоянное количество фермента (PDE 1-11) в присутствии различных концентраций субстрата и низкой концентрации субстрата, так, чтобы IC50 был приблизительно равен Ki (цГМФ или цАМФ с отношением 3:1 немеченого к меченому [3H] с концентрацией 1/3 Km). Конечный объем в анализе доводили до 100 мкл буфером для анализа [20 мМ Tris-HCl pH 7,4, 5 мМ MgCl2, 1 мг/мл альбумин бычьей сыворотки]. Взаимодействие инициировали ферментом, инкубировали в течение 30-60 минут при 30°C с получением <30% метаболизма субстрата и заканчивали добавлением 50 мкл SPA бус из силиката иттрия (Amersham) (содержащий 3 мМ соответствующего немеченого циклического нуклеотида для PDE 9 и 11). Плашки закрывали и трясли в течение 20 минут, после чего бусы оставляли отстаиваться в течение 30 минут в темноте и затем подсчитывали на счетчике для плашек TopCount (Packard, Meriden, CT). Единицы радиоактивности могут быть преобразованы в процент активности неингибированного контроля (100%), представленного против концентрации ингибитора, и значения IC50 ингибитора могут быть получены с использованием «Эмпирической кривой» с разрешением Microsoft Excel.
Одним из примеров селективных ингибиторов PDE10 является папаверин (1-[(3,4-диметоксифенил)метил]-6,7-диметоксиизохинолин). Папаверин известен как эффективный релаксант гладких мышц, использующийся для лечения спазма сосудов мозга и сердца, а также для лечения эректильной дисфункции. Хотя механизм этой терапевтической активности до конца не понятен, обычно папаверину приписывают активность неселективного ингибитора фосфодиэстеразы (The Pharmacological Basis of Therapeutics; Sixth Edition; A.G. Gilman, L.S. Goodman, A. Gilman (eds.) Macmillan Publishing Co., New York, 1980, с. 830). Хотя папаверин представляет собой природный растительный алкалоид, его полный биосинтез описан, например, в Brochmann-Hanssen et al., J. Pharm. Sci. 60:1672, 1971, как приведено здесь в качестве ссылки.
Селективный ингибитор PDE10 может вводиться по настоящему изобретению либо самостоятельно, либо в сочетании с фармацевтически приемлемыми носителями, в виде одной дозы или множественными дозами. Подходящие фармакологические носители включают в себя инертные твердые разбавители или наполнители, стерильные водные растворы и различные органические растворители. Фармацевтические композиции, полученные таким образом, могут затем легко вводиться в различных дозируемых формах, таких как таблетки, порошки, лепешки, сиропы, растворы для инъекций и тому подобное. Эти фармацевтические композиции могут, в случае необходимости, содержать дополнительные ингредиенты, такие как ароматизаторы, связующие агенты, эксципиенты и тому подобное.
Таким образом, для перорального введения могут использоваться таблетки, содержащие различные эксципиенты, такие как цитрат натрия, карбонат кальция и фосфат кальция, вместе с различными дезинтегрирующими агентами, такими как крахмал, метилцеллюлоза, альгиновая кислота и некоторые кремниевые комплексы, и связующими агентами, такими как поливинилпирролидон, сахароза, желатин и гуммиарабик. Кроме того, также часто при получении таблеток используют смазывающие агенты, такие как стеарат магния, лаурилсульфат натрия и тальк. Твердые композиции сходного типа также могут использоваться в виде наполнителей в мягких и твердых наполняемых желатиновых капсулах. Предпочтительными веществами для этой цели является лактоза или молочный сахар и высокомолекулярные полиэтиленгликоли. Если для перорального введения желательны водные суспензии или эликсиры, то основной активный ингредиент в них может сочетаться с различными подсластителями или ароматизаторами, красящим веществом или красителями и, в случае необходимости, эмульгирующими или суспендирующими агентами, вместе с разбавителями, такими как вода, этанол, пропиленгликоль, глицерин и их сочетания.
Для парентерального введения могут использоваться растворы, содержащие селективные ингибиторы PDE10 в кунжутном или арахисовом масле, водном пропиленгликоле или в стерильном водном растворе. Такие водные растворы, в случае необходимости, могут быть подходящим образом забуферены, а жидкий растворитель изотонизирован с помощью подходящего солевого раствора или глюкозы. Эти конкретные водные растворы особенно подходят для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. Используемую стерильную водную среду можно с легкостью получить стандартными способами, известными специалисту в данной области.
Селективный ингибитор PDE10 может вводиться в терапевтических способах по изобретению перорально, чрезкожно (например, при использовании пластыря), парентерально (например, внутривенно), ректально или местно. В целом, дневная доза ингибитора PDE10 для лечения расстройства или состояния в соответствии с описанными здесь способами обычно находится в области от около 0,01 до около 100 мг/кг массы тела пациента, которому вводят ингибитор PDE10. В качестве примера, селективный ингибитор PDE10 может вводиться для лечения, например, психического расстройства или болезни Гентингтона, взрослому больному со средним весом (около 70 кг) в дозе в области от около 1 мг до около 7000 мг в сутки, предпочтительно от около 1 мг до около 1000 мг в сутки, одной порцией или порцией, разделенной на несколько частей (то есть множественной порцией). Варьирования дозы на основании вышеуказанной области дозирования могут быть сделаны врачом данной области с учетом известных параметров, таких как масса, возраст, состояние пациента, подвергающегося лечению, серьезности заболевания и конкретного выбранного пути введения.
