Изобретение относится к области водной токсикологии и санитарной гидробиологии, а именно к способам экологического контроля качества поверхностных пресных вод с помощью методов биотестирования. Предлагаемый способ может быть использован для диагностики токсичности сточных вод различного происхождения и для оценки качества природных пресных вод разных категорий водопользования.
Ассортимент методов биологического тестирования, применяемых в настоящее время в исследовательских целях, достаточно обширен, однако список методов, используемых для практического биотестирования природных и сточных вод в РФ, ограничен буквально несколькими наименованиями [1, 2]. В производственном контроле основным тестируемым параметром при установлении острой токсичности сточных вод по-прежнему является выживаемость, хотя подобные тесты зачастую позволяют выявить лишь приблизительную (и весьма заниженную) токсичность исследуемых проб, особенно если в качестве тест-организмов используются рыбы [2, 3]. В этом отношении более персепективным представляется использование биохимических тест-функций, поскольку любому физиологическому, а тем более морфологическому отклонению от нормы предшествуют определенные биохимические процессы, которые являются первопричиной изменения физиологического состояния тест-организма, ухудшения качества его потомства или гибели. К биохимическим маркерам токсического загрязнения воды следует отнести, прежде всего, ферменты как универсальные катализаторы и регуляторы обменных процессов в клетках живых организмов. В настоящее время использование таких биохимических показателей, как изменение активности ацетилхолинэстеразы и микросомальных монооксигеназ гидробионтов уже находит себе место в практике мониторинга определенных групп токсикантов [4-6].
Известен «Способ определения цианидов в сточных водах» (а.с. №1576860, G01N 33/18, опубл. 07.07.90, Бюлл. №25), включающий отбор пробы воды, содержание в пей в течение 1-5 суток пресноводных водорослей, гомогенизацию растительных образцов, центрифугирование гомогенатов и получение экстракта белков, внесение экстракта в приготовленную инкубационную среду, инкубацию сред и фотоколориметрическое определение в них активности β-цианоаланинсинтазы; о присутствии в исследуемой воде цианидов судят по изменению активности фермента у опытных растений по сравнению с контрольными (взятыми из чистого водоема). Главным недостатком этого способа, сильно ограничивающим область его применения, является возможность тестирования только одной специфической группы загрязнителей поверхностных вод - цианидов.
Известен также «Способ определения концентраций меркаптанов в водной среде» (а.c. №1784913, G01N 33/18, опубл. 30,12.92, Бюлл. №48), основанный на определении активности β-цианоаланинсинтазы у бентосных растений, произрастающих в загрязненных водоемах. Способ предусматривает отбор проб (растений), их гомогенизацию, центрифугирование гомогенатов и получение экстракта белков, добавление к экстракту реагентов, инкубацию реакционных сред, измерение их оптической плотности при 360 нм и расчет активности β-цианоаланинсинтазы, установление концентрации меркаптанов в исследуемых пробах по калибровочному графику, построенному на основании предварительно полученных данных об изменении активности β-цианоаланинсинтазы у водных растений, содержавшихся в течение 24-120 ч и растворах меркаптана концентрацией от 10-3 до 10-7 моль/л. Характеризуясь высокой чувствительностью и экспрессивностью, данный способ имеет два существенных недостатка. Во-первых, он позволяет проводить диагностику загрязнения только вод природных водоемов и не позволяет тестировать сточные воды. Во-вторых, из всего многообразия загрязняющих веществ определению по данному способу подвергаются лишь меркаптаны.
Идея предлагаемого изобретения заключается в использовании для биотестирования токсичности вод таких энзиматических реакций гидробионтов, которые имеют неспецифический характер, т.е. достаточно однотипны (универсальны) в отношении действия широкого спектра токсикантов. Наиболее близким по своей сущности к предлагаемому является «Способ определения токсичности промышленных сточных вод» (а.с. №1751670, G01N 33/18, опубл. 30.07.92, Бюлл. №28), основанный на определении активности множественных форм малатдегидрогеназы (МДГ) микроорганизмов активного ила, помещенных в исследуемую воду. Согласно этому способу отбирают по 200 мл иловой суспензии действующих очистных сооружений в колбы, одну из которых оставляют в качестве контроля, а в другие добавляют компоненты сточных вод. Колбы помещают на магнитные мешалки и проводят инкубацию в течение 1 ч при 37°С, после чего из контрольных и опытных образцов отбирают по 10 мл иловой суспензии, 3-кратно промывают фосфатным буфером (рН 7,0) и центрифугируют 10 мин при 5000 об/мин. Полученные осадки дезинтегрируют в механическом гомогенизаторе при 4°С в течение 5 мин, гомогенаты переносят в чистые колбы и добавляют к ним по 5 мл фосфатного буфера (рН 7,0) и Тритона Х-100 до конечной концентрации 20 мг/мл. Пробы на 2 ч помещают на магнитные мешалки при 37°С, после чего центрифугируют 10 мин при 8000 об/мин. В супернатантах определяют активность МДГ методом электрофореза в плоских блоках полиакриламидного геля (ПААГ) с 1 М трис-ЭДТА-боратным буфером (рН 9,2) в качестве электродного буферного раствора. На линию старта наносят по 50 мкл анализируемых образцов в смеси с 40%-ным раствором сахарозы и проводят электрофорез при силе тока 5 мА/см в первые 30 мин и затем, вплоть до окончания, 10 мА/см. По окончании электрофореза гелевые блоки помещают в инкубационную среду (0,1 М трис-HCl буфер, рН 7,1, - 15 мл, 1 М раствор малата натрия - 10 мл, НАД - 30 мг, нитросиний тетразолиевый - 10 мг, феназинметасульфат - 2 мг, вода дистиллированная - 70 мл) и инкубируют 2 ч при 37°С; множественные формы МДГ выявляются в виде темно-синих зон. Путем денситометрии определяют относительную активность множественных форм МДГ в контрольном и опытных образцах; за критерий токсичности принимают снижение активности хотя бы одной из форм МДГ более чем на 20% по сравнению с контролем.
Обладая рядом достоинств (экспрессивность, универсальность в отношении диагностики сточных вод разного состава), названный способ не лишен существенных недостатков, являющихся следствием использования в качестве тест-объекта микроорганизмов активного ила. Во-первых, способ применим только в условиях производств, имеющих сооружения биологической очистки. Во-вторых, указанный метод не обладает высокой чувствительностью, так как экосистема активного ила адаптирована к высоким концентрациям многих загрязнителей (см. а.с. №1751670, G01N 33/18); действительно, в качестве примеров использования изобретения приведены факты тестирования токсикантов в очень высоких концентрациях, превышающих значения ПДК на несколько порядков. В этом отношении более обоснованным представляется использование данного способа по второму заявленному назначению - для оценки функционального состояния экосистем активного ила на сооружениях биологической очистки.
Задачей, на решение которой направлено предлагаемое изобретение, является разработка нового способа биологического тестирования загрязнения вод, применимого в условиях существующих лабораторий токсикологического контроля и характеризующегося высокой чувствительностью, экспрессивностью, универсальностью и интегральностью (то есть возможностью определения общей токсичности воды с учетом синергизма и антагонизма всех присутствующих в ней химических веществ). Техническим результатом, достигаемым при использовании способа, является:
а) возможность диагностики как сточных, так и природных вод разного назначения;
б) широкий измерительный диапазон, позволяющий тестировать присутствие в водной среде различных токсикантов в концентрациях от 0,1 до 100 ПДК и выше;
в) сокращение времени анализа по сравнению с большинством официально рекомендованных [1] методов биотестирования. Технический результат достигается тем, что в качестве тест-объектов используют пресноводных моллюсков Viviparus viviparus L., а токсичность исследуемой воды диагностируют по изменению общей активности и активности отдельных множественных форм кислой фосфатазы (КФ) и дезоксирибонуклеазы (ДНКазы) в тканях подопытных животных.