Настоящее изобретение иллюстрируют нижеследующие примеры. Тем не менее, понятно, что настоящее изобретение, подробно описанное здесь и в формуле изобретения, не ограничивается деталями следующих примеров.
ПРИМЕРЫ
Пример 1. Селективные ингибиторы PDE10: Папаверин
Папаверин скринировали на ингибирование гидролиза циклических нуклеотидов ферментом PDE10 и различными PDE других семейств генов. Концентрация субстрата циклических нуклеотидов в используемом анализе для каждой отдельной PDE составляла 1/3 от концентрации Km. Это давало возможность сравнивать значения IC50 различных ферментов.
Активность PDE измеряли, используя анализ с бусами SPA из силиката иттрия, описанного выше в подробном описании. Единицы радиоактивности преобразовывали в процент активности неингибированного контроля (100%), представленного против концентрации ингибитора, и получали значения IC50 ингибитора, используя «Эмпирическую кривую» с разрешением Microsoft Excel.
Было отмечено, что папаверин был особенно эффективным конкурентным ингибитором PDE10 при значении IC50, равном 18 нМ (таблица 1). Папаверин был значительно менее эффективен в отношении всех других исследуемых PDE. После PDE10 ферментом, который папаверин ингибировал наиболее сильно, был PDE4D, у которого значение IC50 составляло 320 нМ, что в 19 раз меньше, чем IC50 PDE10. Таким образом, эти данные впервые показали, что папаверин является селективным ингибитором PDE10 и что это соединение может использоваться при исследовании физиологии этого фермента.
(IC50/IC50 PDE10)
Пример 2. Эффекты селективного ингибитора PDE10 на метаболизм циклических нуклеотидов в шиповатых нейронах среднего размера
Был рассмотрен эффект папаверина, селективного ингибитора PDE10, что было определено в примере 1, на метаболизм циклических нуклеотидов в первичной культуре шиповатых нейронов среднего размера крыс.
Нейроны культивировали из полосатого тела эмбрионов крыс E17 в присутствии BDNF, проявляющих фенотип, очень сходный с фенотипом, описанным ранее (Ventimiglia et al., Eur. J. Neurosci. 7 (1995) 213-222). Приблизительно 50% этих нейронов окрашивались положительно на GABA-иммуннореактивность, что подтверждало присутствие шиповатых нейронов среднего размера в культурах. Экспрессия PDE-10 транскрипта в этих культурах при 4-6 DIV была подтверждена анализом защиты от РНКазы.
Культуры клеток полосатого тела получали, как описано ранее (Ventimiglia et al., Eur. J. Neurosci. 7: 213-222, 1995). Кратко, полосатое тело (хвостатое ядро и скорлупа), препарированное у крыс E17, отделяли для продукции суспензии единичных клеток и помещали с плотностью 5x104 нейронов/лунка в мультилуночные плашки, покрытые поли-L-орнитином/ламинином. Клетки помешали в среду Neurobasal с добавлением B27 и BDNF (100 нг/мл). Эксперименты обычно проводили через 4 дня in vitro. Шиповатые нейроны среднего размера составляли основную часть клеток этих культур (50-60%, что было подтверждено GABA-иммунореактивностью).
Для анализа защиты от РНКазы, РНК получали из этих первичных культур шиповатых нейронов среднего размера крыс центрифугированием при 150000 x g при 20°C в течение 21 часа с градиентом 5,7 M хлорида цезия, как ранее описано (Iredale, PA, et al., Mol. Pharmacol.50: 1103-1110, 1996). Осадок РНК ресуспендировали в 0,3 M ацетате натрия, pH 5,2, осаждали этанолом и концентрацию определяли спектрофотометрией. Зонд для РНК гибридизации PDE10 получали ПЦР-амплификацией 914 п.н. фрагмента, выделенного из кДНК мыши (соответствует 380-1294 п.н.). Этот фрагмент затем клонировали в pGEM3Zf. Вектор линеаризировали и для синтеза [32P]-меченого антисмыслового зонда для РНК гибридизации использовали РНК-полимеразу Т7. Анализ защиты от РНКазы осуществляли, используя набор RPAII (Ambion). Кратко, 5 мкг всего клеточного РНК гибридизировали с [32P]-меченым PDE10 зондом для РНК гибридизации (˜105 число импульсов в минуту/образец) в течение ночи при 42°C. На следующий день образцы инкубировали с РНКазой A и T1 в течение 30 минут при 37°C и защищенные двухнитевые фрагменты РНК затем осаждали и прогоняли на 6% полиакриламидном геле, содержащем мочевину.