В основе предлагаемого способа лежит представление о неспецифическом адаптационном синдроме (НАС); полагают, что механизмы развития НАС универсальны для всего животного мира в отношении действия самых разных стрессовых факторов, в том числе токсических веществ [7]. НАС представляет собой ряд однотипных изменений метаболизма, направленных на устранение последствий травмирующего воздействия. В частности, показано, что у гидробионтов разных систематических групп воздействие токсикантов вызывает, наряду со специфическими изменениями (как результат прямого влияния веществ определенной химической структуры на определенный биохимический процесс), целый комплекс неспецифических метаболических реакций [8], и установлена общность биохимических механизмов адаптации к токсическому воздействию у рыб и водных беспозвоночных [9]. Большое значение в развитии адаптационно-компенсаторных процессов придают, в частности, кислым гидролазам, связывая их функциональную роль с деятельностью лизосомального аппарата клетки, который принимает самое активное участие в индуцированных стрессом внутриклеточных перестройках [7]. Положение о существовании НАС позволяет развивать идею о биохимической диагностике токсического загрязнения любой, в том числе неизвестной заранее, природы.
Моллюски в качестве организмов, в адаптации которых к изменяющимся условиям внешней среды ведущую роль играют процессы, происходящие на уровне клеточного и тканевого метаболизма [10], являются удобными объектами для целей энзиматического тестирования загрязнения вод. Живородка речная (Viviparus viviparus L.) - широко распространенный брюхоногий моллюск, ареал обитания которого включает Европу, Закавказье и Малую Азию [11]; встречается практически в любых гидробиоценозах, вне зависимости от силы антропогенной нагрузки на них [12, 13], что позволяет рассматривать данный вид как обладающий широким адаптивным потенциалом к гидрохимическому режиму; характеризуется доступностью для сбора в природных условиях и простотой лабораторного содержания.
Способ осуществляют следующим образом.
Моллюсков собирают в природных водоемах, после чего не менее 1 месяца адаптируют к лабораторным условиям. В течение этого периода улиток содержат в аквариумах (примерно 150-200 особей на аквариум объемом 60 л) при стабильном режиме освещенности, аэрации, температуры и регулярной, раз в 3-4 суток, смене части культивационной воды (на 1/3 объема отстоянной водопроводной воды). По окончании периода акклимации моллюсков используют в опытах, для чего группы особей примерно одного размера (достаточно 5 животных на одно определение) помещают в стеклянные емкости, наполненные тестируемой водой (не менее 1 л). Контрольную группу помещают в такую же емкость с аквариумной водой. Все группы содержат в одинаковых условиях, при прежнем режиме аэрации и освещения. По истечении времени экспозиции, необходимого для острого опыта, моллюсков извлекают из воды и немедленно препарируют, раскалывая раковину и отделяя скальпелем печень (пищеварительная железа, или hepatopancreas; у речной живородки эта железа расположена в 2-3 завитках вершины раковины). Препарированные органы промывают 0,15 М раствором NaCl, объединяют вместе материал, отобранный у всех моллюсков одной группы, взвешивают с точностью до 100 мг и гомогенизируют 5 мин растиранием в охлажденной до 4-10°С фарфоровой ступке с 0,5%-ным раствором Тритона Х-100, взятом в объеме 10 мл на каждый 1 г массы материала. Полученные гомогенаты центрифугируют 30 мин при 10000 g и 4°С, водную фазу без плавающих на поверхности пробирки жиров осторожно отбирают, не допуская взмучивания осадка, и используют для дальнейшего анализа в качестве экстракта белков печени живородки речной.
Концентрацию белка в экстрактах определяют любым доступным методом. В первом варианте способа токсичность исследуемой воды диагностируют, используя в качестве тест-функции величину активности КФ, которую определяют в экстракте белков спсктрофотометрически по скорости гидролиза модельного субстрата р-нитрофенилфосфата. Для этого 0,1 мл экстракта белков, разбавленного предварительно в 30-50 раз дистиллированной водой, вносят в инкубационную среду, содержащую 1,5 мг динатриевой соли р-нитрофенилфосфата в 1,4 мл 0,05 М ацетатного буфера (рН 4,1). Пробы инкубируют 20 мин при 37°С, затем реакцию останавливают прибавлением 2 мл охлажденного до 4-10°С 0,05 М раствора NaOH. В качестве оптического контроля для спсктрофотомерии используют пробу, в которую экстракт белков вносят после прибавления 0,05 М раствора NaOH. Измеряют оптическую плотность проб при длине волны 415 нм. За единицу активности КФ принимают такое количество фермента, которое приводит к увеличению оптической плотности на 1 ед. при 415 нм (по сравнению с оптическим контролем) за 20 мин инкубации при 37°С. Рассчитывают удельную активность КФ в единицах активности на 1 мг белка (Еудельн), исходя из того количества белка, которое было взято для инкубации. Определяют достоверность различий величин удельной активности КФ между контрольной и каждой из опытных групп моллюсков, применяя t критерий Стьюдента для уровня значимости Р=0,05. Тестируемую воду считают токсичной в случае выявления статистически достоверного изменения удельной активности КФ печени живородки речной в опытном варианте по сравнению с контролем.
Во втором варианте способа наличие токсического действия воды устанавливают на основании достоверности различий активности ДНКазы у контрольной и опытной групп моллюсков. Активность ДНКазы определяют спектрофотометрически по приросту кислоторастворимых продуктов деградации ДНК. В качестве субстрата используют 0,3%-ный раствор высокополимерной ДНК, денатурированной нагреванием готового раствора до 100°С в течение 15 мин с последующим быстрым охлаждением в ледяной бане. 0,1 мл экстракта белков, предварительно разбавленного в 20-30 раз дистиллированной водой, вносят в инкубационную среду, содержащую 0,1 мл раствора субстрата и 0,8 мл 0,05М ацетатного буфера (рН 4,6), и инкубируют 1 ч при 37°С, затем охлаждают 10 мин в бытовом холодильнике (4-10°С), после чего негидролизованную ДНК осаждают добавлением 2 мл 5%-ного раствора HClO4. В качестве оптического контроля для спектрофотомерии используют пробу, в которую экстракт белков вносят после прибавления раствора HClO4. Пробы выдерживают 20 мин при 0°С и центрифугируют 15 мин при 8000 g, после чего аккуратно отбирают водную фазу и измеряют ее оптическую плотность при 260 нм. За единицу активности ДНКазы принимают такое количество фермента, которое приводит к увеличению оптической плотности на 1 ед. при 260 нм (по сравнению с оптическим контролем) за 1 ч инкубации при 37°С. Рассчитывают удельную активность ДНКазы в единицах активности на 1 мг белка (Еудельн), исходя из того количества белка, которое было взято для инкубации. Определяют достоверность различий величин удельной активности ДНКазы между контрольной и каждой из опытных групп моллюсков, применяя t критерий Стьюдента для уровня значимости Р=0,05. Тестируемую воду считают токсичной в случае выявления статистически достоверного изменения удельной активности ДНКазы печени живородки речной в опытном варианте по сравнению с контролем.
О токсичности проб воды в третьем варианте способа судят по энзимограммам ДНКазы (составу множественных форм), получаемых методом электрофореза в ПААГ по Дэвису с последующим окрашиванием гелей метиленовым синим. Электрофорез проводят в блоках (колонках или пластинах) ПААГ. В разделяющий гель в ходе приготовления вместо воды вносят водный раствор высокополимерной ДНК, предварительно денатурированной нагреванием (см. выше) из расчета 540 мкг ДНК на 1 мл ПААГ. Концентрация разделяющего ПААГ должна составлять 7,2%, отношение концентраций метиленбисакриламида и суммы мономеров в его составе 2,6%; для концентрирующего ПААГ эти величины должны равняться соответственно 3,9 и 20%. Перед нанесением экстракты белков разбавляют 40%-ным раствором сахарозы до конечной концентрации белка 1-2 мг/мл. При использовании колонок ПААГ высотой 8 см и диаметром 0,4 см на каждую наносят по 0,1 мл разбавленного экстракта, при использовании пластины ПААГ размером 15×6 см и толщиной 2 мм в ячейки сечением 1×5 мм и глубиной 10 мм вносят по 50 мкл образца. Электрофорез проводят в 0,032 М трис-глициновом буфере (рН 8,9) с бромфеноловым синим в качестве метчика, поддерживая температуру на уровне 5-7°С. Первые 10-15 мин (до вхождения белков в разделяющий гель) силу тока устанавливают равной 1,5 мА на колонку (1,5 мА/см ширины пластины), затем, до окончания электрофореза, ее увеличивают до 2,5 мА на колонку (2,5 мА/см пластины). Окончанием электрофореза считают смещение фронта, окрашенного метчиком, до 0,5-1 см от нижнего края блока ПААГ.
По окончании электрофореза блоки ПААГ ополаскивают дистиллированной водой и полностью погружают в охлажденный до 4-10°С 0,05 М ацетатный буфер (рН 4,6). Через 30 мин буферный раствор заменяют на свежую порцию, в которой инкубируют гели в течение 90 мин при 37°С. Затем буферный раствор заменяют на 1 М уксусную кислоту, блоки ПААГ выдерживают в ней 10 мин, ополаскивают дистиллированной водой и погружают на 30 мин в 0,2%-ный раствор метиленового синего, приготовленный на 0,05 М ацетатном буфере (рН 4,6). Избыток красителя вымывают из блоков ПААГ сначала 1,5-2,5 ч 5%-ным раствором уксусной кислоты, заменяя его порцию на свежую каждые 30 мин, а затем дистиллированной водой до появления четких бесцветных зон деградации ДНК (зон ДНКазной активности) на синем фоне геля. Полученную окраску энзимограмм фиксируют, погружая ПААГ в 5%-ный раствор HClO4. В указанных условиях у контрольных (содержавшихся в чистой аквариумной воде) моллюсков на энзимограммах ДНКазы выявляются 5 зон активности (множественные формы ДНКазы) с Rf 0,76, 0,71, 0,34, 0,30 и 0,09. Тестируемую воду считают токсичной в случае выявления на энзимограммах ДНКазы печени живородки речной в остром опыте одной или нескольких зон активности, не характерных для моллюсков контрольной группы.
Токсичность тестируемой воды в четвертом варианте способа определяют по энзимограммам КФ. Электрофорез проводят в блоках (колонках или пластинах) ПААГ по методике Дэвиса; концентрация разделяющего геля должна составлять 5,7%, отношение концентраций метиленбисакриламида и суммы мономеров в его составе 2,6%, для концентрирующего геля эти величины должны равняться соответственно 3,9 и 20%. Перед нанесением экстракты белков разбавляют 40%-ным раствором сахарозы до конечной концентрации белка 1-1,5 мг/мл. При использовании колонок ПААГ высотой 8 см и диаметром 0,4 см на каждую наносят по 0,1 мл разбавленного экстракта, при использовании пластины ПААГ размером 15×6 см и толщиной 2 мм в ячейки сечением 1×5 мм и глубиной 10 мм вносят по 50 мкл образца. Электрофорез проводят в 0,032 М трис-глициновом буфере (рН 8,9) с бромфеноловым синим в качестве метчика, поддерживая температуру на уровне 5-7°С. Первые 10-15 мин (до вхождения белков в разделяющий гель) силу тока устанавливают на уровне 1 мА на колонку (1 мА/см ширины пластины), затем, до окончания электрофореза, ее увеличивают до 2 мА на колонку (2 мА/см пластины). Окончанием электрофореза считают смещение фронта, окрашенного метчиком, до 0,5-1 см от нижнего края блока ПААГ.
Активность КФ в ПААГ обнаруживают по реакции гидролиза α-нафтилфосфата с последующим азосочетанием высвобождающегося нафтола с прочным синим Б. Для этого блоки ПААГ по окончании электрофореза погружают в 0,2 М ацетатный буфер (рН 4,1), где выдерживают 20 мин при 37°С. Затем ПААГ переносят в инкубационную среду, 10 мМ раствор динатриевой соли α-нафтилфосфата в 0,2 М ацетатном буфере (рН 4,1), и инкубируют 20 мин при 37°С. Для визуализации зон активности КФ на энзимограмме в инкубационную среду добавляют водный раствор красителя прочный синий Б до конечной концентрации 0,5 мг/мл и выдерживают ПААГ при комнатной температуре еще 10-20 мин или до появления четких зон активности фермента темно-малинового цвета. Полученную окраску энзимограмм фиксируют, полностью погружая ПААГ в 7%-ный раствор уксусной кислоты. В указанных условиях моллюски контрольной (содержавшейся в чистой аквариумной воде) группы характеризуются пятью зонами активности на энзимограммах КФ (множественные формы КФ) с Rf 0,67, 0,53, 0,41, 0,32 и 0,23. Тестируемую воду считают токсичной в случае выявления на энзимограммах КФ печени живородки речной в остром опыте хотя бы одной зоны активности, не обнаруживаемой в контроле, и/или отсутствия одной или нескольких зон активности, характерных для моллюсков контрольной группы.
Пример 1. Установление токсичности сточной воды неизвестного состава и определение уровня ее безопасного разбавления.
Исследуемая вода представляла собой стоки ливневой канализации предприятия «Лакокраска» (г.Ярославль), поступающие непосредственно в природный водоем (р.Которосль). Пробы воды отбирали в соответствии с действующими правилами отбора проб сточных вод [1]. Для токсикологического эксперимента использовали особей живородки речной, собранных в прибрежной зоне р.Которосль в районе пос.Починки (Ярославская область). Моллюсков (около 150 особей) в течение месяца содержали в аквариуме объемом 60 л при круглосуточной аэрации воды, освещении естественным светом и регулярной (раз в трое суток) смене 1/3 объема аквариумной воды на отстоянную водопроводную воду. В рамках острого опыта акклимированных моллюсков группами по 8 особей помещали в стеклянные емкости объемом 1 л, наполненные профильтрованной аквариумной водой с примесью определенных количеств тестируемых стоков. Экспозиция опыта (время содержания моллюсков в тестируемой воде) составляла 96 ч, после чего моллюсков извлекали из воды и препарировали у них печень (2-3 верхних завитка тела животного). Ткани промывали 0,15 М раствором NaCl и, объединяя органы от животных одной группы, проводили гомогенизацию материала растиранием в течение 5 мин в охлажденной фарфоровой ступке с 0,5%-ным раствором Тритона Х-100, прибавляя его в десятикратном по отношению к навеске ткани объеме. Гомогенаты центрифугировали на рефрижераторной центрифуге К-24 в течение 30 мин при 10000 g и 4°С. Концентрацию общего белка в полученных экстрактах определяли методом Бредфорд.
Токсичность воды оценивали по первому варианту способа. Для этого 0,1 мл разбавленного экстракта, содержащего от 50 до 100 мкг общего белка, вносили в инкубационную среду, содержащую в 1,4 мл 0,05 М ацетатного буфера (рН 4,1) 1,5 мг динатриевой соли р-нитрофенилфосфата («Sigma», США). Пробы инкубировали в течение 20 мин при 37°С, после чего реакцию останавливали, прибавляя 2 мл охлажденного 0,05 М NaOH; в контрольные пробы экстракт вносили после прибавления раствора щелочи. Оптическую плотность полученной окраски измеряли на спектрофотометре СФ-26 при длине волны 415 нм. За единицу активности фермента принимали такое его количество, которое вызывало прирост оптической плотности на 1 ед. при 415 нм за 20 мин инкубации при 37°С. Рассчитывали удельную активность КФ в единицах активности на 1 мг белка (Е/мг), после чего определяли достоверность различий величин удельной активности фермента в контрольном и опытных вариантах, применяя t критерий Стьюдента для уровня значимости Р=0,05.
Полученные данные представлены в табл.1. Качество чистой аквариумной воды (контроль) и сточной воды в серии разбавлений дополнительно оценивали по ряду гидрохимических показателей, таких как концентрация кислорода (по Винклеру), бихроматная окисляемость воды (ХПК), биохимическое потребление кислорода за 5 сут (БПК5), а также с помощью токсикологического теста на дафниях Daphnia magna L. продолжительностью 96 ч [1].