Для оценки эффектов папаверина на циклические нуклеотиды клетки культуры полосатого тела через четыре дня in vitro промывали физиологическим раствором с фосфатным буфером без Ca2+/Mg+ и преинкубировали в течение четырех часов в физиологическом растворе с фосфатным буфером без Ca2+/Mg+ 30 мМ HEPES, CaCl2 1 мМ, декстрозу 1 мг/мл и MgCl2 5мМ. На клетки полосатого ядра воздействовали ингибиторами фосфодиэстеразы и инкубировали в течение двадцати минут при 37 градусах Цельсия. Затем измеряли цГМФ, нейроны стимулировали нитропруссидом натрия, источником окиси азота, в течение двух минут, а затем 20 минут инкубировали с соединением. Затем измеряли цАМР, нейроны стимулировали форсколином, активатором аденилатциклазы в течение двадцатиминутной инкубации с соединением. Клетки лизировали, используя сочетание 9:1 буфера для анализа цАМР SPA прямого скрининга (0,05 M ацетат с 0,01% азидом натрия) и буфера A (133 мг/мл бромид додецилтриметиламмония), и лизаты замораживали в сухом льду. Для определения концентрации соответствующего циклического нуклеотида в клеточном лизате использовали цГМФ [I125] или цАМФ [I125] систему сцинциляционного анализа (SPA) (Amersham код RPA 540 и RPA 559, соответственно).
Папаверин самостоятельно не давал измеримых изменений основного уровня либо цАМР, либо цГМФ в культуре клеток полосатого тела. Следовательно, были исследованы эффекты соединения в условиях, когда синтез цАМФ или цГМФ стимулировали форсколином или донором NO, нитропруссидом натрия (SNP), соответственно. Стимулирование культур форсколином (0,1-10 мкМ) в течение 20 минут приводило к повышению уровней цАМФ в зависимости от концентрации. Аналогично, краткое воздействие на культуры SNP (3-1000 мкМ) в течение 2 минут приводило к повышению уровней цГМФ в зависимости от концентрации. Форсколин, введенный в чистом виде (10 мкМ), не влиял на концентрации цГМФ, а SNP (300 мкМ) не увеличивал уровни цАМФ. Для определения эффектов папаверина на метаболизм цАМФ и цГМФ, культуры клеток полосатого тела инкубировали с различными концентрациями соединения и затем стимулировали с субмаксимальными эффективными концентрациями либо форсколина (1 мкМ), либо SNP (100 мкМ). Эти концентрации форсколина или SNP вызывали 2-3-кратное увеличение уровня цАМФ и цГМФ по сравнению с основным уровнем, соответственно. Папаверин вызывал увеличение накопления цГМФ при стимулировании SNP в зависимости от концентрации со значением EC200 (концентрация ингибитора, повышающая уровень в 2 раза), равным 11,7 мкМ (таблица 2). Максимальный эффект наблюдался при 100 мкМ, при котором уровни цГМФ повышались в 5 раз по сравнению с уровнем цГМФ культуры, стимулированной только SNP. Папаверин также вызывал увеличение накопления цАМФ в культурах, стимулированных форсколином. Однако соединение было в 3,3 раза менее эффективно при активации повышения цАМФ, чем цГМФ. Эффекты папаверина в культурах клеток полосатого тела сравнивали с другими ингибиторами PDE, обладающими различной селективностью (таблица 2). Неселективный ингибитор IBMX вызывал повышение накопления как цГМФ, так и цАМФ в культурах, стимулированных SNP или форсколином, в зависимости от концентрации (3-100 мкМ) со значениями EC200 19 и 30 мкМ, соответственно. Селективный ингибитор PDE4 ролипрам повышал накопление цАМФ при стимулировании форсколином со значением EC200, равным 2,5 мкМ, и для двухкратного увеличения накопления цГМФ требовалось 10-кратное увеличение его концентраций. Ингибитор PDE, активный в отношении цГМФ запринаст, удваивал уровни цАМФ в этих нейронах при концентрации 98 мкМ. Однако 100 мкМ этого соединения не совсем вдвое увеличивали уровень цГМФ. Эти данные впервые показали, что папаверин обладает уникальным эффектом в отношении регуляции циклических нуклеотидов шиповатых нейронов среднего размера и что этот эффект обусловлен селективностью в отношении PDE10.
Пример 3. Эффект селективного ингибитора PDE 10 на животной модели функционирования базальных ганглиев
Изучение у млекопитающих и человека показало, что базальные ганглии регулируют уровень моторной активности, а также когнитивного и эмоционального/пищевого поведения (Graybiel, A.M. Current Biology 10 (14):R509-11, 2000). Были разработаны экспериментальные модели грызунов, которые можно использовать для оценки эффектов соединений на функцию базальных ганглиев. Было обнаружено, что папаверин обладает ранее неизвестными уникальными свойствами на поведенческие реакции в двух таких моделях.
Эффект папаверина, вводимого самостоятельно и в сочетании с галоперидолом исследовали на способность индуцировать каталепсию у самцов крыс CD. Эта животная модель использовалась для анализа эффектов соединений на эффективность базального ганглия. Папаверин (1,0, 3,2, 10 или 32 мг/кг) или носитель вводили подкожно. В нескольких экспериментах вслед за ним немедленно вводили галоперидол. Через тридцать минут после введения препарата(ов) количественно измеряли степень каталепсии, помещая передние лапы животных на приподнятый (10 см) брусок (1 см диаметром), и определяли латентный период до снятия обеих передних лап с бруска с порогом латентного периода в 30 с. Латентные периоды упорядочивали в пределах каждой обрабатываемой группы для сравнения с помощью анализа изменения по Крускаллу-Уоллису. Post-hoc-анализ проводили с помощью U теста Манн-Витнея.