О2, мг/л
Гидрохимические и токсикологические характеристики сточной воды свидетельствовали о ее неудовлетворительном качестве и выраженных токсических свойствах. Судя по высокому значению ХПК и крайне низкой концентрации растворенного кислорода, исследуемые стоки содержали большое количество недоокисленных компонентов, а небольшое БПК этой воды указывало на то, что эти компоненты, скорее всего, имели небиотическую природу. Неразбавленная сточная вода вызывала 100%-ную гибель дафний, причем ее токсические свойства по отношению к дафниям сохранялись вплоть до разбавления в 5 раз (табл.1).
Биотестирование качества воды по предлагаемому способу выявило увеличение активности КФ тест-организмов (табл.1). В опытах с минимальной концентрацией стоков (разбавление в 50 и 25 раз) эти отклонения были статистически не достоверны, но с уменьшением разбавления тестируемой воды следовал практически линейный рост активности КФ, вплоть до опыта с 50%-ной сточной водой. Действие неразбавленных стоков также вызвало достоверное увеличение активности тест-фермента, но оно оказалось не столь существенным, как можно было ожидать (явление, известное в биохимической токсикологии как «депрессия»; его причины будут раскрыты при обсуждении последующих примеров).
Таким образом, была выявлена корреляция между гидрохимическими показателями качества воды, данными дафниевого теста и результатами энзиматического тестирования (предлагаемый способ). Два примененных метода биотестирования обнаружили сопоставимую чувствительность, при этом достоверное изменение активности КФ моллюсков вызывалось более низкой концентрацией стоков (табл.1). При разбавлении сточной воды в 25 раз не наблюдалось значимых отклонений ни одной из тест-функций; именно этот уровень разбавления исследованной воды можно признать безопасным для жизнедеятельности гидробионтов.
В последующих примерах приведены данные по биологической оценке качества вод, содержащих определенные токсические вещества в известных (задаваемых в опыте) концентрациях. Токсиканты вносили однократно в содержащие 1 л аквариумной воды стеклянные сосуды, куда затем помещали подопытных моллюсков (Viviparus viviparus L.). Обычно использовали несколько концентраций загрязнителя, одна из которых соответствовала величине ПДК для данного вещества; контрольную группу моллюсков помещали в стеклянный сосуд с 1 л незагрязненной аквариумной воды. В опытах использовали вещества, относящиеся к основным группам загрязнителей поверхностных вод (тяжелые металлы, фенолы, нефтепродукты, СПАВ, хлорорганические соединения); значения ПДКводн для этих токсикантов приведены в табл.2.
Пример 2. Исследовали действие хлорбензола в двух сериях опытов (концентрация токсиканта 0,0001, 0,0015,0,0050 мг/л и 0,001, 0,015, 0,050 мг/л) при экспозиции 72 ч; токсичность воды диагностировали параллельно по первому и второму варианту способа. Моллюсков собирали в прибрежной зоне р.Вязь в районе деревни Тишково Московской области и в течение месяца содержали в аквариуме при круглосуточной аэрации воды, освещении естественным светом и регулярной (раз в трое суток) смене 1/3 объема аквариумной воды на отстоянную водопроводную воду. В рамках токсикологического эксперимента в стеклянные сосуды с 1 л тестируемой воды помещали группы моллюсков по 5 особей примерно одного размера, по истечении времени экспозиции у животных извлекали печень и получали экстракт белков согласно приведенной в примере 1 прописи. Концентрацию общего белка в экстрактах определяли методом Лоури. Активность КФ определяли спектрофотометрически аналогично описанному в примере 1. Для определения активности ДНКазы в качестве субстрата использовали 0,3%-ный раствор высокополимерной ДНК из эритроцитов цыпленка («Reanal», Венгрия), денатурированной нагреванием (15 мин, 100°С) с последующим быстрым охлаждением в ледяной бане. Готовили инкубационную смесь, содержащую 0,8 мл 0,05 М ацетатного буфера (рН 4,6), 0,1 мл субстрата и 0,1 мл разбавленного в 20-30 раз экстракта белков, и проводили инкубацию в течение 1 ч при 37°С. Затем пробы на 10 мин помещали в холодильник, после чего негидролизованную ДНК осаждали добавлением 2 мл 5%-ного раствора хлорной кислоты; в контроль экстракт вносили после прибавления раствора HClO4. Пробы 20 мин выдерживали при 0°С, затем центрифугировали в течение 15 мин при 8000 g на центрифуге марки ОПН-ИУХП.4.2, после чего измеряли оптическую плотность супернатантов при 260 нм на спектрофотометре СФ-26. За единицу активности принимали такое количество фермента, которое вызывало прирост оптической плотности на 1 ед. при 260 нм за 1 ч инкубации при 37°С. Рассчитывали удельную активность ДНКазы в единицах активности на 1 мг белка (Е/мг), после чего проводили статистическую обработку результатов, используя t критерий Стьюдента для уровня значимости Р=0,05. Полученные результаты приведены в таблице 3.
Воздействие хлорбензола через 72 ч опыта привело к возрастанию активности ДНКазы и КФ в печени моллюсков. Динамика увеличения ферментативной активности в зависимости от концентрации токсиканта в среде носила нелинейный характер, схожий для двух исследованных гидролаз, при этом отклонения активности ДНКазы и КФ от контрольного уровня были статистически достоверными во всех шести опытных вариантах (табл.3). Таким образом, загрязнение воды хлорбензолом в концентрациях, соответствующих 0,1-50 величинам рыбохозяйственной ПДК, может быть диагностировано энзиматически по предлагаемому способу.
Пример 3. Исследовали действие хлорида кадмия в концентрациях (в пересчете на Cd2+) 0,01 и 1,0 мг/л (экспозиция опыта 12, 24, 48, 72 ч) и 0,05 мг/л (при экспозиции 12, 24, 48, 72, 96 ч). Токсичность тестируемой воды в первом случае оценивали по четвертому варианту способа, во втором случае - по третьему варианту. Моллюсков собирали в прибрежной зоне р.Клязьма в районе ж/д платформы «Тарасовская» (Московская обл.). До опыта животных 1 месяц содержали в лаборатории, как это описано в примере 1, после чего группами по 15-20 особей помещали в стеклянные сосуды с 1 л тестируемой воды, по истечении определенного времени экспозиции извлекали и немедленно препарировали по 5 особей. Процедуры выделения тканей и получения экстракта белков проводили согласно описанию в примере 1. Концентрацию общего белка в экстрактах определяли методом Лоури. Для выявления множественных форм ферментов использовали прибор для вертикального электрофореза на колонках ПААГ высотой 8 см и диаметром 0,4 см, применяя в качестве источника тока универсальный источник питания УИП-1. В случае определения активности ДНКазы концентрация разделяющего геля составляла 7,2%, отношение концентраций метиленбисакриламида и суммы мономеров 2,6%; для концентрирующего геля эти величины равнялись соответственно 3,9 и 20%. При приготовлении ПААГ в него вносили денатурированную ДНК (процедура денатурации описана в предыдущем примере) из расчета 540 мкг на 1 мл раствора разделяющего геля. На каждую колонку ПААГ наносили по 0,1 мл экстракта, предварительно разбавленного 40%-ным раствором сахарозы до концентрации белка 1-2 мг/мл. Электрофорез проводили в 0,032 М трис-глициновом буфере, рН 8,9, с бромфеноловым синим в качестве метчика, поддерживая температуру на уровне 5-7°С. Первые 10-15 мин, до вхождения белков в сепарирующий гель, силу тока устанавливали на уровне 1,5 мА на колонку, затем ее увеличивали до 2,5 мА на колонку; в таком режиме электрофорез продолжался 60-80 мин. По окончании электрофореза колонки ПААГ промывали дистиллированной водой и погружали в холодный инкубационный буфер (0,05 М ацетатный буфер, рН 4,6). Через 30 мин буферный раствор заменяли на свежий и проводили инкубацию гелей при 37°С в течение 90 мин. Реакцию останавливали, заменяя буфер на 1 М уксусную кислоту. Через 10 мин колонки промывали дистиллированной водой и помещали на 30 мин в 0,2%-ный раствор метиленового синего, приготовленный на инкубационном буфере. Избыток красителя отмывали 5%-ной уксусной кислотой в течение 1,5-2,5 ч, заменяя ее раствор на свежий каждые 30 мин, а затем - дистиллированной водой, до появления четких бесцветных зон ДНКазной активности на синем фоне геля. Полученные энзимограммы фиксировали в 5%-ной HClO4.
При определении активности КФ концентрация разделяющего геля составляла 5,7% (отношение концентраций метиленбисакриламида и суммы мономеров 2,6%), концентрирующего геля 3,9 (отношение концентраций метиленбисакриламида и суммы мономеров 20%). На каждую колонку ПААГ наносили по 0,1 мл экстракта, предварительно разбавленного 40%-ным раствором сахарозы до концентрации белка 1-1,5 мг/мл. Электрофорез проводили в 0,032 М трис-глициновом буфере (рН 8,9) с добавлением бромфенолового синего при температуре 5-7°С. Первые 10-15 мин, до вхождения белков в разделяющий гель, силу тока поддерживали на уровне 1 мА на колонку, затем ее увеличивали до 2 мА на колонку; в таком режиме электрофорез продолжался 90-100 мин. По окончании электрофореза колонки ПААГ инкубировали 20 мин при 37°С без субстрата в 0,2 М ацетатном буфере (рН 4,1), после чего переносили их в инкубационную среду - 10 мМ раствор динатриевой соли α-нафтилфосфата («Lachema», Чехия) в 0,2 М ацетатном буфере, где инкубировали при 37°С 20 мин. Затем в инкубационную среду добавляли водный раствор красителя прочный синий Б (Fast Blue В, "Reanal", Венгрия) до его конечной концентрации 0,5 мг/мл и инкубировали гель при комнатной температуре еще 10-20 мин, до появления четких зон активности КФ темно-малинового цвета. Окраску фиксировали, помещая колонки ПААГ в 7%-ный раствор уксусной кислоты. Для количественной характеристики ферментативной активности множественных форм КФ электрофоретические спектры КФ в блоках ПААГ сканировали на денситометре Ultroscan XL ("LKB", Швеция); по результатам денситометрии получали значения активности отдельных множественных форм, выраженные в процентах к суммарной активности КФ.
Исследование состава множественных форм КФ живородок показало, что на эпзимограммах контрольных моллюсков при любой экспозиции опыта обнаруживалось 5 форм фермента (Rf 0,67, 0,53, 0,41, 0,32 и 0,23), проявлявших стабильный уровень активности. Воздействие хлорида кадмия вызывало ряд закономерных изменений ферментативной активности форм КФ, которые отражены в табл.4.
Так, при интоксикации моллюсков монотонно возрастала доля относительной активности формы КФ с Rf 0,67, активность же формы с Rf 0,41 постепенно снижалась вплоть до полного угнетения; инактивирование этой формы КФ в печени улиток происходило через 24 или 48 ч при разной интенсивности токсического воздействия и имело необратимый характер. Относительная активность низкоподвижных форм КФ (Rf 0,32 и 0,23) не претерпевала существенных изменений. Наконец, на определенной стадии опыта обнаруживалась новая форма КФ с Rf 0,21, не проявлявшая активности у контрольных животных. Такие изменения, как необратимая инактивация и индукция активности определенных форм фермента, являются качественными характеристиками интоксикации тест-организмов, что позволяет осуществлять тестирование токсичности воды по предлагаемому способу, в отличие от прототипа, без применения денситометрии энзимограмм. При этом в качестве диагностического признака выступают наблюдаемые визуально изменения в электрофоретическом спектре КФ (полное инактивирование хотя бы одной из активных в норме форм фермента и/или появление форм КФ, не обнаруживаемых в контроле). Исходя из выраженности изменений экспозиция (время содержания моллюсков в тестируемой воде) такого опыта может находиться в пределах 48-72 ч.
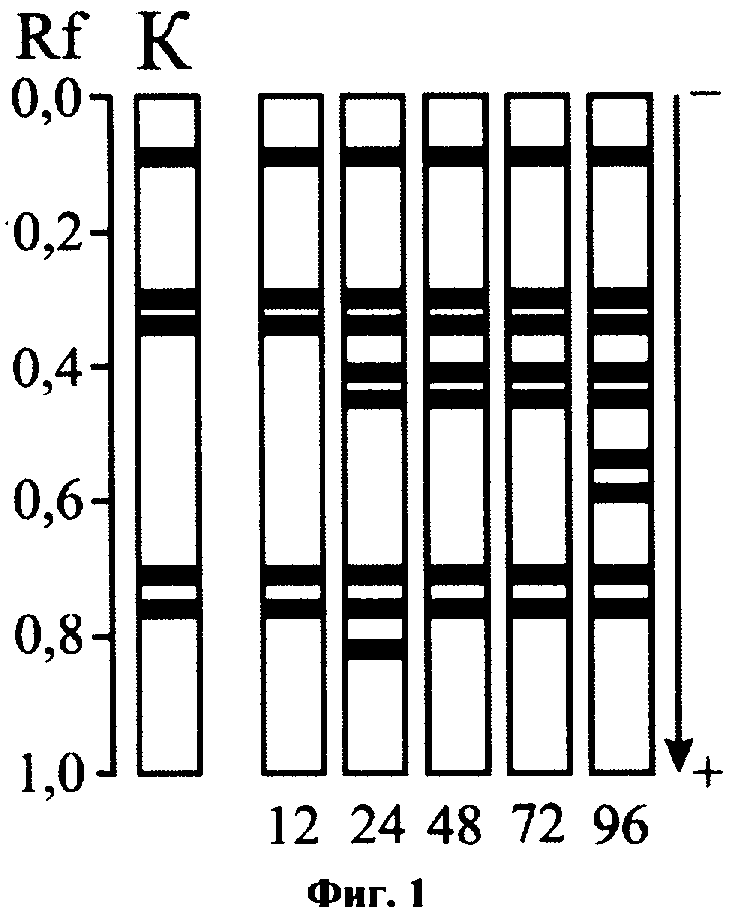
Как в данном опыте, так и во всех последующих токсикологических экспериментах на энзимограммах контрольных моллюсков отмечались 5 зон ДНКазной активности (Rf 0,76, 0,71, 0,34, 0,30 и 0,09). Эти формы ДНКазы всегда присутствовали в электрофоретических спектрах фермента и в условиях токсического воздействия на моллюсков. При этом действие токсикантов приводило к индукции активности дополнительных, не обнаруживаемых у контрольных животных, форм фермента, что позволяло диагностировать интоксикацию моллюсков (определять токсическое загрязнение воды) по числу множественных форм ДНКазы визуально, без применения аппаратуры для количественной обработки энзимограмм. Так, например, присутствие в воде хлорида кадмия к 12 ч экспозиции не сказывалось на гетерогенности ДНКазы, но затем наблюдалась последовательная активация ряда «индуцибельных» форм фермента (см. фиг.1: К - контроль; 12, 24, 48, 72, 96 - экспозиция опыта, ч). Начиная с 24 ч опыта, действие токсиканта приводило к активации латентных в норме форм с Rf 0,45 и 0,41, через 24 ч экспозиции на энзимограммах подопытных моллюсков дополнительно выявлялась форма с Rf 0,82, через 96 ч - формы с Rf 0,59 и 0,54. Таким образом, учитывая выраженность наблюдаемых изменений и фактор экспрессивности, тестирование обусловленной ионами Cd2+токсичности воды можно осуществлять по третьему варианту способа при экспозиции опыта, составляющей 24 ч.
Пример 4. Исследовали действие фенола (карболовой кислоты) в концентрациях 10-4 и 10-3 мг/л (0,1 и 1 ПДКводн соответственно) при экспозиции опыта 24, 48 и 72 ч. Токсичность определяли по второму и третьему вариантам способа. Моллюсков собирали в прибрежной зоне р.Вязь в районе деревни Тишково Московской области и в течение месяца акклимировали их к лабораторным условиям (см. пример 1). Условия токсикологического эксперимента, процедуры получения экстракта белков и определения активности кислой ДНКазы методом электрофореза были аналогичны изложенным в примере 3; спектрофотометрическое определение активности ДНКазы проводили в соответствии с описанием, приведенным в примере 2.