Как было описано ранее (Chartoff, E et al., J Pharmacol. Exp. Ther. 291:531-537, 1999), антипсихотический агент галоперидол давал устойчивую каталепсию в этой модели. Было обнаружено, что максимально эффективная доза галоперидола составляла 1 мг/кг, п.к. Наоборот, папаверин не вызывал каталепсию при самостоятельном введении с дозой до 32 мг/кг, п.к. (p = 0,86). Однако, как показано на фигуре 1, папаверин усиливал каталептический эффект субмаксимальной дозы галоперидола (0,32 мг/кг, п.к. с 0,3% винной кислотой) (p < 0,001). Минимальная эффективная доза папаверина для усиления каталепсии, вызванной галоперидолом, составляет 3,2 мг/кг, п.к. Этот эксперимент показал, что папаверин может изменять функцию базального ганглия, что соответствует антипсихотическим характеристикам.
Пример 4. Эффект селективного ингибитора PDE 10 в животных моделях психоза
Далее был рассмотрен эффект папаверина на локомоторную активность крыс, измеряемую в челночной коробке. Известно, что снижение локомоции, стимулированной PCP, у грызунов является первым тестом первого уровня при поиске новых антипсихотических агентов. Новейшие нетипичные антипсихотические агенты обычно демонстрируют избирательное ингибирование локомоторной активности, стимулированной PCP, по сравнению с локомоторной активностью, стимулированной амфетамином. Взрослые самцы крыс Спрэгью-Даули (250-300 г) получали от Charles River (Wilmington, MA). Локомоторную активность оценивали по смене направления в коммерчески доступных челночных коробках (Coulbourn Instruments, Allentown, PA). Данные получали с 5-минутными интервалами в течение 1 часа после введения препарата. Животным вводили либо носитель (5% DMSO, 5% эмульфор, 90% физиологический раствор), либо фенциклидин (PCP, Sigma Chem. Co.), либо амфетамин сульфат (RBI), затем сразу вводили либо носитель, либо исследуемое соединение. Статистические анализы осуществляли с помощью критерия Стьюдента.
Оба психостимулятора, амфетамин и фенциклидин (PCP), давали значительное увеличение локомоторной активности в этих моделях. Папаверин, при самостоятельном введении (32 мг/кг, в.б.), давал небольшое уменьшение локомоторной активности, которая в некоторых исследованиях имела статистическую важность (фигура 2). Однако в тех же дозах папаверин давал значительное снижение локомоторной активности, стимулированное введением фенциклидина 3,2 мг/кг, в.б., не влияя на поведение, стимулированное введением такой же дозы амфетамина (1 мг/кг, в.б.).
В другом эксперименте, используя такой же анализ локомоторной активности, папаверин вводили одновременно с амфетамином (1 мг/кг, п.к.) или PCP (3,2 мг/кг, п.к.) и локомоторную активность измеряли в течение 30 минут. В этом эксперименте папаверин в обоих случаях эффективно ингибировал локомоторную активность, стимулированную амфетамином и PCP.
Результаты обоих этих экспериментов показали, что папаверин оказывает поведенческий эффект на локомоторную активность, что соответствует антипсихотическим характеристикам.
В примерах 5-7, ниже, определяли селективный ингибитор PDE10 и селективный ингибитор PDE1B в соответствии с анализом, описанным в подробном описании данного изобретения (на таблице 3 показан IC50 в мкМ селективного ингибитора PDE10 в отношении PDE 1, 2, 3, 4, 5, 7, 8, 9, 10 и 11):
Пример 5. Эффекты ингибиторов PDE на накопление цАМФ и цГМФ в шиповатых нейронах среднего размера
Культуры медиальных шиповатых нейронов получали, как описано в примере 2, из полосатого тела эмбрионов крыс E17 или E18. Полосатое тело гидролизировали трипсином и диссоциированные клетки помещали на плашки, покрытые поли-L-омитин/ламинином в среду Neurobasal, содержащую добавку B27. Для анализа продукции циклических нуклеотидов и фосфорилирования CREB к нейронам также добавляли 50 нг/мл BDNF (нейротрофический фактор головного мозга) и использовали при 6 DIV. При этом приблизительно 90% клеток имели нейрональную морфологию и 50% положительную окраску на GABA.
В культуре медиальных шиповатых нейронов было обнаружено, что селективные ингибиторы PDE10 и PDE1B и ролипрам (который селективен в отношении PDE4) усиливают повышение накопления цАМФ (фиг. 3) или цГМФ (фиг. 4), стимулированное форсколином или SNAP, соответственно. Однако не было заметных изменений уровней цАМФ или цГМФ при добавлении соединений в отсутствие раздражителей.
Ингибиторы PDE дифференцировали по эффективности увеличения уровней цАМФ по сравнению с уровнями цГМФ (таблица 4). В таблице 4 эффективность выражена как EC200, то есть как концентрация ингибитора PDE, которая повышает уровни цАМФ или цГМФ, индуцированные форсколином или SNAP, соответственно, на 200%.