Данные, полученные при тестировании токсичности воды по второму варианту способа, свидетельствуют, что загрязнение фенолом привело к резкому увеличению активности ДНКазы в печени моллюсков, при этом уровень ее мог превышать контрольные значения почти в 7 раз (табл.5).
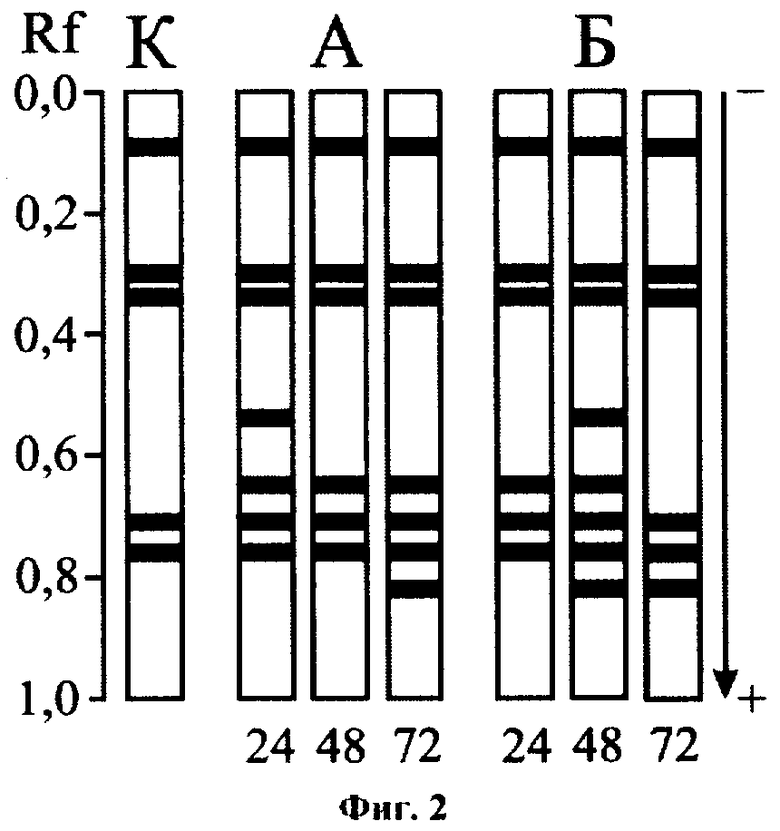
Определение загрязнения по третьему варианту способа также позволяло выявить токсический эффект по индукции активности некоторых латентных в норме форм ДНКазы (Rf 0,82, 0,65 и 0,54), наблюдавшейся в динамике токсического воздействия (фиг.2: К - контроль; А, Б - действие фенола в концентрациях 10-4 и 10-3 мг/л соответственно; 24,48, 72 - экспозиция опыта, ч). Заметим, что определение фенола в воде методами биотестирования наталкивается на существенные трудности. Предлагаемые способы позволяют тестировать токсикант в довольно высоких концентрациях, превышающих ПДК зачастую на несколько порядков [16-18]. Приведенные данные свидетельствуют о высокой чувствительности ДНКазы живородки речной к присутствию в среде достаточно низких концентраций фенола, что позволяет осуществлять энзиматическую диагностику данного вида загрязнений.
Пример 5. Исследовали действие синтетического моющего средства (CMC) «Ariel» в концентрациях 0,02, 0,2 и 2,0 мг/л при экспозиции опыта 24, 48, 72 ч. Токсичность диагностировали по второму варианту способа; необходимые для этого операции проводили так, как это описано в предыдущем примере. Полученные результаты приведены в табл.6.
Динамика изменения активности ДНКазы моллюсков под воздействием CMC имела циклический характер, однотипный для действия различных концентраций загрязнителя: ингибирование ферментативной активности на первые сутки экспозиции, ее рост на вторые сутки и повторное падение ниже контрольных величин - на третьи. Эти изменения могут рассматриваться в качестве маркера интоксикации моллюсков, что позволяет тестировать загрязнение воды CMC.
Пример 6. Исследовали действие сульфата меди (II) в концентрациях (в перерасчете на Cu2+) 0,001, 0,01 и 0,1 мг/л. Токсичность диагностировали по второму и третьему вариантам способа. Экспозиция опыта составляла от 3 до 72 ч, все остальные условия соответствовали изложенным в примере 4.
У контрольных моллюсков стабильно проявляли активность 5 форм ДНКазы (Rf 0,76, 0,71, 0,34,0,30 и 0,09); через 3-12 ч токсического воздействия изменений в электрофоретическом спектре фермента не происходило. Однако к 18 ч пребывания моллюсков в тестируемой воде во всех вариантах опыта наблюдалась активация ряда «индуцибельных» форм ДНКазы, характеризующихся Rf 0,82, 0,65, 0,59 и 0,54 (фиг.4: К - контроль; А, Б, В - действие Cu2+в концентрациях 0,001, 0,01, 0,1 мг/л соответственно, 6, 12, 18, 24, 48, 72 - экспозиция опыта, ч). Выраженность подобных изменений в период 18-72 ч экспозиции позволяет диагностировать присутствие в воде меди по увеличению активных форм ДНКазы.
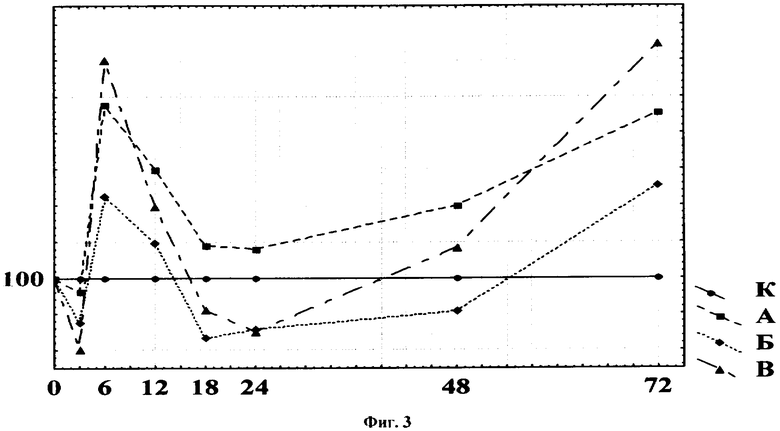
Данные по изменению общей активности ДНКазы при действии на моллюсков сульфата меди (II) приведены в табл.7. Уже через 3 ч опыта происходили статистически достоверные отклонения значений тест-функции под воздействием токсиканта в концентрациях 0,1 и 0,01 мг/л, через 6 ч - во всех трех опытных вариантах. На всем протяжении эксперимента активность ДНКазы претерпевала существенные изменения, которые выражались как в снижении, так и в возрастании ее значений сверх контрольного уровня. При графическом отображении полученных результатов (фиг.3: по оси абсцисс - время экспозиции, ч; по оси ординат - удельная активность, % к контролю) нетрудно заметить, что кривые зависимости активности ДНКазы от продолжительности токсического воздействия имеют форму синусоиды, признанную в практике токсикологии в качестве стандартной и имеющую строгое теоретическое обоснование с точки зрения термодинамики биологических процессов адаптации [19-21]. При анализе полученных графиков четко выявляются периоды, соответствующие трем «классическим» стадиям развития адаптационного синдрома: угнетение (3 ч), стимуляция (6 ч), повторное снижение интенсивности метаболизма (12-24 ч), которое может либо сменяться серией волнообразных изменений функциональных показателей (48, 72 ч), либо продолжаться длительное время. Данный опыт выявил ранний ответ используемой тест-функции на токсическое воздействие, что позволяет обнаруживать загрязнение воды ионами Cu2+в концентрациях на уровне 1-100 ПДК при 3-6 ч экспозиции опыта.
Пример 7. Исследовали действие нефтепродуктов (бензин) в концентрациях 0,005, 0,05 и 0,5 мг/л при экспозиции 3-72 ч. Токсичность диагностировали параллельно по первому, второму и третьему вариантам предлагаемого способа. Определение активности КФ проводили так, как это описано в примере 1; все остальные процедуры и операции были аналогичны описанным в примере 4.