Медиальные шиповатые нейроны, EC200, мкМ
Пример 6. Эффект ингибиторов PDE на фосфорилирование CREB в медиальных шиповатых нейронах
цАМФ и цГМФ активируют протеинкиназы PKA и PKG, соответственно. Оба типа киназ способны фосфорилировать регулятор транскрипции CREB. Изучались эффекты селективных ингибиторов PDE таблицы 3 на фосфорилирование CREB как дальнейшего события сигнального каскада циклических нуклеотидов.
Стимуляция форсколином давала устойчивое увеличение фосфорилирования CREB, что было измерено Вестерн-блоттингом. Селективный ингибитор PDE 10 и ролипрам также увеличивали фосфорилирование CREB, что было измерено Вестерн-блоттингом. Сравнение эффекта селективного ингибитора PDE 10 и ролипрама показано на фиг. 5. По упорядоченным в зависимости от эффективности повышения фосфорилирования CREB данным было определено, что действие форсколина > действия селективного ингибитора PDE 10 > действия ролипрама. Селективный ингибитор PDE 1B был неактивен в отношении увеличения фосфорилирования CREB.
Пример 7. Эффект ингибиторов PDE на дифференциацию медиальных шиповатых нейронов
Транскрипция, активированная вслед за фосфорилированием CREB, вовлечена в выживание и дифференциацию нейронов. Были проведены исследования, оказывают ли ингибиторы PDE таблицы 3 эффект на выживание и дифференциацию медиальных шиповатых нейронов. Эти эксперименты проводили в соответствии с протоколом, использованным Ventimiglia et al. (см. Ventimiglia et al., 1995, supra) для анализа эффективности BDNF на эти процессы в медиальных шиповатых нейронах. Более конкретно, ингибиторы PDE добавляли в культуру медиальных шиповатых нейронов во время их посева и затем измеряли при 6 DIV различных параметрах, связанных с нейрональным выживанием и дифференциацией, используя Array Scan System от Cellomics, Inc (Pittsburgh, PA, USA).
На основании исследованных параметров было обнаружено, что селективный ингибитор PDE 10 поразительно увеличивал число GABA-эргических нейронов. Клетки могут быть окрашены следующим образом: голубые ядра; зеленые нейроны; красные нейроны, окрашивались положительно на GABA. Селективный ингибитор PDE 10 действовал как BDNF, тогда как ролипрам и селективный ингибитор PDE 1B не оказывали никакого эффекта (фиг. 6).
Обсуждение
На основании гибридизации in situ ранее сообщалось о высокой экспрессии мРНК PDE10 в полосатом теле, прилежащем ядре и обонятельных бугорках (Seeger, T.F. Et al., supra). Используя моноклональные антитела в отношении белка PDE10 также в этих областях мозга был обнаружен соответствующий высокий уровень белка PDE10 (Menniti, F.S., Strick, C.A., Seeger, T.F., и Ryan, A.M., Immunihistochemical localization of PDE10 in the rat brain, supra). В медиальных шиповатых нейронах полосатого тела и прилежащем ядре был обнаружен высокий уровень экспрессии мРНК PDE10. Медиальные шиповатые нейроны являются выходными нейронами полосатого тела, прилежащего ядра и обонятельных бугорков и представляют собой приблизительно 95% всех нейронов этих мозговых структур. Более того, высокий уровень белка PDE10 наблюдали в проекциях (аксоны и окончания) медиальных шиповатых нейронов полосатого тела, прилежащего ядра и обонятельных бугорков в другие области мозга, включая бледные шары и черную субстанцию. Эти последние области мозга имеют низкие или неопределяемые уровни мРНК PDE10. Следовательно, высокий уровень белка PDE10 в этих областях возникает из аксонов и окончаний медиальных шиповатых нейронов. Кроме того, низкие уровни экспрессии мРНК PDE10 и белка обнаруживаются в нейронах других областей мозга, включая кору, гиппокамп и мозжечок.
Высокие уровни экспрессии PDE10 в полосатом теле и прилежащем ядре являются, в частности, интересным наблюдением, так как эти области являются главными кортикальными входными ядрами базальных ганглиев, а также основными концевыми областями допаминергическими проекциями среднего мозга. Полосатое тело и его вентральная протяженность, прилежащее ядро, получают глютаматергические афферентные нейроны фактически из каждой области коры головного мозга и действуют как субкортикальная интеграционная область большого разнообразия активности коры. Дорзальный отдел полосатого тела обычно рассматривается как отдел, вовлеченный в регуляцию моторной деятельности, тогда как вентральные отделы, включая прилежащее ядро, действуют на регуляцию эмоционального/пищевого поведения. Таким образом, вероятно, PDE10 участвует в сигнальных путях, регулирующих большое число этих основных физиологических процессов.
В действительности, было обнаружено, что ингибирование PDE10 влияет на метаболизм циклических нуклеотидов и сигнальный путь CREB в медиальных шиповатых нейронах, в отличие от эффектов, возникающих при ингибировании PDE 4 или PDE 1, других основных PDE, экспрессируемых этими нейронами. Также было обнаружено, что ингибиторы PDE10 действуют на функцию базальных ганглиев in vivo.
Каждый селективный ингибитор PDE10, 4 и 1 увеличивает накопление цГМФ и/или цАМФ в медиальных шиповатых нейронах, стимулированных SNAP или форсколином, соответственно (фиг. 3 и 4). Однако ингибиторы различаются по коэффициенту эффективности в отношении двух циклических нуклеотидов (таблица 3). Эти различия вероятно возникают в результате присущей аффиности PDE 10, 4 и 1B в отношении двух циклических нуклеотидов, а также разного доступа различных PDE к пулам циклических нуклеотидов. Примечательно, что эти ингибиторы не оказывают измеримых эффектов на уровни цАМФ и цГМФ в отсутствие стимуляции. Фосфорилирование CREB является одним из последствий при активировании сигнальных каскадов циклических нуклеотидов. Заявителями было показано, что селективный ингибитор PDE10 и селективный ингибитор PDE 4 увеличивают фосфорилирование CREB, при этом селективный ингибитор PDE 10 является более сильным и эффективным. Эти эффекты возникали тогда, когда соединения добавляли в отсутствие других стимуляторов и, следовательно, в отсутствие определяемых изменений уровней циклических нуклеотидов. Было показано, что селективный ингибитор PDE 1B является неактивным. Эти результаты показали, что PDE10 играет важную роль в сигнальных путях циклических нуклеотидов в медиальных шиповатых нейронах и, в частности, по видимому, PDE10 связан с регуляцией фосфорилирования CREB.
Определенные эффекты ингибирования PDE10, показанные в системах in vitro, соответствуют уникальным эффектам ингибирования PDE10 на функцию базальных ганглиев in vivo. Было показано, что селективный ингибитор PDE10 папаверин усиливает каталептический эффект антагониста допаминового рецептора D2 галоперидола, не вызывая его при самостоятельном введении. Более того, это соединение снижает локомоторную гиперактивность, вызванную антагонистом рецептора NMDA фенциклидином. Наличие такого фармакологического свойства папаверина показывает, что он и все ингибиторы PDE10 могут быть эффективны при лечении неврологических и психиатрических расстройств, при которых наблюдается дисфункция базального ганглия, как описано ниже.
Кортикальный сигнал полосатому телу дает первичный возбуждающий стимул GABA-эргическим медиальным шиповатым нейронам. Глютаматергическая активация медиальных шиповатых нейронов в свою очередь регулируется большим допаминергическим выбросом в срединный мозг. Антагонистическая природа этих двух афферентных систем была показана во многих исследованиях. Например, локомоторная активность у лабораторных животных может стимулироваться либо агонистами рецептора допамина, либо антагонистами глютаматного рецептора подтипа NMDA (Carlsson, M.L. and Carlsson, A. Trends Neurosci.13:272-276, 1990). Каталептический эффект антагонистов рецептора допамина D2, такого как галоперидол, снижается антагонистами рецептора NMDA, как генная экспрессия, индуцированная галоперидолом (Chartoff, E et al., J Pharmacol. Exp. Ther. 291:531-537, 1999). Недавно было показано, что блокада рецепторов допамина D2 приводит к увеличению фосфорилирования или к активации рецепторов NMDA полосатого тела (Leveque et al., Journal of Neuroscience 20(11):4011-4020, 2000).
Понимание того, что все клинически эффективные антипсихотические препараты обладают сильной антагонистической активностью D2, привело к оригинальной гипотезе, что симптомы шизофрении являются результатом чрезмерной активности мезолимбической допаминовой системы. Способность химического препарата снижать стимулирующие свойства прямого или непрямого агониста допамина является важнейшим лабораторным исследованием при поиске новых антипсихотических агентов. В последнее время способность антагонистов рецептора NMDA, таких как PCP, точно повторять положительные, отрицательные и когнитивные симптомы шизофрении у человека (Luby et al., 1959; Rosenbaum et al, 1959; Krystal et al. 1994) привела к созданию гипотезы гипофронтальности шизофрении. В двух словах, в соответствии с этой гипотезой ингибирование поведения, связанное с полосатым телом, оказывается недостаточным при шизофрении в результате пониженной глутаматергической и специфической, опосредованной рецептором NMDA, нейропередачи. Эта гипотеза полностью согласуется с известным антипсихотическим эффектом антагонистов рецептора допамина D2, способных дезингибировать напрямую или опосредовано сигнал в полосатое тело (что описано выше). Точность, с которой PCP имитирует симптомы шизофрении у человека, привела к использованию локомоторной активности, стимулированной PCP, у грызунов как теста первого уровня при исследовании новых антипсихотических агентов. Было показано, что новые и вероятно более эффективные нетипические антипсихотические агенты обладают активностью в отношении локомоторной активности, стимулированной PCP, по сравнению с активностью, стимулированной амфетамином, подтверждают этот подход (Gleason S.D. and Shannon H.E. Psychopharmacol. 129:79-84,1997).