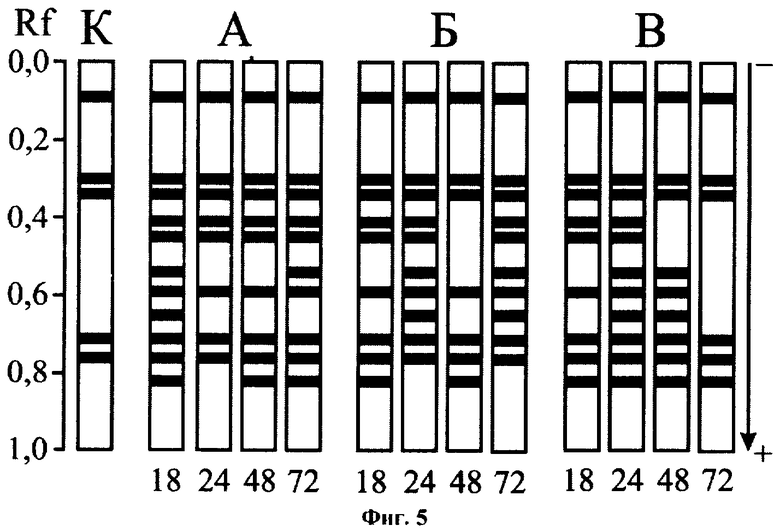
Результаты биотестирования по третьему варианту способа отражены на фиг.5 (К - контроль; А, Б, В - воздействие бензина в концентрациях 0,005, 0,05 и 0,5 мг/л соответственно; 18, 24, 48, 72 - экспозиция опыта, ч). Как и при действии сульфата меди, изменения в электрофоретическом спектре ДНКазы моллюсков под влиянием бензина происходили, начиная с 18 ч экспозиции опыта. К этому моменту набор форм фермента подопытных животных значительно обогащался за счет активации ряда «индуцибельных» форм, характеризующихся Rf 0,82, 0,65, 0,59, 0,54, 0,45 и 0,41. В дальнейшем, при продолжающемся токсическом воздействии, состав комплекса ДНКаз претерпевал существенные изменения, что позволяло констатировать загрязнение воды на основании визуальной оценки полученных энзимограмм.
Данные по определению токсичности воды по первому и второму вариантам способа приведены в табл.8 и 9; они свидетельствуют, что тестирование данного вида загрязнения по достоверному изменению активности ДНКазы и КФ моллюсков возможно уже через 3-6 ч экспозиции опыта, а наиболее выраженные изменения тест-функции наблюдаются через 12 ч экспозиции.
При этом был выявлен схожий характер изменения активности гидролаз в результате действия токсикантов различной химической природы. Как и в случае действия сульфата меди, реакция тест-ферментов в ответ на загрязнение воды нефтепродуктами укладывалась в рамки развития адаптационно-компенсаторных процессов. При этом на более поздних стадиях опытов динамика изменения активности ДНКазы и КФ обнаруживала наиболее выраженное сходство, а в первые часы токсического воздействия разной интенсивности активность гидролаз могла меняться разнонаправлено. Так, присутствие в среде более высоких концентраций меди и бензина приводило вначале к угнетению активности ДНКазы, а наименьшие из использованных концентраций токсикантов не вызывали подобной реакции (табл.7, 8). Как известно, приспособительный смысл первоначального снижения уровня обмена веществ («эффект затаивания») заключается в ограничении контакта животного со средой с целью уменьшения количества поступающего извне токсиканта и избегания таким образом его повреждающего действия [19, 20]. Известно также, что эта стадия приспособительной реакции может быть выражена слабо или совсем отсутствовать, если сила внешнего воздействия была относительно небольшой [20]. Вероятно, при низких концентрациях токсических веществ в среде уровень их накопления в тканях моллюсков в первое время слишком мал для того, чтобы вызвать выраженную реакцию «затаивания» в масштабах целого организма. Отсутствие адаптивной реакции приводит к быстрой кумуляции токсикантов в тканях и накоплению вызванных ими повреждений, в связи с чем, как и при действии высоких концентраций загрязнителей, происходит развитие компенсаторного ответа в виде эффекта стимуляции (т.е. увеличения интенсивности энергетического и пластического обмена с целью ускоренного синтеза имеющихся в резерве генотипа белков для компенсации травмирующего воздействия [20]). Этот эффект является составляющей единого неспецифического адаптационного синдрома клетки (НАС) [7]. В рамках НАС и происходил резкий скачок активности тест-ферментов после начального периода угнетения либо «безразличия» (табл.7, 8, 9), особенно ярко выраженный в случае ДНКазы. Затем наблюдалось не менее резкое угнетение ферментативной активности и падение ее уровня ниже контрольных величин, сменявшееся повторным ростом.
Обобщая изложенные данные, резюмируем, что изменения общей активности ДНКазы и КФ при действии на моллюсков разных токсических веществ имеют схожую динамику, совпадающую со стадиями развития адаптационного синдрома и, следовательно, являются свидетельством интоксикации животных. Таким образом, достоверное отклонение величин активности ДНКазы и КФ подопытных моллюсков от контрольного уровня является маркером присутствия в тестируемой воде токсических веществ в концентрациях, представляющих опасность для жизнедеятельности гидробионтов. С помощью этого показателя можно обнаруживать загрязнение воды как отдельными химическими веществами, так многокомпонентными смесями, в том числе неизвестного заранее состава.
В то же время особенности развития адаптационно-компенсаторных процессов при интоксикации делают возможной ситуацию, когда на момент измерения активность тест-фермента в том или ином опытном варианте приблизится к контрольному уровню, что следовало бы принять за свидетельство приемлемого качества воды. Для исключения возможности ошибки при диагностике по 1 и 2 вариантам способа следует, во-первых, осуществлять тестирование воды в серии разбавлений (что обязательно и при биотестировании традиционными методами, см. [2]), во-вторых, проводить измерения последовательно при двух экспозициях опыта, при этом достоверное изменение значений тест-функции хотя бы в одном из двух опытных вариантов будет являться показателем токсичности исследуемой воды. Учитывая ранний отклик тест-функции на токсическое воздействие, подобную диагностику целесообразно проводить при 6 и 12 ч экспозиции, что обеспечит высокую экспрессивность теста.
По сравнению с общей активностью кислых гидролаз моллюсков изменения в составе множественных форм ДНКазы и КФ возникают позднее и носят более специфический характер. В частности, для действия каждого из использованных нами токсикантов характерна собственная, специфическая динамика активации «индуцибельных» форм ДНКазы. Формирующийся спектр функционирующих форм зависит, видимо, как от природы токсического вещества (особенностей его биотоксического эффекта), так и от силы (концентрации и длительности) его травмирующего воздействия. Принципиальным моментом является то, что состав множественных форм и ДНКазы, и КФ живородки речной отличается стабильностью характеристик в норме и не обнаруживает отличий у моллюсков, собранных в разных местах обитания, благодаря чему возможно четкое определение нормы. Таким образом, любое качественное изменение в электрофоретическом спектре ДНКазы или КФ подопытных моллюсков является свидетельством действия пороговых или подпороговых концентраций токсикантов, что позволяет достаточно просто диагностировать токсическое загрязнение воды на основании визуальной оценки энзимограмм. При этом достаточно провести измерение при одной экспозиции опыта, так как изменения в спектре множественных форм тест-ферментов, возникнув при токсическом воздействии достаточной интенсивности, характеризуются развернутостью во времени (см. табл.4, фиг.1, 2, 4, 5). Экспозиция подобных тестов может находиться в пределах 18-48 ч (третий вариант способа) и 48-72 ч (четвертый вариант способа).
СПИСОК ЛИТЕРАТУРЫ
1. Методическое руководство по биотестированию воды. РД - 118-02-90. М.: Наука, 1991.
2. Жмур Н.С.Государственный и производственный контроль токсичности вод методами биотестирования в России. М.: Международный Дом Сотрудничества, 1997. 117 с.
3. Данильченко О.П. // Обобщенные показатели качества вод - 83. Практические вопросы биотестирования и биоиндикации. Всесоюзный симпозиум (Черноголовка, 2-4 февраля 1983 г.). Тез. докл. Черноголовка, 1983. С.13-17.