Хотя используемые в настоящее время способы антипсихотической терапии обычно направлены на мембранные рецепторы, полагают, что внутриклеточные действия PDE10 в медиальных шиповатых нейронах также могут давать антипсихотический эффект. Известно, что увеличение активности цАМФ и PKA увеличивает ответ нейронов полосатого тела на антагонисты глютамата, включая NMDA (Colwell, C.S. and M.S. Levine, J Neuroscience 15(3)1704-1713, 1995). Нейролептическое действие галоперидола также связано с увеличением уровней цАМФ (Ward, R.P. and D.M. Dorsa, Neuroscience 89(3):927-938, 1999) и активности PKA (Adams, M.R. et al., Proc Natl Acad Sci USA 94:12157-12161, 1997). Уровни цГМФ нейронов полосатого тела также увеличиваются при блокаде рецепторов D2 (Altar, C. A. et al., Eur J. Pharmacol. 181:17-21, 1990), и известно, что PKG фосфорилируют некоторые последующие субстраты, такие как PKA, в том числе эндогенный ингибитор протеинфосфатазы I, DARP (Greengard P et al., Brain Res. Rev. 26:274-284, 1998). Следовательно, была выдвинута гипотеза, в соответствии с которой логично предполагать, что агенты, способные селективно увеличивать уровни циклических нуклеотидов в медиальных шиповатых нейронах полосатого тела, могут усиливать функции полосатого тела с возникновением антипсихотического эффекта и что ингибитор PDE10 может иметь терапевтическую эффективность при лечении психоза, так как такое соединение может ингибировать метаболизм цАМФ и цГМФ, катализируемый PDE10, увеличивая уровни этих циклических нуклеотидов в медиальных шиповатых нейронах.
Кроме психоза, патологическая функция базального ганглия вовлечена в различные нейропсихические состояния, включая синдром дефицита внимания/гиперактивности (ADHD) и сходные расстройства дефицита (Seeman, P. et al., Molecular Psychiatry 3:386-96, 1998), депрессию (Kapur, S., Biol. Psychiatry 32:1-17, 1992; Willner, P., Brain Res. 287:225-236, 1983), обсессивно/компульсивные расстройства, включая синдром Туретта и другие тики (Graybiel AM. Rauch SL. Toward a neurobiology of obsessive-compulsive disorder. Neuron. 28(2):343-7, 2000) и злоупотребление алкоголем или наркотиками (Self, D.W. Annals of Med. 30:379-389, 1998). Некоторые расстройства, включая болезнь Паркинсона, синдром «беспокойных ног» (Hening, W. et al., Sleep 22:970-999, 1999) и болезнь Гентингтона (Vonsattel JP et al., Neuropathological classification of Huntington's disease. J. Neuropathol. Exp. Neurol. 44:559-577. 1985) также связаны с дисфункцией базального ганглия. Таким образом, на основании исследований, описанных здесь, полагают, что ингибитор PDE10 может иметь терапевтическое воздействие на такие расстройства.
Фосфорилирование CREB вызывает транскрипцию различных генов, которые по-разному влияют на нейрональную функцию, в том числе увеличивая выживание и/или дифференциацию нейронов. Было обнаружено, что селективные ингибиторы PDE10 могут увеличивать дифференциацию медиальных шиповатых нейронов до GABA-эргического фенотипа (фиг. 6). Ролипрам (селективный ингибитор PDE4) и селективный ингибитор PDE 1B не демонстрировали такой активности (фиг. 7).
Эффект ингибирования PDE10 на фосфорилирование CREB является чрезвычайно важным в отношении лечения нейродегенеративных состояний, таких как болезнь Гентингтона.
Также фосфорилирование CREB в медиальных шиповатых нейронах и дифференциация медиальных шиповатых нейронов до GABA-эргического фенотипа являются эффективными для идентификации соединений, имеющих активность селективных ингибиторов PDE 10.
Приведенные здесь данные показывают уникальную роль PDE10 в дифференциации и/или выживании медиальных шиповатых нейронов. Эти нейроны выборочно уязвимы при болезни Гентингтона и было сделано предположение, что этот факт может быть результатом потери трофической поддержки этих нейронов (Zuccato et al., Loss of Huntingtin-mediated BDNF gene transcription in Huntington's disease. Science. 293:493-498, 2001). Был сделан вывод, что селективный ингибитор PDE 10 обладает нейротрофической активностью в отношении медиальных шиповатых нейронов. Более того, был сделан вывод, что ингибиторы PDE 10 вероятно обладают нейротрофической активностью в отношении любых нейронов, экспрессирующих PDE 10, и что ингибиторы PDE 10, таким образом, эффективны для лечения нейродегенеративных заболеваний, включая, но ими не ограничиваясь, нейродегенеративные заболевания, представленные здесь.
Наконец, мРНК PDE10 и белок экспрессируются также в нейронах гипокампа и коры головного мозга. Так как когнитивные процессы связаны с функцией гипокампа и коры, предполагают, что PDE10 также играют роль в когнитивных процессах и что ингибитор PDE10 также может быть эффективен для лечения расстройств с характерным компонентом нарушения внимания и/или когнитивной функции, таких как болезнь Альцгеймера и возрастное снижение когнитивных функций (ARCD).
| название | год | авторы | номер документа |
|---|---|---|---|
| СОЛЬ СУКЦИНАТ 2-((4-(1-МЕТИЛ-4-(ПИРИДИН-4-ИЛ)-1Н-ПИРАЗОЛ-3-ИЛ)ФЕНОКСИ)МЕТИЛ)ХИНОЛИНА | 2007 |
|
RU2430918C2 |
| ОРГАНИЧЕСКИЕ СОЕДИНЕНИЯ | 2014 |
|
RU2679142C2 |
| ХИНОЛИНОВЫЕ ПРОИЗВОДНЫЕ КАК ИНГИБИТОРЫ ФЕРМЕНТА PDE10A | 2012 |
|
RU2624440C2 |
| ПОЛИПЕПТИД, ИМЕЮЩИЙ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ ФОСФОДИЭСТЕРАЗЫ | 1998 |
|
RU2272841C2 |
| ОРГАНИЧЕСКИЕ СОЕДИНЕНИЯ | 2015 |
|
RU2709786C2 |
| КОМБИНАЦИИ, СОДЕРЖАЩИЕ ИНГИБИТОРЫ PDE 2, ТАКИЕ КАК 1-АРИЛ-4-МЕТИЛ-[1,2,4]ТРИАЗОЛО[4,3-А]ХИНОКСАЛИНОВЫЕ СОЕДИНЕНИЯ, И ИНГИБИТОРЫ PDE 10, ДЛЯ ПРИМЕНЕНИЯ В ЛЕЧЕНИИ НЕВРОЛОГИЧЕСКИХ ИЛИ МЕТАБОЛИЧЕСКИХ РАССТРОЙСТВ | 2013 |
|
RU2657540C2 |
| СОЕДИНЕНИЯ И СПОСОБЫ | 2015 |
|
RU2711442C2 |
| НОВЫЙ ПЕПТИД-ИНГИБИТОР PI3Kγ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ОРГАНОВ ДЫХАНИЯ | 2015 |
|
RU2704826C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПОВЫШЕНИЯ СОДЕРЖАНИЯ И ДОСТУПНОСТИ ЦИКЛИЧЕСКОГО АДЕНОЗИНМОНОФОСФАТА В ОРГАНИЗМЕ И ЕЕ ПОЛУЧЕНИЕ | 2012 |
|
RU2625765C2 |
| ТРАНСГЕННОЕ ЖИВОТНОЕ, ОТЛИЧНОЕ ОТ ЧЕЛОВЕКА, И ЕГО ПРИМЕНЕНИЯ | 2010 |
|
RU2579701C2 |

Настоящее изобретение относится к лечению определенных неврологических и психических расстройств у млекопитающих, включая человека, которое включает в себя введение селективного ингибитора PDE10, в частности настоящее изобретение относится к способам идентификации химического соединения, которое обладает эффективностью селективного ингибитора PDE10. Технический результат: разработка нового способа скрининга соединений на наличие способности селективно ингибировать PDE10. 2 н.п. ф-лы, 6 ил., 4 табл.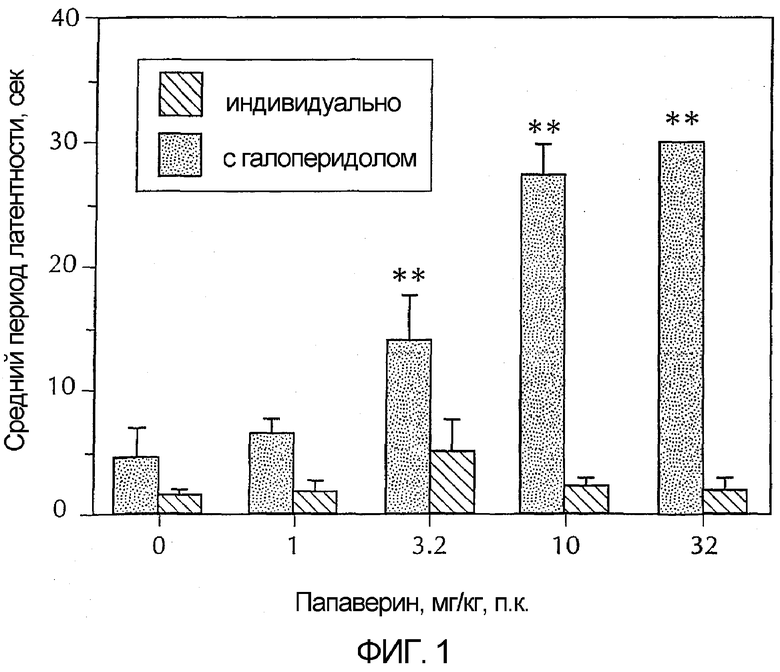
a) введение химического соединения в культуру медиальных шиповатых нейронов; и
b) измерение возможного повышения фосфорилирования CREB в культуре; повышение фосфорилирования CREB, тем самым, показывает, что соединение, использованное в стадии (а) обладает селективной ингибиторной активностью в отношении PDE10.
a) введение химического соединения в культуру медиальных шиповатых нейронов; и
b) измерение возможного повышения количества продуцируемого GABA медиальными шиповатыми нейронами в указанной культуре; повышение продукции GABA указанными медиальными шиповатыми нейронами, тем самым, показывает, что соединение, использованное в стадии (а) обладает селективной ингибиторной активностью в отношении PDE10.
| Fujishige К | |||
| et al | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| - European Journal of Biochemistry, 12.1999; Vol.266, no 3, p.1118-1127, abstract | |||
| DE 3224100 A1, 29.12.1983 | |||
| ЛЕКАРСТВЕННЫЙ ПРЕПАРАТ ДЛЯ НЕЙРОПРОТЕКЦИИ (ВАРИАНТЫ) | 1994 |
|
RU2135162C1 |
| Soderling S.H | |||
| et al | |||
| Isolation and characterization of a dual-substrate phosphodiesterase | |||