4. Гуляева Л.Ф., Гришанова А.Ю., Громова О.А., Слынько Н.Н., Вавилин В.А., Ляхович В.В. Микросомная монооксигеназная система живых организмов в биомониторинге окружающей среды: Аналитический обзор. Новосибирск: Наука, 1994. 100 с.
5. Козловская В.И., Чуйко Г.М., Мензикова О.В., Подгорная В.А. // Биология внутренних вод. Информ. бюл. СПб.: Наука, 1996. №100. С.65-72.
6. Котелевцев С. В., Степанова Л.И., Пономарева Л.В., Бейм A.M. // Второй симпозиум по экологической биохимии рыб (Ростов Великий, декабрь 1990). Тез. докл. Ярославль, 1990. С.129-130.
7. Браун А.Д., Моженок Т.П. Неспецифический адаптационный синдром клеточной системы. Л.: Наука, 1987. 232 с.
8. Маляревская А.Я. // Теоретические проблемы водной токсикологии: норма и патология. М.: Наука, 1983. С.99-105.
9. Маляревская А.Я. // Гидробиол. журн. 1985. Т.21. №3. С.70-80.
10. Хлебович В.В., Бергер В.Я. // Журн. общ. биол. 1975. Т.36. №1. С.11-25.
11. Определитель пресноводных беспозвоночных Европейской части СССР. Л.: Гидрометеоиздат, 1977. 512 с.
12. Брагинский Л.П. // Пробл. анал. химии. 1977. Т.5. С.27-38.
13. Строганов Н.С.// Методики биологических исследований по водной токсикологии. М.: Наука, 1971. С.14-60.
14. Перечень предельно допустимых концентраций и ориентировочно безопасных уровней воздействия вредных веществ для воды рыбохозяйственных водоемов. / Комитет РФ по рыболовству. М.: Мединор, 1995. 220 с.
15. Справочник предельно допустимых концентраций вредных веществ в пищевых продуктах и среде обитания. М., 1993. 142 с.
16. Колупаев Б.И. // Гидробиол. журн. 1989. Т.25. №5. С.52-54.
17. Сазонова В.Е., Панькина Е.В. // Вод. ресурсы. 1998. Т.25. №1. С.80-84.
18. Туманов А.А., Постнов И.Е., Осипова Н.И., Зимин А.Б. // Методы биотестирования вод. Черноголовка: ОИХФ, 1988. С.104-106.
19. Бергер В.Я. // Вопросы теории адаптации. Труды Зоол. ин-та АН СССР. Л., 1987. Т.160, С.13-30.
20. Голиков А.Н., Голиков Н.В. // Вопросы теории адаптации. Труды Зоол. ин-та АН СССР. Л., 1987. Т.160. С.4-12.
21. Филенко О.Ф. Некоторые универсальные закономерности действия химических агентов на водные организмы: Автореф. дисс. д-ра биол. наук. М.: МГУ, 1990. 36 с.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОЦЕНКИ ТОКСИЧНОСТИ ЗАГРЯЗНИТЕЛЕЙ ВОД ДАЛЬНЕВОСТОЧНЫХ МОРЕЙ | 2001 |
|
RU2215290C2 |
| СПОСОБ ОЦЕНКИ ТОКСИЧНОСТИ ЗАГРЯЗНИТЕЛЕЙ ВОД ДАЛЬНЕВОСТОЧНЫХ МОРЕЙ | 2001 |
|
RU2220415C2 |
| СПОСОБ ОЦЕНКИ ТОКСИЧНОСТИ КОМПОНЕНТОВ СРЕДЫ АЗОВСКОГО И ЧЕРНОГО МОРЕЙ | 2013 |
|
RU2519070C1 |
| СПОСОБ БИОТЕСТИРОВАНИЯ ТОКСИЧНОСТИ ВОДНОЙ СРЕДЫ | 2011 |
|
RU2462707C1 |
| ТЕСТ ДЛЯ ИНТЕГРАЛЬНОЙ ОЦЕНКИ СОСТОЯНИЯ ЗАГРЯЗНЕНИЯ МОРСКОЙ И ПРЕСНОЙ ВОДЫ | 1997 |
|
RU2131925C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ТОКСИЧНОСТИ КОНТАКТНОЙ ЗОНЫ "ГРУНТ-ВОДА" | 2007 |
|
RU2358264C2 |
| Способ определения токсичности водных сред | 1985 |
|
SU1328756A1 |
| СПОСОБ ОЦЕНКИ ТОКСИЧНОСТИ ЗАГРЯЗНИТЕЛЕЙ ВОД АЗОВО-ЧЕРНОМОРСКОГО БАССЕЙНА | 2013 |
|
RU2563283C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ТОКСИЧНОСТИ ПОЧВЫ МЕТОДОМ БИОТЕСТИРОВАНИЯ С ИСПОЛЬЗОВАНИЕМ РАВНОРЕСНИЧНЫХ ИНФУЗОРИЙ PARAMECIUM CAUDATUM EHRENBERG | 2011 |
|
RU2482478C2 |
| Способ определения токсичности промышленных сточных вод | 1989 |
|
SU1751670A1 |

Изобретение относится к области водной токсикологии и санитарной гидробиологии и может быть использовано для оценки токсичности воды при биологическом тестировании сточных и природных пресных вод. Способ предусматривает помещение групп моллюсков (Viviparus viviparus L.) в исследуемую (опыт) и чистую (контроль) воду, препарирование у них по истечении времени экспозиции печени, гомогенизацию извлеченных органов, получение экстракта белков печени и определение в нем активности гидролитических ферментов - кислой фосфатазы или дезоксирибонуклеазы (ДНКазы). За показатель токсичности тестируемой воды принимают достоверное изменение общей активности тест-фермента или хотя бы одно качественное изменение в составе его множественных форм в опытном варианте по сравнению с контролем. Способ, характеризуясь экспрессивностью и универсальностью в отношении различных токсикантов, позволяет обнаруживать загрязнение воды опасными веществами в концентрациях на уровне 0,1-100 ПДК и выше. 9 табл., 5 ил.
Способ определения токсического загрязнения сточных и природных пресных вод, предусматривающий измерение показателей ферментативной активности у водных организмов, предварительно выдержанных в чистой (контроль) и исследуемой (опыт) воде, и отнесение тестируемой воды к токсичной в случае достоверного изменения общей активности фермента или при наличии хотя бы одного качественного изменения в его электрофоретическом спектре в опытном варианте по сравнению с контрольным, отличающийся тем, что в качестве водных организмов используют моллюсков Viviparus viviparus L., а в качестве тест-ферментов - дезоксирибонуклеазу и/или кислую фосфатазу.
| Способ определения токсичности промышленных сточных вод | 1989 |
|
SU1751670A1 |
| Способ определения токсичности водных сред | 1984 |
|
SU1270699A1 |
| Способ определения токсичности водных сред | 1985 |
|
SU1328756A1 |
| СПОСОБ ОЦЕНКИ ТОКСИЧНОСТИ ХИМИЧЕСКИХ ВЕЩЕСТВ ДЛЯ ВОДНЫХ ОРГАНИЗМОВ | 0 |
|
SU381336A1 |
| Способ определения токсичности водных сред | 1982 |
|
SU1112276A1 |
| Способ определения содержания меди в водной среде | 1991 |
|
SU1835234A1 |
| БИОЛОГИЧЕСКИЙ СПОСОБ ОПРЕДЕЛЕНИЯ СТЕПЕНИ ОБЩЕЙ ТОКСИЧНОСТИ И ОСНОВНЫХ ТОКСИКАНТОВ ВОДНОЙ СРЕДЫ (ВАРИАНТЫ) | 1996 |
|
RU2110067C1 |
| ЭКСПРЕСС-СПОСОБ БИОТЕСТИРОВАНИЯ ПРЕСНЫХ ВОД "ПОВЕДЕНЧЕСКИЕ РЕАКЦИИ МОЛЛЮСКОВ" ("ПРМ-ТЕСТ") | 1992 |
|
RU2082167C1 |

